INSTITUTE OF PHYSICS PUBLISHING NETWORK: COMPUTATION IN NEURAL SYSTEMS Network: Comput. Neural Syst. 14 (2003) 703–732 PII: S0954-898X(03)55346-5 Modulation of excitatory synaptic coupling facilitates synchronization and complex dynamics in a biophysical model of neuronal dynamics Michael Breakspear 1,2 , John R Terry 3 and Karl J Friston 4 1 Brain Dynamics Centre, Westmead Hospital, Westmead, NSW, 2145, Australia 2 School of Physics, A28, University of Sydney, NSW, 2006, Australia 5 3 Department of Mathematical Sciences, Loughborough University, Loughborough, UK 4 Wellcome Department of Imaging Neuroscience, University College London, UK E-mail: [email protected] Received 17 October 2002, accepted for publication 4 July 2003 Published 8 August 2003 Online at stacks.iop.org/Network/14/703 Abstract In this paper, complex dynamical synchronization in a non-linear model of a neural system is studied, and the computational significance of the behaviours is explored. The local neural dynamics is determined by voltage- and ligand- gated ion channels and feedback between densely interconnected excitatory and inhibitory neurons. A mesoscopic array of local networks is modelled by introducing coupling between the local networks via weak excitatory- to-excitatory connectivity. It is shown that with modulation of this long- range synaptic coupling, the system undergoes a transition from independent oscillations to stable chaotic synchronization. Between these states exists a ‘weakly’ stable state associated with complex, intermittent behaviour in the temporal domain and clusters of synchronous regions in the spatial domain. The paper concludes with a discussion of the putative relevance of such processes in the brain, including the role of neuromodulatory systems and the mechanisms underlying sensory perception, adaptation, computation and complexity. 1. Introduction The mechanisms and function of cooperative behaviour in large-scale neuronal systems currently form a very active area of research. Synchronous oscillations between neurons have been proposed as a mechanism for perceptual ‘binding’ through the coupling of distinct neuronal populations to form dynamic cell assemblies, each encoding various aspects of a perceived object (Singer 1995). Evidence of coherent oscillations has been demonstrated in neurophysiological data (e.g. Eckhorn et al 1988, Gray et al 1989, Engel et al 1991, Rodriguez 5 Address for correspondence. 0954-898X/03/040703+30$30.00 © 2003 IOP Publishing Ltd Printed in the UK 703

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

INSTITUTE OF PHYSICS PUBLISHING NETWORK COMPUTATION IN NEURAL SYSTEMS
Network Comput Neural Syst 14 (2003) 703ndash732 PII S0954-898X(03)55346-5
Modulation of excitatory synaptic coupling facilitatessynchronization and complex dynamics in abiophysical model of neuronal dynamics
Michael Breakspear12 John R Terry3 and Karl J Friston4
1 Brain Dynamics Centre Westmead Hospital Westmead NSW 2145 Australia2 School of Physics A28 University of Sydney NSW 2006 Australia5
3 Department of Mathematical Sciences Loughborough University Loughborough UK4 Wellcome Department of Imaging Neuroscience University College London UK
E-mail mbreakphysicsusydeduau
Received 17 October 2002 accepted for publication 4 July 2003Published 8 August 2003Online at stacksioporgNetwork14703
AbstractIn this paper complex dynamical synchronization in a non-linear model of aneural system is studied and the computational significance of the behavioursis explored The local neural dynamics is determined by voltage- and ligand-gated ion channels and feedback between densely interconnected excitatoryand inhibitory neurons A mesoscopic array of local networks is modelledby introducing coupling between the local networks via weak excitatory-to-excitatory connectivity It is shown that with modulation of this long-range synaptic coupling the system undergoes a transition from independentoscillations to stable chaotic synchronization Between these states exists alsquoweaklyrsquo stable state associated with complex intermittent behaviour in thetemporal domain and clusters of synchronous regions in the spatial domain Thepaper concludes with a discussion of the putative relevance of such processes inthe brain including the role of neuromodulatory systems and the mechanismsunderlying sensory perception adaptation computation and complexity
1 Introduction
The mechanisms and function of cooperative behaviour in large-scale neuronal systemscurrently form a very active area of research Synchronous oscillations between neuronshave been proposed as a mechanism for perceptual lsquobindingrsquo through the coupling of distinctneuronal populations to form dynamic cell assemblies each encoding various aspects of aperceived object (Singer 1995) Evidence of coherent oscillations has been demonstrated inneurophysiological data (eg Eckhorn et al 1988 Gray et al 1989 Engel et al 1991 Rodriguez
5 Address for correspondence
0954-898X03040703+30$3000 copy 2003 IOP Publishing Ltd Printed in the UK 703
704 M Breakspear et al
et al 1999 Haig et al 2000 Tallon-Baudry et al 2001) and computational models of the cortex(eg Lumer et al 1997a 1997b Robinson et al 1998) However in much of this researchonly linear measures of synergistic activity such as the coherence function are employedThe non-linear properties of excitable membranes motivate the study of non-linear aspectsof synchronicity in neural systems The development of computing resources and non-lineartechniques renders such studies increasingly feasible and may offer a meaningful contributionto the understanding of brain function
In this paper we study non-linear interdependence in a model neural system consisting ofan array of coupled small-scale neural subsystems Each subsystem is constituted by denselyinterconnected excitatory and inhibitory neurons These are then coupled together throughweak long-range excitatory projections to form a larger (lsquomesoscopicrsquo) array The dynamicswithin and between the subsystems derives from the properties of voltage-gated ion channelsand competition at post-synaptic ligand-gated channels The properties of the voltage-gatedchannels confer non-linear properties to the dynamics We study the types and stability ofnon-linear synchronization and consider the possible role of these behaviours in adaptive brainfunction
The rest of the paper is structured as follows In the next section we develop the evolutionequations for the neural dynamics firstly in local networks and subsequently across the entirearray In section 3 the local dynamics (in the absence of long-range coupling) of the systemis explored In section 4 the effect of increasing the strength of long-range coupling on theemergent cooperative dynamics is studied Section 5 focuses on the complex behaviours thatare observed in the presence of marginal stability of the synchronous dynamics In the finalsection the possible computational significance of the observed behaviours is discussed
2 Evolution equations
The approach adopted in this study is to consider the behaviour of local ensembles ofneurons with dynamical variables and parameters representing local ensemble averagesThe scale of each ensemble is taken as the extension of the pyramidal cell dendritic arbors(the size of the cortical columns) which is approximately 100ndash300 microm This approach ismotivated by the observations that neurons within columns tend to share the same physiologicalproperties exhibit dense reciprocal interconnectivity and show strong dynamical correlationsin comparison to neurons in different columns (Szentagothai 1983 Nunez 1995 Douglas et al1996 Mountcastle 1997 1998) For these reasons they have been proposed as comprisingthe brainrsquos lsquoelementary functional modulesrsquo (Kandel et al 2000) The next-largest spatialscale of neural organization is achieved through weak long-range excitatory-to-excitatoryintercolumnar connectivity A further discussion of the neurobiological basis of this approachhas been presented in Breakspear and Terry (2002a)
21 Local ensemble dynamics
The dynamical variables studied are the mean membrane potential of local pyramidal cells V and inhibitory interneurons Z and the average number of lsquoopenrsquo potassium ion channels W The evolution equations are adapted from a study of epileptic seizures in hippocampal slices(Larter et al 1999) consisting of coupled differential equations which in turn are derived fromthe model of Morris and Lecar (1981) The main focus is on the mean cell membrane potentialof the pyramidal cells governed by the conductance of sodium potassium and calcium ionsthrough voltage-gated channels plus the passive conductance of lsquoleakyrsquo ions We begin withthe total current flow across the local pyramidal cell membranes considered as capacitors
Synchronization in a model of neuronal dynamics 705
CdV
dt= minusgCamCa(V minus VCa)minus gNamNa(V minus VNa)minus gKW (V minus VK)minus gL(V minus VL) (1)
where gion is the maximum conductance of each population of ion species if all channelsare open m ion is the fraction of channels open and Vion is the Nernst potential for that ionspecies Henceforth following Larter et al (1999) and Destexhe et al (2001) all equationsand parameters are non-dimensional and normalized to neural capacitance C = 1
Each voltage-gated channel acts as a switch which opens when the membrane potentialpasses above a given threshold Tion Hence for a single channel the relationship betweenmembrane voltage and channel conductance is given by a step function Averaging over apopulation of such channels and assuming a Gaussian distribution of Tion around a mean foreach ion species gives the sigmoid-shaped lsquoneural activation functionsrsquo
m ion = 05
(1 + tanh
(V minus Tion
δion
)) (2)
where δion incorporates the variance of this distribution As discussed in Larter et al (1999) theprincipal effect of moving from a microscopic (single-neuron) to a mesoscopic (local network)spatial frame is to further increase the variance of the threshold potentials (consistent with anincrease in their physical heterogeneity) which has the effect of flattening this curve
The fraction of open potassium channels is slightly more complicated The channelslsquorelaxrsquo from one state to another at an exponential rate This is achieved if W is governed by
dW
dt= φ(mK minus W )
τ (3)
where φ is a temperature scaling factor and τ is the lsquorelaxationrsquo time constantThe second modification required for a network formulation is introducing synaptic
interactions between neurons within the same column This is achieved by calculating theaverage cell firing rates and introducing feedback terms subsequent to cell firing to representneurotransmitter release At the cell soma the membrane potential triggers an action potential ifit exceeds a threshold determined chiefly by the sodium channel threshold potential Averagingthis over the ensemble of neurons and assuming once again a Gaussian distribution we obtainthe cell firing rates
QV = 05 times QV max
(1 + tanh
(V minus VT
δV
)) (4)
QZ = 05 times QZ max
(1 + tanh
(Z minus ZT
δZ
)) (5)
where the Qmax are the maximum rates of firing of the excitatory and inhibitory neuronsrespectively
The firing of each of these cell populations feeds back onto the ensemble through synapticcoupling to open ligand-gated channels and raise or lower the membrane potential accordinglyIn the case of excitatory-to-inhibitory and inhibitory-to-excitatory connections this is modelledas additional inputs to the flow of ions across the membrane channel weighted by functionalsynaptic factors aei and aie respectively6 In the case of excitatory-to-excitatory connectionsthe mechanism of this increase in ion conductance is modelled with greater physiological detailincorporating some of the specific characteristics of the ligand-gated channel populations Themean rate of firing QV is assumed to lead to a proportional release of glutamate neurotransmitteracross the synapse which diffuses onto two classes of ligand-gated ion channels One class
6 Note that the feedback terms are multiplied by the potential of the cell of origin This incorporates the potentiatingeffect of residual pre-synaptic intracellular calcium which increases with the pre-synaptic potential (Magleby 1987)
706 M Breakspear et al
Figure 1 A schematic representation of connectivity within cortical columns Panel (a) depictsdense local connectivity between excitatory (EN) and inhibitory (IN) neurons Panel (b) depictsparameters representing strengths of synaptic connectivity between neurons within each columnplus input from ascending neurons within the reticular formation (RF)
embodying AMPA channels open an additional population of sodium channels thus increasingthe net conductance of sodium flow The second class representing NMDA receptors openan additional population of voltage-gated calcium channels Hence this acts to increase themaximum conductance of voltage-gated calcium channels Finally lsquononspecificrsquo subcorticalexcitation is added to both populations of neurons with amplitude I modulated by a randomnoise component of amplitude δ Thus we obtain
dV
dt= minus(gCa + rNMDAaee QV )mCa(V minus VCa)minus (gNamNa + aee QV )(V minus VNa)
minus gKW (V minus VK)minus gL(V minus VL) + aie Z QZ + ane Iδ (6)dZ
dt= b(ani Iδ + aeiV QV ) (7)
where rNMDA denotes the number of NMDA receptors relative to that of AMPA receptors ane
and ani parametrize non-specific input to excitatory and inhibitory cells and aee parametrizesthe local excitatory-to-excitatory synaptic strength Note the difference in the coefficientscontrolling the effect of local excitatory feedback onto NMDA receptors compared to thosefor AMPA receptors A schematic representation of the local network and the parametrizedsynaptic connectivity is presented in figure 1
22 Mesoscopic array dynamics
Each set of equations (2)ndash(7) governs the dynamics within a local cell assembly Couplingbetween N nodes can be introduced parsimoniously as competitive agonist excitatory actionbetween pyramidal cells in different columns However these long-range projections aremodelled to target the same populations of NMDA and AMPA receptors as are targeted by theshort-range excitatory projections Representing each node with a superscript this is achievedby modifying equation (6) to
dV i
dt= minus(gCa + (1 minus C)rNMDAaee Qi
V + CrNMDAaee〈QV 〉)mCa(Vi minus VCa)
minus gKW (V i minus VK)minus gL(Vi minus VL)
minus (gNamNa + (1 minus C)aee QiV + Caee〈QV 〉)(V i minus VNa) + aie Z Qi
Z + ane Iδ (8)
Synchronization in a model of neuronal dynamics 707
where i = 1 N and 〈 〉 represents spatial (lsquomean-fieldrsquo) averaging over cell assembliesC parametrizes the strength of excitatory coupling between cortical columns If C = 0the systems evolve independently C gt 0 introduces interdependences between consecutivecolumns C = 1 corresponds to maximum coupling with excitatory input from outside eachcolumn exceeding excitatory input from within each column
The long-range coupling is lsquoweakrsquo in the sense that we investigate parameter values forwhich C lt aee and C aie etc This either implies that the functional synaptic strength of thelong-range connections are weaker than local connections or that the strengths are similar butthe density of projections is comparatively sparse We prefer the latter interpretation whichis more consistent with other modelling approaches
23 Parameter values and physiological validity
In the sections that follow we set all physiologically measurable parameters (the maximumconductances threshold potentials and Nernst potentials) to within their accepted ranges (Larteret al 1999) Other parameters such as I the functional synaptic strengths and maximum firingrates were chosen to generate dynamically plausible behaviour including the limit cycle andaperiodic dynamics The parameter values are as given in table 1 (unless otherwise stated)There may be significant variability in many of these parameters In this respect it is crucial tonote that the behaviours described below occur in a large region of parameter space Hence theemergent behaviours discussed do not only occur for the parameter values used in this studyBecause as noted above the parameters are dimensionless and normalized to a capacitance ofone it is also important to note that the time units are arbitrary Noting however that the cellcapacitance is typically estimated as close to or equal to 1 microF cmminus2 (eg Destexhe et al 2001)it is reasonable to interpret the time evolution units as milliseconds (as done for the diffusivecoupling parameter employed in Larter et al (1999))
The principal aim of this paper is a quantitative study of non-linear synchronization ina biophysically realistic model of a neural system The model incorporates some aspects ofneural systems with significant detail such as the conductance of K+ glutamate-mediatedneurotransmission and the relatively sparse nature of long-range connectivity This detailallows manipulation of parameters that model pharmacological mechanisms of interest suchas glutamate receptor subclasses Other features such as local inhibitory feedback and thekinetics of Ca2+ and Na+ channels have been incorporated in a more simplified manner Thislimits the complexity of the model but ensures that numerical analysis can be supplementedby quantitative examination of for example the Lyapunov exponents Other biophysicalproperties could be made the subject of future study by changing relevant aspects of theequations
3 Behaviour of local lsquouncoupledrsquo columns
In this section a brief overview of the behaviours exhibited by autonomous local columns(ie C = 0) is presented As indicated in table 1 the dynamics of the model is explored byfixing all parameters except for δV the variance of the excitatory threshold which is varied inthe region [055 07] A similar range of behaviours is also observed if δV is fixed and anothercontrol parameter such as aee or I is varied
31 Fixed point attractors
For values of δV lt 055 the dynamics of a single column settles onto a solitary fixed pointattractor As shown in figure 2 transient dynamics persists for a number of seconds
708 M Breakspear et al
Figure 2 The time series for a fixed point attractor with VT = 054
Table 1 Parameter values unless otherwise stated Adapted from Larter et al (1999)
Parameter Description Value
TCa Threshold value for Ca channels minus001δCa Variance of Ca channel threshold 015gCa Conductance of population of Ca channels 11VCa Ca Nernst potential 1TK Threshold value for K channels 00δK Variance of K channel threshold 030gK Conductance of population of K channels 20VK K Nernst potential minus07TNa Threshold value for Na channels 03δNa Variance of Na channel threshold 015gNa Conductance of population of Na channels 67VNa Na Nernst potential 053VL Nernst potential leak channels minus05gL Conductance of population of leak channels 05VT Threshold potential for excitatory neurons 00δV Variance of excitatory threshold VariedZT Threshold potential for inhibitory neurons 00δZ Variance of inhibitory threshold 070I Subcortical input strength 030aee Excitatory-to-excitatory synaptic strength 04aei Excitatory-to-inhibitory synaptic strength 2aie Inhibitory-to-excitatory synaptic strength 2ane Non-specific-to-excitatory synaptic strength 1ani Non-specific-to-inhibitory synaptic strength 04b Time constant scaling factor 010ϕ Temperature scaling factor 07τ Time constant for K relaxation time 1rNMDA Ratio of NMDA to AMPA receptors 025β Random modulation of subcortical input 0
32 Limit cycle attractors
For 055 lt δV lt 059 the system undergoes a series of period-doubling bifurcations Theseresult in both simple and complex periodic behaviours A limit cycle occurring when δV = 056
Synchronization in a model of neuronal dynamics 709
Figure 3 The approach to a limit cycle attractor in phase space with VT = 056
is illustrated in figure 3 Other parameter variations produce more complicated higher-orderperiodic orbits For example a period-9 orbit occurring when δV = 06 aee = 05 aie = 05gNa = 0 and I = 0165 is presented in figure 4
33 Chaotic attractors
Setting δV gt 059 and keeping all other parameters as per table 1 produces complex aperiodicbehaviour in the uncoupled columns An example of this with δV = 065 is illustrated infigure 5 Panels (a) and (b) illustrate a phase space portrait and a time series plot of thissystem respectively They show an aperiodic but highly structured system In panel (c) afirst-return lsquoPoincarersquo section is presented This was obtained by taking a cross-section throughthe attractor in (a) by setting W = 032 and then plotting V (t) against V (t +l)with a time delayof l = 1 A topographic similarity to the simple logistic map is evident It has been provenanalytically that in restricted regions of parameter space dynamical systems incorporatingthe neural activation function (4) (5) are topologically conjugate to the logistic map (Wang1991) The Poincare map presented here shows that although a 11 map may not exist in moregeneral parameter regions the global morphology of the logistic map is still evident
To determine whether or not these aperiodic oscillations are in fact chaotic the Lyapunovexponents were estimated numerically using an algorithm based upon GrammndashSchmidtorthonormalization For the particular case illustrated the exponents were found to be
λ1 = 0015
λ2 = 00
λ3 = minus026
(9)
A principal Lyapunov exponent greater than zero is consistent with chaotic dynamics Thistogether with the topography of the first-return map and the existence of period-doublingbifurcations provides compelling evidence of chaotic behaviour in the dynamics of the singlecolumn The second exponent reflects the rate of exponential separation of adjacent orbits in
710 M Breakspear et al
Figure 4 A period-9 limit cycle attractor (a) A phase space portrait (b) A representative timeseries
the direction tangent to the flow hence a value of zero is to be expected The sum of the threeexponents is negative implying that the system is volume contracting which is to be expectedin the setting of a non-equilibrium (non-Hamiltonian) physiological system
It is interesting to note that this diversity of behaviours can be obtained throughmanipulation of a single state parameter From a functional perspective this permits eachcortical column to express a variety of qualitatively distinct dynamics through modificationof a single physiological parameter such as through modulation of the functional strength ofthe local synaptic connectivity Conversely it also implies that resetting of a single parametersuch as an abnormal δV resulting from loss of physiological heterogeneity could be associatedwith qualitative changes in local cortical behaviour
4 Cooperative behaviour of coupled columns
Whilst it is interesting to observe the range of dynamic behaviours possible in a single columnwe are primarily interested in the behaviour of many such interacting columns In this section
Synchronization in a model of neuronal dynamics 711
1
Figure 5 Chaotic behaviour of the autonomous system with VT = 065 The other parameters areas per table 1 (a) An attractor in phase space (b) A time series for V (t) (c) A Poincare sectionobtained by setting W = 032 and plotting V (t) against V (t + 1)
712 M Breakspear et al
we study the emergence of cooperative behaviour that results from the introduction of couplingbetween columns Three types of interdependent behaviour of two systems are describedidentical generalized and phase synchronization Synchronous behaviours in larger arrays arealso illustrated
41 Identical chaotic synchronization
411 Transition to identical chaotic synchronization The most straightforward extensionand the one that we consider first is that of two coupled cortical columns where all parametersare in the first case assumed identical7 Trivially the two systems will evolve identicallywhen started with identical initial conditions even if C = 0 This is simply a statement ofthe deterministic nature of the evolution equations In general the random initial conditionswill not be equal in which case it is hardly surprising that the different columns will evolveindependently An example is presented in figure 6 with VT = 06 in both systems C = 0and independent random initial conditions In figure 6(a) we show a segment of the timeseries for each of the two subsystems where it can be seen that the time series are chaoticand asynchronous In figure 6(b) the concurrent values of V 1 and V 2 are plotted against eachother showing that they bear no obvious interrelationship
With the introduction of coupling C gt 0 the systems begin to show epochs ofsynchronized behaviour At C sim= 01 there is a transition to stable fully synchronizedmotion as evident in an exemplar time series shown in figure 7(a) where C = 015 Infigure 7(b) we present the concurrent values of V 1 versus V 2 illustrating that in contrast tothe case in figure 6(b) the systems have become embedded in the low-dimensional lsquosymmetrymanifoldrsquo V 1 = V 2 Panel (c) shows a log-linear plot of the temporal evolution of the distanceof the coupled system from this manifold It can be seen that on average the two systemsapproach at an exponential rate with constant time-averaged exponent Whilst this phenomenon(lsquoidentical chaotic synchronizationrsquo) has been described in a variety of other systems since itsfirst description (Fujisaka and Yamada 1983 Pecora and Carroll 1990) to our knowledge thisis its first quantitative8 description in a detailed physiological model of a neural system
The symmetry of the dynamical system obtained by setting the state parameters in eachcolumn identical forces the invariance of the symmetry manifold V 1 = V 2 Moreover thedynamics within this manifold is governed by the same equations of motion as for a singlecortical column This manifold which contains the orbits of the synchronized system is calledthe synchronization manifold The coupling parameter C does not affect the dynamics withinthe synchronization manifold and as such is termed a normal parameter
412 lsquoBlowout bifurcationrsquo An important issue is the stability of the orbits on thesynchronization manifold This can be determined by analysing the Lyapunov spectraBecause the two-column system evolves in a six-dimensional phase space there are sixLyapunov exponents However for the synchronized system it is helpful to subdivide theseinto those that describe the dynamics within the synchronization manifold and those thatdescribe the dynamics in the transverse direction The latter describe the rate of convergenceor divergence of the coupled systems toward or away from this manifoldmdashand hence thetendency for the two systems to synchronize or separate For the time series in figure 7 this is
7 Whilst it is of course physiologically unrealistic to assume that two columns would have entirely identical structuresit is mathematically simpler to start analysing the system in this situation8 A qualitative description in a neural system with a different form of coupling (potassium diffusion) is evident infigure 8 of Larter et al (1999) Synchronous chaos in an abstract model of a neural system was also described inWennekers and Pasemann (1996)
Synchronization in a model of neuronal dynamics 713
1
2
2
1
Figure 6 Independent oscillations in a two-column system with identical parameters butindependent initial conditions (a) The time series (b) A plot of concurrent mean excitatorymembrane potentials V 1 against V 2
related to the slope of the plot in panel (c)9 For this strength of coupling the largest transverseexponent is negative (minus002) indicating that perturbations away from this state of identicalsynchronization decay and thus that the state of identical chaotic synchronization is stableagainst noise (ie dynamically stable)
To calculate the relationship between coupling strength and the Lyapunov exponents weused an algorithm which measures the growth of perturbations of the variational equations (thelsquoJacobianrsquo) of the system (we do not define these equations here as they are cumbersome anddo not enhance the understanding of the system) We calculated these exponents for increasingvalues of the coupling parameter C from 0 to 02 in increments of 0000 25 The results are
9 Only two of the six dependent variables are shown in figure 7 The actual transverse Lyapunov exponents reflectthe temporal evolution of all six and hence are somewhat more complex than that given by the slope of panel (c)
714 M Breakspear et al
1
2
Figure 7 Identical synchronization in the same system as in figure 7 but with C = 02 (a) Thetime series (b) A plot of concurrent mean excitatory membrane potentials V 1 against V 2 (c) Thelogarithm of the Euclidean distance between V 1 and V 2 versus time (linear)
Synchronization in a model of neuronal dynamics 715
Figure 8 Lyapunov spectra for a coupled system with δV = 07 and C varying from 0 to 02 insteps of 0000 33 At each value of C all Lyapunov exponents are calculated Least-squares linesof best fit were then fitted to the results for each exponent Solid lines indicate transverse exponents(λ) and dashed lines indicate the tangential exponents () Panel (a) shows all three transverseand three tangential exponents Panel (b) only shows the largest two of each
illustrated in figure 8 Panel (a) shows all exponents For illustrative purposes only the mostpositive four are plotted in panel (b) In both panels the tangential exponents () are fitted (inthe least-squares sense) with a dashed line and the transverse ones (λ) are fitted with a solidline They are labelled 1ndash3 in order of decreasing positivity The important features are asfollows For the range of coupling strengths that we consider the largest tangential Lyapunovexponent is positive which indicates the presence of chaotic behaviour within the manifoldWhen examining the line of best fit for this exponent we see that this exponent does not varywhen altering the coupling parameter This is true for all three tangential exponents and verifiesthat the changes in the coupling parameter do not alter the dynamics on the synchronizationmanifold (that is the coupling parameter is normal)
716 M Breakspear et al
The largest transverse Lyapunov exponent (denoted as λ1 in the figure) is a particularlyimportant exponent since it governs the stability of the attractor in the synchronization manifoldwith respect to perturbations out of the manifold If this exponent is negative then we expectsmall perturbations of trajectories to reconverge to the manifold in finite time If however thisexponent is positive then we would instead expect perturbed trajectories to diverge from themanifold This change in local behaviour is termed a bifurcation and the point at which itoccurs the bifurcation point It is apparent in figure 8 that the zero crossing of this exponentmdashthe bifurcation pointmdashoccurs at approximately C = 01 in the present system This typeof bifurcation has been termed a blowout bifurcation since once transverse stability is losttrajectories are lsquoblown outrsquo from the low-dimensional synchronization manifold into the fullphase space (Ott and Sommerer 1994) Hence in the present setting a blowout bifurcation isobserved if C falls below 01
We note two other observations Increasing rNMDA which corresponds to increasing thepopulation of NMDA receptors decreases the coupling strength required to achieve stablesynchronization On the other hand increasing the overall excitatory-to-excitatory synapticconnectivity (aee) has the opposite effect This is because the effect of local excitatory feedbackincreases the tangential Lyapunov exponents which has the effect of lsquosplittingrsquo the synchronizeddynamics
In the above simulations all the parameters of both systems were equal In realphysiological systems such symmetry between systems is clearly impossible We nowconsider the case in which the two columns have different structures A simple way toincorporate this whilst at the same time preserving (as much as possible) simple dynamics is tovary the parameter δV for each column This has the effect of destroying any symmetry-forcedinvariant manifolds and so the simple case of synchronization corresponding to identicallyevolving dynamics will not exist in this case However more exotic forms of synchronizationare still possible and we discuss these next
42 Generalized chaotic synchronization
In order to maintain consistency with the literature we study non-linear interdependence in thesetting where the systems are asymmetrical but the coupling is unidirectional This is easilyachieved in the present system by making C directed and setting C21 = 0 In this settingwe refer to the lsquofreersquo system as the driver and the lsquoslavedrsquo system as the response We firstask the question what is the effect of introducing a slight parameter mismatch into a systemexhibiting stable chaos synchronization An example of this is presented in figure 9 whereC12 = 04 and parameter mismatch has been incorporated by setting δV = 07 in the driversystem and δV = 0699 in the response system In panel (a) the time series appear qualitativelyvery similar However in panel (b) where the difference between the systems is plotted it isapparent that the separation between the two systems is of the same order of magnitude as thesize of each attractor In panel (c) the effect of the parameter mismatch on the synchronizationmanifold can be seen as a transformation away from the symmetry manifold
It has been proposed that in such a setting there exists a continuous functional relationshipbetween the dynamics of the driver and response systems
x2(t) = ψ(x1(t)) (10)
where xi = V i W i Z i This situation has been referred to in the literature as generalizedsynchronization (Afraimovich et al 1986 Rulkov et al 1995) The properties of ψ andthe topology and stability of the synchronization manifold in this context have not yetbeen fully elucidated although some significant results have been achieved For example
Synchronization in a model of neuronal dynamics 717
1
2
2
1
Figure 9 Generalized synchronization in an asymmetrical driverndashresponse system (a) The timeseries of the two systems (b) The distance between the two systems (c) A plot of concurrent meanexcitatory membrane potentials V 1 against V 2 The diagonal represents the symmetry manifold
718 M Breakspear et al
Hunt et al (1997) Josic (1998) Kocarev et al (2000) have shown that the generalizedsynchronization manifold is smooth (differentiable) if and only if it is normally hyperbolic(ie the largest transverse Lyapunov exponent is less than the smallest tangential Lyapunovexponent prior to the introduction of parameter mismatch) Such a condition is met in thepresent setting when C = 04 hence the smoothness of the manifold in panel (c)
However subsequent to the introduction of parameter mismatch we will not be able touse transverse and tangential Lyapunov exponents to detect such synchronization since thesynchronization manifold may possess a complex topology for which it is a non-trivial (atbest) or impossible (at worse) task to define transverse and tangential directions For examplethe manifold may be smooth as in figure 9(c) but have a fractal topology If the dynamics arenot normally hyperbolic then the manifold may be fractal but also non-differentiable (Huntet al 1997) and hence will not have a unique tangent direction at any point Consequently wewill not be able to compute tangential or transverse Lyapunov exponents and hence determinethe stability of the synchronization manifold However other techniques for detecting thepresence of generalized synchronization in complex systems have evolved including theauxiliary system approach (Abarbanel et al 1996) and non-linear interdependence (Schiffet al 1996) which are now explored
421 The auxiliary system approach The auxiliary system is a third subsystem which isa replica of the original response system but coupled independently to the driver system andcommencing from independent initial conditions It is possible to demonstrate that showingthe existence of generalized synchronizationmdashthat equation (10) is satisfiedmdashis equivalent toshowing that the response system and the auxiliary system evolve identically (even though theyare not directly coupled) To determine whether or not this is the case reduces to an analysisof the Lyapunov exponents transverse to the symmetry manifold with respect to the responsesystem and the auxiliary system
Whilst this is a somewhat artificial test for generalized synchronization since theproduction of an auxiliary system for a physical system would be impossible such a testis useful in numerical and theoretical studies In figure 10 we present an example of theauxiliary system approach where the parameter δV = 07 for column 1 and 068 for column 2We consider two values of the coupling C = 0025 and 01 The auxiliary system is simplya copy of the second column beginning from an arbitrary initial state In the first instancewhere C = 0025 we notice that even after allowing for an initial transient state the secondcolumn and the auxiliary do not behave in identical fashions (figure 10(a)) However whenC = 01 we observe (figure 10(b)) that the second column and the auxiliary do behave innear-identical manners after an initial transient and hence conclude that for the particularvalue of the coupling strength considered equation (10) is satisfied and hence generalizedsynchronization between the first and second columns occurs
422 Nonlinear interdependence The question remains as to how we may detect thepresence of generalized synchronization in a practical setting As we mentioned aboveit is not possible to use an auxiliary-system-based approach to detect the presence ofgeneralized synchronization Instead we must consider techniques for detecting suchbehaviour from experimental or numerical time series data Algorithms of this type areoften referred to as detecting non-linear interdependence Nonlinear interdependence is ageneral term encompassing systems where the behaviour of one non-linear system affectsthe behaviour of another More specifically it can be defined as occurring if the nullhypothesis that the interdependence between two signals is purely linear can be rejectedat conventional levels (usually 95) of statistical significance hence it is an experimental
Synchronization in a model of neuronal dynamics 719
2
1
2
1
Figure 10 Concurrent evolution of the response system V 2 and lsquoauxiliaryrsquo system V 1 (a) C =0025 (b) C = 01
term (eg Schiff et al 1996) Whilst this does not strictly correspond to the existence of afunctional relationship between the two systems such a relationship is typically used tomotivate the techniques for its detection Schiff et al (1996) extended the logic of non-linearprediction algorithms from autonomous to coupled systems by exploiting the continuity ofsuch a relationship and hence devised the first method of detecting non-linear interdependencein time series data The basis of the technique is using the forward evolution of a phase spacevectorrsquos neighbours to predict the forward evolution of that vector and the forward evolutionof the concurrent vector of the other (possibly interdependent) system If the error of such aprocedure is better on average than chance (the chance error is normalized to equal one) thennon-linear interdependence is said to be present
720 M Breakspear et al
V1
V2
Figure 11 Experimental and numerical examples of weak generalized non-linear synchronizationin the presence of measurement noise (a) Human scalp EEG data from posterior bipolar scalpelectrodes (O1P3ndashO2P4) (b) Weakly coupled aperiodic data in the present model with additivewhite noise (with 5 of the amplitude of the model signal) Parameters δ1
V = 0605 δ2V = 061
aee = 036 C = 007
A variation of this algorithm optimized to detect weak non-linear interdependence (Terryand Breakspear 2003) was used recently to demonstrate that weak non-linear interdependence isoccasionally present in the scalp EEG of healthy resting human subjects (Breakspear and Terry2002a 2002b 2002c) An example of an epoch of lsquonon-linearly independentrsquo EEG obtainedusing this technique is presented in figure 11(a) For comparison output from the currentmodel is presented in figure 11(b) The model parameters were set such that the dynamicswas still aperiodic but the intermittent large-amplitude bursts were suppressed and had weakmutual coupling (C = 007) Extraneous noise to simulate the effect of extracranial muscleactivity contaminating the EEG data has been added to this time series It should be notedthat the amplitudes and time scales of the modelrsquos variables are dimensionless Nonethelessthe model facilitates qualititative interpretation of experimental findings
Synchronization in a model of neuronal dynamics 721
43 Chaotic phase synchronization
Whereas the concept of generalized synchronization was mathematically motivated and assuch more suited to theoretical investigations of systems the concept of phase synchronizationderives from a more practical signal processing approach Phase synchronization occurs inboth symmetric and asymmetric settings but is often more useful in the asymmetric case (andin experimental data)
Unfortunately the phase of a chaotic signal is typically ambiguous and can be defined in anumber of different ways For example in the case of the coherent Rossler attractor (Osipovet al 1997) the phase may be defined as the arctangent of two of the variables of the systemMore generally it can sometimes be defined as the angle that an orbit subtends with the centreof an attractor It is also possible to extract the phase of a chaotic signal from time seriesdata by use of the Hilbert transform This has enabled the demonstration of chaotic phasesynchronization in a number of coupled asymmetrical systems (Rosenblum et al 1996 1997Zaks et al 1999) Although such an approach does not guarantee that the phase will be uniqueit does permit the practical analysis of phase in noisy time series (Tass et al 1998)
To illustrate phase synchronization in two coupled columns we again vary the parameterδV which if the columns are viewed as oscillators acts to detune them from a commonfrequency An important question is that of the criteria on the basis of which we concludewhether phase synchronization occurs We use a simple locking criterion whereby phasesynchronization is said to occur if
|θ1(t)minus θ2(t)| lt K (11)
where is θi the phase of column i and K is a constant Notice that this does not mean thatsynchronization implies equality of phases merely that their difference is bounded
An example of phase synchronization with δV = 065 for column 1 and 066 for column 2is given in figure 12 In this figure rNMDA = 03 Qualitatively some sort of phase coherenceis evident in the time series panel (a) This was confirmed by plotting the sine of thephase difference which has been extracted using the Hilbert transform panel (b) Clearly|sin(θ1 minus θ2)| lt K and hence |θ1 minus θ2| is bounded It can also be seen that the phase-synchronized dynamics occurs on a highly structured manifold depicted in figure 12(c)However unlike the case for the generalized synchronization manifold depicted in figure 9 itis evident that this manifold is not a simple transformation of the symmetry manifold
It is also possible to see lsquobimodalrsquo stability whereby a system jumps betweenapproximately zero phase difference and a phase lag phase synchronization of π An exampleis depicted in figure 12(d) At approximately 2000 s the system can be seen to jumpspontaneously from the first to the second of these modes The Z -variable is plotted forgraphical purposes
5 Intermittency and desynchronization
51 Desynchronization in an array of two systems
Diminishing the strength of coupling across the blowout bifurcation does not lead to a simpletransition from synchronous to asynchronous oscillations but is instead associated withcomplex temporal behaviours Specifically when the largest transverse Lyapunov exponent isclose to zero the coupled systems exhibit epochs of synchronous behaviours punctuated bysharp bursts of desynchronization This is observed in systems exhibiting identicalgeneralizedand phase synchronization An example of such a phenomenon is given in figure 13 The timeseries panel (a) shows synchronous dynamics except at time = 1000 where it can be seenthat the two time series desynchronize briefly This is evident in panel (b) as a disruption in the
722 M Breakspear et al
2
1
Figure 12 Phase-synchronized coupled columns (a) The time series of V 1 (solid) and V 2 (b) Thesine of the phase difference obtained through Hilbert transformation of the time series in panel (a)(c) The phase synchronization manifold in the V 1ndashV 2 plane (d) The time series of Z (solid) andZ showing a phase slip of π between two modes
Synchronization in a model of neuronal dynamics 723
Figure 13 An example of phase desynchronization in coupled asymmetric systems with C = 01Time series plots of (a) each V -variable and (b) their Euclidean distance (c) The temporal evolutionof the phase difference between the two systems (d) An example of the phase difference evolutionover a longer time interval
724 M Breakspear et al
orderly evolution of the distance between the two systems and in panel (c) where the evolutionof the phase difference reveals a brief violation of the criterion |φ1 minus φ2| lt constant Anexample of the phase difference evolution over a longer time interval is shown in figure 13(d)Note that this is a three-dimensional plot with sin(φ1 minus φ2) and cos(φ1 minus φ2) as they- and z-axes Hence phase desynchronizations are evident as complete revolutions aroundthe cylindrical phase space in which this variable evolves It can be seen that these phenomenaoccur at irregular intervals in the temporal domain
This behaviour can be explained by considering the nature of a chaotic attractor In thestatistical sense an arbitrary initial condition is expected to lead to the chaotic behaviour onthe attractor Consequently when we calculate the Lyapunov exponents of the system theseexponents correspond to those of the chaotic attractor Typically however there are an infinitenumber of unstable periodic orbits embedded within the attractor If the path of a trajectorybecomes close to one of these orbits it may lsquoshadowrsquo the orbit for a brief period of time Eachof these orbits has a corresponding set of Lyapunov exponents which (like the attractor) can bedivided into tangential and transverse exponents These also vary when we change the couplingparameter but are typically not the same as the exponents of the attractor Hence the largesttransverse exponent of the attractor may be negative yet there may be positive transverseexponents associated with periodic orbits Considering all of these exponents together wewould therefore only expect to observe complete stability of the synchronization manifold inthe case where all of the transverse Lyapunov exponents are negative Bursting behaviourwill be observed when the one of these exponents becomes positive typically correspondingto a periodic orbit of low order (Pikovsky and Grassberger 1991 Rulkov and Sushchik 1997Maistrenko et al 1998 Heagy et al 1998) Due to the complex structure of such periodic orbitsthe desynchronous bursts occur across a wide range of timescales
In figure 14 we present the results of a long integration (16times105 time units) in which thelengths of sequential synchronous epochs of phase synchronization are studied The parametervalues were δV = 065 for column 1 and 0645 for column 2 with coupling strength10 equal to0125 In panel (a) the power spectrum of these sequential lengths is plotted The whiteness ofthis spectrum indicates that the lengths of consecutive synchronous epochs are independentThis is due to the high-dimensional lsquomixingrsquo effect of desynchronization In panel (b) thelengths of the synchronous epochs have been sorted again into the order of increasing lengthThe results for two coupling strengths are plotted C = 0125 (solid) and 0135 (dashed)Several features of this plot are of note Firstly the lengths of the synchronous epochsare distributed over almost three orders of magnitude Secondly for both plots there is anapproximately log-linear relationship between the length of an epoch and its relative orderThis reflects an approximately exponential relationship between the length of a synchronousepoch and its likelihood of occurrence Thirdly this relationship is skewed toward longersynchronous epochs for the stronger coupling strengths
This type of signal with patches of lsquoorderrsquo occurring over many timescales interruptedby bursts of disorder can be shown to have a high information capacity (Gatlin 1972)Moreover complex phase desynchronization may make an infrequent yet robust contributionto the dynamical pattern of the alpha rhythm in scalp EEG data for resting human subjects(Breakspear 2002) It is tempting to postulate that such a lsquoweakly stablersquo dynamics may assistin the generation of the rich and fluid mental associations that spontaneously emerge in restingcognitive states Although this phenomenon has been previously described in a variety of othersystems to our knowledge this is the first description in a biophysical model of neocortex
10 The introduction of parameter mismatch causes an increase in the strength of coupling required for stablesynchronization above the value of 01 required in the symmetric case
Synchronization in a model of neuronal dynamics 725
Figure 14 Properties of sequential epochs of phase-synchronous behaviour over a long integrationtime δ1
V = 065 δ2V = 0645 aee = 04 (a) The flat (white) power spectrum (b) Synchronous
epochs sorted in order of increasing length (solid C = 0125 dashed C = 0134)
52 Synchronization and complex dynamics in larger arrays
The synchronous and complex phenomena described above are also observed in arrays ofcoupled systems with N gt 2 Expanding the size of the array permits complex dynamicsin the spatial in addition to the temporal domain However the basic principles governingthe type and stability of synchronized motions are the same In this section a few of thepossible behaviours are discussed Physiological variation across the array is modelled bymodifying the parameter δV = 07 in each array by adding a random number of maximummagnitude 001
In figure 15(a) an array of N = 20 uncoupled (C = 0) systems is shown The lack ofsynchronization in this time series is evident By comparison the rapid transition to completesynchronization is evident in figure 15(b) when C = 03 In fact 19 of the 20 subsystems
726 M Breakspear et al
(b)
(c)
(a)
Figure 15 Synchronization phenomena in an array of coupled cortical columns (a) N = 20C = 0 (b) N = 20 C = 03 (c) N = 6 C = 01 (d) N = 12 C = 009 (e) N = 12C = 012 A small perturbation has been applied to one of the columns at t = 2500 s (f) Evolutionof the phase difference between two of the columns in panel (e) For clarity the W -variable isplotted in panel (c) and the Z -variable in (e)
have phase synchronized within one single cycle Intermediate values of coupling strength areassociated with a rich variety of phenomena For example subsystems with the most similarparameter values typically synchronize into clusters of various sizes An example of this isillustrated in figure 15(c) Note that there is no synchronization between the two differentclusters each containing three synchronized subsystems
Another complex behaviour occurs when subsystems transiently synchronize into largeclusters then spontaneously desynchronize again Hence the dynamics of the entire arrayjumps between different transient dynamics characterized by varying degrees of freedom Anexample of this phenomenon is given in figure 15(d) In addition it is possible to inducedesynchronization across an entire array by applying a small perturbation to one columnfor one time step An example is illustrated in figure 15(e) A small perturbation has been
Synchronization in a model of neuronal dynamics 727
Figure 15 (Continued)
added to one column after 2500 time units It can be seen that the entire array subsequentlydesynchronizes This illustrates the sensitivity of the dynamics to external stimuli Finally it ispossible to lsquoinducersquo synchronization between selected subsystems by increasing the strength ofcoupling between them selectively This allows an extra level of complexity to be introducedinto the model For example it may allow targeted information transfer between a partialnetwork of the entire array
As discussed in section 51 a univariate signal which jumps between epochs of lowentropy (high repetition) and bursts of high entropy (high variability) can be shown to havehigh information capacity This is because the repetition of certain patterns (as in the English
728 M Breakspear et al
language) permits error detectability Bursts of high entropy act in the opposite directionto increase message variability With a multivariate signal in addition to optimizing theinformation capacity within each signal the overall information capacity can be increasedthrough the type of itinerant synchronization briefly illustrated in this section Synchronizationacross an entire array increases the message reliability (since a single lsquoerrant letterrsquo in onesignal can be easily detected) Conversely desynchronization into multiple clusters increasesmessage variability since each cluster will produce independent information Hence a neuralarray which switches dynamically between a range of cluster configurations can be seen asadaptive in the sense that high synchronous states can be expressed when error detectabilityis a priority (such as during quick and reliable execution of a familiar task) but the array canrapidly desynchronize if variability becomes salient (for example if the environment suddenlybecomes novel) A formal operationalized demonstration of this principle is to be the subjectof future work
6 Discussion
In this paper we present a dynamical model for activity in an array of weakly coupledlocal neural assemblies The model incorporates the fundamental principles governing cellmembrane potential as determined by voltage- and ligand-gated ion channels In this wayit allows a reasonable amount of physiological complexity to be incorporated includingsynaptic activation and receptor subtypeswithout recourse to overwhelming detail Numericalanalysis of this model reveals a transition from independent oscillations to stable chaoticsynchronization depending on the balance of local versus long-range connectivity andbiophysical heterogeneity These excitatory couplings are subject to ascending subcorticalneuromodulation arising principally from monoaminergic nuclei such as Locus Coeruleus(Silberstein 1995) Neuromodulation of extrinsic connections was incorporated into the modelvia a coupling parameter C The stability of the synchronization is determined by the valueof the largest transverse Lyapunov exponent which in turn depends on the coupling parameterC If the coupling is weakened with the result that this exponent becomes positive a blowoutbifurcation occurs and independent oscillations re-emerge In the vicinity of the transitionnon-linear desynchronizations with complex temporal and spatial properties are observed
It is pleasing that complex and itinerant behaviour reminiscent of real neuronaldynamics emerges through such a simple manipulation of a single dynamic control parameterFurthermore the ubiquitous projections of classical neuromodulatory systems in real brainssuggest a clear biological mechanism by which the brain could self-organize throughmoderating the complexity of its own dynamics There are many reasons that the complicateddynamics of the sort demonstrated in this paper may be neurobiologically important Wemention four
First the presence of multiple clusters of phase-synchronized subsystems as discussedabove may be a uniquely non-linear mechanism facilitating the lsquocarriagersquo and distinction ofmultiple concurrent dynamic cell assemblies (and hence cognitive processing) This may beimportant for perceptual grouping binding and transitions as suggested by others (eg von derMalsburg 1985 Gray and Singer 1989)
Second the facility to decouple and reconfigure dynamic cell assemblies througha neuromodulatory control parameter may be important for avoiding low-entropysynchronization manifolds In the absence of itinerancy synchronization would renderneuronal dynamics incapable of representing changes in sensory input or indeed cognitive stateFor example in the model it was observed that small perturbations to one column were ableto induce a global change in neuronal dynamics by disrupting a state of weak synchronization
Synchronization in a model of neuronal dynamics 729
Inevitably alternative dynamic configurations emerge epitomizing the importance of lsquolabilersquobrain dynamics (Friston 2000a)
Thirdly from a purely functionalist point of view the balance between functionalspecialization and functional integration may rely upon the sorts of mechanism modelledin this paper The brain appears to adhere to two principles of functional organization namelysegregation of function within cortical areas and sub-areas and the integration of these areasin perceptual synthesis cognition and sensorimotor integration (Tononi et al 1994 Fristonet al 1995) The balance between preserving specialized dynamics unique to each arearequired by specialization and the coupling among these areas mediating functional integrationmay be achieved by the neuromodulatory mechanisms described above Significantlythe spontaneous variation in the numbers of degrees of freedom (for example whenlow-dimensional synchronization manifolds are shadowed) imparts a dynamic characteristicto this perspective that is precluded by linear mechanisms alone
Finally from a mechanistic perspective complexity and diversity of brain dynamics arerequired for selection of adaptive responses (Friston 2000b) There are many examples ofselective mechanisms operating within the brain All can be seen as performing an ascenton some objective function or fitness landscape These mechanisms require complex anditinerant dynamics to facilitate a thorough exploration of the landscape These landscapesmay range from those involved in perception (where the landscape is defined by predictionerror see eg Rao and Ballard (1998)) to those involved in value learning (where thelandscape is determined by the outcome of a behaviour eg in temporal difference models (seeeg Sutton and Barto 1990)) In short complexity is necessary to maintain an information-rich environment in which the brain can respond to sensory inputs An essential componentof this is the maintenance of diversity which is a prerequisite for selectionist mechanismsat many scales (Edelman 1993) Understanding the mechanistic basis of complex dynamicsas measured electrophysiologically may well depend on the sort of biologically plausiblemodels used in this paper This understanding may be important for both basic and clinicalneuroscience (eg Tass 2002)
The present study supplements and advances on other research in this field The studyof complex phase synchronization and clustering in neural oscillator networks similar tothe present model has previously been the subject of formal mathematical study Forexample Terman and Wang (1995) studied synchronization of blocks of neural oscillatorsthat were subject to perturbation by connected lsquostimulirsquo Somers and Kopell (1995) studiedthe emergence of both synchronization and travelling waves in networks of coupled neuraloscillators In the work of Terman and Wang the local dynamics were based on neurallsquocaricaturersquo models such as the FitzHughndashNagumo equations and are hence more abstract thanthe current study The approach of Somers and Kopell only required that the local dynamicsexhibit fast and relaxation phases and is hence more abstract still In both studies onlyperiodic dynamics are studied Complex patterns of chaos synchronization in neural arrayshave been characterized by nonlinear methods (Kaneko 1998) and their putative contributionto both learning (Nakajima and Ueda 1996) and perception (Zhao and Macau 2001 Breakspear2001) explored However these studies employed models such as the logistic equation andtent map which are quite abstract in comparison to the present study On the other handHansel and Sompolinsky (1996) studied the role of chaos synchronization in a model ofvisual perception in a cortical hypercolumn using a very similar approach to that adoptedhere but with the additional complexity that synaptic delays were incorporated However inthis and earlier work (Hansel and Sompolinsky 1992) only linear measures of dynamics andsynchrony (specifically the auto- and cross-correlation functions) were employed The analysisof chaotic synchrony in the study of Larter et al (1999) was based only upon observation of
730 M Breakspear et al
the Euclidean difference between variables from two nodes within a larger array Hence thesignificance of the present work is that it provides a quantitative nonlinear analysis of complexchaos synchronization in a biophysical model of cortical dynamics Such research is requiredin order to bridge the gap between quantitative nonlinear research into synchronization andcomputation in abstract models of neural arrays and qualitative or linear analysis in morebiophysically plausible models
Acknowledgments
The authors would like to thank Peter Robinson for assistance with the manuscript and R Larterfor helpful advice MB is a recipient of a NSW Institute of Psychiatry Research Grant anda University of Sydney SESQUI postdoctoral fellowship KJF is supported by the WellcomeTrust
References
Abarbanel H Rulkov N and Sushchik M 1996 Generalised synchronization of chaos the auxiliary system approachPhys Rev E 53 4528ndash35
Afraimovich V Verichev N and Rabinovich M 1986 Stochastic synchronization of oscillation in dissipative systemsRadiophys Quantum Electron 29 795ndash801
Breakspear M 2001 Perception of odours by a nonlinear model of the olfactory bulb Int J Neural Syst 11 101ndash24Breakspear M 2002 Nonlinear phase desynchronisation in human electroencephalographic data Human Brain Mapping
15 175ndash98Breakspear M and Terry J R 2002a Nonlinear interdependence in neural systems motivation theory and relevance
Int J Neurosci 112 1163ndash84Breakspear M and Terry J R 2002b Detection and description of nonlinear interdependence in normal multichannel
human EEG Clin Neurophysiol 113 735ndash53Breakspear M and Terry J R 2002c Topographic organization of nonlinear interdependence in multichannel human
EEG Neuroimage 16 822ndash35Destexhe A Rudolph M Fellous J M and Sejnowski T J 2001 Fluctuating synaptic conductances recreate in vivo-like
activity in neocortical neurons Neuroscience 107 13ndash24Douglas R J Mahowald M A and Martin K A 1996 Microarchitecture of neocortical columns Brain TheorymdashBiological
Basis and Computational Principles ed A Aertsen and V Braitenberg (Amsterdam Elsevier)Eckhorn R Bauer R Jordan W Brosch M Kruse W Munk M and Reitboeck H J 1988 Coherent oscillations a
mechanism of feature linking in the visual cortex Multiple electrode and correlation analyses in the cat BiolCybern 60 121ndash30
Edelman G M 1993 Neural Darwinism selection and reentrant signalling in higher brain function Neuron 10 115ndash25Engel A K Kreiter A K Konig P and Singer W 1991 Synchronization of oscillatory neuronal responses between
striate and extrastriate visual cortical areas of the cat Proc Natl Acad Sci USA 88 6048ndash52Friston K J 2000a The labile brain I Neuronal transients and nonlinear coupling Phil Trans R Soc B 355 215ndash36Friston K J 2000b The labile brain II Transients complexity and selection Phil Trans R Soc B 355 237ndash52Friston K J Tononi G Sporns O and Edelman G 1995 Characterising the complexity of neuronal interactions Human
Brain Mapping 3 302ndash14Fujisaka H and Yamada T 1983 Stability theory of synchronized motion in coupled-oscillator system Prog Theor
Phys 69 32ndash47Gatlin L 1972 Information Theory and the Living System (New York Columbia University Press)Gray C M Konig P Engel A and Singer W 1989 Oscillatory responses in cat visual cortex exhibit inter-columnar
synchronization which reflects global stimulus properties Science 338 334ndash7Gray C M and Singer W 1989 Stimulus specific neuronal oscillations in orientation columns of cat visual cortex Proc
Natl Acad Sci USA 86 1698ndash702Haig A R Gordon E Wright J J Meares R A and Bahramali H 2000 Synchronous cortical gamma-band activity in
task-relevant cognition Neuroreport 11 669ndash75Hansel D and Sompolinsky H 1992 Synchronization and computation in a chaotic neural network Phys Rev Lett 68
718ndash21
Synchronization in a model of neuronal dynamics 731
Hansel D and Sompolinsky H 1996 Chaos and synchrony in a model of a hypercolumn in visual cortex J ComputNeurosci 3 7ndash4
Heagy J Carroll T and Pecora L 1998 Desynchronization by periodic orbits Phys Rev E 57 R1253ndash6Hunt B Ott E and Yorke J 1997 Differentiable synchronization of chaos Phys Rev E 55 4029ndash34Josic K 1998 Invariant manifolds and synchronization of coupled dynamical systems Phys Rev Lett 80 3053ndash6Kandel E Schwartz J and Jessell T 2000 Principles of Neural Science (New York McGraw-Hill)Kaneko K 1998 On the strength of attractors in a high-dimensional system Physica D 124 322ndash44Kocarev L Parlitz U and Brown R 2000 Robust synchronization of chaotic systems Phys Rev E 61 3716ndash20Larter R Speelman B and Worth R M 1999 A coupled ordinary differential equation lattice model for the simulation
of epileptic seizures Chaos 9 795ndash804Lumer E Edelman G and Tononi G 1997a Neural dynamics in a model of the thalamocortical system I Layers loops
and the emergence of fast synchronous oscillations Cereb Cortex 7 207ndash27Lumer E Edelman G and Tononi G 1997b Neural dynamics in a model of the thalamocortical system II The role of
neural synchrony tested through perturbations of spike timing Cereb Cortex 7 228ndash36Magleby K 1987 Short-term modulation at synapses Synaptic Function ed G M Edelman W E Gall and
W M Cowan (New York Wiley)Maistrenko Y Maistrenko V Popovich A and Mosekilde E 1998 Transverse instability and riddled basins in a system
of two coupled logistic maps Phys Rev E 57 2713ndash24Morris C and Lecar H 1981 Voltage oscillations in the barnacle giant muscle fiber Biophys J 35 193ndash213Mountcastle V B 1997 The columnar organization of the neocortex Brain 120 701ndash22Mountcastle V B 1998 The Cerebral Cortex (Cambridge MA Harvard University Press)Nakajima H and Ueda Y 1996 Riddled basins of the optimal states in learning dynamical systems Physica D 99 35ndash44Nunez P L 1995 Neocortical Dynamics and Human Brain Rhythms (Oxford Oxford University Press)Osipov G V Pikovsky A S Rosenblum M G and Kurths J 1997 Phase synchronization effects in a lattice of nonidentical
Rossler oscillators Phys Rev E 55 2353ndash61Ott E and Sommerer J 1994 Blowout bifurcations the occurrence of riddled basins and onndashoff intermittency Phys
Lett A 188 39ndash47Pecora L and Carroll T 1990 Synchronization in chaotic systems Phys Rev Lett 64 821ndash4Pikovsky A and Grassberger P 1991 Symmetry breaking bifurcation for coupled chaotic attractors J Phys A Math
Gen 24 4587ndash97Rao R P and Ballard D H 1998 Predictive coding in the visual cortex a functional interpretation of some extra-classical
receptive field effects Nature Neurosci 2 79ndash87Robinson P A Wright J J and Rennie C R 1998 Synchronous oscillations in the cerebral cortex Phys Rev E 57
4578ndash88Rodriguez E George N Lachauz J P Martinerie J Renault B and Varela F J 1999 Perceptionrsquos shadow long distance
synchronization of human brain activity Nature 397 430ndash3Rosenblum M Pikovsky A and Kurths J 1996 Phase synchronization of chaotic oscillators Phys Rev Lett 76 1804ndash7Rosenblum M Pikovsky A and Kurths J 1997 From phase to lag synchronization in coupled chaotic oscillators Phys
Rev Lett 78 4193ndash6Rulkov N and Sushchik M 1997 Robustness of synchronized chaotic oscillations Int J Bifurc Chaos 7 625ndash43Rulkov N Sushchik M Tsimring L and Abarbanel H 1995 Generalized synchronization of chaos in unidirectionally
coupled chaotic systems Phys Rev E 51 980ndash94Schiff S So P Chang T Burke R and Sauer T 1996 Detecting dynamical interdependence and generalized synchrony
through mutual prediction in a neural ensemble Phys Rev E 54 6708ndash24Silberstein R B 1995 Neuromodulations of neocortical dynamics Neocortical Dynamics and Human EEG Rhythms
ed P L Nunez (Oxford Oxford University Press)Singer W 1995 Putative functions of temporal correlations in neocortical processing Large-Scale Neuronal Theories
of the Brain ed C Koch and J Davis (London MIT Press)Somers D and Kopell N 1995 Waves and synchrony in networks of oscillators of relaxation and non-relaxation type
Physica D 89 169ndash83Sutton R S and Barto A G 1990 Time derivative models of Pavlovian reinforcement Learning and Computational
Neuroscience Foundations of Adaptive Networks ed M Gabriel and J Moore (Cambridge MA MIT Press) pp497ndash538
Szentagothai J 1983 The modular architectonic principle of neural centers Rev Physiol Biochem Pharmacol 9811ndash61
Tallon-Baudry C Betrand O and Fischer C 2001 Oscillatory synchrony between human extrastriate areas during visualshort-term memory maintenance J Neurosci 21 RC177(1ndash5)
Tass P A 2002 Desynchronization of brain rhythms with soft phase-resetting techniques Biol Cybern 87 102ndash15
732 M Breakspear et al
Tass P Rosenblum M Weule J Kurths J Pikovsky A Volkmann J Schnitzler A and Freund H 1998 Detection of nmphase locking from noisy data application to magnetoencephalography Phys Rev Lett 81 3291ndash4
Terman D and Wang D 1995 Global competition and local cooperation in a network of neural oscillators Physica D81 148ndash76
Terry J and Breakspear M 2003 An improved algorithm for the detection of dynamical interdependence in bivariatetime-series Biol Cybern 88 129ndash36
Tononi G Sporns O and Edelman G M 1994 A measure for brain complexity relating functional segregation andintegration in the nervous system Proc Natl Acad Sci USA 91 5033ndash7
von der Malsburg C 1985 Nervous structures with dynamical links Phys Chem 89 703ndash10Wang X 1991 Period-doublings to chaos in a simple neural network an analytic proof Complex Syst 5 425ndash41Wennekers T and Pasemann F 1996 Synchronous chaos in high dimensional modular neural networks Int J Bifurc
Chaos 6 2055ndash67Zaks M A Parks E H Rosenblum M G and Kurths J 1999 Alternative locking ratios in imperfect phase synchronization
Phys Rev Lett 82 4228ndash31Zhao L and Macau E N 2001 A network of dynamically coupled chaotic maps for scene segmentation IEEE Trans
Neural Netw 12 1375ndash85

704 M Breakspear et al
et al 1999 Haig et al 2000 Tallon-Baudry et al 2001) and computational models of the cortex(eg Lumer et al 1997a 1997b Robinson et al 1998) However in much of this researchonly linear measures of synergistic activity such as the coherence function are employedThe non-linear properties of excitable membranes motivate the study of non-linear aspectsof synchronicity in neural systems The development of computing resources and non-lineartechniques renders such studies increasingly feasible and may offer a meaningful contributionto the understanding of brain function
In this paper we study non-linear interdependence in a model neural system consisting ofan array of coupled small-scale neural subsystems Each subsystem is constituted by denselyinterconnected excitatory and inhibitory neurons These are then coupled together throughweak long-range excitatory projections to form a larger (lsquomesoscopicrsquo) array The dynamicswithin and between the subsystems derives from the properties of voltage-gated ion channelsand competition at post-synaptic ligand-gated channels The properties of the voltage-gatedchannels confer non-linear properties to the dynamics We study the types and stability ofnon-linear synchronization and consider the possible role of these behaviours in adaptive brainfunction
The rest of the paper is structured as follows In the next section we develop the evolutionequations for the neural dynamics firstly in local networks and subsequently across the entirearray In section 3 the local dynamics (in the absence of long-range coupling) of the systemis explored In section 4 the effect of increasing the strength of long-range coupling on theemergent cooperative dynamics is studied Section 5 focuses on the complex behaviours thatare observed in the presence of marginal stability of the synchronous dynamics In the finalsection the possible computational significance of the observed behaviours is discussed
2 Evolution equations
The approach adopted in this study is to consider the behaviour of local ensembles ofneurons with dynamical variables and parameters representing local ensemble averagesThe scale of each ensemble is taken as the extension of the pyramidal cell dendritic arbors(the size of the cortical columns) which is approximately 100ndash300 microm This approach ismotivated by the observations that neurons within columns tend to share the same physiologicalproperties exhibit dense reciprocal interconnectivity and show strong dynamical correlationsin comparison to neurons in different columns (Szentagothai 1983 Nunez 1995 Douglas et al1996 Mountcastle 1997 1998) For these reasons they have been proposed as comprisingthe brainrsquos lsquoelementary functional modulesrsquo (Kandel et al 2000) The next-largest spatialscale of neural organization is achieved through weak long-range excitatory-to-excitatoryintercolumnar connectivity A further discussion of the neurobiological basis of this approachhas been presented in Breakspear and Terry (2002a)
21 Local ensemble dynamics
The dynamical variables studied are the mean membrane potential of local pyramidal cells V and inhibitory interneurons Z and the average number of lsquoopenrsquo potassium ion channels W The evolution equations are adapted from a study of epileptic seizures in hippocampal slices(Larter et al 1999) consisting of coupled differential equations which in turn are derived fromthe model of Morris and Lecar (1981) The main focus is on the mean cell membrane potentialof the pyramidal cells governed by the conductance of sodium potassium and calcium ionsthrough voltage-gated channels plus the passive conductance of lsquoleakyrsquo ions We begin withthe total current flow across the local pyramidal cell membranes considered as capacitors
Synchronization in a model of neuronal dynamics 705
CdV
dt= minusgCamCa(V minus VCa)minus gNamNa(V minus VNa)minus gKW (V minus VK)minus gL(V minus VL) (1)
where gion is the maximum conductance of each population of ion species if all channelsare open m ion is the fraction of channels open and Vion is the Nernst potential for that ionspecies Henceforth following Larter et al (1999) and Destexhe et al (2001) all equationsand parameters are non-dimensional and normalized to neural capacitance C = 1
Each voltage-gated channel acts as a switch which opens when the membrane potentialpasses above a given threshold Tion Hence for a single channel the relationship betweenmembrane voltage and channel conductance is given by a step function Averaging over apopulation of such channels and assuming a Gaussian distribution of Tion around a mean foreach ion species gives the sigmoid-shaped lsquoneural activation functionsrsquo
m ion = 05
(1 + tanh
(V minus Tion
δion
)) (2)
where δion incorporates the variance of this distribution As discussed in Larter et al (1999) theprincipal effect of moving from a microscopic (single-neuron) to a mesoscopic (local network)spatial frame is to further increase the variance of the threshold potentials (consistent with anincrease in their physical heterogeneity) which has the effect of flattening this curve
The fraction of open potassium channels is slightly more complicated The channelslsquorelaxrsquo from one state to another at an exponential rate This is achieved if W is governed by
dW
dt= φ(mK minus W )
τ (3)
where φ is a temperature scaling factor and τ is the lsquorelaxationrsquo time constantThe second modification required for a network formulation is introducing synaptic
interactions between neurons within the same column This is achieved by calculating theaverage cell firing rates and introducing feedback terms subsequent to cell firing to representneurotransmitter release At the cell soma the membrane potential triggers an action potential ifit exceeds a threshold determined chiefly by the sodium channel threshold potential Averagingthis over the ensemble of neurons and assuming once again a Gaussian distribution we obtainthe cell firing rates
QV = 05 times QV max
(1 + tanh
(V minus VT
δV
)) (4)
QZ = 05 times QZ max
(1 + tanh
(Z minus ZT
δZ
)) (5)
where the Qmax are the maximum rates of firing of the excitatory and inhibitory neuronsrespectively
The firing of each of these cell populations feeds back onto the ensemble through synapticcoupling to open ligand-gated channels and raise or lower the membrane potential accordinglyIn the case of excitatory-to-inhibitory and inhibitory-to-excitatory connections this is modelledas additional inputs to the flow of ions across the membrane channel weighted by functionalsynaptic factors aei and aie respectively6 In the case of excitatory-to-excitatory connectionsthe mechanism of this increase in ion conductance is modelled with greater physiological detailincorporating some of the specific characteristics of the ligand-gated channel populations Themean rate of firing QV is assumed to lead to a proportional release of glutamate neurotransmitteracross the synapse which diffuses onto two classes of ligand-gated ion channels One class
6 Note that the feedback terms are multiplied by the potential of the cell of origin This incorporates the potentiatingeffect of residual pre-synaptic intracellular calcium which increases with the pre-synaptic potential (Magleby 1987)
706 M Breakspear et al
Figure 1 A schematic representation of connectivity within cortical columns Panel (a) depictsdense local connectivity between excitatory (EN) and inhibitory (IN) neurons Panel (b) depictsparameters representing strengths of synaptic connectivity between neurons within each columnplus input from ascending neurons within the reticular formation (RF)
embodying AMPA channels open an additional population of sodium channels thus increasingthe net conductance of sodium flow The second class representing NMDA receptors openan additional population of voltage-gated calcium channels Hence this acts to increase themaximum conductance of voltage-gated calcium channels Finally lsquononspecificrsquo subcorticalexcitation is added to both populations of neurons with amplitude I modulated by a randomnoise component of amplitude δ Thus we obtain
dV
dt= minus(gCa + rNMDAaee QV )mCa(V minus VCa)minus (gNamNa + aee QV )(V minus VNa)
minus gKW (V minus VK)minus gL(V minus VL) + aie Z QZ + ane Iδ (6)dZ
dt= b(ani Iδ + aeiV QV ) (7)
where rNMDA denotes the number of NMDA receptors relative to that of AMPA receptors ane
and ani parametrize non-specific input to excitatory and inhibitory cells and aee parametrizesthe local excitatory-to-excitatory synaptic strength Note the difference in the coefficientscontrolling the effect of local excitatory feedback onto NMDA receptors compared to thosefor AMPA receptors A schematic representation of the local network and the parametrizedsynaptic connectivity is presented in figure 1
22 Mesoscopic array dynamics
Each set of equations (2)ndash(7) governs the dynamics within a local cell assembly Couplingbetween N nodes can be introduced parsimoniously as competitive agonist excitatory actionbetween pyramidal cells in different columns However these long-range projections aremodelled to target the same populations of NMDA and AMPA receptors as are targeted by theshort-range excitatory projections Representing each node with a superscript this is achievedby modifying equation (6) to
dV i
dt= minus(gCa + (1 minus C)rNMDAaee Qi
V + CrNMDAaee〈QV 〉)mCa(Vi minus VCa)
minus gKW (V i minus VK)minus gL(Vi minus VL)
minus (gNamNa + (1 minus C)aee QiV + Caee〈QV 〉)(V i minus VNa) + aie Z Qi
Z + ane Iδ (8)
Synchronization in a model of neuronal dynamics 707
where i = 1 N and 〈 〉 represents spatial (lsquomean-fieldrsquo) averaging over cell assembliesC parametrizes the strength of excitatory coupling between cortical columns If C = 0the systems evolve independently C gt 0 introduces interdependences between consecutivecolumns C = 1 corresponds to maximum coupling with excitatory input from outside eachcolumn exceeding excitatory input from within each column
The long-range coupling is lsquoweakrsquo in the sense that we investigate parameter values forwhich C lt aee and C aie etc This either implies that the functional synaptic strength of thelong-range connections are weaker than local connections or that the strengths are similar butthe density of projections is comparatively sparse We prefer the latter interpretation whichis more consistent with other modelling approaches
23 Parameter values and physiological validity
In the sections that follow we set all physiologically measurable parameters (the maximumconductances threshold potentials and Nernst potentials) to within their accepted ranges (Larteret al 1999) Other parameters such as I the functional synaptic strengths and maximum firingrates were chosen to generate dynamically plausible behaviour including the limit cycle andaperiodic dynamics The parameter values are as given in table 1 (unless otherwise stated)There may be significant variability in many of these parameters In this respect it is crucial tonote that the behaviours described below occur in a large region of parameter space Hence theemergent behaviours discussed do not only occur for the parameter values used in this studyBecause as noted above the parameters are dimensionless and normalized to a capacitance ofone it is also important to note that the time units are arbitrary Noting however that the cellcapacitance is typically estimated as close to or equal to 1 microF cmminus2 (eg Destexhe et al 2001)it is reasonable to interpret the time evolution units as milliseconds (as done for the diffusivecoupling parameter employed in Larter et al (1999))
The principal aim of this paper is a quantitative study of non-linear synchronization ina biophysically realistic model of a neural system The model incorporates some aspects ofneural systems with significant detail such as the conductance of K+ glutamate-mediatedneurotransmission and the relatively sparse nature of long-range connectivity This detailallows manipulation of parameters that model pharmacological mechanisms of interest suchas glutamate receptor subclasses Other features such as local inhibitory feedback and thekinetics of Ca2+ and Na+ channels have been incorporated in a more simplified manner Thislimits the complexity of the model but ensures that numerical analysis can be supplementedby quantitative examination of for example the Lyapunov exponents Other biophysicalproperties could be made the subject of future study by changing relevant aspects of theequations
3 Behaviour of local lsquouncoupledrsquo columns
In this section a brief overview of the behaviours exhibited by autonomous local columns(ie C = 0) is presented As indicated in table 1 the dynamics of the model is explored byfixing all parameters except for δV the variance of the excitatory threshold which is varied inthe region [055 07] A similar range of behaviours is also observed if δV is fixed and anothercontrol parameter such as aee or I is varied
31 Fixed point attractors
For values of δV lt 055 the dynamics of a single column settles onto a solitary fixed pointattractor As shown in figure 2 transient dynamics persists for a number of seconds
708 M Breakspear et al
Figure 2 The time series for a fixed point attractor with VT = 054
Table 1 Parameter values unless otherwise stated Adapted from Larter et al (1999)
Parameter Description Value
TCa Threshold value for Ca channels minus001δCa Variance of Ca channel threshold 015gCa Conductance of population of Ca channels 11VCa Ca Nernst potential 1TK Threshold value for K channels 00δK Variance of K channel threshold 030gK Conductance of population of K channels 20VK K Nernst potential minus07TNa Threshold value for Na channels 03δNa Variance of Na channel threshold 015gNa Conductance of population of Na channels 67VNa Na Nernst potential 053VL Nernst potential leak channels minus05gL Conductance of population of leak channels 05VT Threshold potential for excitatory neurons 00δV Variance of excitatory threshold VariedZT Threshold potential for inhibitory neurons 00δZ Variance of inhibitory threshold 070I Subcortical input strength 030aee Excitatory-to-excitatory synaptic strength 04aei Excitatory-to-inhibitory synaptic strength 2aie Inhibitory-to-excitatory synaptic strength 2ane Non-specific-to-excitatory synaptic strength 1ani Non-specific-to-inhibitory synaptic strength 04b Time constant scaling factor 010ϕ Temperature scaling factor 07τ Time constant for K relaxation time 1rNMDA Ratio of NMDA to AMPA receptors 025β Random modulation of subcortical input 0
32 Limit cycle attractors
For 055 lt δV lt 059 the system undergoes a series of period-doubling bifurcations Theseresult in both simple and complex periodic behaviours A limit cycle occurring when δV = 056
Synchronization in a model of neuronal dynamics 709
Figure 3 The approach to a limit cycle attractor in phase space with VT = 056
is illustrated in figure 3 Other parameter variations produce more complicated higher-orderperiodic orbits For example a period-9 orbit occurring when δV = 06 aee = 05 aie = 05gNa = 0 and I = 0165 is presented in figure 4
33 Chaotic attractors
Setting δV gt 059 and keeping all other parameters as per table 1 produces complex aperiodicbehaviour in the uncoupled columns An example of this with δV = 065 is illustrated infigure 5 Panels (a) and (b) illustrate a phase space portrait and a time series plot of thissystem respectively They show an aperiodic but highly structured system In panel (c) afirst-return lsquoPoincarersquo section is presented This was obtained by taking a cross-section throughthe attractor in (a) by setting W = 032 and then plotting V (t) against V (t +l)with a time delayof l = 1 A topographic similarity to the simple logistic map is evident It has been provenanalytically that in restricted regions of parameter space dynamical systems incorporatingthe neural activation function (4) (5) are topologically conjugate to the logistic map (Wang1991) The Poincare map presented here shows that although a 11 map may not exist in moregeneral parameter regions the global morphology of the logistic map is still evident
To determine whether or not these aperiodic oscillations are in fact chaotic the Lyapunovexponents were estimated numerically using an algorithm based upon GrammndashSchmidtorthonormalization For the particular case illustrated the exponents were found to be
λ1 = 0015
λ2 = 00
λ3 = minus026
(9)
A principal Lyapunov exponent greater than zero is consistent with chaotic dynamics Thistogether with the topography of the first-return map and the existence of period-doublingbifurcations provides compelling evidence of chaotic behaviour in the dynamics of the singlecolumn The second exponent reflects the rate of exponential separation of adjacent orbits in
710 M Breakspear et al
Figure 4 A period-9 limit cycle attractor (a) A phase space portrait (b) A representative timeseries
the direction tangent to the flow hence a value of zero is to be expected The sum of the threeexponents is negative implying that the system is volume contracting which is to be expectedin the setting of a non-equilibrium (non-Hamiltonian) physiological system
It is interesting to note that this diversity of behaviours can be obtained throughmanipulation of a single state parameter From a functional perspective this permits eachcortical column to express a variety of qualitatively distinct dynamics through modificationof a single physiological parameter such as through modulation of the functional strength ofthe local synaptic connectivity Conversely it also implies that resetting of a single parametersuch as an abnormal δV resulting from loss of physiological heterogeneity could be associatedwith qualitative changes in local cortical behaviour
4 Cooperative behaviour of coupled columns
Whilst it is interesting to observe the range of dynamic behaviours possible in a single columnwe are primarily interested in the behaviour of many such interacting columns In this section
Synchronization in a model of neuronal dynamics 711
1
Figure 5 Chaotic behaviour of the autonomous system with VT = 065 The other parameters areas per table 1 (a) An attractor in phase space (b) A time series for V (t) (c) A Poincare sectionobtained by setting W = 032 and plotting V (t) against V (t + 1)
712 M Breakspear et al
we study the emergence of cooperative behaviour that results from the introduction of couplingbetween columns Three types of interdependent behaviour of two systems are describedidentical generalized and phase synchronization Synchronous behaviours in larger arrays arealso illustrated
41 Identical chaotic synchronization
411 Transition to identical chaotic synchronization The most straightforward extensionand the one that we consider first is that of two coupled cortical columns where all parametersare in the first case assumed identical7 Trivially the two systems will evolve identicallywhen started with identical initial conditions even if C = 0 This is simply a statement ofthe deterministic nature of the evolution equations In general the random initial conditionswill not be equal in which case it is hardly surprising that the different columns will evolveindependently An example is presented in figure 6 with VT = 06 in both systems C = 0and independent random initial conditions In figure 6(a) we show a segment of the timeseries for each of the two subsystems where it can be seen that the time series are chaoticand asynchronous In figure 6(b) the concurrent values of V 1 and V 2 are plotted against eachother showing that they bear no obvious interrelationship
With the introduction of coupling C gt 0 the systems begin to show epochs ofsynchronized behaviour At C sim= 01 there is a transition to stable fully synchronizedmotion as evident in an exemplar time series shown in figure 7(a) where C = 015 Infigure 7(b) we present the concurrent values of V 1 versus V 2 illustrating that in contrast tothe case in figure 6(b) the systems have become embedded in the low-dimensional lsquosymmetrymanifoldrsquo V 1 = V 2 Panel (c) shows a log-linear plot of the temporal evolution of the distanceof the coupled system from this manifold It can be seen that on average the two systemsapproach at an exponential rate with constant time-averaged exponent Whilst this phenomenon(lsquoidentical chaotic synchronizationrsquo) has been described in a variety of other systems since itsfirst description (Fujisaka and Yamada 1983 Pecora and Carroll 1990) to our knowledge thisis its first quantitative8 description in a detailed physiological model of a neural system
The symmetry of the dynamical system obtained by setting the state parameters in eachcolumn identical forces the invariance of the symmetry manifold V 1 = V 2 Moreover thedynamics within this manifold is governed by the same equations of motion as for a singlecortical column This manifold which contains the orbits of the synchronized system is calledthe synchronization manifold The coupling parameter C does not affect the dynamics withinthe synchronization manifold and as such is termed a normal parameter
412 lsquoBlowout bifurcationrsquo An important issue is the stability of the orbits on thesynchronization manifold This can be determined by analysing the Lyapunov spectraBecause the two-column system evolves in a six-dimensional phase space there are sixLyapunov exponents However for the synchronized system it is helpful to subdivide theseinto those that describe the dynamics within the synchronization manifold and those thatdescribe the dynamics in the transverse direction The latter describe the rate of convergenceor divergence of the coupled systems toward or away from this manifoldmdashand hence thetendency for the two systems to synchronize or separate For the time series in figure 7 this is
7 Whilst it is of course physiologically unrealistic to assume that two columns would have entirely identical structuresit is mathematically simpler to start analysing the system in this situation8 A qualitative description in a neural system with a different form of coupling (potassium diffusion) is evident infigure 8 of Larter et al (1999) Synchronous chaos in an abstract model of a neural system was also described inWennekers and Pasemann (1996)
Synchronization in a model of neuronal dynamics 713
1
2
2
1
Figure 6 Independent oscillations in a two-column system with identical parameters butindependent initial conditions (a) The time series (b) A plot of concurrent mean excitatorymembrane potentials V 1 against V 2
related to the slope of the plot in panel (c)9 For this strength of coupling the largest transverseexponent is negative (minus002) indicating that perturbations away from this state of identicalsynchronization decay and thus that the state of identical chaotic synchronization is stableagainst noise (ie dynamically stable)
To calculate the relationship between coupling strength and the Lyapunov exponents weused an algorithm which measures the growth of perturbations of the variational equations (thelsquoJacobianrsquo) of the system (we do not define these equations here as they are cumbersome anddo not enhance the understanding of the system) We calculated these exponents for increasingvalues of the coupling parameter C from 0 to 02 in increments of 0000 25 The results are
9 Only two of the six dependent variables are shown in figure 7 The actual transverse Lyapunov exponents reflectthe temporal evolution of all six and hence are somewhat more complex than that given by the slope of panel (c)
714 M Breakspear et al
1
2
Figure 7 Identical synchronization in the same system as in figure 7 but with C = 02 (a) Thetime series (b) A plot of concurrent mean excitatory membrane potentials V 1 against V 2 (c) Thelogarithm of the Euclidean distance between V 1 and V 2 versus time (linear)
Synchronization in a model of neuronal dynamics 715
Figure 8 Lyapunov spectra for a coupled system with δV = 07 and C varying from 0 to 02 insteps of 0000 33 At each value of C all Lyapunov exponents are calculated Least-squares linesof best fit were then fitted to the results for each exponent Solid lines indicate transverse exponents(λ) and dashed lines indicate the tangential exponents () Panel (a) shows all three transverseand three tangential exponents Panel (b) only shows the largest two of each
illustrated in figure 8 Panel (a) shows all exponents For illustrative purposes only the mostpositive four are plotted in panel (b) In both panels the tangential exponents () are fitted (inthe least-squares sense) with a dashed line and the transverse ones (λ) are fitted with a solidline They are labelled 1ndash3 in order of decreasing positivity The important features are asfollows For the range of coupling strengths that we consider the largest tangential Lyapunovexponent is positive which indicates the presence of chaotic behaviour within the manifoldWhen examining the line of best fit for this exponent we see that this exponent does not varywhen altering the coupling parameter This is true for all three tangential exponents and verifiesthat the changes in the coupling parameter do not alter the dynamics on the synchronizationmanifold (that is the coupling parameter is normal)
716 M Breakspear et al
The largest transverse Lyapunov exponent (denoted as λ1 in the figure) is a particularlyimportant exponent since it governs the stability of the attractor in the synchronization manifoldwith respect to perturbations out of the manifold If this exponent is negative then we expectsmall perturbations of trajectories to reconverge to the manifold in finite time If however thisexponent is positive then we would instead expect perturbed trajectories to diverge from themanifold This change in local behaviour is termed a bifurcation and the point at which itoccurs the bifurcation point It is apparent in figure 8 that the zero crossing of this exponentmdashthe bifurcation pointmdashoccurs at approximately C = 01 in the present system This typeof bifurcation has been termed a blowout bifurcation since once transverse stability is losttrajectories are lsquoblown outrsquo from the low-dimensional synchronization manifold into the fullphase space (Ott and Sommerer 1994) Hence in the present setting a blowout bifurcation isobserved if C falls below 01
We note two other observations Increasing rNMDA which corresponds to increasing thepopulation of NMDA receptors decreases the coupling strength required to achieve stablesynchronization On the other hand increasing the overall excitatory-to-excitatory synapticconnectivity (aee) has the opposite effect This is because the effect of local excitatory feedbackincreases the tangential Lyapunov exponents which has the effect of lsquosplittingrsquo the synchronizeddynamics
In the above simulations all the parameters of both systems were equal In realphysiological systems such symmetry between systems is clearly impossible We nowconsider the case in which the two columns have different structures A simple way toincorporate this whilst at the same time preserving (as much as possible) simple dynamics is tovary the parameter δV for each column This has the effect of destroying any symmetry-forcedinvariant manifolds and so the simple case of synchronization corresponding to identicallyevolving dynamics will not exist in this case However more exotic forms of synchronizationare still possible and we discuss these next
42 Generalized chaotic synchronization
In order to maintain consistency with the literature we study non-linear interdependence in thesetting where the systems are asymmetrical but the coupling is unidirectional This is easilyachieved in the present system by making C directed and setting C21 = 0 In this settingwe refer to the lsquofreersquo system as the driver and the lsquoslavedrsquo system as the response We firstask the question what is the effect of introducing a slight parameter mismatch into a systemexhibiting stable chaos synchronization An example of this is presented in figure 9 whereC12 = 04 and parameter mismatch has been incorporated by setting δV = 07 in the driversystem and δV = 0699 in the response system In panel (a) the time series appear qualitativelyvery similar However in panel (b) where the difference between the systems is plotted it isapparent that the separation between the two systems is of the same order of magnitude as thesize of each attractor In panel (c) the effect of the parameter mismatch on the synchronizationmanifold can be seen as a transformation away from the symmetry manifold
It has been proposed that in such a setting there exists a continuous functional relationshipbetween the dynamics of the driver and response systems
x2(t) = ψ(x1(t)) (10)
where xi = V i W i Z i This situation has been referred to in the literature as generalizedsynchronization (Afraimovich et al 1986 Rulkov et al 1995) The properties of ψ andthe topology and stability of the synchronization manifold in this context have not yetbeen fully elucidated although some significant results have been achieved For example
Synchronization in a model of neuronal dynamics 717
1
2
2
1
Figure 9 Generalized synchronization in an asymmetrical driverndashresponse system (a) The timeseries of the two systems (b) The distance between the two systems (c) A plot of concurrent meanexcitatory membrane potentials V 1 against V 2 The diagonal represents the symmetry manifold
718 M Breakspear et al
Hunt et al (1997) Josic (1998) Kocarev et al (2000) have shown that the generalizedsynchronization manifold is smooth (differentiable) if and only if it is normally hyperbolic(ie the largest transverse Lyapunov exponent is less than the smallest tangential Lyapunovexponent prior to the introduction of parameter mismatch) Such a condition is met in thepresent setting when C = 04 hence the smoothness of the manifold in panel (c)
However subsequent to the introduction of parameter mismatch we will not be able touse transverse and tangential Lyapunov exponents to detect such synchronization since thesynchronization manifold may possess a complex topology for which it is a non-trivial (atbest) or impossible (at worse) task to define transverse and tangential directions For examplethe manifold may be smooth as in figure 9(c) but have a fractal topology If the dynamics arenot normally hyperbolic then the manifold may be fractal but also non-differentiable (Huntet al 1997) and hence will not have a unique tangent direction at any point Consequently wewill not be able to compute tangential or transverse Lyapunov exponents and hence determinethe stability of the synchronization manifold However other techniques for detecting thepresence of generalized synchronization in complex systems have evolved including theauxiliary system approach (Abarbanel et al 1996) and non-linear interdependence (Schiffet al 1996) which are now explored
421 The auxiliary system approach The auxiliary system is a third subsystem which isa replica of the original response system but coupled independently to the driver system andcommencing from independent initial conditions It is possible to demonstrate that showingthe existence of generalized synchronizationmdashthat equation (10) is satisfiedmdashis equivalent toshowing that the response system and the auxiliary system evolve identically (even though theyare not directly coupled) To determine whether or not this is the case reduces to an analysisof the Lyapunov exponents transverse to the symmetry manifold with respect to the responsesystem and the auxiliary system
Whilst this is a somewhat artificial test for generalized synchronization since theproduction of an auxiliary system for a physical system would be impossible such a testis useful in numerical and theoretical studies In figure 10 we present an example of theauxiliary system approach where the parameter δV = 07 for column 1 and 068 for column 2We consider two values of the coupling C = 0025 and 01 The auxiliary system is simplya copy of the second column beginning from an arbitrary initial state In the first instancewhere C = 0025 we notice that even after allowing for an initial transient state the secondcolumn and the auxiliary do not behave in identical fashions (figure 10(a)) However whenC = 01 we observe (figure 10(b)) that the second column and the auxiliary do behave innear-identical manners after an initial transient and hence conclude that for the particularvalue of the coupling strength considered equation (10) is satisfied and hence generalizedsynchronization between the first and second columns occurs
422 Nonlinear interdependence The question remains as to how we may detect thepresence of generalized synchronization in a practical setting As we mentioned aboveit is not possible to use an auxiliary-system-based approach to detect the presence ofgeneralized synchronization Instead we must consider techniques for detecting suchbehaviour from experimental or numerical time series data Algorithms of this type areoften referred to as detecting non-linear interdependence Nonlinear interdependence is ageneral term encompassing systems where the behaviour of one non-linear system affectsthe behaviour of another More specifically it can be defined as occurring if the nullhypothesis that the interdependence between two signals is purely linear can be rejectedat conventional levels (usually 95) of statistical significance hence it is an experimental
Synchronization in a model of neuronal dynamics 719
2
1
2
1
Figure 10 Concurrent evolution of the response system V 2 and lsquoauxiliaryrsquo system V 1 (a) C =0025 (b) C = 01
term (eg Schiff et al 1996) Whilst this does not strictly correspond to the existence of afunctional relationship between the two systems such a relationship is typically used tomotivate the techniques for its detection Schiff et al (1996) extended the logic of non-linearprediction algorithms from autonomous to coupled systems by exploiting the continuity ofsuch a relationship and hence devised the first method of detecting non-linear interdependencein time series data The basis of the technique is using the forward evolution of a phase spacevectorrsquos neighbours to predict the forward evolution of that vector and the forward evolutionof the concurrent vector of the other (possibly interdependent) system If the error of such aprocedure is better on average than chance (the chance error is normalized to equal one) thennon-linear interdependence is said to be present
720 M Breakspear et al
V1
V2
Figure 11 Experimental and numerical examples of weak generalized non-linear synchronizationin the presence of measurement noise (a) Human scalp EEG data from posterior bipolar scalpelectrodes (O1P3ndashO2P4) (b) Weakly coupled aperiodic data in the present model with additivewhite noise (with 5 of the amplitude of the model signal) Parameters δ1
V = 0605 δ2V = 061
aee = 036 C = 007
A variation of this algorithm optimized to detect weak non-linear interdependence (Terryand Breakspear 2003) was used recently to demonstrate that weak non-linear interdependence isoccasionally present in the scalp EEG of healthy resting human subjects (Breakspear and Terry2002a 2002b 2002c) An example of an epoch of lsquonon-linearly independentrsquo EEG obtainedusing this technique is presented in figure 11(a) For comparison output from the currentmodel is presented in figure 11(b) The model parameters were set such that the dynamicswas still aperiodic but the intermittent large-amplitude bursts were suppressed and had weakmutual coupling (C = 007) Extraneous noise to simulate the effect of extracranial muscleactivity contaminating the EEG data has been added to this time series It should be notedthat the amplitudes and time scales of the modelrsquos variables are dimensionless Nonethelessthe model facilitates qualititative interpretation of experimental findings
Synchronization in a model of neuronal dynamics 721
43 Chaotic phase synchronization
Whereas the concept of generalized synchronization was mathematically motivated and assuch more suited to theoretical investigations of systems the concept of phase synchronizationderives from a more practical signal processing approach Phase synchronization occurs inboth symmetric and asymmetric settings but is often more useful in the asymmetric case (andin experimental data)
Unfortunately the phase of a chaotic signal is typically ambiguous and can be defined in anumber of different ways For example in the case of the coherent Rossler attractor (Osipovet al 1997) the phase may be defined as the arctangent of two of the variables of the systemMore generally it can sometimes be defined as the angle that an orbit subtends with the centreof an attractor It is also possible to extract the phase of a chaotic signal from time seriesdata by use of the Hilbert transform This has enabled the demonstration of chaotic phasesynchronization in a number of coupled asymmetrical systems (Rosenblum et al 1996 1997Zaks et al 1999) Although such an approach does not guarantee that the phase will be uniqueit does permit the practical analysis of phase in noisy time series (Tass et al 1998)
To illustrate phase synchronization in two coupled columns we again vary the parameterδV which if the columns are viewed as oscillators acts to detune them from a commonfrequency An important question is that of the criteria on the basis of which we concludewhether phase synchronization occurs We use a simple locking criterion whereby phasesynchronization is said to occur if
|θ1(t)minus θ2(t)| lt K (11)
where is θi the phase of column i and K is a constant Notice that this does not mean thatsynchronization implies equality of phases merely that their difference is bounded
An example of phase synchronization with δV = 065 for column 1 and 066 for column 2is given in figure 12 In this figure rNMDA = 03 Qualitatively some sort of phase coherenceis evident in the time series panel (a) This was confirmed by plotting the sine of thephase difference which has been extracted using the Hilbert transform panel (b) Clearly|sin(θ1 minus θ2)| lt K and hence |θ1 minus θ2| is bounded It can also be seen that the phase-synchronized dynamics occurs on a highly structured manifold depicted in figure 12(c)However unlike the case for the generalized synchronization manifold depicted in figure 9 itis evident that this manifold is not a simple transformation of the symmetry manifold
It is also possible to see lsquobimodalrsquo stability whereby a system jumps betweenapproximately zero phase difference and a phase lag phase synchronization of π An exampleis depicted in figure 12(d) At approximately 2000 s the system can be seen to jumpspontaneously from the first to the second of these modes The Z -variable is plotted forgraphical purposes
5 Intermittency and desynchronization
51 Desynchronization in an array of two systems
Diminishing the strength of coupling across the blowout bifurcation does not lead to a simpletransition from synchronous to asynchronous oscillations but is instead associated withcomplex temporal behaviours Specifically when the largest transverse Lyapunov exponent isclose to zero the coupled systems exhibit epochs of synchronous behaviours punctuated bysharp bursts of desynchronization This is observed in systems exhibiting identicalgeneralizedand phase synchronization An example of such a phenomenon is given in figure 13 The timeseries panel (a) shows synchronous dynamics except at time = 1000 where it can be seenthat the two time series desynchronize briefly This is evident in panel (b) as a disruption in the
722 M Breakspear et al
2
1
Figure 12 Phase-synchronized coupled columns (a) The time series of V 1 (solid) and V 2 (b) Thesine of the phase difference obtained through Hilbert transformation of the time series in panel (a)(c) The phase synchronization manifold in the V 1ndashV 2 plane (d) The time series of Z (solid) andZ showing a phase slip of π between two modes
Synchronization in a model of neuronal dynamics 723
Figure 13 An example of phase desynchronization in coupled asymmetric systems with C = 01Time series plots of (a) each V -variable and (b) their Euclidean distance (c) The temporal evolutionof the phase difference between the two systems (d) An example of the phase difference evolutionover a longer time interval
724 M Breakspear et al
orderly evolution of the distance between the two systems and in panel (c) where the evolutionof the phase difference reveals a brief violation of the criterion |φ1 minus φ2| lt constant Anexample of the phase difference evolution over a longer time interval is shown in figure 13(d)Note that this is a three-dimensional plot with sin(φ1 minus φ2) and cos(φ1 minus φ2) as they- and z-axes Hence phase desynchronizations are evident as complete revolutions aroundthe cylindrical phase space in which this variable evolves It can be seen that these phenomenaoccur at irregular intervals in the temporal domain
This behaviour can be explained by considering the nature of a chaotic attractor In thestatistical sense an arbitrary initial condition is expected to lead to the chaotic behaviour onthe attractor Consequently when we calculate the Lyapunov exponents of the system theseexponents correspond to those of the chaotic attractor Typically however there are an infinitenumber of unstable periodic orbits embedded within the attractor If the path of a trajectorybecomes close to one of these orbits it may lsquoshadowrsquo the orbit for a brief period of time Eachof these orbits has a corresponding set of Lyapunov exponents which (like the attractor) can bedivided into tangential and transverse exponents These also vary when we change the couplingparameter but are typically not the same as the exponents of the attractor Hence the largesttransverse exponent of the attractor may be negative yet there may be positive transverseexponents associated with periodic orbits Considering all of these exponents together wewould therefore only expect to observe complete stability of the synchronization manifold inthe case where all of the transverse Lyapunov exponents are negative Bursting behaviourwill be observed when the one of these exponents becomes positive typically correspondingto a periodic orbit of low order (Pikovsky and Grassberger 1991 Rulkov and Sushchik 1997Maistrenko et al 1998 Heagy et al 1998) Due to the complex structure of such periodic orbitsthe desynchronous bursts occur across a wide range of timescales
In figure 14 we present the results of a long integration (16times105 time units) in which thelengths of sequential synchronous epochs of phase synchronization are studied The parametervalues were δV = 065 for column 1 and 0645 for column 2 with coupling strength10 equal to0125 In panel (a) the power spectrum of these sequential lengths is plotted The whiteness ofthis spectrum indicates that the lengths of consecutive synchronous epochs are independentThis is due to the high-dimensional lsquomixingrsquo effect of desynchronization In panel (b) thelengths of the synchronous epochs have been sorted again into the order of increasing lengthThe results for two coupling strengths are plotted C = 0125 (solid) and 0135 (dashed)Several features of this plot are of note Firstly the lengths of the synchronous epochsare distributed over almost three orders of magnitude Secondly for both plots there is anapproximately log-linear relationship between the length of an epoch and its relative orderThis reflects an approximately exponential relationship between the length of a synchronousepoch and its likelihood of occurrence Thirdly this relationship is skewed toward longersynchronous epochs for the stronger coupling strengths
This type of signal with patches of lsquoorderrsquo occurring over many timescales interruptedby bursts of disorder can be shown to have a high information capacity (Gatlin 1972)Moreover complex phase desynchronization may make an infrequent yet robust contributionto the dynamical pattern of the alpha rhythm in scalp EEG data for resting human subjects(Breakspear 2002) It is tempting to postulate that such a lsquoweakly stablersquo dynamics may assistin the generation of the rich and fluid mental associations that spontaneously emerge in restingcognitive states Although this phenomenon has been previously described in a variety of othersystems to our knowledge this is the first description in a biophysical model of neocortex
10 The introduction of parameter mismatch causes an increase in the strength of coupling required for stablesynchronization above the value of 01 required in the symmetric case
Synchronization in a model of neuronal dynamics 725
Figure 14 Properties of sequential epochs of phase-synchronous behaviour over a long integrationtime δ1
V = 065 δ2V = 0645 aee = 04 (a) The flat (white) power spectrum (b) Synchronous
epochs sorted in order of increasing length (solid C = 0125 dashed C = 0134)
52 Synchronization and complex dynamics in larger arrays
The synchronous and complex phenomena described above are also observed in arrays ofcoupled systems with N gt 2 Expanding the size of the array permits complex dynamicsin the spatial in addition to the temporal domain However the basic principles governingthe type and stability of synchronized motions are the same In this section a few of thepossible behaviours are discussed Physiological variation across the array is modelled bymodifying the parameter δV = 07 in each array by adding a random number of maximummagnitude 001
In figure 15(a) an array of N = 20 uncoupled (C = 0) systems is shown The lack ofsynchronization in this time series is evident By comparison the rapid transition to completesynchronization is evident in figure 15(b) when C = 03 In fact 19 of the 20 subsystems
726 M Breakspear et al
(b)
(c)
(a)
Figure 15 Synchronization phenomena in an array of coupled cortical columns (a) N = 20C = 0 (b) N = 20 C = 03 (c) N = 6 C = 01 (d) N = 12 C = 009 (e) N = 12C = 012 A small perturbation has been applied to one of the columns at t = 2500 s (f) Evolutionof the phase difference between two of the columns in panel (e) For clarity the W -variable isplotted in panel (c) and the Z -variable in (e)
have phase synchronized within one single cycle Intermediate values of coupling strength areassociated with a rich variety of phenomena For example subsystems with the most similarparameter values typically synchronize into clusters of various sizes An example of this isillustrated in figure 15(c) Note that there is no synchronization between the two differentclusters each containing three synchronized subsystems
Another complex behaviour occurs when subsystems transiently synchronize into largeclusters then spontaneously desynchronize again Hence the dynamics of the entire arrayjumps between different transient dynamics characterized by varying degrees of freedom Anexample of this phenomenon is given in figure 15(d) In addition it is possible to inducedesynchronization across an entire array by applying a small perturbation to one columnfor one time step An example is illustrated in figure 15(e) A small perturbation has been
Synchronization in a model of neuronal dynamics 727
Figure 15 (Continued)
added to one column after 2500 time units It can be seen that the entire array subsequentlydesynchronizes This illustrates the sensitivity of the dynamics to external stimuli Finally it ispossible to lsquoinducersquo synchronization between selected subsystems by increasing the strength ofcoupling between them selectively This allows an extra level of complexity to be introducedinto the model For example it may allow targeted information transfer between a partialnetwork of the entire array
As discussed in section 51 a univariate signal which jumps between epochs of lowentropy (high repetition) and bursts of high entropy (high variability) can be shown to havehigh information capacity This is because the repetition of certain patterns (as in the English
728 M Breakspear et al
language) permits error detectability Bursts of high entropy act in the opposite directionto increase message variability With a multivariate signal in addition to optimizing theinformation capacity within each signal the overall information capacity can be increasedthrough the type of itinerant synchronization briefly illustrated in this section Synchronizationacross an entire array increases the message reliability (since a single lsquoerrant letterrsquo in onesignal can be easily detected) Conversely desynchronization into multiple clusters increasesmessage variability since each cluster will produce independent information Hence a neuralarray which switches dynamically between a range of cluster configurations can be seen asadaptive in the sense that high synchronous states can be expressed when error detectabilityis a priority (such as during quick and reliable execution of a familiar task) but the array canrapidly desynchronize if variability becomes salient (for example if the environment suddenlybecomes novel) A formal operationalized demonstration of this principle is to be the subjectof future work
6 Discussion
In this paper we present a dynamical model for activity in an array of weakly coupledlocal neural assemblies The model incorporates the fundamental principles governing cellmembrane potential as determined by voltage- and ligand-gated ion channels In this wayit allows a reasonable amount of physiological complexity to be incorporated includingsynaptic activation and receptor subtypeswithout recourse to overwhelming detail Numericalanalysis of this model reveals a transition from independent oscillations to stable chaoticsynchronization depending on the balance of local versus long-range connectivity andbiophysical heterogeneity These excitatory couplings are subject to ascending subcorticalneuromodulation arising principally from monoaminergic nuclei such as Locus Coeruleus(Silberstein 1995) Neuromodulation of extrinsic connections was incorporated into the modelvia a coupling parameter C The stability of the synchronization is determined by the valueof the largest transverse Lyapunov exponent which in turn depends on the coupling parameterC If the coupling is weakened with the result that this exponent becomes positive a blowoutbifurcation occurs and independent oscillations re-emerge In the vicinity of the transitionnon-linear desynchronizations with complex temporal and spatial properties are observed
It is pleasing that complex and itinerant behaviour reminiscent of real neuronaldynamics emerges through such a simple manipulation of a single dynamic control parameterFurthermore the ubiquitous projections of classical neuromodulatory systems in real brainssuggest a clear biological mechanism by which the brain could self-organize throughmoderating the complexity of its own dynamics There are many reasons that the complicateddynamics of the sort demonstrated in this paper may be neurobiologically important Wemention four
First the presence of multiple clusters of phase-synchronized subsystems as discussedabove may be a uniquely non-linear mechanism facilitating the lsquocarriagersquo and distinction ofmultiple concurrent dynamic cell assemblies (and hence cognitive processing) This may beimportant for perceptual grouping binding and transitions as suggested by others (eg von derMalsburg 1985 Gray and Singer 1989)
Second the facility to decouple and reconfigure dynamic cell assemblies througha neuromodulatory control parameter may be important for avoiding low-entropysynchronization manifolds In the absence of itinerancy synchronization would renderneuronal dynamics incapable of representing changes in sensory input or indeed cognitive stateFor example in the model it was observed that small perturbations to one column were ableto induce a global change in neuronal dynamics by disrupting a state of weak synchronization
Synchronization in a model of neuronal dynamics 729
Inevitably alternative dynamic configurations emerge epitomizing the importance of lsquolabilersquobrain dynamics (Friston 2000a)
Thirdly from a purely functionalist point of view the balance between functionalspecialization and functional integration may rely upon the sorts of mechanism modelledin this paper The brain appears to adhere to two principles of functional organization namelysegregation of function within cortical areas and sub-areas and the integration of these areasin perceptual synthesis cognition and sensorimotor integration (Tononi et al 1994 Fristonet al 1995) The balance between preserving specialized dynamics unique to each arearequired by specialization and the coupling among these areas mediating functional integrationmay be achieved by the neuromodulatory mechanisms described above Significantlythe spontaneous variation in the numbers of degrees of freedom (for example whenlow-dimensional synchronization manifolds are shadowed) imparts a dynamic characteristicto this perspective that is precluded by linear mechanisms alone
Finally from a mechanistic perspective complexity and diversity of brain dynamics arerequired for selection of adaptive responses (Friston 2000b) There are many examples ofselective mechanisms operating within the brain All can be seen as performing an ascenton some objective function or fitness landscape These mechanisms require complex anditinerant dynamics to facilitate a thorough exploration of the landscape These landscapesmay range from those involved in perception (where the landscape is defined by predictionerror see eg Rao and Ballard (1998)) to those involved in value learning (where thelandscape is determined by the outcome of a behaviour eg in temporal difference models (seeeg Sutton and Barto 1990)) In short complexity is necessary to maintain an information-rich environment in which the brain can respond to sensory inputs An essential componentof this is the maintenance of diversity which is a prerequisite for selectionist mechanismsat many scales (Edelman 1993) Understanding the mechanistic basis of complex dynamicsas measured electrophysiologically may well depend on the sort of biologically plausiblemodels used in this paper This understanding may be important for both basic and clinicalneuroscience (eg Tass 2002)
