This article was originally published in a journal published by Elsevier, and the attached copy is provided by Elsevier for the author’s benefit and for the benefit of the author’s institution, for non-commercial research and educational use including without limitation use in instruction at your institution, sending it to specific colleagues that you know, and providing a copy to your institution’s administrator. All other uses, reproduction and distribution, including without limitation commercial reprints, selling or licensing copies or access, or posting on open internet sites, your personal or institution’s website or repository, are prohibited. For exceptions, permission may be sought for such use through Elsevier’s permissions site at: http://www.elsevier.com/locate/permissionusematerial

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

This article was originally published in a journal published byElsevier, and the attached copy is provided by Elsevier for the
author’s benefit and for the benefit of the author’s institution, fornon-commercial research and educational use including without
limitation use in instruction at your institution, sending it to specificcolleagues that you know, and providing a copy to your institution’s
administrator.
All other uses, reproduction and distribution, including withoutlimitation commercial reprints, selling or licensing copies or access,
or posting on open internet sites, your personal or institution’swebsite or repository, are prohibited. For exceptions, permission
may be sought for such use through Elsevier’s permissions site at:
http://www.elsevier.com/locate/permissionusematerial

Autho
r's
pers
onal
co
py
Molecular characterisation of BSR4, a novel bradyzoite-specificgene from Neospora caninum q
V. Risco-Castillo a, A. Fernandez-Garcıa a, A. Zaballos b, A. Aguado-Martınez a,A. Hemphill c, A. Rodrıguez-Bertos a, G. Alvarez-Garcıa a, L.M. Ortega-Mora a,*
a SALUVET. Dpto. de Sanidad Animal, Facultad de Veterinaria, Universidad Complutense de Madrid, 28040 Madrid, Spainb Dpto. de Inmunologıa y Oncologıa, Centro Nacional de Biotecnologıa, Universidad Autonoma de Madrid, Campus de Cantoblanco, 28049 Madrid, Spain
c Institute of Parasitology, Faculties of Veterinary Medicine and Medicine, University of Bern, Langass-Strasse 122, CH-3012 Berne, Switzerland
Received 4 December 2006; received in revised form 26 January 2007; accepted 4 February 2007
Abstract
Here we present the identification and cloning of the NcBSR4 gene, the putative Neospora caninum orthologue to the Toxoplasma
gondii TgBSR4 gene. To isolate NcBSR4, genome walking PCR was performed on N. caninum genomic DNA using the expressedsequence tag NcEST3c28h02.y1 sequence, which shares a 44% identity with the TgBSR4 gene, as a framework. Nucleotide sequencingof amplified DNA fragments revealed a single uninterrupted 1227 bp open reading frame that encodes a protein of 408 amino acids with66% similarity to the TgBSR4 antigen. A putative 39-residue signal peptide was found at the NH2-terminus, followed by a hydrophilicregion. At the COOH-terminus, a potential site for a glycosylphosphatidylinositol anchor was identified at amino acid 379. A polyclonalserum against recombinant NcBSR4 protein was raised in rabbits, and immunolabelling demonstrated stage-specific expression of theNcBSR4 antigen in N. caninum bradyzoites produced in vitro and in vivo. Furthermore, RT-PCR analysis showed a slight increaseof NcBSR4 transcripts in bradyzoites generated during in vitro tachyzoite-to-bradyzoite stage-conversion, suggesting that this gene isspecifically expressed at the bradyzoite stage and that its transcription relies on the switch to this stage.� 2007 Australian Society for Parasitology Inc. Published by Elsevier Ltd. All rights reserved.
Keywords: Neospora caninum; Bradyzoite; NcBSR4 gene; NcBSR4 protein; Immunohistochemistry
1. Introduction
Neospora caninum is an apicomplexan protozoan para-site causing neosporosis whose most well-known featuresare to cause repeated abortion, stillbirth and birth of con-genitally infected calves in dairy and beef cattle (Andersonet al., 2000), as well as neuromuscular disorders in dogsand, eventually, death (Dubey et al., 1988; Lindsay andDubey, 1989). This parasite is able to infect a broad rangeof hosts (Dubey, 2003; Chavez-Velasquez et al., 2004;Rodrigues et al., 2004; Gondim, 2006), similar to what
has been observed in the closely related apicomplexan Tox-
oplasma gondii. However, biological, morphological,molecular and antigenic differences between N. caninum
and T. gondii have been described (Mugridge et al., 1999;Speer et al., 1999; Naguleswaran et al., 2002), confirmingthey are separate entities.
Like T. gondii, N. caninum is able to persist in immuno-competent hosts in cyst form containing the dormant brad-yzoites within brain and muscle tissues. Those cysts remainquiescent while awaiting an appropriate immunologicalscenario wherein they switch into the fast replicating tach-yzoites, which disseminate and cause a potentially fatal dis-ease, resulting in abortion (Buxton et al., 2002). In vitromodels have been developed, which allow the study ofmechanisms by which switching between tachyzoites andbradyzoites occurs (Vonlaufen et al., 2004; Risco-Castillo
0020-7519/$30.00 � 2007 Australian Society for Parasitology Inc. Published by Elsevier Ltd. All rights reserved.
doi:10.1016/j.ijpara.2007.02.003
q Note. Nucleotide sequence data reported in this paper are available inGenBank under Accession No. EF151130.
* Corresponding author. Tel.: +34 913944069; fax: +34 913943908.E-mail address: [email protected] (L.M. Ortega-Mora).
www.elsevier.com/locate/ijpara
International Journal for Parasitology 37 (2007) 887–896

Autho
r's
pers
onal
co
py
et al., 2004). In that regard, the study of stage-specific anti-gens, by means of their cloning and further characterisa-tion, becomes critical in order to discover mechanismsinvolved in parasite transformation and immune responseevasion, both crucial events in N. caninum pathogenesis.
With regard to stage-specifically expressed Neosporasurface proteins, two immunodominant tachyzoite surfaceantigens have been characterised so far, namely NcSAG1(Hemphill et al., 1997), whose gene has been cloned (Howeet al., 1998; Nishikawa et al., 2000), and NcSRS2 (Hemp-hill and Gottstein, 1996; Nishikawa et al., 2002).
However, compared with data on N. caninum tachyzoiteantigen composition, little is known about bradyzoite anti-gens, due mainly to low bradyzoite harvesting levels in vivo(McGuire et al., 1997) and in vitro (Weiss et al., 1999;Tunev et al., 2002). Methods have recently been optimisedfor efficient production of N. caninum bradyzoites in cellculture (Vonlaufen et al., 2002; Risco-Castillo et al.,2004), allowing analysis of bradyzoite molecular biology.
McAllister et al. (1996) showed that a polyclonal serumto recombinant TgBAG1 protein cross-reacts with N. cani-
num bradyzoites but not with tachyzoites, indicating thelikelihood of similarly expressed proteins in T. gondii andN. caninum. Also, it has been shown that Neospora densegranule proteins GRA1, GRA2 and GRA7 are differen-tially expressed in tachyzoites compared with bradyzoites(Vonlaufen et al., 2004). Furthermore, the cloning andexpression of NcSAG4, the first N. caninum bradyzoite-spe-cific gene to be characterised, has recently been reported(Fernandez-Garcia et al., 2006).
