Lodish • Berk • Kaiser • Krieger • scott • Bretscher • Ploegh • Matsudaira MOLECULAR CELL BIOLOGY SEVENTH EDITION CHAPTER 21 Stem Cells, Cell Asymmetry, and Cell Death Copyright © 2013 by W. H. Freeman and Company

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Lodish • Berk • Kaiser • Krieger • scott • Bretscher • Ploegh • Matsudaira
MOLECULAR CELL BIOLOGY SEVENTH EDITION
CHAPTER 21 Stem Cells, Cell Asymmetry,
and Cell Death
Copyr ight © 2013 by W. H. Freeman and Company


Cell death Programmed cell death is a counterintuitive, but essential, cell fate. During embryogensis, death of specific cells keeps our hands from being webbed, our embryonic tails from persisting, and our brain from being filled with useless nerve connections. In fact, the majority of cells generated during brain development subsequently die. 1. The demise of cells by programmed cell death is marked by a well-defined sequence of morphological
changes, collectively referred to as apoptosis, a Greek word that means “dropping off” or “falling off”, as leaves from a tree. A cell that undergoes apoptosis dies neatly, without damaging its neighbors. The cell shrinks and condenses. The cytoskeleton collapses, the nuclear envelope disassembles, and the nuclear DNA breaks up into fragments. Most importantly, the cell surface is altered, displaying properties that cause the dying cell to be rapidly phagocytosed, either by a neighboring cell or by a macrophage (a specialized phagocytic cell), before any leakage of its contents occurs. This not only avoids the damaging consequences of cell necrosis but also allows the organic components of the dead cell to be recycled by the cell that ingests it (http://www.ncbi.nlm.nih.gov/books/NBK26873/).
2. When cells are placed under stress such as conditions of starvation, they have the capacity to recycle macromolecules for use as nutrients in a process of lysosomal degradation known as autophagy (“eating oneself”). The autophagic pathway involves the formation of a flattened double-membrane cup-shaped structure that envelops a region of the cytosol or an entire organelle (e.g., mitochondrion), forming an autophagosome, or autophagic vesicle.
3. In contrast to apoptosis, cells that die in response to tissue damage exhibit very different morphological changes, referred to as necrosis. Typically, cells that undergo this process swell and burst, releasing their intracellular contents, which can damage surrounding cells and frequently causes inflammation. Apoptosis or Type I cell-death, and Autophagy or Type II cell-death are both forms of programmed cell death, while necrosis is a non-physiological process that occurs as a result of infection or injury. Necrosis is cell death caused by external factors such as trauma or infection, and occurs in several different forms. Recently a form of programmed necrosis, called necroptosis, has been recognized as an alternate form of programmed cell death. It is hypothesized that necroptosis can serve as a cell-death backup to apoptosis when the apoptosis signaling is blocked by endogenous or exogenous factors such as viruses or mutations (Wikipedia).
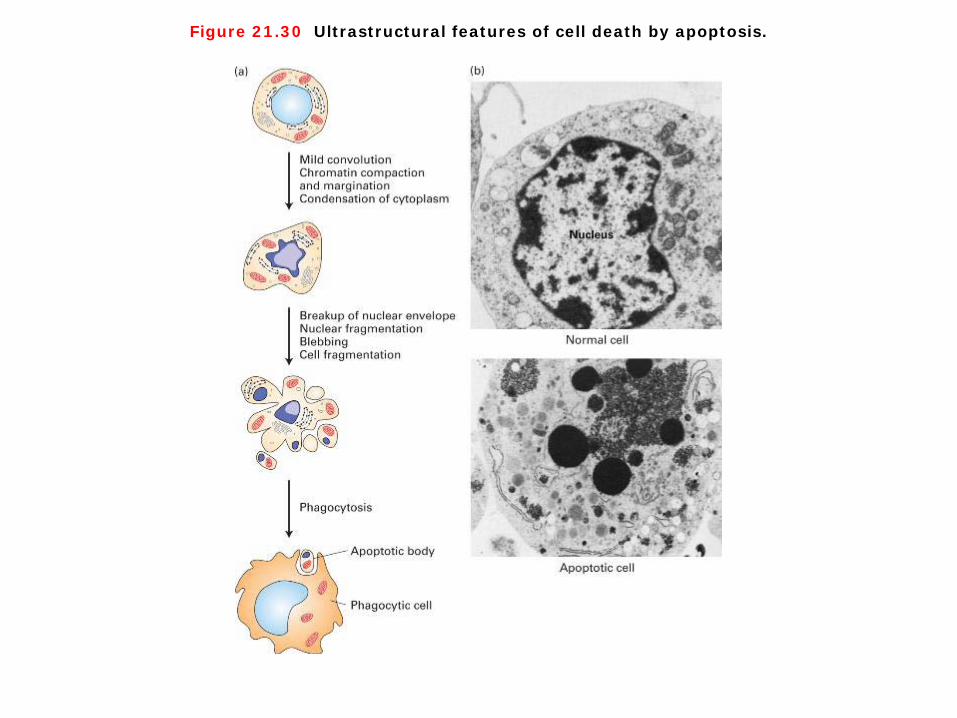
Figure 21.30 Ultrastructural features of cell death by apoptosis.

