The Biochemical Basis of Life History Adaptation: Molecular and Enzymological Causes of NADP 1 -Isocitrate Dehydrogenase Activity Differences Between Morphs of Gryllus firmus That Differ in Lipid Biosynthesis and Life History Rudolf J. Schilder, Anthony J. Zera,*à Christine Black,à Mallary Hoidal,à and Cody Wehrkampà School of Biological Sciences, University of Nebraska Present address: Department of Cellular and Molecular Physiology, Penn State College of Medicine àPresent address: School of Biological Sciences, University of Nebraska *Corresponding author: E-mail: [email protected]. Associate editor: John H. McDonald Abstract Although whole-organism aspects of life-history physiology are well studied and molecular information (e.g., transcript abundance) on life-history variation is accumulating rapidly, much less information is available on the biochemical (enzymological) basis of life-history adaptation. The present study investigated the biochemical and molecular causes of specific activity differences of the lipogenic enzyme, NADP þ -isocitrate dehydrogenase, between genetic lines of the wing- polymorphic cricket, Gryllus firmus, which differ in lipid biosynthesis and life history. With one exception, variation among 21 Nadp þ -Idh genomic sequences, which spanned the entire coding sequence of the gene, was restricted to a few synonymous substitutions within and among replicate flight-capable or flightless lines. No NADP þ -IDH electromorph variation was observed among individuals within or among lines as determined by polyacrylamide gel electrophoresis. Nor did any NADP þ -IDH kinetic or stability parameter, such as K M for substrate or cofactor, k cat , or thermal denaturation, differ between flight-capable and flightless lines. By contrast, line differences in NADP þ -IDH–specific activity strongly covaried with transcript abundance and enzyme protein concentration. These results demonstrate that NADP þ -IDH–specific activity differences between artificially selected lines of G. firmus are due primarily, if not exclusively, to genetic variation in regulators of NADP þ -IDH gene expression, with no observed contribution from altered catalytic efficiency of the enzyme due to changes in amino acid sequence or posttranslational modification. Kinetic analyses indicate that in vitro differences in enzyme-specific activity between flight-capable and flightless lines likely occur in vivo. This study constitutes the most comprehensive analysis to date of the biochemical and molecular causes of naturally occurring genetic variation in enzyme activity that covaries strongly with life history. Key words: life history, wing polymorphism, NADP þ -isocitrate dehydrogenase, biochemical adaptation, enzyme kinetics, transcript abundance, gene regulation. Introduction The extent to which evolutionary modification of metab- olism contributes to life-history adaptation has been a long-standing topic of research in life-history evolution (Townsend and Calow 1981; Rose and Bradley 1998; Zera and Harshman 2001, 2009, 2011). At the level of whole- organism physiology, numerous studies have identified enhanced accumulation of specific macromolecules (e.g., somatic lipid) important for particular life-history traits (e.g., dispersal, extended longevity). A few studies have documented changes in flux through pathways of interme- diary metabolism responsible for adaptive accumulation of these metabolic end products (O#Brien et al. 2002; Zhao and Zera 2002; Zera and Zhao 2006; Zera 2011). At the other extreme, an increasing number of studies have re- ported molecular correlates (i.e., altered gene expression, transcript abundance) of life-history adaptation (Arking et al. 2000; Pelcher et al. 2005; St-Cyr et al. 2008; Wheat et al. 2011). However, what is sorely lacking is information on biochemical (e.g., enzymological) aspects of life-history adaptation. Enzymological data are essential to identify the functional significance of altered sequence or expres- sion of genes encoding metabolic enzymes and to identify the causes of altered pathway function. In short, biochem- ical information provides the functional link between molecular and whole-organism physiological aspects of life-history variation. Genetic variation in enzyme activity can result from a va- riety of causes the most important of which are 1) altered gene and amino acid sequence resulting in changes in cat- alytic efficiency or enzyme stability; 2) changes in gene ex- pression due to altered rates of transcription, transcript © The Author 2011. Published by Oxford University Press on behalf of the Society for Molecular Biology and Evolution. All rights reserved. For permissions, please e-mail: [email protected] Mol. Biol. Evol. 28(12):3381–3393. 2011 doi:10.1093/molbev/msr171 Advance Access publication June 24, 2011 3381 Research article Downloaded from https://academic.oup.com/mbe/article/28/12/3381/1031099 by guest on 18 March 2022

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

The Biochemical Basis of Life History Adaptation: Molecularand Enzymological Causes of NADP1-IsocitrateDehydrogenase Activity Differences Between Morphs ofGryllus firmus That Differ in Lipid Biosynthesis and LifeHistory
Rudolf J. Schilder,� Anthony J. Zera,*� Christine Black,� Mallary Hoidal,� and Cody Wehrkamp�School of Biological Sciences, University of Nebraska
�Present address: Department of Cellular and Molecular Physiology, Penn State College of Medicine
�Present address: School of Biological Sciences, University of Nebraska
*Corresponding author: E-mail: [email protected].
Associate editor: John H. McDonald
Abstract
Although whole-organism aspects of life-history physiology are well studied and molecular information (e.g., transcriptabundance) on life-history variation is accumulating rapidly, much less information is available on the biochemical(enzymological) basis of life-history adaptation. The present study investigated the biochemical and molecular causes ofspecific activity differences of the lipogenic enzyme, NADPþ-isocitrate dehydrogenase, between genetic lines of the wing-polymorphic cricket, Gryllus firmus, which differ in lipid biosynthesis and life history. With one exception, variation among21 Nadpþ-Idh genomic sequences, which spanned the entire coding sequence of the gene, was restricted to a fewsynonymous substitutions within and among replicate flight-capable or flightless lines. No NADPþ-IDH electromorphvariation was observed among individuals within or among lines as determined by polyacrylamide gel electrophoresis. Nordid any NADPþ-IDH kinetic or stability parameter, such as KM for substrate or cofactor, kcat, or thermal denaturation, differbetween flight-capable and flightless lines. By contrast, line differences in NADPþ-IDH–specific activity strongly covariedwith transcript abundance and enzyme protein concentration. These results demonstrate that NADPþ-IDH–specificactivity differences between artificially selected lines of G. firmus are due primarily, if not exclusively, to genetic variation inregulators of NADPþ-IDH gene expression, with no observed contribution from altered catalytic efficiency of the enzymedue to changes in amino acid sequence or posttranslational modification. Kinetic analyses indicate that in vitro differencesin enzyme-specific activity between flight-capable and flightless lines likely occur in vivo. This study constitutes the mostcomprehensive analysis to date of the biochemical and molecular causes of naturally occurring genetic variation in enzymeactivity that covaries strongly with life history.
Key words: life history, wing polymorphism, NADPþ-isocitrate dehydrogenase, biochemical adaptation, enzyme kinetics,transcript abundance, gene regulation.
IntroductionThe extent to which evolutionary modification of metab-olism contributes to life-history adaptation has been along-standing topic of research in life-history evolution(Townsend and Calow 1981; Rose and Bradley 1998; Zeraand Harshman 2001, 2009, 2011). At the level of whole-organism physiology, numerous studies have identifiedenhanced accumulation of specific macromolecules (e.g.,somatic lipid) important for particular life-history traits(e.g., dispersal, extended longevity). A few studies havedocumented changes in flux through pathways of interme-diary metabolism responsible for adaptive accumulation ofthese metabolic end products (O#Brien et al. 2002; Zhaoand Zera 2002; Zera and Zhao 2006; Zera 2011). At theother extreme, an increasing number of studies have re-ported molecular correlates (i.e., altered gene expression,
transcript abundance) of life-history adaptation (Arkinget al. 2000; Pelcher et al. 2005; St-Cyr et al. 2008; Wheatet al. 2011). However, what is sorely lacking is informationon biochemical (e.g., enzymological) aspects of life-historyadaptation. Enzymological data are essential to identifythe functional significance of altered sequence or expres-sion of genes encoding metabolic enzymes and to identifythe causes of altered pathway function. In short, biochem-ical information provides the functional link betweenmolecular and whole-organism physiological aspects oflife-history variation.
Genetic variation in enzyme activity can result from a va-riety of causes the most important of which are 1) alteredgene and amino acid sequence resulting in changes in cat-alytic efficiency or enzyme stability; 2) changes in gene ex-pression due to altered rates of transcription, transcript
© The Author 2011. Published by Oxford University Press on behalf of the Society for Molecular Biology and Evolution. All rights reserved. For permissions, pleasee-mail: [email protected]
Mol. Biol. Evol. 28(12):3381–3393. 2011 doi:10.1093/molbev/msr171 Advance Access publication June 24, 2011 3381
Research
articleD
ownloaded from
https://academic.oup.com
/mbe/article/28/12/3381/1031099 by guest on 18 M
arch 2022

stability, or translation efficiency giving rise to variation inenzyme protein concentration; or 3) various posttransla-tional modifications (PTMs) such as phosphorylation thatalter catalytic efficiency or enzyme stability between enzymeswithout altering amino acid sequence (fig. 1; Eanes 1999;Storz and Zera 2011). Importantly, these different sourcesof variation can result from alterations in very different clas-ses of genes. Although change in catalytic efficiency due toaltered amino acid sequence of the enzyme must result fromvariation in the structural gene that encodes the enzyme,change in enzyme concentration can have a variety of causes:altered regulatory regions that can be 1) tightly linked to butoutside of the structural locus (e.g., cis-acting regulators), 2)within introns of the coding sequence of the structural locus,or 3) in trans-acting regions unlinked to the structural locus,as well as due to changes in the amino acid sequence of theenzyme itself that affects in vivo stability (Chambers 1988;Laurie and Stam 1988; Schulte 2001; reviewed in Storz andZera 2011). Finally, post-translational modification (PTM)typically results from the action of enzymes such as proteinkinases (e.g., MacDonald 2004; Plaxton 2004) whose encod-ing genes are not expected to be linked to the structurallocus that they modify. No study has yet identified the ex-tent to which natural genetic variation in each of thesecauses contributes to adaptive variation in enzyme activityassociated with variation in life history.
Multilevel investigations that simultaneously integratemolecular, biochemical, and physiological studies are espe-cially useful for identifying the functional causes and adap-tive significance of molecular–biochemical variation (Koehnet al. 1983, Feder and Watt 1992; Zera and Harshman 2009,2011; Storz and Zera 2011). The wing-polymorphic cricket,Gryllus firmus, has developed into a powerful experimentalmodel for investigating evolutionary alterations in interme-diarymetabolism that underlie important life-history special-izations for dispersal (somatic function) versus reproductiveoutput (Zera and Harshman 2001, 2009, 2011).
