MOLECULAR CELL BIOLOGY SIXTH EDITION MOLECULAR CELL BIOLOGY SIXTH EDITION Copyright 2008 © W. H. Freeman and Company CHAPTER 11 Transmembrane Transport of Ions and Small Molecules CHAPTER 11 Transmembrane Transport of Ions and Small Molecules Lodish • Berk • Kaiser • Krieger • Scott • Bretscher •Ploegh • Matsudaira © 2008 W. H. Freeman and Company

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

MOLECULAR CELL BIOLOGY
SIXTH EDITION
MOLECULAR CELL BIOLOGY
SIXTH EDITION
Copyright 2008 copy
W H Freeman and Company
CHAPTER 11Transmembrane Transport of Ions
and Small Molecules
CHAPTER 11Transmembrane Transport of Ions
and Small Molecules
Lodish bull Berk bull Kaiser bull Krieger bull Scott bull Bretscher bullPloegh bull Matsudaira
copy 2008 W H Freeman and Company
A study of mutant zebrafish with pale stripes led to the identification of a sodiumcalcium transporter that regulates the darkness of human skin
The phospholipid bilayer is a barrier that controls the transport of molecules in and out of the cell
Gases diffuse freely no proteins required
Water diffuses fast enough that proteins arenrsquot required for transport
Sugars diffuse very slowly so proteins are involved in transport
Charged molecules are virtually impermeable
Studies of synthetic lipid bilayers
help define which types of transport will require the activity of a protein Hence transport of an ion should require
a protein
Only small hydrophobic molecules cross membrane
The bilayer is permeable toSmall hydrophobic moleculesSmall uncharged polar molecules
The bilayer is impermeable toIonsLarge polar molecules
THEREFORE need membrane proteins to transport most molecules and all ions across biomembranes
KEY CONCEPTS
Selective transport across the lipid membrane requires transport proteins
Transport proteins are integral membrane proteins that move molecules and ions
There are two classes of transport proteins transporters (pumps) and channels
Most small molecules did not across membrane
Transporter protein
Also called Na+K+ ATPasesodium-amino acid transporter
Three main class of membrane protein1ATP- power pump( carrier permease)
couple with energy source for active transportbinding of specific solute to transporter which
undergo conformation change2 Channel protein (ion channel)
formation of hydrophilic pore allow passive movement of small inorganic molecule
3 Transportersuniportsymportantiport
Partition Coefficient
Permeability coefficients (in cmsec) through synthetic lipid bilayers
Product of the concentration difference (in molcm3) and permeability coefficient (in cmsec) gives the flow of solute in moles per second per square centimeter of membrane
油品分配係數
Cell membrane
bullBarrier to the passage of most polar molecule
bullMaintain concentration of solute
Diffusion rate depends on
1 Concentration gradient or electrochemical gradient
2 Hydrophobicity
ie higher partition coefficient
3 Particle size
Membrane proteins mediated transport of most molecules and all ions across biomembrane
The rate at which a molecule diffuses across a synthetic lipid bilayer depend on its size and solubilityThe smaller the molecule and the less polar it is the more rapidly it diffuses across the bilayer
Overview of membrane transport proteins
1 All transmembrane
proteins
2 Some transport has ATP binding sites
3 Move molecules uphill (向上) against its gradient
Differences1 Transporters=
-uniporters transport a single molecule down its gradient (passive)
-co-transporters couple movement of a molecule down itsgradient with moving a molecule up its gradient (active)
2 Pumps = hydrolyze ATP to move small moleculesions up a concentration gradient or electric potential (active)
3 Channels = transport waterionssmall molecules down their concentration gradients or electric potentials (passive)
The four mechanisms of small molecules and ions are transported cross cellular membranes
Ion rarr force
促進 主動
If transport substance carries a net charge its movement is influenced by both its concentration gradient and the membrane potential the electric potential (voltage) across the membrane
Substance concentration + electric potential = electrochemical gradient Determines the energetically favorable direction of transport a charged molecule across a membrane
Copyright copy 2009 Dr Salme Taagepera PhD All rights reserved
Passive transport driven by
Concentration gradient(affects both uncharged and
charged solutes)
Electrical gradient(affects only charged solutes)
ldquoElectrochemical gradientrdquo
+
被動 Facilitated Diffusion
Passive transport no metabolic energy is needed because the solute is moving down its concentration gradient
bullIn the case of an uncharged solute the concentration of the solute on each side of the membrane dictates the direction of passive transport
Active transport metabolic energy is used to transport a solute from the side of low concentration to the side of high concentration
Two types of transport are defined by whether metabolic energy is expended to move a solute across the membrane
Facilitated Diffusion
Free DiffusionA Non-channel mediated
ndash lipids gasses (O2 CO2) waterB Channel mediated
ndash ions charged molecules
Facilitated diffusionCarrier mediated
ndash glucose amino acids
[ECF]
Extracellular fluid
Facilitated Diffusion
Rate of diffusion is determined by1 concentration gradient 2 amount of carrier protein3 rate of associationdissociation
Unique features for Uniport transport
1 Higher diffusion rate for uniport than passive diffusion
2 Transported molecules never enter membrane and Irrelevant (無關) to the partition coefficient (did not cross membrane)
3 Transport rate reach Vmaxwhen each uniport working at its maximal rate
4 Transport is specific Each uniport transports only a single species of molecules or single or closely related molecules
Several feature distinguish uniport transport from passive diffusion
GLUT glucose transport
Need a carrier protein
Uniport transport of glucose and water
Glucose utilize glucose as a source for ATP production
Water utilize aquaporins to increase the rate of water movement
H2O hybrophilic did across membrane
Families of GLUT proteins(1-12)Highly homologous in sequence and contain 12 membrane-spanning α
-
helicesDifferent isoforms rarr different cell type expression and different functionGLUT2 express in liver cell ( glucose storage)
and szlig cell( glucose uptake) pancreasGLUT4 found in intracellular membrane
increase expression by insulin for remove the glucose from blood to cellGLUT5 tansport fructoseOther isoforms
Glucose transporter (GLUT) Facilitated Diffusion
Mammalian glucose transporters
Name Tissue distribution Proposed function
Glut1 all fetal and adult tissues basal glucose transport
Glut2 hepatocytes pancreatic β-cells transepithelial transport intestine kidney from and to the blood
Glut3 widely distributed basal glucose transportmostly in brain
Glut4 skeletal muscle heart adipocytes insulin-dependent transport
Glut5 intestine lesser amounts few others fructose transport
Glut7 gluconeogenic tissues mediates flux across endoplasmic reticulum
Glut8 preimplantation blastocyst embryonic insulin- dependent transport
Oded Meyuhas
Each GLUT protein contains 12 membrane-spanning alpha helicesDifferentially expressed
ndash EXAMPLE GLUT4 is only expressed in fat and muscle cells
bull Fat and muscle cells respond to insulin by increasing their uptake of glucose thereby removing glucose from the blood
bull In absence of insulin = GLUT4 on intracellular membranesIn presence of insulin = GLUT4 found on cell surface
bull QUESTION Defects in directing GLUT4 to the cell surface can cause what common disease
Type II diabetes high blood glucose
Copyright copy 2009 Dr Salme Taagepera PhD All rights reserved
GLUT1 is responsible for transporting glucose across the blood- brain barrier rarr
GLUT1 provides glucose for the brain
GLUT1 deficiency syndromebull Brain does not obtain enough glucose from the bloodbull Symptoms seizures developmental delay motor disordersbull Treatment ketogenic diet (high fatlow carb diet)
GLUT1 deficiency syndrome
Glucose
Transporter proteins (membrane protein) can be enriched within artificial membrane
Chloroform and methanol (31)
Phospholipid spontaneously form bilayers
Liposome containing a single type of transport protein are very useful in studying functional properties of transport proteins
bull It is a major experimental tool to study the biochemistry of transport protein function in vitro
bull Widely used as a drug delivery system and for gene transfection
Movement of waterOsmosis movement of water across semipermeable
membraneOsmotic pressure hydrostatic pressure uses to stop
the net flow of water
When CB concentration gt CA
Osmotic pressure π=RT( CB -CA )
Hypertonic solution the concentration is higher than cytosolIsotonic solution equal to cytosolHypotonic solution lower and water move to cytosol
Animal cell ace a problem in maintaining their cell volume within a limited range thereby avoiding lysis
Plant cell has cell wall rarr prevent cell shapeTurgor pressure (膨壓) osmotic pressure plasma membrane against water into the cytosol and then into the vacuole
turgor pressure supplies rigidityThe large forces of turgor pressure are resisted by the strength of cellulose microfibrils in the cell wall
Hypertonic external solution concentration of water is low relative to its concentration inside the cell
Water moves out down its concentration gradientand the cell shrinks
Hypotonic external solution concentration of water is highrelative to its concentration inside the cell
Water moves in down its concentration gradientand the cell swells
When the Na+ ndash K+ pump stops Na+ goes into the cell along its concentration gradient
This adds to the solute concentration in the cytosol
Water moves into the cell along its concentration gradient and the cell bursts
Water draw is equal inside and outside
Water cross membrane is very lowerWater cross membrane via specific channel- aquaporins
Aquaporins increase the water permeability of cell membrane
Aquaporin 1 erythrocyte
Aquaporin2 kidney cells resorb water from urine mutation rarr diabetes insipidus rarr large volume urine
Expression of aquaporin by frog oocytes increases their permeability
control
Injection aquaporin mRNA
Egg move to hypotonic environment
尿崩病
Water channel protein ( aquaporin)
Tetrameric protein
6 α-helices for each subunit
2-nm-long water selective gate
028nm gate widthHighly conserved arginine and histidine in the gateH2 O for HO bonding
with cystein
AquaporinsAquaporins are membrane water channels that play critical are membrane water channels that play critical roles in controlling the roles in controlling the waterwater contents of cellscontents of cells
Water Water crosses the crosses the hydrophobic membranehydrophobic membrane either by simple either by simple diffusion or through a facilitative transport mechanism diffusion or through a facilitative transport mechanism mediated by these mediated by these specialized proteinsspecialized proteins
These protein channels are These protein channels are widely distributedwidely distributed in all kingdoms of in all kingdoms of life including bacteria plants and mammalslife including bacteria plants and mammals
Important in Important in osmotic regulationosmotic regulation acting to prevent bursting of acting to prevent bursting of the cells whenever there are changes of the exterior salt the cells whenever there are changes of the exterior salt concentrationconcentration
ATP powered pump1 P- class
2α 2β
subunit can phosphorylationie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-classbull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-classmaintain low pH in plant vacuolesimilar to F-class
4 ABC (ATP-binding cassete) superfamilyseveral hundred different transport protein
Different classes of pumps exhibit characteristic structure and functional properties
Specific Ion binding site
Must phosphorylation
Transport process requires ATP hydrolysis in which the free energy is liberated by breakdown of ATP into ADP and phosphate
V-class H+ ATP ase pump protons across lysosomal and vacuolar membrane
inside
inside
ATP powered pump
1 P- class
2α 2β
subunit
ie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-class
bull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-class
maintain low pH in plant vacuole
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

A study of mutant zebrafish with pale stripes led to the identification of a sodiumcalcium transporter that regulates the darkness of human skin
The phospholipid bilayer is a barrier that controls the transport of molecules in and out of the cell
Gases diffuse freely no proteins required
Water diffuses fast enough that proteins arenrsquot required for transport
Sugars diffuse very slowly so proteins are involved in transport
Charged molecules are virtually impermeable
Studies of synthetic lipid bilayers
help define which types of transport will require the activity of a protein Hence transport of an ion should require
a protein
Only small hydrophobic molecules cross membrane
The bilayer is permeable toSmall hydrophobic moleculesSmall uncharged polar molecules
The bilayer is impermeable toIonsLarge polar molecules
THEREFORE need membrane proteins to transport most molecules and all ions across biomembranes
KEY CONCEPTS
Selective transport across the lipid membrane requires transport proteins
Transport proteins are integral membrane proteins that move molecules and ions
There are two classes of transport proteins transporters (pumps) and channels
Most small molecules did not across membrane
Transporter protein
Also called Na+K+ ATPasesodium-amino acid transporter
Three main class of membrane protein1ATP- power pump( carrier permease)
couple with energy source for active transportbinding of specific solute to transporter which
undergo conformation change2 Channel protein (ion channel)
formation of hydrophilic pore allow passive movement of small inorganic molecule
3 Transportersuniportsymportantiport
Partition Coefficient
Permeability coefficients (in cmsec) through synthetic lipid bilayers
Product of the concentration difference (in molcm3) and permeability coefficient (in cmsec) gives the flow of solute in moles per second per square centimeter of membrane
油品分配係數
Cell membrane
bullBarrier to the passage of most polar molecule
bullMaintain concentration of solute
Diffusion rate depends on
1 Concentration gradient or electrochemical gradient
2 Hydrophobicity
ie higher partition coefficient
3 Particle size
Membrane proteins mediated transport of most molecules and all ions across biomembrane
The rate at which a molecule diffuses across a synthetic lipid bilayer depend on its size and solubilityThe smaller the molecule and the less polar it is the more rapidly it diffuses across the bilayer
Overview of membrane transport proteins
1 All transmembrane
proteins
2 Some transport has ATP binding sites
3 Move molecules uphill (向上) against its gradient
Differences1 Transporters=
-uniporters transport a single molecule down its gradient (passive)
-co-transporters couple movement of a molecule down itsgradient with moving a molecule up its gradient (active)
2 Pumps = hydrolyze ATP to move small moleculesions up a concentration gradient or electric potential (active)
3 Channels = transport waterionssmall molecules down their concentration gradients or electric potentials (passive)
The four mechanisms of small molecules and ions are transported cross cellular membranes
Ion rarr force
促進 主動
If transport substance carries a net charge its movement is influenced by both its concentration gradient and the membrane potential the electric potential (voltage) across the membrane
Substance concentration + electric potential = electrochemical gradient Determines the energetically favorable direction of transport a charged molecule across a membrane
Copyright copy 2009 Dr Salme Taagepera PhD All rights reserved
Passive transport driven by
Concentration gradient(affects both uncharged and
charged solutes)
Electrical gradient(affects only charged solutes)
ldquoElectrochemical gradientrdquo
+
被動 Facilitated Diffusion
Passive transport no metabolic energy is needed because the solute is moving down its concentration gradient
bullIn the case of an uncharged solute the concentration of the solute on each side of the membrane dictates the direction of passive transport
Active transport metabolic energy is used to transport a solute from the side of low concentration to the side of high concentration
Two types of transport are defined by whether metabolic energy is expended to move a solute across the membrane
Facilitated Diffusion
Free DiffusionA Non-channel mediated
ndash lipids gasses (O2 CO2) waterB Channel mediated
ndash ions charged molecules
Facilitated diffusionCarrier mediated
ndash glucose amino acids
[ECF]
Extracellular fluid
Facilitated Diffusion
Rate of diffusion is determined by1 concentration gradient 2 amount of carrier protein3 rate of associationdissociation
Unique features for Uniport transport
1 Higher diffusion rate for uniport than passive diffusion
2 Transported molecules never enter membrane and Irrelevant (無關) to the partition coefficient (did not cross membrane)
3 Transport rate reach Vmaxwhen each uniport working at its maximal rate
4 Transport is specific Each uniport transports only a single species of molecules or single or closely related molecules
Several feature distinguish uniport transport from passive diffusion
GLUT glucose transport
Need a carrier protein
Uniport transport of glucose and water
Glucose utilize glucose as a source for ATP production
Water utilize aquaporins to increase the rate of water movement
H2O hybrophilic did across membrane
Families of GLUT proteins(1-12)Highly homologous in sequence and contain 12 membrane-spanning α
-
helicesDifferent isoforms rarr different cell type expression and different functionGLUT2 express in liver cell ( glucose storage)
and szlig cell( glucose uptake) pancreasGLUT4 found in intracellular membrane
increase expression by insulin for remove the glucose from blood to cellGLUT5 tansport fructoseOther isoforms
Glucose transporter (GLUT) Facilitated Diffusion
Mammalian glucose transporters
Name Tissue distribution Proposed function
Glut1 all fetal and adult tissues basal glucose transport
Glut2 hepatocytes pancreatic β-cells transepithelial transport intestine kidney from and to the blood
Glut3 widely distributed basal glucose transportmostly in brain
Glut4 skeletal muscle heart adipocytes insulin-dependent transport
Glut5 intestine lesser amounts few others fructose transport
Glut7 gluconeogenic tissues mediates flux across endoplasmic reticulum
Glut8 preimplantation blastocyst embryonic insulin- dependent transport
Oded Meyuhas
Each GLUT protein contains 12 membrane-spanning alpha helicesDifferentially expressed
ndash EXAMPLE GLUT4 is only expressed in fat and muscle cells
bull Fat and muscle cells respond to insulin by increasing their uptake of glucose thereby removing glucose from the blood
bull In absence of insulin = GLUT4 on intracellular membranesIn presence of insulin = GLUT4 found on cell surface
bull QUESTION Defects in directing GLUT4 to the cell surface can cause what common disease
Type II diabetes high blood glucose
Copyright copy 2009 Dr Salme Taagepera PhD All rights reserved
GLUT1 is responsible for transporting glucose across the blood- brain barrier rarr
GLUT1 provides glucose for the brain
GLUT1 deficiency syndromebull Brain does not obtain enough glucose from the bloodbull Symptoms seizures developmental delay motor disordersbull Treatment ketogenic diet (high fatlow carb diet)
GLUT1 deficiency syndrome
Glucose
Transporter proteins (membrane protein) can be enriched within artificial membrane
Chloroform and methanol (31)
Phospholipid spontaneously form bilayers
Liposome containing a single type of transport protein are very useful in studying functional properties of transport proteins
bull It is a major experimental tool to study the biochemistry of transport protein function in vitro
bull Widely used as a drug delivery system and for gene transfection
Movement of waterOsmosis movement of water across semipermeable
membraneOsmotic pressure hydrostatic pressure uses to stop
the net flow of water
When CB concentration gt CA
Osmotic pressure π=RT( CB -CA )
Hypertonic solution the concentration is higher than cytosolIsotonic solution equal to cytosolHypotonic solution lower and water move to cytosol
Animal cell ace a problem in maintaining their cell volume within a limited range thereby avoiding lysis
Plant cell has cell wall rarr prevent cell shapeTurgor pressure (膨壓) osmotic pressure plasma membrane against water into the cytosol and then into the vacuole
turgor pressure supplies rigidityThe large forces of turgor pressure are resisted by the strength of cellulose microfibrils in the cell wall
Hypertonic external solution concentration of water is low relative to its concentration inside the cell
Water moves out down its concentration gradientand the cell shrinks
Hypotonic external solution concentration of water is highrelative to its concentration inside the cell
Water moves in down its concentration gradientand the cell swells
When the Na+ ndash K+ pump stops Na+ goes into the cell along its concentration gradient
This adds to the solute concentration in the cytosol
Water moves into the cell along its concentration gradient and the cell bursts
Water draw is equal inside and outside
Water cross membrane is very lowerWater cross membrane via specific channel- aquaporins
Aquaporins increase the water permeability of cell membrane
Aquaporin 1 erythrocyte
Aquaporin2 kidney cells resorb water from urine mutation rarr diabetes insipidus rarr large volume urine
Expression of aquaporin by frog oocytes increases their permeability
control
Injection aquaporin mRNA
Egg move to hypotonic environment
尿崩病
Water channel protein ( aquaporin)
Tetrameric protein
6 α-helices for each subunit
2-nm-long water selective gate
028nm gate widthHighly conserved arginine and histidine in the gateH2 O for HO bonding
with cystein
AquaporinsAquaporins are membrane water channels that play critical are membrane water channels that play critical roles in controlling the roles in controlling the waterwater contents of cellscontents of cells
Water Water crosses the crosses the hydrophobic membranehydrophobic membrane either by simple either by simple diffusion or through a facilitative transport mechanism diffusion or through a facilitative transport mechanism mediated by these mediated by these specialized proteinsspecialized proteins
These protein channels are These protein channels are widely distributedwidely distributed in all kingdoms of in all kingdoms of life including bacteria plants and mammalslife including bacteria plants and mammals
Important in Important in osmotic regulationosmotic regulation acting to prevent bursting of acting to prevent bursting of the cells whenever there are changes of the exterior salt the cells whenever there are changes of the exterior salt concentrationconcentration
ATP powered pump1 P- class
2α 2β
subunit can phosphorylationie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-classbull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-classmaintain low pH in plant vacuolesimilar to F-class
4 ABC (ATP-binding cassete) superfamilyseveral hundred different transport protein
Different classes of pumps exhibit characteristic structure and functional properties
Specific Ion binding site
Must phosphorylation
Transport process requires ATP hydrolysis in which the free energy is liberated by breakdown of ATP into ADP and phosphate
V-class H+ ATP ase pump protons across lysosomal and vacuolar membrane
inside
inside
ATP powered pump
1 P- class
2α 2β
subunit
ie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-class
bull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-class
maintain low pH in plant vacuole
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

The phospholipid bilayer is a barrier that controls the transport of molecules in and out of the cell
Gases diffuse freely no proteins required
Water diffuses fast enough that proteins arenrsquot required for transport
Sugars diffuse very slowly so proteins are involved in transport
Charged molecules are virtually impermeable
Studies of synthetic lipid bilayers
help define which types of transport will require the activity of a protein Hence transport of an ion should require
a protein
Only small hydrophobic molecules cross membrane
The bilayer is permeable toSmall hydrophobic moleculesSmall uncharged polar molecules
The bilayer is impermeable toIonsLarge polar molecules
THEREFORE need membrane proteins to transport most molecules and all ions across biomembranes
KEY CONCEPTS
Selective transport across the lipid membrane requires transport proteins
Transport proteins are integral membrane proteins that move molecules and ions
There are two classes of transport proteins transporters (pumps) and channels
Most small molecules did not across membrane
Transporter protein
Also called Na+K+ ATPasesodium-amino acid transporter
Three main class of membrane protein1ATP- power pump( carrier permease)
couple with energy source for active transportbinding of specific solute to transporter which
undergo conformation change2 Channel protein (ion channel)
formation of hydrophilic pore allow passive movement of small inorganic molecule
3 Transportersuniportsymportantiport
Partition Coefficient
Permeability coefficients (in cmsec) through synthetic lipid bilayers
Product of the concentration difference (in molcm3) and permeability coefficient (in cmsec) gives the flow of solute in moles per second per square centimeter of membrane
油品分配係數
Cell membrane
bullBarrier to the passage of most polar molecule
bullMaintain concentration of solute
Diffusion rate depends on
1 Concentration gradient or electrochemical gradient
2 Hydrophobicity
ie higher partition coefficient
3 Particle size
Membrane proteins mediated transport of most molecules and all ions across biomembrane
The rate at which a molecule diffuses across a synthetic lipid bilayer depend on its size and solubilityThe smaller the molecule and the less polar it is the more rapidly it diffuses across the bilayer
Overview of membrane transport proteins
1 All transmembrane
proteins
2 Some transport has ATP binding sites
3 Move molecules uphill (向上) against its gradient
Differences1 Transporters=
-uniporters transport a single molecule down its gradient (passive)
-co-transporters couple movement of a molecule down itsgradient with moving a molecule up its gradient (active)
2 Pumps = hydrolyze ATP to move small moleculesions up a concentration gradient or electric potential (active)
3 Channels = transport waterionssmall molecules down their concentration gradients or electric potentials (passive)
The four mechanisms of small molecules and ions are transported cross cellular membranes
Ion rarr force
促進 主動
If transport substance carries a net charge its movement is influenced by both its concentration gradient and the membrane potential the electric potential (voltage) across the membrane
Substance concentration + electric potential = electrochemical gradient Determines the energetically favorable direction of transport a charged molecule across a membrane
Copyright copy 2009 Dr Salme Taagepera PhD All rights reserved
Passive transport driven by
Concentration gradient(affects both uncharged and
charged solutes)
Electrical gradient(affects only charged solutes)
ldquoElectrochemical gradientrdquo
+
被動 Facilitated Diffusion
Passive transport no metabolic energy is needed because the solute is moving down its concentration gradient
bullIn the case of an uncharged solute the concentration of the solute on each side of the membrane dictates the direction of passive transport
Active transport metabolic energy is used to transport a solute from the side of low concentration to the side of high concentration
Two types of transport are defined by whether metabolic energy is expended to move a solute across the membrane
Facilitated Diffusion
Free DiffusionA Non-channel mediated
ndash lipids gasses (O2 CO2) waterB Channel mediated
ndash ions charged molecules
Facilitated diffusionCarrier mediated
ndash glucose amino acids
[ECF]
Extracellular fluid
Facilitated Diffusion
Rate of diffusion is determined by1 concentration gradient 2 amount of carrier protein3 rate of associationdissociation
Unique features for Uniport transport
1 Higher diffusion rate for uniport than passive diffusion
2 Transported molecules never enter membrane and Irrelevant (無關) to the partition coefficient (did not cross membrane)
3 Transport rate reach Vmaxwhen each uniport working at its maximal rate
4 Transport is specific Each uniport transports only a single species of molecules or single or closely related molecules
Several feature distinguish uniport transport from passive diffusion
GLUT glucose transport
Need a carrier protein
Uniport transport of glucose and water
Glucose utilize glucose as a source for ATP production
Water utilize aquaporins to increase the rate of water movement
H2O hybrophilic did across membrane
Families of GLUT proteins(1-12)Highly homologous in sequence and contain 12 membrane-spanning α
-
helicesDifferent isoforms rarr different cell type expression and different functionGLUT2 express in liver cell ( glucose storage)
and szlig cell( glucose uptake) pancreasGLUT4 found in intracellular membrane
increase expression by insulin for remove the glucose from blood to cellGLUT5 tansport fructoseOther isoforms
Glucose transporter (GLUT) Facilitated Diffusion
Mammalian glucose transporters
Name Tissue distribution Proposed function
Glut1 all fetal and adult tissues basal glucose transport
Glut2 hepatocytes pancreatic β-cells transepithelial transport intestine kidney from and to the blood
Glut3 widely distributed basal glucose transportmostly in brain
Glut4 skeletal muscle heart adipocytes insulin-dependent transport
Glut5 intestine lesser amounts few others fructose transport
Glut7 gluconeogenic tissues mediates flux across endoplasmic reticulum
Glut8 preimplantation blastocyst embryonic insulin- dependent transport
Oded Meyuhas
Each GLUT protein contains 12 membrane-spanning alpha helicesDifferentially expressed
ndash EXAMPLE GLUT4 is only expressed in fat and muscle cells
bull Fat and muscle cells respond to insulin by increasing their uptake of glucose thereby removing glucose from the blood
bull In absence of insulin = GLUT4 on intracellular membranesIn presence of insulin = GLUT4 found on cell surface
bull QUESTION Defects in directing GLUT4 to the cell surface can cause what common disease
Type II diabetes high blood glucose
Copyright copy 2009 Dr Salme Taagepera PhD All rights reserved
GLUT1 is responsible for transporting glucose across the blood- brain barrier rarr
GLUT1 provides glucose for the brain
GLUT1 deficiency syndromebull Brain does not obtain enough glucose from the bloodbull Symptoms seizures developmental delay motor disordersbull Treatment ketogenic diet (high fatlow carb diet)
GLUT1 deficiency syndrome
Glucose
Transporter proteins (membrane protein) can be enriched within artificial membrane
Chloroform and methanol (31)
Phospholipid spontaneously form bilayers
Liposome containing a single type of transport protein are very useful in studying functional properties of transport proteins
bull It is a major experimental tool to study the biochemistry of transport protein function in vitro
bull Widely used as a drug delivery system and for gene transfection
Movement of waterOsmosis movement of water across semipermeable
membraneOsmotic pressure hydrostatic pressure uses to stop
the net flow of water
When CB concentration gt CA
Osmotic pressure π=RT( CB -CA )
Hypertonic solution the concentration is higher than cytosolIsotonic solution equal to cytosolHypotonic solution lower and water move to cytosol
Animal cell ace a problem in maintaining their cell volume within a limited range thereby avoiding lysis
Plant cell has cell wall rarr prevent cell shapeTurgor pressure (膨壓) osmotic pressure plasma membrane against water into the cytosol and then into the vacuole
turgor pressure supplies rigidityThe large forces of turgor pressure are resisted by the strength of cellulose microfibrils in the cell wall
Hypertonic external solution concentration of water is low relative to its concentration inside the cell
Water moves out down its concentration gradientand the cell shrinks
Hypotonic external solution concentration of water is highrelative to its concentration inside the cell
Water moves in down its concentration gradientand the cell swells
When the Na+ ndash K+ pump stops Na+ goes into the cell along its concentration gradient
This adds to the solute concentration in the cytosol
Water moves into the cell along its concentration gradient and the cell bursts
Water draw is equal inside and outside
Water cross membrane is very lowerWater cross membrane via specific channel- aquaporins
Aquaporins increase the water permeability of cell membrane
Aquaporin 1 erythrocyte
Aquaporin2 kidney cells resorb water from urine mutation rarr diabetes insipidus rarr large volume urine
Expression of aquaporin by frog oocytes increases their permeability
control
Injection aquaporin mRNA
Egg move to hypotonic environment
尿崩病
Water channel protein ( aquaporin)
Tetrameric protein
6 α-helices for each subunit
2-nm-long water selective gate
028nm gate widthHighly conserved arginine and histidine in the gateH2 O for HO bonding
with cystein
AquaporinsAquaporins are membrane water channels that play critical are membrane water channels that play critical roles in controlling the roles in controlling the waterwater contents of cellscontents of cells
Water Water crosses the crosses the hydrophobic membranehydrophobic membrane either by simple either by simple diffusion or through a facilitative transport mechanism diffusion or through a facilitative transport mechanism mediated by these mediated by these specialized proteinsspecialized proteins
These protein channels are These protein channels are widely distributedwidely distributed in all kingdoms of in all kingdoms of life including bacteria plants and mammalslife including bacteria plants and mammals
Important in Important in osmotic regulationosmotic regulation acting to prevent bursting of acting to prevent bursting of the cells whenever there are changes of the exterior salt the cells whenever there are changes of the exterior salt concentrationconcentration
ATP powered pump1 P- class
2α 2β
subunit can phosphorylationie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-classbull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-classmaintain low pH in plant vacuolesimilar to F-class
4 ABC (ATP-binding cassete) superfamilyseveral hundred different transport protein
Different classes of pumps exhibit characteristic structure and functional properties
Specific Ion binding site
Must phosphorylation
Transport process requires ATP hydrolysis in which the free energy is liberated by breakdown of ATP into ADP and phosphate
V-class H+ ATP ase pump protons across lysosomal and vacuolar membrane
inside
inside
ATP powered pump
1 P- class
2α 2β
subunit
ie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-class
bull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-class
maintain low pH in plant vacuole
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