The present study supplements and advances on other research in this field The studyof complex phase synchronization and clustering in neural oscillator networks similar tothe present model has previously been the subject of formal mathematical study Forexample Terman and Wang (1995) studied synchronization of blocks of neural oscillatorsthat were subject to perturbation by connected lsquostimulirsquo Somers and Kopell (1995) studiedthe emergence of both synchronization and travelling waves in networks of coupled neuraloscillators In the work of Terman and Wang the local dynamics were based on neurallsquocaricaturersquo models such as the FitzHughndashNagumo equations and are hence more abstract thanthe current study The approach of Somers and Kopell only required that the local dynamicsexhibit fast and relaxation phases and is hence more abstract still In both studies onlyperiodic dynamics are studied Complex patterns of chaos synchronization in neural arrayshave been characterized by nonlinear methods (Kaneko 1998) and their putative contributionto both learning (Nakajima and Ueda 1996) and perception (Zhao and Macau 2001 Breakspear2001) explored However these studies employed models such as the logistic equation andtent map which are quite abstract in comparison to the present study On the other handHansel and Sompolinsky (1996) studied the role of chaos synchronization in a model ofvisual perception in a cortical hypercolumn using a very similar approach to that adoptedhere but with the additional complexity that synaptic delays were incorporated However inthis and earlier work (Hansel and Sompolinsky 1992) only linear measures of dynamics andsynchrony (specifically the auto- and cross-correlation functions) were employed The analysisof chaotic synchrony in the study of Larter et al (1999) was based only upon observation of
730 M Breakspear et al
the Euclidean difference between variables from two nodes within a larger array Hence thesignificance of the present work is that it provides a quantitative nonlinear analysis of complexchaos synchronization in a biophysical model of cortical dynamics Such research is requiredin order to bridge the gap between quantitative nonlinear research into synchronization andcomputation in abstract models of neural arrays and qualitative or linear analysis in morebiophysically plausible models
Acknowledgments
The authors would like to thank Peter Robinson for assistance with the manuscript and R Larterfor helpful advice MB is a recipient of a NSW Institute of Psychiatry Research Grant anda University of Sydney SESQUI postdoctoral fellowship KJF is supported by the WellcomeTrust
References
Abarbanel H Rulkov N and Sushchik M 1996 Generalised synchronization of chaos the auxiliary system approachPhys Rev E 53 4528ndash35
Afraimovich V Verichev N and Rabinovich M 1986 Stochastic synchronization of oscillation in dissipative systemsRadiophys Quantum Electron 29 795ndash801
Breakspear M 2001 Perception of odours by a nonlinear model of the olfactory bulb Int J Neural Syst 11 101ndash24Breakspear M 2002 Nonlinear phase desynchronisation in human electroencephalographic data Human Brain Mapping
15 175ndash98Breakspear M and Terry J R 2002a Nonlinear interdependence in neural systems motivation theory and relevance
Int J Neurosci 112 1163ndash84Breakspear M and Terry J R 2002b Detection and description of nonlinear interdependence in normal multichannel
human EEG Clin Neurophysiol 113 735ndash53Breakspear M and Terry J R 2002c Topographic organization of nonlinear interdependence in multichannel human
EEG Neuroimage 16 822ndash35Destexhe A Rudolph M Fellous J M and Sejnowski T J 2001 Fluctuating synaptic conductances recreate in vivo-like
activity in neocortical neurons Neuroscience 107 13ndash24Douglas R J Mahowald M A and Martin K A 1996 Microarchitecture of neocortical columns Brain TheorymdashBiological
Basis and Computational Principles ed A Aertsen and V Braitenberg (Amsterdam Elsevier)Eckhorn R Bauer R Jordan W Brosch M Kruse W Munk M and Reitboeck H J 1988 Coherent oscillations a
mechanism of feature linking in the visual cortex Multiple electrode and correlation analyses in the cat BiolCybern 60 121ndash30
Edelman G M 1993 Neural Darwinism selection and reentrant signalling in higher brain function Neuron 10 115ndash25Engel A K Kreiter A K Konig P and Singer W 1991 Synchronization of oscillatory neuronal responses between
striate and extrastriate visual cortical areas of the cat Proc Natl Acad Sci USA 88 6048ndash52Friston K J 2000a The labile brain I Neuronal transients and nonlinear coupling Phil Trans R Soc B 355 215ndash36Friston K J 2000b The labile brain II Transients complexity and selection Phil Trans R Soc B 355 237ndash52Friston K J Tononi G Sporns O and Edelman G 1995 Characterising the complexity of neuronal interactions Human
Brain Mapping 3 302ndash14Fujisaka H and Yamada T 1983 Stability theory of synchronized motion in coupled-oscillator system Prog Theor
Phys 69 32ndash47Gatlin L 1972 Information Theory and the Living System (New York Columbia University Press)Gray C M Konig P Engel A and Singer W 1989 Oscillatory responses in cat visual cortex exhibit inter-columnar
synchronization which reflects global stimulus properties Science 338 334ndash7Gray C M and Singer W 1989 Stimulus specific neuronal oscillations in orientation columns of cat visual cortex Proc
Natl Acad Sci USA 86 1698ndash702Haig A R Gordon E Wright J J Meares R A and Bahramali H 2000 Synchronous cortical gamma-band activity in
task-relevant cognition Neuroreport 11 669ndash75Hansel D and Sompolinsky H 1992 Synchronization and computation in a chaotic neural network Phys Rev Lett 68
718ndash21
Synchronization in a model of neuronal dynamics 731
Hansel D and Sompolinsky H 1996 Chaos and synchrony in a model of a hypercolumn in visual cortex J ComputNeurosci 3 7ndash4
Heagy J Carroll T and Pecora L 1998 Desynchronization by periodic orbits Phys Rev E 57 R1253ndash6Hunt B Ott E and Yorke J 1997 Differentiable synchronization of chaos Phys Rev E 55 4029ndash34Josic K 1998 Invariant manifolds and synchronization of coupled dynamical systems Phys Rev Lett 80 3053ndash6Kandel E Schwartz J and Jessell T 2000 Principles of Neural Science (New York McGraw-Hill)Kaneko K 1998 On the strength of attractors in a high-dimensional system Physica D 124 322ndash44Kocarev L Parlitz U and Brown R 2000 Robust synchronization of chaotic systems Phys Rev E 61 3716ndash20Larter R Speelman B and Worth R M 1999 A coupled ordinary differential equation lattice model for the simulation
of epileptic seizures Chaos 9 795ndash804Lumer E Edelman G and Tononi G 1997a Neural dynamics in a model of the thalamocortical system I Layers loops
and the emergence of fast synchronous oscillations Cereb Cortex 7 207ndash27Lumer E Edelman G and Tononi G 1997b Neural dynamics in a model of the thalamocortical system II The role of
neural synchrony tested through perturbations of spike timing Cereb Cortex 7 228ndash36Magleby K 1987 Short-term modulation at synapses Synaptic Function ed G M Edelman W E Gall and
W M Cowan (New York Wiley)Maistrenko Y Maistrenko V Popovich A and Mosekilde E 1998 Transverse instability and riddled basins in a system
of two coupled logistic maps Phys Rev E 57 2713ndash24Morris C and Lecar H 1981 Voltage oscillations in the barnacle giant muscle fiber Biophys J 35 193ndash213Mountcastle V B 1997 The columnar organization of the neocortex Brain 120 701ndash22Mountcastle V B 1998 The Cerebral Cortex (Cambridge MA Harvard University Press)Nakajima H and Ueda Y 1996 Riddled basins of the optimal states in learning dynamical systems Physica D 99 35ndash44Nunez P L 1995 Neocortical Dynamics and Human Brain Rhythms (Oxford Oxford University Press)Osipov G V Pikovsky A S Rosenblum M G and Kurths J 1997 Phase synchronization effects in a lattice of nonidentical
Rossler oscillators Phys Rev E 55 2353ndash61Ott E and Sommerer J 1994 Blowout bifurcations the occurrence of riddled basins and onndashoff intermittency Phys
Lett A 188 39ndash47Pecora L and Carroll T 1990 Synchronization in chaotic systems Phys Rev Lett 64 821ndash4Pikovsky A and Grassberger P 1991 Symmetry breaking bifurcation for coupled chaotic attractors J Phys A Math
Gen 24 4587ndash97Rao R P and Ballard D H 1998 Predictive coding in the visual cortex a functional interpretation of some extra-classical
receptive field effects Nature Neurosci 2 79ndash87Robinson P A Wright J J and Rennie C R 1998 Synchronous oscillations in the cerebral cortex Phys Rev E 57
4578ndash88Rodriguez E George N Lachauz J P Martinerie J Renault B and Varela F J 1999 Perceptionrsquos shadow long distance
synchronization of human brain activity Nature 397 430ndash3Rosenblum M Pikovsky A and Kurths J 1996 Phase synchronization of chaotic oscillators Phys Rev Lett 76 1804ndash7Rosenblum M Pikovsky A and Kurths J 1997 From phase to lag synchronization in coupled chaotic oscillators Phys
Rev Lett 78 4193ndash6Rulkov N and Sushchik M 1997 Robustness of synchronized chaotic oscillations Int J Bifurc Chaos 7 625ndash43Rulkov N Sushchik M Tsimring L and Abarbanel H 1995 Generalized synchronization of chaos in unidirectionally
coupled chaotic systems Phys Rev E 51 980ndash94Schiff S So P Chang T Burke R and Sauer T 1996 Detecting dynamical interdependence and generalized synchrony
through mutual prediction in a neural ensemble Phys Rev E 54 6708ndash24Silberstein R B 1995 Neuromodulations of neocortical dynamics Neocortical Dynamics and Human EEG Rhythms
ed P L Nunez (Oxford Oxford University Press)Singer W 1995 Putative functions of temporal correlations in neocortical processing Large-Scale Neuronal Theories
of the Brain ed C Koch and J Davis (London MIT Press)Somers D and Kopell N 1995 Waves and synchrony in networks of oscillators of relaxation and non-relaxation type
Physica D 89 169ndash83Sutton R S and Barto A G 1990 Time derivative models of Pavlovian reinforcement Learning and Computational
Neuroscience Foundations of Adaptive Networks ed M Gabriel and J Moore (Cambridge MA MIT Press) pp497ndash538
Szentagothai J 1983 The modular architectonic principle of neural centers Rev Physiol Biochem Pharmacol 9811ndash61
Tallon-Baudry C Betrand O and Fischer C 2001 Oscillatory synchrony between human extrastriate areas during visualshort-term memory maintenance J Neurosci 21 RC177(1ndash5)
Tass P A 2002 Desynchronization of brain rhythms with soft phase-resetting techniques Biol Cybern 87 102ndash15
732 M Breakspear et al
Tass P Rosenblum M Weule J Kurths J Pikovsky A Volkmann J Schnitzler A and Freund H 1998 Detection of nmphase locking from noisy data application to magnetoencephalography Phys Rev Lett 81 3291ndash4
Terman D and Wang D 1995 Global competition and local cooperation in a network of neural oscillators Physica D81 148ndash76
Terry J and Breakspear M 2003 An improved algorithm for the detection of dynamical interdependence in bivariatetime-series Biol Cybern 88 129ndash36
Tononi G Sporns O and Edelman G M 1994 A measure for brain complexity relating functional segregation andintegration in the nervous system Proc Natl Acad Sci USA 91 5033ndash7
von der Malsburg C 1985 Nervous structures with dynamical links Phys Chem 89 703ndash10Wang X 1991 Period-doublings to chaos in a simple neural network an analytic proof Complex Syst 5 425ndash41Wennekers T and Pasemann F 1996 Synchronous chaos in high dimensional modular neural networks Int J Bifurc
Chaos 6 2055ndash67Zaks M A Parks E H Rosenblum M G and Kurths J 1999 Alternative locking ratios in imperfect phase synchronization
Phys Rev Lett 82 4228ndash31Zhao L and Macau E N 2001 A network of dynamically coupled chaotic maps for scene segmentation IEEE Trans
Neural Netw 12 1375ndash85

Synchronization in a model of neuronal dynamics 705
CdV
dt= minusgCamCa(V minus VCa)minus gNamNa(V minus VNa)minus gKW (V minus VK)minus gL(V minus VL) (1)
where gion is the maximum conductance of each population of ion species if all channelsare open m ion is the fraction of channels open and Vion is the Nernst potential for that ionspecies Henceforth following Larter et al (1999) and Destexhe et al (2001) all equationsand parameters are non-dimensional and normalized to neural capacitance C = 1
Each voltage-gated channel acts as a switch which opens when the membrane potentialpasses above a given threshold Tion Hence for a single channel the relationship betweenmembrane voltage and channel conductance is given by a step function Averaging over apopulation of such channels and assuming a Gaussian distribution of Tion around a mean foreach ion species gives the sigmoid-shaped lsquoneural activation functionsrsquo
m ion = 05
(1 + tanh
(V minus Tion
δion
)) (2)
where δion incorporates the variance of this distribution As discussed in Larter et al (1999) theprincipal effect of moving from a microscopic (single-neuron) to a mesoscopic (local network)spatial frame is to further increase the variance of the threshold potentials (consistent with anincrease in their physical heterogeneity) which has the effect of flattening this curve
The fraction of open potassium channels is slightly more complicated The channelslsquorelaxrsquo from one state to another at an exponential rate This is achieved if W is governed by
dW
dt= φ(mK minus W )
τ (3)
where φ is a temperature scaling factor and τ is the lsquorelaxationrsquo time constantThe second modification required for a network formulation is introducing synaptic
interactions between neurons within the same column This is achieved by calculating theaverage cell firing rates and introducing feedback terms subsequent to cell firing to representneurotransmitter release At the cell soma the membrane potential triggers an action potential ifit exceeds a threshold determined chiefly by the sodium channel threshold potential Averagingthis over the ensemble of neurons and assuming once again a Gaussian distribution we obtainthe cell firing rates
QV = 05 times QV max
(1 + tanh
(V minus VT
δV
)) (4)
QZ = 05 times QZ max
(1 + tanh
(Z minus ZT
δZ
)) (5)
where the Qmax are the maximum rates of firing of the excitatory and inhibitory neuronsrespectively
The firing of each of these cell populations feeds back onto the ensemble through synapticcoupling to open ligand-gated channels and raise or lower the membrane potential accordinglyIn the case of excitatory-to-inhibitory and inhibitory-to-excitatory connections this is modelledas additional inputs to the flow of ions across the membrane channel weighted by functionalsynaptic factors aei and aie respectively6 In the case of excitatory-to-excitatory connectionsthe mechanism of this increase in ion conductance is modelled with greater physiological detailincorporating some of the specific characteristics of the ligand-gated channel populations Themean rate of firing QV is assumed to lead to a proportional release of glutamate neurotransmitteracross the synapse which diffuses onto two classes of ligand-gated ion channels One class
6 Note that the feedback terms are multiplied by the potential of the cell of origin This incorporates the potentiatingeffect of residual pre-synaptic intracellular calcium which increases with the pre-synaptic potential (Magleby 1987)
706 M Breakspear et al
Figure 1 A schematic representation of connectivity within cortical columns Panel (a) depictsdense local connectivity between excitatory (EN) and inhibitory (IN) neurons Panel (b) depictsparameters representing strengths of synaptic connectivity between neurons within each columnplus input from ascending neurons within the reticular formation (RF)
embodying AMPA channels open an additional population of sodium channels thus increasingthe net conductance of sodium flow The second class representing NMDA receptors openan additional population of voltage-gated calcium channels Hence this acts to increase themaximum conductance of voltage-gated calcium channels Finally lsquononspecificrsquo subcorticalexcitation is added to both populations of neurons with amplitude I modulated by a randomnoise component of amplitude δ Thus we obtain
dV
dt= minus(gCa + rNMDAaee QV )mCa(V minus VCa)minus (gNamNa + aee QV )(V minus VNa)
minus gKW (V minus VK)minus gL(V minus VL) + aie Z QZ + ane Iδ (6)dZ
dt= b(ani Iδ + aeiV QV ) (7)
where rNMDA denotes the number of NMDA receptors relative to that of AMPA receptors ane
and ani parametrize non-specific input to excitatory and inhibitory cells and aee parametrizesthe local excitatory-to-excitatory synaptic strength Note the difference in the coefficientscontrolling the effect of local excitatory feedback onto NMDA receptors compared to thosefor AMPA receptors A schematic representation of the local network and the parametrizedsynaptic connectivity is presented in figure 1
22 Mesoscopic array dynamics
Each set of equations (2)ndash(7) governs the dynamics within a local cell assembly Couplingbetween N nodes can be introduced parsimoniously as competitive agonist excitatory actionbetween pyramidal cells in different columns However these long-range projections aremodelled to target the same populations of NMDA and AMPA receptors as are targeted by theshort-range excitatory projections Representing each node with a superscript this is achievedby modifying equation (6) to
dV i
dt= minus(gCa + (1 minus C)rNMDAaee Qi
V + CrNMDAaee〈QV 〉)mCa(Vi minus VCa)
minus gKW (V i minus VK)minus gL(Vi minus VL)
minus (gNamNa + (1 minus C)aee QiV + Caee〈QV 〉)(V i minus VNa) + aie Z Qi
Z + ane Iδ (8)
Synchronization in a model of neuronal dynamics 707
where i = 1 N and 〈 〉 represents spatial (lsquomean-fieldrsquo) averaging over cell assembliesC parametrizes the strength of excitatory coupling between cortical columns If C = 0the systems evolve independently C gt 0 introduces interdependences between consecutivecolumns C = 1 corresponds to maximum coupling with excitatory input from outside eachcolumn exceeding excitatory input from within each column
The long-range coupling is lsquoweakrsquo in the sense that we investigate parameter values forwhich C lt aee and C aie etc This either implies that the functional synaptic strength of thelong-range connections are weaker than local connections or that the strengths are similar butthe density of projections is comparatively sparse We prefer the latter interpretation whichis more consistent with other modelling approaches
23 Parameter values and physiological validity
In the sections that follow we set all physiologically measurable parameters (the maximumconductances threshold potentials and Nernst potentials) to within their accepted ranges (Larteret al 1999) Other parameters such as I the functional synaptic strengths and maximum firingrates were chosen to generate dynamically plausible behaviour including the limit cycle andaperiodic dynamics The parameter values are as given in table 1 (unless otherwise stated)There may be significant variability in many of these parameters In this respect it is crucial tonote that the behaviours described below occur in a large region of parameter space Hence theemergent behaviours discussed do not only occur for the parameter values used in this studyBecause as noted above the parameters are dimensionless and normalized to a capacitance ofone it is also important to note that the time units are arbitrary Noting however that the cellcapacitance is typically estimated as close to or equal to 1 microF cmminus2 (eg Destexhe et al 2001)it is reasonable to interpret the time evolution units as milliseconds (as done for the diffusivecoupling parameter employed in Larter et al (1999))
The principal aim of this paper is a quantitative study of non-linear synchronization ina biophysically realistic model of a neural system The model incorporates some aspects ofneural systems with significant detail such as the conductance of K+ glutamate-mediatedneurotransmission and the relatively sparse nature of long-range connectivity This detailallows manipulation of parameters that model pharmacological mechanisms of interest suchas glutamate receptor subclasses Other features such as local inhibitory feedback and thekinetics of Ca2+ and Na+ channels have been incorporated in a more simplified manner Thislimits the complexity of the model but ensures that numerical analysis can be supplementedby quantitative examination of for example the Lyapunov exponents Other biophysicalproperties could be made the subject of future study by changing relevant aspects of theequations
3 Behaviour of local lsquouncoupledrsquo columns
In this section a brief overview of the behaviours exhibited by autonomous local columns(ie C = 0) is presented As indicated in table 1 the dynamics of the model is explored byfixing all parameters except for δV the variance of the excitatory threshold which is varied inthe region [055 07] A similar range of behaviours is also observed if δV is fixed and anothercontrol parameter such as aee or I is varied
31 Fixed point attractors
For values of δV lt 055 the dynamics of a single column settles onto a solitary fixed pointattractor As shown in figure 2 transient dynamics persists for a number of seconds
708 M Breakspear et al
Figure 2 The time series for a fixed point attractor with VT = 054
Table 1 Parameter values unless otherwise stated Adapted from Larter et al (1999)
Parameter Description Value
TCa Threshold value for Ca channels minus001δCa Variance of Ca channel threshold 015gCa Conductance of population of Ca channels 11VCa Ca Nernst potential 1TK Threshold value for K channels 00δK Variance of K channel threshold 030gK Conductance of population of K channels 20VK K Nernst potential minus07TNa Threshold value for Na channels 03δNa Variance of Na channel threshold 015gNa Conductance of population of Na channels 67VNa Na Nernst potential 053VL Nernst potential leak channels minus05gL Conductance of population of leak channels 05VT Threshold potential for excitatory neurons 00δV Variance of excitatory threshold VariedZT Threshold potential for inhibitory neurons 00δZ Variance of inhibitory threshold 070I Subcortical input strength 030aee Excitatory-to-excitatory synaptic strength 04aei Excitatory-to-inhibitory synaptic strength 2aie Inhibitory-to-excitatory synaptic strength 2ane Non-specific-to-excitatory synaptic strength 1ani Non-specific-to-inhibitory synaptic strength 04b Time constant scaling factor 010ϕ Temperature scaling factor 07τ Time constant for K relaxation time 1rNMDA Ratio of NMDA to AMPA receptors 025β Random modulation of subcortical input 0
32 Limit cycle attractors
For 055 lt δV lt 059 the system undergoes a series of period-doubling bifurcations Theseresult in both simple and complex periodic behaviours A limit cycle occurring when δV = 056
Synchronization in a model of neuronal dynamics 709
Figure 3 The approach to a limit cycle attractor in phase space with VT = 056
is illustrated in figure 3 Other parameter variations produce more complicated higher-orderperiodic orbits For example a period-9 orbit occurring when δV = 06 aee = 05 aie = 05gNa = 0 and I = 0165 is presented in figure 4
33 Chaotic attractors
Setting δV gt 059 and keeping all other parameters as per table 1 produces complex aperiodicbehaviour in the uncoupled columns An example of this with δV = 065 is illustrated infigure 5 Panels (a) and (b) illustrate a phase space portrait and a time series plot of thissystem respectively They show an aperiodic but highly structured system In panel (c) afirst-return lsquoPoincarersquo section is presented This was obtained by taking a cross-section throughthe attractor in (a) by setting W = 032 and then plotting V (t) against V (t +l)with a time delayof l = 1 A topographic similarity to the simple logistic map is evident It has been provenanalytically that in restricted regions of parameter space dynamical systems incorporatingthe neural activation function (4) (5) are topologically conjugate to the logistic map (Wang1991) The Poincare map presented here shows that although a 11 map may not exist in moregeneral parameter regions the global morphology of the logistic map is still evident
To determine whether or not these aperiodic oscillations are in fact chaotic the Lyapunovexponents were estimated numerically using an algorithm based upon GrammndashSchmidtorthonormalization For the particular case illustrated the exponents were found to be
λ1 = 0015
λ2 = 00
λ3 = minus026
(9)
A principal Lyapunov exponent greater than zero is consistent with chaotic dynamics Thistogether with the topography of the first-return map and the existence of period-doublingbifurcations provides compelling evidence of chaotic behaviour in the dynamics of the singlecolumn The second exponent reflects the rate of exponential separation of adjacent orbits in
710 M Breakspear et al
Figure 4 A period-9 limit cycle attractor (a) A phase space portrait (b) A representative timeseries
the direction tangent to the flow hence a value of zero is to be expected The sum of the threeexponents is negative implying that the system is volume contracting which is to be expectedin the setting of a non-equilibrium (non-Hamiltonian) physiological system
It is interesting to note that this diversity of behaviours can be obtained throughmanipulation of a single state parameter From a functional perspective this permits eachcortical column to express a variety of qualitatively distinct dynamics through modificationof a single physiological parameter such as through modulation of the functional strength ofthe local synaptic connectivity Conversely it also implies that resetting of a single parametersuch as an abnormal δV resulting from loss of physiological heterogeneity could be associatedwith qualitative changes in local cortical behaviour
4 Cooperative behaviour of coupled columns
Whilst it is interesting to observe the range of dynamic behaviours possible in a single columnwe are primarily interested in the behaviour of many such interacting columns In this section
Synchronization in a model of neuronal dynamics 711
1
Figure 5 Chaotic behaviour of the autonomous system with VT = 065 The other parameters areas per table 1 (a) An attractor in phase space (b) A time series for V (t) (c) A Poincare sectionobtained by setting W = 032 and plotting V (t) against V (t + 1)
712 M Breakspear et al
we study the emergence of cooperative behaviour that results from the introduction of couplingbetween columns Three types of interdependent behaviour of two systems are describedidentical generalized and phase synchronization Synchronous behaviours in larger arrays arealso illustrated
41 Identical chaotic synchronization
411 Transition to identical chaotic synchronization The most straightforward extensionand the one that we consider first is that of two coupled cortical columns where all parametersare in the first case assumed identical7 Trivially the two systems will evolve identicallywhen started with identical initial conditions even if C = 0 This is simply a statement ofthe deterministic nature of the evolution equations In general the random initial conditionswill not be equal in which case it is hardly surprising that the different columns will evolveindependently An example is presented in figure 6 with VT = 06 in both systems C = 0and independent random initial conditions In figure 6(a) we show a segment of the timeseries for each of the two subsystems where it can be seen that the time series are chaoticand asynchronous In figure 6(b) the concurrent values of V 1 and V 2 are plotted against eachother showing that they bear no obvious interrelationship
With the introduction of coupling C gt 0 the systems begin to show epochs ofsynchronized behaviour At C sim= 01 there is a transition to stable fully synchronizedmotion as evident in an exemplar time series shown in figure 7(a) where C = 015 Infigure 7(b) we present the concurrent values of V 1 versus V 2 illustrating that in contrast tothe case in figure 6(b) the systems have become embedded in the low-dimensional lsquosymmetrymanifoldrsquo V 1 = V 2 Panel (c) shows a log-linear plot of the temporal evolution of the distanceof the coupled system from this manifold It can be seen that on average the two systemsapproach at an exponential rate with constant time-averaged exponent Whilst this phenomenon(lsquoidentical chaotic synchronizationrsquo) has been described in a variety of other systems since itsfirst description (Fujisaka and Yamada 1983 Pecora and Carroll 1990) to our knowledge thisis its first quantitative8 description in a detailed physiological model of a neural system
The symmetry of the dynamical system obtained by setting the state parameters in eachcolumn identical forces the invariance of the symmetry manifold V 1 = V 2 Moreover thedynamics within this manifold is governed by the same equations of motion as for a singlecortical column This manifold which contains the orbits of the synchronized system is calledthe synchronization manifold The coupling parameter C does not affect the dynamics withinthe synchronization manifold and as such is termed a normal parameter
412 lsquoBlowout bifurcationrsquo An important issue is the stability of the orbits on thesynchronization manifold This can be determined by analysing the Lyapunov spectraBecause the two-column system evolves in a six-dimensional phase space there are sixLyapunov exponents However for the synchronized system it is helpful to subdivide theseinto those that describe the dynamics within the synchronization manifold and those thatdescribe the dynamics in the transverse direction The latter describe the rate of convergenceor divergence of the coupled systems toward or away from this manifoldmdashand hence thetendency for the two systems to synchronize or separate For the time series in figure 7 this is
7 Whilst it is of course physiologically unrealistic to assume that two columns would have entirely identical structuresit is mathematically simpler to start analysing the system in this situation8 A qualitative description in a neural system with a different form of coupling (potassium diffusion) is evident infigure 8 of Larter et al (1999) Synchronous chaos in an abstract model of a neural system was also described inWennekers and Pasemann (1996)
Synchronization in a model of neuronal dynamics 713
1
2
2
1
Figure 6 Independent oscillations in a two-column system with identical parameters butindependent initial conditions (a) The time series (b) A plot of concurrent mean excitatorymembrane potentials V 1 against V 2
related to the slope of the plot in panel (c)9 For this strength of coupling the largest transverseexponent is negative (minus002) indicating that perturbations away from this state of identicalsynchronization decay and thus that the state of identical chaotic synchronization is stableagainst noise (ie dynamically stable)
To calculate the relationship between coupling strength and the Lyapunov exponents weused an algorithm which measures the growth of perturbations of the variational equations (thelsquoJacobianrsquo) of the system (we do not define these equations here as they are cumbersome anddo not enhance the understanding of the system) We calculated these exponents for increasingvalues of the coupling parameter C from 0 to 02 in increments of 0000 25 The results are
9 Only two of the six dependent variables are shown in figure 7 The actual transverse Lyapunov exponents reflectthe temporal evolution of all six and hence are somewhat more complex than that given by the slope of panel (c)
714 M Breakspear et al
1
2
Figure 7 Identical synchronization in the same system as in figure 7 but with C = 02 (a) Thetime series (b) A plot of concurrent mean excitatory membrane potentials V 1 against V 2 (c) Thelogarithm of the Euclidean distance between V 1 and V 2 versus time (linear)
Synchronization in a model of neuronal dynamics 715
Figure 8 Lyapunov spectra for a coupled system with δV = 07 and C varying from 0 to 02 insteps of 0000 33 At each value of C all Lyapunov exponents are calculated Least-squares linesof best fit were then fitted to the results for each exponent Solid lines indicate transverse exponents(λ) and dashed lines indicate the tangential exponents () Panel (a) shows all three transverseand three tangential exponents Panel (b) only shows the largest two of each
illustrated in figure 8 Panel (a) shows all exponents For illustrative purposes only the mostpositive four are plotted in panel (b) In both panels the tangential exponents () are fitted (inthe least-squares sense) with a dashed line and the transverse ones (λ) are fitted with a solidline They are labelled 1ndash3 in order of decreasing positivity The important features are asfollows For the range of coupling strengths that we consider the largest tangential Lyapunovexponent is positive which indicates the presence of chaotic behaviour within the manifoldWhen examining the line of best fit for this exponent we see that this exponent does not varywhen altering the coupling parameter This is true for all three tangential exponents and verifiesthat the changes in the coupling parameter do not alter the dynamics on the synchronizationmanifold (that is the coupling parameter is normal)
716 M Breakspear et al
The largest transverse Lyapunov exponent (denoted as λ1 in the figure) is a particularlyimportant exponent since it governs the stability of the attractor in the synchronization manifoldwith respect to perturbations out of the manifold If this exponent is negative then we expectsmall perturbations of trajectories to reconverge to the manifold in finite time If however thisexponent is positive then we would instead expect perturbed trajectories to diverge from themanifold This change in local behaviour is termed a bifurcation and the point at which itoccurs the bifurcation point It is apparent in figure 8 that the zero crossing of this exponentmdashthe bifurcation pointmdashoccurs at approximately C = 01 in the present system This typeof bifurcation has been termed a blowout bifurcation since once transverse stability is losttrajectories are lsquoblown outrsquo from the low-dimensional synchronization manifold into the fullphase space (Ott and Sommerer 1994) Hence in the present setting a blowout bifurcation isobserved if C falls below 01
We note two other observations Increasing rNMDA which corresponds to increasing thepopulation of NMDA receptors decreases the coupling strength required to achieve stablesynchronization On the other hand increasing the overall excitatory-to-excitatory synapticconnectivity (aee) has the opposite effect This is because the effect of local excitatory feedbackincreases the tangential Lyapunov exponents which has the effect of lsquosplittingrsquo the synchronizeddynamics
In the above simulations all the parameters of both systems were equal In realphysiological systems such symmetry between systems is clearly impossible We nowconsider the case in which the two columns have different structures A simple way toincorporate this whilst at the same time preserving (as much as possible) simple dynamics is tovary the parameter δV for each column This has the effect of destroying any symmetry-forcedinvariant manifolds and so the simple case of synchronization corresponding to identicallyevolving dynamics will not exist in this case However more exotic forms of synchronizationare still possible and we discuss these next
42 Generalized chaotic synchronization
In order to maintain consistency with the literature we study non-linear interdependence in thesetting where the systems are asymmetrical but the coupling is unidirectional This is easilyachieved in the present system by making C directed and setting C21 = 0 In this settingwe refer to the lsquofreersquo system as the driver and the lsquoslavedrsquo system as the response We firstask the question what is the effect of introducing a slight parameter mismatch into a systemexhibiting stable chaos synchronization An example of this is presented in figure 9 whereC12 = 04 and parameter mismatch has been incorporated by setting δV = 07 in the driversystem and δV = 0699 in the response system In panel (a) the time series appear qualitativelyvery similar However in panel (b) where the difference between the systems is plotted it isapparent that the separation between the two systems is of the same order of magnitude as thesize of each attractor In panel (c) the effect of the parameter mismatch on the synchronizationmanifold can be seen as a transformation away from the symmetry manifold
It has been proposed that in such a setting there exists a continuous functional relationshipbetween the dynamics of the driver and response systems
x2(t) = ψ(x1(t)) (10)
where xi = V i W i Z i This situation has been referred to in the literature as generalizedsynchronization (Afraimovich et al 1986 Rulkov et al 1995) The properties of ψ andthe topology and stability of the synchronization manifold in this context have not yetbeen fully elucidated although some significant results have been achieved For example
Synchronization in a model of neuronal dynamics 717
1
2
2
1
Figure 9 Generalized synchronization in an asymmetrical driverndashresponse system (a) The timeseries of the two systems (b) The distance between the two systems (c) A plot of concurrent meanexcitatory membrane potentials V 1 against V 2 The diagonal represents the symmetry manifold
718 M Breakspear et al
Hunt et al (1997) Josic (1998) Kocarev et al (2000) have shown that the generalizedsynchronization manifold is smooth (differentiable) if and only if it is normally hyperbolic(ie the largest transverse Lyapunov exponent is less than the smallest tangential Lyapunovexponent prior to the introduction of parameter mismatch) Such a condition is met in thepresent setting when C = 04 hence the smoothness of the manifold in panel (c)
However subsequent to the introduction of parameter mismatch we will not be able touse transverse and tangential Lyapunov exponents to detect such synchronization since thesynchronization manifold may possess a complex topology for which it is a non-trivial (atbest) or impossible (at worse) task to define transverse and tangential directions For examplethe manifold may be smooth as in figure 9(c) but have a fractal topology If the dynamics arenot normally hyperbolic then the manifold may be fractal but also non-differentiable (Huntet al 1997) and hence will not have a unique tangent direction at any point Consequently wewill not be able to compute tangential or transverse Lyapunov exponents and hence determinethe stability of the synchronization manifold However other techniques for detecting thepresence of generalized synchronization in complex systems have evolved including theauxiliary system approach (Abarbanel et al 1996) and non-linear interdependence (Schiffet al 1996) which are now explored
421 The auxiliary system approach The auxiliary system is a third subsystem which isa replica of the original response system but coupled independently to the driver system andcommencing from independent initial conditions It is possible to demonstrate that showingthe existence of generalized synchronizationmdashthat equation (10) is satisfiedmdashis equivalent toshowing that the response system and the auxiliary system evolve identically (even though theyare not directly coupled) To determine whether or not this is the case reduces to an analysisof the Lyapunov exponents transverse to the symmetry manifold with respect to the responsesystem and the auxiliary system
Whilst this is a somewhat artificial test for generalized synchronization since theproduction of an auxiliary system for a physical system would be impossible such a testis useful in numerical and theoretical studies In figure 10 we present an example of theauxiliary system approach where the parameter δV = 07 for column 1 and 068 for column 2We consider two values of the coupling C = 0025 and 01 The auxiliary system is simplya copy of the second column beginning from an arbitrary initial state In the first instancewhere C = 0025 we notice that even after allowing for an initial transient state the secondcolumn and the auxiliary do not behave in identical fashions (figure 10(a)) However whenC = 01 we observe (figure 10(b)) that the second column and the auxiliary do behave innear-identical manners after an initial transient and hence conclude that for the particularvalue of the coupling strength considered equation (10) is satisfied and hence generalizedsynchronization between the first and second columns occurs
422 Nonlinear interdependence The question remains as to how we may detect thepresence of generalized synchronization in a practical setting As we mentioned aboveit is not possible to use an auxiliary-system-based approach to detect the presence ofgeneralized synchronization Instead we must consider techniques for detecting suchbehaviour from experimental or numerical time series data Algorithms of this type areoften referred to as detecting non-linear interdependence Nonlinear interdependence is ageneral term encompassing systems where the behaviour of one non-linear system affectsthe behaviour of another More specifically it can be defined as occurring if the nullhypothesis that the interdependence between two signals is purely linear can be rejectedat conventional levels (usually 95) of statistical significance hence it is an experimental
Synchronization in a model of neuronal dynamics 719
2
1
2
1
Figure 10 Concurrent evolution of the response system V 2 and lsquoauxiliaryrsquo system V 1 (a) C =0025 (b) C = 01
term (eg Schiff et al 1996) Whilst this does not strictly correspond to the existence of afunctional relationship between the two systems such a relationship is typically used tomotivate the techniques for its detection Schiff et al (1996) extended the logic of non-linearprediction algorithms from autonomous to coupled systems by exploiting the continuity ofsuch a relationship and hence devised the first method of detecting non-linear interdependencein time series data The basis of the technique is using the forward evolution of a phase spacevectorrsquos neighbours to predict the forward evolution of that vector and the forward evolutionof the concurrent vector of the other (possibly interdependent) system If the error of such aprocedure is better on average than chance (the chance error is normalized to equal one) thennon-linear interdependence is said to be present
720 M Breakspear et al
V1
V2
Figure 11 Experimental and numerical examples of weak generalized non-linear synchronizationin the presence of measurement noise (a) Human scalp EEG data from posterior bipolar scalpelectrodes (O1P3ndashO2P4) (b) Weakly coupled aperiodic data in the present model with additivewhite noise (with 5 of the amplitude of the model signal) Parameters δ1
V = 0605 δ2V = 061
aee = 036 C = 007
A variation of this algorithm optimized to detect weak non-linear interdependence (Terryand Breakspear 2003) was used recently to demonstrate that weak non-linear interdependence isoccasionally present in the scalp EEG of healthy resting human subjects (Breakspear and Terry2002a 2002b 2002c) An example of an epoch of lsquonon-linearly independentrsquo EEG obtainedusing this technique is presented in figure 11(a) For comparison output from the currentmodel is presented in figure 11(b) The model parameters were set such that the dynamicswas still aperiodic but the intermittent large-amplitude bursts were suppressed and had weakmutual coupling (C = 007) Extraneous noise to simulate the effect of extracranial muscleactivity contaminating the EEG data has been added to this time series It should be notedthat the amplitudes and time scales of the modelrsquos variables are dimensionless Nonethelessthe model facilitates qualititative interpretation of experimental findings
Synchronization in a model of neuronal dynamics 721
43 Chaotic phase synchronization
Whereas the concept of generalized synchronization was mathematically motivated and assuch more suited to theoretical investigations of systems the concept of phase synchronizationderives from a more practical signal processing approach Phase synchronization occurs inboth symmetric and asymmetric settings but is often more useful in the asymmetric case (andin experimental data)
Unfortunately the phase of a chaotic signal is typically ambiguous and can be defined in anumber of different ways For example in the case of the coherent Rossler attractor (Osipovet al 1997) the phase may be defined as the arctangent of two of the variables of the systemMore generally it can sometimes be defined as the angle that an orbit subtends with the centreof an attractor It is also possible to extract the phase of a chaotic signal from time seriesdata by use of the Hilbert transform This has enabled the demonstration of chaotic phasesynchronization in a number of coupled asymmetrical systems (Rosenblum et al 1996 1997Zaks et al 1999) Although such an approach does not guarantee that the phase will be uniqueit does permit the practical analysis of phase in noisy time series (Tass et al 1998)
To illustrate phase synchronization in two coupled columns we again vary the parameterδV which if the columns are viewed as oscillators acts to detune them from a commonfrequency An important question is that of the criteria on the basis of which we concludewhether phase synchronization occurs We use a simple locking criterion whereby phasesynchronization is said to occur if
|θ1(t)minus θ2(t)| lt K (11)
where is θi the phase of column i and K is a constant Notice that this does not mean thatsynchronization implies equality of phases merely that their difference is bounded
An example of phase synchronization with δV = 065 for column 1 and 066 for column 2is given in figure 12 In this figure rNMDA = 03 Qualitatively some sort of phase coherenceis evident in the time series panel (a) This was confirmed by plotting the sine of thephase difference which has been extracted using the Hilbert transform panel (b) Clearly|sin(θ1 minus θ2)| lt K and hence |θ1 minus θ2| is bounded It can also be seen that the phase-synchronized dynamics occurs on a highly structured manifold depicted in figure 12(c)However unlike the case for the generalized synchronization manifold depicted in figure 9 itis evident that this manifold is not a simple transformation of the symmetry manifold
It is also possible to see lsquobimodalrsquo stability whereby a system jumps betweenapproximately zero phase difference and a phase lag phase synchronization of π An exampleis depicted in figure 12(d) At approximately 2000 s the system can be seen to jumpspontaneously from the first to the second of these modes The Z -variable is plotted forgraphical purposes
5 Intermittency and desynchronization
51 Desynchronization in an array of two systems
Diminishing the strength of coupling across the blowout bifurcation does not lead to a simpletransition from synchronous to asynchronous oscillations but is instead associated withcomplex temporal behaviours Specifically when the largest transverse Lyapunov exponent isclose to zero the coupled systems exhibit epochs of synchronous behaviours punctuated bysharp bursts of desynchronization This is observed in systems exhibiting identicalgeneralizedand phase synchronization An example of such a phenomenon is given in figure 13 The timeseries panel (a) shows synchronous dynamics except at time = 1000 where it can be seenthat the two time series desynchronize briefly This is evident in panel (b) as a disruption in the
722 M Breakspear et al
2
1
Figure 12 Phase-synchronized coupled columns (a) The time series of V 1 (solid) and V 2 (b) Thesine of the phase difference obtained through Hilbert transformation of the time series in panel (a)(c) The phase synchronization manifold in the V 1ndashV 2 plane (d) The time series of Z (solid) andZ showing a phase slip of π between two modes
Synchronization in a model of neuronal dynamics 723
Figure 13 An example of phase desynchronization in coupled asymmetric systems with C = 01Time series plots of (a) each V -variable and (b) their Euclidean distance (c) The temporal evolutionof the phase difference between the two systems (d) An example of the phase difference evolutionover a longer time interval
724 M Breakspear et al
orderly evolution of the distance between the two systems and in panel (c) where the evolutionof the phase difference reveals a brief violation of the criterion |φ1 minus φ2| lt constant Anexample of the phase difference evolution over a longer time interval is shown in figure 13(d)Note that this is a three-dimensional plot with sin(φ1 minus φ2) and cos(φ1 minus φ2) as they- and z-axes Hence phase desynchronizations are evident as complete revolutions aroundthe cylindrical phase space in which this variable evolves It can be seen that these phenomenaoccur at irregular intervals in the temporal domain
This behaviour can be explained by considering the nature of a chaotic attractor In thestatistical sense an arbitrary initial condition is expected to lead to the chaotic behaviour onthe attractor Consequently when we calculate the Lyapunov exponents of the system theseexponents correspond to those of the chaotic attractor Typically however there are an infinitenumber of unstable periodic orbits embedded within the attractor If the path of a trajectorybecomes close to one of these orbits it may lsquoshadowrsquo the orbit for a brief period of time Eachof these orbits has a corresponding set of Lyapunov exponents which (like the attractor) can bedivided into tangential and transverse exponents These also vary when we change the couplingparameter but are typically not the same as the exponents of the attractor Hence the largesttransverse exponent of the attractor may be negative yet there may be positive transverseexponents associated with periodic orbits Considering all of these exponents together wewould therefore only expect to observe complete stability of the synchronization manifold inthe case where all of the transverse Lyapunov exponents are negative Bursting behaviourwill be observed when the one of these exponents becomes positive typically correspondingto a periodic orbit of low order (Pikovsky and Grassberger 1991 Rulkov and Sushchik 1997Maistrenko et al 1998 Heagy et al 1998) Due to the complex structure of such periodic orbitsthe desynchronous bursts occur across a wide range of timescales
In figure 14 we present the results of a long integration (16times105 time units) in which thelengths of sequential synchronous epochs of phase synchronization are studied The parametervalues were δV = 065 for column 1 and 0645 for column 2 with coupling strength10 equal to0125 In panel (a) the power spectrum of these sequential lengths is plotted The whiteness ofthis spectrum indicates that the lengths of consecutive synchronous epochs are independentThis is due to the high-dimensional lsquomixingrsquo effect of desynchronization In panel (b) thelengths of the synchronous epochs have been sorted again into the order of increasing lengthThe results for two coupling strengths are plotted C = 0125 (solid) and 0135 (dashed)Several features of this plot are of note Firstly the lengths of the synchronous epochsare distributed over almost three orders of magnitude Secondly for both plots there is anapproximately log-linear relationship between the length of an epoch and its relative orderThis reflects an approximately exponential relationship between the length of a synchronousepoch and its likelihood of occurrence Thirdly this relationship is skewed toward longersynchronous epochs for the stronger coupling strengths
This type of signal with patches of lsquoorderrsquo occurring over many timescales interruptedby bursts of disorder can be shown to have a high information capacity (Gatlin 1972)Moreover complex phase desynchronization may make an infrequent yet robust contributionto the dynamical pattern of the alpha rhythm in scalp EEG data for resting human subjects(Breakspear 2002) It is tempting to postulate that such a lsquoweakly stablersquo dynamics may assistin the generation of the rich and fluid mental associations that spontaneously emerge in restingcognitive states Although this phenomenon has been previously described in a variety of othersystems to our knowledge this is the first description in a biophysical model of neocortex
10 The introduction of parameter mismatch causes an increase in the strength of coupling required for stablesynchronization above the value of 01 required in the symmetric case
Synchronization in a model of neuronal dynamics 725
Figure 14 Properties of sequential epochs of phase-synchronous behaviour over a long integrationtime δ1
V = 065 δ2V = 0645 aee = 04 (a) The flat (white) power spectrum (b) Synchronous
epochs sorted in order of increasing length (solid C = 0125 dashed C = 0134)
52 Synchronization and complex dynamics in larger arrays
The synchronous and complex phenomena described above are also observed in arrays ofcoupled systems with N gt 2 Expanding the size of the array permits complex dynamicsin the spatial in addition to the temporal domain However the basic principles governingthe type and stability of synchronized motions are the same In this section a few of thepossible behaviours are discussed Physiological variation across the array is modelled bymodifying the parameter δV = 07 in each array by adding a random number of maximummagnitude 001
In figure 15(a) an array of N = 20 uncoupled (C = 0) systems is shown The lack ofsynchronization in this time series is evident By comparison the rapid transition to completesynchronization is evident in figure 15(b) when C = 03 In fact 19 of the 20 subsystems
726 M Breakspear et al
(b)
(c)
(a)
Figure 15 Synchronization phenomena in an array of coupled cortical columns (a) N = 20C = 0 (b) N = 20 C = 03 (c) N = 6 C = 01 (d) N = 12 C = 009 (e) N = 12C = 012 A small perturbation has been applied to one of the columns at t = 2500 s (f) Evolutionof the phase difference between two of the columns in panel (e) For clarity the W -variable isplotted in panel (c) and the Z -variable in (e)
have phase synchronized within one single cycle Intermediate values of coupling strength areassociated with a rich variety of phenomena For example subsystems with the most similarparameter values typically synchronize into clusters of various sizes An example of this isillustrated in figure 15(c) Note that there is no synchronization between the two differentclusters each containing three synchronized subsystems
Another complex behaviour occurs when subsystems transiently synchronize into largeclusters then spontaneously desynchronize again Hence the dynamics of the entire arrayjumps between different transient dynamics characterized by varying degrees of freedom Anexample of this phenomenon is given in figure 15(d) In addition it is possible to inducedesynchronization across an entire array by applying a small perturbation to one columnfor one time step An example is illustrated in figure 15(e) A small perturbation has been
Synchronization in a model of neuronal dynamics 727
Figure 15 (Continued)
added to one column after 2500 time units It can be seen that the entire array subsequentlydesynchronizes This illustrates the sensitivity of the dynamics to external stimuli Finally it ispossible to lsquoinducersquo synchronization between selected subsystems by increasing the strength ofcoupling between them selectively This allows an extra level of complexity to be introducedinto the model For example it may allow targeted information transfer between a partialnetwork of the entire array
As discussed in section 51 a univariate signal which jumps between epochs of lowentropy (high repetition) and bursts of high entropy (high variability) can be shown to havehigh information capacity This is because the repetition of certain patterns (as in the English
728 M Breakspear et al
language) permits error detectability Bursts of high entropy act in the opposite directionto increase message variability With a multivariate signal in addition to optimizing theinformation capacity within each signal the overall information capacity can be increasedthrough the type of itinerant synchronization briefly illustrated in this section Synchronizationacross an entire array increases the message reliability (since a single lsquoerrant letterrsquo in onesignal can be easily detected) Conversely desynchronization into multiple clusters increasesmessage variability since each cluster will produce independent information Hence a neuralarray which switches dynamically between a range of cluster configurations can be seen asadaptive in the sense that high synchronous states can be expressed when error detectabilityis a priority (such as during quick and reliable execution of a familiar task) but the array canrapidly desynchronize if variability becomes salient (for example if the environment suddenlybecomes novel) A formal operationalized demonstration of this principle is to be the subjectof future work
6 Discussion
In this paper we present a dynamical model for activity in an array of weakly coupledlocal neural assemblies The model incorporates the fundamental principles governing cellmembrane potential as determined by voltage- and ligand-gated ion channels In this wayit allows a reasonable amount of physiological complexity to be incorporated includingsynaptic activation and receptor subtypeswithout recourse to overwhelming detail Numericalanalysis of this model reveals a transition from independent oscillations to stable chaoticsynchronization depending on the balance of local versus long-range connectivity andbiophysical heterogeneity These excitatory couplings are subject to ascending subcorticalneuromodulation arising principally from monoaminergic nuclei such as Locus Coeruleus(Silberstein 1995) Neuromodulation of extrinsic connections was incorporated into the modelvia a coupling parameter C The stability of the synchronization is determined by the valueof the largest transverse Lyapunov exponent which in turn depends on the coupling parameterC If the coupling is weakened with the result that this exponent becomes positive a blowoutbifurcation occurs and independent oscillations re-emerge In the vicinity of the transitionnon-linear desynchronizations with complex temporal and spatial properties are observed
It is pleasing that complex and itinerant behaviour reminiscent of real neuronaldynamics emerges through such a simple manipulation of a single dynamic control parameterFurthermore the ubiquitous projections of classical neuromodulatory systems in real brainssuggest a clear biological mechanism by which the brain could self-organize throughmoderating the complexity of its own dynamics There are many reasons that the complicateddynamics of the sort demonstrated in this paper may be neurobiologically important Wemention four
First the presence of multiple clusters of phase-synchronized subsystems as discussedabove may be a uniquely non-linear mechanism facilitating the lsquocarriagersquo and distinction ofmultiple concurrent dynamic cell assemblies (and hence cognitive processing) This may beimportant for perceptual grouping binding and transitions as suggested by others (eg von derMalsburg 1985 Gray and Singer 1989)
Second the facility to decouple and reconfigure dynamic cell assemblies througha neuromodulatory control parameter may be important for avoiding low-entropysynchronization manifolds In the absence of itinerancy synchronization would renderneuronal dynamics incapable of representing changes in sensory input or indeed cognitive stateFor example in the model it was observed that small perturbations to one column were ableto induce a global change in neuronal dynamics by disrupting a state of weak synchronization
Synchronization in a model of neuronal dynamics 729
Inevitably alternative dynamic configurations emerge epitomizing the importance of lsquolabilersquobrain dynamics (Friston 2000a)
Thirdly from a purely functionalist point of view the balance between functionalspecialization and functional integration may rely upon the sorts of mechanism modelledin this paper The brain appears to adhere to two principles of functional organization namelysegregation of function within cortical areas and sub-areas and the integration of these areasin perceptual synthesis cognition and sensorimotor integration (Tononi et al 1994 Fristonet al 1995) The balance between preserving specialized dynamics unique to each arearequired by specialization and the coupling among these areas mediating functional integrationmay be achieved by the neuromodulatory mechanisms described above Significantlythe spontaneous variation in the numbers of degrees of freedom (for example whenlow-dimensional synchronization manifolds are shadowed) imparts a dynamic characteristicto this perspective that is precluded by linear mechanisms alone
Finally from a mechanistic perspective complexity and diversity of brain dynamics arerequired for selection of adaptive responses (Friston 2000b) There are many examples ofselective mechanisms operating within the brain All can be seen as performing an ascenton some objective function or fitness landscape These mechanisms require complex anditinerant dynamics to facilitate a thorough exploration of the landscape These landscapesmay range from those involved in perception (where the landscape is defined by predictionerror see eg Rao and Ballard (1998)) to those involved in value learning (where thelandscape is determined by the outcome of a behaviour eg in temporal difference models (seeeg Sutton and Barto 1990)) In short complexity is necessary to maintain an information-rich environment in which the brain can respond to sensory inputs An essential componentof this is the maintenance of diversity which is a prerequisite for selectionist mechanismsat many scales (Edelman 1993) Understanding the mechanistic basis of complex dynamicsas measured electrophysiologically may well depend on the sort of biologically plausiblemodels used in this paper This understanding may be important for both basic and clinicalneuroscience (eg Tass 2002)
The present study supplements and advances on other research in this field The studyof complex phase synchronization and clustering in neural oscillator networks similar tothe present model has previously been the subject of formal mathematical study Forexample Terman and Wang (1995) studied synchronization of blocks of neural oscillatorsthat were subject to perturbation by connected lsquostimulirsquo Somers and Kopell (1995) studiedthe emergence of both synchronization and travelling waves in networks of coupled neuraloscillators In the work of Terman and Wang the local dynamics were based on neurallsquocaricaturersquo models such as the FitzHughndashNagumo equations and are hence more abstract thanthe current study The approach of Somers and Kopell only required that the local dynamicsexhibit fast and relaxation phases and is hence more abstract still In both studies onlyperiodic dynamics are studied Complex patterns of chaos synchronization in neural arrayshave been characterized by nonlinear methods (Kaneko 1998) and their putative contributionto both learning (Nakajima and Ueda 1996) and perception (Zhao and Macau 2001 Breakspear2001) explored However these studies employed models such as the logistic equation andtent map which are quite abstract in comparison to the present study On the other handHansel and Sompolinsky (1996) studied the role of chaos synchronization in a model ofvisual perception in a cortical hypercolumn using a very similar approach to that adoptedhere but with the additional complexity that synaptic delays were incorporated However inthis and earlier work (Hansel and Sompolinsky 1992) only linear measures of dynamics andsynchrony (specifically the auto- and cross-correlation functions) were employed The analysisof chaotic synchrony in the study of Larter et al (1999) was based only upon observation of
730 M Breakspear et al
the Euclidean difference between variables from two nodes within a larger array Hence thesignificance of the present work is that it provides a quantitative nonlinear analysis of complexchaos synchronization in a biophysical model of cortical dynamics Such research is requiredin order to bridge the gap between quantitative nonlinear research into synchronization andcomputation in abstract models of neural arrays and qualitative or linear analysis in morebiophysically plausible models
Acknowledgments
The authors would like to thank Peter Robinson for assistance with the manuscript and R Larterfor helpful advice MB is a recipient of a NSW Institute of Psychiatry Research Grant anda University of Sydney SESQUI postdoctoral fellowship KJF is supported by the WellcomeTrust
References
Abarbanel H Rulkov N and Sushchik M 1996 Generalised synchronization of chaos the auxiliary system approachPhys Rev E 53 4528ndash35
Afraimovich V Verichev N and Rabinovich M 1986 Stochastic synchronization of oscillation in dissipative systemsRadiophys Quantum Electron 29 795ndash801
Breakspear M 2001 Perception of odours by a nonlinear model of the olfactory bulb Int J Neural Syst 11 101ndash24Breakspear M 2002 Nonlinear phase desynchronisation in human electroencephalographic data Human Brain Mapping
15 175ndash98Breakspear M and Terry J R 2002a Nonlinear interdependence in neural systems motivation theory and relevance
Int J Neurosci 112 1163ndash84Breakspear M and Terry J R 2002b Detection and description of nonlinear interdependence in normal multichannel
human EEG Clin Neurophysiol 113 735ndash53Breakspear M and Terry J R 2002c Topographic organization of nonlinear interdependence in multichannel human
EEG Neuroimage 16 822ndash35Destexhe A Rudolph M Fellous J M and Sejnowski T J 2001 Fluctuating synaptic conductances recreate in vivo-like
activity in neocortical neurons Neuroscience 107 13ndash24Douglas R J Mahowald M A and Martin K A 1996 Microarchitecture of neocortical columns Brain TheorymdashBiological
Basis and Computational Principles ed A Aertsen and V Braitenberg (Amsterdam Elsevier)Eckhorn R Bauer R Jordan W Brosch M Kruse W Munk M and Reitboeck H J 1988 Coherent oscillations a
mechanism of feature linking in the visual cortex Multiple electrode and correlation analyses in the cat BiolCybern 60 121ndash30
Edelman G M 1993 Neural Darwinism selection and reentrant signalling in higher brain function Neuron 10 115ndash25Engel A K Kreiter A K Konig P and Singer W 1991 Synchronization of oscillatory neuronal responses between
striate and extrastriate visual cortical areas of the cat Proc Natl Acad Sci USA 88 6048ndash52Friston K J 2000a The labile brain I Neuronal transients and nonlinear coupling Phil Trans R Soc B 355 215ndash36Friston K J 2000b The labile brain II Transients complexity and selection Phil Trans R Soc B 355 237ndash52Friston K J Tononi G Sporns O and Edelman G 1995 Characterising the complexity of neuronal interactions Human
Brain Mapping 3 302ndash14Fujisaka H and Yamada T 1983 Stability theory of synchronized motion in coupled-oscillator system Prog Theor
Phys 69 32ndash47Gatlin L 1972 Information Theory and the Living System (New York Columbia University Press)Gray C M Konig P Engel A and Singer W 1989 Oscillatory responses in cat visual cortex exhibit inter-columnar
synchronization which reflects global stimulus properties Science 338 334ndash7Gray C M and Singer W 1989 Stimulus specific neuronal oscillations in orientation columns of cat visual cortex Proc
Natl Acad Sci USA 86 1698ndash702Haig A R Gordon E Wright J J Meares R A and Bahramali H 2000 Synchronous cortical gamma-band activity in
task-relevant cognition Neuroreport 11 669ndash75Hansel D and Sompolinsky H 1992 Synchronization and computation in a chaotic neural network Phys Rev Lett 68
718ndash21
Synchronization in a model of neuronal dynamics 731
Hansel D and Sompolinsky H 1996 Chaos and synchrony in a model of a hypercolumn in visual cortex J ComputNeurosci 3 7ndash4
Heagy J Carroll T and Pecora L 1998 Desynchronization by periodic orbits Phys Rev E 57 R1253ndash6Hunt B Ott E and Yorke J 1997 Differentiable synchronization of chaos Phys Rev E 55 4029ndash34Josic K 1998 Invariant manifolds and synchronization of coupled dynamical systems Phys Rev Lett 80 3053ndash6Kandel E Schwartz J and Jessell T 2000 Principles of Neural Science (New York McGraw-Hill)Kaneko K 1998 On the strength of attractors in a high-dimensional system Physica D 124 322ndash44Kocarev L Parlitz U and Brown R 2000 Robust synchronization of chaotic systems Phys Rev E 61 3716ndash20Larter R Speelman B and Worth R M 1999 A coupled ordinary differential equation lattice model for the simulation
of epileptic seizures Chaos 9 795ndash804Lumer E Edelman G and Tononi G 1997a Neural dynamics in a model of the thalamocortical system I Layers loops
and the emergence of fast synchronous oscillations Cereb Cortex 7 207ndash27Lumer E Edelman G and Tononi G 1997b Neural dynamics in a model of the thalamocortical system II The role of
neural synchrony tested through perturbations of spike timing Cereb Cortex 7 228ndash36Magleby K 1987 Short-term modulation at synapses Synaptic Function ed G M Edelman W E Gall and
W M Cowan (New York Wiley)Maistrenko Y Maistrenko V Popovich A and Mosekilde E 1998 Transverse instability and riddled basins in a system
of two coupled logistic maps Phys Rev E 57 2713ndash24Morris C and Lecar H 1981 Voltage oscillations in the barnacle giant muscle fiber Biophys J 35 193ndash213Mountcastle V B 1997 The columnar organization of the neocortex Brain 120 701ndash22Mountcastle V B 1998 The Cerebral Cortex (Cambridge MA Harvard University Press)Nakajima H and Ueda Y 1996 Riddled basins of the optimal states in learning dynamical systems Physica D 99 35ndash44Nunez P L 1995 Neocortical Dynamics and Human Brain Rhythms (Oxford Oxford University Press)Osipov G V Pikovsky A S Rosenblum M G and Kurths J 1997 Phase synchronization effects in a lattice of nonidentical
Rossler oscillators Phys Rev E 55 2353ndash61Ott E and Sommerer J 1994 Blowout bifurcations the occurrence of riddled basins and onndashoff intermittency Phys
Lett A 188 39ndash47Pecora L and Carroll T 1990 Synchronization in chaotic systems Phys Rev Lett 64 821ndash4Pikovsky A and Grassberger P 1991 Symmetry breaking bifurcation for coupled chaotic attractors J Phys A Math
Gen 24 4587ndash97Rao R P and Ballard D H 1998 Predictive coding in the visual cortex a functional interpretation of some extra-classical
receptive field effects Nature Neurosci 2 79ndash87Robinson P A Wright J J and Rennie C R 1998 Synchronous oscillations in the cerebral cortex Phys Rev E 57
4578ndash88Rodriguez E George N Lachauz J P Martinerie J Renault B and Varela F J 1999 Perceptionrsquos shadow long distance
synchronization of human brain activity Nature 397 430ndash3Rosenblum M Pikovsky A and Kurths J 1996 Phase synchronization of chaotic oscillators Phys Rev Lett 76 1804ndash7Rosenblum M Pikovsky A and Kurths J 1997 From phase to lag synchronization in coupled chaotic oscillators Phys
Rev Lett 78 4193ndash6Rulkov N and Sushchik M 1997 Robustness of synchronized chaotic oscillations Int J Bifurc Chaos 7 625ndash43Rulkov N Sushchik M Tsimring L and Abarbanel H 1995 Generalized synchronization of chaos in unidirectionally
coupled chaotic systems Phys Rev E 51 980ndash94Schiff S So P Chang T Burke R and Sauer T 1996 Detecting dynamical interdependence and generalized synchrony
through mutual prediction in a neural ensemble Phys Rev E 54 6708ndash24Silberstein R B 1995 Neuromodulations of neocortical dynamics Neocortical Dynamics and Human EEG Rhythms
ed P L Nunez (Oxford Oxford University Press)Singer W 1995 Putative functions of temporal correlations in neocortical processing Large-Scale Neuronal Theories
of the Brain ed C Koch and J Davis (London MIT Press)Somers D and Kopell N 1995 Waves and synchrony in networks of oscillators of relaxation and non-relaxation type
Physica D 89 169ndash83Sutton R S and Barto A G 1990 Time derivative models of Pavlovian reinforcement Learning and Computational
Neuroscience Foundations of Adaptive Networks ed M Gabriel and J Moore (Cambridge MA MIT Press) pp497ndash538
Szentagothai J 1983 The modular architectonic principle of neural centers Rev Physiol Biochem Pharmacol 9811ndash61
Tallon-Baudry C Betrand O and Fischer C 2001 Oscillatory synchrony between human extrastriate areas during visualshort-term memory maintenance J Neurosci 21 RC177(1ndash5)
Tass P A 2002 Desynchronization of brain rhythms with soft phase-resetting techniques Biol Cybern 87 102ndash15
732 M Breakspear et al
Tass P Rosenblum M Weule J Kurths J Pikovsky A Volkmann J Schnitzler A and Freund H 1998 Detection of nmphase locking from noisy data application to magnetoencephalography Phys Rev Lett 81 3291ndash4
Terman D and Wang D 1995 Global competition and local cooperation in a network of neural oscillators Physica D81 148ndash76
Terry J and Breakspear M 2003 An improved algorithm for the detection of dynamical interdependence in bivariatetime-series Biol Cybern 88 129ndash36
Tononi G Sporns O and Edelman G M 1994 A measure for brain complexity relating functional segregation andintegration in the nervous system Proc Natl Acad Sci USA 91 5033ndash7
von der Malsburg C 1985 Nervous structures with dynamical links Phys Chem 89 703ndash10Wang X 1991 Period-doublings to chaos in a simple neural network an analytic proof Complex Syst 5 425ndash41Wennekers T and Pasemann F 1996 Synchronous chaos in high dimensional modular neural networks Int J Bifurc
Chaos 6 2055ndash67Zaks M A Parks E H Rosenblum M G and Kurths J 1999 Alternative locking ratios in imperfect phase synchronization
Phys Rev Lett 82 4228ndash31Zhao L and Macau E N 2001 A network of dynamically coupled chaotic maps for scene segmentation IEEE Trans
Neural Netw 12 1375ndash85

706 M Breakspear et al
Figure 1 A schematic representation of connectivity within cortical columns Panel (a) depictsdense local connectivity between excitatory (EN) and inhibitory (IN) neurons Panel (b) depictsparameters representing strengths of synaptic connectivity between neurons within each columnplus input from ascending neurons within the reticular formation (RF)
embodying AMPA channels open an additional population of sodium channels thus increasingthe net conductance of sodium flow The second class representing NMDA receptors openan additional population of voltage-gated calcium channels Hence this acts to increase themaximum conductance of voltage-gated calcium channels Finally lsquononspecificrsquo subcorticalexcitation is added to both populations of neurons with amplitude I modulated by a randomnoise component of amplitude δ Thus we obtain
dV
dt= minus(gCa + rNMDAaee QV )mCa(V minus VCa)minus (gNamNa + aee QV )(V minus VNa)
minus gKW (V minus VK)minus gL(V minus VL) + aie Z QZ + ane Iδ (6)dZ
dt= b(ani Iδ + aeiV QV ) (7)
where rNMDA denotes the number of NMDA receptors relative to that of AMPA receptors ane
and ani parametrize non-specific input to excitatory and inhibitory cells and aee parametrizesthe local excitatory-to-excitatory synaptic strength Note the difference in the coefficientscontrolling the effect of local excitatory feedback onto NMDA receptors compared to thosefor AMPA receptors A schematic representation of the local network and the parametrizedsynaptic connectivity is presented in figure 1
22 Mesoscopic array dynamics
Each set of equations (2)ndash(7) governs the dynamics within a local cell assembly Couplingbetween N nodes can be introduced parsimoniously as competitive agonist excitatory actionbetween pyramidal cells in different columns However these long-range projections aremodelled to target the same populations of NMDA and AMPA receptors as are targeted by theshort-range excitatory projections Representing each node with a superscript this is achievedby modifying equation (6) to
dV i
dt= minus(gCa + (1 minus C)rNMDAaee Qi
V + CrNMDAaee〈QV 〉)mCa(Vi minus VCa)
minus gKW (V i minus VK)minus gL(Vi minus VL)
minus (gNamNa + (1 minus C)aee QiV + Caee〈QV 〉)(V i minus VNa) + aie Z Qi
Z + ane Iδ (8)
Synchronization in a model of neuronal dynamics 707
where i = 1 N and 〈 〉 represents spatial (lsquomean-fieldrsquo) averaging over cell assembliesC parametrizes the strength of excitatory coupling between cortical columns If C = 0the systems evolve independently C gt 0 introduces interdependences between consecutivecolumns C = 1 corresponds to maximum coupling with excitatory input from outside eachcolumn exceeding excitatory input from within each column
The long-range coupling is lsquoweakrsquo in the sense that we investigate parameter values forwhich C lt aee and C aie etc This either implies that the functional synaptic strength of thelong-range connections are weaker than local connections or that the strengths are similar butthe density of projections is comparatively sparse We prefer the latter interpretation whichis more consistent with other modelling approaches
23 Parameter values and physiological validity
In the sections that follow we set all physiologically measurable parameters (the maximumconductances threshold potentials and Nernst potentials) to within their accepted ranges (Larteret al 1999) Other parameters such as I the functional synaptic strengths and maximum firingrates were chosen to generate dynamically plausible behaviour including the limit cycle andaperiodic dynamics The parameter values are as given in table 1 (unless otherwise stated)There may be significant variability in many of these parameters In this respect it is crucial tonote that the behaviours described below occur in a large region of parameter space Hence theemergent behaviours discussed do not only occur for the parameter values used in this studyBecause as noted above the parameters are dimensionless and normalized to a capacitance ofone it is also important to note that the time units are arbitrary Noting however that the cellcapacitance is typically estimated as close to or equal to 1 microF cmminus2 (eg Destexhe et al 2001)it is reasonable to interpret the time evolution units as milliseconds (as done for the diffusivecoupling parameter employed in Larter et al (1999))
The principal aim of this paper is a quantitative study of non-linear synchronization ina biophysically realistic model of a neural system The model incorporates some aspects ofneural systems with significant detail such as the conductance of K+ glutamate-mediatedneurotransmission and the relatively sparse nature of long-range connectivity This detailallows manipulation of parameters that model pharmacological mechanisms of interest suchas glutamate receptor subclasses Other features such as local inhibitory feedback and thekinetics of Ca2+ and Na+ channels have been incorporated in a more simplified manner Thislimits the complexity of the model but ensures that numerical analysis can be supplementedby quantitative examination of for example the Lyapunov exponents Other biophysicalproperties could be made the subject of future study by changing relevant aspects of theequations
3 Behaviour of local lsquouncoupledrsquo columns
In this section a brief overview of the behaviours exhibited by autonomous local columns(ie C = 0) is presented As indicated in table 1 the dynamics of the model is explored byfixing all parameters except for δV the variance of the excitatory threshold which is varied inthe region [055 07] A similar range of behaviours is also observed if δV is fixed and anothercontrol parameter such as aee or I is varied
31 Fixed point attractors
For values of δV lt 055 the dynamics of a single column settles onto a solitary fixed pointattractor As shown in figure 2 transient dynamics persists for a number of seconds
708 M Breakspear et al
Figure 2 The time series for a fixed point attractor with VT = 054
Table 1 Parameter values unless otherwise stated Adapted from Larter et al (1999)
Parameter Description Value
TCa Threshold value for Ca channels minus001δCa Variance of Ca channel threshold 015gCa Conductance of population of Ca channels 11VCa Ca Nernst potential 1TK Threshold value for K channels 00δK Variance of K channel threshold 030gK Conductance of population of K channels 20VK K Nernst potential minus07TNa Threshold value for Na channels 03δNa Variance of Na channel threshold 015gNa Conductance of population of Na channels 67VNa Na Nernst potential 053VL Nernst potential leak channels minus05gL Conductance of population of leak channels 05VT Threshold potential for excitatory neurons 00δV Variance of excitatory threshold VariedZT Threshold potential for inhibitory neurons 00δZ Variance of inhibitory threshold 070I Subcortical input strength 030aee Excitatory-to-excitatory synaptic strength 04aei Excitatory-to-inhibitory synaptic strength 2aie Inhibitory-to-excitatory synaptic strength 2ane Non-specific-to-excitatory synaptic strength 1ani Non-specific-to-inhibitory synaptic strength 04b Time constant scaling factor 010ϕ Temperature scaling factor 07τ Time constant for K relaxation time 1rNMDA Ratio of NMDA to AMPA receptors 025β Random modulation of subcortical input 0
32 Limit cycle attractors
For 055 lt δV lt 059 the system undergoes a series of period-doubling bifurcations Theseresult in both simple and complex periodic behaviours A limit cycle occurring when δV = 056
Synchronization in a model of neuronal dynamics 709
Figure 3 The approach to a limit cycle attractor in phase space with VT = 056
is illustrated in figure 3 Other parameter variations produce more complicated higher-orderperiodic orbits For example a period-9 orbit occurring when δV = 06 aee = 05 aie = 05gNa = 0 and I = 0165 is presented in figure 4
33 Chaotic attractors
Setting δV gt 059 and keeping all other parameters as per table 1 produces complex aperiodicbehaviour in the uncoupled columns An example of this with δV = 065 is illustrated infigure 5 Panels (a) and (b) illustrate a phase space portrait and a time series plot of thissystem respectively They show an aperiodic but highly structured system In panel (c) afirst-return lsquoPoincarersquo section is presented This was obtained by taking a cross-section throughthe attractor in (a) by setting W = 032 and then plotting V (t) against V (t +l)with a time delayof l = 1 A topographic similarity to the simple logistic map is evident It has been provenanalytically that in restricted regions of parameter space dynamical systems incorporatingthe neural activation function (4) (5) are topologically conjugate to the logistic map (Wang1991) The Poincare map presented here shows that although a 11 map may not exist in moregeneral parameter regions the global morphology of the logistic map is still evident
To determine whether or not these aperiodic oscillations are in fact chaotic the Lyapunovexponents were estimated numerically using an algorithm based upon GrammndashSchmidtorthonormalization For the particular case illustrated the exponents were found to be
λ1 = 0015
λ2 = 00
λ3 = minus026
(9)
A principal Lyapunov exponent greater than zero is consistent with chaotic dynamics Thistogether with the topography of the first-return map and the existence of period-doublingbifurcations provides compelling evidence of chaotic behaviour in the dynamics of the singlecolumn The second exponent reflects the rate of exponential separation of adjacent orbits in
710 M Breakspear et al
Figure 4 A period-9 limit cycle attractor (a) A phase space portrait (b) A representative timeseries
the direction tangent to the flow hence a value of zero is to be expected The sum of the threeexponents is negative implying that the system is volume contracting which is to be expectedin the setting of a non-equilibrium (non-Hamiltonian) physiological system
It is interesting to note that this diversity of behaviours can be obtained throughmanipulation of a single state parameter From a functional perspective this permits eachcortical column to express a variety of qualitatively distinct dynamics through modificationof a single physiological parameter such as through modulation of the functional strength ofthe local synaptic connectivity Conversely it also implies that resetting of a single parametersuch as an abnormal δV resulting from loss of physiological heterogeneity could be associatedwith qualitative changes in local cortical behaviour
4 Cooperative behaviour of coupled columns
Whilst it is interesting to observe the range of dynamic behaviours possible in a single columnwe are primarily interested in the behaviour of many such interacting columns In this section
Synchronization in a model of neuronal dynamics 711
1
Figure 5 Chaotic behaviour of the autonomous system with VT = 065 The other parameters areas per table 1 (a) An attractor in phase space (b) A time series for V (t) (c) A Poincare sectionobtained by setting W = 032 and plotting V (t) against V (t + 1)
712 M Breakspear et al
we study the emergence of cooperative behaviour that results from the introduction of couplingbetween columns Three types of interdependent behaviour of two systems are describedidentical generalized and phase synchronization Synchronous behaviours in larger arrays arealso illustrated
41 Identical chaotic synchronization
411 Transition to identical chaotic synchronization The most straightforward extensionand the one that we consider first is that of two coupled cortical columns where all parametersare in the first case assumed identical7 Trivially the two systems will evolve identicallywhen started with identical initial conditions even if C = 0 This is simply a statement ofthe deterministic nature of the evolution equations In general the random initial conditionswill not be equal in which case it is hardly surprising that the different columns will evolveindependently An example is presented in figure 6 with VT = 06 in both systems C = 0and independent random initial conditions In figure 6(a) we show a segment of the timeseries for each of the two subsystems where it can be seen that the time series are chaoticand asynchronous In figure 6(b) the concurrent values of V 1 and V 2 are plotted against eachother showing that they bear no obvious interrelationship
With the introduction of coupling C gt 0 the systems begin to show epochs ofsynchronized behaviour At C sim= 01 there is a transition to stable fully synchronizedmotion as evident in an exemplar time series shown in figure 7(a) where C = 015 Infigure 7(b) we present the concurrent values of V 1 versus V 2 illustrating that in contrast tothe case in figure 6(b) the systems have become embedded in the low-dimensional lsquosymmetrymanifoldrsquo V 1 = V 2 Panel (c) shows a log-linear plot of the temporal evolution of the distanceof the coupled system from this manifold It can be seen that on average the two systemsapproach at an exponential rate with constant time-averaged exponent Whilst this phenomenon(lsquoidentical chaotic synchronizationrsquo) has been described in a variety of other systems since itsfirst description (Fujisaka and Yamada 1983 Pecora and Carroll 1990) to our knowledge thisis its first quantitative8 description in a detailed physiological model of a neural system
The symmetry of the dynamical system obtained by setting the state parameters in eachcolumn identical forces the invariance of the symmetry manifold V 1 = V 2 Moreover thedynamics within this manifold is governed by the same equations of motion as for a singlecortical column This manifold which contains the orbits of the synchronized system is calledthe synchronization manifold The coupling parameter C does not affect the dynamics withinthe synchronization manifold and as such is termed a normal parameter
412 lsquoBlowout bifurcationrsquo An important issue is the stability of the orbits on thesynchronization manifold This can be determined by analysing the Lyapunov spectraBecause the two-column system evolves in a six-dimensional phase space there are sixLyapunov exponents However for the synchronized system it is helpful to subdivide theseinto those that describe the dynamics within the synchronization manifold and those thatdescribe the dynamics in the transverse direction The latter describe the rate of convergenceor divergence of the coupled systems toward or away from this manifoldmdashand hence thetendency for the two systems to synchronize or separate For the time series in figure 7 this is
7 Whilst it is of course physiologically unrealistic to assume that two columns would have entirely identical structuresit is mathematically simpler to start analysing the system in this situation8 A qualitative description in a neural system with a different form of coupling (potassium diffusion) is evident infigure 8 of Larter et al (1999) Synchronous chaos in an abstract model of a neural system was also described inWennekers and Pasemann (1996)
Synchronization in a model of neuronal dynamics 713
1
2
2
1
Figure 6 Independent oscillations in a two-column system with identical parameters butindependent initial conditions (a) The time series (b) A plot of concurrent mean excitatorymembrane potentials V 1 against V 2
related to the slope of the plot in panel (c)9 For this strength of coupling the largest transverseexponent is negative (minus002) indicating that perturbations away from this state of identicalsynchronization decay and thus that the state of identical chaotic synchronization is stableagainst noise (ie dynamically stable)
To calculate the relationship between coupling strength and the Lyapunov exponents weused an algorithm which measures the growth of perturbations of the variational equations (thelsquoJacobianrsquo) of the system (we do not define these equations here as they are cumbersome anddo not enhance the understanding of the system) We calculated these exponents for increasingvalues of the coupling parameter C from 0 to 02 in increments of 0000 25 The results are
9 Only two of the six dependent variables are shown in figure 7 The actual transverse Lyapunov exponents reflectthe temporal evolution of all six and hence are somewhat more complex than that given by the slope of panel (c)
714 M Breakspear et al
1
2
Figure 7 Identical synchronization in the same system as in figure 7 but with C = 02 (a) Thetime series (b) A plot of concurrent mean excitatory membrane potentials V 1 against V 2 (c) Thelogarithm of the Euclidean distance between V 1 and V 2 versus time (linear)
Synchronization in a model of neuronal dynamics 715
Figure 8 Lyapunov spectra for a coupled system with δV = 07 and C varying from 0 to 02 insteps of 0000 33 At each value of C all Lyapunov exponents are calculated Least-squares linesof best fit were then fitted to the results for each exponent Solid lines indicate transverse exponents(λ) and dashed lines indicate the tangential exponents () Panel (a) shows all three transverseand three tangential exponents Panel (b) only shows the largest two of each
illustrated in figure 8 Panel (a) shows all exponents For illustrative purposes only the mostpositive four are plotted in panel (b) In both panels the tangential exponents () are fitted (inthe least-squares sense) with a dashed line and the transverse ones (λ) are fitted with a solidline They are labelled 1ndash3 in order of decreasing positivity The important features are asfollows For the range of coupling strengths that we consider the largest tangential Lyapunovexponent is positive which indicates the presence of chaotic behaviour within the manifoldWhen examining the line of best fit for this exponent we see that this exponent does not varywhen altering the coupling parameter This is true for all three tangential exponents and verifiesthat the changes in the coupling parameter do not alter the dynamics on the synchronizationmanifold (that is the coupling parameter is normal)
716 M Breakspear et al
The largest transverse Lyapunov exponent (denoted as λ1 in the figure) is a particularlyimportant exponent since it governs the stability of the attractor in the synchronization manifoldwith respect to perturbations out of the manifold If this exponent is negative then we expectsmall perturbations of trajectories to reconverge to the manifold in finite time If however thisexponent is positive then we would instead expect perturbed trajectories to diverge from themanifold This change in local behaviour is termed a bifurcation and the point at which itoccurs the bifurcation point It is apparent in figure 8 that the zero crossing of this exponentmdashthe bifurcation pointmdashoccurs at approximately C = 01 in the present system This typeof bifurcation has been termed a blowout bifurcation since once transverse stability is losttrajectories are lsquoblown outrsquo from the low-dimensional synchronization manifold into the fullphase space (Ott and Sommerer 1994) Hence in the present setting a blowout bifurcation isobserved if C falls below 01
We note two other observations Increasing rNMDA which corresponds to increasing thepopulation of NMDA receptors decreases the coupling strength required to achieve stablesynchronization On the other hand increasing the overall excitatory-to-excitatory synapticconnectivity (aee) has the opposite effect This is because the effect of local excitatory feedbackincreases the tangential Lyapunov exponents which has the effect of lsquosplittingrsquo the synchronizeddynamics
In the above simulations all the parameters of both systems were equal In realphysiological systems such symmetry between systems is clearly impossible We nowconsider the case in which the two columns have different structures A simple way toincorporate this whilst at the same time preserving (as much as possible) simple dynamics is tovary the parameter δV for each column This has the effect of destroying any symmetry-forcedinvariant manifolds and so the simple case of synchronization corresponding to identicallyevolving dynamics will not exist in this case However more exotic forms of synchronizationare still possible and we discuss these next
42 Generalized chaotic synchronization
In order to maintain consistency with the literature we study non-linear interdependence in thesetting where the systems are asymmetrical but the coupling is unidirectional This is easilyachieved in the present system by making C directed and setting C21 = 0 In this settingwe refer to the lsquofreersquo system as the driver and the lsquoslavedrsquo system as the response We firstask the question what is the effect of introducing a slight parameter mismatch into a systemexhibiting stable chaos synchronization An example of this is presented in figure 9 whereC12 = 04 and parameter mismatch has been incorporated by setting δV = 07 in the driversystem and δV = 0699 in the response system In panel (a) the time series appear qualitativelyvery similar However in panel (b) where the difference between the systems is plotted it isapparent that the separation between the two systems is of the same order of magnitude as thesize of each attractor In panel (c) the effect of the parameter mismatch on the synchronizationmanifold can be seen as a transformation away from the symmetry manifold
It has been proposed that in such a setting there exists a continuous functional relationshipbetween the dynamics of the driver and response systems
x2(t) = ψ(x1(t)) (10)
where xi = V i W i Z i This situation has been referred to in the literature as generalizedsynchronization (Afraimovich et al 1986 Rulkov et al 1995) The properties of ψ andthe topology and stability of the synchronization manifold in this context have not yetbeen fully elucidated although some significant results have been achieved For example
Synchronization in a model of neuronal dynamics 717
1
2
2
1
Figure 9 Generalized synchronization in an asymmetrical driverndashresponse system (a) The timeseries of the two systems (b) The distance between the two systems (c) A plot of concurrent meanexcitatory membrane potentials V 1 against V 2 The diagonal represents the symmetry manifold
718 M Breakspear et al
Hunt et al (1997) Josic (1998) Kocarev et al (2000) have shown that the generalizedsynchronization manifold is smooth (differentiable) if and only if it is normally hyperbolic(ie the largest transverse Lyapunov exponent is less than the smallest tangential Lyapunovexponent prior to the introduction of parameter mismatch) Such a condition is met in thepresent setting when C = 04 hence the smoothness of the manifold in panel (c)
However subsequent to the introduction of parameter mismatch we will not be able touse transverse and tangential Lyapunov exponents to detect such synchronization since thesynchronization manifold may possess a complex topology for which it is a non-trivial (atbest) or impossible (at worse) task to define transverse and tangential directions For examplethe manifold may be smooth as in figure 9(c) but have a fractal topology If the dynamics arenot normally hyperbolic then the manifold may be fractal but also non-differentiable (Huntet al 1997) and hence will not have a unique tangent direction at any point Consequently wewill not be able to compute tangential or transverse Lyapunov exponents and hence determinethe stability of the synchronization manifold However other techniques for detecting thepresence of generalized synchronization in complex systems have evolved including theauxiliary system approach (Abarbanel et al 1996) and non-linear interdependence (Schiffet al 1996) which are now explored
421 The auxiliary system approach The auxiliary system is a third subsystem which isa replica of the original response system but coupled independently to the driver system andcommencing from independent initial conditions It is possible to demonstrate that showingthe existence of generalized synchronizationmdashthat equation (10) is satisfiedmdashis equivalent toshowing that the response system and the auxiliary system evolve identically (even though theyare not directly coupled) To determine whether or not this is the case reduces to an analysisof the Lyapunov exponents transverse to the symmetry manifold with respect to the responsesystem and the auxiliary system
Whilst this is a somewhat artificial test for generalized synchronization since theproduction of an auxiliary system for a physical system would be impossible such a testis useful in numerical and theoretical studies In figure 10 we present an example of theauxiliary system approach where the parameter δV = 07 for column 1 and 068 for column 2We consider two values of the coupling C = 0025 and 01 The auxiliary system is simplya copy of the second column beginning from an arbitrary initial state In the first instancewhere C = 0025 we notice that even after allowing for an initial transient state the secondcolumn and the auxiliary do not behave in identical fashions (figure 10(a)) However whenC = 01 we observe (figure 10(b)) that the second column and the auxiliary do behave innear-identical manners after an initial transient and hence conclude that for the particularvalue of the coupling strength considered equation (10) is satisfied and hence generalizedsynchronization between the first and second columns occurs
422 Nonlinear interdependence The question remains as to how we may detect thepresence of generalized synchronization in a practical setting As we mentioned aboveit is not possible to use an auxiliary-system-based approach to detect the presence ofgeneralized synchronization Instead we must consider techniques for detecting suchbehaviour from experimental or numerical time series data Algorithms of this type areoften referred to as detecting non-linear interdependence Nonlinear interdependence is ageneral term encompassing systems where the behaviour of one non-linear system affectsthe behaviour of another More specifically it can be defined as occurring if the nullhypothesis that the interdependence between two signals is purely linear can be rejectedat conventional levels (usually 95) of statistical significance hence it is an experimental
Synchronization in a model of neuronal dynamics 719
2
1
2
1
Figure 10 Concurrent evolution of the response system V 2 and lsquoauxiliaryrsquo system V 1 (a) C =0025 (b) C = 01
term (eg Schiff et al 1996) Whilst this does not strictly correspond to the existence of afunctional relationship between the two systems such a relationship is typically used tomotivate the techniques for its detection Schiff et al (1996) extended the logic of non-linearprediction algorithms from autonomous to coupled systems by exploiting the continuity ofsuch a relationship and hence devised the first method of detecting non-linear interdependencein time series data The basis of the technique is using the forward evolution of a phase spacevectorrsquos neighbours to predict the forward evolution of that vector and the forward evolutionof the concurrent vector of the other (possibly interdependent) system If the error of such aprocedure is better on average than chance (the chance error is normalized to equal one) thennon-linear interdependence is said to be present
720 M Breakspear et al
V1
V2
Figure 11 Experimental and numerical examples of weak generalized non-linear synchronizationin the presence of measurement noise (a) Human scalp EEG data from posterior bipolar scalpelectrodes (O1P3ndashO2P4) (b) Weakly coupled aperiodic data in the present model with additivewhite noise (with 5 of the amplitude of the model signal) Parameters δ1
V = 0605 δ2V = 061
aee = 036 C = 007
A variation of this algorithm optimized to detect weak non-linear interdependence (Terryand Breakspear 2003) was used recently to demonstrate that weak non-linear interdependence isoccasionally present in the scalp EEG of healthy resting human subjects (Breakspear and Terry2002a 2002b 2002c) An example of an epoch of lsquonon-linearly independentrsquo EEG obtainedusing this technique is presented in figure 11(a) For comparison output from the currentmodel is presented in figure 11(b) The model parameters were set such that the dynamicswas still aperiodic but the intermittent large-amplitude bursts were suppressed and had weakmutual coupling (C = 007) Extraneous noise to simulate the effect of extracranial muscleactivity contaminating the EEG data has been added to this time series It should be notedthat the amplitudes and time scales of the modelrsquos variables are dimensionless Nonethelessthe model facilitates qualititative interpretation of experimental findings
Synchronization in a model of neuronal dynamics 721
43 Chaotic phase synchronization
Whereas the concept of generalized synchronization was mathematically motivated and assuch more suited to theoretical investigations of systems the concept of phase synchronizationderives from a more practical signal processing approach Phase synchronization occurs inboth symmetric and asymmetric settings but is often more useful in the asymmetric case (andin experimental data)
Unfortunately the phase of a chaotic signal is typically ambiguous and can be defined in anumber of different ways For example in the case of the coherent Rossler attractor (Osipovet al 1997) the phase may be defined as the arctangent of two of the variables of the systemMore generally it can sometimes be defined as the angle that an orbit subtends with the centreof an attractor It is also possible to extract the phase of a chaotic signal from time seriesdata by use of the Hilbert transform This has enabled the demonstration of chaotic phasesynchronization in a number of coupled asymmetrical systems (Rosenblum et al 1996 1997Zaks et al 1999) Although such an approach does not guarantee that the phase will be uniqueit does permit the practical analysis of phase in noisy time series (Tass et al 1998)
To illustrate phase synchronization in two coupled columns we again vary the parameterδV which if the columns are viewed as oscillators acts to detune them from a commonfrequency An important question is that of the criteria on the basis of which we concludewhether phase synchronization occurs We use a simple locking criterion whereby phasesynchronization is said to occur if
|θ1(t)minus θ2(t)| lt K (11)
where is θi the phase of column i and K is a constant Notice that this does not mean thatsynchronization implies equality of phases merely that their difference is bounded
An example of phase synchronization with δV = 065 for column 1 and 066 for column 2is given in figure 12 In this figure rNMDA = 03 Qualitatively some sort of phase coherenceis evident in the time series panel (a) This was confirmed by plotting the sine of thephase difference which has been extracted using the Hilbert transform panel (b) Clearly|sin(θ1 minus θ2)| lt K and hence |θ1 minus θ2| is bounded It can also be seen that the phase-synchronized dynamics occurs on a highly structured manifold depicted in figure 12(c)However unlike the case for the generalized synchronization manifold depicted in figure 9 itis evident that this manifold is not a simple transformation of the symmetry manifold
It is also possible to see lsquobimodalrsquo stability whereby a system jumps betweenapproximately zero phase difference and a phase lag phase synchronization of π An exampleis depicted in figure 12(d) At approximately 2000 s the system can be seen to jumpspontaneously from the first to the second of these modes The Z -variable is plotted forgraphical purposes
5 Intermittency and desynchronization
51 Desynchronization in an array of two systems
Diminishing the strength of coupling across the blowout bifurcation does not lead to a simpletransition from synchronous to asynchronous oscillations but is instead associated withcomplex temporal behaviours Specifically when the largest transverse Lyapunov exponent isclose to zero the coupled systems exhibit epochs of synchronous behaviours punctuated bysharp bursts of desynchronization This is observed in systems exhibiting identicalgeneralizedand phase synchronization An example of such a phenomenon is given in figure 13 The timeseries panel (a) shows synchronous dynamics except at time = 1000 where it can be seenthat the two time series desynchronize briefly This is evident in panel (b) as a disruption in the
722 M Breakspear et al
2
1
Figure 12 Phase-synchronized coupled columns (a) The time series of V 1 (solid) and V 2 (b) Thesine of the phase difference obtained through Hilbert transformation of the time series in panel (a)(c) The phase synchronization manifold in the V 1ndashV 2 plane (d) The time series of Z (solid) andZ showing a phase slip of π between two modes
Synchronization in a model of neuronal dynamics 723
Figure 13 An example of phase desynchronization in coupled asymmetric systems with C = 01Time series plots of (a) each V -variable and (b) their Euclidean distance (c) The temporal evolutionof the phase difference between the two systems (d) An example of the phase difference evolutionover a longer time interval
724 M Breakspear et al
orderly evolution of the distance between the two systems and in panel (c) where the evolutionof the phase difference reveals a brief violation of the criterion |φ1 minus φ2| lt constant Anexample of the phase difference evolution over a longer time interval is shown in figure 13(d)Note that this is a three-dimensional plot with sin(φ1 minus φ2) and cos(φ1 minus φ2) as they- and z-axes Hence phase desynchronizations are evident as complete revolutions aroundthe cylindrical phase space in which this variable evolves It can be seen that these phenomenaoccur at irregular intervals in the temporal domain
This behaviour can be explained by considering the nature of a chaotic attractor In thestatistical sense an arbitrary initial condition is expected to lead to the chaotic behaviour onthe attractor Consequently when we calculate the Lyapunov exponents of the system theseexponents correspond to those of the chaotic attractor Typically however there are an infinitenumber of unstable periodic orbits embedded within the attractor If the path of a trajectorybecomes close to one of these orbits it may lsquoshadowrsquo the orbit for a brief period of time Eachof these orbits has a corresponding set of Lyapunov exponents which (like the attractor) can bedivided into tangential and transverse exponents These also vary when we change the couplingparameter but are typically not the same as the exponents of the attractor Hence the largesttransverse exponent of the attractor may be negative yet there may be positive transverseexponents associated with periodic orbits Considering all of these exponents together wewould therefore only expect to observe complete stability of the synchronization manifold inthe case where all of the transverse Lyapunov exponents are negative Bursting behaviourwill be observed when the one of these exponents becomes positive typically correspondingto a periodic orbit of low order (Pikovsky and Grassberger 1991 Rulkov and Sushchik 1997Maistrenko et al 1998 Heagy et al 1998) Due to the complex structure of such periodic orbitsthe desynchronous bursts occur across a wide range of timescales
In figure 14 we present the results of a long integration (16times105 time units) in which thelengths of sequential synchronous epochs of phase synchronization are studied The parametervalues were δV = 065 for column 1 and 0645 for column 2 with coupling strength10 equal to0125 In panel (a) the power spectrum of these sequential lengths is plotted The whiteness ofthis spectrum indicates that the lengths of consecutive synchronous epochs are independentThis is due to the high-dimensional lsquomixingrsquo effect of desynchronization In panel (b) thelengths of the synchronous epochs have been sorted again into the order of increasing lengthThe results for two coupling strengths are plotted C = 0125 (solid) and 0135 (dashed)Several features of this plot are of note Firstly the lengths of the synchronous epochsare distributed over almost three orders of magnitude Secondly for both plots there is anapproximately log-linear relationship between the length of an epoch and its relative orderThis reflects an approximately exponential relationship between the length of a synchronousepoch and its likelihood of occurrence Thirdly this relationship is skewed toward longersynchronous epochs for the stronger coupling strengths
This type of signal with patches of lsquoorderrsquo occurring over many timescales interruptedby bursts of disorder can be shown to have a high information capacity (Gatlin 1972)Moreover complex phase desynchronization may make an infrequent yet robust contributionto the dynamical pattern of the alpha rhythm in scalp EEG data for resting human subjects(Breakspear 2002) It is tempting to postulate that such a lsquoweakly stablersquo dynamics may assistin the generation of the rich and fluid mental associations that spontaneously emerge in restingcognitive states Although this phenomenon has been previously described in a variety of othersystems to our knowledge this is the first description in a biophysical model of neocortex
10 The introduction of parameter mismatch causes an increase in the strength of coupling required for stablesynchronization above the value of 01 required in the symmetric case
Synchronization in a model of neuronal dynamics 725
Figure 14 Properties of sequential epochs of phase-synchronous behaviour over a long integrationtime δ1
V = 065 δ2V = 0645 aee = 04 (a) The flat (white) power spectrum (b) Synchronous
epochs sorted in order of increasing length (solid C = 0125 dashed C = 0134)
52 Synchronization and complex dynamics in larger arrays
The synchronous and complex phenomena described above are also observed in arrays ofcoupled systems with N gt 2 Expanding the size of the array permits complex dynamicsin the spatial in addition to the temporal domain However the basic principles governingthe type and stability of synchronized motions are the same In this section a few of thepossible behaviours are discussed Physiological variation across the array is modelled bymodifying the parameter δV = 07 in each array by adding a random number of maximummagnitude 001
In figure 15(a) an array of N = 20 uncoupled (C = 0) systems is shown The lack ofsynchronization in this time series is evident By comparison the rapid transition to completesynchronization is evident in figure 15(b) when C = 03 In fact 19 of the 20 subsystems
726 M Breakspear et al
(b)
(c)
(a)
Figure 15 Synchronization phenomena in an array of coupled cortical columns (a) N = 20C = 0 (b) N = 20 C = 03 (c) N = 6 C = 01 (d) N = 12 C = 009 (e) N = 12C = 012 A small perturbation has been applied to one of the columns at t = 2500 s (f) Evolutionof the phase difference between two of the columns in panel (e) For clarity the W -variable isplotted in panel (c) and the Z -variable in (e)
have phase synchronized within one single cycle Intermediate values of coupling strength areassociated with a rich variety of phenomena For example subsystems with the most similarparameter values typically synchronize into clusters of various sizes An example of this isillustrated in figure 15(c) Note that there is no synchronization between the two differentclusters each containing three synchronized subsystems
Another complex behaviour occurs when subsystems transiently synchronize into largeclusters then spontaneously desynchronize again Hence the dynamics of the entire arrayjumps between different transient dynamics characterized by varying degrees of freedom Anexample of this phenomenon is given in figure 15(d) In addition it is possible to inducedesynchronization across an entire array by applying a small perturbation to one columnfor one time step An example is illustrated in figure 15(e) A small perturbation has been
Synchronization in a model of neuronal dynamics 727
Figure 15 (Continued)
added to one column after 2500 time units It can be seen that the entire array subsequentlydesynchronizes This illustrates the sensitivity of the dynamics to external stimuli Finally it ispossible to lsquoinducersquo synchronization between selected subsystems by increasing the strength ofcoupling between them selectively This allows an extra level of complexity to be introducedinto the model For example it may allow targeted information transfer between a partialnetwork of the entire array
As discussed in section 51 a univariate signal which jumps between epochs of lowentropy (high repetition) and bursts of high entropy (high variability) can be shown to havehigh information capacity This is because the repetition of certain patterns (as in the English
728 M Breakspear et al
language) permits error detectability Bursts of high entropy act in the opposite directionto increase message variability With a multivariate signal in addition to optimizing theinformation capacity within each signal the overall information capacity can be increasedthrough the type of itinerant synchronization briefly illustrated in this section Synchronizationacross an entire array increases the message reliability (since a single lsquoerrant letterrsquo in onesignal can be easily detected) Conversely desynchronization into multiple clusters increasesmessage variability since each cluster will produce independent information Hence a neuralarray which switches dynamically between a range of cluster configurations can be seen asadaptive in the sense that high synchronous states can be expressed when error detectabilityis a priority (such as during quick and reliable execution of a familiar task) but the array canrapidly desynchronize if variability becomes salient (for example if the environment suddenlybecomes novel) A formal operationalized demonstration of this principle is to be the subjectof future work
6 Discussion
In this paper we present a dynamical model for activity in an array of weakly coupledlocal neural assemblies The model incorporates the fundamental principles governing cellmembrane potential as determined by voltage- and ligand-gated ion channels In this wayit allows a reasonable amount of physiological complexity to be incorporated includingsynaptic activation and receptor subtypeswithout recourse to overwhelming detail Numericalanalysis of this model reveals a transition from independent oscillations to stable chaoticsynchronization depending on the balance of local versus long-range connectivity andbiophysical heterogeneity These excitatory couplings are subject to ascending subcorticalneuromodulation arising principally from monoaminergic nuclei such as Locus Coeruleus(Silberstein 1995) Neuromodulation of extrinsic connections was incorporated into the modelvia a coupling parameter C The stability of the synchronization is determined by the valueof the largest transverse Lyapunov exponent which in turn depends on the coupling parameterC If the coupling is weakened with the result that this exponent becomes positive a blowoutbifurcation occurs and independent oscillations re-emerge In the vicinity of the transitionnon-linear desynchronizations with complex temporal and spatial properties are observed
It is pleasing that complex and itinerant behaviour reminiscent of real neuronaldynamics emerges through such a simple manipulation of a single dynamic control parameterFurthermore the ubiquitous projections of classical neuromodulatory systems in real brainssuggest a clear biological mechanism by which the brain could self-organize throughmoderating the complexity of its own dynamics There are many reasons that the complicateddynamics of the sort demonstrated in this paper may be neurobiologically important Wemention four
First the presence of multiple clusters of phase-synchronized subsystems as discussedabove may be a uniquely non-linear mechanism facilitating the lsquocarriagersquo and distinction ofmultiple concurrent dynamic cell assemblies (and hence cognitive processing) This may beimportant for perceptual grouping binding and transitions as suggested by others (eg von derMalsburg 1985 Gray and Singer 1989)
Second the facility to decouple and reconfigure dynamic cell assemblies througha neuromodulatory control parameter may be important for avoiding low-entropysynchronization manifolds In the absence of itinerancy synchronization would renderneuronal dynamics incapable of representing changes in sensory input or indeed cognitive stateFor example in the model it was observed that small perturbations to one column were ableto induce a global change in neuronal dynamics by disrupting a state of weak synchronization
Synchronization in a model of neuronal dynamics 729
Inevitably alternative dynamic configurations emerge epitomizing the importance of lsquolabilersquobrain dynamics (Friston 2000a)
Thirdly from a purely functionalist point of view the balance between functionalspecialization and functional integration may rely upon the sorts of mechanism modelledin this paper The brain appears to adhere to two principles of functional organization namelysegregation of function within cortical areas and sub-areas and the integration of these areasin perceptual synthesis cognition and sensorimotor integration (Tononi et al 1994 Fristonet al 1995) The balance between preserving specialized dynamics unique to each arearequired by specialization and the coupling among these areas mediating functional integrationmay be achieved by the neuromodulatory mechanisms described above Significantlythe spontaneous variation in the numbers of degrees of freedom (for example whenlow-dimensional synchronization manifolds are shadowed) imparts a dynamic characteristicto this perspective that is precluded by linear mechanisms alone
Finally from a mechanistic perspective complexity and diversity of brain dynamics arerequired for selection of adaptive responses (Friston 2000b) There are many examples ofselective mechanisms operating within the brain All can be seen as performing an ascenton some objective function or fitness landscape These mechanisms require complex anditinerant dynamics to facilitate a thorough exploration of the landscape These landscapesmay range from those involved in perception (where the landscape is defined by predictionerror see eg Rao and Ballard (1998)) to those involved in value learning (where thelandscape is determined by the outcome of a behaviour eg in temporal difference models (seeeg Sutton and Barto 1990)) In short complexity is necessary to maintain an information-rich environment in which the brain can respond to sensory inputs An essential componentof this is the maintenance of diversity which is a prerequisite for selectionist mechanismsat many scales (Edelman 1993) Understanding the mechanistic basis of complex dynamicsas measured electrophysiologically may well depend on the sort of biologically plausiblemodels used in this paper This understanding may be important for both basic and clinicalneuroscience (eg Tass 2002)
The present study supplements and advances on other research in this field The studyof complex phase synchronization and clustering in neural oscillator networks similar tothe present model has previously been the subject of formal mathematical study Forexample Terman and Wang (1995) studied synchronization of blocks of neural oscillatorsthat were subject to perturbation by connected lsquostimulirsquo Somers and Kopell (1995) studiedthe emergence of both synchronization and travelling waves in networks of coupled neuraloscillators In the work of Terman and Wang the local dynamics were based on neurallsquocaricaturersquo models such as the FitzHughndashNagumo equations and are hence more abstract thanthe current study The approach of Somers and Kopell only required that the local dynamicsexhibit fast and relaxation phases and is hence more abstract still In both studies onlyperiodic dynamics are studied Complex patterns of chaos synchronization in neural arrayshave been characterized by nonlinear methods (Kaneko 1998) and their putative contributionto both learning (Nakajima and Ueda 1996) and perception (Zhao and Macau 2001 Breakspear2001) explored However these studies employed models such as the logistic equation andtent map which are quite abstract in comparison to the present study On the other handHansel and Sompolinsky (1996) studied the role of chaos synchronization in a model ofvisual perception in a cortical hypercolumn using a very similar approach to that adoptedhere but with the additional complexity that synaptic delays were incorporated However inthis and earlier work (Hansel and Sompolinsky 1992) only linear measures of dynamics andsynchrony (specifically the auto- and cross-correlation functions) were employed The analysisof chaotic synchrony in the study of Larter et al (1999) was based only upon observation of
730 M Breakspear et al
the Euclidean difference between variables from two nodes within a larger array Hence thesignificance of the present work is that it provides a quantitative nonlinear analysis of complexchaos synchronization in a biophysical model of cortical dynamics Such research is requiredin order to bridge the gap between quantitative nonlinear research into synchronization andcomputation in abstract models of neural arrays and qualitative or linear analysis in morebiophysically plausible models
Acknowledgments
The authors would like to thank Peter Robinson for assistance with the manuscript and R Larterfor helpful advice MB is a recipient of a NSW Institute of Psychiatry Research Grant anda University of Sydney SESQUI postdoctoral fellowship KJF is supported by the WellcomeTrust
References
Abarbanel H Rulkov N and Sushchik M 1996 Generalised synchronization of chaos the auxiliary system approachPhys Rev E 53 4528ndash35
Afraimovich V Verichev N and Rabinovich M 1986 Stochastic synchronization of oscillation in dissipative systemsRadiophys Quantum Electron 29 795ndash801
Breakspear M 2001 Perception of odours by a nonlinear model of the olfactory bulb Int J Neural Syst 11 101ndash24Breakspear M 2002 Nonlinear phase desynchronisation in human electroencephalographic data Human Brain Mapping
15 175ndash98Breakspear M and Terry J R 2002a Nonlinear interdependence in neural systems motivation theory and relevance
Int J Neurosci 112 1163ndash84Breakspear M and Terry J R 2002b Detection and description of nonlinear interdependence in normal multichannel
human EEG Clin Neurophysiol 113 735ndash53Breakspear M and Terry J R 2002c Topographic organization of nonlinear interdependence in multichannel human
EEG Neuroimage 16 822ndash35Destexhe A Rudolph M Fellous J M and Sejnowski T J 2001 Fluctuating synaptic conductances recreate in vivo-like
activity in neocortical neurons Neuroscience 107 13ndash24Douglas R J Mahowald M A and Martin K A 1996 Microarchitecture of neocortical columns Brain TheorymdashBiological
Basis and Computational Principles ed A Aertsen and V Braitenberg (Amsterdam Elsevier)Eckhorn R Bauer R Jordan W Brosch M Kruse W Munk M and Reitboeck H J 1988 Coherent oscillations a
mechanism of feature linking in the visual cortex Multiple electrode and correlation analyses in the cat BiolCybern 60 121ndash30
Edelman G M 1993 Neural Darwinism selection and reentrant signalling in higher brain function Neuron 10 115ndash25Engel A K Kreiter A K Konig P and Singer W 1991 Synchronization of oscillatory neuronal responses between
striate and extrastriate visual cortical areas of the cat Proc Natl Acad Sci USA 88 6048ndash52Friston K J 2000a The labile brain I Neuronal transients and nonlinear coupling Phil Trans R Soc B 355 215ndash36Friston K J 2000b The labile brain II Transients complexity and selection Phil Trans R Soc B 355 237ndash52Friston K J Tononi G Sporns O and Edelman G 1995 Characterising the complexity of neuronal interactions Human
Brain Mapping 3 302ndash14Fujisaka H and Yamada T 1983 Stability theory of synchronized motion in coupled-oscillator system Prog Theor
Phys 69 32ndash47Gatlin L 1972 Information Theory and the Living System (New York Columbia University Press)Gray C M Konig P Engel A and Singer W 1989 Oscillatory responses in cat visual cortex exhibit inter-columnar
synchronization which reflects global stimulus properties Science 338 334ndash7Gray C M and Singer W 1989 Stimulus specific neuronal oscillations in orientation columns of cat visual cortex Proc