Wing polymorphism is a phylogenetically widespreadecologically important polymorphism in insects (Harrison1980; Zera and Denno 1997; Guerra 2011). In G. firmus, thepolymorphism consists of a flight-capable morph [LW(f)]that delays reproduction and a flightless morph (SW) withenhanced early-age fecundity. In G. firmus, relative to theSW morph, the LW(f), dispersing morph exhibits a geneti-cally elevated rate of de novo fatty acid biosynthesis, ele-vated specific activities of lipogenic enzymes, and enhancedproduction of triglyceride flight fuel, a key adaptation fordispersal (Zhao and Zera 2002; Zera and Zhao 2003; Zeraand Harshman 2009, 2011). However, the causes of the in-creased activity of the lipogenic enzymes in LW(f) G. firmusremain unknown.
Here, we focus on NADPþ-isocitrate dehydrogenase(NADPþ-IDH), an enzyme that produces NADPH requiredfor de novo fatty acid biosynthesis and which is known toplay an important role in lipogenesis in numerous organ-isms (Geer et al. 1978; Koh et al. 2004; Liu et al. 2006). Pre-vious studies using artificially selected lines of G. firmushave shown that NADPþ-IDH–specific activity is substan-tially elevated in LW(f) lines that exhibit elevated rate oflipid biosynthesis and elevated triglyceride level duringthe first week of adulthood (Zhao and Zera 2002; Zeraand Zhao 2003; Zera 2005). Recent sequencing of theNadpþ-Idh gene and homogeneous purification of theNADPþ-IDH enzyme from G. firmus (Zera et al. 2011) haveprovided the tools required to investigate the molecularand biochemical causes of the adaptive difference betweenmorphs (genotypes) in enzyme activity, which is the sub-ject of the present study.
Materials and Methods
Background on G. firmus: Morphs, Selected Lines,and Rearing ConditionsGryllus firmus, the sand field cricket, occurs in natural pop-ulations in the southeastern United States as a long-winged(LW) morph, some of which are capable of flight, or asa short-winged (SW) morph that is obligatorily flightless(Veazy et al. 1976; Zera et al. 1997). Except for a few rarecases, all SW females molt into adults with white, nonfunc-tional underdeveloped flight muscles. All LW females ini-tially have fully developed (pink) flight muscles at orshortly after the adult molt [denoted LW(f)]; however,flight muscles degenerate in some of these females render-ing them flightless. In artificially selected lines used in thepresent study, LW(f) females typically (.95%) retainedtheir flight muscles by the end of the first week of adult-hood when biochemical studies were performed. Thus, theLW flightless morph is not considered here (see Zera et al.1997; Zhao and Zera 2001 for physiological and biochem-ical information on this morph).
Studies were conducted on three pairs (blocks; indepen-dent selection trials) of LW- and SW-selected lines. Numberfollowing LW or SW (e.g., LW-1) indicates the block towhich the line belongs. All lines had been derived fromone and same base population and had been raised under
enzymeactivity
enzymeconcentration
geneexpression
structural gene
unlinked regulators(e.g. hormones)
linked regulators(e.g. promoters, enhancers)
enzyme turnoverenzymecatalytic efficiency
primary amino acidsequence
post-translationalmodification
FIG. 1. Factors potentially contributing to variation in enzymeactivity via affects on enzyme concentration (boxes with solid lines)versus catalytic efficiency (boxes with dashed lines). Redrawn fromZera and Harshman (2011).
Schilder et al. · doi:10.1093/molbev/msr171 MBE
3382
Dow
nloaded from https://academ
ic.oup.com/m
be/article/28/12/3381/1031099 by guest on 18 March 2022

the same temperature (28 ± 1 �C), photoperiod (16 light: 8dark), and had been fed the standard (100%) diet sincetheir inception (see Zera 2005 and references therein fordetails of artificial selection). Lines were close to being purebreeding when experiments were conducted (i.e., produced. 95% of the selected morph) and were the same linesused in previous studies of lipid metabolism and life-historyevolution (Zera 2005; Zera and Harshman 2009, 2011) (seeDiscussion).
Tissue Collection and StorageWe collected tissue from adult female G. firmus, on the dayof molt to adulthood (D0) or 5 days post-adult eclosion(D5). These are the same days of adulthood that were stud-ied previously for various physiological differences betweenLW(f) and SW morphs (e.g., Zhao and Zera 2002; Zera andZhao 2003). Individuals were chilled on ice and samples offat body, the primary site of lipid biosynthesis, were re-moved under ice-cold saline (0.15 M NaCl). Samples wereblotted dry, placed in pre-weighed 1.5-ml Eppendorf tubes,flash frozen in liquid nitrogen, and stored at �80 �C.NADPþ-IDH–specific enzyme activity, mRNA transcriptabundance (quantitative polymerase chain reaction[qPCR]), and enzyme protein concentration (see below)were measured from fat body samples taken from the sameindividual. Additional fat body samples or whole bodieswere used for various other studies as described below.Fat body samples were removed from all individuals at ap-proximately the same time of day (i.e., between 11 and 12AM) to eliminate any potential circadian variation in en-zyme activity.
Real-Time qPCR of G. firmus Fat Body Nadpþ-IdhRNA Extraction and Reverse TranscriptionRNA extraction from frozen fat body samples was per-formed using the Nucleospin RNA/Protein Kit (Machery-Nagel), according to manufacturer instructions, whichincluded treatment with DNase that was later removed us-ing a Nucleospin RNA Clean-up Kit (Machery-Nagel). RNAshowed consistently high purity (i.e., 260:280 ratio .1.9)and was stored in 40 ll RNase/DNase-free water at�80 �C. One microgram of total RNA was reverse tran-scribed using 200 U of Superscript III Reverse Transcriptase(Invitrogen) and 2.5 lM modified oligo dT primer (5#-ATTCTAGAGGCCGAGGCGGCCGACATG(t) 30VN-3#) ina total volume of 20 ll. Reverse transcription reactionswere incubated as follows in order to minimize interferencedue to secondary structure: 15 min at 25 �C, 1 h at 36 �C, 1h at 50 �C, 1 h at 55 �C, and 15 min at 70 �C. cDNA sampleswere diluted 1:3 in RNase/DNase-free water and stored at�20 �C until used.
Transcript Abundance of Nadpþ-Idh Using Real-Time
qPCRWe used Primer Express (Applied Biosystems) softwareto design an Nadpþ-Idh Taqman probe (5#- 6-FAM/CCTCCACCAACCCCATTGCTTCC/TAMRA-3#) and prim-er set (forward: 5#-TTCCATCAACAAGGCAAGGAA-3#,
reverse: 5#-CTTTGGTCCAGGCGAAAA-3#) based on thedeposited G. firmus Nadpþ-Idh–coding sequence (GenBankaccession number DQ886272; fig. 1 of Zera et al. 2011). Thisprimer–probe combination results in a 64 nt amplicon cor-responding to nucleotides 1,032–1,096 within DQ88672. Wenormalized Nadpþ-Idh transcript abundances to that ofb-Actin (cytoplasmic) using the following probe andprimer sequences, respectively: 5#- 6-FAM/CCTGTTCTC-CTGACTGAAGCCCCT/TAMRA-3# and forward primer:5#- GAGTTGCCCCTGAGGAGCA-3#, reverse primer: 5#-TTTCCCTGTTGGCTTTAGGGT-3#. These probe andprimer sequences were designed from the b-Actin nucleo-tide sequence of the congener G. assimilis (Anand et al.2008). The b-Actin nucleotide sequence is highly conservedin Gryllus, with the G. assimilis sequence exhibiting 96%identity with the corresponding nucleotide sequence ofG. bimaculatus, a species outside the North American Gryl-lus clade that contains both G. assimilis and G. firmus(Anand et al. 2008). Thus, G. firmus and G. assimilis are ex-pected to exhibit.96% actin sequence identity. This probeand primer combination resulted in a 69 nt amplicon whichis the expected amplification size based on the G. bimacu-latus and G. assimilis sequences (Anand et al. 2008). Exten-sive testing of the Nadpþ-Idh and b-Actin qPCR primer pairsusing standard PCR, but with cycling conditions andmaster-mix the same as that used during the actual qPCR assays,consistently yielded single amplicons of the expected size.Melting curves generated during preliminary qPCR assaysindicated the presence of single amplicons for bothNadpþIdh and b-Actin qPCRs.
Sodium Dodecyl Sulfate–Polyacrylamide GelElectrophoresis, Native PAGE, and Starch GelElectrophoresisSodium dodecyl sulfate–polyacrylamide gel electrophoresis(SDS PAGE) was performed using 10% T gels (Garfin 1990),while native PAGE was performed using 7.5% T, precast BI-ORAD Ready Gels (Tris–HCl buffer, pH 8.2), as described inZera et al. (2011). Horizontal starch gel electrophoresis wasperformed using gels made of 11% hydrolyzed starch dis-solved in 0.013 M Tris–citrate buffer, essentially as de-scribed in Murphy et al. (1996). Tank buffer was 0.135M Tris–citrate, pH 7.3, and gels were run overnight at150 V. A previous native PAGE study (Zera et al. 2011) iden-tified one and the same NADPþ-IDH electromorph in ho-mogenates of whole bodies or various organs. SDS gels werestained for protein using either Coomassie Blue or by silverstaining (Garfin 1990; Zera et al. 2011). Native PAGE orstarch gels were stained for NADPþ-IDH activity (Murphyet al. 1996), as described in Zera et al. (2011). Native starchgels were run in addition to native PAGE gels because, insome instances, phenotypic variation not observed in na-tive PAGE gels is seen in starch gels (Coyne et al. 1979).
Fat Body NADPþ-Isocitrate Dehydrogenase–Specific ActivityFat body samples from single individuals were homoge-nized and assayed for NADPþ-IDH activity as described
Causes of NADPþ-IDH Activity Variation in Gryllus · doi:10.1093/molbev/msr171 MBE
3383
Dow
nloaded from https://academ
ic.oup.com/m
be/article/28/12/3381/1031099 by guest on 18 March 2022

previously (Zhao and Zera 2001; Zera and Zhao 2003).The assay cocktail consisted of 6.0 mM DL-isocitric acidtrisodium salt, 4.0 mM MgCl2, and 0.4 mM NADPþ in 50mM MOPS (3-[N-Morpholino]propane sulfonic acid)buffer. NADPþ-IDH activities were measured in duplicateat 28 �C using a 96-well microtiter plate reader (FLUOstarOmega; BMG Labtech) in 200 ll reaction volumes. Back-ground experiments demonstrated no loss in enzyme ac-tivity due to flash freezing fat body in liquid nitrogen andstorage at �80 �C for several months or during the 1-hperiod when supernatants were kept on ice prior to as-say. Previous studies (described in Zhao and Zera 2001)demonstrated that NADPþ-IDH activities are propor-tional to enzyme concentration and are thus initial rates(no detectable decrease in activity during the assay pe-riod). NADPþ-IDH activities were normalized to totalprotein concentration, measured by the Bradford proteinassay (Bio-Rad) using bovine serum albumin (BSA) asa standard.
