Facies (2012) 58:637–647 DOI 10.1007/s10347-012-0297-1 123 ORIGINAL ARTICLE Mohlerina basiliensis (Mohler, 1938): a Middle Jurassic–Early Cretaceous facultative (?) epilithic benthic foraminifer Felix Schlagintweit Received: 26 November 2011 / Accepted: 25 January 2012 / Published online: 17 February 2012 © Springer-Verlag 2012 Abstract Mohlerina basiliensis (Mohler, 1938) repre- sents a common, cosmopolitan, shallow-water Middle Jurassic–Early Cretaceous trochospiral foraminifer. Given the numerous illustrations from thin-section specimens in the literature displaying tests dispersed within the matrix (micritic and sparitic), a free, vagile benthic mode of life is generally suggested. As an exceptional case, specimens are found with their tests attached to a variety of hard sub- strates, thus creating a special elevated microhabitat. The Wxation is due to a calcitic cement. With its facultative ses- sile mode of life, Mohlerina can be compared with several modern calcareous trochospiral rotaliacean foraminifera (e.g., Ammonia, Cibicides, Discorbis). The rare preserva- tion of Wxed Mohlerina in situ is most likely due to physical taphonomic degradation (e.g., breakage). Tests of Mohlerina attached to hard substrates were found in both low- and high- energy paleoenvironments. The high test variability of Mohlerina might be inXuenced by the diVerent settling strategies and substrate surface topographies within diVer- ent paleohabitats. Keywords Mohlerina · Benthic foraminifera · Rotaliacea · Microhabitat · Taphonomy · Paleoecology Introduction Benthic foraminifera are typical constituents of Paleozoic– Cenozoic shallow-water platform carbonates where a large variety of diVerent paleoenvironmental settings oVered a plenty of ecological niches for settlement. A common, cos- mopolitan taxon of Middle Jurassic to Early Cretaceous carbonates is Mohlerina basiliensis (Mohler) originally described by Mohler (1938) from the Upper Oxfordian of Switzerland. According to Bassoullet (1997), it has a rather long stratigraphic range from the Middle/Upper Bathonian until the Valanginian. Originally, this species was described as Conicospirillina basiliensis by Mohler. As Bernier (1984, p. 526) stated, however, due to its plurilocu- lar test it cannot belong to Conicospirillina Cushman which consists of a proloculus followed by a single trochospirally coiled chamber. In addition, Conicospirillina builds its test from calcite that is in crystallographic continuity, whereas the species described by Mohler (1938) exhibits a lamellar wall structure. Finally, Bucur et al. (1996) made a taxo- nomic revision with the creation of the new genus Mohle- rina and the new combination M. basiliensis (Mohler). It is worth mentioning, however, that representatives of the genus Conicospirillina are well represented in the Upper Jurassic (e.g., Quilty 2011). Mohlerina basiliensis was reported preferentially from high-energy facies, but also occurs in micritic sediments, from inner platform to upper slope depositional settings (Védrine 2008, with references therein). Given that the numerous illustrations from thin-section specimens in the literature show tests dispersed within the matrix (micritic and sparitic), a motile benthic mode of life can be expected. Based on specimens from the Upper Jurassic of the Northern Calcareous Alps of Austria, this study describes an alternative facultative Wxo-sessile mode of life of Mohlerina permitting additional comparisons with modern rotaliacean foraminifers along with consider- ations about the suprageneric taxonomic position of the genus Mohlerina. F. Schlagintweit (&) Lerchenauerstr. 167, 80935 Munich, Germany e-mail: [email protected]

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Facies (2012) 58:637–647
DOI 10.1007/s10347-012-0297-1ORIGINAL ARTICLE
Mohlerina basiliensis (Mohler, 1938): a Middle Jurassic–Early Cretaceous facultative (?) epilithic benthic foraminifer
Felix Schlagintweit
Received: 26 November 2011 / Accepted: 25 January 2012 / Published online: 17 February 2012© Springer-Verlag 2012
Abstract Mohlerina basiliensis (Mohler, 1938) repre-sents a common, cosmopolitan, shallow-water MiddleJurassic–Early Cretaceous trochospiral foraminifer. Giventhe numerous illustrations from thin-section specimens inthe literature displaying tests dispersed within the matrix(micritic and sparitic), a free, vagile benthic mode of life isgenerally suggested. As an exceptional case, specimens arefound with their tests attached to a variety of hard sub-strates, thus creating a special elevated microhabitat. TheWxation is due to a calcitic cement. With its facultative ses-sile mode of life, Mohlerina can be compared with severalmodern calcareous trochospiral rotaliacean foraminifera(e.g., Ammonia, Cibicides, Discorbis). The rare preserva-tion of Wxed Mohlerina in situ is most likely due to physicaltaphonomic degradation (e.g., breakage). Tests of Mohlerinaattached to hard substrates were found in both low- and high-energy paleoenvironments. The high test variability ofMohlerina might be inXuenced by the diVerent settlingstrategies and substrate surface topographies within diVer-ent paleohabitats.
Keywords Mohlerina · Benthic foraminifera · Rotaliacea · Microhabitat · Taphonomy · Paleoecology
Introduction
Benthic foraminifera are typical constituents of Paleozoic–Cenozoic shallow-water platform carbonates where a large
variety of diVerent paleoenvironmental settings oVered aplenty of ecological niches for settlement. A common, cos-mopolitan taxon of Middle Jurassic to Early Cretaceouscarbonates is Mohlerina basiliensis (Mohler) originallydescribed by Mohler (1938) from the Upper Oxfordian ofSwitzerland. According to Bassoullet (1997), it has a ratherlong stratigraphic range from the Middle/Upper Bathonianuntil the Valanginian. Originally, this species wasdescribed as Conicospirillina basiliensis by Mohler. AsBernier (1984, p. 526) stated, however, due to its plurilocu-lar test it cannot belong to Conicospirillina Cushman whichconsists of a proloculus followed by a single trochospirallycoiled chamber. In addition, Conicospirillina builds its testfrom calcite that is in crystallographic continuity, whereasthe species described by Mohler (1938) exhibits a lamellarwall structure. Finally, Bucur et al. (1996) made a taxo-nomic revision with the creation of the new genus Mohle-rina and the new combination M. basiliensis (Mohler). It isworth mentioning, however, that representatives of thegenus Conicospirillina are well represented in the UpperJurassic (e.g., Quilty 2011).
Mohlerina basiliensis was reported preferentially fromhigh-energy facies, but also occurs in micritic sediments,from inner platform to upper slope depositional settings(Védrine 2008, with references therein). Given that thenumerous illustrations from thin-section specimens in theliterature show tests dispersed within the matrix (micriticand sparitic), a motile benthic mode of life can beexpected. Based on specimens from the Upper Jurassic ofthe Northern Calcareous Alps of Austria, this studydescribes an alternative facultative Wxo-sessile mode oflife of Mohlerina permitting additional comparisons withmodern rotaliacean foraminifers along with consider-ations about the suprageneric taxonomic position of thegenus Mohlerina.