Figure 18-2 Molecular Biology of the Cell (© Garland Science 2008)

Cell death by apoptosis 1. Programmed cell death occurs through apoptosis. Dying cells shrink, condense, and then fragment, releasing small membrane bound apoptotic bodies, which generally are then engulfed by other cells (Figure 21-30). In dying cells, nuclei condense and the DNA is fragmented. Importantly, the intracellular constituents are not released into the extracellular milieu, where they might have deleterious effects on neighboring cells. The stereotypical changes that occur in cells during apoptosis, such as condensation of the nucleus and engulfment by surrounding cells, suggested to early scientists that this type of cell death was under the control of a strict program. This program is critical during both embryonic and adult life to maintain normal cell number and composition. 2. The amount of apoptosis that occurs in developing and adult animal tissues can be astonishing. In the developing vertebrate nervous system, for example, up to half or more of the nerve cells normally die soon after they are formed. In a healthy adult human, billions of cells die in the bone marrow and intestine every hour. It seems remarkably wasteful for so many cells to die, especially as the vast majority are perfectly healthy at the time they kill themselves. What purposes does this massive cell death serve? In some cases, the answers are clear. Mouse paws, for example, are sculpted by cell death during embryonic development: they start out as spadelike structures, and the individual digits separate only as the cells between them die (Figure 18-2 Molecular Biology of the Cell). In other cases, cells die when the structure they form is no longer needed. When a tadpole changes into a frog, the cells in the tail die, and the tail, which is not needed in the frog, disappears. In many other cases, cell death helps regulate cell numbers. In the developing nervous system, for example, cell death adjusts the number of nerve cells to match the number of target cells that require innervation. In all these cases, the cells die by apoptosis. (http://www.ncbi.nlm.nih.gov/books/NBK26873/)

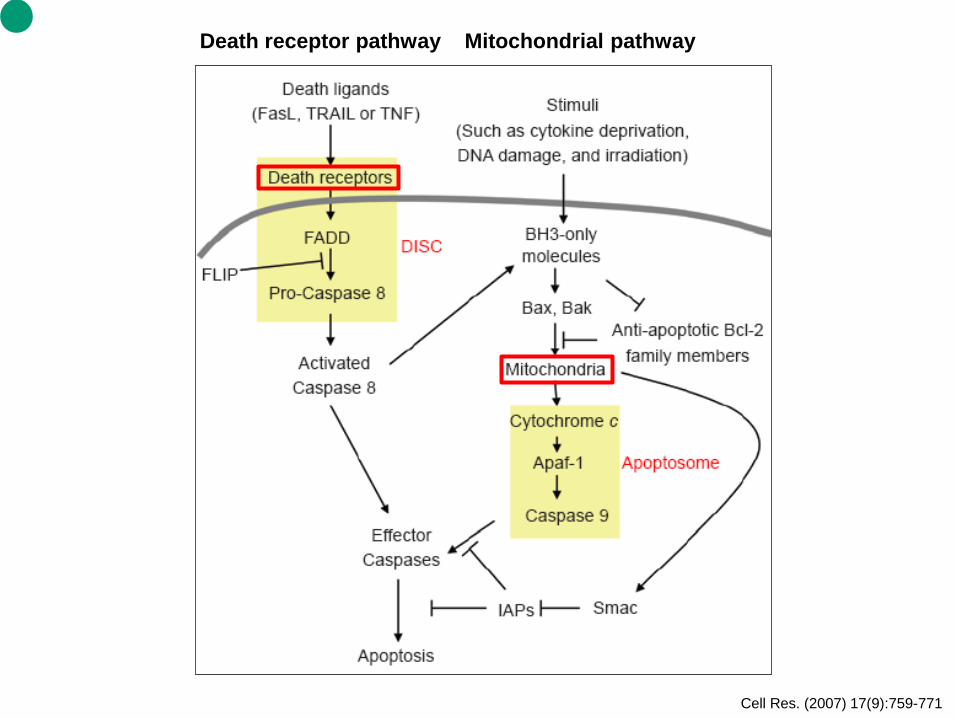
Cell Res. (2007) 17(9):759-771
Death receptor pathway Mitochondrial pathway