Generation and Characterization of a PolyclonalAntibody Against G. firmus NADPþ-IDHInjection of homogeneously purified NADPþ-IDH from G.firmus into New Zealand rabbits failed to produce any de-tectable polyclonal antibodies to the protein. Thus, anti-body against G. firmus NADPþ-IDH was generated(GenScript Corp.) using the synthetic peptide ‘‘YRFHQQG-KETSTNPC’’, which corresponds (except for the final ‘‘C’’crosslinking amino acid) to AA 315-328 of G. firmusNADPþ-IDH (nucleotides 1,026–1,067 of GenBank acces-sion number DQ886272; fig. 1 of Zera et al. 2011). This pep-tide, conjugated to a carrier protein, was injectedsubcutaneously (four injections each several weeks apart)into each of two New Zealand rabbits. The fourth bleed,following the third injection, exhibited a high antibody titerin each rabbit as determined by enzyme-linked immuno-sorbent assay (ELISA) (GenScript Corp.) and was used inall subsequent studies. Antibody bound strongly toNADPþ-IDH purified to homogeneity from G. firmusand subjected to SDS PAGE and western blotting (see sup-plementary methods and supplementary fig. S1, Supple-mentary Material online). Antibody also bound toNADPþ-IDH in crude fat body, although much lessstrongly, and bound to additional proteins (supplementaryfig. S1, Supplementary Material online). Binding of antibodyto NADPþ-IDH in fat body homogenate was more appar-ent when the enzyme was concentrated by differential pre-cipitation with NH4SO4 (resuspended fraction between40% and 80% NH4SO4 saturation), which eliminated theadditional cross-reacting proteins (supplementary fig. S1,Supplementary Material online).
Quantification of G. firmus Fat Body NADPþ-IDHEnzyme Protein ConcentrationNADPþ-IDH enzyme protein concentration in crude fatbody homogenate from LW(f) and SW G. firmus was quan-tified using rocket immunoelectrophoresis (RIE) employing
the G. firmus anti-NADPþ-IDH polyclonal antibody de-scribed above. RIE, which has been used previously to quan-tify numerous proteins in crude tissue homogenates,including genetic variants of NADPþ-IDH (Bentley et al.1983), basically involves incorporation of an antibody intoan agarose gel. Antigen (crude fat body homogenate con-taining NADPþ-IDH in this case) is subjected to electro-phoresis and movement of antigen through the gelcontacts antibody, forming an arc (rocket), the total inten-sity of which is proportional to the concentration of anti-gen (supplementary fig. S2, Supplementary Materialonline). RIE is particularly advantageous for quantifying en-zyme concentration compared with the more commonlyused ELISA when the antibody that is used reacts with mul-tiple antigens, as is the case for the NADPþ-IDH antibodydescribed above. A histochemical stain, specific for the en-zyme in question, can be used to identify the rocket cor-responding to the enzyme of interest (see supplementaryfig. S2, Supplementary Material online). RIE was performedusing a 1% agarose (low-melting) slab gel containing 60mM Tris–HCl, 20 mM Tricine, 1 mM lithium lactate,and 3 mM sodium azide, 10% NADPþ-IDH antiserum, withpH adjusted to 8.6 (modified from Monthony et al. 1978).Five microliters of fat body homogenate of LW(f) or SW G.firmus, diluted 1:8 with the same MOPS buffer used in theenzyme assay, was pipetted into 3- to 4-mm wells punchedinto the gels. The electrode buffer was 60 mM Tris–HCl and20 mM Tricine (pH 8.6). Gels were run for 5 h, at 150 V(;15 mA), under constant cooling at 4 �C. The NADP-IDH enzyme–antibody complexes were stained using a so-lution containing 0.1 M Tris–HCl buffer (pH 8.0) with 2.8mM MgCl2, 3.4 mM Na-isocitrate, 0.15 mM NADPþ, and 5mg MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazo-lium bromide)/ml and a small amount of phenazine me-thosulphate. The gel was incubated at 28 �C in the darkfor approximately 15min. RIEs were scanned using a flatbedscanner (Canon CanoScan 8400F), and total rocket stainingintensities were quantified using ImageJ (NIH) software. Nostained rockets were observed in gels without antibody. Seesupplementary methods (Supplementary Material online)for additional details. A highly linear standard curve (r2 50.985) was obtained using a serially diluted homogenate offat body pooled from three LW(f) and three SW G. firmus(supplementary fig. S2, Supplementary Material online). Toremove gel-to-gel variation in rocket intensity, aliquots ofthe same fat body homogenate were run on all gels ana-lyzed, and rocket staining intensities were standardized tothat homogenate.
Estimation of Kinetic Constants for NADPþ-IDHMichaelis constants (KM) for DL � Naþ-isocitrate andNADPþ as well as the turnover number, kcat, for NADP
þ-IDH were estimated at pH 8.0 using enzyme purified tohomogeneity from whole bodies of crickets from LW- orSW-selected lines of Block 2 (purified separately for eachline). Enzyme was purified to homogeneity as described pre-viously and exhibited a single band of the expected molec-ular mass on SDS gels stained for protein using Coomassie
Schilder et al. · doi:10.1093/molbev/msr171 MBE
3384
Dow
nloaded from https://academ
ic.oup.com/m
be/article/28/12/3381/1031099 by guest on 18 March 2022
blue or silver staining (see Materials and Methods and fig.5 of Zera et al. 2011). Prior kinetic studies (Zera et al.2011) provided KM estimates and saturating substrateconcentrations used to design the kinetic analysis inthe present study. Vmax and KM for a particular substrate(Naþ-isocitrate or NADPþ) were estimated by measuringreaction rates in triplicate for six concentrations of thatsubstrate which bracketed the expected KM while keepingthe other substrate constant under expected saturatingconditions (for additional details, see Zera et al. 2011;for a similar approach, see Place and Powers 1979).Michaelis constants and Vmax were estimated from theuntransformed reaction rates using the nonlinear regres-sion program Enzfitter (Biosoft). kcat was determined bydividing reaction rates by the enzyme protein concentra-tion. In addition, Michaelis constants for each substratewere measured on LW(f) and SW crickets from each ofthe six selected lines. KMs were estimated using superna-tants of crude fat body homogenates (15,000 � g spin for15 min) at pH 8.0, the pH at which enzyme assays havetypically been performed, as well as at pH 7.0, theexpected in vivo pH at the rearing temperature of thecrickets (28 �C). Because of the large number of KM es-timates, we used the two-substrate method of Duggleby(1979) as described in Zera et al. (2011).
Thermal Denaturation ExperimentNADPþ-IDH from whole bodies of LW(f)- and SW-selectedlines of Block 2 were partially purified (each line separately)by anion exchange chromatography to remove substancesthat might influence thermal stability. Enzyme solutionswere diluted in 50 mM Kþ-phosphate buffer, pH 7.8, con-taining 1 mg/ml BSA, 0.1% b-mercaptoethanol, and 1%glycerol, and were incubated for varying amounts of timeat 46 �C. Percent activity remaining was determined bycomparison of incubated samples with enzyme dilutedto the same degree in identical buffer and maintainedon ice during the incubation period.
Statistical Assessment of Phenotypic and GeneticDifferences Between Morphs for VariousBiochemical TraitsThe existence of overall (phenotypic) differences amongLW(f) and SW morphs for a particular trait (e.g., transcriptabundance) was assessed using a standard two-way analysisof variance (ANOVA) of the entire data set (i.e., all lines inall blocks) with morph (fixed effect) and block (randomeffect) as main effects. Significant phenotypic differencesbetween morphs do not necessarily mean that morphs dif-fer genetically for the trait under consideration. Genetic dif-ferences between morphs should be manifest as differencesbetween LW(f)- and SW-selected lines within a block thatare consistent across the blocks (i.e., higher mean transcriptabundance in LW vs. SW lines in each of the three blocks).Thus, within each block, the means of a particular trait werecompared between LW- and SW-selected lines using a stan-dard t-test, and the consistency of differences across blocks
was assessed essentially as performed previously (e.g., seeZhao and Zera 2002; Zera and Zhao 2003, 2006).