Natl Acad Sci USA 86 1698ndash702Haig A R Gordon E Wright J J Meares R A and Bahramali H 2000 Synchronous cortical gamma-band activity in
task-relevant cognition Neuroreport 11 669ndash75Hansel D and Sompolinsky H 1992 Synchronization and computation in a chaotic neural network Phys Rev Lett 68
718ndash21
Synchronization in a model of neuronal dynamics 731
Hansel D and Sompolinsky H 1996 Chaos and synchrony in a model of a hypercolumn in visual cortex J ComputNeurosci 3 7ndash4
Heagy J Carroll T and Pecora L 1998 Desynchronization by periodic orbits Phys Rev E 57 R1253ndash6Hunt B Ott E and Yorke J 1997 Differentiable synchronization of chaos Phys Rev E 55 4029ndash34Josic K 1998 Invariant manifolds and synchronization of coupled dynamical systems Phys Rev Lett 80 3053ndash6Kandel E Schwartz J and Jessell T 2000 Principles of Neural Science (New York McGraw-Hill)Kaneko K 1998 On the strength of attractors in a high-dimensional system Physica D 124 322ndash44Kocarev L Parlitz U and Brown R 2000 Robust synchronization of chaotic systems Phys Rev E 61 3716ndash20Larter R Speelman B and Worth R M 1999 A coupled ordinary differential equation lattice model for the simulation
of epileptic seizures Chaos 9 795ndash804Lumer E Edelman G and Tononi G 1997a Neural dynamics in a model of the thalamocortical system I Layers loops
and the emergence of fast synchronous oscillations Cereb Cortex 7 207ndash27Lumer E Edelman G and Tononi G 1997b Neural dynamics in a model of the thalamocortical system II The role of
neural synchrony tested through perturbations of spike timing Cereb Cortex 7 228ndash36Magleby K 1987 Short-term modulation at synapses Synaptic Function ed G M Edelman W E Gall and
W M Cowan (New York Wiley)Maistrenko Y Maistrenko V Popovich A and Mosekilde E 1998 Transverse instability and riddled basins in a system
of two coupled logistic maps Phys Rev E 57 2713ndash24Morris C and Lecar H 1981 Voltage oscillations in the barnacle giant muscle fiber Biophys J 35 193ndash213Mountcastle V B 1997 The columnar organization of the neocortex Brain 120 701ndash22Mountcastle V B 1998 The Cerebral Cortex (Cambridge MA Harvard University Press)Nakajima H and Ueda Y 1996 Riddled basins of the optimal states in learning dynamical systems Physica D 99 35ndash44Nunez P L 1995 Neocortical Dynamics and Human Brain Rhythms (Oxford Oxford University Press)Osipov G V Pikovsky A S Rosenblum M G and Kurths J 1997 Phase synchronization effects in a lattice of nonidentical
Rossler oscillators Phys Rev E 55 2353ndash61Ott E and Sommerer J 1994 Blowout bifurcations the occurrence of riddled basins and onndashoff intermittency Phys
Lett A 188 39ndash47Pecora L and Carroll T 1990 Synchronization in chaotic systems Phys Rev Lett 64 821ndash4Pikovsky A and Grassberger P 1991 Symmetry breaking bifurcation for coupled chaotic attractors J Phys A Math
Gen 24 4587ndash97Rao R P and Ballard D H 1998 Predictive coding in the visual cortex a functional interpretation of some extra-classical
receptive field effects Nature Neurosci 2 79ndash87Robinson P A Wright J J and Rennie C R 1998 Synchronous oscillations in the cerebral cortex Phys Rev E 57
4578ndash88Rodriguez E George N Lachauz J P Martinerie J Renault B and Varela F J 1999 Perceptionrsquos shadow long distance
synchronization of human brain activity Nature 397 430ndash3Rosenblum M Pikovsky A and Kurths J 1996 Phase synchronization of chaotic oscillators Phys Rev Lett 76 1804ndash7Rosenblum M Pikovsky A and Kurths J 1997 From phase to lag synchronization in coupled chaotic oscillators Phys
Rev Lett 78 4193ndash6Rulkov N and Sushchik M 1997 Robustness of synchronized chaotic oscillations Int J Bifurc Chaos 7 625ndash43Rulkov N Sushchik M Tsimring L and Abarbanel H 1995 Generalized synchronization of chaos in unidirectionally
coupled chaotic systems Phys Rev E 51 980ndash94Schiff S So P Chang T Burke R and Sauer T 1996 Detecting dynamical interdependence and generalized synchrony
through mutual prediction in a neural ensemble Phys Rev E 54 6708ndash24Silberstein R B 1995 Neuromodulations of neocortical dynamics Neocortical Dynamics and Human EEG Rhythms
ed P L Nunez (Oxford Oxford University Press)Singer W 1995 Putative functions of temporal correlations in neocortical processing Large-Scale Neuronal Theories
of the Brain ed C Koch and J Davis (London MIT Press)Somers D and Kopell N 1995 Waves and synchrony in networks of oscillators of relaxation and non-relaxation type
Physica D 89 169ndash83Sutton R S and Barto A G 1990 Time derivative models of Pavlovian reinforcement Learning and Computational
Neuroscience Foundations of Adaptive Networks ed M Gabriel and J Moore (Cambridge MA MIT Press) pp497ndash538
Szentagothai J 1983 The modular architectonic principle of neural centers Rev Physiol Biochem Pharmacol 9811ndash61
Tallon-Baudry C Betrand O and Fischer C 2001 Oscillatory synchrony between human extrastriate areas during visualshort-term memory maintenance J Neurosci 21 RC177(1ndash5)
Tass P A 2002 Desynchronization of brain rhythms with soft phase-resetting techniques Biol Cybern 87 102ndash15
732 M Breakspear et al
Tass P Rosenblum M Weule J Kurths J Pikovsky A Volkmann J Schnitzler A and Freund H 1998 Detection of nmphase locking from noisy data application to magnetoencephalography Phys Rev Lett 81 3291ndash4
Terman D and Wang D 1995 Global competition and local cooperation in a network of neural oscillators Physica D81 148ndash76
Terry J and Breakspear M 2003 An improved algorithm for the detection of dynamical interdependence in bivariatetime-series Biol Cybern 88 129ndash36
Tononi G Sporns O and Edelman G M 1994 A measure for brain complexity relating functional segregation andintegration in the nervous system Proc Natl Acad Sci USA 91 5033ndash7
von der Malsburg C 1985 Nervous structures with dynamical links Phys Chem 89 703ndash10Wang X 1991 Period-doublings to chaos in a simple neural network an analytic proof Complex Syst 5 425ndash41Wennekers T and Pasemann F 1996 Synchronous chaos in high dimensional modular neural networks Int J Bifurc
Chaos 6 2055ndash67Zaks M A Parks E H Rosenblum M G and Kurths J 1999 Alternative locking ratios in imperfect phase synchronization
Phys Rev Lett 82 4228ndash31Zhao L and Macau E N 2001 A network of dynamically coupled chaotic maps for scene segmentation IEEE Trans
Neural Netw 12 1375ndash85

Synchronization in a model of neuronal dynamics 707
where i = 1 N and 〈 〉 represents spatial (lsquomean-fieldrsquo) averaging over cell assembliesC parametrizes the strength of excitatory coupling between cortical columns If C = 0the systems evolve independently C gt 0 introduces interdependences between consecutivecolumns C = 1 corresponds to maximum coupling with excitatory input from outside eachcolumn exceeding excitatory input from within each column
The long-range coupling is lsquoweakrsquo in the sense that we investigate parameter values forwhich C lt aee and C aie etc This either implies that the functional synaptic strength of thelong-range connections are weaker than local connections or that the strengths are similar butthe density of projections is comparatively sparse We prefer the latter interpretation whichis more consistent with other modelling approaches
23 Parameter values and physiological validity
In the sections that follow we set all physiologically measurable parameters (the maximumconductances threshold potentials and Nernst potentials) to within their accepted ranges (Larteret al 1999) Other parameters such as I the functional synaptic strengths and maximum firingrates were chosen to generate dynamically plausible behaviour including the limit cycle andaperiodic dynamics The parameter values are as given in table 1 (unless otherwise stated)There may be significant variability in many of these parameters In this respect it is crucial tonote that the behaviours described below occur in a large region of parameter space Hence theemergent behaviours discussed do not only occur for the parameter values used in this studyBecause as noted above the parameters are dimensionless and normalized to a capacitance ofone it is also important to note that the time units are arbitrary Noting however that the cellcapacitance is typically estimated as close to or equal to 1 microF cmminus2 (eg Destexhe et al 2001)it is reasonable to interpret the time evolution units as milliseconds (as done for the diffusivecoupling parameter employed in Larter et al (1999))
The principal aim of this paper is a quantitative study of non-linear synchronization ina biophysically realistic model of a neural system The model incorporates some aspects ofneural systems with significant detail such as the conductance of K+ glutamate-mediatedneurotransmission and the relatively sparse nature of long-range connectivity This detailallows manipulation of parameters that model pharmacological mechanisms of interest suchas glutamate receptor subclasses Other features such as local inhibitory feedback and thekinetics of Ca2+ and Na+ channels have been incorporated in a more simplified manner Thislimits the complexity of the model but ensures that numerical analysis can be supplementedby quantitative examination of for example the Lyapunov exponents Other biophysicalproperties could be made the subject of future study by changing relevant aspects of theequations
3 Behaviour of local lsquouncoupledrsquo columns
In this section a brief overview of the behaviours exhibited by autonomous local columns(ie C = 0) is presented As indicated in table 1 the dynamics of the model is explored byfixing all parameters except for δV the variance of the excitatory threshold which is varied inthe region [055 07] A similar range of behaviours is also observed if δV is fixed and anothercontrol parameter such as aee or I is varied
31 Fixed point attractors
For values of δV lt 055 the dynamics of a single column settles onto a solitary fixed pointattractor As shown in figure 2 transient dynamics persists for a number of seconds
708 M Breakspear et al
Figure 2 The time series for a fixed point attractor with VT = 054
Table 1 Parameter values unless otherwise stated Adapted from Larter et al (1999)
Parameter Description Value
TCa Threshold value for Ca channels minus001δCa Variance of Ca channel threshold 015gCa Conductance of population of Ca channels 11VCa Ca Nernst potential 1TK Threshold value for K channels 00δK Variance of K channel threshold 030gK Conductance of population of K channels 20VK K Nernst potential minus07TNa Threshold value for Na channels 03δNa Variance of Na channel threshold 015gNa Conductance of population of Na channels 67VNa Na Nernst potential 053VL Nernst potential leak channels minus05gL Conductance of population of leak channels 05VT Threshold potential for excitatory neurons 00δV Variance of excitatory threshold VariedZT Threshold potential for inhibitory neurons 00δZ Variance of inhibitory threshold 070I Subcortical input strength 030aee Excitatory-to-excitatory synaptic strength 04aei Excitatory-to-inhibitory synaptic strength 2aie Inhibitory-to-excitatory synaptic strength 2ane Non-specific-to-excitatory synaptic strength 1ani Non-specific-to-inhibitory synaptic strength 04b Time constant scaling factor 010ϕ Temperature scaling factor 07τ Time constant for K relaxation time 1rNMDA Ratio of NMDA to AMPA receptors 025β Random modulation of subcortical input 0
32 Limit cycle attractors
For 055 lt δV lt 059 the system undergoes a series of period-doubling bifurcations Theseresult in both simple and complex periodic behaviours A limit cycle occurring when δV = 056
Synchronization in a model of neuronal dynamics 709
Figure 3 The approach to a limit cycle attractor in phase space with VT = 056
is illustrated in figure 3 Other parameter variations produce more complicated higher-orderperiodic orbits For example a period-9 orbit occurring when δV = 06 aee = 05 aie = 05gNa = 0 and I = 0165 is presented in figure 4
33 Chaotic attractors
Setting δV gt 059 and keeping all other parameters as per table 1 produces complex aperiodicbehaviour in the uncoupled columns An example of this with δV = 065 is illustrated infigure 5 Panels (a) and (b) illustrate a phase space portrait and a time series plot of thissystem respectively They show an aperiodic but highly structured system In panel (c) afirst-return lsquoPoincarersquo section is presented This was obtained by taking a cross-section throughthe attractor in (a) by setting W = 032 and then plotting V (t) against V (t +l)with a time delayof l = 1 A topographic similarity to the simple logistic map is evident It has been provenanalytically that in restricted regions of parameter space dynamical systems incorporatingthe neural activation function (4) (5) are topologically conjugate to the logistic map (Wang1991) The Poincare map presented here shows that although a 11 map may not exist in moregeneral parameter regions the global morphology of the logistic map is still evident
To determine whether or not these aperiodic oscillations are in fact chaotic the Lyapunovexponents were estimated numerically using an algorithm based upon GrammndashSchmidtorthonormalization For the particular case illustrated the exponents were found to be
λ1 = 0015
λ2 = 00
λ3 = minus026
(9)
A principal Lyapunov exponent greater than zero is consistent with chaotic dynamics Thistogether with the topography of the first-return map and the existence of period-doublingbifurcations provides compelling evidence of chaotic behaviour in the dynamics of the singlecolumn The second exponent reflects the rate of exponential separation of adjacent orbits in
710 M Breakspear et al
Figure 4 A period-9 limit cycle attractor (a) A phase space portrait (b) A representative timeseries
the direction tangent to the flow hence a value of zero is to be expected The sum of the threeexponents is negative implying that the system is volume contracting which is to be expectedin the setting of a non-equilibrium (non-Hamiltonian) physiological system
It is interesting to note that this diversity of behaviours can be obtained throughmanipulation of a single state parameter From a functional perspective this permits eachcortical column to express a variety of qualitatively distinct dynamics through modificationof a single physiological parameter such as through modulation of the functional strength ofthe local synaptic connectivity Conversely it also implies that resetting of a single parametersuch as an abnormal δV resulting from loss of physiological heterogeneity could be associatedwith qualitative changes in local cortical behaviour
4 Cooperative behaviour of coupled columns
Whilst it is interesting to observe the range of dynamic behaviours possible in a single columnwe are primarily interested in the behaviour of many such interacting columns In this section
Synchronization in a model of neuronal dynamics 711
1
Figure 5 Chaotic behaviour of the autonomous system with VT = 065 The other parameters areas per table 1 (a) An attractor in phase space (b) A time series for V (t) (c) A Poincare sectionobtained by setting W = 032 and plotting V (t) against V (t + 1)
712 M Breakspear et al
we study the emergence of cooperative behaviour that results from the introduction of couplingbetween columns Three types of interdependent behaviour of two systems are describedidentical generalized and phase synchronization Synchronous behaviours in larger arrays arealso illustrated
41 Identical chaotic synchronization
411 Transition to identical chaotic synchronization The most straightforward extensionand the one that we consider first is that of two coupled cortical columns where all parametersare in the first case assumed identical7 Trivially the two systems will evolve identicallywhen started with identical initial conditions even if C = 0 This is simply a statement ofthe deterministic nature of the evolution equations In general the random initial conditionswill not be equal in which case it is hardly surprising that the different columns will evolveindependently An example is presented in figure 6 with VT = 06 in both systems C = 0and independent random initial conditions In figure 6(a) we show a segment of the timeseries for each of the two subsystems where it can be seen that the time series are chaoticand asynchronous In figure 6(b) the concurrent values of V 1 and V 2 are plotted against eachother showing that they bear no obvious interrelationship
With the introduction of coupling C gt 0 the systems begin to show epochs ofsynchronized behaviour At C sim= 01 there is a transition to stable fully synchronizedmotion as evident in an exemplar time series shown in figure 7(a) where C = 015 Infigure 7(b) we present the concurrent values of V 1 versus V 2 illustrating that in contrast tothe case in figure 6(b) the systems have become embedded in the low-dimensional lsquosymmetrymanifoldrsquo V 1 = V 2 Panel (c) shows a log-linear plot of the temporal evolution of the distanceof the coupled system from this manifold It can be seen that on average the two systemsapproach at an exponential rate with constant time-averaged exponent Whilst this phenomenon(lsquoidentical chaotic synchronizationrsquo) has been described in a variety of other systems since itsfirst description (Fujisaka and Yamada 1983 Pecora and Carroll 1990) to our knowledge thisis its first quantitative8 description in a detailed physiological model of a neural system
The symmetry of the dynamical system obtained by setting the state parameters in eachcolumn identical forces the invariance of the symmetry manifold V 1 = V 2 Moreover thedynamics within this manifold is governed by the same equations of motion as for a singlecortical column This manifold which contains the orbits of the synchronized system is calledthe synchronization manifold The coupling parameter C does not affect the dynamics withinthe synchronization manifold and as such is termed a normal parameter
412 lsquoBlowout bifurcationrsquo An important issue is the stability of the orbits on thesynchronization manifold This can be determined by analysing the Lyapunov spectraBecause the two-column system evolves in a six-dimensional phase space there are sixLyapunov exponents However for the synchronized system it is helpful to subdivide theseinto those that describe the dynamics within the synchronization manifold and those thatdescribe the dynamics in the transverse direction The latter describe the rate of convergenceor divergence of the coupled systems toward or away from this manifoldmdashand hence thetendency for the two systems to synchronize or separate For the time series in figure 7 this is
7 Whilst it is of course physiologically unrealistic to assume that two columns would have entirely identical structuresit is mathematically simpler to start analysing the system in this situation8 A qualitative description in a neural system with a different form of coupling (potassium diffusion) is evident infigure 8 of Larter et al (1999) Synchronous chaos in an abstract model of a neural system was also described inWennekers and Pasemann (1996)
Synchronization in a model of neuronal dynamics 713
1
2
2
1
Figure 6 Independent oscillations in a two-column system with identical parameters butindependent initial conditions (a) The time series (b) A plot of concurrent mean excitatorymembrane potentials V 1 against V 2
related to the slope of the plot in panel (c)9 For this strength of coupling the largest transverseexponent is negative (minus002) indicating that perturbations away from this state of identicalsynchronization decay and thus that the state of identical chaotic synchronization is stableagainst noise (ie dynamically stable)
To calculate the relationship between coupling strength and the Lyapunov exponents weused an algorithm which measures the growth of perturbations of the variational equations (thelsquoJacobianrsquo) of the system (we do not define these equations here as they are cumbersome anddo not enhance the understanding of the system) We calculated these exponents for increasingvalues of the coupling parameter C from 0 to 02 in increments of 0000 25 The results are
9 Only two of the six dependent variables are shown in figure 7 The actual transverse Lyapunov exponents reflectthe temporal evolution of all six and hence are somewhat more complex than that given by the slope of panel (c)
714 M Breakspear et al
1
2
Figure 7 Identical synchronization in the same system as in figure 7 but with C = 02 (a) Thetime series (b) A plot of concurrent mean excitatory membrane potentials V 1 against V 2 (c) Thelogarithm of the Euclidean distance between V 1 and V 2 versus time (linear)
Synchronization in a model of neuronal dynamics 715
Figure 8 Lyapunov spectra for a coupled system with δV = 07 and C varying from 0 to 02 insteps of 0000 33 At each value of C all Lyapunov exponents are calculated Least-squares linesof best fit were then fitted to the results for each exponent Solid lines indicate transverse exponents(λ) and dashed lines indicate the tangential exponents () Panel (a) shows all three transverseand three tangential exponents Panel (b) only shows the largest two of each
illustrated in figure 8 Panel (a) shows all exponents For illustrative purposes only the mostpositive four are plotted in panel (b) In both panels the tangential exponents () are fitted (inthe least-squares sense) with a dashed line and the transverse ones (λ) are fitted with a solidline They are labelled 1ndash3 in order of decreasing positivity The important features are asfollows For the range of coupling strengths that we consider the largest tangential Lyapunovexponent is positive which indicates the presence of chaotic behaviour within the manifoldWhen examining the line of best fit for this exponent we see that this exponent does not varywhen altering the coupling parameter This is true for all three tangential exponents and verifiesthat the changes in the coupling parameter do not alter the dynamics on the synchronizationmanifold (that is the coupling parameter is normal)
716 M Breakspear et al
The largest transverse Lyapunov exponent (denoted as λ1 in the figure) is a particularlyimportant exponent since it governs the stability of the attractor in the synchronization manifoldwith respect to perturbations out of the manifold If this exponent is negative then we expectsmall perturbations of trajectories to reconverge to the manifold in finite time If however thisexponent is positive then we would instead expect perturbed trajectories to diverge from themanifold This change in local behaviour is termed a bifurcation and the point at which itoccurs the bifurcation point It is apparent in figure 8 that the zero crossing of this exponentmdashthe bifurcation pointmdashoccurs at approximately C = 01 in the present system This typeof bifurcation has been termed a blowout bifurcation since once transverse stability is losttrajectories are lsquoblown outrsquo from the low-dimensional synchronization manifold into the fullphase space (Ott and Sommerer 1994) Hence in the present setting a blowout bifurcation isobserved if C falls below 01
We note two other observations Increasing rNMDA which corresponds to increasing thepopulation of NMDA receptors decreases the coupling strength required to achieve stablesynchronization On the other hand increasing the overall excitatory-to-excitatory synapticconnectivity (aee) has the opposite effect This is because the effect of local excitatory feedbackincreases the tangential Lyapunov exponents which has the effect of lsquosplittingrsquo the synchronizeddynamics
In the above simulations all the parameters of both systems were equal In realphysiological systems such symmetry between systems is clearly impossible We nowconsider the case in which the two columns have different structures A simple way toincorporate this whilst at the same time preserving (as much as possible) simple dynamics is tovary the parameter δV for each column This has the effect of destroying any symmetry-forcedinvariant manifolds and so the simple case of synchronization corresponding to identicallyevolving dynamics will not exist in this case However more exotic forms of synchronizationare still possible and we discuss these next
42 Generalized chaotic synchronization
In order to maintain consistency with the literature we study non-linear interdependence in thesetting where the systems are asymmetrical but the coupling is unidirectional This is easilyachieved in the present system by making C directed and setting C21 = 0 In this settingwe refer to the lsquofreersquo system as the driver and the lsquoslavedrsquo system as the response We firstask the question what is the effect of introducing a slight parameter mismatch into a systemexhibiting stable chaos synchronization An example of this is presented in figure 9 whereC12 = 04 and parameter mismatch has been incorporated by setting δV = 07 in the driversystem and δV = 0699 in the response system In panel (a) the time series appear qualitativelyvery similar However in panel (b) where the difference between the systems is plotted it isapparent that the separation between the two systems is of the same order of magnitude as thesize of each attractor In panel (c) the effect of the parameter mismatch on the synchronizationmanifold can be seen as a transformation away from the symmetry manifold
It has been proposed that in such a setting there exists a continuous functional relationshipbetween the dynamics of the driver and response systems
x2(t) = ψ(x1(t)) (10)
where xi = V i W i Z i This situation has been referred to in the literature as generalizedsynchronization (Afraimovich et al 1986 Rulkov et al 1995) The properties of ψ andthe topology and stability of the synchronization manifold in this context have not yetbeen fully elucidated although some significant results have been achieved For example
Synchronization in a model of neuronal dynamics 717
1
2
2
1
Figure 9 Generalized synchronization in an asymmetrical driverndashresponse system (a) The timeseries of the two systems (b) The distance between the two systems (c) A plot of concurrent meanexcitatory membrane potentials V 1 against V 2 The diagonal represents the symmetry manifold
718 M Breakspear et al
Hunt et al (1997) Josic (1998) Kocarev et al (2000) have shown that the generalizedsynchronization manifold is smooth (differentiable) if and only if it is normally hyperbolic(ie the largest transverse Lyapunov exponent is less than the smallest tangential Lyapunovexponent prior to the introduction of parameter mismatch) Such a condition is met in thepresent setting when C = 04 hence the smoothness of the manifold in panel (c)
However subsequent to the introduction of parameter mismatch we will not be able touse transverse and tangential Lyapunov exponents to detect such synchronization since thesynchronization manifold may possess a complex topology for which it is a non-trivial (atbest) or impossible (at worse) task to define transverse and tangential directions For examplethe manifold may be smooth as in figure 9(c) but have a fractal topology If the dynamics arenot normally hyperbolic then the manifold may be fractal but also non-differentiable (Huntet al 1997) and hence will not have a unique tangent direction at any point Consequently wewill not be able to compute tangential or transverse Lyapunov exponents and hence determinethe stability of the synchronization manifold However other techniques for detecting thepresence of generalized synchronization in complex systems have evolved including theauxiliary system approach (Abarbanel et al 1996) and non-linear interdependence (Schiffet al 1996) which are now explored
421 The auxiliary system approach The auxiliary system is a third subsystem which isa replica of the original response system but coupled independently to the driver system andcommencing from independent initial conditions It is possible to demonstrate that showingthe existence of generalized synchronizationmdashthat equation (10) is satisfiedmdashis equivalent toshowing that the response system and the auxiliary system evolve identically (even though theyare not directly coupled) To determine whether or not this is the case reduces to an analysisof the Lyapunov exponents transverse to the symmetry manifold with respect to the responsesystem and the auxiliary system
Whilst this is a somewhat artificial test for generalized synchronization since theproduction of an auxiliary system for a physical system would be impossible such a testis useful in numerical and theoretical studies In figure 10 we present an example of theauxiliary system approach where the parameter δV = 07 for column 1 and 068 for column 2We consider two values of the coupling C = 0025 and 01 The auxiliary system is simplya copy of the second column beginning from an arbitrary initial state In the first instancewhere C = 0025 we notice that even after allowing for an initial transient state the secondcolumn and the auxiliary do not behave in identical fashions (figure 10(a)) However whenC = 01 we observe (figure 10(b)) that the second column and the auxiliary do behave innear-identical manners after an initial transient and hence conclude that for the particularvalue of the coupling strength considered equation (10) is satisfied and hence generalizedsynchronization between the first and second columns occurs
422 Nonlinear interdependence The question remains as to how we may detect thepresence of generalized synchronization in a practical setting As we mentioned aboveit is not possible to use an auxiliary-system-based approach to detect the presence ofgeneralized synchronization Instead we must consider techniques for detecting suchbehaviour from experimental or numerical time series data Algorithms of this type areoften referred to as detecting non-linear interdependence Nonlinear interdependence is ageneral term encompassing systems where the behaviour of one non-linear system affectsthe behaviour of another More specifically it can be defined as occurring if the nullhypothesis that the interdependence between two signals is purely linear can be rejectedat conventional levels (usually 95) of statistical significance hence it is an experimental
Synchronization in a model of neuronal dynamics 719
2
1
2
1
Figure 10 Concurrent evolution of the response system V 2 and lsquoauxiliaryrsquo system V 1 (a) C =0025 (b) C = 01
term (eg Schiff et al 1996) Whilst this does not strictly correspond to the existence of afunctional relationship between the two systems such a relationship is typically used tomotivate the techniques for its detection Schiff et al (1996) extended the logic of non-linearprediction algorithms from autonomous to coupled systems by exploiting the continuity ofsuch a relationship and hence devised the first method of detecting non-linear interdependencein time series data The basis of the technique is using the forward evolution of a phase spacevectorrsquos neighbours to predict the forward evolution of that vector and the forward evolutionof the concurrent vector of the other (possibly interdependent) system If the error of such aprocedure is better on average than chance (the chance error is normalized to equal one) thennon-linear interdependence is said to be present
720 M Breakspear et al
V1
V2
Figure 11 Experimental and numerical examples of weak generalized non-linear synchronizationin the presence of measurement noise (a) Human scalp EEG data from posterior bipolar scalpelectrodes (O1P3ndashO2P4) (b) Weakly coupled aperiodic data in the present model with additivewhite noise (with 5 of the amplitude of the model signal) Parameters δ1
V = 0605 δ2V = 061
aee = 036 C = 007
A variation of this algorithm optimized to detect weak non-linear interdependence (Terryand Breakspear 2003) was used recently to demonstrate that weak non-linear interdependence isoccasionally present in the scalp EEG of healthy resting human subjects (Breakspear and Terry2002a 2002b 2002c) An example of an epoch of lsquonon-linearly independentrsquo EEG obtainedusing this technique is presented in figure 11(a) For comparison output from the currentmodel is presented in figure 11(b) The model parameters were set such that the dynamicswas still aperiodic but the intermittent large-amplitude bursts were suppressed and had weakmutual coupling (C = 007) Extraneous noise to simulate the effect of extracranial muscleactivity contaminating the EEG data has been added to this time series It should be notedthat the amplitudes and time scales of the modelrsquos variables are dimensionless Nonethelessthe model facilitates qualititative interpretation of experimental findings
Synchronization in a model of neuronal dynamics 721
43 Chaotic phase synchronization
Whereas the concept of generalized synchronization was mathematically motivated and assuch more suited to theoretical investigations of systems the concept of phase synchronizationderives from a more practical signal processing approach Phase synchronization occurs inboth symmetric and asymmetric settings but is often more useful in the asymmetric case (andin experimental data)
Unfortunately the phase of a chaotic signal is typically ambiguous and can be defined in anumber of different ways For example in the case of the coherent Rossler attractor (Osipovet al 1997) the phase may be defined as the arctangent of two of the variables of the systemMore generally it can sometimes be defined as the angle that an orbit subtends with the centreof an attractor It is also possible to extract the phase of a chaotic signal from time seriesdata by use of the Hilbert transform This has enabled the demonstration of chaotic phasesynchronization in a number of coupled asymmetrical systems (Rosenblum et al 1996 1997Zaks et al 1999) Although such an approach does not guarantee that the phase will be uniqueit does permit the practical analysis of phase in noisy time series (Tass et al 1998)
To illustrate phase synchronization in two coupled columns we again vary the parameterδV which if the columns are viewed as oscillators acts to detune them from a commonfrequency An important question is that of the criteria on the basis of which we concludewhether phase synchronization occurs We use a simple locking criterion whereby phasesynchronization is said to occur if
|θ1(t)minus θ2(t)| lt K (11)
where is θi the phase of column i and K is a constant Notice that this does not mean thatsynchronization implies equality of phases merely that their difference is bounded
An example of phase synchronization with δV = 065 for column 1 and 066 for column 2is given in figure 12 In this figure rNMDA = 03 Qualitatively some sort of phase coherenceis evident in the time series panel (a) This was confirmed by plotting the sine of thephase difference which has been extracted using the Hilbert transform panel (b) Clearly|sin(θ1 minus θ2)| lt K and hence |θ1 minus θ2| is bounded It can also be seen that the phase-synchronized dynamics occurs on a highly structured manifold depicted in figure 12(c)However unlike the case for the generalized synchronization manifold depicted in figure 9 itis evident that this manifold is not a simple transformation of the symmetry manifold
It is also possible to see lsquobimodalrsquo stability whereby a system jumps betweenapproximately zero phase difference and a phase lag phase synchronization of π An exampleis depicted in figure 12(d) At approximately 2000 s the system can be seen to jumpspontaneously from the first to the second of these modes The Z -variable is plotted forgraphical purposes
5 Intermittency and desynchronization
51 Desynchronization in an array of two systems
Diminishing the strength of coupling across the blowout bifurcation does not lead to a simpletransition from synchronous to asynchronous oscillations but is instead associated withcomplex temporal behaviours Specifically when the largest transverse Lyapunov exponent isclose to zero the coupled systems exhibit epochs of synchronous behaviours punctuated bysharp bursts of desynchronization This is observed in systems exhibiting identicalgeneralizedand phase synchronization An example of such a phenomenon is given in figure 13 The timeseries panel (a) shows synchronous dynamics except at time = 1000 where it can be seenthat the two time series desynchronize briefly This is evident in panel (b) as a disruption in the
722 M Breakspear et al
2
1
Figure 12 Phase-synchronized coupled columns (a) The time series of V 1 (solid) and V 2 (b) Thesine of the phase difference obtained through Hilbert transformation of the time series in panel (a)(c) The phase synchronization manifold in the V 1ndashV 2 plane (d) The time series of Z (solid) andZ showing a phase slip of π between two modes
Synchronization in a model of neuronal dynamics 723
Figure 13 An example of phase desynchronization in coupled asymmetric systems with C = 01Time series plots of (a) each V -variable and (b) their Euclidean distance (c) The temporal evolutionof the phase difference between the two systems (d) An example of the phase difference evolutionover a longer time interval
724 M Breakspear et al
orderly evolution of the distance between the two systems and in panel (c) where the evolutionof the phase difference reveals a brief violation of the criterion |φ1 minus φ2| lt constant Anexample of the phase difference evolution over a longer time interval is shown in figure 13(d)Note that this is a three-dimensional plot with sin(φ1 minus φ2) and cos(φ1 minus φ2) as they- and z-axes Hence phase desynchronizations are evident as complete revolutions aroundthe cylindrical phase space in which this variable evolves It can be seen that these phenomenaoccur at irregular intervals in the temporal domain
This behaviour can be explained by considering the nature of a chaotic attractor In thestatistical sense an arbitrary initial condition is expected to lead to the chaotic behaviour onthe attractor Consequently when we calculate the Lyapunov exponents of the system theseexponents correspond to those of the chaotic attractor Typically however there are an infinitenumber of unstable periodic orbits embedded within the attractor If the path of a trajectorybecomes close to one of these orbits it may lsquoshadowrsquo the orbit for a brief period of time Eachof these orbits has a corresponding set of Lyapunov exponents which (like the attractor) can bedivided into tangential and transverse exponents These also vary when we change the couplingparameter but are typically not the same as the exponents of the attractor Hence the largesttransverse exponent of the attractor may be negative yet there may be positive transverseexponents associated with periodic orbits Considering all of these exponents together wewould therefore only expect to observe complete stability of the synchronization manifold inthe case where all of the transverse Lyapunov exponents are negative Bursting behaviourwill be observed when the one of these exponents becomes positive typically correspondingto a periodic orbit of low order (Pikovsky and Grassberger 1991 Rulkov and Sushchik 1997Maistrenko et al 1998 Heagy et al 1998) Due to the complex structure of such periodic orbitsthe desynchronous bursts occur across a wide range of timescales
In figure 14 we present the results of a long integration (16times105 time units) in which thelengths of sequential synchronous epochs of phase synchronization are studied The parametervalues were δV = 065 for column 1 and 0645 for column 2 with coupling strength10 equal to0125 In panel (a) the power spectrum of these sequential lengths is plotted The whiteness ofthis spectrum indicates that the lengths of consecutive synchronous epochs are independentThis is due to the high-dimensional lsquomixingrsquo effect of desynchronization In panel (b) thelengths of the synchronous epochs have been sorted again into the order of increasing lengthThe results for two coupling strengths are plotted C = 0125 (solid) and 0135 (dashed)Several features of this plot are of note Firstly the lengths of the synchronous epochsare distributed over almost three orders of magnitude Secondly for both plots there is anapproximately log-linear relationship between the length of an epoch and its relative orderThis reflects an approximately exponential relationship between the length of a synchronousepoch and its likelihood of occurrence Thirdly this relationship is skewed toward longersynchronous epochs for the stronger coupling strengths
This type of signal with patches of lsquoorderrsquo occurring over many timescales interruptedby bursts of disorder can be shown to have a high information capacity (Gatlin 1972)Moreover complex phase desynchronization may make an infrequent yet robust contributionto the dynamical pattern of the alpha rhythm in scalp EEG data for resting human subjects(Breakspear 2002) It is tempting to postulate that such a lsquoweakly stablersquo dynamics may assistin the generation of the rich and fluid mental associations that spontaneously emerge in restingcognitive states Although this phenomenon has been previously described in a variety of othersystems to our knowledge this is the first description in a biophysical model of neocortex
10 The introduction of parameter mismatch causes an increase in the strength of coupling required for stablesynchronization above the value of 01 required in the symmetric case
Synchronization in a model of neuronal dynamics 725
Figure 14 Properties of sequential epochs of phase-synchronous behaviour over a long integrationtime δ1
V = 065 δ2V = 0645 aee = 04 (a) The flat (white) power spectrum (b) Synchronous
epochs sorted in order of increasing length (solid C = 0125 dashed C = 0134)
52 Synchronization and complex dynamics in larger arrays
The synchronous and complex phenomena described above are also observed in arrays ofcoupled systems with N gt 2 Expanding the size of the array permits complex dynamicsin the spatial in addition to the temporal domain However the basic principles governingthe type and stability of synchronized motions are the same In this section a few of thepossible behaviours are discussed Physiological variation across the array is modelled bymodifying the parameter δV = 07 in each array by adding a random number of maximummagnitude 001
In figure 15(a) an array of N = 20 uncoupled (C = 0) systems is shown The lack ofsynchronization in this time series is evident By comparison the rapid transition to completesynchronization is evident in figure 15(b) when C = 03 In fact 19 of the 20 subsystems
726 M Breakspear et al
(b)
(c)
(a)
Figure 15 Synchronization phenomena in an array of coupled cortical columns (a) N = 20C = 0 (b) N = 20 C = 03 (c) N = 6 C = 01 (d) N = 12 C = 009 (e) N = 12C = 012 A small perturbation has been applied to one of the columns at t = 2500 s (f) Evolutionof the phase difference between two of the columns in panel (e) For clarity the W -variable isplotted in panel (c) and the Z -variable in (e)
have phase synchronized within one single cycle Intermediate values of coupling strength areassociated with a rich variety of phenomena For example subsystems with the most similarparameter values typically synchronize into clusters of various sizes An example of this isillustrated in figure 15(c) Note that there is no synchronization between the two differentclusters each containing three synchronized subsystems
Another complex behaviour occurs when subsystems transiently synchronize into largeclusters then spontaneously desynchronize again Hence the dynamics of the entire arrayjumps between different transient dynamics characterized by varying degrees of freedom Anexample of this phenomenon is given in figure 15(d) In addition it is possible to inducedesynchronization across an entire array by applying a small perturbation to one columnfor one time step An example is illustrated in figure 15(e) A small perturbation has been
Synchronization in a model of neuronal dynamics 727
Figure 15 (Continued)
added to one column after 2500 time units It can be seen that the entire array subsequentlydesynchronizes This illustrates the sensitivity of the dynamics to external stimuli Finally it ispossible to lsquoinducersquo synchronization between selected subsystems by increasing the strength ofcoupling between them selectively This allows an extra level of complexity to be introducedinto the model For example it may allow targeted information transfer between a partialnetwork of the entire array
As discussed in section 51 a univariate signal which jumps between epochs of lowentropy (high repetition) and bursts of high entropy (high variability) can be shown to havehigh information capacity This is because the repetition of certain patterns (as in the English
728 M Breakspear et al
language) permits error detectability Bursts of high entropy act in the opposite directionto increase message variability With a multivariate signal in addition to optimizing theinformation capacity within each signal the overall information capacity can be increasedthrough the type of itinerant synchronization briefly illustrated in this section Synchronizationacross an entire array increases the message reliability (since a single lsquoerrant letterrsquo in onesignal can be easily detected) Conversely desynchronization into multiple clusters increasesmessage variability since each cluster will produce independent information Hence a neuralarray which switches dynamically between a range of cluster configurations can be seen asadaptive in the sense that high synchronous states can be expressed when error detectabilityis a priority (such as during quick and reliable execution of a familiar task) but the array canrapidly desynchronize if variability becomes salient (for example if the environment suddenlybecomes novel) A formal operationalized demonstration of this principle is to be the subjectof future work
6 Discussion
In this paper we present a dynamical model for activity in an array of weakly coupledlocal neural assemblies The model incorporates the fundamental principles governing cellmembrane potential as determined by voltage- and ligand-gated ion channels In this wayit allows a reasonable amount of physiological complexity to be incorporated includingsynaptic activation and receptor subtypeswithout recourse to overwhelming detail Numericalanalysis of this model reveals a transition from independent oscillations to stable chaoticsynchronization depending on the balance of local versus long-range connectivity andbiophysical heterogeneity These excitatory couplings are subject to ascending subcorticalneuromodulation arising principally from monoaminergic nuclei such as Locus Coeruleus(Silberstein 1995) Neuromodulation of extrinsic connections was incorporated into the modelvia a coupling parameter C The stability of the synchronization is determined by the valueof the largest transverse Lyapunov exponent which in turn depends on the coupling parameterC If the coupling is weakened with the result that this exponent becomes positive a blowoutbifurcation occurs and independent oscillations re-emerge In the vicinity of the transitionnon-linear desynchronizations with complex temporal and spatial properties are observed
It is pleasing that complex and itinerant behaviour reminiscent of real neuronaldynamics emerges through such a simple manipulation of a single dynamic control parameterFurthermore the ubiquitous projections of classical neuromodulatory systems in real brainssuggest a clear biological mechanism by which the brain could self-organize throughmoderating the complexity of its own dynamics There are many reasons that the complicateddynamics of the sort demonstrated in this paper may be neurobiologically important Wemention four
First the presence of multiple clusters of phase-synchronized subsystems as discussedabove may be a uniquely non-linear mechanism facilitating the lsquocarriagersquo and distinction ofmultiple concurrent dynamic cell assemblies (and hence cognitive processing) This may beimportant for perceptual grouping binding and transitions as suggested by others (eg von derMalsburg 1985 Gray and Singer 1989)
Second the facility to decouple and reconfigure dynamic cell assemblies througha neuromodulatory control parameter may be important for avoiding low-entropysynchronization manifolds In the absence of itinerancy synchronization would renderneuronal dynamics incapable of representing changes in sensory input or indeed cognitive stateFor example in the model it was observed that small perturbations to one column were ableto induce a global change in neuronal dynamics by disrupting a state of weak synchronization
Synchronization in a model of neuronal dynamics 729
Inevitably alternative dynamic configurations emerge epitomizing the importance of lsquolabilersquobrain dynamics (Friston 2000a)
Thirdly from a purely functionalist point of view the balance between functionalspecialization and functional integration may rely upon the sorts of mechanism modelledin this paper The brain appears to adhere to two principles of functional organization namelysegregation of function within cortical areas and sub-areas and the integration of these areasin perceptual synthesis cognition and sensorimotor integration (Tononi et al 1994 Fristonet al 1995) The balance between preserving specialized dynamics unique to each arearequired by specialization and the coupling among these areas mediating functional integrationmay be achieved by the neuromodulatory mechanisms described above Significantlythe spontaneous variation in the numbers of degrees of freedom (for example whenlow-dimensional synchronization manifolds are shadowed) imparts a dynamic characteristicto this perspective that is precluded by linear mechanisms alone
Finally from a mechanistic perspective complexity and diversity of brain dynamics arerequired for selection of adaptive responses (Friston 2000b) There are many examples ofselective mechanisms operating within the brain All can be seen as performing an ascenton some objective function or fitness landscape These mechanisms require complex anditinerant dynamics to facilitate a thorough exploration of the landscape These landscapesmay range from those involved in perception (where the landscape is defined by predictionerror see eg Rao and Ballard (1998)) to those involved in value learning (where thelandscape is determined by the outcome of a behaviour eg in temporal difference models (seeeg Sutton and Barto 1990)) In short complexity is necessary to maintain an information-rich environment in which the brain can respond to sensory inputs An essential componentof this is the maintenance of diversity which is a prerequisite for selectionist mechanismsat many scales (Edelman 1993) Understanding the mechanistic basis of complex dynamicsas measured electrophysiologically may well depend on the sort of biologically plausiblemodels used in this paper This understanding may be important for both basic and clinicalneuroscience (eg Tass 2002)
The present study supplements and advances on other research in this field The studyof complex phase synchronization and clustering in neural oscillator networks similar tothe present model has previously been the subject of formal mathematical study Forexample Terman and Wang (1995) studied synchronization of blocks of neural oscillatorsthat were subject to perturbation by connected lsquostimulirsquo Somers and Kopell (1995) studiedthe emergence of both synchronization and travelling waves in networks of coupled neuraloscillators In the work of Terman and Wang the local dynamics were based on neurallsquocaricaturersquo models such as the FitzHughndashNagumo equations and are hence more abstract thanthe current study The approach of Somers and Kopell only required that the local dynamicsexhibit fast and relaxation phases and is hence more abstract still In both studies onlyperiodic dynamics are studied Complex patterns of chaos synchronization in neural arrayshave been characterized by nonlinear methods (Kaneko 1998) and their putative contributionto both learning (Nakajima and Ueda 1996) and perception (Zhao and Macau 2001 Breakspear2001) explored However these studies employed models such as the logistic equation andtent map which are quite abstract in comparison to the present study On the other handHansel and Sompolinsky (1996) studied the role of chaos synchronization in a model ofvisual perception in a cortical hypercolumn using a very similar approach to that adoptedhere but with the additional complexity that synaptic delays were incorporated However inthis and earlier work (Hansel and Sompolinsky 1992) only linear measures of dynamics andsynchrony (specifically the auto- and cross-correlation functions) were employed The analysisof chaotic synchrony in the study of Larter et al (1999) was based only upon observation of
730 M Breakspear et al
the Euclidean difference between variables from two nodes within a larger array Hence thesignificance of the present work is that it provides a quantitative nonlinear analysis of complexchaos synchronization in a biophysical model of cortical dynamics Such research is requiredin order to bridge the gap between quantitative nonlinear research into synchronization andcomputation in abstract models of neural arrays and qualitative or linear analysis in morebiophysically plausible models
Acknowledgments
The authors would like to thank Peter Robinson for assistance with the manuscript and R Larterfor helpful advice MB is a recipient of a NSW Institute of Psychiatry Research Grant anda University of Sydney SESQUI postdoctoral fellowship KJF is supported by the WellcomeTrust
References
Abarbanel H Rulkov N and Sushchik M 1996 Generalised synchronization of chaos the auxiliary system approachPhys Rev E 53 4528ndash35
Afraimovich V Verichev N and Rabinovich M 1986 Stochastic synchronization of oscillation in dissipative systemsRadiophys Quantum Electron 29 795ndash801
Breakspear M 2001 Perception of odours by a nonlinear model of the olfactory bulb Int J Neural Syst 11 101ndash24Breakspear M 2002 Nonlinear phase desynchronisation in human electroencephalographic data Human Brain Mapping
15 175ndash98Breakspear M and Terry J R 2002a Nonlinear interdependence in neural systems motivation theory and relevance
Int J Neurosci 112 1163ndash84Breakspear M and Terry J R 2002b Detection and description of nonlinear interdependence in normal multichannel
human EEG Clin Neurophysiol 113 735ndash53Breakspear M and Terry J R 2002c Topographic organization of nonlinear interdependence in multichannel human
EEG Neuroimage 16 822ndash35Destexhe A Rudolph M Fellous J M and Sejnowski T J 2001 Fluctuating synaptic conductances recreate in vivo-like
activity in neocortical neurons Neuroscience 107 13ndash24Douglas R J Mahowald M A and Martin K A 1996 Microarchitecture of neocortical columns Brain TheorymdashBiological
Basis and Computational Principles ed A Aertsen and V Braitenberg (Amsterdam Elsevier)Eckhorn R Bauer R Jordan W Brosch M Kruse W Munk M and Reitboeck H J 1988 Coherent oscillations a
mechanism of feature linking in the visual cortex Multiple electrode and correlation analyses in the cat BiolCybern 60 121ndash30
Edelman G M 1993 Neural Darwinism selection and reentrant signalling in higher brain function Neuron 10 115ndash25Engel A K Kreiter A K Konig P and Singer W 1991 Synchronization of oscillatory neuronal responses between
striate and extrastriate visual cortical areas of the cat Proc Natl Acad Sci USA 88 6048ndash52Friston K J 2000a The labile brain I Neuronal transients and nonlinear coupling Phil Trans R Soc B 355 215ndash36Friston K J 2000b The labile brain II Transients complexity and selection Phil Trans R Soc B 355 237ndash52Friston K J Tononi G Sporns O and Edelman G 1995 Characterising the complexity of neuronal interactions Human
Brain Mapping 3 302ndash14Fujisaka H and Yamada T 1983 Stability theory of synchronized motion in coupled-oscillator system Prog Theor
Phys 69 32ndash47Gatlin L 1972 Information Theory and the Living System (New York Columbia University Press)Gray C M Konig P Engel A and Singer W 1989 Oscillatory responses in cat visual cortex exhibit inter-columnar
synchronization which reflects global stimulus properties Science 338 334ndash7Gray C M and Singer W 1989 Stimulus specific neuronal oscillations in orientation columns of cat visual cortex Proc
Natl Acad Sci USA 86 1698ndash702Haig A R Gordon E Wright J J Meares R A and Bahramali H 2000 Synchronous cortical gamma-band activity in
task-relevant cognition Neuroreport 11 669ndash75Hansel D and Sompolinsky H 1992 Synchronization and computation in a chaotic neural network Phys Rev Lett 68
718ndash21
Synchronization in a model of neuronal dynamics 731
Hansel D and Sompolinsky H 1996 Chaos and synchrony in a model of a hypercolumn in visual cortex J ComputNeurosci 3 7ndash4
Heagy J Carroll T and Pecora L 1998 Desynchronization by periodic orbits Phys Rev E 57 R1253ndash6Hunt B Ott E and Yorke J 1997 Differentiable synchronization of chaos Phys Rev E 55 4029ndash34Josic K 1998 Invariant manifolds and synchronization of coupled dynamical systems Phys Rev Lett 80 3053ndash6Kandel E Schwartz J and Jessell T 2000 Principles of Neural Science (New York McGraw-Hill)Kaneko K 1998 On the strength of attractors in a high-dimensional system Physica D 124 322ndash44Kocarev L Parlitz U and Brown R 2000 Robust synchronization of chaotic systems Phys Rev E 61 3716ndash20Larter R Speelman B and Worth R M 1999 A coupled ordinary differential equation lattice model for the simulation
of epileptic seizures Chaos 9 795ndash804Lumer E Edelman G and Tononi G 1997a Neural dynamics in a model of the thalamocortical system I Layers loops
and the emergence of fast synchronous oscillations Cereb Cortex 7 207ndash27Lumer E Edelman G and Tononi G 1997b Neural dynamics in a model of the thalamocortical system II The role of
neural synchrony tested through perturbations of spike timing Cereb Cortex 7 228ndash36Magleby K 1987 Short-term modulation at synapses Synaptic Function ed G M Edelman W E Gall and
W M Cowan (New York Wiley)Maistrenko Y Maistrenko V Popovich A and Mosekilde E 1998 Transverse instability and riddled basins in a system
of two coupled logistic maps Phys Rev E 57 2713ndash24Morris C and Lecar H 1981 Voltage oscillations in the barnacle giant muscle fiber Biophys J 35 193ndash213Mountcastle V B 1997 The columnar organization of the neocortex Brain 120 701ndash22Mountcastle V B 1998 The Cerebral Cortex (Cambridge MA Harvard University Press)Nakajima H and Ueda Y 1996 Riddled basins of the optimal states in learning dynamical systems Physica D 99 35ndash44Nunez P L 1995 Neocortical Dynamics and Human Brain Rhythms (Oxford Oxford University Press)Osipov G V Pikovsky A S Rosenblum M G and Kurths J 1997 Phase synchronization effects in a lattice of nonidentical
Rossler oscillators Phys Rev E 55 2353ndash61Ott E and Sommerer J 1994 Blowout bifurcations the occurrence of riddled basins and onndashoff intermittency Phys
Lett A 188 39ndash47Pecora L and Carroll T 1990 Synchronization in chaotic systems Phys Rev Lett 64 821ndash4Pikovsky A and Grassberger P 1991 Symmetry breaking bifurcation for coupled chaotic attractors J Phys A Math
Gen 24 4587ndash97Rao R P and Ballard D H 1998 Predictive coding in the visual cortex a functional interpretation of some extra-classical
receptive field effects Nature Neurosci 2 79ndash87Robinson P A Wright J J and Rennie C R 1998 Synchronous oscillations in the cerebral cortex Phys Rev E 57
4578ndash88Rodriguez E George N Lachauz J P Martinerie J Renault B and Varela F J 1999 Perceptionrsquos shadow long distance
synchronization of human brain activity Nature 397 430ndash3Rosenblum M Pikovsky A and Kurths J 1996 Phase synchronization of chaotic oscillators Phys Rev Lett 76 1804ndash7Rosenblum M Pikovsky A and Kurths J 1997 From phase to lag synchronization in coupled chaotic oscillators Phys
Rev Lett 78 4193ndash6Rulkov N and Sushchik M 1997 Robustness of synchronized chaotic oscillations Int J Bifurc Chaos 7 625ndash43Rulkov N Sushchik M Tsimring L and Abarbanel H 1995 Generalized synchronization of chaos in unidirectionally
coupled chaotic systems Phys Rev E 51 980ndash94Schiff S So P Chang T Burke R and Sauer T 1996 Detecting dynamical interdependence and generalized synchrony
through mutual prediction in a neural ensemble Phys Rev E 54 6708ndash24Silberstein R B 1995 Neuromodulations of neocortical dynamics Neocortical Dynamics and Human EEG Rhythms
ed P L Nunez (Oxford Oxford University Press)Singer W 1995 Putative functions of temporal correlations in neocortical processing Large-Scale Neuronal Theories
of the Brain ed C Koch and J Davis (London MIT Press)Somers D and Kopell N 1995 Waves and synchrony in networks of oscillators of relaxation and non-relaxation type
Physica D 89 169ndash83Sutton R S and Barto A G 1990 Time derivative models of Pavlovian reinforcement Learning and Computational
Neuroscience Foundations of Adaptive Networks ed M Gabriel and J Moore (Cambridge MA MIT Press) pp497ndash538
Szentagothai J 1983 The modular architectonic principle of neural centers Rev Physiol Biochem Pharmacol 9811ndash61
Tallon-Baudry C Betrand O and Fischer C 2001 Oscillatory synchrony between human extrastriate areas during visualshort-term memory maintenance J Neurosci 21 RC177(1ndash5)
Tass P A 2002 Desynchronization of brain rhythms with soft phase-resetting techniques Biol Cybern 87 102ndash15
732 M Breakspear et al
Tass P Rosenblum M Weule J Kurths J Pikovsky A Volkmann J Schnitzler A and Freund H 1998 Detection of nmphase locking from noisy data application to magnetoencephalography Phys Rev Lett 81 3291ndash4
Terman D and Wang D 1995 Global competition and local cooperation in a network of neural oscillators Physica D81 148ndash76
Terry J and Breakspear M 2003 An improved algorithm for the detection of dynamical interdependence in bivariatetime-series Biol Cybern 88 129ndash36
Tononi G Sporns O and Edelman G M 1994 A measure for brain complexity relating functional segregation andintegration in the nervous system Proc Natl Acad Sci USA 91 5033ndash7
von der Malsburg C 1985 Nervous structures with dynamical links Phys Chem 89 703ndash10Wang X 1991 Period-doublings to chaos in a simple neural network an analytic proof Complex Syst 5 425ndash41Wennekers T and Pasemann F 1996 Synchronous chaos in high dimensional modular neural networks Int J Bifurc
Chaos 6 2055ndash67Zaks M A Parks E H Rosenblum M G and Kurths J 1999 Alternative locking ratios in imperfect phase synchronization
Phys Rev Lett 82 4228ndash31Zhao L and Macau E N 2001 A network of dynamically coupled chaotic maps for scene segmentation IEEE Trans
Neural Netw 12 1375ndash85
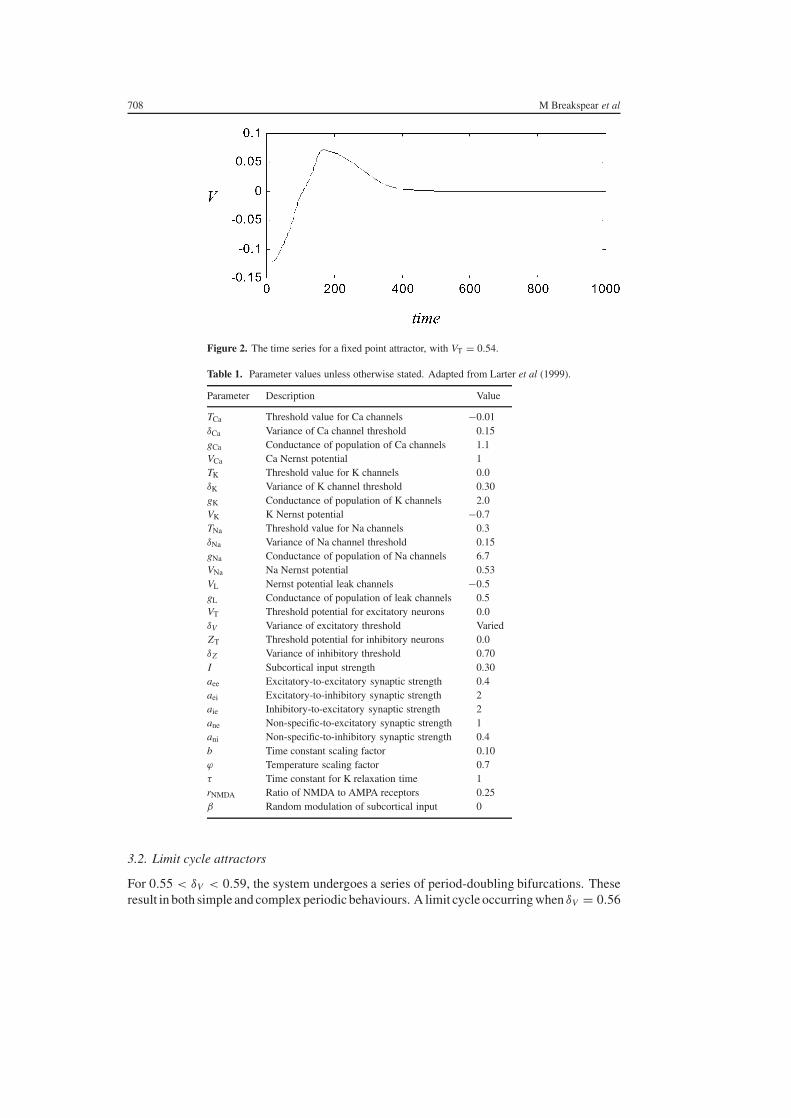
708 M Breakspear et al
Figure 2 The time series for a fixed point attractor with VT = 054
Table 1 Parameter values unless otherwise stated Adapted from Larter et al (1999)
Parameter Description Value
TCa Threshold value for Ca channels minus001δCa Variance of Ca channel threshold 015gCa Conductance of population of Ca channels 11VCa Ca Nernst potential 1TK Threshold value for K channels 00δK Variance of K channel threshold 030gK Conductance of population of K channels 20VK K Nernst potential minus07TNa Threshold value for Na channels 03δNa Variance of Na channel threshold 015gNa Conductance of population of Na channels 67VNa Na Nernst potential 053VL Nernst potential leak channels minus05gL Conductance of population of leak channels 05VT Threshold potential for excitatory neurons 00δV Variance of excitatory threshold VariedZT Threshold potential for inhibitory neurons 00δZ Variance of inhibitory threshold 070I Subcortical input strength 030aee Excitatory-to-excitatory synaptic strength 04aei Excitatory-to-inhibitory synaptic strength 2aie Inhibitory-to-excitatory synaptic strength 2ane Non-specific-to-excitatory synaptic strength 1ani Non-specific-to-inhibitory synaptic strength 04b Time constant scaling factor 010ϕ Temperature scaling factor 07τ Time constant for K relaxation time 1rNMDA Ratio of NMDA to AMPA receptors 025β Random modulation of subcortical input 0
32 Limit cycle attractors
For 055 lt δV lt 059 the system undergoes a series of period-doubling bifurcations Theseresult in both simple and complex periodic behaviours A limit cycle occurring when δV = 056
Synchronization in a model of neuronal dynamics 709
Figure 3 The approach to a limit cycle attractor in phase space with VT = 056
is illustrated in figure 3 Other parameter variations produce more complicated higher-orderperiodic orbits For example a period-9 orbit occurring when δV = 06 aee = 05 aie = 05gNa = 0 and I = 0165 is presented in figure 4
33 Chaotic attractors
Setting δV gt 059 and keeping all other parameters as per table 1 produces complex aperiodicbehaviour in the uncoupled columns An example of this with δV = 065 is illustrated infigure 5 Panels (a) and (b) illustrate a phase space portrait and a time series plot of thissystem respectively They show an aperiodic but highly structured system In panel (c) afirst-return lsquoPoincarersquo section is presented This was obtained by taking a cross-section throughthe attractor in (a) by setting W = 032 and then plotting V (t) against V (t +l)with a time delayof l = 1 A topographic similarity to the simple logistic map is evident It has been provenanalytically that in restricted regions of parameter space dynamical systems incorporatingthe neural activation function (4) (5) are topologically conjugate to the logistic map (Wang1991) The Poincare map presented here shows that although a 11 map may not exist in moregeneral parameter regions the global morphology of the logistic map is still evident
To determine whether or not these aperiodic oscillations are in fact chaotic the Lyapunovexponents were estimated numerically using an algorithm based upon GrammndashSchmidtorthonormalization For the particular case illustrated the exponents were found to be
λ1 = 0015
λ2 = 00
λ3 = minus026
(9)
A principal Lyapunov exponent greater than zero is consistent with chaotic dynamics Thistogether with the topography of the first-return map and the existence of period-doublingbifurcations provides compelling evidence of chaotic behaviour in the dynamics of the singlecolumn The second exponent reflects the rate of exponential separation of adjacent orbits in
710 M Breakspear et al
Figure 4 A period-9 limit cycle attractor (a) A phase space portrait (b) A representative timeseries
the direction tangent to the flow hence a value of zero is to be expected The sum of the threeexponents is negative implying that the system is volume contracting which is to be expectedin the setting of a non-equilibrium (non-Hamiltonian) physiological system
It is interesting to note that this diversity of behaviours can be obtained throughmanipulation of a single state parameter From a functional perspective this permits eachcortical column to express a variety of qualitatively distinct dynamics through modificationof a single physiological parameter such as through modulation of the functional strength ofthe local synaptic connectivity Conversely it also implies that resetting of a single parametersuch as an abnormal δV resulting from loss of physiological heterogeneity could be associatedwith qualitative changes in local cortical behaviour
4 Cooperative behaviour of coupled columns
Whilst it is interesting to observe the range of dynamic behaviours possible in a single columnwe are primarily interested in the behaviour of many such interacting columns In this section
Synchronization in a model of neuronal dynamics 711
1
Figure 5 Chaotic behaviour of the autonomous system with VT = 065 The other parameters areas per table 1 (a) An attractor in phase space (b) A time series for V (t) (c) A Poincare sectionobtained by setting W = 032 and plotting V (t) against V (t + 1)
712 M Breakspear et al
we study the emergence of cooperative behaviour that results from the introduction of couplingbetween columns Three types of interdependent behaviour of two systems are describedidentical generalized and phase synchronization Synchronous behaviours in larger arrays arealso illustrated
41 Identical chaotic synchronization
411 Transition to identical chaotic synchronization The most straightforward extensionand the one that we consider first is that of two coupled cortical columns where all parametersare in the first case assumed identical7 Trivially the two systems will evolve identicallywhen started with identical initial conditions even if C = 0 This is simply a statement ofthe deterministic nature of the evolution equations In general the random initial conditionswill not be equal in which case it is hardly surprising that the different columns will evolveindependently An example is presented in figure 6 with VT = 06 in both systems C = 0and independent random initial conditions In figure 6(a) we show a segment of the timeseries for each of the two subsystems where it can be seen that the time series are chaoticand asynchronous In figure 6(b) the concurrent values of V 1 and V 2 are plotted against eachother showing that they bear no obvious interrelationship
With the introduction of coupling C gt 0 the systems begin to show epochs ofsynchronized behaviour At C sim= 01 there is a transition to stable fully synchronizedmotion as evident in an exemplar time series shown in figure 7(a) where C = 015 Infigure 7(b) we present the concurrent values of V 1 versus V 2 illustrating that in contrast tothe case in figure 6(b) the systems have become embedded in the low-dimensional lsquosymmetrymanifoldrsquo V 1 = V 2 Panel (c) shows a log-linear plot of the temporal evolution of the distanceof the coupled system from this manifold It can be seen that on average the two systemsapproach at an exponential rate with constant time-averaged exponent Whilst this phenomenon(lsquoidentical chaotic synchronizationrsquo) has been described in a variety of other systems since itsfirst description (Fujisaka and Yamada 1983 Pecora and Carroll 1990) to our knowledge thisis its first quantitative8 description in a detailed physiological model of a neural system
The symmetry of the dynamical system obtained by setting the state parameters in eachcolumn identical forces the invariance of the symmetry manifold V 1 = V 2 Moreover thedynamics within this manifold is governed by the same equations of motion as for a singlecortical column This manifold which contains the orbits of the synchronized system is calledthe synchronization manifold The coupling parameter C does not affect the dynamics withinthe synchronization manifold and as such is termed a normal parameter
412 lsquoBlowout bifurcationrsquo An important issue is the stability of the orbits on thesynchronization manifold This can be determined by analysing the Lyapunov spectraBecause the two-column system evolves in a six-dimensional phase space there are sixLyapunov exponents However for the synchronized system it is helpful to subdivide theseinto those that describe the dynamics within the synchronization manifold and those thatdescribe the dynamics in the transverse direction The latter describe the rate of convergenceor divergence of the coupled systems toward or away from this manifoldmdashand hence thetendency for the two systems to synchronize or separate For the time series in figure 7 this is
7 Whilst it is of course physiologically unrealistic to assume that two columns would have entirely identical structuresit is mathematically simpler to start analysing the system in this situation8 A qualitative description in a neural system with a different form of coupling (potassium diffusion) is evident infigure 8 of Larter et al (1999) Synchronous chaos in an abstract model of a neural system was also described inWennekers and Pasemann (1996)
Synchronization in a model of neuronal dynamics 713
1
2
2
1
Figure 6 Independent oscillations in a two-column system with identical parameters butindependent initial conditions (a) The time series (b) A plot of concurrent mean excitatorymembrane potentials V 1 against V 2
related to the slope of the plot in panel (c)9 For this strength of coupling the largest transverseexponent is negative (minus002) indicating that perturbations away from this state of identicalsynchronization decay and thus that the state of identical chaotic synchronization is stableagainst noise (ie dynamically stable)
To calculate the relationship between coupling strength and the Lyapunov exponents weused an algorithm which measures the growth of perturbations of the variational equations (thelsquoJacobianrsquo) of the system (we do not define these equations here as they are cumbersome anddo not enhance the understanding of the system) We calculated these exponents for increasingvalues of the coupling parameter C from 0 to 02 in increments of 0000 25 The results are
9 Only two of the six dependent variables are shown in figure 7 The actual transverse Lyapunov exponents reflectthe temporal evolution of all six and hence are somewhat more complex than that given by the slope of panel (c)
714 M Breakspear et al
1
2
Figure 7 Identical synchronization in the same system as in figure 7 but with C = 02 (a) Thetime series (b) A plot of concurrent mean excitatory membrane potentials V 1 against V 2 (c) Thelogarithm of the Euclidean distance between V 1 and V 2 versus time (linear)
Synchronization in a model of neuronal dynamics 715
Figure 8 Lyapunov spectra for a coupled system with δV = 07 and C varying from 0 to 02 insteps of 0000 33 At each value of C all Lyapunov exponents are calculated Least-squares linesof best fit were then fitted to the results for each exponent Solid lines indicate transverse exponents(λ) and dashed lines indicate the tangential exponents () Panel (a) shows all three transverseand three tangential exponents Panel (b) only shows the largest two of each
illustrated in figure 8 Panel (a) shows all exponents For illustrative purposes only the mostpositive four are plotted in panel (b) In both panels the tangential exponents () are fitted (inthe least-squares sense) with a dashed line and the transverse ones (λ) are fitted with a solidline They are labelled 1ndash3 in order of decreasing positivity The important features are asfollows For the range of coupling strengths that we consider the largest tangential Lyapunovexponent is positive which indicates the presence of chaotic behaviour within the manifoldWhen examining the line of best fit for this exponent we see that this exponent does not varywhen altering the coupling parameter This is true for all three tangential exponents and verifiesthat the changes in the coupling parameter do not alter the dynamics on the synchronizationmanifold (that is the coupling parameter is normal)
716 M Breakspear et al
The largest transverse Lyapunov exponent (denoted as λ1 in the figure) is a particularlyimportant exponent since it governs the stability of the attractor in the synchronization manifoldwith respect to perturbations out of the manifold If this exponent is negative then we expectsmall perturbations of trajectories to reconverge to the manifold in finite time If however thisexponent is positive then we would instead expect perturbed trajectories to diverge from themanifold This change in local behaviour is termed a bifurcation and the point at which itoccurs the bifurcation point It is apparent in figure 8 that the zero crossing of this exponentmdashthe bifurcation pointmdashoccurs at approximately C = 01 in the present system This typeof bifurcation has been termed a blowout bifurcation since once transverse stability is losttrajectories are lsquoblown outrsquo from the low-dimensional synchronization manifold into the fullphase space (Ott and Sommerer 1994) Hence in the present setting a blowout bifurcation isobserved if C falls below 01
We note two other observations Increasing rNMDA which corresponds to increasing thepopulation of NMDA receptors decreases the coupling strength required to achieve stablesynchronization On the other hand increasing the overall excitatory-to-excitatory synapticconnectivity (aee) has the opposite effect This is because the effect of local excitatory feedbackincreases the tangential Lyapunov exponents which has the effect of lsquosplittingrsquo the synchronizeddynamics
In the above simulations all the parameters of both systems were equal In realphysiological systems such symmetry between systems is clearly impossible We nowconsider the case in which the two columns have different structures A simple way toincorporate this whilst at the same time preserving (as much as possible) simple dynamics is tovary the parameter δV for each column This has the effect of destroying any symmetry-forcedinvariant manifolds and so the simple case of synchronization corresponding to identicallyevolving dynamics will not exist in this case However more exotic forms of synchronizationare still possible and we discuss these next
42 Generalized chaotic synchronization
In order to maintain consistency with the literature we study non-linear interdependence in thesetting where the systems are asymmetrical but the coupling is unidirectional This is easilyachieved in the present system by making C directed and setting C21 = 0 In this settingwe refer to the lsquofreersquo system as the driver and the lsquoslavedrsquo system as the response We firstask the question what is the effect of introducing a slight parameter mismatch into a systemexhibiting stable chaos synchronization An example of this is presented in figure 9 whereC12 = 04 and parameter mismatch has been incorporated by setting δV = 07 in the driversystem and δV = 0699 in the response system In panel (a) the time series appear qualitativelyvery similar However in panel (b) where the difference between the systems is plotted it isapparent that the separation between the two systems is of the same order of magnitude as thesize of each attractor In panel (c) the effect of the parameter mismatch on the synchronizationmanifold can be seen as a transformation away from the symmetry manifold
It has been proposed that in such a setting there exists a continuous functional relationshipbetween the dynamics of the driver and response systems
x2(t) = ψ(x1(t)) (10)
where xi = V i W i Z i This situation has been referred to in the literature as generalizedsynchronization (Afraimovich et al 1986 Rulkov et al 1995) The properties of ψ andthe topology and stability of the synchronization manifold in this context have not yetbeen fully elucidated although some significant results have been achieved For example
Synchronization in a model of neuronal dynamics 717
1
2
2
1
Figure 9 Generalized synchronization in an asymmetrical driverndashresponse system (a) The timeseries of the two systems (b) The distance between the two systems (c) A plot of concurrent meanexcitatory membrane potentials V 1 against V 2 The diagonal represents the symmetry manifold
718 M Breakspear et al
Hunt et al (1997) Josic (1998) Kocarev et al (2000) have shown that the generalizedsynchronization manifold is smooth (differentiable) if and only if it is normally hyperbolic(ie the largest transverse Lyapunov exponent is less than the smallest tangential Lyapunovexponent prior to the introduction of parameter mismatch) Such a condition is met in thepresent setting when C = 04 hence the smoothness of the manifold in panel (c)
However subsequent to the introduction of parameter mismatch we will not be able touse transverse and tangential Lyapunov exponents to detect such synchronization since thesynchronization manifold may possess a complex topology for which it is a non-trivial (atbest) or impossible (at worse) task to define transverse and tangential directions For examplethe manifold may be smooth as in figure 9(c) but have a fractal topology If the dynamics arenot normally hyperbolic then the manifold may be fractal but also non-differentiable (Huntet al 1997) and hence will not have a unique tangent direction at any point Consequently wewill not be able to compute tangential or transverse Lyapunov exponents and hence determinethe stability of the synchronization manifold However other techniques for detecting thepresence of generalized synchronization in complex systems have evolved including theauxiliary system approach (Abarbanel et al 1996) and non-linear interdependence (Schiffet al 1996) which are now explored
421 The auxiliary system approach The auxiliary system is a third subsystem which isa replica of the original response system but coupled independently to the driver system andcommencing from independent initial conditions It is possible to demonstrate that showingthe existence of generalized synchronizationmdashthat equation (10) is satisfiedmdashis equivalent toshowing that the response system and the auxiliary system evolve identically (even though theyare not directly coupled) To determine whether or not this is the case reduces to an analysisof the Lyapunov exponents transverse to the symmetry manifold with respect to the responsesystem and the auxiliary system
Whilst this is a somewhat artificial test for generalized synchronization since theproduction of an auxiliary system for a physical system would be impossible such a testis useful in numerical and theoretical studies In figure 10 we present an example of theauxiliary system approach where the parameter δV = 07 for column 1 and 068 for column 2We consider two values of the coupling C = 0025 and 01 The auxiliary system is simplya copy of the second column beginning from an arbitrary initial state In the first instancewhere C = 0025 we notice that even after allowing for an initial transient state the secondcolumn and the auxiliary do not behave in identical fashions (figure 10(a)) However whenC = 01 we observe (figure 10(b)) that the second column and the auxiliary do behave innear-identical manners after an initial transient and hence conclude that for the particularvalue of the coupling strength considered equation (10) is satisfied and hence generalizedsynchronization between the first and second columns occurs
422 Nonlinear interdependence The question remains as to how we may detect thepresence of generalized synchronization in a practical setting As we mentioned aboveit is not possible to use an auxiliary-system-based approach to detect the presence ofgeneralized synchronization Instead we must consider techniques for detecting suchbehaviour from experimental or numerical time series data Algorithms of this type areoften referred to as detecting non-linear interdependence Nonlinear interdependence is ageneral term encompassing systems where the behaviour of one non-linear system affectsthe behaviour of another More specifically it can be defined as occurring if the nullhypothesis that the interdependence between two signals is purely linear can be rejectedat conventional levels (usually 95) of statistical significance hence it is an experimental
Synchronization in a model of neuronal dynamics 719
2
1
2
1
Figure 10 Concurrent evolution of the response system V 2 and lsquoauxiliaryrsquo system V 1 (a) C =0025 (b) C = 01
term (eg Schiff et al 1996) Whilst this does not strictly correspond to the existence of afunctional relationship between the two systems such a relationship is typically used tomotivate the techniques for its detection Schiff et al (1996) extended the logic of non-linearprediction algorithms from autonomous to coupled systems by exploiting the continuity ofsuch a relationship and hence devised the first method of detecting non-linear interdependencein time series data The basis of the technique is using the forward evolution of a phase spacevectorrsquos neighbours to predict the forward evolution of that vector and the forward evolutionof the concurrent vector of the other (possibly interdependent) system If the error of such aprocedure is better on average than chance (the chance error is normalized to equal one) thennon-linear interdependence is said to be present
720 M Breakspear et al
V1
V2
Figure 11 Experimental and numerical examples of weak generalized non-linear synchronizationin the presence of measurement noise (a) Human scalp EEG data from posterior bipolar scalpelectrodes (O1P3ndashO2P4) (b) Weakly coupled aperiodic data in the present model with additivewhite noise (with 5 of the amplitude of the model signal) Parameters δ1
V = 0605 δ2V = 061
aee = 036 C = 007
A variation of this algorithm optimized to detect weak non-linear interdependence (Terryand Breakspear 2003) was used recently to demonstrate that weak non-linear interdependence isoccasionally present in the scalp EEG of healthy resting human subjects (Breakspear and Terry2002a 2002b 2002c) An example of an epoch of lsquonon-linearly independentrsquo EEG obtainedusing this technique is presented in figure 11(a) For comparison output from the currentmodel is presented in figure 11(b) The model parameters were set such that the dynamicswas still aperiodic but the intermittent large-amplitude bursts were suppressed and had weakmutual coupling (C = 007) Extraneous noise to simulate the effect of extracranial muscleactivity contaminating the EEG data has been added to this time series It should be notedthat the amplitudes and time scales of the modelrsquos variables are dimensionless Nonethelessthe model facilitates qualititative interpretation of experimental findings
Synchronization in a model of neuronal dynamics 721
43 Chaotic phase synchronization
Whereas the concept of generalized synchronization was mathematically motivated and assuch more suited to theoretical investigations of systems the concept of phase synchronizationderives from a more practical signal processing approach Phase synchronization occurs inboth symmetric and asymmetric settings but is often more useful in the asymmetric case (andin experimental data)
Unfortunately the phase of a chaotic signal is typically ambiguous and can be defined in anumber of different ways For example in the case of the coherent Rossler attractor (Osipovet al 1997) the phase may be defined as the arctangent of two of the variables of the systemMore generally it can sometimes be defined as the angle that an orbit subtends with the centreof an attractor It is also possible to extract the phase of a chaotic signal from time seriesdata by use of the Hilbert transform This has enabled the demonstration of chaotic phasesynchronization in a number of coupled asymmetrical systems (Rosenblum et al 1996 1997Zaks et al 1999) Although such an approach does not guarantee that the phase will be uniqueit does permit the practical analysis of phase in noisy time series (Tass et al 1998)
To illustrate phase synchronization in two coupled columns we again vary the parameterδV which if the columns are viewed as oscillators acts to detune them from a commonfrequency An important question is that of the criteria on the basis of which we concludewhether phase synchronization occurs We use a simple locking criterion whereby phasesynchronization is said to occur if
|θ1(t)minus θ2(t)| lt K (11)
where is θi the phase of column i and K is a constant Notice that this does not mean thatsynchronization implies equality of phases merely that their difference is bounded
An example of phase synchronization with δV = 065 for column 1 and 066 for column 2is given in figure 12 In this figure rNMDA = 03 Qualitatively some sort of phase coherenceis evident in the time series panel (a) This was confirmed by plotting the sine of thephase difference which has been extracted using the Hilbert transform panel (b) Clearly|sin(θ1 minus θ2)| lt K and hence |θ1 minus θ2| is bounded It can also be seen that the phase-synchronized dynamics occurs on a highly structured manifold depicted in figure 12(c)However unlike the case for the generalized synchronization manifold depicted in figure 9 itis evident that this manifold is not a simple transformation of the symmetry manifold
It is also possible to see lsquobimodalrsquo stability whereby a system jumps betweenapproximately zero phase difference and a phase lag phase synchronization of π An exampleis depicted in figure 12(d) At approximately 2000 s the system can be seen to jumpspontaneously from the first to the second of these modes The Z -variable is plotted forgraphical purposes
5 Intermittency and desynchronization
51 Desynchronization in an array of two systems
Diminishing the strength of coupling across the blowout bifurcation does not lead to a simpletransition from synchronous to asynchronous oscillations but is instead associated withcomplex temporal behaviours Specifically when the largest transverse Lyapunov exponent isclose to zero the coupled systems exhibit epochs of synchronous behaviours punctuated bysharp bursts of desynchronization This is observed in systems exhibiting identicalgeneralizedand phase synchronization An example of such a phenomenon is given in figure 13 The timeseries panel (a) shows synchronous dynamics except at time = 1000 where it can be seenthat the two time series desynchronize briefly This is evident in panel (b) as a disruption in the
722 M Breakspear et al
2
1
Figure 12 Phase-synchronized coupled columns (a) The time series of V 1 (solid) and V 2 (b) Thesine of the phase difference obtained through Hilbert transformation of the time series in panel (a)(c) The phase synchronization manifold in the V 1ndashV 2 plane (d) The time series of Z (solid) andZ showing a phase slip of π between two modes
Synchronization in a model of neuronal dynamics 723
Figure 13 An example of phase desynchronization in coupled asymmetric systems with C = 01Time series plots of (a) each V -variable and (b) their Euclidean distance (c) The temporal evolutionof the phase difference between the two systems (d) An example of the phase difference evolutionover a longer time interval
724 M Breakspear et al
orderly evolution of the distance between the two systems and in panel (c) where the evolutionof the phase difference reveals a brief violation of the criterion |φ1 minus φ2| lt constant Anexample of the phase difference evolution over a longer time interval is shown in figure 13(d)Note that this is a three-dimensional plot with sin(φ1 minus φ2) and cos(φ1 minus φ2) as they- and z-axes Hence phase desynchronizations are evident as complete revolutions aroundthe cylindrical phase space in which this variable evolves It can be seen that these phenomenaoccur at irregular intervals in the temporal domain
This behaviour can be explained by considering the nature of a chaotic attractor In thestatistical sense an arbitrary initial condition is expected to lead to the chaotic behaviour onthe attractor Consequently when we calculate the Lyapunov exponents of the system theseexponents correspond to those of the chaotic attractor Typically however there are an infinitenumber of unstable periodic orbits embedded within the attractor If the path of a trajectorybecomes close to one of these orbits it may lsquoshadowrsquo the orbit for a brief period of time Eachof these orbits has a corresponding set of Lyapunov exponents which (like the attractor) can bedivided into tangential and transverse exponents These also vary when we change the couplingparameter but are typically not the same as the exponents of the attractor Hence the largesttransverse exponent of the attractor may be negative yet there may be positive transverseexponents associated with periodic orbits Considering all of these exponents together wewould therefore only expect to observe complete stability of the synchronization manifold inthe case where all of the transverse Lyapunov exponents are negative Bursting behaviourwill be observed when the one of these exponents becomes positive typically correspondingto a periodic orbit of low order (Pikovsky and Grassberger 1991 Rulkov and Sushchik 1997Maistrenko et al 1998 Heagy et al 1998) Due to the complex structure of such periodic orbitsthe desynchronous bursts occur across a wide range of timescales
In figure 14 we present the results of a long integration (16times105 time units) in which thelengths of sequential synchronous epochs of phase synchronization are studied The parametervalues were δV = 065 for column 1 and 0645 for column 2 with coupling strength10 equal to0125 In panel (a) the power spectrum of these sequential lengths is plotted The whiteness ofthis spectrum indicates that the lengths of consecutive synchronous epochs are independentThis is due to the high-dimensional lsquomixingrsquo effect of desynchronization In panel (b) thelengths of the synchronous epochs have been sorted again into the order of increasing lengthThe results for two coupling strengths are plotted C = 0125 (solid) and 0135 (dashed)Several features of this plot are of note Firstly the lengths of the synchronous epochsare distributed over almost three orders of magnitude Secondly for both plots there is anapproximately log-linear relationship between the length of an epoch and its relative orderThis reflects an approximately exponential relationship between the length of a synchronousepoch and its likelihood of occurrence Thirdly this relationship is skewed toward longersynchronous epochs for the stronger coupling strengths
This type of signal with patches of lsquoorderrsquo occurring over many timescales interruptedby bursts of disorder can be shown to have a high information capacity (Gatlin 1972)Moreover complex phase desynchronization may make an infrequent yet robust contributionto the dynamical pattern of the alpha rhythm in scalp EEG data for resting human subjects(Breakspear 2002) It is tempting to postulate that such a lsquoweakly stablersquo dynamics may assistin the generation of the rich and fluid mental associations that spontaneously emerge in restingcognitive states Although this phenomenon has been previously described in a variety of othersystems to our knowledge this is the first description in a biophysical model of neocortex
10 The introduction of parameter mismatch causes an increase in the strength of coupling required for stablesynchronization above the value of 01 required in the symmetric case
Synchronization in a model of neuronal dynamics 725
Figure 14 Properties of sequential epochs of phase-synchronous behaviour over a long integrationtime δ1
V = 065 δ2V = 0645 aee = 04 (a) The flat (white) power spectrum (b) Synchronous
epochs sorted in order of increasing length (solid C = 0125 dashed C = 0134)
52 Synchronization and complex dynamics in larger arrays
The synchronous and complex phenomena described above are also observed in arrays ofcoupled systems with N gt 2 Expanding the size of the array permits complex dynamicsin the spatial in addition to the temporal domain However the basic principles governingthe type and stability of synchronized motions are the same In this section a few of thepossible behaviours are discussed Physiological variation across the array is modelled bymodifying the parameter δV = 07 in each array by adding a random number of maximummagnitude 001
In figure 15(a) an array of N = 20 uncoupled (C = 0) systems is shown The lack ofsynchronization in this time series is evident By comparison the rapid transition to completesynchronization is evident in figure 15(b) when C = 03 In fact 19 of the 20 subsystems
726 M Breakspear et al
(b)
(c)
(a)
Figure 15 Synchronization phenomena in an array of coupled cortical columns (a) N = 20C = 0 (b) N = 20 C = 03 (c) N = 6 C = 01 (d) N = 12 C = 009 (e) N = 12C = 012 A small perturbation has been applied to one of the columns at t = 2500 s (f) Evolutionof the phase difference between two of the columns in panel (e) For clarity the W -variable isplotted in panel (c) and the Z -variable in (e)
have phase synchronized within one single cycle Intermediate values of coupling strength areassociated with a rich variety of phenomena For example subsystems with the most similarparameter values typically synchronize into clusters of various sizes An example of this isillustrated in figure 15(c) Note that there is no synchronization between the two differentclusters each containing three synchronized subsystems
Another complex behaviour occurs when subsystems transiently synchronize into largeclusters then spontaneously desynchronize again Hence the dynamics of the entire arrayjumps between different transient dynamics characterized by varying degrees of freedom Anexample of this phenomenon is given in figure 15(d) In addition it is possible to inducedesynchronization across an entire array by applying a small perturbation to one columnfor one time step An example is illustrated in figure 15(e) A small perturbation has been
Synchronization in a model of neuronal dynamics 727
Figure 15 (Continued)
added to one column after 2500 time units It can be seen that the entire array subsequentlydesynchronizes This illustrates the sensitivity of the dynamics to external stimuli Finally it ispossible to lsquoinducersquo synchronization between selected subsystems by increasing the strength ofcoupling between them selectively This allows an extra level of complexity to be introducedinto the model For example it may allow targeted information transfer between a partialnetwork of the entire array
As discussed in section 51 a univariate signal which jumps between epochs of lowentropy (high repetition) and bursts of high entropy (high variability) can be shown to havehigh information capacity This is because the repetition of certain patterns (as in the English
728 M Breakspear et al
language) permits error detectability Bursts of high entropy act in the opposite directionto increase message variability With a multivariate signal in addition to optimizing theinformation capacity within each signal the overall information capacity can be increasedthrough the type of itinerant synchronization briefly illustrated in this section Synchronizationacross an entire array increases the message reliability (since a single lsquoerrant letterrsquo in onesignal can be easily detected) Conversely desynchronization into multiple clusters increasesmessage variability since each cluster will produce independent information Hence a neuralarray which switches dynamically between a range of cluster configurations can be seen asadaptive in the sense that high synchronous states can be expressed when error detectabilityis a priority (such as during quick and reliable execution of a familiar task) but the array canrapidly desynchronize if variability becomes salient (for example if the environment suddenlybecomes novel) A formal operationalized demonstration of this principle is to be the subjectof future work
6 Discussion
In this paper we present a dynamical model for activity in an array of weakly coupledlocal neural assemblies The model incorporates the fundamental principles governing cellmembrane potential as determined by voltage- and ligand-gated ion channels In this wayit allows a reasonable amount of physiological complexity to be incorporated includingsynaptic activation and receptor subtypeswithout recourse to overwhelming detail Numericalanalysis of this model reveals a transition from independent oscillations to stable chaoticsynchronization depending on the balance of local versus long-range connectivity andbiophysical heterogeneity These excitatory couplings are subject to ascending subcorticalneuromodulation arising principally from monoaminergic nuclei such as Locus Coeruleus(Silberstein 1995) Neuromodulation of extrinsic connections was incorporated into the modelvia a coupling parameter C The stability of the synchronization is determined by the valueof the largest transverse Lyapunov exponent which in turn depends on the coupling parameterC If the coupling is weakened with the result that this exponent becomes positive a blowoutbifurcation occurs and independent oscillations re-emerge In the vicinity of the transitionnon-linear desynchronizations with complex temporal and spatial properties are observed
It is pleasing that complex and itinerant behaviour reminiscent of real neuronaldynamics emerges through such a simple manipulation of a single dynamic control parameterFurthermore the ubiquitous projections of classical neuromodulatory systems in real brainssuggest a clear biological mechanism by which the brain could self-organize throughmoderating the complexity of its own dynamics There are many reasons that the complicateddynamics of the sort demonstrated in this paper may be neurobiologically important Wemention four
First the presence of multiple clusters of phase-synchronized subsystems as discussedabove may be a uniquely non-linear mechanism facilitating the lsquocarriagersquo and distinction ofmultiple concurrent dynamic cell assemblies (and hence cognitive processing) This may beimportant for perceptual grouping binding and transitions as suggested by others (eg von derMalsburg 1985 Gray and Singer 1989)
Second the facility to decouple and reconfigure dynamic cell assemblies througha neuromodulatory control parameter may be important for avoiding low-entropysynchronization manifolds In the absence of itinerancy synchronization would renderneuronal dynamics incapable of representing changes in sensory input or indeed cognitive stateFor example in the model it was observed that small perturbations to one column were ableto induce a global change in neuronal dynamics by disrupting a state of weak synchronization
Synchronization in a model of neuronal dynamics 729
Inevitably alternative dynamic configurations emerge epitomizing the importance of lsquolabilersquobrain dynamics (Friston 2000a)
Thirdly from a purely functionalist point of view the balance between functionalspecialization and functional integration may rely upon the sorts of mechanism modelledin this paper The brain appears to adhere to two principles of functional organization namelysegregation of function within cortical areas and sub-areas and the integration of these areasin perceptual synthesis cognition and sensorimotor integration (Tononi et al 1994 Fristonet al 1995) The balance between preserving specialized dynamics unique to each arearequired by specialization and the coupling among these areas mediating functional integrationmay be achieved by the neuromodulatory mechanisms described above Significantlythe spontaneous variation in the numbers of degrees of freedom (for example whenlow-dimensional synchronization manifolds are shadowed) imparts a dynamic characteristicto this perspective that is precluded by linear mechanisms alone
Finally from a mechanistic perspective complexity and diversity of brain dynamics arerequired for selection of adaptive responses (Friston 2000b) There are many examples ofselective mechanisms operating within the brain All can be seen as performing an ascenton some objective function or fitness landscape These mechanisms require complex anditinerant dynamics to facilitate a thorough exploration of the landscape These landscapesmay range from those involved in perception (where the landscape is defined by predictionerror see eg Rao and Ballard (1998)) to those involved in value learning (where thelandscape is determined by the outcome of a behaviour eg in temporal difference models (seeeg Sutton and Barto 1990)) In short complexity is necessary to maintain an information-rich environment in which the brain can respond to sensory inputs An essential componentof this is the maintenance of diversity which is a prerequisite for selectionist mechanismsat many scales (Edelman 1993) Understanding the mechanistic basis of complex dynamicsas measured electrophysiologically may well depend on the sort of biologically plausiblemodels used in this paper This understanding may be important for both basic and clinicalneuroscience (eg Tass 2002)
The present study supplements and advances on other research in this field The studyof complex phase synchronization and clustering in neural oscillator networks similar tothe present model has previously been the subject of formal mathematical study Forexample Terman and Wang (1995) studied synchronization of blocks of neural oscillatorsthat were subject to perturbation by connected lsquostimulirsquo Somers and Kopell (1995) studiedthe emergence of both synchronization and travelling waves in networks of coupled neuraloscillators In the work of Terman and Wang the local dynamics were based on neurallsquocaricaturersquo models such as the FitzHughndashNagumo equations and are hence more abstract thanthe current study The approach of Somers and Kopell only required that the local dynamicsexhibit fast and relaxation phases and is hence more abstract still In both studies onlyperiodic dynamics are studied Complex patterns of chaos synchronization in neural arrayshave been characterized by nonlinear methods (Kaneko 1998) and their putative contributionto both learning (Nakajima and Ueda 1996) and perception (Zhao and Macau 2001 Breakspear2001) explored However these studies employed models such as the logistic equation andtent map which are quite abstract in comparison to the present study On the other handHansel and Sompolinsky (1996) studied the role of chaos synchronization in a model ofvisual perception in a cortical hypercolumn using a very similar approach to that adoptedhere but with the additional complexity that synaptic delays were incorporated However inthis and earlier work (Hansel and Sompolinsky 1992) only linear measures of dynamics andsynchrony (specifically the auto- and cross-correlation functions) were employed The analysisof chaotic synchrony in the study of Larter et al (1999) was based only upon observation of
730 M Breakspear et al
the Euclidean difference between variables from two nodes within a larger array Hence thesignificance of the present work is that it provides a quantitative nonlinear analysis of complexchaos synchronization in a biophysical model of cortical dynamics Such research is requiredin order to bridge the gap between quantitative nonlinear research into synchronization andcomputation in abstract models of neural arrays and qualitative or linear analysis in morebiophysically plausible models
Acknowledgments
The authors would like to thank Peter Robinson for assistance with the manuscript and R Larterfor helpful advice MB is a recipient of a NSW Institute of Psychiatry Research Grant anda University of Sydney SESQUI postdoctoral fellowship KJF is supported by the WellcomeTrust
References
Abarbanel H Rulkov N and Sushchik M 1996 Generalised synchronization of chaos the auxiliary system approachPhys Rev E 53 4528ndash35
Afraimovich V Verichev N and Rabinovich M 1986 Stochastic synchronization of oscillation in dissipative systemsRadiophys Quantum Electron 29 795ndash801
Breakspear M 2001 Perception of odours by a nonlinear model of the olfactory bulb Int J Neural Syst 11 101ndash24Breakspear M 2002 Nonlinear phase desynchronisation in human electroencephalographic data Human Brain Mapping
15 175ndash98Breakspear M and Terry J R 2002a Nonlinear interdependence in neural systems motivation theory and relevance
Int J Neurosci 112 1163ndash84Breakspear M and Terry J R 2002b Detection and description of nonlinear interdependence in normal multichannel
human EEG Clin Neurophysiol 113 735ndash53Breakspear M and Terry J R 2002c Topographic organization of nonlinear interdependence in multichannel human
EEG Neuroimage 16 822ndash35Destexhe A Rudolph M Fellous J M and Sejnowski T J 2001 Fluctuating synaptic conductances recreate in vivo-like
activity in neocortical neurons Neuroscience 107 13ndash24Douglas R J Mahowald M A and Martin K A 1996 Microarchitecture of neocortical columns Brain TheorymdashBiological
Basis and Computational Principles ed A Aertsen and V Braitenberg (Amsterdam Elsevier)Eckhorn R Bauer R Jordan W Brosch M Kruse W Munk M and Reitboeck H J 1988 Coherent oscillations a
mechanism of feature linking in the visual cortex Multiple electrode and correlation analyses in the cat BiolCybern 60 121ndash30
Edelman G M 1993 Neural Darwinism selection and reentrant signalling in higher brain function Neuron 10 115ndash25Engel A K Kreiter A K Konig P and Singer W 1991 Synchronization of oscillatory neuronal responses between
striate and extrastriate visual cortical areas of the cat Proc Natl Acad Sci USA 88 6048ndash52Friston K J 2000a The labile brain I Neuronal transients and nonlinear coupling Phil Trans R Soc B 355 215ndash36Friston K J 2000b The labile brain II Transients complexity and selection Phil Trans R Soc B 355 237ndash52Friston K J Tononi G Sporns O and Edelman G 1995 Characterising the complexity of neuronal interactions Human
Brain Mapping 3 302ndash14Fujisaka H and Yamada T 1983 Stability theory of synchronized motion in coupled-oscillator system Prog Theor
Phys 69 32ndash47Gatlin L 1972 Information Theory and the Living System (New York Columbia University Press)Gray C M Konig P Engel A and Singer W 1989 Oscillatory responses in cat visual cortex exhibit inter-columnar
synchronization which reflects global stimulus properties Science 338 334ndash7Gray C M and Singer W 1989 Stimulus specific neuronal oscillations in orientation columns of cat visual cortex Proc
Natl Acad Sci USA 86 1698ndash702Haig A R Gordon E Wright J J Meares R A and Bahramali H 2000 Synchronous cortical gamma-band activity in
task-relevant cognition Neuroreport 11 669ndash75Hansel D and Sompolinsky H 1992 Synchronization and computation in a chaotic neural network Phys Rev Lett 68
718ndash21
Synchronization in a model of neuronal dynamics 731
Hansel D and Sompolinsky H 1996 Chaos and synchrony in a model of a hypercolumn in visual cortex J ComputNeurosci 3 7ndash4
Heagy J Carroll T and Pecora L 1998 Desynchronization by periodic orbits Phys Rev E 57 R1253ndash6Hunt B Ott E and Yorke J 1997 Differentiable synchronization of chaos Phys Rev E 55 4029ndash34Josic K 1998 Invariant manifolds and synchronization of coupled dynamical systems Phys Rev Lett 80 3053ndash6Kandel E Schwartz J and Jessell T 2000 Principles of Neural Science (New York McGraw-Hill)Kaneko K 1998 On the strength of attractors in a high-dimensional system Physica D 124 322ndash44Kocarev L Parlitz U and Brown R 2000 Robust synchronization of chaotic systems Phys Rev E 61 3716ndash20Larter R Speelman B and Worth R M 1999 A coupled ordinary differential equation lattice model for the simulation
of epileptic seizures Chaos 9 795ndash804Lumer E Edelman G and Tononi G 1997a Neural dynamics in a model of the thalamocortical system I Layers loops
and the emergence of fast synchronous oscillations Cereb Cortex 7 207ndash27Lumer E Edelman G and Tononi G 1997b Neural dynamics in a model of the thalamocortical system II The role of
neural synchrony tested through perturbations of spike timing Cereb Cortex 7 228ndash36Magleby K 1987 Short-term modulation at synapses Synaptic Function ed G M Edelman W E Gall and
W M Cowan (New York Wiley)Maistrenko Y Maistrenko V Popovich A and Mosekilde E 1998 Transverse instability and riddled basins in a system
of two coupled logistic maps Phys Rev E 57 2713ndash24Morris C and Lecar H 1981 Voltage oscillations in the barnacle giant muscle fiber Biophys J 35 193ndash213Mountcastle V B 1997 The columnar organization of the neocortex Brain 120 701ndash22Mountcastle V B 1998 The Cerebral Cortex (Cambridge MA Harvard University Press)Nakajima H and Ueda Y 1996 Riddled basins of the optimal states in learning dynamical systems Physica D 99 35ndash44Nunez P L 1995 Neocortical Dynamics and Human Brain Rhythms (Oxford Oxford University Press)Osipov G V Pikovsky A S Rosenblum M G and Kurths J 1997 Phase synchronization effects in a lattice of nonidentical
Rossler oscillators Phys Rev E 55 2353ndash61Ott E and Sommerer J 1994 Blowout bifurcations the occurrence of riddled basins and onndashoff intermittency Phys
Lett A 188 39ndash47Pecora L and Carroll T 1990 Synchronization in chaotic systems Phys Rev Lett 64 821ndash4Pikovsky A and Grassberger P 1991 Symmetry breaking bifurcation for coupled chaotic attractors J Phys A Math
Gen 24 4587ndash97Rao R P and Ballard D H 1998 Predictive coding in the visual cortex a functional interpretation of some extra-classical
receptive field effects Nature Neurosci 2 79ndash87Robinson P A Wright J J and Rennie C R 1998 Synchronous oscillations in the cerebral cortex Phys Rev E 57
4578ndash88Rodriguez E George N Lachauz J P Martinerie J Renault B and Varela F J 1999 Perceptionrsquos shadow long distance
synchronization of human brain activity Nature 397 430ndash3Rosenblum M Pikovsky A and Kurths J 1996 Phase synchronization of chaotic oscillators Phys Rev Lett 76 1804ndash7Rosenblum M Pikovsky A and Kurths J 1997 From phase to lag synchronization in coupled chaotic oscillators Phys
Rev Lett 78 4193ndash6Rulkov N and Sushchik M 1997 Robustness of synchronized chaotic oscillations Int J Bifurc Chaos 7 625ndash43Rulkov N Sushchik M Tsimring L and Abarbanel H 1995 Generalized synchronization of chaos in unidirectionally
coupled chaotic systems Phys Rev E 51 980ndash94Schiff S So P Chang T Burke R and Sauer T 1996 Detecting dynamical interdependence and generalized synchrony
through mutual prediction in a neural ensemble Phys Rev E 54 6708ndash24Silberstein R B 1995 Neuromodulations of neocortical dynamics Neocortical Dynamics and Human EEG Rhythms
ed P L Nunez (Oxford Oxford University Press)Singer W 1995 Putative functions of temporal correlations in neocortical processing Large-Scale Neuronal Theories
of the Brain ed C Koch and J Davis (London MIT Press)Somers D and Kopell N 1995 Waves and synchrony in networks of oscillators of relaxation and non-relaxation type
Physica D 89 169ndash83Sutton R S and Barto A G 1990 Time derivative models of Pavlovian reinforcement Learning and Computational
Neuroscience Foundations of Adaptive Networks ed M Gabriel and J Moore (Cambridge MA MIT Press) pp497ndash538
Szentagothai J 1983 The modular architectonic principle of neural centers Rev Physiol Biochem Pharmacol 9811ndash61
Tallon-Baudry C Betrand O and Fischer C 2001 Oscillatory synchrony between human extrastriate areas during visualshort-term memory maintenance J Neurosci 21 RC177(1ndash5)
Tass P A 2002 Desynchronization of brain rhythms with soft phase-resetting techniques Biol Cybern 87 102ndash15
732 M Breakspear et al
Tass P Rosenblum M Weule J Kurths J Pikovsky A Volkmann J Schnitzler A and Freund H 1998 Detection of nmphase locking from noisy data application to magnetoencephalography Phys Rev Lett 81 3291ndash4
Terman D and Wang D 1995 Global competition and local cooperation in a network of neural oscillators Physica D81 148ndash76
Terry J and Breakspear M 2003 An improved algorithm for the detection of dynamical interdependence in bivariatetime-series Biol Cybern 88 129ndash36
Tononi G Sporns O and Edelman G M 1994 A measure for brain complexity relating functional segregation andintegration in the nervous system Proc Natl Acad Sci USA 91 5033ndash7
von der Malsburg C 1985 Nervous structures with dynamical links Phys Chem 89 703ndash10Wang X 1991 Period-doublings to chaos in a simple neural network an analytic proof Complex Syst 5 425ndash41Wennekers T and Pasemann F 1996 Synchronous chaos in high dimensional modular neural networks Int J Bifurc
Chaos 6 2055ndash67Zaks M A Parks E H Rosenblum M G and Kurths J 1999 Alternative locking ratios in imperfect phase synchronization
Phys Rev Lett 82 4228ndash31Zhao L and Macau E N 2001 A network of dynamically coupled chaotic maps for scene segmentation IEEE Trans
Neural Netw 12 1375ndash85
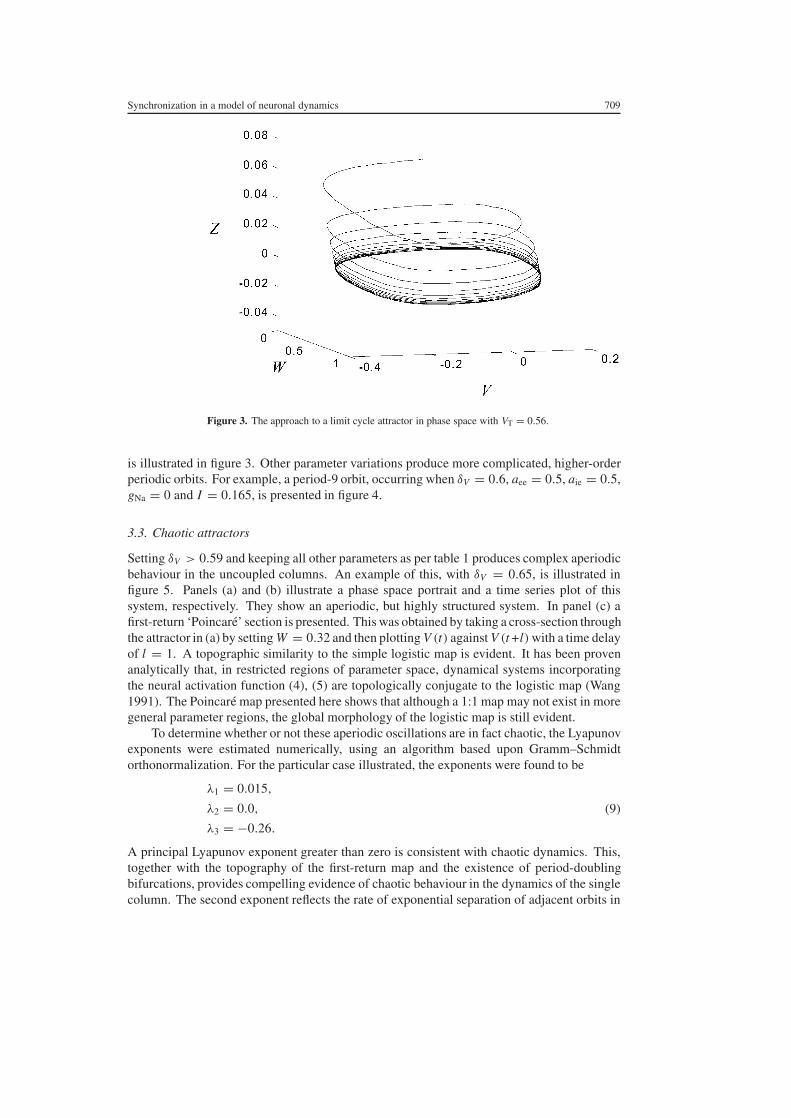
Synchronization in a model of neuronal dynamics 709
Figure 3 The approach to a limit cycle attractor in phase space with VT = 056
is illustrated in figure 3 Other parameter variations produce more complicated higher-orderperiodic orbits For example a period-9 orbit occurring when δV = 06 aee = 05 aie = 05gNa = 0 and I = 0165 is presented in figure 4
33 Chaotic attractors
Setting δV gt 059 and keeping all other parameters as per table 1 produces complex aperiodicbehaviour in the uncoupled columns An example of this with δV = 065 is illustrated infigure 5 Panels (a) and (b) illustrate a phase space portrait and a time series plot of thissystem respectively They show an aperiodic but highly structured system In panel (c) afirst-return lsquoPoincarersquo section is presented This was obtained by taking a cross-section throughthe attractor in (a) by setting W = 032 and then plotting V (t) against V (t +l)with a time delayof l = 1 A topographic similarity to the simple logistic map is evident It has been provenanalytically that in restricted regions of parameter space dynamical systems incorporatingthe neural activation function (4) (5) are topologically conjugate to the logistic map (Wang1991) The Poincare map presented here shows that although a 11 map may not exist in moregeneral parameter regions the global morphology of the logistic map is still evident
To determine whether or not these aperiodic oscillations are in fact chaotic the Lyapunovexponents were estimated numerically using an algorithm based upon GrammndashSchmidtorthonormalization For the particular case illustrated the exponents were found to be
λ1 = 0015
λ2 = 00
λ3 = minus026
(9)
A principal Lyapunov exponent greater than zero is consistent with chaotic dynamics Thistogether with the topography of the first-return map and the existence of period-doublingbifurcations provides compelling evidence of chaotic behaviour in the dynamics of the singlecolumn The second exponent reflects the rate of exponential separation of adjacent orbits in
710 M Breakspear et al
Figure 4 A period-9 limit cycle attractor (a) A phase space portrait (b) A representative timeseries
the direction tangent to the flow hence a value of zero is to be expected The sum of the threeexponents is negative implying that the system is volume contracting which is to be expectedin the setting of a non-equilibrium (non-Hamiltonian) physiological system
It is interesting to note that this diversity of behaviours can be obtained throughmanipulation of a single state parameter From a functional perspective this permits eachcortical column to express a variety of qualitatively distinct dynamics through modificationof a single physiological parameter such as through modulation of the functional strength ofthe local synaptic connectivity Conversely it also implies that resetting of a single parametersuch as an abnormal δV resulting from loss of physiological heterogeneity could be associatedwith qualitative changes in local cortical behaviour
4 Cooperative behaviour of coupled columns
Whilst it is interesting to observe the range of dynamic behaviours possible in a single columnwe are primarily interested in the behaviour of many such interacting columns In this section
Synchronization in a model of neuronal dynamics 711
1
Figure 5 Chaotic behaviour of the autonomous system with VT = 065 The other parameters areas per table 1 (a) An attractor in phase space (b) A time series for V (t) (c) A Poincare sectionobtained by setting W = 032 and plotting V (t) against V (t + 1)
712 M Breakspear et al
we study the emergence of cooperative behaviour that results from the introduction of couplingbetween columns Three types of interdependent behaviour of two systems are describedidentical generalized and phase synchronization Synchronous behaviours in larger arrays arealso illustrated
41 Identical chaotic synchronization
411 Transition to identical chaotic synchronization The most straightforward extensionand the one that we consider first is that of two coupled cortical columns where all parametersare in the first case assumed identical7 Trivially the two systems will evolve identicallywhen started with identical initial conditions even if C = 0 This is simply a statement ofthe deterministic nature of the evolution equations In general the random initial conditionswill not be equal in which case it is hardly surprising that the different columns will evolveindependently An example is presented in figure 6 with VT = 06 in both systems C = 0and independent random initial conditions In figure 6(a) we show a segment of the timeseries for each of the two subsystems where it can be seen that the time series are chaoticand asynchronous In figure 6(b) the concurrent values of V 1 and V 2 are plotted against eachother showing that they bear no obvious interrelationship
With the introduction of coupling C gt 0 the systems begin to show epochs ofsynchronized behaviour At C sim= 01 there is a transition to stable fully synchronizedmotion as evident in an exemplar time series shown in figure 7(a) where C = 015 Infigure 7(b) we present the concurrent values of V 1 versus V 2 illustrating that in contrast tothe case in figure 6(b) the systems have become embedded in the low-dimensional lsquosymmetrymanifoldrsquo V 1 = V 2 Panel (c) shows a log-linear plot of the temporal evolution of the distanceof the coupled system from this manifold It can be seen that on average the two systemsapproach at an exponential rate with constant time-averaged exponent Whilst this phenomenon(lsquoidentical chaotic synchronizationrsquo) has been described in a variety of other systems since itsfirst description (Fujisaka and Yamada 1983 Pecora and Carroll 1990) to our knowledge thisis its first quantitative8 description in a detailed physiological model of a neural system
The symmetry of the dynamical system obtained by setting the state parameters in eachcolumn identical forces the invariance of the symmetry manifold V 1 = V 2 Moreover thedynamics within this manifold is governed by the same equations of motion as for a singlecortical column This manifold which contains the orbits of the synchronized system is calledthe synchronization manifold The coupling parameter C does not affect the dynamics withinthe synchronization manifold and as such is termed a normal parameter
412 lsquoBlowout bifurcationrsquo An important issue is the stability of the orbits on thesynchronization manifold This can be determined by analysing the Lyapunov spectraBecause the two-column system evolves in a six-dimensional phase space there are sixLyapunov exponents However for the synchronized system it is helpful to subdivide theseinto those that describe the dynamics within the synchronization manifold and those thatdescribe the dynamics in the transverse direction The latter describe the rate of convergenceor divergence of the coupled systems toward or away from this manifoldmdashand hence thetendency for the two systems to synchronize or separate For the time series in figure 7 this is
7 Whilst it is of course physiologically unrealistic to assume that two columns would have entirely identical structuresit is mathematically simpler to start analysing the system in this situation8 A qualitative description in a neural system with a different form of coupling (potassium diffusion) is evident infigure 8 of Larter et al (1999) Synchronous chaos in an abstract model of a neural system was also described inWennekers and Pasemann (1996)
Synchronization in a model of neuronal dynamics 713
1
2
2
1
Figure 6 Independent oscillations in a two-column system with identical parameters butindependent initial conditions (a) The time series (b) A plot of concurrent mean excitatorymembrane potentials V 1 against V 2
related to the slope of the plot in panel (c)9 For this strength of coupling the largest transverseexponent is negative (minus002) indicating that perturbations away from this state of identicalsynchronization decay and thus that the state of identical chaotic synchronization is stableagainst noise (ie dynamically stable)
To calculate the relationship between coupling strength and the Lyapunov exponents weused an algorithm which measures the growth of perturbations of the variational equations (thelsquoJacobianrsquo) of the system (we do not define these equations here as they are cumbersome anddo not enhance the understanding of the system) We calculated these exponents for increasingvalues of the coupling parameter C from 0 to 02 in increments of 0000 25 The results are
9 Only two of the six dependent variables are shown in figure 7 The actual transverse Lyapunov exponents reflectthe temporal evolution of all six and hence are somewhat more complex than that given by the slope of panel (c)
714 M Breakspear et al
1
2
Figure 7 Identical synchronization in the same system as in figure 7 but with C = 02 (a) Thetime series (b) A plot of concurrent mean excitatory membrane potentials V 1 against V 2 (c) Thelogarithm of the Euclidean distance between V 1 and V 2 versus time (linear)
Synchronization in a model of neuronal dynamics 715
Figure 8 Lyapunov spectra for a coupled system with δV = 07 and C varying from 0 to 02 insteps of 0000 33 At each value of C all Lyapunov exponents are calculated Least-squares linesof best fit were then fitted to the results for each exponent Solid lines indicate transverse exponents(λ) and dashed lines indicate the tangential exponents () Panel (a) shows all three transverseand three tangential exponents Panel (b) only shows the largest two of each
illustrated in figure 8 Panel (a) shows all exponents For illustrative purposes only the mostpositive four are plotted in panel (b) In both panels the tangential exponents () are fitted (inthe least-squares sense) with a dashed line and the transverse ones (λ) are fitted with a solidline They are labelled 1ndash3 in order of decreasing positivity The important features are asfollows For the range of coupling strengths that we consider the largest tangential Lyapunovexponent is positive which indicates the presence of chaotic behaviour within the manifoldWhen examining the line of best fit for this exponent we see that this exponent does not varywhen altering the coupling parameter This is true for all three tangential exponents and verifiesthat the changes in the coupling parameter do not alter the dynamics on the synchronizationmanifold (that is the coupling parameter is normal)