Two major pathways leading to apoptosis There are two major pathways leading to apoptosis in the mammalian system: an extrinsic pathway initiated by death receptors and an intrinsic pathway that occurs through the mitochondria (Figure 1). The extrinsic pathway depends on binding of appropriate exogenous mediators to death receptors at the cell surface. In contrast, the intrinsic pathway responds to signals from within the cell, such as damages caused by radiation and various chemotherapeutic agents, to induce apoptotic signaling via the release of mitochondrial factors (Cell Res. (2007) 17(9):759-771).
1. Extrinsic apoptosis signaling pathways. The extrinsic pathway of apoptosis signaling is initiated
when death receptors at the cell surface encounter specific cognate “death ligands,” inducing a conformational change that is transmitted through the cell membrane. These receptors can activate caspases within seconds of ligand binding and lead to apoptotic cell death in a matter of hours.
2. Intrinsic signaling pathways. The mitochondrial pathway of apoptosis in mammalian cells centers on a key event: mitochondrial outer membrane permeabilization (MOMP), considered the point-of-noreturn in apoptosis induction. Release of certain proteins from the mitochondrial intermembrane space due to MOMP triggers a cascade of caspase activation that results in irreversible events culminating in apoptosis.

Figure 21.33 Evolutionary conservation of apoptosis pathways.

Evolutionary conservation of apoptosis pathways The first mammalian apoptotic gene to be cloned, bcl-2, was isolated from human follicular lymphomas. A mutant form of this gene was created in lymphoma cells; a chromosomal rearrangement joined the protein-coding region of the bcl-2 gene to an immunoglobulin gene enhancer. The combination results in overproduction of the Bcl-2 protein that keeps these cancer cells alive when otherwise they would become programmed to die. 1. The human Bcl-2 protein and worm CED-9 protein are homologous; even though the two proteins are
only 23 percent identical in sequence, a bcl-2 transgene can block the extensive cell death found in ced-9 mutant worms. Thus both proteins act as regulators that suppress the apoptotic pathway (Figure 21-33). In addition, both proteins contain a single transmembrane domain and are localized mainly to the outer mitochondrial membrane, where they serve as sensors that control the apoptotic pathway in response to external stimuli. As we discuss next, other regulators promote apoptosis.
2. In the worm apoptotic pathway, CED-3 (caspase-9 in mammals) is required to destroy cell components during apoptosis. CED-4 (Apaf-1) is a protease-activating factor that cause autocleavage of (and by) the CED-3 precursor protein, creating an active CED-3 (caspase-9) protease that initiates cell death (Figures 21-33). Cell death does not occur in ced-3 and ced-4 single mutants or in ced-9/ced-3 double mutants, whereas all cells die during embryonic life in ced-9 mutants, so the adult form never develops. These genetic studies indicate that CED-3 and CED-4 are “killer” proteins required for cell death, that CED-9 (Bcl-2) suppresses apoptosis, and that the apoptotic pathway can be activated in all cells. Moreover, the absence of cell death in ced-9/ced-3 double mutants suggests that CED-9 acts “upstream” of CED-3 to suppress the apoptotic pathway.
3. The mechanism by which CED-9 (Bcl-2) controls CED-3 (caspase-9) is now known. CED-9 protein, which is normally tethered to the outside of mitochondria, forms a complex with an asymmetric CED-4 (Apaf-1) dimer, thereby preventing activation of CED-3 by CED-4. As a result, the cell survives. This mechanism fits with the genetics, which shows that the absence of CED-9 has no effect if CED-3 is also missing (ced-3/ced-9 double mutants have no cell death). The three-dimensional structure of the trimeric CED-4/CED-9 complex reveals a huge contact surface between each of the two CED-4 molecules and the single CED-9 molecule; the large contact surface makes the association highly specific, but in such a way that the dissociation of the complex can be regulated.
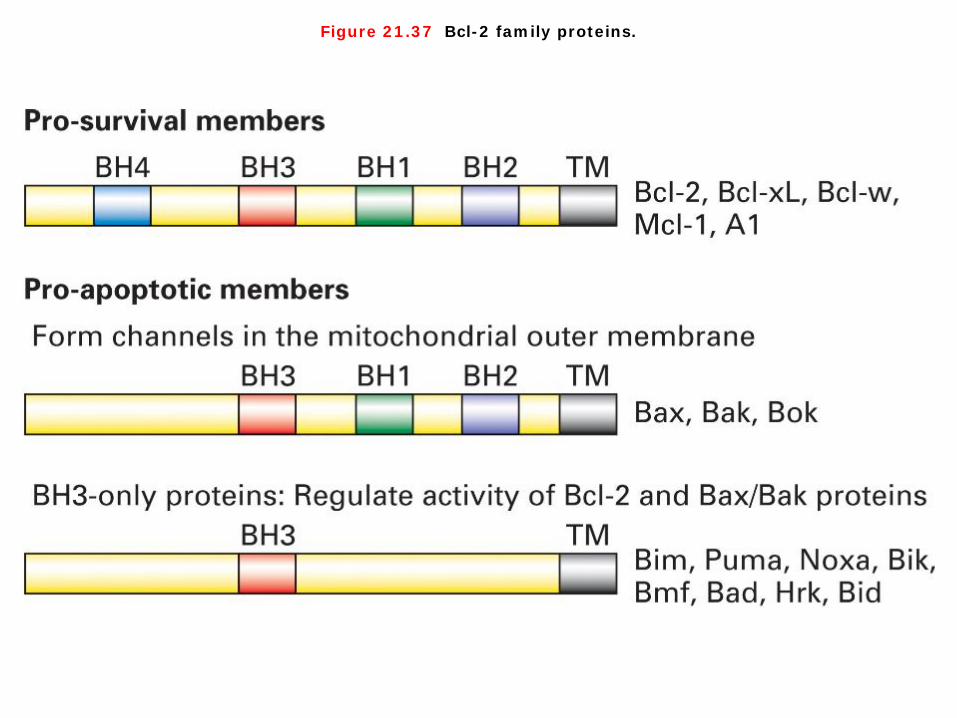
Figure 21.37 Bcl-2 family proteins.