In this study, we report the identification, cloning,expression and molecular characterisation of a bradyzo-ite-specific gene of N. caninum, which represents an ortho-logue to TgBSR4 (Tomavo et al., 1991; Knoll andBoothroyd, 1998).
2. Materials and methods
2.1. Production of N. caninum bradyzoites
Neospora caninum parasites were maintained in vitro byserial passage in MARC 145 cells. In vitro induction ofstage conversion was carried out in MARC-145 cellsinfected with tachyzoites at a host:parasite ratio of 2:1,and cultured in either 75 cm2 flasks or coverslips in 24-wellplates. At 24 h p.i., medium was replaced and 70 lMsodium nitroprusside (Sigma) was added and renewed dailyfor 7 days, as described previously (Risco-Castillo et al.,2004). Infected cells cultured in medium at pH 7.2 wereused as controls. Coverslips were fixed daily with 4% form-aldehyde and 4% saccharose in PBS up to day 3 and atdays 5 and 7 after stress, whereas cultures on flasks werecollected at the same intervals for RNA extraction. Para-sites were subsequently purified from the cell monolayerby rupture with a 25-gauge needle, followed by centrifuga-tion (1350g, 4 �C, 15 min). The pellet was resuspended incold PBS, washed once by centrifugation, and parasites
separated from intact host cells on PD-10 columns (GEHealthcare Life Sciences).
Flasks containing confluent keratinocyte monolayerswere infected with N. caninum parasites and stressed for 8days for bradyzoite conversion, essentially as describedby Vonlaufen et al. (2002).
2.2. Nucleic acid isolation and sequencing
Total DNA from purified tachyzoites was isolated usinga GenomicPrep� Cells and Tissue DNA Isolation kit (GEHealthcare Life Sciences). Total RNA from parasites wasisolated using the NucleoSpin II RNA purification kit(Clontech) following the manufacturer’s recommendations.Samples were then treated with 3 U of DNase I (Roche)during 30 min at 37 �C and 5 min at 95 �C to inactivateleftover DNA.
On the basis of the N. caninum expressed sequence tag(NcEST) 3c28h02.y1 sequence found by means of the BasicLocal Alignment Search Tool (BLAST–http://www.ncbi.nlm.nih.gov/BLAST/), which exhibited a 44% identity withthe TgBSR4 gene, we used the Universal Genome Walker kit(Clontech) to isolate the NcBSR4 gene. Briefly, a N. caninumgenomic DNA library was built as previously described (Fer-nandez-Garcia et al., 2006). NcBSR4 gene-specific primersF1BSR4, F2BSR4, R1BSR4 and R2BSR4 (Table 1) weredesigned using Primer Express software (v.2.0, Applied Bio-systems). Amplification was performed by nested PCR usinga DNA polymerase with exonuclease activity (Ecotaq Plus,ECOGEN).
The Expand High Fidelity PCR System (Roche) wasused to confirm boundaries of the NcEST that was primar-ily used as a template. Primers NcESTF, NcESTR,FNcEST5 and RNcEST3 were derived from 3 0 and 5 0
flanking sequences of the NcEST 3c28h02.y1 sequence(Table 1). Briefly, the PCR conditions were 94 �C for2 min, 30 cycles at 94 �C for 15 s, 60 �C for 30 s and72 �C for 1 min, and a final elongation at 72 �C for 7 min.
PCR products were analyzed by electrophoresis on a1.5% low melting agarose gel, excised and purified withthe GENECLEAN Turbo nucleic acid purification kit(Q-BIOgene). Purified DNA fragments were sequencedwith an ABI Prism 377 DNA sequencer (Applied Biosys-tems), using primers AP1, AP2, F1BSR4, F2BSR4,R1BSR4, R2BSR4, NcESTF, NcESTR and RNcEST3.To confirm fidelity of sequencing, at least two sequencesfrom separate PCRs were aligned with 100% consensus.
2.3. Sequence analysis
General similarity between DNA and protein sequenceswere assessed against non-redundant GenBank databasesthrough BLAST NCBI (http://www.ncbi.nlm.nih.gov/BLAST/) searches. Multiple and pairwise sequence align-ment was achieved using ClustalW analysis (http://www.e-bi.ac.uk/clustalw/) and the BioEdit program. We alsosearched the sequence obtained against the T. gondii and
888 V. Risco-Castillo et al. / International Journal for Parasitology 37 (2007) 887–896

Autho
r's
pers
onal
co
pyN. caninum TIGR GenBank (http://tigrblast.tigr.org/tgi/)and the ToxoDB database (http://www.toxodb.org/toxo/home.jsp). Deduction of some physico-chemical propertiesand functional motifs from the NcBSR4 sequence werecomputed using ExPASy proteomics tools (http://au.exp-asy.org/). Hydrophobic domains were identified using boththe Kyte–Doolittle hydropathy scale (Kyte and Doolittle,1982) and the TMHMM predictor (http://www.cbs.dtu.dk/). The N-terminal signal peptide was identified withSignalP v3.0, while the COOH-terminal glycosylphosphat-idylinositol anchor (GPI) was sought by http://men-del.imp.univie.ac.at/sat/gpi/gpi_server.html. To predicttertiary structures in the amino acid sequence, we usedthe CYSPRED prediction method (http://www.predictprotein.org).
2.4. RT-PCR
Specific primers were designed for NcBSR4 (NcBSR4F3and NcBSR4R3), while primers for NcSAG1, NcSAG4 and18S ribosomal RNA (Nc18sR) genes were as described byFernandez-Garcia et al. (2006) (Table 1). ComplementaryDNA was synthesised from 200 ng of total parasite RNAusing Superscript II RNase H minus Reverse Transcriptase(2 U, Invitrogen), random hexamers (12.5 nM, AppliedBiosystems) and RNase inhibitor (40 U, Ambion) in 20 llof reaction mixture, according to the manufacturer’sinstructions.
Quantitative PCR was performed in the ABI Prism 7300Sequence Detector (Applied Biosystems), using the DNAbinding dye SYBR Green I (Platinum SYBR Green qPCR
SuperMix UDG, Invitrogen) (Fernandez-Garcia et al.,2006). Four independent experiments were performed withcDNA obtained from three separate bradyzoite yields pro-duced in vitro. All samples were processed in triplicate. Foramplification, data acquisition and data analysis, we usedSequence Detection System Software v.1.6. (AppliedBiosystems).
The mean Ct value between replicate samples did notexceed a standard deviation of 0.5. Results were expressedas x-fold induction calculated by the 2�DDCt formula. Therelative quantitation of NcSAG1, NcSAG4 and NcBSR4expression was carried out using the comparative Ct
method, using N. caninum tachyzoites as the referenceand the 18S ribosomal RNA as the normaliser.