716 M Breakspear et al
The largest transverse Lyapunov exponent (denoted as λ1 in the figure) is a particularlyimportant exponent since it governs the stability of the attractor in the synchronization manifoldwith respect to perturbations out of the manifold If this exponent is negative then we expectsmall perturbations of trajectories to reconverge to the manifold in finite time If however thisexponent is positive then we would instead expect perturbed trajectories to diverge from themanifold This change in local behaviour is termed a bifurcation and the point at which itoccurs the bifurcation point It is apparent in figure 8 that the zero crossing of this exponentmdashthe bifurcation pointmdashoccurs at approximately C = 01 in the present system This typeof bifurcation has been termed a blowout bifurcation since once transverse stability is losttrajectories are lsquoblown outrsquo from the low-dimensional synchronization manifold into the fullphase space (Ott and Sommerer 1994) Hence in the present setting a blowout bifurcation isobserved if C falls below 01
We note two other observations Increasing rNMDA which corresponds to increasing thepopulation of NMDA receptors decreases the coupling strength required to achieve stablesynchronization On the other hand increasing the overall excitatory-to-excitatory synapticconnectivity (aee) has the opposite effect This is because the effect of local excitatory feedbackincreases the tangential Lyapunov exponents which has the effect of lsquosplittingrsquo the synchronizeddynamics
In the above simulations all the parameters of both systems were equal In realphysiological systems such symmetry between systems is clearly impossible We nowconsider the case in which the two columns have different structures A simple way toincorporate this whilst at the same time preserving (as much as possible) simple dynamics is tovary the parameter δV for each column This has the effect of destroying any symmetry-forcedinvariant manifolds and so the simple case of synchronization corresponding to identicallyevolving dynamics will not exist in this case However more exotic forms of synchronizationare still possible and we discuss these next
42 Generalized chaotic synchronization
In order to maintain consistency with the literature we study non-linear interdependence in thesetting where the systems are asymmetrical but the coupling is unidirectional This is easilyachieved in the present system by making C directed and setting C21 = 0 In this settingwe refer to the lsquofreersquo system as the driver and the lsquoslavedrsquo system as the response We firstask the question what is the effect of introducing a slight parameter mismatch into a systemexhibiting stable chaos synchronization An example of this is presented in figure 9 whereC12 = 04 and parameter mismatch has been incorporated by setting δV = 07 in the driversystem and δV = 0699 in the response system In panel (a) the time series appear qualitativelyvery similar However in panel (b) where the difference between the systems is plotted it isapparent that the separation between the two systems is of the same order of magnitude as thesize of each attractor In panel (c) the effect of the parameter mismatch on the synchronizationmanifold can be seen as a transformation away from the symmetry manifold
It has been proposed that in such a setting there exists a continuous functional relationshipbetween the dynamics of the driver and response systems
x2(t) = ψ(x1(t)) (10)
where xi = V i W i Z i This situation has been referred to in the literature as generalizedsynchronization (Afraimovich et al 1986 Rulkov et al 1995) The properties of ψ andthe topology and stability of the synchronization manifold in this context have not yetbeen fully elucidated although some significant results have been achieved For example
Synchronization in a model of neuronal dynamics 717
1
2
2
1
Figure 9 Generalized synchronization in an asymmetrical driverndashresponse system (a) The timeseries of the two systems (b) The distance between the two systems (c) A plot of concurrent meanexcitatory membrane potentials V 1 against V 2 The diagonal represents the symmetry manifold
718 M Breakspear et al
Hunt et al (1997) Josic (1998) Kocarev et al (2000) have shown that the generalizedsynchronization manifold is smooth (differentiable) if and only if it is normally hyperbolic(ie the largest transverse Lyapunov exponent is less than the smallest tangential Lyapunovexponent prior to the introduction of parameter mismatch) Such a condition is met in thepresent setting when C = 04 hence the smoothness of the manifold in panel (c)
However subsequent to the introduction of parameter mismatch we will not be able touse transverse and tangential Lyapunov exponents to detect such synchronization since thesynchronization manifold may possess a complex topology for which it is a non-trivial (atbest) or impossible (at worse) task to define transverse and tangential directions For examplethe manifold may be smooth as in figure 9(c) but have a fractal topology If the dynamics arenot normally hyperbolic then the manifold may be fractal but also non-differentiable (Huntet al 1997) and hence will not have a unique tangent direction at any point Consequently wewill not be able to compute tangential or transverse Lyapunov exponents and hence determinethe stability of the synchronization manifold However other techniques for detecting thepresence of generalized synchronization in complex systems have evolved including theauxiliary system approach (Abarbanel et al 1996) and non-linear interdependence (Schiffet al 1996) which are now explored
421 The auxiliary system approach The auxiliary system is a third subsystem which isa replica of the original response system but coupled independently to the driver system andcommencing from independent initial conditions It is possible to demonstrate that showingthe existence of generalized synchronizationmdashthat equation (10) is satisfiedmdashis equivalent toshowing that the response system and the auxiliary system evolve identically (even though theyare not directly coupled) To determine whether or not this is the case reduces to an analysisof the Lyapunov exponents transverse to the symmetry manifold with respect to the responsesystem and the auxiliary system
Whilst this is a somewhat artificial test for generalized synchronization since theproduction of an auxiliary system for a physical system would be impossible such a testis useful in numerical and theoretical studies In figure 10 we present an example of theauxiliary system approach where the parameter δV = 07 for column 1 and 068 for column 2We consider two values of the coupling C = 0025 and 01 The auxiliary system is simplya copy of the second column beginning from an arbitrary initial state In the first instancewhere C = 0025 we notice that even after allowing for an initial transient state the secondcolumn and the auxiliary do not behave in identical fashions (figure 10(a)) However whenC = 01 we observe (figure 10(b)) that the second column and the auxiliary do behave innear-identical manners after an initial transient and hence conclude that for the particularvalue of the coupling strength considered equation (10) is satisfied and hence generalizedsynchronization between the first and second columns occurs
422 Nonlinear interdependence The question remains as to how we may detect thepresence of generalized synchronization in a practical setting As we mentioned aboveit is not possible to use an auxiliary-system-based approach to detect the presence ofgeneralized synchronization Instead we must consider techniques for detecting suchbehaviour from experimental or numerical time series data Algorithms of this type areoften referred to as detecting non-linear interdependence Nonlinear interdependence is ageneral term encompassing systems where the behaviour of one non-linear system affectsthe behaviour of another More specifically it can be defined as occurring if the nullhypothesis that the interdependence between two signals is purely linear can be rejectedat conventional levels (usually 95) of statistical significance hence it is an experimental
Synchronization in a model of neuronal dynamics 719
2
1
2
1
Figure 10 Concurrent evolution of the response system V 2 and lsquoauxiliaryrsquo system V 1 (a) C =0025 (b) C = 01
term (eg Schiff et al 1996) Whilst this does not strictly correspond to the existence of afunctional relationship between the two systems such a relationship is typically used tomotivate the techniques for its detection Schiff et al (1996) extended the logic of non-linearprediction algorithms from autonomous to coupled systems by exploiting the continuity ofsuch a relationship and hence devised the first method of detecting non-linear interdependencein time series data The basis of the technique is using the forward evolution of a phase spacevectorrsquos neighbours to predict the forward evolution of that vector and the forward evolutionof the concurrent vector of the other (possibly interdependent) system If the error of such aprocedure is better on average than chance (the chance error is normalized to equal one) thennon-linear interdependence is said to be present
720 M Breakspear et al
V1
V2
Figure 11 Experimental and numerical examples of weak generalized non-linear synchronizationin the presence of measurement noise (a) Human scalp EEG data from posterior bipolar scalpelectrodes (O1P3ndashO2P4) (b) Weakly coupled aperiodic data in the present model with additivewhite noise (with 5 of the amplitude of the model signal) Parameters δ1
V = 0605 δ2V = 061
aee = 036 C = 007
A variation of this algorithm optimized to detect weak non-linear interdependence (Terryand Breakspear 2003) was used recently to demonstrate that weak non-linear interdependence isoccasionally present in the scalp EEG of healthy resting human subjects (Breakspear and Terry2002a 2002b 2002c) An example of an epoch of lsquonon-linearly independentrsquo EEG obtainedusing this technique is presented in figure 11(a) For comparison output from the currentmodel is presented in figure 11(b) The model parameters were set such that the dynamicswas still aperiodic but the intermittent large-amplitude bursts were suppressed and had weakmutual coupling (C = 007) Extraneous noise to simulate the effect of extracranial muscleactivity contaminating the EEG data has been added to this time series It should be notedthat the amplitudes and time scales of the modelrsquos variables are dimensionless Nonethelessthe model facilitates qualititative interpretation of experimental findings
Synchronization in a model of neuronal dynamics 721
43 Chaotic phase synchronization
Whereas the concept of generalized synchronization was mathematically motivated and assuch more suited to theoretical investigations of systems the concept of phase synchronizationderives from a more practical signal processing approach Phase synchronization occurs inboth symmetric and asymmetric settings but is often more useful in the asymmetric case (andin experimental data)
Unfortunately the phase of a chaotic signal is typically ambiguous and can be defined in anumber of different ways For example in the case of the coherent Rossler attractor (Osipovet al 1997) the phase may be defined as the arctangent of two of the variables of the systemMore generally it can sometimes be defined as the angle that an orbit subtends with the centreof an attractor It is also possible to extract the phase of a chaotic signal from time seriesdata by use of the Hilbert transform This has enabled the demonstration of chaotic phasesynchronization in a number of coupled asymmetrical systems (Rosenblum et al 1996 1997Zaks et al 1999) Although such an approach does not guarantee that the phase will be uniqueit does permit the practical analysis of phase in noisy time series (Tass et al 1998)
To illustrate phase synchronization in two coupled columns we again vary the parameterδV which if the columns are viewed as oscillators acts to detune them from a commonfrequency An important question is that of the criteria on the basis of which we concludewhether phase synchronization occurs We use a simple locking criterion whereby phasesynchronization is said to occur if
|θ1(t)minus θ2(t)| lt K (11)
where is θi the phase of column i and K is a constant Notice that this does not mean thatsynchronization implies equality of phases merely that their difference is bounded
An example of phase synchronization with δV = 065 for column 1 and 066 for column 2is given in figure 12 In this figure rNMDA = 03 Qualitatively some sort of phase coherenceis evident in the time series panel (a) This was confirmed by plotting the sine of thephase difference which has been extracted using the Hilbert transform panel (b) Clearly|sin(θ1 minus θ2)| lt K and hence |θ1 minus θ2| is bounded It can also be seen that the phase-synchronized dynamics occurs on a highly structured manifold depicted in figure 12(c)However unlike the case for the generalized synchronization manifold depicted in figure 9 itis evident that this manifold is not a simple transformation of the symmetry manifold
It is also possible to see lsquobimodalrsquo stability whereby a system jumps betweenapproximately zero phase difference and a phase lag phase synchronization of π An exampleis depicted in figure 12(d) At approximately 2000 s the system can be seen to jumpspontaneously from the first to the second of these modes The Z -variable is plotted forgraphical purposes
5 Intermittency and desynchronization
51 Desynchronization in an array of two systems
Diminishing the strength of coupling across the blowout bifurcation does not lead to a simpletransition from synchronous to asynchronous oscillations but is instead associated withcomplex temporal behaviours Specifically when the largest transverse Lyapunov exponent isclose to zero the coupled systems exhibit epochs of synchronous behaviours punctuated bysharp bursts of desynchronization This is observed in systems exhibiting identicalgeneralizedand phase synchronization An example of such a phenomenon is given in figure 13 The timeseries panel (a) shows synchronous dynamics except at time = 1000 where it can be seenthat the two time series desynchronize briefly This is evident in panel (b) as a disruption in the
722 M Breakspear et al
2
1
Figure 12 Phase-synchronized coupled columns (a) The time series of V 1 (solid) and V 2 (b) Thesine of the phase difference obtained through Hilbert transformation of the time series in panel (a)(c) The phase synchronization manifold in the V 1ndashV 2 plane (d) The time series of Z (solid) andZ showing a phase slip of π between two modes
Synchronization in a model of neuronal dynamics 723
Figure 13 An example of phase desynchronization in coupled asymmetric systems with C = 01Time series plots of (a) each V -variable and (b) their Euclidean distance (c) The temporal evolutionof the phase difference between the two systems (d) An example of the phase difference evolutionover a longer time interval
724 M Breakspear et al
orderly evolution of the distance between the two systems and in panel (c) where the evolutionof the phase difference reveals a brief violation of the criterion |φ1 minus φ2| lt constant Anexample of the phase difference evolution over a longer time interval is shown in figure 13(d)Note that this is a three-dimensional plot with sin(φ1 minus φ2) and cos(φ1 minus φ2) as they- and z-axes Hence phase desynchronizations are evident as complete revolutions aroundthe cylindrical phase space in which this variable evolves It can be seen that these phenomenaoccur at irregular intervals in the temporal domain
This behaviour can be explained by considering the nature of a chaotic attractor In thestatistical sense an arbitrary initial condition is expected to lead to the chaotic behaviour onthe attractor Consequently when we calculate the Lyapunov exponents of the system theseexponents correspond to those of the chaotic attractor Typically however there are an infinitenumber of unstable periodic orbits embedded within the attractor If the path of a trajectorybecomes close to one of these orbits it may lsquoshadowrsquo the orbit for a brief period of time Eachof these orbits has a corresponding set of Lyapunov exponents which (like the attractor) can bedivided into tangential and transverse exponents These also vary when we change the couplingparameter but are typically not the same as the exponents of the attractor Hence the largesttransverse exponent of the attractor may be negative yet there may be positive transverseexponents associated with periodic orbits Considering all of these exponents together wewould therefore only expect to observe complete stability of the synchronization manifold inthe case where all of the transverse Lyapunov exponents are negative Bursting behaviourwill be observed when the one of these exponents becomes positive typically correspondingto a periodic orbit of low order (Pikovsky and Grassberger 1991 Rulkov and Sushchik 1997Maistrenko et al 1998 Heagy et al 1998) Due to the complex structure of such periodic orbitsthe desynchronous bursts occur across a wide range of timescales
In figure 14 we present the results of a long integration (16times105 time units) in which thelengths of sequential synchronous epochs of phase synchronization are studied The parametervalues were δV = 065 for column 1 and 0645 for column 2 with coupling strength10 equal to0125 In panel (a) the power spectrum of these sequential lengths is plotted The whiteness ofthis spectrum indicates that the lengths of consecutive synchronous epochs are independentThis is due to the high-dimensional lsquomixingrsquo effect of desynchronization In panel (b) thelengths of the synchronous epochs have been sorted again into the order of increasing lengthThe results for two coupling strengths are plotted C = 0125 (solid) and 0135 (dashed)Several features of this plot are of note Firstly the lengths of the synchronous epochsare distributed over almost three orders of magnitude Secondly for both plots there is anapproximately log-linear relationship between the length of an epoch and its relative orderThis reflects an approximately exponential relationship between the length of a synchronousepoch and its likelihood of occurrence Thirdly this relationship is skewed toward longersynchronous epochs for the stronger coupling strengths
This type of signal with patches of lsquoorderrsquo occurring over many timescales interruptedby bursts of disorder can be shown to have a high information capacity (Gatlin 1972)Moreover complex phase desynchronization may make an infrequent yet robust contributionto the dynamical pattern of the alpha rhythm in scalp EEG data for resting human subjects(Breakspear 2002) It is tempting to postulate that such a lsquoweakly stablersquo dynamics may assistin the generation of the rich and fluid mental associations that spontaneously emerge in restingcognitive states Although this phenomenon has been previously described in a variety of othersystems to our knowledge this is the first description in a biophysical model of neocortex
10 The introduction of parameter mismatch causes an increase in the strength of coupling required for stablesynchronization above the value of 01 required in the symmetric case
Synchronization in a model of neuronal dynamics 725
Figure 14 Properties of sequential epochs of phase-synchronous behaviour over a long integrationtime δ1
V = 065 δ2V = 0645 aee = 04 (a) The flat (white) power spectrum (b) Synchronous
epochs sorted in order of increasing length (solid C = 0125 dashed C = 0134)
52 Synchronization and complex dynamics in larger arrays
The synchronous and complex phenomena described above are also observed in arrays ofcoupled systems with N gt 2 Expanding the size of the array permits complex dynamicsin the spatial in addition to the temporal domain However the basic principles governingthe type and stability of synchronized motions are the same In this section a few of thepossible behaviours are discussed Physiological variation across the array is modelled bymodifying the parameter δV = 07 in each array by adding a random number of maximummagnitude 001
In figure 15(a) an array of N = 20 uncoupled (C = 0) systems is shown The lack ofsynchronization in this time series is evident By comparison the rapid transition to completesynchronization is evident in figure 15(b) when C = 03 In fact 19 of the 20 subsystems
726 M Breakspear et al
(b)
(c)
(a)
Figure 15 Synchronization phenomena in an array of coupled cortical columns (a) N = 20C = 0 (b) N = 20 C = 03 (c) N = 6 C = 01 (d) N = 12 C = 009 (e) N = 12C = 012 A small perturbation has been applied to one of the columns at t = 2500 s (f) Evolutionof the phase difference between two of the columns in panel (e) For clarity the W -variable isplotted in panel (c) and the Z -variable in (e)
have phase synchronized within one single cycle Intermediate values of coupling strength areassociated with a rich variety of phenomena For example subsystems with the most similarparameter values typically synchronize into clusters of various sizes An example of this isillustrated in figure 15(c) Note that there is no synchronization between the two differentclusters each containing three synchronized subsystems
Another complex behaviour occurs when subsystems transiently synchronize into largeclusters then spontaneously desynchronize again Hence the dynamics of the entire arrayjumps between different transient dynamics characterized by varying degrees of freedom Anexample of this phenomenon is given in figure 15(d) In addition it is possible to inducedesynchronization across an entire array by applying a small perturbation to one columnfor one time step An example is illustrated in figure 15(e) A small perturbation has been
Synchronization in a model of neuronal dynamics 727
Figure 15 (Continued)
added to one column after 2500 time units It can be seen that the entire array subsequentlydesynchronizes This illustrates the sensitivity of the dynamics to external stimuli Finally it ispossible to lsquoinducersquo synchronization between selected subsystems by increasing the strength ofcoupling between them selectively This allows an extra level of complexity to be introducedinto the model For example it may allow targeted information transfer between a partialnetwork of the entire array
As discussed in section 51 a univariate signal which jumps between epochs of lowentropy (high repetition) and bursts of high entropy (high variability) can be shown to havehigh information capacity This is because the repetition of certain patterns (as in the English
728 M Breakspear et al
language) permits error detectability Bursts of high entropy act in the opposite directionto increase message variability With a multivariate signal in addition to optimizing theinformation capacity within each signal the overall information capacity can be increasedthrough the type of itinerant synchronization briefly illustrated in this section Synchronizationacross an entire array increases the message reliability (since a single lsquoerrant letterrsquo in onesignal can be easily detected) Conversely desynchronization into multiple clusters increasesmessage variability since each cluster will produce independent information Hence a neuralarray which switches dynamically between a range of cluster configurations can be seen asadaptive in the sense that high synchronous states can be expressed when error detectabilityis a priority (such as during quick and reliable execution of a familiar task) but the array canrapidly desynchronize if variability becomes salient (for example if the environment suddenlybecomes novel) A formal operationalized demonstration of this principle is to be the subjectof future work
6 Discussion
In this paper we present a dynamical model for activity in an array of weakly coupledlocal neural assemblies The model incorporates the fundamental principles governing cellmembrane potential as determined by voltage- and ligand-gated ion channels In this wayit allows a reasonable amount of physiological complexity to be incorporated includingsynaptic activation and receptor subtypeswithout recourse to overwhelming detail Numericalanalysis of this model reveals a transition from independent oscillations to stable chaoticsynchronization depending on the balance of local versus long-range connectivity andbiophysical heterogeneity These excitatory couplings are subject to ascending subcorticalneuromodulation arising principally from monoaminergic nuclei such as Locus Coeruleus(Silberstein 1995) Neuromodulation of extrinsic connections was incorporated into the modelvia a coupling parameter C The stability of the synchronization is determined by the valueof the largest transverse Lyapunov exponent which in turn depends on the coupling parameterC If the coupling is weakened with the result that this exponent becomes positive a blowoutbifurcation occurs and independent oscillations re-emerge In the vicinity of the transitionnon-linear desynchronizations with complex temporal and spatial properties are observed
It is pleasing that complex and itinerant behaviour reminiscent of real neuronaldynamics emerges through such a simple manipulation of a single dynamic control parameterFurthermore the ubiquitous projections of classical neuromodulatory systems in real brainssuggest a clear biological mechanism by which the brain could self-organize throughmoderating the complexity of its own dynamics There are many reasons that the complicateddynamics of the sort demonstrated in this paper may be neurobiologically important Wemention four
First the presence of multiple clusters of phase-synchronized subsystems as discussedabove may be a uniquely non-linear mechanism facilitating the lsquocarriagersquo and distinction ofmultiple concurrent dynamic cell assemblies (and hence cognitive processing) This may beimportant for perceptual grouping binding and transitions as suggested by others (eg von derMalsburg 1985 Gray and Singer 1989)
Second the facility to decouple and reconfigure dynamic cell assemblies througha neuromodulatory control parameter may be important for avoiding low-entropysynchronization manifolds In the absence of itinerancy synchronization would renderneuronal dynamics incapable of representing changes in sensory input or indeed cognitive stateFor example in the model it was observed that small perturbations to one column were ableto induce a global change in neuronal dynamics by disrupting a state of weak synchronization
Synchronization in a model of neuronal dynamics 729
Inevitably alternative dynamic configurations emerge epitomizing the importance of lsquolabilersquobrain dynamics (Friston 2000a)
Thirdly from a purely functionalist point of view the balance between functionalspecialization and functional integration may rely upon the sorts of mechanism modelledin this paper The brain appears to adhere to two principles of functional organization namelysegregation of function within cortical areas and sub-areas and the integration of these areasin perceptual synthesis cognition and sensorimotor integration (Tononi et al 1994 Fristonet al 1995) The balance between preserving specialized dynamics unique to each arearequired by specialization and the coupling among these areas mediating functional integrationmay be achieved by the neuromodulatory mechanisms described above Significantlythe spontaneous variation in the numbers of degrees of freedom (for example whenlow-dimensional synchronization manifolds are shadowed) imparts a dynamic characteristicto this perspective that is precluded by linear mechanisms alone
Finally from a mechanistic perspective complexity and diversity of brain dynamics arerequired for selection of adaptive responses (Friston 2000b) There are many examples ofselective mechanisms operating within the brain All can be seen as performing an ascenton some objective function or fitness landscape These mechanisms require complex anditinerant dynamics to facilitate a thorough exploration of the landscape These landscapesmay range from those involved in perception (where the landscape is defined by predictionerror see eg Rao and Ballard (1998)) to those involved in value learning (where thelandscape is determined by the outcome of a behaviour eg in temporal difference models (seeeg Sutton and Barto 1990)) In short complexity is necessary to maintain an information-rich environment in which the brain can respond to sensory inputs An essential componentof this is the maintenance of diversity which is a prerequisite for selectionist mechanismsat many scales (Edelman 1993) Understanding the mechanistic basis of complex dynamicsas measured electrophysiologically may well depend on the sort of biologically plausiblemodels used in this paper This understanding may be important for both basic and clinicalneuroscience (eg Tass 2002)
The present study supplements and advances on other research in this field The studyof complex phase synchronization and clustering in neural oscillator networks similar tothe present model has previously been the subject of formal mathematical study Forexample Terman and Wang (1995) studied synchronization of blocks of neural oscillatorsthat were subject to perturbation by connected lsquostimulirsquo Somers and Kopell (1995) studiedthe emergence of both synchronization and travelling waves in networks of coupled neuraloscillators In the work of Terman and Wang the local dynamics were based on neurallsquocaricaturersquo models such as the FitzHughndashNagumo equations and are hence more abstract thanthe current study The approach of Somers and Kopell only required that the local dynamicsexhibit fast and relaxation phases and is hence more abstract still In both studies onlyperiodic dynamics are studied Complex patterns of chaos synchronization in neural arrayshave been characterized by nonlinear methods (Kaneko 1998) and their putative contributionto both learning (Nakajima and Ueda 1996) and perception (Zhao and Macau 2001 Breakspear2001) explored However these studies employed models such as the logistic equation andtent map which are quite abstract in comparison to the present study On the other handHansel and Sompolinsky (1996) studied the role of chaos synchronization in a model ofvisual perception in a cortical hypercolumn using a very similar approach to that adoptedhere but with the additional complexity that synaptic delays were incorporated However inthis and earlier work (Hansel and Sompolinsky 1992) only linear measures of dynamics andsynchrony (specifically the auto- and cross-correlation functions) were employed The analysisof chaotic synchrony in the study of Larter et al (1999) was based only upon observation of
730 M Breakspear et al
the Euclidean difference between variables from two nodes within a larger array Hence thesignificance of the present work is that it provides a quantitative nonlinear analysis of complexchaos synchronization in a biophysical model of cortical dynamics Such research is requiredin order to bridge the gap between quantitative nonlinear research into synchronization andcomputation in abstract models of neural arrays and qualitative or linear analysis in morebiophysically plausible models
Acknowledgments
The authors would like to thank Peter Robinson for assistance with the manuscript and R Larterfor helpful advice MB is a recipient of a NSW Institute of Psychiatry Research Grant anda University of Sydney SESQUI postdoctoral fellowship KJF is supported by the WellcomeTrust
References
Abarbanel H Rulkov N and Sushchik M 1996 Generalised synchronization of chaos the auxiliary system approachPhys Rev E 53 4528ndash35
Afraimovich V Verichev N and Rabinovich M 1986 Stochastic synchronization of oscillation in dissipative systemsRadiophys Quantum Electron 29 795ndash801
Breakspear M 2001 Perception of odours by a nonlinear model of the olfactory bulb Int J Neural Syst 11 101ndash24Breakspear M 2002 Nonlinear phase desynchronisation in human electroencephalographic data Human Brain Mapping
15 175ndash98Breakspear M and Terry J R 2002a Nonlinear interdependence in neural systems motivation theory and relevance
Int J Neurosci 112 1163ndash84Breakspear M and Terry J R 2002b Detection and description of nonlinear interdependence in normal multichannel
human EEG Clin Neurophysiol 113 735ndash53Breakspear M and Terry J R 2002c Topographic organization of nonlinear interdependence in multichannel human
EEG Neuroimage 16 822ndash35Destexhe A Rudolph M Fellous J M and Sejnowski T J 2001 Fluctuating synaptic conductances recreate in vivo-like
activity in neocortical neurons Neuroscience 107 13ndash24Douglas R J Mahowald M A and Martin K A 1996 Microarchitecture of neocortical columns Brain TheorymdashBiological
Basis and Computational Principles ed A Aertsen and V Braitenberg (Amsterdam Elsevier)Eckhorn R Bauer R Jordan W Brosch M Kruse W Munk M and Reitboeck H J 1988 Coherent oscillations a
mechanism of feature linking in the visual cortex Multiple electrode and correlation analyses in the cat BiolCybern 60 121ndash30
Edelman G M 1993 Neural Darwinism selection and reentrant signalling in higher brain function Neuron 10 115ndash25Engel A K Kreiter A K Konig P and Singer W 1991 Synchronization of oscillatory neuronal responses between
striate and extrastriate visual cortical areas of the cat Proc Natl Acad Sci USA 88 6048ndash52Friston K J 2000a The labile brain I Neuronal transients and nonlinear coupling Phil Trans R Soc B 355 215ndash36Friston K J 2000b The labile brain II Transients complexity and selection Phil Trans R Soc B 355 237ndash52Friston K J Tononi G Sporns O and Edelman G 1995 Characterising the complexity of neuronal interactions Human
Brain Mapping 3 302ndash14Fujisaka H and Yamada T 1983 Stability theory of synchronized motion in coupled-oscillator system Prog Theor
Phys 69 32ndash47Gatlin L 1972 Information Theory and the Living System (New York Columbia University Press)Gray C M Konig P Engel A and Singer W 1989 Oscillatory responses in cat visual cortex exhibit inter-columnar
synchronization which reflects global stimulus properties Science 338 334ndash7Gray C M and Singer W 1989 Stimulus specific neuronal oscillations in orientation columns of cat visual cortex Proc
Natl Acad Sci USA 86 1698ndash702Haig A R Gordon E Wright J J Meares R A and Bahramali H 2000 Synchronous cortical gamma-band activity in
task-relevant cognition Neuroreport 11 669ndash75Hansel D and Sompolinsky H 1992 Synchronization and computation in a chaotic neural network Phys Rev Lett 68
718ndash21
Synchronization in a model of neuronal dynamics 731
Hansel D and Sompolinsky H 1996 Chaos and synchrony in a model of a hypercolumn in visual cortex J ComputNeurosci 3 7ndash4
Heagy J Carroll T and Pecora L 1998 Desynchronization by periodic orbits Phys Rev E 57 R1253ndash6Hunt B Ott E and Yorke J 1997 Differentiable synchronization of chaos Phys Rev E 55 4029ndash34Josic K 1998 Invariant manifolds and synchronization of coupled dynamical systems Phys Rev Lett 80 3053ndash6Kandel E Schwartz J and Jessell T 2000 Principles of Neural Science (New York McGraw-Hill)Kaneko K 1998 On the strength of attractors in a high-dimensional system Physica D 124 322ndash44Kocarev L Parlitz U and Brown R 2000 Robust synchronization of chaotic systems Phys Rev E 61 3716ndash20Larter R Speelman B and Worth R M 1999 A coupled ordinary differential equation lattice model for the simulation
of epileptic seizures Chaos 9 795ndash804Lumer E Edelman G and Tononi G 1997a Neural dynamics in a model of the thalamocortical system I Layers loops
and the emergence of fast synchronous oscillations Cereb Cortex 7 207ndash27Lumer E Edelman G and Tononi G 1997b Neural dynamics in a model of the thalamocortical system II The role of
neural synchrony tested through perturbations of spike timing Cereb Cortex 7 228ndash36Magleby K 1987 Short-term modulation at synapses Synaptic Function ed G M Edelman W E Gall and
W M Cowan (New York Wiley)Maistrenko Y Maistrenko V Popovich A and Mosekilde E 1998 Transverse instability and riddled basins in a system
of two coupled logistic maps Phys Rev E 57 2713ndash24Morris C and Lecar H 1981 Voltage oscillations in the barnacle giant muscle fiber Biophys J 35 193ndash213Mountcastle V B 1997 The columnar organization of the neocortex Brain 120 701ndash22Mountcastle V B 1998 The Cerebral Cortex (Cambridge MA Harvard University Press)Nakajima H and Ueda Y 1996 Riddled basins of the optimal states in learning dynamical systems Physica D 99 35ndash44Nunez P L 1995 Neocortical Dynamics and Human Brain Rhythms (Oxford Oxford University Press)Osipov G V Pikovsky A S Rosenblum M G and Kurths J 1997 Phase synchronization effects in a lattice of nonidentical
Rossler oscillators Phys Rev E 55 2353ndash61Ott E and Sommerer J 1994 Blowout bifurcations the occurrence of riddled basins and onndashoff intermittency Phys
Lett A 188 39ndash47Pecora L and Carroll T 1990 Synchronization in chaotic systems Phys Rev Lett 64 821ndash4Pikovsky A and Grassberger P 1991 Symmetry breaking bifurcation for coupled chaotic attractors J Phys A Math
Gen 24 4587ndash97Rao R P and Ballard D H 1998 Predictive coding in the visual cortex a functional interpretation of some extra-classical
receptive field effects Nature Neurosci 2 79ndash87Robinson P A Wright J J and Rennie C R 1998 Synchronous oscillations in the cerebral cortex Phys Rev E 57
4578ndash88Rodriguez E George N Lachauz J P Martinerie J Renault B and Varela F J 1999 Perceptionrsquos shadow long distance
synchronization of human brain activity Nature 397 430ndash3Rosenblum M Pikovsky A and Kurths J 1996 Phase synchronization of chaotic oscillators Phys Rev Lett 76 1804ndash7Rosenblum M Pikovsky A and Kurths J 1997 From phase to lag synchronization in coupled chaotic oscillators Phys
Rev Lett 78 4193ndash6Rulkov N and Sushchik M 1997 Robustness of synchronized chaotic oscillations Int J Bifurc Chaos 7 625ndash43Rulkov N Sushchik M Tsimring L and Abarbanel H 1995 Generalized synchronization of chaos in unidirectionally
coupled chaotic systems Phys Rev E 51 980ndash94Schiff S So P Chang T Burke R and Sauer T 1996 Detecting dynamical interdependence and generalized synchrony
through mutual prediction in a neural ensemble Phys Rev E 54 6708ndash24Silberstein R B 1995 Neuromodulations of neocortical dynamics Neocortical Dynamics and Human EEG Rhythms
ed P L Nunez (Oxford Oxford University Press)Singer W 1995 Putative functions of temporal correlations in neocortical processing Large-Scale Neuronal Theories
of the Brain ed C Koch and J Davis (London MIT Press)Somers D and Kopell N 1995 Waves and synchrony in networks of oscillators of relaxation and non-relaxation type
Physica D 89 169ndash83Sutton R S and Barto A G 1990 Time derivative models of Pavlovian reinforcement Learning and Computational
Neuroscience Foundations of Adaptive Networks ed M Gabriel and J Moore (Cambridge MA MIT Press) pp497ndash538
Szentagothai J 1983 The modular architectonic principle of neural centers Rev Physiol Biochem Pharmacol 9811ndash61
Tallon-Baudry C Betrand O and Fischer C 2001 Oscillatory synchrony between human extrastriate areas during visualshort-term memory maintenance J Neurosci 21 RC177(1ndash5)
Tass P A 2002 Desynchronization of brain rhythms with soft phase-resetting techniques Biol Cybern 87 102ndash15
732 M Breakspear et al
Tass P Rosenblum M Weule J Kurths J Pikovsky A Volkmann J Schnitzler A and Freund H 1998 Detection of nmphase locking from noisy data application to magnetoencephalography Phys Rev Lett 81 3291ndash4
Terman D and Wang D 1995 Global competition and local cooperation in a network of neural oscillators Physica D81 148ndash76
Terry J and Breakspear M 2003 An improved algorithm for the detection of dynamical interdependence in bivariatetime-series Biol Cybern 88 129ndash36
Tononi G Sporns O and Edelman G M 1994 A measure for brain complexity relating functional segregation andintegration in the nervous system Proc Natl Acad Sci USA 91 5033ndash7
von der Malsburg C 1985 Nervous structures with dynamical links Phys Chem 89 703ndash10Wang X 1991 Period-doublings to chaos in a simple neural network an analytic proof Complex Syst 5 425ndash41Wennekers T and Pasemann F 1996 Synchronous chaos in high dimensional modular neural networks Int J Bifurc
Chaos 6 2055ndash67Zaks M A Parks E H Rosenblum M G and Kurths J 1999 Alternative locking ratios in imperfect phase synchronization
Phys Rev Lett 82 4228ndash31Zhao L and Macau E N 2001 A network of dynamically coupled chaotic maps for scene segmentation IEEE Trans
Neural Netw 12 1375ndash85
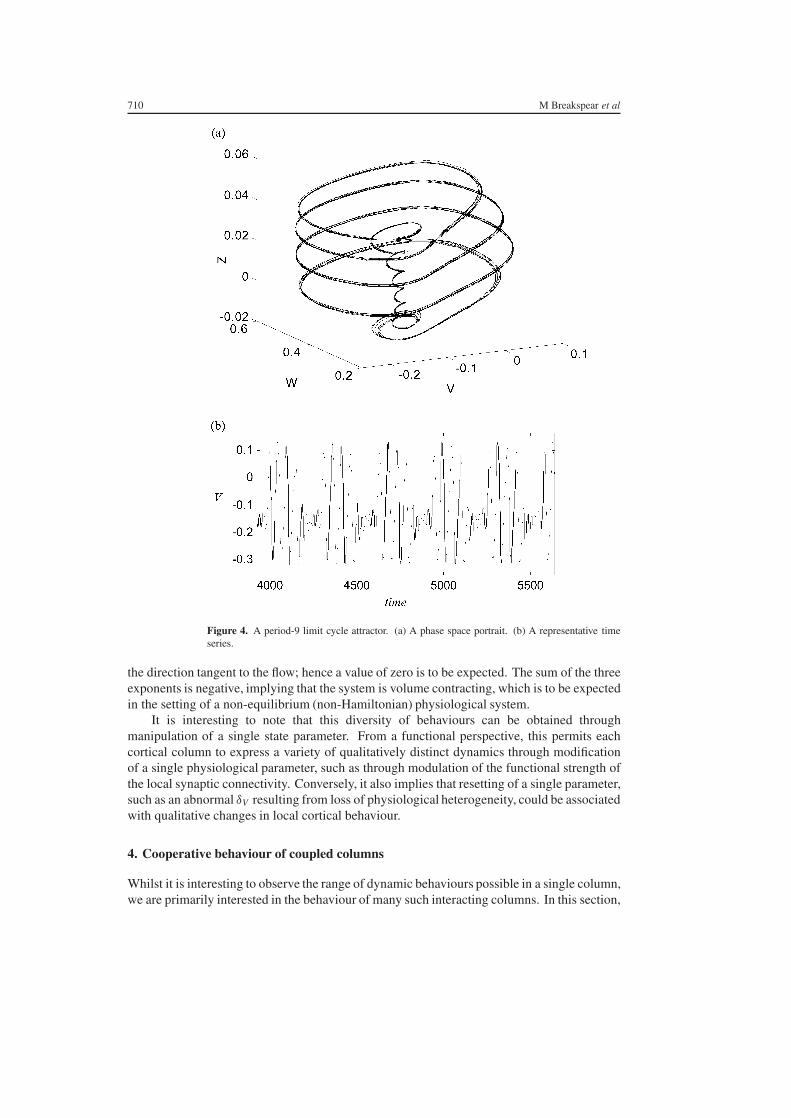
710 M Breakspear et al
Figure 4 A period-9 limit cycle attractor (a) A phase space portrait (b) A representative timeseries
the direction tangent to the flow hence a value of zero is to be expected The sum of the threeexponents is negative implying that the system is volume contracting which is to be expectedin the setting of a non-equilibrium (non-Hamiltonian) physiological system
It is interesting to note that this diversity of behaviours can be obtained throughmanipulation of a single state parameter From a functional perspective this permits eachcortical column to express a variety of qualitatively distinct dynamics through modificationof a single physiological parameter such as through modulation of the functional strength ofthe local synaptic connectivity Conversely it also implies that resetting of a single parametersuch as an abnormal δV resulting from loss of physiological heterogeneity could be associatedwith qualitative changes in local cortical behaviour
4 Cooperative behaviour of coupled columns
Whilst it is interesting to observe the range of dynamic behaviours possible in a single columnwe are primarily interested in the behaviour of many such interacting columns In this section
Synchronization in a model of neuronal dynamics 711
1
Figure 5 Chaotic behaviour of the autonomous system with VT = 065 The other parameters areas per table 1 (a) An attractor in phase space (b) A time series for V (t) (c) A Poincare sectionobtained by setting W = 032 and plotting V (t) against V (t + 1)
712 M Breakspear et al
we study the emergence of cooperative behaviour that results from the introduction of couplingbetween columns Three types of interdependent behaviour of two systems are describedidentical generalized and phase synchronization Synchronous behaviours in larger arrays arealso illustrated
41 Identical chaotic synchronization
411 Transition to identical chaotic synchronization The most straightforward extensionand the one that we consider first is that of two coupled cortical columns where all parametersare in the first case assumed identical7 Trivially the two systems will evolve identicallywhen started with identical initial conditions even if C = 0 This is simply a statement ofthe deterministic nature of the evolution equations In general the random initial conditionswill not be equal in which case it is hardly surprising that the different columns will evolveindependently An example is presented in figure 6 with VT = 06 in both systems C = 0and independent random initial conditions In figure 6(a) we show a segment of the timeseries for each of the two subsystems where it can be seen that the time series are chaoticand asynchronous In figure 6(b) the concurrent values of V 1 and V 2 are plotted against eachother showing that they bear no obvious interrelationship
With the introduction of coupling C gt 0 the systems begin to show epochs ofsynchronized behaviour At C sim= 01 there is a transition to stable fully synchronizedmotion as evident in an exemplar time series shown in figure 7(a) where C = 015 Infigure 7(b) we present the concurrent values of V 1 versus V 2 illustrating that in contrast tothe case in figure 6(b) the systems have become embedded in the low-dimensional lsquosymmetrymanifoldrsquo V 1 = V 2 Panel (c) shows a log-linear plot of the temporal evolution of the distanceof the coupled system from this manifold It can be seen that on average the two systemsapproach at an exponential rate with constant time-averaged exponent Whilst this phenomenon(lsquoidentical chaotic synchronizationrsquo) has been described in a variety of other systems since itsfirst description (Fujisaka and Yamada 1983 Pecora and Carroll 1990) to our knowledge thisis its first quantitative8 description in a detailed physiological model of a neural system
The symmetry of the dynamical system obtained by setting the state parameters in eachcolumn identical forces the invariance of the symmetry manifold V 1 = V 2 Moreover thedynamics within this manifold is governed by the same equations of motion as for a singlecortical column This manifold which contains the orbits of the synchronized system is calledthe synchronization manifold The coupling parameter C does not affect the dynamics withinthe synchronization manifold and as such is termed a normal parameter
412 lsquoBlowout bifurcationrsquo An important issue is the stability of the orbits on thesynchronization manifold This can be determined by analysing the Lyapunov spectraBecause the two-column system evolves in a six-dimensional phase space there are sixLyapunov exponents However for the synchronized system it is helpful to subdivide theseinto those that describe the dynamics within the synchronization manifold and those thatdescribe the dynamics in the transverse direction The latter describe the rate of convergenceor divergence of the coupled systems toward or away from this manifoldmdashand hence thetendency for the two systems to synchronize or separate For the time series in figure 7 this is
7 Whilst it is of course physiologically unrealistic to assume that two columns would have entirely identical structuresit is mathematically simpler to start analysing the system in this situation8 A qualitative description in a neural system with a different form of coupling (potassium diffusion) is evident infigure 8 of Larter et al (1999) Synchronous chaos in an abstract model of a neural system was also described inWennekers and Pasemann (1996)
Synchronization in a model of neuronal dynamics 713
1
2
2
1
Figure 6 Independent oscillations in a two-column system with identical parameters butindependent initial conditions (a) The time series (b) A plot of concurrent mean excitatorymembrane potentials V 1 against V 2
related to the slope of the plot in panel (c)9 For this strength of coupling the largest transverseexponent is negative (minus002) indicating that perturbations away from this state of identicalsynchronization decay and thus that the state of identical chaotic synchronization is stableagainst noise (ie dynamically stable)
To calculate the relationship between coupling strength and the Lyapunov exponents weused an algorithm which measures the growth of perturbations of the variational equations (thelsquoJacobianrsquo) of the system (we do not define these equations here as they are cumbersome anddo not enhance the understanding of the system) We calculated these exponents for increasingvalues of the coupling parameter C from 0 to 02 in increments of 0000 25 The results are
9 Only two of the six dependent variables are shown in figure 7 The actual transverse Lyapunov exponents reflectthe temporal evolution of all six and hence are somewhat more complex than that given by the slope of panel (c)
714 M Breakspear et al
1
2
Figure 7 Identical synchronization in the same system as in figure 7 but with C = 02 (a) Thetime series (b) A plot of concurrent mean excitatory membrane potentials V 1 against V 2 (c) Thelogarithm of the Euclidean distance between V 1 and V 2 versus time (linear)
Synchronization in a model of neuronal dynamics 715
Figure 8 Lyapunov spectra for a coupled system with δV = 07 and C varying from 0 to 02 insteps of 0000 33 At each value of C all Lyapunov exponents are calculated Least-squares linesof best fit were then fitted to the results for each exponent Solid lines indicate transverse exponents(λ) and dashed lines indicate the tangential exponents () Panel (a) shows all three transverseand three tangential exponents Panel (b) only shows the largest two of each
illustrated in figure 8 Panel (a) shows all exponents For illustrative purposes only the mostpositive four are plotted in panel (b) In both panels the tangential exponents () are fitted (inthe least-squares sense) with a dashed line and the transverse ones (λ) are fitted with a solidline They are labelled 1ndash3 in order of decreasing positivity The important features are asfollows For the range of coupling strengths that we consider the largest tangential Lyapunovexponent is positive which indicates the presence of chaotic behaviour within the manifoldWhen examining the line of best fit for this exponent we see that this exponent does not varywhen altering the coupling parameter This is true for all three tangential exponents and verifiesthat the changes in the coupling parameter do not alter the dynamics on the synchronizationmanifold (that is the coupling parameter is normal)
716 M Breakspear et al
The largest transverse Lyapunov exponent (denoted as λ1 in the figure) is a particularlyimportant exponent since it governs the stability of the attractor in the synchronization manifoldwith respect to perturbations out of the manifold If this exponent is negative then we expectsmall perturbations of trajectories to reconverge to the manifold in finite time If however thisexponent is positive then we would instead expect perturbed trajectories to diverge from themanifold This change in local behaviour is termed a bifurcation and the point at which itoccurs the bifurcation point It is apparent in figure 8 that the zero crossing of this exponentmdashthe bifurcation pointmdashoccurs at approximately C = 01 in the present system This typeof bifurcation has been termed a blowout bifurcation since once transverse stability is losttrajectories are lsquoblown outrsquo from the low-dimensional synchronization manifold into the fullphase space (Ott and Sommerer 1994) Hence in the present setting a blowout bifurcation isobserved if C falls below 01
We note two other observations Increasing rNMDA which corresponds to increasing thepopulation of NMDA receptors decreases the coupling strength required to achieve stablesynchronization On the other hand increasing the overall excitatory-to-excitatory synapticconnectivity (aee) has the opposite effect This is because the effect of local excitatory feedbackincreases the tangential Lyapunov exponents which has the effect of lsquosplittingrsquo the synchronizeddynamics
In the above simulations all the parameters of both systems were equal In realphysiological systems such symmetry between systems is clearly impossible We nowconsider the case in which the two columns have different structures A simple way toincorporate this whilst at the same time preserving (as much as possible) simple dynamics is tovary the parameter δV for each column This has the effect of destroying any symmetry-forcedinvariant manifolds and so the simple case of synchronization corresponding to identicallyevolving dynamics will not exist in this case However more exotic forms of synchronizationare still possible and we discuss these next
42 Generalized chaotic synchronization
In order to maintain consistency with the literature we study non-linear interdependence in thesetting where the systems are asymmetrical but the coupling is unidirectional This is easilyachieved in the present system by making C directed and setting C21 = 0 In this settingwe refer to the lsquofreersquo system as the driver and the lsquoslavedrsquo system as the response We firstask the question what is the effect of introducing a slight parameter mismatch into a systemexhibiting stable chaos synchronization An example of this is presented in figure 9 whereC12 = 04 and parameter mismatch has been incorporated by setting δV = 07 in the driversystem and δV = 0699 in the response system In panel (a) the time series appear qualitativelyvery similar However in panel (b) where the difference between the systems is plotted it isapparent that the separation between the two systems is of the same order of magnitude as thesize of each attractor In panel (c) the effect of the parameter mismatch on the synchronizationmanifold can be seen as a transformation away from the symmetry manifold
It has been proposed that in such a setting there exists a continuous functional relationshipbetween the dynamics of the driver and response systems
x2(t) = ψ(x1(t)) (10)
where xi = V i W i Z i This situation has been referred to in the literature as generalizedsynchronization (Afraimovich et al 1986 Rulkov et al 1995) The properties of ψ andthe topology and stability of the synchronization manifold in this context have not yetbeen fully elucidated although some significant results have been achieved For example
Synchronization in a model of neuronal dynamics 717
1
2
2
1
Figure 9 Generalized synchronization in an asymmetrical driverndashresponse system (a) The timeseries of the two systems (b) The distance between the two systems (c) A plot of concurrent meanexcitatory membrane potentials V 1 against V 2 The diagonal represents the symmetry manifold
718 M Breakspear et al
Hunt et al (1997) Josic (1998) Kocarev et al (2000) have shown that the generalizedsynchronization manifold is smooth (differentiable) if and only if it is normally hyperbolic(ie the largest transverse Lyapunov exponent is less than the smallest tangential Lyapunovexponent prior to the introduction of parameter mismatch) Such a condition is met in thepresent setting when C = 04 hence the smoothness of the manifold in panel (c)
However subsequent to the introduction of parameter mismatch we will not be able touse transverse and tangential Lyapunov exponents to detect such synchronization since thesynchronization manifold may possess a complex topology for which it is a non-trivial (atbest) or impossible (at worse) task to define transverse and tangential directions For examplethe manifold may be smooth as in figure 9(c) but have a fractal topology If the dynamics arenot normally hyperbolic then the manifold may be fractal but also non-differentiable (Huntet al 1997) and hence will not have a unique tangent direction at any point Consequently wewill not be able to compute tangential or transverse Lyapunov exponents and hence determinethe stability of the synchronization manifold However other techniques for detecting thepresence of generalized synchronization in complex systems have evolved including theauxiliary system approach (Abarbanel et al 1996) and non-linear interdependence (Schiffet al 1996) which are now explored
421 The auxiliary system approach The auxiliary system is a third subsystem which isa replica of the original response system but coupled independently to the driver system andcommencing from independent initial conditions It is possible to demonstrate that showingthe existence of generalized synchronizationmdashthat equation (10) is satisfiedmdashis equivalent toshowing that the response system and the auxiliary system evolve identically (even though theyare not directly coupled) To determine whether or not this is the case reduces to an analysisof the Lyapunov exponents transverse to the symmetry manifold with respect to the responsesystem and the auxiliary system
Whilst this is a somewhat artificial test for generalized synchronization since theproduction of an auxiliary system for a physical system would be impossible such a testis useful in numerical and theoretical studies In figure 10 we present an example of theauxiliary system approach where the parameter δV = 07 for column 1 and 068 for column 2We consider two values of the coupling C = 0025 and 01 The auxiliary system is simplya copy of the second column beginning from an arbitrary initial state In the first instancewhere C = 0025 we notice that even after allowing for an initial transient state the secondcolumn and the auxiliary do not behave in identical fashions (figure 10(a)) However whenC = 01 we observe (figure 10(b)) that the second column and the auxiliary do behave innear-identical manners after an initial transient and hence conclude that for the particularvalue of the coupling strength considered equation (10) is satisfied and hence generalizedsynchronization between the first and second columns occurs
422 Nonlinear interdependence The question remains as to how we may detect thepresence of generalized synchronization in a practical setting As we mentioned aboveit is not possible to use an auxiliary-system-based approach to detect the presence ofgeneralized synchronization Instead we must consider techniques for detecting suchbehaviour from experimental or numerical time series data Algorithms of this type areoften referred to as detecting non-linear interdependence Nonlinear interdependence is ageneral term encompassing systems where the behaviour of one non-linear system affectsthe behaviour of another More specifically it can be defined as occurring if the nullhypothesis that the interdependence between two signals is purely linear can be rejectedat conventional levels (usually 95) of statistical significance hence it is an experimental
Synchronization in a model of neuronal dynamics 719
2
1
2
1
Figure 10 Concurrent evolution of the response system V 2 and lsquoauxiliaryrsquo system V 1 (a) C =0025 (b) C = 01
term (eg Schiff et al 1996) Whilst this does not strictly correspond to the existence of afunctional relationship between the two systems such a relationship is typically used tomotivate the techniques for its detection Schiff et al (1996) extended the logic of non-linearprediction algorithms from autonomous to coupled systems by exploiting the continuity ofsuch a relationship and hence devised the first method of detecting non-linear interdependencein time series data The basis of the technique is using the forward evolution of a phase spacevectorrsquos neighbours to predict the forward evolution of that vector and the forward evolutionof the concurrent vector of the other (possibly interdependent) system If the error of such aprocedure is better on average than chance (the chance error is normalized to equal one) thennon-linear interdependence is said to be present
720 M Breakspear et al
V1
V2
Figure 11 Experimental and numerical examples of weak generalized non-linear synchronizationin the presence of measurement noise (a) Human scalp EEG data from posterior bipolar scalpelectrodes (O1P3ndashO2P4) (b) Weakly coupled aperiodic data in the present model with additivewhite noise (with 5 of the amplitude of the model signal) Parameters δ1
V = 0605 δ2V = 061
aee = 036 C = 007
A variation of this algorithm optimized to detect weak non-linear interdependence (Terryand Breakspear 2003) was used recently to demonstrate that weak non-linear interdependence isoccasionally present in the scalp EEG of healthy resting human subjects (Breakspear and Terry2002a 2002b 2002c) An example of an epoch of lsquonon-linearly independentrsquo EEG obtainedusing this technique is presented in figure 11(a) For comparison output from the currentmodel is presented in figure 11(b) The model parameters were set such that the dynamicswas still aperiodic but the intermittent large-amplitude bursts were suppressed and had weakmutual coupling (C = 007) Extraneous noise to simulate the effect of extracranial muscleactivity contaminating the EEG data has been added to this time series It should be notedthat the amplitudes and time scales of the modelrsquos variables are dimensionless Nonethelessthe model facilitates qualititative interpretation of experimental findings
Synchronization in a model of neuronal dynamics 721
43 Chaotic phase synchronization
Whereas the concept of generalized synchronization was mathematically motivated and assuch more suited to theoretical investigations of systems the concept of phase synchronizationderives from a more practical signal processing approach Phase synchronization occurs inboth symmetric and asymmetric settings but is often more useful in the asymmetric case (andin experimental data)
Unfortunately the phase of a chaotic signal is typically ambiguous and can be defined in anumber of different ways For example in the case of the coherent Rossler attractor (Osipovet al 1997) the phase may be defined as the arctangent of two of the variables of the systemMore generally it can sometimes be defined as the angle that an orbit subtends with the centreof an attractor It is also possible to extract the phase of a chaotic signal from time seriesdata by use of the Hilbert transform This has enabled the demonstration of chaotic phasesynchronization in a number of coupled asymmetrical systems (Rosenblum et al 1996 1997Zaks et al 1999) Although such an approach does not guarantee that the phase will be uniqueit does permit the practical analysis of phase in noisy time series (Tass et al 1998)
To illustrate phase synchronization in two coupled columns we again vary the parameterδV which if the columns are viewed as oscillators acts to detune them from a commonfrequency An important question is that of the criteria on the basis of which we concludewhether phase synchronization occurs We use a simple locking criterion whereby phasesynchronization is said to occur if
|θ1(t)minus θ2(t)| lt K (11)
where is θi the phase of column i and K is a constant Notice that this does not mean thatsynchronization implies equality of phases merely that their difference is bounded
An example of phase synchronization with δV = 065 for column 1 and 066 for column 2is given in figure 12 In this figure rNMDA = 03 Qualitatively some sort of phase coherenceis evident in the time series panel (a) This was confirmed by plotting the sine of thephase difference which has been extracted using the Hilbert transform panel (b) Clearly|sin(θ1 minus θ2)| lt K and hence |θ1 minus θ2| is bounded It can also be seen that the phase-synchronized dynamics occurs on a highly structured manifold depicted in figure 12(c)However unlike the case for the generalized synchronization manifold depicted in figure 9 itis evident that this manifold is not a simple transformation of the symmetry manifold
It is also possible to see lsquobimodalrsquo stability whereby a system jumps betweenapproximately zero phase difference and a phase lag phase synchronization of π An exampleis depicted in figure 12(d) At approximately 2000 s the system can be seen to jumpspontaneously from the first to the second of these modes The Z -variable is plotted forgraphical purposes
5 Intermittency and desynchronization
51 Desynchronization in an array of two systems
Diminishing the strength of coupling across the blowout bifurcation does not lead to a simpletransition from synchronous to asynchronous oscillations but is instead associated withcomplex temporal behaviours Specifically when the largest transverse Lyapunov exponent isclose to zero the coupled systems exhibit epochs of synchronous behaviours punctuated bysharp bursts of desynchronization This is observed in systems exhibiting identicalgeneralizedand phase synchronization An example of such a phenomenon is given in figure 13 The timeseries panel (a) shows synchronous dynamics except at time = 1000 where it can be seenthat the two time series desynchronize briefly This is evident in panel (b) as a disruption in the
722 M Breakspear et al
2
1
Figure 12 Phase-synchronized coupled columns (a) The time series of V 1 (solid) and V 2 (b) Thesine of the phase difference obtained through Hilbert transformation of the time series in panel (a)(c) The phase synchronization manifold in the V 1ndashV 2 plane (d) The time series of Z (solid) andZ showing a phase slip of π between two modes
Synchronization in a model of neuronal dynamics 723
Figure 13 An example of phase desynchronization in coupled asymmetric systems with C = 01Time series plots of (a) each V -variable and (b) their Euclidean distance (c) The temporal evolutionof the phase difference between the two systems (d) An example of the phase difference evolutionover a longer time interval
724 M Breakspear et al
orderly evolution of the distance between the two systems and in panel (c) where the evolutionof the phase difference reveals a brief violation of the criterion |φ1 minus φ2| lt constant Anexample of the phase difference evolution over a longer time interval is shown in figure 13(d)Note that this is a three-dimensional plot with sin(φ1 minus φ2) and cos(φ1 minus φ2) as they- and z-axes Hence phase desynchronizations are evident as complete revolutions aroundthe cylindrical phase space in which this variable evolves It can be seen that these phenomenaoccur at irregular intervals in the temporal domain
This behaviour can be explained by considering the nature of a chaotic attractor In thestatistical sense an arbitrary initial condition is expected to lead to the chaotic behaviour onthe attractor Consequently when we calculate the Lyapunov exponents of the system theseexponents correspond to those of the chaotic attractor Typically however there are an infinitenumber of unstable periodic orbits embedded within the attractor If the path of a trajectorybecomes close to one of these orbits it may lsquoshadowrsquo the orbit for a brief period of time Eachof these orbits has a corresponding set of Lyapunov exponents which (like the attractor) can bedivided into tangential and transverse exponents These also vary when we change the couplingparameter but are typically not the same as the exponents of the attractor Hence the largesttransverse exponent of the attractor may be negative yet there may be positive transverseexponents associated with periodic orbits Considering all of these exponents together wewould therefore only expect to observe complete stability of the synchronization manifold inthe case where all of the transverse Lyapunov exponents are negative Bursting behaviourwill be observed when the one of these exponents becomes positive typically correspondingto a periodic orbit of low order (Pikovsky and Grassberger 1991 Rulkov and Sushchik 1997Maistrenko et al 1998 Heagy et al 1998) Due to the complex structure of such periodic orbitsthe desynchronous bursts occur across a wide range of timescales
In figure 14 we present the results of a long integration (16times105 time units) in which thelengths of sequential synchronous epochs of phase synchronization are studied The parametervalues were δV = 065 for column 1 and 0645 for column 2 with coupling strength10 equal to0125 In panel (a) the power spectrum of these sequential lengths is plotted The whiteness ofthis spectrum indicates that the lengths of consecutive synchronous epochs are independentThis is due to the high-dimensional lsquomixingrsquo effect of desynchronization In panel (b) thelengths of the synchronous epochs have been sorted again into the order of increasing lengthThe results for two coupling strengths are plotted C = 0125 (solid) and 0135 (dashed)Several features of this plot are of note Firstly the lengths of the synchronous epochsare distributed over almost three orders of magnitude Secondly for both plots there is anapproximately log-linear relationship between the length of an epoch and its relative orderThis reflects an approximately exponential relationship between the length of a synchronousepoch and its likelihood of occurrence Thirdly this relationship is skewed toward longersynchronous epochs for the stronger coupling strengths
This type of signal with patches of lsquoorderrsquo occurring over many timescales interruptedby bursts of disorder can be shown to have a high information capacity (Gatlin 1972)Moreover complex phase desynchronization may make an infrequent yet robust contributionto the dynamical pattern of the alpha rhythm in scalp EEG data for resting human subjects(Breakspear 2002) It is tempting to postulate that such a lsquoweakly stablersquo dynamics may assistin the generation of the rich and fluid mental associations that spontaneously emerge in restingcognitive states Although this phenomenon has been previously described in a variety of othersystems to our knowledge this is the first description in a biophysical model of neocortex
10 The introduction of parameter mismatch causes an increase in the strength of coupling required for stablesynchronization above the value of 01 required in the symmetric case
Synchronization in a model of neuronal dynamics 725
Figure 14 Properties of sequential epochs of phase-synchronous behaviour over a long integrationtime δ1
V = 065 δ2V = 0645 aee = 04 (a) The flat (white) power spectrum (b) Synchronous
epochs sorted in order of increasing length (solid C = 0125 dashed C = 0134)
52 Synchronization and complex dynamics in larger arrays
The synchronous and complex phenomena described above are also observed in arrays ofcoupled systems with N gt 2 Expanding the size of the array permits complex dynamicsin the spatial in addition to the temporal domain However the basic principles governingthe type and stability of synchronized motions are the same In this section a few of thepossible behaviours are discussed Physiological variation across the array is modelled bymodifying the parameter δV = 07 in each array by adding a random number of maximummagnitude 001
In figure 15(a) an array of N = 20 uncoupled (C = 0) systems is shown The lack ofsynchronization in this time series is evident By comparison the rapid transition to completesynchronization is evident in figure 15(b) when C = 03 In fact 19 of the 20 subsystems
726 M Breakspear et al
(b)
(c)
(a)
Figure 15 Synchronization phenomena in an array of coupled cortical columns (a) N = 20C = 0 (b) N = 20 C = 03 (c) N = 6 C = 01 (d) N = 12 C = 009 (e) N = 12C = 012 A small perturbation has been applied to one of the columns at t = 2500 s (f) Evolutionof the phase difference between two of the columns in panel (e) For clarity the W -variable isplotted in panel (c) and the Z -variable in (e)
have phase synchronized within one single cycle Intermediate values of coupling strength areassociated with a rich variety of phenomena For example subsystems with the most similarparameter values typically synchronize into clusters of various sizes An example of this isillustrated in figure 15(c) Note that there is no synchronization between the two differentclusters each containing three synchronized subsystems
Another complex behaviour occurs when subsystems transiently synchronize into largeclusters then spontaneously desynchronize again Hence the dynamics of the entire arrayjumps between different transient dynamics characterized by varying degrees of freedom Anexample of this phenomenon is given in figure 15(d) In addition it is possible to inducedesynchronization across an entire array by applying a small perturbation to one columnfor one time step An example is illustrated in figure 15(e) A small perturbation has been
Synchronization in a model of neuronal dynamics 727
Figure 15 (Continued)
added to one column after 2500 time units It can be seen that the entire array subsequentlydesynchronizes This illustrates the sensitivity of the dynamics to external stimuli Finally it ispossible to lsquoinducersquo synchronization between selected subsystems by increasing the strength ofcoupling between them selectively This allows an extra level of complexity to be introducedinto the model For example it may allow targeted information transfer between a partialnetwork of the entire array
As discussed in section 51 a univariate signal which jumps between epochs of lowentropy (high repetition) and bursts of high entropy (high variability) can be shown to havehigh information capacity This is because the repetition of certain patterns (as in the English
728 M Breakspear et al
language) permits error detectability Bursts of high entropy act in the opposite directionto increase message variability With a multivariate signal in addition to optimizing theinformation capacity within each signal the overall information capacity can be increasedthrough the type of itinerant synchronization briefly illustrated in this section Synchronizationacross an entire array increases the message reliability (since a single lsquoerrant letterrsquo in onesignal can be easily detected) Conversely desynchronization into multiple clusters increasesmessage variability since each cluster will produce independent information Hence a neuralarray which switches dynamically between a range of cluster configurations can be seen asadaptive in the sense that high synchronous states can be expressed when error detectabilityis a priority (such as during quick and reliable execution of a familiar task) but the array canrapidly desynchronize if variability becomes salient (for example if the environment suddenlybecomes novel) A formal operationalized demonstration of this principle is to be the subjectof future work
6 Discussion
In this paper we present a dynamical model for activity in an array of weakly coupledlocal neural assemblies The model incorporates the fundamental principles governing cellmembrane potential as determined by voltage- and ligand-gated ion channels In this wayit allows a reasonable amount of physiological complexity to be incorporated includingsynaptic activation and receptor subtypeswithout recourse to overwhelming detail Numericalanalysis of this model reveals a transition from independent oscillations to stable chaoticsynchronization depending on the balance of local versus long-range connectivity andbiophysical heterogeneity These excitatory couplings are subject to ascending subcorticalneuromodulation arising principally from monoaminergic nuclei such as Locus Coeruleus(Silberstein 1995) Neuromodulation of extrinsic connections was incorporated into the modelvia a coupling parameter C The stability of the synchronization is determined by the valueof the largest transverse Lyapunov exponent which in turn depends on the coupling parameterC If the coupling is weakened with the result that this exponent becomes positive a blowoutbifurcation occurs and independent oscillations re-emerge In the vicinity of the transitionnon-linear desynchronizations with complex temporal and spatial properties are observed
It is pleasing that complex and itinerant behaviour reminiscent of real neuronaldynamics emerges through such a simple manipulation of a single dynamic control parameterFurthermore the ubiquitous projections of classical neuromodulatory systems in real brainssuggest a clear biological mechanism by which the brain could self-organize throughmoderating the complexity of its own dynamics There are many reasons that the complicateddynamics of the sort demonstrated in this paper may be neurobiologically important Wemention four
First the presence of multiple clusters of phase-synchronized subsystems as discussedabove may be a uniquely non-linear mechanism facilitating the lsquocarriagersquo and distinction ofmultiple concurrent dynamic cell assemblies (and hence cognitive processing) This may beimportant for perceptual grouping binding and transitions as suggested by others (eg von derMalsburg 1985 Gray and Singer 1989)
Second the facility to decouple and reconfigure dynamic cell assemblies througha neuromodulatory control parameter may be important for avoiding low-entropysynchronization manifolds In the absence of itinerancy synchronization would renderneuronal dynamics incapable of representing changes in sensory input or indeed cognitive stateFor example in the model it was observed that small perturbations to one column were ableto induce a global change in neuronal dynamics by disrupting a state of weak synchronization
Synchronization in a model of neuronal dynamics 729
Inevitably alternative dynamic configurations emerge epitomizing the importance of lsquolabilersquobrain dynamics (Friston 2000a)
Thirdly from a purely functionalist point of view the balance between functionalspecialization and functional integration may rely upon the sorts of mechanism modelledin this paper The brain appears to adhere to two principles of functional organization namelysegregation of function within cortical areas and sub-areas and the integration of these areasin perceptual synthesis cognition and sensorimotor integration (Tononi et al 1994 Fristonet al 1995) The balance between preserving specialized dynamics unique to each arearequired by specialization and the coupling among these areas mediating functional integrationmay be achieved by the neuromodulatory mechanisms described above Significantlythe spontaneous variation in the numbers of degrees of freedom (for example whenlow-dimensional synchronization manifolds are shadowed) imparts a dynamic characteristicto this perspective that is precluded by linear mechanisms alone
Finally from a mechanistic perspective complexity and diversity of brain dynamics arerequired for selection of adaptive responses (Friston 2000b) There are many examples ofselective mechanisms operating within the brain All can be seen as performing an ascenton some objective function or fitness landscape These mechanisms require complex anditinerant dynamics to facilitate a thorough exploration of the landscape These landscapesmay range from those involved in perception (where the landscape is defined by predictionerror see eg Rao and Ballard (1998)) to those involved in value learning (where thelandscape is determined by the outcome of a behaviour eg in temporal difference models (seeeg Sutton and Barto 1990)) In short complexity is necessary to maintain an information-rich environment in which the brain can respond to sensory inputs An essential componentof this is the maintenance of diversity which is a prerequisite for selectionist mechanismsat many scales (Edelman 1993) Understanding the mechanistic basis of complex dynamicsas measured electrophysiologically may well depend on the sort of biologically plausiblemodels used in this paper This understanding may be important for both basic and clinicalneuroscience (eg Tass 2002)
The present study supplements and advances on other research in this field The studyof complex phase synchronization and clustering in neural oscillator networks similar tothe present model has previously been the subject of formal mathematical study Forexample Terman and Wang (1995) studied synchronization of blocks of neural oscillatorsthat were subject to perturbation by connected lsquostimulirsquo Somers and Kopell (1995) studiedthe emergence of both synchronization and travelling waves in networks of coupled neuraloscillators In the work of Terman and Wang the local dynamics were based on neurallsquocaricaturersquo models such as the FitzHughndashNagumo equations and are hence more abstract thanthe current study The approach of Somers and Kopell only required that the local dynamicsexhibit fast and relaxation phases and is hence more abstract still In both studies onlyperiodic dynamics are studied Complex patterns of chaos synchronization in neural arrayshave been characterized by nonlinear methods (Kaneko 1998) and their putative contributionto both learning (Nakajima and Ueda 1996) and perception (Zhao and Macau 2001 Breakspear2001) explored However these studies employed models such as the logistic equation andtent map which are quite abstract in comparison to the present study On the other handHansel and Sompolinsky (1996) studied the role of chaos synchronization in a model ofvisual perception in a cortical hypercolumn using a very similar approach to that adoptedhere but with the additional complexity that synaptic delays were incorporated However inthis and earlier work (Hansel and Sompolinsky 1992) only linear measures of dynamics andsynchrony (specifically the auto- and cross-correlation functions) were employed The analysisof chaotic synchrony in the study of Larter et al (1999) was based only upon observation of
730 M Breakspear et al
the Euclidean difference between variables from two nodes within a larger array Hence thesignificance of the present work is that it provides a quantitative nonlinear analysis of complexchaos synchronization in a biophysical model of cortical dynamics Such research is requiredin order to bridge the gap between quantitative nonlinear research into synchronization andcomputation in abstract models of neural arrays and qualitative or linear analysis in morebiophysically plausible models
Acknowledgments
The authors would like to thank Peter Robinson for assistance with the manuscript and R Larterfor helpful advice MB is a recipient of a NSW Institute of Psychiatry Research Grant anda University of Sydney SESQUI postdoctoral fellowship KJF is supported by the WellcomeTrust
References
Abarbanel H Rulkov N and Sushchik M 1996 Generalised synchronization of chaos the auxiliary system approachPhys Rev E 53 4528ndash35
Afraimovich V Verichev N and Rabinovich M 1986 Stochastic synchronization of oscillation in dissipative systemsRadiophys Quantum Electron 29 795ndash801
Breakspear M 2001 Perception of odours by a nonlinear model of the olfactory bulb Int J Neural Syst 11 101ndash24Breakspear M 2002 Nonlinear phase desynchronisation in human electroencephalographic data Human Brain Mapping
15 175ndash98Breakspear M and Terry J R 2002a Nonlinear interdependence in neural systems motivation theory and relevance
Int J Neurosci 112 1163ndash84Breakspear M and Terry J R 2002b Detection and description of nonlinear interdependence in normal multichannel
human EEG Clin Neurophysiol 113 735ndash53Breakspear M and Terry J R 2002c Topographic organization of nonlinear interdependence in multichannel human
EEG Neuroimage 16 822ndash35Destexhe A Rudolph M Fellous J M and Sejnowski T J 2001 Fluctuating synaptic conductances recreate in vivo-like
activity in neocortical neurons Neuroscience 107 13ndash24Douglas R J Mahowald M A and Martin K A 1996 Microarchitecture of neocortical columns Brain TheorymdashBiological
Basis and Computational Principles ed A Aertsen and V Braitenberg (Amsterdam Elsevier)Eckhorn R Bauer R Jordan W Brosch M Kruse W Munk M and Reitboeck H J 1988 Coherent oscillations a
mechanism of feature linking in the visual cortex Multiple electrode and correlation analyses in the cat BiolCybern 60 121ndash30
Edelman G M 1993 Neural Darwinism selection and reentrant signalling in higher brain function Neuron 10 115ndash25Engel A K Kreiter A K Konig P and Singer W 1991 Synchronization of oscillatory neuronal responses between
striate and extrastriate visual cortical areas of the cat Proc Natl Acad Sci USA 88 6048ndash52Friston K J 2000a The labile brain I Neuronal transients and nonlinear coupling Phil Trans R Soc B 355 215ndash36Friston K J 2000b The labile brain II Transients complexity and selection Phil Trans R Soc B 355 237ndash52Friston K J Tononi G Sporns O and Edelman G 1995 Characterising the complexity of neuronal interactions Human
Brain Mapping 3 302ndash14Fujisaka H and Yamada T 1983 Stability theory of synchronized motion in coupled-oscillator system Prog Theor
Phys 69 32ndash47Gatlin L 1972 Information Theory and the Living System (New York Columbia University Press)Gray C M Konig P Engel A and Singer W 1989 Oscillatory responses in cat visual cortex exhibit inter-columnar
synchronization which reflects global stimulus properties Science 338 334ndash7Gray C M and Singer W 1989 Stimulus specific neuronal oscillations in orientation columns of cat visual cortex Proc
Natl Acad Sci USA 86 1698ndash702Haig A R Gordon E Wright J J Meares R A and Bahramali H 2000 Synchronous cortical gamma-band activity in
task-relevant cognition Neuroreport 11 669ndash75Hansel D and Sompolinsky H 1992 Synchronization and computation in a chaotic neural network Phys Rev Lett 68
718ndash21
Synchronization in a model of neuronal dynamics 731
Hansel D and Sompolinsky H 1996 Chaos and synchrony in a model of a hypercolumn in visual cortex J ComputNeurosci 3 7ndash4
Heagy J Carroll T and Pecora L 1998 Desynchronization by periodic orbits Phys Rev E 57 R1253ndash6Hunt B Ott E and Yorke J 1997 Differentiable synchronization of chaos Phys Rev E 55 4029ndash34Josic K 1998 Invariant manifolds and synchronization of coupled dynamical systems Phys Rev Lett 80 3053ndash6Kandel E Schwartz J and Jessell T 2000 Principles of Neural Science (New York McGraw-Hill)Kaneko K 1998 On the strength of attractors in a high-dimensional system Physica D 124 322ndash44Kocarev L Parlitz U and Brown R 2000 Robust synchronization of chaotic systems Phys Rev E 61 3716ndash20Larter R Speelman B and Worth R M 1999 A coupled ordinary differential equation lattice model for the simulation
of epileptic seizures Chaos 9 795ndash804Lumer E Edelman G and Tononi G 1997a Neural dynamics in a model of the thalamocortical system I Layers loops
and the emergence of fast synchronous oscillations Cereb Cortex 7 207ndash27Lumer E Edelman G and Tononi G 1997b Neural dynamics in a model of the thalamocortical system II The role of
neural synchrony tested through perturbations of spike timing Cereb Cortex 7 228ndash36Magleby K 1987 Short-term modulation at synapses Synaptic Function ed G M Edelman W E Gall and
W M Cowan (New York Wiley)Maistrenko Y Maistrenko V Popovich A and Mosekilde E 1998 Transverse instability and riddled basins in a system
of two coupled logistic maps Phys Rev E 57 2713ndash24Morris C and Lecar H 1981 Voltage oscillations in the barnacle giant muscle fiber Biophys J 35 193ndash213Mountcastle V B 1997 The columnar organization of the neocortex Brain 120 701ndash22Mountcastle V B 1998 The Cerebral Cortex (Cambridge MA Harvard University Press)Nakajima H and Ueda Y 1996 Riddled basins of the optimal states in learning dynamical systems Physica D 99 35ndash44Nunez P L 1995 Neocortical Dynamics and Human Brain Rhythms (Oxford Oxford University Press)Osipov G V Pikovsky A S Rosenblum M G and Kurths J 1997 Phase synchronization effects in a lattice of nonidentical
Rossler oscillators Phys Rev E 55 2353ndash61Ott E and Sommerer J 1994 Blowout bifurcations the occurrence of riddled basins and onndashoff intermittency Phys
Lett A 188 39ndash47Pecora L and Carroll T 1990 Synchronization in chaotic systems Phys Rev Lett 64 821ndash4Pikovsky A and Grassberger P 1991 Symmetry breaking bifurcation for coupled chaotic attractors J Phys A Math
Gen 24 4587ndash97Rao R P and Ballard D H 1998 Predictive coding in the visual cortex a functional interpretation of some extra-classical
receptive field effects Nature Neurosci 2 79ndash87Robinson P A Wright J J and Rennie C R 1998 Synchronous oscillations in the cerebral cortex Phys Rev E 57
4578ndash88Rodriguez E George N Lachauz J P Martinerie J Renault B and Varela F J 1999 Perceptionrsquos shadow long distance
synchronization of human brain activity Nature 397 430ndash3Rosenblum M Pikovsky A and Kurths J 1996 Phase synchronization of chaotic oscillators Phys Rev Lett 76 1804ndash7Rosenblum M Pikovsky A and Kurths J 1997 From phase to lag synchronization in coupled chaotic oscillators Phys
Rev Lett 78 4193ndash6Rulkov N and Sushchik M 1997 Robustness of synchronized chaotic oscillations Int J Bifurc Chaos 7 625ndash43Rulkov N Sushchik M Tsimring L and Abarbanel H 1995 Generalized synchronization of chaos in unidirectionally
coupled chaotic systems Phys Rev E 51 980ndash94Schiff S So P Chang T Burke R and Sauer T 1996 Detecting dynamical interdependence and generalized synchrony
through mutual prediction in a neural ensemble Phys Rev E 54 6708ndash24Silberstein R B 1995 Neuromodulations of neocortical dynamics Neocortical Dynamics and Human EEG Rhythms
ed P L Nunez (Oxford Oxford University Press)Singer W 1995 Putative functions of temporal correlations in neocortical processing Large-Scale Neuronal Theories
of the Brain ed C Koch and J Davis (London MIT Press)Somers D and Kopell N 1995 Waves and synchrony in networks of oscillators of relaxation and non-relaxation type
Physica D 89 169ndash83Sutton R S and Barto A G 1990 Time derivative models of Pavlovian reinforcement Learning and Computational
Neuroscience Foundations of Adaptive Networks ed M Gabriel and J Moore (Cambridge MA MIT Press) pp497ndash538
Szentagothai J 1983 The modular architectonic principle of neural centers Rev Physiol Biochem Pharmacol 9811ndash61
Tallon-Baudry C Betrand O and Fischer C 2001 Oscillatory synchrony between human extrastriate areas during visualshort-term memory maintenance J Neurosci 21 RC177(1ndash5)
Tass P A 2002 Desynchronization of brain rhythms with soft phase-resetting techniques Biol Cybern 87 102ndash15
732 M Breakspear et al
Tass P Rosenblum M Weule J Kurths J Pikovsky A Volkmann J Schnitzler A and Freund H 1998 Detection of nmphase locking from noisy data application to magnetoencephalography Phys Rev Lett 81 3291ndash4
Terman D and Wang D 1995 Global competition and local cooperation in a network of neural oscillators Physica D81 148ndash76
Terry J and Breakspear M 2003 An improved algorithm for the detection of dynamical interdependence in bivariatetime-series Biol Cybern 88 129ndash36
Tononi G Sporns O and Edelman G M 1994 A measure for brain complexity relating functional segregation andintegration in the nervous system Proc Natl Acad Sci USA 91 5033ndash7
von der Malsburg C 1985 Nervous structures with dynamical links Phys Chem 89 703ndash10Wang X 1991 Period-doublings to chaos in a simple neural network an analytic proof Complex Syst 5 425ndash41Wennekers T and Pasemann F 1996 Synchronous chaos in high dimensional modular neural networks Int J Bifurc
Chaos 6 2055ndash67Zaks M A Parks E H Rosenblum M G and Kurths J 1999 Alternative locking ratios in imperfect phase synchronization
Phys Rev Lett 82 4228ndash31Zhao L and Macau E N 2001 A network of dynamically coupled chaotic maps for scene segmentation IEEE Trans
Neural Netw 12 1375ndash85
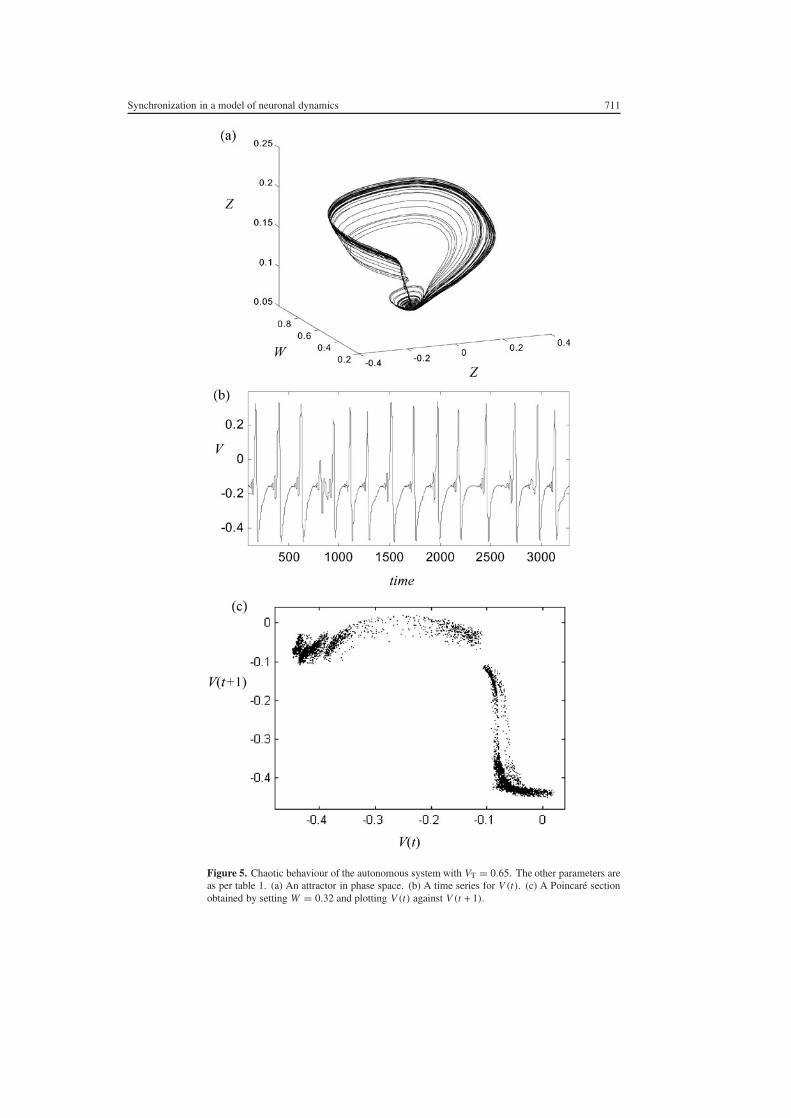
Synchronization in a model of neuronal dynamics 711
1
Figure 5 Chaotic behaviour of the autonomous system with VT = 065 The other parameters areas per table 1 (a) An attractor in phase space (b) A time series for V (t) (c) A Poincare sectionobtained by setting W = 032 and plotting V (t) against V (t + 1)
712 M Breakspear et al
we study the emergence of cooperative behaviour that results from the introduction of couplingbetween columns Three types of interdependent behaviour of two systems are describedidentical generalized and phase synchronization Synchronous behaviours in larger arrays arealso illustrated
41 Identical chaotic synchronization
411 Transition to identical chaotic synchronization The most straightforward extensionand the one that we consider first is that of two coupled cortical columns where all parametersare in the first case assumed identical7 Trivially the two systems will evolve identicallywhen started with identical initial conditions even if C = 0 This is simply a statement ofthe deterministic nature of the evolution equations In general the random initial conditionswill not be equal in which case it is hardly surprising that the different columns will evolveindependently An example is presented in figure 6 with VT = 06 in both systems C = 0and independent random initial conditions In figure 6(a) we show a segment of the timeseries for each of the two subsystems where it can be seen that the time series are chaoticand asynchronous In figure 6(b) the concurrent values of V 1 and V 2 are plotted against eachother showing that they bear no obvious interrelationship
With the introduction of coupling C gt 0 the systems begin to show epochs ofsynchronized behaviour At C sim= 01 there is a transition to stable fully synchronizedmotion as evident in an exemplar time series shown in figure 7(a) where C = 015 Infigure 7(b) we present the concurrent values of V 1 versus V 2 illustrating that in contrast tothe case in figure 6(b) the systems have become embedded in the low-dimensional lsquosymmetrymanifoldrsquo V 1 = V 2 Panel (c) shows a log-linear plot of the temporal evolution of the distanceof the coupled system from this manifold It can be seen that on average the two systemsapproach at an exponential rate with constant time-averaged exponent Whilst this phenomenon(lsquoidentical chaotic synchronizationrsquo) has been described in a variety of other systems since itsfirst description (Fujisaka and Yamada 1983 Pecora and Carroll 1990) to our knowledge thisis its first quantitative8 description in a detailed physiological model of a neural system
The symmetry of the dynamical system obtained by setting the state parameters in eachcolumn identical forces the invariance of the symmetry manifold V 1 = V 2 Moreover thedynamics within this manifold is governed by the same equations of motion as for a singlecortical column This manifold which contains the orbits of the synchronized system is calledthe synchronization manifold The coupling parameter C does not affect the dynamics withinthe synchronization manifold and as such is termed a normal parameter
412 lsquoBlowout bifurcationrsquo An important issue is the stability of the orbits on thesynchronization manifold This can be determined by analysing the Lyapunov spectraBecause the two-column system evolves in a six-dimensional phase space there are sixLyapunov exponents However for the synchronized system it is helpful to subdivide theseinto those that describe the dynamics within the synchronization manifold and those thatdescribe the dynamics in the transverse direction The latter describe the rate of convergenceor divergence of the coupled systems toward or away from this manifoldmdashand hence thetendency for the two systems to synchronize or separate For the time series in figure 7 this is
7 Whilst it is of course physiologically unrealistic to assume that two columns would have entirely identical structuresit is mathematically simpler to start analysing the system in this situation8 A qualitative description in a neural system with a different form of coupling (potassium diffusion) is evident infigure 8 of Larter et al (1999) Synchronous chaos in an abstract model of a neural system was also described inWennekers and Pasemann (1996)
Synchronization in a model of neuronal dynamics 713
1
2
2
1
Figure 6 Independent oscillations in a two-column system with identical parameters butindependent initial conditions (a) The time series (b) A plot of concurrent mean excitatorymembrane potentials V 1 against V 2
related to the slope of the plot in panel (c)9 For this strength of coupling the largest transverseexponent is negative (minus002) indicating that perturbations away from this state of identicalsynchronization decay and thus that the state of identical chaotic synchronization is stableagainst noise (ie dynamically stable)
To calculate the relationship between coupling strength and the Lyapunov exponents weused an algorithm which measures the growth of perturbations of the variational equations (thelsquoJacobianrsquo) of the system (we do not define these equations here as they are cumbersome anddo not enhance the understanding of the system) We calculated these exponents for increasingvalues of the coupling parameter C from 0 to 02 in increments of 0000 25 The results are
9 Only two of the six dependent variables are shown in figure 7 The actual transverse Lyapunov exponents reflectthe temporal evolution of all six and hence are somewhat more complex than that given by the slope of panel (c)
714 M Breakspear et al
1
2
Figure 7 Identical synchronization in the same system as in figure 7 but with C = 02 (a) Thetime series (b) A plot of concurrent mean excitatory membrane potentials V 1 against V 2 (c) Thelogarithm of the Euclidean distance between V 1 and V 2 versus time (linear)
Synchronization in a model of neuronal dynamics 715
Figure 8 Lyapunov spectra for a coupled system with δV = 07 and C varying from 0 to 02 insteps of 0000 33 At each value of C all Lyapunov exponents are calculated Least-squares linesof best fit were then fitted to the results for each exponent Solid lines indicate transverse exponents(λ) and dashed lines indicate the tangential exponents () Panel (a) shows all three transverseand three tangential exponents Panel (b) only shows the largest two of each
illustrated in figure 8 Panel (a) shows all exponents For illustrative purposes only the mostpositive four are plotted in panel (b) In both panels the tangential exponents () are fitted (inthe least-squares sense) with a dashed line and the transverse ones (λ) are fitted with a solidline They are labelled 1ndash3 in order of decreasing positivity The important features are asfollows For the range of coupling strengths that we consider the largest tangential Lyapunovexponent is positive which indicates the presence of chaotic behaviour within the manifoldWhen examining the line of best fit for this exponent we see that this exponent does not varywhen altering the coupling parameter This is true for all three tangential exponents and verifiesthat the changes in the coupling parameter do not alter the dynamics on the synchronizationmanifold (that is the coupling parameter is normal)
716 M Breakspear et al
The largest transverse Lyapunov exponent (denoted as λ1 in the figure) is a particularlyimportant exponent since it governs the stability of the attractor in the synchronization manifoldwith respect to perturbations out of the manifold If this exponent is negative then we expectsmall perturbations of trajectories to reconverge to the manifold in finite time If however thisexponent is positive then we would instead expect perturbed trajectories to diverge from themanifold This change in local behaviour is termed a bifurcation and the point at which itoccurs the bifurcation point It is apparent in figure 8 that the zero crossing of this exponentmdashthe bifurcation pointmdashoccurs at approximately C = 01 in the present system This typeof bifurcation has been termed a blowout bifurcation since once transverse stability is losttrajectories are lsquoblown outrsquo from the low-dimensional synchronization manifold into the fullphase space (Ott and Sommerer 1994) Hence in the present setting a blowout bifurcation isobserved if C falls below 01
We note two other observations Increasing rNMDA which corresponds to increasing thepopulation of NMDA receptors decreases the coupling strength required to achieve stablesynchronization On the other hand increasing the overall excitatory-to-excitatory synapticconnectivity (aee) has the opposite effect This is because the effect of local excitatory feedbackincreases the tangential Lyapunov exponents which has the effect of lsquosplittingrsquo the synchronizeddynamics
In the above simulations all the parameters of both systems were equal In realphysiological systems such symmetry between systems is clearly impossible We nowconsider the case in which the two columns have different structures A simple way toincorporate this whilst at the same time preserving (as much as possible) simple dynamics is tovary the parameter δV for each column This has the effect of destroying any symmetry-forcedinvariant manifolds and so the simple case of synchronization corresponding to identicallyevolving dynamics will not exist in this case However more exotic forms of synchronizationare still possible and we discuss these next
42 Generalized chaotic synchronization
In order to maintain consistency with the literature we study non-linear interdependence in thesetting where the systems are asymmetrical but the coupling is unidirectional This is easilyachieved in the present system by making C directed and setting C21 = 0 In this settingwe refer to the lsquofreersquo system as the driver and the lsquoslavedrsquo system as the response We firstask the question what is the effect of introducing a slight parameter mismatch into a systemexhibiting stable chaos synchronization An example of this is presented in figure 9 whereC12 = 04 and parameter mismatch has been incorporated by setting δV = 07 in the driversystem and δV = 0699 in the response system In panel (a) the time series appear qualitativelyvery similar However in panel (b) where the difference between the systems is plotted it isapparent that the separation between the two systems is of the same order of magnitude as thesize of each attractor In panel (c) the effect of the parameter mismatch on the synchronizationmanifold can be seen as a transformation away from the symmetry manifold
It has been proposed that in such a setting there exists a continuous functional relationshipbetween the dynamics of the driver and response systems
x2(t) = ψ(x1(t)) (10)
where xi = V i W i Z i This situation has been referred to in the literature as generalizedsynchronization (Afraimovich et al 1986 Rulkov et al 1995) The properties of ψ andthe topology and stability of the synchronization manifold in this context have not yetbeen fully elucidated although some significant results have been achieved For example
Synchronization in a model of neuronal dynamics 717
1
2
2
1
Figure 9 Generalized synchronization in an asymmetrical driverndashresponse system (a) The timeseries of the two systems (b) The distance between the two systems (c) A plot of concurrent meanexcitatory membrane potentials V 1 against V 2 The diagonal represents the symmetry manifold
718 M Breakspear et al
Hunt et al (1997) Josic (1998) Kocarev et al (2000) have shown that the generalizedsynchronization manifold is smooth (differentiable) if and only if it is normally hyperbolic(ie the largest transverse Lyapunov exponent is less than the smallest tangential Lyapunovexponent prior to the introduction of parameter mismatch) Such a condition is met in thepresent setting when C = 04 hence the smoothness of the manifold in panel (c)
However subsequent to the introduction of parameter mismatch we will not be able touse transverse and tangential Lyapunov exponents to detect such synchronization since thesynchronization manifold may possess a complex topology for which it is a non-trivial (atbest) or impossible (at worse) task to define transverse and tangential directions For examplethe manifold may be smooth as in figure 9(c) but have a fractal topology If the dynamics arenot normally hyperbolic then the manifold may be fractal but also non-differentiable (Huntet al 1997) and hence will not have a unique tangent direction at any point Consequently wewill not be able to compute tangential or transverse Lyapunov exponents and hence determinethe stability of the synchronization manifold However other techniques for detecting thepresence of generalized synchronization in complex systems have evolved including theauxiliary system approach (Abarbanel et al 1996) and non-linear interdependence (Schiffet al 1996) which are now explored
421 The auxiliary system approach The auxiliary system is a third subsystem which isa replica of the original response system but coupled independently to the driver system andcommencing from independent initial conditions It is possible to demonstrate that showingthe existence of generalized synchronizationmdashthat equation (10) is satisfiedmdashis equivalent toshowing that the response system and the auxiliary system evolve identically (even though theyare not directly coupled) To determine whether or not this is the case reduces to an analysisof the Lyapunov exponents transverse to the symmetry manifold with respect to the responsesystem and the auxiliary system
Whilst this is a somewhat artificial test for generalized synchronization since theproduction of an auxiliary system for a physical system would be impossible such a testis useful in numerical and theoretical studies In figure 10 we present an example of theauxiliary system approach where the parameter δV = 07 for column 1 and 068 for column 2We consider two values of the coupling C = 0025 and 01 The auxiliary system is simplya copy of the second column beginning from an arbitrary initial state In the first instancewhere C = 0025 we notice that even after allowing for an initial transient state the secondcolumn and the auxiliary do not behave in identical fashions (figure 10(a)) However whenC = 01 we observe (figure 10(b)) that the second column and the auxiliary do behave innear-identical manners after an initial transient and hence conclude that for the particularvalue of the coupling strength considered equation (10) is satisfied and hence generalizedsynchronization between the first and second columns occurs
422 Nonlinear interdependence The question remains as to how we may detect thepresence of generalized synchronization in a practical setting As we mentioned aboveit is not possible to use an auxiliary-system-based approach to detect the presence ofgeneralized synchronization Instead we must consider techniques for detecting suchbehaviour from experimental or numerical time series data Algorithms of this type areoften referred to as detecting non-linear interdependence Nonlinear interdependence is ageneral term encompassing systems where the behaviour of one non-linear system affectsthe behaviour of another More specifically it can be defined as occurring if the nullhypothesis that the interdependence between two signals is purely linear can be rejectedat conventional levels (usually 95) of statistical significance hence it is an experimental
Synchronization in a model of neuronal dynamics 719
2
1
2
1
Figure 10 Concurrent evolution of the response system V 2 and lsquoauxiliaryrsquo system V 1 (a) C =0025 (b) C = 01
term (eg Schiff et al 1996) Whilst this does not strictly correspond to the existence of afunctional relationship between the two systems such a relationship is typically used tomotivate the techniques for its detection Schiff et al (1996) extended the logic of non-linearprediction algorithms from autonomous to coupled systems by exploiting the continuity ofsuch a relationship and hence devised the first method of detecting non-linear interdependencein time series data The basis of the technique is using the forward evolution of a phase spacevectorrsquos neighbours to predict the forward evolution of that vector and the forward evolutionof the concurrent vector of the other (possibly interdependent) system If the error of such aprocedure is better on average than chance (the chance error is normalized to equal one) thennon-linear interdependence is said to be present
720 M Breakspear et al
V1
V2
Figure 11 Experimental and numerical examples of weak generalized non-linear synchronizationin the presence of measurement noise (a) Human scalp EEG data from posterior bipolar scalpelectrodes (O1P3ndashO2P4) (b) Weakly coupled aperiodic data in the present model with additivewhite noise (with 5 of the amplitude of the model signal) Parameters δ1
V = 0605 δ2V = 061
aee = 036 C = 007
A variation of this algorithm optimized to detect weak non-linear interdependence (Terryand Breakspear 2003) was used recently to demonstrate that weak non-linear interdependence isoccasionally present in the scalp EEG of healthy resting human subjects (Breakspear and Terry2002a 2002b 2002c) An example of an epoch of lsquonon-linearly independentrsquo EEG obtainedusing this technique is presented in figure 11(a) For comparison output from the currentmodel is presented in figure 11(b) The model parameters were set such that the dynamicswas still aperiodic but the intermittent large-amplitude bursts were suppressed and had weakmutual coupling (C = 007) Extraneous noise to simulate the effect of extracranial muscleactivity contaminating the EEG data has been added to this time series It should be notedthat the amplitudes and time scales of the modelrsquos variables are dimensionless Nonethelessthe model facilitates qualititative interpretation of experimental findings
Synchronization in a model of neuronal dynamics 721
43 Chaotic phase synchronization
Whereas the concept of generalized synchronization was mathematically motivated and assuch more suited to theoretical investigations of systems the concept of phase synchronizationderives from a more practical signal processing approach Phase synchronization occurs inboth symmetric and asymmetric settings but is often more useful in the asymmetric case (andin experimental data)
Unfortunately the phase of a chaotic signal is typically ambiguous and can be defined in anumber of different ways For example in the case of the coherent Rossler attractor (Osipovet al 1997) the phase may be defined as the arctangent of two of the variables of the systemMore generally it can sometimes be defined as the angle that an orbit subtends with the centreof an attractor It is also possible to extract the phase of a chaotic signal from time seriesdata by use of the Hilbert transform This has enabled the demonstration of chaotic phasesynchronization in a number of coupled asymmetrical systems (Rosenblum et al 1996 1997Zaks et al 1999) Although such an approach does not guarantee that the phase will be uniqueit does permit the practical analysis of phase in noisy time series (Tass et al 1998)
To illustrate phase synchronization in two coupled columns we again vary the parameterδV which if the columns are viewed as oscillators acts to detune them from a commonfrequency An important question is that of the criteria on the basis of which we concludewhether phase synchronization occurs We use a simple locking criterion whereby phasesynchronization is said to occur if
|θ1(t)minus θ2(t)| lt K (11)
where is θi the phase of column i and K is a constant Notice that this does not mean thatsynchronization implies equality of phases merely that their difference is bounded
An example of phase synchronization with δV = 065 for column 1 and 066 for column 2is given in figure 12 In this figure rNMDA = 03 Qualitatively some sort of phase coherenceis evident in the time series panel (a) This was confirmed by plotting the sine of thephase difference which has been extracted using the Hilbert transform panel (b) Clearly|sin(θ1 minus θ2)| lt K and hence |θ1 minus θ2| is bounded It can also be seen that the phase-synchronized dynamics occurs on a highly structured manifold depicted in figure 12(c)However unlike the case for the generalized synchronization manifold depicted in figure 9 itis evident that this manifold is not a simple transformation of the symmetry manifold
It is also possible to see lsquobimodalrsquo stability whereby a system jumps betweenapproximately zero phase difference and a phase lag phase synchronization of π An exampleis depicted in figure 12(d) At approximately 2000 s the system can be seen to jumpspontaneously from the first to the second of these modes The Z -variable is plotted forgraphical purposes
5 Intermittency and desynchronization
51 Desynchronization in an array of two systems
Diminishing the strength of coupling across the blowout bifurcation does not lead to a simpletransition from synchronous to asynchronous oscillations but is instead associated withcomplex temporal behaviours Specifically when the largest transverse Lyapunov exponent isclose to zero the coupled systems exhibit epochs of synchronous behaviours punctuated bysharp bursts of desynchronization This is observed in systems exhibiting identicalgeneralizedand phase synchronization An example of such a phenomenon is given in figure 13 The timeseries panel (a) shows synchronous dynamics except at time = 1000 where it can be seenthat the two time series desynchronize briefly This is evident in panel (b) as a disruption in the
722 M Breakspear et al
2
1
Figure 12 Phase-synchronized coupled columns (a) The time series of V 1 (solid) and V 2 (b) Thesine of the phase difference obtained through Hilbert transformation of the time series in panel (a)(c) The phase synchronization manifold in the V 1ndashV 2 plane (d) The time series of Z (solid) andZ showing a phase slip of π between two modes
Synchronization in a model of neuronal dynamics 723
Figure 13 An example of phase desynchronization in coupled asymmetric systems with C = 01Time series plots of (a) each V -variable and (b) their Euclidean distance (c) The temporal evolutionof the phase difference between the two systems (d) An example of the phase difference evolutionover a longer time interval
724 M Breakspear et al
orderly evolution of the distance between the two systems and in panel (c) where the evolutionof the phase difference reveals a brief violation of the criterion |φ1 minus φ2| lt constant Anexample of the phase difference evolution over a longer time interval is shown in figure 13(d)Note that this is a three-dimensional plot with sin(φ1 minus φ2) and cos(φ1 minus φ2) as they- and z-axes Hence phase desynchronizations are evident as complete revolutions aroundthe cylindrical phase space in which this variable evolves It can be seen that these phenomenaoccur at irregular intervals in the temporal domain
This behaviour can be explained by considering the nature of a chaotic attractor In thestatistical sense an arbitrary initial condition is expected to lead to the chaotic behaviour onthe attractor Consequently when we calculate the Lyapunov exponents of the system theseexponents correspond to those of the chaotic attractor Typically however there are an infinitenumber of unstable periodic orbits embedded within the attractor If the path of a trajectorybecomes close to one of these orbits it may lsquoshadowrsquo the orbit for a brief period of time Eachof these orbits has a corresponding set of Lyapunov exponents which (like the attractor) can bedivided into tangential and transverse exponents These also vary when we change the couplingparameter but are typically not the same as the exponents of the attractor Hence the largesttransverse exponent of the attractor may be negative yet there may be positive transverseexponents associated with periodic orbits Considering all of these exponents together wewould therefore only expect to observe complete stability of the synchronization manifold inthe case where all of the transverse Lyapunov exponents are negative Bursting behaviourwill be observed when the one of these exponents becomes positive typically correspondingto a periodic orbit of low order (Pikovsky and Grassberger 1991 Rulkov and Sushchik 1997Maistrenko et al 1998 Heagy et al 1998) Due to the complex structure of such periodic orbitsthe desynchronous bursts occur across a wide range of timescales
In figure 14 we present the results of a long integration (16times105 time units) in which thelengths of sequential synchronous epochs of phase synchronization are studied The parametervalues were δV = 065 for column 1 and 0645 for column 2 with coupling strength10 equal to0125 In panel (a) the power spectrum of these sequential lengths is plotted The whiteness ofthis spectrum indicates that the lengths of consecutive synchronous epochs are independentThis is due to the high-dimensional lsquomixingrsquo effect of desynchronization In panel (b) thelengths of the synchronous epochs have been sorted again into the order of increasing lengthThe results for two coupling strengths are plotted C = 0125 (solid) and 0135 (dashed)Several features of this plot are of note Firstly the lengths of the synchronous epochsare distributed over almost three orders of magnitude Secondly for both plots there is anapproximately log-linear relationship between the length of an epoch and its relative orderThis reflects an approximately exponential relationship between the length of a synchronousepoch and its likelihood of occurrence Thirdly this relationship is skewed toward longersynchronous epochs for the stronger coupling strengths
This type of signal with patches of lsquoorderrsquo occurring over many timescales interruptedby bursts of disorder can be shown to have a high information capacity (Gatlin 1972)Moreover complex phase desynchronization may make an infrequent yet robust contributionto the dynamical pattern of the alpha rhythm in scalp EEG data for resting human subjects(Breakspear 2002) It is tempting to postulate that such a lsquoweakly stablersquo dynamics may assistin the generation of the rich and fluid mental associations that spontaneously emerge in restingcognitive states Although this phenomenon has been previously described in a variety of othersystems to our knowledge this is the first description in a biophysical model of neocortex
10 The introduction of parameter mismatch causes an increase in the strength of coupling required for stablesynchronization above the value of 01 required in the symmetric case
Synchronization in a model of neuronal dynamics 725
Figure 14 Properties of sequential epochs of phase-synchronous behaviour over a long integrationtime δ1
V = 065 δ2V = 0645 aee = 04 (a) The flat (white) power spectrum (b) Synchronous
epochs sorted in order of increasing length (solid C = 0125 dashed C = 0134)
52 Synchronization and complex dynamics in larger arrays
The synchronous and complex phenomena described above are also observed in arrays ofcoupled systems with N gt 2 Expanding the size of the array permits complex dynamicsin the spatial in addition to the temporal domain However the basic principles governingthe type and stability of synchronized motions are the same In this section a few of thepossible behaviours are discussed Physiological variation across the array is modelled bymodifying the parameter δV = 07 in each array by adding a random number of maximummagnitude 001
In figure 15(a) an array of N = 20 uncoupled (C = 0) systems is shown The lack ofsynchronization in this time series is evident By comparison the rapid transition to completesynchronization is evident in figure 15(b) when C = 03 In fact 19 of the 20 subsystems
726 M Breakspear et al
(b)
(c)
(a)
Figure 15 Synchronization phenomena in an array of coupled cortical columns (a) N = 20C = 0 (b) N = 20 C = 03 (c) N = 6 C = 01 (d) N = 12 C = 009 (e) N = 12C = 012 A small perturbation has been applied to one of the columns at t = 2500 s (f) Evolutionof the phase difference between two of the columns in panel (e) For clarity the W -variable isplotted in panel (c) and the Z -variable in (e)
have phase synchronized within one single cycle Intermediate values of coupling strength areassociated with a rich variety of phenomena For example subsystems with the most similarparameter values typically synchronize into clusters of various sizes An example of this isillustrated in figure 15(c) Note that there is no synchronization between the two differentclusters each containing three synchronized subsystems
Another complex behaviour occurs when subsystems transiently synchronize into largeclusters then spontaneously desynchronize again Hence the dynamics of the entire arrayjumps between different transient dynamics characterized by varying degrees of freedom Anexample of this phenomenon is given in figure 15(d) In addition it is possible to inducedesynchronization across an entire array by applying a small perturbation to one columnfor one time step An example is illustrated in figure 15(e) A small perturbation has been
Synchronization in a model of neuronal dynamics 727
Figure 15 (Continued)
added to one column after 2500 time units It can be seen that the entire array subsequentlydesynchronizes This illustrates the sensitivity of the dynamics to external stimuli Finally it ispossible to lsquoinducersquo synchronization between selected subsystems by increasing the strength ofcoupling between them selectively This allows an extra level of complexity to be introducedinto the model For example it may allow targeted information transfer between a partialnetwork of the entire array
As discussed in section 51 a univariate signal which jumps between epochs of lowentropy (high repetition) and bursts of high entropy (high variability) can be shown to havehigh information capacity This is because the repetition of certain patterns (as in the English
728 M Breakspear et al
language) permits error detectability Bursts of high entropy act in the opposite directionto increase message variability With a multivariate signal in addition to optimizing theinformation capacity within each signal the overall information capacity can be increasedthrough the type of itinerant synchronization briefly illustrated in this section Synchronizationacross an entire array increases the message reliability (since a single lsquoerrant letterrsquo in onesignal can be easily detected) Conversely desynchronization into multiple clusters increasesmessage variability since each cluster will produce independent information Hence a neuralarray which switches dynamically between a range of cluster configurations can be seen asadaptive in the sense that high synchronous states can be expressed when error detectabilityis a priority (such as during quick and reliable execution of a familiar task) but the array canrapidly desynchronize if variability becomes salient (for example if the environment suddenlybecomes novel) A formal operationalized demonstration of this principle is to be the subjectof future work
6 Discussion
In this paper we present a dynamical model for activity in an array of weakly coupledlocal neural assemblies The model incorporates the fundamental principles governing cellmembrane potential as determined by voltage- and ligand-gated ion channels In this wayit allows a reasonable amount of physiological complexity to be incorporated includingsynaptic activation and receptor subtypeswithout recourse to overwhelming detail Numericalanalysis of this model reveals a transition from independent oscillations to stable chaoticsynchronization depending on the balance of local versus long-range connectivity andbiophysical heterogeneity These excitatory couplings are subject to ascending subcorticalneuromodulation arising principally from monoaminergic nuclei such as Locus Coeruleus(Silberstein 1995) Neuromodulation of extrinsic connections was incorporated into the modelvia a coupling parameter C The stability of the synchronization is determined by the valueof the largest transverse Lyapunov exponent which in turn depends on the coupling parameterC If the coupling is weakened with the result that this exponent becomes positive a blowoutbifurcation occurs and independent oscillations re-emerge In the vicinity of the transitionnon-linear desynchronizations with complex temporal and spatial properties are observed
It is pleasing that complex and itinerant behaviour reminiscent of real neuronaldynamics emerges through such a simple manipulation of a single dynamic control parameterFurthermore the ubiquitous projections of classical neuromodulatory systems in real brainssuggest a clear biological mechanism by which the brain could self-organize throughmoderating the complexity of its own dynamics There are many reasons that the complicateddynamics of the sort demonstrated in this paper may be neurobiologically important Wemention four
First the presence of multiple clusters of phase-synchronized subsystems as discussedabove may be a uniquely non-linear mechanism facilitating the lsquocarriagersquo and distinction ofmultiple concurrent dynamic cell assemblies (and hence cognitive processing) This may beimportant for perceptual grouping binding and transitions as suggested by others (eg von derMalsburg 1985 Gray and Singer 1989)
Second the facility to decouple and reconfigure dynamic cell assemblies througha neuromodulatory control parameter may be important for avoiding low-entropysynchronization manifolds In the absence of itinerancy synchronization would renderneuronal dynamics incapable of representing changes in sensory input or indeed cognitive stateFor example in the model it was observed that small perturbations to one column were ableto induce a global change in neuronal dynamics by disrupting a state of weak synchronization
Synchronization in a model of neuronal dynamics 729
Inevitably alternative dynamic configurations emerge epitomizing the importance of lsquolabilersquobrain dynamics (Friston 2000a)
Thirdly from a purely functionalist point of view the balance between functionalspecialization and functional integration may rely upon the sorts of mechanism modelledin this paper The brain appears to adhere to two principles of functional organization namelysegregation of function within cortical areas and sub-areas and the integration of these areasin perceptual synthesis cognition and sensorimotor integration (Tononi et al 1994 Fristonet al 1995) The balance between preserving specialized dynamics unique to each arearequired by specialization and the coupling among these areas mediating functional integrationmay be achieved by the neuromodulatory mechanisms described above Significantlythe spontaneous variation in the numbers of degrees of freedom (for example whenlow-dimensional synchronization manifolds are shadowed) imparts a dynamic characteristicto this perspective that is precluded by linear mechanisms alone