Bcl-2 family proteins 1. In order to explain how Bcl-2 carries out this function and how Bcl-2 activity is regulated by trophic factors and by many environmental stimuli, we need to introduce several other important members of the Bcl-2 family of proteins. All members of the Bcl-2 family share a close homology in up to four characteristic regions termed the Bcl-2 homology domains (BH1-4 domains; Figure 21-37). Each protein has either a pro-survival or a pro-apoptotic function. Many members of this family are single-pass transmembrane proteins and all participate in oligomeric interactions. 2. In vertebrate cells, Bax or Bak is required for mitochondrial damage and induction of apoptosis. These two similar pro-apoptotic proteins contain several of the BH1-4 domains (Figure 21-37) and have a three-dimensional structure very similar to that of the pro-survival members of the family. As evidence for their role in promoting apoptosis, most mice lacking both Bax and Bak die in utero. Those that survive show significant developmental defects, including the persistence of interdigital webs and accumulation of extra cells in the central nervous and hematopoietic systems. Cells isolated from these mice are resistant to virtually all apoptotic stimuli. Conversely, overproduction of Bax in cultured cells induces death. Bax and Bak reside in the outer mitochondrial membrane, normally tightly bound to Bcl-2. When released from Bcl-2 either by being present in excess, by being displaced by binding of certain BH3-only proteins to Bcl-2, or by direct binding to other BH3-only proteins-Bax and Bak form oligomers that generate pores in the outer mitochondrial membrane. This allow release into the cytosol of mitochondrial proteins such as cytochrome c that, in normal healthy cells, are localized in the space between the inner and outer mitochondrial membrane.


Figure 21.38 Integration of multiple signaling pathways in vertebrate cells that regulate mitochondrial outer membrane permeability and apoptosis.