F. Schlagintweit (&)Lerchenauerstr. 167, 80935 Munich, Germanye-mail: [email protected]
123

638 Facies (2012) 58:637–647
Geological overview and material
In the Northern Calcareous Alps, M. basiliensis (Mohler)represents a widely distributed foraminifer in the LateJurassic-earliest Cretaceous Plassen Carbonate Platform(Fenninger and Hötzl 1967; Steiger and Wurm 1980;Schlagintweit and Ebli 1999; Schlagintweit et al. 2005).With a preference for well-agitated outer platform settings,it predominantly occurs in the Upper Kimmeridgian andUpper Tithonian-lowermost Berriasian parts of the PlassenCarbonate Platform. In the resedimented Barmstein Lime-stone with its dominance of clasts of the Plassen CarbonatePlatform, M. basiliensis is one of the most characteristicforaminifera, besides protopeneroplids and some largeragglutinating lituolids (Steiger 1981; Gawlick et al. 2005).The species is especially abundant as individual bioclasts incalciturbidites (grain- to pack-stones), more rarely it occursin lithiWed clasts of the Barmstein megabreccias character-ized by their mainly clast-supported fabric. The occur-rences of these Upper Jurassic limestones are concentratedin the Salzkammergut area. The material studied originatesfrom the following localities (Fig. 1):
Barmstein limestone
Mount Höherstein Plateau, Salzkammergut (Gawlicket al. 2007).Weitenau syncline, Osterhorn Mountains of the Salz-burg Calcareous Alps (Plöchinger 1984; Krische et al.2011).Mount Zwerchwand, Salzkammergut (Gawlick et al.2010).
Plassen Carbonate Platform
Mount Rettenstein (Auer et al. 2009).Mount Trisselwand (Schlagintweit and Ebli 1999).
Paleontologic part
Order Foraminiferida Eichwald, 1830Suborder Rotaliina Delage and Hérouard, 1986? Family Rosalinidae Reiss, 1963
Remarks Following a suggestion of Bernier (1984),Bucur et al. (1996) placed Mohlerina within the familyDiscorbidae Ehrenberg. According to Loeblich and Tappan(1988, p. 557) the family Discorbidae is known from theMiddle Eocene–Holocene, the superfamily Discorbaceafrom the Middle Jurassic (Aalenian) to Holocene. Accord-ing to the revision of Levy et al. (1986), the heterogeneousfamily Discorbidae should be replaced by Rosalinidae(Hansen and Revets 1992, for details) whose range onlyrecently has been extended into the Albian (Revets 2002).This proposal, however, has not been followed by any sub-sequent worker who without exception still retain the Disc-orbidae (e.g., Foresi et al. 2004).
In any case, the ability of Mohlerina to attach itself tohard substrates as demonstrated here provides further evi-dence for the inclusion of the genus within the calcareousRotaliina as the modern trochospirally coiled and attachedcalcareous taxa belong to this group. However, the exactsuprageneric position still needs further clariWcation. TheDiscorbidae are characterized by “an umbilical aperturewith a distinct Xap extending over the umbilical region”(Loeblich and Tappan 1988, p. 557), a feature so far notseen in Mohlerina.
Genus Mohlerina Bucur, Senowbari-Daryan andAbate, 1996M. basiliensis (Mohler, 1938) (Figs. 2, 3 and 4, ?5b,?5d, 6, 7d)
Synonymy See Bernier (1984), Bucur et al. (1996),Schlagintweit et al. (2005), and Védrine (2008).
Description Test large (up to 2.04 mm in diameter) troc-hospirally enrolled with a highly variable apical angle rang-ing from 77–150° (Table 1). Depending on the apical angle,the outline in axial views is almost discoidal to high-conicalwith rounded peripheral margins. In transverse sections, theoutline is almost circular (Fig. 2e). The spiral (dorsal) sideis convex, the umbilical (ventral) side may be Xat or exhib-its a concave depression with or without a distinct convexbulge (Fig. 2a, i, j). In axial sections chamber shapes aresubglobular becoming elongated hemi-spherical (Fig. 2a).In large specimens, there may be up to 13 crescent-shapedchambers in the Wnal whorl visible in transverse sections(Fig. 2b, c, e). The foramina are represented by a low inte-riomarginal slit on the umbilical side. Details of theaperture (e.g., lip, accessory openings) are unknown. Thetest wall is calcareous bilamellar with an inner thin micro-crystalline (“inner lining”, Hansen and Reiss 1971) and an
Fig. 1 Geographic position of the samples studied within the NorthernCalcareous Alps of Austria. 1 Weitenau, 2 Mount Höherstein Plateauand Mount Zwerchwand, 3 Mount Trisselwand, 4 Mount Rettenstein
123
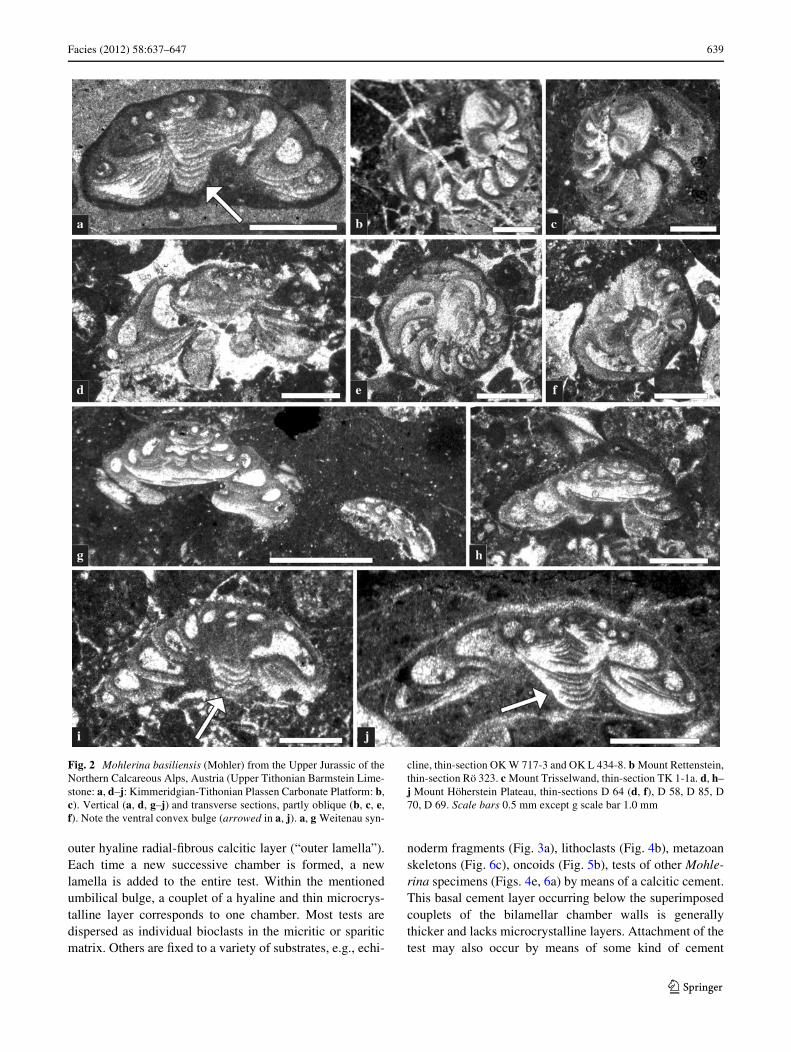
Facies (2012) 58:637–647 639
outer hyaline radial-Wbrous calcitic layer (“outer lamella”).Each time a new successive chamber is formed, a newlamella is added to the entire test. Within the mentionedumbilical bulge, a couplet of a hyaline and thin microcrys-talline layer corresponds to one chamber. Most tests aredispersed as individual bioclasts in the micritic or spariticmatrix. Others are Wxed to a variety of substrates, e.g., echi-
noderm fragments (Fig. 3a), lithoclasts (Fig. 4b), metazoanskeletons (Fig. 6c), oncoids (Fig. 5b), tests of other Mohle-rina specimens (Figs. 4e, 6a) by means of a calcitic cement.This basal cement layer occurring below the superimposedcouplets of the bilamellar chamber walls is generallythicker and lacks microcrystalline layers. Attachment of thetest may also occur by means of some kind of cement
Fig. 2 Mohlerina basiliensis (Mohler) from the Upper Jurassic of theNorthern Calcareous Alps, Austria (Upper Tithonian Barmstein Lime-stone: a, d–j: Kimmeridgian-Tithonian Plassen Carbonate Platform: b,c). Vertical (a, d, g–j) and transverse sections, partly oblique (b, c, e,f). Note the ventral convex bulge (arrowed in a, j). a, g Weitenau syn-
cline, thin-section OK W 717-3 and OK L 434-8. b Mount Rettenstein,thin-section Rö 323. c Mount Trisselwand, thin-section TK 1-1a. d, h–j Mount Höherstein Plateau, thin-sections D 64 (d, f), D 58, D 85, D70, D 69. Scale bars 0.5 mm except g scale bar 1.0 mm
123

640 Facies (2012) 58:637–647
appendages of the test margins towards the substrate andthus creating an elevated position above the latter (Fig. 3a).