Finally from a mechanistic perspective complexity and diversity of brain dynamics arerequired for selection of adaptive responses (Friston 2000b) There are many examples ofselective mechanisms operating within the brain All can be seen as performing an ascenton some objective function or fitness landscape These mechanisms require complex anditinerant dynamics to facilitate a thorough exploration of the landscape These landscapesmay range from those involved in perception (where the landscape is defined by predictionerror see eg Rao and Ballard (1998)) to those involved in value learning (where thelandscape is determined by the outcome of a behaviour eg in temporal difference models (seeeg Sutton and Barto 1990)) In short complexity is necessary to maintain an information-rich environment in which the brain can respond to sensory inputs An essential componentof this is the maintenance of diversity which is a prerequisite for selectionist mechanismsat many scales (Edelman 1993) Understanding the mechanistic basis of complex dynamicsas measured electrophysiologically may well depend on the sort of biologically plausiblemodels used in this paper This understanding may be important for both basic and clinicalneuroscience (eg Tass 2002)
The present study supplements and advances on other research in this field The studyof complex phase synchronization and clustering in neural oscillator networks similar tothe present model has previously been the subject of formal mathematical study Forexample Terman and Wang (1995) studied synchronization of blocks of neural oscillatorsthat were subject to perturbation by connected lsquostimulirsquo Somers and Kopell (1995) studiedthe emergence of both synchronization and travelling waves in networks of coupled neuraloscillators In the work of Terman and Wang the local dynamics were based on neurallsquocaricaturersquo models such as the FitzHughndashNagumo equations and are hence more abstract thanthe current study The approach of Somers and Kopell only required that the local dynamicsexhibit fast and relaxation phases and is hence more abstract still In both studies onlyperiodic dynamics are studied Complex patterns of chaos synchronization in neural arrayshave been characterized by nonlinear methods (Kaneko 1998) and their putative contributionto both learning (Nakajima and Ueda 1996) and perception (Zhao and Macau 2001 Breakspear2001) explored However these studies employed models such as the logistic equation andtent map which are quite abstract in comparison to the present study On the other handHansel and Sompolinsky (1996) studied the role of chaos synchronization in a model ofvisual perception in a cortical hypercolumn using a very similar approach to that adoptedhere but with the additional complexity that synaptic delays were incorporated However inthis and earlier work (Hansel and Sompolinsky 1992) only linear measures of dynamics andsynchrony (specifically the auto- and cross-correlation functions) were employed The analysisof chaotic synchrony in the study of Larter et al (1999) was based only upon observation of
730 M Breakspear et al
the Euclidean difference between variables from two nodes within a larger array Hence thesignificance of the present work is that it provides a quantitative nonlinear analysis of complexchaos synchronization in a biophysical model of cortical dynamics Such research is requiredin order to bridge the gap between quantitative nonlinear research into synchronization andcomputation in abstract models of neural arrays and qualitative or linear analysis in morebiophysically plausible models
Acknowledgments
The authors would like to thank Peter Robinson for assistance with the manuscript and R Larterfor helpful advice MB is a recipient of a NSW Institute of Psychiatry Research Grant anda University of Sydney SESQUI postdoctoral fellowship KJF is supported by the WellcomeTrust
References
Abarbanel H Rulkov N and Sushchik M 1996 Generalised synchronization of chaos the auxiliary system approachPhys Rev E 53 4528ndash35
Afraimovich V Verichev N and Rabinovich M 1986 Stochastic synchronization of oscillation in dissipative systemsRadiophys Quantum Electron 29 795ndash801
Breakspear M 2001 Perception of odours by a nonlinear model of the olfactory bulb Int J Neural Syst 11 101ndash24Breakspear M 2002 Nonlinear phase desynchronisation in human electroencephalographic data Human Brain Mapping
15 175ndash98Breakspear M and Terry J R 2002a Nonlinear interdependence in neural systems motivation theory and relevance
Int J Neurosci 112 1163ndash84Breakspear M and Terry J R 2002b Detection and description of nonlinear interdependence in normal multichannel
human EEG Clin Neurophysiol 113 735ndash53Breakspear M and Terry J R 2002c Topographic organization of nonlinear interdependence in multichannel human
EEG Neuroimage 16 822ndash35Destexhe A Rudolph M Fellous J M and Sejnowski T J 2001 Fluctuating synaptic conductances recreate in vivo-like
activity in neocortical neurons Neuroscience 107 13ndash24Douglas R J Mahowald M A and Martin K A 1996 Microarchitecture of neocortical columns Brain TheorymdashBiological
Basis and Computational Principles ed A Aertsen and V Braitenberg (Amsterdam Elsevier)Eckhorn R Bauer R Jordan W Brosch M Kruse W Munk M and Reitboeck H J 1988 Coherent oscillations a
mechanism of feature linking in the visual cortex Multiple electrode and correlation analyses in the cat BiolCybern 60 121ndash30
Edelman G M 1993 Neural Darwinism selection and reentrant signalling in higher brain function Neuron 10 115ndash25Engel A K Kreiter A K Konig P and Singer W 1991 Synchronization of oscillatory neuronal responses between
striate and extrastriate visual cortical areas of the cat Proc Natl Acad Sci USA 88 6048ndash52Friston K J 2000a The labile brain I Neuronal transients and nonlinear coupling Phil Trans R Soc B 355 215ndash36Friston K J 2000b The labile brain II Transients complexity and selection Phil Trans R Soc B 355 237ndash52Friston K J Tononi G Sporns O and Edelman G 1995 Characterising the complexity of neuronal interactions Human
Brain Mapping 3 302ndash14Fujisaka H and Yamada T 1983 Stability theory of synchronized motion in coupled-oscillator system Prog Theor
Phys 69 32ndash47Gatlin L 1972 Information Theory and the Living System (New York Columbia University Press)Gray C M Konig P Engel A and Singer W 1989 Oscillatory responses in cat visual cortex exhibit inter-columnar
synchronization which reflects global stimulus properties Science 338 334ndash7Gray C M and Singer W 1989 Stimulus specific neuronal oscillations in orientation columns of cat visual cortex Proc
Natl Acad Sci USA 86 1698ndash702Haig A R Gordon E Wright J J Meares R A and Bahramali H 2000 Synchronous cortical gamma-band activity in
task-relevant cognition Neuroreport 11 669ndash75Hansel D and Sompolinsky H 1992 Synchronization and computation in a chaotic neural network Phys Rev Lett 68
718ndash21
Synchronization in a model of neuronal dynamics 731
Hansel D and Sompolinsky H 1996 Chaos and synchrony in a model of a hypercolumn in visual cortex J ComputNeurosci 3 7ndash4
Heagy J Carroll T and Pecora L 1998 Desynchronization by periodic orbits Phys Rev E 57 R1253ndash6Hunt B Ott E and Yorke J 1997 Differentiable synchronization of chaos Phys Rev E 55 4029ndash34Josic K 1998 Invariant manifolds and synchronization of coupled dynamical systems Phys Rev Lett 80 3053ndash6Kandel E Schwartz J and Jessell T 2000 Principles of Neural Science (New York McGraw-Hill)Kaneko K 1998 On the strength of attractors in a high-dimensional system Physica D 124 322ndash44Kocarev L Parlitz U and Brown R 2000 Robust synchronization of chaotic systems Phys Rev E 61 3716ndash20Larter R Speelman B and Worth R M 1999 A coupled ordinary differential equation lattice model for the simulation
of epileptic seizures Chaos 9 795ndash804Lumer E Edelman G and Tononi G 1997a Neural dynamics in a model of the thalamocortical system I Layers loops
and the emergence of fast synchronous oscillations Cereb Cortex 7 207ndash27Lumer E Edelman G and Tononi G 1997b Neural dynamics in a model of the thalamocortical system II The role of
neural synchrony tested through perturbations of spike timing Cereb Cortex 7 228ndash36Magleby K 1987 Short-term modulation at synapses Synaptic Function ed G M Edelman W E Gall and
W M Cowan (New York Wiley)Maistrenko Y Maistrenko V Popovich A and Mosekilde E 1998 Transverse instability and riddled basins in a system
of two coupled logistic maps Phys Rev E 57 2713ndash24Morris C and Lecar H 1981 Voltage oscillations in the barnacle giant muscle fiber Biophys J 35 193ndash213Mountcastle V B 1997 The columnar organization of the neocortex Brain 120 701ndash22Mountcastle V B 1998 The Cerebral Cortex (Cambridge MA Harvard University Press)Nakajima H and Ueda Y 1996 Riddled basins of the optimal states in learning dynamical systems Physica D 99 35ndash44Nunez P L 1995 Neocortical Dynamics and Human Brain Rhythms (Oxford Oxford University Press)Osipov G V Pikovsky A S Rosenblum M G and Kurths J 1997 Phase synchronization effects in a lattice of nonidentical
Rossler oscillators Phys Rev E 55 2353ndash61Ott E and Sommerer J 1994 Blowout bifurcations the occurrence of riddled basins and onndashoff intermittency Phys
Lett A 188 39ndash47Pecora L and Carroll T 1990 Synchronization in chaotic systems Phys Rev Lett 64 821ndash4Pikovsky A and Grassberger P 1991 Symmetry breaking bifurcation for coupled chaotic attractors J Phys A Math
Gen 24 4587ndash97Rao R P and Ballard D H 1998 Predictive coding in the visual cortex a functional interpretation of some extra-classical
receptive field effects Nature Neurosci 2 79ndash87Robinson P A Wright J J and Rennie C R 1998 Synchronous oscillations in the cerebral cortex Phys Rev E 57
4578ndash88Rodriguez E George N Lachauz J P Martinerie J Renault B and Varela F J 1999 Perceptionrsquos shadow long distance
synchronization of human brain activity Nature 397 430ndash3Rosenblum M Pikovsky A and Kurths J 1996 Phase synchronization of chaotic oscillators Phys Rev Lett 76 1804ndash7Rosenblum M Pikovsky A and Kurths J 1997 From phase to lag synchronization in coupled chaotic oscillators Phys
Rev Lett 78 4193ndash6Rulkov N and Sushchik M 1997 Robustness of synchronized chaotic oscillations Int J Bifurc Chaos 7 625ndash43Rulkov N Sushchik M Tsimring L and Abarbanel H 1995 Generalized synchronization of chaos in unidirectionally
coupled chaotic systems Phys Rev E 51 980ndash94Schiff S So P Chang T Burke R and Sauer T 1996 Detecting dynamical interdependence and generalized synchrony
through mutual prediction in a neural ensemble Phys Rev E 54 6708ndash24Silberstein R B 1995 Neuromodulations of neocortical dynamics Neocortical Dynamics and Human EEG Rhythms
ed P L Nunez (Oxford Oxford University Press)Singer W 1995 Putative functions of temporal correlations in neocortical processing Large-Scale Neuronal Theories
of the Brain ed C Koch and J Davis (London MIT Press)Somers D and Kopell N 1995 Waves and synchrony in networks of oscillators of relaxation and non-relaxation type
Physica D 89 169ndash83Sutton R S and Barto A G 1990 Time derivative models of Pavlovian reinforcement Learning and Computational
Neuroscience Foundations of Adaptive Networks ed M Gabriel and J Moore (Cambridge MA MIT Press) pp497ndash538
Szentagothai J 1983 The modular architectonic principle of neural centers Rev Physiol Biochem Pharmacol 9811ndash61
Tallon-Baudry C Betrand O and Fischer C 2001 Oscillatory synchrony between human extrastriate areas during visualshort-term memory maintenance J Neurosci 21 RC177(1ndash5)
Tass P A 2002 Desynchronization of brain rhythms with soft phase-resetting techniques Biol Cybern 87 102ndash15
732 M Breakspear et al
Tass P Rosenblum M Weule J Kurths J Pikovsky A Volkmann J Schnitzler A and Freund H 1998 Detection of nmphase locking from noisy data application to magnetoencephalography Phys Rev Lett 81 3291ndash4
Terman D and Wang D 1995 Global competition and local cooperation in a network of neural oscillators Physica D81 148ndash76
Terry J and Breakspear M 2003 An improved algorithm for the detection of dynamical interdependence in bivariatetime-series Biol Cybern 88 129ndash36
Tononi G Sporns O and Edelman G M 1994 A measure for brain complexity relating functional segregation andintegration in the nervous system Proc Natl Acad Sci USA 91 5033ndash7
von der Malsburg C 1985 Nervous structures with dynamical links Phys Chem 89 703ndash10Wang X 1991 Period-doublings to chaos in a simple neural network an analytic proof Complex Syst 5 425ndash41Wennekers T and Pasemann F 1996 Synchronous chaos in high dimensional modular neural networks Int J Bifurc
Chaos 6 2055ndash67Zaks M A Parks E H Rosenblum M G and Kurths J 1999 Alternative locking ratios in imperfect phase synchronization
Phys Rev Lett 82 4228ndash31Zhao L and Macau E N 2001 A network of dynamically coupled chaotic maps for scene segmentation IEEE Trans
Neural Netw 12 1375ndash85

712 M Breakspear et al
we study the emergence of cooperative behaviour that results from the introduction of couplingbetween columns Three types of interdependent behaviour of two systems are describedidentical generalized and phase synchronization Synchronous behaviours in larger arrays arealso illustrated
41 Identical chaotic synchronization
411 Transition to identical chaotic synchronization The most straightforward extensionand the one that we consider first is that of two coupled cortical columns where all parametersare in the first case assumed identical7 Trivially the two systems will evolve identicallywhen started with identical initial conditions even if C = 0 This is simply a statement ofthe deterministic nature of the evolution equations In general the random initial conditionswill not be equal in which case it is hardly surprising that the different columns will evolveindependently An example is presented in figure 6 with VT = 06 in both systems C = 0and independent random initial conditions In figure 6(a) we show a segment of the timeseries for each of the two subsystems where it can be seen that the time series are chaoticand asynchronous In figure 6(b) the concurrent values of V 1 and V 2 are plotted against eachother showing that they bear no obvious interrelationship
With the introduction of coupling C gt 0 the systems begin to show epochs ofsynchronized behaviour At C sim= 01 there is a transition to stable fully synchronizedmotion as evident in an exemplar time series shown in figure 7(a) where C = 015 Infigure 7(b) we present the concurrent values of V 1 versus V 2 illustrating that in contrast tothe case in figure 6(b) the systems have become embedded in the low-dimensional lsquosymmetrymanifoldrsquo V 1 = V 2 Panel (c) shows a log-linear plot of the temporal evolution of the distanceof the coupled system from this manifold It can be seen that on average the two systemsapproach at an exponential rate with constant time-averaged exponent Whilst this phenomenon(lsquoidentical chaotic synchronizationrsquo) has been described in a variety of other systems since itsfirst description (Fujisaka and Yamada 1983 Pecora and Carroll 1990) to our knowledge thisis its first quantitative8 description in a detailed physiological model of a neural system
The symmetry of the dynamical system obtained by setting the state parameters in eachcolumn identical forces the invariance of the symmetry manifold V 1 = V 2 Moreover thedynamics within this manifold is governed by the same equations of motion as for a singlecortical column This manifold which contains the orbits of the synchronized system is calledthe synchronization manifold The coupling parameter C does not affect the dynamics withinthe synchronization manifold and as such is termed a normal parameter
412 lsquoBlowout bifurcationrsquo An important issue is the stability of the orbits on thesynchronization manifold This can be determined by analysing the Lyapunov spectraBecause the two-column system evolves in a six-dimensional phase space there are sixLyapunov exponents However for the synchronized system it is helpful to subdivide theseinto those that describe the dynamics within the synchronization manifold and those thatdescribe the dynamics in the transverse direction The latter describe the rate of convergenceor divergence of the coupled systems toward or away from this manifoldmdashand hence thetendency for the two systems to synchronize or separate For the time series in figure 7 this is
7 Whilst it is of course physiologically unrealistic to assume that two columns would have entirely identical structuresit is mathematically simpler to start analysing the system in this situation8 A qualitative description in a neural system with a different form of coupling (potassium diffusion) is evident infigure 8 of Larter et al (1999) Synchronous chaos in an abstract model of a neural system was also described inWennekers and Pasemann (1996)
Synchronization in a model of neuronal dynamics 713
1
2
2
1
Figure 6 Independent oscillations in a two-column system with identical parameters butindependent initial conditions (a) The time series (b) A plot of concurrent mean excitatorymembrane potentials V 1 against V 2
related to the slope of the plot in panel (c)9 For this strength of coupling the largest transverseexponent is negative (minus002) indicating that perturbations away from this state of identicalsynchronization decay and thus that the state of identical chaotic synchronization is stableagainst noise (ie dynamically stable)
To calculate the relationship between coupling strength and the Lyapunov exponents weused an algorithm which measures the growth of perturbations of the variational equations (thelsquoJacobianrsquo) of the system (we do not define these equations here as they are cumbersome anddo not enhance the understanding of the system) We calculated these exponents for increasingvalues of the coupling parameter C from 0 to 02 in increments of 0000 25 The results are
9 Only two of the six dependent variables are shown in figure 7 The actual transverse Lyapunov exponents reflectthe temporal evolution of all six and hence are somewhat more complex than that given by the slope of panel (c)
714 M Breakspear et al
1
2
Figure 7 Identical synchronization in the same system as in figure 7 but with C = 02 (a) Thetime series (b) A plot of concurrent mean excitatory membrane potentials V 1 against V 2 (c) Thelogarithm of the Euclidean distance between V 1 and V 2 versus time (linear)
Synchronization in a model of neuronal dynamics 715
Figure 8 Lyapunov spectra for a coupled system with δV = 07 and C varying from 0 to 02 insteps of 0000 33 At each value of C all Lyapunov exponents are calculated Least-squares linesof best fit were then fitted to the results for each exponent Solid lines indicate transverse exponents(λ) and dashed lines indicate the tangential exponents () Panel (a) shows all three transverseand three tangential exponents Panel (b) only shows the largest two of each
illustrated in figure 8 Panel (a) shows all exponents For illustrative purposes only the mostpositive four are plotted in panel (b) In both panels the tangential exponents () are fitted (inthe least-squares sense) with a dashed line and the transverse ones (λ) are fitted with a solidline They are labelled 1ndash3 in order of decreasing positivity The important features are asfollows For the range of coupling strengths that we consider the largest tangential Lyapunovexponent is positive which indicates the presence of chaotic behaviour within the manifoldWhen examining the line of best fit for this exponent we see that this exponent does not varywhen altering the coupling parameter This is true for all three tangential exponents and verifiesthat the changes in the coupling parameter do not alter the dynamics on the synchronizationmanifold (that is the coupling parameter is normal)
716 M Breakspear et al
The largest transverse Lyapunov exponent (denoted as λ1 in the figure) is a particularlyimportant exponent since it governs the stability of the attractor in the synchronization manifoldwith respect to perturbations out of the manifold If this exponent is negative then we expectsmall perturbations of trajectories to reconverge to the manifold in finite time If however thisexponent is positive then we would instead expect perturbed trajectories to diverge from themanifold This change in local behaviour is termed a bifurcation and the point at which itoccurs the bifurcation point It is apparent in figure 8 that the zero crossing of this exponentmdashthe bifurcation pointmdashoccurs at approximately C = 01 in the present system This typeof bifurcation has been termed a blowout bifurcation since once transverse stability is losttrajectories are lsquoblown outrsquo from the low-dimensional synchronization manifold into the fullphase space (Ott and Sommerer 1994) Hence in the present setting a blowout bifurcation isobserved if C falls below 01
We note two other observations Increasing rNMDA which corresponds to increasing thepopulation of NMDA receptors decreases the coupling strength required to achieve stablesynchronization On the other hand increasing the overall excitatory-to-excitatory synapticconnectivity (aee) has the opposite effect This is because the effect of local excitatory feedbackincreases the tangential Lyapunov exponents which has the effect of lsquosplittingrsquo the synchronizeddynamics
In the above simulations all the parameters of both systems were equal In realphysiological systems such symmetry between systems is clearly impossible We nowconsider the case in which the two columns have different structures A simple way toincorporate this whilst at the same time preserving (as much as possible) simple dynamics is tovary the parameter δV for each column This has the effect of destroying any symmetry-forcedinvariant manifolds and so the simple case of synchronization corresponding to identicallyevolving dynamics will not exist in this case However more exotic forms of synchronizationare still possible and we discuss these next
42 Generalized chaotic synchronization
In order to maintain consistency with the literature we study non-linear interdependence in thesetting where the systems are asymmetrical but the coupling is unidirectional This is easilyachieved in the present system by making C directed and setting C21 = 0 In this settingwe refer to the lsquofreersquo system as the driver and the lsquoslavedrsquo system as the response We firstask the question what is the effect of introducing a slight parameter mismatch into a systemexhibiting stable chaos synchronization An example of this is presented in figure 9 whereC12 = 04 and parameter mismatch has been incorporated by setting δV = 07 in the driversystem and δV = 0699 in the response system In panel (a) the time series appear qualitativelyvery similar However in panel (b) where the difference between the systems is plotted it isapparent that the separation between the two systems is of the same order of magnitude as thesize of each attractor In panel (c) the effect of the parameter mismatch on the synchronizationmanifold can be seen as a transformation away from the symmetry manifold
It has been proposed that in such a setting there exists a continuous functional relationshipbetween the dynamics of the driver and response systems
x2(t) = ψ(x1(t)) (10)
where xi = V i W i Z i This situation has been referred to in the literature as generalizedsynchronization (Afraimovich et al 1986 Rulkov et al 1995) The properties of ψ andthe topology and stability of the synchronization manifold in this context have not yetbeen fully elucidated although some significant results have been achieved For example
Synchronization in a model of neuronal dynamics 717
1
2
2
1
Figure 9 Generalized synchronization in an asymmetrical driverndashresponse system (a) The timeseries of the two systems (b) The distance between the two systems (c) A plot of concurrent meanexcitatory membrane potentials V 1 against V 2 The diagonal represents the symmetry manifold
718 M Breakspear et al
Hunt et al (1997) Josic (1998) Kocarev et al (2000) have shown that the generalizedsynchronization manifold is smooth (differentiable) if and only if it is normally hyperbolic(ie the largest transverse Lyapunov exponent is less than the smallest tangential Lyapunovexponent prior to the introduction of parameter mismatch) Such a condition is met in thepresent setting when C = 04 hence the smoothness of the manifold in panel (c)
However subsequent to the introduction of parameter mismatch we will not be able touse transverse and tangential Lyapunov exponents to detect such synchronization since thesynchronization manifold may possess a complex topology for which it is a non-trivial (atbest) or impossible (at worse) task to define transverse and tangential directions For examplethe manifold may be smooth as in figure 9(c) but have a fractal topology If the dynamics arenot normally hyperbolic then the manifold may be fractal but also non-differentiable (Huntet al 1997) and hence will not have a unique tangent direction at any point Consequently wewill not be able to compute tangential or transverse Lyapunov exponents and hence determinethe stability of the synchronization manifold However other techniques for detecting thepresence of generalized synchronization in complex systems have evolved including theauxiliary system approach (Abarbanel et al 1996) and non-linear interdependence (Schiffet al 1996) which are now explored
421 The auxiliary system approach The auxiliary system is a third subsystem which isa replica of the original response system but coupled independently to the driver system andcommencing from independent initial conditions It is possible to demonstrate that showingthe existence of generalized synchronizationmdashthat equation (10) is satisfiedmdashis equivalent toshowing that the response system and the auxiliary system evolve identically (even though theyare not directly coupled) To determine whether or not this is the case reduces to an analysisof the Lyapunov exponents transverse to the symmetry manifold with respect to the responsesystem and the auxiliary system
Whilst this is a somewhat artificial test for generalized synchronization since theproduction of an auxiliary system for a physical system would be impossible such a testis useful in numerical and theoretical studies In figure 10 we present an example of theauxiliary system approach where the parameter δV = 07 for column 1 and 068 for column 2We consider two values of the coupling C = 0025 and 01 The auxiliary system is simplya copy of the second column beginning from an arbitrary initial state In the first instancewhere C = 0025 we notice that even after allowing for an initial transient state the secondcolumn and the auxiliary do not behave in identical fashions (figure 10(a)) However whenC = 01 we observe (figure 10(b)) that the second column and the auxiliary do behave innear-identical manners after an initial transient and hence conclude that for the particularvalue of the coupling strength considered equation (10) is satisfied and hence generalizedsynchronization between the first and second columns occurs
422 Nonlinear interdependence The question remains as to how we may detect thepresence of generalized synchronization in a practical setting As we mentioned aboveit is not possible to use an auxiliary-system-based approach to detect the presence ofgeneralized synchronization Instead we must consider techniques for detecting suchbehaviour from experimental or numerical time series data Algorithms of this type areoften referred to as detecting non-linear interdependence Nonlinear interdependence is ageneral term encompassing systems where the behaviour of one non-linear system affectsthe behaviour of another More specifically it can be defined as occurring if the nullhypothesis that the interdependence between two signals is purely linear can be rejectedat conventional levels (usually 95) of statistical significance hence it is an experimental
Synchronization in a model of neuronal dynamics 719
2
1
2
1
Figure 10 Concurrent evolution of the response system V 2 and lsquoauxiliaryrsquo system V 1 (a) C =0025 (b) C = 01
term (eg Schiff et al 1996) Whilst this does not strictly correspond to the existence of afunctional relationship between the two systems such a relationship is typically used tomotivate the techniques for its detection Schiff et al (1996) extended the logic of non-linearprediction algorithms from autonomous to coupled systems by exploiting the continuity ofsuch a relationship and hence devised the first method of detecting non-linear interdependencein time series data The basis of the technique is using the forward evolution of a phase spacevectorrsquos neighbours to predict the forward evolution of that vector and the forward evolutionof the concurrent vector of the other (possibly interdependent) system If the error of such aprocedure is better on average than chance (the chance error is normalized to equal one) thennon-linear interdependence is said to be present
720 M Breakspear et al
V1
V2
Figure 11 Experimental and numerical examples of weak generalized non-linear synchronizationin the presence of measurement noise (a) Human scalp EEG data from posterior bipolar scalpelectrodes (O1P3ndashO2P4) (b) Weakly coupled aperiodic data in the present model with additivewhite noise (with 5 of the amplitude of the model signal) Parameters δ1
V = 0605 δ2V = 061
aee = 036 C = 007
A variation of this algorithm optimized to detect weak non-linear interdependence (Terryand Breakspear 2003) was used recently to demonstrate that weak non-linear interdependence isoccasionally present in the scalp EEG of healthy resting human subjects (Breakspear and Terry2002a 2002b 2002c) An example of an epoch of lsquonon-linearly independentrsquo EEG obtainedusing this technique is presented in figure 11(a) For comparison output from the currentmodel is presented in figure 11(b) The model parameters were set such that the dynamicswas still aperiodic but the intermittent large-amplitude bursts were suppressed and had weakmutual coupling (C = 007) Extraneous noise to simulate the effect of extracranial muscleactivity contaminating the EEG data has been added to this time series It should be notedthat the amplitudes and time scales of the modelrsquos variables are dimensionless Nonethelessthe model facilitates qualititative interpretation of experimental findings
Synchronization in a model of neuronal dynamics 721
43 Chaotic phase synchronization
Whereas the concept of generalized synchronization was mathematically motivated and assuch more suited to theoretical investigations of systems the concept of phase synchronizationderives from a more practical signal processing approach Phase synchronization occurs inboth symmetric and asymmetric settings but is often more useful in the asymmetric case (andin experimental data)
Unfortunately the phase of a chaotic signal is typically ambiguous and can be defined in anumber of different ways For example in the case of the coherent Rossler attractor (Osipovet al 1997) the phase may be defined as the arctangent of two of the variables of the systemMore generally it can sometimes be defined as the angle that an orbit subtends with the centreof an attractor It is also possible to extract the phase of a chaotic signal from time seriesdata by use of the Hilbert transform This has enabled the demonstration of chaotic phasesynchronization in a number of coupled asymmetrical systems (Rosenblum et al 1996 1997Zaks et al 1999) Although such an approach does not guarantee that the phase will be uniqueit does permit the practical analysis of phase in noisy time series (Tass et al 1998)
To illustrate phase synchronization in two coupled columns we again vary the parameterδV which if the columns are viewed as oscillators acts to detune them from a commonfrequency An important question is that of the criteria on the basis of which we concludewhether phase synchronization occurs We use a simple locking criterion whereby phasesynchronization is said to occur if
|θ1(t)minus θ2(t)| lt K (11)
where is θi the phase of column i and K is a constant Notice that this does not mean thatsynchronization implies equality of phases merely that their difference is bounded
An example of phase synchronization with δV = 065 for column 1 and 066 for column 2is given in figure 12 In this figure rNMDA = 03 Qualitatively some sort of phase coherenceis evident in the time series panel (a) This was confirmed by plotting the sine of thephase difference which has been extracted using the Hilbert transform panel (b) Clearly|sin(θ1 minus θ2)| lt K and hence |θ1 minus θ2| is bounded It can also be seen that the phase-synchronized dynamics occurs on a highly structured manifold depicted in figure 12(c)However unlike the case for the generalized synchronization manifold depicted in figure 9 itis evident that this manifold is not a simple transformation of the symmetry manifold
It is also possible to see lsquobimodalrsquo stability whereby a system jumps betweenapproximately zero phase difference and a phase lag phase synchronization of π An exampleis depicted in figure 12(d) At approximately 2000 s the system can be seen to jumpspontaneously from the first to the second of these modes The Z -variable is plotted forgraphical purposes
5 Intermittency and desynchronization
51 Desynchronization in an array of two systems
Diminishing the strength of coupling across the blowout bifurcation does not lead to a simpletransition from synchronous to asynchronous oscillations but is instead associated withcomplex temporal behaviours Specifically when the largest transverse Lyapunov exponent isclose to zero the coupled systems exhibit epochs of synchronous behaviours punctuated bysharp bursts of desynchronization This is observed in systems exhibiting identicalgeneralizedand phase synchronization An example of such a phenomenon is given in figure 13 The timeseries panel (a) shows synchronous dynamics except at time = 1000 where it can be seenthat the two time series desynchronize briefly This is evident in panel (b) as a disruption in the
722 M Breakspear et al
2
1
Figure 12 Phase-synchronized coupled columns (a) The time series of V 1 (solid) and V 2 (b) Thesine of the phase difference obtained through Hilbert transformation of the time series in panel (a)(c) The phase synchronization manifold in the V 1ndashV 2 plane (d) The time series of Z (solid) andZ showing a phase slip of π between two modes
Synchronization in a model of neuronal dynamics 723
Figure 13 An example of phase desynchronization in coupled asymmetric systems with C = 01Time series plots of (a) each V -variable and (b) their Euclidean distance (c) The temporal evolutionof the phase difference between the two systems (d) An example of the phase difference evolutionover a longer time interval
724 M Breakspear et al
orderly evolution of the distance between the two systems and in panel (c) where the evolutionof the phase difference reveals a brief violation of the criterion |φ1 minus φ2| lt constant Anexample of the phase difference evolution over a longer time interval is shown in figure 13(d)Note that this is a three-dimensional plot with sin(φ1 minus φ2) and cos(φ1 minus φ2) as they- and z-axes Hence phase desynchronizations are evident as complete revolutions aroundthe cylindrical phase space in which this variable evolves It can be seen that these phenomenaoccur at irregular intervals in the temporal domain
This behaviour can be explained by considering the nature of a chaotic attractor In thestatistical sense an arbitrary initial condition is expected to lead to the chaotic behaviour onthe attractor Consequently when we calculate the Lyapunov exponents of the system theseexponents correspond to those of the chaotic attractor Typically however there are an infinitenumber of unstable periodic orbits embedded within the attractor If the path of a trajectorybecomes close to one of these orbits it may lsquoshadowrsquo the orbit for a brief period of time Eachof these orbits has a corresponding set of Lyapunov exponents which (like the attractor) can bedivided into tangential and transverse exponents These also vary when we change the couplingparameter but are typically not the same as the exponents of the attractor Hence the largesttransverse exponent of the attractor may be negative yet there may be positive transverseexponents associated with periodic orbits Considering all of these exponents together wewould therefore only expect to observe complete stability of the synchronization manifold inthe case where all of the transverse Lyapunov exponents are negative Bursting behaviourwill be observed when the one of these exponents becomes positive typically correspondingto a periodic orbit of low order (Pikovsky and Grassberger 1991 Rulkov and Sushchik 1997Maistrenko et al 1998 Heagy et al 1998) Due to the complex structure of such periodic orbitsthe desynchronous bursts occur across a wide range of timescales
In figure 14 we present the results of a long integration (16times105 time units) in which thelengths of sequential synchronous epochs of phase synchronization are studied The parametervalues were δV = 065 for column 1 and 0645 for column 2 with coupling strength10 equal to0125 In panel (a) the power spectrum of these sequential lengths is plotted The whiteness ofthis spectrum indicates that the lengths of consecutive synchronous epochs are independentThis is due to the high-dimensional lsquomixingrsquo effect of desynchronization In panel (b) thelengths of the synchronous epochs have been sorted again into the order of increasing lengthThe results for two coupling strengths are plotted C = 0125 (solid) and 0135 (dashed)Several features of this plot are of note Firstly the lengths of the synchronous epochsare distributed over almost three orders of magnitude Secondly for both plots there is anapproximately log-linear relationship between the length of an epoch and its relative orderThis reflects an approximately exponential relationship between the length of a synchronousepoch and its likelihood of occurrence Thirdly this relationship is skewed toward longersynchronous epochs for the stronger coupling strengths
This type of signal with patches of lsquoorderrsquo occurring over many timescales interruptedby bursts of disorder can be shown to have a high information capacity (Gatlin 1972)Moreover complex phase desynchronization may make an infrequent yet robust contributionto the dynamical pattern of the alpha rhythm in scalp EEG data for resting human subjects(Breakspear 2002) It is tempting to postulate that such a lsquoweakly stablersquo dynamics may assistin the generation of the rich and fluid mental associations that spontaneously emerge in restingcognitive states Although this phenomenon has been previously described in a variety of othersystems to our knowledge this is the first description in a biophysical model of neocortex
10 The introduction of parameter mismatch causes an increase in the strength of coupling required for stablesynchronization above the value of 01 required in the symmetric case
Synchronization in a model of neuronal dynamics 725
Figure 14 Properties of sequential epochs of phase-synchronous behaviour over a long integrationtime δ1
V = 065 δ2V = 0645 aee = 04 (a) The flat (white) power spectrum (b) Synchronous
epochs sorted in order of increasing length (solid C = 0125 dashed C = 0134)
52 Synchronization and complex dynamics in larger arrays
The synchronous and complex phenomena described above are also observed in arrays ofcoupled systems with N gt 2 Expanding the size of the array permits complex dynamicsin the spatial in addition to the temporal domain However the basic principles governingthe type and stability of synchronized motions are the same In this section a few of thepossible behaviours are discussed Physiological variation across the array is modelled bymodifying the parameter δV = 07 in each array by adding a random number of maximummagnitude 001
In figure 15(a) an array of N = 20 uncoupled (C = 0) systems is shown The lack ofsynchronization in this time series is evident By comparison the rapid transition to completesynchronization is evident in figure 15(b) when C = 03 In fact 19 of the 20 subsystems
726 M Breakspear et al
(b)
(c)
(a)
Figure 15 Synchronization phenomena in an array of coupled cortical columns (a) N = 20C = 0 (b) N = 20 C = 03 (c) N = 6 C = 01 (d) N = 12 C = 009 (e) N = 12C = 012 A small perturbation has been applied to one of the columns at t = 2500 s (f) Evolutionof the phase difference between two of the columns in panel (e) For clarity the W -variable isplotted in panel (c) and the Z -variable in (e)
have phase synchronized within one single cycle Intermediate values of coupling strength areassociated with a rich variety of phenomena For example subsystems with the most similarparameter values typically synchronize into clusters of various sizes An example of this isillustrated in figure 15(c) Note that there is no synchronization between the two differentclusters each containing three synchronized subsystems
Another complex behaviour occurs when subsystems transiently synchronize into largeclusters then spontaneously desynchronize again Hence the dynamics of the entire arrayjumps between different transient dynamics characterized by varying degrees of freedom Anexample of this phenomenon is given in figure 15(d) In addition it is possible to inducedesynchronization across an entire array by applying a small perturbation to one columnfor one time step An example is illustrated in figure 15(e) A small perturbation has been
Synchronization in a model of neuronal dynamics 727
Figure 15 (Continued)
added to one column after 2500 time units It can be seen that the entire array subsequentlydesynchronizes This illustrates the sensitivity of the dynamics to external stimuli Finally it ispossible to lsquoinducersquo synchronization between selected subsystems by increasing the strength ofcoupling between them selectively This allows an extra level of complexity to be introducedinto the model For example it may allow targeted information transfer between a partialnetwork of the entire array
As discussed in section 51 a univariate signal which jumps between epochs of lowentropy (high repetition) and bursts of high entropy (high variability) can be shown to havehigh information capacity This is because the repetition of certain patterns (as in the English
728 M Breakspear et al
language) permits error detectability Bursts of high entropy act in the opposite directionto increase message variability With a multivariate signal in addition to optimizing theinformation capacity within each signal the overall information capacity can be increasedthrough the type of itinerant synchronization briefly illustrated in this section Synchronizationacross an entire array increases the message reliability (since a single lsquoerrant letterrsquo in onesignal can be easily detected) Conversely desynchronization into multiple clusters increasesmessage variability since each cluster will produce independent information Hence a neuralarray which switches dynamically between a range of cluster configurations can be seen asadaptive in the sense that high synchronous states can be expressed when error detectabilityis a priority (such as during quick and reliable execution of a familiar task) but the array canrapidly desynchronize if variability becomes salient (for example if the environment suddenlybecomes novel) A formal operationalized demonstration of this principle is to be the subjectof future work
6 Discussion
In this paper we present a dynamical model for activity in an array of weakly coupledlocal neural assemblies The model incorporates the fundamental principles governing cellmembrane potential as determined by voltage- and ligand-gated ion channels In this wayit allows a reasonable amount of physiological complexity to be incorporated includingsynaptic activation and receptor subtypeswithout recourse to overwhelming detail Numericalanalysis of this model reveals a transition from independent oscillations to stable chaoticsynchronization depending on the balance of local versus long-range connectivity andbiophysical heterogeneity These excitatory couplings are subject to ascending subcorticalneuromodulation arising principally from monoaminergic nuclei such as Locus Coeruleus(Silberstein 1995) Neuromodulation of extrinsic connections was incorporated into the modelvia a coupling parameter C The stability of the synchronization is determined by the valueof the largest transverse Lyapunov exponent which in turn depends on the coupling parameterC If the coupling is weakened with the result that this exponent becomes positive a blowoutbifurcation occurs and independent oscillations re-emerge In the vicinity of the transitionnon-linear desynchronizations with complex temporal and spatial properties are observed
It is pleasing that complex and itinerant behaviour reminiscent of real neuronaldynamics emerges through such a simple manipulation of a single dynamic control parameterFurthermore the ubiquitous projections of classical neuromodulatory systems in real brainssuggest a clear biological mechanism by which the brain could self-organize throughmoderating the complexity of its own dynamics There are many reasons that the complicateddynamics of the sort demonstrated in this paper may be neurobiologically important Wemention four
First the presence of multiple clusters of phase-synchronized subsystems as discussedabove may be a uniquely non-linear mechanism facilitating the lsquocarriagersquo and distinction ofmultiple concurrent dynamic cell assemblies (and hence cognitive processing) This may beimportant for perceptual grouping binding and transitions as suggested by others (eg von derMalsburg 1985 Gray and Singer 1989)
Second the facility to decouple and reconfigure dynamic cell assemblies througha neuromodulatory control parameter may be important for avoiding low-entropysynchronization manifolds In the absence of itinerancy synchronization would renderneuronal dynamics incapable of representing changes in sensory input or indeed cognitive stateFor example in the model it was observed that small perturbations to one column were ableto induce a global change in neuronal dynamics by disrupting a state of weak synchronization
Synchronization in a model of neuronal dynamics 729
Inevitably alternative dynamic configurations emerge epitomizing the importance of lsquolabilersquobrain dynamics (Friston 2000a)
Thirdly from a purely functionalist point of view the balance between functionalspecialization and functional integration may rely upon the sorts of mechanism modelledin this paper The brain appears to adhere to two principles of functional organization namelysegregation of function within cortical areas and sub-areas and the integration of these areasin perceptual synthesis cognition and sensorimotor integration (Tononi et al 1994 Fristonet al 1995) The balance between preserving specialized dynamics unique to each arearequired by specialization and the coupling among these areas mediating functional integrationmay be achieved by the neuromodulatory mechanisms described above Significantlythe spontaneous variation in the numbers of degrees of freedom (for example whenlow-dimensional synchronization manifolds are shadowed) imparts a dynamic characteristicto this perspective that is precluded by linear mechanisms alone
Finally from a mechanistic perspective complexity and diversity of brain dynamics arerequired for selection of adaptive responses (Friston 2000b) There are many examples ofselective mechanisms operating within the brain All can be seen as performing an ascenton some objective function or fitness landscape These mechanisms require complex anditinerant dynamics to facilitate a thorough exploration of the landscape These landscapesmay range from those involved in perception (where the landscape is defined by predictionerror see eg Rao and Ballard (1998)) to those involved in value learning (where thelandscape is determined by the outcome of a behaviour eg in temporal difference models (seeeg Sutton and Barto 1990)) In short complexity is necessary to maintain an information-rich environment in which the brain can respond to sensory inputs An essential componentof this is the maintenance of diversity which is a prerequisite for selectionist mechanismsat many scales (Edelman 1993) Understanding the mechanistic basis of complex dynamicsas measured electrophysiologically may well depend on the sort of biologically plausiblemodels used in this paper This understanding may be important for both basic and clinicalneuroscience (eg Tass 2002)
The present study supplements and advances on other research in this field The studyof complex phase synchronization and clustering in neural oscillator networks similar tothe present model has previously been the subject of formal mathematical study Forexample Terman and Wang (1995) studied synchronization of blocks of neural oscillatorsthat were subject to perturbation by connected lsquostimulirsquo Somers and Kopell (1995) studiedthe emergence of both synchronization and travelling waves in networks of coupled neuraloscillators In the work of Terman and Wang the local dynamics were based on neurallsquocaricaturersquo models such as the FitzHughndashNagumo equations and are hence more abstract thanthe current study The approach of Somers and Kopell only required that the local dynamicsexhibit fast and relaxation phases and is hence more abstract still In both studies onlyperiodic dynamics are studied Complex patterns of chaos synchronization in neural arrayshave been characterized by nonlinear methods (Kaneko 1998) and their putative contributionto both learning (Nakajima and Ueda 1996) and perception (Zhao and Macau 2001 Breakspear2001) explored However these studies employed models such as the logistic equation andtent map which are quite abstract in comparison to the present study On the other handHansel and Sompolinsky (1996) studied the role of chaos synchronization in a model ofvisual perception in a cortical hypercolumn using a very similar approach to that adoptedhere but with the additional complexity that synaptic delays were incorporated However inthis and earlier work (Hansel and Sompolinsky 1992) only linear measures of dynamics andsynchrony (specifically the auto- and cross-correlation functions) were employed The analysisof chaotic synchrony in the study of Larter et al (1999) was based only upon observation of
730 M Breakspear et al
the Euclidean difference between variables from two nodes within a larger array Hence thesignificance of the present work is that it provides a quantitative nonlinear analysis of complexchaos synchronization in a biophysical model of cortical dynamics Such research is requiredin order to bridge the gap between quantitative nonlinear research into synchronization andcomputation in abstract models of neural arrays and qualitative or linear analysis in morebiophysically plausible models
Acknowledgments
The authors would like to thank Peter Robinson for assistance with the manuscript and R Larterfor helpful advice MB is a recipient of a NSW Institute of Psychiatry Research Grant anda University of Sydney SESQUI postdoctoral fellowship KJF is supported by the WellcomeTrust
References
Abarbanel H Rulkov N and Sushchik M 1996 Generalised synchronization of chaos the auxiliary system approachPhys Rev E 53 4528ndash35
Afraimovich V Verichev N and Rabinovich M 1986 Stochastic synchronization of oscillation in dissipative systemsRadiophys Quantum Electron 29 795ndash801
Breakspear M 2001 Perception of odours by a nonlinear model of the olfactory bulb Int J Neural Syst 11 101ndash24Breakspear M 2002 Nonlinear phase desynchronisation in human electroencephalographic data Human Brain Mapping
15 175ndash98Breakspear M and Terry J R 2002a Nonlinear interdependence in neural systems motivation theory and relevance
Int J Neurosci 112 1163ndash84Breakspear M and Terry J R 2002b Detection and description of nonlinear interdependence in normal multichannel
human EEG Clin Neurophysiol 113 735ndash53Breakspear M and Terry J R 2002c Topographic organization of nonlinear interdependence in multichannel human
EEG Neuroimage 16 822ndash35Destexhe A Rudolph M Fellous J M and Sejnowski T J 2001 Fluctuating synaptic conductances recreate in vivo-like
activity in neocortical neurons Neuroscience 107 13ndash24Douglas R J Mahowald M A and Martin K A 1996 Microarchitecture of neocortical columns Brain TheorymdashBiological
Basis and Computational Principles ed A Aertsen and V Braitenberg (Amsterdam Elsevier)Eckhorn R Bauer R Jordan W Brosch M Kruse W Munk M and Reitboeck H J 1988 Coherent oscillations a
mechanism of feature linking in the visual cortex Multiple electrode and correlation analyses in the cat BiolCybern 60 121ndash30
Edelman G M 1993 Neural Darwinism selection and reentrant signalling in higher brain function Neuron 10 115ndash25Engel A K Kreiter A K Konig P and Singer W 1991 Synchronization of oscillatory neuronal responses between
striate and extrastriate visual cortical areas of the cat Proc Natl Acad Sci USA 88 6048ndash52Friston K J 2000a The labile brain I Neuronal transients and nonlinear coupling Phil Trans R Soc B 355 215ndash36Friston K J 2000b The labile brain II Transients complexity and selection Phil Trans R Soc B 355 237ndash52Friston K J Tononi G Sporns O and Edelman G 1995 Characterising the complexity of neuronal interactions Human
Brain Mapping 3 302ndash14Fujisaka H and Yamada T 1983 Stability theory of synchronized motion in coupled-oscillator system Prog Theor
Phys 69 32ndash47Gatlin L 1972 Information Theory and the Living System (New York Columbia University Press)Gray C M Konig P Engel A and Singer W 1989 Oscillatory responses in cat visual cortex exhibit inter-columnar
synchronization which reflects global stimulus properties Science 338 334ndash7Gray C M and Singer W 1989 Stimulus specific neuronal oscillations in orientation columns of cat visual cortex Proc
Natl Acad Sci USA 86 1698ndash702Haig A R Gordon E Wright J J Meares R A and Bahramali H 2000 Synchronous cortical gamma-band activity in
task-relevant cognition Neuroreport 11 669ndash75Hansel D and Sompolinsky H 1992 Synchronization and computation in a chaotic neural network Phys Rev Lett 68
718ndash21
Synchronization in a model of neuronal dynamics 731
Hansel D and Sompolinsky H 1996 Chaos and synchrony in a model of a hypercolumn in visual cortex J ComputNeurosci 3 7ndash4
Heagy J Carroll T and Pecora L 1998 Desynchronization by periodic orbits Phys Rev E 57 R1253ndash6Hunt B Ott E and Yorke J 1997 Differentiable synchronization of chaos Phys Rev E 55 4029ndash34Josic K 1998 Invariant manifolds and synchronization of coupled dynamical systems Phys Rev Lett 80 3053ndash6Kandel E Schwartz J and Jessell T 2000 Principles of Neural Science (New York McGraw-Hill)Kaneko K 1998 On the strength of attractors in a high-dimensional system Physica D 124 322ndash44Kocarev L Parlitz U and Brown R 2000 Robust synchronization of chaotic systems Phys Rev E 61 3716ndash20Larter R Speelman B and Worth R M 1999 A coupled ordinary differential equation lattice model for the simulation
of epileptic seizures Chaos 9 795ndash804Lumer E Edelman G and Tononi G 1997a Neural dynamics in a model of the thalamocortical system I Layers loops
and the emergence of fast synchronous oscillations Cereb Cortex 7 207ndash27Lumer E Edelman G and Tononi G 1997b Neural dynamics in a model of the thalamocortical system II The role of
neural synchrony tested through perturbations of spike timing Cereb Cortex 7 228ndash36Magleby K 1987 Short-term modulation at synapses Synaptic Function ed G M Edelman W E Gall and
W M Cowan (New York Wiley)Maistrenko Y Maistrenko V Popovich A and Mosekilde E 1998 Transverse instability and riddled basins in a system
of two coupled logistic maps Phys Rev E 57 2713ndash24Morris C and Lecar H 1981 Voltage oscillations in the barnacle giant muscle fiber Biophys J 35 193ndash213Mountcastle V B 1997 The columnar organization of the neocortex Brain 120 701ndash22Mountcastle V B 1998 The Cerebral Cortex (Cambridge MA Harvard University Press)Nakajima H and Ueda Y 1996 Riddled basins of the optimal states in learning dynamical systems Physica D 99 35ndash44Nunez P L 1995 Neocortical Dynamics and Human Brain Rhythms (Oxford Oxford University Press)Osipov G V Pikovsky A S Rosenblum M G and Kurths J 1997 Phase synchronization effects in a lattice of nonidentical
Rossler oscillators Phys Rev E 55 2353ndash61Ott E and Sommerer J 1994 Blowout bifurcations the occurrence of riddled basins and onndashoff intermittency Phys
Lett A 188 39ndash47Pecora L and Carroll T 1990 Synchronization in chaotic systems Phys Rev Lett 64 821ndash4Pikovsky A and Grassberger P 1991 Symmetry breaking bifurcation for coupled chaotic attractors J Phys A Math
Gen 24 4587ndash97Rao R P and Ballard D H 1998 Predictive coding in the visual cortex a functional interpretation of some extra-classical
receptive field effects Nature Neurosci 2 79ndash87Robinson P A Wright J J and Rennie C R 1998 Synchronous oscillations in the cerebral cortex Phys Rev E 57
4578ndash88Rodriguez E George N Lachauz J P Martinerie J Renault B and Varela F J 1999 Perceptionrsquos shadow long distance
synchronization of human brain activity Nature 397 430ndash3Rosenblum M Pikovsky A and Kurths J 1996 Phase synchronization of chaotic oscillators Phys Rev Lett 76 1804ndash7Rosenblum M Pikovsky A and Kurths J 1997 From phase to lag synchronization in coupled chaotic oscillators Phys
Rev Lett 78 4193ndash6Rulkov N and Sushchik M 1997 Robustness of synchronized chaotic oscillations Int J Bifurc Chaos 7 625ndash43Rulkov N Sushchik M Tsimring L and Abarbanel H 1995 Generalized synchronization of chaos in unidirectionally
coupled chaotic systems Phys Rev E 51 980ndash94Schiff S So P Chang T Burke R and Sauer T 1996 Detecting dynamical interdependence and generalized synchrony
through mutual prediction in a neural ensemble Phys Rev E 54 6708ndash24Silberstein R B 1995 Neuromodulations of neocortical dynamics Neocortical Dynamics and Human EEG Rhythms
ed P L Nunez (Oxford Oxford University Press)Singer W 1995 Putative functions of temporal correlations in neocortical processing Large-Scale Neuronal Theories
of the Brain ed C Koch and J Davis (London MIT Press)Somers D and Kopell N 1995 Waves and synchrony in networks of oscillators of relaxation and non-relaxation type
Physica D 89 169ndash83Sutton R S and Barto A G 1990 Time derivative models of Pavlovian reinforcement Learning and Computational
Neuroscience Foundations of Adaptive Networks ed M Gabriel and J Moore (Cambridge MA MIT Press) pp497ndash538
Szentagothai J 1983 The modular architectonic principle of neural centers Rev Physiol Biochem Pharmacol 9811ndash61
Tallon-Baudry C Betrand O and Fischer C 2001 Oscillatory synchrony between human extrastriate areas during visualshort-term memory maintenance J Neurosci 21 RC177(1ndash5)
Tass P A 2002 Desynchronization of brain rhythms with soft phase-resetting techniques Biol Cybern 87 102ndash15
732 M Breakspear et al
Tass P Rosenblum M Weule J Kurths J Pikovsky A Volkmann J Schnitzler A and Freund H 1998 Detection of nmphase locking from noisy data application to magnetoencephalography Phys Rev Lett 81 3291ndash4
Terman D and Wang D 1995 Global competition and local cooperation in a network of neural oscillators Physica D81 148ndash76
Terry J and Breakspear M 2003 An improved algorithm for the detection of dynamical interdependence in bivariatetime-series Biol Cybern 88 129ndash36
Tononi G Sporns O and Edelman G M 1994 A measure for brain complexity relating functional segregation andintegration in the nervous system Proc Natl Acad Sci USA 91 5033ndash7
von der Malsburg C 1985 Nervous structures with dynamical links Phys Chem 89 703ndash10Wang X 1991 Period-doublings to chaos in a simple neural network an analytic proof Complex Syst 5 425ndash41Wennekers T and Pasemann F 1996 Synchronous chaos in high dimensional modular neural networks Int J Bifurc
Chaos 6 2055ndash67Zaks M A Parks E H Rosenblum M G and Kurths J 1999 Alternative locking ratios in imperfect phase synchronization
Phys Rev Lett 82 4228ndash31Zhao L and Macau E N 2001 A network of dynamically coupled chaotic maps for scene segmentation IEEE Trans
Neural Netw 12 1375ndash85
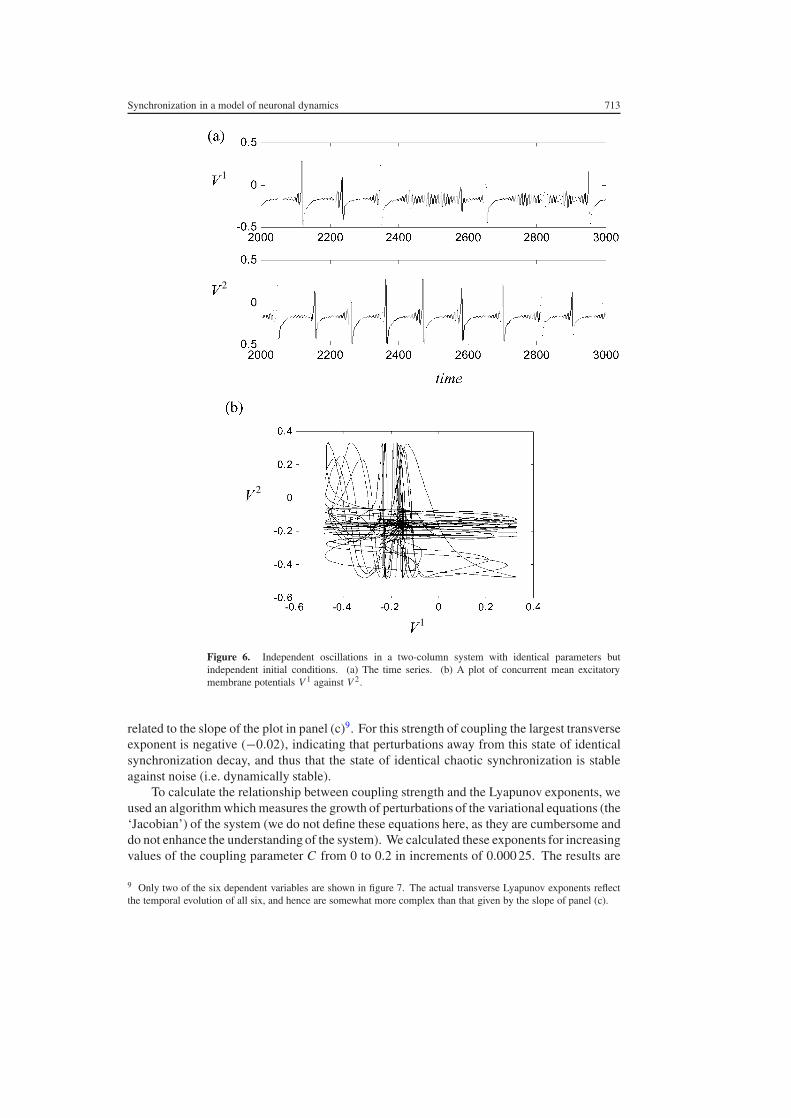
Synchronization in a model of neuronal dynamics 713
1
2
2
1
Figure 6 Independent oscillations in a two-column system with identical parameters butindependent initial conditions (a) The time series (b) A plot of concurrent mean excitatorymembrane potentials V 1 against V 2
related to the slope of the plot in panel (c)9 For this strength of coupling the largest transverseexponent is negative (minus002) indicating that perturbations away from this state of identicalsynchronization decay and thus that the state of identical chaotic synchronization is stableagainst noise (ie dynamically stable)
To calculate the relationship between coupling strength and the Lyapunov exponents weused an algorithm which measures the growth of perturbations of the variational equations (thelsquoJacobianrsquo) of the system (we do not define these equations here as they are cumbersome anddo not enhance the understanding of the system) We calculated these exponents for increasingvalues of the coupling parameter C from 0 to 02 in increments of 0000 25 The results are
9 Only two of the six dependent variables are shown in figure 7 The actual transverse Lyapunov exponents reflectthe temporal evolution of all six and hence are somewhat more complex than that given by the slope of panel (c)
714 M Breakspear et al
1
2
Figure 7 Identical synchronization in the same system as in figure 7 but with C = 02 (a) Thetime series (b) A plot of concurrent mean excitatory membrane potentials V 1 against V 2 (c) Thelogarithm of the Euclidean distance between V 1 and V 2 versus time (linear)
Synchronization in a model of neuronal dynamics 715
Figure 8 Lyapunov spectra for a coupled system with δV = 07 and C varying from 0 to 02 insteps of 0000 33 At each value of C all Lyapunov exponents are calculated Least-squares linesof best fit were then fitted to the results for each exponent Solid lines indicate transverse exponents(λ) and dashed lines indicate the tangential exponents () Panel (a) shows all three transverseand three tangential exponents Panel (b) only shows the largest two of each
illustrated in figure 8 Panel (a) shows all exponents For illustrative purposes only the mostpositive four are plotted in panel (b) In both panels the tangential exponents () are fitted (inthe least-squares sense) with a dashed line and the transverse ones (λ) are fitted with a solidline They are labelled 1ndash3 in order of decreasing positivity The important features are asfollows For the range of coupling strengths that we consider the largest tangential Lyapunovexponent is positive which indicates the presence of chaotic behaviour within the manifoldWhen examining the line of best fit for this exponent we see that this exponent does not varywhen altering the coupling parameter This is true for all three tangential exponents and verifiesthat the changes in the coupling parameter do not alter the dynamics on the synchronizationmanifold (that is the coupling parameter is normal)
716 M Breakspear et al
The largest transverse Lyapunov exponent (denoted as λ1 in the figure) is a particularlyimportant exponent since it governs the stability of the attractor in the synchronization manifoldwith respect to perturbations out of the manifold If this exponent is negative then we expectsmall perturbations of trajectories to reconverge to the manifold in finite time If however thisexponent is positive then we would instead expect perturbed trajectories to diverge from themanifold This change in local behaviour is termed a bifurcation and the point at which itoccurs the bifurcation point It is apparent in figure 8 that the zero crossing of this exponentmdashthe bifurcation pointmdashoccurs at approximately C = 01 in the present system This typeof bifurcation has been termed a blowout bifurcation since once transverse stability is losttrajectories are lsquoblown outrsquo from the low-dimensional synchronization manifold into the fullphase space (Ott and Sommerer 1994) Hence in the present setting a blowout bifurcation isobserved if C falls below 01
We note two other observations Increasing rNMDA which corresponds to increasing thepopulation of NMDA receptors decreases the coupling strength required to achieve stablesynchronization On the other hand increasing the overall excitatory-to-excitatory synapticconnectivity (aee) has the opposite effect This is because the effect of local excitatory feedbackincreases the tangential Lyapunov exponents which has the effect of lsquosplittingrsquo the synchronizeddynamics
In the above simulations all the parameters of both systems were equal In realphysiological systems such symmetry between systems is clearly impossible We nowconsider the case in which the two columns have different structures A simple way toincorporate this whilst at the same time preserving (as much as possible) simple dynamics is tovary the parameter δV for each column This has the effect of destroying any symmetry-forcedinvariant manifolds and so the simple case of synchronization corresponding to identicallyevolving dynamics will not exist in this case However more exotic forms of synchronizationare still possible and we discuss these next
42 Generalized chaotic synchronization
In order to maintain consistency with the literature we study non-linear interdependence in thesetting where the systems are asymmetrical but the coupling is unidirectional This is easilyachieved in the present system by making C directed and setting C21 = 0 In this settingwe refer to the lsquofreersquo system as the driver and the lsquoslavedrsquo system as the response We firstask the question what is the effect of introducing a slight parameter mismatch into a systemexhibiting stable chaos synchronization An example of this is presented in figure 9 whereC12 = 04 and parameter mismatch has been incorporated by setting δV = 07 in the driversystem and δV = 0699 in the response system In panel (a) the time series appear qualitativelyvery similar However in panel (b) where the difference between the systems is plotted it isapparent that the separation between the two systems is of the same order of magnitude as thesize of each attractor In panel (c) the effect of the parameter mismatch on the synchronizationmanifold can be seen as a transformation away from the symmetry manifold
It has been proposed that in such a setting there exists a continuous functional relationshipbetween the dynamics of the driver and response systems
x2(t) = ψ(x1(t)) (10)
where xi = V i W i Z i This situation has been referred to in the literature as generalizedsynchronization (Afraimovich et al 1986 Rulkov et al 1995) The properties of ψ andthe topology and stability of the synchronization manifold in this context have not yetbeen fully elucidated although some significant results have been achieved For example
Synchronization in a model of neuronal dynamics 717
1
2
2
1
Figure 9 Generalized synchronization in an asymmetrical driverndashresponse system (a) The timeseries of the two systems (b) The distance between the two systems (c) A plot of concurrent meanexcitatory membrane potentials V 1 against V 2 The diagonal represents the symmetry manifold
718 M Breakspear et al
Hunt et al (1997) Josic (1998) Kocarev et al (2000) have shown that the generalizedsynchronization manifold is smooth (differentiable) if and only if it is normally hyperbolic(ie the largest transverse Lyapunov exponent is less than the smallest tangential Lyapunovexponent prior to the introduction of parameter mismatch) Such a condition is met in thepresent setting when C = 04 hence the smoothness of the manifold in panel (c)
However subsequent to the introduction of parameter mismatch we will not be able touse transverse and tangential Lyapunov exponents to detect such synchronization since thesynchronization manifold may possess a complex topology for which it is a non-trivial (atbest) or impossible (at worse) task to define transverse and tangential directions For examplethe manifold may be smooth as in figure 9(c) but have a fractal topology If the dynamics arenot normally hyperbolic then the manifold may be fractal but also non-differentiable (Huntet al 1997) and hence will not have a unique tangent direction at any point Consequently wewill not be able to compute tangential or transverse Lyapunov exponents and hence determinethe stability of the synchronization manifold However other techniques for detecting thepresence of generalized synchronization in complex systems have evolved including theauxiliary system approach (Abarbanel et al 1996) and non-linear interdependence (Schiffet al 1996) which are now explored
421 The auxiliary system approach The auxiliary system is a third subsystem which isa replica of the original response system but coupled independently to the driver system andcommencing from independent initial conditions It is possible to demonstrate that showingthe existence of generalized synchronizationmdashthat equation (10) is satisfiedmdashis equivalent toshowing that the response system and the auxiliary system evolve identically (even though theyare not directly coupled) To determine whether or not this is the case reduces to an analysisof the Lyapunov exponents transverse to the symmetry manifold with respect to the responsesystem and the auxiliary system
Whilst this is a somewhat artificial test for generalized synchronization since theproduction of an auxiliary system for a physical system would be impossible such a testis useful in numerical and theoretical studies In figure 10 we present an example of theauxiliary system approach where the parameter δV = 07 for column 1 and 068 for column 2We consider two values of the coupling C = 0025 and 01 The auxiliary system is simplya copy of the second column beginning from an arbitrary initial state In the first instancewhere C = 0025 we notice that even after allowing for an initial transient state the secondcolumn and the auxiliary do not behave in identical fashions (figure 10(a)) However whenC = 01 we observe (figure 10(b)) that the second column and the auxiliary do behave innear-identical manners after an initial transient and hence conclude that for the particularvalue of the coupling strength considered equation (10) is satisfied and hence generalizedsynchronization between the first and second columns occurs
422 Nonlinear interdependence The question remains as to how we may detect thepresence of generalized synchronization in a practical setting As we mentioned aboveit is not possible to use an auxiliary-system-based approach to detect the presence ofgeneralized synchronization Instead we must consider techniques for detecting suchbehaviour from experimental or numerical time series data Algorithms of this type areoften referred to as detecting non-linear interdependence Nonlinear interdependence is ageneral term encompassing systems where the behaviour of one non-linear system affectsthe behaviour of another More specifically it can be defined as occurring if the nullhypothesis that the interdependence between two signals is purely linear can be rejectedat conventional levels (usually 95) of statistical significance hence it is an experimental
Synchronization in a model of neuronal dynamics 719
2
1
2
1
Figure 10 Concurrent evolution of the response system V 2 and lsquoauxiliaryrsquo system V 1 (a) C =0025 (b) C = 01
term (eg Schiff et al 1996) Whilst this does not strictly correspond to the existence of afunctional relationship between the two systems such a relationship is typically used tomotivate the techniques for its detection Schiff et al (1996) extended the logic of non-linearprediction algorithms from autonomous to coupled systems by exploiting the continuity ofsuch a relationship and hence devised the first method of detecting non-linear interdependencein time series data The basis of the technique is using the forward evolution of a phase spacevectorrsquos neighbours to predict the forward evolution of that vector and the forward evolutionof the concurrent vector of the other (possibly interdependent) system If the error of such aprocedure is better on average than chance (the chance error is normalized to equal one) thennon-linear interdependence is said to be present
720 M Breakspear et al
V1
V2
Figure 11 Experimental and numerical examples of weak generalized non-linear synchronizationin the presence of measurement noise (a) Human scalp EEG data from posterior bipolar scalpelectrodes (O1P3ndashO2P4) (b) Weakly coupled aperiodic data in the present model with additivewhite noise (with 5 of the amplitude of the model signal) Parameters δ1
V = 0605 δ2V = 061
aee = 036 C = 007
A variation of this algorithm optimized to detect weak non-linear interdependence (Terryand Breakspear 2003) was used recently to demonstrate that weak non-linear interdependence isoccasionally present in the scalp EEG of healthy resting human subjects (Breakspear and Terry2002a 2002b 2002c) An example of an epoch of lsquonon-linearly independentrsquo EEG obtainedusing this technique is presented in figure 11(a) For comparison output from the currentmodel is presented in figure 11(b) The model parameters were set such that the dynamicswas still aperiodic but the intermittent large-amplitude bursts were suppressed and had weakmutual coupling (C = 007) Extraneous noise to simulate the effect of extracranial muscleactivity contaminating the EEG data has been added to this time series It should be notedthat the amplitudes and time scales of the modelrsquos variables are dimensionless Nonethelessthe model facilitates qualititative interpretation of experimental findings
Synchronization in a model of neuronal dynamics 721
43 Chaotic phase synchronization
Whereas the concept of generalized synchronization was mathematically motivated and assuch more suited to theoretical investigations of systems the concept of phase synchronizationderives from a more practical signal processing approach Phase synchronization occurs inboth symmetric and asymmetric settings but is often more useful in the asymmetric case (andin experimental data)
Unfortunately the phase of a chaotic signal is typically ambiguous and can be defined in anumber of different ways For example in the case of the coherent Rossler attractor (Osipovet al 1997) the phase may be defined as the arctangent of two of the variables of the systemMore generally it can sometimes be defined as the angle that an orbit subtends with the centreof an attractor It is also possible to extract the phase of a chaotic signal from time seriesdata by use of the Hilbert transform This has enabled the demonstration of chaotic phasesynchronization in a number of coupled asymmetrical systems (Rosenblum et al 1996 1997Zaks et al 1999) Although such an approach does not guarantee that the phase will be uniqueit does permit the practical analysis of phase in noisy time series (Tass et al 1998)
To illustrate phase synchronization in two coupled columns we again vary the parameterδV which if the columns are viewed as oscillators acts to detune them from a commonfrequency An important question is that of the criteria on the basis of which we concludewhether phase synchronization occurs We use a simple locking criterion whereby phasesynchronization is said to occur if
|θ1(t)minus θ2(t)| lt K (11)
where is θi the phase of column i and K is a constant Notice that this does not mean thatsynchronization implies equality of phases merely that their difference is bounded
An example of phase synchronization with δV = 065 for column 1 and 066 for column 2is given in figure 12 In this figure rNMDA = 03 Qualitatively some sort of phase coherenceis evident in the time series panel (a) This was confirmed by plotting the sine of thephase difference which has been extracted using the Hilbert transform panel (b) Clearly|sin(θ1 minus θ2)| lt K and hence |θ1 minus θ2| is bounded It can also be seen that the phase-synchronized dynamics occurs on a highly structured manifold depicted in figure 12(c)However unlike the case for the generalized synchronization manifold depicted in figure 9 itis evident that this manifold is not a simple transformation of the symmetry manifold
It is also possible to see lsquobimodalrsquo stability whereby a system jumps betweenapproximately zero phase difference and a phase lag phase synchronization of π An exampleis depicted in figure 12(d) At approximately 2000 s the system can be seen to jumpspontaneously from the first to the second of these modes The Z -variable is plotted forgraphical purposes
5 Intermittency and desynchronization
51 Desynchronization in an array of two systems
Diminishing the strength of coupling across the blowout bifurcation does not lead to a simpletransition from synchronous to asynchronous oscillations but is instead associated withcomplex temporal behaviours Specifically when the largest transverse Lyapunov exponent isclose to zero the coupled systems exhibit epochs of synchronous behaviours punctuated bysharp bursts of desynchronization This is observed in systems exhibiting identicalgeneralizedand phase synchronization An example of such a phenomenon is given in figure 13 The timeseries panel (a) shows synchronous dynamics except at time = 1000 where it can be seenthat the two time series desynchronize briefly This is evident in panel (b) as a disruption in the
722 M Breakspear et al
2
1
Figure 12 Phase-synchronized coupled columns (a) The time series of V 1 (solid) and V 2 (b) Thesine of the phase difference obtained through Hilbert transformation of the time series in panel (a)(c) The phase synchronization manifold in the V 1ndashV 2 plane (d) The time series of Z (solid) andZ showing a phase slip of π between two modes
Synchronization in a model of neuronal dynamics 723
Figure 13 An example of phase desynchronization in coupled asymmetric systems with C = 01Time series plots of (a) each V -variable and (b) their Euclidean distance (c) The temporal evolutionof the phase difference between the two systems (d) An example of the phase difference evolutionover a longer time interval
724 M Breakspear et al
orderly evolution of the distance between the two systems and in panel (c) where the evolutionof the phase difference reveals a brief violation of the criterion |φ1 minus φ2| lt constant Anexample of the phase difference evolution over a longer time interval is shown in figure 13(d)Note that this is a three-dimensional plot with sin(φ1 minus φ2) and cos(φ1 minus φ2) as they- and z-axes Hence phase desynchronizations are evident as complete revolutions aroundthe cylindrical phase space in which this variable evolves It can be seen that these phenomenaoccur at irregular intervals in the temporal domain
This behaviour can be explained by considering the nature of a chaotic attractor In thestatistical sense an arbitrary initial condition is expected to lead to the chaotic behaviour onthe attractor Consequently when we calculate the Lyapunov exponents of the system theseexponents correspond to those of the chaotic attractor Typically however there are an infinitenumber of unstable periodic orbits embedded within the attractor If the path of a trajectorybecomes close to one of these orbits it may lsquoshadowrsquo the orbit for a brief period of time Eachof these orbits has a corresponding set of Lyapunov exponents which (like the attractor) can bedivided into tangential and transverse exponents These also vary when we change the couplingparameter but are typically not the same as the exponents of the attractor Hence the largesttransverse exponent of the attractor may be negative yet there may be positive transverseexponents associated with periodic orbits Considering all of these exponents together wewould therefore only expect to observe complete stability of the synchronization manifold inthe case where all of the transverse Lyapunov exponents are negative Bursting behaviourwill be observed when the one of these exponents becomes positive typically correspondingto a periodic orbit of low order (Pikovsky and Grassberger 1991 Rulkov and Sushchik 1997Maistrenko et al 1998 Heagy et al 1998) Due to the complex structure of such periodic orbitsthe desynchronous bursts occur across a wide range of timescales
In figure 14 we present the results of a long integration (16times105 time units) in which thelengths of sequential synchronous epochs of phase synchronization are studied The parametervalues were δV = 065 for column 1 and 0645 for column 2 with coupling strength10 equal to0125 In panel (a) the power spectrum of these sequential lengths is plotted The whiteness ofthis spectrum indicates that the lengths of consecutive synchronous epochs are independentThis is due to the high-dimensional lsquomixingrsquo effect of desynchronization In panel (b) thelengths of the synchronous epochs have been sorted again into the order of increasing lengthThe results for two coupling strengths are plotted C = 0125 (solid) and 0135 (dashed)Several features of this plot are of note Firstly the lengths of the synchronous epochsare distributed over almost three orders of magnitude Secondly for both plots there is anapproximately log-linear relationship between the length of an epoch and its relative orderThis reflects an approximately exponential relationship between the length of a synchronousepoch and its likelihood of occurrence Thirdly this relationship is skewed toward longersynchronous epochs for the stronger coupling strengths
This type of signal with patches of lsquoorderrsquo occurring over many timescales interruptedby bursts of disorder can be shown to have a high information capacity (Gatlin 1972)Moreover complex phase desynchronization may make an infrequent yet robust contributionto the dynamical pattern of the alpha rhythm in scalp EEG data for resting human subjects(Breakspear 2002) It is tempting to postulate that such a lsquoweakly stablersquo dynamics may assistin the generation of the rich and fluid mental associations that spontaneously emerge in restingcognitive states Although this phenomenon has been previously described in a variety of othersystems to our knowledge this is the first description in a biophysical model of neocortex
10 The introduction of parameter mismatch causes an increase in the strength of coupling required for stablesynchronization above the value of 01 required in the symmetric case
Synchronization in a model of neuronal dynamics 725
Figure 14 Properties of sequential epochs of phase-synchronous behaviour over a long integrationtime δ1
V = 065 δ2V = 0645 aee = 04 (a) The flat (white) power spectrum (b) Synchronous
epochs sorted in order of increasing length (solid C = 0125 dashed C = 0134)
52 Synchronization and complex dynamics in larger arrays
The synchronous and complex phenomena described above are also observed in arrays ofcoupled systems with N gt 2 Expanding the size of the array permits complex dynamicsin the spatial in addition to the temporal domain However the basic principles governingthe type and stability of synchronized motions are the same In this section a few of thepossible behaviours are discussed Physiological variation across the array is modelled bymodifying the parameter δV = 07 in each array by adding a random number of maximummagnitude 001
In figure 15(a) an array of N = 20 uncoupled (C = 0) systems is shown The lack ofsynchronization in this time series is evident By comparison the rapid transition to completesynchronization is evident in figure 15(b) when C = 03 In fact 19 of the 20 subsystems
726 M Breakspear et al
(b)
(c)
(a)
Figure 15 Synchronization phenomena in an array of coupled cortical columns (a) N = 20C = 0 (b) N = 20 C = 03 (c) N = 6 C = 01 (d) N = 12 C = 009 (e) N = 12C = 012 A small perturbation has been applied to one of the columns at t = 2500 s (f) Evolutionof the phase difference between two of the columns in panel (e) For clarity the W -variable isplotted in panel (c) and the Z -variable in (e)
have phase synchronized within one single cycle Intermediate values of coupling strength areassociated with a rich variety of phenomena For example subsystems with the most similarparameter values typically synchronize into clusters of various sizes An example of this isillustrated in figure 15(c) Note that there is no synchronization between the two differentclusters each containing three synchronized subsystems
Another complex behaviour occurs when subsystems transiently synchronize into largeclusters then spontaneously desynchronize again Hence the dynamics of the entire arrayjumps between different transient dynamics characterized by varying degrees of freedom Anexample of this phenomenon is given in figure 15(d) In addition it is possible to inducedesynchronization across an entire array by applying a small perturbation to one columnfor one time step An example is illustrated in figure 15(e) A small perturbation has been
Synchronization in a model of neuronal dynamics 727
Figure 15 (Continued)
added to one column after 2500 time units It can be seen that the entire array subsequentlydesynchronizes This illustrates the sensitivity of the dynamics to external stimuli Finally it ispossible to lsquoinducersquo synchronization between selected subsystems by increasing the strength ofcoupling between them selectively This allows an extra level of complexity to be introducedinto the model For example it may allow targeted information transfer between a partialnetwork of the entire array
As discussed in section 51 a univariate signal which jumps between epochs of lowentropy (high repetition) and bursts of high entropy (high variability) can be shown to havehigh information capacity This is because the repetition of certain patterns (as in the English
728 M Breakspear et al
language) permits error detectability Bursts of high entropy act in the opposite directionto increase message variability With a multivariate signal in addition to optimizing theinformation capacity within each signal the overall information capacity can be increasedthrough the type of itinerant synchronization briefly illustrated in this section Synchronizationacross an entire array increases the message reliability (since a single lsquoerrant letterrsquo in onesignal can be easily detected) Conversely desynchronization into multiple clusters increasesmessage variability since each cluster will produce independent information Hence a neuralarray which switches dynamically between a range of cluster configurations can be seen asadaptive in the sense that high synchronous states can be expressed when error detectabilityis a priority (such as during quick and reliable execution of a familiar task) but the array canrapidly desynchronize if variability becomes salient (for example if the environment suddenlybecomes novel) A formal operationalized demonstration of this principle is to be the subjectof future work
6 Discussion
In this paper we present a dynamical model for activity in an array of weakly coupledlocal neural assemblies The model incorporates the fundamental principles governing cellmembrane potential as determined by voltage- and ligand-gated ion channels In this wayit allows a reasonable amount of physiological complexity to be incorporated includingsynaptic activation and receptor subtypeswithout recourse to overwhelming detail Numericalanalysis of this model reveals a transition from independent oscillations to stable chaoticsynchronization depending on the balance of local versus long-range connectivity andbiophysical heterogeneity These excitatory couplings are subject to ascending subcorticalneuromodulation arising principally from monoaminergic nuclei such as Locus Coeruleus(Silberstein 1995) Neuromodulation of extrinsic connections was incorporated into the modelvia a coupling parameter C The stability of the synchronization is determined by the valueof the largest transverse Lyapunov exponent which in turn depends on the coupling parameterC If the coupling is weakened with the result that this exponent becomes positive a blowoutbifurcation occurs and independent oscillations re-emerge In the vicinity of the transitionnon-linear desynchronizations with complex temporal and spatial properties are observed
It is pleasing that complex and itinerant behaviour reminiscent of real neuronaldynamics emerges through such a simple manipulation of a single dynamic control parameterFurthermore the ubiquitous projections of classical neuromodulatory systems in real brainssuggest a clear biological mechanism by which the brain could self-organize throughmoderating the complexity of its own dynamics There are many reasons that the complicateddynamics of the sort demonstrated in this paper may be neurobiologically important Wemention four
First the presence of multiple clusters of phase-synchronized subsystems as discussedabove may be a uniquely non-linear mechanism facilitating the lsquocarriagersquo and distinction ofmultiple concurrent dynamic cell assemblies (and hence cognitive processing) This may beimportant for perceptual grouping binding and transitions as suggested by others (eg von derMalsburg 1985 Gray and Singer 1989)
Second the facility to decouple and reconfigure dynamic cell assemblies througha neuromodulatory control parameter may be important for avoiding low-entropysynchronization manifolds In the absence of itinerancy synchronization would renderneuronal dynamics incapable of representing changes in sensory input or indeed cognitive stateFor example in the model it was observed that small perturbations to one column were ableto induce a global change in neuronal dynamics by disrupting a state of weak synchronization
Synchronization in a model of neuronal dynamics 729
Inevitably alternative dynamic configurations emerge epitomizing the importance of lsquolabilersquobrain dynamics (Friston 2000a)
Thirdly from a purely functionalist point of view the balance between functionalspecialization and functional integration may rely upon the sorts of mechanism modelledin this paper The brain appears to adhere to two principles of functional organization namelysegregation of function within cortical areas and sub-areas and the integration of these areasin perceptual synthesis cognition and sensorimotor integration (Tononi et al 1994 Fristonet al 1995) The balance between preserving specialized dynamics unique to each arearequired by specialization and the coupling among these areas mediating functional integrationmay be achieved by the neuromodulatory mechanisms described above Significantlythe spontaneous variation in the numbers of degrees of freedom (for example whenlow-dimensional synchronization manifolds are shadowed) imparts a dynamic characteristicto this perspective that is precluded by linear mechanisms alone
Finally from a mechanistic perspective complexity and diversity of brain dynamics arerequired for selection of adaptive responses (Friston 2000b) There are many examples ofselective mechanisms operating within the brain All can be seen as performing an ascenton some objective function or fitness landscape These mechanisms require complex anditinerant dynamics to facilitate a thorough exploration of the landscape These landscapesmay range from those involved in perception (where the landscape is defined by predictionerror see eg Rao and Ballard (1998)) to those involved in value learning (where thelandscape is determined by the outcome of a behaviour eg in temporal difference models (seeeg Sutton and Barto 1990)) In short complexity is necessary to maintain an information-rich environment in which the brain can respond to sensory inputs An essential componentof this is the maintenance of diversity which is a prerequisite for selectionist mechanismsat many scales (Edelman 1993) Understanding the mechanistic basis of complex dynamicsas measured electrophysiologically may well depend on the sort of biologically plausiblemodels used in this paper This understanding may be important for both basic and clinicalneuroscience (eg Tass 2002)
The present study supplements and advances on other research in this field The studyof complex phase synchronization and clustering in neural oscillator networks similar tothe present model has previously been the subject of formal mathematical study Forexample Terman and Wang (1995) studied synchronization of blocks of neural oscillatorsthat were subject to perturbation by connected lsquostimulirsquo Somers and Kopell (1995) studiedthe emergence of both synchronization and travelling waves in networks of coupled neuraloscillators In the work of Terman and Wang the local dynamics were based on neurallsquocaricaturersquo models such as the FitzHughndashNagumo equations and are hence more abstract thanthe current study The approach of Somers and Kopell only required that the local dynamicsexhibit fast and relaxation phases and is hence more abstract still In both studies onlyperiodic dynamics are studied Complex patterns of chaos synchronization in neural arrayshave been characterized by nonlinear methods (Kaneko 1998) and their putative contributionto both learning (Nakajima and Ueda 1996) and perception (Zhao and Macau 2001 Breakspear2001) explored However these studies employed models such as the logistic equation andtent map which are quite abstract in comparison to the present study On the other handHansel and Sompolinsky (1996) studied the role of chaos synchronization in a model ofvisual perception in a cortical hypercolumn using a very similar approach to that adoptedhere but with the additional complexity that synaptic delays were incorporated However inthis and earlier work (Hansel and Sompolinsky 1992) only linear measures of dynamics andsynchrony (specifically the auto- and cross-correlation functions) were employed The analysisof chaotic synchrony in the study of Larter et al (1999) was based only upon observation of
730 M Breakspear et al
the Euclidean difference between variables from two nodes within a larger array Hence thesignificance of the present work is that it provides a quantitative nonlinear analysis of complexchaos synchronization in a biophysical model of cortical dynamics Such research is requiredin order to bridge the gap between quantitative nonlinear research into synchronization andcomputation in abstract models of neural arrays and qualitative or linear analysis in morebiophysically plausible models
Acknowledgments
The authors would like to thank Peter Robinson for assistance with the manuscript and R Larterfor helpful advice MB is a recipient of a NSW Institute of Psychiatry Research Grant anda University of Sydney SESQUI postdoctoral fellowship KJF is supported by the WellcomeTrust
References
Abarbanel H Rulkov N and Sushchik M 1996 Generalised synchronization of chaos the auxiliary system approachPhys Rev E 53 4528ndash35
Afraimovich V Verichev N and Rabinovich M 1986 Stochastic synchronization of oscillation in dissipative systemsRadiophys Quantum Electron 29 795ndash801
Breakspear M 2001 Perception of odours by a nonlinear model of the olfactory bulb Int J Neural Syst 11 101ndash24Breakspear M 2002 Nonlinear phase desynchronisation in human electroencephalographic data Human Brain Mapping
15 175ndash98Breakspear M and Terry J R 2002a Nonlinear interdependence in neural systems motivation theory and relevance
Int J Neurosci 112 1163ndash84Breakspear M and Terry J R 2002b Detection and description of nonlinear interdependence in normal multichannel
human EEG Clin Neurophysiol 113 735ndash53Breakspear M and Terry J R 2002c Topographic organization of nonlinear interdependence in multichannel human
EEG Neuroimage 16 822ndash35Destexhe A Rudolph M Fellous J M and Sejnowski T J 2001 Fluctuating synaptic conductances recreate in vivo-like
activity in neocortical neurons Neuroscience 107 13ndash24Douglas R J Mahowald M A and Martin K A 1996 Microarchitecture of neocortical columns Brain TheorymdashBiological
Basis and Computational Principles ed A Aertsen and V Braitenberg (Amsterdam Elsevier)Eckhorn R Bauer R Jordan W Brosch M Kruse W Munk M and Reitboeck H J 1988 Coherent oscillations a
mechanism of feature linking in the visual cortex Multiple electrode and correlation analyses in the cat BiolCybern 60 121ndash30
Edelman G M 1993 Neural Darwinism selection and reentrant signalling in higher brain function Neuron 10 115ndash25Engel A K Kreiter A K Konig P and Singer W 1991 Synchronization of oscillatory neuronal responses between
striate and extrastriate visual cortical areas of the cat Proc Natl Acad Sci USA 88 6048ndash52Friston K J 2000a The labile brain I Neuronal transients and nonlinear coupling Phil Trans R Soc B 355 215ndash36Friston K J 2000b The labile brain II Transients complexity and selection Phil Trans R Soc B 355 237ndash52Friston K J Tononi G Sporns O and Edelman G 1995 Characterising the complexity of neuronal interactions Human
Brain Mapping 3 302ndash14Fujisaka H and Yamada T 1983 Stability theory of synchronized motion in coupled-oscillator system Prog Theor
Phys 69 32ndash47Gatlin L 1972 Information Theory and the Living System (New York Columbia University Press)Gray C M Konig P Engel A and Singer W 1989 Oscillatory responses in cat visual cortex exhibit inter-columnar
synchronization which reflects global stimulus properties Science 338 334ndash7Gray C M and Singer W 1989 Stimulus specific neuronal oscillations in orientation columns of cat visual cortex Proc
Natl Acad Sci USA 86 1698ndash702Haig A R Gordon E Wright J J Meares R A and Bahramali H 2000 Synchronous cortical gamma-band activity in
task-relevant cognition Neuroreport 11 669ndash75Hansel D and Sompolinsky H 1992 Synchronization and computation in a chaotic neural network Phys Rev Lett 68
718ndash21
Synchronization in a model of neuronal dynamics 731
Hansel D and Sompolinsky H 1996 Chaos and synchrony in a model of a hypercolumn in visual cortex J ComputNeurosci 3 7ndash4
Heagy J Carroll T and Pecora L 1998 Desynchronization by periodic orbits Phys Rev E 57 R1253ndash6Hunt B Ott E and Yorke J 1997 Differentiable synchronization of chaos Phys Rev E 55 4029ndash34Josic K 1998 Invariant manifolds and synchronization of coupled dynamical systems Phys Rev Lett 80 3053ndash6Kandel E Schwartz J and Jessell T 2000 Principles of Neural Science (New York McGraw-Hill)Kaneko K 1998 On the strength of attractors in a high-dimensional system Physica D 124 322ndash44Kocarev L Parlitz U and Brown R 2000 Robust synchronization of chaotic systems Phys Rev E 61 3716ndash20Larter R Speelman B and Worth R M 1999 A coupled ordinary differential equation lattice model for the simulation
of epileptic seizures Chaos 9 795ndash804Lumer E Edelman G and Tononi G 1997a Neural dynamics in a model of the thalamocortical system I Layers loops
and the emergence of fast synchronous oscillations Cereb Cortex 7 207ndash27Lumer E Edelman G and Tononi G 1997b Neural dynamics in a model of the thalamocortical system II The role of
neural synchrony tested through perturbations of spike timing Cereb Cortex 7 228ndash36Magleby K 1987 Short-term modulation at synapses Synaptic Function ed G M Edelman W E Gall and
W M Cowan (New York Wiley)Maistrenko Y Maistrenko V Popovich A and Mosekilde E 1998 Transverse instability and riddled basins in a system
of two coupled logistic maps Phys Rev E 57 2713ndash24Morris C and Lecar H 1981 Voltage oscillations in the barnacle giant muscle fiber Biophys J 35 193ndash213Mountcastle V B 1997 The columnar organization of the neocortex Brain 120 701ndash22Mountcastle V B 1998 The Cerebral Cortex (Cambridge MA Harvard University Press)Nakajima H and Ueda Y 1996 Riddled basins of the optimal states in learning dynamical systems Physica D 99 35ndash44Nunez P L 1995 Neocortical Dynamics and Human Brain Rhythms (Oxford Oxford University Press)Osipov G V Pikovsky A S Rosenblum M G and Kurths J 1997 Phase synchronization effects in a lattice of nonidentical
Rossler oscillators Phys Rev E 55 2353ndash61Ott E and Sommerer J 1994 Blowout bifurcations the occurrence of riddled basins and onndashoff intermittency Phys
Lett A 188 39ndash47Pecora L and Carroll T 1990 Synchronization in chaotic systems Phys Rev Lett 64 821ndash4Pikovsky A and Grassberger P 1991 Symmetry breaking bifurcation for coupled chaotic attractors J Phys A Math
Gen 24 4587ndash97Rao R P and Ballard D H 1998 Predictive coding in the visual cortex a functional interpretation of some extra-classical
receptive field effects Nature Neurosci 2 79ndash87Robinson P A Wright J J and Rennie C R 1998 Synchronous oscillations in the cerebral cortex Phys Rev E 57
4578ndash88Rodriguez E George N Lachauz J P Martinerie J Renault B and Varela F J 1999 Perceptionrsquos shadow long distance
synchronization of human brain activity Nature 397 430ndash3Rosenblum M Pikovsky A and Kurths J 1996 Phase synchronization of chaotic oscillators Phys Rev Lett 76 1804ndash7Rosenblum M Pikovsky A and Kurths J 1997 From phase to lag synchronization in coupled chaotic oscillators Phys
Rev Lett 78 4193ndash6Rulkov N and Sushchik M 1997 Robustness of synchronized chaotic oscillations Int J Bifurc Chaos 7 625ndash43Rulkov N Sushchik M Tsimring L and Abarbanel H 1995 Generalized synchronization of chaos in unidirectionally
coupled chaotic systems Phys Rev E 51 980ndash94Schiff S So P Chang T Burke R and Sauer T 1996 Detecting dynamical interdependence and generalized synchrony
through mutual prediction in a neural ensemble Phys Rev E 54 6708ndash24Silberstein R B 1995 Neuromodulations of neocortical dynamics Neocortical Dynamics and Human EEG Rhythms
ed P L Nunez (Oxford Oxford University Press)Singer W 1995 Putative functions of temporal correlations in neocortical processing Large-Scale Neuronal Theories
of the Brain ed C Koch and J Davis (London MIT Press)Somers D and Kopell N 1995 Waves and synchrony in networks of oscillators of relaxation and non-relaxation type
Physica D 89 169ndash83Sutton R S and Barto A G 1990 Time derivative models of Pavlovian reinforcement Learning and Computational
Neuroscience Foundations of Adaptive Networks ed M Gabriel and J Moore (Cambridge MA MIT Press) pp497ndash538
Szentagothai J 1983 The modular architectonic principle of neural centers Rev Physiol Biochem Pharmacol 9811ndash61
Tallon-Baudry C Betrand O and Fischer C 2001 Oscillatory synchrony between human extrastriate areas during visualshort-term memory maintenance J Neurosci 21 RC177(1ndash5)
Tass P A 2002 Desynchronization of brain rhythms with soft phase-resetting techniques Biol Cybern 87 102ndash15
732 M Breakspear et al
Tass P Rosenblum M Weule J Kurths J Pikovsky A Volkmann J Schnitzler A and Freund H 1998 Detection of nmphase locking from noisy data application to magnetoencephalography Phys Rev Lett 81 3291ndash4
Terman D and Wang D 1995 Global competition and local cooperation in a network of neural oscillators Physica D81 148ndash76
Terry J and Breakspear M 2003 An improved algorithm for the detection of dynamical interdependence in bivariatetime-series Biol Cybern 88 129ndash36
Tononi G Sporns O and Edelman G M 1994 A measure for brain complexity relating functional segregation andintegration in the nervous system Proc Natl Acad Sci USA 91 5033ndash7
von der Malsburg C 1985 Nervous structures with dynamical links Phys Chem 89 703ndash10Wang X 1991 Period-doublings to chaos in a simple neural network an analytic proof Complex Syst 5 425ndash41Wennekers T and Pasemann F 1996 Synchronous chaos in high dimensional modular neural networks Int J Bifurc
Chaos 6 2055ndash67Zaks M A Parks E H Rosenblum M G and Kurths J 1999 Alternative locking ratios in imperfect phase synchronization
Phys Rev Lett 82 4228ndash31Zhao L and Macau E N 2001 A network of dynamically coupled chaotic maps for scene segmentation IEEE Trans
Neural Netw 12 1375ndash85
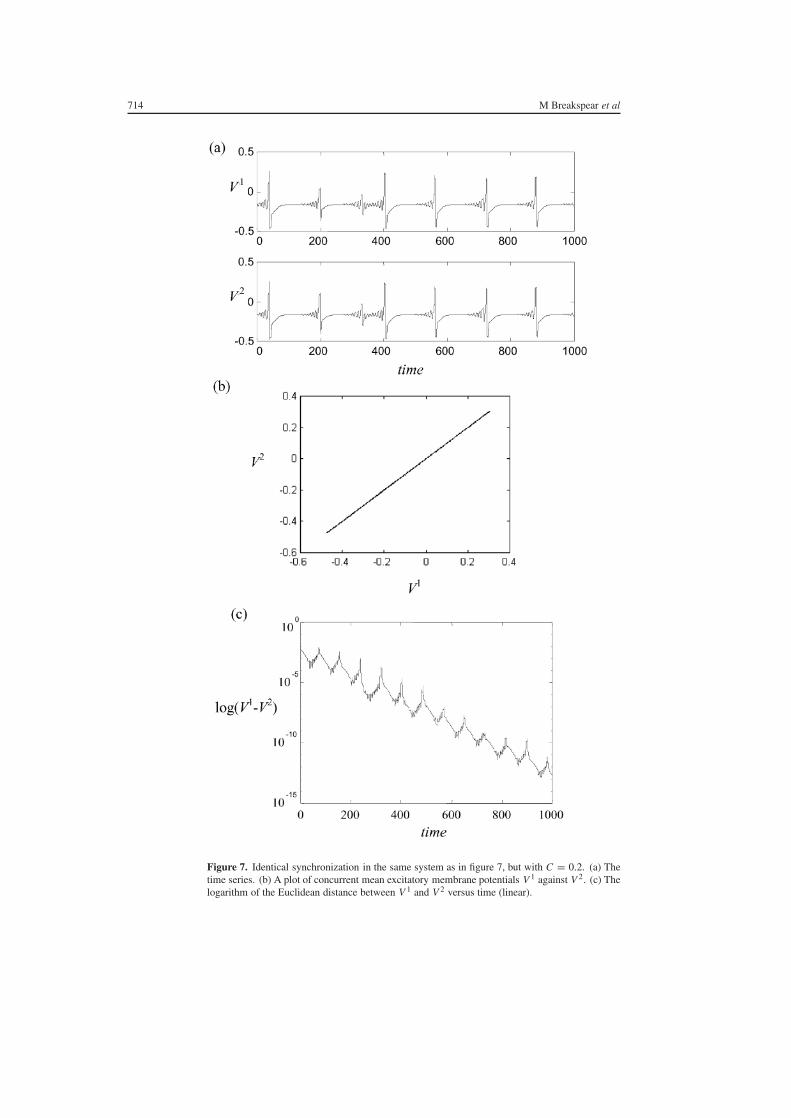
714 M Breakspear et al
1
2
Figure 7 Identical synchronization in the same system as in figure 7 but with C = 02 (a) Thetime series (b) A plot of concurrent mean excitatory membrane potentials V 1 against V 2 (c) Thelogarithm of the Euclidean distance between V 1 and V 2 versus time (linear)
Synchronization in a model of neuronal dynamics 715
Figure 8 Lyapunov spectra for a coupled system with δV = 07 and C varying from 0 to 02 insteps of 0000 33 At each value of C all Lyapunov exponents are calculated Least-squares linesof best fit were then fitted to the results for each exponent Solid lines indicate transverse exponents(λ) and dashed lines indicate the tangential exponents () Panel (a) shows all three transverseand three tangential exponents Panel (b) only shows the largest two of each
illustrated in figure 8 Panel (a) shows all exponents For illustrative purposes only the mostpositive four are plotted in panel (b) In both panels the tangential exponents () are fitted (inthe least-squares sense) with a dashed line and the transverse ones (λ) are fitted with a solidline They are labelled 1ndash3 in order of decreasing positivity The important features are asfollows For the range of coupling strengths that we consider the largest tangential Lyapunovexponent is positive which indicates the presence of chaotic behaviour within the manifoldWhen examining the line of best fit for this exponent we see that this exponent does not varywhen altering the coupling parameter This is true for all three tangential exponents and verifiesthat the changes in the coupling parameter do not alter the dynamics on the synchronizationmanifold (that is the coupling parameter is normal)
716 M Breakspear et al
The largest transverse Lyapunov exponent (denoted as λ1 in the figure) is a particularlyimportant exponent since it governs the stability of the attractor in the synchronization manifoldwith respect to perturbations out of the manifold If this exponent is negative then we expectsmall perturbations of trajectories to reconverge to the manifold in finite time If however thisexponent is positive then we would instead expect perturbed trajectories to diverge from themanifold This change in local behaviour is termed a bifurcation and the point at which itoccurs the bifurcation point It is apparent in figure 8 that the zero crossing of this exponentmdashthe bifurcation pointmdashoccurs at approximately C = 01 in the present system This typeof bifurcation has been termed a blowout bifurcation since once transverse stability is losttrajectories are lsquoblown outrsquo from the low-dimensional synchronization manifold into the fullphase space (Ott and Sommerer 1994) Hence in the present setting a blowout bifurcation isobserved if C falls below 01
We note two other observations Increasing rNMDA which corresponds to increasing thepopulation of NMDA receptors decreases the coupling strength required to achieve stablesynchronization On the other hand increasing the overall excitatory-to-excitatory synapticconnectivity (aee) has the opposite effect This is because the effect of local excitatory feedbackincreases the tangential Lyapunov exponents which has the effect of lsquosplittingrsquo the synchronizeddynamics
In the above simulations all the parameters of both systems were equal In realphysiological systems such symmetry between systems is clearly impossible We nowconsider the case in which the two columns have different structures A simple way toincorporate this whilst at the same time preserving (as much as possible) simple dynamics is tovary the parameter δV for each column This has the effect of destroying any symmetry-forcedinvariant manifolds and so the simple case of synchronization corresponding to identicallyevolving dynamics will not exist in this case However more exotic forms of synchronizationare still possible and we discuss these next
42 Generalized chaotic synchronization
In order to maintain consistency with the literature we study non-linear interdependence in thesetting where the systems are asymmetrical but the coupling is unidirectional This is easilyachieved in the present system by making C directed and setting C21 = 0 In this settingwe refer to the lsquofreersquo system as the driver and the lsquoslavedrsquo system as the response We firstask the question what is the effect of introducing a slight parameter mismatch into a systemexhibiting stable chaos synchronization An example of this is presented in figure 9 whereC12 = 04 and parameter mismatch has been incorporated by setting δV = 07 in the driversystem and δV = 0699 in the response system In panel (a) the time series appear qualitativelyvery similar However in panel (b) where the difference between the systems is plotted it isapparent that the separation between the two systems is of the same order of magnitude as thesize of each attractor In panel (c) the effect of the parameter mismatch on the synchronizationmanifold can be seen as a transformation away from the symmetry manifold
It has been proposed that in such a setting there exists a continuous functional relationshipbetween the dynamics of the driver and response systems
x2(t) = ψ(x1(t)) (10)
where xi = V i W i Z i This situation has been referred to in the literature as generalizedsynchronization (Afraimovich et al 1986 Rulkov et al 1995) The properties of ψ andthe topology and stability of the synchronization manifold in this context have not yetbeen fully elucidated although some significant results have been achieved For example
Synchronization in a model of neuronal dynamics 717
1
2
2
1
Figure 9 Generalized synchronization in an asymmetrical driverndashresponse system (a) The timeseries of the two systems (b) The distance between the two systems (c) A plot of concurrent meanexcitatory membrane potentials V 1 against V 2 The diagonal represents the symmetry manifold
718 M Breakspear et al
Hunt et al (1997) Josic (1998) Kocarev et al (2000) have shown that the generalizedsynchronization manifold is smooth (differentiable) if and only if it is normally hyperbolic(ie the largest transverse Lyapunov exponent is less than the smallest tangential Lyapunovexponent prior to the introduction of parameter mismatch) Such a condition is met in thepresent setting when C = 04 hence the smoothness of the manifold in panel (c)
However subsequent to the introduction of parameter mismatch we will not be able touse transverse and tangential Lyapunov exponents to detect such synchronization since thesynchronization manifold may possess a complex topology for which it is a non-trivial (atbest) or impossible (at worse) task to define transverse and tangential directions For examplethe manifold may be smooth as in figure 9(c) but have a fractal topology If the dynamics arenot normally hyperbolic then the manifold may be fractal but also non-differentiable (Huntet al 1997) and hence will not have a unique tangent direction at any point Consequently wewill not be able to compute tangential or transverse Lyapunov exponents and hence determinethe stability of the synchronization manifold However other techniques for detecting thepresence of generalized synchronization in complex systems have evolved including theauxiliary system approach (Abarbanel et al 1996) and non-linear interdependence (Schiffet al 1996) which are now explored
421 The auxiliary system approach The auxiliary system is a third subsystem which isa replica of the original response system but coupled independently to the driver system andcommencing from independent initial conditions It is possible to demonstrate that showingthe existence of generalized synchronizationmdashthat equation (10) is satisfiedmdashis equivalent toshowing that the response system and the auxiliary system evolve identically (even though theyare not directly coupled) To determine whether or not this is the case reduces to an analysisof the Lyapunov exponents transverse to the symmetry manifold with respect to the responsesystem and the auxiliary system
Whilst this is a somewhat artificial test for generalized synchronization since theproduction of an auxiliary system for a physical system would be impossible such a testis useful in numerical and theoretical studies In figure 10 we present an example of theauxiliary system approach where the parameter δV = 07 for column 1 and 068 for column 2We consider two values of the coupling C = 0025 and 01 The auxiliary system is simplya copy of the second column beginning from an arbitrary initial state In the first instancewhere C = 0025 we notice that even after allowing for an initial transient state the secondcolumn and the auxiliary do not behave in identical fashions (figure 10(a)) However whenC = 01 we observe (figure 10(b)) that the second column and the auxiliary do behave innear-identical manners after an initial transient and hence conclude that for the particularvalue of the coupling strength considered equation (10) is satisfied and hence generalizedsynchronization between the first and second columns occurs
422 Nonlinear interdependence The question remains as to how we may detect thepresence of generalized synchronization in a practical setting As we mentioned aboveit is not possible to use an auxiliary-system-based approach to detect the presence ofgeneralized synchronization Instead we must consider techniques for detecting suchbehaviour from experimental or numerical time series data Algorithms of this type areoften referred to as detecting non-linear interdependence Nonlinear interdependence is ageneral term encompassing systems where the behaviour of one non-linear system affectsthe behaviour of another More specifically it can be defined as occurring if the nullhypothesis that the interdependence between two signals is purely linear can be rejectedat conventional levels (usually 95) of statistical significance hence it is an experimental
Synchronization in a model of neuronal dynamics 719
2
1
2
1
Figure 10 Concurrent evolution of the response system V 2 and lsquoauxiliaryrsquo system V 1 (a) C =0025 (b) C = 01
term (eg Schiff et al 1996) Whilst this does not strictly correspond to the existence of afunctional relationship between the two systems such a relationship is typically used tomotivate the techniques for its detection Schiff et al (1996) extended the logic of non-linearprediction algorithms from autonomous to coupled systems by exploiting the continuity ofsuch a relationship and hence devised the first method of detecting non-linear interdependencein time series data The basis of the technique is using the forward evolution of a phase spacevectorrsquos neighbours to predict the forward evolution of that vector and the forward evolutionof the concurrent vector of the other (possibly interdependent) system If the error of such aprocedure is better on average than chance (the chance error is normalized to equal one) thennon-linear interdependence is said to be present
720 M Breakspear et al
V1
V2
Figure 11 Experimental and numerical examples of weak generalized non-linear synchronizationin the presence of measurement noise (a) Human scalp EEG data from posterior bipolar scalpelectrodes (O1P3ndashO2P4) (b) Weakly coupled aperiodic data in the present model with additivewhite noise (with 5 of the amplitude of the model signal) Parameters δ1
V = 0605 δ2V = 061
aee = 036 C = 007
A variation of this algorithm optimized to detect weak non-linear interdependence (Terryand Breakspear 2003) was used recently to demonstrate that weak non-linear interdependence isoccasionally present in the scalp EEG of healthy resting human subjects (Breakspear and Terry2002a 2002b 2002c) An example of an epoch of lsquonon-linearly independentrsquo EEG obtainedusing this technique is presented in figure 11(a) For comparison output from the currentmodel is presented in figure 11(b) The model parameters were set such that the dynamicswas still aperiodic but the intermittent large-amplitude bursts were suppressed and had weakmutual coupling (C = 007) Extraneous noise to simulate the effect of extracranial muscleactivity contaminating the EEG data has been added to this time series It should be notedthat the amplitudes and time scales of the modelrsquos variables are dimensionless Nonethelessthe model facilitates qualititative interpretation of experimental findings
Synchronization in a model of neuronal dynamics 721
43 Chaotic phase synchronization
Whereas the concept of generalized synchronization was mathematically motivated and assuch more suited to theoretical investigations of systems the concept of phase synchronizationderives from a more practical signal processing approach Phase synchronization occurs inboth symmetric and asymmetric settings but is often more useful in the asymmetric case (andin experimental data)
Unfortunately the phase of a chaotic signal is typically ambiguous and can be defined in anumber of different ways For example in the case of the coherent Rossler attractor (Osipovet al 1997) the phase may be defined as the arctangent of two of the variables of the systemMore generally it can sometimes be defined as the angle that an orbit subtends with the centreof an attractor It is also possible to extract the phase of a chaotic signal from time seriesdata by use of the Hilbert transform This has enabled the demonstration of chaotic phasesynchronization in a number of coupled asymmetrical systems (Rosenblum et al 1996 1997Zaks et al 1999) Although such an approach does not guarantee that the phase will be uniqueit does permit the practical analysis of phase in noisy time series (Tass et al 1998)
To illustrate phase synchronization in two coupled columns we again vary the parameterδV which if the columns are viewed as oscillators acts to detune them from a commonfrequency An important question is that of the criteria on the basis of which we concludewhether phase synchronization occurs We use a simple locking criterion whereby phasesynchronization is said to occur if
|θ1(t)minus θ2(t)| lt K (11)
where is θi the phase of column i and K is a constant Notice that this does not mean thatsynchronization implies equality of phases merely that their difference is bounded
An example of phase synchronization with δV = 065 for column 1 and 066 for column 2is given in figure 12 In this figure rNMDA = 03 Qualitatively some sort of phase coherenceis evident in the time series panel (a) This was confirmed by plotting the sine of thephase difference which has been extracted using the Hilbert transform panel (b) Clearly|sin(θ1 minus θ2)| lt K and hence |θ1 minus θ2| is bounded It can also be seen that the phase-synchronized dynamics occurs on a highly structured manifold depicted in figure 12(c)However unlike the case for the generalized synchronization manifold depicted in figure 9 itis evident that this manifold is not a simple transformation of the symmetry manifold
It is also possible to see lsquobimodalrsquo stability whereby a system jumps betweenapproximately zero phase difference and a phase lag phase synchronization of π An exampleis depicted in figure 12(d) At approximately 2000 s the system can be seen to jumpspontaneously from the first to the second of these modes The Z -variable is plotted forgraphical purposes
5 Intermittency and desynchronization
51 Desynchronization in an array of two systems
Diminishing the strength of coupling across the blowout bifurcation does not lead to a simpletransition from synchronous to asynchronous oscillations but is instead associated withcomplex temporal behaviours Specifically when the largest transverse Lyapunov exponent isclose to zero the coupled systems exhibit epochs of synchronous behaviours punctuated bysharp bursts of desynchronization This is observed in systems exhibiting identicalgeneralizedand phase synchronization An example of such a phenomenon is given in figure 13 The timeseries panel (a) shows synchronous dynamics except at time = 1000 where it can be seenthat the two time series desynchronize briefly This is evident in panel (b) as a disruption in the
722 M Breakspear et al
2
1
Figure 12 Phase-synchronized coupled columns (a) The time series of V 1 (solid) and V 2 (b) Thesine of the phase difference obtained through Hilbert transformation of the time series in panel (a)(c) The phase synchronization manifold in the V 1ndashV 2 plane (d) The time series of Z (solid) andZ showing a phase slip of π between two modes
Synchronization in a model of neuronal dynamics 723
Figure 13 An example of phase desynchronization in coupled asymmetric systems with C = 01Time series plots of (a) each V -variable and (b) their Euclidean distance (c) The temporal evolutionof the phase difference between the two systems (d) An example of the phase difference evolutionover a longer time interval
724 M Breakspear et al
orderly evolution of the distance between the two systems and in panel (c) where the evolutionof the phase difference reveals a brief violation of the criterion |φ1 minus φ2| lt constant Anexample of the phase difference evolution over a longer time interval is shown in figure 13(d)Note that this is a three-dimensional plot with sin(φ1 minus φ2) and cos(φ1 minus φ2) as they- and z-axes Hence phase desynchronizations are evident as complete revolutions aroundthe cylindrical phase space in which this variable evolves It can be seen that these phenomenaoccur at irregular intervals in the temporal domain
This behaviour can be explained by considering the nature of a chaotic attractor In thestatistical sense an arbitrary initial condition is expected to lead to the chaotic behaviour onthe attractor Consequently when we calculate the Lyapunov exponents of the system theseexponents correspond to those of the chaotic attractor Typically however there are an infinitenumber of unstable periodic orbits embedded within the attractor If the path of a trajectorybecomes close to one of these orbits it may lsquoshadowrsquo the orbit for a brief period of time Eachof these orbits has a corresponding set of Lyapunov exponents which (like the attractor) can bedivided into tangential and transverse exponents These also vary when we change the couplingparameter but are typically not the same as the exponents of the attractor Hence the largesttransverse exponent of the attractor may be negative yet there may be positive transverseexponents associated with periodic orbits Considering all of these exponents together wewould therefore only expect to observe complete stability of the synchronization manifold inthe case where all of the transverse Lyapunov exponents are negative Bursting behaviourwill be observed when the one of these exponents becomes positive typically correspondingto a periodic orbit of low order (Pikovsky and Grassberger 1991 Rulkov and Sushchik 1997Maistrenko et al 1998 Heagy et al 1998) Due to the complex structure of such periodic orbitsthe desynchronous bursts occur across a wide range of timescales
In figure 14 we present the results of a long integration (16times105 time units) in which thelengths of sequential synchronous epochs of phase synchronization are studied The parametervalues were δV = 065 for column 1 and 0645 for column 2 with coupling strength10 equal to0125 In panel (a) the power spectrum of these sequential lengths is plotted The whiteness ofthis spectrum indicates that the lengths of consecutive synchronous epochs are independentThis is due to the high-dimensional lsquomixingrsquo effect of desynchronization In panel (b) thelengths of the synchronous epochs have been sorted again into the order of increasing lengthThe results for two coupling strengths are plotted C = 0125 (solid) and 0135 (dashed)Several features of this plot are of note Firstly the lengths of the synchronous epochsare distributed over almost three orders of magnitude Secondly for both plots there is anapproximately log-linear relationship between the length of an epoch and its relative orderThis reflects an approximately exponential relationship between the length of a synchronousepoch and its likelihood of occurrence Thirdly this relationship is skewed toward longersynchronous epochs for the stronger coupling strengths
This type of signal with patches of lsquoorderrsquo occurring over many timescales interruptedby bursts of disorder can be shown to have a high information capacity (Gatlin 1972)Moreover complex phase desynchronization may make an infrequent yet robust contributionto the dynamical pattern of the alpha rhythm in scalp EEG data for resting human subjects(Breakspear 2002) It is tempting to postulate that such a lsquoweakly stablersquo dynamics may assistin the generation of the rich and fluid mental associations that spontaneously emerge in restingcognitive states Although this phenomenon has been previously described in a variety of othersystems to our knowledge this is the first description in a biophysical model of neocortex
10 The introduction of parameter mismatch causes an increase in the strength of coupling required for stablesynchronization above the value of 01 required in the symmetric case
Synchronization in a model of neuronal dynamics 725
Figure 14 Properties of sequential epochs of phase-synchronous behaviour over a long integrationtime δ1
V = 065 δ2V = 0645 aee = 04 (a) The flat (white) power spectrum (b) Synchronous
epochs sorted in order of increasing length (solid C = 0125 dashed C = 0134)
52 Synchronization and complex dynamics in larger arrays
The synchronous and complex phenomena described above are also observed in arrays ofcoupled systems with N gt 2 Expanding the size of the array permits complex dynamicsin the spatial in addition to the temporal domain However the basic principles governingthe type and stability of synchronized motions are the same In this section a few of thepossible behaviours are discussed Physiological variation across the array is modelled bymodifying the parameter δV = 07 in each array by adding a random number of maximummagnitude 001
