Volume 12 Number 24 1984 Nucleic Acids Research MoleculaT (Honing and carboxyl-propeptide analysis of human type III procollagen Helen R.Loidl 1 , Jane M.Brinker 1 , Mary May 2 , Taina Pihlajaniemi 3 , Scott Morrow 2 , Joel Rosenbloom 2 and Jeanne C.Myers 1 -" •Connective Tissue Research Institute and 'School of Dental Medicine, University of Pennsylvania, Philadelphia, PA 19104, JDept. Biochemistry, Rutgers Medical School, Piscataway, NJ 08854, and 4 Dept. Medicine and Human Genetics, University of Pennsylvania, Philadelphia, PA 19104, USA Received 10 September 1984; Accepted 14 November 1984 ABSTRACT Two human cDNA libraries prepared from normal fibroblast (GM3348) and rhabdomyosarcoma (CCL136) mRNAs were screened under cross hybridization conditions with a genomic fragment coding for exons 2 and 3 of the avian type III procollagen C00H-propept1de (Yamada, Y., Mudryj, M., Sullivan, M. and deCrombrugghe, B. (1983) J. B1ol. Chem. 258, 2758-2761). One cDNA clone containing a 1.12 kb Insert was Isolated from the CCL136 l i b r a r y and later used to Identify a GM3348 derived clone with a 2.4 kb Insert. Comparison of the human and avian type III C-term1nal propeptides revealed much more divergence 1n the first 54 amino adds following the terminal cysteine of the triple helical region than 1s present 1n the a l ( i ) and a2(I) procollagen chains of these species. Analysis of poly (A + ) RNA from normal human fibroblast and tumor cell lines showed that they differed greatly 1n the relative amounts of al(l), a2(l), and al(lll) mRNAs. Furthermore, as we previously reported for the al(i) and a 2(1) transcripts, multiple mRNAs also hybridize to the cloned a l ( I I I ) DNA. INTRODUCTION Five major and at least five minor types of collagen comprise the most abundant protein family found 1n vertebrates (reviews 1,2; and ref. 3-7). In general, collagen 1s synthesized in the form of procollagen molecules composed of three polypeptide chains consisting of a central triple helical domain with the repeating Gly-X-Y sequence, and shorter carboxy and amino propeptides. The different chains are the products of probably more than fifteen genes which have recently begun to be Isolated from several species (reviews 8,9; and ref. 10-17). Determination of the DNA sequences of the genes and their encoded amino add sequences are fundamental to under- standing the function of the various types of collagen and the factors controlling their synthesis. Previously, human cDNA clones coding for the al and a2 chains of the type I procollagen heterotrimer (pro al >2 (pro a2) were Identified (18,19) and used to Isolate overlapping genomic fragments (20,21). Analysis of these two genes, which are located on different chromosomes (22), showed that the coding region 1s frequently Interrupted © IRL Press Limited, Oxford, England. 9383 by guest on August 24, 2016 http://nar.oxfordjournals.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Volume 12 Number 24 1984 Nucleic Ac ids Research
MoleculaT (Honing and carboxyl-propeptide analysis of human type III procollagen
Helen R.Loidl1, Jane M.Brinker1, Mary May2, Taina Pihlajaniemi3, Scott Morrow2, JoelRosenbloom2 and Jeanne C.Myers1-"
•Connective Tissue Research Institute and 'School of Dental Medicine, University of Pennsylvania,Philadelphia, PA 19104, JDept. Biochemistry, Rutgers Medical School, Piscataway, NJ 08854, and4Dept. Medicine and Human Genetics, University of Pennsylvania, Philadelphia, PA 19104, USA
Received 10 September 1984; Accepted 14 November 1984
ABSTRACT
Two human cDNA l i b r a r i e s prepared from normal f i b r o b l a s t (GM3348) andrhabdomyosarcoma (CCL136) mRNAs were screened under cross h y b r i d i z a t i o ncond i t i ons w i th a genomic fragment coding f o r exons 2 and 3 o f the aviantype I I I procol lagen C00H-propept1de (Yamada, Y . , Mudryj , M., S u l l i v a n , M.and deCrombrugghe, B. (1983) J . B1o l . Chem. 258, 2758-2761). One cDNAclone con ta in ing a 1.12 kb I n s e r t was I s o l a t e d from the CCL136 l i b r a r y andl a t e r used to I d e n t i f y a GM3348 der ived clone w i t h a 2.4 kb I n s e r t .Comparison o f the human and avian type I I I C-term1nal propept ides revealedmuch more divergence 1n the f i r s t 54 amino a d d s f o l l ow ing the terminalcys te ine o f the t r i p l e h e l i c a l region than 1s present 1n the a l ( i ) anda 2 ( I ) proco l lagen chains of these spec ies. Ana lys is of poly (A+) RNA fromnormal human f i b r o b l a s t and tumor c e l l l i n e s showed tha t they d i f f e r e dg r e a t l y 1n the r e l a t i v e amounts of a l ( l ) , a 2 ( l ) , and a l ( l l l ) mRNAs.Furthermore, as we p rev ious ly repor ted f o r the a l ( i ) and a 2(1)t ranscr ip ts , mul t ip le mRNAs also hybridize to the cloned a l ( I I I ) DNA.
INTRODUCTION
Five major and at least f i ve minor types of collagen comprise the most
abundant protein family found 1n vertebrates (reviews 1,2; and ref. 3-7).