2.5. Expression and purification of recombinant NcBSR4
The expression vector system pRSET C (Invitrogen) wasused to produce a truncated form of the putative NcBSR4(amino acids 40–408) as recombinant protein. An NcBSR4
gene fragment corresponding to nucleotides 1084–2195 wasamplified from genomic DNA of N. caninum by PCR witholigonucleotides FBamHNcBSR4 and RHindIIINcBSR4(Table 1). PCR was performed on 100 ng of genomicDNA using the Expand High Fidelity Plus PCR System(Roche). The amplified DNA fragment was purified,digested with BamHI and HindIII and cloned in thepRSET-C plasmid previously linearised with the sameendonucleases.
The cloned sequence was expressed in Rosetta (DE3)pLysS Escherichia coli cells (Novagen) as polyhistidine
Table 1Oligonucleotides used for PCR, RT-PCR, sequencing and genome walking
Primer name Primer sequence 5 0–3 0a Location (nt) Direction
AP1 gtaatacgactcactatagggc BothAP2 actatagggcactatagggc BothF1BSR4 tcggcacagaaacagagctga 1411–1431 SenseF2BSR4 aagggttcagcgcaaacga 1593–1611 SenseR1BSR4 aaccacgattctggagtccttcat 1096–1119 AntisenseR2BSR4 aactccaaccacgccaactg 1036–1055 AntisenseNcESTF gtgccgcttgaggagattga 803–822 SenseNcESTR aagtcgtacggaaagtcggaaga 1865–1887 AntisenseNcESTR2 ccgaagaaacctacctcgaagc 2308–2329 AntisenseFNcEST5 agacaatcgcgaacctccaa 519–538 SenseRNcEST3 gttgtgccggtgactgacag 1152–1171 AntisenseNcBSR4F3 ccatgaaggactccagaatcg 1094–1114 SenseNcBSR4R3 tcgctggaaacgcacattta 1175–1194 AntisenseNc18sF gatacagaaccaacccaccttcc 189–211b SenseNc18sR agaccgaagtcaaacgcgatc 265–245b AntisenseNcSAG1F cggtgtcgcaatgtgctctt 721–739c SenseNcSAG1R acggtcgtcccagaacaaac 921–901c AntisenseNcSAG4F gatttcaagaagccgctgga 339–358d SenseNcSAG4R tgagaacttgtgtgtcgcctgtt 468–446d AntisenseFBamHNcBSR4 tataggatcccagatgatctccatgaaggactc 1084–1106 SenseRHindIIINcBSR4 acttaagcttttacaaggagccaacaatgagc 2174–2195 Antisense
a Underlined restriction sites for cloning of NcBSR4.b Nucleotide (nt) position in Nc18sR.c nt position in NcSAG1.d nt position in NcSAG4.
V. Risco-Castillo et al. / International Journal for Parasitology 37 (2007) 887–896 889

Autho
r's
pers
onal
co
py
(His6) fusion proteins as previously described (Fernandez-Garcia et al., 2006), with some modifications: 1% glucosewas added to the selective medium and cell lysis wasperformed with 1· BugBuster protein extraction reagent(Novagen) containing 8· sodium phosphate buffer, pH 7.4(5 ml g�1 bacteria); benzonase (1 ll ml�1 sample, Novagen)and lysozyme (1 KU ml�1 sample). Cell lysis was enhancedby gentle shaking (1 h, room temperature). Inclusion bodieswere washed four times in BugBuster reagent and lysozymeand finally suspended in 0.1· BugBuster solutions.
Denatured recombinant NcBSR4 (rNcBSR4) was solu-bilised by incubating for 2 h at room temperature with abinding buffer containing 8 M Urea and 40 mM imidazolein phosphate buffer (pH 7.4). A HisTrap� HP column wasplaced onto an AktaPrime purification system (GE Health-care Life Sciences), washed with dH2O and equilibratedwith binding buffer as mentioned above. The column wasthen loaded with the histidine-tagged protein, and His6-tagged rNcBSR4 was eluted using an imidazole ascendinggradient of up to 500 mM.
Protein concentration was estimated by Coomassiestaining using serial dilutions of BSA as a standard. Proteinconcentration was measured using Quantity One software(v.7.2, BioRad). Results were confirmed through an absor-bance assay which uses the molar extinction coefficient ofrNcBSR4 calculated from its amino acid composition (Gilland von Hippel, 1989).
2.6. Mass spectrometry analysis
Protein corresponding to rNcBSR4 was manuallyexcised from prepared Coomassie-stained 1-D gels. Stainedgel pieces were destained with acetonitrile (ACN), washedtwice with 50% ACN in 25 mM ammonium bicarbonate(AmBic), and remaining solvent was removed by dryingthe gel pieces in a SpeedVac. The band was reduced with10 mM dithiothreitol in 25 mM AmBic for 30 min at56 �C and subsequently alkylated with 55 mM iodoaceta-mide in 25 mM AmBic for 20 min in the dark. After wash-ing with 25 mm AmBic and ACN, it was vacuum-dried andincubated with 12.5 ng/ll sequencing grade trypsin(Roche) in 25 mM AmBic overnight at 37 �C. After diges-tion, the crude extract was separated and peptidesextracted into 50% CAN, 1% trifluoroacetic acid (TFA)and then into 100% ACN. All extracts were pooled, andthe volume was reduced by SpeedVac.
Samples for mass spectrometry (MS/MS) sequencingwere mixed 1:1 with a-cyano matrix (5 mg/ml in 50%ACN/0.3% TFA) and spotted on the stained steel plate.MS/MS and de novo sequencing analyses were carriedout using the MALDI-tandem time-of-flight mass spec-trometer 4700 Proteomics Analyzer (Applied Biosystem).
2.7. Polyclonal rabbit anti-rNcBSR4 sera production
Two New Zealand rabbits were each immunisedintradermally with a 200 lg dose of denatured rNcBSR4
purified by IMAC (Vaitukaitis, 1981). The inoculum wasprepared in FCA (Difco Laboratories) according to stan-dard methods. Another five inoculations were performedat 3-week intervals i.m. with 100 lg of rNcBSR4 preparedin Freund’s incomplete adjuvant. Sera were collected 18weeks after the first immunisation. All procedures involv-ing animal handling met the terms established by the EUlegislation.
2.8. Immunofluorescence
Fixed coverslips were double-labelled with the monoclo-nal mouse anti-NcSAG1 antibody (aSAG1; 1:1500)(Bjorkman and Hemphill, 1998) and rabbit polyclonalanti-rNcBSR4 serum (aBSR4; 1:500). To confirm tachyzo-ite-bradyzoite conversion, polyclonal rabbit anti-BAG1serum (aBAG1; 1:500 dilution) (McAllister et al., 1996)was used together with aSAG1. Goat anti-rabbit and goatanti-mouse secondary antibodies conjugated to Alexa 488and Alexa 594, respectively, (Molecular Probes) were usedas secondary antibodies. Parasite and host cell nuclei were4 0,6-diamidino-2-phenylindole (DAPI)-labelled. Photomi-crographs were taken with a 100· oil-immersion objectiveon a fluorescence-inverted microscope (Nikon EclipseTE200) connected to a digital camera. Duplicates for eachday from the stage-conversion assay were tested.