The bilayer is permeable toSmall hydrophobic moleculesSmall uncharged polar molecules
The bilayer is impermeable toIonsLarge polar molecules
THEREFORE need membrane proteins to transport most molecules and all ions across biomembranes
KEY CONCEPTS
Selective transport across the lipid membrane requires transport proteins
Transport proteins are integral membrane proteins that move molecules and ions
There are two classes of transport proteins transporters (pumps) and channels
Most small molecules did not across membrane
Transporter protein
Also called Na+K+ ATPasesodium-amino acid transporter
Three main class of membrane protein1ATP- power pump( carrier permease)
couple with energy source for active transportbinding of specific solute to transporter which
undergo conformation change2 Channel protein (ion channel)
formation of hydrophilic pore allow passive movement of small inorganic molecule
3 Transportersuniportsymportantiport
Partition Coefficient
Permeability coefficients (in cmsec) through synthetic lipid bilayers
Product of the concentration difference (in molcm3) and permeability coefficient (in cmsec) gives the flow of solute in moles per second per square centimeter of membrane
油品分配係數
Cell membrane
bullBarrier to the passage of most polar molecule
bullMaintain concentration of solute
Diffusion rate depends on
1 Concentration gradient or electrochemical gradient
2 Hydrophobicity
ie higher partition coefficient
3 Particle size
Membrane proteins mediated transport of most molecules and all ions across biomembrane
The rate at which a molecule diffuses across a synthetic lipid bilayer depend on its size and solubilityThe smaller the molecule and the less polar it is the more rapidly it diffuses across the bilayer
Overview of membrane transport proteins
1 All transmembrane
proteins
2 Some transport has ATP binding sites
3 Move molecules uphill (向上) against its gradient
Differences1 Transporters=
-uniporters transport a single molecule down its gradient (passive)
-co-transporters couple movement of a molecule down itsgradient with moving a molecule up its gradient (active)
2 Pumps = hydrolyze ATP to move small moleculesions up a concentration gradient or electric potential (active)
3 Channels = transport waterionssmall molecules down their concentration gradients or electric potentials (passive)
The four mechanisms of small molecules and ions are transported cross cellular membranes
Ion rarr force
促進 主動
If transport substance carries a net charge its movement is influenced by both its concentration gradient and the membrane potential the electric potential (voltage) across the membrane
Substance concentration + electric potential = electrochemical gradient Determines the energetically favorable direction of transport a charged molecule across a membrane
Copyright copy 2009 Dr Salme Taagepera PhD All rights reserved
Passive transport driven by
Concentration gradient(affects both uncharged and
charged solutes)
Electrical gradient(affects only charged solutes)
ldquoElectrochemical gradientrdquo
+
被動 Facilitated Diffusion
Passive transport no metabolic energy is needed because the solute is moving down its concentration gradient
bullIn the case of an uncharged solute the concentration of the solute on each side of the membrane dictates the direction of passive transport
Active transport metabolic energy is used to transport a solute from the side of low concentration to the side of high concentration
Two types of transport are defined by whether metabolic energy is expended to move a solute across the membrane
Facilitated Diffusion
Free DiffusionA Non-channel mediated
ndash lipids gasses (O2 CO2) waterB Channel mediated
ndash ions charged molecules
Facilitated diffusionCarrier mediated
ndash glucose amino acids
[ECF]
Extracellular fluid
Facilitated Diffusion
Rate of diffusion is determined by1 concentration gradient 2 amount of carrier protein3 rate of associationdissociation
Unique features for Uniport transport
1 Higher diffusion rate for uniport than passive diffusion
2 Transported molecules never enter membrane and Irrelevant (無關) to the partition coefficient (did not cross membrane)
3 Transport rate reach Vmaxwhen each uniport working at its maximal rate
4 Transport is specific Each uniport transports only a single species of molecules or single or closely related molecules
Several feature distinguish uniport transport from passive diffusion
GLUT glucose transport
Need a carrier protein
Uniport transport of glucose and water
Glucose utilize glucose as a source for ATP production
Water utilize aquaporins to increase the rate of water movement
H2O hybrophilic did across membrane
Families of GLUT proteins(1-12)Highly homologous in sequence and contain 12 membrane-spanning α
-
helicesDifferent isoforms rarr different cell type expression and different functionGLUT2 express in liver cell ( glucose storage)
and szlig cell( glucose uptake) pancreasGLUT4 found in intracellular membrane
increase expression by insulin for remove the glucose from blood to cellGLUT5 tansport fructoseOther isoforms
Glucose transporter (GLUT) Facilitated Diffusion
Mammalian glucose transporters
Name Tissue distribution Proposed function
Glut1 all fetal and adult tissues basal glucose transport
Glut2 hepatocytes pancreatic β-cells transepithelial transport intestine kidney from and to the blood
Glut3 widely distributed basal glucose transportmostly in brain
Glut4 skeletal muscle heart adipocytes insulin-dependent transport
Glut5 intestine lesser amounts few others fructose transport
Glut7 gluconeogenic tissues mediates flux across endoplasmic reticulum
Glut8 preimplantation blastocyst embryonic insulin- dependent transport
Oded Meyuhas
Each GLUT protein contains 12 membrane-spanning alpha helicesDifferentially expressed
ndash EXAMPLE GLUT4 is only expressed in fat and muscle cells
bull Fat and muscle cells respond to insulin by increasing their uptake of glucose thereby removing glucose from the blood
bull In absence of insulin = GLUT4 on intracellular membranesIn presence of insulin = GLUT4 found on cell surface
bull QUESTION Defects in directing GLUT4 to the cell surface can cause what common disease
Type II diabetes high blood glucose
Copyright copy 2009 Dr Salme Taagepera PhD All rights reserved
GLUT1 is responsible for transporting glucose across the blood- brain barrier rarr
GLUT1 provides glucose for the brain
GLUT1 deficiency syndromebull Brain does not obtain enough glucose from the bloodbull Symptoms seizures developmental delay motor disordersbull Treatment ketogenic diet (high fatlow carb diet)
GLUT1 deficiency syndrome
Glucose
Transporter proteins (membrane protein) can be enriched within artificial membrane
Chloroform and methanol (31)
Phospholipid spontaneously form bilayers
Liposome containing a single type of transport protein are very useful in studying functional properties of transport proteins
bull It is a major experimental tool to study the biochemistry of transport protein function in vitro
bull Widely used as a drug delivery system and for gene transfection
Movement of waterOsmosis movement of water across semipermeable
membraneOsmotic pressure hydrostatic pressure uses to stop
the net flow of water
When CB concentration gt CA
Osmotic pressure π=RT( CB -CA )
Hypertonic solution the concentration is higher than cytosolIsotonic solution equal to cytosolHypotonic solution lower and water move to cytosol
Animal cell ace a problem in maintaining their cell volume within a limited range thereby avoiding lysis
Plant cell has cell wall rarr prevent cell shapeTurgor pressure (膨壓) osmotic pressure plasma membrane against water into the cytosol and then into the vacuole
turgor pressure supplies rigidityThe large forces of turgor pressure are resisted by the strength of cellulose microfibrils in the cell wall
Hypertonic external solution concentration of water is low relative to its concentration inside the cell
Water moves out down its concentration gradientand the cell shrinks
Hypotonic external solution concentration of water is highrelative to its concentration inside the cell
Water moves in down its concentration gradientand the cell swells
When the Na+ ndash K+ pump stops Na+ goes into the cell along its concentration gradient
This adds to the solute concentration in the cytosol
Water moves into the cell along its concentration gradient and the cell bursts
Water draw is equal inside and outside
Water cross membrane is very lowerWater cross membrane via specific channel- aquaporins
Aquaporins increase the water permeability of cell membrane
Aquaporin 1 erythrocyte
Aquaporin2 kidney cells resorb water from urine mutation rarr diabetes insipidus rarr large volume urine
Expression of aquaporin by frog oocytes increases their permeability
control
Injection aquaporin mRNA
Egg move to hypotonic environment
尿崩病
Water channel protein ( aquaporin)
Tetrameric protein
6 α-helices for each subunit
2-nm-long water selective gate
028nm gate widthHighly conserved arginine and histidine in the gateH2 O for HO bonding
with cystein
AquaporinsAquaporins are membrane water channels that play critical are membrane water channels that play critical roles in controlling the roles in controlling the waterwater contents of cellscontents of cells
Water Water crosses the crosses the hydrophobic membranehydrophobic membrane either by simple either by simple diffusion or through a facilitative transport mechanism diffusion or through a facilitative transport mechanism mediated by these mediated by these specialized proteinsspecialized proteins
These protein channels are These protein channels are widely distributedwidely distributed in all kingdoms of in all kingdoms of life including bacteria plants and mammalslife including bacteria plants and mammals
Important in Important in osmotic regulationosmotic regulation acting to prevent bursting of acting to prevent bursting of the cells whenever there are changes of the exterior salt the cells whenever there are changes of the exterior salt concentrationconcentration
ATP powered pump1 P- class
2α 2β
subunit can phosphorylationie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-classbull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-classmaintain low pH in plant vacuolesimilar to F-class
4 ABC (ATP-binding cassete) superfamilyseveral hundred different transport protein
Different classes of pumps exhibit characteristic structure and functional properties
Specific Ion binding site
Must phosphorylation
Transport process requires ATP hydrolysis in which the free energy is liberated by breakdown of ATP into ADP and phosphate
V-class H+ ATP ase pump protons across lysosomal and vacuolar membrane
inside
inside
ATP powered pump
1 P- class
2α 2β
subunit
ie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-class
bull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-class
maintain low pH in plant vacuole
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