In general, collagen 1s synthesized in the form of procollagen molecules
composed of three polypeptide chains consist ing of a central t r i p l e hel ical
domain with the repeating Gly-X-Y sequence, and shorter carboxy and amino
propeptides. The d i f fe rent chains are the products of probably more than
f i f teen genes which have recently begun to be Isolated from several species
(reviews 8,9; and ref . 10-17). Determination of the DNA sequences of the
genes and thei r encoded amino add sequences are fundamental to under-
standing the function of the various types of collagen and the factors
cont ro l l ing the i r synthesis. Previously, human cDNA clones coding for the
al and a2 chains of the type I procollagen heterotrimer (pro al >2 (pro a2)
were Ident i f ied (18,19) and used to Isolate overlapping genomic fragments
(20,21). Analysis of these two genes, which are located on d i f fe ren t
chromosomes (22), showed that the coding region 1s frequently Interrupted
© IRL Press Limited, Oxford, England. 9383
by guest on August 24, 2016
http://nar.oxfordjournals.org/D
ownloaded from

Nucleic Acids Research
by about 50 sometimes very la rge Int rons (20,21) . Although the mRNA
t ransc r i p t s of these genes are normally found at a two to one r a t i o
r e f l e c t i n g the type I collagen subunit composit ion, the leve ls can vary 1n
disease states (23) .
To study the coordinate and d i f f e r e n t i a l expression of several
procol l agens, we constructed two cDNA l i b r a r i e s contain ing sequences
corresponding to d i f f e r e n t types of procollagen 1n add i t ion to type I .
Here we report the I s o l a t i o n and charac te r i za t ion of cDNA clones coding for
about ha l f of the human a l ( m ) procollagen cha in . From the DNA sequence
of these c lones, part of the t r i p l e he l i ca l and 3' untranslated region and
the en t i re C-term1nal propeptide was der ived. While the te lopept lde and
adjacent 51 par t of the C00H-propept1de of avian (24,25) and human type I I I
are more divergent than seen 1n the ccl(I) and a2 ( I ) chains of these
species (26-28) , the major par t of the C00H-propept1de 1s conserved.
Comparison of t h i s domain 1n three human procollagen chains also shows
s i g n i f i c a n t homology In the regions where the cysteine residues and
g lycos lya t ion attachment s i te are loca ted . Together with e luc ida t ing the
s t i l l undetermined ro le o f type I I I , these probes w i l l be valuable 1n
def in ing mutations and regulatory abnormal i t ies 1n genetic and acquired
diseases of connective t i s sue .
MATERIALS AND METHODS
Enzymes and Isotopes
Res t r i c t i on endonucleases were purchased from New England Bio labs, DNA
polymerase I and proteinase K from Boehringer Mannheim, T4 DNA Ugase and
rabb i t r e t i cu locy te lysate from Bethesda Research Laborator ies. A l l
enzymes were used according to manufacturers suggestions. Formamide was
obtained from Fluka, o l i go dT ce l l u l ose and primers for DNA sequencing from
Col laborat ive Research, and n i t r o c e l l u l o s e f i l t e r s from Schleicher S
Schuei l . Labeled Isotopes were purchased from Amersham. Cell l i n e 013348
was obtained from the Human Genetic Mutant Cell Repository, Camden, New
Jersey, and CCL136 and HT1080 from the American Type Culture C o l l e c t i o n ,
Rockv i l le , Maryland.
RNA P u r i f i c a t i o n and cONA Cloning
Poly (A+) RNA was Iso lated by o l i go dT ce l lu lose chromatography and
size f rac t iona ted on a sucrose-SDS gradient (19). F i r s t strand DNA
synthesis was performed 1n a 150 ul react ion contain ing 5ug of mRNA, 0.7ug
o l i go dT 12-18 primer, 1.30mH dGTP, .53CTM TTP, .32 mM dCTP, .27mM dATP and
55 uni ts /ml of AMV reverse t ranscr ip tase p u r i f i e d by DEAE and CM sepharose
9384
by guest on August 24, 2016
http://nar.oxfordjournals.org/D
ownloaded from

Nucleic Acids Research
chromatography (29). Second strand DNA was synthesized with the Klenow
fragment of DNA polymerase I after heating the reaction mix at 100° for 2
minutes (30). Following Incubation for 1 hr. at 38*, the reaction was
passed through a G50 sephadex column and the peak fractions were ethanol
precipitated. The double-stranded DNA (lOOng) was treated with 3500
units/ml of Sj nuclease (Miles) at 25* for 30 minutes, and then size
fractionated on a 5-20% neutral sucrose-SDS gradient. The pool containing
the largest DNA was dC-ta1led for 6 minutes at 11° with terminal
transferase. 3ng of DNA was annealed with 5ng of pBR322 which had been
cleaved with PstI and dG-ta1led (New England Nuclear). The hybrid
molecules were used to transform Escher1ch1a col l strain MC1061 (31).
Screening of the Bacterial Colonies
The clones were replica plated onto nitrocellulose f i l t e r s , grown for
four hours and then the plasmid DNA was amplified by overnight Incubation
of the f i l t e r s at 37* on Luria plates containing 12.5ug/ml tetracycline and
150 ug/ml chloremphenicol. The f i l t e r s were processed (30), baked at 76*
for 2 hours, and hybridized under low stringency (35% formaraide, 6xSSPE,
34* for 40 hours) or high stringency (50% forniamide, 5xSSPE, 40* for 20
hours). Final washing conditions were lxSSC, 55° or .2xSSC, 65°
respectively. The DNA probe used for Isolating the 1n1tal human type I I I
clone was lOng/ml of the Insert from a subcloned genomic fragment
containing exons 2 and 3 coding for part of the C00H-propept1de of avian
type I I I procollagen (24,25) provided by Y. Yamada. The human type I I I
cDNA clone, E6, was Isolated using 2ng/ml of the 5' PstI restr ict ion
fragment of RJ5 which was Isolated by electroel ution from an agarose gel.
Inserts and plasmids were nick translated with 32p nucleotides to a
specific act iv i ty of 4-8xlO8 cptrt/ug.
DNA Sequencing
Plasmid DNA was restricted with PstI , or PstI and EcoRI and the
fragments were Isolated by electroel ution after electrophoresis 1n 1%
agarose gels. The DNA was extracted with phenol /chloroform and then cloned
Into the phage M13mp8,9 system (32). Restriction fragments derived from
the cDNA Inserts, now contained 1n the recombinant phage, were sequenced by
the Sanger dideoxy method using a universal primer of 17 nucleotides.