Results
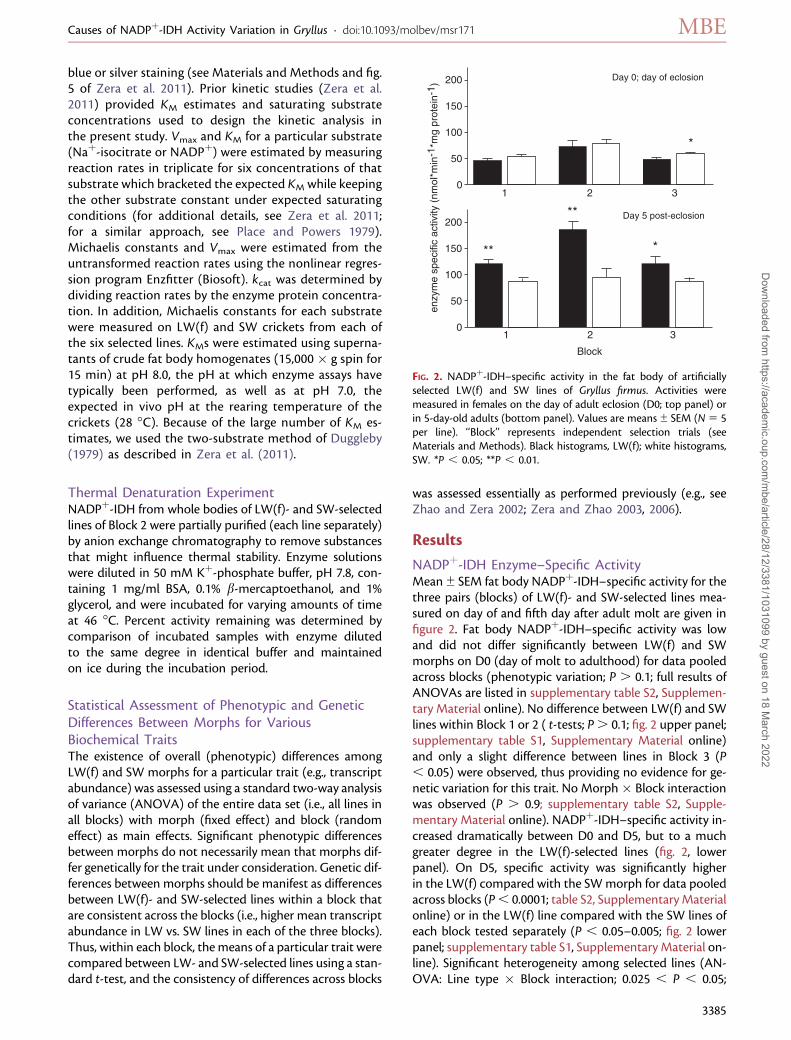
NADPþ-IDH Enzyme–Specific ActivityMean ± SEM fat body NADPþ-IDH–specific activity for thethree pairs (blocks) of LW(f)- and SW-selected lines mea-sured on day of and fifth day after adult molt are given infigure 2. Fat body NADPþ-IDH–specific activity was lowand did not differ significantly between LW(f) and SWmorphs on D0 (day of molt to adulthood) for data pooledacross blocks (phenotypic variation; P . 0.1; full results ofANOVAs are listed in supplementary table S2, Supplemen-tary Material online). No difference between LW(f) and SWlines within Block 1 or 2 ( t-tests; P. 0.1; fig. 2 upper panel;supplementary table S1, Supplementary Material online)and only a slight difference between lines in Block 3 (P, 0.05) were observed, thus providing no evidence for ge-netic variation for this trait. No Morph� Block interactionwas observed (P . 0.9; supplementary table S2, Supple-mentary Material online). NADPþ-IDH–specific activity in-creased dramatically between D0 and D5, but to a muchgreater degree in the LW(f)-selected lines (fig. 2, lowerpanel). On D5, specific activity was significantly higherin the LW(f) compared with the SWmorph for data pooledacross blocks (P, 0.0001; table S2, Supplementary Materialonline) or in the LW(f) line compared with the SW lines ofeach block tested separately (P , 0.05–0.005; fig. 2 lowerpanel; supplementary table S1, Supplementary Material on-line). Significant heterogeneity among selected lines (AN-OVA: Line type � Block interaction; 0.025 , P , 0.05;
150
0
50
100
200
1 2 3
150
0
50
100
200
1 2 3
Day 5 post-eclosion
Block
**
**
*
Day 0; day of eclosion
enzy
me
spec
ific
activ
ity (
nmol
*min
-1*m
g pr
otei
n-1 )
*
FIG. 2. NADPþ-IDH–specific activity in the fat body of artificiallyselected LW(f) and SW lines of Gryllus firmus. Activities weremeasured in females on the day of adult eclosion (D0; top panel) orin 5-day-old adults (bottom panel). Values are means ± SEM (N5 5per line). ‘‘Block’’ represents independent selection trials (seeMaterials and Methods). Black histograms, LW(f); white histograms,SW. *P , 0.05; **P , 0.01.
Causes of NADPþ-IDH Activity Variation in Gryllus · doi:10.1093/molbev/msr171 MBE
3385
Dow
nloaded from https://academ
ic.oup.com/m
be/article/28/12/3381/1031099 by guest on 18 March 2022
supplementary table S2 Supplementary Material online)appeared to be due to the greater difference in specific ac-tivity between LW(f) and SW lines of Block 2 comparedwith lines of the other blocks. In summary, substantial ge-netic elevation of NADPþ-IDH–specific activity was ob-served in LW(f) versus SW lines on D5 but not on D0.
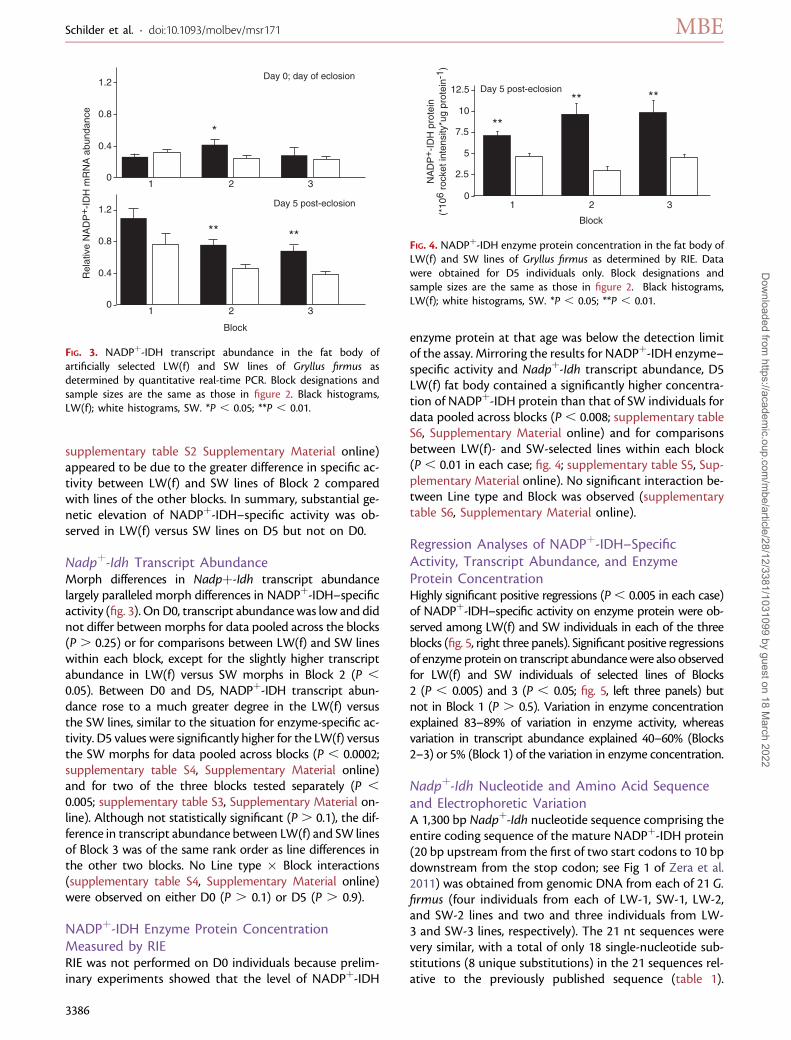
Nadpþ-Idh Transcript AbundanceMorph differences in Nadpþ-Idh transcript abundancelargely paralleled morph differences in NADPþ-IDH–specificactivity (fig. 3). On D0, transcript abundancewas low and didnot differ between morphs for data pooled across the blocks(P . 0.25) or for comparisons between LW(f) and SW lineswithin each block, except for the slightly higher transcriptabundance in LW(f) versus SW morphs in Block 2 (P ,
0.05). Between D0 and D5, NADPþ-IDH transcript abun-dance rose to a much greater degree in the LW(f) versusthe SW lines, similar to the situation for enzyme-specific ac-tivity. D5 values were significantly higher for the LW(f) versusthe SW morphs for data pooled across blocks (P , 0.0002;supplementary table S4, Supplementary Material online)and for two of the three blocks tested separately (P ,
0.005; supplementary table S3, Supplementary Material on-line). Although not statistically significant (P. 0.1), the dif-ference in transcript abundance between LW(f) and SW linesof Block 3 was of the same rank order as line differences inthe other two blocks. No Line type � Block interactions(supplementary table S4, Supplementary Material online)were observed on either D0 (P . 0.1) or D5 (P . 0.9).
NADPþ-IDH Enzyme Protein ConcentrationMeasured by RIERIE was not performed on D0 individuals because prelim-inary experiments showed that the level of NADPþ-IDH
enzyme protein at that age was below the detection limitof the assay. Mirroring the results for NADPþ-IDH enzyme–specific activity and Nadpþ-Idh transcript abundance, D5LW(f) fat body contained a significantly higher concentra-tion of NADPþ-IDH protein than that of SW individuals fordata pooled across blocks (P, 0.008; supplementary tableS6, Supplementary Material online) and for comparisonsbetween LW(f)- and SW-selected lines within each block(P , 0.01 in each case; fig. 4; supplementary table S5, Sup-plementary Material online). No significant interaction be-tween Line type and Block was observed (supplementarytable S6, Supplementary Material online).
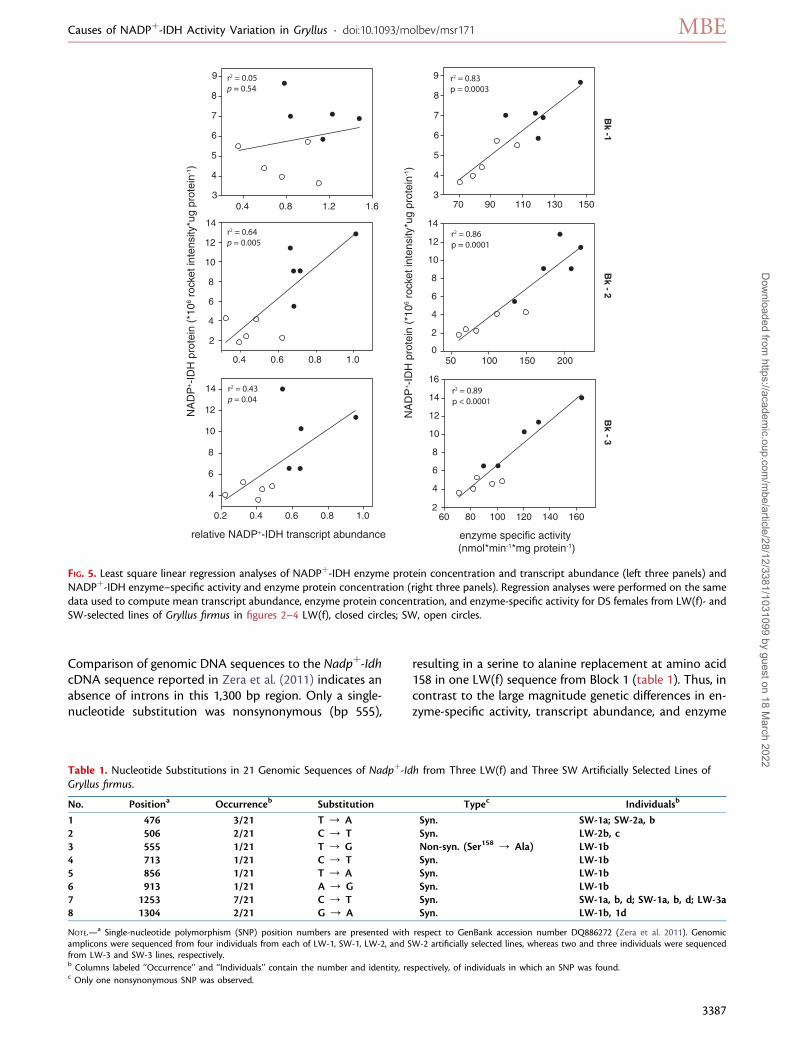
Regression Analyses of NADPþ-IDH–SpecificActivity, Transcript Abundance, and EnzymeProtein ConcentrationHighly significant positive regressions (P, 0.005 in each case)of NADPþ-IDH–specific activity on enzyme protein were ob-served among LW(f) and SW individuals in each of the threeblocks (fig. 5, right three panels). Significant positive regressionsof enzyme protein on transcript abundancewere also observedfor LW(f) and SW individuals of selected lines of Blocks2 (P , 0.005) and 3 (P , 0.05; fig. 5, left three panels) butnot in Block 1 (P . 0.5). Variation in enzyme concentrationexplained 83–89% of variation in enzyme activity, whereasvariation in transcript abundance explained 40–60% (Blocks2–3) or 5% (Block 1) of the variation in enzyme concentration.