Remarks Besides low trochospiral to almost discoidalforms, the Alpine material also includes high conicalforms. From the measured biometric data of the Alpinespecimens, it is apparent that the test height and the apicalangle have the widest range (Table 1). The greatestobserved test height (from the apex to the plane connect-ing the distal margins of the Wnal whorl) is about threetimes the lowest measured value. The highest apical angle(150°) is nearly double the lowest value (77°). Rememberthat the apical angle of Mohler’s holotype (115°) lies inbetween these values. Recently, a second representativeof Mohlerina was described as Mohlerina balcanica byDragastan and Richter (2003) from the Upper Valangin-ian of Greece. M. balcanica is reported to diVer fromM. basiliensis by the “shape of the test, high conical tro-chospiral, by the deep concave umbilical side, more coil-ing spirals (6–7) and by the robust and thick walls withinner and outer lamellae” (Dragastan and Richter 2003,p. 91). Given that the high test variability was most proba-bly controlled by its specialized mode of life within diVer-
ent habitats combined with specimens of diVerentontogenetic stages (juvenile vs. adult), the discriminationof the species M. balcanica Dragastan and Richter is chal-lenged. The three broken specimens and one randomoblique section of M. balcanica illustrated in its originaldescription do not allow a clear assessment of the testdiameters to be made (0.55–0.8 mm according to Draga-stan and Richter 2003). Apart from this, the number ofwhorls is clearly related to the test size as concluded fromTable 1. Until a clear and statistically based analysis of amultitude of specimens that would enable an objectivespeciWc discrimination is undertaken, M. balcanica ishere considered a junior synonym of M. basiliensis. Oth-erwise, taking the above criteria, most of the Upper Juras-sic high trochospiral specimens with 6–7 coiling spiralswould belong to the Lower Cretaceous M. balcanica. Theco-existence of two distinct species within a population ina given stratigraphic interval, here the Late Jurassic, addi-tionally challenges its validity. The two specimens ofMohlerina? n. sp. reported recently by Dragastan (2011,pl. 4/17–18) from the Lower Berriasian of the EasternCarpathians, most likely represent oblique sections of
Fig. 3 Mohlerina basiliensis (Mohler) from the Upper TithonianBarmstein Limestone of the Northern Calcareous Alps, Austria (a–b),the Kimmeridgian of Romania (c), and the Tithonian of Sicily/Italy(d modiWed from Bucur et al. 1996, pl. 4/5). a, b Specimen Wxed to arandomly corroded echinoderm fragment by means of a slightly yel-lowish calcitic cement. In the present case, the cement does not appearto cover the whole ventral side, but seems to form palisades or a ring-like construction (arrows in a) in order to attain an elevated position ofthe test towards the substrate. Weitenau syncline, Osterhorn Moun-
tains, thin-section OK W 347-1. c Specimen, possibly broken from theattaching substrate. The dark, irregular micritic remains that are inclose contact with the lowermost sparitic layer (?cement, arrowed) areinterpreted as substrate remains. The outline of the ultimate chamberwhorl on the left side is marked by the white dashed line (modiWedfrom Bucur et al. 2010, pl. 62/67). d Specimen possibly attached to acompletely recrystallized basal substrate (S?) (modiWed from Bucuret al. 1996, pl. 4/5)
123
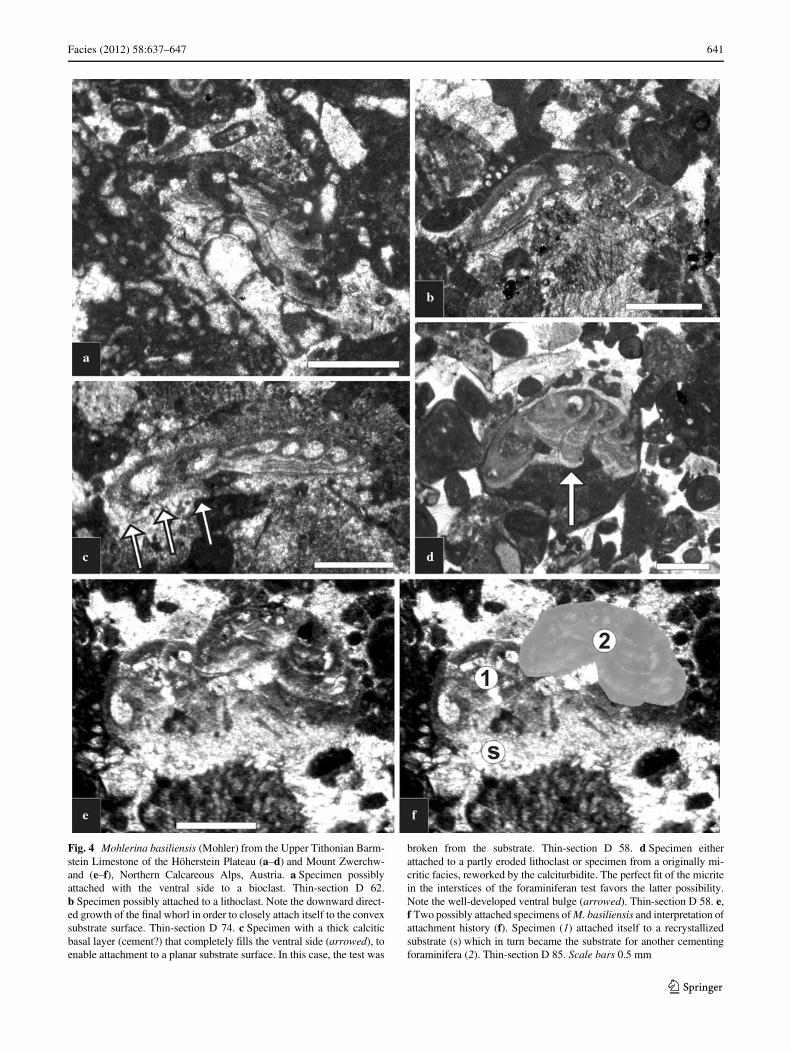
Facies (2012) 58:637–647 641
Fig. 4 Mohlerina basiliensis (Mohler) from the Upper Tithonian Barm-stein Limestone of the Höherstein Plateau (a–d) and Mount Zwerchw-and (e–f), Northern Calcareous Alps, Austria. a Specimen possiblyattached with the ventral side to a bioclast. Thin-section D 62.b Specimen possibly attached to a lithoclast. Note the downward direct-ed growth of the Wnal whorl in order to closely attach itself to the convexsubstrate surface. Thin-section D 74. c Specimen with a thick calciticbasal layer (cement?) that completely Wlls the ventral side (arrowed), toenable attachment to a planar substrate surface. In this case, the test was