KEY CONCEPTS
Selective transport across the lipid membrane requires transport proteins
Transport proteins are integral membrane proteins that move molecules and ions
There are two classes of transport proteins transporters (pumps) and channels
Most small molecules did not across membrane
Transporter protein
Also called Na+K+ ATPasesodium-amino acid transporter
Three main class of membrane protein1ATP- power pump( carrier permease)
couple with energy source for active transportbinding of specific solute to transporter which
undergo conformation change2 Channel protein (ion channel)
formation of hydrophilic pore allow passive movement of small inorganic molecule
3 Transportersuniportsymportantiport
Partition Coefficient
Permeability coefficients (in cmsec) through synthetic lipid bilayers
Product of the concentration difference (in molcm3) and permeability coefficient (in cmsec) gives the flow of solute in moles per second per square centimeter of membrane
油品分配係數
Cell membrane
bullBarrier to the passage of most polar molecule
bullMaintain concentration of solute
Diffusion rate depends on
1 Concentration gradient or electrochemical gradient
2 Hydrophobicity
ie higher partition coefficient
3 Particle size
Membrane proteins mediated transport of most molecules and all ions across biomembrane
The rate at which a molecule diffuses across a synthetic lipid bilayer depend on its size and solubilityThe smaller the molecule and the less polar it is the more rapidly it diffuses across the bilayer
Overview of membrane transport proteins
1 All transmembrane
proteins
2 Some transport has ATP binding sites
3 Move molecules uphill (向上) against its gradient
Differences1 Transporters=
-uniporters transport a single molecule down its gradient (passive)
-co-transporters couple movement of a molecule down itsgradient with moving a molecule up its gradient (active)
2 Pumps = hydrolyze ATP to move small moleculesions up a concentration gradient or electric potential (active)
3 Channels = transport waterionssmall molecules down their concentration gradients or electric potentials (passive)
The four mechanisms of small molecules and ions are transported cross cellular membranes
Ion rarr force
促進 主動
If transport substance carries a net charge its movement is influenced by both its concentration gradient and the membrane potential the electric potential (voltage) across the membrane
Substance concentration + electric potential = electrochemical gradient Determines the energetically favorable direction of transport a charged molecule across a membrane
Copyright copy 2009 Dr Salme Taagepera PhD All rights reserved
Passive transport driven by
Concentration gradient(affects both uncharged and
charged solutes)
Electrical gradient(affects only charged solutes)
ldquoElectrochemical gradientrdquo
+
被動 Facilitated Diffusion
Passive transport no metabolic energy is needed because the solute is moving down its concentration gradient
bullIn the case of an uncharged solute the concentration of the solute on each side of the membrane dictates the direction of passive transport
Active transport metabolic energy is used to transport a solute from the side of low concentration to the side of high concentration
Two types of transport are defined by whether metabolic energy is expended to move a solute across the membrane
Facilitated Diffusion
Free DiffusionA Non-channel mediated
ndash lipids gasses (O2 CO2) waterB Channel mediated
ndash ions charged molecules
Facilitated diffusionCarrier mediated
ndash glucose amino acids
[ECF]
Extracellular fluid
Facilitated Diffusion
Rate of diffusion is determined by1 concentration gradient 2 amount of carrier protein3 rate of associationdissociation
Unique features for Uniport transport
1 Higher diffusion rate for uniport than passive diffusion
2 Transported molecules never enter membrane and Irrelevant (無關) to the partition coefficient (did not cross membrane)
3 Transport rate reach Vmaxwhen each uniport working at its maximal rate
4 Transport is specific Each uniport transports only a single species of molecules or single or closely related molecules
Several feature distinguish uniport transport from passive diffusion
GLUT glucose transport
Need a carrier protein
Uniport transport of glucose and water
Glucose utilize glucose as a source for ATP production
Water utilize aquaporins to increase the rate of water movement
H2O hybrophilic did across membrane
Families of GLUT proteins(1-12)Highly homologous in sequence and contain 12 membrane-spanning α
-
helicesDifferent isoforms rarr different cell type expression and different functionGLUT2 express in liver cell ( glucose storage)
and szlig cell( glucose uptake) pancreasGLUT4 found in intracellular membrane
increase expression by insulin for remove the glucose from blood to cellGLUT5 tansport fructoseOther isoforms
Glucose transporter (GLUT) Facilitated Diffusion
Mammalian glucose transporters
Name Tissue distribution Proposed function
Glut1 all fetal and adult tissues basal glucose transport
Glut2 hepatocytes pancreatic β-cells transepithelial transport intestine kidney from and to the blood
Glut3 widely distributed basal glucose transportmostly in brain
Glut4 skeletal muscle heart adipocytes insulin-dependent transport
Glut5 intestine lesser amounts few others fructose transport
Glut7 gluconeogenic tissues mediates flux across endoplasmic reticulum
Glut8 preimplantation blastocyst embryonic insulin- dependent transport
Oded Meyuhas
Each GLUT protein contains 12 membrane-spanning alpha helicesDifferentially expressed
ndash EXAMPLE GLUT4 is only expressed in fat and muscle cells
bull Fat and muscle cells respond to insulin by increasing their uptake of glucose thereby removing glucose from the blood
bull In absence of insulin = GLUT4 on intracellular membranesIn presence of insulin = GLUT4 found on cell surface
bull QUESTION Defects in directing GLUT4 to the cell surface can cause what common disease
Type II diabetes high blood glucose
Copyright copy 2009 Dr Salme Taagepera PhD All rights reserved
GLUT1 is responsible for transporting glucose across the blood- brain barrier rarr
GLUT1 provides glucose for the brain
GLUT1 deficiency syndromebull Brain does not obtain enough glucose from the bloodbull Symptoms seizures developmental delay motor disordersbull Treatment ketogenic diet (high fatlow carb diet)
GLUT1 deficiency syndrome
Glucose
Transporter proteins (membrane protein) can be enriched within artificial membrane
Chloroform and methanol (31)
Phospholipid spontaneously form bilayers
Liposome containing a single type of transport protein are very useful in studying functional properties of transport proteins
bull It is a major experimental tool to study the biochemistry of transport protein function in vitro
bull Widely used as a drug delivery system and for gene transfection
Movement of waterOsmosis movement of water across semipermeable
membraneOsmotic pressure hydrostatic pressure uses to stop
the net flow of water
When CB concentration gt CA
Osmotic pressure π=RT( CB -CA )
Hypertonic solution the concentration is higher than cytosolIsotonic solution equal to cytosolHypotonic solution lower and water move to cytosol
Animal cell ace a problem in maintaining their cell volume within a limited range thereby avoiding lysis
Plant cell has cell wall rarr prevent cell shapeTurgor pressure (膨壓) osmotic pressure plasma membrane against water into the cytosol and then into the vacuole
turgor pressure supplies rigidityThe large forces of turgor pressure are resisted by the strength of cellulose microfibrils in the cell wall
Hypertonic external solution concentration of water is low relative to its concentration inside the cell
Water moves out down its concentration gradientand the cell shrinks
Hypotonic external solution concentration of water is highrelative to its concentration inside the cell
Water moves in down its concentration gradientand the cell swells
When the Na+ ndash K+ pump stops Na+ goes into the cell along its concentration gradient
This adds to the solute concentration in the cytosol
Water moves into the cell along its concentration gradient and the cell bursts
Water draw is equal inside and outside
Water cross membrane is very lowerWater cross membrane via specific channel- aquaporins
Aquaporins increase the water permeability of cell membrane
Aquaporin 1 erythrocyte
Aquaporin2 kidney cells resorb water from urine mutation rarr diabetes insipidus rarr large volume urine
Expression of aquaporin by frog oocytes increases their permeability
control
Injection aquaporin mRNA
Egg move to hypotonic environment
尿崩病
Water channel protein ( aquaporin)
Tetrameric protein
6 α-helices for each subunit
2-nm-long water selective gate
028nm gate widthHighly conserved arginine and histidine in the gateH2 O for HO bonding
with cystein
AquaporinsAquaporins are membrane water channels that play critical are membrane water channels that play critical roles in controlling the roles in controlling the waterwater contents of cellscontents of cells
Water Water crosses the crosses the hydrophobic membranehydrophobic membrane either by simple either by simple diffusion or through a facilitative transport mechanism diffusion or through a facilitative transport mechanism mediated by these mediated by these specialized proteinsspecialized proteins
These protein channels are These protein channels are widely distributedwidely distributed in all kingdoms of in all kingdoms of life including bacteria plants and mammalslife including bacteria plants and mammals
Important in Important in osmotic regulationosmotic regulation acting to prevent bursting of acting to prevent bursting of the cells whenever there are changes of the exterior salt the cells whenever there are changes of the exterior salt concentrationconcentration
ATP powered pump1 P- class
2α 2β
subunit can phosphorylationie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-classbull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-classmaintain low pH in plant vacuolesimilar to F-class
4 ABC (ATP-binding cassete) superfamilyseveral hundred different transport protein
Different classes of pumps exhibit characteristic structure and functional properties
Specific Ion binding site
Must phosphorylation
Transport process requires ATP hydrolysis in which the free energy is liberated by breakdown of ATP into ADP and phosphate
V-class H+ ATP ase pump protons across lysosomal and vacuolar membrane
inside
inside
ATP powered pump
1 P- class
2α 2β
subunit
ie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-class
bull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-class
maintain low pH in plant vacuole
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Most small molecules did not across membrane
Transporter protein
Also called Na+K+ ATPasesodium-amino acid transporter
Three main class of membrane protein1ATP- power pump( carrier permease)
couple with energy source for active transportbinding of specific solute to transporter which
undergo conformation change2 Channel protein (ion channel)
formation of hydrophilic pore allow passive movement of small inorganic molecule
3 Transportersuniportsymportantiport
Partition Coefficient
Permeability coefficients (in cmsec) through synthetic lipid bilayers
Product of the concentration difference (in molcm3) and permeability coefficient (in cmsec) gives the flow of solute in moles per second per square centimeter of membrane
油品分配係數
Cell membrane
bullBarrier to the passage of most polar molecule
bullMaintain concentration of solute
Diffusion rate depends on
1 Concentration gradient or electrochemical gradient
2 Hydrophobicity
ie higher partition coefficient
3 Particle size
Membrane proteins mediated transport of most molecules and all ions across biomembrane
The rate at which a molecule diffuses across a synthetic lipid bilayer depend on its size and solubilityThe smaller the molecule and the less polar it is the more rapidly it diffuses across the bilayer
Overview of membrane transport proteins
1 All transmembrane
proteins
2 Some transport has ATP binding sites
3 Move molecules uphill (向上) against its gradient
Differences1 Transporters=
-uniporters transport a single molecule down its gradient (passive)
-co-transporters couple movement of a molecule down itsgradient with moving a molecule up its gradient (active)
2 Pumps = hydrolyze ATP to move small moleculesions up a concentration gradient or electric potential (active)
3 Channels = transport waterionssmall molecules down their concentration gradients or electric potentials (passive)
The four mechanisms of small molecules and ions are transported cross cellular membranes
Ion rarr force
促進 主動
If transport substance carries a net charge its movement is influenced by both its concentration gradient and the membrane potential the electric potential (voltage) across the membrane
Substance concentration + electric potential = electrochemical gradient Determines the energetically favorable direction of transport a charged molecule across a membrane
Copyright copy 2009 Dr Salme Taagepera PhD All rights reserved
Passive transport driven by
Concentration gradient(affects both uncharged and
charged solutes)
Electrical gradient(affects only charged solutes)
ldquoElectrochemical gradientrdquo
+
被動 Facilitated Diffusion
Passive transport no metabolic energy is needed because the solute is moving down its concentration gradient
bullIn the case of an uncharged solute the concentration of the solute on each side of the membrane dictates the direction of passive transport
Active transport metabolic energy is used to transport a solute from the side of low concentration to the side of high concentration
Two types of transport are defined by whether metabolic energy is expended to move a solute across the membrane
Facilitated Diffusion
Free DiffusionA Non-channel mediated
ndash lipids gasses (O2 CO2) waterB Channel mediated
ndash ions charged molecules
Facilitated diffusionCarrier mediated
ndash glucose amino acids
[ECF]
Extracellular fluid
Facilitated Diffusion
Rate of diffusion is determined by1 concentration gradient 2 amount of carrier protein3 rate of associationdissociation
Unique features for Uniport transport
1 Higher diffusion rate for uniport than passive diffusion
2 Transported molecules never enter membrane and Irrelevant (無關) to the partition coefficient (did not cross membrane)
3 Transport rate reach Vmaxwhen each uniport working at its maximal rate
4 Transport is specific Each uniport transports only a single species of molecules or single or closely related molecules
Several feature distinguish uniport transport from passive diffusion
GLUT glucose transport
Need a carrier protein
Uniport transport of glucose and water
Glucose utilize glucose as a source for ATP production
Water utilize aquaporins to increase the rate of water movement
H2O hybrophilic did across membrane
Families of GLUT proteins(1-12)Highly homologous in sequence and contain 12 membrane-spanning α
-
helicesDifferent isoforms rarr different cell type expression and different functionGLUT2 express in liver cell ( glucose storage)
and szlig cell( glucose uptake) pancreasGLUT4 found in intracellular membrane
increase expression by insulin for remove the glucose from blood to cellGLUT5 tansport fructoseOther isoforms
Glucose transporter (GLUT) Facilitated Diffusion
Mammalian glucose transporters
Name Tissue distribution Proposed function
Glut1 all fetal and adult tissues basal glucose transport
Glut2 hepatocytes pancreatic β-cells transepithelial transport intestine kidney from and to the blood
Glut3 widely distributed basal glucose transportmostly in brain
Glut4 skeletal muscle heart adipocytes insulin-dependent transport
Glut5 intestine lesser amounts few others fructose transport
Glut7 gluconeogenic tissues mediates flux across endoplasmic reticulum
Glut8 preimplantation blastocyst embryonic insulin- dependent transport
Oded Meyuhas
Each GLUT protein contains 12 membrane-spanning alpha helicesDifferentially expressed
ndash EXAMPLE GLUT4 is only expressed in fat and muscle cells
bull Fat and muscle cells respond to insulin by increasing their uptake of glucose thereby removing glucose from the blood
bull In absence of insulin = GLUT4 on intracellular membranesIn presence of insulin = GLUT4 found on cell surface
bull QUESTION Defects in directing GLUT4 to the cell surface can cause what common disease
Type II diabetes high blood glucose
Copyright copy 2009 Dr Salme Taagepera PhD All rights reserved
GLUT1 is responsible for transporting glucose across the blood- brain barrier rarr
GLUT1 provides glucose for the brain
GLUT1 deficiency syndromebull Brain does not obtain enough glucose from the bloodbull Symptoms seizures developmental delay motor disordersbull Treatment ketogenic diet (high fatlow carb diet)
GLUT1 deficiency syndrome
Glucose
Transporter proteins (membrane protein) can be enriched within artificial membrane
Chloroform and methanol (31)
Phospholipid spontaneously form bilayers
Liposome containing a single type of transport protein are very useful in studying functional properties of transport proteins
bull It is a major experimental tool to study the biochemistry of transport protein function in vitro
bull Widely used as a drug delivery system and for gene transfection
Movement of waterOsmosis movement of water across semipermeable
membraneOsmotic pressure hydrostatic pressure uses to stop
the net flow of water
When CB concentration gt CA
Osmotic pressure π=RT( CB -CA )
Hypertonic solution the concentration is higher than cytosolIsotonic solution equal to cytosolHypotonic solution lower and water move to cytosol
Animal cell ace a problem in maintaining their cell volume within a limited range thereby avoiding lysis
Plant cell has cell wall rarr prevent cell shapeTurgor pressure (膨壓) osmotic pressure plasma membrane against water into the cytosol and then into the vacuole
turgor pressure supplies rigidityThe large forces of turgor pressure are resisted by the strength of cellulose microfibrils in the cell wall
Hypertonic external solution concentration of water is low relative to its concentration inside the cell
Water moves out down its concentration gradientand the cell shrinks
Hypotonic external solution concentration of water is highrelative to its concentration inside the cell
Water moves in down its concentration gradientand the cell swells
When the Na+ ndash K+ pump stops Na+ goes into the cell along its concentration gradient
This adds to the solute concentration in the cytosol
Water moves into the cell along its concentration gradient and the cell bursts
Water draw is equal inside and outside
Water cross membrane is very lowerWater cross membrane via specific channel- aquaporins
Aquaporins increase the water permeability of cell membrane
Aquaporin 1 erythrocyte
Aquaporin2 kidney cells resorb water from urine mutation rarr diabetes insipidus rarr large volume urine
Expression of aquaporin by frog oocytes increases their permeability
control
Injection aquaporin mRNA
Egg move to hypotonic environment
尿崩病
Water channel protein ( aquaporin)
Tetrameric protein
6 α-helices for each subunit
2-nm-long water selective gate
028nm gate widthHighly conserved arginine and histidine in the gateH2 O for HO bonding
with cystein
AquaporinsAquaporins are membrane water channels that play critical are membrane water channels that play critical roles in controlling the roles in controlling the waterwater contents of cellscontents of cells
Water Water crosses the crosses the hydrophobic membranehydrophobic membrane either by simple either by simple diffusion or through a facilitative transport mechanism diffusion or through a facilitative transport mechanism mediated by these mediated by these specialized proteinsspecialized proteins
These protein channels are These protein channels are widely distributedwidely distributed in all kingdoms of in all kingdoms of life including bacteria plants and mammalslife including bacteria plants and mammals
Important in Important in osmotic regulationosmotic regulation acting to prevent bursting of acting to prevent bursting of the cells whenever there are changes of the exterior salt the cells whenever there are changes of the exterior salt concentrationconcentration
ATP powered pump1 P- class
2α 2β
subunit can phosphorylationie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-classbull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-classmaintain low pH in plant vacuolesimilar to F-class
4 ABC (ATP-binding cassete) superfamilyseveral hundred different transport protein
Different classes of pumps exhibit characteristic structure and functional properties
Specific Ion binding site
Must phosphorylation
Transport process requires ATP hydrolysis in which the free energy is liberated by breakdown of ATP into ADP and phosphate
V-class H+ ATP ase pump protons across lysosomal and vacuolar membrane
inside
inside
ATP powered pump
1 P- class
2α 2β
subunit
ie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-class
bull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-class
maintain low pH in plant vacuole
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Three main class of membrane protein1ATP- power pump( carrier permease)
couple with energy source for active transportbinding of specific solute to transporter which
undergo conformation change2 Channel protein (ion channel)
formation of hydrophilic pore allow passive movement of small inorganic molecule
3 Transportersuniportsymportantiport
Partition Coefficient
Permeability coefficients (in cmsec) through synthetic lipid bilayers
Product of the concentration difference (in molcm3) and permeability coefficient (in cmsec) gives the flow of solute in moles per second per square centimeter of membrane
油品分配係數
Cell membrane
bullBarrier to the passage of most polar molecule
bullMaintain concentration of solute
Diffusion rate depends on
1 Concentration gradient or electrochemical gradient
2 Hydrophobicity
ie higher partition coefficient
3 Particle size
Membrane proteins mediated transport of most molecules and all ions across biomembrane
The rate at which a molecule diffuses across a synthetic lipid bilayer depend on its size and solubilityThe smaller the molecule and the less polar it is the more rapidly it diffuses across the bilayer
Overview of membrane transport proteins
1 All transmembrane
proteins
2 Some transport has ATP binding sites
3 Move molecules uphill (向上) against its gradient
Differences1 Transporters=
-uniporters transport a single molecule down its gradient (passive)
-co-transporters couple movement of a molecule down itsgradient with moving a molecule up its gradient (active)
2 Pumps = hydrolyze ATP to move small moleculesions up a concentration gradient or electric potential (active)
3 Channels = transport waterionssmall molecules down their concentration gradients or electric potentials (passive)
The four mechanisms of small molecules and ions are transported cross cellular membranes
Ion rarr force
促進 主動
If transport substance carries a net charge its movement is influenced by both its concentration gradient and the membrane potential the electric potential (voltage) across the membrane
Substance concentration + electric potential = electrochemical gradient Determines the energetically favorable direction of transport a charged molecule across a membrane
Copyright copy 2009 Dr Salme Taagepera PhD All rights reserved
Passive transport driven by
Concentration gradient(affects both uncharged and
charged solutes)
Electrical gradient(affects only charged solutes)
ldquoElectrochemical gradientrdquo
+
被動 Facilitated Diffusion
Passive transport no metabolic energy is needed because the solute is moving down its concentration gradient
bullIn the case of an uncharged solute the concentration of the solute on each side of the membrane dictates the direction of passive transport
Active transport metabolic energy is used to transport a solute from the side of low concentration to the side of high concentration
Two types of transport are defined by whether metabolic energy is expended to move a solute across the membrane
Facilitated Diffusion
Free DiffusionA Non-channel mediated
ndash lipids gasses (O2 CO2) waterB Channel mediated
ndash ions charged molecules
Facilitated diffusionCarrier mediated
ndash glucose amino acids
[ECF]
Extracellular fluid
Facilitated Diffusion
Rate of diffusion is determined by1 concentration gradient 2 amount of carrier protein3 rate of associationdissociation
Unique features for Uniport transport
1 Higher diffusion rate for uniport than passive diffusion
2 Transported molecules never enter membrane and Irrelevant (無關) to the partition coefficient (did not cross membrane)
3 Transport rate reach Vmaxwhen each uniport working at its maximal rate
4 Transport is specific Each uniport transports only a single species of molecules or single or closely related molecules
Several feature distinguish uniport transport from passive diffusion
GLUT glucose transport
Need a carrier protein
Uniport transport of glucose and water
Glucose utilize glucose as a source for ATP production
Water utilize aquaporins to increase the rate of water movement
H2O hybrophilic did across membrane
Families of GLUT proteins(1-12)Highly homologous in sequence and contain 12 membrane-spanning α
-
helicesDifferent isoforms rarr different cell type expression and different functionGLUT2 express in liver cell ( glucose storage)
and szlig cell( glucose uptake) pancreasGLUT4 found in intracellular membrane
increase expression by insulin for remove the glucose from blood to cellGLUT5 tansport fructoseOther isoforms
Glucose transporter (GLUT) Facilitated Diffusion
Mammalian glucose transporters
Name Tissue distribution Proposed function
Glut1 all fetal and adult tissues basal glucose transport
Glut2 hepatocytes pancreatic β-cells transepithelial transport intestine kidney from and to the blood
Glut3 widely distributed basal glucose transportmostly in brain
Glut4 skeletal muscle heart adipocytes insulin-dependent transport
Glut5 intestine lesser amounts few others fructose transport
Glut7 gluconeogenic tissues mediates flux across endoplasmic reticulum
Glut8 preimplantation blastocyst embryonic insulin- dependent transport
Oded Meyuhas
Each GLUT protein contains 12 membrane-spanning alpha helicesDifferentially expressed
ndash EXAMPLE GLUT4 is only expressed in fat and muscle cells
bull Fat and muscle cells respond to insulin by increasing their uptake of glucose thereby removing glucose from the blood
bull In absence of insulin = GLUT4 on intracellular membranesIn presence of insulin = GLUT4 found on cell surface
bull QUESTION Defects in directing GLUT4 to the cell surface can cause what common disease
Type II diabetes high blood glucose
Copyright copy 2009 Dr Salme Taagepera PhD All rights reserved
GLUT1 is responsible for transporting glucose across the blood- brain barrier rarr
GLUT1 provides glucose for the brain
GLUT1 deficiency syndromebull Brain does not obtain enough glucose from the bloodbull Symptoms seizures developmental delay motor disordersbull Treatment ketogenic diet (high fatlow carb diet)
GLUT1 deficiency syndrome
Glucose
Transporter proteins (membrane protein) can be enriched within artificial membrane
Chloroform and methanol (31)
Phospholipid spontaneously form bilayers
Liposome containing a single type of transport protein are very useful in studying functional properties of transport proteins
bull It is a major experimental tool to study the biochemistry of transport protein function in vitro
bull Widely used as a drug delivery system and for gene transfection
Movement of waterOsmosis movement of water across semipermeable
membraneOsmotic pressure hydrostatic pressure uses to stop
the net flow of water
When CB concentration gt CA
Osmotic pressure π=RT( CB -CA )
Hypertonic solution the concentration is higher than cytosolIsotonic solution equal to cytosolHypotonic solution lower and water move to cytosol
Animal cell ace a problem in maintaining their cell volume within a limited range thereby avoiding lysis
Plant cell has cell wall rarr prevent cell shapeTurgor pressure (膨壓) osmotic pressure plasma membrane against water into the cytosol and then into the vacuole
turgor pressure supplies rigidityThe large forces of turgor pressure are resisted by the strength of cellulose microfibrils in the cell wall
Hypertonic external solution concentration of water is low relative to its concentration inside the cell
Water moves out down its concentration gradientand the cell shrinks
Hypotonic external solution concentration of water is highrelative to its concentration inside the cell
Water moves in down its concentration gradientand the cell swells
When the Na+ ndash K+ pump stops Na+ goes into the cell along its concentration gradient
This adds to the solute concentration in the cytosol
Water moves into the cell along its concentration gradient and the cell bursts
Water draw is equal inside and outside
Water cross membrane is very lowerWater cross membrane via specific channel- aquaporins
Aquaporins increase the water permeability of cell membrane
Aquaporin 1 erythrocyte
Aquaporin2 kidney cells resorb water from urine mutation rarr diabetes insipidus rarr large volume urine
Expression of aquaporin by frog oocytes increases their permeability
control
Injection aquaporin mRNA
Egg move to hypotonic environment
尿崩病
Water channel protein ( aquaporin)
Tetrameric protein
6 α-helices for each subunit
2-nm-long water selective gate
028nm gate widthHighly conserved arginine and histidine in the gateH2 O for HO bonding
with cystein
AquaporinsAquaporins are membrane water channels that play critical are membrane water channels that play critical roles in controlling the roles in controlling the waterwater contents of cellscontents of cells
Water Water crosses the crosses the hydrophobic membranehydrophobic membrane either by simple either by simple diffusion or through a facilitative transport mechanism diffusion or through a facilitative transport mechanism mediated by these mediated by these specialized proteinsspecialized proteins
These protein channels are These protein channels are widely distributedwidely distributed in all kingdoms of in all kingdoms of life including bacteria plants and mammalslife including bacteria plants and mammals
Important in Important in osmotic regulationosmotic regulation acting to prevent bursting of acting to prevent bursting of the cells whenever there are changes of the exterior salt the cells whenever there are changes of the exterior salt concentrationconcentration
ATP powered pump1 P- class
2α 2β
subunit can phosphorylationie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-classbull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-classmaintain low pH in plant vacuolesimilar to F-class
4 ABC (ATP-binding cassete) superfamilyseveral hundred different transport protein
Different classes of pumps exhibit characteristic structure and functional properties
Specific Ion binding site
Must phosphorylation
Transport process requires ATP hydrolysis in which the free energy is liberated by breakdown of ATP into ADP and phosphate
V-class H+ ATP ase pump protons across lysosomal and vacuolar membrane
inside
inside
ATP powered pump
1 P- class
2α 2β
subunit
ie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-class
bull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-class
maintain low pH in plant vacuole
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Partition Coefficient
Permeability coefficients (in cmsec) through synthetic lipid bilayers
Product of the concentration difference (in molcm3) and permeability coefficient (in cmsec) gives the flow of solute in moles per second per square centimeter of membrane
油品分配係數
Cell membrane
bullBarrier to the passage of most polar molecule
bullMaintain concentration of solute
Diffusion rate depends on
1 Concentration gradient or electrochemical gradient
2 Hydrophobicity
ie higher partition coefficient
3 Particle size
Membrane proteins mediated transport of most molecules and all ions across biomembrane
The rate at which a molecule diffuses across a synthetic lipid bilayer depend on its size and solubilityThe smaller the molecule and the less polar it is the more rapidly it diffuses across the bilayer
Overview of membrane transport proteins
1 All transmembrane
proteins
2 Some transport has ATP binding sites
3 Move molecules uphill (向上) against its gradient
Differences1 Transporters=
-uniporters transport a single molecule down its gradient (passive)
-co-transporters couple movement of a molecule down itsgradient with moving a molecule up its gradient (active)
2 Pumps = hydrolyze ATP to move small moleculesions up a concentration gradient or electric potential (active)
3 Channels = transport waterionssmall molecules down their concentration gradients or electric potentials (passive)
The four mechanisms of small molecules and ions are transported cross cellular membranes
Ion rarr force
促進 主動
If transport substance carries a net charge its movement is influenced by both its concentration gradient and the membrane potential the electric potential (voltage) across the membrane
Substance concentration + electric potential = electrochemical gradient Determines the energetically favorable direction of transport a charged molecule across a membrane
Copyright copy 2009 Dr Salme Taagepera PhD All rights reserved
Passive transport driven by
Concentration gradient(affects both uncharged and
charged solutes)
Electrical gradient(affects only charged solutes)
ldquoElectrochemical gradientrdquo
+
被動 Facilitated Diffusion
Passive transport no metabolic energy is needed because the solute is moving down its concentration gradient
bullIn the case of an uncharged solute the concentration of the solute on each side of the membrane dictates the direction of passive transport
Active transport metabolic energy is used to transport a solute from the side of low concentration to the side of high concentration
Two types of transport are defined by whether metabolic energy is expended to move a solute across the membrane
Facilitated Diffusion
Free DiffusionA Non-channel mediated
ndash lipids gasses (O2 CO2) waterB Channel mediated
ndash ions charged molecules
Facilitated diffusionCarrier mediated
ndash glucose amino acids
[ECF]
Extracellular fluid
Facilitated Diffusion
Rate of diffusion is determined by1 concentration gradient 2 amount of carrier protein3 rate of associationdissociation
Unique features for Uniport transport
1 Higher diffusion rate for uniport than passive diffusion
2 Transported molecules never enter membrane and Irrelevant (無關) to the partition coefficient (did not cross membrane)
3 Transport rate reach Vmaxwhen each uniport working at its maximal rate
4 Transport is specific Each uniport transports only a single species of molecules or single or closely related molecules
Several feature distinguish uniport transport from passive diffusion
GLUT glucose transport
Need a carrier protein
Uniport transport of glucose and water
Glucose utilize glucose as a source for ATP production
Water utilize aquaporins to increase the rate of water movement
H2O hybrophilic did across membrane
Families of GLUT proteins(1-12)Highly homologous in sequence and contain 12 membrane-spanning α
-
helicesDifferent isoforms rarr different cell type expression and different functionGLUT2 express in liver cell ( glucose storage)
and szlig cell( glucose uptake) pancreasGLUT4 found in intracellular membrane
increase expression by insulin for remove the glucose from blood to cellGLUT5 tansport fructoseOther isoforms
Glucose transporter (GLUT) Facilitated Diffusion
Mammalian glucose transporters
Name Tissue distribution Proposed function
Glut1 all fetal and adult tissues basal glucose transport
Glut2 hepatocytes pancreatic β-cells transepithelial transport intestine kidney from and to the blood
Glut3 widely distributed basal glucose transportmostly in brain
Glut4 skeletal muscle heart adipocytes insulin-dependent transport
Glut5 intestine lesser amounts few others fructose transport
Glut7 gluconeogenic tissues mediates flux across endoplasmic reticulum
Glut8 preimplantation blastocyst embryonic insulin- dependent transport
Oded Meyuhas
Each GLUT protein contains 12 membrane-spanning alpha helicesDifferentially expressed
ndash EXAMPLE GLUT4 is only expressed in fat and muscle cells
bull Fat and muscle cells respond to insulin by increasing their uptake of glucose thereby removing glucose from the blood
bull In absence of insulin = GLUT4 on intracellular membranesIn presence of insulin = GLUT4 found on cell surface
bull QUESTION Defects in directing GLUT4 to the cell surface can cause what common disease
Type II diabetes high blood glucose
Copyright copy 2009 Dr Salme Taagepera PhD All rights reserved
GLUT1 is responsible for transporting glucose across the blood- brain barrier rarr
GLUT1 provides glucose for the brain
GLUT1 deficiency syndromebull Brain does not obtain enough glucose from the bloodbull Symptoms seizures developmental delay motor disordersbull Treatment ketogenic diet (high fatlow carb diet)
GLUT1 deficiency syndrome
Glucose
Transporter proteins (membrane protein) can be enriched within artificial membrane
Chloroform and methanol (31)
Phospholipid spontaneously form bilayers
Liposome containing a single type of transport protein are very useful in studying functional properties of transport proteins
bull It is a major experimental tool to study the biochemistry of transport protein function in vitro
bull Widely used as a drug delivery system and for gene transfection
Movement of waterOsmosis movement of water across semipermeable
membraneOsmotic pressure hydrostatic pressure uses to stop
the net flow of water
When CB concentration gt CA
Osmotic pressure π=RT( CB -CA )
Hypertonic solution the concentration is higher than cytosolIsotonic solution equal to cytosolHypotonic solution lower and water move to cytosol
Animal cell ace a problem in maintaining their cell volume within a limited range thereby avoiding lysis
Plant cell has cell wall rarr prevent cell shapeTurgor pressure (膨壓) osmotic pressure plasma membrane against water into the cytosol and then into the vacuole
turgor pressure supplies rigidityThe large forces of turgor pressure are resisted by the strength of cellulose microfibrils in the cell wall
Hypertonic external solution concentration of water is low relative to its concentration inside the cell
Water moves out down its concentration gradientand the cell shrinks
Hypotonic external solution concentration of water is highrelative to its concentration inside the cell
Water moves in down its concentration gradientand the cell swells
When the Na+ ndash K+ pump stops Na+ goes into the cell along its concentration gradient
This adds to the solute concentration in the cytosol
Water moves into the cell along its concentration gradient and the cell bursts
Water draw is equal inside and outside
Water cross membrane is very lowerWater cross membrane via specific channel- aquaporins
Aquaporins increase the water permeability of cell membrane
Aquaporin 1 erythrocyte
Aquaporin2 kidney cells resorb water from urine mutation rarr diabetes insipidus rarr large volume urine
Expression of aquaporin by frog oocytes increases their permeability
control
Injection aquaporin mRNA
Egg move to hypotonic environment
尿崩病
Water channel protein ( aquaporin)
Tetrameric protein
6 α-helices for each subunit
2-nm-long water selective gate
028nm gate widthHighly conserved arginine and histidine in the gateH2 O for HO bonding
with cystein
AquaporinsAquaporins are membrane water channels that play critical are membrane water channels that play critical roles in controlling the roles in controlling the waterwater contents of cellscontents of cells
Water Water crosses the crosses the hydrophobic membranehydrophobic membrane either by simple either by simple diffusion or through a facilitative transport mechanism diffusion or through a facilitative transport mechanism mediated by these mediated by these specialized proteinsspecialized proteins
These protein channels are These protein channels are widely distributedwidely distributed in all kingdoms of in all kingdoms of life including bacteria plants and mammalslife including bacteria plants and mammals
Important in Important in osmotic regulationosmotic regulation acting to prevent bursting of acting to prevent bursting of the cells whenever there are changes of the exterior salt the cells whenever there are changes of the exterior salt concentrationconcentration
ATP powered pump1 P- class
2α 2β
subunit can phosphorylationie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-classbull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-classmaintain low pH in plant vacuolesimilar to F-class
4 ABC (ATP-binding cassete) superfamilyseveral hundred different transport protein
Different classes of pumps exhibit characteristic structure and functional properties
Specific Ion binding site
Must phosphorylation
Transport process requires ATP hydrolysis in which the free energy is liberated by breakdown of ATP into ADP and phosphate
V-class H+ ATP ase pump protons across lysosomal and vacuolar membrane
inside
inside
ATP powered pump
1 P- class
2α 2β
subunit
ie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-class
bull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-class
maintain low pH in plant vacuole
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Cell membrane
bullBarrier to the passage of most polar molecule
bullMaintain concentration of solute
Diffusion rate depends on
1 Concentration gradient or electrochemical gradient
2 Hydrophobicity
ie higher partition coefficient
3 Particle size
Membrane proteins mediated transport of most molecules and all ions across biomembrane
The rate at which a molecule diffuses across a synthetic lipid bilayer depend on its size and solubilityThe smaller the molecule and the less polar it is the more rapidly it diffuses across the bilayer
Overview of membrane transport proteins
1 All transmembrane
proteins
2 Some transport has ATP binding sites
3 Move molecules uphill (向上) against its gradient
Differences1 Transporters=
-uniporters transport a single molecule down its gradient (passive)
-co-transporters couple movement of a molecule down itsgradient with moving a molecule up its gradient (active)
2 Pumps = hydrolyze ATP to move small moleculesions up a concentration gradient or electric potential (active)
3 Channels = transport waterionssmall molecules down their concentration gradients or electric potentials (passive)
The four mechanisms of small molecules and ions are transported cross cellular membranes
Ion rarr force
促進 主動
If transport substance carries a net charge its movement is influenced by both its concentration gradient and the membrane potential the electric potential (voltage) across the membrane
Substance concentration + electric potential = electrochemical gradient Determines the energetically favorable direction of transport a charged molecule across a membrane
Copyright copy 2009 Dr Salme Taagepera PhD All rights reserved
Passive transport driven by
Concentration gradient(affects both uncharged and
charged solutes)
Electrical gradient(affects only charged solutes)
ldquoElectrochemical gradientrdquo
+
被動 Facilitated Diffusion
Passive transport no metabolic energy is needed because the solute is moving down its concentration gradient
bullIn the case of an uncharged solute the concentration of the solute on each side of the membrane dictates the direction of passive transport
Active transport metabolic energy is used to transport a solute from the side of low concentration to the side of high concentration
Two types of transport are defined by whether metabolic energy is expended to move a solute across the membrane
Facilitated Diffusion
Free DiffusionA Non-channel mediated
ndash lipids gasses (O2 CO2) waterB Channel mediated
ndash ions charged molecules
Facilitated diffusionCarrier mediated
ndash glucose amino acids
[ECF]
Extracellular fluid
Facilitated Diffusion
Rate of diffusion is determined by1 concentration gradient 2 amount of carrier protein3 rate of associationdissociation
Unique features for Uniport transport
1 Higher diffusion rate for uniport than passive diffusion
2 Transported molecules never enter membrane and Irrelevant (無關) to the partition coefficient (did not cross membrane)
3 Transport rate reach Vmaxwhen each uniport working at its maximal rate
4 Transport is specific Each uniport transports only a single species of molecules or single or closely related molecules
Several feature distinguish uniport transport from passive diffusion
GLUT glucose transport
Need a carrier protein
Uniport transport of glucose and water
Glucose utilize glucose as a source for ATP production
Water utilize aquaporins to increase the rate of water movement
H2O hybrophilic did across membrane
Families of GLUT proteins(1-12)Highly homologous in sequence and contain 12 membrane-spanning α
-
helicesDifferent isoforms rarr different cell type expression and different functionGLUT2 express in liver cell ( glucose storage)
and szlig cell( glucose uptake) pancreasGLUT4 found in intracellular membrane
increase expression by insulin for remove the glucose from blood to cellGLUT5 tansport fructoseOther isoforms
Glucose transporter (GLUT) Facilitated Diffusion
Mammalian glucose transporters
Name Tissue distribution Proposed function
Glut1 all fetal and adult tissues basal glucose transport
Glut2 hepatocytes pancreatic β-cells transepithelial transport intestine kidney from and to the blood
Glut3 widely distributed basal glucose transportmostly in brain
Glut4 skeletal muscle heart adipocytes insulin-dependent transport
Glut5 intestine lesser amounts few others fructose transport
Glut7 gluconeogenic tissues mediates flux across endoplasmic reticulum
Glut8 preimplantation blastocyst embryonic insulin- dependent transport
Oded Meyuhas
Each GLUT protein contains 12 membrane-spanning alpha helicesDifferentially expressed
ndash EXAMPLE GLUT4 is only expressed in fat and muscle cells
bull Fat and muscle cells respond to insulin by increasing their uptake of glucose thereby removing glucose from the blood
bull In absence of insulin = GLUT4 on intracellular membranesIn presence of insulin = GLUT4 found on cell surface
bull QUESTION Defects in directing GLUT4 to the cell surface can cause what common disease
Type II diabetes high blood glucose
Copyright copy 2009 Dr Salme Taagepera PhD All rights reserved
GLUT1 is responsible for transporting glucose across the blood- brain barrier rarr
GLUT1 provides glucose for the brain
GLUT1 deficiency syndromebull Brain does not obtain enough glucose from the bloodbull Symptoms seizures developmental delay motor disordersbull Treatment ketogenic diet (high fatlow carb diet)
GLUT1 deficiency syndrome
Glucose
Transporter proteins (membrane protein) can be enriched within artificial membrane
Chloroform and methanol (31)
Phospholipid spontaneously form bilayers
Liposome containing a single type of transport protein are very useful in studying functional properties of transport proteins
bull It is a major experimental tool to study the biochemistry of transport protein function in vitro
bull Widely used as a drug delivery system and for gene transfection
Movement of waterOsmosis movement of water across semipermeable
membraneOsmotic pressure hydrostatic pressure uses to stop
the net flow of water
When CB concentration gt CA
Osmotic pressure π=RT( CB -CA )
Hypertonic solution the concentration is higher than cytosolIsotonic solution equal to cytosolHypotonic solution lower and water move to cytosol
Animal cell ace a problem in maintaining their cell volume within a limited range thereby avoiding lysis
Plant cell has cell wall rarr prevent cell shapeTurgor pressure (膨壓) osmotic pressure plasma membrane against water into the cytosol and then into the vacuole
turgor pressure supplies rigidityThe large forces of turgor pressure are resisted by the strength of cellulose microfibrils in the cell wall
Hypertonic external solution concentration of water is low relative to its concentration inside the cell
Water moves out down its concentration gradientand the cell shrinks
Hypotonic external solution concentration of water is highrelative to its concentration inside the cell
Water moves in down its concentration gradientand the cell swells
When the Na+ ndash K+ pump stops Na+ goes into the cell along its concentration gradient
This adds to the solute concentration in the cytosol
Water moves into the cell along its concentration gradient and the cell bursts
Water draw is equal inside and outside
Water cross membrane is very lowerWater cross membrane via specific channel- aquaporins
Aquaporins increase the water permeability of cell membrane
Aquaporin 1 erythrocyte
Aquaporin2 kidney cells resorb water from urine mutation rarr diabetes insipidus rarr large volume urine
Expression of aquaporin by frog oocytes increases their permeability
control
Injection aquaporin mRNA
Egg move to hypotonic environment
尿崩病
Water channel protein ( aquaporin)
Tetrameric protein
6 α-helices for each subunit
2-nm-long water selective gate
028nm gate widthHighly conserved arginine and histidine in the gateH2 O for HO bonding
with cystein
AquaporinsAquaporins are membrane water channels that play critical are membrane water channels that play critical roles in controlling the roles in controlling the waterwater contents of cellscontents of cells
Water Water crosses the crosses the hydrophobic membranehydrophobic membrane either by simple either by simple diffusion or through a facilitative transport mechanism diffusion or through a facilitative transport mechanism mediated by these mediated by these specialized proteinsspecialized proteins
These protein channels are These protein channels are widely distributedwidely distributed in all kingdoms of in all kingdoms of life including bacteria plants and mammalslife including bacteria plants and mammals
Important in Important in osmotic regulationosmotic regulation acting to prevent bursting of acting to prevent bursting of the cells whenever there are changes of the exterior salt the cells whenever there are changes of the exterior salt concentrationconcentration
ATP powered pump1 P- class
2α 2β
subunit can phosphorylationie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-classbull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-classmaintain low pH in plant vacuolesimilar to F-class
4 ABC (ATP-binding cassete) superfamilyseveral hundred different transport protein
Different classes of pumps exhibit characteristic structure and functional properties
Specific Ion binding site
Must phosphorylation
Transport process requires ATP hydrolysis in which the free energy is liberated by breakdown of ATP into ADP and phosphate
V-class H+ ATP ase pump protons across lysosomal and vacuolar membrane
inside
inside
ATP powered pump
1 P- class
2α 2β
subunit
ie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-class
bull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-class
maintain low pH in plant vacuole
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Overview of membrane transport proteins
1 All transmembrane
proteins
2 Some transport has ATP binding sites
3 Move molecules uphill (向上) against its gradient
Differences1 Transporters=
-uniporters transport a single molecule down its gradient (passive)
-co-transporters couple movement of a molecule down itsgradient with moving a molecule up its gradient (active)
2 Pumps = hydrolyze ATP to move small moleculesions up a concentration gradient or electric potential (active)
3 Channels = transport waterionssmall molecules down their concentration gradients or electric potentials (passive)
The four mechanisms of small molecules and ions are transported cross cellular membranes
Ion rarr force
促進 主動
If transport substance carries a net charge its movement is influenced by both its concentration gradient and the membrane potential the electric potential (voltage) across the membrane
Substance concentration + electric potential = electrochemical gradient Determines the energetically favorable direction of transport a charged molecule across a membrane
Copyright copy 2009 Dr Salme Taagepera PhD All rights reserved
Passive transport driven by
Concentration gradient(affects both uncharged and
charged solutes)
Electrical gradient(affects only charged solutes)
ldquoElectrochemical gradientrdquo
+
被動 Facilitated Diffusion
Passive transport no metabolic energy is needed because the solute is moving down its concentration gradient
bullIn the case of an uncharged solute the concentration of the solute on each side of the membrane dictates the direction of passive transport
Active transport metabolic energy is used to transport a solute from the side of low concentration to the side of high concentration
Two types of transport are defined by whether metabolic energy is expended to move a solute across the membrane
Facilitated Diffusion
Free DiffusionA Non-channel mediated
ndash lipids gasses (O2 CO2) waterB Channel mediated
ndash ions charged molecules
Facilitated diffusionCarrier mediated
ndash glucose amino acids
[ECF]
Extracellular fluid
Facilitated Diffusion
Rate of diffusion is determined by1 concentration gradient 2 amount of carrier protein3 rate of associationdissociation
Unique features for Uniport transport
1 Higher diffusion rate for uniport than passive diffusion
2 Transported molecules never enter membrane and Irrelevant (無關) to the partition coefficient (did not cross membrane)
3 Transport rate reach Vmaxwhen each uniport working at its maximal rate
4 Transport is specific Each uniport transports only a single species of molecules or single or closely related molecules
Several feature distinguish uniport transport from passive diffusion
GLUT glucose transport
Need a carrier protein
Uniport transport of glucose and water
Glucose utilize glucose as a source for ATP production
Water utilize aquaporins to increase the rate of water movement
H2O hybrophilic did across membrane
Families of GLUT proteins(1-12)Highly homologous in sequence and contain 12 membrane-spanning α
-
helicesDifferent isoforms rarr different cell type expression and different functionGLUT2 express in liver cell ( glucose storage)
and szlig cell( glucose uptake) pancreasGLUT4 found in intracellular membrane
increase expression by insulin for remove the glucose from blood to cellGLUT5 tansport fructoseOther isoforms
Glucose transporter (GLUT) Facilitated Diffusion
Mammalian glucose transporters
Name Tissue distribution Proposed function
Glut1 all fetal and adult tissues basal glucose transport
Glut2 hepatocytes pancreatic β-cells transepithelial transport intestine kidney from and to the blood
Glut3 widely distributed basal glucose transportmostly in brain
Glut4 skeletal muscle heart adipocytes insulin-dependent transport
Glut5 intestine lesser amounts few others fructose transport
Glut7 gluconeogenic tissues mediates flux across endoplasmic reticulum
Glut8 preimplantation blastocyst embryonic insulin- dependent transport
Oded Meyuhas
Each GLUT protein contains 12 membrane-spanning alpha helicesDifferentially expressed
ndash EXAMPLE GLUT4 is only expressed in fat and muscle cells
bull Fat and muscle cells respond to insulin by increasing their uptake of glucose thereby removing glucose from the blood
bull In absence of insulin = GLUT4 on intracellular membranesIn presence of insulin = GLUT4 found on cell surface
bull QUESTION Defects in directing GLUT4 to the cell surface can cause what common disease
Type II diabetes high blood glucose
Copyright copy 2009 Dr Salme Taagepera PhD All rights reserved
GLUT1 is responsible for transporting glucose across the blood- brain barrier rarr
GLUT1 provides glucose for the brain
GLUT1 deficiency syndromebull Brain does not obtain enough glucose from the bloodbull Symptoms seizures developmental delay motor disordersbull Treatment ketogenic diet (high fatlow carb diet)
GLUT1 deficiency syndrome
Glucose
Transporter proteins (membrane protein) can be enriched within artificial membrane
Chloroform and methanol (31)
Phospholipid spontaneously form bilayers
Liposome containing a single type of transport protein are very useful in studying functional properties of transport proteins
bull It is a major experimental tool to study the biochemistry of transport protein function in vitro
bull Widely used as a drug delivery system and for gene transfection
Movement of waterOsmosis movement of water across semipermeable
membraneOsmotic pressure hydrostatic pressure uses to stop
the net flow of water
When CB concentration gt CA
Osmotic pressure π=RT( CB -CA )
Hypertonic solution the concentration is higher than cytosolIsotonic solution equal to cytosolHypotonic solution lower and water move to cytosol
Animal cell ace a problem in maintaining their cell volume within a limited range thereby avoiding lysis
Plant cell has cell wall rarr prevent cell shapeTurgor pressure (膨壓) osmotic pressure plasma membrane against water into the cytosol and then into the vacuole
turgor pressure supplies rigidityThe large forces of turgor pressure are resisted by the strength of cellulose microfibrils in the cell wall
Hypertonic external solution concentration of water is low relative to its concentration inside the cell
Water moves out down its concentration gradientand the cell shrinks
Hypotonic external solution concentration of water is highrelative to its concentration inside the cell
Water moves in down its concentration gradientand the cell swells
When the Na+ ndash K+ pump stops Na+ goes into the cell along its concentration gradient
This adds to the solute concentration in the cytosol
Water moves into the cell along its concentration gradient and the cell bursts
Water draw is equal inside and outside
Water cross membrane is very lowerWater cross membrane via specific channel- aquaporins
Aquaporins increase the water permeability of cell membrane
Aquaporin 1 erythrocyte
Aquaporin2 kidney cells resorb water from urine mutation rarr diabetes insipidus rarr large volume urine
Expression of aquaporin by frog oocytes increases their permeability
control
Injection aquaporin mRNA
Egg move to hypotonic environment
尿崩病
Water channel protein ( aquaporin)
Tetrameric protein
6 α-helices for each subunit
2-nm-long water selective gate
028nm gate widthHighly conserved arginine and histidine in the gateH2 O for HO bonding
with cystein
AquaporinsAquaporins are membrane water channels that play critical are membrane water channels that play critical roles in controlling the roles in controlling the waterwater contents of cellscontents of cells
Water Water crosses the crosses the hydrophobic membranehydrophobic membrane either by simple either by simple diffusion or through a facilitative transport mechanism diffusion or through a facilitative transport mechanism mediated by these mediated by these specialized proteinsspecialized proteins
These protein channels are These protein channels are widely distributedwidely distributed in all kingdoms of in all kingdoms of life including bacteria plants and mammalslife including bacteria plants and mammals
Important in Important in osmotic regulationosmotic regulation acting to prevent bursting of acting to prevent bursting of the cells whenever there are changes of the exterior salt the cells whenever there are changes of the exterior salt concentrationconcentration
ATP powered pump1 P- class
2α 2β
subunit can phosphorylationie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-classbull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-classmaintain low pH in plant vacuolesimilar to F-class
4 ABC (ATP-binding cassete) superfamilyseveral hundred different transport protein
Different classes of pumps exhibit characteristic structure and functional properties
Specific Ion binding site
Must phosphorylation
Transport process requires ATP hydrolysis in which the free energy is liberated by breakdown of ATP into ADP and phosphate
V-class H+ ATP ase pump protons across lysosomal and vacuolar membrane
inside
inside
ATP powered pump
1 P- class
2α 2β
subunit
ie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-class
bull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-class
maintain low pH in plant vacuole
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Differences1 Transporters=
-uniporters transport a single molecule down its gradient (passive)
-co-transporters couple movement of a molecule down itsgradient with moving a molecule up its gradient (active)
2 Pumps = hydrolyze ATP to move small moleculesions up a concentration gradient or electric potential (active)
3 Channels = transport waterionssmall molecules down their concentration gradients or electric potentials (passive)
The four mechanisms of small molecules and ions are transported cross cellular membranes
Ion rarr force
促進 主動
If transport substance carries a net charge its movement is influenced by both its concentration gradient and the membrane potential the electric potential (voltage) across the membrane
Substance concentration + electric potential = electrochemical gradient Determines the energetically favorable direction of transport a charged molecule across a membrane
Copyright copy 2009 Dr Salme Taagepera PhD All rights reserved
Passive transport driven by
Concentration gradient(affects both uncharged and
charged solutes)
Electrical gradient(affects only charged solutes)
ldquoElectrochemical gradientrdquo
+
被動 Facilitated Diffusion
Passive transport no metabolic energy is needed because the solute is moving down its concentration gradient
bullIn the case of an uncharged solute the concentration of the solute on each side of the membrane dictates the direction of passive transport
Active transport metabolic energy is used to transport a solute from the side of low concentration to the side of high concentration
Two types of transport are defined by whether metabolic energy is expended to move a solute across the membrane
Facilitated Diffusion
Free DiffusionA Non-channel mediated
ndash lipids gasses (O2 CO2) waterB Channel mediated
ndash ions charged molecules
Facilitated diffusionCarrier mediated
ndash glucose amino acids
[ECF]
Extracellular fluid
Facilitated Diffusion
Rate of diffusion is determined by1 concentration gradient 2 amount of carrier protein3 rate of associationdissociation
Unique features for Uniport transport
1 Higher diffusion rate for uniport than passive diffusion
2 Transported molecules never enter membrane and Irrelevant (無關) to the partition coefficient (did not cross membrane)
3 Transport rate reach Vmaxwhen each uniport working at its maximal rate
4 Transport is specific Each uniport transports only a single species of molecules or single or closely related molecules
Several feature distinguish uniport transport from passive diffusion
GLUT glucose transport
Need a carrier protein
Uniport transport of glucose and water
Glucose utilize glucose as a source for ATP production
Water utilize aquaporins to increase the rate of water movement
H2O hybrophilic did across membrane
Families of GLUT proteins(1-12)Highly homologous in sequence and contain 12 membrane-spanning α
-
helicesDifferent isoforms rarr different cell type expression and different functionGLUT2 express in liver cell ( glucose storage)
and szlig cell( glucose uptake) pancreasGLUT4 found in intracellular membrane
increase expression by insulin for remove the glucose from blood to cellGLUT5 tansport fructoseOther isoforms
Glucose transporter (GLUT) Facilitated Diffusion
Mammalian glucose transporters
Name Tissue distribution Proposed function
Glut1 all fetal and adult tissues basal glucose transport
Glut2 hepatocytes pancreatic β-cells transepithelial transport intestine kidney from and to the blood
Glut3 widely distributed basal glucose transportmostly in brain
Glut4 skeletal muscle heart adipocytes insulin-dependent transport
Glut5 intestine lesser amounts few others fructose transport
Glut7 gluconeogenic tissues mediates flux across endoplasmic reticulum
Glut8 preimplantation blastocyst embryonic insulin- dependent transport
Oded Meyuhas
Each GLUT protein contains 12 membrane-spanning alpha helicesDifferentially expressed
ndash EXAMPLE GLUT4 is only expressed in fat and muscle cells
bull Fat and muscle cells respond to insulin by increasing their uptake of glucose thereby removing glucose from the blood
bull In absence of insulin = GLUT4 on intracellular membranesIn presence of insulin = GLUT4 found on cell surface
bull QUESTION Defects in directing GLUT4 to the cell surface can cause what common disease
Type II diabetes high blood glucose
Copyright copy 2009 Dr Salme Taagepera PhD All rights reserved
GLUT1 is responsible for transporting glucose across the blood- brain barrier rarr
GLUT1 provides glucose for the brain
GLUT1 deficiency syndromebull Brain does not obtain enough glucose from the bloodbull Symptoms seizures developmental delay motor disordersbull Treatment ketogenic diet (high fatlow carb diet)
GLUT1 deficiency syndrome
Glucose
Transporter proteins (membrane protein) can be enriched within artificial membrane
Chloroform and methanol (31)
Phospholipid spontaneously form bilayers
Liposome containing a single type of transport protein are very useful in studying functional properties of transport proteins
bull It is a major experimental tool to study the biochemistry of transport protein function in vitro
bull Widely used as a drug delivery system and for gene transfection
Movement of waterOsmosis movement of water across semipermeable
membraneOsmotic pressure hydrostatic pressure uses to stop
the net flow of water
When CB concentration gt CA
Osmotic pressure π=RT( CB -CA )
Hypertonic solution the concentration is higher than cytosolIsotonic solution equal to cytosolHypotonic solution lower and water move to cytosol
Animal cell ace a problem in maintaining their cell volume within a limited range thereby avoiding lysis
Plant cell has cell wall rarr prevent cell shapeTurgor pressure (膨壓) osmotic pressure plasma membrane against water into the cytosol and then into the vacuole
turgor pressure supplies rigidityThe large forces of turgor pressure are resisted by the strength of cellulose microfibrils in the cell wall
Hypertonic external solution concentration of water is low relative to its concentration inside the cell
Water moves out down its concentration gradientand the cell shrinks
Hypotonic external solution concentration of water is highrelative to its concentration inside the cell
Water moves in down its concentration gradientand the cell swells
When the Na+ ndash K+ pump stops Na+ goes into the cell along its concentration gradient
This adds to the solute concentration in the cytosol
Water moves into the cell along its concentration gradient and the cell bursts
Water draw is equal inside and outside
Water cross membrane is very lowerWater cross membrane via specific channel- aquaporins
Aquaporins increase the water permeability of cell membrane
Aquaporin 1 erythrocyte
Aquaporin2 kidney cells resorb water from urine mutation rarr diabetes insipidus rarr large volume urine
Expression of aquaporin by frog oocytes increases their permeability
control
Injection aquaporin mRNA
Egg move to hypotonic environment
尿崩病
Water channel protein ( aquaporin)
Tetrameric protein
6 α-helices for each subunit
2-nm-long water selective gate
028nm gate widthHighly conserved arginine and histidine in the gateH2 O for HO bonding
with cystein
AquaporinsAquaporins are membrane water channels that play critical are membrane water channels that play critical roles in controlling the roles in controlling the waterwater contents of cellscontents of cells
Water Water crosses the crosses the hydrophobic membranehydrophobic membrane either by simple either by simple diffusion or through a facilitative transport mechanism diffusion or through a facilitative transport mechanism mediated by these mediated by these specialized proteinsspecialized proteins
These protein channels are These protein channels are widely distributedwidely distributed in all kingdoms of in all kingdoms of life including bacteria plants and mammalslife including bacteria plants and mammals
Important in Important in osmotic regulationosmotic regulation acting to prevent bursting of acting to prevent bursting of the cells whenever there are changes of the exterior salt the cells whenever there are changes of the exterior salt concentrationconcentration
ATP powered pump1 P- class
2α 2β
subunit can phosphorylationie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-classbull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-classmaintain low pH in plant vacuolesimilar to F-class
4 ABC (ATP-binding cassete) superfamilyseveral hundred different transport protein
Different classes of pumps exhibit characteristic structure and functional properties
Specific Ion binding site
Must phosphorylation
Transport process requires ATP hydrolysis in which the free energy is liberated by breakdown of ATP into ADP and phosphate
V-class H+ ATP ase pump protons across lysosomal and vacuolar membrane
inside
inside
ATP powered pump
1 P- class
2α 2β
subunit
ie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-class
bull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-class
maintain low pH in plant vacuole
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