2.9. Immunogold TEM
Keratinocyte cultures containing N. caninum bradyzo-ites were fixed and LR-White embedded and on-sectionlabelled as described previously (Hemphill et al., 2004).Sections were loaded onto formvar-carbon coated gridsand non-specific binding sites were blocked for 2 h in 3%BSA in PBS. They were then incubated in a wet chamberwith aBSR4 diluted 1:100 in 0.1% BSA in PBS for 1 h.After five washes of PBS (5 min each), the goat anti-rabbitantibody conjugated to 10 nm diameter gold particles(Pharmacia) was used at a dilution of 1:6 in PBS and0.3% BSA. The negative control was incubated with anti-rabbit conjugate only. After extensive washing in PBS,grids were air dried and finally stained with uranyl acetateand lead citrate. Specimens were viewed on a Philips 400TEM operating at 80 kV.
2.10. Immunohistochemistry
A naturally Neospora-infected calf was blood-sampledprior to feeding with colostrum and tested by the IndirectFluorescence Antibody Test (IFAT). After confirmationof its seropositivity, brain slices containing N. caninum tis-sue cysts were examined by immunohistochemistry to eval-uate immune reactivity of anti-rNcBSR4 against maturebradyzoites produced in vivo. Immunolabelling was per-formed by the streptavidin-biotin-peroxidase technique.Polyclonal rabbit anti-rNcBSR4 serum was used at 1:100
890 V. Risco-Castillo et al. / International Journal for Parasitology 37 (2007) 887–896

Autho
r's
pers
onal
co
py
dilution, following standard methods previously described(Pereira-Bueno et al., 2003).
3. Results
3.1. Isolation and characterisation of the NcBSR4 gene
In this study, we isolated N. caninum NcBSR4, an ortho-logue of T. gondii TgBSR4, coding for a bradyzoite-specificprotein. Amplification of the sequence named NcEST3c28h02.y1 at both 3 0 and 5 0 ends by the Genome Walkingmethod allowed us to obtain a 2676 nucleotide sequence inwhich a continuous open reading frame (ORF) of 1227nucleotides was present. Similarity of its encoded proteinto TgBSR4 is 66%, sharing a total of 273 residues. Dueto the fact that the T. gondii genome is completelysequenced and no other sequence showed higher similarity,the N. caninum gene identified is the most likely orthologueof the TgBSR4 gene. After ORF translation, a 408 aminoacid protein (Fig. 1) was predicted with a molecular massof 42940.9 Da.
DNA from three N. caninum isolates (NC-1, NC-Liv,Nc-SweB1) was amplified using NcESTF and NcESTR2oligonucleotides (Table 1), and sequenced to confirm com-position of the complete NcBSR4 ORF. No differenceamong isolates was observed. We propose the denomina-tion NcBSR4 for this gene, and NcBSR4 for the protein
encoded, in accordance with existing nomenclature (Howeand Sibley, 1999).
Two hydrophobic domains were identified at the NH2
and COOH terminal ends of NcBSR4 protein using theKyte–Doolittle hydrophobicity scale. A 39-amino acidputative signal peptide was found at the N-terminus anda potential GPI anchor was found 30 amino acids upstreamof the C-terminus, in accordance with the x and x + 2 rule(Gerber et al., 1992). The ScanProsite tool, which displaysfunctional motifs, found an N-glycosylation site at the 258–261 position. Furthermore, the CYSPRED predictionmethod found 12 conserved cysteine residues of the SRSantigen family (Fig. 1).
3.2. Production and immunoreactivity of the NcBSR4
recombinant protein (rNcBSR4)
The NcBSR4 protein (amino acids 40–408) was pro-duced as a recombinant protein (rNcBSR4) in the prokary-otic expression vector system pRSET C (Invitrogen), andexpressed as a fusion protein containing an N-terminalhexahistidine tag. Recombinant NcBSR4 accumulated asinclusion bodies (Fig. 2) that were denatured and then puri-fied by IMAC. The apparent molecular mass of rNcBSR4protein as determined by SDS–PAGE was 51 kDa, whilethe predicted mass was around 43 kDa. The molecularmass of rNcBSR4 was confirmed by mass spectrometric
Fig. 1. Deduced polypeptide sequence of NcBSR4 and alignment produced by ClustalW with the homologous protein in Toxoplasma gondii (TgBSR4).The amino acid sequence of NcBSR4 protein contains a polypeptide of 408 amino acids. The putative signal peptide at the amino end is in bold and italics,while an arrow indicates the potential glycosylphosphatidylinositol anchor cleavage at the carboxyl end. Boxes indicate conserved cysteines and thepredicted N-glycosylation site is shaded.
V. Risco-Castillo et al. / International Journal for Parasitology 37 (2007) 887–896 891

Autho
r's
pers
onal
co
pyanalysis and MS/MS analyses corroborated the composi-tion of NcBSR4 as deduced by nucleic acid translation.
Furthermore, purified rNcBSR4 was used in SDS–PAGE under reducing conditions and challenged againstthe polyclonal rabbit anti-serum developed against thisprotein, reacting specifically as expected.
3.3. Stage-specific NcBSR4 expression
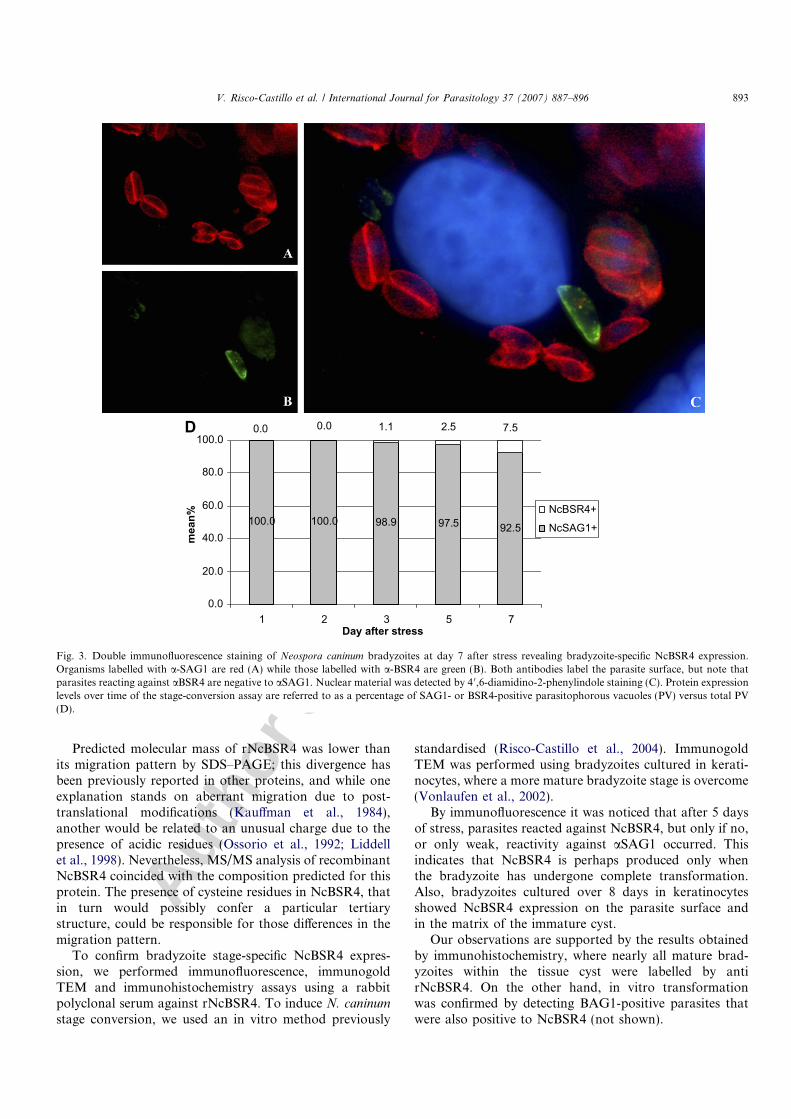
Transformation of tachyzoites into bradyzoites along atime course was confirmed by BAG1 staining, observingup to 37% of BAG1-positives parasitophorous vacuoles(PVs) at day 7 after stress (data not shown). Double immu-nofluorescence assays carried out at days 5 and 7 afterstress revealed bradyzoite-specific NcBSR4 expression infew bradyzoites (Fig. 3A and C) that had left to expressSAG1 (Fig. 3B and C). The mean percentage of BSR4-positive PVs at day 7 after stress was 7.5% (Fig. 3D). Incontrol cultures containing unstressed tachyzoites none ofthe parasites reacted against aNcBSR4, nor were anyPVs stained with aBAG1.