broken from the substrate. Thin-section D 58. d Specimen eitherattached to a partly eroded lithoclast or specimen from a originally mi-critic facies, reworked by the calciturbidite. The perfect Wt of the micritein the interstices of the foraminiferan test favors the latter possibility.Note the well-developed ventral bulge (arrowed). Thin-section D 58. e,f Two possibly attached specimens of M. basiliensis and interpretation ofattachment history (f). Specimen (1) attached itself to a recrystallizedsubstrate (s) which in turn became the substrate for another cementingforaminifera (2). Thin-section D 85. Scale bars 0.5 mm
123
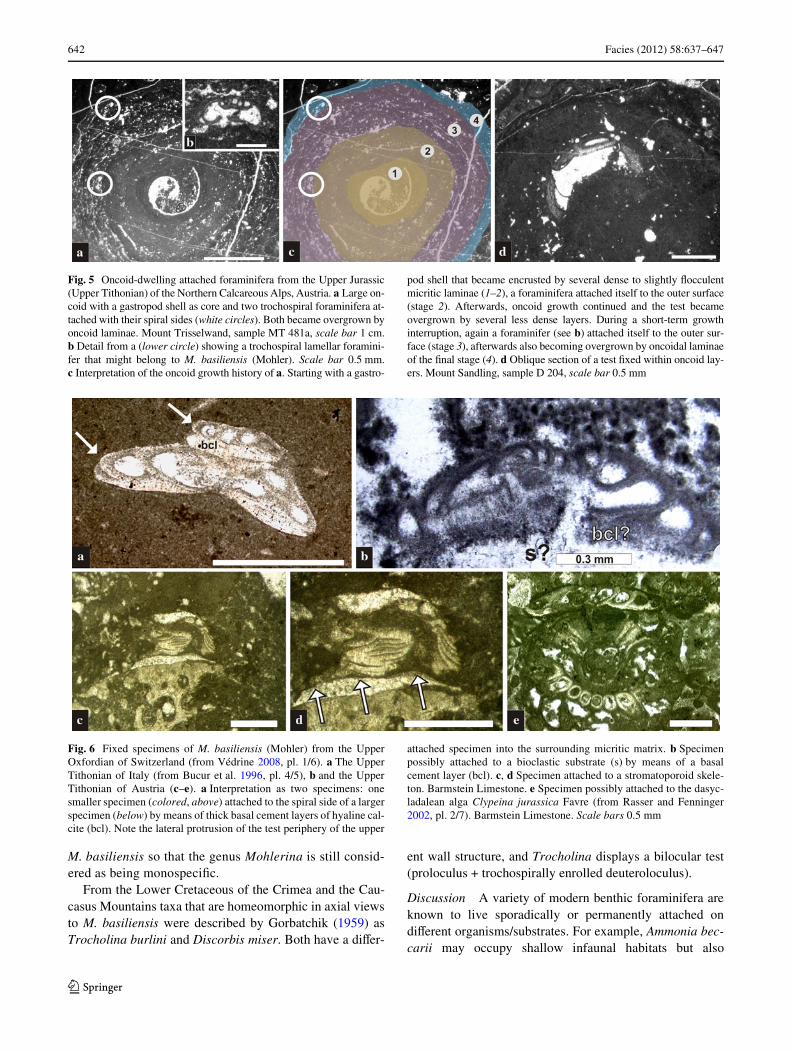
642 Facies (2012) 58:637–647
M. basiliensis so that the genus Mohlerina is still consid-ered as being monospeciWc.
From the Lower Cretaceous of the Crimea and the Cau-casus Mountains taxa that are homeomorphic in axial viewsto M. basiliensis were described by Gorbatchik (1959) asTrocholina burlini and Discorbis miser. Both have a diVer-
ent wall structure, and Trocholina displays a bilocular test(proloculus + trochospirally enrolled deuteroloculus).
Discussion A variety of modern benthic foraminifera areknown to live sporadically or permanently attached ondiVerent organisms/substrates. For example, Ammonia bec-carii may occupy shallow infaunal habitats but also
Fig. 5 Oncoid-dwelling attached foraminifera from the Upper Jurassic(Upper Tithonian) of the Northern Calcareous Alps, Austria. a Large on-coid with a gastropod shell as core and two trochospiral foraminifera at-tached with their spiral sides (white circles). Both became overgrown byoncoid laminae. Mount Trisselwand, sample MT 481a, scale bar 1 cm.b Detail from a (lower circle) showing a trochospiral lamellar foramini-fer that might belong to M. basiliensis (Mohler). Scale bar 0.5 mm.c Interpretation of the oncoid growth history of a. Starting with a gastro-
pod shell that became encrusted by several dense to slightly Xocculentmicritic laminae (1–2), a foraminifera attached itself to the outer surface(stage 2). Afterwards, oncoid growth continued and the test becameovergrown by several less dense layers. During a short-term growthinterruption, again a foraminifer (see b) attached itself to the outer sur-face (stage 3), afterwards also becoming overgrown by oncoidal laminaeof the Wnal stage (4). d Oblique section of a test Wxed within oncoid lay-ers. Mount Sandling, sample D 204, scale bar 0.5 mm
Fig. 6 Fixed specimens of M. basiliensis (Mohler) from the UpperOxfordian of Switzerland (from Védrine 2008, pl. 1/6). a The UpperTithonian of Italy (from Bucur et al. 1996, pl. 4/5), b and the UpperTithonian of Austria (c–e). a Interpretation as two specimens: onesmaller specimen (colored, above) attached to the spiral side of a largerspecimen (below) by means of thick basal cement layers of hyaline cal-cite (bcl). Note the lateral protrusion of the test periphery of the upper
attached specimen into the surrounding micritic matrix. b Specimenpossibly attached to a bioclastic substrate (s) by means of a basalcement layer (bcl). c, d Specimen attached to a stromatoporoid skele-ton. Barmstein Limestone. e Specimen possibly attached to the dasyc-ladalean alga Clypeina jurassica Favre (from Rasser and Fenninger2002, pl. 2/7). Barmstein Limestone. Scale bars 0.5 mm
123

Facies (2012) 58:637–647 643
attaches to hard substrates (e.g., Takata et al. 2010). Mostrecords are from foraminiferans that Wx their test to plants,so-called epiphytic taxa (Brasier 1975; Röttger and Krüger1990; Langer 1993; Kitazato 1994; Ribes et al. 2000;Wilson and Ramsook 2007; among many others). Epiphyticforaminifera attach to sea-grass in order to feed on diatoms,fungi, and bacteria that form a bioWlm on the plant surface(Langer 1993). Other groups attach to living sessile andvagile metazoans, e.g., crustaceans, pelecypods (e.g.,Dobson and Haynes 1973; Hayward and Haynes 1976;Zumwalt and Delaca 1980; Langer and Bagi 1994; Alexanderand DeLaca 1987). These associations of epizoic foraminif-era and invertebrates are usually suggested as representingexamples of commensalism. Among the epiphytic taxa,Langer (1993) distinguished permanently sessile (morpho-type A) and temporarily motile species (morphotype B). Inthe fossil record it is often diYcult to judge whether thespecimens were attached during the life-time of the host orwere colonizing dead bioclastic substrates.