The four mechanisms of small molecules and ions are transported cross cellular membranes
Ion rarr force
促進 主動
If transport substance carries a net charge its movement is influenced by both its concentration gradient and the membrane potential the electric potential (voltage) across the membrane
Substance concentration + electric potential = electrochemical gradient Determines the energetically favorable direction of transport a charged molecule across a membrane
Copyright copy 2009 Dr Salme Taagepera PhD All rights reserved
Passive transport driven by
Concentration gradient(affects both uncharged and
charged solutes)
Electrical gradient(affects only charged solutes)
ldquoElectrochemical gradientrdquo
+
被動 Facilitated Diffusion
Passive transport no metabolic energy is needed because the solute is moving down its concentration gradient
bullIn the case of an uncharged solute the concentration of the solute on each side of the membrane dictates the direction of passive transport
Active transport metabolic energy is used to transport a solute from the side of low concentration to the side of high concentration
Two types of transport are defined by whether metabolic energy is expended to move a solute across the membrane
Facilitated Diffusion
Free DiffusionA Non-channel mediated
ndash lipids gasses (O2 CO2) waterB Channel mediated
ndash ions charged molecules
Facilitated diffusionCarrier mediated
ndash glucose amino acids
[ECF]
Extracellular fluid
Facilitated Diffusion
Rate of diffusion is determined by1 concentration gradient 2 amount of carrier protein3 rate of associationdissociation
Unique features for Uniport transport
1 Higher diffusion rate for uniport than passive diffusion
2 Transported molecules never enter membrane and Irrelevant (無關) to the partition coefficient (did not cross membrane)
3 Transport rate reach Vmaxwhen each uniport working at its maximal rate
4 Transport is specific Each uniport transports only a single species of molecules or single or closely related molecules
Several feature distinguish uniport transport from passive diffusion
GLUT glucose transport
Need a carrier protein
Uniport transport of glucose and water
Glucose utilize glucose as a source for ATP production
Water utilize aquaporins to increase the rate of water movement
H2O hybrophilic did across membrane
Families of GLUT proteins(1-12)Highly homologous in sequence and contain 12 membrane-spanning α
-
helicesDifferent isoforms rarr different cell type expression and different functionGLUT2 express in liver cell ( glucose storage)
and szlig cell( glucose uptake) pancreasGLUT4 found in intracellular membrane
increase expression by insulin for remove the glucose from blood to cellGLUT5 tansport fructoseOther isoforms
Glucose transporter (GLUT) Facilitated Diffusion
Mammalian glucose transporters
Name Tissue distribution Proposed function
Glut1 all fetal and adult tissues basal glucose transport
Glut2 hepatocytes pancreatic β-cells transepithelial transport intestine kidney from and to the blood
Glut3 widely distributed basal glucose transportmostly in brain
Glut4 skeletal muscle heart adipocytes insulin-dependent transport
Glut5 intestine lesser amounts few others fructose transport
Glut7 gluconeogenic tissues mediates flux across endoplasmic reticulum
Glut8 preimplantation blastocyst embryonic insulin- dependent transport
Oded Meyuhas
Each GLUT protein contains 12 membrane-spanning alpha helicesDifferentially expressed
ndash EXAMPLE GLUT4 is only expressed in fat and muscle cells
bull Fat and muscle cells respond to insulin by increasing their uptake of glucose thereby removing glucose from the blood
bull In absence of insulin = GLUT4 on intracellular membranesIn presence of insulin = GLUT4 found on cell surface
bull QUESTION Defects in directing GLUT4 to the cell surface can cause what common disease
Type II diabetes high blood glucose
Copyright copy 2009 Dr Salme Taagepera PhD All rights reserved
GLUT1 is responsible for transporting glucose across the blood- brain barrier rarr
GLUT1 provides glucose for the brain
GLUT1 deficiency syndromebull Brain does not obtain enough glucose from the bloodbull Symptoms seizures developmental delay motor disordersbull Treatment ketogenic diet (high fatlow carb diet)
GLUT1 deficiency syndrome
Glucose
Transporter proteins (membrane protein) can be enriched within artificial membrane
Chloroform and methanol (31)
Phospholipid spontaneously form bilayers
Liposome containing a single type of transport protein are very useful in studying functional properties of transport proteins
bull It is a major experimental tool to study the biochemistry of transport protein function in vitro
bull Widely used as a drug delivery system and for gene transfection
Movement of waterOsmosis movement of water across semipermeable
membraneOsmotic pressure hydrostatic pressure uses to stop
the net flow of water
When CB concentration gt CA
Osmotic pressure π=RT( CB -CA )
Hypertonic solution the concentration is higher than cytosolIsotonic solution equal to cytosolHypotonic solution lower and water move to cytosol
Animal cell ace a problem in maintaining their cell volume within a limited range thereby avoiding lysis
Plant cell has cell wall rarr prevent cell shapeTurgor pressure (膨壓) osmotic pressure plasma membrane against water into the cytosol and then into the vacuole
turgor pressure supplies rigidityThe large forces of turgor pressure are resisted by the strength of cellulose microfibrils in the cell wall
Hypertonic external solution concentration of water is low relative to its concentration inside the cell
Water moves out down its concentration gradientand the cell shrinks
Hypotonic external solution concentration of water is highrelative to its concentration inside the cell
Water moves in down its concentration gradientand the cell swells
When the Na+ ndash K+ pump stops Na+ goes into the cell along its concentration gradient
This adds to the solute concentration in the cytosol
Water moves into the cell along its concentration gradient and the cell bursts
Water draw is equal inside and outside
Water cross membrane is very lowerWater cross membrane via specific channel- aquaporins
Aquaporins increase the water permeability of cell membrane
Aquaporin 1 erythrocyte
Aquaporin2 kidney cells resorb water from urine mutation rarr diabetes insipidus rarr large volume urine
Expression of aquaporin by frog oocytes increases their permeability
control
Injection aquaporin mRNA
Egg move to hypotonic environment
尿崩病
Water channel protein ( aquaporin)
Tetrameric protein
6 α-helices for each subunit
2-nm-long water selective gate
028nm gate widthHighly conserved arginine and histidine in the gateH2 O for HO bonding
with cystein
AquaporinsAquaporins are membrane water channels that play critical are membrane water channels that play critical roles in controlling the roles in controlling the waterwater contents of cellscontents of cells
Water Water crosses the crosses the hydrophobic membranehydrophobic membrane either by simple either by simple diffusion or through a facilitative transport mechanism diffusion or through a facilitative transport mechanism mediated by these mediated by these specialized proteinsspecialized proteins
These protein channels are These protein channels are widely distributedwidely distributed in all kingdoms of in all kingdoms of life including bacteria plants and mammalslife including bacteria plants and mammals
Important in Important in osmotic regulationosmotic regulation acting to prevent bursting of acting to prevent bursting of the cells whenever there are changes of the exterior salt the cells whenever there are changes of the exterior salt concentrationconcentration
ATP powered pump1 P- class
2α 2β
subunit can phosphorylationie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-classbull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-classmaintain low pH in plant vacuolesimilar to F-class
4 ABC (ATP-binding cassete) superfamilyseveral hundred different transport protein
Different classes of pumps exhibit characteristic structure and functional properties
Specific Ion binding site
Must phosphorylation
Transport process requires ATP hydrolysis in which the free energy is liberated by breakdown of ATP into ADP and phosphate
V-class H+ ATP ase pump protons across lysosomal and vacuolar membrane
inside
inside
ATP powered pump
1 P- class
2α 2β
subunit
ie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-class
bull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-class
maintain low pH in plant vacuole
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