In figure 15(a) an array of N = 20 uncoupled (C = 0) systems is shown The lack ofsynchronization in this time series is evident By comparison the rapid transition to completesynchronization is evident in figure 15(b) when C = 03 In fact 19 of the 20 subsystems
726 M Breakspear et al
(b)
(c)
(a)
Figure 15 Synchronization phenomena in an array of coupled cortical columns (a) N = 20C = 0 (b) N = 20 C = 03 (c) N = 6 C = 01 (d) N = 12 C = 009 (e) N = 12C = 012 A small perturbation has been applied to one of the columns at t = 2500 s (f) Evolutionof the phase difference between two of the columns in panel (e) For clarity the W -variable isplotted in panel (c) and the Z -variable in (e)
have phase synchronized within one single cycle Intermediate values of coupling strength areassociated with a rich variety of phenomena For example subsystems with the most similarparameter values typically synchronize into clusters of various sizes An example of this isillustrated in figure 15(c) Note that there is no synchronization between the two differentclusters each containing three synchronized subsystems
Another complex behaviour occurs when subsystems transiently synchronize into largeclusters then spontaneously desynchronize again Hence the dynamics of the entire arrayjumps between different transient dynamics characterized by varying degrees of freedom Anexample of this phenomenon is given in figure 15(d) In addition it is possible to inducedesynchronization across an entire array by applying a small perturbation to one columnfor one time step An example is illustrated in figure 15(e) A small perturbation has been
Synchronization in a model of neuronal dynamics 727
Figure 15 (Continued)
added to one column after 2500 time units It can be seen that the entire array subsequentlydesynchronizes This illustrates the sensitivity of the dynamics to external stimuli Finally it ispossible to lsquoinducersquo synchronization between selected subsystems by increasing the strength ofcoupling between them selectively This allows an extra level of complexity to be introducedinto the model For example it may allow targeted information transfer between a partialnetwork of the entire array
As discussed in section 51 a univariate signal which jumps between epochs of lowentropy (high repetition) and bursts of high entropy (high variability) can be shown to havehigh information capacity This is because the repetition of certain patterns (as in the English
728 M Breakspear et al
language) permits error detectability Bursts of high entropy act in the opposite directionto increase message variability With a multivariate signal in addition to optimizing theinformation capacity within each signal the overall information capacity can be increasedthrough the type of itinerant synchronization briefly illustrated in this section Synchronizationacross an entire array increases the message reliability (since a single lsquoerrant letterrsquo in onesignal can be easily detected) Conversely desynchronization into multiple clusters increasesmessage variability since each cluster will produce independent information Hence a neuralarray which switches dynamically between a range of cluster configurations can be seen asadaptive in the sense that high synchronous states can be expressed when error detectabilityis a priority (such as during quick and reliable execution of a familiar task) but the array canrapidly desynchronize if variability becomes salient (for example if the environment suddenlybecomes novel) A formal operationalized demonstration of this principle is to be the subjectof future work
6 Discussion
In this paper we present a dynamical model for activity in an array of weakly coupledlocal neural assemblies The model incorporates the fundamental principles governing cellmembrane potential as determined by voltage- and ligand-gated ion channels In this wayit allows a reasonable amount of physiological complexity to be incorporated includingsynaptic activation and receptor subtypeswithout recourse to overwhelming detail Numericalanalysis of this model reveals a transition from independent oscillations to stable chaoticsynchronization depending on the balance of local versus long-range connectivity andbiophysical heterogeneity These excitatory couplings are subject to ascending subcorticalneuromodulation arising principally from monoaminergic nuclei such as Locus Coeruleus(Silberstein 1995) Neuromodulation of extrinsic connections was incorporated into the modelvia a coupling parameter C The stability of the synchronization is determined by the valueof the largest transverse Lyapunov exponent which in turn depends on the coupling parameterC If the coupling is weakened with the result that this exponent becomes positive a blowoutbifurcation occurs and independent oscillations re-emerge In the vicinity of the transitionnon-linear desynchronizations with complex temporal and spatial properties are observed
It is pleasing that complex and itinerant behaviour reminiscent of real neuronaldynamics emerges through such a simple manipulation of a single dynamic control parameterFurthermore the ubiquitous projections of classical neuromodulatory systems in real brainssuggest a clear biological mechanism by which the brain could self-organize throughmoderating the complexity of its own dynamics There are many reasons that the complicateddynamics of the sort demonstrated in this paper may be neurobiologically important Wemention four
First the presence of multiple clusters of phase-synchronized subsystems as discussedabove may be a uniquely non-linear mechanism facilitating the lsquocarriagersquo and distinction ofmultiple concurrent dynamic cell assemblies (and hence cognitive processing) This may beimportant for perceptual grouping binding and transitions as suggested by others (eg von derMalsburg 1985 Gray and Singer 1989)
Second the facility to decouple and reconfigure dynamic cell assemblies througha neuromodulatory control parameter may be important for avoiding low-entropysynchronization manifolds In the absence of itinerancy synchronization would renderneuronal dynamics incapable of representing changes in sensory input or indeed cognitive stateFor example in the model it was observed that small perturbations to one column were ableto induce a global change in neuronal dynamics by disrupting a state of weak synchronization
Synchronization in a model of neuronal dynamics 729
Inevitably alternative dynamic configurations emerge epitomizing the importance of lsquolabilersquobrain dynamics (Friston 2000a)
Thirdly from a purely functionalist point of view the balance between functionalspecialization and functional integration may rely upon the sorts of mechanism modelledin this paper The brain appears to adhere to two principles of functional organization namelysegregation of function within cortical areas and sub-areas and the integration of these areasin perceptual synthesis cognition and sensorimotor integration (Tononi et al 1994 Fristonet al 1995) The balance between preserving specialized dynamics unique to each arearequired by specialization and the coupling among these areas mediating functional integrationmay be achieved by the neuromodulatory mechanisms described above Significantlythe spontaneous variation in the numbers of degrees of freedom (for example whenlow-dimensional synchronization manifolds are shadowed) imparts a dynamic characteristicto this perspective that is precluded by linear mechanisms alone
Finally from a mechanistic perspective complexity and diversity of brain dynamics arerequired for selection of adaptive responses (Friston 2000b) There are many examples ofselective mechanisms operating within the brain All can be seen as performing an ascenton some objective function or fitness landscape These mechanisms require complex anditinerant dynamics to facilitate a thorough exploration of the landscape These landscapesmay range from those involved in perception (where the landscape is defined by predictionerror see eg Rao and Ballard (1998)) to those involved in value learning (where thelandscape is determined by the outcome of a behaviour eg in temporal difference models (seeeg Sutton and Barto 1990)) In short complexity is necessary to maintain an information-rich environment in which the brain can respond to sensory inputs An essential componentof this is the maintenance of diversity which is a prerequisite for selectionist mechanismsat many scales (Edelman 1993) Understanding the mechanistic basis of complex dynamicsas measured electrophysiologically may well depend on the sort of biologically plausiblemodels used in this paper This understanding may be important for both basic and clinicalneuroscience (eg Tass 2002)
The present study supplements and advances on other research in this field The studyof complex phase synchronization and clustering in neural oscillator networks similar tothe present model has previously been the subject of formal mathematical study Forexample Terman and Wang (1995) studied synchronization of blocks of neural oscillatorsthat were subject to perturbation by connected lsquostimulirsquo Somers and Kopell (1995) studiedthe emergence of both synchronization and travelling waves in networks of coupled neuraloscillators In the work of Terman and Wang the local dynamics were based on neurallsquocaricaturersquo models such as the FitzHughndashNagumo equations and are hence more abstract thanthe current study The approach of Somers and Kopell only required that the local dynamicsexhibit fast and relaxation phases and is hence more abstract still In both studies onlyperiodic dynamics are studied Complex patterns of chaos synchronization in neural arrayshave been characterized by nonlinear methods (Kaneko 1998) and their putative contributionto both learning (Nakajima and Ueda 1996) and perception (Zhao and Macau 2001 Breakspear2001) explored However these studies employed models such as the logistic equation andtent map which are quite abstract in comparison to the present study On the other handHansel and Sompolinsky (1996) studied the role of chaos synchronization in a model ofvisual perception in a cortical hypercolumn using a very similar approach to that adoptedhere but with the additional complexity that synaptic delays were incorporated However inthis and earlier work (Hansel and Sompolinsky 1992) only linear measures of dynamics andsynchrony (specifically the auto- and cross-correlation functions) were employed The analysisof chaotic synchrony in the study of Larter et al (1999) was based only upon observation of
730 M Breakspear et al
the Euclidean difference between variables from two nodes within a larger array Hence thesignificance of the present work is that it provides a quantitative nonlinear analysis of complexchaos synchronization in a biophysical model of cortical dynamics Such research is requiredin order to bridge the gap between quantitative nonlinear research into synchronization andcomputation in abstract models of neural arrays and qualitative or linear analysis in morebiophysically plausible models
Acknowledgments
The authors would like to thank Peter Robinson for assistance with the manuscript and R Larterfor helpful advice MB is a recipient of a NSW Institute of Psychiatry Research Grant anda University of Sydney SESQUI postdoctoral fellowship KJF is supported by the WellcomeTrust
References
Abarbanel H Rulkov N and Sushchik M 1996 Generalised synchronization of chaos the auxiliary system approachPhys Rev E 53 4528ndash35
Afraimovich V Verichev N and Rabinovich M 1986 Stochastic synchronization of oscillation in dissipative systemsRadiophys Quantum Electron 29 795ndash801
Breakspear M 2001 Perception of odours by a nonlinear model of the olfactory bulb Int J Neural Syst 11 101ndash24Breakspear M 2002 Nonlinear phase desynchronisation in human electroencephalographic data Human Brain Mapping
15 175ndash98Breakspear M and Terry J R 2002a Nonlinear interdependence in neural systems motivation theory and relevance
Int J Neurosci 112 1163ndash84Breakspear M and Terry J R 2002b Detection and description of nonlinear interdependence in normal multichannel
human EEG Clin Neurophysiol 113 735ndash53Breakspear M and Terry J R 2002c Topographic organization of nonlinear interdependence in multichannel human
EEG Neuroimage 16 822ndash35Destexhe A Rudolph M Fellous J M and Sejnowski T J 2001 Fluctuating synaptic conductances recreate in vivo-like
activity in neocortical neurons Neuroscience 107 13ndash24Douglas R J Mahowald M A and Martin K A 1996 Microarchitecture of neocortical columns Brain TheorymdashBiological
Basis and Computational Principles ed A Aertsen and V Braitenberg (Amsterdam Elsevier)Eckhorn R Bauer R Jordan W Brosch M Kruse W Munk M and Reitboeck H J 1988 Coherent oscillations a
mechanism of feature linking in the visual cortex Multiple electrode and correlation analyses in the cat BiolCybern 60 121ndash30
Edelman G M 1993 Neural Darwinism selection and reentrant signalling in higher brain function Neuron 10 115ndash25Engel A K Kreiter A K Konig P and Singer W 1991 Synchronization of oscillatory neuronal responses between
striate and extrastriate visual cortical areas of the cat Proc Natl Acad Sci USA 88 6048ndash52Friston K J 2000a The labile brain I Neuronal transients and nonlinear coupling Phil Trans R Soc B 355 215ndash36Friston K J 2000b The labile brain II Transients complexity and selection Phil Trans R Soc B 355 237ndash52Friston K J Tononi G Sporns O and Edelman G 1995 Characterising the complexity of neuronal interactions Human
Brain Mapping 3 302ndash14Fujisaka H and Yamada T 1983 Stability theory of synchronized motion in coupled-oscillator system Prog Theor
Phys 69 32ndash47Gatlin L 1972 Information Theory and the Living System (New York Columbia University Press)Gray C M Konig P Engel A and Singer W 1989 Oscillatory responses in cat visual cortex exhibit inter-columnar
synchronization which reflects global stimulus properties Science 338 334ndash7Gray C M and Singer W 1989 Stimulus specific neuronal oscillations in orientation columns of cat visual cortex Proc
Natl Acad Sci USA 86 1698ndash702Haig A R Gordon E Wright J J Meares R A and Bahramali H 2000 Synchronous cortical gamma-band activity in
task-relevant cognition Neuroreport 11 669ndash75Hansel D and Sompolinsky H 1992 Synchronization and computation in a chaotic neural network Phys Rev Lett 68
718ndash21
Synchronization in a model of neuronal dynamics 731
Hansel D and Sompolinsky H 1996 Chaos and synchrony in a model of a hypercolumn in visual cortex J ComputNeurosci 3 7ndash4
Heagy J Carroll T and Pecora L 1998 Desynchronization by periodic orbits Phys Rev E 57 R1253ndash6Hunt B Ott E and Yorke J 1997 Differentiable synchronization of chaos Phys Rev E 55 4029ndash34Josic K 1998 Invariant manifolds and synchronization of coupled dynamical systems Phys Rev Lett 80 3053ndash6Kandel E Schwartz J and Jessell T 2000 Principles of Neural Science (New York McGraw-Hill)Kaneko K 1998 On the strength of attractors in a high-dimensional system Physica D 124 322ndash44Kocarev L Parlitz U and Brown R 2000 Robust synchronization of chaotic systems Phys Rev E 61 3716ndash20Larter R Speelman B and Worth R M 1999 A coupled ordinary differential equation lattice model for the simulation
of epileptic seizures Chaos 9 795ndash804Lumer E Edelman G and Tononi G 1997a Neural dynamics in a model of the thalamocortical system I Layers loops
and the emergence of fast synchronous oscillations Cereb Cortex 7 207ndash27Lumer E Edelman G and Tononi G 1997b Neural dynamics in a model of the thalamocortical system II The role of
neural synchrony tested through perturbations of spike timing Cereb Cortex 7 228ndash36Magleby K 1987 Short-term modulation at synapses Synaptic Function ed G M Edelman W E Gall and
W M Cowan (New York Wiley)Maistrenko Y Maistrenko V Popovich A and Mosekilde E 1998 Transverse instability and riddled basins in a system
of two coupled logistic maps Phys Rev E 57 2713ndash24Morris C and Lecar H 1981 Voltage oscillations in the barnacle giant muscle fiber Biophys J 35 193ndash213Mountcastle V B 1997 The columnar organization of the neocortex Brain 120 701ndash22Mountcastle V B 1998 The Cerebral Cortex (Cambridge MA Harvard University Press)Nakajima H and Ueda Y 1996 Riddled basins of the optimal states in learning dynamical systems Physica D 99 35ndash44Nunez P L 1995 Neocortical Dynamics and Human Brain Rhythms (Oxford Oxford University Press)Osipov G V Pikovsky A S Rosenblum M G and Kurths J 1997 Phase synchronization effects in a lattice of nonidentical
Rossler oscillators Phys Rev E 55 2353ndash61Ott E and Sommerer J 1994 Blowout bifurcations the occurrence of riddled basins and onndashoff intermittency Phys
Lett A 188 39ndash47Pecora L and Carroll T 1990 Synchronization in chaotic systems Phys Rev Lett 64 821ndash4Pikovsky A and Grassberger P 1991 Symmetry breaking bifurcation for coupled chaotic attractors J Phys A Math
Gen 24 4587ndash97Rao R P and Ballard D H 1998 Predictive coding in the visual cortex a functional interpretation of some extra-classical
receptive field effects Nature Neurosci 2 79ndash87Robinson P A Wright J J and Rennie C R 1998 Synchronous oscillations in the cerebral cortex Phys Rev E 57
4578ndash88Rodriguez E George N Lachauz J P Martinerie J Renault B and Varela F J 1999 Perceptionrsquos shadow long distance
synchronization of human brain activity Nature 397 430ndash3Rosenblum M Pikovsky A and Kurths J 1996 Phase synchronization of chaotic oscillators Phys Rev Lett 76 1804ndash7Rosenblum M Pikovsky A and Kurths J 1997 From phase to lag synchronization in coupled chaotic oscillators Phys
Rev Lett 78 4193ndash6Rulkov N and Sushchik M 1997 Robustness of synchronized chaotic oscillations Int J Bifurc Chaos 7 625ndash43Rulkov N Sushchik M Tsimring L and Abarbanel H 1995 Generalized synchronization of chaos in unidirectionally
coupled chaotic systems Phys Rev E 51 980ndash94Schiff S So P Chang T Burke R and Sauer T 1996 Detecting dynamical interdependence and generalized synchrony
through mutual prediction in a neural ensemble Phys Rev E 54 6708ndash24Silberstein R B 1995 Neuromodulations of neocortical dynamics Neocortical Dynamics and Human EEG Rhythms
ed P L Nunez (Oxford Oxford University Press)Singer W 1995 Putative functions of temporal correlations in neocortical processing Large-Scale Neuronal Theories
of the Brain ed C Koch and J Davis (London MIT Press)Somers D and Kopell N 1995 Waves and synchrony in networks of oscillators of relaxation and non-relaxation type
Physica D 89 169ndash83Sutton R S and Barto A G 1990 Time derivative models of Pavlovian reinforcement Learning and Computational
Neuroscience Foundations of Adaptive Networks ed M Gabriel and J Moore (Cambridge MA MIT Press) pp497ndash538
Szentagothai J 1983 The modular architectonic principle of neural centers Rev Physiol Biochem Pharmacol 9811ndash61
Tallon-Baudry C Betrand O and Fischer C 2001 Oscillatory synchrony between human extrastriate areas during visualshort-term memory maintenance J Neurosci 21 RC177(1ndash5)
Tass P A 2002 Desynchronization of brain rhythms with soft phase-resetting techniques Biol Cybern 87 102ndash15
732 M Breakspear et al
Tass P Rosenblum M Weule J Kurths J Pikovsky A Volkmann J Schnitzler A and Freund H 1998 Detection of nmphase locking from noisy data application to magnetoencephalography Phys Rev Lett 81 3291ndash4
Terman D and Wang D 1995 Global competition and local cooperation in a network of neural oscillators Physica D81 148ndash76
Terry J and Breakspear M 2003 An improved algorithm for the detection of dynamical interdependence in bivariatetime-series Biol Cybern 88 129ndash36
Tononi G Sporns O and Edelman G M 1994 A measure for brain complexity relating functional segregation andintegration in the nervous system Proc Natl Acad Sci USA 91 5033ndash7
von der Malsburg C 1985 Nervous structures with dynamical links Phys Chem 89 703ndash10Wang X 1991 Period-doublings to chaos in a simple neural network an analytic proof Complex Syst 5 425ndash41Wennekers T and Pasemann F 1996 Synchronous chaos in high dimensional modular neural networks Int J Bifurc
Chaos 6 2055ndash67Zaks M A Parks E H Rosenblum M G and Kurths J 1999 Alternative locking ratios in imperfect phase synchronization
Phys Rev Lett 82 4228ndash31Zhao L and Macau E N 2001 A network of dynamically coupled chaotic maps for scene segmentation IEEE Trans
Neural Netw 12 1375ndash85
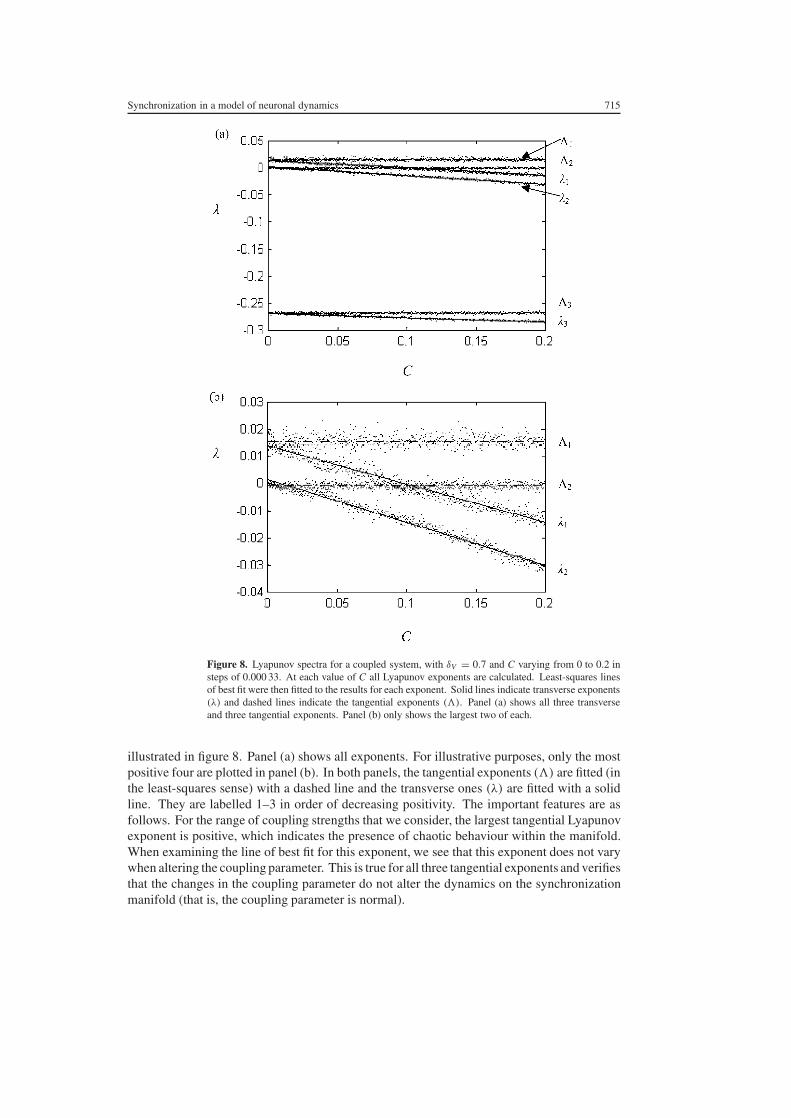
Synchronization in a model of neuronal dynamics 715
Figure 8 Lyapunov spectra for a coupled system with δV = 07 and C varying from 0 to 02 insteps of 0000 33 At each value of C all Lyapunov exponents are calculated Least-squares linesof best fit were then fitted to the results for each exponent Solid lines indicate transverse exponents(λ) and dashed lines indicate the tangential exponents () Panel (a) shows all three transverseand three tangential exponents Panel (b) only shows the largest two of each
illustrated in figure 8 Panel (a) shows all exponents For illustrative purposes only the mostpositive four are plotted in panel (b) In both panels the tangential exponents () are fitted (inthe least-squares sense) with a dashed line and the transverse ones (λ) are fitted with a solidline They are labelled 1ndash3 in order of decreasing positivity The important features are asfollows For the range of coupling strengths that we consider the largest tangential Lyapunovexponent is positive which indicates the presence of chaotic behaviour within the manifoldWhen examining the line of best fit for this exponent we see that this exponent does not varywhen altering the coupling parameter This is true for all three tangential exponents and verifiesthat the changes in the coupling parameter do not alter the dynamics on the synchronizationmanifold (that is the coupling parameter is normal)
716 M Breakspear et al
The largest transverse Lyapunov exponent (denoted as λ1 in the figure) is a particularlyimportant exponent since it governs the stability of the attractor in the synchronization manifoldwith respect to perturbations out of the manifold If this exponent is negative then we expectsmall perturbations of trajectories to reconverge to the manifold in finite time If however thisexponent is positive then we would instead expect perturbed trajectories to diverge from themanifold This change in local behaviour is termed a bifurcation and the point at which itoccurs the bifurcation point It is apparent in figure 8 that the zero crossing of this exponentmdashthe bifurcation pointmdashoccurs at approximately C = 01 in the present system This typeof bifurcation has been termed a blowout bifurcation since once transverse stability is losttrajectories are lsquoblown outrsquo from the low-dimensional synchronization manifold into the fullphase space (Ott and Sommerer 1994) Hence in the present setting a blowout bifurcation isobserved if C falls below 01
We note two other observations Increasing rNMDA which corresponds to increasing thepopulation of NMDA receptors decreases the coupling strength required to achieve stablesynchronization On the other hand increasing the overall excitatory-to-excitatory synapticconnectivity (aee) has the opposite effect This is because the effect of local excitatory feedbackincreases the tangential Lyapunov exponents which has the effect of lsquosplittingrsquo the synchronizeddynamics
In the above simulations all the parameters of both systems were equal In realphysiological systems such symmetry between systems is clearly impossible We nowconsider the case in which the two columns have different structures A simple way toincorporate this whilst at the same time preserving (as much as possible) simple dynamics is tovary the parameter δV for each column This has the effect of destroying any symmetry-forcedinvariant manifolds and so the simple case of synchronization corresponding to identicallyevolving dynamics will not exist in this case However more exotic forms of synchronizationare still possible and we discuss these next
42 Generalized chaotic synchronization
In order to maintain consistency with the literature we study non-linear interdependence in thesetting where the systems are asymmetrical but the coupling is unidirectional This is easilyachieved in the present system by making C directed and setting C21 = 0 In this settingwe refer to the lsquofreersquo system as the driver and the lsquoslavedrsquo system as the response We firstask the question what is the effect of introducing a slight parameter mismatch into a systemexhibiting stable chaos synchronization An example of this is presented in figure 9 whereC12 = 04 and parameter mismatch has been incorporated by setting δV = 07 in the driversystem and δV = 0699 in the response system In panel (a) the time series appear qualitativelyvery similar However in panel (b) where the difference between the systems is plotted it isapparent that the separation between the two systems is of the same order of magnitude as thesize of each attractor In panel (c) the effect of the parameter mismatch on the synchronizationmanifold can be seen as a transformation away from the symmetry manifold
It has been proposed that in such a setting there exists a continuous functional relationshipbetween the dynamics of the driver and response systems
x2(t) = ψ(x1(t)) (10)
where xi = V i W i Z i This situation has been referred to in the literature as generalizedsynchronization (Afraimovich et al 1986 Rulkov et al 1995) The properties of ψ andthe topology and stability of the synchronization manifold in this context have not yetbeen fully elucidated although some significant results have been achieved For example
Synchronization in a model of neuronal dynamics 717
1
2
2
1
Figure 9 Generalized synchronization in an asymmetrical driverndashresponse system (a) The timeseries of the two systems (b) The distance between the two systems (c) A plot of concurrent meanexcitatory membrane potentials V 1 against V 2 The diagonal represents the symmetry manifold
718 M Breakspear et al
Hunt et al (1997) Josic (1998) Kocarev et al (2000) have shown that the generalizedsynchronization manifold is smooth (differentiable) if and only if it is normally hyperbolic(ie the largest transverse Lyapunov exponent is less than the smallest tangential Lyapunovexponent prior to the introduction of parameter mismatch) Such a condition is met in thepresent setting when C = 04 hence the smoothness of the manifold in panel (c)
However subsequent to the introduction of parameter mismatch we will not be able touse transverse and tangential Lyapunov exponents to detect such synchronization since thesynchronization manifold may possess a complex topology for which it is a non-trivial (atbest) or impossible (at worse) task to define transverse and tangential directions For examplethe manifold may be smooth as in figure 9(c) but have a fractal topology If the dynamics arenot normally hyperbolic then the manifold may be fractal but also non-differentiable (Huntet al 1997) and hence will not have a unique tangent direction at any point Consequently wewill not be able to compute tangential or transverse Lyapunov exponents and hence determinethe stability of the synchronization manifold However other techniques for detecting thepresence of generalized synchronization in complex systems have evolved including theauxiliary system approach (Abarbanel et al 1996) and non-linear interdependence (Schiffet al 1996) which are now explored
421 The auxiliary system approach The auxiliary system is a third subsystem which isa replica of the original response system but coupled independently to the driver system andcommencing from independent initial conditions It is possible to demonstrate that showingthe existence of generalized synchronizationmdashthat equation (10) is satisfiedmdashis equivalent toshowing that the response system and the auxiliary system evolve identically (even though theyare not directly coupled) To determine whether or not this is the case reduces to an analysisof the Lyapunov exponents transverse to the symmetry manifold with respect to the responsesystem and the auxiliary system
Whilst this is a somewhat artificial test for generalized synchronization since theproduction of an auxiliary system for a physical system would be impossible such a testis useful in numerical and theoretical studies In figure 10 we present an example of theauxiliary system approach where the parameter δV = 07 for column 1 and 068 for column 2We consider two values of the coupling C = 0025 and 01 The auxiliary system is simplya copy of the second column beginning from an arbitrary initial state In the first instancewhere C = 0025 we notice that even after allowing for an initial transient state the secondcolumn and the auxiliary do not behave in identical fashions (figure 10(a)) However whenC = 01 we observe (figure 10(b)) that the second column and the auxiliary do behave innear-identical manners after an initial transient and hence conclude that for the particularvalue of the coupling strength considered equation (10) is satisfied and hence generalizedsynchronization between the first and second columns occurs
422 Nonlinear interdependence The question remains as to how we may detect thepresence of generalized synchronization in a practical setting As we mentioned aboveit is not possible to use an auxiliary-system-based approach to detect the presence ofgeneralized synchronization Instead we must consider techniques for detecting suchbehaviour from experimental or numerical time series data Algorithms of this type areoften referred to as detecting non-linear interdependence Nonlinear interdependence is ageneral term encompassing systems where the behaviour of one non-linear system affectsthe behaviour of another More specifically it can be defined as occurring if the nullhypothesis that the interdependence between two signals is purely linear can be rejectedat conventional levels (usually 95) of statistical significance hence it is an experimental
Synchronization in a model of neuronal dynamics 719
2
1
2
1
Figure 10 Concurrent evolution of the response system V 2 and lsquoauxiliaryrsquo system V 1 (a) C =0025 (b) C = 01
term (eg Schiff et al 1996) Whilst this does not strictly correspond to the existence of afunctional relationship between the two systems such a relationship is typically used tomotivate the techniques for its detection Schiff et al (1996) extended the logic of non-linearprediction algorithms from autonomous to coupled systems by exploiting the continuity ofsuch a relationship and hence devised the first method of detecting non-linear interdependencein time series data The basis of the technique is using the forward evolution of a phase spacevectorrsquos neighbours to predict the forward evolution of that vector and the forward evolutionof the concurrent vector of the other (possibly interdependent) system If the error of such aprocedure is better on average than chance (the chance error is normalized to equal one) thennon-linear interdependence is said to be present
720 M Breakspear et al
V1
V2
Figure 11 Experimental and numerical examples of weak generalized non-linear synchronizationin the presence of measurement noise (a) Human scalp EEG data from posterior bipolar scalpelectrodes (O1P3ndashO2P4) (b) Weakly coupled aperiodic data in the present model with additivewhite noise (with 5 of the amplitude of the model signal) Parameters δ1
V = 0605 δ2V = 061
aee = 036 C = 007
A variation of this algorithm optimized to detect weak non-linear interdependence (Terryand Breakspear 2003) was used recently to demonstrate that weak non-linear interdependence isoccasionally present in the scalp EEG of healthy resting human subjects (Breakspear and Terry2002a 2002b 2002c) An example of an epoch of lsquonon-linearly independentrsquo EEG obtainedusing this technique is presented in figure 11(a) For comparison output from the currentmodel is presented in figure 11(b) The model parameters were set such that the dynamicswas still aperiodic but the intermittent large-amplitude bursts were suppressed and had weakmutual coupling (C = 007) Extraneous noise to simulate the effect of extracranial muscleactivity contaminating the EEG data has been added to this time series It should be notedthat the amplitudes and time scales of the modelrsquos variables are dimensionless Nonethelessthe model facilitates qualititative interpretation of experimental findings
Synchronization in a model of neuronal dynamics 721
43 Chaotic phase synchronization
Whereas the concept of generalized synchronization was mathematically motivated and assuch more suited to theoretical investigations of systems the concept of phase synchronizationderives from a more practical signal processing approach Phase synchronization occurs inboth symmetric and asymmetric settings but is often more useful in the asymmetric case (andin experimental data)
Unfortunately the phase of a chaotic signal is typically ambiguous and can be defined in anumber of different ways For example in the case of the coherent Rossler attractor (Osipovet al 1997) the phase may be defined as the arctangent of two of the variables of the systemMore generally it can sometimes be defined as the angle that an orbit subtends with the centreof an attractor It is also possible to extract the phase of a chaotic signal from time seriesdata by use of the Hilbert transform This has enabled the demonstration of chaotic phasesynchronization in a number of coupled asymmetrical systems (Rosenblum et al 1996 1997Zaks et al 1999) Although such an approach does not guarantee that the phase will be uniqueit does permit the practical analysis of phase in noisy time series (Tass et al 1998)
To illustrate phase synchronization in two coupled columns we again vary the parameterδV which if the columns are viewed as oscillators acts to detune them from a commonfrequency An important question is that of the criteria on the basis of which we concludewhether phase synchronization occurs We use a simple locking criterion whereby phasesynchronization is said to occur if
|θ1(t)minus θ2(t)| lt K (11)
where is θi the phase of column i and K is a constant Notice that this does not mean thatsynchronization implies equality of phases merely that their difference is bounded
An example of phase synchronization with δV = 065 for column 1 and 066 for column 2is given in figure 12 In this figure rNMDA = 03 Qualitatively some sort of phase coherenceis evident in the time series panel (a) This was confirmed by plotting the sine of thephase difference which has been extracted using the Hilbert transform panel (b) Clearly|sin(θ1 minus θ2)| lt K and hence |θ1 minus θ2| is bounded It can also be seen that the phase-synchronized dynamics occurs on a highly structured manifold depicted in figure 12(c)However unlike the case for the generalized synchronization manifold depicted in figure 9 itis evident that this manifold is not a simple transformation of the symmetry manifold
It is also possible to see lsquobimodalrsquo stability whereby a system jumps betweenapproximately zero phase difference and a phase lag phase synchronization of π An exampleis depicted in figure 12(d) At approximately 2000 s the system can be seen to jumpspontaneously from the first to the second of these modes The Z -variable is plotted forgraphical purposes
5 Intermittency and desynchronization
51 Desynchronization in an array of two systems
Diminishing the strength of coupling across the blowout bifurcation does not lead to a simpletransition from synchronous to asynchronous oscillations but is instead associated withcomplex temporal behaviours Specifically when the largest transverse Lyapunov exponent isclose to zero the coupled systems exhibit epochs of synchronous behaviours punctuated bysharp bursts of desynchronization This is observed in systems exhibiting identicalgeneralizedand phase synchronization An example of such a phenomenon is given in figure 13 The timeseries panel (a) shows synchronous dynamics except at time = 1000 where it can be seenthat the two time series desynchronize briefly This is evident in panel (b) as a disruption in the
722 M Breakspear et al
2
1
Figure 12 Phase-synchronized coupled columns (a) The time series of V 1 (solid) and V 2 (b) Thesine of the phase difference obtained through Hilbert transformation of the time series in panel (a)(c) The phase synchronization manifold in the V 1ndashV 2 plane (d) The time series of Z (solid) andZ showing a phase slip of π between two modes
Synchronization in a model of neuronal dynamics 723
Figure 13 An example of phase desynchronization in coupled asymmetric systems with C = 01Time series plots of (a) each V -variable and (b) their Euclidean distance (c) The temporal evolutionof the phase difference between the two systems (d) An example of the phase difference evolutionover a longer time interval
724 M Breakspear et al
orderly evolution of the distance between the two systems and in panel (c) where the evolutionof the phase difference reveals a brief violation of the criterion |φ1 minus φ2| lt constant Anexample of the phase difference evolution over a longer time interval is shown in figure 13(d)Note that this is a three-dimensional plot with sin(φ1 minus φ2) and cos(φ1 minus φ2) as they- and z-axes Hence phase desynchronizations are evident as complete revolutions aroundthe cylindrical phase space in which this variable evolves It can be seen that these phenomenaoccur at irregular intervals in the temporal domain
This behaviour can be explained by considering the nature of a chaotic attractor In thestatistical sense an arbitrary initial condition is expected to lead to the chaotic behaviour onthe attractor Consequently when we calculate the Lyapunov exponents of the system theseexponents correspond to those of the chaotic attractor Typically however there are an infinitenumber of unstable periodic orbits embedded within the attractor If the path of a trajectorybecomes close to one of these orbits it may lsquoshadowrsquo the orbit for a brief period of time Eachof these orbits has a corresponding set of Lyapunov exponents which (like the attractor) can bedivided into tangential and transverse exponents These also vary when we change the couplingparameter but are typically not the same as the exponents of the attractor Hence the largesttransverse exponent of the attractor may be negative yet there may be positive transverseexponents associated with periodic orbits Considering all of these exponents together wewould therefore only expect to observe complete stability of the synchronization manifold inthe case where all of the transverse Lyapunov exponents are negative Bursting behaviourwill be observed when the one of these exponents becomes positive typically correspondingto a periodic orbit of low order (Pikovsky and Grassberger 1991 Rulkov and Sushchik 1997Maistrenko et al 1998 Heagy et al 1998) Due to the complex structure of such periodic orbitsthe desynchronous bursts occur across a wide range of timescales
In figure 14 we present the results of a long integration (16times105 time units) in which thelengths of sequential synchronous epochs of phase synchronization are studied The parametervalues were δV = 065 for column 1 and 0645 for column 2 with coupling strength10 equal to0125 In panel (a) the power spectrum of these sequential lengths is plotted The whiteness ofthis spectrum indicates that the lengths of consecutive synchronous epochs are independentThis is due to the high-dimensional lsquomixingrsquo effect of desynchronization In panel (b) thelengths of the synchronous epochs have been sorted again into the order of increasing lengthThe results for two coupling strengths are plotted C = 0125 (solid) and 0135 (dashed)Several features of this plot are of note Firstly the lengths of the synchronous epochsare distributed over almost three orders of magnitude Secondly for both plots there is anapproximately log-linear relationship between the length of an epoch and its relative orderThis reflects an approximately exponential relationship between the length of a synchronousepoch and its likelihood of occurrence Thirdly this relationship is skewed toward longersynchronous epochs for the stronger coupling strengths
This type of signal with patches of lsquoorderrsquo occurring over many timescales interruptedby bursts of disorder can be shown to have a high information capacity (Gatlin 1972)Moreover complex phase desynchronization may make an infrequent yet robust contributionto the dynamical pattern of the alpha rhythm in scalp EEG data for resting human subjects(Breakspear 2002) It is tempting to postulate that such a lsquoweakly stablersquo dynamics may assistin the generation of the rich and fluid mental associations that spontaneously emerge in restingcognitive states Although this phenomenon has been previously described in a variety of othersystems to our knowledge this is the first description in a biophysical model of neocortex
10 The introduction of parameter mismatch causes an increase in the strength of coupling required for stablesynchronization above the value of 01 required in the symmetric case
Synchronization in a model of neuronal dynamics 725
Figure 14 Properties of sequential epochs of phase-synchronous behaviour over a long integrationtime δ1
V = 065 δ2V = 0645 aee = 04 (a) The flat (white) power spectrum (b) Synchronous
epochs sorted in order of increasing length (solid C = 0125 dashed C = 0134)
52 Synchronization and complex dynamics in larger arrays
The synchronous and complex phenomena described above are also observed in arrays ofcoupled systems with N gt 2 Expanding the size of the array permits complex dynamicsin the spatial in addition to the temporal domain However the basic principles governingthe type and stability of synchronized motions are the same In this section a few of thepossible behaviours are discussed Physiological variation across the array is modelled bymodifying the parameter δV = 07 in each array by adding a random number of maximummagnitude 001
In figure 15(a) an array of N = 20 uncoupled (C = 0) systems is shown The lack ofsynchronization in this time series is evident By comparison the rapid transition to completesynchronization is evident in figure 15(b) when C = 03 In fact 19 of the 20 subsystems
726 M Breakspear et al
(b)
(c)
(a)
Figure 15 Synchronization phenomena in an array of coupled cortical columns (a) N = 20C = 0 (b) N = 20 C = 03 (c) N = 6 C = 01 (d) N = 12 C = 009 (e) N = 12C = 012 A small perturbation has been applied to one of the columns at t = 2500 s (f) Evolutionof the phase difference between two of the columns in panel (e) For clarity the W -variable isplotted in panel (c) and the Z -variable in (e)
have phase synchronized within one single cycle Intermediate values of coupling strength areassociated with a rich variety of phenomena For example subsystems with the most similarparameter values typically synchronize into clusters of various sizes An example of this isillustrated in figure 15(c) Note that there is no synchronization between the two differentclusters each containing three synchronized subsystems
Another complex behaviour occurs when subsystems transiently synchronize into largeclusters then spontaneously desynchronize again Hence the dynamics of the entire arrayjumps between different transient dynamics characterized by varying degrees of freedom Anexample of this phenomenon is given in figure 15(d) In addition it is possible to inducedesynchronization across an entire array by applying a small perturbation to one columnfor one time step An example is illustrated in figure 15(e) A small perturbation has been
Synchronization in a model of neuronal dynamics 727
Figure 15 (Continued)
added to one column after 2500 time units It can be seen that the entire array subsequentlydesynchronizes This illustrates the sensitivity of the dynamics to external stimuli Finally it ispossible to lsquoinducersquo synchronization between selected subsystems by increasing the strength ofcoupling between them selectively This allows an extra level of complexity to be introducedinto the model For example it may allow targeted information transfer between a partialnetwork of the entire array
As discussed in section 51 a univariate signal which jumps between epochs of lowentropy (high repetition) and bursts of high entropy (high variability) can be shown to havehigh information capacity This is because the repetition of certain patterns (as in the English
728 M Breakspear et al
language) permits error detectability Bursts of high entropy act in the opposite directionto increase message variability With a multivariate signal in addition to optimizing theinformation capacity within each signal the overall information capacity can be increasedthrough the type of itinerant synchronization briefly illustrated in this section Synchronizationacross an entire array increases the message reliability (since a single lsquoerrant letterrsquo in onesignal can be easily detected) Conversely desynchronization into multiple clusters increasesmessage variability since each cluster will produce independent information Hence a neuralarray which switches dynamically between a range of cluster configurations can be seen asadaptive in the sense that high synchronous states can be expressed when error detectabilityis a priority (such as during quick and reliable execution of a familiar task) but the array canrapidly desynchronize if variability becomes salient (for example if the environment suddenlybecomes novel) A formal operationalized demonstration of this principle is to be the subjectof future work
6 Discussion
In this paper we present a dynamical model for activity in an array of weakly coupledlocal neural assemblies The model incorporates the fundamental principles governing cellmembrane potential as determined by voltage- and ligand-gated ion channels In this wayit allows a reasonable amount of physiological complexity to be incorporated includingsynaptic activation and receptor subtypeswithout recourse to overwhelming detail Numericalanalysis of this model reveals a transition from independent oscillations to stable chaoticsynchronization depending on the balance of local versus long-range connectivity andbiophysical heterogeneity These excitatory couplings are subject to ascending subcorticalneuromodulation arising principally from monoaminergic nuclei such as Locus Coeruleus(Silberstein 1995) Neuromodulation of extrinsic connections was incorporated into the modelvia a coupling parameter C The stability of the synchronization is determined by the valueof the largest transverse Lyapunov exponent which in turn depends on the coupling parameterC If the coupling is weakened with the result that this exponent becomes positive a blowoutbifurcation occurs and independent oscillations re-emerge In the vicinity of the transitionnon-linear desynchronizations with complex temporal and spatial properties are observed
It is pleasing that complex and itinerant behaviour reminiscent of real neuronaldynamics emerges through such a simple manipulation of a single dynamic control parameterFurthermore the ubiquitous projections of classical neuromodulatory systems in real brainssuggest a clear biological mechanism by which the brain could self-organize throughmoderating the complexity of its own dynamics There are many reasons that the complicateddynamics of the sort demonstrated in this paper may be neurobiologically important Wemention four
First the presence of multiple clusters of phase-synchronized subsystems as discussedabove may be a uniquely non-linear mechanism facilitating the lsquocarriagersquo and distinction ofmultiple concurrent dynamic cell assemblies (and hence cognitive processing) This may beimportant for perceptual grouping binding and transitions as suggested by others (eg von derMalsburg 1985 Gray and Singer 1989)
Second the facility to decouple and reconfigure dynamic cell assemblies througha neuromodulatory control parameter may be important for avoiding low-entropysynchronization manifolds In the absence of itinerancy synchronization would renderneuronal dynamics incapable of representing changes in sensory input or indeed cognitive stateFor example in the model it was observed that small perturbations to one column were ableto induce a global change in neuronal dynamics by disrupting a state of weak synchronization
Synchronization in a model of neuronal dynamics 729
Inevitably alternative dynamic configurations emerge epitomizing the importance of lsquolabilersquobrain dynamics (Friston 2000a)
Thirdly from a purely functionalist point of view the balance between functionalspecialization and functional integration may rely upon the sorts of mechanism modelledin this paper The brain appears to adhere to two principles of functional organization namelysegregation of function within cortical areas and sub-areas and the integration of these areasin perceptual synthesis cognition and sensorimotor integration (Tononi et al 1994 Fristonet al 1995) The balance between preserving specialized dynamics unique to each arearequired by specialization and the coupling among these areas mediating functional integrationmay be achieved by the neuromodulatory mechanisms described above Significantlythe spontaneous variation in the numbers of degrees of freedom (for example whenlow-dimensional synchronization manifolds are shadowed) imparts a dynamic characteristicto this perspective that is precluded by linear mechanisms alone
Finally from a mechanistic perspective complexity and diversity of brain dynamics arerequired for selection of adaptive responses (Friston 2000b) There are many examples ofselective mechanisms operating within the brain All can be seen as performing an ascenton some objective function or fitness landscape These mechanisms require complex anditinerant dynamics to facilitate a thorough exploration of the landscape These landscapesmay range from those involved in perception (where the landscape is defined by predictionerror see eg Rao and Ballard (1998)) to those involved in value learning (where thelandscape is determined by the outcome of a behaviour eg in temporal difference models (seeeg Sutton and Barto 1990)) In short complexity is necessary to maintain an information-rich environment in which the brain can respond to sensory inputs An essential componentof this is the maintenance of diversity which is a prerequisite for selectionist mechanismsat many scales (Edelman 1993) Understanding the mechanistic basis of complex dynamicsas measured electrophysiologically may well depend on the sort of biologically plausiblemodels used in this paper This understanding may be important for both basic and clinicalneuroscience (eg Tass 2002)
The present study supplements and advances on other research in this field The studyof complex phase synchronization and clustering in neural oscillator networks similar tothe present model has previously been the subject of formal mathematical study Forexample Terman and Wang (1995) studied synchronization of blocks of neural oscillatorsthat were subject to perturbation by connected lsquostimulirsquo Somers and Kopell (1995) studiedthe emergence of both synchronization and travelling waves in networks of coupled neuraloscillators In the work of Terman and Wang the local dynamics were based on neurallsquocaricaturersquo models such as the FitzHughndashNagumo equations and are hence more abstract thanthe current study The approach of Somers and Kopell only required that the local dynamicsexhibit fast and relaxation phases and is hence more abstract still In both studies onlyperiodic dynamics are studied Complex patterns of chaos synchronization in neural arrayshave been characterized by nonlinear methods (Kaneko 1998) and their putative contributionto both learning (Nakajima and Ueda 1996) and perception (Zhao and Macau 2001 Breakspear2001) explored However these studies employed models such as the logistic equation andtent map which are quite abstract in comparison to the present study On the other handHansel and Sompolinsky (1996) studied the role of chaos synchronization in a model ofvisual perception in a cortical hypercolumn using a very similar approach to that adoptedhere but with the additional complexity that synaptic delays were incorporated However inthis and earlier work (Hansel and Sompolinsky 1992) only linear measures of dynamics andsynchrony (specifically the auto- and cross-correlation functions) were employed The analysisof chaotic synchrony in the study of Larter et al (1999) was based only upon observation of
730 M Breakspear et al
the Euclidean difference between variables from two nodes within a larger array Hence thesignificance of the present work is that it provides a quantitative nonlinear analysis of complexchaos synchronization in a biophysical model of cortical dynamics Such research is requiredin order to bridge the gap between quantitative nonlinear research into synchronization andcomputation in abstract models of neural arrays and qualitative or linear analysis in morebiophysically plausible models
Acknowledgments
The authors would like to thank Peter Robinson for assistance with the manuscript and R Larterfor helpful advice MB is a recipient of a NSW Institute of Psychiatry Research Grant anda University of Sydney SESQUI postdoctoral fellowship KJF is supported by the WellcomeTrust
References
Abarbanel H Rulkov N and Sushchik M 1996 Generalised synchronization of chaos the auxiliary system approachPhys Rev E 53 4528ndash35
Afraimovich V Verichev N and Rabinovich M 1986 Stochastic synchronization of oscillation in dissipative systemsRadiophys Quantum Electron 29 795ndash801
Breakspear M 2001 Perception of odours by a nonlinear model of the olfactory bulb Int J Neural Syst 11 101ndash24Breakspear M 2002 Nonlinear phase desynchronisation in human electroencephalographic data Human Brain Mapping
15 175ndash98Breakspear M and Terry J R 2002a Nonlinear interdependence in neural systems motivation theory and relevance
Int J Neurosci 112 1163ndash84Breakspear M and Terry J R 2002b Detection and description of nonlinear interdependence in normal multichannel
human EEG Clin Neurophysiol 113 735ndash53Breakspear M and Terry J R 2002c Topographic organization of nonlinear interdependence in multichannel human
EEG Neuroimage 16 822ndash35Destexhe A Rudolph M Fellous J M and Sejnowski T J 2001 Fluctuating synaptic conductances recreate in vivo-like
activity in neocortical neurons Neuroscience 107 13ndash24Douglas R J Mahowald M A and Martin K A 1996 Microarchitecture of neocortical columns Brain TheorymdashBiological
Basis and Computational Principles ed A Aertsen and V Braitenberg (Amsterdam Elsevier)Eckhorn R Bauer R Jordan W Brosch M Kruse W Munk M and Reitboeck H J 1988 Coherent oscillations a
mechanism of feature linking in the visual cortex Multiple electrode and correlation analyses in the cat BiolCybern 60 121ndash30
Edelman G M 1993 Neural Darwinism selection and reentrant signalling in higher brain function Neuron 10 115ndash25Engel A K Kreiter A K Konig P and Singer W 1991 Synchronization of oscillatory neuronal responses between
striate and extrastriate visual cortical areas of the cat Proc Natl Acad Sci USA 88 6048ndash52Friston K J 2000a The labile brain I Neuronal transients and nonlinear coupling Phil Trans R Soc B 355 215ndash36Friston K J 2000b The labile brain II Transients complexity and selection Phil Trans R Soc B 355 237ndash52Friston K J Tononi G Sporns O and Edelman G 1995 Characterising the complexity of neuronal interactions Human
Brain Mapping 3 302ndash14Fujisaka H and Yamada T 1983 Stability theory of synchronized motion in coupled-oscillator system Prog Theor
Phys 69 32ndash47Gatlin L 1972 Information Theory and the Living System (New York Columbia University Press)Gray C M Konig P Engel A and Singer W 1989 Oscillatory responses in cat visual cortex exhibit inter-columnar
synchronization which reflects global stimulus properties Science 338 334ndash7Gray C M and Singer W 1989 Stimulus specific neuronal oscillations in orientation columns of cat visual cortex Proc
Natl Acad Sci USA 86 1698ndash702Haig A R Gordon E Wright J J Meares R A and Bahramali H 2000 Synchronous cortical gamma-band activity in
task-relevant cognition Neuroreport 11 669ndash75Hansel D and Sompolinsky H 1992 Synchronization and computation in a chaotic neural network Phys Rev Lett 68
718ndash21
Synchronization in a model of neuronal dynamics 731
Hansel D and Sompolinsky H 1996 Chaos and synchrony in a model of a hypercolumn in visual cortex J ComputNeurosci 3 7ndash4
Heagy J Carroll T and Pecora L 1998 Desynchronization by periodic orbits Phys Rev E 57 R1253ndash6Hunt B Ott E and Yorke J 1997 Differentiable synchronization of chaos Phys Rev E 55 4029ndash34Josic K 1998 Invariant manifolds and synchronization of coupled dynamical systems Phys Rev Lett 80 3053ndash6Kandel E Schwartz J and Jessell T 2000 Principles of Neural Science (New York McGraw-Hill)Kaneko K 1998 On the strength of attractors in a high-dimensional system Physica D 124 322ndash44Kocarev L Parlitz U and Brown R 2000 Robust synchronization of chaotic systems Phys Rev E 61 3716ndash20Larter R Speelman B and Worth R M 1999 A coupled ordinary differential equation lattice model for the simulation
of epileptic seizures Chaos 9 795ndash804Lumer E Edelman G and Tononi G 1997a Neural dynamics in a model of the thalamocortical system I Layers loops
and the emergence of fast synchronous oscillations Cereb Cortex 7 207ndash27Lumer E Edelman G and Tononi G 1997b Neural dynamics in a model of the thalamocortical system II The role of
neural synchrony tested through perturbations of spike timing Cereb Cortex 7 228ndash36Magleby K 1987 Short-term modulation at synapses Synaptic Function ed G M Edelman W E Gall and
W M Cowan (New York Wiley)Maistrenko Y Maistrenko V Popovich A and Mosekilde E 1998 Transverse instability and riddled basins in a system
of two coupled logistic maps Phys Rev E 57 2713ndash24Morris C and Lecar H 1981 Voltage oscillations in the barnacle giant muscle fiber Biophys J 35 193ndash213Mountcastle V B 1997 The columnar organization of the neocortex Brain 120 701ndash22Mountcastle V B 1998 The Cerebral Cortex (Cambridge MA Harvard University Press)Nakajima H and Ueda Y 1996 Riddled basins of the optimal states in learning dynamical systems Physica D 99 35ndash44Nunez P L 1995 Neocortical Dynamics and Human Brain Rhythms (Oxford Oxford University Press)Osipov G V Pikovsky A S Rosenblum M G and Kurths J 1997 Phase synchronization effects in a lattice of nonidentical
Rossler oscillators Phys Rev E 55 2353ndash61Ott E and Sommerer J 1994 Blowout bifurcations the occurrence of riddled basins and onndashoff intermittency Phys
Lett A 188 39ndash47Pecora L and Carroll T 1990 Synchronization in chaotic systems Phys Rev Lett 64 821ndash4Pikovsky A and Grassberger P 1991 Symmetry breaking bifurcation for coupled chaotic attractors J Phys A Math
Gen 24 4587ndash97Rao R P and Ballard D H 1998 Predictive coding in the visual cortex a functional interpretation of some extra-classical
receptive field effects Nature Neurosci 2 79ndash87Robinson P A Wright J J and Rennie C R 1998 Synchronous oscillations in the cerebral cortex Phys Rev E 57
4578ndash88Rodriguez E George N Lachauz J P Martinerie J Renault B and Varela F J 1999 Perceptionrsquos shadow long distance
synchronization of human brain activity Nature 397 430ndash3Rosenblum M Pikovsky A and Kurths J 1996 Phase synchronization of chaotic oscillators Phys Rev Lett 76 1804ndash7Rosenblum M Pikovsky A and Kurths J 1997 From phase to lag synchronization in coupled chaotic oscillators Phys
Rev Lett 78 4193ndash6Rulkov N and Sushchik M 1997 Robustness of synchronized chaotic oscillations Int J Bifurc Chaos 7 625ndash43Rulkov N Sushchik M Tsimring L and Abarbanel H 1995 Generalized synchronization of chaos in unidirectionally
coupled chaotic systems Phys Rev E 51 980ndash94Schiff S So P Chang T Burke R and Sauer T 1996 Detecting dynamical interdependence and generalized synchrony
through mutual prediction in a neural ensemble Phys Rev E 54 6708ndash24Silberstein R B 1995 Neuromodulations of neocortical dynamics Neocortical Dynamics and Human EEG Rhythms
ed P L Nunez (Oxford Oxford University Press)Singer W 1995 Putative functions of temporal correlations in neocortical processing Large-Scale Neuronal Theories
of the Brain ed C Koch and J Davis (London MIT Press)Somers D and Kopell N 1995 Waves and synchrony in networks of oscillators of relaxation and non-relaxation type
Physica D 89 169ndash83Sutton R S and Barto A G 1990 Time derivative models of Pavlovian reinforcement Learning and Computational
Neuroscience Foundations of Adaptive Networks ed M Gabriel and J Moore (Cambridge MA MIT Press) pp497ndash538
Szentagothai J 1983 The modular architectonic principle of neural centers Rev Physiol Biochem Pharmacol 9811ndash61
Tallon-Baudry C Betrand O and Fischer C 2001 Oscillatory synchrony between human extrastriate areas during visualshort-term memory maintenance J Neurosci 21 RC177(1ndash5)
Tass P A 2002 Desynchronization of brain rhythms with soft phase-resetting techniques Biol Cybern 87 102ndash15
732 M Breakspear et al
Tass P Rosenblum M Weule J Kurths J Pikovsky A Volkmann J Schnitzler A and Freund H 1998 Detection of nmphase locking from noisy data application to magnetoencephalography Phys Rev Lett 81 3291ndash4
Terman D and Wang D 1995 Global competition and local cooperation in a network of neural oscillators Physica D81 148ndash76
Terry J and Breakspear M 2003 An improved algorithm for the detection of dynamical interdependence in bivariatetime-series Biol Cybern 88 129ndash36
Tononi G Sporns O and Edelman G M 1994 A measure for brain complexity relating functional segregation andintegration in the nervous system Proc Natl Acad Sci USA 91 5033ndash7
von der Malsburg C 1985 Nervous structures with dynamical links Phys Chem 89 703ndash10Wang X 1991 Period-doublings to chaos in a simple neural network an analytic proof Complex Syst 5 425ndash41Wennekers T and Pasemann F 1996 Synchronous chaos in high dimensional modular neural networks Int J Bifurc
Chaos 6 2055ndash67Zaks M A Parks E H Rosenblum M G and Kurths J 1999 Alternative locking ratios in imperfect phase synchronization
Phys Rev Lett 82 4228ndash31Zhao L and Macau E N 2001 A network of dynamically coupled chaotic maps for scene segmentation IEEE Trans
Neural Netw 12 1375ndash85

716 M Breakspear et al
The largest transverse Lyapunov exponent (denoted as λ1 in the figure) is a particularlyimportant exponent since it governs the stability of the attractor in the synchronization manifoldwith respect to perturbations out of the manifold If this exponent is negative then we expectsmall perturbations of trajectories to reconverge to the manifold in finite time If however thisexponent is positive then we would instead expect perturbed trajectories to diverge from themanifold This change in local behaviour is termed a bifurcation and the point at which itoccurs the bifurcation point It is apparent in figure 8 that the zero crossing of this exponentmdashthe bifurcation pointmdashoccurs at approximately C = 01 in the present system This typeof bifurcation has been termed a blowout bifurcation since once transverse stability is losttrajectories are lsquoblown outrsquo from the low-dimensional synchronization manifold into the fullphase space (Ott and Sommerer 1994) Hence in the present setting a blowout bifurcation isobserved if C falls below 01
We note two other observations Increasing rNMDA which corresponds to increasing thepopulation of NMDA receptors decreases the coupling strength required to achieve stablesynchronization On the other hand increasing the overall excitatory-to-excitatory synapticconnectivity (aee) has the opposite effect This is because the effect of local excitatory feedbackincreases the tangential Lyapunov exponents which has the effect of lsquosplittingrsquo the synchronizeddynamics
In the above simulations all the parameters of both systems were equal In realphysiological systems such symmetry between systems is clearly impossible We nowconsider the case in which the two columns have different structures A simple way toincorporate this whilst at the same time preserving (as much as possible) simple dynamics is tovary the parameter δV for each column This has the effect of destroying any symmetry-forcedinvariant manifolds and so the simple case of synchronization corresponding to identicallyevolving dynamics will not exist in this case However more exotic forms of synchronizationare still possible and we discuss these next
42 Generalized chaotic synchronization
In order to maintain consistency with the literature we study non-linear interdependence in thesetting where the systems are asymmetrical but the coupling is unidirectional This is easilyachieved in the present system by making C directed and setting C21 = 0 In this settingwe refer to the lsquofreersquo system as the driver and the lsquoslavedrsquo system as the response We firstask the question what is the effect of introducing a slight parameter mismatch into a systemexhibiting stable chaos synchronization An example of this is presented in figure 9 whereC12 = 04 and parameter mismatch has been incorporated by setting δV = 07 in the driversystem and δV = 0699 in the response system In panel (a) the time series appear qualitativelyvery similar However in panel (b) where the difference between the systems is plotted it isapparent that the separation between the two systems is of the same order of magnitude as thesize of each attractor In panel (c) the effect of the parameter mismatch on the synchronizationmanifold can be seen as a transformation away from the symmetry manifold
It has been proposed that in such a setting there exists a continuous functional relationshipbetween the dynamics of the driver and response systems
x2(t) = ψ(x1(t)) (10)
where xi = V i W i Z i This situation has been referred to in the literature as generalizedsynchronization (Afraimovich et al 1986 Rulkov et al 1995) The properties of ψ andthe topology and stability of the synchronization manifold in this context have not yetbeen fully elucidated although some significant results have been achieved For example
Synchronization in a model of neuronal dynamics 717
1
2
2
1
Figure 9 Generalized synchronization in an asymmetrical driverndashresponse system (a) The timeseries of the two systems (b) The distance between the two systems (c) A plot of concurrent meanexcitatory membrane potentials V 1 against V 2 The diagonal represents the symmetry manifold
718 M Breakspear et al
Hunt et al (1997) Josic (1998) Kocarev et al (2000) have shown that the generalizedsynchronization manifold is smooth (differentiable) if and only if it is normally hyperbolic(ie the largest transverse Lyapunov exponent is less than the smallest tangential Lyapunovexponent prior to the introduction of parameter mismatch) Such a condition is met in thepresent setting when C = 04 hence the smoothness of the manifold in panel (c)
However subsequent to the introduction of parameter mismatch we will not be able touse transverse and tangential Lyapunov exponents to detect such synchronization since thesynchronization manifold may possess a complex topology for which it is a non-trivial (atbest) or impossible (at worse) task to define transverse and tangential directions For examplethe manifold may be smooth as in figure 9(c) but have a fractal topology If the dynamics arenot normally hyperbolic then the manifold may be fractal but also non-differentiable (Huntet al 1997) and hence will not have a unique tangent direction at any point Consequently wewill not be able to compute tangential or transverse Lyapunov exponents and hence determinethe stability of the synchronization manifold However other techniques for detecting thepresence of generalized synchronization in complex systems have evolved including theauxiliary system approach (Abarbanel et al 1996) and non-linear interdependence (Schiffet al 1996) which are now explored
421 The auxiliary system approach The auxiliary system is a third subsystem which isa replica of the original response system but coupled independently to the driver system andcommencing from independent initial conditions It is possible to demonstrate that showingthe existence of generalized synchronizationmdashthat equation (10) is satisfiedmdashis equivalent toshowing that the response system and the auxiliary system evolve identically (even though theyare not directly coupled) To determine whether or not this is the case reduces to an analysisof the Lyapunov exponents transverse to the symmetry manifold with respect to the responsesystem and the auxiliary system
Whilst this is a somewhat artificial test for generalized synchronization since theproduction of an auxiliary system for a physical system would be impossible such a testis useful in numerical and theoretical studies In figure 10 we present an example of theauxiliary system approach where the parameter δV = 07 for column 1 and 068 for column 2We consider two values of the coupling C = 0025 and 01 The auxiliary system is simplya copy of the second column beginning from an arbitrary initial state In the first instancewhere C = 0025 we notice that even after allowing for an initial transient state the secondcolumn and the auxiliary do not behave in identical fashions (figure 10(a)) However whenC = 01 we observe (figure 10(b)) that the second column and the auxiliary do behave innear-identical manners after an initial transient and hence conclude that for the particularvalue of the coupling strength considered equation (10) is satisfied and hence generalizedsynchronization between the first and second columns occurs
422 Nonlinear interdependence The question remains as to how we may detect thepresence of generalized synchronization in a practical setting As we mentioned aboveit is not possible to use an auxiliary-system-based approach to detect the presence ofgeneralized synchronization Instead we must consider techniques for detecting suchbehaviour from experimental or numerical time series data Algorithms of this type areoften referred to as detecting non-linear interdependence Nonlinear interdependence is ageneral term encompassing systems where the behaviour of one non-linear system affectsthe behaviour of another More specifically it can be defined as occurring if the nullhypothesis that the interdependence between two signals is purely linear can be rejectedat conventional levels (usually 95) of statistical significance hence it is an experimental
Synchronization in a model of neuronal dynamics 719
2
1
2
1
Figure 10 Concurrent evolution of the response system V 2 and lsquoauxiliaryrsquo system V 1 (a) C =0025 (b) C = 01
term (eg Schiff et al 1996) Whilst this does not strictly correspond to the existence of afunctional relationship between the two systems such a relationship is typically used tomotivate the techniques for its detection Schiff et al (1996) extended the logic of non-linearprediction algorithms from autonomous to coupled systems by exploiting the continuity ofsuch a relationship and hence devised the first method of detecting non-linear interdependencein time series data The basis of the technique is using the forward evolution of a phase spacevectorrsquos neighbours to predict the forward evolution of that vector and the forward evolutionof the concurrent vector of the other (possibly interdependent) system If the error of such aprocedure is better on average than chance (the chance error is normalized to equal one) thennon-linear interdependence is said to be present
720 M Breakspear et al
V1
V2
Figure 11 Experimental and numerical examples of weak generalized non-linear synchronizationin the presence of measurement noise (a) Human scalp EEG data from posterior bipolar scalpelectrodes (O1P3ndashO2P4) (b) Weakly coupled aperiodic data in the present model with additivewhite noise (with 5 of the amplitude of the model signal) Parameters δ1
V = 0605 δ2V = 061
aee = 036 C = 007
A variation of this algorithm optimized to detect weak non-linear interdependence (Terryand Breakspear 2003) was used recently to demonstrate that weak non-linear interdependence isoccasionally present in the scalp EEG of healthy resting human subjects (Breakspear and Terry2002a 2002b 2002c) An example of an epoch of lsquonon-linearly independentrsquo EEG obtainedusing this technique is presented in figure 11(a) For comparison output from the currentmodel is presented in figure 11(b) The model parameters were set such that the dynamicswas still aperiodic but the intermittent large-amplitude bursts were suppressed and had weakmutual coupling (C = 007) Extraneous noise to simulate the effect of extracranial muscleactivity contaminating the EEG data has been added to this time series It should be notedthat the amplitudes and time scales of the modelrsquos variables are dimensionless Nonethelessthe model facilitates qualititative interpretation of experimental findings
Synchronization in a model of neuronal dynamics 721
43 Chaotic phase synchronization
Whereas the concept of generalized synchronization was mathematically motivated and assuch more suited to theoretical investigations of systems the concept of phase synchronizationderives from a more practical signal processing approach Phase synchronization occurs inboth symmetric and asymmetric settings but is often more useful in the asymmetric case (andin experimental data)
Unfortunately the phase of a chaotic signal is typically ambiguous and can be defined in anumber of different ways For example in the case of the coherent Rossler attractor (Osipovet al 1997) the phase may be defined as the arctangent of two of the variables of the systemMore generally it can sometimes be defined as the angle that an orbit subtends with the centreof an attractor It is also possible to extract the phase of a chaotic signal from time seriesdata by use of the Hilbert transform This has enabled the demonstration of chaotic phasesynchronization in a number of coupled asymmetrical systems (Rosenblum et al 1996 1997Zaks et al 1999) Although such an approach does not guarantee that the phase will be uniqueit does permit the practical analysis of phase in noisy time series (Tass et al 1998)
To illustrate phase synchronization in two coupled columns we again vary the parameterδV which if the columns are viewed as oscillators acts to detune them from a commonfrequency An important question is that of the criteria on the basis of which we concludewhether phase synchronization occurs We use a simple locking criterion whereby phasesynchronization is said to occur if
|θ1(t)minus θ2(t)| lt K (11)
where is θi the phase of column i and K is a constant Notice that this does not mean thatsynchronization implies equality of phases merely that their difference is bounded
An example of phase synchronization with δV = 065 for column 1 and 066 for column 2is given in figure 12 In this figure rNMDA = 03 Qualitatively some sort of phase coherenceis evident in the time series panel (a) This was confirmed by plotting the sine of thephase difference which has been extracted using the Hilbert transform panel (b) Clearly|sin(θ1 minus θ2)| lt K and hence |θ1 minus θ2| is bounded It can also be seen that the phase-synchronized dynamics occurs on a highly structured manifold depicted in figure 12(c)However unlike the case for the generalized synchronization manifold depicted in figure 9 itis evident that this manifold is not a simple transformation of the symmetry manifold
It is also possible to see lsquobimodalrsquo stability whereby a system jumps betweenapproximately zero phase difference and a phase lag phase synchronization of π An exampleis depicted in figure 12(d) At approximately 2000 s the system can be seen to jumpspontaneously from the first to the second of these modes The Z -variable is plotted forgraphical purposes
5 Intermittency and desynchronization
51 Desynchronization in an array of two systems
Diminishing the strength of coupling across the blowout bifurcation does not lead to a simpletransition from synchronous to asynchronous oscillations but is instead associated withcomplex temporal behaviours Specifically when the largest transverse Lyapunov exponent isclose to zero the coupled systems exhibit epochs of synchronous behaviours punctuated bysharp bursts of desynchronization This is observed in systems exhibiting identicalgeneralizedand phase synchronization An example of such a phenomenon is given in figure 13 The timeseries panel (a) shows synchronous dynamics except at time = 1000 where it can be seenthat the two time series desynchronize briefly This is evident in panel (b) as a disruption in the
722 M Breakspear et al
2
1
Figure 12 Phase-synchronized coupled columns (a) The time series of V 1 (solid) and V 2 (b) Thesine of the phase difference obtained through Hilbert transformation of the time series in panel (a)(c) The phase synchronization manifold in the V 1ndashV 2 plane (d) The time series of Z (solid) andZ showing a phase slip of π between two modes
Synchronization in a model of neuronal dynamics 723
Figure 13 An example of phase desynchronization in coupled asymmetric systems with C = 01Time series plots of (a) each V -variable and (b) their Euclidean distance (c) The temporal evolutionof the phase difference between the two systems (d) An example of the phase difference evolutionover a longer time interval
724 M Breakspear et al
orderly evolution of the distance between the two systems and in panel (c) where the evolutionof the phase difference reveals a brief violation of the criterion |φ1 minus φ2| lt constant Anexample of the phase difference evolution over a longer time interval is shown in figure 13(d)Note that this is a three-dimensional plot with sin(φ1 minus φ2) and cos(φ1 minus φ2) as they- and z-axes Hence phase desynchronizations are evident as complete revolutions aroundthe cylindrical phase space in which this variable evolves It can be seen that these phenomenaoccur at irregular intervals in the temporal domain
This behaviour can be explained by considering the nature of a chaotic attractor In thestatistical sense an arbitrary initial condition is expected to lead to the chaotic behaviour onthe attractor Consequently when we calculate the Lyapunov exponents of the system theseexponents correspond to those of the chaotic attractor Typically however there are an infinitenumber of unstable periodic orbits embedded within the attractor If the path of a trajectorybecomes close to one of these orbits it may lsquoshadowrsquo the orbit for a brief period of time Eachof these orbits has a corresponding set of Lyapunov exponents which (like the attractor) can bedivided into tangential and transverse exponents These also vary when we change the couplingparameter but are typically not the same as the exponents of the attractor Hence the largesttransverse exponent of the attractor may be negative yet there may be positive transverseexponents associated with periodic orbits Considering all of these exponents together wewould therefore only expect to observe complete stability of the synchronization manifold inthe case where all of the transverse Lyapunov exponents are negative Bursting behaviourwill be observed when the one of these exponents becomes positive typically correspondingto a periodic orbit of low order (Pikovsky and Grassberger 1991 Rulkov and Sushchik 1997Maistrenko et al 1998 Heagy et al 1998) Due to the complex structure of such periodic orbitsthe desynchronous bursts occur across a wide range of timescales
In figure 14 we present the results of a long integration (16times105 time units) in which thelengths of sequential synchronous epochs of phase synchronization are studied The parametervalues were δV = 065 for column 1 and 0645 for column 2 with coupling strength10 equal to0125 In panel (a) the power spectrum of these sequential lengths is plotted The whiteness ofthis spectrum indicates that the lengths of consecutive synchronous epochs are independentThis is due to the high-dimensional lsquomixingrsquo effect of desynchronization In panel (b) thelengths of the synchronous epochs have been sorted again into the order of increasing lengthThe results for two coupling strengths are plotted C = 0125 (solid) and 0135 (dashed)Several features of this plot are of note Firstly the lengths of the synchronous epochsare distributed over almost three orders of magnitude Secondly for both plots there is anapproximately log-linear relationship between the length of an epoch and its relative orderThis reflects an approximately exponential relationship between the length of a synchronousepoch and its likelihood of occurrence Thirdly this relationship is skewed toward longersynchronous epochs for the stronger coupling strengths
This type of signal with patches of lsquoorderrsquo occurring over many timescales interruptedby bursts of disorder can be shown to have a high information capacity (Gatlin 1972)Moreover complex phase desynchronization may make an infrequent yet robust contributionto the dynamical pattern of the alpha rhythm in scalp EEG data for resting human subjects(Breakspear 2002) It is tempting to postulate that such a lsquoweakly stablersquo dynamics may assistin the generation of the rich and fluid mental associations that spontaneously emerge in restingcognitive states Although this phenomenon has been previously described in a variety of othersystems to our knowledge this is the first description in a biophysical model of neocortex
10 The introduction of parameter mismatch causes an increase in the strength of coupling required for stablesynchronization above the value of 01 required in the symmetric case
Synchronization in a model of neuronal dynamics 725
Figure 14 Properties of sequential epochs of phase-synchronous behaviour over a long integrationtime δ1
V = 065 δ2V = 0645 aee = 04 (a) The flat (white) power spectrum (b) Synchronous
epochs sorted in order of increasing length (solid C = 0125 dashed C = 0134)
52 Synchronization and complex dynamics in larger arrays
The synchronous and complex phenomena described above are also observed in arrays ofcoupled systems with N gt 2 Expanding the size of the array permits complex dynamicsin the spatial in addition to the temporal domain However the basic principles governingthe type and stability of synchronized motions are the same In this section a few of thepossible behaviours are discussed Physiological variation across the array is modelled bymodifying the parameter δV = 07 in each array by adding a random number of maximummagnitude 001
In figure 15(a) an array of N = 20 uncoupled (C = 0) systems is shown The lack ofsynchronization in this time series is evident By comparison the rapid transition to completesynchronization is evident in figure 15(b) when C = 03 In fact 19 of the 20 subsystems
726 M Breakspear et al
(b)
(c)
(a)
Figure 15 Synchronization phenomena in an array of coupled cortical columns (a) N = 20C = 0 (b) N = 20 C = 03 (c) N = 6 C = 01 (d) N = 12 C = 009 (e) N = 12C = 012 A small perturbation has been applied to one of the columns at t = 2500 s (f) Evolutionof the phase difference between two of the columns in panel (e) For clarity the W -variable isplotted in panel (c) and the Z -variable in (e)
have phase synchronized within one single cycle Intermediate values of coupling strength areassociated with a rich variety of phenomena For example subsystems with the most similarparameter values typically synchronize into clusters of various sizes An example of this isillustrated in figure 15(c) Note that there is no synchronization between the two differentclusters each containing three synchronized subsystems
Another complex behaviour occurs when subsystems transiently synchronize into largeclusters then spontaneously desynchronize again Hence the dynamics of the entire arrayjumps between different transient dynamics characterized by varying degrees of freedom Anexample of this phenomenon is given in figure 15(d) In addition it is possible to inducedesynchronization across an entire array by applying a small perturbation to one columnfor one time step An example is illustrated in figure 15(e) A small perturbation has been
Synchronization in a model of neuronal dynamics 727
Figure 15 (Continued)
added to one column after 2500 time units It can be seen that the entire array subsequentlydesynchronizes This illustrates the sensitivity of the dynamics to external stimuli Finally it ispossible to lsquoinducersquo synchronization between selected subsystems by increasing the strength ofcoupling between them selectively This allows an extra level of complexity to be introducedinto the model For example it may allow targeted information transfer between a partialnetwork of the entire array
As discussed in section 51 a univariate signal which jumps between epochs of lowentropy (high repetition) and bursts of high entropy (high variability) can be shown to havehigh information capacity This is because the repetition of certain patterns (as in the English
728 M Breakspear et al
language) permits error detectability Bursts of high entropy act in the opposite directionto increase message variability With a multivariate signal in addition to optimizing theinformation capacity within each signal the overall information capacity can be increasedthrough the type of itinerant synchronization briefly illustrated in this section Synchronizationacross an entire array increases the message reliability (since a single lsquoerrant letterrsquo in onesignal can be easily detected) Conversely desynchronization into multiple clusters increasesmessage variability since each cluster will produce independent information Hence a neuralarray which switches dynamically between a range of cluster configurations can be seen asadaptive in the sense that high synchronous states can be expressed when error detectabilityis a priority (such as during quick and reliable execution of a familiar task) but the array canrapidly desynchronize if variability becomes salient (for example if the environment suddenlybecomes novel) A formal operationalized demonstration of this principle is to be the subjectof future work
6 Discussion
In this paper we present a dynamical model for activity in an array of weakly coupledlocal neural assemblies The model incorporates the fundamental principles governing cellmembrane potential as determined by voltage- and ligand-gated ion channels In this wayit allows a reasonable amount of physiological complexity to be incorporated includingsynaptic activation and receptor subtypeswithout recourse to overwhelming detail Numericalanalysis of this model reveals a transition from independent oscillations to stable chaoticsynchronization depending on the balance of local versus long-range connectivity andbiophysical heterogeneity These excitatory couplings are subject to ascending subcorticalneuromodulation arising principally from monoaminergic nuclei such as Locus Coeruleus(Silberstein 1995) Neuromodulation of extrinsic connections was incorporated into the modelvia a coupling parameter C The stability of the synchronization is determined by the valueof the largest transverse Lyapunov exponent which in turn depends on the coupling parameterC If the coupling is weakened with the result that this exponent becomes positive a blowoutbifurcation occurs and independent oscillations re-emerge In the vicinity of the transitionnon-linear desynchronizations with complex temporal and spatial properties are observed
It is pleasing that complex and itinerant behaviour reminiscent of real neuronaldynamics emerges through such a simple manipulation of a single dynamic control parameterFurthermore the ubiquitous projections of classical neuromodulatory systems in real brainssuggest a clear biological mechanism by which the brain could self-organize throughmoderating the complexity of its own dynamics There are many reasons that the complicateddynamics of the sort demonstrated in this paper may be neurobiologically important Wemention four
First the presence of multiple clusters of phase-synchronized subsystems as discussedabove may be a uniquely non-linear mechanism facilitating the lsquocarriagersquo and distinction ofmultiple concurrent dynamic cell assemblies (and hence cognitive processing) This may beimportant for perceptual grouping binding and transitions as suggested by others (eg von derMalsburg 1985 Gray and Singer 1989)
Second the facility to decouple and reconfigure dynamic cell assemblies througha neuromodulatory control parameter may be important for avoiding low-entropysynchronization manifolds In the absence of itinerancy synchronization would renderneuronal dynamics incapable of representing changes in sensory input or indeed cognitive stateFor example in the model it was observed that small perturbations to one column were ableto induce a global change in neuronal dynamics by disrupting a state of weak synchronization
Synchronization in a model of neuronal dynamics 729
Inevitably alternative dynamic configurations emerge epitomizing the importance of lsquolabilersquobrain dynamics (Friston 2000a)
Thirdly from a purely functionalist point of view the balance between functionalspecialization and functional integration may rely upon the sorts of mechanism modelledin this paper The brain appears to adhere to two principles of functional organization namelysegregation of function within cortical areas and sub-areas and the integration of these areasin perceptual synthesis cognition and sensorimotor integration (Tononi et al 1994 Fristonet al 1995) The balance between preserving specialized dynamics unique to each arearequired by specialization and the coupling among these areas mediating functional integrationmay be achieved by the neuromodulatory mechanisms described above Significantlythe spontaneous variation in the numbers of degrees of freedom (for example whenlow-dimensional synchronization manifolds are shadowed) imparts a dynamic characteristicto this perspective that is precluded by linear mechanisms alone
Finally from a mechanistic perspective complexity and diversity of brain dynamics arerequired for selection of adaptive responses (Friston 2000b) There are many examples ofselective mechanisms operating within the brain All can be seen as performing an ascenton some objective function or fitness landscape These mechanisms require complex anditinerant dynamics to facilitate a thorough exploration of the landscape These landscapesmay range from those involved in perception (where the landscape is defined by predictionerror see eg Rao and Ballard (1998)) to those involved in value learning (where thelandscape is determined by the outcome of a behaviour eg in temporal difference models (seeeg Sutton and Barto 1990)) In short complexity is necessary to maintain an information-rich environment in which the brain can respond to sensory inputs An essential componentof this is the maintenance of diversity which is a prerequisite for selectionist mechanismsat many scales (Edelman 1993) Understanding the mechanistic basis of complex dynamicsas measured electrophysiologically may well depend on the sort of biologically plausiblemodels used in this paper This understanding may be important for both basic and clinicalneuroscience (eg Tass 2002)
The present study supplements and advances on other research in this field The studyof complex phase synchronization and clustering in neural oscillator networks similar tothe present model has previously been the subject of formal mathematical study Forexample Terman and Wang (1995) studied synchronization of blocks of neural oscillatorsthat were subject to perturbation by connected lsquostimulirsquo Somers and Kopell (1995) studiedthe emergence of both synchronization and travelling waves in networks of coupled neuraloscillators In the work of Terman and Wang the local dynamics were based on neurallsquocaricaturersquo models such as the FitzHughndashNagumo equations and are hence more abstract thanthe current study The approach of Somers and Kopell only required that the local dynamicsexhibit fast and relaxation phases and is hence more abstract still In both studies onlyperiodic dynamics are studied Complex patterns of chaos synchronization in neural arrayshave been characterized by nonlinear methods (Kaneko 1998) and their putative contributionto both learning (Nakajima and Ueda 1996) and perception (Zhao and Macau 2001 Breakspear2001) explored However these studies employed models such as the logistic equation andtent map which are quite abstract in comparison to the present study On the other handHansel and Sompolinsky (1996) studied the role of chaos synchronization in a model ofvisual perception in a cortical hypercolumn using a very similar approach to that adoptedhere but with the additional complexity that synaptic delays were incorporated However inthis and earlier work (Hansel and Sompolinsky 1992) only linear measures of dynamics andsynchrony (specifically the auto- and cross-correlation functions) were employed The analysisof chaotic synchrony in the study of Larter et al (1999) was based only upon observation of
730 M Breakspear et al
the Euclidean difference between variables from two nodes within a larger array Hence thesignificance of the present work is that it provides a quantitative nonlinear analysis of complexchaos synchronization in a biophysical model of cortical dynamics Such research is requiredin order to bridge the gap between quantitative nonlinear research into synchronization andcomputation in abstract models of neural arrays and qualitative or linear analysis in morebiophysically plausible models
Acknowledgments
The authors would like to thank Peter Robinson for assistance with the manuscript and R Larterfor helpful advice MB is a recipient of a NSW Institute of Psychiatry Research Grant anda University of Sydney SESQUI postdoctoral fellowship KJF is supported by the WellcomeTrust
References
Abarbanel H Rulkov N and Sushchik M 1996 Generalised synchronization of chaos the auxiliary system approachPhys Rev E 53 4528ndash35
Afraimovich V Verichev N and Rabinovich M 1986 Stochastic synchronization of oscillation in dissipative systemsRadiophys Quantum Electron 29 795ndash801
Breakspear M 2001 Perception of odours by a nonlinear model of the olfactory bulb Int J Neural Syst 11 101ndash24Breakspear M 2002 Nonlinear phase desynchronisation in human electroencephalographic data Human Brain Mapping
15 175ndash98Breakspear M and Terry J R 2002a Nonlinear interdependence in neural systems motivation theory and relevance
Int J Neurosci 112 1163ndash84Breakspear M and Terry J R 2002b Detection and description of nonlinear interdependence in normal multichannel
human EEG Clin Neurophysiol 113 735ndash53Breakspear M and Terry J R 2002c Topographic organization of nonlinear interdependence in multichannel human
EEG Neuroimage 16 822ndash35Destexhe A Rudolph M Fellous J M and Sejnowski T J 2001 Fluctuating synaptic conductances recreate in vivo-like
activity in neocortical neurons Neuroscience 107 13ndash24Douglas R J Mahowald M A and Martin K A 1996 Microarchitecture of neocortical columns Brain TheorymdashBiological
Basis and Computational Principles ed A Aertsen and V Braitenberg (Amsterdam Elsevier)Eckhorn R Bauer R Jordan W Brosch M Kruse W Munk M and Reitboeck H J 1988 Coherent oscillations a
mechanism of feature linking in the visual cortex Multiple electrode and correlation analyses in the cat BiolCybern 60 121ndash30
Edelman G M 1993 Neural Darwinism selection and reentrant signalling in higher brain function Neuron 10 115ndash25Engel A K Kreiter A K Konig P and Singer W 1991 Synchronization of oscillatory neuronal responses between
striate and extrastriate visual cortical areas of the cat Proc Natl Acad Sci USA 88 6048ndash52Friston K J 2000a The labile brain I Neuronal transients and nonlinear coupling Phil Trans R Soc B 355 215ndash36Friston K J 2000b The labile brain II Transients complexity and selection Phil Trans R Soc B 355 237ndash52Friston K J Tononi G Sporns O and Edelman G 1995 Characterising the complexity of neuronal interactions Human
Brain Mapping 3 302ndash14Fujisaka H and Yamada T 1983 Stability theory of synchronized motion in coupled-oscillator system Prog Theor
Phys 69 32ndash47Gatlin L 1972 Information Theory and the Living System (New York Columbia University Press)Gray C M Konig P Engel A and Singer W 1989 Oscillatory responses in cat visual cortex exhibit inter-columnar
synchronization which reflects global stimulus properties Science 338 334ndash7Gray C M and Singer W 1989 Stimulus specific neuronal oscillations in orientation columns of cat visual cortex Proc
Natl Acad Sci USA 86 1698ndash702Haig A R Gordon E Wright J J Meares R A and Bahramali H 2000 Synchronous cortical gamma-band activity in
task-relevant cognition Neuroreport 11 669ndash75Hansel D and Sompolinsky H 1992 Synchronization and computation in a chaotic neural network Phys Rev Lett 68
718ndash21
Synchronization in a model of neuronal dynamics 731
Hansel D and Sompolinsky H 1996 Chaos and synchrony in a model of a hypercolumn in visual cortex J ComputNeurosci 3 7ndash4
Heagy J Carroll T and Pecora L 1998 Desynchronization by periodic orbits Phys Rev E 57 R1253ndash6Hunt B Ott E and Yorke J 1997 Differentiable synchronization of chaos Phys Rev E 55 4029ndash34Josic K 1998 Invariant manifolds and synchronization of coupled dynamical systems Phys Rev Lett 80 3053ndash6Kandel E Schwartz J and Jessell T 2000 Principles of Neural Science (New York McGraw-Hill)Kaneko K 1998 On the strength of attractors in a high-dimensional system Physica D 124 322ndash44Kocarev L Parlitz U and Brown R 2000 Robust synchronization of chaotic systems Phys Rev E 61 3716ndash20Larter R Speelman B and Worth R M 1999 A coupled ordinary differential equation lattice model for the simulation
of epileptic seizures Chaos 9 795ndash804Lumer E Edelman G and Tononi G 1997a Neural dynamics in a model of the thalamocortical system I Layers loops
and the emergence of fast synchronous oscillations Cereb Cortex 7 207ndash27Lumer E Edelman G and Tononi G 1997b Neural dynamics in a model of the thalamocortical system II The role of
neural synchrony tested through perturbations of spike timing Cereb Cortex 7 228ndash36Magleby K 1987 Short-term modulation at synapses Synaptic Function ed G M Edelman W E Gall and
W M Cowan (New York Wiley)Maistrenko Y Maistrenko V Popovich A and Mosekilde E 1998 Transverse instability and riddled basins in a system
of two coupled logistic maps Phys Rev E 57 2713ndash24Morris C and Lecar H 1981 Voltage oscillations in the barnacle giant muscle fiber Biophys J 35 193ndash213Mountcastle V B 1997 The columnar organization of the neocortex Brain 120 701ndash22Mountcastle V B 1998 The Cerebral Cortex (Cambridge MA Harvard University Press)Nakajima H and Ueda Y 1996 Riddled basins of the optimal states in learning dynamical systems Physica D 99 35ndash44Nunez P L 1995 Neocortical Dynamics and Human Brain Rhythms (Oxford Oxford University Press)Osipov G V Pikovsky A S Rosenblum M G and Kurths J 1997 Phase synchronization effects in a lattice of nonidentical
Rossler oscillators Phys Rev E 55 2353ndash61Ott E and Sommerer J 1994 Blowout bifurcations the occurrence of riddled basins and onndashoff intermittency Phys
Lett A 188 39ndash47Pecora L and Carroll T 1990 Synchronization in chaotic systems Phys Rev Lett 64 821ndash4Pikovsky A and Grassberger P 1991 Symmetry breaking bifurcation for coupled chaotic attractors J Phys A Math
Gen 24 4587ndash97Rao R P and Ballard D H 1998 Predictive coding in the visual cortex a functional interpretation of some extra-classical
receptive field effects Nature Neurosci 2 79ndash87Robinson P A Wright J J and Rennie C R 1998 Synchronous oscillations in the cerebral cortex Phys Rev E 57
4578ndash88Rodriguez E George N Lachauz J P Martinerie J Renault B and Varela F J 1999 Perceptionrsquos shadow long distance
synchronization of human brain activity Nature 397 430ndash3Rosenblum M Pikovsky A and Kurths J 1996 Phase synchronization of chaotic oscillators Phys Rev Lett 76 1804ndash7Rosenblum M Pikovsky A and Kurths J 1997 From phase to lag synchronization in coupled chaotic oscillators Phys
Rev Lett 78 4193ndash6Rulkov N and Sushchik M 1997 Robustness of synchronized chaotic oscillations Int J Bifurc Chaos 7 625ndash43Rulkov N Sushchik M Tsimring L and Abarbanel H 1995 Generalized synchronization of chaos in unidirectionally
coupled chaotic systems Phys Rev E 51 980ndash94Schiff S So P Chang T Burke R and Sauer T 1996 Detecting dynamical interdependence and generalized synchrony
through mutual prediction in a neural ensemble Phys Rev E 54 6708ndash24Silberstein R B 1995 Neuromodulations of neocortical dynamics Neocortical Dynamics and Human EEG Rhythms
ed P L Nunez (Oxford Oxford University Press)Singer W 1995 Putative functions of temporal correlations in neocortical processing Large-Scale Neuronal Theories
of the Brain ed C Koch and J Davis (London MIT Press)Somers D and Kopell N 1995 Waves and synchrony in networks of oscillators of relaxation and non-relaxation type
Physica D 89 169ndash83Sutton R S and Barto A G 1990 Time derivative models of Pavlovian reinforcement Learning and Computational
Neuroscience Foundations of Adaptive Networks ed M Gabriel and J Moore (Cambridge MA MIT Press) pp497ndash538
Szentagothai J 1983 The modular architectonic principle of neural centers Rev Physiol Biochem Pharmacol 9811ndash61
Tallon-Baudry C Betrand O and Fischer C 2001 Oscillatory synchrony between human extrastriate areas during visualshort-term memory maintenance J Neurosci 21 RC177(1ndash5)
Tass P A 2002 Desynchronization of brain rhythms with soft phase-resetting techniques Biol Cybern 87 102ndash15
732 M Breakspear et al
Tass P Rosenblum M Weule J Kurths J Pikovsky A Volkmann J Schnitzler A and Freund H 1998 Detection of nmphase locking from noisy data application to magnetoencephalography Phys Rev Lett 81 3291ndash4
Terman D and Wang D 1995 Global competition and local cooperation in a network of neural oscillators Physica D81 148ndash76
Terry J and Breakspear M 2003 An improved algorithm for the detection of dynamical interdependence in bivariatetime-series Biol Cybern 88 129ndash36
Tononi G Sporns O and Edelman G M 1994 A measure for brain complexity relating functional segregation andintegration in the nervous system Proc Natl Acad Sci USA 91 5033ndash7
von der Malsburg C 1985 Nervous structures with dynamical links Phys Chem 89 703ndash10Wang X 1991 Period-doublings to chaos in a simple neural network an analytic proof Complex Syst 5 425ndash41Wennekers T and Pasemann F 1996 Synchronous chaos in high dimensional modular neural networks Int J Bifurc
Chaos 6 2055ndash67Zaks M A Parks E H Rosenblum M G and Kurths J 1999 Alternative locking ratios in imperfect phase synchronization
Phys Rev Lett 82 4228ndash31Zhao L and Macau E N 2001 A network of dynamically coupled chaotic maps for scene segmentation IEEE Trans
Neural Netw 12 1375ndash85
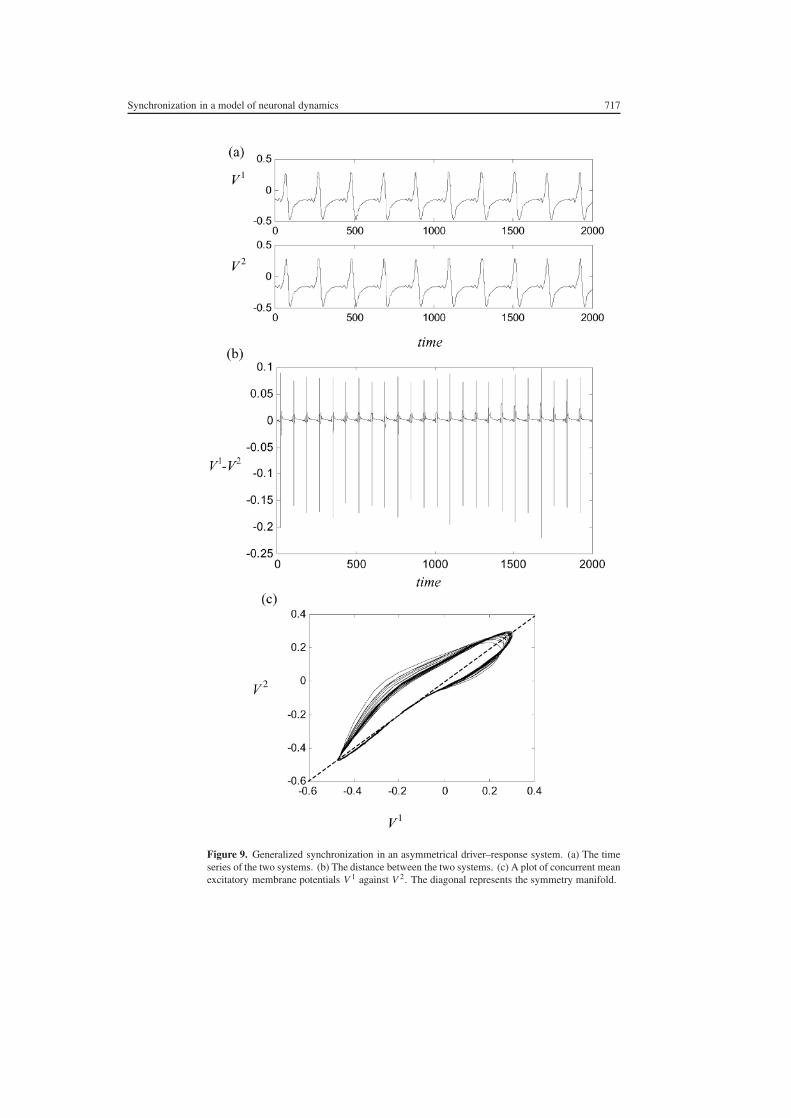
Synchronization in a model of neuronal dynamics 717
1
2
2
1
Figure 9 Generalized synchronization in an asymmetrical driverndashresponse system (a) The timeseries of the two systems (b) The distance between the two systems (c) A plot of concurrent meanexcitatory membrane potentials V 1 against V 2 The diagonal represents the symmetry manifold
718 M Breakspear et al
Hunt et al (1997) Josic (1998) Kocarev et al (2000) have shown that the generalizedsynchronization manifold is smooth (differentiable) if and only if it is normally hyperbolic(ie the largest transverse Lyapunov exponent is less than the smallest tangential Lyapunovexponent prior to the introduction of parameter mismatch) Such a condition is met in thepresent setting when C = 04 hence the smoothness of the manifold in panel (c)
However subsequent to the introduction of parameter mismatch we will not be able touse transverse and tangential Lyapunov exponents to detect such synchronization since thesynchronization manifold may possess a complex topology for which it is a non-trivial (atbest) or impossible (at worse) task to define transverse and tangential directions For examplethe manifold may be smooth as in figure 9(c) but have a fractal topology If the dynamics arenot normally hyperbolic then the manifold may be fractal but also non-differentiable (Huntet al 1997) and hence will not have a unique tangent direction at any point Consequently wewill not be able to compute tangential or transverse Lyapunov exponents and hence determinethe stability of the synchronization manifold However other techniques for detecting thepresence of generalized synchronization in complex systems have evolved including theauxiliary system approach (Abarbanel et al 1996) and non-linear interdependence (Schiffet al 1996) which are now explored
421 The auxiliary system approach The auxiliary system is a third subsystem which isa replica of the original response system but coupled independently to the driver system andcommencing from independent initial conditions It is possible to demonstrate that showingthe existence of generalized synchronizationmdashthat equation (10) is satisfiedmdashis equivalent toshowing that the response system and the auxiliary system evolve identically (even though theyare not directly coupled) To determine whether or not this is the case reduces to an analysisof the Lyapunov exponents transverse to the symmetry manifold with respect to the responsesystem and the auxiliary system
Whilst this is a somewhat artificial test for generalized synchronization since theproduction of an auxiliary system for a physical system would be impossible such a testis useful in numerical and theoretical studies In figure 10 we present an example of theauxiliary system approach where the parameter δV = 07 for column 1 and 068 for column 2We consider two values of the coupling C = 0025 and 01 The auxiliary system is simplya copy of the second column beginning from an arbitrary initial state In the first instancewhere C = 0025 we notice that even after allowing for an initial transient state the secondcolumn and the auxiliary do not behave in identical fashions (figure 10(a)) However whenC = 01 we observe (figure 10(b)) that the second column and the auxiliary do behave innear-identical manners after an initial transient and hence conclude that for the particularvalue of the coupling strength considered equation (10) is satisfied and hence generalizedsynchronization between the first and second columns occurs
422 Nonlinear interdependence The question remains as to how we may detect thepresence of generalized synchronization in a practical setting As we mentioned aboveit is not possible to use an auxiliary-system-based approach to detect the presence ofgeneralized synchronization Instead we must consider techniques for detecting suchbehaviour from experimental or numerical time series data Algorithms of this type areoften referred to as detecting non-linear interdependence Nonlinear interdependence is ageneral term encompassing systems where the behaviour of one non-linear system affectsthe behaviour of another More specifically it can be defined as occurring if the nullhypothesis that the interdependence between two signals is purely linear can be rejectedat conventional levels (usually 95) of statistical significance hence it is an experimental
Synchronization in a model of neuronal dynamics 719
2
1
2
1
Figure 10 Concurrent evolution of the response system V 2 and lsquoauxiliaryrsquo system V 1 (a) C =0025 (b) C = 01
term (eg Schiff et al 1996) Whilst this does not strictly correspond to the existence of afunctional relationship between the two systems such a relationship is typically used tomotivate the techniques for its detection Schiff et al (1996) extended the logic of non-linearprediction algorithms from autonomous to coupled systems by exploiting the continuity ofsuch a relationship and hence devised the first method of detecting non-linear interdependencein time series data The basis of the technique is using the forward evolution of a phase spacevectorrsquos neighbours to predict the forward evolution of that vector and the forward evolutionof the concurrent vector of the other (possibly interdependent) system If the error of such aprocedure is better on average than chance (the chance error is normalized to equal one) thennon-linear interdependence is said to be present
720 M Breakspear et al
V1
V2
Figure 11 Experimental and numerical examples of weak generalized non-linear synchronizationin the presence of measurement noise (a) Human scalp EEG data from posterior bipolar scalpelectrodes (O1P3ndashO2P4) (b) Weakly coupled aperiodic data in the present model with additivewhite noise (with 5 of the amplitude of the model signal) Parameters δ1
V = 0605 δ2V = 061
aee = 036 C = 007
A variation of this algorithm optimized to detect weak non-linear interdependence (Terryand Breakspear 2003) was used recently to demonstrate that weak non-linear interdependence isoccasionally present in the scalp EEG of healthy resting human subjects (Breakspear and Terry2002a 2002b 2002c) An example of an epoch of lsquonon-linearly independentrsquo EEG obtainedusing this technique is presented in figure 11(a) For comparison output from the currentmodel is presented in figure 11(b) The model parameters were set such that the dynamicswas still aperiodic but the intermittent large-amplitude bursts were suppressed and had weakmutual coupling (C = 007) Extraneous noise to simulate the effect of extracranial muscleactivity contaminating the EEG data has been added to this time series It should be notedthat the amplitudes and time scales of the modelrsquos variables are dimensionless Nonethelessthe model facilitates qualititative interpretation of experimental findings
Synchronization in a model of neuronal dynamics 721
43 Chaotic phase synchronization
Whereas the concept of generalized synchronization was mathematically motivated and assuch more suited to theoretical investigations of systems the concept of phase synchronizationderives from a more practical signal processing approach Phase synchronization occurs inboth symmetric and asymmetric settings but is often more useful in the asymmetric case (andin experimental data)
Unfortunately the phase of a chaotic signal is typically ambiguous and can be defined in anumber of different ways For example in the case of the coherent Rossler attractor (Osipovet al 1997) the phase may be defined as the arctangent of two of the variables of the systemMore generally it can sometimes be defined as the angle that an orbit subtends with the centreof an attractor It is also possible to extract the phase of a chaotic signal from time seriesdata by use of the Hilbert transform This has enabled the demonstration of chaotic phasesynchronization in a number of coupled asymmetrical systems (Rosenblum et al 1996 1997Zaks et al 1999) Although such an approach does not guarantee that the phase will be uniqueit does permit the practical analysis of phase in noisy time series (Tass et al 1998)
To illustrate phase synchronization in two coupled columns we again vary the parameterδV which if the columns are viewed as oscillators acts to detune them from a commonfrequency An important question is that of the criteria on the basis of which we concludewhether phase synchronization occurs We use a simple locking criterion whereby phasesynchronization is said to occur if
|θ1(t)minus θ2(t)| lt K (11)
where is θi the phase of column i and K is a constant Notice that this does not mean thatsynchronization implies equality of phases merely that their difference is bounded
An example of phase synchronization with δV = 065 for column 1 and 066 for column 2is given in figure 12 In this figure rNMDA = 03 Qualitatively some sort of phase coherenceis evident in the time series panel (a) This was confirmed by plotting the sine of thephase difference which has been extracted using the Hilbert transform panel (b) Clearly|sin(θ1 minus θ2)| lt K and hence |θ1 minus θ2| is bounded It can also be seen that the phase-synchronized dynamics occurs on a highly structured manifold depicted in figure 12(c)However unlike the case for the generalized synchronization manifold depicted in figure 9 itis evident that this manifold is not a simple transformation of the symmetry manifold
It is also possible to see lsquobimodalrsquo stability whereby a system jumps betweenapproximately zero phase difference and a phase lag phase synchronization of π An exampleis depicted in figure 12(d) At approximately 2000 s the system can be seen to jumpspontaneously from the first to the second of these modes The Z -variable is plotted forgraphical purposes
5 Intermittency and desynchronization
51 Desynchronization in an array of two systems
Diminishing the strength of coupling across the blowout bifurcation does not lead to a simpletransition from synchronous to asynchronous oscillations but is instead associated withcomplex temporal behaviours Specifically when the largest transverse Lyapunov exponent isclose to zero the coupled systems exhibit epochs of synchronous behaviours punctuated bysharp bursts of desynchronization This is observed in systems exhibiting identicalgeneralizedand phase synchronization An example of such a phenomenon is given in figure 13 The timeseries panel (a) shows synchronous dynamics except at time = 1000 where it can be seenthat the two time series desynchronize briefly This is evident in panel (b) as a disruption in the
722 M Breakspear et al
2
1
Figure 12 Phase-synchronized coupled columns (a) The time series of V 1 (solid) and V 2 (b) Thesine of the phase difference obtained through Hilbert transformation of the time series in panel (a)(c) The phase synchronization manifold in the V 1ndashV 2 plane (d) The time series of Z (solid) andZ showing a phase slip of π between two modes
Synchronization in a model of neuronal dynamics 723
Figure 13 An example of phase desynchronization in coupled asymmetric systems with C = 01Time series plots of (a) each V -variable and (b) their Euclidean distance (c) The temporal evolutionof the phase difference between the two systems (d) An example of the phase difference evolutionover a longer time interval
724 M Breakspear et al
orderly evolution of the distance between the two systems and in panel (c) where the evolutionof the phase difference reveals a brief violation of the criterion |φ1 minus φ2| lt constant Anexample of the phase difference evolution over a longer time interval is shown in figure 13(d)Note that this is a three-dimensional plot with sin(φ1 minus φ2) and cos(φ1 minus φ2) as they- and z-axes Hence phase desynchronizations are evident as complete revolutions aroundthe cylindrical phase space in which this variable evolves It can be seen that these phenomenaoccur at irregular intervals in the temporal domain
This behaviour can be explained by considering the nature of a chaotic attractor In thestatistical sense an arbitrary initial condition is expected to lead to the chaotic behaviour onthe attractor Consequently when we calculate the Lyapunov exponents of the system theseexponents correspond to those of the chaotic attractor Typically however there are an infinitenumber of unstable periodic orbits embedded within the attractor If the path of a trajectorybecomes close to one of these orbits it may lsquoshadowrsquo the orbit for a brief period of time Eachof these orbits has a corresponding set of Lyapunov exponents which (like the attractor) can bedivided into tangential and transverse exponents These also vary when we change the couplingparameter but are typically not the same as the exponents of the attractor Hence the largesttransverse exponent of the attractor may be negative yet there may be positive transverseexponents associated with periodic orbits Considering all of these exponents together wewould therefore only expect to observe complete stability of the synchronization manifold inthe case where all of the transverse Lyapunov exponents are negative Bursting behaviourwill be observed when the one of these exponents becomes positive typically correspondingto a periodic orbit of low order (Pikovsky and Grassberger 1991 Rulkov and Sushchik 1997Maistrenko et al 1998 Heagy et al 1998) Due to the complex structure of such periodic orbitsthe desynchronous bursts occur across a wide range of timescales
In figure 14 we present the results of a long integration (16times105 time units) in which thelengths of sequential synchronous epochs of phase synchronization are studied The parametervalues were δV = 065 for column 1 and 0645 for column 2 with coupling strength10 equal to0125 In panel (a) the power spectrum of these sequential lengths is plotted The whiteness ofthis spectrum indicates that the lengths of consecutive synchronous epochs are independentThis is due to the high-dimensional lsquomixingrsquo effect of desynchronization In panel (b) thelengths of the synchronous epochs have been sorted again into the order of increasing lengthThe results for two coupling strengths are plotted C = 0125 (solid) and 0135 (dashed)Several features of this plot are of note Firstly the lengths of the synchronous epochsare distributed over almost three orders of magnitude Secondly for both plots there is anapproximately log-linear relationship between the length of an epoch and its relative orderThis reflects an approximately exponential relationship between the length of a synchronousepoch and its likelihood of occurrence Thirdly this relationship is skewed toward longersynchronous epochs for the stronger coupling strengths
This type of signal with patches of lsquoorderrsquo occurring over many timescales interruptedby bursts of disorder can be shown to have a high information capacity (Gatlin 1972)Moreover complex phase desynchronization may make an infrequent yet robust contributionto the dynamical pattern of the alpha rhythm in scalp EEG data for resting human subjects(Breakspear 2002) It is tempting to postulate that such a lsquoweakly stablersquo dynamics may assistin the generation of the rich and fluid mental associations that spontaneously emerge in restingcognitive states Although this phenomenon has been previously described in a variety of othersystems to our knowledge this is the first description in a biophysical model of neocortex
10 The introduction of parameter mismatch causes an increase in the strength of coupling required for stablesynchronization above the value of 01 required in the symmetric case
Synchronization in a model of neuronal dynamics 725
Figure 14 Properties of sequential epochs of phase-synchronous behaviour over a long integrationtime δ1
V = 065 δ2V = 0645 aee = 04 (a) The flat (white) power spectrum (b) Synchronous
epochs sorted in order of increasing length (solid C = 0125 dashed C = 0134)
52 Synchronization and complex dynamics in larger arrays
The synchronous and complex phenomena described above are also observed in arrays ofcoupled systems with N gt 2 Expanding the size of the array permits complex dynamicsin the spatial in addition to the temporal domain However the basic principles governingthe type and stability of synchronized motions are the same In this section a few of thepossible behaviours are discussed Physiological variation across the array is modelled bymodifying the parameter δV = 07 in each array by adding a random number of maximummagnitude 001
In figure 15(a) an array of N = 20 uncoupled (C = 0) systems is shown The lack ofsynchronization in this time series is evident By comparison the rapid transition to completesynchronization is evident in figure 15(b) when C = 03 In fact 19 of the 20 subsystems
726 M Breakspear et al
(b)
(c)
(a)
Figure 15 Synchronization phenomena in an array of coupled cortical columns (a) N = 20C = 0 (b) N = 20 C = 03 (c) N = 6 C = 01 (d) N = 12 C = 009 (e) N = 12C = 012 A small perturbation has been applied to one of the columns at t = 2500 s (f) Evolutionof the phase difference between two of the columns in panel (e) For clarity the W -variable isplotted in panel (c) and the Z -variable in (e)
have phase synchronized within one single cycle Intermediate values of coupling strength areassociated with a rich variety of phenomena For example subsystems with the most similarparameter values typically synchronize into clusters of various sizes An example of this isillustrated in figure 15(c) Note that there is no synchronization between the two differentclusters each containing three synchronized subsystems
Another complex behaviour occurs when subsystems transiently synchronize into largeclusters then spontaneously desynchronize again Hence the dynamics of the entire arrayjumps between different transient dynamics characterized by varying degrees of freedom Anexample of this phenomenon is given in figure 15(d) In addition it is possible to inducedesynchronization across an entire array by applying a small perturbation to one columnfor one time step An example is illustrated in figure 15(e) A small perturbation has been
Synchronization in a model of neuronal dynamics 727
Figure 15 (Continued)
added to one column after 2500 time units It can be seen that the entire array subsequentlydesynchronizes This illustrates the sensitivity of the dynamics to external stimuli Finally it ispossible to lsquoinducersquo synchronization between selected subsystems by increasing the strength ofcoupling between them selectively This allows an extra level of complexity to be introducedinto the model For example it may allow targeted information transfer between a partialnetwork of the entire array
As discussed in section 51 a univariate signal which jumps between epochs of lowentropy (high repetition) and bursts of high entropy (high variability) can be shown to havehigh information capacity This is because the repetition of certain patterns (as in the English
728 M Breakspear et al
language) permits error detectability Bursts of high entropy act in the opposite directionto increase message variability With a multivariate signal in addition to optimizing theinformation capacity within each signal the overall information capacity can be increasedthrough the type of itinerant synchronization briefly illustrated in this section Synchronizationacross an entire array increases the message reliability (since a single lsquoerrant letterrsquo in onesignal can be easily detected) Conversely desynchronization into multiple clusters increasesmessage variability since each cluster will produce independent information Hence a neuralarray which switches dynamically between a range of cluster configurations can be seen asadaptive in the sense that high synchronous states can be expressed when error detectabilityis a priority (such as during quick and reliable execution of a familiar task) but the array canrapidly desynchronize if variability becomes salient (for example if the environment suddenlybecomes novel) A formal operationalized demonstration of this principle is to be the subjectof future work
6 Discussion
In this paper we present a dynamical model for activity in an array of weakly coupledlocal neural assemblies The model incorporates the fundamental principles governing cellmembrane potential as determined by voltage- and ligand-gated ion channels In this wayit allows a reasonable amount of physiological complexity to be incorporated includingsynaptic activation and receptor subtypeswithout recourse to overwhelming detail Numericalanalysis of this model reveals a transition from independent oscillations to stable chaoticsynchronization depending on the balance of local versus long-range connectivity andbiophysical heterogeneity These excitatory couplings are subject to ascending subcorticalneuromodulation arising principally from monoaminergic nuclei such as Locus Coeruleus(Silberstein 1995) Neuromodulation of extrinsic connections was incorporated into the modelvia a coupling parameter C The stability of the synchronization is determined by the valueof the largest transverse Lyapunov exponent which in turn depends on the coupling parameterC If the coupling is weakened with the result that this exponent becomes positive a blowoutbifurcation occurs and independent oscillations re-emerge In the vicinity of the transitionnon-linear desynchronizations with complex temporal and spatial properties are observed
It is pleasing that complex and itinerant behaviour reminiscent of real neuronaldynamics emerges through such a simple manipulation of a single dynamic control parameterFurthermore the ubiquitous projections of classical neuromodulatory systems in real brainssuggest a clear biological mechanism by which the brain could self-organize throughmoderating the complexity of its own dynamics There are many reasons that the complicateddynamics of the sort demonstrated in this paper may be neurobiologically important Wemention four
First the presence of multiple clusters of phase-synchronized subsystems as discussedabove may be a uniquely non-linear mechanism facilitating the lsquocarriagersquo and distinction ofmultiple concurrent dynamic cell assemblies (and hence cognitive processing) This may beimportant for perceptual grouping binding and transitions as suggested by others (eg von derMalsburg 1985 Gray and Singer 1989)