There are diVerent ways by which these foraminiferantaxa Wx their tests to the host substrate. They can simplyattach without aVecting the substrate, pit the surface leavingattachment scars (DeLaca and Lipps 1972; Bromley andHeinberg 2006; Wisshak and Rüggeberg 2006) or evendeeply bioerode and thus penetrate the substrate (Todd1965; Vénec-Peyre 1987, 1996, with references therein).Substrate-etching foraminifera are mostly not Wxed bycement and therefore often fall oV after death. Benthic
foraminifera attach to substrates by organic Wlms/cements,e.g., epiphytic taxa, or an inorganic calcareous (aragoniticor calcitic) or siliceous cement (e.g., Delaca and Lipps1972; Lipps 1973; Poag 1982; Röttger and Krüger 1990;Langer 1992, 1993; Zampi et al. 1996). All of the arena-ceous epizoic foraminifera attach themselves by cementthat is composed of both secreted and foreign material that,in the case of agglutinating hosts, often is diYcult to dis-cern (Langer and Bagi 1994). All these attaching taxa gen-erally exhibit a rather large morphological variabilityreferring to ecophenotypic responses (e.g., Langer and Bagi1994; Holzmann and Pawlowski 2000).
Modern attached calcareous benthic foraminifera aredominated by trochospiral rotaliids, e.g., Ammonia, Cibi-cides, Discorbis, Rosalina (see Ribes et al. 2000). Theseprovide a good cross-reference source for comparisons withthe Late Jurassic-Early Cretaceous Mohlerina. It can beassumed that in quiet-water lagoonal wackestones, speci-mens of Mohlerina cemented to any kind of available hardsubstrate (e.g., pelecypod shells, stromatoporoid skeletons,oncoids etc.) would have a much better potential for preser-vation in life position than in high-energy settings. In thelatter case, attached specimens are likely to be rare due tophysical taphonomic degradation (e.g., breakage). Thisassumption, however, would indicate that there was no hab-itat preference for the attaching strategy. Therefore it isunknown to what extent specimens of Mohlerina were orig-inally cemented or not and whether, for instance, the sessile
Fig. 7 a Schematic drawing of a Paleozoic Tetrataxis specimen raisedabove the substrate with formation of a micro-aquarium between thetest and the substrate (modiWed from Vachard et al. 2010, Fig. 12/14)(without scale). Arrows in a–d indicate water Xow direction.b Attached tetrataxid foraminifer from the Carboniferous of England.
Note the basal pod of sparry calcite (arrowed) (slightly modiWed fromCossey and Mundy 1990, Fig. 3b). c Specimen of Tetrataxis exhibitinga micro-aquarium. Early Carboniferous of Montagne Noir, S-France(Pille 2008). d Specimen of M. basiliensis (Mohler) exhibiting a mi-croaquarium. Upper Jurassic of Northern Calcareous Alps, Austria
123
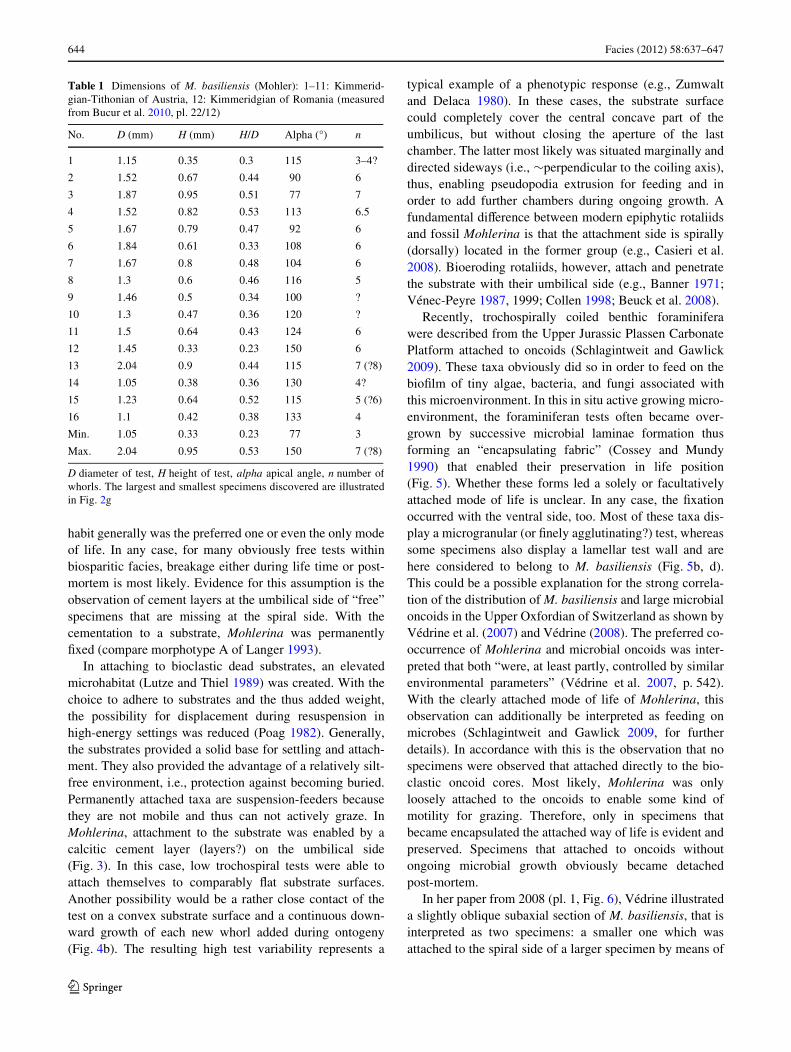
644 Facies (2012) 58:637–647
habit generally was the preferred one or even the only modeof life. In any case, for many obviously free tests withinbiosparitic facies, breakage either during life time or post-mortem is most likely. Evidence for this assumption is theobservation of cement layers at the umbilical side of “free”specimens that are missing at the spiral side. With thecementation to a substrate, Mohlerina was permanentlyWxed (compare morphotype A of Langer 1993).
In attaching to bioclastic dead substrates, an elevatedmicrohabitat (Lutze and Thiel 1989) was created. With thechoice to adhere to substrates and the thus added weight,the possibility for displacement during resuspension inhigh-energy settings was reduced (Poag 1982). Generally,the substrates provided a solid base for settling and attach-ment. They also provided the advantage of a relatively silt-free environment, i.e., protection against becoming buried.Permanently attached taxa are suspension-feeders becausethey are not mobile and thus can not actively graze. InMohlerina, attachment to the substrate was enabled by acalcitic cement layer (layers?) on the umbilical side(Fig. 3). In this case, low trochospiral tests were able toattach themselves to comparably Xat substrate surfaces.Another possibility would be a rather close contact of thetest on a convex substrate surface and a continuous down-ward growth of each new whorl added during ontogeny(Fig. 4b). The resulting high test variability represents a
typical example of a phenotypic response (e.g., Zumwaltand Delaca 1980). In these cases, the substrate surfacecould completely cover the central concave part of theumbilicus, but without closing the aperture of the lastchamber. The latter most likely was situated marginally anddirected sideways (i.e., »perpendicular to the coiling axis),thus, enabling pseudopodia extrusion for feeding and inorder to add further chambers during ongoing growth. Afundamental diVerence between modern epiphytic rotaliidsand fossil Mohlerina is that the attachment side is spirally(dorsally) located in the former group (e.g., Casieri et al.2008). Bioeroding rotaliids, however, attach and penetratethe substrate with their umbilical side (e.g., Banner 1971;Vénec-Peyre 1987, 1999; Collen 1998; Beuck et al. 2008).