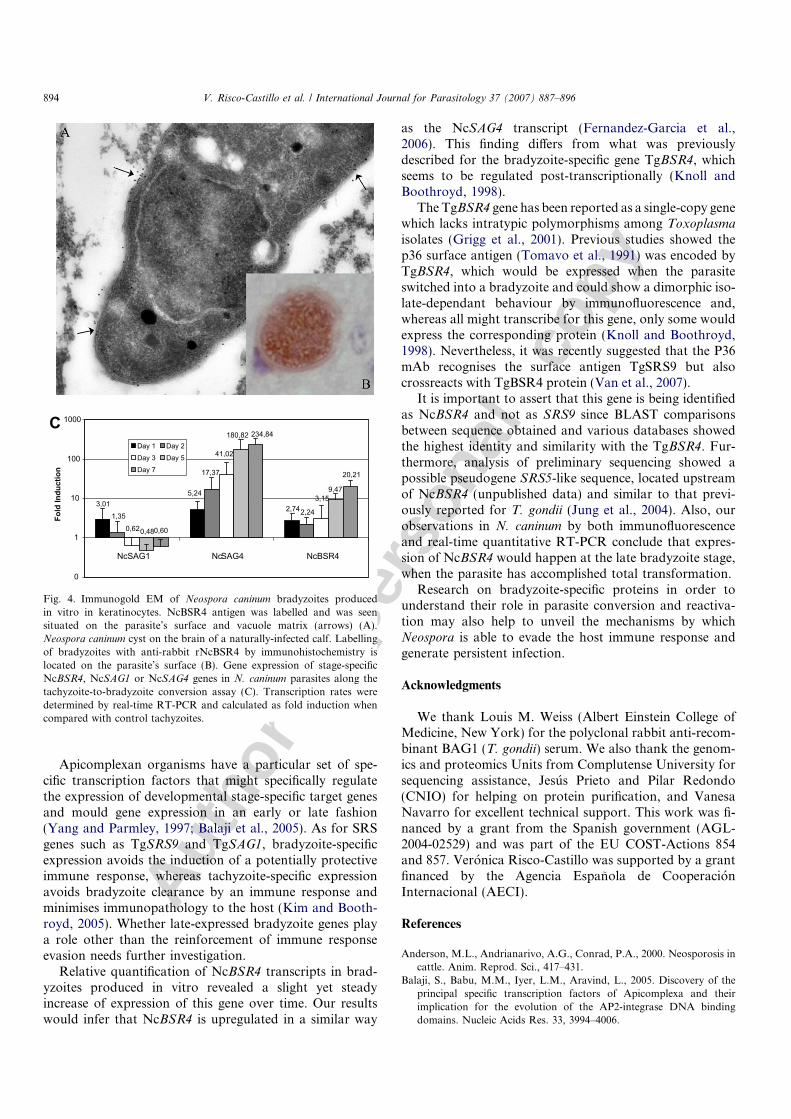
Observations by immunogold TEM of N. caninum-infected keratinocytes that were treated with SNP for 8days revealed labelling of NcBSR4, localizing on the sur-face of bradyzoites (Fig. 4A). Moreover, reaction of anti-recNcNSR4 by immunohistochemistry against maturebradyzoites on paraffin sections of brain tissue obtainedfrom an infected calf was confirmed, and aNcBSR4 waslabelled all bradyzoites contained in a tissue cyst (Fig. 4B).
On the other hand, real-time quantitative RT-PCR wasused to compare mRNA levels of NcBSR4, NcSAG1 or
NcSAG4 genes in N. caninum parasites obtained duringthe stage-conversion assay (1, 2, 3, 5 and 7 days). Compar-ison of Ct values for each triplicate sample showed minimalvariation when the standard deviation was calculated (datanot shown). When compared with the reference and nor-malised to 18S ribosomal RNA, NcSAG1 mRNA expres-sion levels showed minor variations, although decreasingwith time. When analyzing the expression of NcSAG4, abradyzoite-specific gene, transcript levels reached a meanvalue of 235-fold induction at day 7 after stress, whileNcBSR4 transcript levels increased up to 20-fold(Fig. 4C). Absence of DNA was confirmed by directPCR of total RNA from each sample.
4. Discussion
To date, the precise role of GPI anchored surface pro-teins, also known as the SRS superfamily, from apicom-plexan still needs to be disclosed. For N. caninum
bradyzoites, as for T. gondii, they might act as merely pro-tective barriers, as receptors that ease invasion of a broadrange of cell types or even as mediators for immuneresponse evasion, facilitating survival and propagation ofthe parasite (Boothroyd et al., 1998; Lekutis et al., 2001).Given that information regarding genes coding for suchproteins are more extensively depicted in T. gondii, ourapproach relied on the use of the bradyzoite-specificTgBSR4 gene as a framework for the application of thegenome walking method.
This paper reports the identification, cloning, expressionand molecular characterisation of NcBSR4, a novel surfacegene of N. caninum homologous to T. gondii TgBSR4,which is specifically expressed at the bradyzoite stage.