Northern Blot Hybridization
Electrophoresis of poly (A+) RNA 1n .7% agarose, 2M formaldehyde gels,
transfer to nitrocellulose and hybridization conditions were essentially as
described (19,33).
9385
by guest on August 24, 2016
http://nar.oxfordjournals.org/D
ownloaded from

Nucleic Acids Research
. . ( IH)Uon jTI, COOH-PROPEPTIDE .3 '
5' i_
Figure 1 . Restr ic t ion Map of Human Type I I I cOHA Clones.The regions of the pro al ( I I I ) chain encoded by the cDNA clones are
shown on the top of the f igure. RJ5 terminates 5' at residue 892 of thet r i p l e hel ical region (35) and by comparison E6 extends to about 592. Onthe 3' side, RJ5 1s missing the last 30 amino adds of the COOH propeptidewhich have been derived from E6. The human type I I I cDNA clones, RJ5 andE6, were Independently analyzed for the presence of endonucleaseres t r i c t ion s i tes . Al l sites localized 1n RJ5 by res t r i c t i on mapping werealso found at the same position 1n E6: EcoRI, Ps t I , Aval/Xhol, Aval, andPvuII. Other enzymes that were determined negative with both clones wereBamHI, Xbal, H i n d l l l , EcoRV, Smal, C la l , Sa i l , and B g l l l . The 51 1100 bpand 3' 930 bp PstI fragments of E6 were separately hybridized to Southernblots of RJ5 cleaved with the di f ferent res t r i c t i on enzymes in order tocompletely ver i fy the Ident i f icat ion and regions of overlap. Both clonesare Inserted Into the PstI site of pBR322 1n the u or ienta t ion. The bottompart of the f igure shows the strategy for the DNA sequencing of RJ5 and E6.Both strands of the four fragments of RJ5, 51 Ps t I , middle Ps t I , and 3'Pstl-EcoRI and EcoRI-PstI were completely sequenced. The las t 90nucleotides coding for the C00H-propept1de and 78 nucleotides of the 3'untranslated region were determined from the 3' EcoRI-PstI fragment of E6.
RESULTS
Isolat ion of cONA Clones
Two sources of mRNA were investigated for the cloning of human type I I I
procollagen sequences. One was a normal f ibrob last cel l l i ne , GM3348, and
the other a human rhabdomyosarcoma cel l l i n e , CCL136, that was reported to
synthesize type I I I , and to a lesser extent types IV and V (34). Poly (A+)
RNA was size fract ionated and the 28-35S RNA analyzed by ce l l free
translat ion for the presence of type I I I procollagen. The gradient
f ract ion giving the highest y ie ld of a protein that was sensitive to
bacterial collagenase and which was s l i gh t l y larger 1n size than an a l ( I )
standard by polyacrylamide gel electrophoresis was used as template for AMV
reverse transcMptase. Double stranded cDNA was prepared and Inserted Into
the PstI s i te of the plasmid PBR322. Transformation of the recombinant
molecules was carried out using E. co l l s t ra in MC1061 (31) which gave an
eff ic iency of 0.5- lx lO8 colonies per ug of supercoiled pBR322. The I n i t i a l
screening of approximately 8000 colonies under cross hybridization
9386
by guest on August 24, 2016
http://nar.oxfordjournals.org/D
ownloaded from
Nucleic Acids Research
I '<L
al(m) al(I) a2(I)A B C
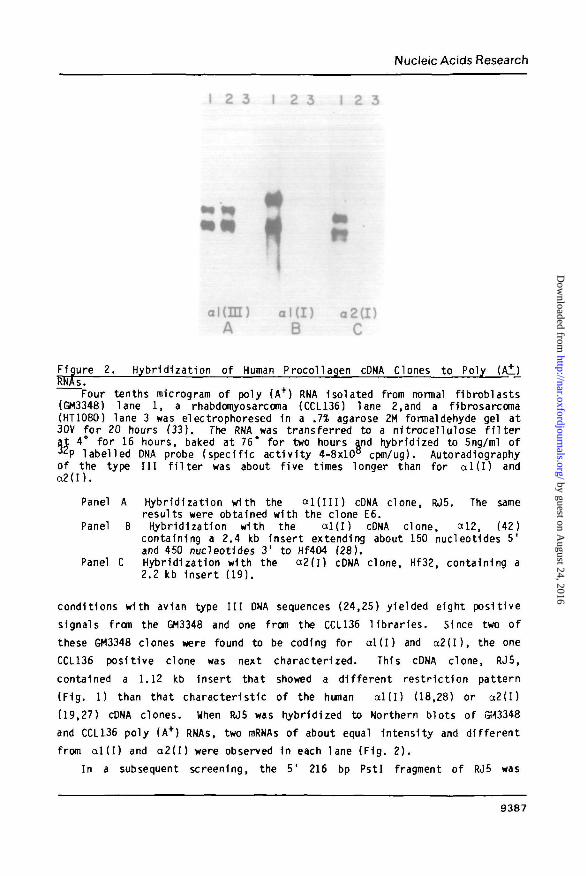
Figure 2. Hybridization of Human Procoilagen cDNA Clones to PolyRNAs.
Four tenths microgram of poly (A+) RNA Isolated from normal fibroblasts(GM3348) lane 1, a rhabdomyosarcoma (CCL136) lane 2,and a fibrosarcoma(HT1080) lane 3 was electrophoresed 1n a .1% agarose 2M formaldehyde gel at30V for 20 hours (33). The RNA was transferred to a nitrocellulose f i l t e rat 4° for 16 hours, baked at 76* for two hours and hybridized to 5ng/ml of32P labelled DNA probe (specific act iv i ty 4-8xlO8 cpm/ug). Autoradiographyof the type I I I f i l t e r was about f ive times longer than for a l ( I ) ando2( I ) .