Recently, trochospirally coiled benthic foraminiferawere described from the Upper Jurassic Plassen CarbonatePlatform attached to oncoids (Schlagintweit and Gawlick2009). These taxa obviously did so in order to feed on thebioWlm of tiny algae, bacteria, and fungi associated withthis microenvironment. In this in situ active growing micro-environment, the foraminiferan tests often became over-grown by successive microbial laminae formation thusforming an “encapsulating fabric” (Cossey and Mundy1990) that enabled their preservation in life position(Fig. 5). Whether these forms led a solely or facultativelyattached mode of life is unclear. In any case, the Wxationoccurred with the ventral side, too. Most of these taxa dis-play a microgranular (or Wnely agglutinating?) test, whereassome specimens also display a lamellar test wall and arehere considered to belong to M. basiliensis (Fig. 5b, d).This could be a possible explanation for the strong correla-tion of the distribution of M. basiliensis and large microbialoncoids in the Upper Oxfordian of Switzerland as shown byVédrine et al. (2007) and Védrine (2008). The preferred co-occurrence of Mohlerina and microbial oncoids was inter-preted that both “were, at least partly, controlled by similarenvironmental parameters” (Védrine et al. 2007, p. 542).With the clearly attached mode of life of Mohlerina, thisobservation can additionally be interpreted as feeding onmicrobes (Schlagintweit and Gawlick 2009, for furtherdetails). In accordance with this is the observation that nospecimens were observed that attached directly to the bio-clastic oncoid cores. Most likely, Mohlerina was onlyloosely attached to the oncoids to enable some kind ofmotility for grazing. Therefore, only in specimens thatbecame encapsulated the attached way of life is evident andpreserved. Specimens that attached to oncoids withoutongoing microbial growth obviously became detachedpost-mortem.
In her paper from 2008 (pl. 1, Fig. 6), Védrine illustrateda slightly oblique subaxial section of M. basiliensis, that isinterpreted as two specimens: a smaller one which wasattached to the spiral side of a larger specimen by means of
Table 1 Dimensions of M. basiliensis (Mohler): 1–11: Kimmerid-gian-Tithonian of Austria, 12: Kimmeridgian of Romania (measuredfrom Bucur et al. 2010, pl. 22/12)
D diameter of test, H height of test, alpha apical angle, n number ofwhorls. The largest and smallest specimens discovered are illustratedin Fig. 2g
No. D (mm) H (mm) H/D Alpha (°) n
1 1.15 0.35 0.3 115 3–4?
2 1.52 0.67 0.44 90 6
3 1.87 0.95 0.51 77 7
4 1.52 0.82 0.53 113 6.5
5 1.67 0.79 0.47 92 6
6 1.84 0.61 0.33 108 6
7 1.67 0.8 0.48 104 6
8 1.3 0.6 0.46 116 5
9 1.46 0.5 0.34 100 ?
10 1.3 0.47 0.36 120 ?
11 1.5 0.64 0.43 124 6
12 1.45 0.33 0.23 150 6
13 2.04 0.9 0.44 115 7 (?8)
14 1.05 0.38 0.36 130 4?
15 1.23 0.64 0.52 115 5 (?6)
16 1.1 0.42 0.38 133 4
Min. 1.05 0.33 0.23 77 3
Max. 2.04 0.95 0.53 150 7 (?8)
123

Facies (2012) 58:637–647 645
comparable thick basal cement layers. The test periphery ofthe attaching specimens is clearly distinguished by its lat-eral protrusion into the surrounding micritic matrix. Theseexamples clearly display that Mohlerina was also attachingin quiet water, lagoonal settings. Besides the substrateavailability, the reason for the attachment to other forami-niferan tests is not evident.
There are other examples of in situ preserved attachedforaminifera that were generally considered to representfree living forms (Fig. 7). Loosely attached Tetrataxis spec-imens were described by Cossey and Mundy (1990) fromthe Carboniferous of England. These forms were observedWxed to corals by their umbilical surfaces by means of acalcareous pod (Fig. 7b), and that later on became encapsu-lated in life position by the subsequent overgrowth ofmicrobial laminae. The diVerent modes of life of Tetrataxiswere summarized by Vachard et al. (2010, Fig. 12). A spe-cial case is the interposition of a so-called micro-aquarium(=“corridor” in Vachard and Krainer 2001, Fig. 8) betweenthe foraminiferan test and substrate (Vachard et al. 2010)(Fig. 7a, c), observed also in one specimen of M. basiliensis(Fig. 7d). Here, the raising of the test above the substratetook place by means of marginal cement appendages. Thephylogenetic relationship between Tetrataxis and Mohle-rina, both with two-layered walls is unknown. Tetrataxispossesses an outer dark microgranular layer and a hyalineWbrous inner layer, whereas in Mohlerina it is just the otherway round. Triassic forms assigned to Tetrataxis are said tohave an agglutinated wall (e.g., Fugagnoli 1996) and there-fore “are unrelated to the … genus” Tetrataxis (Loeblichand Tappan 1988, p. 249), meaning that they must belongto another taxon.
Another example includes the calcareous trochospiralforaminifer Trocholina blaui described by Senowbari-Daryan et al. (2010) from the Late Triassic of Iran. Thisspecies lived with its umbilical side attached to the thalli ofsolenoporacean algae but may occur also free within themicritic matrix. An attachment also with the umbilical sideto basal substrates is reported from trochospiral arenaceoustaxa such as Cretaceous dictyopsellids (Arnaud-Vanneau1980, pl. 66/67). Some arenaceous crust-forming taxa suchas the Cretaceous Bdelloidina Carter develop an initial tro-chospiral part in which the aperture is interio-marginal(instead of multiple terminal in the rectilinear portion)(Hofker 1965; Frieg and Kaever 1975). This initial part alsoattached itself with the umbilical side to the substrate.
Conclusions
Mohlerina basiliensis (Mohler) is a trochospirally coiledrotaliid foraminifer that may exhibit a motile or attachedmode of life in both high- and low-energy depositional set-
tings. Due to taphonomic uncertainties, it is unknownwhether the attachment actually was the preferred (oronly?) mode of life. Fixation to bioclastic substrates tookplace by means of a basal calcitic cement layer or marginalappendages creating an elevated test position. In the lattercase, a free space remains between the test and substrate, aso-called micro-aquarium, reported also from Late Paleo-zoic Tetrataxis. When M. basiliensis attached itself tomicrobial crusts (oncoids), a cement layer is not detectableso that only a loose contact can be assumed. This might bean adaptation to maintain a certain kind of motility in orderto feed on microbes.
Mohlerina can be compared with modern trochospiralrotaliids living solely or facultatively attached, e.g., manyepiphytic taxa. In contrast to these forms, Mohlerina wasattached by its umbilical side, whereas the major part ofmodern rotaliids Wxes themselves to the substrate with theirspiral sides. Other than in cases where tests were attachedto oncoid surfaces, that later became overgrown by micro-bial laminae, there are no indications of a preferential sub-strate selection.
The genus Mohlerina Bucur, Senowbari-Daryan andAbate is considered monospeciWc with the single speciesM. basiliensis (Mohler). The Late Jurassic populations withadult and juvenile specimens, perhaps including also sexualand asexual generations, coupled with a high intraspeciWcvariability (morphospecies) connected to diVerent habitats,e.g., attached or (periodically) free, also include morpho-logically similar Lower Cretaceous forms described asMohlerina balkanica Dragastan and Richter. The high testvariability can therefore be considered as a typical exampleof an ecophenotypic response.
Acknowledgments Most of the thin-sections used in this study werekindly provided by Hans-Jürgen Gawlick and Oliver Krische (bothUniversity of Leoben). Thanks to Ioan Bucur (Romania), Patrick Cos-sey (StaVordshire), David Mundy (Calgary), Michael Rasser (Stutt-gart) and Stéphanie Védrine (Orléans) for providing photographs oftheir previous works and the kind permission to use them. The Wgurefrom the PhD thesis of Pille (2008) was provided by Daniel Vachard(Lille) who also gave the permission for illustration. I am also gratefulto Mike Kaminski (Dharan) for helping with some literature andreviewing the paper. The helpful remarks of the second reviewerAntonietta Cherchi (Cagliari) are also acknowledged. Patrick Cossey(StaVordshire) is additionally thanked for critically reading the manu-script and improving the English.