Intrinsic signaling pathways 1. We saw earlier that neurotrophins such as nerve growth factor protect neurons from cell death; this is mediated by the BH3-only protein called Bad. ① In the absence of trophic factors, Bad is unphosphorylated and binds to Bcl-2 or the closely
related anti-apoptotic protein Bcl-xL at the mitochondrial membrane (Figure 21-38). This inhibits the ability of Bcl-2 and Bcl-xL to bind Bax and Bak, thereby allowing Bak and Bax channels to form and promoting cell death. Phosphorylated Bad, however, cannot bind to Bcl-2/Bcl-xL and is found in the cytosol complexed to the phosphoserine-binding protein 14-3-3.
② A number of trophic factors including NGF have been shown to trigger the PI-3 kinase signaling pathway, leading to activation of a downstream kinase called PKB. Activated PKB phosphorylates Bad at sites known to inhibit its pro-apoptotic activity. These findings support the mechanism for the survival action of trophic factors depicted in Figure 21-38. In other cell types, different trophic factors may promote cell survival through post-translational modification of other components of the cell-death machinery.
2. DNA damage or ultraviolet irradiation leads to induction of the BH3-only Puma protein. Puma binds to Bax and Bak, allowing them to form oligomeric channels. 3. Removal of a cell from its usbstratum disrupts integrin signaling, leading to release of the BH3-only Bim protein from the cytoskeleton. Bim also binds to Bax and Bak to promote channel formation.
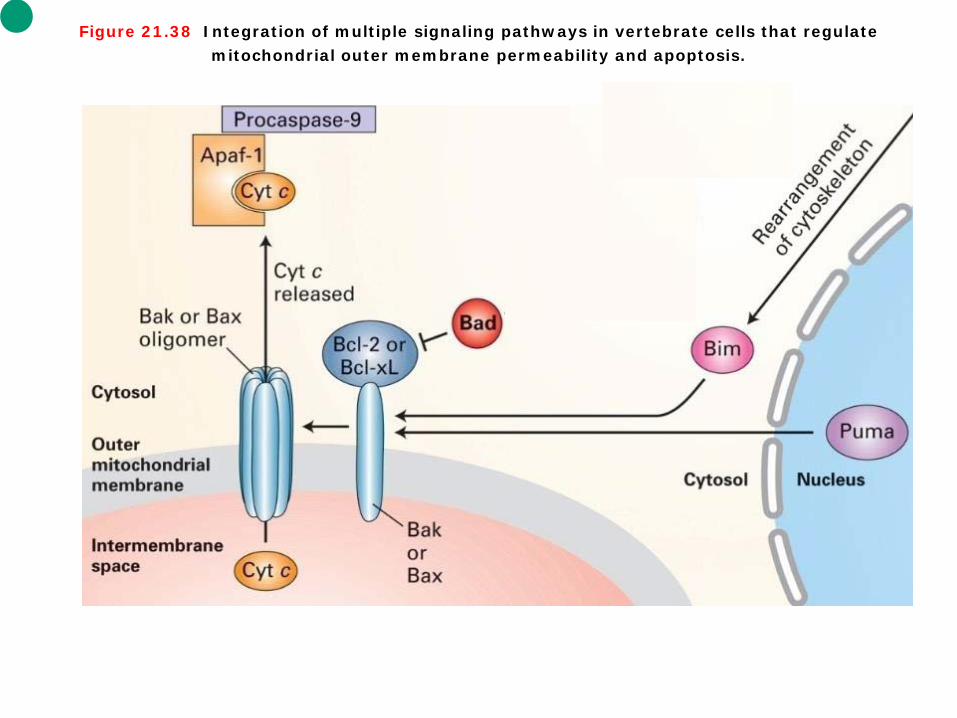
Figure 21.38 Integration of multiple signaling pathways in vertebrate cells that regulate mitochondrial outer membrane permeability and apoptosis.

Intrinsic signaling pathways 1. Some Bcl-2 family members preserve or disrupt the integrity of mitochondria, thereby controlling release of mitochondrial proteins such as cytochrome c. In normal healthy cells, cytochrome c is localized between the inner and outer mitochondrial membrane, but in cells undergoing apoptosis, cytochrome c is released into the cytosol. A variety of death-inducing stimuli cause Bax monomers to move from the cytosol to the outer mitochondrial membrane where they oligomerize. The principal way cytochrome c in the cytosol activates apoptosis is by binding Apaf-1, the mammalian homolog of CED-4. After binding cytochrome c, Apaf-1 cleaves its bound dATP into dADP and undergoes a dramatic assembly process into a disk-shaped heptamer, a 1.4-megadalton wheel of death called the apoptosome. The apoptosome seves as an activation machine for the initiator caspase, caspase-9, which is monomeric in the inactive signals yet should not be activatable in an irreversible manner because accidental activation would lead to an undesirable snowball effect and rapid cell death. Significantly, caspase-9 does not require cleavage to become activated, but rather it is activated by dimerization following binding to the apoptosome. Caspase-9 then cleaves multiple molecules of effector caspases, such as caspase-3, leading to destruction of cell proteins. 2. In mammals and flies, but not worms, apoptosis is regulated by several other proteins (see Figure 21-33). A family of inhibitor of apoptosis proteins (IAPs) provides another way to restrain both initiator and effector caspases. IAPs have one or more zinc-binding domains that can bind directly to caspases and inhibit their protease activity. The inhibition of caspases by IAPs, however, creates a problem when a cell needs to undergo apoptosis. Mitochondria enter the picture once again since they are the source of a family of proteins, called SMAC/DIABLOs, which inhibit IAPs. Assembly of Bax/Bak channels leads to release of SMAC/DIABLOs from mitochondria. SMAC/DIABLOs then binds to IAPs in the cytosol, thereby blocking the IAPs from binding to caspases. By relieving IAP-mediated inhibition, SMAC/DIABLOs promote caspase activity and cell death.