In analyzing the encoded protein, two hydrophobicdomains were found revealing an N-terminal signal peptideand a GPI anchor at the COOH-end. The presence of aGPI anchor, which in the case of NcBSR4 is at the 379amino acid position, correlates with what was expectedfor a surface protein, allowing the protein to be tetheredto the outer surface membrane of the parasite (Cross,1990; Knoll and Boothroyd, 1998; Jung et al., 2004),although slight GPI anchor modifications in N. caninum
have been reported (Schares et al., 2000).Howe et al. (1998) reported two N. caninum tachyzoite
surface antigens, NcSAG1 and NcSRS2, bearing 12 con-served cysteine residues previously identified in SAG1,SAG3 and SRS antigens of T. gondii. Their conservationindicates that they are probably involved in intramoleculardisulfide bonding and therefore all these proteins have asimilar folding pattern (Howe and Sibley, 1999). Twelveconserved cysteine residues have also been found inNcBSR4 when compared with the TgBSR4 amino acidicsequence; therefore elucidation of the presence of thesemodifications on the N. caninum bradyzoite surface, as wellas their implication in the parasite’s interaction with thehost immune response, is a field that needs to be morethoughtfully studied.
Fig. 2. SDS–PAGE and Coomassie staining of the recombinant NcBSR4protein from induced Escherichia coli lysates. Lanes 1 and 3, inclusionbodies; lanes 2 and 4, soluble proteins. Arrow indicates the position of therecombinant NcBSR4 protein. The molecular weight markers are shownon the right side.
892 V. Risco-Castillo et al. / International Journal for Parasitology 37 (2007) 887–896
Autho
r's
pers
onal
co
py
Predicted molecular mass of rNcBSR4 was lower thanits migration pattern by SDS–PAGE; this divergence hasbeen previously reported in other proteins, and while oneexplanation stands on aberrant migration due to post-translational modifications (Kauffman et al., 1984),another would be related to an unusual charge due to thepresence of acidic residues (Ossorio et al., 1992; Liddellet al., 1998). Nevertheless, MS/MS analysis of recombinantNcBSR4 coincided with the composition predicted for thisprotein. The presence of cysteine residues in NcBSR4, thatin turn would possibly confer a particular tertiarystructure, could be responsible for those differences in themigration pattern.
To confirm bradyzoite stage-specific NcBSR4 expres-sion, we performed immunofluorescence, immunogoldTEM and immunohistochemistry assays using a rabbitpolyclonal serum against rNcBSR4. To induce N. caninum
stage conversion, we used an in vitro method previously
standardised (Risco-Castillo et al., 2004). ImmunogoldTEM was performed using bradyzoites cultured in kerati-nocytes, where a more mature bradyzoite stage is overcome(Vonlaufen et al., 2002).
By immunofluorescence it was noticed that after 5 daysof stress, parasites reacted against NcBSR4, but only if no,or only weak, reactivity against aSAG1 occurred. Thisindicates that NcBSR4 is perhaps produced only whenthe bradyzoite has undergone complete transformation.Also, bradyzoites cultured over 8 days in keratinocytesshowed NcBSR4 expression on the parasite surface andin the matrix of the immature cyst.
Our observations are supported by the results obtainedby immunohistochemistry, where nearly all mature brad-yzoites within the tissue cyst were labelled by antirNcBSR4. On the other hand, in vitro transformationwas confirmed by detecting BAG1-positive parasites thatwere also positive to NcBSR4 (not shown).
100.0 100.0 98.9 97.5 92.5
0.0 0.0 1.1 2.5 7.5
0.0
20.0
40.0
60.0
80.0
100.0
1 2 3 5 7Day after stress
mea
n%
NcBSR4+
NcSAG1+
D
Fig. 3. Double immunofluorescence staining of Neospora caninum bradyzoites at day 7 after stress revealing bradyzoite-specific NcBSR4 expression.Organisms labelled with a-SAG1 are red (A) while those labelled with a-BSR4 are green (B). Both antibodies label the parasite surface, but note thatparasites reacting against aBSR4 are negative to aSAG1. Nuclear material was detected by 4 0,6-diamidino-2-phenylindole staining (C). Protein expressionlevels over time of the stage-conversion assay are referred to as a percentage of SAG1- or BSR4-positive parasitophorous vacuoles (PV) versus total PV(D).
V. Risco-Castillo et al. / International Journal for Parasitology 37 (2007) 887–896 893
Autho
r's
pers
onal
co
py
Apicomplexan organisms have a particular set of spe-cific transcription factors that might specifically regulatethe expression of developmental stage-specific target genesand mould gene expression in an early or late fashion(Yang and Parmley, 1997; Balaji et al., 2005). As for SRSgenes such as TgSRS9 and TgSAG1, bradyzoite-specificexpression avoids the induction of a potentially protectiveimmune response, whereas tachyzoite-specific expressionavoids bradyzoite clearance by an immune response andminimises immunopathology to the host (Kim and Booth-royd, 2005). Whether late-expressed bradyzoite genes playa role other than the reinforcement of immune responseevasion needs further investigation.
Relative quantification of NcBSR4 transcripts in brad-yzoites produced in vitro revealed a slight yet steadyincrease of expression of this gene over time. Our resultswould infer that NcBSR4 is upregulated in a similar way
as the NcSAG4 transcript (Fernandez-Garcia et al.,2006). This finding differs from what was previouslydescribed for the bradyzoite-specific gene TgBSR4, whichseems to be regulated post-transcriptionally (Knoll andBoothroyd, 1998).
The TgBSR4 gene has been reported as a single-copy genewhich lacks intratypic polymorphisms among Toxoplasma
isolates (Grigg et al., 2001). Previous studies showed thep36 surface antigen (Tomavo et al., 1991) was encoded byTgBSR4, which would be expressed when the parasiteswitched into a bradyzoite and could show a dimorphic iso-late-dependant behaviour by immunofluorescence and,whereas all might transcribe for this gene, only some wouldexpress the corresponding protein (Knoll and Boothroyd,1998). Nevertheless, it was recently suggested that the P36mAb recognises the surface antigen TgSRS9 but alsocrossreacts with TgBSR4 protein (Van et al., 2007).
It is important to assert that this gene is being identifiedas NcBSR4 and not as SRS9 since BLAST comparisonsbetween sequence obtained and various databases showedthe highest identity and similarity with the TgBSR4. Fur-thermore, analysis of preliminary sequencing showed apossible pseudogene SRS5-like sequence, located upstreamof NcBSR4 (unpublished data) and similar to that previ-ously reported for T. gondii (Jung et al., 2004). Also, ourobservations in N. caninum by both immunofluorescenceand real-time quantitative RT-PCR conclude that expres-sion of NcBSR4 would happen at the late bradyzoite stage,when the parasite has accomplished total transformation.
Research on bradyzoite-specific proteins in order tounderstand their role in parasite conversion and reactiva-tion may also help to unveil the mechanisms by whichNeospora is able to evade the host immune response andgenerate persistent infection.