Second the facility to decouple and reconfigure dynamic cell assemblies througha neuromodulatory control parameter may be important for avoiding low-entropysynchronization manifolds In the absence of itinerancy synchronization would renderneuronal dynamics incapable of representing changes in sensory input or indeed cognitive stateFor example in the model it was observed that small perturbations to one column were ableto induce a global change in neuronal dynamics by disrupting a state of weak synchronization
Synchronization in a model of neuronal dynamics 729
Inevitably alternative dynamic configurations emerge epitomizing the importance of lsquolabilersquobrain dynamics (Friston 2000a)
Thirdly from a purely functionalist point of view the balance between functionalspecialization and functional integration may rely upon the sorts of mechanism modelledin this paper The brain appears to adhere to two principles of functional organization namelysegregation of function within cortical areas and sub-areas and the integration of these areasin perceptual synthesis cognition and sensorimotor integration (Tononi et al 1994 Fristonet al 1995) The balance between preserving specialized dynamics unique to each arearequired by specialization and the coupling among these areas mediating functional integrationmay be achieved by the neuromodulatory mechanisms described above Significantlythe spontaneous variation in the numbers of degrees of freedom (for example whenlow-dimensional synchronization manifolds are shadowed) imparts a dynamic characteristicto this perspective that is precluded by linear mechanisms alone
Finally from a mechanistic perspective complexity and diversity of brain dynamics arerequired for selection of adaptive responses (Friston 2000b) There are many examples ofselective mechanisms operating within the brain All can be seen as performing an ascenton some objective function or fitness landscape These mechanisms require complex anditinerant dynamics to facilitate a thorough exploration of the landscape These landscapesmay range from those involved in perception (where the landscape is defined by predictionerror see eg Rao and Ballard (1998)) to those involved in value learning (where thelandscape is determined by the outcome of a behaviour eg in temporal difference models (seeeg Sutton and Barto 1990)) In short complexity is necessary to maintain an information-rich environment in which the brain can respond to sensory inputs An essential componentof this is the maintenance of diversity which is a prerequisite for selectionist mechanismsat many scales (Edelman 1993) Understanding the mechanistic basis of complex dynamicsas measured electrophysiologically may well depend on the sort of biologically plausiblemodels used in this paper This understanding may be important for both basic and clinicalneuroscience (eg Tass 2002)
The present study supplements and advances on other research in this field The studyof complex phase synchronization and clustering in neural oscillator networks similar tothe present model has previously been the subject of formal mathematical study Forexample Terman and Wang (1995) studied synchronization of blocks of neural oscillatorsthat were subject to perturbation by connected lsquostimulirsquo Somers and Kopell (1995) studiedthe emergence of both synchronization and travelling waves in networks of coupled neuraloscillators In the work of Terman and Wang the local dynamics were based on neurallsquocaricaturersquo models such as the FitzHughndashNagumo equations and are hence more abstract thanthe current study The approach of Somers and Kopell only required that the local dynamicsexhibit fast and relaxation phases and is hence more abstract still In both studies onlyperiodic dynamics are studied Complex patterns of chaos synchronization in neural arrayshave been characterized by nonlinear methods (Kaneko 1998) and their putative contributionto both learning (Nakajima and Ueda 1996) and perception (Zhao and Macau 2001 Breakspear2001) explored However these studies employed models such as the logistic equation andtent map which are quite abstract in comparison to the present study On the other handHansel and Sompolinsky (1996) studied the role of chaos synchronization in a model ofvisual perception in a cortical hypercolumn using a very similar approach to that adoptedhere but with the additional complexity that synaptic delays were incorporated However inthis and earlier work (Hansel and Sompolinsky 1992) only linear measures of dynamics andsynchrony (specifically the auto- and cross-correlation functions) were employed The analysisof chaotic synchrony in the study of Larter et al (1999) was based only upon observation of
730 M Breakspear et al
the Euclidean difference between variables from two nodes within a larger array Hence thesignificance of the present work is that it provides a quantitative nonlinear analysis of complexchaos synchronization in a biophysical model of cortical dynamics Such research is requiredin order to bridge the gap between quantitative nonlinear research into synchronization andcomputation in abstract models of neural arrays and qualitative or linear analysis in morebiophysically plausible models
Acknowledgments
The authors would like to thank Peter Robinson for assistance with the manuscript and R Larterfor helpful advice MB is a recipient of a NSW Institute of Psychiatry Research Grant anda University of Sydney SESQUI postdoctoral fellowship KJF is supported by the WellcomeTrust
References
Abarbanel H Rulkov N and Sushchik M 1996 Generalised synchronization of chaos the auxiliary system approachPhys Rev E 53 4528ndash35
Afraimovich V Verichev N and Rabinovich M 1986 Stochastic synchronization of oscillation in dissipative systemsRadiophys Quantum Electron 29 795ndash801
Breakspear M 2001 Perception of odours by a nonlinear model of the olfactory bulb Int J Neural Syst 11 101ndash24Breakspear M 2002 Nonlinear phase desynchronisation in human electroencephalographic data Human Brain Mapping
15 175ndash98Breakspear M and Terry J R 2002a Nonlinear interdependence in neural systems motivation theory and relevance
Int J Neurosci 112 1163ndash84Breakspear M and Terry J R 2002b Detection and description of nonlinear interdependence in normal multichannel
human EEG Clin Neurophysiol 113 735ndash53Breakspear M and Terry J R 2002c Topographic organization of nonlinear interdependence in multichannel human
EEG Neuroimage 16 822ndash35Destexhe A Rudolph M Fellous J M and Sejnowski T J 2001 Fluctuating synaptic conductances recreate in vivo-like
activity in neocortical neurons Neuroscience 107 13ndash24Douglas R J Mahowald M A and Martin K A 1996 Microarchitecture of neocortical columns Brain TheorymdashBiological
Basis and Computational Principles ed A Aertsen and V Braitenberg (Amsterdam Elsevier)Eckhorn R Bauer R Jordan W Brosch M Kruse W Munk M and Reitboeck H J 1988 Coherent oscillations a
mechanism of feature linking in the visual cortex Multiple electrode and correlation analyses in the cat BiolCybern 60 121ndash30
Edelman G M 1993 Neural Darwinism selection and reentrant signalling in higher brain function Neuron 10 115ndash25Engel A K Kreiter A K Konig P and Singer W 1991 Synchronization of oscillatory neuronal responses between
striate and extrastriate visual cortical areas of the cat Proc Natl Acad Sci USA 88 6048ndash52Friston K J 2000a The labile brain I Neuronal transients and nonlinear coupling Phil Trans R Soc B 355 215ndash36Friston K J 2000b The labile brain II Transients complexity and selection Phil Trans R Soc B 355 237ndash52Friston K J Tononi G Sporns O and Edelman G 1995 Characterising the complexity of neuronal interactions Human
Brain Mapping 3 302ndash14Fujisaka H and Yamada T 1983 Stability theory of synchronized motion in coupled-oscillator system Prog Theor
Phys 69 32ndash47Gatlin L 1972 Information Theory and the Living System (New York Columbia University Press)Gray C M Konig P Engel A and Singer W 1989 Oscillatory responses in cat visual cortex exhibit inter-columnar
synchronization which reflects global stimulus properties Science 338 334ndash7Gray C M and Singer W 1989 Stimulus specific neuronal oscillations in orientation columns of cat visual cortex Proc
Natl Acad Sci USA 86 1698ndash702Haig A R Gordon E Wright J J Meares R A and Bahramali H 2000 Synchronous cortical gamma-band activity in
task-relevant cognition Neuroreport 11 669ndash75Hansel D and Sompolinsky H 1992 Synchronization and computation in a chaotic neural network Phys Rev Lett 68
718ndash21
Synchronization in a model of neuronal dynamics 731
Hansel D and Sompolinsky H 1996 Chaos and synchrony in a model of a hypercolumn in visual cortex J ComputNeurosci 3 7ndash4
Heagy J Carroll T and Pecora L 1998 Desynchronization by periodic orbits Phys Rev E 57 R1253ndash6Hunt B Ott E and Yorke J 1997 Differentiable synchronization of chaos Phys Rev E 55 4029ndash34Josic K 1998 Invariant manifolds and synchronization of coupled dynamical systems Phys Rev Lett 80 3053ndash6Kandel E Schwartz J and Jessell T 2000 Principles of Neural Science (New York McGraw-Hill)Kaneko K 1998 On the strength of attractors in a high-dimensional system Physica D 124 322ndash44Kocarev L Parlitz U and Brown R 2000 Robust synchronization of chaotic systems Phys Rev E 61 3716ndash20Larter R Speelman B and Worth R M 1999 A coupled ordinary differential equation lattice model for the simulation
of epileptic seizures Chaos 9 795ndash804Lumer E Edelman G and Tononi G 1997a Neural dynamics in a model of the thalamocortical system I Layers loops
and the emergence of fast synchronous oscillations Cereb Cortex 7 207ndash27Lumer E Edelman G and Tononi G 1997b Neural dynamics in a model of the thalamocortical system II The role of
neural synchrony tested through perturbations of spike timing Cereb Cortex 7 228ndash36Magleby K 1987 Short-term modulation at synapses Synaptic Function ed G M Edelman W E Gall and
W M Cowan (New York Wiley)Maistrenko Y Maistrenko V Popovich A and Mosekilde E 1998 Transverse instability and riddled basins in a system
of two coupled logistic maps Phys Rev E 57 2713ndash24Morris C and Lecar H 1981 Voltage oscillations in the barnacle giant muscle fiber Biophys J 35 193ndash213Mountcastle V B 1997 The columnar organization of the neocortex Brain 120 701ndash22Mountcastle V B 1998 The Cerebral Cortex (Cambridge MA Harvard University Press)Nakajima H and Ueda Y 1996 Riddled basins of the optimal states in learning dynamical systems Physica D 99 35ndash44Nunez P L 1995 Neocortical Dynamics and Human Brain Rhythms (Oxford Oxford University Press)Osipov G V Pikovsky A S Rosenblum M G and Kurths J 1997 Phase synchronization effects in a lattice of nonidentical
Rossler oscillators Phys Rev E 55 2353ndash61Ott E and Sommerer J 1994 Blowout bifurcations the occurrence of riddled basins and onndashoff intermittency Phys
Lett A 188 39ndash47Pecora L and Carroll T 1990 Synchronization in chaotic systems Phys Rev Lett 64 821ndash4Pikovsky A and Grassberger P 1991 Symmetry breaking bifurcation for coupled chaotic attractors J Phys A Math
Gen 24 4587ndash97Rao R P and Ballard D H 1998 Predictive coding in the visual cortex a functional interpretation of some extra-classical
receptive field effects Nature Neurosci 2 79ndash87Robinson P A Wright J J and Rennie C R 1998 Synchronous oscillations in the cerebral cortex Phys Rev E 57
4578ndash88Rodriguez E George N Lachauz J P Martinerie J Renault B and Varela F J 1999 Perceptionrsquos shadow long distance
synchronization of human brain activity Nature 397 430ndash3Rosenblum M Pikovsky A and Kurths J 1996 Phase synchronization of chaotic oscillators Phys Rev Lett 76 1804ndash7Rosenblum M Pikovsky A and Kurths J 1997 From phase to lag synchronization in coupled chaotic oscillators Phys
Rev Lett 78 4193ndash6Rulkov N and Sushchik M 1997 Robustness of synchronized chaotic oscillations Int J Bifurc Chaos 7 625ndash43Rulkov N Sushchik M Tsimring L and Abarbanel H 1995 Generalized synchronization of chaos in unidirectionally
coupled chaotic systems Phys Rev E 51 980ndash94Schiff S So P Chang T Burke R and Sauer T 1996 Detecting dynamical interdependence and generalized synchrony
through mutual prediction in a neural ensemble Phys Rev E 54 6708ndash24Silberstein R B 1995 Neuromodulations of neocortical dynamics Neocortical Dynamics and Human EEG Rhythms
ed P L Nunez (Oxford Oxford University Press)Singer W 1995 Putative functions of temporal correlations in neocortical processing Large-Scale Neuronal Theories
of the Brain ed C Koch and J Davis (London MIT Press)Somers D and Kopell N 1995 Waves and synchrony in networks of oscillators of relaxation and non-relaxation type
Physica D 89 169ndash83Sutton R S and Barto A G 1990 Time derivative models of Pavlovian reinforcement Learning and Computational
Neuroscience Foundations of Adaptive Networks ed M Gabriel and J Moore (Cambridge MA MIT Press) pp497ndash538
Szentagothai J 1983 The modular architectonic principle of neural centers Rev Physiol Biochem Pharmacol 9811ndash61
Tallon-Baudry C Betrand O and Fischer C 2001 Oscillatory synchrony between human extrastriate areas during visualshort-term memory maintenance J Neurosci 21 RC177(1ndash5)
Tass P A 2002 Desynchronization of brain rhythms with soft phase-resetting techniques Biol Cybern 87 102ndash15
732 M Breakspear et al
Tass P Rosenblum M Weule J Kurths J Pikovsky A Volkmann J Schnitzler A and Freund H 1998 Detection of nmphase locking from noisy data application to magnetoencephalography Phys Rev Lett 81 3291ndash4
Terman D and Wang D 1995 Global competition and local cooperation in a network of neural oscillators Physica D81 148ndash76
Terry J and Breakspear M 2003 An improved algorithm for the detection of dynamical interdependence in bivariatetime-series Biol Cybern 88 129ndash36
Tononi G Sporns O and Edelman G M 1994 A measure for brain complexity relating functional segregation andintegration in the nervous system Proc Natl Acad Sci USA 91 5033ndash7
von der Malsburg C 1985 Nervous structures with dynamical links Phys Chem 89 703ndash10Wang X 1991 Period-doublings to chaos in a simple neural network an analytic proof Complex Syst 5 425ndash41Wennekers T and Pasemann F 1996 Synchronous chaos in high dimensional modular neural networks Int J Bifurc
Chaos 6 2055ndash67Zaks M A Parks E H Rosenblum M G and Kurths J 1999 Alternative locking ratios in imperfect phase synchronization
Phys Rev Lett 82 4228ndash31Zhao L and Macau E N 2001 A network of dynamically coupled chaotic maps for scene segmentation IEEE Trans
Neural Netw 12 1375ndash85

718 M Breakspear et al
Hunt et al (1997) Josic (1998) Kocarev et al (2000) have shown that the generalizedsynchronization manifold is smooth (differentiable) if and only if it is normally hyperbolic(ie the largest transverse Lyapunov exponent is less than the smallest tangential Lyapunovexponent prior to the introduction of parameter mismatch) Such a condition is met in thepresent setting when C = 04 hence the smoothness of the manifold in panel (c)
However subsequent to the introduction of parameter mismatch we will not be able touse transverse and tangential Lyapunov exponents to detect such synchronization since thesynchronization manifold may possess a complex topology for which it is a non-trivial (atbest) or impossible (at worse) task to define transverse and tangential directions For examplethe manifold may be smooth as in figure 9(c) but have a fractal topology If the dynamics arenot normally hyperbolic then the manifold may be fractal but also non-differentiable (Huntet al 1997) and hence will not have a unique tangent direction at any point Consequently wewill not be able to compute tangential or transverse Lyapunov exponents and hence determinethe stability of the synchronization manifold However other techniques for detecting thepresence of generalized synchronization in complex systems have evolved including theauxiliary system approach (Abarbanel et al 1996) and non-linear interdependence (Schiffet al 1996) which are now explored
421 The auxiliary system approach The auxiliary system is a third subsystem which isa replica of the original response system but coupled independently to the driver system andcommencing from independent initial conditions It is possible to demonstrate that showingthe existence of generalized synchronizationmdashthat equation (10) is satisfiedmdashis equivalent toshowing that the response system and the auxiliary system evolve identically (even though theyare not directly coupled) To determine whether or not this is the case reduces to an analysisof the Lyapunov exponents transverse to the symmetry manifold with respect to the responsesystem and the auxiliary system
Whilst this is a somewhat artificial test for generalized synchronization since theproduction of an auxiliary system for a physical system would be impossible such a testis useful in numerical and theoretical studies In figure 10 we present an example of theauxiliary system approach where the parameter δV = 07 for column 1 and 068 for column 2We consider two values of the coupling C = 0025 and 01 The auxiliary system is simplya copy of the second column beginning from an arbitrary initial state In the first instancewhere C = 0025 we notice that even after allowing for an initial transient state the secondcolumn and the auxiliary do not behave in identical fashions (figure 10(a)) However whenC = 01 we observe (figure 10(b)) that the second column and the auxiliary do behave innear-identical manners after an initial transient and hence conclude that for the particularvalue of the coupling strength considered equation (10) is satisfied and hence generalizedsynchronization between the first and second columns occurs
422 Nonlinear interdependence The question remains as to how we may detect thepresence of generalized synchronization in a practical setting As we mentioned aboveit is not possible to use an auxiliary-system-based approach to detect the presence ofgeneralized synchronization Instead we must consider techniques for detecting suchbehaviour from experimental or numerical time series data Algorithms of this type areoften referred to as detecting non-linear interdependence Nonlinear interdependence is ageneral term encompassing systems where the behaviour of one non-linear system affectsthe behaviour of another More specifically it can be defined as occurring if the nullhypothesis that the interdependence between two signals is purely linear can be rejectedat conventional levels (usually 95) of statistical significance hence it is an experimental
Synchronization in a model of neuronal dynamics 719
2
1
2
1
Figure 10 Concurrent evolution of the response system V 2 and lsquoauxiliaryrsquo system V 1 (a) C =0025 (b) C = 01
term (eg Schiff et al 1996) Whilst this does not strictly correspond to the existence of afunctional relationship between the two systems such a relationship is typically used tomotivate the techniques for its detection Schiff et al (1996) extended the logic of non-linearprediction algorithms from autonomous to coupled systems by exploiting the continuity ofsuch a relationship and hence devised the first method of detecting non-linear interdependencein time series data The basis of the technique is using the forward evolution of a phase spacevectorrsquos neighbours to predict the forward evolution of that vector and the forward evolutionof the concurrent vector of the other (possibly interdependent) system If the error of such aprocedure is better on average than chance (the chance error is normalized to equal one) thennon-linear interdependence is said to be present
720 M Breakspear et al
V1
V2
Figure 11 Experimental and numerical examples of weak generalized non-linear synchronizationin the presence of measurement noise (a) Human scalp EEG data from posterior bipolar scalpelectrodes (O1P3ndashO2P4) (b) Weakly coupled aperiodic data in the present model with additivewhite noise (with 5 of the amplitude of the model signal) Parameters δ1
V = 0605 δ2V = 061
aee = 036 C = 007
A variation of this algorithm optimized to detect weak non-linear interdependence (Terryand Breakspear 2003) was used recently to demonstrate that weak non-linear interdependence isoccasionally present in the scalp EEG of healthy resting human subjects (Breakspear and Terry2002a 2002b 2002c) An example of an epoch of lsquonon-linearly independentrsquo EEG obtainedusing this technique is presented in figure 11(a) For comparison output from the currentmodel is presented in figure 11(b) The model parameters were set such that the dynamicswas still aperiodic but the intermittent large-amplitude bursts were suppressed and had weakmutual coupling (C = 007) Extraneous noise to simulate the effect of extracranial muscleactivity contaminating the EEG data has been added to this time series It should be notedthat the amplitudes and time scales of the modelrsquos variables are dimensionless Nonethelessthe model facilitates qualititative interpretation of experimental findings
Synchronization in a model of neuronal dynamics 721
43 Chaotic phase synchronization
Whereas the concept of generalized synchronization was mathematically motivated and assuch more suited to theoretical investigations of systems the concept of phase synchronizationderives from a more practical signal processing approach Phase synchronization occurs inboth symmetric and asymmetric settings but is often more useful in the asymmetric case (andin experimental data)
Unfortunately the phase of a chaotic signal is typically ambiguous and can be defined in anumber of different ways For example in the case of the coherent Rossler attractor (Osipovet al 1997) the phase may be defined as the arctangent of two of the variables of the systemMore generally it can sometimes be defined as the angle that an orbit subtends with the centreof an attractor It is also possible to extract the phase of a chaotic signal from time seriesdata by use of the Hilbert transform This has enabled the demonstration of chaotic phasesynchronization in a number of coupled asymmetrical systems (Rosenblum et al 1996 1997Zaks et al 1999) Although such an approach does not guarantee that the phase will be uniqueit does permit the practical analysis of phase in noisy time series (Tass et al 1998)
To illustrate phase synchronization in two coupled columns we again vary the parameterδV which if the columns are viewed as oscillators acts to detune them from a commonfrequency An important question is that of the criteria on the basis of which we concludewhether phase synchronization occurs We use a simple locking criterion whereby phasesynchronization is said to occur if
|θ1(t)minus θ2(t)| lt K (11)
where is θi the phase of column i and K is a constant Notice that this does not mean thatsynchronization implies equality of phases merely that their difference is bounded
An example of phase synchronization with δV = 065 for column 1 and 066 for column 2is given in figure 12 In this figure rNMDA = 03 Qualitatively some sort of phase coherenceis evident in the time series panel (a) This was confirmed by plotting the sine of thephase difference which has been extracted using the Hilbert transform panel (b) Clearly|sin(θ1 minus θ2)| lt K and hence |θ1 minus θ2| is bounded It can also be seen that the phase-synchronized dynamics occurs on a highly structured manifold depicted in figure 12(c)However unlike the case for the generalized synchronization manifold depicted in figure 9 itis evident that this manifold is not a simple transformation of the symmetry manifold
It is also possible to see lsquobimodalrsquo stability whereby a system jumps betweenapproximately zero phase difference and a phase lag phase synchronization of π An exampleis depicted in figure 12(d) At approximately 2000 s the system can be seen to jumpspontaneously from the first to the second of these modes The Z -variable is plotted forgraphical purposes
5 Intermittency and desynchronization
51 Desynchronization in an array of two systems
Diminishing the strength of coupling across the blowout bifurcation does not lead to a simpletransition from synchronous to asynchronous oscillations but is instead associated withcomplex temporal behaviours Specifically when the largest transverse Lyapunov exponent isclose to zero the coupled systems exhibit epochs of synchronous behaviours punctuated bysharp bursts of desynchronization This is observed in systems exhibiting identicalgeneralizedand phase synchronization An example of such a phenomenon is given in figure 13 The timeseries panel (a) shows synchronous dynamics except at time = 1000 where it can be seenthat the two time series desynchronize briefly This is evident in panel (b) as a disruption in the
722 M Breakspear et al
2
1
Figure 12 Phase-synchronized coupled columns (a) The time series of V 1 (solid) and V 2 (b) Thesine of the phase difference obtained through Hilbert transformation of the time series in panel (a)(c) The phase synchronization manifold in the V 1ndashV 2 plane (d) The time series of Z (solid) andZ showing a phase slip of π between two modes
Synchronization in a model of neuronal dynamics 723
Figure 13 An example of phase desynchronization in coupled asymmetric systems with C = 01Time series plots of (a) each V -variable and (b) their Euclidean distance (c) The temporal evolutionof the phase difference between the two systems (d) An example of the phase difference evolutionover a longer time interval
724 M Breakspear et al
orderly evolution of the distance between the two systems and in panel (c) where the evolutionof the phase difference reveals a brief violation of the criterion |φ1 minus φ2| lt constant Anexample of the phase difference evolution over a longer time interval is shown in figure 13(d)Note that this is a three-dimensional plot with sin(φ1 minus φ2) and cos(φ1 minus φ2) as they- and z-axes Hence phase desynchronizations are evident as complete revolutions aroundthe cylindrical phase space in which this variable evolves It can be seen that these phenomenaoccur at irregular intervals in the temporal domain
This behaviour can be explained by considering the nature of a chaotic attractor In thestatistical sense an arbitrary initial condition is expected to lead to the chaotic behaviour onthe attractor Consequently when we calculate the Lyapunov exponents of the system theseexponents correspond to those of the chaotic attractor Typically however there are an infinitenumber of unstable periodic orbits embedded within the attractor If the path of a trajectorybecomes close to one of these orbits it may lsquoshadowrsquo the orbit for a brief period of time Eachof these orbits has a corresponding set of Lyapunov exponents which (like the attractor) can bedivided into tangential and transverse exponents These also vary when we change the couplingparameter but are typically not the same as the exponents of the attractor Hence the largesttransverse exponent of the attractor may be negative yet there may be positive transverseexponents associated with periodic orbits Considering all of these exponents together wewould therefore only expect to observe complete stability of the synchronization manifold inthe case where all of the transverse Lyapunov exponents are negative Bursting behaviourwill be observed when the one of these exponents becomes positive typically correspondingto a periodic orbit of low order (Pikovsky and Grassberger 1991 Rulkov and Sushchik 1997Maistrenko et al 1998 Heagy et al 1998) Due to the complex structure of such periodic orbitsthe desynchronous bursts occur across a wide range of timescales
In figure 14 we present the results of a long integration (16times105 time units) in which thelengths of sequential synchronous epochs of phase synchronization are studied The parametervalues were δV = 065 for column 1 and 0645 for column 2 with coupling strength10 equal to0125 In panel (a) the power spectrum of these sequential lengths is plotted The whiteness ofthis spectrum indicates that the lengths of consecutive synchronous epochs are independentThis is due to the high-dimensional lsquomixingrsquo effect of desynchronization In panel (b) thelengths of the synchronous epochs have been sorted again into the order of increasing lengthThe results for two coupling strengths are plotted C = 0125 (solid) and 0135 (dashed)Several features of this plot are of note Firstly the lengths of the synchronous epochsare distributed over almost three orders of magnitude Secondly for both plots there is anapproximately log-linear relationship between the length of an epoch and its relative orderThis reflects an approximately exponential relationship between the length of a synchronousepoch and its likelihood of occurrence Thirdly this relationship is skewed toward longersynchronous epochs for the stronger coupling strengths
This type of signal with patches of lsquoorderrsquo occurring over many timescales interruptedby bursts of disorder can be shown to have a high information capacity (Gatlin 1972)Moreover complex phase desynchronization may make an infrequent yet robust contributionto the dynamical pattern of the alpha rhythm in scalp EEG data for resting human subjects(Breakspear 2002) It is tempting to postulate that such a lsquoweakly stablersquo dynamics may assistin the generation of the rich and fluid mental associations that spontaneously emerge in restingcognitive states Although this phenomenon has been previously described in a variety of othersystems to our knowledge this is the first description in a biophysical model of neocortex
10 The introduction of parameter mismatch causes an increase in the strength of coupling required for stablesynchronization above the value of 01 required in the symmetric case
Synchronization in a model of neuronal dynamics 725
Figure 14 Properties of sequential epochs of phase-synchronous behaviour over a long integrationtime δ1
V = 065 δ2V = 0645 aee = 04 (a) The flat (white) power spectrum (b) Synchronous
epochs sorted in order of increasing length (solid C = 0125 dashed C = 0134)
52 Synchronization and complex dynamics in larger arrays
The synchronous and complex phenomena described above are also observed in arrays ofcoupled systems with N gt 2 Expanding the size of the array permits complex dynamicsin the spatial in addition to the temporal domain However the basic principles governingthe type and stability of synchronized motions are the same In this section a few of thepossible behaviours are discussed Physiological variation across the array is modelled bymodifying the parameter δV = 07 in each array by adding a random number of maximummagnitude 001
In figure 15(a) an array of N = 20 uncoupled (C = 0) systems is shown The lack ofsynchronization in this time series is evident By comparison the rapid transition to completesynchronization is evident in figure 15(b) when C = 03 In fact 19 of the 20 subsystems
726 M Breakspear et al
(b)
(c)
(a)
Figure 15 Synchronization phenomena in an array of coupled cortical columns (a) N = 20C = 0 (b) N = 20 C = 03 (c) N = 6 C = 01 (d) N = 12 C = 009 (e) N = 12C = 012 A small perturbation has been applied to one of the columns at t = 2500 s (f) Evolutionof the phase difference between two of the columns in panel (e) For clarity the W -variable isplotted in panel (c) and the Z -variable in (e)
have phase synchronized within one single cycle Intermediate values of coupling strength areassociated with a rich variety of phenomena For example subsystems with the most similarparameter values typically synchronize into clusters of various sizes An example of this isillustrated in figure 15(c) Note that there is no synchronization between the two differentclusters each containing three synchronized subsystems
Another complex behaviour occurs when subsystems transiently synchronize into largeclusters then spontaneously desynchronize again Hence the dynamics of the entire arrayjumps between different transient dynamics characterized by varying degrees of freedom Anexample of this phenomenon is given in figure 15(d) In addition it is possible to inducedesynchronization across an entire array by applying a small perturbation to one columnfor one time step An example is illustrated in figure 15(e) A small perturbation has been
Synchronization in a model of neuronal dynamics 727
Figure 15 (Continued)
added to one column after 2500 time units It can be seen that the entire array subsequentlydesynchronizes This illustrates the sensitivity of the dynamics to external stimuli Finally it ispossible to lsquoinducersquo synchronization between selected subsystems by increasing the strength ofcoupling between them selectively This allows an extra level of complexity to be introducedinto the model For example it may allow targeted information transfer between a partialnetwork of the entire array
As discussed in section 51 a univariate signal which jumps between epochs of lowentropy (high repetition) and bursts of high entropy (high variability) can be shown to havehigh information capacity This is because the repetition of certain patterns (as in the English
728 M Breakspear et al
language) permits error detectability Bursts of high entropy act in the opposite directionto increase message variability With a multivariate signal in addition to optimizing theinformation capacity within each signal the overall information capacity can be increasedthrough the type of itinerant synchronization briefly illustrated in this section Synchronizationacross an entire array increases the message reliability (since a single lsquoerrant letterrsquo in onesignal can be easily detected) Conversely desynchronization into multiple clusters increasesmessage variability since each cluster will produce independent information Hence a neuralarray which switches dynamically between a range of cluster configurations can be seen asadaptive in the sense that high synchronous states can be expressed when error detectabilityis a priority (such as during quick and reliable execution of a familiar task) but the array canrapidly desynchronize if variability becomes salient (for example if the environment suddenlybecomes novel) A formal operationalized demonstration of this principle is to be the subjectof future work
6 Discussion
In this paper we present a dynamical model for activity in an array of weakly coupledlocal neural assemblies The model incorporates the fundamental principles governing cellmembrane potential as determined by voltage- and ligand-gated ion channels In this wayit allows a reasonable amount of physiological complexity to be incorporated includingsynaptic activation and receptor subtypeswithout recourse to overwhelming detail Numericalanalysis of this model reveals a transition from independent oscillations to stable chaoticsynchronization depending on the balance of local versus long-range connectivity andbiophysical heterogeneity These excitatory couplings are subject to ascending subcorticalneuromodulation arising principally from monoaminergic nuclei such as Locus Coeruleus(Silberstein 1995) Neuromodulation of extrinsic connections was incorporated into the modelvia a coupling parameter C The stability of the synchronization is determined by the valueof the largest transverse Lyapunov exponent which in turn depends on the coupling parameterC If the coupling is weakened with the result that this exponent becomes positive a blowoutbifurcation occurs and independent oscillations re-emerge In the vicinity of the transitionnon-linear desynchronizations with complex temporal and spatial properties are observed
It is pleasing that complex and itinerant behaviour reminiscent of real neuronaldynamics emerges through such a simple manipulation of a single dynamic control parameterFurthermore the ubiquitous projections of classical neuromodulatory systems in real brainssuggest a clear biological mechanism by which the brain could self-organize throughmoderating the complexity of its own dynamics There are many reasons that the complicateddynamics of the sort demonstrated in this paper may be neurobiologically important Wemention four
First the presence of multiple clusters of phase-synchronized subsystems as discussedabove may be a uniquely non-linear mechanism facilitating the lsquocarriagersquo and distinction ofmultiple concurrent dynamic cell assemblies (and hence cognitive processing) This may beimportant for perceptual grouping binding and transitions as suggested by others (eg von derMalsburg 1985 Gray and Singer 1989)
Second the facility to decouple and reconfigure dynamic cell assemblies througha neuromodulatory control parameter may be important for avoiding low-entropysynchronization manifolds In the absence of itinerancy synchronization would renderneuronal dynamics incapable of representing changes in sensory input or indeed cognitive stateFor example in the model it was observed that small perturbations to one column were ableto induce a global change in neuronal dynamics by disrupting a state of weak synchronization
Synchronization in a model of neuronal dynamics 729
Inevitably alternative dynamic configurations emerge epitomizing the importance of lsquolabilersquobrain dynamics (Friston 2000a)
Thirdly from a purely functionalist point of view the balance between functionalspecialization and functional integration may rely upon the sorts of mechanism modelledin this paper The brain appears to adhere to two principles of functional organization namelysegregation of function within cortical areas and sub-areas and the integration of these areasin perceptual synthesis cognition and sensorimotor integration (Tononi et al 1994 Fristonet al 1995) The balance between preserving specialized dynamics unique to each arearequired by specialization and the coupling among these areas mediating functional integrationmay be achieved by the neuromodulatory mechanisms described above Significantlythe spontaneous variation in the numbers of degrees of freedom (for example whenlow-dimensional synchronization manifolds are shadowed) imparts a dynamic characteristicto this perspective that is precluded by linear mechanisms alone
Finally from a mechanistic perspective complexity and diversity of brain dynamics arerequired for selection of adaptive responses (Friston 2000b) There are many examples ofselective mechanisms operating within the brain All can be seen as performing an ascenton some objective function or fitness landscape These mechanisms require complex anditinerant dynamics to facilitate a thorough exploration of the landscape These landscapesmay range from those involved in perception (where the landscape is defined by predictionerror see eg Rao and Ballard (1998)) to those involved in value learning (where thelandscape is determined by the outcome of a behaviour eg in temporal difference models (seeeg Sutton and Barto 1990)) In short complexity is necessary to maintain an information-rich environment in which the brain can respond to sensory inputs An essential componentof this is the maintenance of diversity which is a prerequisite for selectionist mechanismsat many scales (Edelman 1993) Understanding the mechanistic basis of complex dynamicsas measured electrophysiologically may well depend on the sort of biologically plausiblemodels used in this paper This understanding may be important for both basic and clinicalneuroscience (eg Tass 2002)
The present study supplements and advances on other research in this field The studyof complex phase synchronization and clustering in neural oscillator networks similar tothe present model has previously been the subject of formal mathematical study Forexample Terman and Wang (1995) studied synchronization of blocks of neural oscillatorsthat were subject to perturbation by connected lsquostimulirsquo Somers and Kopell (1995) studiedthe emergence of both synchronization and travelling waves in networks of coupled neuraloscillators In the work of Terman and Wang the local dynamics were based on neurallsquocaricaturersquo models such as the FitzHughndashNagumo equations and are hence more abstract thanthe current study The approach of Somers and Kopell only required that the local dynamicsexhibit fast and relaxation phases and is hence more abstract still In both studies onlyperiodic dynamics are studied Complex patterns of chaos synchronization in neural arrayshave been characterized by nonlinear methods (Kaneko 1998) and their putative contributionto both learning (Nakajima and Ueda 1996) and perception (Zhao and Macau 2001 Breakspear2001) explored However these studies employed models such as the logistic equation andtent map which are quite abstract in comparison to the present study On the other handHansel and Sompolinsky (1996) studied the role of chaos synchronization in a model ofvisual perception in a cortical hypercolumn using a very similar approach to that adoptedhere but with the additional complexity that synaptic delays were incorporated However inthis and earlier work (Hansel and Sompolinsky 1992) only linear measures of dynamics andsynchrony (specifically the auto- and cross-correlation functions) were employed The analysisof chaotic synchrony in the study of Larter et al (1999) was based only upon observation of
730 M Breakspear et al
the Euclidean difference between variables from two nodes within a larger array Hence thesignificance of the present work is that it provides a quantitative nonlinear analysis of complexchaos synchronization in a biophysical model of cortical dynamics Such research is requiredin order to bridge the gap between quantitative nonlinear research into synchronization andcomputation in abstract models of neural arrays and qualitative or linear analysis in morebiophysically plausible models
Acknowledgments
The authors would like to thank Peter Robinson for assistance with the manuscript and R Larterfor helpful advice MB is a recipient of a NSW Institute of Psychiatry Research Grant anda University of Sydney SESQUI postdoctoral fellowship KJF is supported by the WellcomeTrust
References
Abarbanel H Rulkov N and Sushchik M 1996 Generalised synchronization of chaos the auxiliary system approachPhys Rev E 53 4528ndash35
Afraimovich V Verichev N and Rabinovich M 1986 Stochastic synchronization of oscillation in dissipative systemsRadiophys Quantum Electron 29 795ndash801
Breakspear M 2001 Perception of odours by a nonlinear model of the olfactory bulb Int J Neural Syst 11 101ndash24Breakspear M 2002 Nonlinear phase desynchronisation in human electroencephalographic data Human Brain Mapping
15 175ndash98Breakspear M and Terry J R 2002a Nonlinear interdependence in neural systems motivation theory and relevance
Int J Neurosci 112 1163ndash84Breakspear M and Terry J R 2002b Detection and description of nonlinear interdependence in normal multichannel
human EEG Clin Neurophysiol 113 735ndash53Breakspear M and Terry J R 2002c Topographic organization of nonlinear interdependence in multichannel human
EEG Neuroimage 16 822ndash35Destexhe A Rudolph M Fellous J M and Sejnowski T J 2001 Fluctuating synaptic conductances recreate in vivo-like
activity in neocortical neurons Neuroscience 107 13ndash24Douglas R J Mahowald M A and Martin K A 1996 Microarchitecture of neocortical columns Brain TheorymdashBiological
Basis and Computational Principles ed A Aertsen and V Braitenberg (Amsterdam Elsevier)Eckhorn R Bauer R Jordan W Brosch M Kruse W Munk M and Reitboeck H J 1988 Coherent oscillations a
mechanism of feature linking in the visual cortex Multiple electrode and correlation analyses in the cat BiolCybern 60 121ndash30
Edelman G M 1993 Neural Darwinism selection and reentrant signalling in higher brain function Neuron 10 115ndash25Engel A K Kreiter A K Konig P and Singer W 1991 Synchronization of oscillatory neuronal responses between
striate and extrastriate visual cortical areas of the cat Proc Natl Acad Sci USA 88 6048ndash52Friston K J 2000a The labile brain I Neuronal transients and nonlinear coupling Phil Trans R Soc B 355 215ndash36Friston K J 2000b The labile brain II Transients complexity and selection Phil Trans R Soc B 355 237ndash52Friston K J Tononi G Sporns O and Edelman G 1995 Characterising the complexity of neuronal interactions Human
Brain Mapping 3 302ndash14Fujisaka H and Yamada T 1983 Stability theory of synchronized motion in coupled-oscillator system Prog Theor
Phys 69 32ndash47Gatlin L 1972 Information Theory and the Living System (New York Columbia University Press)Gray C M Konig P Engel A and Singer W 1989 Oscillatory responses in cat visual cortex exhibit inter-columnar
synchronization which reflects global stimulus properties Science 338 334ndash7Gray C M and Singer W 1989 Stimulus specific neuronal oscillations in orientation columns of cat visual cortex Proc
Natl Acad Sci USA 86 1698ndash702Haig A R Gordon E Wright J J Meares R A and Bahramali H 2000 Synchronous cortical gamma-band activity in
task-relevant cognition Neuroreport 11 669ndash75Hansel D and Sompolinsky H 1992 Synchronization and computation in a chaotic neural network Phys Rev Lett 68
718ndash21
Synchronization in a model of neuronal dynamics 731
Hansel D and Sompolinsky H 1996 Chaos and synchrony in a model of a hypercolumn in visual cortex J ComputNeurosci 3 7ndash4
Heagy J Carroll T and Pecora L 1998 Desynchronization by periodic orbits Phys Rev E 57 R1253ndash6Hunt B Ott E and Yorke J 1997 Differentiable synchronization of chaos Phys Rev E 55 4029ndash34Josic K 1998 Invariant manifolds and synchronization of coupled dynamical systems Phys Rev Lett 80 3053ndash6Kandel E Schwartz J and Jessell T 2000 Principles of Neural Science (New York McGraw-Hill)Kaneko K 1998 On the strength of attractors in a high-dimensional system Physica D 124 322ndash44Kocarev L Parlitz U and Brown R 2000 Robust synchronization of chaotic systems Phys Rev E 61 3716ndash20Larter R Speelman B and Worth R M 1999 A coupled ordinary differential equation lattice model for the simulation
of epileptic seizures Chaos 9 795ndash804Lumer E Edelman G and Tononi G 1997a Neural dynamics in a model of the thalamocortical system I Layers loops
and the emergence of fast synchronous oscillations Cereb Cortex 7 207ndash27Lumer E Edelman G and Tononi G 1997b Neural dynamics in a model of the thalamocortical system II The role of
neural synchrony tested through perturbations of spike timing Cereb Cortex 7 228ndash36Magleby K 1987 Short-term modulation at synapses Synaptic Function ed G M Edelman W E Gall and
W M Cowan (New York Wiley)Maistrenko Y Maistrenko V Popovich A and Mosekilde E 1998 Transverse instability and riddled basins in a system
of two coupled logistic maps Phys Rev E 57 2713ndash24Morris C and Lecar H 1981 Voltage oscillations in the barnacle giant muscle fiber Biophys J 35 193ndash213Mountcastle V B 1997 The columnar organization of the neocortex Brain 120 701ndash22Mountcastle V B 1998 The Cerebral Cortex (Cambridge MA Harvard University Press)Nakajima H and Ueda Y 1996 Riddled basins of the optimal states in learning dynamical systems Physica D 99 35ndash44Nunez P L 1995 Neocortical Dynamics and Human Brain Rhythms (Oxford Oxford University Press)Osipov G V Pikovsky A S Rosenblum M G and Kurths J 1997 Phase synchronization effects in a lattice of nonidentical
Rossler oscillators Phys Rev E 55 2353ndash61Ott E and Sommerer J 1994 Blowout bifurcations the occurrence of riddled basins and onndashoff intermittency Phys
Lett A 188 39ndash47Pecora L and Carroll T 1990 Synchronization in chaotic systems Phys Rev Lett 64 821ndash4Pikovsky A and Grassberger P 1991 Symmetry breaking bifurcation for coupled chaotic attractors J Phys A Math
Gen 24 4587ndash97Rao R P and Ballard D H 1998 Predictive coding in the visual cortex a functional interpretation of some extra-classical
receptive field effects Nature Neurosci 2 79ndash87Robinson P A Wright J J and Rennie C R 1998 Synchronous oscillations in the cerebral cortex Phys Rev E 57
4578ndash88Rodriguez E George N Lachauz J P Martinerie J Renault B and Varela F J 1999 Perceptionrsquos shadow long distance
synchronization of human brain activity Nature 397 430ndash3Rosenblum M Pikovsky A and Kurths J 1996 Phase synchronization of chaotic oscillators Phys Rev Lett 76 1804ndash7Rosenblum M Pikovsky A and Kurths J 1997 From phase to lag synchronization in coupled chaotic oscillators Phys
Rev Lett 78 4193ndash6Rulkov N and Sushchik M 1997 Robustness of synchronized chaotic oscillations Int J Bifurc Chaos 7 625ndash43Rulkov N Sushchik M Tsimring L and Abarbanel H 1995 Generalized synchronization of chaos in unidirectionally
coupled chaotic systems Phys Rev E 51 980ndash94Schiff S So P Chang T Burke R and Sauer T 1996 Detecting dynamical interdependence and generalized synchrony
through mutual prediction in a neural ensemble Phys Rev E 54 6708ndash24Silberstein R B 1995 Neuromodulations of neocortical dynamics Neocortical Dynamics and Human EEG Rhythms
ed P L Nunez (Oxford Oxford University Press)Singer W 1995 Putative functions of temporal correlations in neocortical processing Large-Scale Neuronal Theories
of the Brain ed C Koch and J Davis (London MIT Press)Somers D and Kopell N 1995 Waves and synchrony in networks of oscillators of relaxation and non-relaxation type
Physica D 89 169ndash83Sutton R S and Barto A G 1990 Time derivative models of Pavlovian reinforcement Learning and Computational
Neuroscience Foundations of Adaptive Networks ed M Gabriel and J Moore (Cambridge MA MIT Press) pp497ndash538
Szentagothai J 1983 The modular architectonic principle of neural centers Rev Physiol Biochem Pharmacol 9811ndash61
Tallon-Baudry C Betrand O and Fischer C 2001 Oscillatory synchrony between human extrastriate areas during visualshort-term memory maintenance J Neurosci 21 RC177(1ndash5)
Tass P A 2002 Desynchronization of brain rhythms with soft phase-resetting techniques Biol Cybern 87 102ndash15
732 M Breakspear et al
Tass P Rosenblum M Weule J Kurths J Pikovsky A Volkmann J Schnitzler A and Freund H 1998 Detection of nmphase locking from noisy data application to magnetoencephalography Phys Rev Lett 81 3291ndash4
Terman D and Wang D 1995 Global competition and local cooperation in a network of neural oscillators Physica D81 148ndash76
Terry J and Breakspear M 2003 An improved algorithm for the detection of dynamical interdependence in bivariatetime-series Biol Cybern 88 129ndash36
Tononi G Sporns O and Edelman G M 1994 A measure for brain complexity relating functional segregation andintegration in the nervous system Proc Natl Acad Sci USA 91 5033ndash7
von der Malsburg C 1985 Nervous structures with dynamical links Phys Chem 89 703ndash10Wang X 1991 Period-doublings to chaos in a simple neural network an analytic proof Complex Syst 5 425ndash41Wennekers T and Pasemann F 1996 Synchronous chaos in high dimensional modular neural networks Int J Bifurc
Chaos 6 2055ndash67Zaks M A Parks E H Rosenblum M G and Kurths J 1999 Alternative locking ratios in imperfect phase synchronization
Phys Rev Lett 82 4228ndash31Zhao L and Macau E N 2001 A network of dynamically coupled chaotic maps for scene segmentation IEEE Trans
Neural Netw 12 1375ndash85
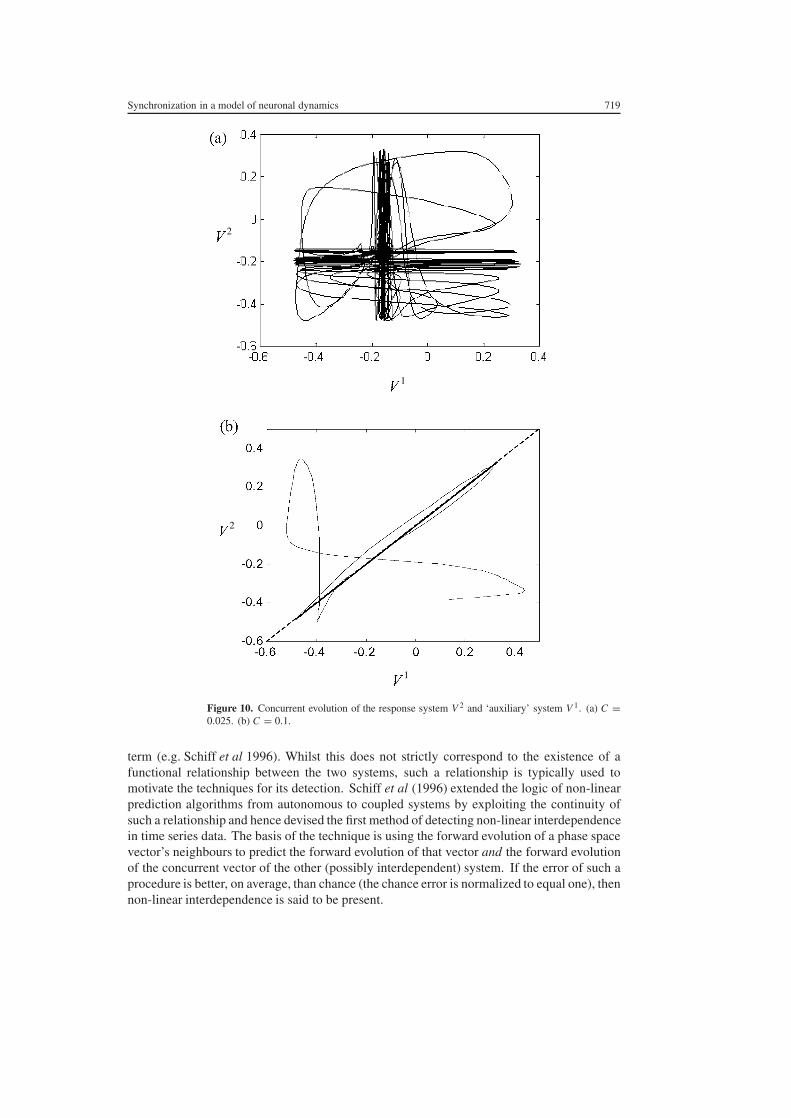
Synchronization in a model of neuronal dynamics 719
2
1
2
1
Figure 10 Concurrent evolution of the response system V 2 and lsquoauxiliaryrsquo system V 1 (a) C =0025 (b) C = 01
term (eg Schiff et al 1996) Whilst this does not strictly correspond to the existence of afunctional relationship between the two systems such a relationship is typically used tomotivate the techniques for its detection Schiff et al (1996) extended the logic of non-linearprediction algorithms from autonomous to coupled systems by exploiting the continuity ofsuch a relationship and hence devised the first method of detecting non-linear interdependencein time series data The basis of the technique is using the forward evolution of a phase spacevectorrsquos neighbours to predict the forward evolution of that vector and the forward evolutionof the concurrent vector of the other (possibly interdependent) system If the error of such aprocedure is better on average than chance (the chance error is normalized to equal one) thennon-linear interdependence is said to be present
720 M Breakspear et al
V1
V2
Figure 11 Experimental and numerical examples of weak generalized non-linear synchronizationin the presence of measurement noise (a) Human scalp EEG data from posterior bipolar scalpelectrodes (O1P3ndashO2P4) (b) Weakly coupled aperiodic data in the present model with additivewhite noise (with 5 of the amplitude of the model signal) Parameters δ1
V = 0605 δ2V = 061
aee = 036 C = 007
A variation of this algorithm optimized to detect weak non-linear interdependence (Terryand Breakspear 2003) was used recently to demonstrate that weak non-linear interdependence isoccasionally present in the scalp EEG of healthy resting human subjects (Breakspear and Terry2002a 2002b 2002c) An example of an epoch of lsquonon-linearly independentrsquo EEG obtainedusing this technique is presented in figure 11(a) For comparison output from the currentmodel is presented in figure 11(b) The model parameters were set such that the dynamicswas still aperiodic but the intermittent large-amplitude bursts were suppressed and had weakmutual coupling (C = 007) Extraneous noise to simulate the effect of extracranial muscleactivity contaminating the EEG data has been added to this time series It should be notedthat the amplitudes and time scales of the modelrsquos variables are dimensionless Nonethelessthe model facilitates qualititative interpretation of experimental findings
Synchronization in a model of neuronal dynamics 721
43 Chaotic phase synchronization
Whereas the concept of generalized synchronization was mathematically motivated and assuch more suited to theoretical investigations of systems the concept of phase synchronizationderives from a more practical signal processing approach Phase synchronization occurs inboth symmetric and asymmetric settings but is often more useful in the asymmetric case (andin experimental data)
Unfortunately the phase of a chaotic signal is typically ambiguous and can be defined in anumber of different ways For example in the case of the coherent Rossler attractor (Osipovet al 1997) the phase may be defined as the arctangent of two of the variables of the systemMore generally it can sometimes be defined as the angle that an orbit subtends with the centreof an attractor It is also possible to extract the phase of a chaotic signal from time seriesdata by use of the Hilbert transform This has enabled the demonstration of chaotic phasesynchronization in a number of coupled asymmetrical systems (Rosenblum et al 1996 1997Zaks et al 1999) Although such an approach does not guarantee that the phase will be uniqueit does permit the practical analysis of phase in noisy time series (Tass et al 1998)
To illustrate phase synchronization in two coupled columns we again vary the parameterδV which if the columns are viewed as oscillators acts to detune them from a commonfrequency An important question is that of the criteria on the basis of which we concludewhether phase synchronization occurs We use a simple locking criterion whereby phasesynchronization is said to occur if
|θ1(t)minus θ2(t)| lt K (11)
where is θi the phase of column i and K is a constant Notice that this does not mean thatsynchronization implies equality of phases merely that their difference is bounded
An example of phase synchronization with δV = 065 for column 1 and 066 for column 2is given in figure 12 In this figure rNMDA = 03 Qualitatively some sort of phase coherenceis evident in the time series panel (a) This was confirmed by plotting the sine of thephase difference which has been extracted using the Hilbert transform panel (b) Clearly|sin(θ1 minus θ2)| lt K and hence |θ1 minus θ2| is bounded It can also be seen that the phase-synchronized dynamics occurs on a highly structured manifold depicted in figure 12(c)However unlike the case for the generalized synchronization manifold depicted in figure 9 itis evident that this manifold is not a simple transformation of the symmetry manifold
It is also possible to see lsquobimodalrsquo stability whereby a system jumps betweenapproximately zero phase difference and a phase lag phase synchronization of π An exampleis depicted in figure 12(d) At approximately 2000 s the system can be seen to jumpspontaneously from the first to the second of these modes The Z -variable is plotted forgraphical purposes
5 Intermittency and desynchronization
51 Desynchronization in an array of two systems
Diminishing the strength of coupling across the blowout bifurcation does not lead to a simpletransition from synchronous to asynchronous oscillations but is instead associated withcomplex temporal behaviours Specifically when the largest transverse Lyapunov exponent isclose to zero the coupled systems exhibit epochs of synchronous behaviours punctuated bysharp bursts of desynchronization This is observed in systems exhibiting identicalgeneralizedand phase synchronization An example of such a phenomenon is given in figure 13 The timeseries panel (a) shows synchronous dynamics except at time = 1000 where it can be seenthat the two time series desynchronize briefly This is evident in panel (b) as a disruption in the
722 M Breakspear et al
2
1
Figure 12 Phase-synchronized coupled columns (a) The time series of V 1 (solid) and V 2 (b) Thesine of the phase difference obtained through Hilbert transformation of the time series in panel (a)(c) The phase synchronization manifold in the V 1ndashV 2 plane (d) The time series of Z (solid) andZ showing a phase slip of π between two modes
Synchronization in a model of neuronal dynamics 723
Figure 13 An example of phase desynchronization in coupled asymmetric systems with C = 01Time series plots of (a) each V -variable and (b) their Euclidean distance (c) The temporal evolutionof the phase difference between the two systems (d) An example of the phase difference evolutionover a longer time interval
724 M Breakspear et al
orderly evolution of the distance between the two systems and in panel (c) where the evolutionof the phase difference reveals a brief violation of the criterion |φ1 minus φ2| lt constant Anexample of the phase difference evolution over a longer time interval is shown in figure 13(d)Note that this is a three-dimensional plot with sin(φ1 minus φ2) and cos(φ1 minus φ2) as they- and z-axes Hence phase desynchronizations are evident as complete revolutions aroundthe cylindrical phase space in which this variable evolves It can be seen that these phenomenaoccur at irregular intervals in the temporal domain
This behaviour can be explained by considering the nature of a chaotic attractor In thestatistical sense an arbitrary initial condition is expected to lead to the chaotic behaviour onthe attractor Consequently when we calculate the Lyapunov exponents of the system theseexponents correspond to those of the chaotic attractor Typically however there are an infinitenumber of unstable periodic orbits embedded within the attractor If the path of a trajectorybecomes close to one of these orbits it may lsquoshadowrsquo the orbit for a brief period of time Eachof these orbits has a corresponding set of Lyapunov exponents which (like the attractor) can bedivided into tangential and transverse exponents These also vary when we change the couplingparameter but are typically not the same as the exponents of the attractor Hence the largesttransverse exponent of the attractor may be negative yet there may be positive transverseexponents associated with periodic orbits Considering all of these exponents together wewould therefore only expect to observe complete stability of the synchronization manifold inthe case where all of the transverse Lyapunov exponents are negative Bursting behaviourwill be observed when the one of these exponents becomes positive typically correspondingto a periodic orbit of low order (Pikovsky and Grassberger 1991 Rulkov and Sushchik 1997Maistrenko et al 1998 Heagy et al 1998) Due to the complex structure of such periodic orbitsthe desynchronous bursts occur across a wide range of timescales
In figure 14 we present the results of a long integration (16times105 time units) in which thelengths of sequential synchronous epochs of phase synchronization are studied The parametervalues were δV = 065 for column 1 and 0645 for column 2 with coupling strength10 equal to0125 In panel (a) the power spectrum of these sequential lengths is plotted The whiteness ofthis spectrum indicates that the lengths of consecutive synchronous epochs are independentThis is due to the high-dimensional lsquomixingrsquo effect of desynchronization In panel (b) thelengths of the synchronous epochs have been sorted again into the order of increasing lengthThe results for two coupling strengths are plotted C = 0125 (solid) and 0135 (dashed)Several features of this plot are of note Firstly the lengths of the synchronous epochsare distributed over almost three orders of magnitude Secondly for both plots there is anapproximately log-linear relationship between the length of an epoch and its relative orderThis reflects an approximately exponential relationship between the length of a synchronousepoch and its likelihood of occurrence Thirdly this relationship is skewed toward longersynchronous epochs for the stronger coupling strengths
This type of signal with patches of lsquoorderrsquo occurring over many timescales interruptedby bursts of disorder can be shown to have a high information capacity (Gatlin 1972)Moreover complex phase desynchronization may make an infrequent yet robust contributionto the dynamical pattern of the alpha rhythm in scalp EEG data for resting human subjects(Breakspear 2002) It is tempting to postulate that such a lsquoweakly stablersquo dynamics may assistin the generation of the rich and fluid mental associations that spontaneously emerge in restingcognitive states Although this phenomenon has been previously described in a variety of othersystems to our knowledge this is the first description in a biophysical model of neocortex
10 The introduction of parameter mismatch causes an increase in the strength of coupling required for stablesynchronization above the value of 01 required in the symmetric case
Synchronization in a model of neuronal dynamics 725
Figure 14 Properties of sequential epochs of phase-synchronous behaviour over a long integrationtime δ1
V = 065 δ2V = 0645 aee = 04 (a) The flat (white) power spectrum (b) Synchronous
epochs sorted in order of increasing length (solid C = 0125 dashed C = 0134)
52 Synchronization and complex dynamics in larger arrays
The synchronous and complex phenomena described above are also observed in arrays ofcoupled systems with N gt 2 Expanding the size of the array permits complex dynamicsin the spatial in addition to the temporal domain However the basic principles governingthe type and stability of synchronized motions are the same In this section a few of thepossible behaviours are discussed Physiological variation across the array is modelled bymodifying the parameter δV = 07 in each array by adding a random number of maximummagnitude 001
In figure 15(a) an array of N = 20 uncoupled (C = 0) systems is shown The lack ofsynchronization in this time series is evident By comparison the rapid transition to completesynchronization is evident in figure 15(b) when C = 03 In fact 19 of the 20 subsystems
726 M Breakspear et al
(b)
(c)
(a)
Figure 15 Synchronization phenomena in an array of coupled cortical columns (a) N = 20C = 0 (b) N = 20 C = 03 (c) N = 6 C = 01 (d) N = 12 C = 009 (e) N = 12C = 012 A small perturbation has been applied to one of the columns at t = 2500 s (f) Evolutionof the phase difference between two of the columns in panel (e) For clarity the W -variable isplotted in panel (c) and the Z -variable in (e)
have phase synchronized within one single cycle Intermediate values of coupling strength areassociated with a rich variety of phenomena For example subsystems with the most similarparameter values typically synchronize into clusters of various sizes An example of this isillustrated in figure 15(c) Note that there is no synchronization between the two differentclusters each containing three synchronized subsystems
Another complex behaviour occurs when subsystems transiently synchronize into largeclusters then spontaneously desynchronize again Hence the dynamics of the entire arrayjumps between different transient dynamics characterized by varying degrees of freedom Anexample of this phenomenon is given in figure 15(d) In addition it is possible to inducedesynchronization across an entire array by applying a small perturbation to one columnfor one time step An example is illustrated in figure 15(e) A small perturbation has been
Synchronization in a model of neuronal dynamics 727
Figure 15 (Continued)
added to one column after 2500 time units It can be seen that the entire array subsequentlydesynchronizes This illustrates the sensitivity of the dynamics to external stimuli Finally it ispossible to lsquoinducersquo synchronization between selected subsystems by increasing the strength ofcoupling between them selectively This allows an extra level of complexity to be introducedinto the model For example it may allow targeted information transfer between a partialnetwork of the entire array
As discussed in section 51 a univariate signal which jumps between epochs of lowentropy (high repetition) and bursts of high entropy (high variability) can be shown to havehigh information capacity This is because the repetition of certain patterns (as in the English
728 M Breakspear et al
language) permits error detectability Bursts of high entropy act in the opposite directionto increase message variability With a multivariate signal in addition to optimizing theinformation capacity within each signal the overall information capacity can be increasedthrough the type of itinerant synchronization briefly illustrated in this section Synchronizationacross an entire array increases the message reliability (since a single lsquoerrant letterrsquo in onesignal can be easily detected) Conversely desynchronization into multiple clusters increasesmessage variability since each cluster will produce independent information Hence a neuralarray which switches dynamically between a range of cluster configurations can be seen asadaptive in the sense that high synchronous states can be expressed when error detectabilityis a priority (such as during quick and reliable execution of a familiar task) but the array canrapidly desynchronize if variability becomes salient (for example if the environment suddenlybecomes novel) A formal operationalized demonstration of this principle is to be the subjectof future work
6 Discussion
In this paper we present a dynamical model for activity in an array of weakly coupledlocal neural assemblies The model incorporates the fundamental principles governing cellmembrane potential as determined by voltage- and ligand-gated ion channels In this wayit allows a reasonable amount of physiological complexity to be incorporated includingsynaptic activation and receptor subtypeswithout recourse to overwhelming detail Numericalanalysis of this model reveals a transition from independent oscillations to stable chaoticsynchronization depending on the balance of local versus long-range connectivity andbiophysical heterogeneity These excitatory couplings are subject to ascending subcorticalneuromodulation arising principally from monoaminergic nuclei such as Locus Coeruleus(Silberstein 1995) Neuromodulation of extrinsic connections was incorporated into the modelvia a coupling parameter C The stability of the synchronization is determined by the valueof the largest transverse Lyapunov exponent which in turn depends on the coupling parameterC If the coupling is weakened with the result that this exponent becomes positive a blowoutbifurcation occurs and independent oscillations re-emerge In the vicinity of the transitionnon-linear desynchronizations with complex temporal and spatial properties are observed
It is pleasing that complex and itinerant behaviour reminiscent of real neuronaldynamics emerges through such a simple manipulation of a single dynamic control parameterFurthermore the ubiquitous projections of classical neuromodulatory systems in real brainssuggest a clear biological mechanism by which the brain could self-organize throughmoderating the complexity of its own dynamics There are many reasons that the complicateddynamics of the sort demonstrated in this paper may be neurobiologically important Wemention four
First the presence of multiple clusters of phase-synchronized subsystems as discussedabove may be a uniquely non-linear mechanism facilitating the lsquocarriagersquo and distinction ofmultiple concurrent dynamic cell assemblies (and hence cognitive processing) This may beimportant for perceptual grouping binding and transitions as suggested by others (eg von derMalsburg 1985 Gray and Singer 1989)
Second the facility to decouple and reconfigure dynamic cell assemblies througha neuromodulatory control parameter may be important for avoiding low-entropysynchronization manifolds In the absence of itinerancy synchronization would renderneuronal dynamics incapable of representing changes in sensory input or indeed cognitive stateFor example in the model it was observed that small perturbations to one column were ableto induce a global change in neuronal dynamics by disrupting a state of weak synchronization
Synchronization in a model of neuronal dynamics 729
Inevitably alternative dynamic configurations emerge epitomizing the importance of lsquolabilersquobrain dynamics (Friston 2000a)
Thirdly from a purely functionalist point of view the balance between functionalspecialization and functional integration may rely upon the sorts of mechanism modelledin this paper The brain appears to adhere to two principles of functional organization namelysegregation of function within cortical areas and sub-areas and the integration of these areasin perceptual synthesis cognition and sensorimotor integration (Tononi et al 1994 Fristonet al 1995) The balance between preserving specialized dynamics unique to each arearequired by specialization and the coupling among these areas mediating functional integrationmay be achieved by the neuromodulatory mechanisms described above Significantlythe spontaneous variation in the numbers of degrees of freedom (for example whenlow-dimensional synchronization manifolds are shadowed) imparts a dynamic characteristicto this perspective that is precluded by linear mechanisms alone
Finally from a mechanistic perspective complexity and diversity of brain dynamics arerequired for selection of adaptive responses (Friston 2000b) There are many examples ofselective mechanisms operating within the brain All can be seen as performing an ascenton some objective function or fitness landscape These mechanisms require complex anditinerant dynamics to facilitate a thorough exploration of the landscape These landscapesmay range from those involved in perception (where the landscape is defined by predictionerror see eg Rao and Ballard (1998)) to those involved in value learning (where thelandscape is determined by the outcome of a behaviour eg in temporal difference models (seeeg Sutton and Barto 1990)) In short complexity is necessary to maintain an information-rich environment in which the brain can respond to sensory inputs An essential componentof this is the maintenance of diversity which is a prerequisite for selectionist mechanismsat many scales (Edelman 1993) Understanding the mechanistic basis of complex dynamicsas measured electrophysiologically may well depend on the sort of biologically plausiblemodels used in this paper This understanding may be important for both basic and clinicalneuroscience (eg Tass 2002)
The present study supplements and advances on other research in this field The studyof complex phase synchronization and clustering in neural oscillator networks similar tothe present model has previously been the subject of formal mathematical study Forexample Terman and Wang (1995) studied synchronization of blocks of neural oscillatorsthat were subject to perturbation by connected lsquostimulirsquo Somers and Kopell (1995) studiedthe emergence of both synchronization and travelling waves in networks of coupled neuraloscillators In the work of Terman and Wang the local dynamics were based on neurallsquocaricaturersquo models such as the FitzHughndashNagumo equations and are hence more abstract thanthe current study The approach of Somers and Kopell only required that the local dynamicsexhibit fast and relaxation phases and is hence more abstract still In both studies onlyperiodic dynamics are studied Complex patterns of chaos synchronization in neural arrayshave been characterized by nonlinear methods (Kaneko 1998) and their putative contributionto both learning (Nakajima and Ueda 1996) and perception (Zhao and Macau 2001 Breakspear2001) explored However these studies employed models such as the logistic equation andtent map which are quite abstract in comparison to the present study On the other handHansel and Sompolinsky (1996) studied the role of chaos synchronization in a model ofvisual perception in a cortical hypercolumn using a very similar approach to that adoptedhere but with the additional complexity that synaptic delays were incorporated However inthis and earlier work (Hansel and Sompolinsky 1992) only linear measures of dynamics andsynchrony (specifically the auto- and cross-correlation functions) were employed The analysisof chaotic synchrony in the study of Larter et al (1999) was based only upon observation of
730 M Breakspear et al
the Euclidean difference between variables from two nodes within a larger array Hence thesignificance of the present work is that it provides a quantitative nonlinear analysis of complexchaos synchronization in a biophysical model of cortical dynamics Such research is requiredin order to bridge the gap between quantitative nonlinear research into synchronization andcomputation in abstract models of neural arrays and qualitative or linear analysis in morebiophysically plausible models
Acknowledgments
The authors would like to thank Peter Robinson for assistance with the manuscript and R Larterfor helpful advice MB is a recipient of a NSW Institute of Psychiatry Research Grant anda University of Sydney SESQUI postdoctoral fellowship KJF is supported by the WellcomeTrust
References
Abarbanel H Rulkov N and Sushchik M 1996 Generalised synchronization of chaos the auxiliary system approachPhys Rev E 53 4528ndash35
Afraimovich V Verichev N and Rabinovich M 1986 Stochastic synchronization of oscillation in dissipative systemsRadiophys Quantum Electron 29 795ndash801
Breakspear M 2001 Perception of odours by a nonlinear model of the olfactory bulb Int J Neural Syst 11 101ndash24Breakspear M 2002 Nonlinear phase desynchronisation in human electroencephalographic data Human Brain Mapping
15 175ndash98Breakspear M and Terry J R 2002a Nonlinear interdependence in neural systems motivation theory and relevance
Int J Neurosci 112 1163ndash84Breakspear M and Terry J R 2002b Detection and description of nonlinear interdependence in normal multichannel
human EEG Clin Neurophysiol 113 735ndash53Breakspear M and Terry J R 2002c Topographic organization of nonlinear interdependence in multichannel human
EEG Neuroimage 16 822ndash35Destexhe A Rudolph M Fellous J M and Sejnowski T J 2001 Fluctuating synaptic conductances recreate in vivo-like
activity in neocortical neurons Neuroscience 107 13ndash24Douglas R J Mahowald M A and Martin K A 1996 Microarchitecture of neocortical columns Brain TheorymdashBiological
Basis and Computational Principles ed A Aertsen and V Braitenberg (Amsterdam Elsevier)Eckhorn R Bauer R Jordan W Brosch M Kruse W Munk M and Reitboeck H J 1988 Coherent oscillations a
mechanism of feature linking in the visual cortex Multiple electrode and correlation analyses in the cat BiolCybern 60 121ndash30
Edelman G M 1993 Neural Darwinism selection and reentrant signalling in higher brain function Neuron 10 115ndash25Engel A K Kreiter A K Konig P and Singer W 1991 Synchronization of oscillatory neuronal responses between
striate and extrastriate visual cortical areas of the cat Proc Natl Acad Sci USA 88 6048ndash52Friston K J 2000a The labile brain I Neuronal transients and nonlinear coupling Phil Trans R Soc B 355 215ndash36Friston K J 2000b The labile brain II Transients complexity and selection Phil Trans R Soc B 355 237ndash52Friston K J Tononi G Sporns O and Edelman G 1995 Characterising the complexity of neuronal interactions Human
Brain Mapping 3 302ndash14Fujisaka H and Yamada T 1983 Stability theory of synchronized motion in coupled-oscillator system Prog Theor
Phys 69 32ndash47Gatlin L 1972 Information Theory and the Living System (New York Columbia University Press)Gray C M Konig P Engel A and Singer W 1989 Oscillatory responses in cat visual cortex exhibit inter-columnar
synchronization which reflects global stimulus properties Science 338 334ndash7Gray C M and Singer W 1989 Stimulus specific neuronal oscillations in orientation columns of cat visual cortex Proc
Natl Acad Sci USA 86 1698ndash702Haig A R Gordon E Wright J J Meares R A and Bahramali H 2000 Synchronous cortical gamma-band activity in
task-relevant cognition Neuroreport 11 669ndash75Hansel D and Sompolinsky H 1992 Synchronization and computation in a chaotic neural network Phys Rev Lett 68
718ndash21
Synchronization in a model of neuronal dynamics 731
Hansel D and Sompolinsky H 1996 Chaos and synchrony in a model of a hypercolumn in visual cortex J ComputNeurosci 3 7ndash4
Heagy J Carroll T and Pecora L 1998 Desynchronization by periodic orbits Phys Rev E 57 R1253ndash6Hunt B Ott E and Yorke J 1997 Differentiable synchronization of chaos Phys Rev E 55 4029ndash34Josic K 1998 Invariant manifolds and synchronization of coupled dynamical systems Phys Rev Lett 80 3053ndash6Kandel E Schwartz J and Jessell T 2000 Principles of Neural Science (New York McGraw-Hill)Kaneko K 1998 On the strength of attractors in a high-dimensional system Physica D 124 322ndash44Kocarev L Parlitz U and Brown R 2000 Robust synchronization of chaotic systems Phys Rev E 61 3716ndash20Larter R Speelman B and Worth R M 1999 A coupled ordinary differential equation lattice model for the simulation
of epileptic seizures Chaos 9 795ndash804Lumer E Edelman G and Tononi G 1997a Neural dynamics in a model of the thalamocortical system I Layers loops
and the emergence of fast synchronous oscillations Cereb Cortex 7 207ndash27Lumer E Edelman G and Tononi G 1997b Neural dynamics in a model of the thalamocortical system II The role of
neural synchrony tested through perturbations of spike timing Cereb Cortex 7 228ndash36Magleby K 1987 Short-term modulation at synapses Synaptic Function ed G M Edelman W E Gall and
W M Cowan (New York Wiley)Maistrenko Y Maistrenko V Popovich A and Mosekilde E 1998 Transverse instability and riddled basins in a system
of two coupled logistic maps Phys Rev E 57 2713ndash24Morris C and Lecar H 1981 Voltage oscillations in the barnacle giant muscle fiber Biophys J 35 193ndash213Mountcastle V B 1997 The columnar organization of the neocortex Brain 120 701ndash22Mountcastle V B 1998 The Cerebral Cortex (Cambridge MA Harvard University Press)Nakajima H and Ueda Y 1996 Riddled basins of the optimal states in learning dynamical systems Physica D 99 35ndash44Nunez P L 1995 Neocortical Dynamics and Human Brain Rhythms (Oxford Oxford University Press)Osipov G V Pikovsky A S Rosenblum M G and Kurths J 1997 Phase synchronization effects in a lattice of nonidentical
Rossler oscillators Phys Rev E 55 2353ndash61Ott E and Sommerer J 1994 Blowout bifurcations the occurrence of riddled basins and onndashoff intermittency Phys
Lett A 188 39ndash47Pecora L and Carroll T 1990 Synchronization in chaotic systems Phys Rev Lett 64 821ndash4Pikovsky A and Grassberger P 1991 Symmetry breaking bifurcation for coupled chaotic attractors J Phys A Math
Gen 24 4587ndash97Rao R P and Ballard D H 1998 Predictive coding in the visual cortex a functional interpretation of some extra-classical
receptive field effects Nature Neurosci 2 79ndash87Robinson P A Wright J J and Rennie C R 1998 Synchronous oscillations in the cerebral cortex Phys Rev E 57
4578ndash88Rodriguez E George N Lachauz J P Martinerie J Renault B and Varela F J 1999 Perceptionrsquos shadow long distance
synchronization of human brain activity Nature 397 430ndash3Rosenblum M Pikovsky A and Kurths J 1996 Phase synchronization of chaotic oscillators Phys Rev Lett 76 1804ndash7Rosenblum M Pikovsky A and Kurths J 1997 From phase to lag synchronization in coupled chaotic oscillators Phys
Rev Lett 78 4193ndash6Rulkov N and Sushchik M 1997 Robustness of synchronized chaotic oscillations Int J Bifurc Chaos 7 625ndash43Rulkov N Sushchik M Tsimring L and Abarbanel H 1995 Generalized synchronization of chaos in unidirectionally
coupled chaotic systems Phys Rev E 51 980ndash94Schiff S So P Chang T Burke R and Sauer T 1996 Detecting dynamical interdependence and generalized synchrony
through mutual prediction in a neural ensemble Phys Rev E 54 6708ndash24Silberstein R B 1995 Neuromodulations of neocortical dynamics Neocortical Dynamics and Human EEG Rhythms
ed P L Nunez (Oxford Oxford University Press)Singer W 1995 Putative functions of temporal correlations in neocortical processing Large-Scale Neuronal Theories
of the Brain ed C Koch and J Davis (London MIT Press)Somers D and Kopell N 1995 Waves and synchrony in networks of oscillators of relaxation and non-relaxation type
Physica D 89 169ndash83Sutton R S and Barto A G 1990 Time derivative models of Pavlovian reinforcement Learning and Computational
Neuroscience Foundations of Adaptive Networks ed M Gabriel and J Moore (Cambridge MA MIT Press) pp497ndash538
Szentagothai J 1983 The modular architectonic principle of neural centers Rev Physiol Biochem Pharmacol 9811ndash61
Tallon-Baudry C Betrand O and Fischer C 2001 Oscillatory synchrony between human extrastriate areas during visualshort-term memory maintenance J Neurosci 21 RC177(1ndash5)
Tass P A 2002 Desynchronization of brain rhythms with soft phase-resetting techniques Biol Cybern 87 102ndash15
732 M Breakspear et al
Tass P Rosenblum M Weule J Kurths J Pikovsky A Volkmann J Schnitzler A and Freund H 1998 Detection of nmphase locking from noisy data application to magnetoencephalography Phys Rev Lett 81 3291ndash4
Terman D and Wang D 1995 Global competition and local cooperation in a network of neural oscillators Physica D81 148ndash76
Terry J and Breakspear M 2003 An improved algorithm for the detection of dynamical interdependence in bivariatetime-series Biol Cybern 88 129ndash36
Tononi G Sporns O and Edelman G M 1994 A measure for brain complexity relating functional segregation andintegration in the nervous system Proc Natl Acad Sci USA 91 5033ndash7
von der Malsburg C 1985 Nervous structures with dynamical links Phys Chem 89 703ndash10Wang X 1991 Period-doublings to chaos in a simple neural network an analytic proof Complex Syst 5 425ndash41Wennekers T and Pasemann F 1996 Synchronous chaos in high dimensional modular neural networks Int J Bifurc
Chaos 6 2055ndash67Zaks M A Parks E H Rosenblum M G and Kurths J 1999 Alternative locking ratios in imperfect phase synchronization
Phys Rev Lett 82 4228ndash31Zhao L and Macau E N 2001 A network of dynamically coupled chaotic maps for scene segmentation IEEE Trans
Neural Netw 12 1375ndash85
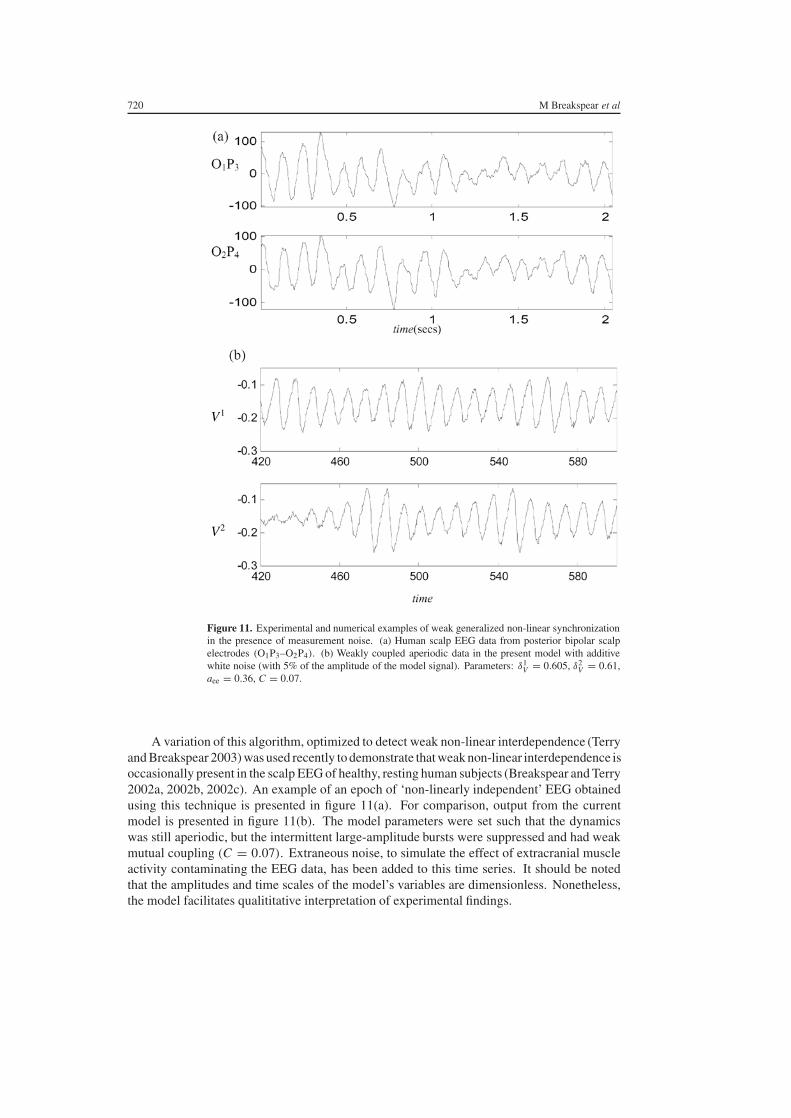
720 M Breakspear et al
V1
V2
Figure 11 Experimental and numerical examples of weak generalized non-linear synchronizationin the presence of measurement noise (a) Human scalp EEG data from posterior bipolar scalpelectrodes (O1P3ndashO2P4) (b) Weakly coupled aperiodic data in the present model with additivewhite noise (with 5 of the amplitude of the model signal) Parameters δ1
V = 0605 δ2V = 061
aee = 036 C = 007
A variation of this algorithm optimized to detect weak non-linear interdependence (Terryand Breakspear 2003) was used recently to demonstrate that weak non-linear interdependence isoccasionally present in the scalp EEG of healthy resting human subjects (Breakspear and Terry2002a 2002b 2002c) An example of an epoch of lsquonon-linearly independentrsquo EEG obtainedusing this technique is presented in figure 11(a) For comparison output from the currentmodel is presented in figure 11(b) The model parameters were set such that the dynamicswas still aperiodic but the intermittent large-amplitude bursts were suppressed and had weakmutual coupling (C = 007) Extraneous noise to simulate the effect of extracranial muscleactivity contaminating the EEG data has been added to this time series It should be notedthat the amplitudes and time scales of the modelrsquos variables are dimensionless Nonethelessthe model facilitates qualititative interpretation of experimental findings
Synchronization in a model of neuronal dynamics 721
43 Chaotic phase synchronization
Whereas the concept of generalized synchronization was mathematically motivated and assuch more suited to theoretical investigations of systems the concept of phase synchronizationderives from a more practical signal processing approach Phase synchronization occurs inboth symmetric and asymmetric settings but is often more useful in the asymmetric case (andin experimental data)
Unfortunately the phase of a chaotic signal is typically ambiguous and can be defined in anumber of different ways For example in the case of the coherent Rossler attractor (Osipovet al 1997) the phase may be defined as the arctangent of two of the variables of the systemMore generally it can sometimes be defined as the angle that an orbit subtends with the centreof an attractor It is also possible to extract the phase of a chaotic signal from time seriesdata by use of the Hilbert transform This has enabled the demonstration of chaotic phasesynchronization in a number of coupled asymmetrical systems (Rosenblum et al 1996 1997Zaks et al 1999) Although such an approach does not guarantee that the phase will be uniqueit does permit the practical analysis of phase in noisy time series (Tass et al 1998)
To illustrate phase synchronization in two coupled columns we again vary the parameterδV which if the columns are viewed as oscillators acts to detune them from a commonfrequency An important question is that of the criteria on the basis of which we concludewhether phase synchronization occurs We use a simple locking criterion whereby phasesynchronization is said to occur if
|θ1(t)minus θ2(t)| lt K (11)
where is θi the phase of column i and K is a constant Notice that this does not mean thatsynchronization implies equality of phases merely that their difference is bounded
An example of phase synchronization with δV = 065 for column 1 and 066 for column 2is given in figure 12 In this figure rNMDA = 03 Qualitatively some sort of phase coherenceis evident in the time series panel (a) This was confirmed by plotting the sine of thephase difference which has been extracted using the Hilbert transform panel (b) Clearly|sin(θ1 minus θ2)| lt K and hence |θ1 minus θ2| is bounded It can also be seen that the phase-synchronized dynamics occurs on a highly structured manifold depicted in figure 12(c)However unlike the case for the generalized synchronization manifold depicted in figure 9 itis evident that this manifold is not a simple transformation of the symmetry manifold
It is also possible to see lsquobimodalrsquo stability whereby a system jumps betweenapproximately zero phase difference and a phase lag phase synchronization of π An exampleis depicted in figure 12(d) At approximately 2000 s the system can be seen to jumpspontaneously from the first to the second of these modes The Z -variable is plotted forgraphical purposes
5 Intermittency and desynchronization
51 Desynchronization in an array of two systems
Diminishing the strength of coupling across the blowout bifurcation does not lead to a simpletransition from synchronous to asynchronous oscillations but is instead associated withcomplex temporal behaviours Specifically when the largest transverse Lyapunov exponent isclose to zero the coupled systems exhibit epochs of synchronous behaviours punctuated bysharp bursts of desynchronization This is observed in systems exhibiting identicalgeneralizedand phase synchronization An example of such a phenomenon is given in figure 13 The timeseries panel (a) shows synchronous dynamics except at time = 1000 where it can be seenthat the two time series desynchronize briefly This is evident in panel (b) as a disruption in the
722 M Breakspear et al
2
1
Figure 12 Phase-synchronized coupled columns (a) The time series of V 1 (solid) and V 2 (b) Thesine of the phase difference obtained through Hilbert transformation of the time series in panel (a)(c) The phase synchronization manifold in the V 1ndashV 2 plane (d) The time series of Z (solid) andZ showing a phase slip of π between two modes
Synchronization in a model of neuronal dynamics 723
Figure 13 An example of phase desynchronization in coupled asymmetric systems with C = 01Time series plots of (a) each V -variable and (b) their Euclidean distance (c) The temporal evolutionof the phase difference between the two systems (d) An example of the phase difference evolutionover a longer time interval
724 M Breakspear et al
orderly evolution of the distance between the two systems and in panel (c) where the evolutionof the phase difference reveals a brief violation of the criterion |φ1 minus φ2| lt constant Anexample of the phase difference evolution over a longer time interval is shown in figure 13(d)Note that this is a three-dimensional plot with sin(φ1 minus φ2) and cos(φ1 minus φ2) as they- and z-axes Hence phase desynchronizations are evident as complete revolutions aroundthe cylindrical phase space in which this variable evolves It can be seen that these phenomenaoccur at irregular intervals in the temporal domain
This behaviour can be explained by considering the nature of a chaotic attractor In thestatistical sense an arbitrary initial condition is expected to lead to the chaotic behaviour onthe attractor Consequently when we calculate the Lyapunov exponents of the system theseexponents correspond to those of the chaotic attractor Typically however there are an infinitenumber of unstable periodic orbits embedded within the attractor If the path of a trajectorybecomes close to one of these orbits it may lsquoshadowrsquo the orbit for a brief period of time Eachof these orbits has a corresponding set of Lyapunov exponents which (like the attractor) can bedivided into tangential and transverse exponents These also vary when we change the couplingparameter but are typically not the same as the exponents of the attractor Hence the largesttransverse exponent of the attractor may be negative yet there may be positive transverseexponents associated with periodic orbits Considering all of these exponents together wewould therefore only expect to observe complete stability of the synchronization manifold inthe case where all of the transverse Lyapunov exponents are negative Bursting behaviourwill be observed when the one of these exponents becomes positive typically correspondingto a periodic orbit of low order (Pikovsky and Grassberger 1991 Rulkov and Sushchik 1997Maistrenko et al 1998 Heagy et al 1998) Due to the complex structure of such periodic orbitsthe desynchronous bursts occur across a wide range of timescales
In figure 14 we present the results of a long integration (16times105 time units) in which thelengths of sequential synchronous epochs of phase synchronization are studied The parametervalues were δV = 065 for column 1 and 0645 for column 2 with coupling strength10 equal to0125 In panel (a) the power spectrum of these sequential lengths is plotted The whiteness ofthis spectrum indicates that the lengths of consecutive synchronous epochs are independentThis is due to the high-dimensional lsquomixingrsquo effect of desynchronization In panel (b) thelengths of the synchronous epochs have been sorted again into the order of increasing lengthThe results for two coupling strengths are plotted C = 0125 (solid) and 0135 (dashed)Several features of this plot are of note Firstly the lengths of the synchronous epochsare distributed over almost three orders of magnitude Secondly for both plots there is anapproximately log-linear relationship between the length of an epoch and its relative orderThis reflects an approximately exponential relationship between the length of a synchronousepoch and its likelihood of occurrence Thirdly this relationship is skewed toward longersynchronous epochs for the stronger coupling strengths
This type of signal with patches of lsquoorderrsquo occurring over many timescales interruptedby bursts of disorder can be shown to have a high information capacity (Gatlin 1972)Moreover complex phase desynchronization may make an infrequent yet robust contributionto the dynamical pattern of the alpha rhythm in scalp EEG data for resting human subjects(Breakspear 2002) It is tempting to postulate that such a lsquoweakly stablersquo dynamics may assistin the generation of the rich and fluid mental associations that spontaneously emerge in restingcognitive states Although this phenomenon has been previously described in a variety of othersystems to our knowledge this is the first description in a biophysical model of neocortex
10 The introduction of parameter mismatch causes an increase in the strength of coupling required for stablesynchronization above the value of 01 required in the symmetric case
Synchronization in a model of neuronal dynamics 725
Figure 14 Properties of sequential epochs of phase-synchronous behaviour over a long integrationtime δ1
V = 065 δ2V = 0645 aee = 04 (a) The flat (white) power spectrum (b) Synchronous
epochs sorted in order of increasing length (solid C = 0125 dashed C = 0134)
52 Synchronization and complex dynamics in larger arrays
The synchronous and complex phenomena described above are also observed in arrays ofcoupled systems with N gt 2 Expanding the size of the array permits complex dynamicsin the spatial in addition to the temporal domain However the basic principles governingthe type and stability of synchronized motions are the same In this section a few of thepossible behaviours are discussed Physiological variation across the array is modelled bymodifying the parameter δV = 07 in each array by adding a random number of maximummagnitude 001
In figure 15(a) an array of N = 20 uncoupled (C = 0) systems is shown The lack ofsynchronization in this time series is evident By comparison the rapid transition to completesynchronization is evident in figure 15(b) when C = 03 In fact 19 of the 20 subsystems
726 M Breakspear et al
(b)
(c)
(a)
Figure 15 Synchronization phenomena in an array of coupled cortical columns (a) N = 20C = 0 (b) N = 20 C = 03 (c) N = 6 C = 01 (d) N = 12 C = 009 (e) N = 12C = 012 A small perturbation has been applied to one of the columns at t = 2500 s (f) Evolutionof the phase difference between two of the columns in panel (e) For clarity the W -variable isplotted in panel (c) and the Z -variable in (e)
have phase synchronized within one single cycle Intermediate values of coupling strength areassociated with a rich variety of phenomena For example subsystems with the most similarparameter values typically synchronize into clusters of various sizes An example of this isillustrated in figure 15(c) Note that there is no synchronization between the two differentclusters each containing three synchronized subsystems
Another complex behaviour occurs when subsystems transiently synchronize into largeclusters then spontaneously desynchronize again Hence the dynamics of the entire arrayjumps between different transient dynamics characterized by varying degrees of freedom Anexample of this phenomenon is given in figure 15(d) In addition it is possible to inducedesynchronization across an entire array by applying a small perturbation to one columnfor one time step An example is illustrated in figure 15(e) A small perturbation has been
Synchronization in a model of neuronal dynamics 727
Figure 15 (Continued)
added to one column after 2500 time units It can be seen that the entire array subsequentlydesynchronizes This illustrates the sensitivity of the dynamics to external stimuli Finally it ispossible to lsquoinducersquo synchronization between selected subsystems by increasing the strength ofcoupling between them selectively This allows an extra level of complexity to be introducedinto the model For example it may allow targeted information transfer between a partialnetwork of the entire array
As discussed in section 51 a univariate signal which jumps between epochs of lowentropy (high repetition) and bursts of high entropy (high variability) can be shown to havehigh information capacity This is because the repetition of certain patterns (as in the English
728 M Breakspear et al
language) permits error detectability Bursts of high entropy act in the opposite directionto increase message variability With a multivariate signal in addition to optimizing theinformation capacity within each signal the overall information capacity can be increasedthrough the type of itinerant synchronization briefly illustrated in this section Synchronizationacross an entire array increases the message reliability (since a single lsquoerrant letterrsquo in onesignal can be easily detected) Conversely desynchronization into multiple clusters increasesmessage variability since each cluster will produce independent information Hence a neuralarray which switches dynamically between a range of cluster configurations can be seen asadaptive in the sense that high synchronous states can be expressed when error detectabilityis a priority (such as during quick and reliable execution of a familiar task) but the array canrapidly desynchronize if variability becomes salient (for example if the environment suddenlybecomes novel) A formal operationalized demonstration of this principle is to be the subjectof future work
6 Discussion
In this paper we present a dynamical model for activity in an array of weakly coupledlocal neural assemblies The model incorporates the fundamental principles governing cellmembrane potential as determined by voltage- and ligand-gated ion channels In this wayit allows a reasonable amount of physiological complexity to be incorporated includingsynaptic activation and receptor subtypeswithout recourse to overwhelming detail Numericalanalysis of this model reveals a transition from independent oscillations to stable chaoticsynchronization depending on the balance of local versus long-range connectivity andbiophysical heterogeneity These excitatory couplings are subject to ascending subcorticalneuromodulation arising principally from monoaminergic nuclei such as Locus Coeruleus(Silberstein 1995) Neuromodulation of extrinsic connections was incorporated into the modelvia a coupling parameter C The stability of the synchronization is determined by the valueof the largest transverse Lyapunov exponent which in turn depends on the coupling parameterC If the coupling is weakened with the result that this exponent becomes positive a blowoutbifurcation occurs and independent oscillations re-emerge In the vicinity of the transitionnon-linear desynchronizations with complex temporal and spatial properties are observed
It is pleasing that complex and itinerant behaviour reminiscent of real neuronaldynamics emerges through such a simple manipulation of a single dynamic control parameterFurthermore the ubiquitous projections of classical neuromodulatory systems in real brainssuggest a clear biological mechanism by which the brain could self-organize throughmoderating the complexity of its own dynamics There are many reasons that the complicateddynamics of the sort demonstrated in this paper may be neurobiologically important Wemention four
First the presence of multiple clusters of phase-synchronized subsystems as discussedabove may be a uniquely non-linear mechanism facilitating the lsquocarriagersquo and distinction ofmultiple concurrent dynamic cell assemblies (and hence cognitive processing) This may beimportant for perceptual grouping binding and transitions as suggested by others (eg von derMalsburg 1985 Gray and Singer 1989)
Second the facility to decouple and reconfigure dynamic cell assemblies througha neuromodulatory control parameter may be important for avoiding low-entropysynchronization manifolds In the absence of itinerancy synchronization would renderneuronal dynamics incapable of representing changes in sensory input or indeed cognitive stateFor example in the model it was observed that small perturbations to one column were ableto induce a global change in neuronal dynamics by disrupting a state of weak synchronization
Synchronization in a model of neuronal dynamics 729
Inevitably alternative dynamic configurations emerge epitomizing the importance of lsquolabilersquobrain dynamics (Friston 2000a)
Thirdly from a purely functionalist point of view the balance between functionalspecialization and functional integration may rely upon the sorts of mechanism modelledin this paper The brain appears to adhere to two principles of functional organization namelysegregation of function within cortical areas and sub-areas and the integration of these areasin perceptual synthesis cognition and sensorimotor integration (Tononi et al 1994 Fristonet al 1995) The balance between preserving specialized dynamics unique to each arearequired by specialization and the coupling among these areas mediating functional integrationmay be achieved by the neuromodulatory mechanisms described above Significantlythe spontaneous variation in the numbers of degrees of freedom (for example whenlow-dimensional synchronization manifolds are shadowed) imparts a dynamic characteristicto this perspective that is precluded by linear mechanisms alone
Finally from a mechanistic perspective complexity and diversity of brain dynamics arerequired for selection of adaptive responses (Friston 2000b) There are many examples ofselective mechanisms operating within the brain All can be seen as performing an ascenton some objective function or fitness landscape These mechanisms require complex anditinerant dynamics to facilitate a thorough exploration of the landscape These landscapesmay range from those involved in perception (where the landscape is defined by predictionerror see eg Rao and Ballard (1998)) to those involved in value learning (where thelandscape is determined by the outcome of a behaviour eg in temporal difference models (seeeg Sutton and Barto 1990)) In short complexity is necessary to maintain an information-rich environment in which the brain can respond to sensory inputs An essential componentof this is the maintenance of diversity which is a prerequisite for selectionist mechanismsat many scales (Edelman 1993) Understanding the mechanistic basis of complex dynamicsas measured electrophysiologically may well depend on the sort of biologically plausiblemodels used in this paper This understanding may be important for both basic and clinicalneuroscience (eg Tass 2002)
The present study supplements and advances on other research in this field The studyof complex phase synchronization and clustering in neural oscillator networks similar tothe present model has previously been the subject of formal mathematical study Forexample Terman and Wang (1995) studied synchronization of blocks of neural oscillatorsthat were subject to perturbation by connected lsquostimulirsquo Somers and Kopell (1995) studiedthe emergence of both synchronization and travelling waves in networks of coupled neuraloscillators In the work of Terman and Wang the local dynamics were based on neurallsquocaricaturersquo models such as the FitzHughndashNagumo equations and are hence more abstract thanthe current study The approach of Somers and Kopell only required that the local dynamicsexhibit fast and relaxation phases and is hence more abstract still In both studies onlyperiodic dynamics are studied Complex patterns of chaos synchronization in neural arrayshave been characterized by nonlinear methods (Kaneko 1998) and their putative contributionto both learning (Nakajima and Ueda 1996) and perception (Zhao and Macau 2001 Breakspear2001) explored However these studies employed models such as the logistic equation andtent map which are quite abstract in comparison to the present study On the other handHansel and Sompolinsky (1996) studied the role of chaos synchronization in a model ofvisual perception in a cortical hypercolumn using a very similar approach to that adoptedhere but with the additional complexity that synaptic delays were incorporated However inthis and earlier work (Hansel and Sompolinsky 1992) only linear measures of dynamics andsynchrony (specifically the auto- and cross-correlation functions) were employed The analysisof chaotic synchrony in the study of Larter et al (1999) was based only upon observation of
730 M Breakspear et al
the Euclidean difference between variables from two nodes within a larger array Hence thesignificance of the present work is that it provides a quantitative nonlinear analysis of complexchaos synchronization in a biophysical model of cortical dynamics Such research is requiredin order to bridge the gap between quantitative nonlinear research into synchronization andcomputation in abstract models of neural arrays and qualitative or linear analysis in morebiophysically plausible models
Acknowledgments
The authors would like to thank Peter Robinson for assistance with the manuscript and R Larterfor helpful advice MB is a recipient of a NSW Institute of Psychiatry Research Grant anda University of Sydney SESQUI postdoctoral fellowship KJF is supported by the WellcomeTrust
References
Abarbanel H Rulkov N and Sushchik M 1996 Generalised synchronization of chaos the auxiliary system approachPhys Rev E 53 4528ndash35
Afraimovich V Verichev N and Rabinovich M 1986 Stochastic synchronization of oscillation in dissipative systemsRadiophys Quantum Electron 29 795ndash801
Breakspear M 2001 Perception of odours by a nonlinear model of the olfactory bulb Int J Neural Syst 11 101ndash24Breakspear M 2002 Nonlinear phase desynchronisation in human electroencephalographic data Human Brain Mapping
15 175ndash98Breakspear M and Terry J R 2002a Nonlinear interdependence in neural systems motivation theory and relevance
Int J Neurosci 112 1163ndash84Breakspear M and Terry J R 2002b Detection and description of nonlinear interdependence in normal multichannel
human EEG Clin Neurophysiol 113 735ndash53Breakspear M and Terry J R 2002c Topographic organization of nonlinear interdependence in multichannel human
EEG Neuroimage 16 822ndash35Destexhe A Rudolph M Fellous J M and Sejnowski T J 2001 Fluctuating synaptic conductances recreate in vivo-like
activity in neocortical neurons Neuroscience 107 13ndash24Douglas R J Mahowald M A and Martin K A 1996 Microarchitecture of neocortical columns Brain TheorymdashBiological
Basis and Computational Principles ed A Aertsen and V Braitenberg (Amsterdam Elsevier)Eckhorn R Bauer R Jordan W Brosch M Kruse W Munk M and Reitboeck H J 1988 Coherent oscillations a
mechanism of feature linking in the visual cortex Multiple electrode and correlation analyses in the cat BiolCybern 60 121ndash30
Edelman G M 1993 Neural Darwinism selection and reentrant signalling in higher brain function Neuron 10 115ndash25Engel A K Kreiter A K Konig P and Singer W 1991 Synchronization of oscillatory neuronal responses between
striate and extrastriate visual cortical areas of the cat Proc Natl Acad Sci USA 88 6048ndash52Friston K J 2000a The labile brain I Neuronal transients and nonlinear coupling Phil Trans R Soc B 355 215ndash36Friston K J 2000b The labile brain II Transients complexity and selection Phil Trans R Soc B 355 237ndash52Friston K J Tononi G Sporns O and Edelman G 1995 Characterising the complexity of neuronal interactions Human
Brain Mapping 3 302ndash14Fujisaka H and Yamada T 1983 Stability theory of synchronized motion in coupled-oscillator system Prog Theor
Phys 69 32ndash47Gatlin L 1972 Information Theory and the Living System (New York Columbia University Press)Gray C M Konig P Engel A and Singer W 1989 Oscillatory responses in cat visual cortex exhibit inter-columnar
synchronization which reflects global stimulus properties Science 338 334ndash7Gray C M and Singer W 1989 Stimulus specific neuronal oscillations in orientation columns of cat visual cortex Proc
Natl Acad Sci USA 86 1698ndash702Haig A R Gordon E Wright J J Meares R A and Bahramali H 2000 Synchronous cortical gamma-band activity in
task-relevant cognition Neuroreport 11 669ndash75Hansel D and Sompolinsky H 1992 Synchronization and computation in a chaotic neural network Phys Rev Lett 68
718ndash21
Synchronization in a model of neuronal dynamics 731
Hansel D and Sompolinsky H 1996 Chaos and synchrony in a model of a hypercolumn in visual cortex J ComputNeurosci 3 7ndash4
Heagy J Carroll T and Pecora L 1998 Desynchronization by periodic orbits Phys Rev E 57 R1253ndash6Hunt B Ott E and Yorke J 1997 Differentiable synchronization of chaos Phys Rev E 55 4029ndash34Josic K 1998 Invariant manifolds and synchronization of coupled dynamical systems Phys Rev Lett 80 3053ndash6Kandel E Schwartz J and Jessell T 2000 Principles of Neural Science (New York McGraw-Hill)Kaneko K 1998 On the strength of attractors in a high-dimensional system Physica D 124 322ndash44Kocarev L Parlitz U and Brown R 2000 Robust synchronization of chaotic systems Phys Rev E 61 3716ndash20Larter R Speelman B and Worth R M 1999 A coupled ordinary differential equation lattice model for the simulation
of epileptic seizures Chaos 9 795ndash804Lumer E Edelman G and Tononi G 1997a Neural dynamics in a model of the thalamocortical system I Layers loops
and the emergence of fast synchronous oscillations Cereb Cortex 7 207ndash27Lumer E Edelman G and Tononi G 1997b Neural dynamics in a model of the thalamocortical system II The role of
neural synchrony tested through perturbations of spike timing Cereb Cortex 7 228ndash36Magleby K 1987 Short-term modulation at synapses Synaptic Function ed G M Edelman W E Gall and
W M Cowan (New York Wiley)Maistrenko Y Maistrenko V Popovich A and Mosekilde E 1998 Transverse instability and riddled basins in a system
of two coupled logistic maps Phys Rev E 57 2713ndash24Morris C and Lecar H 1981 Voltage oscillations in the barnacle giant muscle fiber Biophys J 35 193ndash213Mountcastle V B 1997 The columnar organization of the neocortex Brain 120 701ndash22Mountcastle V B 1998 The Cerebral Cortex (Cambridge MA Harvard University Press)Nakajima H and Ueda Y 1996 Riddled basins of the optimal states in learning dynamical systems Physica D 99 35ndash44Nunez P L 1995 Neocortical Dynamics and Human Brain Rhythms (Oxford Oxford University Press)Osipov G V Pikovsky A S Rosenblum M G and Kurths J 1997 Phase synchronization effects in a lattice of nonidentical
Rossler oscillators Phys Rev E 55 2353ndash61Ott E and Sommerer J 1994 Blowout bifurcations the occurrence of riddled basins and onndashoff intermittency Phys
Lett A 188 39ndash47Pecora L and Carroll T 1990 Synchronization in chaotic systems Phys Rev Lett 64 821ndash4Pikovsky A and Grassberger P 1991 Symmetry breaking bifurcation for coupled chaotic attractors J Phys A Math
Gen 24 4587ndash97Rao R P and Ballard D H 1998 Predictive coding in the visual cortex a functional interpretation of some extra-classical
receptive field effects Nature Neurosci 2 79ndash87Robinson P A Wright J J and Rennie C R 1998 Synchronous oscillations in the cerebral cortex Phys Rev E 57
4578ndash88Rodriguez E George N Lachauz J P Martinerie J Renault B and Varela F J 1999 Perceptionrsquos shadow long distance
synchronization of human brain activity Nature 397 430ndash3Rosenblum M Pikovsky A and Kurths J 1996 Phase synchronization of chaotic oscillators Phys Rev Lett 76 1804ndash7Rosenblum M Pikovsky A and Kurths J 1997 From phase to lag synchronization in coupled chaotic oscillators Phys
Rev Lett 78 4193ndash6Rulkov N and Sushchik M 1997 Robustness of synchronized chaotic oscillations Int J Bifurc Chaos 7 625ndash43Rulkov N Sushchik M Tsimring L and Abarbanel H 1995 Generalized synchronization of chaos in unidirectionally
coupled chaotic systems Phys Rev E 51 980ndash94Schiff S So P Chang T Burke R and Sauer T 1996 Detecting dynamical interdependence and generalized synchrony
through mutual prediction in a neural ensemble Phys Rev E 54 6708ndash24Silberstein R B 1995 Neuromodulations of neocortical dynamics Neocortical Dynamics and Human EEG Rhythms
ed P L Nunez (Oxford Oxford University Press)Singer W 1995 Putative functions of temporal correlations in neocortical processing Large-Scale Neuronal Theories
of the Brain ed C Koch and J Davis (London MIT Press)Somers D and Kopell N 1995 Waves and synchrony in networks of oscillators of relaxation and non-relaxation type
Physica D 89 169ndash83Sutton R S and Barto A G 1990 Time derivative models of Pavlovian reinforcement Learning and Computational
Neuroscience Foundations of Adaptive Networks ed M Gabriel and J Moore (Cambridge MA MIT Press) pp497ndash538
Szentagothai J 1983 The modular architectonic principle of neural centers Rev Physiol Biochem Pharmacol 9811ndash61
Tallon-Baudry C Betrand O and Fischer C 2001 Oscillatory synchrony between human extrastriate areas during visualshort-term memory maintenance J Neurosci 21 RC177(1ndash5)
Tass P A 2002 Desynchronization of brain rhythms with soft phase-resetting techniques Biol Cybern 87 102ndash15
732 M Breakspear et al
Tass P Rosenblum M Weule J Kurths J Pikovsky A Volkmann J Schnitzler A and Freund H 1998 Detection of nmphase locking from noisy data application to magnetoencephalography Phys Rev Lett 81 3291ndash4
Terman D and Wang D 1995 Global competition and local cooperation in a network of neural oscillators Physica D81 148ndash76
Terry J and Breakspear M 2003 An improved algorithm for the detection of dynamical interdependence in bivariatetime-series Biol Cybern 88 129ndash36
Tononi G Sporns O and Edelman G M 1994 A measure for brain complexity relating functional segregation andintegration in the nervous system Proc Natl Acad Sci USA 91 5033ndash7
von der Malsburg C 1985 Nervous structures with dynamical links Phys Chem 89 703ndash10Wang X 1991 Period-doublings to chaos in a simple neural network an analytic proof Complex Syst 5 425ndash41Wennekers T and Pasemann F 1996 Synchronous chaos in high dimensional modular neural networks Int J Bifurc
Chaos 6 2055ndash67Zaks M A Parks E H Rosenblum M G and Kurths J 1999 Alternative locking ratios in imperfect phase synchronization
Phys Rev Lett 82 4228ndash31Zhao L and Macau E N 2001 A network of dynamically coupled chaotic maps for scene segmentation IEEE Trans
Neural Netw 12 1375ndash85

Synchronization in a model of neuronal dynamics 721
43 Chaotic phase synchronization
Whereas the concept of generalized synchronization was mathematically motivated and assuch more suited to theoretical investigations of systems the concept of phase synchronizationderives from a more practical signal processing approach Phase synchronization occurs inboth symmetric and asymmetric settings but is often more useful in the asymmetric case (andin experimental data)
Unfortunately the phase of a chaotic signal is typically ambiguous and can be defined in anumber of different ways For example in the case of the coherent Rossler attractor (Osipovet al 1997) the phase may be defined as the arctangent of two of the variables of the systemMore generally it can sometimes be defined as the angle that an orbit subtends with the centreof an attractor It is also possible to extract the phase of a chaotic signal from time seriesdata by use of the Hilbert transform This has enabled the demonstration of chaotic phasesynchronization in a number of coupled asymmetrical systems (Rosenblum et al 1996 1997Zaks et al 1999) Although such an approach does not guarantee that the phase will be uniqueit does permit the practical analysis of phase in noisy time series (Tass et al 1998)
To illustrate phase synchronization in two coupled columns we again vary the parameterδV which if the columns are viewed as oscillators acts to detune them from a commonfrequency An important question is that of the criteria on the basis of which we concludewhether phase synchronization occurs We use a simple locking criterion whereby phasesynchronization is said to occur if
|θ1(t)minus θ2(t)| lt K (11)
where is θi the phase of column i and K is a constant Notice that this does not mean thatsynchronization implies equality of phases merely that their difference is bounded
An example of phase synchronization with δV = 065 for column 1 and 066 for column 2is given in figure 12 In this figure rNMDA = 03 Qualitatively some sort of phase coherenceis evident in the time series panel (a) This was confirmed by plotting the sine of thephase difference which has been extracted using the Hilbert transform panel (b) Clearly|sin(θ1 minus θ2)| lt K and hence |θ1 minus θ2| is bounded It can also be seen that the phase-synchronized dynamics occurs on a highly structured manifold depicted in figure 12(c)However unlike the case for the generalized synchronization manifold depicted in figure 9 itis evident that this manifold is not a simple transformation of the symmetry manifold
It is also possible to see lsquobimodalrsquo stability whereby a system jumps betweenapproximately zero phase difference and a phase lag phase synchronization of π An exampleis depicted in figure 12(d) At approximately 2000 s the system can be seen to jumpspontaneously from the first to the second of these modes The Z -variable is plotted forgraphical purposes
5 Intermittency and desynchronization
51 Desynchronization in an array of two systems
Diminishing the strength of coupling across the blowout bifurcation does not lead to a simpletransition from synchronous to asynchronous oscillations but is instead associated withcomplex temporal behaviours Specifically when the largest transverse Lyapunov exponent isclose to zero the coupled systems exhibit epochs of synchronous behaviours punctuated bysharp bursts of desynchronization This is observed in systems exhibiting identicalgeneralizedand phase synchronization An example of such a phenomenon is given in figure 13 The timeseries panel (a) shows synchronous dynamics except at time = 1000 where it can be seenthat the two time series desynchronize briefly This is evident in panel (b) as a disruption in the
722 M Breakspear et al
2
1
Figure 12 Phase-synchronized coupled columns (a) The time series of V 1 (solid) and V 2 (b) Thesine of the phase difference obtained through Hilbert transformation of the time series in panel (a)(c) The phase synchronization manifold in the V 1ndashV 2 plane (d) The time series of Z (solid) andZ showing a phase slip of π between two modes
Synchronization in a model of neuronal dynamics 723
Figure 13 An example of phase desynchronization in coupled asymmetric systems with C = 01Time series plots of (a) each V -variable and (b) their Euclidean distance (c) The temporal evolutionof the phase difference between the two systems (d) An example of the phase difference evolutionover a longer time interval
724 M Breakspear et al
orderly evolution of the distance between the two systems and in panel (c) where the evolutionof the phase difference reveals a brief violation of the criterion |φ1 minus φ2| lt constant Anexample of the phase difference evolution over a longer time interval is shown in figure 13(d)Note that this is a three-dimensional plot with sin(φ1 minus φ2) and cos(φ1 minus φ2) as they- and z-axes Hence phase desynchronizations are evident as complete revolutions aroundthe cylindrical phase space in which this variable evolves It can be seen that these phenomenaoccur at irregular intervals in the temporal domain
This behaviour can be explained by considering the nature of a chaotic attractor In thestatistical sense an arbitrary initial condition is expected to lead to the chaotic behaviour onthe attractor Consequently when we calculate the Lyapunov exponents of the system theseexponents correspond to those of the chaotic attractor Typically however there are an infinitenumber of unstable periodic orbits embedded within the attractor If the path of a trajectorybecomes close to one of these orbits it may lsquoshadowrsquo the orbit for a brief period of time Eachof these orbits has a corresponding set of Lyapunov exponents which (like the attractor) can bedivided into tangential and transverse exponents These also vary when we change the couplingparameter but are typically not the same as the exponents of the attractor Hence the largesttransverse exponent of the attractor may be negative yet there may be positive transverseexponents associated with periodic orbits Considering all of these exponents together wewould therefore only expect to observe complete stability of the synchronization manifold inthe case where all of the transverse Lyapunov exponents are negative Bursting behaviourwill be observed when the one of these exponents becomes positive typically correspondingto a periodic orbit of low order (Pikovsky and Grassberger 1991 Rulkov and Sushchik 1997Maistrenko et al 1998 Heagy et al 1998) Due to the complex structure of such periodic orbitsthe desynchronous bursts occur across a wide range of timescales
In figure 14 we present the results of a long integration (16times105 time units) in which thelengths of sequential synchronous epochs of phase synchronization are studied The parametervalues were δV = 065 for column 1 and 0645 for column 2 with coupling strength10 equal to0125 In panel (a) the power spectrum of these sequential lengths is plotted The whiteness ofthis spectrum indicates that the lengths of consecutive synchronous epochs are independentThis is due to the high-dimensional lsquomixingrsquo effect of desynchronization In panel (b) thelengths of the synchronous epochs have been sorted again into the order of increasing lengthThe results for two coupling strengths are plotted C = 0125 (solid) and 0135 (dashed)Several features of this plot are of note Firstly the lengths of the synchronous epochsare distributed over almost three orders of magnitude Secondly for both plots there is anapproximately log-linear relationship between the length of an epoch and its relative orderThis reflects an approximately exponential relationship between the length of a synchronousepoch and its likelihood of occurrence Thirdly this relationship is skewed toward longersynchronous epochs for the stronger coupling strengths
This type of signal with patches of lsquoorderrsquo occurring over many timescales interruptedby bursts of disorder can be shown to have a high information capacity (Gatlin 1972)Moreover complex phase desynchronization may make an infrequent yet robust contributionto the dynamical pattern of the alpha rhythm in scalp EEG data for resting human subjects(Breakspear 2002) It is tempting to postulate that such a lsquoweakly stablersquo dynamics may assistin the generation of the rich and fluid mental associations that spontaneously emerge in restingcognitive states Although this phenomenon has been previously described in a variety of othersystems to our knowledge this is the first description in a biophysical model of neocortex
10 The introduction of parameter mismatch causes an increase in the strength of coupling required for stablesynchronization above the value of 01 required in the symmetric case
Synchronization in a model of neuronal dynamics 725
Figure 14 Properties of sequential epochs of phase-synchronous behaviour over a long integrationtime δ1
V = 065 δ2V = 0645 aee = 04 (a) The flat (white) power spectrum (b) Synchronous
epochs sorted in order of increasing length (solid C = 0125 dashed C = 0134)
52 Synchronization and complex dynamics in larger arrays
The synchronous and complex phenomena described above are also observed in arrays ofcoupled systems with N gt 2 Expanding the size of the array permits complex dynamicsin the spatial in addition to the temporal domain However the basic principles governingthe type and stability of synchronized motions are the same In this section a few of thepossible behaviours are discussed Physiological variation across the array is modelled bymodifying the parameter δV = 07 in each array by adding a random number of maximummagnitude 001
In figure 15(a) an array of N = 20 uncoupled (C = 0) systems is shown The lack ofsynchronization in this time series is evident By comparison the rapid transition to completesynchronization is evident in figure 15(b) when C = 03 In fact 19 of the 20 subsystems
726 M Breakspear et al
(b)
(c)
(a)
Figure 15 Synchronization phenomena in an array of coupled cortical columns (a) N = 20C = 0 (b) N = 20 C = 03 (c) N = 6 C = 01 (d) N = 12 C = 009 (e) N = 12C = 012 A small perturbation has been applied to one of the columns at t = 2500 s (f) Evolutionof the phase difference between two of the columns in panel (e) For clarity the W -variable isplotted in panel (c) and the Z -variable in (e)