Panel A Hybridization with the a l ( I I I ) cDNA clone, RJ5. The sameresults were obtained with the clone E6.
Panel B Hybridization with the ccl(i) cDNA clone, a 12, (42)containing a 2.4 kb insert extending about 150 nucleotides 5'and 450 nucleotides 31 to Hf404 (28).
Panel C Hybridization with the a2( i) cDNA clone, Hf32, containing a2.2 kb Insert (19).
conditions with avian type I I I DNA sequences (24,25) yielded eight positive
signals from the GM3348 and one from the CCL136 l ibrar ies. Since two of
these GM3348 clones were found to be coding for a l ( I ) and a2( I ) , the one
CCL136 positive clone was next characterized. This cDNA clone, RJ5,
contained a 1.12 kb insert that showed a different restr ict ion pattern
(F1g. 1) than that characteristic of the human a l ( I ) (18,28) or a2(I)
(19,27) cDNA clones. When RJ5 was hybridized to Northern blots of S43348
and CCL136 poly (A+) RNAs, two mRNAs of about equal Intensity and different
from a l ( I ) and a2(I) were observed in each lane (Fig. 2).
In a subsequent screening, the 5' 216 bp PstI fragment of RJ5 was
9387
by guest on August 24, 2016
http://nar.oxfordjournals.org/D
ownloaded from

Nucleic Acids Research
( U i u ACT i t c CCT H T CCC CCT K I CCT K l CCC CCT CCT K l ( 1 1 TCC C U CCT CCT CCI CCT CCT C U CCC C M CCI CCT CAC AAACLY CLI SER CL1 PRO U t O.Y PRO ALA CLY AlA PIO CLT PRO ALA 6LT SCR ARC CLT ALA PRO CLY PRO CLR CLT PRO ARC CLY ASP LTS
«J »51U I CAA ACA U T U A CCT CCA CCT CCT U C ATC AA> U A CAT CCA WA TTC CCT H T AAT CCA CCT CCC CCA CtT TCT C O CCC CCT CCTCLT CLU THA GLT CLI ARG CLY ALA ALA CLT ILE LTS CLT HIS ARC CLT PHI PRO GLT AS! PRO CLT ALA PRO CLT SCR PRO CLT PRO ALA
PRO
til MlCCT CAC CAG CGT CCA ATC CCC ACT CCA CCA CCT CCA GCC CCC ACA CCA CCT CTT CCA CCC ACT CCA CCT CCT CCC AAA CAT CCA ACC AGTCtT U I C L I CL< ALA H E U.1 SCR PRO CLT PRO ALA t L I PRO ARC CLT PKO »AL CLT PRO SCR 111 PRO H O CLT LTS ASP U T THR SCR
ALA THR
M; ionCCA CAT CCA CCT CCC ATT CCA CCA CCA GCC CCT CCA CCT AAC AGA CCT CAA ACA C W TCT CAC CCC TCC CCA CCC CAC CCA CCC CAA CCACLT HIS PRO CLT PRO ILE CLT PRO PflO CLY PKO AJIG CLT AS! ARC CLT CLU ARC CLT SCR CLU CLT SER PRO CLY HIS PRO CLT CLH PRO
SCR
1 0 1 * 1013GCC CCT CCT CCA CCT CCT CCT CCC CCT U T CCT TCCGLT PRO PRO CLT PRO PRO CLY ALA PRO CLT PRO CTS
•
Figure 3. The DMA Sequence Encoding 132 Amino Adds of the Triple HelicalRegion of Human Type I I I Procollagen.
The protein sequence obtained from the DNA sequence of the type I I Iclones, RJ5 and E6, was compared with the published human type I I I proteinsequence derived by Edman degradation (35). Only four differences werefound and they are shown on the bottom l ine . These occur at residues 924,980, 981 and 983.
puri f ied and hybridized under stringent conditions to the same set of
f i l t e r s and to about 4000 new colonies from a later transformation. Eight
positive clones were Identif ied and the largest clone, and the one
extending most 51 , E6, was later characterized by restr ict ion mapping (Fig.
1) and Southern blot hybridization. E6 contains a 2.4 kb Insert that
overlaps RJ5 about 400 bases 3' and 900 bases 5 ' , and therefore entirely
encompases the original Isolate. The Inserts of the other type I I I clones
terminate 3' to the end of E6 as does one other positive from the original
eight GM3348 clones and these were not examined further.
Comparison to Human Amino Add Sequence Determined by Protein Sequencing
The 5' 216 bp PstI fragment of RJ5 contained entirely Gly-X-Y sequences
as did 180 nucleotides of the 5' region of the adjacent 382 bp PstI
fragment. Immediately at the 3' end of the 180 nucleotides were found two
adjacent codons, TGC and TGT, that designate the cysteine residues
responsible for formation of the disulf ide bonds of type I I I collagen.
According to the published amino add sequences of type I I I , the t r i p le
helical region contains 1023 residues terminating at the 3' end with the
f i r s t of the double cysteines (35). Therefore, the most 5' amino add
encoded by RJ5 1s residue 892 (F1g. 3) and by comparison E6 extends to
about 592.