Nat Rev Cancer (2007) 7(11):819-822

p53 in regulating cell proliferation and cell death The p53 protein is a sensor essential for the checkpoint control that arrests cells with damaged DNA in G1. Although p53 has several functions, its ability to activate transcription of certain genes is most relevant to its tumor-suppressing function. The activity of p53 normally is kept low by a protein called Mdm2. When Mdm2 is bound to p53, it inhibits the transcription-activating ability of p53 and catalyzes the addition of ubiquitin molecules, thus targeting p53 for proteasomal degradation. Phosphorylation of p53 by ATM displaces bound Mdm2 from p53, thereby stabilizing it. The activity of p53 is not limited to inducing cell-cycle arrest. In addition, this multipurpose tumor suppressor stimulates production of pro-apoptotic proteins and DNA-repair enzymes. Among the genes activated by p53 are several encoding pro-apoptotic proteins such as Bax. When most cells suffer extensive DNA damage, the p53-induced expression of proapoptotic proteins leads to their quick demise.

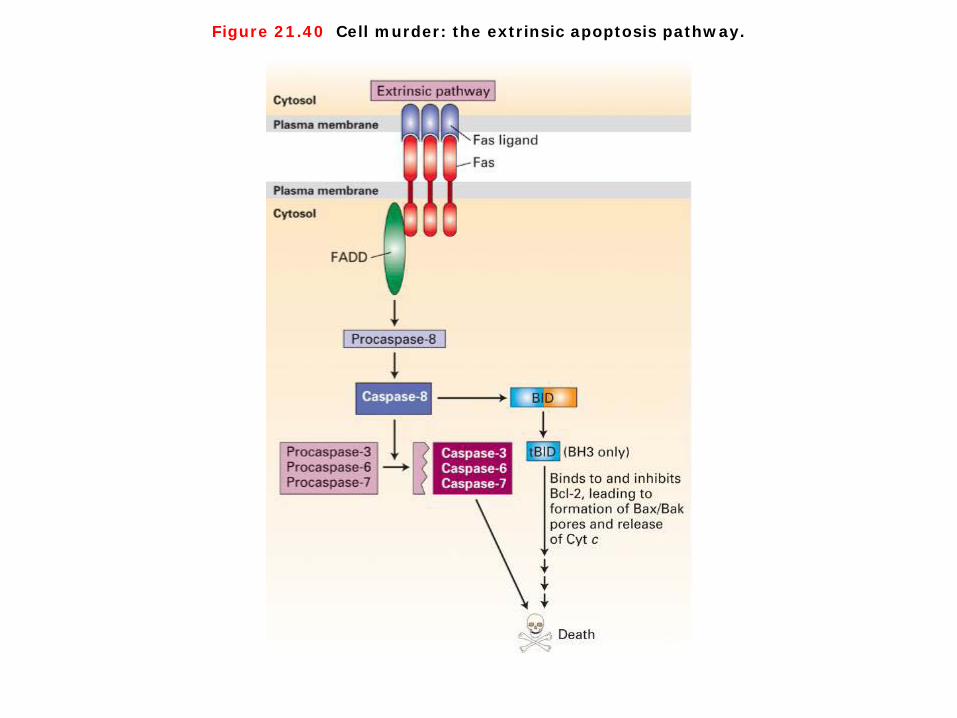
Figure 21.40 Cell murder: the extrinsic apoptosis pathway.

Cell murder: the extrinsic apoptosis pathway 1. Although cell death can arise as a default in the absence of survival factors, apoptosis can also be stimulated by positively acting “death” signals. For instance, tumor necrosis factor (TNF), which is released by macrophages, triggers the cell death and tissue destruction seen in certain chronic inflammatory diseases. Another important death-inducing signal, the Fas ligand, is a cell-surface protein expressed by activated natural killer cells and cytotoxic T lymphocytes. This signal can trigger death of virus-infected cells, some tumor cells, and foreign graft cells. 2. Both TNF and Fas ligand act through cell-surface “death” receptors that have a single transmembrane domain and are activated when ligand binding brings three receptor molecules into close proximity. The trimeric receptor complex attracts a protein called FADD (Fas-associated death domain), which serves as an adapter to recruit and in some way activate caspase-8, an initiator caspase, in cells receiving a death signal. The death domain found in FADD is a sequence that is present in a number of proteins involved in apoptosis. Once activated, caspase-8 activates other caspases and the amplification cascade begins. To test the ability of the Fas receptor to induce cell death, researchers incubated cells with antibodies against the receptor. These antibodies, which bind and cross-link Fas receptors, were found to stimulate cell death, indicating that activation of the Fas receptor is sufficient to trigger apoptosis.