If transport substance carries a net charge its movement is influenced by both its concentration gradient and the membrane potential the electric potential (voltage) across the membrane
Substance concentration + electric potential = electrochemical gradient Determines the energetically favorable direction of transport a charged molecule across a membrane
Copyright copy 2009 Dr Salme Taagepera PhD All rights reserved
Passive transport driven by
Concentration gradient(affects both uncharged and
charged solutes)
Electrical gradient(affects only charged solutes)
ldquoElectrochemical gradientrdquo
+
被動 Facilitated Diffusion
Passive transport no metabolic energy is needed because the solute is moving down its concentration gradient
bullIn the case of an uncharged solute the concentration of the solute on each side of the membrane dictates the direction of passive transport
Active transport metabolic energy is used to transport a solute from the side of low concentration to the side of high concentration
Two types of transport are defined by whether metabolic energy is expended to move a solute across the membrane
Facilitated Diffusion
Free DiffusionA Non-channel mediated
ndash lipids gasses (O2 CO2) waterB Channel mediated
ndash ions charged molecules
Facilitated diffusionCarrier mediated
ndash glucose amino acids
[ECF]
Extracellular fluid
Facilitated Diffusion
Rate of diffusion is determined by1 concentration gradient 2 amount of carrier protein3 rate of associationdissociation
Unique features for Uniport transport
1 Higher diffusion rate for uniport than passive diffusion
2 Transported molecules never enter membrane and Irrelevant (無關) to the partition coefficient (did not cross membrane)
3 Transport rate reach Vmaxwhen each uniport working at its maximal rate
4 Transport is specific Each uniport transports only a single species of molecules or single or closely related molecules
Several feature distinguish uniport transport from passive diffusion
GLUT glucose transport
Need a carrier protein
Uniport transport of glucose and water
Glucose utilize glucose as a source for ATP production
Water utilize aquaporins to increase the rate of water movement
H2O hybrophilic did across membrane
Families of GLUT proteins(1-12)Highly homologous in sequence and contain 12 membrane-spanning α
-
helicesDifferent isoforms rarr different cell type expression and different functionGLUT2 express in liver cell ( glucose storage)
and szlig cell( glucose uptake) pancreasGLUT4 found in intracellular membrane
increase expression by insulin for remove the glucose from blood to cellGLUT5 tansport fructoseOther isoforms
Glucose transporter (GLUT) Facilitated Diffusion
Mammalian glucose transporters
Name Tissue distribution Proposed function
Glut1 all fetal and adult tissues basal glucose transport
Glut2 hepatocytes pancreatic β-cells transepithelial transport intestine kidney from and to the blood
Glut3 widely distributed basal glucose transportmostly in brain
Glut4 skeletal muscle heart adipocytes insulin-dependent transport
Glut5 intestine lesser amounts few others fructose transport
Glut7 gluconeogenic tissues mediates flux across endoplasmic reticulum
Glut8 preimplantation blastocyst embryonic insulin- dependent transport
Oded Meyuhas
Each GLUT protein contains 12 membrane-spanning alpha helicesDifferentially expressed
ndash EXAMPLE GLUT4 is only expressed in fat and muscle cells
bull Fat and muscle cells respond to insulin by increasing their uptake of glucose thereby removing glucose from the blood
bull In absence of insulin = GLUT4 on intracellular membranesIn presence of insulin = GLUT4 found on cell surface
bull QUESTION Defects in directing GLUT4 to the cell surface can cause what common disease
Type II diabetes high blood glucose
Copyright copy 2009 Dr Salme Taagepera PhD All rights reserved
GLUT1 is responsible for transporting glucose across the blood- brain barrier rarr
GLUT1 provides glucose for the brain
GLUT1 deficiency syndromebull Brain does not obtain enough glucose from the bloodbull Symptoms seizures developmental delay motor disordersbull Treatment ketogenic diet (high fatlow carb diet)
GLUT1 deficiency syndrome
Glucose
Transporter proteins (membrane protein) can be enriched within artificial membrane
Chloroform and methanol (31)
Phospholipid spontaneously form bilayers
Liposome containing a single type of transport protein are very useful in studying functional properties of transport proteins
bull It is a major experimental tool to study the biochemistry of transport protein function in vitro
bull Widely used as a drug delivery system and for gene transfection
Movement of waterOsmosis movement of water across semipermeable
membraneOsmotic pressure hydrostatic pressure uses to stop
the net flow of water
When CB concentration gt CA
Osmotic pressure π=RT( CB -CA )
Hypertonic solution the concentration is higher than cytosolIsotonic solution equal to cytosolHypotonic solution lower and water move to cytosol
Animal cell ace a problem in maintaining their cell volume within a limited range thereby avoiding lysis
Plant cell has cell wall rarr prevent cell shapeTurgor pressure (膨壓) osmotic pressure plasma membrane against water into the cytosol and then into the vacuole
turgor pressure supplies rigidityThe large forces of turgor pressure are resisted by the strength of cellulose microfibrils in the cell wall
Hypertonic external solution concentration of water is low relative to its concentration inside the cell
Water moves out down its concentration gradientand the cell shrinks
Hypotonic external solution concentration of water is highrelative to its concentration inside the cell
Water moves in down its concentration gradientand the cell swells
When the Na+ ndash K+ pump stops Na+ goes into the cell along its concentration gradient
This adds to the solute concentration in the cytosol
Water moves into the cell along its concentration gradient and the cell bursts
Water draw is equal inside and outside
Water cross membrane is very lowerWater cross membrane via specific channel- aquaporins
Aquaporins increase the water permeability of cell membrane
Aquaporin 1 erythrocyte
Aquaporin2 kidney cells resorb water from urine mutation rarr diabetes insipidus rarr large volume urine
Expression of aquaporin by frog oocytes increases their permeability
control
Injection aquaporin mRNA
Egg move to hypotonic environment
尿崩病
Water channel protein ( aquaporin)
Tetrameric protein
6 α-helices for each subunit
2-nm-long water selective gate
028nm gate widthHighly conserved arginine and histidine in the gateH2 O for HO bonding
with cystein
AquaporinsAquaporins are membrane water channels that play critical are membrane water channels that play critical roles in controlling the roles in controlling the waterwater contents of cellscontents of cells
Water Water crosses the crosses the hydrophobic membranehydrophobic membrane either by simple either by simple diffusion or through a facilitative transport mechanism diffusion or through a facilitative transport mechanism mediated by these mediated by these specialized proteinsspecialized proteins
These protein channels are These protein channels are widely distributedwidely distributed in all kingdoms of in all kingdoms of life including bacteria plants and mammalslife including bacteria plants and mammals
Important in Important in osmotic regulationosmotic regulation acting to prevent bursting of acting to prevent bursting of the cells whenever there are changes of the exterior salt the cells whenever there are changes of the exterior salt concentrationconcentration
ATP powered pump1 P- class
2α 2β
subunit can phosphorylationie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-classbull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-classmaintain low pH in plant vacuolesimilar to F-class
4 ABC (ATP-binding cassete) superfamilyseveral hundred different transport protein
Different classes of pumps exhibit characteristic structure and functional properties
Specific Ion binding site
Must phosphorylation
Transport process requires ATP hydrolysis in which the free energy is liberated by breakdown of ATP into ADP and phosphate
V-class H+ ATP ase pump protons across lysosomal and vacuolar membrane
inside
inside
ATP powered pump
1 P- class
2α 2β
subunit
ie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-class
bull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-class
maintain low pH in plant vacuole
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Copyright copy 2009 Dr Salme Taagepera PhD All rights reserved
Passive transport driven by
Concentration gradient(affects both uncharged and
charged solutes)
Electrical gradient(affects only charged solutes)
ldquoElectrochemical gradientrdquo
+
被動 Facilitated Diffusion
Passive transport no metabolic energy is needed because the solute is moving down its concentration gradient
bullIn the case of an uncharged solute the concentration of the solute on each side of the membrane dictates the direction of passive transport
Active transport metabolic energy is used to transport a solute from the side of low concentration to the side of high concentration
Two types of transport are defined by whether metabolic energy is expended to move a solute across the membrane
Facilitated Diffusion
Free DiffusionA Non-channel mediated
ndash lipids gasses (O2 CO2) waterB Channel mediated
ndash ions charged molecules
Facilitated diffusionCarrier mediated
ndash glucose amino acids
[ECF]
Extracellular fluid
Facilitated Diffusion
Rate of diffusion is determined by1 concentration gradient 2 amount of carrier protein3 rate of associationdissociation
Unique features for Uniport transport
1 Higher diffusion rate for uniport than passive diffusion
2 Transported molecules never enter membrane and Irrelevant (無關) to the partition coefficient (did not cross membrane)
3 Transport rate reach Vmaxwhen each uniport working at its maximal rate
4 Transport is specific Each uniport transports only a single species of molecules or single or closely related molecules
Several feature distinguish uniport transport from passive diffusion
GLUT glucose transport
Need a carrier protein
Uniport transport of glucose and water
Glucose utilize glucose as a source for ATP production
Water utilize aquaporins to increase the rate of water movement
H2O hybrophilic did across membrane
Families of GLUT proteins(1-12)Highly homologous in sequence and contain 12 membrane-spanning α
-
helicesDifferent isoforms rarr different cell type expression and different functionGLUT2 express in liver cell ( glucose storage)
and szlig cell( glucose uptake) pancreasGLUT4 found in intracellular membrane
increase expression by insulin for remove the glucose from blood to cellGLUT5 tansport fructoseOther isoforms
Glucose transporter (GLUT) Facilitated Diffusion
Mammalian glucose transporters
Name Tissue distribution Proposed function
Glut1 all fetal and adult tissues basal glucose transport
Glut2 hepatocytes pancreatic β-cells transepithelial transport intestine kidney from and to the blood
Glut3 widely distributed basal glucose transportmostly in brain
Glut4 skeletal muscle heart adipocytes insulin-dependent transport
Glut5 intestine lesser amounts few others fructose transport
Glut7 gluconeogenic tissues mediates flux across endoplasmic reticulum
Glut8 preimplantation blastocyst embryonic insulin- dependent transport
Oded Meyuhas
Each GLUT protein contains 12 membrane-spanning alpha helicesDifferentially expressed
ndash EXAMPLE GLUT4 is only expressed in fat and muscle cells
bull Fat and muscle cells respond to insulin by increasing their uptake of glucose thereby removing glucose from the blood
bull In absence of insulin = GLUT4 on intracellular membranesIn presence of insulin = GLUT4 found on cell surface
bull QUESTION Defects in directing GLUT4 to the cell surface can cause what common disease
Type II diabetes high blood glucose
Copyright copy 2009 Dr Salme Taagepera PhD All rights reserved
GLUT1 is responsible for transporting glucose across the blood- brain barrier rarr
GLUT1 provides glucose for the brain
GLUT1 deficiency syndromebull Brain does not obtain enough glucose from the bloodbull Symptoms seizures developmental delay motor disordersbull Treatment ketogenic diet (high fatlow carb diet)
GLUT1 deficiency syndrome
Glucose
Transporter proteins (membrane protein) can be enriched within artificial membrane
Chloroform and methanol (31)
Phospholipid spontaneously form bilayers
Liposome containing a single type of transport protein are very useful in studying functional properties of transport proteins
bull It is a major experimental tool to study the biochemistry of transport protein function in vitro
bull Widely used as a drug delivery system and for gene transfection
Movement of waterOsmosis movement of water across semipermeable
membraneOsmotic pressure hydrostatic pressure uses to stop
the net flow of water
When CB concentration gt CA
Osmotic pressure π=RT( CB -CA )
Hypertonic solution the concentration is higher than cytosolIsotonic solution equal to cytosolHypotonic solution lower and water move to cytosol
Animal cell ace a problem in maintaining their cell volume within a limited range thereby avoiding lysis
Plant cell has cell wall rarr prevent cell shapeTurgor pressure (膨壓) osmotic pressure plasma membrane against water into the cytosol and then into the vacuole
turgor pressure supplies rigidityThe large forces of turgor pressure are resisted by the strength of cellulose microfibrils in the cell wall
Hypertonic external solution concentration of water is low relative to its concentration inside the cell
Water moves out down its concentration gradientand the cell shrinks
Hypotonic external solution concentration of water is highrelative to its concentration inside the cell
Water moves in down its concentration gradientand the cell swells
When the Na+ ndash K+ pump stops Na+ goes into the cell along its concentration gradient
This adds to the solute concentration in the cytosol
Water moves into the cell along its concentration gradient and the cell bursts
Water draw is equal inside and outside
Water cross membrane is very lowerWater cross membrane via specific channel- aquaporins
Aquaporins increase the water permeability of cell membrane
Aquaporin 1 erythrocyte
Aquaporin2 kidney cells resorb water from urine mutation rarr diabetes insipidus rarr large volume urine
Expression of aquaporin by frog oocytes increases their permeability
control
Injection aquaporin mRNA
Egg move to hypotonic environment
尿崩病
Water channel protein ( aquaporin)
Tetrameric protein
6 α-helices for each subunit
2-nm-long water selective gate
028nm gate widthHighly conserved arginine and histidine in the gateH2 O for HO bonding
with cystein
AquaporinsAquaporins are membrane water channels that play critical are membrane water channels that play critical roles in controlling the roles in controlling the waterwater contents of cellscontents of cells
Water Water crosses the crosses the hydrophobic membranehydrophobic membrane either by simple either by simple diffusion or through a facilitative transport mechanism diffusion or through a facilitative transport mechanism mediated by these mediated by these specialized proteinsspecialized proteins
These protein channels are These protein channels are widely distributedwidely distributed in all kingdoms of in all kingdoms of life including bacteria plants and mammalslife including bacteria plants and mammals
Important in Important in osmotic regulationosmotic regulation acting to prevent bursting of acting to prevent bursting of the cells whenever there are changes of the exterior salt the cells whenever there are changes of the exterior salt concentrationconcentration
ATP powered pump1 P- class
2α 2β
subunit can phosphorylationie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-classbull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-classmaintain low pH in plant vacuolesimilar to F-class
4 ABC (ATP-binding cassete) superfamilyseveral hundred different transport protein
Different classes of pumps exhibit characteristic structure and functional properties
Specific Ion binding site
Must phosphorylation
Transport process requires ATP hydrolysis in which the free energy is liberated by breakdown of ATP into ADP and phosphate
V-class H+ ATP ase pump protons across lysosomal and vacuolar membrane
inside
inside
ATP powered pump
1 P- class
2α 2β
subunit
ie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-class
bull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-class
maintain low pH in plant vacuole
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Passive transport no metabolic energy is needed because the solute is moving down its concentration gradient
bullIn the case of an uncharged solute the concentration of the solute on each side of the membrane dictates the direction of passive transport
Active transport metabolic energy is used to transport a solute from the side of low concentration to the side of high concentration
Two types of transport are defined by whether metabolic energy is expended to move a solute across the membrane
Facilitated Diffusion
Free DiffusionA Non-channel mediated
ndash lipids gasses (O2 CO2) waterB Channel mediated
ndash ions charged molecules
Facilitated diffusionCarrier mediated
ndash glucose amino acids
[ECF]
Extracellular fluid
Facilitated Diffusion
Rate of diffusion is determined by1 concentration gradient 2 amount of carrier protein3 rate of associationdissociation
Unique features for Uniport transport
1 Higher diffusion rate for uniport than passive diffusion
2 Transported molecules never enter membrane and Irrelevant (無關) to the partition coefficient (did not cross membrane)
3 Transport rate reach Vmaxwhen each uniport working at its maximal rate
4 Transport is specific Each uniport transports only a single species of molecules or single or closely related molecules
Several feature distinguish uniport transport from passive diffusion
GLUT glucose transport
Need a carrier protein
Uniport transport of glucose and water
Glucose utilize glucose as a source for ATP production
Water utilize aquaporins to increase the rate of water movement
H2O hybrophilic did across membrane
Families of GLUT proteins(1-12)Highly homologous in sequence and contain 12 membrane-spanning α
-
helicesDifferent isoforms rarr different cell type expression and different functionGLUT2 express in liver cell ( glucose storage)
and szlig cell( glucose uptake) pancreasGLUT4 found in intracellular membrane
increase expression by insulin for remove the glucose from blood to cellGLUT5 tansport fructoseOther isoforms
Glucose transporter (GLUT) Facilitated Diffusion
Mammalian glucose transporters
Name Tissue distribution Proposed function
Glut1 all fetal and adult tissues basal glucose transport
Glut2 hepatocytes pancreatic β-cells transepithelial transport intestine kidney from and to the blood
Glut3 widely distributed basal glucose transportmostly in brain
Glut4 skeletal muscle heart adipocytes insulin-dependent transport
Glut5 intestine lesser amounts few others fructose transport
Glut7 gluconeogenic tissues mediates flux across endoplasmic reticulum
Glut8 preimplantation blastocyst embryonic insulin- dependent transport
Oded Meyuhas
Each GLUT protein contains 12 membrane-spanning alpha helicesDifferentially expressed
ndash EXAMPLE GLUT4 is only expressed in fat and muscle cells
bull Fat and muscle cells respond to insulin by increasing their uptake of glucose thereby removing glucose from the blood
bull In absence of insulin = GLUT4 on intracellular membranesIn presence of insulin = GLUT4 found on cell surface
bull QUESTION Defects in directing GLUT4 to the cell surface can cause what common disease
Type II diabetes high blood glucose
Copyright copy 2009 Dr Salme Taagepera PhD All rights reserved
GLUT1 is responsible for transporting glucose across the blood- brain barrier rarr
GLUT1 provides glucose for the brain
GLUT1 deficiency syndromebull Brain does not obtain enough glucose from the bloodbull Symptoms seizures developmental delay motor disordersbull Treatment ketogenic diet (high fatlow carb diet)
GLUT1 deficiency syndrome
Glucose
Transporter proteins (membrane protein) can be enriched within artificial membrane
Chloroform and methanol (31)
Phospholipid spontaneously form bilayers
Liposome containing a single type of transport protein are very useful in studying functional properties of transport proteins
bull It is a major experimental tool to study the biochemistry of transport protein function in vitro
bull Widely used as a drug delivery system and for gene transfection
Movement of waterOsmosis movement of water across semipermeable
membraneOsmotic pressure hydrostatic pressure uses to stop
the net flow of water
When CB concentration gt CA
Osmotic pressure π=RT( CB -CA )
Hypertonic solution the concentration is higher than cytosolIsotonic solution equal to cytosolHypotonic solution lower and water move to cytosol
Animal cell ace a problem in maintaining their cell volume within a limited range thereby avoiding lysis
Plant cell has cell wall rarr prevent cell shapeTurgor pressure (膨壓) osmotic pressure plasma membrane against water into the cytosol and then into the vacuole
turgor pressure supplies rigidityThe large forces of turgor pressure are resisted by the strength of cellulose microfibrils in the cell wall
Hypertonic external solution concentration of water is low relative to its concentration inside the cell
Water moves out down its concentration gradientand the cell shrinks
Hypotonic external solution concentration of water is highrelative to its concentration inside the cell
Water moves in down its concentration gradientand the cell swells
When the Na+ ndash K+ pump stops Na+ goes into the cell along its concentration gradient
This adds to the solute concentration in the cytosol
Water moves into the cell along its concentration gradient and the cell bursts
Water draw is equal inside and outside
Water cross membrane is very lowerWater cross membrane via specific channel- aquaporins
Aquaporins increase the water permeability of cell membrane
Aquaporin 1 erythrocyte
Aquaporin2 kidney cells resorb water from urine mutation rarr diabetes insipidus rarr large volume urine
Expression of aquaporin by frog oocytes increases their permeability
control
Injection aquaporin mRNA
Egg move to hypotonic environment
尿崩病
Water channel protein ( aquaporin)
Tetrameric protein
6 α-helices for each subunit
2-nm-long water selective gate
028nm gate widthHighly conserved arginine and histidine in the gateH2 O for HO bonding
with cystein
AquaporinsAquaporins are membrane water channels that play critical are membrane water channels that play critical roles in controlling the roles in controlling the waterwater contents of cellscontents of cells
Water Water crosses the crosses the hydrophobic membranehydrophobic membrane either by simple either by simple diffusion or through a facilitative transport mechanism diffusion or through a facilitative transport mechanism mediated by these mediated by these specialized proteinsspecialized proteins
These protein channels are These protein channels are widely distributedwidely distributed in all kingdoms of in all kingdoms of life including bacteria plants and mammalslife including bacteria plants and mammals
Important in Important in osmotic regulationosmotic regulation acting to prevent bursting of acting to prevent bursting of the cells whenever there are changes of the exterior salt the cells whenever there are changes of the exterior salt concentrationconcentration
ATP powered pump1 P- class
2α 2β
subunit can phosphorylationie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-classbull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-classmaintain low pH in plant vacuolesimilar to F-class
4 ABC (ATP-binding cassete) superfamilyseveral hundred different transport protein
Different classes of pumps exhibit characteristic structure and functional properties
Specific Ion binding site
Must phosphorylation
Transport process requires ATP hydrolysis in which the free energy is liberated by breakdown of ATP into ADP and phosphate
V-class H+ ATP ase pump protons across lysosomal and vacuolar membrane
inside
inside
ATP powered pump
1 P- class
2α 2β
subunit
ie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-class
bull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-class
maintain low pH in plant vacuole
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Facilitated Diffusion
Free DiffusionA Non-channel mediated
ndash lipids gasses (O2 CO2) waterB Channel mediated
ndash ions charged molecules
Facilitated diffusionCarrier mediated
ndash glucose amino acids
[ECF]
Extracellular fluid
Facilitated Diffusion
Rate of diffusion is determined by1 concentration gradient 2 amount of carrier protein3 rate of associationdissociation
Unique features for Uniport transport
1 Higher diffusion rate for uniport than passive diffusion
2 Transported molecules never enter membrane and Irrelevant (無關) to the partition coefficient (did not cross membrane)
3 Transport rate reach Vmaxwhen each uniport working at its maximal rate
4 Transport is specific Each uniport transports only a single species of molecules or single or closely related molecules
Several feature distinguish uniport transport from passive diffusion
GLUT glucose transport
Need a carrier protein
Uniport transport of glucose and water
Glucose utilize glucose as a source for ATP production
Water utilize aquaporins to increase the rate of water movement
H2O hybrophilic did across membrane
Families of GLUT proteins(1-12)Highly homologous in sequence and contain 12 membrane-spanning α
-
helicesDifferent isoforms rarr different cell type expression and different functionGLUT2 express in liver cell ( glucose storage)
and szlig cell( glucose uptake) pancreasGLUT4 found in intracellular membrane
increase expression by insulin for remove the glucose from blood to cellGLUT5 tansport fructoseOther isoforms
Glucose transporter (GLUT) Facilitated Diffusion
Mammalian glucose transporters
Name Tissue distribution Proposed function
Glut1 all fetal and adult tissues basal glucose transport
Glut2 hepatocytes pancreatic β-cells transepithelial transport intestine kidney from and to the blood
Glut3 widely distributed basal glucose transportmostly in brain
Glut4 skeletal muscle heart adipocytes insulin-dependent transport
Glut5 intestine lesser amounts few others fructose transport
Glut7 gluconeogenic tissues mediates flux across endoplasmic reticulum
Glut8 preimplantation blastocyst embryonic insulin- dependent transport
Oded Meyuhas
Each GLUT protein contains 12 membrane-spanning alpha helicesDifferentially expressed
ndash EXAMPLE GLUT4 is only expressed in fat and muscle cells
bull Fat and muscle cells respond to insulin by increasing their uptake of glucose thereby removing glucose from the blood
bull In absence of insulin = GLUT4 on intracellular membranesIn presence of insulin = GLUT4 found on cell surface
bull QUESTION Defects in directing GLUT4 to the cell surface can cause what common disease
Type II diabetes high blood glucose
Copyright copy 2009 Dr Salme Taagepera PhD All rights reserved
GLUT1 is responsible for transporting glucose across the blood- brain barrier rarr
GLUT1 provides glucose for the brain
GLUT1 deficiency syndromebull Brain does not obtain enough glucose from the bloodbull Symptoms seizures developmental delay motor disordersbull Treatment ketogenic diet (high fatlow carb diet)
GLUT1 deficiency syndrome
Glucose
Transporter proteins (membrane protein) can be enriched within artificial membrane
Chloroform and methanol (31)
Phospholipid spontaneously form bilayers
Liposome containing a single type of transport protein are very useful in studying functional properties of transport proteins
bull It is a major experimental tool to study the biochemistry of transport protein function in vitro
bull Widely used as a drug delivery system and for gene transfection
Movement of waterOsmosis movement of water across semipermeable
membraneOsmotic pressure hydrostatic pressure uses to stop
the net flow of water
When CB concentration gt CA
Osmotic pressure π=RT( CB -CA )
Hypertonic solution the concentration is higher than cytosolIsotonic solution equal to cytosolHypotonic solution lower and water move to cytosol
Animal cell ace a problem in maintaining their cell volume within a limited range thereby avoiding lysis
Plant cell has cell wall rarr prevent cell shapeTurgor pressure (膨壓) osmotic pressure plasma membrane against water into the cytosol and then into the vacuole
turgor pressure supplies rigidityThe large forces of turgor pressure are resisted by the strength of cellulose microfibrils in the cell wall
Hypertonic external solution concentration of water is low relative to its concentration inside the cell
Water moves out down its concentration gradientand the cell shrinks
Hypotonic external solution concentration of water is highrelative to its concentration inside the cell
Water moves in down its concentration gradientand the cell swells
When the Na+ ndash K+ pump stops Na+ goes into the cell along its concentration gradient
This adds to the solute concentration in the cytosol
Water moves into the cell along its concentration gradient and the cell bursts
Water draw is equal inside and outside
Water cross membrane is very lowerWater cross membrane via specific channel- aquaporins
Aquaporins increase the water permeability of cell membrane
Aquaporin 1 erythrocyte
Aquaporin2 kidney cells resorb water from urine mutation rarr diabetes insipidus rarr large volume urine
Expression of aquaporin by frog oocytes increases their permeability
control
Injection aquaporin mRNA
Egg move to hypotonic environment
尿崩病
Water channel protein ( aquaporin)
Tetrameric protein
6 α-helices for each subunit
2-nm-long water selective gate
028nm gate widthHighly conserved arginine and histidine in the gateH2 O for HO bonding
with cystein
AquaporinsAquaporins are membrane water channels that play critical are membrane water channels that play critical roles in controlling the roles in controlling the waterwater contents of cellscontents of cells
Water Water crosses the crosses the hydrophobic membranehydrophobic membrane either by simple either by simple diffusion or through a facilitative transport mechanism diffusion or through a facilitative transport mechanism mediated by these mediated by these specialized proteinsspecialized proteins
These protein channels are These protein channels are widely distributedwidely distributed in all kingdoms of in all kingdoms of life including bacteria plants and mammalslife including bacteria plants and mammals
Important in Important in osmotic regulationosmotic regulation acting to prevent bursting of acting to prevent bursting of the cells whenever there are changes of the exterior salt the cells whenever there are changes of the exterior salt concentrationconcentration
ATP powered pump1 P- class
2α 2β
subunit can phosphorylationie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-classbull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-classmaintain low pH in plant vacuolesimilar to F-class
4 ABC (ATP-binding cassete) superfamilyseveral hundred different transport protein
Different classes of pumps exhibit characteristic structure and functional properties
Specific Ion binding site
Must phosphorylation
Transport process requires ATP hydrolysis in which the free energy is liberated by breakdown of ATP into ADP and phosphate
V-class H+ ATP ase pump protons across lysosomal and vacuolar membrane
inside
inside
ATP powered pump
1 P- class
2α 2β
subunit
ie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-class
bull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-class
maintain low pH in plant vacuole
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Free DiffusionA Non-channel mediated
ndash lipids gasses (O2 CO2) waterB Channel mediated
ndash ions charged molecules
Facilitated diffusionCarrier mediated
ndash glucose amino acids
[ECF]
Extracellular fluid
Facilitated Diffusion
Rate of diffusion is determined by1 concentration gradient 2 amount of carrier protein3 rate of associationdissociation
Unique features for Uniport transport
1 Higher diffusion rate for uniport than passive diffusion
2 Transported molecules never enter membrane and Irrelevant (無關) to the partition coefficient (did not cross membrane)
3 Transport rate reach Vmaxwhen each uniport working at its maximal rate
4 Transport is specific Each uniport transports only a single species of molecules or single or closely related molecules
Several feature distinguish uniport transport from passive diffusion
GLUT glucose transport
Need a carrier protein
Uniport transport of glucose and water
Glucose utilize glucose as a source for ATP production
Water utilize aquaporins to increase the rate of water movement
H2O hybrophilic did across membrane
Families of GLUT proteins(1-12)Highly homologous in sequence and contain 12 membrane-spanning α
-
helicesDifferent isoforms rarr different cell type expression and different functionGLUT2 express in liver cell ( glucose storage)
and szlig cell( glucose uptake) pancreasGLUT4 found in intracellular membrane
increase expression by insulin for remove the glucose from blood to cellGLUT5 tansport fructoseOther isoforms
Glucose transporter (GLUT) Facilitated Diffusion
Mammalian glucose transporters
Name Tissue distribution Proposed function
Glut1 all fetal and adult tissues basal glucose transport
Glut2 hepatocytes pancreatic β-cells transepithelial transport intestine kidney from and to the blood
Glut3 widely distributed basal glucose transportmostly in brain
Glut4 skeletal muscle heart adipocytes insulin-dependent transport
Glut5 intestine lesser amounts few others fructose transport
Glut7 gluconeogenic tissues mediates flux across endoplasmic reticulum
Glut8 preimplantation blastocyst embryonic insulin- dependent transport
Oded Meyuhas
Each GLUT protein contains 12 membrane-spanning alpha helicesDifferentially expressed
ndash EXAMPLE GLUT4 is only expressed in fat and muscle cells
bull Fat and muscle cells respond to insulin by increasing their uptake of glucose thereby removing glucose from the blood
bull In absence of insulin = GLUT4 on intracellular membranesIn presence of insulin = GLUT4 found on cell surface
bull QUESTION Defects in directing GLUT4 to the cell surface can cause what common disease
Type II diabetes high blood glucose
Copyright copy 2009 Dr Salme Taagepera PhD All rights reserved
GLUT1 is responsible for transporting glucose across the blood- brain barrier rarr
GLUT1 provides glucose for the brain
GLUT1 deficiency syndromebull Brain does not obtain enough glucose from the bloodbull Symptoms seizures developmental delay motor disordersbull Treatment ketogenic diet (high fatlow carb diet)
GLUT1 deficiency syndrome
Glucose
Transporter proteins (membrane protein) can be enriched within artificial membrane
Chloroform and methanol (31)
Phospholipid spontaneously form bilayers
Liposome containing a single type of transport protein are very useful in studying functional properties of transport proteins
bull It is a major experimental tool to study the biochemistry of transport protein function in vitro
bull Widely used as a drug delivery system and for gene transfection
Movement of waterOsmosis movement of water across semipermeable
membraneOsmotic pressure hydrostatic pressure uses to stop
the net flow of water
When CB concentration gt CA
Osmotic pressure π=RT( CB -CA )
Hypertonic solution the concentration is higher than cytosolIsotonic solution equal to cytosolHypotonic solution lower and water move to cytosol
Animal cell ace a problem in maintaining their cell volume within a limited range thereby avoiding lysis
Plant cell has cell wall rarr prevent cell shapeTurgor pressure (膨壓) osmotic pressure plasma membrane against water into the cytosol and then into the vacuole
turgor pressure supplies rigidityThe large forces of turgor pressure are resisted by the strength of cellulose microfibrils in the cell wall
Hypertonic external solution concentration of water is low relative to its concentration inside the cell
Water moves out down its concentration gradientand the cell shrinks
Hypotonic external solution concentration of water is highrelative to its concentration inside the cell
Water moves in down its concentration gradientand the cell swells
When the Na+ ndash K+ pump stops Na+ goes into the cell along its concentration gradient
This adds to the solute concentration in the cytosol
Water moves into the cell along its concentration gradient and the cell bursts
Water draw is equal inside and outside
Water cross membrane is very lowerWater cross membrane via specific channel- aquaporins
Aquaporins increase the water permeability of cell membrane
Aquaporin 1 erythrocyte
Aquaporin2 kidney cells resorb water from urine mutation rarr diabetes insipidus rarr large volume urine
Expression of aquaporin by frog oocytes increases their permeability
control
Injection aquaporin mRNA
Egg move to hypotonic environment
尿崩病
Water channel protein ( aquaporin)
Tetrameric protein
6 α-helices for each subunit
2-nm-long water selective gate
028nm gate widthHighly conserved arginine and histidine in the gateH2 O for HO bonding
with cystein
AquaporinsAquaporins are membrane water channels that play critical are membrane water channels that play critical roles in controlling the roles in controlling the waterwater contents of cellscontents of cells
Water Water crosses the crosses the hydrophobic membranehydrophobic membrane either by simple either by simple diffusion or through a facilitative transport mechanism diffusion or through a facilitative transport mechanism mediated by these mediated by these specialized proteinsspecialized proteins
These protein channels are These protein channels are widely distributedwidely distributed in all kingdoms of in all kingdoms of life including bacteria plants and mammalslife including bacteria plants and mammals
Important in Important in osmotic regulationosmotic regulation acting to prevent bursting of acting to prevent bursting of the cells whenever there are changes of the exterior salt the cells whenever there are changes of the exterior salt concentrationconcentration
ATP powered pump1 P- class
2α 2β
subunit can phosphorylationie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-classbull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-classmaintain low pH in plant vacuolesimilar to F-class
4 ABC (ATP-binding cassete) superfamilyseveral hundred different transport protein
Different classes of pumps exhibit characteristic structure and functional properties
Specific Ion binding site
Must phosphorylation
Transport process requires ATP hydrolysis in which the free energy is liberated by breakdown of ATP into ADP and phosphate
V-class H+ ATP ase pump protons across lysosomal and vacuolar membrane
inside
inside
ATP powered pump
1 P- class
2α 2β
subunit
ie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-class
bull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-class
maintain low pH in plant vacuole
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Facilitated Diffusion
Rate of diffusion is determined by1 concentration gradient 2 amount of carrier protein3 rate of associationdissociation
Unique features for Uniport transport
1 Higher diffusion rate for uniport than passive diffusion
2 Transported molecules never enter membrane and Irrelevant (無關) to the partition coefficient (did not cross membrane)
3 Transport rate reach Vmaxwhen each uniport working at its maximal rate
4 Transport is specific Each uniport transports only a single species of molecules or single or closely related molecules
Several feature distinguish uniport transport from passive diffusion
GLUT glucose transport
Need a carrier protein
Uniport transport of glucose and water
Glucose utilize glucose as a source for ATP production
Water utilize aquaporins to increase the rate of water movement
H2O hybrophilic did across membrane
Families of GLUT proteins(1-12)Highly homologous in sequence and contain 12 membrane-spanning α
-
helicesDifferent isoforms rarr different cell type expression and different functionGLUT2 express in liver cell ( glucose storage)
and szlig cell( glucose uptake) pancreasGLUT4 found in intracellular membrane
increase expression by insulin for remove the glucose from blood to cellGLUT5 tansport fructoseOther isoforms
Glucose transporter (GLUT) Facilitated Diffusion
Mammalian glucose transporters
Name Tissue distribution Proposed function
Glut1 all fetal and adult tissues basal glucose transport
Glut2 hepatocytes pancreatic β-cells transepithelial transport intestine kidney from and to the blood
Glut3 widely distributed basal glucose transportmostly in brain
Glut4 skeletal muscle heart adipocytes insulin-dependent transport
Glut5 intestine lesser amounts few others fructose transport
Glut7 gluconeogenic tissues mediates flux across endoplasmic reticulum
Glut8 preimplantation blastocyst embryonic insulin- dependent transport
Oded Meyuhas
Each GLUT protein contains 12 membrane-spanning alpha helicesDifferentially expressed
ndash EXAMPLE GLUT4 is only expressed in fat and muscle cells
bull Fat and muscle cells respond to insulin by increasing their uptake of glucose thereby removing glucose from the blood
bull In absence of insulin = GLUT4 on intracellular membranesIn presence of insulin = GLUT4 found on cell surface
bull QUESTION Defects in directing GLUT4 to the cell surface can cause what common disease
Type II diabetes high blood glucose
Copyright copy 2009 Dr Salme Taagepera PhD All rights reserved
GLUT1 is responsible for transporting glucose across the blood- brain barrier rarr
GLUT1 provides glucose for the brain
GLUT1 deficiency syndromebull Brain does not obtain enough glucose from the bloodbull Symptoms seizures developmental delay motor disordersbull Treatment ketogenic diet (high fatlow carb diet)
GLUT1 deficiency syndrome
Glucose
Transporter proteins (membrane protein) can be enriched within artificial membrane
Chloroform and methanol (31)
Phospholipid spontaneously form bilayers
Liposome containing a single type of transport protein are very useful in studying functional properties of transport proteins
bull It is a major experimental tool to study the biochemistry of transport protein function in vitro
bull Widely used as a drug delivery system and for gene transfection
Movement of waterOsmosis movement of water across semipermeable
membraneOsmotic pressure hydrostatic pressure uses to stop
the net flow of water
When CB concentration gt CA
Osmotic pressure π=RT( CB -CA )
Hypertonic solution the concentration is higher than cytosolIsotonic solution equal to cytosolHypotonic solution lower and water move to cytosol
Animal cell ace a problem in maintaining their cell volume within a limited range thereby avoiding lysis
Plant cell has cell wall rarr prevent cell shapeTurgor pressure (膨壓) osmotic pressure plasma membrane against water into the cytosol and then into the vacuole
turgor pressure supplies rigidityThe large forces of turgor pressure are resisted by the strength of cellulose microfibrils in the cell wall
Hypertonic external solution concentration of water is low relative to its concentration inside the cell
Water moves out down its concentration gradientand the cell shrinks
Hypotonic external solution concentration of water is highrelative to its concentration inside the cell
Water moves in down its concentration gradientand the cell swells
When the Na+ ndash K+ pump stops Na+ goes into the cell along its concentration gradient
This adds to the solute concentration in the cytosol
Water moves into the cell along its concentration gradient and the cell bursts
Water draw is equal inside and outside
Water cross membrane is very lowerWater cross membrane via specific channel- aquaporins
Aquaporins increase the water permeability of cell membrane
Aquaporin 1 erythrocyte
Aquaporin2 kidney cells resorb water from urine mutation rarr diabetes insipidus rarr large volume urine
Expression of aquaporin by frog oocytes increases their permeability
control
Injection aquaporin mRNA
Egg move to hypotonic environment
尿崩病
Water channel protein ( aquaporin)
Tetrameric protein
6 α-helices for each subunit
2-nm-long water selective gate
028nm gate widthHighly conserved arginine and histidine in the gateH2 O for HO bonding
with cystein
AquaporinsAquaporins are membrane water channels that play critical are membrane water channels that play critical roles in controlling the roles in controlling the waterwater contents of cellscontents of cells
Water Water crosses the crosses the hydrophobic membranehydrophobic membrane either by simple either by simple diffusion or through a facilitative transport mechanism diffusion or through a facilitative transport mechanism mediated by these mediated by these specialized proteinsspecialized proteins
These protein channels are These protein channels are widely distributedwidely distributed in all kingdoms of in all kingdoms of life including bacteria plants and mammalslife including bacteria plants and mammals
Important in Important in osmotic regulationosmotic regulation acting to prevent bursting of acting to prevent bursting of the cells whenever there are changes of the exterior salt the cells whenever there are changes of the exterior salt concentrationconcentration
ATP powered pump1 P- class
2α 2β
subunit can phosphorylationie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-classbull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-classmaintain low pH in plant vacuolesimilar to F-class
4 ABC (ATP-binding cassete) superfamilyseveral hundred different transport protein
Different classes of pumps exhibit characteristic structure and functional properties
Specific Ion binding site
Must phosphorylation
Transport process requires ATP hydrolysis in which the free energy is liberated by breakdown of ATP into ADP and phosphate
V-class H+ ATP ase pump protons across lysosomal and vacuolar membrane
inside
inside
ATP powered pump
1 P- class
2α 2β
subunit
ie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-class
bull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-class
maintain low pH in plant vacuole
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Unique features for Uniport transport
1 Higher diffusion rate for uniport than passive diffusion
2 Transported molecules never enter membrane and Irrelevant (無關) to the partition coefficient (did not cross membrane)
3 Transport rate reach Vmaxwhen each uniport working at its maximal rate
4 Transport is specific Each uniport transports only a single species of molecules or single or closely related molecules
Several feature distinguish uniport transport from passive diffusion
GLUT glucose transport
Need a carrier protein
Uniport transport of glucose and water
Glucose utilize glucose as a source for ATP production
Water utilize aquaporins to increase the rate of water movement
H2O hybrophilic did across membrane
Families of GLUT proteins(1-12)Highly homologous in sequence and contain 12 membrane-spanning α
-
helicesDifferent isoforms rarr different cell type expression and different functionGLUT2 express in liver cell ( glucose storage)
and szlig cell( glucose uptake) pancreasGLUT4 found in intracellular membrane
increase expression by insulin for remove the glucose from blood to cellGLUT5 tansport fructoseOther isoforms
Glucose transporter (GLUT) Facilitated Diffusion
Mammalian glucose transporters
Name Tissue distribution Proposed function
Glut1 all fetal and adult tissues basal glucose transport
Glut2 hepatocytes pancreatic β-cells transepithelial transport intestine kidney from and to the blood
Glut3 widely distributed basal glucose transportmostly in brain
Glut4 skeletal muscle heart adipocytes insulin-dependent transport
Glut5 intestine lesser amounts few others fructose transport
Glut7 gluconeogenic tissues mediates flux across endoplasmic reticulum
Glut8 preimplantation blastocyst embryonic insulin- dependent transport
Oded Meyuhas
Each GLUT protein contains 12 membrane-spanning alpha helicesDifferentially expressed
ndash EXAMPLE GLUT4 is only expressed in fat and muscle cells
bull Fat and muscle cells respond to insulin by increasing their uptake of glucose thereby removing glucose from the blood
bull In absence of insulin = GLUT4 on intracellular membranesIn presence of insulin = GLUT4 found on cell surface
bull QUESTION Defects in directing GLUT4 to the cell surface can cause what common disease
Type II diabetes high blood glucose
Copyright copy 2009 Dr Salme Taagepera PhD All rights reserved
GLUT1 is responsible for transporting glucose across the blood- brain barrier rarr
GLUT1 provides glucose for the brain
GLUT1 deficiency syndromebull Brain does not obtain enough glucose from the bloodbull Symptoms seizures developmental delay motor disordersbull Treatment ketogenic diet (high fatlow carb diet)
GLUT1 deficiency syndrome
Glucose
Transporter proteins (membrane protein) can be enriched within artificial membrane
Chloroform and methanol (31)
Phospholipid spontaneously form bilayers
Liposome containing a single type of transport protein are very useful in studying functional properties of transport proteins
bull It is a major experimental tool to study the biochemistry of transport protein function in vitro
bull Widely used as a drug delivery system and for gene transfection
Movement of waterOsmosis movement of water across semipermeable
membraneOsmotic pressure hydrostatic pressure uses to stop
the net flow of water
When CB concentration gt CA
Osmotic pressure π=RT( CB -CA )
Hypertonic solution the concentration is higher than cytosolIsotonic solution equal to cytosolHypotonic solution lower and water move to cytosol
Animal cell ace a problem in maintaining their cell volume within a limited range thereby avoiding lysis
Plant cell has cell wall rarr prevent cell shapeTurgor pressure (膨壓) osmotic pressure plasma membrane against water into the cytosol and then into the vacuole
turgor pressure supplies rigidityThe large forces of turgor pressure are resisted by the strength of cellulose microfibrils in the cell wall
Hypertonic external solution concentration of water is low relative to its concentration inside the cell
Water moves out down its concentration gradientand the cell shrinks
Hypotonic external solution concentration of water is highrelative to its concentration inside the cell
Water moves in down its concentration gradientand the cell swells
When the Na+ ndash K+ pump stops Na+ goes into the cell along its concentration gradient
This adds to the solute concentration in the cytosol
Water moves into the cell along its concentration gradient and the cell bursts
Water draw is equal inside and outside
Water cross membrane is very lowerWater cross membrane via specific channel- aquaporins
Aquaporins increase the water permeability of cell membrane
Aquaporin 1 erythrocyte
Aquaporin2 kidney cells resorb water from urine mutation rarr diabetes insipidus rarr large volume urine
Expression of aquaporin by frog oocytes increases their permeability
control
Injection aquaporin mRNA
Egg move to hypotonic environment
尿崩病
Water channel protein ( aquaporin)
Tetrameric protein
6 α-helices for each subunit
2-nm-long water selective gate
028nm gate widthHighly conserved arginine and histidine in the gateH2 O for HO bonding
with cystein
AquaporinsAquaporins are membrane water channels that play critical are membrane water channels that play critical roles in controlling the roles in controlling the waterwater contents of cellscontents of cells
Water Water crosses the crosses the hydrophobic membranehydrophobic membrane either by simple either by simple diffusion or through a facilitative transport mechanism diffusion or through a facilitative transport mechanism mediated by these mediated by these specialized proteinsspecialized proteins
These protein channels are These protein channels are widely distributedwidely distributed in all kingdoms of in all kingdoms of life including bacteria plants and mammalslife including bacteria plants and mammals
Important in Important in osmotic regulationosmotic regulation acting to prevent bursting of acting to prevent bursting of the cells whenever there are changes of the exterior salt the cells whenever there are changes of the exterior salt concentrationconcentration
ATP powered pump1 P- class
2α 2β
subunit can phosphorylationie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-classbull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-classmaintain low pH in plant vacuolesimilar to F-class
4 ABC (ATP-binding cassete) superfamilyseveral hundred different transport protein
Different classes of pumps exhibit characteristic structure and functional properties
Specific Ion binding site
Must phosphorylation
Transport process requires ATP hydrolysis in which the free energy is liberated by breakdown of ATP into ADP and phosphate
V-class H+ ATP ase pump protons across lysosomal and vacuolar membrane
inside
inside
ATP powered pump
1 P- class
2α 2β
subunit
ie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-class
bull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-class
maintain low pH in plant vacuole
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Uniport transport of glucose and water
Glucose utilize glucose as a source for ATP production
Water utilize aquaporins to increase the rate of water movement
H2O hybrophilic did across membrane
Families of GLUT proteins(1-12)Highly homologous in sequence and contain 12 membrane-spanning α
-
helicesDifferent isoforms rarr different cell type expression and different functionGLUT2 express in liver cell ( glucose storage)
and szlig cell( glucose uptake) pancreasGLUT4 found in intracellular membrane
increase expression by insulin for remove the glucose from blood to cellGLUT5 tansport fructoseOther isoforms
Glucose transporter (GLUT) Facilitated Diffusion
Mammalian glucose transporters
Name Tissue distribution Proposed function
Glut1 all fetal and adult tissues basal glucose transport
Glut2 hepatocytes pancreatic β-cells transepithelial transport intestine kidney from and to the blood
Glut3 widely distributed basal glucose transportmostly in brain
Glut4 skeletal muscle heart adipocytes insulin-dependent transport
Glut5 intestine lesser amounts few others fructose transport
Glut7 gluconeogenic tissues mediates flux across endoplasmic reticulum
Glut8 preimplantation blastocyst embryonic insulin- dependent transport
Oded Meyuhas
Each GLUT protein contains 12 membrane-spanning alpha helicesDifferentially expressed
ndash EXAMPLE GLUT4 is only expressed in fat and muscle cells
bull Fat and muscle cells respond to insulin by increasing their uptake of glucose thereby removing glucose from the blood
bull In absence of insulin = GLUT4 on intracellular membranesIn presence of insulin = GLUT4 found on cell surface
bull QUESTION Defects in directing GLUT4 to the cell surface can cause what common disease
Type II diabetes high blood glucose
Copyright copy 2009 Dr Salme Taagepera PhD All rights reserved
GLUT1 is responsible for transporting glucose across the blood- brain barrier rarr
GLUT1 provides glucose for the brain
GLUT1 deficiency syndromebull Brain does not obtain enough glucose from the bloodbull Symptoms seizures developmental delay motor disordersbull Treatment ketogenic diet (high fatlow carb diet)
GLUT1 deficiency syndrome
Glucose
Transporter proteins (membrane protein) can be enriched within artificial membrane
Chloroform and methanol (31)
Phospholipid spontaneously form bilayers
Liposome containing a single type of transport protein are very useful in studying functional properties of transport proteins
bull It is a major experimental tool to study the biochemistry of transport protein function in vitro
bull Widely used as a drug delivery system and for gene transfection
Movement of waterOsmosis movement of water across semipermeable
membraneOsmotic pressure hydrostatic pressure uses to stop
the net flow of water
When CB concentration gt CA
Osmotic pressure π=RT( CB -CA )
Hypertonic solution the concentration is higher than cytosolIsotonic solution equal to cytosolHypotonic solution lower and water move to cytosol
Animal cell ace a problem in maintaining their cell volume within a limited range thereby avoiding lysis
Plant cell has cell wall rarr prevent cell shapeTurgor pressure (膨壓) osmotic pressure plasma membrane against water into the cytosol and then into the vacuole
turgor pressure supplies rigidityThe large forces of turgor pressure are resisted by the strength of cellulose microfibrils in the cell wall
Hypertonic external solution concentration of water is low relative to its concentration inside the cell
Water moves out down its concentration gradientand the cell shrinks
Hypotonic external solution concentration of water is highrelative to its concentration inside the cell
Water moves in down its concentration gradientand the cell swells
When the Na+ ndash K+ pump stops Na+ goes into the cell along its concentration gradient
This adds to the solute concentration in the cytosol
Water moves into the cell along its concentration gradient and the cell bursts
Water draw is equal inside and outside
Water cross membrane is very lowerWater cross membrane via specific channel- aquaporins
Aquaporins increase the water permeability of cell membrane
Aquaporin 1 erythrocyte
Aquaporin2 kidney cells resorb water from urine mutation rarr diabetes insipidus rarr large volume urine
Expression of aquaporin by frog oocytes increases their permeability
control
Injection aquaporin mRNA
Egg move to hypotonic environment
尿崩病
Water channel protein ( aquaporin)
Tetrameric protein
6 α-helices for each subunit
2-nm-long water selective gate
028nm gate widthHighly conserved arginine and histidine in the gateH2 O for HO bonding
with cystein
AquaporinsAquaporins are membrane water channels that play critical are membrane water channels that play critical roles in controlling the roles in controlling the waterwater contents of cellscontents of cells
Water Water crosses the crosses the hydrophobic membranehydrophobic membrane either by simple either by simple diffusion or through a facilitative transport mechanism diffusion or through a facilitative transport mechanism mediated by these mediated by these specialized proteinsspecialized proteins
These protein channels are These protein channels are widely distributedwidely distributed in all kingdoms of in all kingdoms of life including bacteria plants and mammalslife including bacteria plants and mammals
Important in Important in osmotic regulationosmotic regulation acting to prevent bursting of acting to prevent bursting of the cells whenever there are changes of the exterior salt the cells whenever there are changes of the exterior salt concentrationconcentration
ATP powered pump1 P- class
2α 2β
subunit can phosphorylationie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-classbull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-classmaintain low pH in plant vacuolesimilar to F-class
4 ABC (ATP-binding cassete) superfamilyseveral hundred different transport protein
Different classes of pumps exhibit characteristic structure and functional properties
Specific Ion binding site
Must phosphorylation
Transport process requires ATP hydrolysis in which the free energy is liberated by breakdown of ATP into ADP and phosphate
V-class H+ ATP ase pump protons across lysosomal and vacuolar membrane
inside
inside
ATP powered pump
1 P- class
2α 2β
subunit
ie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-class
bull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-class
maintain low pH in plant vacuole
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Families of GLUT proteins(1-12)Highly homologous in sequence and contain 12 membrane-spanning α
-
helicesDifferent isoforms rarr different cell type expression and different functionGLUT2 express in liver cell ( glucose storage)
and szlig cell( glucose uptake) pancreasGLUT4 found in intracellular membrane
increase expression by insulin for remove the glucose from blood to cellGLUT5 tansport fructoseOther isoforms
Glucose transporter (GLUT) Facilitated Diffusion
Mammalian glucose transporters
Name Tissue distribution Proposed function
Glut1 all fetal and adult tissues basal glucose transport
Glut2 hepatocytes pancreatic β-cells transepithelial transport intestine kidney from and to the blood
Glut3 widely distributed basal glucose transportmostly in brain
Glut4 skeletal muscle heart adipocytes insulin-dependent transport
Glut5 intestine lesser amounts few others fructose transport
Glut7 gluconeogenic tissues mediates flux across endoplasmic reticulum
Glut8 preimplantation blastocyst embryonic insulin- dependent transport
Oded Meyuhas
Each GLUT protein contains 12 membrane-spanning alpha helicesDifferentially expressed
ndash EXAMPLE GLUT4 is only expressed in fat and muscle cells
bull Fat and muscle cells respond to insulin by increasing their uptake of glucose thereby removing glucose from the blood
bull In absence of insulin = GLUT4 on intracellular membranesIn presence of insulin = GLUT4 found on cell surface
bull QUESTION Defects in directing GLUT4 to the cell surface can cause what common disease
Type II diabetes high blood glucose
Copyright copy 2009 Dr Salme Taagepera PhD All rights reserved
GLUT1 is responsible for transporting glucose across the blood- brain barrier rarr
GLUT1 provides glucose for the brain
GLUT1 deficiency syndromebull Brain does not obtain enough glucose from the bloodbull Symptoms seizures developmental delay motor disordersbull Treatment ketogenic diet (high fatlow carb diet)
GLUT1 deficiency syndrome
Glucose
Transporter proteins (membrane protein) can be enriched within artificial membrane
Chloroform and methanol (31)
Phospholipid spontaneously form bilayers
Liposome containing a single type of transport protein are very useful in studying functional properties of transport proteins
bull It is a major experimental tool to study the biochemistry of transport protein function in vitro
bull Widely used as a drug delivery system and for gene transfection
Movement of waterOsmosis movement of water across semipermeable
membraneOsmotic pressure hydrostatic pressure uses to stop
the net flow of water
When CB concentration gt CA
Osmotic pressure π=RT( CB -CA )
Hypertonic solution the concentration is higher than cytosolIsotonic solution equal to cytosolHypotonic solution lower and water move to cytosol
Animal cell ace a problem in maintaining their cell volume within a limited range thereby avoiding lysis
Plant cell has cell wall rarr prevent cell shapeTurgor pressure (膨壓) osmotic pressure plasma membrane against water into the cytosol and then into the vacuole
turgor pressure supplies rigidityThe large forces of turgor pressure are resisted by the strength of cellulose microfibrils in the cell wall
Hypertonic external solution concentration of water is low relative to its concentration inside the cell
Water moves out down its concentration gradientand the cell shrinks
Hypotonic external solution concentration of water is highrelative to its concentration inside the cell
Water moves in down its concentration gradientand the cell swells
When the Na+ ndash K+ pump stops Na+ goes into the cell along its concentration gradient
This adds to the solute concentration in the cytosol
Water moves into the cell along its concentration gradient and the cell bursts
Water draw is equal inside and outside
Water cross membrane is very lowerWater cross membrane via specific channel- aquaporins
Aquaporins increase the water permeability of cell membrane
Aquaporin 1 erythrocyte
Aquaporin2 kidney cells resorb water from urine mutation rarr diabetes insipidus rarr large volume urine
Expression of aquaporin by frog oocytes increases their permeability
control
Injection aquaporin mRNA
Egg move to hypotonic environment
尿崩病
Water channel protein ( aquaporin)
Tetrameric protein
6 α-helices for each subunit
2-nm-long water selective gate
028nm gate widthHighly conserved arginine and histidine in the gateH2 O for HO bonding
with cystein
AquaporinsAquaporins are membrane water channels that play critical are membrane water channels that play critical roles in controlling the roles in controlling the waterwater contents of cellscontents of cells
Water Water crosses the crosses the hydrophobic membranehydrophobic membrane either by simple either by simple diffusion or through a facilitative transport mechanism diffusion or through a facilitative transport mechanism mediated by these mediated by these specialized proteinsspecialized proteins
These protein channels are These protein channels are widely distributedwidely distributed in all kingdoms of in all kingdoms of life including bacteria plants and mammalslife including bacteria plants and mammals
Important in Important in osmotic regulationosmotic regulation acting to prevent bursting of acting to prevent bursting of the cells whenever there are changes of the exterior salt the cells whenever there are changes of the exterior salt concentrationconcentration
ATP powered pump1 P- class
2α 2β
subunit can phosphorylationie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-classbull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-classmaintain low pH in plant vacuolesimilar to F-class
4 ABC (ATP-binding cassete) superfamilyseveral hundred different transport protein
Different classes of pumps exhibit characteristic structure and functional properties
Specific Ion binding site
Must phosphorylation
Transport process requires ATP hydrolysis in which the free energy is liberated by breakdown of ATP into ADP and phosphate
V-class H+ ATP ase pump protons across lysosomal and vacuolar membrane
inside
inside
ATP powered pump
1 P- class
2α 2β
subunit
ie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-class
bull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-class
maintain low pH in plant vacuole
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Mammalian glucose transporters
Name Tissue distribution Proposed function
Glut1 all fetal and adult tissues basal glucose transport
Glut2 hepatocytes pancreatic β-cells transepithelial transport intestine kidney from and to the blood
Glut3 widely distributed basal glucose transportmostly in brain
Glut4 skeletal muscle heart adipocytes insulin-dependent transport
Glut5 intestine lesser amounts few others fructose transport
Glut7 gluconeogenic tissues mediates flux across endoplasmic reticulum
Glut8 preimplantation blastocyst embryonic insulin- dependent transport
Oded Meyuhas
Each GLUT protein contains 12 membrane-spanning alpha helicesDifferentially expressed
ndash EXAMPLE GLUT4 is only expressed in fat and muscle cells
bull Fat and muscle cells respond to insulin by increasing their uptake of glucose thereby removing glucose from the blood
bull In absence of insulin = GLUT4 on intracellular membranesIn presence of insulin = GLUT4 found on cell surface
bull QUESTION Defects in directing GLUT4 to the cell surface can cause what common disease
Type II diabetes high blood glucose
Copyright copy 2009 Dr Salme Taagepera PhD All rights reserved
GLUT1 is responsible for transporting glucose across the blood- brain barrier rarr
GLUT1 provides glucose for the brain
GLUT1 deficiency syndromebull Brain does not obtain enough glucose from the bloodbull Symptoms seizures developmental delay motor disordersbull Treatment ketogenic diet (high fatlow carb diet)
GLUT1 deficiency syndrome
Glucose
Transporter proteins (membrane protein) can be enriched within artificial membrane
Chloroform and methanol (31)
Phospholipid spontaneously form bilayers
Liposome containing a single type of transport protein are very useful in studying functional properties of transport proteins
bull It is a major experimental tool to study the biochemistry of transport protein function in vitro
bull Widely used as a drug delivery system and for gene transfection
Movement of waterOsmosis movement of water across semipermeable
membraneOsmotic pressure hydrostatic pressure uses to stop
the net flow of water
When CB concentration gt CA
Osmotic pressure π=RT( CB -CA )
Hypertonic solution the concentration is higher than cytosolIsotonic solution equal to cytosolHypotonic solution lower and water move to cytosol
Animal cell ace a problem in maintaining their cell volume within a limited range thereby avoiding lysis
Plant cell has cell wall rarr prevent cell shapeTurgor pressure (膨壓) osmotic pressure plasma membrane against water into the cytosol and then into the vacuole
turgor pressure supplies rigidityThe large forces of turgor pressure are resisted by the strength of cellulose microfibrils in the cell wall
Hypertonic external solution concentration of water is low relative to its concentration inside the cell
Water moves out down its concentration gradientand the cell shrinks
Hypotonic external solution concentration of water is highrelative to its concentration inside the cell
Water moves in down its concentration gradientand the cell swells
When the Na+ ndash K+ pump stops Na+ goes into the cell along its concentration gradient
This adds to the solute concentration in the cytosol
Water moves into the cell along its concentration gradient and the cell bursts
Water draw is equal inside and outside
Water cross membrane is very lowerWater cross membrane via specific channel- aquaporins
Aquaporins increase the water permeability of cell membrane
Aquaporin 1 erythrocyte
Aquaporin2 kidney cells resorb water from urine mutation rarr diabetes insipidus rarr large volume urine
Expression of aquaporin by frog oocytes increases their permeability
control
Injection aquaporin mRNA
Egg move to hypotonic environment
尿崩病
Water channel protein ( aquaporin)
Tetrameric protein
6 α-helices for each subunit
2-nm-long water selective gate
028nm gate widthHighly conserved arginine and histidine in the gateH2 O for HO bonding
with cystein
AquaporinsAquaporins are membrane water channels that play critical are membrane water channels that play critical roles in controlling the roles in controlling the waterwater contents of cellscontents of cells
Water Water crosses the crosses the hydrophobic membranehydrophobic membrane either by simple either by simple diffusion or through a facilitative transport mechanism diffusion or through a facilitative transport mechanism mediated by these mediated by these specialized proteinsspecialized proteins
These protein channels are These protein channels are widely distributedwidely distributed in all kingdoms of in all kingdoms of life including bacteria plants and mammalslife including bacteria plants and mammals
Important in Important in osmotic regulationosmotic regulation acting to prevent bursting of acting to prevent bursting of the cells whenever there are changes of the exterior salt the cells whenever there are changes of the exterior salt concentrationconcentration
ATP powered pump1 P- class
2α 2β
subunit can phosphorylationie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-classbull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-classmaintain low pH in plant vacuolesimilar to F-class
4 ABC (ATP-binding cassete) superfamilyseveral hundred different transport protein
Different classes of pumps exhibit characteristic structure and functional properties
Specific Ion binding site
Must phosphorylation
Transport process requires ATP hydrolysis in which the free energy is liberated by breakdown of ATP into ADP and phosphate
V-class H+ ATP ase pump protons across lysosomal and vacuolar membrane
inside
inside
ATP powered pump
1 P- class
2α 2β
subunit
ie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-class
bull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-class
maintain low pH in plant vacuole
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Each GLUT protein contains 12 membrane-spanning alpha helicesDifferentially expressed
ndash EXAMPLE GLUT4 is only expressed in fat and muscle cells
bull Fat and muscle cells respond to insulin by increasing their uptake of glucose thereby removing glucose from the blood
bull In absence of insulin = GLUT4 on intracellular membranesIn presence of insulin = GLUT4 found on cell surface
bull QUESTION Defects in directing GLUT4 to the cell surface can cause what common disease
Type II diabetes high blood glucose
Copyright copy 2009 Dr Salme Taagepera PhD All rights reserved
GLUT1 is responsible for transporting glucose across the blood- brain barrier rarr
GLUT1 provides glucose for the brain
GLUT1 deficiency syndromebull Brain does not obtain enough glucose from the bloodbull Symptoms seizures developmental delay motor disordersbull Treatment ketogenic diet (high fatlow carb diet)
GLUT1 deficiency syndrome
Glucose
Transporter proteins (membrane protein) can be enriched within artificial membrane
Chloroform and methanol (31)
Phospholipid spontaneously form bilayers
Liposome containing a single type of transport protein are very useful in studying functional properties of transport proteins
bull It is a major experimental tool to study the biochemistry of transport protein function in vitro
bull Widely used as a drug delivery system and for gene transfection
Movement of waterOsmosis movement of water across semipermeable
membraneOsmotic pressure hydrostatic pressure uses to stop
the net flow of water
When CB concentration gt CA
Osmotic pressure π=RT( CB -CA )
Hypertonic solution the concentration is higher than cytosolIsotonic solution equal to cytosolHypotonic solution lower and water move to cytosol
Animal cell ace a problem in maintaining their cell volume within a limited range thereby avoiding lysis
Plant cell has cell wall rarr prevent cell shapeTurgor pressure (膨壓) osmotic pressure plasma membrane against water into the cytosol and then into the vacuole
turgor pressure supplies rigidityThe large forces of turgor pressure are resisted by the strength of cellulose microfibrils in the cell wall
Hypertonic external solution concentration of water is low relative to its concentration inside the cell
Water moves out down its concentration gradientand the cell shrinks
Hypotonic external solution concentration of water is highrelative to its concentration inside the cell
Water moves in down its concentration gradientand the cell swells
When the Na+ ndash K+ pump stops Na+ goes into the cell along its concentration gradient
This adds to the solute concentration in the cytosol
Water moves into the cell along its concentration gradient and the cell bursts
Water draw is equal inside and outside
Water cross membrane is very lowerWater cross membrane via specific channel- aquaporins
Aquaporins increase the water permeability of cell membrane
Aquaporin 1 erythrocyte
Aquaporin2 kidney cells resorb water from urine mutation rarr diabetes insipidus rarr large volume urine
Expression of aquaporin by frog oocytes increases their permeability
control
Injection aquaporin mRNA
Egg move to hypotonic environment
尿崩病
Water channel protein ( aquaporin)
Tetrameric protein
6 α-helices for each subunit
2-nm-long water selective gate
028nm gate widthHighly conserved arginine and histidine in the gateH2 O for HO bonding
with cystein
AquaporinsAquaporins are membrane water channels that play critical are membrane water channels that play critical roles in controlling the roles in controlling the waterwater contents of cellscontents of cells
Water Water crosses the crosses the hydrophobic membranehydrophobic membrane either by simple either by simple diffusion or through a facilitative transport mechanism diffusion or through a facilitative transport mechanism mediated by these mediated by these specialized proteinsspecialized proteins
These protein channels are These protein channels are widely distributedwidely distributed in all kingdoms of in all kingdoms of life including bacteria plants and mammalslife including bacteria plants and mammals
Important in Important in osmotic regulationosmotic regulation acting to prevent bursting of acting to prevent bursting of the cells whenever there are changes of the exterior salt the cells whenever there are changes of the exterior salt concentrationconcentration
ATP powered pump1 P- class
2α 2β
subunit can phosphorylationie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-classbull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-classmaintain low pH in plant vacuolesimilar to F-class
4 ABC (ATP-binding cassete) superfamilyseveral hundred different transport protein
Different classes of pumps exhibit characteristic structure and functional properties
Specific Ion binding site
Must phosphorylation
Transport process requires ATP hydrolysis in which the free energy is liberated by breakdown of ATP into ADP and phosphate
V-class H+ ATP ase pump protons across lysosomal and vacuolar membrane
inside
inside
ATP powered pump
1 P- class
2α 2β
subunit
ie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-class
bull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-class
maintain low pH in plant vacuole
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Copyright copy 2009 Dr Salme Taagepera PhD All rights reserved
GLUT1 is responsible for transporting glucose across the blood- brain barrier rarr
GLUT1 provides glucose for the brain
GLUT1 deficiency syndromebull Brain does not obtain enough glucose from the bloodbull Symptoms seizures developmental delay motor disordersbull Treatment ketogenic diet (high fatlow carb diet)
GLUT1 deficiency syndrome
Glucose
Transporter proteins (membrane protein) can be enriched within artificial membrane
Chloroform and methanol (31)
Phospholipid spontaneously form bilayers
Liposome containing a single type of transport protein are very useful in studying functional properties of transport proteins
bull It is a major experimental tool to study the biochemistry of transport protein function in vitro
bull Widely used as a drug delivery system and for gene transfection
Movement of waterOsmosis movement of water across semipermeable
membraneOsmotic pressure hydrostatic pressure uses to stop
the net flow of water
When CB concentration gt CA
Osmotic pressure π=RT( CB -CA )
Hypertonic solution the concentration is higher than cytosolIsotonic solution equal to cytosolHypotonic solution lower and water move to cytosol
Animal cell ace a problem in maintaining their cell volume within a limited range thereby avoiding lysis
Plant cell has cell wall rarr prevent cell shapeTurgor pressure (膨壓) osmotic pressure plasma membrane against water into the cytosol and then into the vacuole
turgor pressure supplies rigidityThe large forces of turgor pressure are resisted by the strength of cellulose microfibrils in the cell wall
Hypertonic external solution concentration of water is low relative to its concentration inside the cell
Water moves out down its concentration gradientand the cell shrinks
Hypotonic external solution concentration of water is highrelative to its concentration inside the cell
Water moves in down its concentration gradientand the cell swells
When the Na+ ndash K+ pump stops Na+ goes into the cell along its concentration gradient
This adds to the solute concentration in the cytosol
Water moves into the cell along its concentration gradient and the cell bursts
Water draw is equal inside and outside
Water cross membrane is very lowerWater cross membrane via specific channel- aquaporins
Aquaporins increase the water permeability of cell membrane
Aquaporin 1 erythrocyte
Aquaporin2 kidney cells resorb water from urine mutation rarr diabetes insipidus rarr large volume urine
Expression of aquaporin by frog oocytes increases their permeability
control
Injection aquaporin mRNA
Egg move to hypotonic environment
尿崩病
Water channel protein ( aquaporin)
Tetrameric protein
6 α-helices for each subunit
2-nm-long water selective gate
028nm gate widthHighly conserved arginine and histidine in the gateH2 O for HO bonding
with cystein
AquaporinsAquaporins are membrane water channels that play critical are membrane water channels that play critical roles in controlling the roles in controlling the waterwater contents of cellscontents of cells
Water Water crosses the crosses the hydrophobic membranehydrophobic membrane either by simple either by simple diffusion or through a facilitative transport mechanism diffusion or through a facilitative transport mechanism mediated by these mediated by these specialized proteinsspecialized proteins
These protein channels are These protein channels are widely distributedwidely distributed in all kingdoms of in all kingdoms of life including bacteria plants and mammalslife including bacteria plants and mammals
Important in Important in osmotic regulationosmotic regulation acting to prevent bursting of acting to prevent bursting of the cells whenever there are changes of the exterior salt the cells whenever there are changes of the exterior salt concentrationconcentration
ATP powered pump1 P- class
2α 2β
subunit can phosphorylationie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-classbull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-classmaintain low pH in plant vacuolesimilar to F-class
4 ABC (ATP-binding cassete) superfamilyseveral hundred different transport protein
Different classes of pumps exhibit characteristic structure and functional properties
Specific Ion binding site
Must phosphorylation
Transport process requires ATP hydrolysis in which the free energy is liberated by breakdown of ATP into ADP and phosphate
V-class H+ ATP ase pump protons across lysosomal and vacuolar membrane
inside
inside
ATP powered pump
1 P- class
2α 2β
subunit
ie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-class
bull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-class
maintain low pH in plant vacuole
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Transporter proteins (membrane protein) can be enriched within artificial membrane
Chloroform and methanol (31)
Phospholipid spontaneously form bilayers
Liposome containing a single type of transport protein are very useful in studying functional properties of transport proteins
bull It is a major experimental tool to study the biochemistry of transport protein function in vitro
bull Widely used as a drug delivery system and for gene transfection
Movement of waterOsmosis movement of water across semipermeable
membraneOsmotic pressure hydrostatic pressure uses to stop
the net flow of water
When CB concentration gt CA
Osmotic pressure π=RT( CB -CA )
Hypertonic solution the concentration is higher than cytosolIsotonic solution equal to cytosolHypotonic solution lower and water move to cytosol
Animal cell ace a problem in maintaining their cell volume within a limited range thereby avoiding lysis
Plant cell has cell wall rarr prevent cell shapeTurgor pressure (膨壓) osmotic pressure plasma membrane against water into the cytosol and then into the vacuole
turgor pressure supplies rigidityThe large forces of turgor pressure are resisted by the strength of cellulose microfibrils in the cell wall
Hypertonic external solution concentration of water is low relative to its concentration inside the cell
Water moves out down its concentration gradientand the cell shrinks
Hypotonic external solution concentration of water is highrelative to its concentration inside the cell
Water moves in down its concentration gradientand the cell swells
When the Na+ ndash K+ pump stops Na+ goes into the cell along its concentration gradient
This adds to the solute concentration in the cytosol
Water moves into the cell along its concentration gradient and the cell bursts
Water draw is equal inside and outside
Water cross membrane is very lowerWater cross membrane via specific channel- aquaporins
Aquaporins increase the water permeability of cell membrane
Aquaporin 1 erythrocyte
Aquaporin2 kidney cells resorb water from urine mutation rarr diabetes insipidus rarr large volume urine
Expression of aquaporin by frog oocytes increases their permeability
control
Injection aquaporin mRNA
Egg move to hypotonic environment
尿崩病
Water channel protein ( aquaporin)
Tetrameric protein
6 α-helices for each subunit
2-nm-long water selective gate
028nm gate widthHighly conserved arginine and histidine in the gateH2 O for HO bonding
with cystein
AquaporinsAquaporins are membrane water channels that play critical are membrane water channels that play critical roles in controlling the roles in controlling the waterwater contents of cellscontents of cells
Water Water crosses the crosses the hydrophobic membranehydrophobic membrane either by simple either by simple diffusion or through a facilitative transport mechanism diffusion or through a facilitative transport mechanism mediated by these mediated by these specialized proteinsspecialized proteins
These protein channels are These protein channels are widely distributedwidely distributed in all kingdoms of in all kingdoms of life including bacteria plants and mammalslife including bacteria plants and mammals
Important in Important in osmotic regulationosmotic regulation acting to prevent bursting of acting to prevent bursting of the cells whenever there are changes of the exterior salt the cells whenever there are changes of the exterior salt concentrationconcentration
ATP powered pump1 P- class
2α 2β
subunit can phosphorylationie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-classbull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-classmaintain low pH in plant vacuolesimilar to F-class
4 ABC (ATP-binding cassete) superfamilyseveral hundred different transport protein
Different classes of pumps exhibit characteristic structure and functional properties
Specific Ion binding site
Must phosphorylation
Transport process requires ATP hydrolysis in which the free energy is liberated by breakdown of ATP into ADP and phosphate
V-class H+ ATP ase pump protons across lysosomal and vacuolar membrane
inside
inside
ATP powered pump
1 P- class
2α 2β
subunit
ie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-class
bull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-class
maintain low pH in plant vacuole
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Liposome containing a single type of transport protein are very useful in studying functional properties of transport proteins
bull It is a major experimental tool to study the biochemistry of transport protein function in vitro
bull Widely used as a drug delivery system and for gene transfection
Movement of waterOsmosis movement of water across semipermeable
membraneOsmotic pressure hydrostatic pressure uses to stop
the net flow of water
When CB concentration gt CA
Osmotic pressure π=RT( CB -CA )
Hypertonic solution the concentration is higher than cytosolIsotonic solution equal to cytosolHypotonic solution lower and water move to cytosol
Animal cell ace a problem in maintaining their cell volume within a limited range thereby avoiding lysis
Plant cell has cell wall rarr prevent cell shapeTurgor pressure (膨壓) osmotic pressure plasma membrane against water into the cytosol and then into the vacuole
turgor pressure supplies rigidityThe large forces of turgor pressure are resisted by the strength of cellulose microfibrils in the cell wall
Hypertonic external solution concentration of water is low relative to its concentration inside the cell
Water moves out down its concentration gradientand the cell shrinks
Hypotonic external solution concentration of water is highrelative to its concentration inside the cell
Water moves in down its concentration gradientand the cell swells
When the Na+ ndash K+ pump stops Na+ goes into the cell along its concentration gradient
This adds to the solute concentration in the cytosol
Water moves into the cell along its concentration gradient and the cell bursts
Water draw is equal inside and outside
Water cross membrane is very lowerWater cross membrane via specific channel- aquaporins
Aquaporins increase the water permeability of cell membrane
Aquaporin 1 erythrocyte
Aquaporin2 kidney cells resorb water from urine mutation rarr diabetes insipidus rarr large volume urine
Expression of aquaporin by frog oocytes increases their permeability
control
Injection aquaporin mRNA
Egg move to hypotonic environment
尿崩病
Water channel protein ( aquaporin)
Tetrameric protein
6 α-helices for each subunit
2-nm-long water selective gate
028nm gate widthHighly conserved arginine and histidine in the gateH2 O for HO bonding
with cystein
AquaporinsAquaporins are membrane water channels that play critical are membrane water channels that play critical roles in controlling the roles in controlling the waterwater contents of cellscontents of cells
Water Water crosses the crosses the hydrophobic membranehydrophobic membrane either by simple either by simple diffusion or through a facilitative transport mechanism diffusion or through a facilitative transport mechanism mediated by these mediated by these specialized proteinsspecialized proteins
These protein channels are These protein channels are widely distributedwidely distributed in all kingdoms of in all kingdoms of life including bacteria plants and mammalslife including bacteria plants and mammals
Important in Important in osmotic regulationosmotic regulation acting to prevent bursting of acting to prevent bursting of the cells whenever there are changes of the exterior salt the cells whenever there are changes of the exterior salt concentrationconcentration
ATP powered pump1 P- class
2α 2β
subunit can phosphorylationie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-classbull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-classmaintain low pH in plant vacuolesimilar to F-class
4 ABC (ATP-binding cassete) superfamilyseveral hundred different transport protein
Different classes of pumps exhibit characteristic structure and functional properties
Specific Ion binding site
Must phosphorylation
Transport process requires ATP hydrolysis in which the free energy is liberated by breakdown of ATP into ADP and phosphate
V-class H+ ATP ase pump protons across lysosomal and vacuolar membrane
inside
inside
ATP powered pump
1 P- class
2α 2β
subunit
ie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-class
bull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-class
maintain low pH in plant vacuole
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Movement of waterOsmosis movement of water across semipermeable
membraneOsmotic pressure hydrostatic pressure uses to stop
the net flow of water
When CB concentration gt CA
Osmotic pressure π=RT( CB -CA )
Hypertonic solution the concentration is higher than cytosolIsotonic solution equal to cytosolHypotonic solution lower and water move to cytosol
Animal cell ace a problem in maintaining their cell volume within a limited range thereby avoiding lysis
Plant cell has cell wall rarr prevent cell shapeTurgor pressure (膨壓) osmotic pressure plasma membrane against water into the cytosol and then into the vacuole
turgor pressure supplies rigidityThe large forces of turgor pressure are resisted by the strength of cellulose microfibrils in the cell wall
Hypertonic external solution concentration of water is low relative to its concentration inside the cell
Water moves out down its concentration gradientand the cell shrinks
Hypotonic external solution concentration of water is highrelative to its concentration inside the cell
Water moves in down its concentration gradientand the cell swells
When the Na+ ndash K+ pump stops Na+ goes into the cell along its concentration gradient
This adds to the solute concentration in the cytosol
Water moves into the cell along its concentration gradient and the cell bursts
Water draw is equal inside and outside
Water cross membrane is very lowerWater cross membrane via specific channel- aquaporins
Aquaporins increase the water permeability of cell membrane
Aquaporin 1 erythrocyte
Aquaporin2 kidney cells resorb water from urine mutation rarr diabetes insipidus rarr large volume urine
Expression of aquaporin by frog oocytes increases their permeability
control
Injection aquaporin mRNA
Egg move to hypotonic environment
尿崩病
Water channel protein ( aquaporin)
Tetrameric protein
6 α-helices for each subunit
2-nm-long water selective gate
028nm gate widthHighly conserved arginine and histidine in the gateH2 O for HO bonding
with cystein
AquaporinsAquaporins are membrane water channels that play critical are membrane water channels that play critical roles in controlling the roles in controlling the waterwater contents of cellscontents of cells
Water Water crosses the crosses the hydrophobic membranehydrophobic membrane either by simple either by simple diffusion or through a facilitative transport mechanism diffusion or through a facilitative transport mechanism mediated by these mediated by these specialized proteinsspecialized proteins
These protein channels are These protein channels are widely distributedwidely distributed in all kingdoms of in all kingdoms of life including bacteria plants and mammalslife including bacteria plants and mammals
Important in Important in osmotic regulationosmotic regulation acting to prevent bursting of acting to prevent bursting of the cells whenever there are changes of the exterior salt the cells whenever there are changes of the exterior salt concentrationconcentration
ATP powered pump1 P- class
2α 2β
subunit can phosphorylationie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-classbull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-classmaintain low pH in plant vacuolesimilar to F-class
4 ABC (ATP-binding cassete) superfamilyseveral hundred different transport protein
Different classes of pumps exhibit characteristic structure and functional properties
Specific Ion binding site
Must phosphorylation
Transport process requires ATP hydrolysis in which the free energy is liberated by breakdown of ATP into ADP and phosphate
V-class H+ ATP ase pump protons across lysosomal and vacuolar membrane
inside
inside
ATP powered pump
1 P- class
2α 2β
subunit
ie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-class
bull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-class
maintain low pH in plant vacuole
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Hypertonic solution the concentration is higher than cytosolIsotonic solution equal to cytosolHypotonic solution lower and water move to cytosol
Animal cell ace a problem in maintaining their cell volume within a limited range thereby avoiding lysis
Plant cell has cell wall rarr prevent cell shapeTurgor pressure (膨壓) osmotic pressure plasma membrane against water into the cytosol and then into the vacuole
turgor pressure supplies rigidityThe large forces of turgor pressure are resisted by the strength of cellulose microfibrils in the cell wall
Hypertonic external solution concentration of water is low relative to its concentration inside the cell
Water moves out down its concentration gradientand the cell shrinks
Hypotonic external solution concentration of water is highrelative to its concentration inside the cell
Water moves in down its concentration gradientand the cell swells
When the Na+ ndash K+ pump stops Na+ goes into the cell along its concentration gradient
This adds to the solute concentration in the cytosol
Water moves into the cell along its concentration gradient and the cell bursts
Water draw is equal inside and outside
Water cross membrane is very lowerWater cross membrane via specific channel- aquaporins
Aquaporins increase the water permeability of cell membrane
Aquaporin 1 erythrocyte
Aquaporin2 kidney cells resorb water from urine mutation rarr diabetes insipidus rarr large volume urine
Expression of aquaporin by frog oocytes increases their permeability
control
Injection aquaporin mRNA
Egg move to hypotonic environment
尿崩病
Water channel protein ( aquaporin)
Tetrameric protein
6 α-helices for each subunit
2-nm-long water selective gate
028nm gate widthHighly conserved arginine and histidine in the gateH2 O for HO bonding
with cystein
AquaporinsAquaporins are membrane water channels that play critical are membrane water channels that play critical roles in controlling the roles in controlling the waterwater contents of cellscontents of cells
Water Water crosses the crosses the hydrophobic membranehydrophobic membrane either by simple either by simple diffusion or through a facilitative transport mechanism diffusion or through a facilitative transport mechanism mediated by these mediated by these specialized proteinsspecialized proteins
These protein channels are These protein channels are widely distributedwidely distributed in all kingdoms of in all kingdoms of life including bacteria plants and mammalslife including bacteria plants and mammals
Important in Important in osmotic regulationosmotic regulation acting to prevent bursting of acting to prevent bursting of the cells whenever there are changes of the exterior salt the cells whenever there are changes of the exterior salt concentrationconcentration
ATP powered pump1 P- class
2α 2β
subunit can phosphorylationie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-classbull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-classmaintain low pH in plant vacuolesimilar to F-class
4 ABC (ATP-binding cassete) superfamilyseveral hundred different transport protein
Different classes of pumps exhibit characteristic structure and functional properties
Specific Ion binding site
Must phosphorylation
Transport process requires ATP hydrolysis in which the free energy is liberated by breakdown of ATP into ADP and phosphate
V-class H+ ATP ase pump protons across lysosomal and vacuolar membrane
inside
inside
ATP powered pump
1 P- class
2α 2β
subunit
ie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-class
bull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-class
maintain low pH in plant vacuole
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Hypertonic external solution concentration of water is low relative to its concentration inside the cell
Water moves out down its concentration gradientand the cell shrinks
Hypotonic external solution concentration of water is highrelative to its concentration inside the cell
Water moves in down its concentration gradientand the cell swells
When the Na+ ndash K+ pump stops Na+ goes into the cell along its concentration gradient
This adds to the solute concentration in the cytosol
Water moves into the cell along its concentration gradient and the cell bursts
Water draw is equal inside and outside
Water cross membrane is very lowerWater cross membrane via specific channel- aquaporins
Aquaporins increase the water permeability of cell membrane
Aquaporin 1 erythrocyte
Aquaporin2 kidney cells resorb water from urine mutation rarr diabetes insipidus rarr large volume urine
Expression of aquaporin by frog oocytes increases their permeability
control
Injection aquaporin mRNA
Egg move to hypotonic environment
尿崩病
Water channel protein ( aquaporin)
Tetrameric protein
6 α-helices for each subunit
2-nm-long water selective gate
028nm gate widthHighly conserved arginine and histidine in the gateH2 O for HO bonding
with cystein
AquaporinsAquaporins are membrane water channels that play critical are membrane water channels that play critical roles in controlling the roles in controlling the waterwater contents of cellscontents of cells
Water Water crosses the crosses the hydrophobic membranehydrophobic membrane either by simple either by simple diffusion or through a facilitative transport mechanism diffusion or through a facilitative transport mechanism mediated by these mediated by these specialized proteinsspecialized proteins
These protein channels are These protein channels are widely distributedwidely distributed in all kingdoms of in all kingdoms of life including bacteria plants and mammalslife including bacteria plants and mammals
Important in Important in osmotic regulationosmotic regulation acting to prevent bursting of acting to prevent bursting of the cells whenever there are changes of the exterior salt the cells whenever there are changes of the exterior salt concentrationconcentration
ATP powered pump1 P- class
2α 2β
subunit can phosphorylationie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-classbull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-classmaintain low pH in plant vacuolesimilar to F-class
4 ABC (ATP-binding cassete) superfamilyseveral hundred different transport protein
Different classes of pumps exhibit characteristic structure and functional properties
Specific Ion binding site
Must phosphorylation
Transport process requires ATP hydrolysis in which the free energy is liberated by breakdown of ATP into ADP and phosphate
V-class H+ ATP ase pump protons across lysosomal and vacuolar membrane
inside
inside
ATP powered pump
1 P- class
2α 2β
subunit
ie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-class
bull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-class
maintain low pH in plant vacuole
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