Acknowledgments
We thank Louis M. Weiss (Albert Einstein College ofMedicine, New York) for the polyclonal rabbit anti-recom-binant BAG1 (T. gondii) serum. We also thank the genom-ics and proteomics Units from Complutense University forsequencing assistance, Jesus Prieto and Pilar Redondo(CNIO) for helping on protein purification, and VanesaNavarro for excellent technical support. This work was fi-nanced by a grant from the Spanish government (AGL-2004-02529) and was part of the EU COST-Actions 854and 857. Veronica Risco-Castillo was supported by a grantfinanced by the Agencia Espanola de CooperacionInternacional (AECI).
References
Anderson, M.L., Andrianarivo, A.G., Conrad, P.A., 2000. Neosporosis incattle. Anim. Reprod. Sci., 417–431.
Balaji, S., Babu, M.M., Iyer, L.M., Aravind, L., 2005. Discovery of theprincipal specific transcription factors of Apicomplexa and theirimplication for the evolution of the AP2-integrase DNA bindingdomains. Nucleic Acids Res. 33, 3994–4006.
9,47
2,74
5,243,01
2,24
17,37
1,35
3,15
41,02
0,62
180,82
0,48
20,21
234,84
0,60
0
1
10
100
1000
NcSAG1 NcSAG4 NcBSR4
Fold
Indu
ctio
n
Day 1 Day 2Day 3 Day 5Day 7
C
Fig. 4. Immunogold EM of Neospora caninum bradyzoites producedin vitro in keratinocytes. NcBSR4 antigen was labelled and was seensituated on the parasite’s surface and vacuole matrix (arrows) (A).Neospora caninum cyst on the brain of a naturally-infected calf. Labellingof bradyzoites with anti-rabbit rNcBSR4 by immunohistochemistry islocated on the parasite’s surface (B). Gene expression of stage-specificNcBSR4, NcSAG1 or NcSAG4 genes in N. caninum parasites along thetachyzoite-to-bradyzoite conversion assay (C). Transcription rates weredetermined by real-time RT-PCR and calculated as fold induction whencompared with control tachyzoites.
894 V. Risco-Castillo et al. / International Journal for Parasitology 37 (2007) 887–896

Autho
r's
pers
onal
co
py
Bjorkman, C., Hemphill, A., 1998. Characterization of Neospora caninum
iscom antigens using monoclonal antibodies. Parasite Immunol. 20,73–80.
Boothroyd, J.C., Hehl, A., Knoll, L.J., Manger, I.D., 1998. The surface ofToxoplasma: more and less. Int. J. Parasitol. 28, 3–9.
Buxton, D., McAllister, M.M., Dubey, J.P., 2002. The comparativepathogenesis of neosporosis. Trends Parasitol. 18, 546–552.
Chavez-Velasquez, A., Alvarez-Garcia, G., Collantes-Fernandez, E.,Casas-Astos, E., Rosadio-Alcantara, R., Serrano-Martinez, E., Orte-ga-Mora, L.M., 2004. First report of Neospora caninum infection inadult alpacas (Vicugna pacos) and llamas (Lama glama). J. Parasitol.90, 864–866.
Cross, G.A., 1990. Glycolipid anchoring of plasma membrane proteins.Annu. Rev. Cell Biol. 6, 1–39.
Dubey, J.P., Carpenter, J.L., Speer, C.A., Topper, M.J., Uggla, A., 1988.Newly recognized fatal protozoan disease of dogs. J. Am. Vet. Med.Assoc. 192, 1269–1285.
Dubey, J.P., 2003. Review of Neospora caninum and neosporosis inanimals. Korean J. Parasitol. 41, 1–16.
Fernandez-Garcia, A., Risco-Castillo, V., Zaballos, A., Alvarez-Garcia,G., Ortega-Mora, L.M., 2006. Identification and molecular cloning ofthe Neospora caninum SAG4 gene specifically expressed at bradyzoitestage. Mol. Biochem. Parasitol. 146, 89–97.
Gerber, L.D., Kodukula, K., Undenfriend, S., 1992. PhospatidylinositolGlycan (Pi-G) anchored membrane-proteins-amino-acid-requirementsadjacent to the site of cleavage and Pi-G attachment in the COOH-terminal signal peptide. J. Biol. Chem. 267, 12168–12173.
Gill, S.C., von Hippel, P.H., 1989. Calculation of protein extinctioncoefficients from amino acid sequence data. Anal. Biochem. 182,319–326.
Gondim, L.F.P., 2006. Neospora caninum in wildlife. Trends Parasitol. 22,247–252.
Grigg, M.E., Bonnefoy, S., Hehl, A.B., Suzuki, Y., Boothroyd, J.C., 2001.Success and virulence in Toxoplasma as the result of sexual recom-bination between two distinct ancestries. Science 294, 161–165.
Hemphill, A., Gottstein, B., 1996. Identification of a major surface proteinon Neospora caninum tachyzoites. Parasitol. Res. 82, 497–504.
Hemphill, A., Felleisen, R., Connolly, B., Gottstein, B., Hentrich, B.,Muller, N., 1997. Characterization of a cDNA-clone encoding Nc-p43,a major Neospora caninum tachyzoite surface protein. Parasitology 115(Pt 6), 581–590.
Hemphill, A., Vonlaufen, N., Naguleswaran, A., Keller, N., Riesen, M.,Guetg, N., Srinivasan, S., Alaeddine, F., 2004. Tissue culture andexplant approaches to studying and visualizing Neospora caninum
and its interactions with the host cell. Microsc. Microanal. 10,602–620.
Howe, D.K., Crawford, A.C., Lindsay, D., Sibley, L.D., 1998. The p29and p35 immunodominant antigens of Neospora caninum tachyzoitesare homologous to the family of surface antigens of Toxoplasma
gondii. Infect. Immun. 66, 5322–5328.Howe, D.K., Sibley, L.D., 1999. Comparison of the major antigens of
Neospora caninum and Toxoplasma gondii. Int. J. Parasitol. 29, 1489–1496.
Jung, C., Lee, C.Y., Grigg, M.E., 2004. The SRS superfamily ofToxoplasma surface proteins. Int. J. Parasitol. 34, 285–296.
Kauffman, E., Geisler, N., Weber, K., 1984. SDS–PAGE stronglyoverstimates the molecular masses of neurofilament proteins. FEBSLett. 170, 81–84.
Kim, S.K., Boothroyd, J.C., 2005. Stage-specific expression of surfaceantigens by Toxoplasma gondii as a mechanism to facilitate parasitepersistence. J. Immunol. 174, 8038–8048.
Knoll, L.J., Boothroyd, J.C., 1998. Isolation of developmentally regulatedgenes from Toxoplasma gondii by a gene trap with the positive andnegative selectable marker hypoxanthine–xanthine–guanine phospho-ribosyltransferase. Mol. Cell Biol. 18, 807–814.
Kyte, J., Doolittle, R.F., 1982. A simple method for displayingthe hydropathic character of a protein. J. Mol. Biol. 157,105–132.
Lekutis, C., Ferguson, D.J., Grigg, M.E., Camps, M., Boothroyd, J.C.,2001. Surface antigens of Toxoplasma gondii: variations on a theme.Int. J. Parasitol. 31, 1285–1292.