Only a few differences were apparent when we compared the type I I I
sequences derived from the clones with the reported human amino acid
sequence determined by Edman degradation (34): at 924 a threonine versus
9388
by guest on August 24, 2016
http://nar.oxfordjournals.org/D
ownloaded from

Nucleic Acids Research
CC I I I C CTC CS A I CCA Tt CSC 1*1 tCH LAA C C AA AICI C*T CST t i l CCA U C K I CCC »TI CCI O l MT U i H I W U l d T d «ST TT7 CCC C t l TAJ-TAT CU UI Ui CU ATC CATCIS (Lr CLT I U CLT ALA ALA ALA III I I I 6LI I U t i l UT CII ITJ » U t i l CIT H I ALA MO [11 ,111 U l . t t f CLI PI© HEI AW• U.1 TU U l LEB UT A A A B.T • » TU UT TTI B.I • £ • AMI I I I UB
U K C CTC C A C r i s A * T C U C « T M A C A A A GITC AAA AIC AAC ACC tAT | « ATI ATC ACT ICA CTC A M TCT CCI AAI U A t U ATA |AA A K CTC ATI A l l CCT CAT U T UT t i l AJUMI US III U l THI UP U l HE «T TKI U l LEI I U HI >U U l 111 U l !U U l HI IEI HE HI Pt© UP UT HI All IUAM CLI Ltl U l IEI «T HE AH IU ll£ l£i
il n
U C LCC CCT A*A AAC TCC MA tAC CT« AAA TTC T « CAT CCI I U CTt AJU U T CtA tAA TAC I t t S l T t « CCI AAC CAA u i I K AAAA H Fttt ALA A l l AW C I I A l t ISP L [ | ITJ PHt CTS HIS m t L I LEI LTI U l U l U f l IT1 H M U U T W A U U l ttt CTJ I U
* • HE +
* A I » I A T ( C C J C j £ c C C A A116 *AT CCI AIC AAI 6IA U C U T AAT *T* CAA ACI U t 1AA ACA I K AIA ACT CCC AAT CCI TT1 JUT %T1 CCA C M AAA O U TCfi T ULEI AIP « .A lit LTJ I.I HI CTJ U l V I U l TW1 U.T U l H I CTI HE S I U * U l Ml U l AM »U « • UC LTI IIJ TIP TlJ" • ' TTI * * Ltl » u TW AM
111 IMT A C C A A A C t A I A I A 6 CC I A C CC
A U CAT ICT ACT CCT U C AAC AAA CAC GIT TCC TTT C U U C IXC ATC CAT GCT CCI I I I CAC I I I I U I U MC M l CCT fcU CTI CCT
THI CLI U l U.T LTS HI UP Alt
Kl INC I C A T C C A C IC CC «C
(UA CAT U C CTT U T CTC CAC CTC CCA TTC CIT C M CTT CTC TCC * « CCA CCT I U CAC AAC ATC ACA TAT CAC I « AAA AAT A K ATTt i e U P m LEI ASP m u i IEI ALA P«E LEO U C LEI LEI U I M I U I ALA H I U I m i n m TTI HII e n LTI ASI H I ILE
SEI CLI ILE •
in noC A A C C T A A A C I O T I A C I A C
OCA TAC ATI SA1 CAI « C AIT «** AAT «TA AAG AAG GCC CTG AAG CTG ATC ttt Tt* U T I U U I O U TIC AAt K I tAA I U AAT A U
U k U l T U T i l U P HE
111 1 «A T C T G T * C T T I C T I C A T A A A
AAA TTC ACI TAC ACA t ! T CTC U C CAT GCT TIC ACS AAA CAC ACT CCC CAA TCC ACC AAA ACA CTC ITT CAA TAT CCA ACA CCC A U tCTU S ME T O TTI m U L LEI U . I U P CLT CTS THI ITS I I S T i l Ctl U l TIP U l LTS TO T U P>E UQ TTI A1G THI A1G LTS U l
TTI Hit ALA * U T T U
t*[ tnA » T * * T A T A T C t ( C C
I » L A H L E I P I O I U T U I V ILE ALA P H T T I A S * ILE U T U T P t O U P t i l U S P * U T * U W T U U . I P H T U C T I P I C I E !PUT T U ILE *
I ( A T CC K T C T I T I C ACC CC K AAC t A T C CIC CAA I t TtC TCC I I C I t i C C I IIAA ACC A U C T t TAT C T I AAA TCC C U CAA A U AAA TTT AAC TCC ATA I * f t I T CCI CTT 6 1 1 CIA ATC T T I ICA ACC
Figure 4. Comparison of the Human and Avian a l ( I I I ) Telo- and C00H-P ropeptide Regi ons~ ~~
The middle two lines are the nucieotides and corresponding amino addsencoded by the human type I I I clones. The nucleotide differences found 1nthe avian are shown on the top Hne, and where these designate a differentamino add 1s shown on the bottom l ine. Amino add 1 1s the most 3' of thedouble cysteines (the 5' cysteine 1s residue 1023 of the t r i p le helicalregion shown 1n figure 2). The symbols Indicate the deletion A 1n the
avian sequence of three amino adds, Insertion v of one amino a d d , thecysteine residues*, and the cleavage site + between the telo- andC00H-propept1de, and the glycosylation attachment site which is underlined.The last 78 nucleotides are the beginning of the 3' untranslated region.
proline, at 980 a threonine versus alanine, at 981 a serine vs. threonine,
and at 983 a h1st1d1ne vs. serine. The remaining 128 amino adds were
Identical.