Nadpþ-Idh Nucleotide and Amino Acid Sequenceand Electrophoretic VariationA 1,300 bp Nadpþ-Idh nucleotide sequence comprising theentire coding sequence of the mature NADPþ-IDH protein(20 bp upstream from the first of two start codons to 10 bpdownstream from the stop codon; see Fig 1 of Zera et al.2011) was obtained from genomic DNA from each of 21 G.firmus (four individuals from each of LW-1, SW-1, LW-2,and SW-2 lines and two and three individuals from LW-3 and SW-3 lines, respectively). The 21 nt sequences werevery similar, with a total of only 18 single-nucleotide sub-stitutions (8 unique substitutions) in the 21 sequences rel-ative to the previously published sequence (table 1).
0.8
0
0.4
1.2
1 2 3
0.8
0
0.4
1.2
1 2 3
Day 5 post-eclosion
Block
** **
Day 0; day of eclosionR
elat
ive
NA
DP
+-I
DH
mR
NA
abu
ndan
ce
*
FIG. 3. NADPþ-IDH transcript abundance in the fat body ofartificially selected LW(f) and SW lines of Gryllus firmus asdetermined by quantitative real-time PCR. Block designations andsample sizes are the same as those in figure 2. Black histograms,LW(f); white histograms, SW. *P , 0.05; **P , 0.01.
Day 5 post-eclosion
0
2.5
5
7.5
10
12.5
NA
DP
+-I
DH
pro
tein
(*10
6 ro
cket
inte
nsity
*ug
prot
ein-
1 )
1 2 3
Block
**
** **
FIG. 4. NADPþ-IDH enzyme protein concentration in the fat body ofLW(f) and SW lines of Gryllus firmus as determined by RIE. Datawere obtained for D5 individuals only. Block designations andsample sizes are the same as those in figure 2. Black histograms,LW(f); white histograms, SW. *P , 0.05; **P , 0.01.
Schilder et al. · doi:10.1093/molbev/msr171 MBE
3386
Dow
nloaded from https://academ
ic.oup.com/m
be/article/28/12/3381/1031099 by guest on 18 March 2022
Comparison of genomic DNA sequences to the Nadpþ-IdhcDNA sequence reported in Zera et al. (2011) indicates anabsence of introns in this 1,300 bp region. Only a single-nucleotide substitution was nonsynonymous (bp 555),
resulting in a serine to alanine replacement at amino acid158 in one LW(f) sequence from Block 1 (table 1). Thus, incontrast to the large magnitude genetic differences in en-zyme-specific activity, transcript abundance, and enzyme
NA
DP
+-I
DH
pro
tein
(*1
06 ro
cket
inte
nsity
*ug
prot
ein-1
)3
4
5
6
7
8
9
0.4 0.8 1.2 1.6
2
4
6
8
10
12
14
relative NADP+-IDH transcript abundance
0.4 0.6 0.8 1.0
4
6
8
10
12
14
0.2 0.4 0.6 0.8 1.0
r2 = 0.05p = 0.54
r2 = 0.64p = 0.005
r2 = 0.43p = 0.04
enzyme specific activity (nmol*min-1*mg protein-1)
3
4
5
6
7
8
9
70 90 110 130 150
NA
DP
+-I
DH
pro
tein
(*1
06 ro
cket
inte
nsity
*ug
prot
ein-1
)
2
4
6
8
10
12
14
16
60 80 100 120 140 160
r2 = 0.83p = 0.0003
0
2
4
6
8
10
12
14
50 100 150 200
r2 = 0.86p = 0.0001
r2 = 0.89p < 0.0001
Bk -1
Bk - 2
Bk - 3
FIG. 5. Least square linear regression analyses of NADPþ-IDH enzyme protein concentration and transcript abundance (left three panels) andNADPþ-IDH enzyme–specific activity and enzyme protein concentration (right three panels). Regression analyses were performed on the samedata used to compute mean transcript abundance, enzyme protein concentration, and enzyme-specific activity for D5 females from LW(f)- andSW-selected lines of Gryllus firmus in figures 2–4 LW(f), closed circles; SW, open circles.
Table 1. Nucleotide Substitutions in 21 Genomic Sequences of Nadpþ-Idh from Three LW(f) and Three SW Artificially Selected Lines ofGryllus firmus.
No. Positiona Occurrenceb Substitution Typec Individualsb
1 476 3/21 T / A Syn. SW-1a; SW-2a, b2 506 2/21 C / T Syn. LW-2b, c3 555 1/21 T / G Non-syn. (Ser158 / Ala) LW-1b4 713 1/21 C / T Syn. LW-1b5 856 1/21 T / A Syn. LW-1b6 913 1/21 A / G Syn. LW-1b7 1253 7/21 C / T Syn. SW-1a, b, d; SW-1a, b, d; LW-3a8 1304 2/21 G / A Syn. LW-1b, 1d
NOTE.—a Single-nucleotide polymorphism (SNP) position numbers are presented with respect to GenBank accession number DQ886272 (Zera et al. 2011). Genomicamplicons were sequenced from four individuals from each of LW-1, SW-1, LW-2, and SW-2 artificially selected lines, whereas two and three individuals were sequencedfrom LW-3 and SW-3 lines, respectively.b Columns labeled ‘‘Occurrence’’ and ‘‘Individuals’’ contain the number and identity, respectively, of individuals in which an SNP was found.c Only one nonsynonymous SNP was observed.
Causes of NADPþ-IDH Activity Variation in Gryllus · doi:10.1093/molbev/msr171 MBE
3387
Dow
nloaded from https://academ
ic.oup.com/m
be/article/28/12/3381/1031099 by guest on 18 March 2022
protein concentration, no major differences in gene oramino acid sequence were observed for NADPþ-IDHbetween LW(f) and SW G. firmus.
No NADPþ-IDH electrophoretic variation was observedfor fat body homogenates from ten females from each ofthe three LW-selected and each of the three SW-selectedlines subjected to native PAGE. In all cases, a single electro-morph was observed (see supplementary fig. S3, Supple-mentary Material online, for a representative gel). Norwas any electrophoretic variation observed among IDHelectromorphs (20–25 individuals per line for each ofthe three LW and three SW lines) using starch gel electro-phoresis (data not shown).
NADPþ-IDH Enzyme Kinetics and ThermostabilityNADPþ-IDH from LW or SW lines of Block 2, purified tohomogeneity using the protocol of Zera et al. (2011), ex-hibited no significant difference in any of the followingkey kinetic constants: turnover number (kcat) or Michaelisconstant (KM) for either Naþ-isocitric acid or NADPþ
(table 2). Nor (with one exception) were any differencesobserved between LW(f) versus SW lines of each of thethree blocks with respect to NADPþ-IDH Michaelis con-stants for either Na-isocitrate or NADPþ (table 3). Thiswas the case for Michaelis constants measured either atthe standard assay pH 8.0 or pH 7.0, the in vivo pH atthe temperature at which crickets were reared (28 �C).The similarity between LW(f) and SW Michaelis constantsmeasured on homogeneously purified enzyme, after exten-sive purification (Block 2 lines; table 2), compared withconstants measured using unpurified fat body homoge-nates (Block 2 lines; table 3) indicates that these constantsare neither altered significantly during purification nor bymaterial in supernatants of crude homogenates. Similarto the situation for nucleotide sequence, amino acid se-quence, and electrophoretic mobility but unlike specificactivity, transcript abundance, and enzyme protein con-centration (with one exception), no phenotypic or geneticdifferences were observed between LW(f) and SW lines inany aspect of enzyme catalytic efficiency.
Purified NADPþ-IDH from LW and SW lines of Block 2exhibited standard first-order (linear) thermal denatur-ation profiles (r2 . 0.97 for each linear regression; supple-mentary fig. S4, Supplementary Material online). Thermaldenaturation rate constantants (kD; slopes of regressionlines) did not differ significantly between LW(f) and SW
NADPþ-IDHs (kD(LW(f):�0.028 ± 0.001, 95% confidence in-terval [CI] 5 �0.030 to �0.025; kD(SW): �0.024 ± 0.001;95% CI 5 �0.026 to �0.022).
DiscussionAlthough physiological aspects of life-history adaptationhave been extensively studied for decades (Townsendand Calow 1981; Rose and Bradley 1998; Zera and Harsh-man 2001, 2009, 2011) and molecular/genomic data on thistopic are accumulating rapidly (e.g., Pelcher et al. 2005; St-Cyr et al. 2008; Wheat et al. 2011), the biochemical basis oflife-history variation and trade-offs, especially in the con-text of allocation trade-offs in outbred populations, re-mains understudied. The present investigation involveda comprehensive analysis of the relative contribution ofchanges in gene expression, primary amino acid sequence,enzyme kinetics and stability, and (indirectly) posttransla-tional modification to genetic differences in the activity ofan enzyme that is associated with adaptive differences inlipid biosynthesis and life-history. The main finding is thatthere appears to be a relatively straightforward molecularcause of genetic variation in NADPþ-IDH–specific activity.Comparison between pairs of artificially selected linesidentified parallel large magnitude elevations in meanNADPþ-IDH transcript abundance, enzyme protein concen-tration, and enzyme-specific activity in LW(f) lines, relativeto SW lines. Furthermore, regression analysis indicated thatvariation in transcript abundance accounted for a significantamount of variation in enzyme protein concentration intwo of three blocks, which, in turn, accounted for a signifi-cant amount of variation in enzyme-specific activity amongindividuals of LW(f) and SW lines within each of the threeblocks (i.e., independent selection trial). These resultsstrongly indicate that the difference in enzyme-specificactivity between LW(f) and SW lines and LW(f) and SW in-dividuals within blocks is due primarily to differences in geneexpression leading to differences in enzyme concentrationand ultimately enzyme-specific activity.