References
Alexander SP, DeLaca TE (1987) Feeding adaptions of the foraminif-eran Cibicides refulgens living epizoically and parasitically on theAntarctic scallop Adamussium colbecki. Biol Bull 173:136–159
Arnaud-Vanneau A (1980) L’Urgonien du Vercors septentrional et dela Chartreuse. Géol Alpine Mém 11(3):1–874
Auer M, Suzuki S, Schlagintweit F, Gawlick HJ (2009) Spatial andtemporal development of siliceous basin and shallow-water
123

646 Facies (2012) 58:637–647
carbonate sedimentation in Oxfordian Northern Calcareous Alps.Facies 55:63–87
Banner FT (1971) A new genus of the Planorbulinidae an endoparasiteof another foraminifer. Rev Espak Micropaleont 3:113–128
Bassoullet JP (1997) Les Grands foraminifères. In: Cariou E, Hantz-pergue P (coords) Biostratigraphie du Jurassique ouest-européenet Méditerranéen: zonations paralléles et distribution et microfos-siles. Bull Centres Rech Explor-Prod Elf-Aquitaine Mém 17:293–304
Bernier P (1984) Les formations carbonatées du Kimmeridgien et duPortlandien dans le Jura méridional. Stratigraphie, micropaléon-tologie, sédimentologie. Doc Lab Geol 92:1–803
Beuck L, Correa ML, Freiwald A (2008) Biogeographical distributionof Hyrrokkin (Rosalinidae, Foraminifera) and its host-speciWcmorphological and textural trace variability. In: Wisshak M,Tapanila L (eds) Current developments in bioerosion. Springer,Berlin, pp 329–360
Brasier MD (1975) Ecology of recent sediment-dwelling and phytalforaminifera from the lagoons around Barbuda, West Indies.J Foram Res 5:193–210
Bromley RG, Heinberg C (2006) Attachment strategies of organismson hard substrates: a palaeontological view. Palaeogeogr Palaeo-climatol Palaeoecol 232:429–453
Bucur II, Senowbari-Daryan B, Abate B (1996) Remarks on someforaminifera from the Upper Jurassic (Tithonian) reef limestoneof Madonie Mountains (Sicily). Boll Soc Paleont Ital 35:65–80
Bucur II, S8s8ran E, Balica C, Beleo D, Bruchental C, Chendeo C,Chendeo O, Hosu A, Laz8r DF, L8p8dat A, Marian AV, MircescuC, Turi V, Ungureanu R (2010) Mesozoic carbonate depositsfrom some areas of the Romanian Carpathians—case studies.Cluj Univ Press, Cluj Napoca, p 203
Casieri S, Frezza V, Mancini S, Carboni MG (2008) Living sessile epi-phytic foraminifera from Posidonia oceanica meadows of Ischiaand Ponza Islands (Tyrrhenian Sea, Italy). Giornate di Paleonto-logia VIII edizione—Simp Soc Paleont Italiana, Siena, 9–13Settembre 2008, Riassunti dei lavori, pp 15–17
Collen JD (1998) Metarotaliella tuvaluensis sp. nov. from FunafutiAtoll, western PaciWc Ocean: relationship to miliolid foraminif-era. J Foram Res 28:66–75
Cossey PJ, Mundy DJC (1990) Tetrataxis: a loosely attached limpet-like foraminifer from the Upper Palaeozoic. Lethaia 23:311–322
Delaca TE, Lipps JH (1972) The mechanism and adaptive signiWcanceof attachment and substrate pitting in the foraminiferan Rosalinaglobularis d’Orbigny. J Foram Res 2:68–72
Delage Y, Hérouard E (1986) Traité de Zoologie Concrète, vol 1, LaCellule et les Protozoaires. Schleicher Frères, Paris, p 584
Dobson M, Haynes J (1973) Association of foraminifera with hydroidson the deep shelf. Micropaleontology 19:78–90
Dragastan O (2011) Early Cretaceous foraminifera, algal nodules andcalpionellids from the Lopoo Valley, Bicaz Gorges (Eastern Car-pathians, Romania). Anal Stiint Univ “Al. I. Cuza” din Iasi sergeol 57:91–113
Dragastan O, Richter DK (2003) Calcareous algae and foraminifersfrom Neocomian limestones of Methana Peninsula, AsprovouniMts. (Greece) and from south Dobrogea (Romania). Ann UnivBucuresti Geol Spec Publ 1:57–128
Ehrenberg CG (1838) Über dem blossen Auge unsichtbare Kalkthier-chen und Kieselthierchen als Hauptbestandteile der Kreidegebir-ge. Bericht über die zu Bekanntmachung geeignetenVerhandlungen königl preuss Akad Wiss Berlin, pp 192–200
Eichwald CE (1830) Zoologia specialis, vol 2. Vilnae, p 233Feldman HR, Brett CE (1998) Epi- and endo-biontic organisms on
Late Jurassic crinoid columns from the Negev Desert, Israel:implications for co-evolution. Lethaia 31:57–71
Fenninger A, Hötzl H (1967) Die Mikrofauna und -Xora des Plassen-und Tressensteinkalkes der Typlokalität (Nördliche Kalkalpen).N Jb Geol Paläont Abh 128:1–37
Foresi LM, Zampi M, Focardi S (2004) Test morphology and organiclayer of Discorinopsis aguayoi (Bermúdez): relationships withenvironmental conditions. Rev Espak Micropaleont 36:323–347
Frieg C, Kaever M (1975) Bau und systematische Stellung von Bdel-loidina cribrosa (Reuss 1846). N Jb Geol Paläont Mh 1975:449–463
Fugagnoli A (1996) On the occurrence of Duotaxis metula Kristan(foraminifera) in the Lower Jurassic (Calcari Grigi, Venetian Pre-alps, Italy). Rev Paléobiol 15:385–392
Gawlick HJ, Schlagintweit F, Missoni S (2005) Die Barmsteinkalkeder Typlokalität nordwestlich Hallein (hohes Tithonium bis tie-feres Berriasium; Salzburger Kalkalpen) - Sedimentologie,Mikrofazies, Stratigraphie und Mikropaläontologie: neueAspekte zur Interpretation der Entwicklungsgeschichte der Ober-Jura-Karbonatplattform und der tektonischen Interpretation derHallstätter Zone von Hallein-Bad Dürrnberg. N Jb Geol PaläontAbh 236:351–421
Gawlick HJ, Schlagintweit F, Suzuki H (2007) Die Ober-Jura bisUnter-Kreide Schichtfolge des Gebietes Sandling-Höherstein(Salzkammergut, Österreich) - Implikationen zur Rekonstruktiondes Block-Puzzles der zentralen Nördlichen Kalkalpen, derGliederung der karbonatklastischen RadiolaritXyschbecken undder Entwicklung der Plassen-Karbonatplattform. N Jb GeolPaläont Abh 243:1–70
Gawlick HJ, Missoni S, Schlagintweit F, Suzuki H (2010) TiefwasserBeckengenese und Initiierung einer Karbonatplattform im Jurades Salzkammergutes (Nördliche Kalkalpen, Österreich).Exkursionsführer PANGEO 2010. J Alp Geol 52:101–171
Gorbatchik TN (1959) New foraminifera from the Lower Cretaceousof the Crimea and NE Caucasus. Paleont J 1:78–83 (in Russian)
Hansen HJ, Reiss Z (1971) Electron microscopy of rotaliacean wallstructures. Bull Geol Soc Den 20:329–346
Hansen HJ, Revets SA (1992) A revision and classiWcation of the Disc-orbidae, Rosalinidae and Rotaliidae. J Foram Res 22:166–180
Hayward JJB, Haynes JR (1976) Chlamys opercularis (Linneaus) as amobile substrate for foraminifera. J Foram Res 6:30–38