Analysis of the Telo- and Carboxy- Propeptides of a l ( I I I ) , a2(I ) and a l ( I )
Alignment of the human and chick type I I I (24,25) sequences beyond the
collagenous region revealed considerable dissimi lar i ty 1n the f i r s t 54
amino adds (Fig. 4) . In the human, three amino adds (Gly I le Gly)
corresponding to residues 11-13 were Inserted and one amino add (Glu)
between the two tryosines at positions 23 and 24 was deleted. Within the
telopeptide (cleavage Is presumed to occur between residues 25 and 26), 12
9389
by guest on August 24, 2016
http://nar.oxfordjournals.org/D
ownloaded from

Nucleic Acids Research
a l ( l l l ) PIO ASP « l l SO AtC ITS Ai« PI0 ALA AM * U CIS UE AIT LCI ITS PHI CIS HIS PIO CLU LEU LIS SCI BLI SLUa l l I) - CLU IK« - - - - U ( LEU SE« - - - IIP S£l - - T i la l l l l - CLU - Tut THI KIT - - SCI ASP TIP -
* *
tl IMa l l I I I ) T l * TIP »AL ASP P«0 AS! U.1 S l l CIS LIS LCU U P ALA 1LC LTS »Al PDC CIS AS! RET U.U Till 6LI U.% TNI CTSa l ( l ) - - III TKI MT CIO - - - - T i l - ASP PHI P«0all l l - lie »si
* • *
B i« 1 M
AlA H I Oi l A i l I H T i l T i l HIS CIS LTS ASK S t l ILC ALA T i l HIT ASPa l l I I I ) K T TCC CM T J O n t l TAT CAC TCC AAA AAT ACC ATT ICA TAC «T0 SATa i l I) --c --T c —c c --ca i m --c -c --c s --c -— 8-e --e c
«AL•
Figure 5. Conserved Regions 1n the COOH Propeptide of Human o t l ( I I I ) ,a2(I) and a_KI).
Figure 5A shows residues 55-106 of the COOH propeptide of type I I Iprocoiiagen (top l ine) and the corresponding region of a2(I) (27) anda l ( i ) (28). Dashes Indicate no difference from the type I I I amino addsequence. Asterisks designate the cysteines found 1n c t l ( I I I ) .
Figure 5B shows residues 168-184 of the C00H-propept1de Including thegiycosylation attachment site (underlined). The two top lines show thetype I I I amino add and nucleotide sequence. The two bottom lines are thenucleotide sequences of the a2( I ) (27) and a l ( I ) (28) cDNA clones.Dashes Indicate no change from the type I I I DMA sequence. Only one change1n the amino acid sequence 1s noted: Val at position 180 1n the ocl(I).
differences were observed between chick and human and one at the
propeptidase cleavage site Involved a charge change, Arg to Gly. In the
region containing residues 26 to 54, 13 additional changes occurred
Including three charge differences, the most notable being a Lys 1n the
human at residue 32 and a Glu 1n the chick. Therefore, a total of 25
changes 1n 54 amino acids (not counting the deletion and Insertion) have
been inst i tu ted. In the corresponding region of the chick (26) and human
a l ( I ) (28) there are only two changes, and eight are present 1n the a2(I)
not Including the four amino acid insertion in the human (26,27). The fact
that the human and avian type I I I genes have diverged more than a l ( I ) or
a2(l) suggests that the lat ter two have evolved more recently or that
changes in this region of ctl ( I I I ) are tolerated.
However, when residues 55-106 were compared, very strong homology
between the type I I I chick and human protein sequence was Immediately
evident (Fig. 4) . Only three amino adds have diverged and none of these
involved a difference in charge. Since 1t seemed Important that the
protein sequence in this area be conserved, we Investigated the comparable
region in the human a l ( I ) (28) and a2(I) (27). As can be seen 1n F1g.
5A, this segment of the three different procollagen COOH-propeptides also
9390
by guest on August 24, 2016
http://nar.oxfordjournals.org/D
ownloaded from

Nucleic Acids Research
shows str iking homology with each other. Interestingly, and perhaps most
signif icant 1s the presence in this domain of the f i r s t f ive of the eight
(seven in a2) cysteine residues 1n a K H I ) , a2( I ) , and a l ( I ) . These
amino adds are Involved 1n both Inter- and 1ntracha1n disulf ide bonds and
thus the assembly of the procollagen molecules (9,36-38). Because of these
functions there 1s apparently a selective pressure for maintaining a
specific conformation.
Previously, other investigators have compared several di f ferent
procollagen chains and found a conserved nucleotide and amino add sequence
surrounding positions 171-173 (Asn He Thr) that designate the
glycosylation attachment site (25). In the DNA sequence coding for amino
acids 170 to 179, there are th i r ty Identical nucleotldes 1n the avian
ocl(III) (25), and human a2(I) (27) and a l ( I ) (28). This homology 1s
probably why we observed hybridization of the chick type I I I genomic
subclone with both human a l ( l ) and cc2(I) mRNAs even at stringent
hybridization and washing conditions (not shown). However, since
hybridization of the avian a l ( H I ) DNA with the human a l ( I I I ) mRNA in the
CCL136 cell l ine was very fa int and inconsistent, we were not surprised to
find that changes in these th i r ty nucleotides of the human type I I I had
occurred. Although four out of th i r ty nucleotides have diverged, the amino
add sequence 1s identical (Fig. 3). This region contains the 6th cysteine
residue. These results Indicate that there does not appear to be
conservation of the DNA sequence as long as the protein is relat ively
unchanged (Fig. 5B).
In the final third of the COOH-propeptide, inclusive of the last two
cysteines, homology between human and avian c t l ( I I I ) (Fig. 4) and human
cd(I) and a2(I) 1s also apparent (25,27,28). Especially notable 1s the
maintenance of the charged residues (Arg, Lys, Glu, Asp). The last f i f teen
amino adds of human and avian a l ( I I I ) are Identical and more l ike the
human a l ( I ) sequence (28) than the o2(I) (27).
Northern Blot Hybridization
Transcription of the human a l ( I ) and a2(I) procollagen genes results
1n multiple mRNAs 1n cultured fibroblasts (18-21). The difference 1n size
(200-1000 nucleotides) is due to the use of different polyadenylation
attachment sequences 1n the 3' untranslated region. To determine whether
this feature also applied to the ocl(II I) gene, we hybridized the cloned
a l ( I I I ) DNA to poly (A+) RNA from normal f ibroblast and rhabdomyosarcoma
cell l ines. As shown 1n Fig. 2A, two mRNAs of about equal intensity are
present. These are similar 1n size to the a2(I) mRNAs which range from
9391
by guest on August 24, 2016
http://nar.oxfordjournals.org/D
ownloaded from

Nucleic Acids Research
5500-6200 nucleotides (Fig. 2C, and ref. 19,20). Whether these dif ferent
procollagen RNA transcripts are related to tissue specif ici ty s t m remains
to be determined. I t 1s unlikely that their occurence 1s fortuitous since
we have recently Identif ied a cDNA clone coding for another human
procollagen chain which also hybridizes to multiple mRNAs. (tyers, J.C. £ t
a i , manuscript in preparation).