When the Na+ ndash K+ pump stops Na+ goes into the cell along its concentration gradient
This adds to the solute concentration in the cytosol
Water moves into the cell along its concentration gradient and the cell bursts
Water draw is equal inside and outside
Water cross membrane is very lowerWater cross membrane via specific channel- aquaporins
Aquaporins increase the water permeability of cell membrane
Aquaporin 1 erythrocyte
Aquaporin2 kidney cells resorb water from urine mutation rarr diabetes insipidus rarr large volume urine
Expression of aquaporin by frog oocytes increases their permeability
control
Injection aquaporin mRNA
Egg move to hypotonic environment
尿崩病
Water channel protein ( aquaporin)
Tetrameric protein
6 α-helices for each subunit
2-nm-long water selective gate
028nm gate widthHighly conserved arginine and histidine in the gateH2 O for HO bonding
with cystein
AquaporinsAquaporins are membrane water channels that play critical are membrane water channels that play critical roles in controlling the roles in controlling the waterwater contents of cellscontents of cells
Water Water crosses the crosses the hydrophobic membranehydrophobic membrane either by simple either by simple diffusion or through a facilitative transport mechanism diffusion or through a facilitative transport mechanism mediated by these mediated by these specialized proteinsspecialized proteins
These protein channels are These protein channels are widely distributedwidely distributed in all kingdoms of in all kingdoms of life including bacteria plants and mammalslife including bacteria plants and mammals
Important in Important in osmotic regulationosmotic regulation acting to prevent bursting of acting to prevent bursting of the cells whenever there are changes of the exterior salt the cells whenever there are changes of the exterior salt concentrationconcentration
ATP powered pump1 P- class
2α 2β
subunit can phosphorylationie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-classbull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-classmaintain low pH in plant vacuolesimilar to F-class
4 ABC (ATP-binding cassete) superfamilyseveral hundred different transport protein
Different classes of pumps exhibit characteristic structure and functional properties
Specific Ion binding site
Must phosphorylation
Transport process requires ATP hydrolysis in which the free energy is liberated by breakdown of ATP into ADP and phosphate
V-class H+ ATP ase pump protons across lysosomal and vacuolar membrane
inside
inside
ATP powered pump
1 P- class
2α 2β
subunit
ie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-class
bull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-class
maintain low pH in plant vacuole
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Water cross membrane is very lowerWater cross membrane via specific channel- aquaporins
Aquaporins increase the water permeability of cell membrane
Aquaporin 1 erythrocyte
Aquaporin2 kidney cells resorb water from urine mutation rarr diabetes insipidus rarr large volume urine
Expression of aquaporin by frog oocytes increases their permeability
control
Injection aquaporin mRNA
Egg move to hypotonic environment
尿崩病
Water channel protein ( aquaporin)
Tetrameric protein
6 α-helices for each subunit
2-nm-long water selective gate
028nm gate widthHighly conserved arginine and histidine in the gateH2 O for HO bonding
with cystein
AquaporinsAquaporins are membrane water channels that play critical are membrane water channels that play critical roles in controlling the roles in controlling the waterwater contents of cellscontents of cells
Water Water crosses the crosses the hydrophobic membranehydrophobic membrane either by simple either by simple diffusion or through a facilitative transport mechanism diffusion or through a facilitative transport mechanism mediated by these mediated by these specialized proteinsspecialized proteins
These protein channels are These protein channels are widely distributedwidely distributed in all kingdoms of in all kingdoms of life including bacteria plants and mammalslife including bacteria plants and mammals
Important in Important in osmotic regulationosmotic regulation acting to prevent bursting of acting to prevent bursting of the cells whenever there are changes of the exterior salt the cells whenever there are changes of the exterior salt concentrationconcentration
ATP powered pump1 P- class
2α 2β
subunit can phosphorylationie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-classbull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-classmaintain low pH in plant vacuolesimilar to F-class
4 ABC (ATP-binding cassete) superfamilyseveral hundred different transport protein
Different classes of pumps exhibit characteristic structure and functional properties
Specific Ion binding site
Must phosphorylation
Transport process requires ATP hydrolysis in which the free energy is liberated by breakdown of ATP into ADP and phosphate
V-class H+ ATP ase pump protons across lysosomal and vacuolar membrane
inside
inside
ATP powered pump
1 P- class
2α 2β
subunit
ie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-class
bull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-class
maintain low pH in plant vacuole
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Aquaporin 1 erythrocyte
Aquaporin2 kidney cells resorb water from urine mutation rarr diabetes insipidus rarr large volume urine
Expression of aquaporin by frog oocytes increases their permeability
control
Injection aquaporin mRNA
Egg move to hypotonic environment
尿崩病
Water channel protein ( aquaporin)
Tetrameric protein
6 α-helices for each subunit
2-nm-long water selective gate
028nm gate widthHighly conserved arginine and histidine in the gateH2 O for HO bonding
with cystein
AquaporinsAquaporins are membrane water channels that play critical are membrane water channels that play critical roles in controlling the roles in controlling the waterwater contents of cellscontents of cells
Water Water crosses the crosses the hydrophobic membranehydrophobic membrane either by simple either by simple diffusion or through a facilitative transport mechanism diffusion or through a facilitative transport mechanism mediated by these mediated by these specialized proteinsspecialized proteins
These protein channels are These protein channels are widely distributedwidely distributed in all kingdoms of in all kingdoms of life including bacteria plants and mammalslife including bacteria plants and mammals
Important in Important in osmotic regulationosmotic regulation acting to prevent bursting of acting to prevent bursting of the cells whenever there are changes of the exterior salt the cells whenever there are changes of the exterior salt concentrationconcentration
ATP powered pump1 P- class
2α 2β
subunit can phosphorylationie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-classbull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-classmaintain low pH in plant vacuolesimilar to F-class
4 ABC (ATP-binding cassete) superfamilyseveral hundred different transport protein
Different classes of pumps exhibit characteristic structure and functional properties
Specific Ion binding site
Must phosphorylation
Transport process requires ATP hydrolysis in which the free energy is liberated by breakdown of ATP into ADP and phosphate
V-class H+ ATP ase pump protons across lysosomal and vacuolar membrane
inside
inside
ATP powered pump
1 P- class
2α 2β
subunit
ie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-class
bull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-class
maintain low pH in plant vacuole
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-
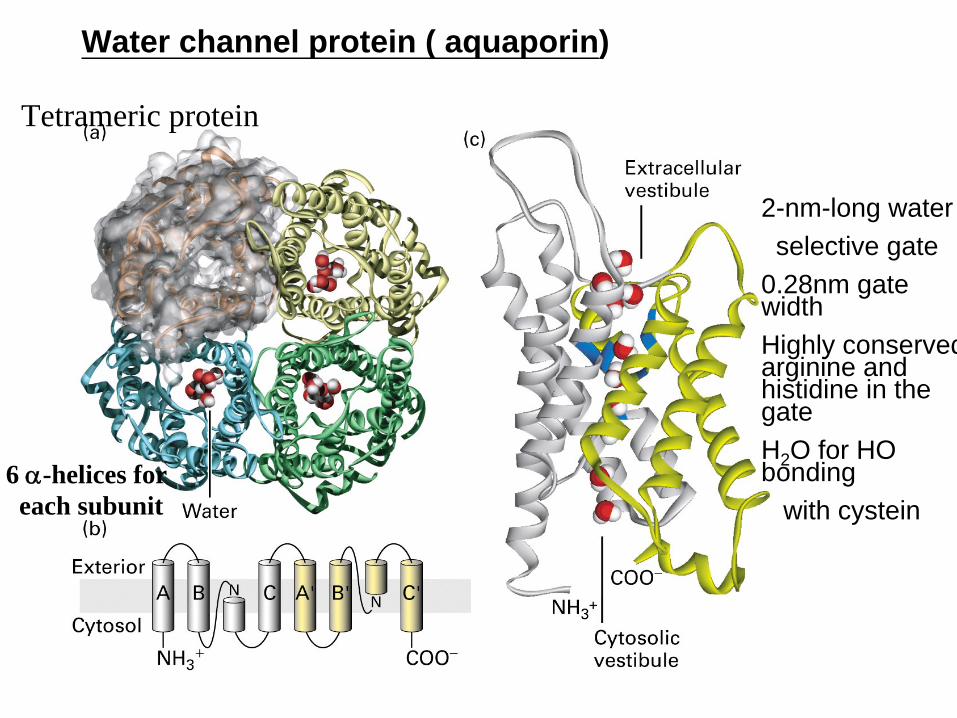
Water channel protein ( aquaporin)
Tetrameric protein
6 α-helices for each subunit
2-nm-long water selective gate
028nm gate widthHighly conserved arginine and histidine in the gateH2 O for HO bonding
with cystein
AquaporinsAquaporins are membrane water channels that play critical are membrane water channels that play critical roles in controlling the roles in controlling the waterwater contents of cellscontents of cells
Water Water crosses the crosses the hydrophobic membranehydrophobic membrane either by simple either by simple diffusion or through a facilitative transport mechanism diffusion or through a facilitative transport mechanism mediated by these mediated by these specialized proteinsspecialized proteins
These protein channels are These protein channels are widely distributedwidely distributed in all kingdoms of in all kingdoms of life including bacteria plants and mammalslife including bacteria plants and mammals
Important in Important in osmotic regulationosmotic regulation acting to prevent bursting of acting to prevent bursting of the cells whenever there are changes of the exterior salt the cells whenever there are changes of the exterior salt concentrationconcentration
ATP powered pump1 P- class
2α 2β
subunit can phosphorylationie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-classbull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-classmaintain low pH in plant vacuolesimilar to F-class
4 ABC (ATP-binding cassete) superfamilyseveral hundred different transport protein
Different classes of pumps exhibit characteristic structure and functional properties
Specific Ion binding site
Must phosphorylation
Transport process requires ATP hydrolysis in which the free energy is liberated by breakdown of ATP into ADP and phosphate
V-class H+ ATP ase pump protons across lysosomal and vacuolar membrane
inside
inside
ATP powered pump
1 P- class
2α 2β
subunit
ie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-class
bull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-class
maintain low pH in plant vacuole
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