Hofker J (1965) Arenaceous foraminifera attached to the walls of holesin the hard grounds of the Lower Md in the quarry Curfs. Natu-urhist Maandbl 54:29–32
Holzmann M, Pawlowski J (2000) Taxonomic relationships in the ge-nus Ammonia (foraminifera) based on ribosomed DNA sequenc-es. J Micropaleont 19:85–95
Kitazato H (1994) Foraminiferal microhabitats in four marine environ-ments around Japan. Mar Micropaleont 24:29–41
Krische O, Kurz W, Gawlick HJ (2011) The Weitenau syncline: not auniform Late Jurassic-Early Cretaceous basin Wll (Salzburg, Aus-tria). In: Bornemann A, Brachert TC, Ehrmann W (eds) Sediment2011, Leipzig June 23–26, 2011, abstracts, p 51
Langer MR (1992) Biosynthesis of glycosaminoglycans in foraminif-era: a review. Mar Micropaleont 19:245–255
Langer MR (1993) Epiphytic foraminifera. Mar Micropaleont 20:235–265
Langer MR, Bagi H (1994) Tubicolous polychaetes as substrates forepizoic foraminifera. J Micropaleont 13:132
Levy A, Mathieu R, Poignant A, Rosset Moulinier M (1986) Discorbi-dae and Rotaliidae: a classiWcation to be revised. J Foram Res22:166–180
Lipps JH (1973) Test structure in foraminifera. Ann Rev Microbiol27:471–486
Loeblich AR, Tappan H (1988) Foraminiferal genera and their classi-Wcation, vols 1 + 2. Van Nostrand Reinhold, New York
123

Facies (2012) 58:637–647 647
Lutze GF, Thiel H (1989) Epibenthic foraminifera from elevatedmicrohabitats: Cibicidoides wuellerstorW and Planulina arimin-ensis. J Foram Res 19:153–158
Mohler W (1938) Mikropaläontologische Untersuchungen in der nord-schweizerischen Juraformation. Abh Schweiz Paläont Ges 60:1–53
Pille L (2008) Foraminifères et algues calcaires du Mississippien sup-érieur (Viséen supérieur-Serpukhovien): rôles biostratigraphique,paléoécologique et paléogéographique aux échelles locale, régio-nale et mondiale. Unpublished Thèse Univ Lille, vol 3, pp 1–226,pp 1–255, 72 pls
Plöchinger B (1984) Zum Nachweis jurassisch-kretazischer Eingleit-ungen von Hallstätter Gesteinsmassen beiderseits des Salzach-Quertals (Salzburg). Geol Rdsch 73:293–304
Poag CW (1982) Environmental implications of test-to-substrate attac-hement among some modern sublittoral foraminifera. GSA Bull93:252–268
Quilty PG (2011) Late Jurassic foraminifera, Wallaby Plateau, oVshoreWestern Australia. J Foram Res 41:182–195
Rasser M, Fenninger A (2002) Jurassic/Cretaceous dasycladalean algalstratigraphy in the Northern Calcareous Alps: a critical reviewand a palaeobiogeographic approach using similarity indices. In:Bucur II, Filipescu S (eds) Research advances in calcareous algaeand microbial carbonates. Proceedings of the 4th internationalIFAA Reg Meet Cluj-Napoca, Cluj University Press, pp 167–190
Reiss Z (1963) ReclassiWcation of perforate foraminifera. Bull GeolSurv Israel 35:1–111
Revets SA (2002) The genus Albertinopsis, gen. nov. and the relation-ships between gavelinellidae and rosalinidae. J Foram Res 32:51–56
Ribes T, Salvadó HJR, Del Pilar Garcia A (2000) Foraminiferal colo-nization on artiWcial sea grass leaves. J Foram Res 30:192–201
Röttger R, Krüger R (1990) Observations on the biology of Calcarini-dae (Foraminiferida). Mar Biol 106:419–425
Schlagintweit F, Ebli O (1999) New results on microfacies, biostratig-raphy and sedimentology of Late Jurassic-Early Cretaceous plat-form carbonates of the Northern Calcareous Alps. Part I:Tressenstein limestone, plassen-formation. Abh Geol BA56:379–418
Schlagintweit F, Gawlick HJ (2009) Oncoid-dwelling foraminiferafrom Late Jurassic shallow-water carbonates of the Northern Cal-careous Alps (Austria and Germany). Facies 55:259–266
Schlagintweit F, Gawlick HJ, Lein R (2005) Mikropaläontologie undBiostratigraphie der Plassen-Karbonatplattform der Typlokalität
(Ober-Jura bis Unter-Kreide, Salzkammergut, Österreich).J Alpine Geol (Mitt Ges Geol Bergbaustud Österr) 47:11–102
Senowbari-Daryan B, Rashidi K, Torabi H (2010) Foraminifera andtheir associations of a possibly Rhaetian section of the NaybandFormation in central Iran, northeast of Esfahan. Facies 56:567–596
Steiger T (1981) Kalkturbidite im Oberjura der Nördlichen Kalkalpen(Barmstein Kalke, Salzburg, Österreich). Facies 4:215–348
Steiger T, Wurm D (1980) Faziesmuster oberjurassischer Plattform-Karbonate (Plassenkalke, Nördliche Kalkalpen, Steirisches Salz-kammergut, Österreich). Facies 2:241–284
Takata H, Seto K, Kurata K, Hiratsuka J, Khim BK (2010) Life historyof Ammonia “beccarii” forma 1 on hard substrate in the OhashiRiver, southeastern Japan. Fundam Appl Limnol 178:81–88
Todd R (1965) A new Rosalina (foraminifera) parasitic on a bivalve.Deep Sea Res 12:831–837
Vachard D, Krainer K (2001) Smaller foraminifers of the UpperCarboniferous from the Auernig Group, Carnic Alps (Austria/Italy). Riv Ital Paleont Stratigr 107:147–168
Vachard D, Pille L, Gaillot J (2010) Palaeozoic foraminifera: system-atics, palaeoecology and responses to global changes. Rev Micro-paléont 53:209–254
Védrine S (2008) Co-occurrence of the foraminifer Mohlerina basili-ensis with Bacinella-Lithocodium oncoids: palaeoenvironmentaland palaeoecological implications (Late Oxfordian, Swiss Jura).J Micropaleont 27:35–44
Védrine S, Strasser A, Hug W (2007) Oncoid growth and distributioncontrolled by sea-level Xuctuations and climate (Late Oxfordian,Swiss Jura Mountains). Facies 53:535–552
Vénec-Peyré MT (1987) Boring foraminifera in French Polynesiancoral reefs. Coral Reefs 5:205–212
Vénec-Peyré MT (1996) Bio-eroding foraminifera: a review. MarMicropaleont 28:19–30
Wilson B, Ramsook A (2007) Population densities and diversitiesof epiphytal foraminifera on nearshore substrates, Nevis, WestIndies. J Foram Res 37:213–222
Wisshak M, Rüggeberg A (2006) Colonisation and bioerosion ofexperimental substrates by benthic foraminiferans from euphoticto aphotic depths (Kosterfjord, SW Sweden). Facies 52:1–17
Zampi M, Benocci S, Focardi S (1996) Epibiont foraminifera ofSertella frigida (Waters) (Bryozoa, Cheilostomata) from TerranovaBay, Ross Sea, Antarctica. Polar Biol 17:363–370
Zumwalt GS, Delaca TE (1980) Utilization of brachiopod feedingcurrents by epizoic foraminifera. J Paleont 54:477–484
123
Related Documents