Hybridization of a l ( I I I ) , a l ( I ) and a2(I) human procollagen cDNA
clones to poly (A+) RNAs purified from normal and tumor cell lines also
showed a signif icant difference 1n their representation (Fig. 2). Although
a l l three procollagen genes are expressed 1n normal human fibroblasts, we
have never detected a2(I) mRNA 1n the CCL136 tumor cell l ine (Fig. 2C) and
only trace levels of a l ( I ) can be seen after long exposure (Fig. 2B). The
amount of type I I I 1s comparable 1n both the normal and CCL136 cell l ines
(F1g. 2A), and 1n the lat ter vastly exceeds the small amount of a l ( l ) .
None of the three genes seem to be expressed 1n a human fibrosarcoma cell
l i ne , HT1080, which synthesizes type IV procollagen (Fig. 2 and ref. 39).
DISCUSSION
Analysis of the type I I I sequences provides a great deal of information
about regions of the protein that have been maintained between species and
the dif ferent procollagen chains. Comparison of the C-term1nal propeptides
of human and avian a l ( I I I ) shows strong homology after the variable 5'
region, but 1n the three human procollagen chains two divergent and two
conserved regions can be seen. The telopeptide and 5' part of the
C00H-propept1de are divergent but the adjacent 3' region where the f i r s t
f ive cysteines (four 1n a2(D) are located has been essentially
maintained. The second variable region 1s followed by the conserved end of
the C00H-propept1de which Includes the other three cysteines and the
glycosylation attachment s i te. The c r i t i ca l role of this domain 1n
selection and association of the procollagen chains 1s probably responsible
for the conservation of these regions (9,32). Support for this concept
comes from our studies of an osteogenesis Imperfecta patient whose DNA
contains a four nucleotide frameshift deletion 1n the pro a2(I) collagen
gene coding for the end of the carboxy-propeptide (38). Change of the last
33 aroino adds Including the position of the final cysteine prevents
Incorporation of pro a2(I) chains Into the type I procollagen molecule and
results exclusively 1n ( 01)3 homotrimer formation (38,40).
Whereas type I 1s the major constituent of bone and tendon (1,2,9),
type I I I seems to predominate 1n tissues rich in smooth muscle cel ls (41).
9392
by guest on August 24, 2016
http://nar.oxfordjournals.org/D
ownloaded from

Nucleic Acids Research
The role of type I I I collagen has not been clearly defined but in another
Inherited disease of connective tissue, Ehlers Danlos type IV, rupture of
viscera, uterus and large blood vessels 1s attributed to reduced amounts
and/or structural ly abnormal type I I I collagen (9,42). Several type I
procollagen mutations resulting 1n skeletal abnormalities have recently
been defined by the use of a l ( I ) and a 2( I ) DNA clones (37,38,43).
Similar studies with these type I I I cDNA probes, and in the future genomic
clones, w i l l certainly elucidate the function and regulation of type I I I
procollagen.
ACKNOWLEDGEMENTS
We are very g r a t e f u l to Dr. Y. Yamada f o r p rov id ing the avian type I I I
genomic subclones. The encouragement and he lp fu l suggest ions of Drs. N.
Ke fa l i des and E.J . Macarak are deeply app rec ia ted . We also thank M. Mason
fo r e x c e l l e n t t yp ing o f the manuscript and DNA sequences. These s tud ies
were supported by Nat ional I n s t i t u t e s of Health Research Grants AM33348,
AM2O553, DE-02623 and HL-29702.
*To whom correspondence should be addressed:Connective Tissue Research I n s t i t u t e and U n i v e r s i t y of Pennsylvania3624 Market S t r e e t , P h i l a d e l p h i a , PA 19104
REFERENCES1 . B o r n s t e i n , P. and Sage, H. (1980) Ann. Rev. Biochem. 49,957-1003.2. M i l l e r , E .J . and Gay, S. (1982) Methods Enzymol. 84.37J2.3. Burgeson, R.E., Hebda, P.A., Mo r r i s , N.P. « H o l H s T e r , D.W. (1982) J .
B1o l . Chem. 257,7852-7856.4 . Jander , R., 'K l u te rbe rg , J . , and G l a n v i l l e , R.W. (1983) Eur . J .
Biochem. 133,39-46.5. Bentz, H.7~Morr1s, N.P. , Murray, L.W., Saka l , L.Y. H o l l i s t e r , D.W. and
Burgeson, R.E. (1983) Proc. N a t l . Acad. Sc1. USA 80,3168-3172.6. Sage, H., Trueb, B. and Borns te in , P. (1983) J . BTol . Chem.
258,13391-13401.7. UTFson, G . J . , K i e l t y , C M . , Garner, C , Schor, S .L . , and Gran t , M.E.