AquaporinsAquaporins are membrane water channels that play critical are membrane water channels that play critical roles in controlling the roles in controlling the waterwater contents of cellscontents of cells
Water Water crosses the crosses the hydrophobic membranehydrophobic membrane either by simple either by simple diffusion or through a facilitative transport mechanism diffusion or through a facilitative transport mechanism mediated by these mediated by these specialized proteinsspecialized proteins
These protein channels are These protein channels are widely distributedwidely distributed in all kingdoms of in all kingdoms of life including bacteria plants and mammalslife including bacteria plants and mammals
Important in Important in osmotic regulationosmotic regulation acting to prevent bursting of acting to prevent bursting of the cells whenever there are changes of the exterior salt the cells whenever there are changes of the exterior salt concentrationconcentration
ATP powered pump1 P- class
2α 2β
subunit can phosphorylationie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-classbull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-classmaintain low pH in plant vacuolesimilar to F-class
4 ABC (ATP-binding cassete) superfamilyseveral hundred different transport protein
Different classes of pumps exhibit characteristic structure and functional properties
Specific Ion binding site
Must phosphorylation
Transport process requires ATP hydrolysis in which the free energy is liberated by breakdown of ATP into ADP and phosphate
V-class H+ ATP ase pump protons across lysosomal and vacuolar membrane
inside
inside
ATP powered pump
1 P- class
2α 2β
subunit
ie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-class
bull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-class
maintain low pH in plant vacuole
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

ATP powered pump1 P- class
2α 2β
subunit can phosphorylationie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-classbull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-classmaintain low pH in plant vacuolesimilar to F-class
4 ABC (ATP-binding cassete) superfamilyseveral hundred different transport protein
Different classes of pumps exhibit characteristic structure and functional properties
Specific Ion binding site
Must phosphorylation
Transport process requires ATP hydrolysis in which the free energy is liberated by breakdown of ATP into ADP and phosphate
V-class H+ ATP ase pump protons across lysosomal and vacuolar membrane
inside
inside
ATP powered pump
1 P- class
2α 2β
subunit
ie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-class
bull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-class
maintain low pH in plant vacuole
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Specific Ion binding site
Must phosphorylation
Transport process requires ATP hydrolysis in which the free energy is liberated by breakdown of ATP into ADP and phosphate
V-class H+ ATP ase pump protons across lysosomal and vacuolar membrane
inside
inside
ATP powered pump
1 P- class
2α 2β
subunit
ie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-class
bull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-class
maintain low pH in plant vacuole
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

V-class H+ ATP ase pump protons across lysosomal and vacuolar membrane
inside
inside
ATP powered pump
1 P- class
2α 2β
subunit
ie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-class
bull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-class
maintain low pH in plant vacuole
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

inside
ATP powered pump
1 P- class
2α 2β
subunit
ie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-class
bull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-class
maintain low pH in plant vacuole
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