DNA sequencing, kinetic and thermostability analyses,and electrophoretic studies of NADPþ-IDH strongly sup-port the aforementioned hypothesis. Although variation
Table 2. Substrate and Cofactor Michaelis Constants (KM) andTurnover Numbers (kcat) for Homogeneously Purified CytoplasmicNADPþ-IDH from LW(f) or SW Lines of Gryllus firmus.
Morpha KM(Isocitrate)b KM(NADP1)
b kcatb
LW(f) 13.8 6 1.9 1.2 6 0.5 29,2166 6 2,786SW 11.9 6 2.6 2.6 6 0.9 34,412 6 2,473
NOTE.—a Enzyme was purified from BK-2 LW(f) or SW individuals (see Materialsand Methods for additional information on selected lines and blocks).b Kinetic constants (KM) were estimated for Naþ-isocitrate and NADPþ at pH 8.0at 28 �C (see Materials and Methods); KM are in lM ± SEM. kcat (s
�1 ± SEM) wasestimated assuming one active site per dimeric NADPþ-IDH molecule, usinga subunit molecular mass of 46.3 kD (see Zera et al. 2011).
Table 3. Substrate and Cofactor Michaelis (KM) Constants forNADPþ-IDH from Gryllus firmus LW(f) and SW Morphs Measuredat Two pHs.
pH, Substratea Morph BK-1 BK-2 BK-3
7.0, Isocitric acidLW(f) 12.6 6 2.4 12.1 6 1.6 13.7 6 3.2SW 9.1 6 1.9 10.2 6 1.8 9.2 6 2.8
7.0, NADP1LW(f) 3.3 6 0.3 4.3 6 1.2 4.6 6 0.8SW 4.0 6 0.4 4.5 6 0.8 3.4 6 0.7
8.0, Isocitric acidLW(f) 17.7 6 1.9 14.1 6 2.0 20.3 6 1.7SW 20.6 6 1.8 14.7 6 1.8 16.8 6 2.3
8.0, NADP1LW(f) 1.6 6 0.4 1.4 6 0.6 1.3 6 0.3SW 4.2 6 0.5* 2.3 6 0.4 1.6 6 0.7
NOTE.—a Kinetic constants (KM) were estimated for isocitrate and NADPþ at pH7.0 and 8.0, at 28 �C. Values are presented in lM ± SEM. Asterisk indicatessignificantly different KM between BK-1 LW(f) and SW (P , 0.05); all other KMdifferences between morphs were nonsignificant (P . 0.1). Constants weremeasured on supernatants of crude homogenates (see text).
Schilder et al. · doi:10.1093/molbev/msr171 MBE
3388
Dow
nloaded from https://academ
ic.oup.com/m
be/article/28/12/3381/1031099 by guest on 18 March 2022

in amino acid sequence is a major cause of adaptive var-iation in enzyme function within species (i.e., allozymepolymorphism; Eanes 1999; Watt and Dean 2000; Storzand Zera 2011), this cannot be the case for activity differ-ences between LW(f) and SW NADPþ-IDHs (see table 1;Results). With one exception, the enzymes from all linesexhibit identical amino acid sequences. Nevertheless,LW(f) and SW enzymes could still differ kinetically dueto PTM, a common mechanism for altering enzyme func-tion in response to changing metabolic conditions (Mac-Donald 2004; Plaxton 2004). The absence of differencesbetween the pairs of LW(f) and SW lines in various keycomponents of catalytic efficiency, namely, Michaelis con-stants for NADPþ and isocitric acid, and enzyme turnovernumber, kcat (Results; see below), as well as thermostability,provides no evidence for kinetic or stability differences be-tween LW(f) and SW NADPþ-IDH due to any cause. Theabsence of electrophoretic variation on native PAGE alsoprovides no evidence for PTM. Although more sophisti-cated analyses, such as mass spectrometric comparisonsof NADPþ-IDH amino acid sequences, might uncoverPTM, the absence of kinetic differences between LW(f)and SW NADPþ-IDHs, indicate that any such modificationwould not likely be functionally significant.
Zera and Zhao (2003) reported significantly elevatedspecific activities of NADPþ-IDH and other lipogenic en-zymes in LW(f) versus SW lines of G. firmus. Kinetic anal-yses undertaken in the present study (tables 2 and 3)allow an assessment of the extent to which these in vitroNADPþ-IDH–specific activities exist in vivo and theirlikely functional causes. These issues are best discussedin the context of the basic Michaelis–Menten equationthat describes the reaction rate for an enzyme that hasa single substrate:
v5Vmax � ½S�KM þ ½S� ;
where v 5 reaction rate, [S] is the substrate concentration,Vmax 5 maximal rate, and KM 5 Michaelis constant (i.e., [S]at which v 5 Vmax/2).
Furthermore Vmax 5 [E] � kcat, where [E] 5 enzymeconcentration, kcat 5 turnover number (number of prod-uct molecules produced per unit time per enzyme mole-cule (i.e., active site).
The above equation can be rearranged to separate outcomponents of the reaction rate due to enzyme concen-tration [E] (left side) and kinetic properties of the enzymedue to KM and kcat (right side):
v5 ½E� � kcat � ½S�KM þ ½S�
Importantly, substrate concentration, [S], influences therelative contribution of kcat and KM to the reaction velocity.When [S] is very high (e.g.,.10 � KM), the KM term in thedenominator can be ignored and the right hand term re-duces to kcat. Thus, v 5 [E] � kcat, with KM exerting noinfluence on the reaction rate. However, when [S] is very
low (, KM/10), [S] in the denominator can be ignored inthe equation above, and the rate equation reduces to v 5[E] � (kcat/KM). At intermediate substrate concentrations,both kcat/KM and kcat contribute to the reaction velocity,with the relative importance of each component being de-termined by [S]. Because in vivo substrate concentrationsfor enzymes of intermediary metabolism, in general, arethought to be near or below the KM (Fersht 1999), kcat/KM alone or both kcat and kcat/KM are thought to contrib-ute significantly to the kinetic component of the reactionvelocity for most of these enzymes (Koehn et al. 1983;Fersht 1999; Hochachka and Somero 2002). The rate equa-tion for an enzyme with two substrates (e.g., dehydro-genases such as NADPþ-IDH which bind cofactor inaddition to a specific substrate) has the same form asthe equation above, except that the equation contains ad-ditional terms on the right side reflecting the binding ofmultiple substrates to the enzyme (Fromm 1975).
The elevated specific activity of NADPþ-IDH in LW(f)versus SW lines of G. firmus reported previously (Zeraand Zhao 2003) and verified in the present study (fig. 2)was measured under saturating substrate conditions, asis standard practice for specific activity measurements.For reasons discussed above, if LW(f) and SW enzymes dif-fered in KM, such a difference would not have been man-ifest under conditions used to measure specific activity butcould potentially influence relative reaction rates undersubsaturating conditions expected in vivo. Demonstrationthat the cofactor or substrate KMs do not differ betweenLW(f) and SW NADPþ-IDHs (tables 2 and 3) indicates thatthe relative difference between these enzymes (Zera andZhao 2003; fig. 2) should be manifest under any substratecondition. Furthermore, the statistically equivalent kcat val-ues for LW(f) and SW NADPþ-IDHs (table 2) indicate thatthe elevated enzyme-specific activity in LW(f) lines is due tohigher enzyme concentration and not elevated catalytic ef-ficiency. This result independently corroborates the higher[E] of LW(f) versus SW NADPþ-IDHs measured via immu-noelectrophoresis (fig. 4).
We found no significant difference in in vitro thermaldenaturation rate between LW(f) and SW NADPþ-IDHs(supplementary fig. S4, Supplementary Material online),providing no evidence for differential enzyme degradationthat might also contribute to the differences in [E] betweenLW(f) and SW lines (fig. 4). In vitro rate of thermal dena-turation is often correlated with in vivo stability and turn-over rate for enzymes (Goldberg and St John 1976;McLendon and Radany 1978). Anderson and McDonald(1983) undertook a more sophisticated investigation of thistopic by measuring in vivo turnover rate of ADH usingpulse-label-chase experiments and also found no differencein alcohol dehydrogenase turnover between ADH-FF andADH-SS genotypes.
Implications for Evolutionary BiochemistryAt present, it is difficult to assess the generality of the majorfindings of the present study regarding the biochemicalmechanisms underlying life-history adaptation. Although
Causes of NADPþ-IDH Activity Variation in Gryllus · doi:10.1093/molbev/msr171 MBE
3389
Dow
nloaded from https://academ
ic.oup.com/m
be/article/28/12/3381/1031099 by guest on 18 March 2022

a number of other studies have reported altered enzymefunction correlated with differences in life history (e.g.,Orr et al. 2005; Legan et al. 2008), these studies have typ-ically used highly inbred laboratory stocks containing mu-tations that exhibit large phenotypic effects and often largenegative pleiotropic effects on fitness. Mutants with thesecharacteristics are likely to be eliminated rapidly from out-bred populations and thus are not expected to contributesignificantly to life-history evolution (Hughes and Reynolds2005; Zera and Harshman 2009, 2011). The biochemical ba-sis of life-history variation in outbred populations remainsan understudied aspect of life-history evolution.
Results of the present study also bear on the more gen-eral issue regarding the causes of intraspecific biochemicaladaptation of enzymes. A comprehensive analysis of themolecular and enzymological causes of adaptive variationin allozyme function requires studies to be conducted onhomogeneously purified enzyme in which both the kineticconstants of the enzyme and enzyme concentration arequantified. Surprisingly, few such studies have been con-ducted to date (reviewed in Storz and Zera 2011). Charac-terizations of unpurified or partially purified enzyme canyield important information on adaptive differences insome key enzymatic traits such as the Michaelis constant,Vmax, Vmax/KM (which is the most important contributor tothe overall rate of catalysis under in vivo conditions formost enzymes), and thermal stability. In particular, studiesby Watt and colleagues using partially purified enzymepreparations (e.g., Watt 1977; Watt and Dean 2000) havereported differences in the KM, thermal stability, and Vmax/KM between PGI allozymes of Colias that correlate with as-pects of performance or fitness. However, homogeneouslypurified enzyme is required to estimate the two key com-ponents of Vmax: [E] (often quantified using antibodiesraised against homogeneously purified enzyme, as wasdone in the present study) and kcat, the turnover numberof the enzyme. Genetic variation in [E] can result from nu-cleotide changes affecting gene expression (due to variablegenes linked or unlinked to the structural locus) or enzymestability (due to changes in the amino acid sequence of theenzyme). The key point is that, without information on kcatand [E], it is not possible to disentangle the relative con-tribution of variation in gene expression versus variation inthe properties of the enzyme per se (i.e., kcat or stability) tovariation in Vmax and hence Vmax/KM.