(1983) Biochem. J . 211,417-426.8. Tate, V . , F i ne r , M . T f o e d t k e r , H. and Doty, P. (1982) Cold Spr ing
Harbor Symp. 47^1039-1049.9. Prockop, D.J. and K1v1r i kko , K . I . (1984) New Eng l . J . Med. 311 ,
376-386.10. Lukens, L .N . , F r i s c h a u f , A .M. , Pawlowski, P . J . , B H e r l e y , G.T. and
Lehrach, H. (1983) Nuc le ic Acids Res. 11,6021-39.11 . Kurk inen, M., Barlow, D.P. , Helfman, D.'HT, W i l l i a m s , J .G. and Hogan,
B.L.M. (1983) Nuc le ic Ac ids . Res. 11,6199-6209.12. Wang, S.Y. , and Gudas, L . J . (1983 )T roc . N a t l . Acad. Sc1. USA
80,5880-5884.13. UTnomuja, Y . , Showal ter , A . , vander Rest, M., Seidah, N.G.. C h r e t i e n ,
M., and Olsen, B.R. (1984) Biochemistry 23,617-624.14. Ninomuja, Y . , and Olsen, B.R. (1984) P r o c T N a t l . Acad. S c i . USA
9393
by guest on August 24, 2016
http://nar.oxfordjournals.org/D
ownloaded from

Nucleic Acids Research
81,3014-3018.15. Strom, CM. and Uphol t , W.B. (1984) Nucleic Acids Res. 12, 1025-1038.16. Sandei l , L . J . , Pren t ice , H.L., Kravis, 0. and Upholt, W7¥. (1984) J .
B i o l . Chem. 259, 7826-7834.17. Yamada, Y. , TTa"u, G., Mudryj , M., Ob ic i , S., and deCrombrugghe, B.
(1984) Nature 31£, 333-337.18. Chu, M.L., Myers, J . C . , Bernard, M.P., Ding, J . F . , and Ramirez, F.
(1982) Nucleic Adds Res. ^£,5925-5934.19. Myers, J . C . , Chu, M.L., Faro, S.H., Clark, H .J . , Prockop, D .J . , and
Ramirez, F. (1981) Proc. Natl . Acad. Sc i . USA 78,3516-3520.20. f lyers, J . C . , Dickson, L.A., de Wet, W.J. , BernaTd", M.P., Chu, M-L.,
D IL ibe r to , M., Pepe, G., Sangiorgi , F.O., and Ramirez, F. (1983) J .B i o l . Chem. 258,10128-10135.
2 1 . Chu, M.-L. , "dT~Wet, W., Bernard M., Ding, J . F . , Morabi to, M., Myers,J .C . , Wi l l iams, C , and Ramirez, F. (1984) Nature 3J£, 337-340.
22. Huerre, C , Junien, C , Weil, D., Chu, M-L, Morabito, M., VanCong, N.,Myers, J . C . , Foubert, C , Gross, M-S., Prockop, D.J . , Boue, J . , Kaplan,J .C . , dela Chapelle, A. , and Ramirez, F. (1982) Proc. Na t l . Acad. Sc i .USA 79_,6627-6630.
23. de Wet, W.J . , Chu, M-L., and Prockop, D.J. (1983) J . B1ol . Chem.258,14385-14389.
24. Tamada, Y. , Mudryj , M., Su l l i van , M. and de Crombrugghe, B. (1983) J .B i o l . Chem. 258,2758-2761.
25. Yamada, Y. , KuFn, K., and de Crombrugghe, B. (1983) Nucleic Adds Res.11,2733-2744.
26. F u l l e r , F. and Boedtker, H. (1981) Biochemistry. 20_,996-1006.27. Bernard, M.P., Myers, J .C. , Chu, M.L., Ramirez, F., Elkenberry, E.F.,
and Prockop, D.J. (1983) Biochemistry 22,1139-1145.28. Bernard, M.P., Chu, M.L., Myers, J .C . , Ramirez, F., Elkenberry, E., and
Prockop, D.J. (1983) Biochemistry 22,5213-23.29. Myers, J . C . , Ramirez, F., Kadan, D.L., Flood, M. and Spiegelman, S.
(1980) Ana l . Biochem. 101,88-96.30. Mania t is , T . , F r i t s c h , TJ., Sambrook, J . (1982) Molecular Cloning: A
Laboratory Manual, Cold Spring Harbor Laboratory, W.3 1 . Casadaban, M.J. and Cohen, S.N. (1980) J . Mol. B i o l . 138,179-207.32. Messing, J . (1983) Methods Enzymol . 101,20-78.33 Thomas, P.S. (1980) Proc. N a t l . A c a d T S d . USA ]]_, 5201-5205.34. Kr ieg , T . , T impl , R., A l i t a l o , K., Kurkinen, M. and Vaher i , A. (1979)
Febs. L e t t . 104,405-410.35. Seyer, J.M. "and'Kang, A.H. (1981) Biochemistry 20_,2621-2627.36. 01 sen, B. and Dickson, L. 1n Chem. and B i o l . Mineral ized Connect. T i ss .
(ed. Ve1s. A.) E lsev ier /Nor th-Hol land, NY (1981) 143-153.37. Dickson, L.A., P1hl a janiemi, T. , Deak, S.B., Pope, F.M., N i cho l l s , A.,
Prockop, D .J . , and Myers, J.C. (1984) Proc. N a t l . Acad. S d . USA, 81_,4524-4528.
38. P1hlajan1em1, T . , Dickson, L.A., Pope, F.M., Korhonen, V.R., N icho l l s ,A. , Prockop, D .J . , and Myers, J.C. (1984) J . B i o l . Chem., 1n press.
39. P1hlajan1em1, T . , M y l l y l a , R., A l i t a l o , K., Vaher i , A. and K i v i r i k k o ,K . I . (1981) Biochemistry 20,7409-7415.
40. Deak, S.B., N i c h o l l s , A . , T o p e , F.M. and Prockop, D.J. (1983) J . B1ol.Chem. 258, 15192-15197.
4 1 . Hayne,~RT, Vail , M.S. and M U l e r , E.J. (1978) Biochemistry 17, 446-452.42. Pye r i t z , R.E., S t o l l e , C.A., Parfrey, N.A. and Myers, J .C. TT984) Amer.
J . Hed. Genet., In press.43. Chu, M.L., Wi l l i ams, C .J . , Pepe, G., H i rsch, J . L . , Prockop, D.J. and
Ramirez, F. (1983) Nature 304, 78-80.
9394
by guest on August 24, 2016
http://nar.oxfordjournals.org/D
ownloaded from
Related Documents