ATP powered pump
1 P- class
2α 2β
subunit
ie Na+-K+ ATP ase Ca+ATP ase H+pump
2 F-class
bull
locate on bacterial membrane chloroplast and mitochondria
bull
pump proton from exoplasmic space to cytosolic for ATP synthesis
3 V-class
maintain low pH in plant vacuole
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Neel large energy RBC need 50 ATP for NaK pump nerve and kidney need 25 for ion transport
Extracellular intracellular Extracellular intracellular
Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Muscle relaxation depends on Ca2+ APTase that pump Ca2+ from the cytosol into the sacroplasmic reticulum (SR)
In muscle cell cytosol Ca2+ 10-7 M (resting state) to 10-6M (contraction)
Most intracellular Ca2+ storage in SR(10-2M)
Ca2+ is released from the sarcoplasmic reticulum through Ca2+ release channels when the muscle contracts
Most cytosol Ca2+ transport into SR via Ca2+ ATPase pumpCa2+ pump comprises 90 of the sarcoplasmic reticulum
membrane proteinResponsible for restoring the Ca2+ gradient (pumps it back into
the sarcoplasmic reticulum
Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Operational model of the Ca2+-ATPase in the SR membrane of skeletal muscle cells
Higher Ca+2
Lower Ca+2
Plays a major role in muscle relaxationby transporting released Ca2+ back into SRA single subunit protein with 10 transmembrane fragmentsIs highly homologous to NaK-ATPase
10-2
10-6
Low affinity for calcium
Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Conformational change
α-helix
Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Ca++
ATP ADP + Pi
Ca++
signal
calmodulin
endoplasmicreticulum
Ca++
Ca++-ATPase
Ca++-release channel
outside of cell
ATP ADP + Pi
Ca++
signal-activated channelcytosol Ca++-ATPase
Ca++
Calmodulin-mediated activation of plasma membrane Ca2+ ATPase leads to rapid Ca2+ export rarr keep cytosolic Ca2+ very low
Allosteric activation
Calmodulin regulates the plasma membrane Ca2+ pump that control cytosolic Ca2+ concentration
ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

ATP-powered ion pumps generate and maintain ionic gradients across cellular membranes
Extracellular intracellular
Na+K+ ATPase maintain the intracellular Na+ and K+ concentration in animal cell
High lowLow high
Na+ transport outK+ transport in
By Na+K+ ATPase
Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Greatest consumer cellular energySets up concentration amp electrical gradientsHydrolysis of 1 ATP moves 2K+ in and 3Na+ out against their concentration gradients
Na+ K+ ATPase(maintain the intracellular Na and K concentration in animal cell)
Higher affinity for Na+
Membrane potential Utilizes 30 cells energy
Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Four major domainsFour major domains
M M -- MembraneMembrane--bound domain which is composed of 10 bound domain which is composed of 10 transmembranetransmembrane segmentssegments
NN-- NucleotideNucleotide--binding domain where adenine moiety of binding domain where adenine moiety of ATP and ADP bindsATP and ADP binds
P P ndashndash Phosphatase domain which contains invariant Asp Phosphatase domain which contains invariant Asp residue which became residue which became phosphorylatedphosphorylated during the ATP during the ATP hydrolysishydrolysis
A domain A domain ndashndash essential for conformational transitions between essential for conformational transitions between E1 and E2 statesE1 and E2 states
Na+K+ ATPase
Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Na+- K+ Pump on the Plasma Membrane
K+ is 10 to 20 X higher inside animal cells than outsideNa+ is 10 to 20 X higher outside animal cells than insideThese concentration gradients are maintained by the Na+ - K+
pump on the plasma membranendash Pump operates as an antiporter pumping K+ in and Na+
outTransport cycle depends on autophosphorylation of the protein
ndash Terminal phosphate of ATP is transferred to an aspartic acid of the pumpbull Ion pumps that autophosphorylate are called P-type
transport ATPases
Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Na+- K+ Pump on the Plasma Membrane
The Na+- K+ pump is electrogenicndash It generates an electrical potential (known as membrane
potential) across the membranebull Reason
ndash Pumps 3 Na+ ions out for every 2 K+ ions it pumps in
ndash Thus the inside of the cell is negative relative to the outside
Electrogenic effect of the pump contributes only ~10 of the membrane potentialndash remaining 90 is only indirectly attributable to the
Na+- K+ pump (discussed later)
Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Effect of proton pumping by V-class ion pumps on H+ concentration gradients and electric potential gradients across cellular membrane
V-class H+ ATPase pump protons across lysosomal an vacuolar membranes
Generation of electrochemical gradient
Electrochemical gradient combines the membrane potential and concentration gradient which work additively to increase the driving force
Only transport H+
ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

ABC TransportersLargest family of membrane transport proteins
ndash 78 genes (5 of genome) encode ABC transporters in E colindash Many more in animal cellsndash Known as the ABC transporter superfamily
They use the energy derived from ATP hydrolysis to transport a variety of small molecules includingndash Amino acids sugars inorganic ions peptides
ABC transporters also catalyze the flipping of lipids between monolayers in membranes
All ABC transporters each contain 2 highly conserved ATP- binding domains
Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Bacterial permease are ABC proteins that import a variety of nutrients from the environment
Structure of the E coil BtuCD protein an ABC transporter mediating vitamin B12 uptake
ATP-binding cassette
ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

ABC transporter
bull2 T ( transmembrane ) domain each has 6 α- helix form pathways for transported substance
bull2A ( ATP- binding domain) 30-40 homology for membranesie bacterial permease
bull
use ATP hydrolysisbull
transport aa sugars vitamines or peptides
bull
inducible depend on the environmental conditionie mammalian ABC transporter ( Multi Drug Resistant)
bull
export drug from cytosol to extracellular mediumbull
mdr gene amplified by drugs stimulation
bull
mostly hydrophobic for MDR proteinscancer cell resistant to drug mechanisms
Bacterial permeases are ABC proteins that import a variety of nutrients from the enviornment
About 50 ABC small-molecule pumps are known in mammals
A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

A section of the double membrane of E coli
Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Auxiliary transport system associated with transport ATPases in bacteria with double membranes
The transport ATPases belong to the ABC transporter supefamily
Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Examples of a few ABC proteins
Nature Structural amp Molecular Biology 11 918 - 926 (2004)
The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

The Multidrug Resistance Protein (MDR)ABC (ATP-binding cassette)
170 Kdalton P-glycoprotein that pumps hydrophobic drugs out of cells in a ATP-dependent fashion
Uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium
It reduces the cytoplasmic concentration of drugs and hence their toxicity It therefore reduces the effectiveness of chemotherapeutic drugs It is overexpressed in some tumour cells Need high concentration to killed cell
It transports a wide range of chemically unrelated proteins including the anthracyclines actinomycine D valinomycin and gramicidin
The approximately 50 mammalian ABC transporters play diverse and important roles in cell and organ physiology
A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

A typical ABC transporter consists of four domains ndash two highly hydrophobicdomains and two ATP-binding catalytic domains
ATP binding leads to dimerization of the two ATP-binding domains and ATPhydrolysis leads to their dissociation
Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Action of the Multi-drug Resistant Transport Protein
Mode of action of MDR1 involves flipping [flippase] or pumping of the lipid soluble drugs that typically have some positive charges across the membrane (ATPase side) to the exterior where the drug is released to the outside
MDR1 is found in high activity in organs like liver and kidney that play a major role in the breakdown of drugs and other toxic substances but unfortunately reaches highly unregulated levels in corresponding tumor cells
Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Structural model of E coli lipid flippase and ABC protein homologous to mammalian MDR1
ABC protein that transport lipid-soluble substrates may operated by a flippase mechanism
Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Proposed mechanisms of action for the MDR1 protein
FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

FlippasesLipids can be moved from one
monolayer to the other by flippase proteins
Some flippases operate passively and do not require an energy source
Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Active flippases can generate membrane asymmetries
Certain ABC proteins ldquo filprdquo phospholipids and other lipid-soluble substrates from one membrane leaflet to opposite leaflet
Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Flippase model of transport by MDR1 and similar ABC protein
Spontaneously
Diffuses laterally
Flips the charged substrate molecule
1 Hydrophobic portion of target molecule spontaneously inserts itself into the inner leaflet
2 Molecule diffuses laterally until it bumps into MDR
3 MDR ldquoflipsrdquo the molecule from the inner to outer leaflet (this step is energetically unfavorable and requires ATP)
4 Molecule diffuses away and
5 Spontaneously moves out of the outer leaflet
Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Diseases linked with ABC proteins1 ALD( X-link adrenoleukodestrophy)
defect in ABC transport protein( ABCD1)
located on peroxisome used for transport for very long fatty acid absence ABCD1rarr fatty acid rarraccumulate cytosol rarr cell damage
2 Tangiers disease
Dificiency in plasma ABCA1 proteins which is used for transport of phospholipis and cholesterol
3 Cystic fibrosis
mutation of CTFR( cystic fibrosis transmenbrane regulator a Cl- transporter in the apical membrane of lung sweat gland and pancrease)
licked it rarr did not resorption of Cl rarr taste saltyrarrThis leads to abnormalities in the pancreas skin intestine sweat glands and lungs
腎上腺腦白質失氧症
脂蛋白缺乏疾病
囊狀纖維化
Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Polycystic kidney disease 多囊性腎病
這個病的病理是腎內生出很 多個小型的囊腫這些囊腫 慢慢的長大產生壓迫力 使周圍的腎組織功能上產生 障礙而致腎衰竭病徵
多囊腎的病徵腎小管不大斷 擴大令腎臟增大以及影 響腎功能
PDK1 or PDK2 mutation
Regulation of ion transport
Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Incorporation of FAs into membrane lipids takes place on organelle membranes
Fig 18-4 Phospholipid synthesis
Fatty acid didnrsquot directly pass membraneAcetyl CoA
-------- saturated fatty acidacetyl-CoA
carboxylasefatty acid synthase
Annexin
V binds to anionic phospholipidslong exposure of exoplasmic face of plasma membsignal for scavenger cells to remove dying cells
Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Flippases move phospholipids from one membrane leaflet to the opposite leaflet
asymmetric distribution of phospholipids
senescence or apoptosis ndash disturb the asymmetric distribution
Phosphatidylserine (PS) and phosphatidylethanolamine cytosolic leaflet
exposure of these anionic phospholipids on the exoplasmic face ndash signal
for scavenger cells to remove and destroy
Annexin V ndash a protein that specifically binds to PS phospholipids
fluorescently labeled annexin Vndash to detect apoptotic cells
flippase ABC superfamily of small molecule pumps
In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

In vitro fluorescence quenching assay can detect phospholipid flippase activity of ABCB4
Yeast sec mutant ndash at nonpermissive temp secretory vesicle cannot fuse with plasma membrane ndash purify the secretory vesicles
(dithionite)
Phospholipid
flippase
activity of ABCB4
有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

有為的青年伸個懶腰讓我們一起繼續前進進入細胞生物學領域另一個高深且符合我們的境界也就是溫老師會考的範圍也就各位研究所考試------可能考的地方之一雖然這幾年國立大學研究所考得不多但勿恃敵之不來恃吾有以待之
Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Nongated ion channels and the resting membrane potentialGated need ligand to activation Non-gated do not need ligand
Ion Channel (non-gate)Generation of electrochemical gradient across plasma membraneie Ca+ gradient
regulation of signal transduction muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells
ie Na+ gradient uptake of aa symport antiport formed membrane potential
ie K+ gradientformed membrane potential
Q how does the electrochemical gradient formedSelective movement of Ions Create a transmembrane electric potential difference
Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Depending on the type of the channel this gating process may be driven by
1 ligand binding (ligand-gated channels)2 changes in electrical potential across cell membrane (voltage-gatedchannels)3 mechanical forces acting on cellular components (mechanosensitivechannels)
Ion gating Channel
Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Gated ion channels respond to different kinds of stimuli
Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Selective movement of ions creates a transmembrane electric potential difference
Ion no move rarr no membrane potentialIon move rarr create membrane potential
各位把它想
成細胞外大
量Na流入
內結果膜
外電壓會出
現負
59mv
The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

The membrane potential in animal cells depends largely on resting K+
channel
Many open K+ channel but few open Na+ Cl- or Ca2+ channels on animal membraneSo major ionic movement across the membrane is K+ it form the inside out ward by
the K+ concentration gradient rarr creating an positive charge on the outside outward flow of K+ ions through these channels also called resting K+ channels
-59mv-59mv
各位把它想
成細胞內大
量K流出去 結果膜外電
壓會出現正
但膜內會
是負
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-
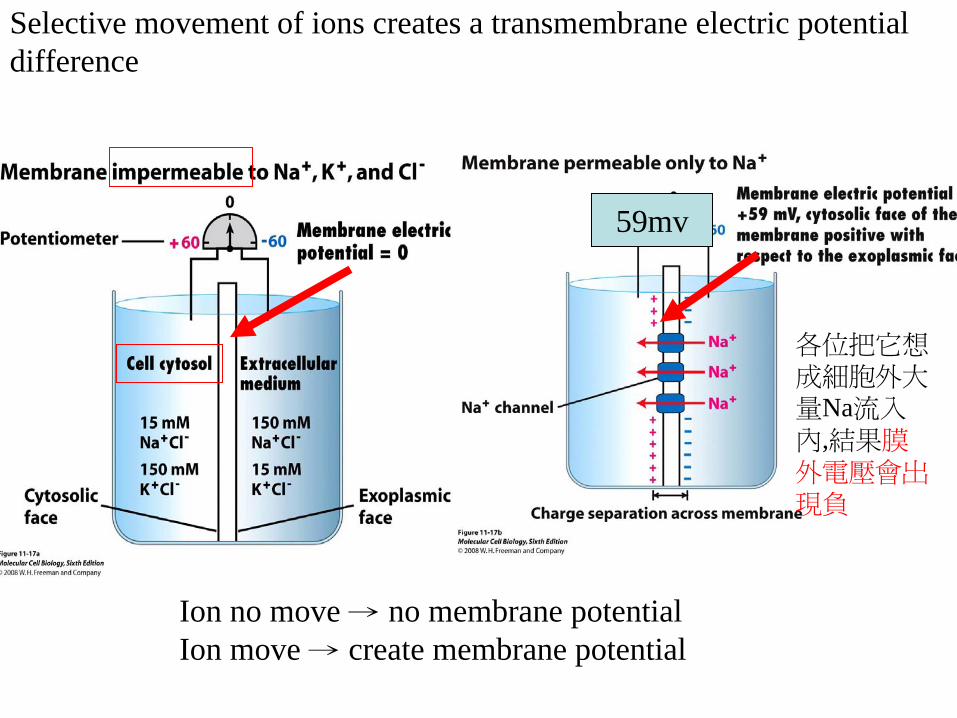
Negative charge on intracellular organic anions balanced by K+
High intracellular [K+] generated by Na+-K+ ATPaseLarge K+ concentration gradient ([K+]i [K+]o asymp
30)
Plasma membrane contains spontaneously active K+ channels rArr K+ move freely out of cell
As K+ moves out of cell leaves negative charge build up rArr opposes further K+ exitAt equilibrium electrical force balances concentration gradient and electrochemical
gradient for K+ is zero (even though there is still a very substantial K+
concentration gradient)Resting membrane potential = flow of positivenegative ions across plasma
membrane precisely balancedMembrane potential measured as voltage difference across membraneFor animal cells resting membrane potential varies between -20 and -200 mVNegative value due to negativity of intracellular compartment compared to
extracellular fluidBecause K+ channels predominate in resting plasma membrane resting membrane
potential mainly due to K+ concentration gradientNernst equation permits calculation of membrane potential (V)
Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Potential difference exists across every cellrsquos plasma membranendash cytoplasm side is negative pole and
extracellular fluid side is positive poleInside of cell negatively charged because
ndash large negatively charged molecules are more abundant inside the cell
ndash sodium potassium ATPase pumpndash resting K+ ion channels (from in to out flow)
Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Ion channels contain a selectivity filter formed from conserved transmembrane a helices and p segmentsIon-selectivity filter
Transmembrane domain
Structure like but function differentStructure of resting K+channel from the bacterium Streptomyces lividans
Ion channels contain a selectivity filter formed from conserved transmembrane α helices and P segment
Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Voltage-gated K+ channels have four subunits each containing six transmembrane α
helices
Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Ion channels are selective pores in the membrane
Ion channels have ion selectivity - they only allow passage of specific molecules
Ion channels are not open continuously conformational changes open and close
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-
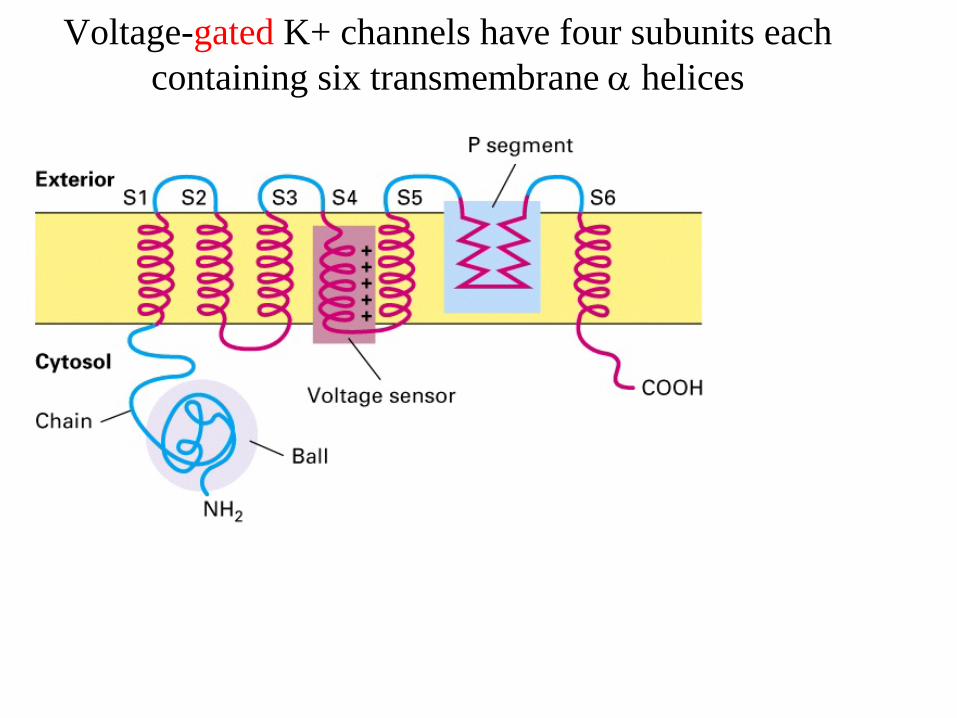
Smaller Na+ does not fit perfectly
Each ion contain eight water molecules
Mechanism of ion selectivity and transport in resting K+ channel
EACH OF THE binding sites closely mimics potassium ions octahedral hydration shell thereby minimizing the energy required to strip off their water coats Because of their smaller size sodium ions dont fit in these binding sites as snugly and thus find the energetic cost of trading their water coat for a spot in the selectivity filter too high
In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

In the vestibule the ions are hydrated In the selectivity filter the carbonyl oxygens areplaced precisely to accommodate a dehydrated K+ ion The dehydration of the K+ ionrequires energy which is precisely balanced by the energy regained by the interactionof the ion with the carbonyl oxygens that serve as surrogate water molecules
Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Loss four of eight H2O
bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

bull 鈉離子通道(sodium channel)03times05大小但更重要的是其內表
面帶有極強的負電荷這些負電荷主要 會把鈉離子拉向通道這是因為脫水後 的鈉離子直徑要比其它離子小
bull 鉀離子通道
03times03的大小但它們不帶有負電 荷 但是鉀的水合離子比鈉的水合離子 要小得多因此體積小的水合鉀離子就
可以很容易地穿過這個較小的通道而 鈉離子則不行
Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Patch clamps permit measurement of ion movements through single channels
effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

effect effectV=I x R
Ion flux through individual Na+ channel
Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Novel ion channels can be characterized by a combination of oocyte expression and path clamping
Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Na+ entry into mammalian cells has a negative change in free energy
Transmembrane forces acting on Na+ ions
CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

CotransportUse the energy stored in Na+ or H+ electrochemical gradient to power the transport of another substance
Symport the transportd molecules and cotransported ion move in the same direction
Antiport the transported molecules move in opposited direction
Depending on how many solute molecules are transported and in what direction carrier proteins are dubbed uniporters symporters or antiporters
Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Coupling of Active Transport to Ion Gradients without energy against gradient
In mammalian cells Na+ electrochemical gradient is maintained across the plasma membrane by active transport of Na+ out of the cell using ATP as an energy sourcendash This electrochemical gradient provides the driving force
for the active transport of a 2nd soluteEg in intestinal and kidney cells symport systems driven by
the Na+ gradient are used to transport sugars and amino acids into the cells
Bigger the Na+ gradient the greater the rate of solute entry
Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Only small hydrophobic molecules cross membrane by simple diffusion
Electrochemical gradient
Membrane potential
Concentration gradient
Ion concentration gradients across the membranes establishes the membrane electric potential
The differences in ion concentrations across the membrane establishes a membrane electrochemical gradient
An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

An electrochemical gradient combines the membrane potential and the concentration gradient
Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Operation Model for the two-Na+one glucose symport
Glucose transport against its gradient in the epithelial cells of intestine
Na+ linked symporters import amino acids and glucose into animal cells against high concentration gradients
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-
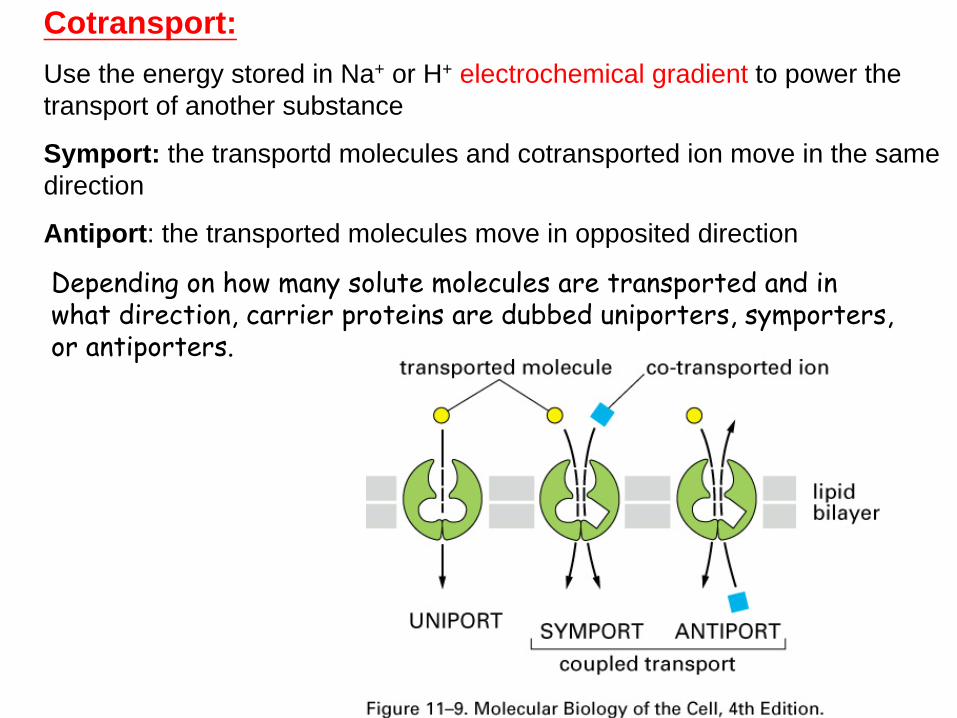
Carrier oscillates betweenstate A and state B
Binding of Na+ and glucoseis cooperative (binding of either ligand induces a conformational change thatenhances binding of the 2nd
ligand)
Since Na+ higher in the extracellular space (amp very low inside) glucosemore likely to bind in A state
AccordinglyNa+ and glucose enter the cell (by an A to B transition) moreoften than they leave the cell
Result is net transport of Na+ and glucose into thecell
The Na+ gradientIs used to driveactive transportof glucose
Na+ pumped outby an ATP-driven pump
Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Three carrier proteins appropriately positioned in the plasma membrane function to transport glucose across the intestinal epithelium
Without ion force
Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Increases PM area
Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Bacterial symporter structure reveals the mechanism of substrate binding
No 3-D Mammalian sodium symporter it similar to bacterial sodium-amino acid transporter
Bind to sodium rarrconformation change rarr bind to amino acid rarr transport substrate
3-D structural of the two Na+ one leucine symporter
In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

In cardiac or muscle Ca2+ uarrrarr contractionNormal cytosol is 10000 fold of Ca2+ concentration than Cardiac (10-6 rarr 10-2 M)Cardiac muscles contain 3Na+ 1 Ca2+ antiporterMovement of three sodium is required to power the export of one calcium
3Na+ outside + Ca+2
inside 3Na+ inside + Ca+2
outside
maintenance of low cytosolic Ca 2+ concentration
ie inhibition of Na+K+ ATPase by Quabain and Digoxin
raises cytosolic Na+
lowers the efficiency of Na+Ca+2 antiport
increases cytosolic Ca+2
( used in cogestive heart failure)
Na+ linked antiport Exports Ca+2 from cardiac Muscle Cells
More contraction
Electrochemical gradient
Normal condition
烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

烏本 毛地黃素
Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Cotransporters that regulate cytosolic pH
H2 CO3 H+ + HCO-
H+ can be neutrolized by
1Na+HCO3-Cl- antiport
2 Cabonic anhydrase
HCO3- CO2 +OH-
3 Na+H+ antiport
Carrier proteins in the plasma membrane regulate cytosolic pH(pHi ) at about 72
There are two mechanisms by which this pH is regulated- H+ is transported out of the cell
Na+-H+ exchanger an antiporter couples the influx of Na+ to an efflux of H+
- HCO3- is brought into the cell to neutralize H+ in the cytosol
Na+ -driven Cl- - HCO3- exchanger uses a combination
of the two mechanisms by coupling an influx of Na+ andHCO3 to an efflux of Cl- and H+
The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

The activity of membrane transport proteins that regulated the cytosolic pH of memmalian cells changes with pH
A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

A putative cation exchange protein plays a key role in evolution of human skin pigmentation
SLC245A5 a human transporter regulated skin color
TEM of skin melanophores
Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Plant vacuole membrane
bull
pH 3mdash6bull
Low pH maintained by
V-class ATP-powered pumppyrophosphate-hydrolyzing proton pump (PPi -powered pump)
The H+ pump inside rarr inside positive rarr
powers mover negative ion move inside High positive inside rarrantiport many ion and sucrose rarr inward
Numerous transport proteins enable plant vacuoles to accumulate metabolites and ions
More positive
Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Trans-epithelial transport
Import of molecules on the lumen side of intestinal epithelial cells and their export on the blood facing sides
Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Transcellular transport of glucose from the intestinal lumen into the blood
1
2
3
4 Cholera toxin activated Cl- secretion
Basolateral Na+ K+ ATPasegenerates Na+ gradient thatdrives the Symporter
Glucose + normal saline rarr co-transport for energy supply
[high]
microvilli
Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Mechanism of Action of Cholera Toxin
Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

Acidification of the stomach lumen by parietal cells in the gastric lining
Parietal cells acidify the stomach contents while maintaining a neutral cytolic pH
P-class1
2
3
bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

bull END
bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-

bull END
- 投影片編號 1
- 投影片編號 2
- 投影片編號 3
- 投影片編號 4
- 投影片編號 5
- 投影片編號 6
- 投影片編號 7
- 投影片編號 8
- 投影片編號 9
- 投影片編號 10
- 投影片編號 11
- 投影片編號 12
- 投影片編號 13
- 投影片編號 14
- 投影片編號 15
- 投影片編號 16
- Free Diffusion
- 投影片編號 18
- 投影片編號 19
- 投影片編號 20
- 投影片編號 21
- 投影片編號 22
- 投影片編號 23
- 投影片編號 24
- 投影片編號 25
- 投影片編號 26
- 投影片編號 27
- 投影片編號 28
- 投影片編號 29
- 投影片編號 30
- 投影片編號 31
- 投影片編號 32
- 投影片編號 33
- 投影片編號 34
- 投影片編號 35
- 投影片編號 36
- 投影片編號 37
- 投影片編號 38
- 投影片編號 39
- 投影片編號 40
- 投影片編號 41
- 投影片編號 42
- 投影片編號 43
- 投影片編號 44
- 投影片編號 45
- 投影片編號 46
- 投影片編號 47
- 投影片編號 48
- Na+- K+ Pump on the Plasma Membrane
- Na+- K+ Pump on the Plasma Membrane
- 投影片編號 51
- ABC Transporters
- 投影片編號 53
- 投影片編號 54
- 投影片編號 55
- 投影片編號 56
- 投影片編號 57
- 投影片編號 58
- 投影片編號 59
- 投影片編號 60
- Action of the Multi-drug Resistant Transport Protein
- 投影片編號 62
- Proposed mechanisms of action for the MDR1 protein
- Flippases
- 投影片編號 65
- 投影片編號 66
- 投影片編號 67
- 投影片編號 68
- 投影片編號 69
- 投影片編號 70
- 投影片編號 71
- 投影片編號 72
- 投影片編號 73
- 投影片編號 74
- 投影片編號 75
- 投影片編號 76
- 投影片編號 77
- 投影片編號 78
- 投影片編號 79
- Voltage-gated K+ channels have four subunits each containing six transmembrane helices
- Ion channels are selective pores in the membrane
- 投影片編號 82
- 投影片編號 83
- 投影片編號 84
- 投影片編號 85
- 投影片編號 86
- 投影片編號 87
- 投影片編號 88
- 投影片編號 89
- 投影片編號 90
- 投影片編號 91
- 投影片編號 92
- 投影片編號 93
- Coupling of Active Transport to Ion Gradients without energy against gradient
- 投影片編號 95
- 投影片編號 96
- 投影片編號 97
- 投影片編號 98
- 投影片編號 99
- 投影片編號 100
- 投影片編號 101
- 投影片編號 102
- 投影片編號 103
- 投影片編號 104
- 投影片編號 105
- A putative cation exchange protein plays a key role in evolution of human skin pigmentation
- 投影片編號 107
- 投影片編號 108
- 投影片編號 109
- 投影片編號 110
- 投影片編號 111
- 投影片編號 112
- 投影片編號 113
-
Related Documents