Nat Rev Mol Cell Biol. (2007) 8(9):741-752
Autophagy

Autophagy Autophagy starts with the stepwise engulfment of cytoplasmic material (cytosol and/or organelles) by the phagophore (also called isolation membrane), which sequesters material in double-membraned vesicles named autophagosomes (also called autophagic vacuoles). 1. In many cellular settings, the first regulatory process (figure, step 1) involves the de-repression of the
mTOR Ser/Thr kinase, which inhibits autophagy by phosphorylating autophagy protein-13 (Atg13). This phosphorylation leads to the dissociation of Atg13 from a protein complex that contains Atg1 kinase and Atg17, and thus attenuates the Atg1 kinase activity. When mTOR is inhibited, re-association of dephosphorylated Atg13 with Atg1 stimulates its catalytic activity and induces autophagy.
2. Among the initial steps of vesicle nucleation is the activation of mammalian Vps34, a class III phosphatidylinositol 3-kinase (PI3K), to generate phosphatidylinositol-3-phosphate (PtdIns3P) (step 2). Vps34 activation depends on the formation of a multiprotein complex in which beclin-1 (Becn1; the mammalian orthologue of Atg6), UVRAG (UV irradiation resistance-associated tumour suppressor gene) and a myristylated kinase (Vps15, or p150 in humans) participate.
3. Two ubiquitin-like conjugation systems are part of the vesicle elongation process (step 3). One pathway involves the covalent conjugation of Atg12 to Atg5, with the help of the E1-like enzyme Atg7 and the E2-like enzyme Atg10. The second pathway involves the conjugation of phosphatidylethanolamine (PE) to LC3/Atg8 (LC3 is one of the mammalian homologues of Atg8) by the sequential action of the protease Atg4, the E1-like enzyme Atg7 and the E2-like enzyme Atg3. Lipid conjugation leads to the conversion of the soluble form of LC3 (named LC3-I) to the autophagic-vesicle-associated form (LC3-II). LC3-II is used as a marker of autophagy because its lipidation and specific recruitment to autophagosomes provides a shift from diffuse to punctate staining of the protein and increases its electrophoretic mobility on gels compared with LC3-I.
4. The mechanism of retrieval in which the Atg9 complex participates is poorly studied (step 4). 5. Autophagosomes undergo maturation by fusion with lysosomes to create autolysosomes (steps 5
and 6). In the autolysosomes, the inner membrane as well as the luminal content of the autophagic vacuoles is degraded by lysosomal enzymes that act optimally within this acidic compartment. Bcl2 and Bcl-XL are regulators of beclin-1.
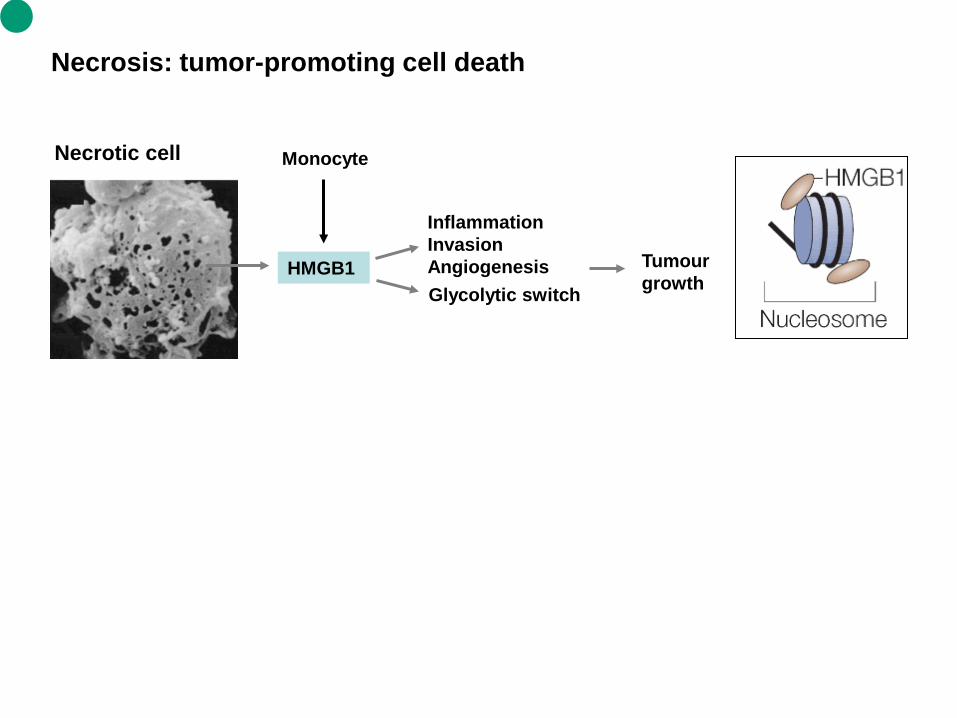
Necrosis: tumor-promoting cell death
HMGB1
Inflammation Invasion Angiogenesis Tumour
growth
Necrotic cell Monocyte
Glycolytic switch