Three notable allozyme studies have focused on the rel-ative importance of variation in kinetic properties of theenzyme versus enzyme concentration to adaptive differen-ces in allozyme function (also see Storz and Zera 2011).Similar to the situation for G. firmus NADPþ-IDHs, differ-ences in enzyme activity between electromorphs (allo-zymes) are completely (LDH, Fundulus heteroclitus) orlargely (ADH, Drosophila melanogaster) due to differencesin enzyme protein concentration, with variation in cata-lytic efficiency playing a much less or no significant role inenzyme adaptation (Eanes 1999; Storz and Zera 2011). Bycontrast, differences in in vivo activity between glucose-6-phosphate dehydrogenase electromorphs appear to be
exclusively or primarily due to variation in allozyme kinet-ics, specifically the KM for glucose-6-phosphate, with noobserved differences in either kcat or [E] (Eanes 1999).The relative importance of kinetic versus regulatory causesof adaptive variation in enzyme activity remainsan unresolved and understudied topic in evolutionarybiochemistry.
In a similar vein, only a few studies have investigatedgenetic or molecular mechanisms responsible for geneticvariation in [E]. Quantitative genetic studies in D. mela-nogaster indicate that genetic variation in [E] is influencedby variable genes on every chromosome (Chambers 1988),implying that a multitude of mechanisms can be involved.Similar to the situation for G. firmus, studies in Fundulushave shown that differences among LDH genotypes in[E] is exclusively due to genotypic differences in LDH tran-script abundance for fish raised at a particular temperature(Crawford and Powers 1989). Subsequent work in F. heter-oclitus has gone on to identify allozyme-associated differ-ences in gene transcription (Schulte 2001) that account forthe variation in transcript abundance. By contrast, studiesin D. melanogaster have identified no difference in tran-script abundance between ADH genotypes that differ in[E] (Laurie and Stam 1988). Interestingly, allelic differencesin [E] for ADH appear to be due to epistatic interactionsbetween a small insertion–deletion polymorphism withinthe first intron of the ADH structural locus that is in stronglinkage disequilibrium with the nucleotide site responsiblefor the fast/slow amino acid substitution (Laurie and Stam1988; Storz and Zera 2011). The mechanism by which thisindel influences [E] in a genotype-specific manner is un-known. The molecular mechanisms responsible formorph-associated differences in NADPþ-IDH transcriptabundance in G. firmus have not been investigated. An-other lipogenic enzyme in G. firmus, 6-phosphogluconatedehydrogenase, differs in specific activity between morphs,similar to the situation for NADPþ-IDH (Schilder R andZera AJ, unpublished data). However, no significant covari-ance between transcript abundance and either specific ac-tivity or 6-PGDH enzyme concentration have beenobserved (Schilder R and Zera AJ, unpublished data). Inshort, for the relatively few studies of genetically variableenzyme activity in outbred populations, no consistent pat-tern has emerged with regard to the correlation betweenenzyme activity and transcript abundance. This findingunderscores the importance of having corroborating pro-tein data when drawing inferences on the functional oradaptive significance of variable transcript abundance(Idaker et al. 2001; Feder and Walser 2005; Zera 2011).
The present study constitutes an important step in iden-tifying the biochemical basis of morph-specific differencesin lipogenic flux that underlies life-history adaptations ofthe LW(f) and SW morphs of G. firmus. Both theoreticalconsiderations and experimental results suggest that themorph difference in NADPþ-IDH activity, by itself, is notthe primary cause of the flux difference in lipogenesis be-tween the morphs. ‘‘Multisite modulation,’’ a key conceptof Metabolic Control Analysis (Kacser and Burns 1979; Fell
Schilder et al. · doi:10.1093/molbev/msr171 MBE
3390
Dow
nloaded from https://academ
ic.oup.com/m
be/article/28/12/3381/1031099 by guest on 18 March 2022

and Thomas 1995; summarized in Fell 1997), proposes thatthe optimal increase in pathway flux should result from anequivalent elevation of activities of multiple enzymes ofa pathway rather than increasing activities of one ora few enzymes. According to this scenario, co-ordinatechange in multiple enzymes in toto cause the flux change,a hypothesis that is supported by many studies reportingco-ordinate changes in multiple enzymes of a metabolicpathway in response to various regulators, such as insulin,glucagon, hypoxia regulator, etc. (summarized in Fell 1997;Hochachka et al. 1998; Zera and Zhao 2003; Zera andHarshman 2011). The coordinately increased activity of nu-merous lipogenic enzymes and lipogenic flux in LW(f) ver-sus SW lines of G. firmus (Zhao and Zera 2002; Zera andZhao 2003) and artificially selected lines of Mus musculus(reviewed in Fell 1997 and in Zera 2011) are also consistentwith this hypothesis. Morph differences in global hor-monal or metabolic regulators (e.g., juvenile hormone, ec-dysteroids, insulin-like peptides, AMP kinase, etc.; Plaxton2004; Zera and Zhao 2004) likely play the key role in co-ordinating the increase in lipogenic enzyme activities inLW(f) versus SW lines. Morph-specific differences inNADPþIDH gene expression, enzyme concentration,and enzyme activity now provide a useful experimentalmodel to investigate the mechanism of action of theseglobal regulators.
Resurgence of Evolutionary EnzymologyBiochemical characterization of allozymes was a key focusof population genetics during the 1970s–1980s (Koehnet al. 1983; Zera et al. 1985; Watt and Dean 2000). Witha few notable exceptions, the focus of experimental pop-ulation genetics largely shifted in subsequent decades fromfunctional studies of variable proteins to statistical analysesof molecular sequence variation (Eanes 1999; Watt andDean 2000; Storz and Zera 2011). The consequence of thisshift is that currently there is a much greater amount ofinformation on genetic variation in DNA sequence, aminoacid sequence, and transcript abundance than informationon enzyme function. Because of this imbalance, functionalstudies of genetically variable enzymes are assuming in-creasing importance because functional information is es-sential to thoroughly identify specific mechanisms ofprotein microevolution (Storz and Zera 2011). Indeed,the extensive information on variation in gene expressionand nucleotide and amino acid sequences now providevaluable background context for functional studies of en-zyme function. For many enzymes, most notably dehydro-genases, standard chromatographic techniques, such asaffinity chromatography, allow relatively straightforwardhomogeneous purification of allozymes (Place and Powers1979; Eanes 1999; Zera et al. 2011). Moreover, newer mo-lecular techniques, such as in vitro protein expression, al-low purified enzyme to be obtained in cases in whichsufficient starting material is unavailable or in which theenzyme is difficult to purify, thus making studies of purifiedenzymes and hence estimation of key parameters such askcat and [E] more feasible. Finally, as mentioned above, it is
becoming increasingly apparent that the extent to whichvariation in gene expression gives rise to variation in pro-tein function or concentration cannot be assumed a prioribut must be established empirically through functionalstudies. The convergence of these factors provides a strongimpetus for a resurgence in functional studies of enzymeadaptation which will likely occur in the near future.
Supplementary MaterialSupplementary method, tables S1–S6, and figures S1–S4are available at Molecular Biology and Evolution online(http://www.mbe.oxfordjournals.org/).
AcknowledgmentsResearch reported here was supported by NSF award IOS-0516973 to AJZ.
ReferencesAnand A, Crone EJ, Zera AJ. 2008. Tissue and stage-specific juvenile
hormone esterase (JHE) and epoxide hydrolase (JHEH) enzymeactivities and Jhe transcript abundance in lines of the cricketGryllus assimilis artificially selected for plasma JHE activity:implications for JHE microevolution. J Insect Physiol.54:1323–1331.
Anderson SM, McDonald JF. 1983. Biochemical and molecularanalysis of naturally-occurring Adh variants in Drosophilamelanogaster. Proc Natl Acad Sci U S A. 80:4798–4802.
Arking R, Burde V, Graves K, et al. (13 co-authors). 2000. Forwardand reverse selection for longevity in Drosophila is characterizedby alteration of antioxidant gene expression and oxidativedamage patterns. Exp Gerontol. 35:167–185.
Bentley MM, Meidinger RG, Williamson JH. 1983. Characterizationof a low-activity allele of Nadpþ-dependent isocitrate de-hydrogenase from Drosophila melanogaster. Biochem Genet.21:725–733.
Chambers GK. 1988. The Drosophila alcohol-dehydrogenase gene-enzyme system—introduction. Adv Genet. 25:39–107.
Coyne JA, Eanes WF, Ramshaw JAM, Koehn RK. 1979. Electropho-retic heterogeneity of alpha-glycerophosphate dehydrogenaseamong many species of Drosophila. Syst Zool. 28:164–175.
Crawford DL, Powers DA. 1989. Molecular basis of evolutionaryadaptation at the lactate dehydrogenase-B locus in thefish Fundulus heteroclitus. Proc Natl Acad Sci U S A. 86:9365–9369.
Duggleby RG. 1979. Experimental designs for estimating the kineticparameters for enzyme-catalyzed reactions. J Theor Biol.81:671–684.
Eanes WF. 1999. Analysis of selection on enzyme polymorphisms.Annu Rev Ecol Syst. 30:301–326.
Feder ME, Walser JC. 2005. The biological limitations of tran-scriptomics in elucidating stress and stress responses. J Evol Biol.18:901–910.
Feder ME, Watt WB. 1992. Functional biology of adaptation. In:Berry RJ, Crawford TJ, Hewitt GM, editors. Genes in ecology.Oxford: Blackwell Scientific Publications. p. 365–392.
Fell D. 1997. Understanding the control of metabolism. London:Portland Press.
Fell D, Thomas T. 1995. Physiological control of metabolic flux: therequirement for multisite modulation. Biochem J. 311:35–39.
Fersht A. 1999. Structure and mechanism in protein science. NewYork: W. H. Freeman and Co.