Liddell, S., Lally, N.C., Jenkins, M.C., Dubey, J.P., 1998. Isolation of thecDNA encoding a dense granule associated antigen (NCDG2) ofNeospora caninum. Mol. Biochem. Parasitol. 93, 153–158.
Lindsay, D.S., Dubey, J.P., 1989. In vitro development of Neospora
caninum (Protozoa: Apicomplexa) from dogs. J. Parasitol. 75, 163–165.McAllister, M.M., Parmley, S.F., Weiss, L.M., Welch, V.J., McGuire,
A.M., 1996. An immunohistochemical method for detecting bradyzo-ite antigen (BAG5) in Toxoplasma gondii-infected tissues cross-reactswith a Neospora caninum bradyzoite antigen. J. Parasitol. 82, 354–355.
McGuire, A.M., McAllister, M.M., Jolley, W.R., Anderson-Sprecher,R.C., 1997. A protocol for the production of Neospora caninum tissuecysts in mice. J. Parasitol. 83, 647–651.
Mugridge, N.B., Morrison, D.A., Heckeroth, A.R., Johnson, A.M.,Tenter, A.M., 1999. Phylogenetic analysis based on full-length largesubunit ribosomal RNA gene sequence comparison reveals thatNeospora caninum is more closely related to Hammondia heydorni
than to Toxoplasma gondii. Int. J. Parasitol. 29, 1545–1556.Naguleswaran, A., Cannas, A., Keller, N., Vonlaufen, N., Bjorkman, C.,
Hemphill, A., 2002. Vero cell surface proteoglycan interaction with themicroneme protein NcMIC(3) mediates adhesion of Neospora caninum
tachyzoites to host cells unlike that in Toxoplasma gondii. Int. J.Parasitol. 32, 695–704.
Nishikawa, Y., Kousaka, Y., Fukumoto, S., Xuan, X., Nagasawa, H.,Igarashi, I., Fujisaki, K., Otsuka, H., Mikami, T., 2000. Delivery ofNeospora caninum surface protein, NcSRS2 (Nc-p43), to mouse usingrecombinant vaccinia virus. Parasitol. Res. 86, 934–939.
Nishikawa, Y., Tragoolpua, K., Makala, L., Xuan, X., Nagasawa, H.,2002. Neospora caninum NcSRS2 is a transmembrane protein thatcontains a glycosylphosphatidylinositol anchor in insect cells. Vet.Parasitol. 109, 191–201.
Ossorio, P.N., Schwartzman, J.D., Boothroyd, J.C., 1992. A Toxoplasma
gondii rhoptry protein associated with host cell penetration hasunusual charge asymmetry. Mol. Biochem. Parasitol. 50, 1–15.
Pereira-Bueno, J., Quintanilla-Gozalo, A., Perez-Perez, V., Espi-Felguer-oso, A., Alvarez-Garcia, G., Collantes-Fernandez, E., Ortega-Mora,L.M., 2003. Evaluation by different diagnostic techniques of bovineabortion associated with Neospora caninum in Spain. Vet. Parasitol.111, 143–152.
Risco-Castillo, V., Fernandez-Garcia, A., Ortega-Mora, L.M., 2004.Comparative analysis of stress agents in a simplified in vitro system ofNeospora caninum bradyzoite production. J. Parasitol. 90, 466–470.
Rodrigues, A.A.R., Gennari, S.M., Aguiar, D.M., Sreekumar, C., Hill,D.E., Miska, K.B., Vianna, M.C.B., Dubey, J.P., 2004. Shedding ofNeospora caninum oocysts by dogs fed tissues from naturally infectedwater buffaloes (Bubalus bubalis) from Brazil. Vet. Parasitol. 124, 139–150.
Schares, G., Zinecker, C.F., Schmidt, J., Azzouz, N., Conraths, F.J.,Gerold, P., Schwarz, R.T., 2000. Structural analysis of free andprotein-bound glycosyl-phosphatidylinositols of Neospora caninum.Mol. Biochem. Parasitol. 105, 155–161.
Speer, C.A., Dubey, J.P., McAllister, M.M., Blixt, J.A., 1999.Comparative ultrastructure of tachyzoites, bradyzoites, and tissuecysts of Neospora caninum. In: Methods in Enzimology, vol. 73Immunochemical Tecniques Part B. Langone, J.J., Van Vunakis, H.(Eds.), Academic Press, New York. pp 46–52. Int. J. Parasitol. 29,1509–1519.
Tomavo, S., Fortier, B., Soete, M., Ansel, C., Camus, D., Dubremetz,J.F., 1991. Characterization of bradyzoite-specific antigens of Toxo-
plasma gondii. Infect. Immun. 59, 3750–3753.Tunev, S.S., McAllister, M.M., Anderson-Sprecher, R.C., Weiss, L.M.,
2002. Neospora caninum in vitro: evidence that the destiny of aparasitophorous vacuole depends on the phenotype of the progenitorzoite. J. Parasitol. 88, 1095–1099.
Vaitukaitis, J., 1981. Production of antisera with small doses of immu-nogen: multiple intradermal injections. In: Langone, J.J., Van Vuna-
V. Risco-Castillo et al. / International Journal for Parasitology 37 (2007) 887–896 895

Autho
r's
pers
onal
co
py
kis (Eds.), Methods in Enzimology, Volume 73 ImmunochemicalTecniques Part B. Academic Press, New York, pp. 46–52.
Van, T.T., Kim, S.K., Camps, M., Boothroyd, J.C., Knoll, L.J., 2007. TheBSR4 protein is up-regulated in Toxoplasma gondii bradyzoites,however the dominant surface antigen recognised by the P36 mono-clonal antibody is SRS9. Int. J. Parasitol., in press, doi:10.1016/j.ijpara.2007.02.001.
Vonlaufen, N., Muller, N., Keller, N., Naguleswaran, A., Bohne, W.,McAllister, M.M., Bjorkman, C., Muller, E., Caldelari, R., Hemphill,A., 2002. Exogenous nitric oxide triggers Neospora caninum tachyzoite-to-bradyzoite stage conversion in murine epidermal keratinocyte cellcultures. Int. J. Parasitol. 32, 1253–1265.
Vonlaufen, N., Guetg, N., Naguleswaran, A., Muller, N., Bjork-man, C., Schares, G., von, B.D., Ellis, J., Hemphill, A., 2004.In vitro induction of Neospora caninum bradyzoites in vero cellsreveals differential antigen expression, localization, and host-cellrecognition of tachyzoites and bradyzoites. Infect. Immun. 72,576–583.
Weiss, L.M., Ma, Y.F., Halonen, S., McAllister, M.M., Zhang, Y.W.,1999. The in vitro development of Neospora caninum bradyzoites. Int.J. Parasitol. 29, 1713–1723.
Yang, S., Parmley, S.F., 1997. Toxoplasma gondii expresses two distinctlactate dehydrogenase homologous genes during its life cycle inintermediate hosts. Gene 184, 1–12.
896 V. Risco-Castillo et al. / International Journal for Parasitology 37 (2007) 887–896
Related Documents