Necrosis 1. Necrosis is caused by factors external to the cell or tissue, such as infection, toxins, or trauma which result in the unregulated digestion of cell components. Cellular death due to necrosis does not follow the apoptotic signal transduction pathway, but rather various receptors are activated, and result in the loss of cell membrane integrity and an uncontrolled release of products of cell death into the extracellular space (Wikipedia). Cells that die due to metabolic, genotoxic or anoxic stress have limited ways of initiating the apoptotic cascade because these important pathways are blocked by endogenous inhibitors, including high concentrations of B-cell lymphoma 2 (BCL-2) and BCL-XL, or members of the IAP family of inhibitors of apoptosis. The apoptotic to necrotic conversion that is associated with unscheduled cell death and the subsequent release of necrotic mediators is supported by clinical observations in which the presence of necrosis is almost always deemed a poor prognostic finding. 2. High-mobility group box 1 protein (HMGB1) is an evolutionarily ancient factor, pre-dating the split in the animal and plant kingdoms. HMGB1 is associated with the regulation and 'bending' of DNA to allow access to various transcriptional complexes. Its release from necrotic cells, but not apoptotic cells, promotes the recruitment of inflammatory cells, thereby indicating that HMGB1 lies at the 'crossroads' of necrosis and apoptosis. During apoptosis, HMGB1 seems to be sequestered and ultimately degraded by the programmed proteolytic process that is unleashed in a coordinate manner in the cell. Whereas apoptotic cell death is the usual form of 'quiet' cell death and is not itself associated with inflammation, necrotic cell death alerts both inflammatory and reparative mechanisms that are important for angiogenesis, stromagenesis and the promotion of epithelial proliferation, all of which are required for tumour growth.



Figure 21.1 Overview of the birth, lineage, and death of cells.

Figure 21.34 Activation of CED-3 protease in C. elegans.
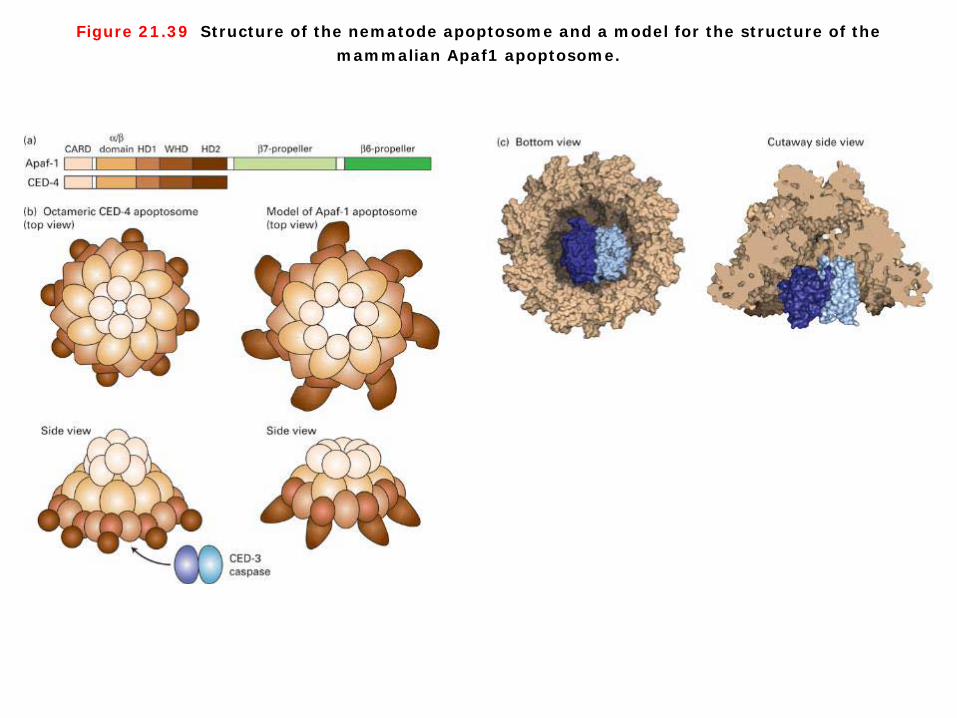
Figure 21.39 Structure of the nematode apoptosome and a model for the structure of the mammalian Apaf1 apoptosome.
Related Documents