Fromm HJ. 1975. Initial rate enzyme kinetics. Berlin: Springer.
Causes of NADPþ-IDH Activity Variation in Gryllus · doi:10.1093/molbev/msr171 MBE
3391
Dow
nloaded from https://academ
ic.oup.com/m
be/article/28/12/3381/1031099 by guest on 18 March 2022

Garfin DA. 1990. One-dimensional gel electrophoresis. MethodsEnzymol. 182:425–441.
Geer BW, Woodward CG, Marshall SD. 1978. Regulation of theoxidative NADP-enzyme tissue levels in Drosophila melanogast-er. II The biochemical basis of dietary carbohydrate and D-glycerate modulation. J Exp Zool. 203:391–402.
Goldberg AL, St John AC. 1976. Intracellular protein degradation inmammalian and bacterial cells. Part 2. Annu Rev Biochem.45:747–803.
Guerra PA. Forthcoming 2011. Evaluating the life history trade-offbetween dispersal capability and reproduction in wing di-morphic insects: a meta-analysis. Biol Rev.
Harrison RG. 1980. Dispersal polymorphisms in insects. Annu RevEcol Syst. 11:95–118.
Hochachka PW, McClelland GB, Burness GP, Staples JF, Suarez RK.1998. Integrating metabolic pathway fluxes with gene-to-enzyme expression rates. Comp Biochem Physiol. 120:17–26.
Hochachka PW, Somero GN. 2002. Biochemical adaptation.Mechanism and process in physiological evolution. Oxford:Oxford University Press.
Hughes K, Reynolds R. 2005. Evolutionary and mechanistic theoriesof ageing. Annu Rev Entomol. 50:421–445.
Idaker T, Thorsson V, Ranish JA, Christmas R, Buhler J, Eng JK,Bumgarner R, Goodlett DR, Aebersold R, Hood L. 2001.Integrated genomic and proteomic analyses of a systemicallyperturbed metabolic network. Science 272:929–934.
Kacser H, Burns JA. 1979. Molecular democracy: who shares thecontrols. Biochem Soc Trans. 7:1149–1160.
Koehn RK, Zera AJ, Hall JG. 1983. Enzyme polymorphism andnatural selection. In: Evolution of genes and proteins. Nei M,Koehn RK, editors. Sunderland (MA): Sinauer p. 115–136
Koh H-J, Lee S-M, Son B-G, et al. (12 co-authors). 2004. CytosolicNADPþ-dependent isocitrate dehydrogenase plays a key role inlipid metabolism. J Biol Chem. 279:39968–39974.
Laurie CC, Stam LF. 1988. Quantitative analysis of RNA produced byslow and fast alleles of Adh in Drosophila melanogaster. ProcNatl Acad Sci U S A. 85:5161–5165.
Legan SK, Rebrin I, Mockett RJ, Radyuk SN, Klichko VI, Sohal RS,Orr WC. 2008. Overexpression of glucose-6-phosphate de-hydrogenase extends the life span of Drosophila melanogaster. JBiol Chem. 283:32492–32499.
Liu W, Capuco AV, Romagnolo DF. 2006. Expression of cytosolicNADPþ-dependent isocitrate dehydrogenase in bovine mam-mary epithelium: modulation by regulators of differentiationand metabolic effectors. Exp Biol Med. 231:599–610.
MacDonald JA. 2004. Signal transduction pathways and the controlof cellular responses to external stimuli. In: Storey KB, editor.Functional metabolism: Regulation and adaptation. Hoboken(NJ): Wiley-Liss. p. 87–123.
McLendon C, Radany E. 1978. Is protein turnover thermodynam-ically controlled? J Biol Chem. 253:6335–6337.
Monthony JF, Wallace EG, Allen DM. 1978. A non-barbital buffer forimmunoelectrophoresis and zone electrophoresis in agarosegels. Clin Chem. 24:1825–1827.
Murphy R, Sites J Jr., Buth D, Haufler C. 1996. Proteins: isozymeelectrophoresis. In: Hillis D, Moritz C, Mable B, editors. Molecularsystematics. Sunderland (MA): Sinauer Associates, Inc. p. 51–120.
O’Brien DM, Marilyn ML, Boggs CL. 2002. Renewable andnonrenewable resources: amino acid turnover and allocationto reproduction. Proc Natl Acad Sci U S A. 99:4413–4418.
Orr WC, Radyuk SN, Prabhudesai L, Toroser D, Benes JJ, Luchak JM,Mockett RJ, Rebrin I, Hubbard JG, Sohal RS. 2005. Over-expression of glutamate-cysteine ligase extends life span inDrosophila melanogaster. J Biol Chem. 280:37331–37338.
Pelcher SD, Macdonald SJ, Marguerie SR, Certa U, Stearns SC,Partridge L. 2005. Genome-wide transcript profiles in aging and
calorically restricted Drosophila melanogaster. Curr Biol.12:712–723.
Place AR, Powers DA. 1979. Genetic variation and relative catalyticefficiencies: lactate dehydrogenase-B allozymes of Fundulusheteroclitus. Proc Natl Acad Sci U S A. 76:2354–2358.
Plaxton WC. 2004. Principles of metabolic control. In: Storey KB,editor. Functional metabolism: regulation and adaptation.Hoboken (NJ): Wiley-Liss. p. 1–23.
Rose MR, Bradley TJ. 1998. Evolutionary physiology of the cost ofreproduction. Oikos 83:443–451.
Schulte P. 2001. Environmental adaptations as windows onmolecular evolution. Comp Biochem Physiol. 128:597–611.
St-Cyr J, Derome N, Bernatchez L. 2008. The transcriptomics of lifehistory trade-offs in whitefish species pairs (Coregonus sp.). MolEcol. 17:1850–1870.
Storz J, Zera AJ. Forthcoming 2011. Experimental approaches forevaluating the contribution of amino acid mutations toadaptive phenotypic evolution. Mol Methods Evol Genet.772:(In Press).
Townsend CR, Calow P. 1981. Physiological ecology. An evolutionaryapproach to resource use. Oxford: Blackwell Scientific Publications.
Veazy JN, Kay CAR, Walker TJ, Whitcomb WH. 1976. Seasonalabundance, sex ratio, and macroptery of field crickets innorthern Florida. Ann Entomol Soc Am. 69:374–380.
Watt WB. 1977. Adaptation at specific loci. I. Natural selection onphosphoglucose isomerase of Colias butterflies: biochemical andpopulational aspects. Genetics 87:177–194.
Watt WB, Dean AM. 2000. Molecular-functional studies of adaptivegenetic variation in prokaryotes and eukaryotes. Annu RevGenet. 34:593–622.
Wheat CW, Fescemyer HW, Kvist J, Tas E, Vera JC, Frilander MJ,Hanski I, Marden JH. 2011. Functional genomics of life historyvariation in a butterfly metapopulation. Mol Ecol. 20:1813–1828.
Zera AJ. 2005. Intermediary metabolism and life history trade-offs:lipid metabolism in lines of the wing-polymorphic cricket,Gryllus firmus, selected for flight capability vs. early agereproduction. Integr Comp Biol. 45:511–524.
Zera AJ. 2011. Microevolution of intermediary metabolism:evolutionary genetics meets metabolic biochemistry. J Exp Biol.214:179–190.
Zera AJ, Denno RF. 1997. Physiology and ecology of dispersalpolymorphism in insects. Annu Rev Entomol. 42:207–231.
Zera AJ, Harshman LG. 2001. Physiology of life history trade-offs inanimals. Annu Rev Ecol Syst. 32:95–126.
Zera AJ, Harshman LG. 2009. Laboratory selection studies of life-history physiology in insects. In: Garland T Jr, Rose MR, editors.Experimental evolution: methods and applications. Berkeley(CA): University of California Press. p. 217–262.
Zera AJ, Harshman LG. 2011. Intermediary metabolism and thebiochemical-molecular basis of life history variation and trade-offs in two insect models. In: Flatt T, Heyland A, editors.Molecular mechanisms underlying life history evolution. Oxford:Oxford University Press. p. 311–328.
Zera AJ, Koehn JG, Hall JG. 1985. Allozymes and biochemicaladaptation. In: Kerkut GA, Gilbert LI, editors. Comprehensiveinsect physiology, biochemistry and pharmacology. Oxford:Pergamon. p. 633–674.
Zera AJ, Newman S, Berkheim D, Black C, Klug L, Crone E.Forthcoming 2011. Purification and characterization of cyto-plasmic NADPþ-isocitrate dehydrogenase, and amplification ofthe Nadpþ-Idh gene from the wing-dimorphic sand fieldcricket, Gryllus firmus. J Insect Sci. 11(53):
Zera AJ, Sall J, Grudzinski K. 1997. Flight-muscle polymorphism inthe cricket Gryllus firmus: muscle characteristics and theirinfluence on the evolution of flightlessness. Physiol Zool.70:519–529.
Schilder et al. · doi:10.1093/molbev/msr171 MBE
3392
Dow
nloaded from https://academ
ic.oup.com/m
be/article/28/12/3381/1031099 by guest on 18 March 2022

Zera AJ, Zhao Z. 2003. Life history evolution and the microevolutionof intermediary metabolism: activities of lipid-metabolizingenzymes in life-history morphs of a wing-dimorphic cricket.Evolution 57:568–596.
Zera AJ, Zhao Z. 2004. Effect of a juvenile hormone analogue onlipid metabolism in a wing-polymorphic cricket: implications forthe biochemical basis of the trade-off between reproduction anddispersal. Biochem Physiol Zool. 77:255–266.
Zera AJ, Zhao Z. 2006. Intermediary metabolism and life-historytrade-offs: differential metabolism of amino acids underlies the
dispersal-reproduction trade-off in a wing-polymorphic cricket.Am Nat. 167:889–900.
Zhao Z, Zera AJ. 2001. Enzymological and radiotracer studies of lipidmetabolism in the flight-capable and flightless morphs of thewing-polymorphic cricket, Gryllus firmus. J Insect Physiol. 47:1337–1347.
Zhao Z, Zera AJ. 2002. Differential lipid biosynthesis underliesa tradeoff between reproduction and flight capability ina wing-polymorphic cricket. Proc Natl Acad Sci U S A. 99:16829–16834.
Causes of NADPþ-IDH Activity Variation in Gryllus · doi:10.1093/molbev/msr171 MBE
3393
Dow
nloaded from https://academ
ic.oup.com/m
be/article/28/12/3381/1031099 by guest on 18 March 2022
Related Documents