have phase synchronized within one single cycle Intermediate values of coupling strength areassociated with a rich variety of phenomena For example subsystems with the most similarparameter values typically synchronize into clusters of various sizes An example of this isillustrated in figure 15(c) Note that there is no synchronization between the two differentclusters each containing three synchronized subsystems
Another complex behaviour occurs when subsystems transiently synchronize into largeclusters then spontaneously desynchronize again Hence the dynamics of the entire arrayjumps between different transient dynamics characterized by varying degrees of freedom Anexample of this phenomenon is given in figure 15(d) In addition it is possible to inducedesynchronization across an entire array by applying a small perturbation to one columnfor one time step An example is illustrated in figure 15(e) A small perturbation has been
Synchronization in a model of neuronal dynamics 727
Figure 15 (Continued)
added to one column after 2500 time units It can be seen that the entire array subsequentlydesynchronizes This illustrates the sensitivity of the dynamics to external stimuli Finally it ispossible to lsquoinducersquo synchronization between selected subsystems by increasing the strength ofcoupling between them selectively This allows an extra level of complexity to be introducedinto the model For example it may allow targeted information transfer between a partialnetwork of the entire array
As discussed in section 51 a univariate signal which jumps between epochs of lowentropy (high repetition) and bursts of high entropy (high variability) can be shown to havehigh information capacity This is because the repetition of certain patterns (as in the English
728 M Breakspear et al
language) permits error detectability Bursts of high entropy act in the opposite directionto increase message variability With a multivariate signal in addition to optimizing theinformation capacity within each signal the overall information capacity can be increasedthrough the type of itinerant synchronization briefly illustrated in this section Synchronizationacross an entire array increases the message reliability (since a single lsquoerrant letterrsquo in onesignal can be easily detected) Conversely desynchronization into multiple clusters increasesmessage variability since each cluster will produce independent information Hence a neuralarray which switches dynamically between a range of cluster configurations can be seen asadaptive in the sense that high synchronous states can be expressed when error detectabilityis a priority (such as during quick and reliable execution of a familiar task) but the array canrapidly desynchronize if variability becomes salient (for example if the environment suddenlybecomes novel) A formal operationalized demonstration of this principle is to be the subjectof future work
6 Discussion
In this paper we present a dynamical model for activity in an array of weakly coupledlocal neural assemblies The model incorporates the fundamental principles governing cellmembrane potential as determined by voltage- and ligand-gated ion channels In this wayit allows a reasonable amount of physiological complexity to be incorporated includingsynaptic activation and receptor subtypeswithout recourse to overwhelming detail Numericalanalysis of this model reveals a transition from independent oscillations to stable chaoticsynchronization depending on the balance of local versus long-range connectivity andbiophysical heterogeneity These excitatory couplings are subject to ascending subcorticalneuromodulation arising principally from monoaminergic nuclei such as Locus Coeruleus(Silberstein 1995) Neuromodulation of extrinsic connections was incorporated into the modelvia a coupling parameter C The stability of the synchronization is determined by the valueof the largest transverse Lyapunov exponent which in turn depends on the coupling parameterC If the coupling is weakened with the result that this exponent becomes positive a blowoutbifurcation occurs and independent oscillations re-emerge In the vicinity of the transitionnon-linear desynchronizations with complex temporal and spatial properties are observed
It is pleasing that complex and itinerant behaviour reminiscent of real neuronaldynamics emerges through such a simple manipulation of a single dynamic control parameterFurthermore the ubiquitous projections of classical neuromodulatory systems in real brainssuggest a clear biological mechanism by which the brain could self-organize throughmoderating the complexity of its own dynamics There are many reasons that the complicateddynamics of the sort demonstrated in this paper may be neurobiologically important Wemention four
First the presence of multiple clusters of phase-synchronized subsystems as discussedabove may be a uniquely non-linear mechanism facilitating the lsquocarriagersquo and distinction ofmultiple concurrent dynamic cell assemblies (and hence cognitive processing) This may beimportant for perceptual grouping binding and transitions as suggested by others (eg von derMalsburg 1985 Gray and Singer 1989)
Second the facility to decouple and reconfigure dynamic cell assemblies througha neuromodulatory control parameter may be important for avoiding low-entropysynchronization manifolds In the absence of itinerancy synchronization would renderneuronal dynamics incapable of representing changes in sensory input or indeed cognitive stateFor example in the model it was observed that small perturbations to one column were ableto induce a global change in neuronal dynamics by disrupting a state of weak synchronization
Synchronization in a model of neuronal dynamics 729
Inevitably alternative dynamic configurations emerge epitomizing the importance of lsquolabilersquobrain dynamics (Friston 2000a)
Thirdly from a purely functionalist point of view the balance between functionalspecialization and functional integration may rely upon the sorts of mechanism modelledin this paper The brain appears to adhere to two principles of functional organization namelysegregation of function within cortical areas and sub-areas and the integration of these areasin perceptual synthesis cognition and sensorimotor integration (Tononi et al 1994 Fristonet al 1995) The balance between preserving specialized dynamics unique to each arearequired by specialization and the coupling among these areas mediating functional integrationmay be achieved by the neuromodulatory mechanisms described above Significantlythe spontaneous variation in the numbers of degrees of freedom (for example whenlow-dimensional synchronization manifolds are shadowed) imparts a dynamic characteristicto this perspective that is precluded by linear mechanisms alone
Finally from a mechanistic perspective complexity and diversity of brain dynamics arerequired for selection of adaptive responses (Friston 2000b) There are many examples ofselective mechanisms operating within the brain All can be seen as performing an ascenton some objective function or fitness landscape These mechanisms require complex anditinerant dynamics to facilitate a thorough exploration of the landscape These landscapesmay range from those involved in perception (where the landscape is defined by predictionerror see eg Rao and Ballard (1998)) to those involved in value learning (where thelandscape is determined by the outcome of a behaviour eg in temporal difference models (seeeg Sutton and Barto 1990)) In short complexity is necessary to maintain an information-rich environment in which the brain can respond to sensory inputs An essential componentof this is the maintenance of diversity which is a prerequisite for selectionist mechanismsat many scales (Edelman 1993) Understanding the mechanistic basis of complex dynamicsas measured electrophysiologically may well depend on the sort of biologically plausiblemodels used in this paper This understanding may be important for both basic and clinicalneuroscience (eg Tass 2002)
The present study supplements and advances on other research in this field The studyof complex phase synchronization and clustering in neural oscillator networks similar tothe present model has previously been the subject of formal mathematical study Forexample Terman and Wang (1995) studied synchronization of blocks of neural oscillatorsthat were subject to perturbation by connected lsquostimulirsquo Somers and Kopell (1995) studiedthe emergence of both synchronization and travelling waves in networks of coupled neuraloscillators In the work of Terman and Wang the local dynamics were based on neurallsquocaricaturersquo models such as the FitzHughndashNagumo equations and are hence more abstract thanthe current study The approach of Somers and Kopell only required that the local dynamicsexhibit fast and relaxation phases and is hence more abstract still In both studies onlyperiodic dynamics are studied Complex patterns of chaos synchronization in neural arrayshave been characterized by nonlinear methods (Kaneko 1998) and their putative contributionto both learning (Nakajima and Ueda 1996) and perception (Zhao and Macau 2001 Breakspear2001) explored However these studies employed models such as the logistic equation andtent map which are quite abstract in comparison to the present study On the other handHansel and Sompolinsky (1996) studied the role of chaos synchronization in a model ofvisual perception in a cortical hypercolumn using a very similar approach to that adoptedhere but with the additional complexity that synaptic delays were incorporated However inthis and earlier work (Hansel and Sompolinsky 1992) only linear measures of dynamics andsynchrony (specifically the auto- and cross-correlation functions) were employed The analysisof chaotic synchrony in the study of Larter et al (1999) was based only upon observation of
730 M Breakspear et al
the Euclidean difference between variables from two nodes within a larger array Hence thesignificance of the present work is that it provides a quantitative nonlinear analysis of complexchaos synchronization in a biophysical model of cortical dynamics Such research is requiredin order to bridge the gap between quantitative nonlinear research into synchronization andcomputation in abstract models of neural arrays and qualitative or linear analysis in morebiophysically plausible models
Acknowledgments
The authors would like to thank Peter Robinson for assistance with the manuscript and R Larterfor helpful advice MB is a recipient of a NSW Institute of Psychiatry Research Grant anda University of Sydney SESQUI postdoctoral fellowship KJF is supported by the WellcomeTrust
References
Abarbanel H Rulkov N and Sushchik M 1996 Generalised synchronization of chaos the auxiliary system approachPhys Rev E 53 4528ndash35
Afraimovich V Verichev N and Rabinovich M 1986 Stochastic synchronization of oscillation in dissipative systemsRadiophys Quantum Electron 29 795ndash801
Breakspear M 2001 Perception of odours by a nonlinear model of the olfactory bulb Int J Neural Syst 11 101ndash24Breakspear M 2002 Nonlinear phase desynchronisation in human electroencephalographic data Human Brain Mapping
15 175ndash98Breakspear M and Terry J R 2002a Nonlinear interdependence in neural systems motivation theory and relevance
Int J Neurosci 112 1163ndash84Breakspear M and Terry J R 2002b Detection and description of nonlinear interdependence in normal multichannel
human EEG Clin Neurophysiol 113 735ndash53Breakspear M and Terry J R 2002c Topographic organization of nonlinear interdependence in multichannel human
EEG Neuroimage 16 822ndash35Destexhe A Rudolph M Fellous J M and Sejnowski T J 2001 Fluctuating synaptic conductances recreate in vivo-like
activity in neocortical neurons Neuroscience 107 13ndash24Douglas R J Mahowald M A and Martin K A 1996 Microarchitecture of neocortical columns Brain TheorymdashBiological
Basis and Computational Principles ed A Aertsen and V Braitenberg (Amsterdam Elsevier)Eckhorn R Bauer R Jordan W Brosch M Kruse W Munk M and Reitboeck H J 1988 Coherent oscillations a
mechanism of feature linking in the visual cortex Multiple electrode and correlation analyses in the cat BiolCybern 60 121ndash30
Edelman G M 1993 Neural Darwinism selection and reentrant signalling in higher brain function Neuron 10 115ndash25Engel A K Kreiter A K Konig P and Singer W 1991 Synchronization of oscillatory neuronal responses between
striate and extrastriate visual cortical areas of the cat Proc Natl Acad Sci USA 88 6048ndash52Friston K J 2000a The labile brain I Neuronal transients and nonlinear coupling Phil Trans R Soc B 355 215ndash36Friston K J 2000b The labile brain II Transients complexity and selection Phil Trans R Soc B 355 237ndash52Friston K J Tononi G Sporns O and Edelman G 1995 Characterising the complexity of neuronal interactions Human
Brain Mapping 3 302ndash14Fujisaka H and Yamada T 1983 Stability theory of synchronized motion in coupled-oscillator system Prog Theor
Phys 69 32ndash47Gatlin L 1972 Information Theory and the Living System (New York Columbia University Press)Gray C M Konig P Engel A and Singer W 1989 Oscillatory responses in cat visual cortex exhibit inter-columnar
synchronization which reflects global stimulus properties Science 338 334ndash7Gray C M and Singer W 1989 Stimulus specific neuronal oscillations in orientation columns of cat visual cortex Proc
Natl Acad Sci USA 86 1698ndash702Haig A R Gordon E Wright J J Meares R A and Bahramali H 2000 Synchronous cortical gamma-band activity in
task-relevant cognition Neuroreport 11 669ndash75Hansel D and Sompolinsky H 1992 Synchronization and computation in a chaotic neural network Phys Rev Lett 68
718ndash21
Synchronization in a model of neuronal dynamics 731
Hansel D and Sompolinsky H 1996 Chaos and synchrony in a model of a hypercolumn in visual cortex J ComputNeurosci 3 7ndash4
Heagy J Carroll T and Pecora L 1998 Desynchronization by periodic orbits Phys Rev E 57 R1253ndash6Hunt B Ott E and Yorke J 1997 Differentiable synchronization of chaos Phys Rev E 55 4029ndash34Josic K 1998 Invariant manifolds and synchronization of coupled dynamical systems Phys Rev Lett 80 3053ndash6Kandel E Schwartz J and Jessell T 2000 Principles of Neural Science (New York McGraw-Hill)Kaneko K 1998 On the strength of attractors in a high-dimensional system Physica D 124 322ndash44Kocarev L Parlitz U and Brown R 2000 Robust synchronization of chaotic systems Phys Rev E 61 3716ndash20Larter R Speelman B and Worth R M 1999 A coupled ordinary differential equation lattice model for the simulation
of epileptic seizures Chaos 9 795ndash804Lumer E Edelman G and Tononi G 1997a Neural dynamics in a model of the thalamocortical system I Layers loops
and the emergence of fast synchronous oscillations Cereb Cortex 7 207ndash27Lumer E Edelman G and Tononi G 1997b Neural dynamics in a model of the thalamocortical system II The role of
neural synchrony tested through perturbations of spike timing Cereb Cortex 7 228ndash36Magleby K 1987 Short-term modulation at synapses Synaptic Function ed G M Edelman W E Gall and
W M Cowan (New York Wiley)Maistrenko Y Maistrenko V Popovich A and Mosekilde E 1998 Transverse instability and riddled basins in a system
of two coupled logistic maps Phys Rev E 57 2713ndash24Morris C and Lecar H 1981 Voltage oscillations in the barnacle giant muscle fiber Biophys J 35 193ndash213Mountcastle V B 1997 The columnar organization of the neocortex Brain 120 701ndash22Mountcastle V B 1998 The Cerebral Cortex (Cambridge MA Harvard University Press)Nakajima H and Ueda Y 1996 Riddled basins of the optimal states in learning dynamical systems Physica D 99 35ndash44Nunez P L 1995 Neocortical Dynamics and Human Brain Rhythms (Oxford Oxford University Press)Osipov G V Pikovsky A S Rosenblum M G and Kurths J 1997 Phase synchronization effects in a lattice of nonidentical
Rossler oscillators Phys Rev E 55 2353ndash61Ott E and Sommerer J 1994 Blowout bifurcations the occurrence of riddled basins and onndashoff intermittency Phys
Lett A 188 39ndash47Pecora L and Carroll T 1990 Synchronization in chaotic systems Phys Rev Lett 64 821ndash4Pikovsky A and Grassberger P 1991 Symmetry breaking bifurcation for coupled chaotic attractors J Phys A Math
Gen 24 4587ndash97Rao R P and Ballard D H 1998 Predictive coding in the visual cortex a functional interpretation of some extra-classical
receptive field effects Nature Neurosci 2 79ndash87Robinson P A Wright J J and Rennie C R 1998 Synchronous oscillations in the cerebral cortex Phys Rev E 57
4578ndash88Rodriguez E George N Lachauz J P Martinerie J Renault B and Varela F J 1999 Perceptionrsquos shadow long distance
synchronization of human brain activity Nature 397 430ndash3Rosenblum M Pikovsky A and Kurths J 1996 Phase synchronization of chaotic oscillators Phys Rev Lett 76 1804ndash7Rosenblum M Pikovsky A and Kurths J 1997 From phase to lag synchronization in coupled chaotic oscillators Phys
Rev Lett 78 4193ndash6Rulkov N and Sushchik M 1997 Robustness of synchronized chaotic oscillations Int J Bifurc Chaos 7 625ndash43Rulkov N Sushchik M Tsimring L and Abarbanel H 1995 Generalized synchronization of chaos in unidirectionally
coupled chaotic systems Phys Rev E 51 980ndash94Schiff S So P Chang T Burke R and Sauer T 1996 Detecting dynamical interdependence and generalized synchrony
through mutual prediction in a neural ensemble Phys Rev E 54 6708ndash24Silberstein R B 1995 Neuromodulations of neocortical dynamics Neocortical Dynamics and Human EEG Rhythms
ed P L Nunez (Oxford Oxford University Press)Singer W 1995 Putative functions of temporal correlations in neocortical processing Large-Scale Neuronal Theories
of the Brain ed C Koch and J Davis (London MIT Press)Somers D and Kopell N 1995 Waves and synchrony in networks of oscillators of relaxation and non-relaxation type
Physica D 89 169ndash83Sutton R S and Barto A G 1990 Time derivative models of Pavlovian reinforcement Learning and Computational
Neuroscience Foundations of Adaptive Networks ed M Gabriel and J Moore (Cambridge MA MIT Press) pp497ndash538
Szentagothai J 1983 The modular architectonic principle of neural centers Rev Physiol Biochem Pharmacol 9811ndash61
Tallon-Baudry C Betrand O and Fischer C 2001 Oscillatory synchrony between human extrastriate areas during visualshort-term memory maintenance J Neurosci 21 RC177(1ndash5)
Tass P A 2002 Desynchronization of brain rhythms with soft phase-resetting techniques Biol Cybern 87 102ndash15
732 M Breakspear et al
Tass P Rosenblum M Weule J Kurths J Pikovsky A Volkmann J Schnitzler A and Freund H 1998 Detection of nmphase locking from noisy data application to magnetoencephalography Phys Rev Lett 81 3291ndash4
Terman D and Wang D 1995 Global competition and local cooperation in a network of neural oscillators Physica D81 148ndash76
Terry J and Breakspear M 2003 An improved algorithm for the detection of dynamical interdependence in bivariatetime-series Biol Cybern 88 129ndash36
Tononi G Sporns O and Edelman G M 1994 A measure for brain complexity relating functional segregation andintegration in the nervous system Proc Natl Acad Sci USA 91 5033ndash7
von der Malsburg C 1985 Nervous structures with dynamical links Phys Chem 89 703ndash10Wang X 1991 Period-doublings to chaos in a simple neural network an analytic proof Complex Syst 5 425ndash41Wennekers T and Pasemann F 1996 Synchronous chaos in high dimensional modular neural networks Int J Bifurc
Chaos 6 2055ndash67Zaks M A Parks E H Rosenblum M G and Kurths J 1999 Alternative locking ratios in imperfect phase synchronization
Phys Rev Lett 82 4228ndash31Zhao L and Macau E N 2001 A network of dynamically coupled chaotic maps for scene segmentation IEEE Trans
Neural Netw 12 1375ndash85
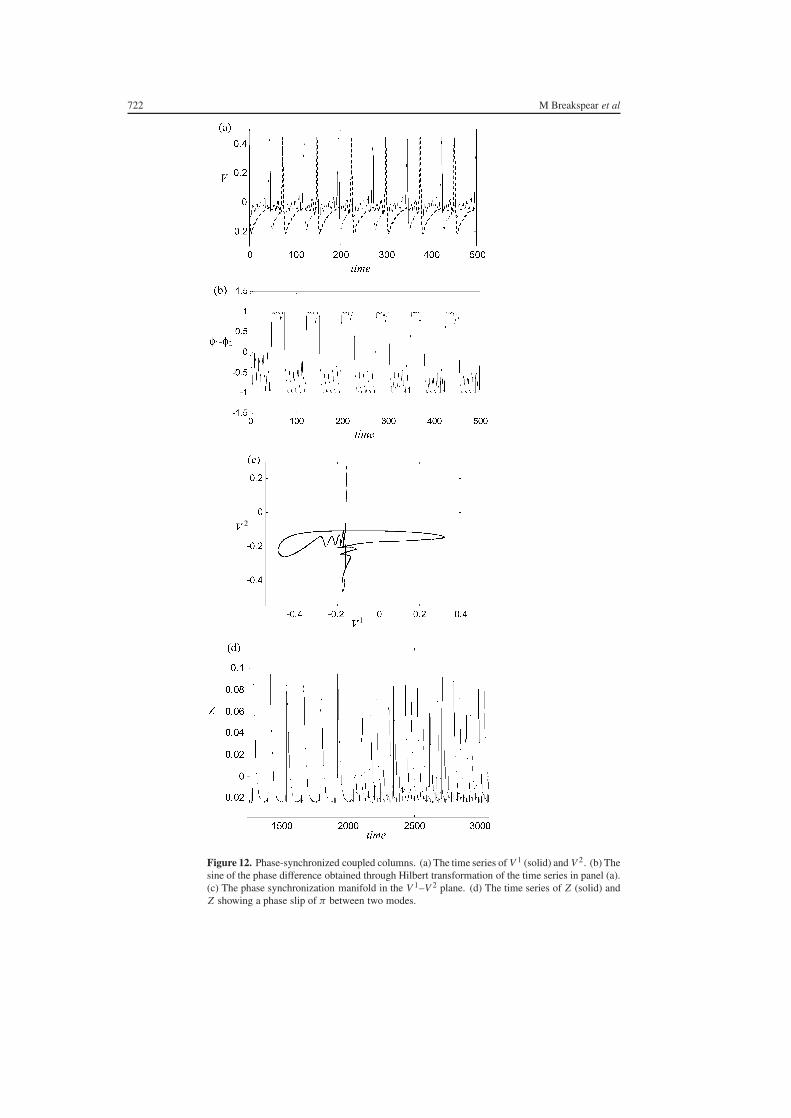
722 M Breakspear et al
2
1
Figure 12 Phase-synchronized coupled columns (a) The time series of V 1 (solid) and V 2 (b) Thesine of the phase difference obtained through Hilbert transformation of the time series in panel (a)(c) The phase synchronization manifold in the V 1ndashV 2 plane (d) The time series of Z (solid) andZ showing a phase slip of π between two modes
Synchronization in a model of neuronal dynamics 723
Figure 13 An example of phase desynchronization in coupled asymmetric systems with C = 01Time series plots of (a) each V -variable and (b) their Euclidean distance (c) The temporal evolutionof the phase difference between the two systems (d) An example of the phase difference evolutionover a longer time interval
724 M Breakspear et al
orderly evolution of the distance between the two systems and in panel (c) where the evolutionof the phase difference reveals a brief violation of the criterion |φ1 minus φ2| lt constant Anexample of the phase difference evolution over a longer time interval is shown in figure 13(d)Note that this is a three-dimensional plot with sin(φ1 minus φ2) and cos(φ1 minus φ2) as they- and z-axes Hence phase desynchronizations are evident as complete revolutions aroundthe cylindrical phase space in which this variable evolves It can be seen that these phenomenaoccur at irregular intervals in the temporal domain
This behaviour can be explained by considering the nature of a chaotic attractor In thestatistical sense an arbitrary initial condition is expected to lead to the chaotic behaviour onthe attractor Consequently when we calculate the Lyapunov exponents of the system theseexponents correspond to those of the chaotic attractor Typically however there are an infinitenumber of unstable periodic orbits embedded within the attractor If the path of a trajectorybecomes close to one of these orbits it may lsquoshadowrsquo the orbit for a brief period of time Eachof these orbits has a corresponding set of Lyapunov exponents which (like the attractor) can bedivided into tangential and transverse exponents These also vary when we change the couplingparameter but are typically not the same as the exponents of the attractor Hence the largesttransverse exponent of the attractor may be negative yet there may be positive transverseexponents associated with periodic orbits Considering all of these exponents together wewould therefore only expect to observe complete stability of the synchronization manifold inthe case where all of the transverse Lyapunov exponents are negative Bursting behaviourwill be observed when the one of these exponents becomes positive typically correspondingto a periodic orbit of low order (Pikovsky and Grassberger 1991 Rulkov and Sushchik 1997Maistrenko et al 1998 Heagy et al 1998) Due to the complex structure of such periodic orbitsthe desynchronous bursts occur across a wide range of timescales
In figure 14 we present the results of a long integration (16times105 time units) in which thelengths of sequential synchronous epochs of phase synchronization are studied The parametervalues were δV = 065 for column 1 and 0645 for column 2 with coupling strength10 equal to0125 In panel (a) the power spectrum of these sequential lengths is plotted The whiteness ofthis spectrum indicates that the lengths of consecutive synchronous epochs are independentThis is due to the high-dimensional lsquomixingrsquo effect of desynchronization In panel (b) thelengths of the synchronous epochs have been sorted again into the order of increasing lengthThe results for two coupling strengths are plotted C = 0125 (solid) and 0135 (dashed)Several features of this plot are of note Firstly the lengths of the synchronous epochsare distributed over almost three orders of magnitude Secondly for both plots there is anapproximately log-linear relationship between the length of an epoch and its relative orderThis reflects an approximately exponential relationship between the length of a synchronousepoch and its likelihood of occurrence Thirdly this relationship is skewed toward longersynchronous epochs for the stronger coupling strengths
This type of signal with patches of lsquoorderrsquo occurring over many timescales interruptedby bursts of disorder can be shown to have a high information capacity (Gatlin 1972)Moreover complex phase desynchronization may make an infrequent yet robust contributionto the dynamical pattern of the alpha rhythm in scalp EEG data for resting human subjects(Breakspear 2002) It is tempting to postulate that such a lsquoweakly stablersquo dynamics may assistin the generation of the rich and fluid mental associations that spontaneously emerge in restingcognitive states Although this phenomenon has been previously described in a variety of othersystems to our knowledge this is the first description in a biophysical model of neocortex
10 The introduction of parameter mismatch causes an increase in the strength of coupling required for stablesynchronization above the value of 01 required in the symmetric case
Synchronization in a model of neuronal dynamics 725
Figure 14 Properties of sequential epochs of phase-synchronous behaviour over a long integrationtime δ1
V = 065 δ2V = 0645 aee = 04 (a) The flat (white) power spectrum (b) Synchronous
epochs sorted in order of increasing length (solid C = 0125 dashed C = 0134)
52 Synchronization and complex dynamics in larger arrays
The synchronous and complex phenomena described above are also observed in arrays ofcoupled systems with N gt 2 Expanding the size of the array permits complex dynamicsin the spatial in addition to the temporal domain However the basic principles governingthe type and stability of synchronized motions are the same In this section a few of thepossible behaviours are discussed Physiological variation across the array is modelled bymodifying the parameter δV = 07 in each array by adding a random number of maximummagnitude 001
In figure 15(a) an array of N = 20 uncoupled (C = 0) systems is shown The lack ofsynchronization in this time series is evident By comparison the rapid transition to completesynchronization is evident in figure 15(b) when C = 03 In fact 19 of the 20 subsystems
726 M Breakspear et al
(b)
(c)
(a)
Figure 15 Synchronization phenomena in an array of coupled cortical columns (a) N = 20C = 0 (b) N = 20 C = 03 (c) N = 6 C = 01 (d) N = 12 C = 009 (e) N = 12C = 012 A small perturbation has been applied to one of the columns at t = 2500 s (f) Evolutionof the phase difference between two of the columns in panel (e) For clarity the W -variable isplotted in panel (c) and the Z -variable in (e)
have phase synchronized within one single cycle Intermediate values of coupling strength areassociated with a rich variety of phenomena For example subsystems with the most similarparameter values typically synchronize into clusters of various sizes An example of this isillustrated in figure 15(c) Note that there is no synchronization between the two differentclusters each containing three synchronized subsystems
Another complex behaviour occurs when subsystems transiently synchronize into largeclusters then spontaneously desynchronize again Hence the dynamics of the entire arrayjumps between different transient dynamics characterized by varying degrees of freedom Anexample of this phenomenon is given in figure 15(d) In addition it is possible to inducedesynchronization across an entire array by applying a small perturbation to one columnfor one time step An example is illustrated in figure 15(e) A small perturbation has been
Synchronization in a model of neuronal dynamics 727
Figure 15 (Continued)
added to one column after 2500 time units It can be seen that the entire array subsequentlydesynchronizes This illustrates the sensitivity of the dynamics to external stimuli Finally it ispossible to lsquoinducersquo synchronization between selected subsystems by increasing the strength ofcoupling between them selectively This allows an extra level of complexity to be introducedinto the model For example it may allow targeted information transfer between a partialnetwork of the entire array
As discussed in section 51 a univariate signal which jumps between epochs of lowentropy (high repetition) and bursts of high entropy (high variability) can be shown to havehigh information capacity This is because the repetition of certain patterns (as in the English
728 M Breakspear et al
language) permits error detectability Bursts of high entropy act in the opposite directionto increase message variability With a multivariate signal in addition to optimizing theinformation capacity within each signal the overall information capacity can be increasedthrough the type of itinerant synchronization briefly illustrated in this section Synchronizationacross an entire array increases the message reliability (since a single lsquoerrant letterrsquo in onesignal can be easily detected) Conversely desynchronization into multiple clusters increasesmessage variability since each cluster will produce independent information Hence a neuralarray which switches dynamically between a range of cluster configurations can be seen asadaptive in the sense that high synchronous states can be expressed when error detectabilityis a priority (such as during quick and reliable execution of a familiar task) but the array canrapidly desynchronize if variability becomes salient (for example if the environment suddenlybecomes novel) A formal operationalized demonstration of this principle is to be the subjectof future work
6 Discussion
In this paper we present a dynamical model for activity in an array of weakly coupledlocal neural assemblies The model incorporates the fundamental principles governing cellmembrane potential as determined by voltage- and ligand-gated ion channels In this wayit allows a reasonable amount of physiological complexity to be incorporated includingsynaptic activation and receptor subtypeswithout recourse to overwhelming detail Numericalanalysis of this model reveals a transition from independent oscillations to stable chaoticsynchronization depending on the balance of local versus long-range connectivity andbiophysical heterogeneity These excitatory couplings are subject to ascending subcorticalneuromodulation arising principally from monoaminergic nuclei such as Locus Coeruleus(Silberstein 1995) Neuromodulation of extrinsic connections was incorporated into the modelvia a coupling parameter C The stability of the synchronization is determined by the valueof the largest transverse Lyapunov exponent which in turn depends on the coupling parameterC If the coupling is weakened with the result that this exponent becomes positive a blowoutbifurcation occurs and independent oscillations re-emerge In the vicinity of the transitionnon-linear desynchronizations with complex temporal and spatial properties are observed
It is pleasing that complex and itinerant behaviour reminiscent of real neuronaldynamics emerges through such a simple manipulation of a single dynamic control parameterFurthermore the ubiquitous projections of classical neuromodulatory systems in real brainssuggest a clear biological mechanism by which the brain could self-organize throughmoderating the complexity of its own dynamics There are many reasons that the complicateddynamics of the sort demonstrated in this paper may be neurobiologically important Wemention four
First the presence of multiple clusters of phase-synchronized subsystems as discussedabove may be a uniquely non-linear mechanism facilitating the lsquocarriagersquo and distinction ofmultiple concurrent dynamic cell assemblies (and hence cognitive processing) This may beimportant for perceptual grouping binding and transitions as suggested by others (eg von derMalsburg 1985 Gray and Singer 1989)
Second the facility to decouple and reconfigure dynamic cell assemblies througha neuromodulatory control parameter may be important for avoiding low-entropysynchronization manifolds In the absence of itinerancy synchronization would renderneuronal dynamics incapable of representing changes in sensory input or indeed cognitive stateFor example in the model it was observed that small perturbations to one column were ableto induce a global change in neuronal dynamics by disrupting a state of weak synchronization
Synchronization in a model of neuronal dynamics 729
Inevitably alternative dynamic configurations emerge epitomizing the importance of lsquolabilersquobrain dynamics (Friston 2000a)
Thirdly from a purely functionalist point of view the balance between functionalspecialization and functional integration may rely upon the sorts of mechanism modelledin this paper The brain appears to adhere to two principles of functional organization namelysegregation of function within cortical areas and sub-areas and the integration of these areasin perceptual synthesis cognition and sensorimotor integration (Tononi et al 1994 Fristonet al 1995) The balance between preserving specialized dynamics unique to each arearequired by specialization and the coupling among these areas mediating functional integrationmay be achieved by the neuromodulatory mechanisms described above Significantlythe spontaneous variation in the numbers of degrees of freedom (for example whenlow-dimensional synchronization manifolds are shadowed) imparts a dynamic characteristicto this perspective that is precluded by linear mechanisms alone
Finally from a mechanistic perspective complexity and diversity of brain dynamics arerequired for selection of adaptive responses (Friston 2000b) There are many examples ofselective mechanisms operating within the brain All can be seen as performing an ascenton some objective function or fitness landscape These mechanisms require complex anditinerant dynamics to facilitate a thorough exploration of the landscape These landscapesmay range from those involved in perception (where the landscape is defined by predictionerror see eg Rao and Ballard (1998)) to those involved in value learning (where thelandscape is determined by the outcome of a behaviour eg in temporal difference models (seeeg Sutton and Barto 1990)) In short complexity is necessary to maintain an information-rich environment in which the brain can respond to sensory inputs An essential componentof this is the maintenance of diversity which is a prerequisite for selectionist mechanismsat many scales (Edelman 1993) Understanding the mechanistic basis of complex dynamicsas measured electrophysiologically may well depend on the sort of biologically plausiblemodels used in this paper This understanding may be important for both basic and clinicalneuroscience (eg Tass 2002)
The present study supplements and advances on other research in this field The studyof complex phase synchronization and clustering in neural oscillator networks similar tothe present model has previously been the subject of formal mathematical study Forexample Terman and Wang (1995) studied synchronization of blocks of neural oscillatorsthat were subject to perturbation by connected lsquostimulirsquo Somers and Kopell (1995) studiedthe emergence of both synchronization and travelling waves in networks of coupled neuraloscillators In the work of Terman and Wang the local dynamics were based on neurallsquocaricaturersquo models such as the FitzHughndashNagumo equations and are hence more abstract thanthe current study The approach of Somers and Kopell only required that the local dynamicsexhibit fast and relaxation phases and is hence more abstract still In both studies onlyperiodic dynamics are studied Complex patterns of chaos synchronization in neural arrayshave been characterized by nonlinear methods (Kaneko 1998) and their putative contributionto both learning (Nakajima and Ueda 1996) and perception (Zhao and Macau 2001 Breakspear2001) explored However these studies employed models such as the logistic equation andtent map which are quite abstract in comparison to the present study On the other handHansel and Sompolinsky (1996) studied the role of chaos synchronization in a model ofvisual perception in a cortical hypercolumn using a very similar approach to that adoptedhere but with the additional complexity that synaptic delays were incorporated However inthis and earlier work (Hansel and Sompolinsky 1992) only linear measures of dynamics andsynchrony (specifically the auto- and cross-correlation functions) were employed The analysisof chaotic synchrony in the study of Larter et al (1999) was based only upon observation of
730 M Breakspear et al
the Euclidean difference between variables from two nodes within a larger array Hence thesignificance of the present work is that it provides a quantitative nonlinear analysis of complexchaos synchronization in a biophysical model of cortical dynamics Such research is requiredin order to bridge the gap between quantitative nonlinear research into synchronization andcomputation in abstract models of neural arrays and qualitative or linear analysis in morebiophysically plausible models
Acknowledgments
The authors would like to thank Peter Robinson for assistance with the manuscript and R Larterfor helpful advice MB is a recipient of a NSW Institute of Psychiatry Research Grant anda University of Sydney SESQUI postdoctoral fellowship KJF is supported by the WellcomeTrust
References
Abarbanel H Rulkov N and Sushchik M 1996 Generalised synchronization of chaos the auxiliary system approachPhys Rev E 53 4528ndash35
Afraimovich V Verichev N and Rabinovich M 1986 Stochastic synchronization of oscillation in dissipative systemsRadiophys Quantum Electron 29 795ndash801
Breakspear M 2001 Perception of odours by a nonlinear model of the olfactory bulb Int J Neural Syst 11 101ndash24Breakspear M 2002 Nonlinear phase desynchronisation in human electroencephalographic data Human Brain Mapping
15 175ndash98Breakspear M and Terry J R 2002a Nonlinear interdependence in neural systems motivation theory and relevance
Int J Neurosci 112 1163ndash84Breakspear M and Terry J R 2002b Detection and description of nonlinear interdependence in normal multichannel
human EEG Clin Neurophysiol 113 735ndash53Breakspear M and Terry J R 2002c Topographic organization of nonlinear interdependence in multichannel human
EEG Neuroimage 16 822ndash35Destexhe A Rudolph M Fellous J M and Sejnowski T J 2001 Fluctuating synaptic conductances recreate in vivo-like
activity in neocortical neurons Neuroscience 107 13ndash24Douglas R J Mahowald M A and Martin K A 1996 Microarchitecture of neocortical columns Brain TheorymdashBiological
Basis and Computational Principles ed A Aertsen and V Braitenberg (Amsterdam Elsevier)Eckhorn R Bauer R Jordan W Brosch M Kruse W Munk M and Reitboeck H J 1988 Coherent oscillations a
mechanism of feature linking in the visual cortex Multiple electrode and correlation analyses in the cat BiolCybern 60 121ndash30
Edelman G M 1993 Neural Darwinism selection and reentrant signalling in higher brain function Neuron 10 115ndash25Engel A K Kreiter A K Konig P and Singer W 1991 Synchronization of oscillatory neuronal responses between
striate and extrastriate visual cortical areas of the cat Proc Natl Acad Sci USA 88 6048ndash52Friston K J 2000a The labile brain I Neuronal transients and nonlinear coupling Phil Trans R Soc B 355 215ndash36Friston K J 2000b The labile brain II Transients complexity and selection Phil Trans R Soc B 355 237ndash52Friston K J Tononi G Sporns O and Edelman G 1995 Characterising the complexity of neuronal interactions Human
Brain Mapping 3 302ndash14Fujisaka H and Yamada T 1983 Stability theory of synchronized motion in coupled-oscillator system Prog Theor
Phys 69 32ndash47Gatlin L 1972 Information Theory and the Living System (New York Columbia University Press)Gray C M Konig P Engel A and Singer W 1989 Oscillatory responses in cat visual cortex exhibit inter-columnar
synchronization which reflects global stimulus properties Science 338 334ndash7Gray C M and Singer W 1989 Stimulus specific neuronal oscillations in orientation columns of cat visual cortex Proc
Natl Acad Sci USA 86 1698ndash702Haig A R Gordon E Wright J J Meares R A and Bahramali H 2000 Synchronous cortical gamma-band activity in
task-relevant cognition Neuroreport 11 669ndash75Hansel D and Sompolinsky H 1992 Synchronization and computation in a chaotic neural network Phys Rev Lett 68
718ndash21
Synchronization in a model of neuronal dynamics 731
Hansel D and Sompolinsky H 1996 Chaos and synchrony in a model of a hypercolumn in visual cortex J ComputNeurosci 3 7ndash4
Heagy J Carroll T and Pecora L 1998 Desynchronization by periodic orbits Phys Rev E 57 R1253ndash6Hunt B Ott E and Yorke J 1997 Differentiable synchronization of chaos Phys Rev E 55 4029ndash34Josic K 1998 Invariant manifolds and synchronization of coupled dynamical systems Phys Rev Lett 80 3053ndash6Kandel E Schwartz J and Jessell T 2000 Principles of Neural Science (New York McGraw-Hill)Kaneko K 1998 On the strength of attractors in a high-dimensional system Physica D 124 322ndash44Kocarev L Parlitz U and Brown R 2000 Robust synchronization of chaotic systems Phys Rev E 61 3716ndash20Larter R Speelman B and Worth R M 1999 A coupled ordinary differential equation lattice model for the simulation
of epileptic seizures Chaos 9 795ndash804Lumer E Edelman G and Tononi G 1997a Neural dynamics in a model of the thalamocortical system I Layers loops
and the emergence of fast synchronous oscillations Cereb Cortex 7 207ndash27Lumer E Edelman G and Tononi G 1997b Neural dynamics in a model of the thalamocortical system II The role of
neural synchrony tested through perturbations of spike timing Cereb Cortex 7 228ndash36Magleby K 1987 Short-term modulation at synapses Synaptic Function ed G M Edelman W E Gall and
W M Cowan (New York Wiley)Maistrenko Y Maistrenko V Popovich A and Mosekilde E 1998 Transverse instability and riddled basins in a system
of two coupled logistic maps Phys Rev E 57 2713ndash24Morris C and Lecar H 1981 Voltage oscillations in the barnacle giant muscle fiber Biophys J 35 193ndash213Mountcastle V B 1997 The columnar organization of the neocortex Brain 120 701ndash22Mountcastle V B 1998 The Cerebral Cortex (Cambridge MA Harvard University Press)Nakajima H and Ueda Y 1996 Riddled basins of the optimal states in learning dynamical systems Physica D 99 35ndash44Nunez P L 1995 Neocortical Dynamics and Human Brain Rhythms (Oxford Oxford University Press)Osipov G V Pikovsky A S Rosenblum M G and Kurths J 1997 Phase synchronization effects in a lattice of nonidentical
Rossler oscillators Phys Rev E 55 2353ndash61Ott E and Sommerer J 1994 Blowout bifurcations the occurrence of riddled basins and onndashoff intermittency Phys
Lett A 188 39ndash47Pecora L and Carroll T 1990 Synchronization in chaotic systems Phys Rev Lett 64 821ndash4Pikovsky A and Grassberger P 1991 Symmetry breaking bifurcation for coupled chaotic attractors J Phys A Math
Gen 24 4587ndash97Rao R P and Ballard D H 1998 Predictive coding in the visual cortex a functional interpretation of some extra-classical
receptive field effects Nature Neurosci 2 79ndash87Robinson P A Wright J J and Rennie C R 1998 Synchronous oscillations in the cerebral cortex Phys Rev E 57
4578ndash88Rodriguez E George N Lachauz J P Martinerie J Renault B and Varela F J 1999 Perceptionrsquos shadow long distance
synchronization of human brain activity Nature 397 430ndash3Rosenblum M Pikovsky A and Kurths J 1996 Phase synchronization of chaotic oscillators Phys Rev Lett 76 1804ndash7Rosenblum M Pikovsky A and Kurths J 1997 From phase to lag synchronization in coupled chaotic oscillators Phys
Rev Lett 78 4193ndash6Rulkov N and Sushchik M 1997 Robustness of synchronized chaotic oscillations Int J Bifurc Chaos 7 625ndash43Rulkov N Sushchik M Tsimring L and Abarbanel H 1995 Generalized synchronization of chaos in unidirectionally
coupled chaotic systems Phys Rev E 51 980ndash94Schiff S So P Chang T Burke R and Sauer T 1996 Detecting dynamical interdependence and generalized synchrony
through mutual prediction in a neural ensemble Phys Rev E 54 6708ndash24Silberstein R B 1995 Neuromodulations of neocortical dynamics Neocortical Dynamics and Human EEG Rhythms
ed P L Nunez (Oxford Oxford University Press)Singer W 1995 Putative functions of temporal correlations in neocortical processing Large-Scale Neuronal Theories
of the Brain ed C Koch and J Davis (London MIT Press)Somers D and Kopell N 1995 Waves and synchrony in networks of oscillators of relaxation and non-relaxation type
Physica D 89 169ndash83Sutton R S and Barto A G 1990 Time derivative models of Pavlovian reinforcement Learning and Computational
Neuroscience Foundations of Adaptive Networks ed M Gabriel and J Moore (Cambridge MA MIT Press) pp497ndash538
Szentagothai J 1983 The modular architectonic principle of neural centers Rev Physiol Biochem Pharmacol 9811ndash61
Tallon-Baudry C Betrand O and Fischer C 2001 Oscillatory synchrony between human extrastriate areas during visualshort-term memory maintenance J Neurosci 21 RC177(1ndash5)
Tass P A 2002 Desynchronization of brain rhythms with soft phase-resetting techniques Biol Cybern 87 102ndash15
732 M Breakspear et al
Tass P Rosenblum M Weule J Kurths J Pikovsky A Volkmann J Schnitzler A and Freund H 1998 Detection of nmphase locking from noisy data application to magnetoencephalography Phys Rev Lett 81 3291ndash4
Terman D and Wang D 1995 Global competition and local cooperation in a network of neural oscillators Physica D81 148ndash76
Terry J and Breakspear M 2003 An improved algorithm for the detection of dynamical interdependence in bivariatetime-series Biol Cybern 88 129ndash36
Tononi G Sporns O and Edelman G M 1994 A measure for brain complexity relating functional segregation andintegration in the nervous system Proc Natl Acad Sci USA 91 5033ndash7
von der Malsburg C 1985 Nervous structures with dynamical links Phys Chem 89 703ndash10Wang X 1991 Period-doublings to chaos in a simple neural network an analytic proof Complex Syst 5 425ndash41Wennekers T and Pasemann F 1996 Synchronous chaos in high dimensional modular neural networks Int J Bifurc
Chaos 6 2055ndash67Zaks M A Parks E H Rosenblum M G and Kurths J 1999 Alternative locking ratios in imperfect phase synchronization
Phys Rev Lett 82 4228ndash31Zhao L and Macau E N 2001 A network of dynamically coupled chaotic maps for scene segmentation IEEE Trans
Neural Netw 12 1375ndash85
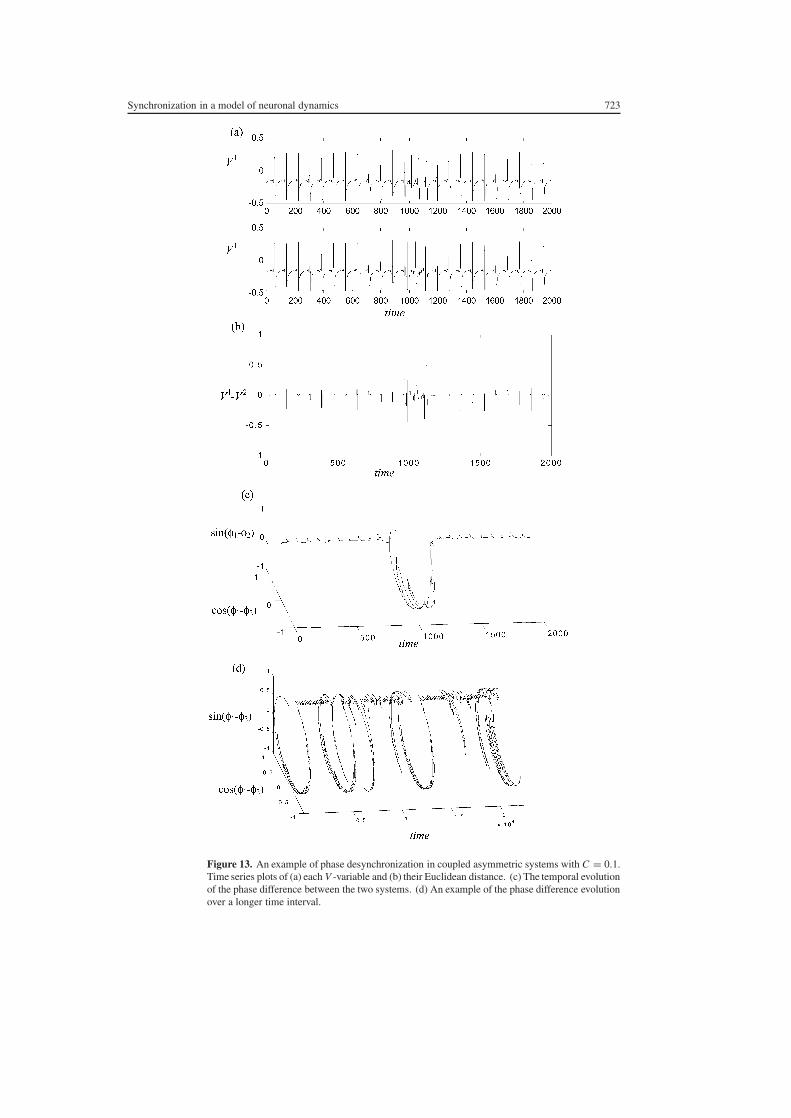
Synchronization in a model of neuronal dynamics 723
Figure 13 An example of phase desynchronization in coupled asymmetric systems with C = 01Time series plots of (a) each V -variable and (b) their Euclidean distance (c) The temporal evolutionof the phase difference between the two systems (d) An example of the phase difference evolutionover a longer time interval
724 M Breakspear et al
orderly evolution of the distance between the two systems and in panel (c) where the evolutionof the phase difference reveals a brief violation of the criterion |φ1 minus φ2| lt constant Anexample of the phase difference evolution over a longer time interval is shown in figure 13(d)Note that this is a three-dimensional plot with sin(φ1 minus φ2) and cos(φ1 minus φ2) as they- and z-axes Hence phase desynchronizations are evident as complete revolutions aroundthe cylindrical phase space in which this variable evolves It can be seen that these phenomenaoccur at irregular intervals in the temporal domain
This behaviour can be explained by considering the nature of a chaotic attractor In thestatistical sense an arbitrary initial condition is expected to lead to the chaotic behaviour onthe attractor Consequently when we calculate the Lyapunov exponents of the system theseexponents correspond to those of the chaotic attractor Typically however there are an infinitenumber of unstable periodic orbits embedded within the attractor If the path of a trajectorybecomes close to one of these orbits it may lsquoshadowrsquo the orbit for a brief period of time Eachof these orbits has a corresponding set of Lyapunov exponents which (like the attractor) can bedivided into tangential and transverse exponents These also vary when we change the couplingparameter but are typically not the same as the exponents of the attractor Hence the largesttransverse exponent of the attractor may be negative yet there may be positive transverseexponents associated with periodic orbits Considering all of these exponents together wewould therefore only expect to observe complete stability of the synchronization manifold inthe case where all of the transverse Lyapunov exponents are negative Bursting behaviourwill be observed when the one of these exponents becomes positive typically correspondingto a periodic orbit of low order (Pikovsky and Grassberger 1991 Rulkov and Sushchik 1997Maistrenko et al 1998 Heagy et al 1998) Due to the complex structure of such periodic orbitsthe desynchronous bursts occur across a wide range of timescales
In figure 14 we present the results of a long integration (16times105 time units) in which thelengths of sequential synchronous epochs of phase synchronization are studied The parametervalues were δV = 065 for column 1 and 0645 for column 2 with coupling strength10 equal to0125 In panel (a) the power spectrum of these sequential lengths is plotted The whiteness ofthis spectrum indicates that the lengths of consecutive synchronous epochs are independentThis is due to the high-dimensional lsquomixingrsquo effect of desynchronization In panel (b) thelengths of the synchronous epochs have been sorted again into the order of increasing lengthThe results for two coupling strengths are plotted C = 0125 (solid) and 0135 (dashed)Several features of this plot are of note Firstly the lengths of the synchronous epochsare distributed over almost three orders of magnitude Secondly for both plots there is anapproximately log-linear relationship between the length of an epoch and its relative orderThis reflects an approximately exponential relationship between the length of a synchronousepoch and its likelihood of occurrence Thirdly this relationship is skewed toward longersynchronous epochs for the stronger coupling strengths
This type of signal with patches of lsquoorderrsquo occurring over many timescales interruptedby bursts of disorder can be shown to have a high information capacity (Gatlin 1972)Moreover complex phase desynchronization may make an infrequent yet robust contributionto the dynamical pattern of the alpha rhythm in scalp EEG data for resting human subjects(Breakspear 2002) It is tempting to postulate that such a lsquoweakly stablersquo dynamics may assistin the generation of the rich and fluid mental associations that spontaneously emerge in restingcognitive states Although this phenomenon has been previously described in a variety of othersystems to our knowledge this is the first description in a biophysical model of neocortex
10 The introduction of parameter mismatch causes an increase in the strength of coupling required for stablesynchronization above the value of 01 required in the symmetric case
Synchronization in a model of neuronal dynamics 725
Figure 14 Properties of sequential epochs of phase-synchronous behaviour over a long integrationtime δ1
V = 065 δ2V = 0645 aee = 04 (a) The flat (white) power spectrum (b) Synchronous
epochs sorted in order of increasing length (solid C = 0125 dashed C = 0134)
52 Synchronization and complex dynamics in larger arrays
The synchronous and complex phenomena described above are also observed in arrays ofcoupled systems with N gt 2 Expanding the size of the array permits complex dynamicsin the spatial in addition to the temporal domain However the basic principles governingthe type and stability of synchronized motions are the same In this section a few of thepossible behaviours are discussed Physiological variation across the array is modelled bymodifying the parameter δV = 07 in each array by adding a random number of maximummagnitude 001
In figure 15(a) an array of N = 20 uncoupled (C = 0) systems is shown The lack ofsynchronization in this time series is evident By comparison the rapid transition to completesynchronization is evident in figure 15(b) when C = 03 In fact 19 of the 20 subsystems
726 M Breakspear et al
(b)
(c)
(a)
Figure 15 Synchronization phenomena in an array of coupled cortical columns (a) N = 20C = 0 (b) N = 20 C = 03 (c) N = 6 C = 01 (d) N = 12 C = 009 (e) N = 12C = 012 A small perturbation has been applied to one of the columns at t = 2500 s (f) Evolutionof the phase difference between two of the columns in panel (e) For clarity the W -variable isplotted in panel (c) and the Z -variable in (e)
have phase synchronized within one single cycle Intermediate values of coupling strength areassociated with a rich variety of phenomena For example subsystems with the most similarparameter values typically synchronize into clusters of various sizes An example of this isillustrated in figure 15(c) Note that there is no synchronization between the two differentclusters each containing three synchronized subsystems
Another complex behaviour occurs when subsystems transiently synchronize into largeclusters then spontaneously desynchronize again Hence the dynamics of the entire arrayjumps between different transient dynamics characterized by varying degrees of freedom Anexample of this phenomenon is given in figure 15(d) In addition it is possible to inducedesynchronization across an entire array by applying a small perturbation to one columnfor one time step An example is illustrated in figure 15(e) A small perturbation has been
Synchronization in a model of neuronal dynamics 727
Figure 15 (Continued)
added to one column after 2500 time units It can be seen that the entire array subsequentlydesynchronizes This illustrates the sensitivity of the dynamics to external stimuli Finally it ispossible to lsquoinducersquo synchronization between selected subsystems by increasing the strength ofcoupling between them selectively This allows an extra level of complexity to be introducedinto the model For example it may allow targeted information transfer between a partialnetwork of the entire array
As discussed in section 51 a univariate signal which jumps between epochs of lowentropy (high repetition) and bursts of high entropy (high variability) can be shown to havehigh information capacity This is because the repetition of certain patterns (as in the English
728 M Breakspear et al
language) permits error detectability Bursts of high entropy act in the opposite directionto increase message variability With a multivariate signal in addition to optimizing theinformation capacity within each signal the overall information capacity can be increasedthrough the type of itinerant synchronization briefly illustrated in this section Synchronizationacross an entire array increases the message reliability (since a single lsquoerrant letterrsquo in onesignal can be easily detected) Conversely desynchronization into multiple clusters increasesmessage variability since each cluster will produce independent information Hence a neuralarray which switches dynamically between a range of cluster configurations can be seen asadaptive in the sense that high synchronous states can be expressed when error detectabilityis a priority (such as during quick and reliable execution of a familiar task) but the array canrapidly desynchronize if variability becomes salient (for example if the environment suddenlybecomes novel) A formal operationalized demonstration of this principle is to be the subjectof future work
6 Discussion
In this paper we present a dynamical model for activity in an array of weakly coupledlocal neural assemblies The model incorporates the fundamental principles governing cellmembrane potential as determined by voltage- and ligand-gated ion channels In this wayit allows a reasonable amount of physiological complexity to be incorporated includingsynaptic activation and receptor subtypeswithout recourse to overwhelming detail Numericalanalysis of this model reveals a transition from independent oscillations to stable chaoticsynchronization depending on the balance of local versus long-range connectivity andbiophysical heterogeneity These excitatory couplings are subject to ascending subcorticalneuromodulation arising principally from monoaminergic nuclei such as Locus Coeruleus(Silberstein 1995) Neuromodulation of extrinsic connections was incorporated into the modelvia a coupling parameter C The stability of the synchronization is determined by the valueof the largest transverse Lyapunov exponent which in turn depends on the coupling parameterC If the coupling is weakened with the result that this exponent becomes positive a blowoutbifurcation occurs and independent oscillations re-emerge In the vicinity of the transitionnon-linear desynchronizations with complex temporal and spatial properties are observed
It is pleasing that complex and itinerant behaviour reminiscent of real neuronaldynamics emerges through such a simple manipulation of a single dynamic control parameterFurthermore the ubiquitous projections of classical neuromodulatory systems in real brainssuggest a clear biological mechanism by which the brain could self-organize throughmoderating the complexity of its own dynamics There are many reasons that the complicateddynamics of the sort demonstrated in this paper may be neurobiologically important Wemention four
First the presence of multiple clusters of phase-synchronized subsystems as discussedabove may be a uniquely non-linear mechanism facilitating the lsquocarriagersquo and distinction ofmultiple concurrent dynamic cell assemblies (and hence cognitive processing) This may beimportant for perceptual grouping binding and transitions as suggested by others (eg von derMalsburg 1985 Gray and Singer 1989)
Second the facility to decouple and reconfigure dynamic cell assemblies througha neuromodulatory control parameter may be important for avoiding low-entropysynchronization manifolds In the absence of itinerancy synchronization would renderneuronal dynamics incapable of representing changes in sensory input or indeed cognitive stateFor example in the model it was observed that small perturbations to one column were ableto induce a global change in neuronal dynamics by disrupting a state of weak synchronization
Synchronization in a model of neuronal dynamics 729
Inevitably alternative dynamic configurations emerge epitomizing the importance of lsquolabilersquobrain dynamics (Friston 2000a)
Thirdly from a purely functionalist point of view the balance between functionalspecialization and functional integration may rely upon the sorts of mechanism modelledin this paper The brain appears to adhere to two principles of functional organization namelysegregation of function within cortical areas and sub-areas and the integration of these areasin perceptual synthesis cognition and sensorimotor integration (Tononi et al 1994 Fristonet al 1995) The balance between preserving specialized dynamics unique to each arearequired by specialization and the coupling among these areas mediating functional integrationmay be achieved by the neuromodulatory mechanisms described above Significantlythe spontaneous variation in the numbers of degrees of freedom (for example whenlow-dimensional synchronization manifolds are shadowed) imparts a dynamic characteristicto this perspective that is precluded by linear mechanisms alone
Finally from a mechanistic perspective complexity and diversity of brain dynamics arerequired for selection of adaptive responses (Friston 2000b) There are many examples ofselective mechanisms operating within the brain All can be seen as performing an ascenton some objective function or fitness landscape These mechanisms require complex anditinerant dynamics to facilitate a thorough exploration of the landscape These landscapesmay range from those involved in perception (where the landscape is defined by predictionerror see eg Rao and Ballard (1998)) to those involved in value learning (where thelandscape is determined by the outcome of a behaviour eg in temporal difference models (seeeg Sutton and Barto 1990)) In short complexity is necessary to maintain an information-rich environment in which the brain can respond to sensory inputs An essential componentof this is the maintenance of diversity which is a prerequisite for selectionist mechanismsat many scales (Edelman 1993) Understanding the mechanistic basis of complex dynamicsas measured electrophysiologically may well depend on the sort of biologically plausiblemodels used in this paper This understanding may be important for both basic and clinicalneuroscience (eg Tass 2002)
The present study supplements and advances on other research in this field The studyof complex phase synchronization and clustering in neural oscillator networks similar tothe present model has previously been the subject of formal mathematical study Forexample Terman and Wang (1995) studied synchronization of blocks of neural oscillatorsthat were subject to perturbation by connected lsquostimulirsquo Somers and Kopell (1995) studiedthe emergence of both synchronization and travelling waves in networks of coupled neuraloscillators In the work of Terman and Wang the local dynamics were based on neurallsquocaricaturersquo models such as the FitzHughndashNagumo equations and are hence more abstract thanthe current study The approach of Somers and Kopell only required that the local dynamicsexhibit fast and relaxation phases and is hence more abstract still In both studies onlyperiodic dynamics are studied Complex patterns of chaos synchronization in neural arrayshave been characterized by nonlinear methods (Kaneko 1998) and their putative contributionto both learning (Nakajima and Ueda 1996) and perception (Zhao and Macau 2001 Breakspear2001) explored However these studies employed models such as the logistic equation andtent map which are quite abstract in comparison to the present study On the other handHansel and Sompolinsky (1996) studied the role of chaos synchronization in a model ofvisual perception in a cortical hypercolumn using a very similar approach to that adoptedhere but with the additional complexity that synaptic delays were incorporated However inthis and earlier work (Hansel and Sompolinsky 1992) only linear measures of dynamics andsynchrony (specifically the auto- and cross-correlation functions) were employed The analysisof chaotic synchrony in the study of Larter et al (1999) was based only upon observation of
730 M Breakspear et al
the Euclidean difference between variables from two nodes within a larger array Hence thesignificance of the present work is that it provides a quantitative nonlinear analysis of complexchaos synchronization in a biophysical model of cortical dynamics Such research is requiredin order to bridge the gap between quantitative nonlinear research into synchronization andcomputation in abstract models of neural arrays and qualitative or linear analysis in morebiophysically plausible models
Acknowledgments
The authors would like to thank Peter Robinson for assistance with the manuscript and R Larterfor helpful advice MB is a recipient of a NSW Institute of Psychiatry Research Grant anda University of Sydney SESQUI postdoctoral fellowship KJF is supported by the WellcomeTrust
References
Abarbanel H Rulkov N and Sushchik M 1996 Generalised synchronization of chaos the auxiliary system approachPhys Rev E 53 4528ndash35
Afraimovich V Verichev N and Rabinovich M 1986 Stochastic synchronization of oscillation in dissipative systemsRadiophys Quantum Electron 29 795ndash801
Breakspear M 2001 Perception of odours by a nonlinear model of the olfactory bulb Int J Neural Syst 11 101ndash24Breakspear M 2002 Nonlinear phase desynchronisation in human electroencephalographic data Human Brain Mapping
15 175ndash98Breakspear M and Terry J R 2002a Nonlinear interdependence in neural systems motivation theory and relevance
Int J Neurosci 112 1163ndash84Breakspear M and Terry J R 2002b Detection and description of nonlinear interdependence in normal multichannel
human EEG Clin Neurophysiol 113 735ndash53Breakspear M and Terry J R 2002c Topographic organization of nonlinear interdependence in multichannel human
EEG Neuroimage 16 822ndash35Destexhe A Rudolph M Fellous J M and Sejnowski T J 2001 Fluctuating synaptic conductances recreate in vivo-like
activity in neocortical neurons Neuroscience 107 13ndash24Douglas R J Mahowald M A and Martin K A 1996 Microarchitecture of neocortical columns Brain TheorymdashBiological
Basis and Computational Principles ed A Aertsen and V Braitenberg (Amsterdam Elsevier)Eckhorn R Bauer R Jordan W Brosch M Kruse W Munk M and Reitboeck H J 1988 Coherent oscillations a
mechanism of feature linking in the visual cortex Multiple electrode and correlation analyses in the cat BiolCybern 60 121ndash30
Edelman G M 1993 Neural Darwinism selection and reentrant signalling in higher brain function Neuron 10 115ndash25Engel A K Kreiter A K Konig P and Singer W 1991 Synchronization of oscillatory neuronal responses between
striate and extrastriate visual cortical areas of the cat Proc Natl Acad Sci USA 88 6048ndash52Friston K J 2000a The labile brain I Neuronal transients and nonlinear coupling Phil Trans R Soc B 355 215ndash36Friston K J 2000b The labile brain II Transients complexity and selection Phil Trans R Soc B 355 237ndash52Friston K J Tononi G Sporns O and Edelman G 1995 Characterising the complexity of neuronal interactions Human
Brain Mapping 3 302ndash14Fujisaka H and Yamada T 1983 Stability theory of synchronized motion in coupled-oscillator system Prog Theor
Phys 69 32ndash47Gatlin L 1972 Information Theory and the Living System (New York Columbia University Press)Gray C M Konig P Engel A and Singer W 1989 Oscillatory responses in cat visual cortex exhibit inter-columnar
synchronization which reflects global stimulus properties Science 338 334ndash7Gray C M and Singer W 1989 Stimulus specific neuronal oscillations in orientation columns of cat visual cortex Proc
Natl Acad Sci USA 86 1698ndash702Haig A R Gordon E Wright J J Meares R A and Bahramali H 2000 Synchronous cortical gamma-band activity in
task-relevant cognition Neuroreport 11 669ndash75Hansel D and Sompolinsky H 1992 Synchronization and computation in a chaotic neural network Phys Rev Lett 68
718ndash21
Synchronization in a model of neuronal dynamics 731
Hansel D and Sompolinsky H 1996 Chaos and synchrony in a model of a hypercolumn in visual cortex J ComputNeurosci 3 7ndash4
Heagy J Carroll T and Pecora L 1998 Desynchronization by periodic orbits Phys Rev E 57 R1253ndash6Hunt B Ott E and Yorke J 1997 Differentiable synchronization of chaos Phys Rev E 55 4029ndash34Josic K 1998 Invariant manifolds and synchronization of coupled dynamical systems Phys Rev Lett 80 3053ndash6Kandel E Schwartz J and Jessell T 2000 Principles of Neural Science (New York McGraw-Hill)Kaneko K 1998 On the strength of attractors in a high-dimensional system Physica D 124 322ndash44Kocarev L Parlitz U and Brown R 2000 Robust synchronization of chaotic systems Phys Rev E 61 3716ndash20Larter R Speelman B and Worth R M 1999 A coupled ordinary differential equation lattice model for the simulation
of epileptic seizures Chaos 9 795ndash804Lumer E Edelman G and Tononi G 1997a Neural dynamics in a model of the thalamocortical system I Layers loops
and the emergence of fast synchronous oscillations Cereb Cortex 7 207ndash27Lumer E Edelman G and Tononi G 1997b Neural dynamics in a model of the thalamocortical system II The role of
neural synchrony tested through perturbations of spike timing Cereb Cortex 7 228ndash36Magleby K 1987 Short-term modulation at synapses Synaptic Function ed G M Edelman W E Gall and
W M Cowan (New York Wiley)Maistrenko Y Maistrenko V Popovich A and Mosekilde E 1998 Transverse instability and riddled basins in a system
of two coupled logistic maps Phys Rev E 57 2713ndash24Morris C and Lecar H 1981 Voltage oscillations in the barnacle giant muscle fiber Biophys J 35 193ndash213Mountcastle V B 1997 The columnar organization of the neocortex Brain 120 701ndash22Mountcastle V B 1998 The Cerebral Cortex (Cambridge MA Harvard University Press)Nakajima H and Ueda Y 1996 Riddled basins of the optimal states in learning dynamical systems Physica D 99 35ndash44Nunez P L 1995 Neocortical Dynamics and Human Brain Rhythms (Oxford Oxford University Press)Osipov G V Pikovsky A S Rosenblum M G and Kurths J 1997 Phase synchronization effects in a lattice of nonidentical
Rossler oscillators Phys Rev E 55 2353ndash61Ott E and Sommerer J 1994 Blowout bifurcations the occurrence of riddled basins and onndashoff intermittency Phys
Lett A 188 39ndash47Pecora L and Carroll T 1990 Synchronization in chaotic systems Phys Rev Lett 64 821ndash4Pikovsky A and Grassberger P 1991 Symmetry breaking bifurcation for coupled chaotic attractors J Phys A Math
Gen 24 4587ndash97Rao R P and Ballard D H 1998 Predictive coding in the visual cortex a functional interpretation of some extra-classical
receptive field effects Nature Neurosci 2 79ndash87Robinson P A Wright J J and Rennie C R 1998 Synchronous oscillations in the cerebral cortex Phys Rev E 57
4578ndash88Rodriguez E George N Lachauz J P Martinerie J Renault B and Varela F J 1999 Perceptionrsquos shadow long distance
synchronization of human brain activity Nature 397 430ndash3Rosenblum M Pikovsky A and Kurths J 1996 Phase synchronization of chaotic oscillators Phys Rev Lett 76 1804ndash7Rosenblum M Pikovsky A and Kurths J 1997 From phase to lag synchronization in coupled chaotic oscillators Phys
Rev Lett 78 4193ndash6Rulkov N and Sushchik M 1997 Robustness of synchronized chaotic oscillations Int J Bifurc Chaos 7 625ndash43Rulkov N Sushchik M Tsimring L and Abarbanel H 1995 Generalized synchronization of chaos in unidirectionally
coupled chaotic systems Phys Rev E 51 980ndash94Schiff S So P Chang T Burke R and Sauer T 1996 Detecting dynamical interdependence and generalized synchrony
through mutual prediction in a neural ensemble Phys Rev E 54 6708ndash24Silberstein R B 1995 Neuromodulations of neocortical dynamics Neocortical Dynamics and Human EEG Rhythms
ed P L Nunez (Oxford Oxford University Press)Singer W 1995 Putative functions of temporal correlations in neocortical processing Large-Scale Neuronal Theories
of the Brain ed C Koch and J Davis (London MIT Press)Somers D and Kopell N 1995 Waves and synchrony in networks of oscillators of relaxation and non-relaxation type
Physica D 89 169ndash83Sutton R S and Barto A G 1990 Time derivative models of Pavlovian reinforcement Learning and Computational
Neuroscience Foundations of Adaptive Networks ed M Gabriel and J Moore (Cambridge MA MIT Press) pp497ndash538
Szentagothai J 1983 The modular architectonic principle of neural centers Rev Physiol Biochem Pharmacol 9811ndash61
Tallon-Baudry C Betrand O and Fischer C 2001 Oscillatory synchrony between human extrastriate areas during visualshort-term memory maintenance J Neurosci 21 RC177(1ndash5)
Tass P A 2002 Desynchronization of brain rhythms with soft phase-resetting techniques Biol Cybern 87 102ndash15
732 M Breakspear et al
Tass P Rosenblum M Weule J Kurths J Pikovsky A Volkmann J Schnitzler A and Freund H 1998 Detection of nmphase locking from noisy data application to magnetoencephalography Phys Rev Lett 81 3291ndash4
Terman D and Wang D 1995 Global competition and local cooperation in a network of neural oscillators Physica D81 148ndash76
Terry J and Breakspear M 2003 An improved algorithm for the detection of dynamical interdependence in bivariatetime-series Biol Cybern 88 129ndash36
Tononi G Sporns O and Edelman G M 1994 A measure for brain complexity relating functional segregation andintegration in the nervous system Proc Natl Acad Sci USA 91 5033ndash7
von der Malsburg C 1985 Nervous structures with dynamical links Phys Chem 89 703ndash10Wang X 1991 Period-doublings to chaos in a simple neural network an analytic proof Complex Syst 5 425ndash41Wennekers T and Pasemann F 1996 Synchronous chaos in high dimensional modular neural networks Int J Bifurc
Chaos 6 2055ndash67Zaks M A Parks E H Rosenblum M G and Kurths J 1999 Alternative locking ratios in imperfect phase synchronization
Phys Rev Lett 82 4228ndash31Zhao L and Macau E N 2001 A network of dynamically coupled chaotic maps for scene segmentation IEEE Trans
Neural Netw 12 1375ndash85

724 M Breakspear et al
orderly evolution of the distance between the two systems and in panel (c) where the evolutionof the phase difference reveals a brief violation of the criterion |φ1 minus φ2| lt constant Anexample of the phase difference evolution over a longer time interval is shown in figure 13(d)Note that this is a three-dimensional plot with sin(φ1 minus φ2) and cos(φ1 minus φ2) as they- and z-axes Hence phase desynchronizations are evident as complete revolutions aroundthe cylindrical phase space in which this variable evolves It can be seen that these phenomenaoccur at irregular intervals in the temporal domain
This behaviour can be explained by considering the nature of a chaotic attractor In thestatistical sense an arbitrary initial condition is expected to lead to the chaotic behaviour onthe attractor Consequently when we calculate the Lyapunov exponents of the system theseexponents correspond to those of the chaotic attractor Typically however there are an infinitenumber of unstable periodic orbits embedded within the attractor If the path of a trajectorybecomes close to one of these orbits it may lsquoshadowrsquo the orbit for a brief period of time Eachof these orbits has a corresponding set of Lyapunov exponents which (like the attractor) can bedivided into tangential and transverse exponents These also vary when we change the couplingparameter but are typically not the same as the exponents of the attractor Hence the largesttransverse exponent of the attractor may be negative yet there may be positive transverseexponents associated with periodic orbits Considering all of these exponents together wewould therefore only expect to observe complete stability of the synchronization manifold inthe case where all of the transverse Lyapunov exponents are negative Bursting behaviourwill be observed when the one of these exponents becomes positive typically correspondingto a periodic orbit of low order (Pikovsky and Grassberger 1991 Rulkov and Sushchik 1997Maistrenko et al 1998 Heagy et al 1998) Due to the complex structure of such periodic orbitsthe desynchronous bursts occur across a wide range of timescales
In figure 14 we present the results of a long integration (16times105 time units) in which thelengths of sequential synchronous epochs of phase synchronization are studied The parametervalues were δV = 065 for column 1 and 0645 for column 2 with coupling strength10 equal to0125 In panel (a) the power spectrum of these sequential lengths is plotted The whiteness ofthis spectrum indicates that the lengths of consecutive synchronous epochs are independentThis is due to the high-dimensional lsquomixingrsquo effect of desynchronization In panel (b) thelengths of the synchronous epochs have been sorted again into the order of increasing lengthThe results for two coupling strengths are plotted C = 0125 (solid) and 0135 (dashed)Several features of this plot are of note Firstly the lengths of the synchronous epochsare distributed over almost three orders of magnitude Secondly for both plots there is anapproximately log-linear relationship between the length of an epoch and its relative orderThis reflects an approximately exponential relationship between the length of a synchronousepoch and its likelihood of occurrence Thirdly this relationship is skewed toward longersynchronous epochs for the stronger coupling strengths
This type of signal with patches of lsquoorderrsquo occurring over many timescales interruptedby bursts of disorder can be shown to have a high information capacity (Gatlin 1972)Moreover complex phase desynchronization may make an infrequent yet robust contributionto the dynamical pattern of the alpha rhythm in scalp EEG data for resting human subjects(Breakspear 2002) It is tempting to postulate that such a lsquoweakly stablersquo dynamics may assistin the generation of the rich and fluid mental associations that spontaneously emerge in restingcognitive states Although this phenomenon has been previously described in a variety of othersystems to our knowledge this is the first description in a biophysical model of neocortex
10 The introduction of parameter mismatch causes an increase in the strength of coupling required for stablesynchronization above the value of 01 required in the symmetric case
Synchronization in a model of neuronal dynamics 725
Figure 14 Properties of sequential epochs of phase-synchronous behaviour over a long integrationtime δ1
V = 065 δ2V = 0645 aee = 04 (a) The flat (white) power spectrum (b) Synchronous
epochs sorted in order of increasing length (solid C = 0125 dashed C = 0134)
52 Synchronization and complex dynamics in larger arrays
The synchronous and complex phenomena described above are also observed in arrays ofcoupled systems with N gt 2 Expanding the size of the array permits complex dynamicsin the spatial in addition to the temporal domain However the basic principles governingthe type and stability of synchronized motions are the same In this section a few of thepossible behaviours are discussed Physiological variation across the array is modelled bymodifying the parameter δV = 07 in each array by adding a random number of maximummagnitude 001
In figure 15(a) an array of N = 20 uncoupled (C = 0) systems is shown The lack ofsynchronization in this time series is evident By comparison the rapid transition to completesynchronization is evident in figure 15(b) when C = 03 In fact 19 of the 20 subsystems
726 M Breakspear et al
(b)
(c)
(a)
Figure 15 Synchronization phenomena in an array of coupled cortical columns (a) N = 20C = 0 (b) N = 20 C = 03 (c) N = 6 C = 01 (d) N = 12 C = 009 (e) N = 12C = 012 A small perturbation has been applied to one of the columns at t = 2500 s (f) Evolutionof the phase difference between two of the columns in panel (e) For clarity the W -variable isplotted in panel (c) and the Z -variable in (e)
have phase synchronized within one single cycle Intermediate values of coupling strength areassociated with a rich variety of phenomena For example subsystems with the most similarparameter values typically synchronize into clusters of various sizes An example of this isillustrated in figure 15(c) Note that there is no synchronization between the two differentclusters each containing three synchronized subsystems
Another complex behaviour occurs when subsystems transiently synchronize into largeclusters then spontaneously desynchronize again Hence the dynamics of the entire arrayjumps between different transient dynamics characterized by varying degrees of freedom Anexample of this phenomenon is given in figure 15(d) In addition it is possible to inducedesynchronization across an entire array by applying a small perturbation to one columnfor one time step An example is illustrated in figure 15(e) A small perturbation has been
Synchronization in a model of neuronal dynamics 727
Figure 15 (Continued)
added to one column after 2500 time units It can be seen that the entire array subsequentlydesynchronizes This illustrates the sensitivity of the dynamics to external stimuli Finally it ispossible to lsquoinducersquo synchronization between selected subsystems by increasing the strength ofcoupling between them selectively This allows an extra level of complexity to be introducedinto the model For example it may allow targeted information transfer between a partialnetwork of the entire array
As discussed in section 51 a univariate signal which jumps between epochs of lowentropy (high repetition) and bursts of high entropy (high variability) can be shown to havehigh information capacity This is because the repetition of certain patterns (as in the English
728 M Breakspear et al
language) permits error detectability Bursts of high entropy act in the opposite directionto increase message variability With a multivariate signal in addition to optimizing theinformation capacity within each signal the overall information capacity can be increasedthrough the type of itinerant synchronization briefly illustrated in this section Synchronizationacross an entire array increases the message reliability (since a single lsquoerrant letterrsquo in onesignal can be easily detected) Conversely desynchronization into multiple clusters increasesmessage variability since each cluster will produce independent information Hence a neuralarray which switches dynamically between a range of cluster configurations can be seen asadaptive in the sense that high synchronous states can be expressed when error detectabilityis a priority (such as during quick and reliable execution of a familiar task) but the array canrapidly desynchronize if variability becomes salient (for example if the environment suddenlybecomes novel) A formal operationalized demonstration of this principle is to be the subjectof future work
6 Discussion
In this paper we present a dynamical model for activity in an array of weakly coupledlocal neural assemblies The model incorporates the fundamental principles governing cellmembrane potential as determined by voltage- and ligand-gated ion channels In this wayit allows a reasonable amount of physiological complexity to be incorporated includingsynaptic activation and receptor subtypeswithout recourse to overwhelming detail Numericalanalysis of this model reveals a transition from independent oscillations to stable chaoticsynchronization depending on the balance of local versus long-range connectivity andbiophysical heterogeneity These excitatory couplings are subject to ascending subcorticalneuromodulation arising principally from monoaminergic nuclei such as Locus Coeruleus(Silberstein 1995) Neuromodulation of extrinsic connections was incorporated into the modelvia a coupling parameter C The stability of the synchronization is determined by the valueof the largest transverse Lyapunov exponent which in turn depends on the coupling parameterC If the coupling is weakened with the result that this exponent becomes positive a blowoutbifurcation occurs and independent oscillations re-emerge In the vicinity of the transitionnon-linear desynchronizations with complex temporal and spatial properties are observed
It is pleasing that complex and itinerant behaviour reminiscent of real neuronaldynamics emerges through such a simple manipulation of a single dynamic control parameterFurthermore the ubiquitous projections of classical neuromodulatory systems in real brainssuggest a clear biological mechanism by which the brain could self-organize throughmoderating the complexity of its own dynamics There are many reasons that the complicateddynamics of the sort demonstrated in this paper may be neurobiologically important Wemention four
First the presence of multiple clusters of phase-synchronized subsystems as discussedabove may be a uniquely non-linear mechanism facilitating the lsquocarriagersquo and distinction ofmultiple concurrent dynamic cell assemblies (and hence cognitive processing) This may beimportant for perceptual grouping binding and transitions as suggested by others (eg von derMalsburg 1985 Gray and Singer 1989)
Second the facility to decouple and reconfigure dynamic cell assemblies througha neuromodulatory control parameter may be important for avoiding low-entropysynchronization manifolds In the absence of itinerancy synchronization would renderneuronal dynamics incapable of representing changes in sensory input or indeed cognitive stateFor example in the model it was observed that small perturbations to one column were ableto induce a global change in neuronal dynamics by disrupting a state of weak synchronization
Synchronization in a model of neuronal dynamics 729
Inevitably alternative dynamic configurations emerge epitomizing the importance of lsquolabilersquobrain dynamics (Friston 2000a)
Thirdly from a purely functionalist point of view the balance between functionalspecialization and functional integration may rely upon the sorts of mechanism modelledin this paper The brain appears to adhere to two principles of functional organization namelysegregation of function within cortical areas and sub-areas and the integration of these areasin perceptual synthesis cognition and sensorimotor integration (Tononi et al 1994 Fristonet al 1995) The balance between preserving specialized dynamics unique to each arearequired by specialization and the coupling among these areas mediating functional integrationmay be achieved by the neuromodulatory mechanisms described above Significantlythe spontaneous variation in the numbers of degrees of freedom (for example whenlow-dimensional synchronization manifolds are shadowed) imparts a dynamic characteristicto this perspective that is precluded by linear mechanisms alone
Finally from a mechanistic perspective complexity and diversity of brain dynamics arerequired for selection of adaptive responses (Friston 2000b) There are many examples ofselective mechanisms operating within the brain All can be seen as performing an ascenton some objective function or fitness landscape These mechanisms require complex anditinerant dynamics to facilitate a thorough exploration of the landscape These landscapesmay range from those involved in perception (where the landscape is defined by predictionerror see eg Rao and Ballard (1998)) to those involved in value learning (where thelandscape is determined by the outcome of a behaviour eg in temporal difference models (seeeg Sutton and Barto 1990)) In short complexity is necessary to maintain an information-rich environment in which the brain can respond to sensory inputs An essential componentof this is the maintenance of diversity which is a prerequisite for selectionist mechanismsat many scales (Edelman 1993) Understanding the mechanistic basis of complex dynamicsas measured electrophysiologically may well depend on the sort of biologically plausiblemodels used in this paper This understanding may be important for both basic and clinicalneuroscience (eg Tass 2002)
The present study supplements and advances on other research in this field The studyof complex phase synchronization and clustering in neural oscillator networks similar tothe present model has previously been the subject of formal mathematical study Forexample Terman and Wang (1995) studied synchronization of blocks of neural oscillatorsthat were subject to perturbation by connected lsquostimulirsquo Somers and Kopell (1995) studiedthe emergence of both synchronization and travelling waves in networks of coupled neuraloscillators In the work of Terman and Wang the local dynamics were based on neurallsquocaricaturersquo models such as the FitzHughndashNagumo equations and are hence more abstract thanthe current study The approach of Somers and Kopell only required that the local dynamicsexhibit fast and relaxation phases and is hence more abstract still In both studies onlyperiodic dynamics are studied Complex patterns of chaos synchronization in neural arrayshave been characterized by nonlinear methods (Kaneko 1998) and their putative contributionto both learning (Nakajima and Ueda 1996) and perception (Zhao and Macau 2001 Breakspear2001) explored However these studies employed models such as the logistic equation andtent map which are quite abstract in comparison to the present study On the other handHansel and Sompolinsky (1996) studied the role of chaos synchronization in a model ofvisual perception in a cortical hypercolumn using a very similar approach to that adoptedhere but with the additional complexity that synaptic delays were incorporated However inthis and earlier work (Hansel and Sompolinsky 1992) only linear measures of dynamics andsynchrony (specifically the auto- and cross-correlation functions) were employed The analysisof chaotic synchrony in the study of Larter et al (1999) was based only upon observation of
730 M Breakspear et al
the Euclidean difference between variables from two nodes within a larger array Hence thesignificance of the present work is that it provides a quantitative nonlinear analysis of complexchaos synchronization in a biophysical model of cortical dynamics Such research is requiredin order to bridge the gap between quantitative nonlinear research into synchronization andcomputation in abstract models of neural arrays and qualitative or linear analysis in morebiophysically plausible models
Acknowledgments
The authors would like to thank Peter Robinson for assistance with the manuscript and R Larterfor helpful advice MB is a recipient of a NSW Institute of Psychiatry Research Grant anda University of Sydney SESQUI postdoctoral fellowship KJF is supported by the WellcomeTrust
References
Abarbanel H Rulkov N and Sushchik M 1996 Generalised synchronization of chaos the auxiliary system approachPhys Rev E 53 4528ndash35
Afraimovich V Verichev N and Rabinovich M 1986 Stochastic synchronization of oscillation in dissipative systemsRadiophys Quantum Electron 29 795ndash801
Breakspear M 2001 Perception of odours by a nonlinear model of the olfactory bulb Int J Neural Syst 11 101ndash24Breakspear M 2002 Nonlinear phase desynchronisation in human electroencephalographic data Human Brain Mapping
15 175ndash98Breakspear M and Terry J R 2002a Nonlinear interdependence in neural systems motivation theory and relevance
Int J Neurosci 112 1163ndash84Breakspear M and Terry J R 2002b Detection and description of nonlinear interdependence in normal multichannel
human EEG Clin Neurophysiol 113 735ndash53Breakspear M and Terry J R 2002c Topographic organization of nonlinear interdependence in multichannel human
EEG Neuroimage 16 822ndash35Destexhe A Rudolph M Fellous J M and Sejnowski T J 2001 Fluctuating synaptic conductances recreate in vivo-like
activity in neocortical neurons Neuroscience 107 13ndash24Douglas R J Mahowald M A and Martin K A 1996 Microarchitecture of neocortical columns Brain TheorymdashBiological
Basis and Computational Principles ed A Aertsen and V Braitenberg (Amsterdam Elsevier)Eckhorn R Bauer R Jordan W Brosch M Kruse W Munk M and Reitboeck H J 1988 Coherent oscillations a
mechanism of feature linking in the visual cortex Multiple electrode and correlation analyses in the cat BiolCybern 60 121ndash30
Edelman G M 1993 Neural Darwinism selection and reentrant signalling in higher brain function Neuron 10 115ndash25Engel A K Kreiter A K Konig P and Singer W 1991 Synchronization of oscillatory neuronal responses between
striate and extrastriate visual cortical areas of the cat Proc Natl Acad Sci USA 88 6048ndash52Friston K J 2000a The labile brain I Neuronal transients and nonlinear coupling Phil Trans R Soc B 355 215ndash36Friston K J 2000b The labile brain II Transients complexity and selection Phil Trans R Soc B 355 237ndash52Friston K J Tononi G Sporns O and Edelman G 1995 Characterising the complexity of neuronal interactions Human
Brain Mapping 3 302ndash14Fujisaka H and Yamada T 1983 Stability theory of synchronized motion in coupled-oscillator system Prog Theor
Phys 69 32ndash47Gatlin L 1972 Information Theory and the Living System (New York Columbia University Press)Gray C M Konig P Engel A and Singer W 1989 Oscillatory responses in cat visual cortex exhibit inter-columnar
synchronization which reflects global stimulus properties Science 338 334ndash7Gray C M and Singer W 1989 Stimulus specific neuronal oscillations in orientation columns of cat visual cortex Proc
Natl Acad Sci USA 86 1698ndash702Haig A R Gordon E Wright J J Meares R A and Bahramali H 2000 Synchronous cortical gamma-band activity in
task-relevant cognition Neuroreport 11 669ndash75Hansel D and Sompolinsky H 1992 Synchronization and computation in a chaotic neural network Phys Rev Lett 68
718ndash21
Synchronization in a model of neuronal dynamics 731
Hansel D and Sompolinsky H 1996 Chaos and synchrony in a model of a hypercolumn in visual cortex J ComputNeurosci 3 7ndash4
Heagy J Carroll T and Pecora L 1998 Desynchronization by periodic orbits Phys Rev E 57 R1253ndash6Hunt B Ott E and Yorke J 1997 Differentiable synchronization of chaos Phys Rev E 55 4029ndash34Josic K 1998 Invariant manifolds and synchronization of coupled dynamical systems Phys Rev Lett 80 3053ndash6Kandel E Schwartz J and Jessell T 2000 Principles of Neural Science (New York McGraw-Hill)Kaneko K 1998 On the strength of attractors in a high-dimensional system Physica D 124 322ndash44Kocarev L Parlitz U and Brown R 2000 Robust synchronization of chaotic systems Phys Rev E 61 3716ndash20Larter R Speelman B and Worth R M 1999 A coupled ordinary differential equation lattice model for the simulation
of epileptic seizures Chaos 9 795ndash804Lumer E Edelman G and Tononi G 1997a Neural dynamics in a model of the thalamocortical system I Layers loops
and the emergence of fast synchronous oscillations Cereb Cortex 7 207ndash27Lumer E Edelman G and Tononi G 1997b Neural dynamics in a model of the thalamocortical system II The role of
neural synchrony tested through perturbations of spike timing Cereb Cortex 7 228ndash36Magleby K 1987 Short-term modulation at synapses Synaptic Function ed G M Edelman W E Gall and
W M Cowan (New York Wiley)Maistrenko Y Maistrenko V Popovich A and Mosekilde E 1998 Transverse instability and riddled basins in a system
of two coupled logistic maps Phys Rev E 57 2713ndash24Morris C and Lecar H 1981 Voltage oscillations in the barnacle giant muscle fiber Biophys J 35 193ndash213Mountcastle V B 1997 The columnar organization of the neocortex Brain 120 701ndash22Mountcastle V B 1998 The Cerebral Cortex (Cambridge MA Harvard University Press)Nakajima H and Ueda Y 1996 Riddled basins of the optimal states in learning dynamical systems Physica D 99 35ndash44Nunez P L 1995 Neocortical Dynamics and Human Brain Rhythms (Oxford Oxford University Press)Osipov G V Pikovsky A S Rosenblum M G and Kurths J 1997 Phase synchronization effects in a lattice of nonidentical
Rossler oscillators Phys Rev E 55 2353ndash61Ott E and Sommerer J 1994 Blowout bifurcations the occurrence of riddled basins and onndashoff intermittency Phys
Lett A 188 39ndash47Pecora L and Carroll T 1990 Synchronization in chaotic systems Phys Rev Lett 64 821ndash4Pikovsky A and Grassberger P 1991 Symmetry breaking bifurcation for coupled chaotic attractors J Phys A Math
Gen 24 4587ndash97Rao R P and Ballard D H 1998 Predictive coding in the visual cortex a functional interpretation of some extra-classical
receptive field effects Nature Neurosci 2 79ndash87Robinson P A Wright J J and Rennie C R 1998 Synchronous oscillations in the cerebral cortex Phys Rev E 57
4578ndash88Rodriguez E George N Lachauz J P Martinerie J Renault B and Varela F J 1999 Perceptionrsquos shadow long distance
synchronization of human brain activity Nature 397 430ndash3Rosenblum M Pikovsky A and Kurths J 1996 Phase synchronization of chaotic oscillators Phys Rev Lett 76 1804ndash7Rosenblum M Pikovsky A and Kurths J 1997 From phase to lag synchronization in coupled chaotic oscillators Phys
Rev Lett 78 4193ndash6Rulkov N and Sushchik M 1997 Robustness of synchronized chaotic oscillations Int J Bifurc Chaos 7 625ndash43Rulkov N Sushchik M Tsimring L and Abarbanel H 1995 Generalized synchronization of chaos in unidirectionally
coupled chaotic systems Phys Rev E 51 980ndash94Schiff S So P Chang T Burke R and Sauer T 1996 Detecting dynamical interdependence and generalized synchrony
through mutual prediction in a neural ensemble Phys Rev E 54 6708ndash24Silberstein R B 1995 Neuromodulations of neocortical dynamics Neocortical Dynamics and Human EEG Rhythms
ed P L Nunez (Oxford Oxford University Press)Singer W 1995 Putative functions of temporal correlations in neocortical processing Large-Scale Neuronal Theories
of the Brain ed C Koch and J Davis (London MIT Press)Somers D and Kopell N 1995 Waves and synchrony in networks of oscillators of relaxation and non-relaxation type
Physica D 89 169ndash83Sutton R S and Barto A G 1990 Time derivative models of Pavlovian reinforcement Learning and Computational
Neuroscience Foundations of Adaptive Networks ed M Gabriel and J Moore (Cambridge MA MIT Press) pp497ndash538
Szentagothai J 1983 The modular architectonic principle of neural centers Rev Physiol Biochem Pharmacol 9811ndash61
Tallon-Baudry C Betrand O and Fischer C 2001 Oscillatory synchrony between human extrastriate areas during visualshort-term memory maintenance J Neurosci 21 RC177(1ndash5)
Tass P A 2002 Desynchronization of brain rhythms with soft phase-resetting techniques Biol Cybern 87 102ndash15
732 M Breakspear et al
Tass P Rosenblum M Weule J Kurths J Pikovsky A Volkmann J Schnitzler A and Freund H 1998 Detection of nmphase locking from noisy data application to magnetoencephalography Phys Rev Lett 81 3291ndash4
Terman D and Wang D 1995 Global competition and local cooperation in a network of neural oscillators Physica D81 148ndash76
Terry J and Breakspear M 2003 An improved algorithm for the detection of dynamical interdependence in bivariatetime-series Biol Cybern 88 129ndash36
Tononi G Sporns O and Edelman G M 1994 A measure for brain complexity relating functional segregation andintegration in the nervous system Proc Natl Acad Sci USA 91 5033ndash7
von der Malsburg C 1985 Nervous structures with dynamical links Phys Chem 89 703ndash10Wang X 1991 Period-doublings to chaos in a simple neural network an analytic proof Complex Syst 5 425ndash41Wennekers T and Pasemann F 1996 Synchronous chaos in high dimensional modular neural networks Int J Bifurc
Chaos 6 2055ndash67Zaks M A Parks E H Rosenblum M G and Kurths J 1999 Alternative locking ratios in imperfect phase synchronization
Phys Rev Lett 82 4228ndash31Zhao L and Macau E N 2001 A network of dynamically coupled chaotic maps for scene segmentation IEEE Trans
Neural Netw 12 1375ndash85
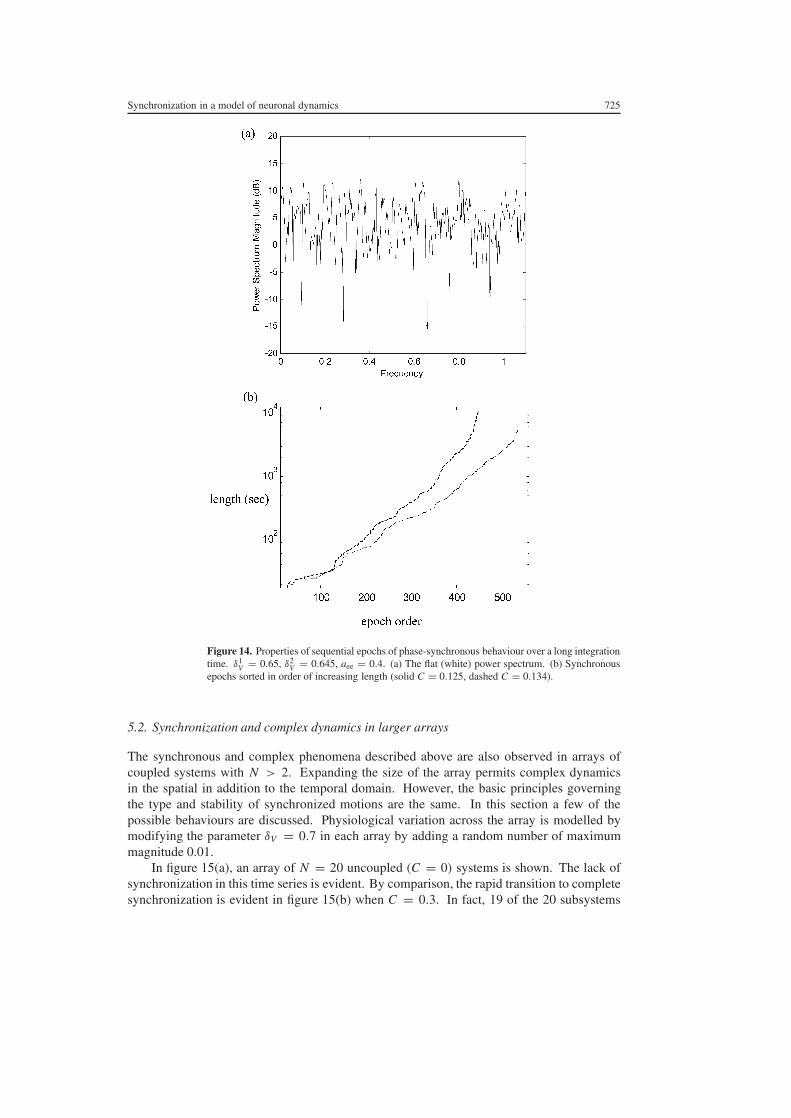
Synchronization in a model of neuronal dynamics 725
Figure 14 Properties of sequential epochs of phase-synchronous behaviour over a long integrationtime δ1
V = 065 δ2V = 0645 aee = 04 (a) The flat (white) power spectrum (b) Synchronous
epochs sorted in order of increasing length (solid C = 0125 dashed C = 0134)
52 Synchronization and complex dynamics in larger arrays
The synchronous and complex phenomena described above are also observed in arrays ofcoupled systems with N gt 2 Expanding the size of the array permits complex dynamicsin the spatial in addition to the temporal domain However the basic principles governingthe type and stability of synchronized motions are the same In this section a few of thepossible behaviours are discussed Physiological variation across the array is modelled bymodifying the parameter δV = 07 in each array by adding a random number of maximummagnitude 001
In figure 15(a) an array of N = 20 uncoupled (C = 0) systems is shown The lack ofsynchronization in this time series is evident By comparison the rapid transition to completesynchronization is evident in figure 15(b) when C = 03 In fact 19 of the 20 subsystems
726 M Breakspear et al
(b)
(c)
(a)
Figure 15 Synchronization phenomena in an array of coupled cortical columns (a) N = 20C = 0 (b) N = 20 C = 03 (c) N = 6 C = 01 (d) N = 12 C = 009 (e) N = 12C = 012 A small perturbation has been applied to one of the columns at t = 2500 s (f) Evolutionof the phase difference between two of the columns in panel (e) For clarity the W -variable isplotted in panel (c) and the Z -variable in (e)
have phase synchronized within one single cycle Intermediate values of coupling strength areassociated with a rich variety of phenomena For example subsystems with the most similarparameter values typically synchronize into clusters of various sizes An example of this isillustrated in figure 15(c) Note that there is no synchronization between the two differentclusters each containing three synchronized subsystems
Another complex behaviour occurs when subsystems transiently synchronize into largeclusters then spontaneously desynchronize again Hence the dynamics of the entire arrayjumps between different transient dynamics characterized by varying degrees of freedom Anexample of this phenomenon is given in figure 15(d) In addition it is possible to inducedesynchronization across an entire array by applying a small perturbation to one columnfor one time step An example is illustrated in figure 15(e) A small perturbation has been
Synchronization in a model of neuronal dynamics 727
Figure 15 (Continued)
added to one column after 2500 time units It can be seen that the entire array subsequentlydesynchronizes This illustrates the sensitivity of the dynamics to external stimuli Finally it ispossible to lsquoinducersquo synchronization between selected subsystems by increasing the strength ofcoupling between them selectively This allows an extra level of complexity to be introducedinto the model For example it may allow targeted information transfer between a partialnetwork of the entire array
As discussed in section 51 a univariate signal which jumps between epochs of lowentropy (high repetition) and bursts of high entropy (high variability) can be shown to havehigh information capacity This is because the repetition of certain patterns (as in the English
728 M Breakspear et al
language) permits error detectability Bursts of high entropy act in the opposite directionto increase message variability With a multivariate signal in addition to optimizing theinformation capacity within each signal the overall information capacity can be increasedthrough the type of itinerant synchronization briefly illustrated in this section Synchronizationacross an entire array increases the message reliability (since a single lsquoerrant letterrsquo in onesignal can be easily detected) Conversely desynchronization into multiple clusters increasesmessage variability since each cluster will produce independent information Hence a neuralarray which switches dynamically between a range of cluster configurations can be seen asadaptive in the sense that high synchronous states can be expressed when error detectabilityis a priority (such as during quick and reliable execution of a familiar task) but the array canrapidly desynchronize if variability becomes salient (for example if the environment suddenlybecomes novel) A formal operationalized demonstration of this principle is to be the subjectof future work
6 Discussion
In this paper we present a dynamical model for activity in an array of weakly coupledlocal neural assemblies The model incorporates the fundamental principles governing cellmembrane potential as determined by voltage- and ligand-gated ion channels In this wayit allows a reasonable amount of physiological complexity to be incorporated includingsynaptic activation and receptor subtypeswithout recourse to overwhelming detail Numericalanalysis of this model reveals a transition from independent oscillations to stable chaoticsynchronization depending on the balance of local versus long-range connectivity andbiophysical heterogeneity These excitatory couplings are subject to ascending subcorticalneuromodulation arising principally from monoaminergic nuclei such as Locus Coeruleus(Silberstein 1995) Neuromodulation of extrinsic connections was incorporated into the modelvia a coupling parameter C The stability of the synchronization is determined by the valueof the largest transverse Lyapunov exponent which in turn depends on the coupling parameterC If the coupling is weakened with the result that this exponent becomes positive a blowoutbifurcation occurs and independent oscillations re-emerge In the vicinity of the transitionnon-linear desynchronizations with complex temporal and spatial properties are observed
It is pleasing that complex and itinerant behaviour reminiscent of real neuronaldynamics emerges through such a simple manipulation of a single dynamic control parameterFurthermore the ubiquitous projections of classical neuromodulatory systems in real brainssuggest a clear biological mechanism by which the brain could self-organize throughmoderating the complexity of its own dynamics There are many reasons that the complicateddynamics of the sort demonstrated in this paper may be neurobiologically important Wemention four
First the presence of multiple clusters of phase-synchronized subsystems as discussedabove may be a uniquely non-linear mechanism facilitating the lsquocarriagersquo and distinction ofmultiple concurrent dynamic cell assemblies (and hence cognitive processing) This may beimportant for perceptual grouping binding and transitions as suggested by others (eg von derMalsburg 1985 Gray and Singer 1989)
Second the facility to decouple and reconfigure dynamic cell assemblies througha neuromodulatory control parameter may be important for avoiding low-entropysynchronization manifolds In the absence of itinerancy synchronization would renderneuronal dynamics incapable of representing changes in sensory input or indeed cognitive stateFor example in the model it was observed that small perturbations to one column were ableto induce a global change in neuronal dynamics by disrupting a state of weak synchronization
Synchronization in a model of neuronal dynamics 729
Inevitably alternative dynamic configurations emerge epitomizing the importance of lsquolabilersquobrain dynamics (Friston 2000a)
Thirdly from a purely functionalist point of view the balance between functionalspecialization and functional integration may rely upon the sorts of mechanism modelledin this paper The brain appears to adhere to two principles of functional organization namelysegregation of function within cortical areas and sub-areas and the integration of these areasin perceptual synthesis cognition and sensorimotor integration (Tononi et al 1994 Fristonet al 1995) The balance between preserving specialized dynamics unique to each arearequired by specialization and the coupling among these areas mediating functional integrationmay be achieved by the neuromodulatory mechanisms described above Significantlythe spontaneous variation in the numbers of degrees of freedom (for example whenlow-dimensional synchronization manifolds are shadowed) imparts a dynamic characteristicto this perspective that is precluded by linear mechanisms alone
Finally from a mechanistic perspective complexity and diversity of brain dynamics arerequired for selection of adaptive responses (Friston 2000b) There are many examples ofselective mechanisms operating within the brain All can be seen as performing an ascenton some objective function or fitness landscape These mechanisms require complex anditinerant dynamics to facilitate a thorough exploration of the landscape These landscapesmay range from those involved in perception (where the landscape is defined by predictionerror see eg Rao and Ballard (1998)) to those involved in value learning (where thelandscape is determined by the outcome of a behaviour eg in temporal difference models (seeeg Sutton and Barto 1990)) In short complexity is necessary to maintain an information-rich environment in which the brain can respond to sensory inputs An essential componentof this is the maintenance of diversity which is a prerequisite for selectionist mechanismsat many scales (Edelman 1993) Understanding the mechanistic basis of complex dynamicsas measured electrophysiologically may well depend on the sort of biologically plausiblemodels used in this paper This understanding may be important for both basic and clinicalneuroscience (eg Tass 2002)
The present study supplements and advances on other research in this field The studyof complex phase synchronization and clustering in neural oscillator networks similar tothe present model has previously been the subject of formal mathematical study Forexample Terman and Wang (1995) studied synchronization of blocks of neural oscillatorsthat were subject to perturbation by connected lsquostimulirsquo Somers and Kopell (1995) studiedthe emergence of both synchronization and travelling waves in networks of coupled neuraloscillators In the work of Terman and Wang the local dynamics were based on neurallsquocaricaturersquo models such as the FitzHughndashNagumo equations and are hence more abstract thanthe current study The approach of Somers and Kopell only required that the local dynamicsexhibit fast and relaxation phases and is hence more abstract still In both studies onlyperiodic dynamics are studied Complex patterns of chaos synchronization in neural arrayshave been characterized by nonlinear methods (Kaneko 1998) and their putative contributionto both learning (Nakajima and Ueda 1996) and perception (Zhao and Macau 2001 Breakspear2001) explored However these studies employed models such as the logistic equation andtent map which are quite abstract in comparison to the present study On the other handHansel and Sompolinsky (1996) studied the role of chaos synchronization in a model ofvisual perception in a cortical hypercolumn using a very similar approach to that adoptedhere but with the additional complexity that synaptic delays were incorporated However inthis and earlier work (Hansel and Sompolinsky 1992) only linear measures of dynamics andsynchrony (specifically the auto- and cross-correlation functions) were employed The analysisof chaotic synchrony in the study of Larter et al (1999) was based only upon observation of
730 M Breakspear et al
the Euclidean difference between variables from two nodes within a larger array Hence thesignificance of the present work is that it provides a quantitative nonlinear analysis of complexchaos synchronization in a biophysical model of cortical dynamics Such research is requiredin order to bridge the gap between quantitative nonlinear research into synchronization andcomputation in abstract models of neural arrays and qualitative or linear analysis in morebiophysically plausible models
Acknowledgments
The authors would like to thank Peter Robinson for assistance with the manuscript and R Larterfor helpful advice MB is a recipient of a NSW Institute of Psychiatry Research Grant anda University of Sydney SESQUI postdoctoral fellowship KJF is supported by the WellcomeTrust
References
Abarbanel H Rulkov N and Sushchik M 1996 Generalised synchronization of chaos the auxiliary system approachPhys Rev E 53 4528ndash35
Afraimovich V Verichev N and Rabinovich M 1986 Stochastic synchronization of oscillation in dissipative systemsRadiophys Quantum Electron 29 795ndash801
Breakspear M 2001 Perception of odours by a nonlinear model of the olfactory bulb Int J Neural Syst 11 101ndash24Breakspear M 2002 Nonlinear phase desynchronisation in human electroencephalographic data Human Brain Mapping
15 175ndash98Breakspear M and Terry J R 2002a Nonlinear interdependence in neural systems motivation theory and relevance
Int J Neurosci 112 1163ndash84Breakspear M and Terry J R 2002b Detection and description of nonlinear interdependence in normal multichannel
human EEG Clin Neurophysiol 113 735ndash53Breakspear M and Terry J R 2002c Topographic organization of nonlinear interdependence in multichannel human
EEG Neuroimage 16 822ndash35Destexhe A Rudolph M Fellous J M and Sejnowski T J 2001 Fluctuating synaptic conductances recreate in vivo-like
activity in neocortical neurons Neuroscience 107 13ndash24Douglas R J Mahowald M A and Martin K A 1996 Microarchitecture of neocortical columns Brain TheorymdashBiological
Basis and Computational Principles ed A Aertsen and V Braitenberg (Amsterdam Elsevier)Eckhorn R Bauer R Jordan W Brosch M Kruse W Munk M and Reitboeck H J 1988 Coherent oscillations a
mechanism of feature linking in the visual cortex Multiple electrode and correlation analyses in the cat BiolCybern 60 121ndash30
Edelman G M 1993 Neural Darwinism selection and reentrant signalling in higher brain function Neuron 10 115ndash25Engel A K Kreiter A K Konig P and Singer W 1991 Synchronization of oscillatory neuronal responses between
striate and extrastriate visual cortical areas of the cat Proc Natl Acad Sci USA 88 6048ndash52Friston K J 2000a The labile brain I Neuronal transients and nonlinear coupling Phil Trans R Soc B 355 215ndash36Friston K J 2000b The labile brain II Transients complexity and selection Phil Trans R Soc B 355 237ndash52Friston K J Tononi G Sporns O and Edelman G 1995 Characterising the complexity of neuronal interactions Human
Brain Mapping 3 302ndash14Fujisaka H and Yamada T 1983 Stability theory of synchronized motion in coupled-oscillator system Prog Theor
Phys 69 32ndash47Gatlin L 1972 Information Theory and the Living System (New York Columbia University Press)Gray C M Konig P Engel A and Singer W 1989 Oscillatory responses in cat visual cortex exhibit inter-columnar
synchronization which reflects global stimulus properties Science 338 334ndash7Gray C M and Singer W 1989 Stimulus specific neuronal oscillations in orientation columns of cat visual cortex Proc
Natl Acad Sci USA 86 1698ndash702Haig A R Gordon E Wright J J Meares R A and Bahramali H 2000 Synchronous cortical gamma-band activity in
task-relevant cognition Neuroreport 11 669ndash75Hansel D and Sompolinsky H 1992 Synchronization and computation in a chaotic neural network Phys Rev Lett 68
718ndash21
Synchronization in a model of neuronal dynamics 731
Hansel D and Sompolinsky H 1996 Chaos and synchrony in a model of a hypercolumn in visual cortex J ComputNeurosci 3 7ndash4
Heagy J Carroll T and Pecora L 1998 Desynchronization by periodic orbits Phys Rev E 57 R1253ndash6Hunt B Ott E and Yorke J 1997 Differentiable synchronization of chaos Phys Rev E 55 4029ndash34Josic K 1998 Invariant manifolds and synchronization of coupled dynamical systems Phys Rev Lett 80 3053ndash6Kandel E Schwartz J and Jessell T 2000 Principles of Neural Science (New York McGraw-Hill)Kaneko K 1998 On the strength of attractors in a high-dimensional system Physica D 124 322ndash44Kocarev L Parlitz U and Brown R 2000 Robust synchronization of chaotic systems Phys Rev E 61 3716ndash20Larter R Speelman B and Worth R M 1999 A coupled ordinary differential equation lattice model for the simulation
of epileptic seizures Chaos 9 795ndash804Lumer E Edelman G and Tononi G 1997a Neural dynamics in a model of the thalamocortical system I Layers loops
and the emergence of fast synchronous oscillations Cereb Cortex 7 207ndash27Lumer E Edelman G and Tononi G 1997b Neural dynamics in a model of the thalamocortical system II The role of
neural synchrony tested through perturbations of spike timing Cereb Cortex 7 228ndash36Magleby K 1987 Short-term modulation at synapses Synaptic Function ed G M Edelman W E Gall and
W M Cowan (New York Wiley)Maistrenko Y Maistrenko V Popovich A and Mosekilde E 1998 Transverse instability and riddled basins in a system
of two coupled logistic maps Phys Rev E 57 2713ndash24Morris C and Lecar H 1981 Voltage oscillations in the barnacle giant muscle fiber Biophys J 35 193ndash213Mountcastle V B 1997 The columnar organization of the neocortex Brain 120 701ndash22Mountcastle V B 1998 The Cerebral Cortex (Cambridge MA Harvard University Press)Nakajima H and Ueda Y 1996 Riddled basins of the optimal states in learning dynamical systems Physica D 99 35ndash44Nunez P L 1995 Neocortical Dynamics and Human Brain Rhythms (Oxford Oxford University Press)Osipov G V Pikovsky A S Rosenblum M G and Kurths J 1997 Phase synchronization effects in a lattice of nonidentical
Rossler oscillators Phys Rev E 55 2353ndash61Ott E and Sommerer J 1994 Blowout bifurcations the occurrence of riddled basins and onndashoff intermittency Phys
Lett A 188 39ndash47Pecora L and Carroll T 1990 Synchronization in chaotic systems Phys Rev Lett 64 821ndash4Pikovsky A and Grassberger P 1991 Symmetry breaking bifurcation for coupled chaotic attractors J Phys A Math
Gen 24 4587ndash97Rao R P and Ballard D H 1998 Predictive coding in the visual cortex a functional interpretation of some extra-classical
receptive field effects Nature Neurosci 2 79ndash87Robinson P A Wright J J and Rennie C R 1998 Synchronous oscillations in the cerebral cortex Phys Rev E 57
4578ndash88Rodriguez E George N Lachauz J P Martinerie J Renault B and Varela F J 1999 Perceptionrsquos shadow long distance
synchronization of human brain activity Nature 397 430ndash3Rosenblum M Pikovsky A and Kurths J 1996 Phase synchronization of chaotic oscillators Phys Rev Lett 76 1804ndash7Rosenblum M Pikovsky A and Kurths J 1997 From phase to lag synchronization in coupled chaotic oscillators Phys
Rev Lett 78 4193ndash6Rulkov N and Sushchik M 1997 Robustness of synchronized chaotic oscillations Int J Bifurc Chaos 7 625ndash43Rulkov N Sushchik M Tsimring L and Abarbanel H 1995 Generalized synchronization of chaos in unidirectionally
coupled chaotic systems Phys Rev E 51 980ndash94Schiff S So P Chang T Burke R and Sauer T 1996 Detecting dynamical interdependence and generalized synchrony
through mutual prediction in a neural ensemble Phys Rev E 54 6708ndash24Silberstein R B 1995 Neuromodulations of neocortical dynamics Neocortical Dynamics and Human EEG Rhythms
ed P L Nunez (Oxford Oxford University Press)Singer W 1995 Putative functions of temporal correlations in neocortical processing Large-Scale Neuronal Theories
of the Brain ed C Koch and J Davis (London MIT Press)Somers D and Kopell N 1995 Waves and synchrony in networks of oscillators of relaxation and non-relaxation type
Physica D 89 169ndash83Sutton R S and Barto A G 1990 Time derivative models of Pavlovian reinforcement Learning and Computational
Neuroscience Foundations of Adaptive Networks ed M Gabriel and J Moore (Cambridge MA MIT Press) pp497ndash538
Szentagothai J 1983 The modular architectonic principle of neural centers Rev Physiol Biochem Pharmacol 9811ndash61
Tallon-Baudry C Betrand O and Fischer C 2001 Oscillatory synchrony between human extrastriate areas during visualshort-term memory maintenance J Neurosci 21 RC177(1ndash5)
Tass P A 2002 Desynchronization of brain rhythms with soft phase-resetting techniques Biol Cybern 87 102ndash15
732 M Breakspear et al
Tass P Rosenblum M Weule J Kurths J Pikovsky A Volkmann J Schnitzler A and Freund H 1998 Detection of nmphase locking from noisy data application to magnetoencephalography Phys Rev Lett 81 3291ndash4
Terman D and Wang D 1995 Global competition and local cooperation in a network of neural oscillators Physica D81 148ndash76
Terry J and Breakspear M 2003 An improved algorithm for the detection of dynamical interdependence in bivariatetime-series Biol Cybern 88 129ndash36
Tononi G Sporns O and Edelman G M 1994 A measure for brain complexity relating functional segregation andintegration in the nervous system Proc Natl Acad Sci USA 91 5033ndash7
von der Malsburg C 1985 Nervous structures with dynamical links Phys Chem 89 703ndash10Wang X 1991 Period-doublings to chaos in a simple neural network an analytic proof Complex Syst 5 425ndash41Wennekers T and Pasemann F 1996 Synchronous chaos in high dimensional modular neural networks Int J Bifurc
Chaos 6 2055ndash67Zaks M A Parks E H Rosenblum M G and Kurths J 1999 Alternative locking ratios in imperfect phase synchronization
Phys Rev Lett 82 4228ndash31Zhao L and Macau E N 2001 A network of dynamically coupled chaotic maps for scene segmentation IEEE Trans
Neural Netw 12 1375ndash85
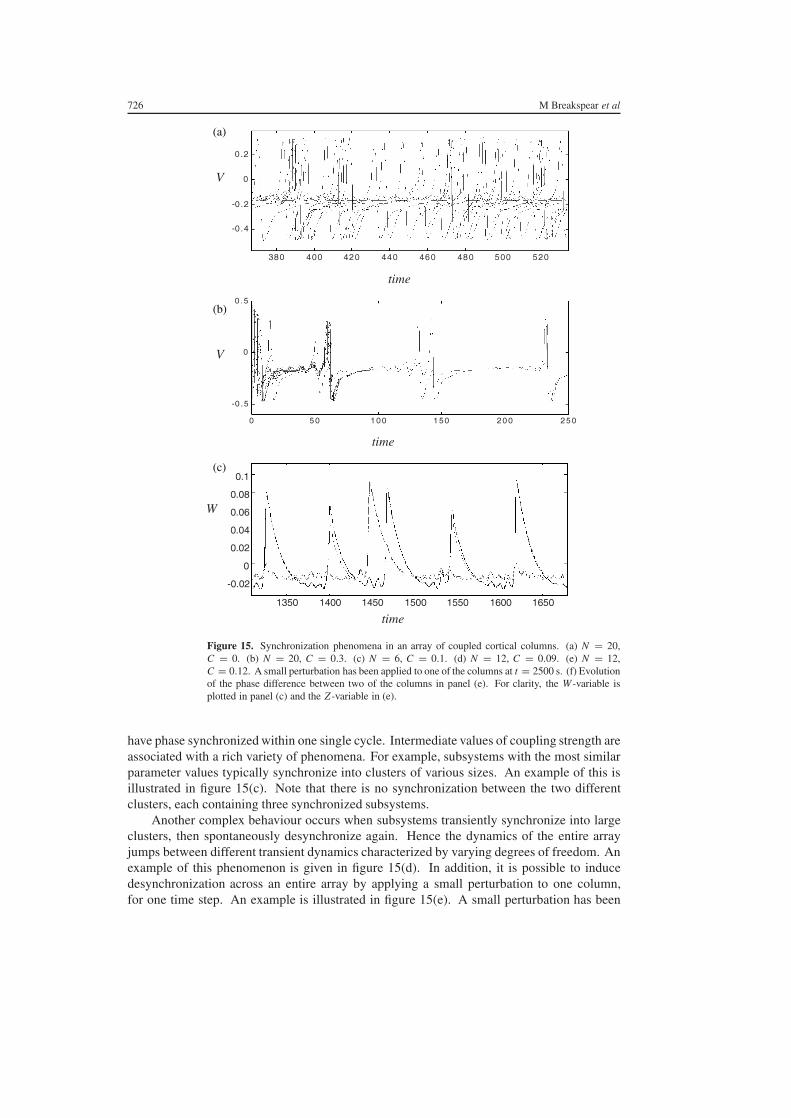
726 M Breakspear et al
(b)
(c)
(a)
Figure 15 Synchronization phenomena in an array of coupled cortical columns (a) N = 20C = 0 (b) N = 20 C = 03 (c) N = 6 C = 01 (d) N = 12 C = 009 (e) N = 12C = 012 A small perturbation has been applied to one of the columns at t = 2500 s (f) Evolutionof the phase difference between two of the columns in panel (e) For clarity the W -variable isplotted in panel (c) and the Z -variable in (e)
have phase synchronized within one single cycle Intermediate values of coupling strength areassociated with a rich variety of phenomena For example subsystems with the most similarparameter values typically synchronize into clusters of various sizes An example of this isillustrated in figure 15(c) Note that there is no synchronization between the two differentclusters each containing three synchronized subsystems
Another complex behaviour occurs when subsystems transiently synchronize into largeclusters then spontaneously desynchronize again Hence the dynamics of the entire arrayjumps between different transient dynamics characterized by varying degrees of freedom Anexample of this phenomenon is given in figure 15(d) In addition it is possible to inducedesynchronization across an entire array by applying a small perturbation to one columnfor one time step An example is illustrated in figure 15(e) A small perturbation has been
Synchronization in a model of neuronal dynamics 727
Figure 15 (Continued)
added to one column after 2500 time units It can be seen that the entire array subsequentlydesynchronizes This illustrates the sensitivity of the dynamics to external stimuli Finally it ispossible to lsquoinducersquo synchronization between selected subsystems by increasing the strength ofcoupling between them selectively This allows an extra level of complexity to be introducedinto the model For example it may allow targeted information transfer between a partialnetwork of the entire array
As discussed in section 51 a univariate signal which jumps between epochs of lowentropy (high repetition) and bursts of high entropy (high variability) can be shown to havehigh information capacity This is because the repetition of certain patterns (as in the English
728 M Breakspear et al
language) permits error detectability Bursts of high entropy act in the opposite directionto increase message variability With a multivariate signal in addition to optimizing theinformation capacity within each signal the overall information capacity can be increasedthrough the type of itinerant synchronization briefly illustrated in this section Synchronizationacross an entire array increases the message reliability (since a single lsquoerrant letterrsquo in onesignal can be easily detected) Conversely desynchronization into multiple clusters increasesmessage variability since each cluster will produce independent information Hence a neuralarray which switches dynamically between a range of cluster configurations can be seen asadaptive in the sense that high synchronous states can be expressed when error detectabilityis a priority (such as during quick and reliable execution of a familiar task) but the array canrapidly desynchronize if variability becomes salient (for example if the environment suddenlybecomes novel) A formal operationalized demonstration of this principle is to be the subjectof future work
6 Discussion
In this paper we present a dynamical model for activity in an array of weakly coupledlocal neural assemblies The model incorporates the fundamental principles governing cellmembrane potential as determined by voltage- and ligand-gated ion channels In this wayit allows a reasonable amount of physiological complexity to be incorporated includingsynaptic activation and receptor subtypeswithout recourse to overwhelming detail Numericalanalysis of this model reveals a transition from independent oscillations to stable chaoticsynchronization depending on the balance of local versus long-range connectivity andbiophysical heterogeneity These excitatory couplings are subject to ascending subcorticalneuromodulation arising principally from monoaminergic nuclei such as Locus Coeruleus(Silberstein 1995) Neuromodulation of extrinsic connections was incorporated into the modelvia a coupling parameter C The stability of the synchronization is determined by the valueof the largest transverse Lyapunov exponent which in turn depends on the coupling parameterC If the coupling is weakened with the result that this exponent becomes positive a blowoutbifurcation occurs and independent oscillations re-emerge In the vicinity of the transitionnon-linear desynchronizations with complex temporal and spatial properties are observed
It is pleasing that complex and itinerant behaviour reminiscent of real neuronaldynamics emerges through such a simple manipulation of a single dynamic control parameterFurthermore the ubiquitous projections of classical neuromodulatory systems in real brainssuggest a clear biological mechanism by which the brain could self-organize throughmoderating the complexity of its own dynamics There are many reasons that the complicateddynamics of the sort demonstrated in this paper may be neurobiologically important Wemention four
First the presence of multiple clusters of phase-synchronized subsystems as discussedabove may be a uniquely non-linear mechanism facilitating the lsquocarriagersquo and distinction ofmultiple concurrent dynamic cell assemblies (and hence cognitive processing) This may beimportant for perceptual grouping binding and transitions as suggested by others (eg von derMalsburg 1985 Gray and Singer 1989)
Second the facility to decouple and reconfigure dynamic cell assemblies througha neuromodulatory control parameter may be important for avoiding low-entropysynchronization manifolds In the absence of itinerancy synchronization would renderneuronal dynamics incapable of representing changes in sensory input or indeed cognitive stateFor example in the model it was observed that small perturbations to one column were ableto induce a global change in neuronal dynamics by disrupting a state of weak synchronization
Synchronization in a model of neuronal dynamics 729
Inevitably alternative dynamic configurations emerge epitomizing the importance of lsquolabilersquobrain dynamics (Friston 2000a)
Thirdly from a purely functionalist point of view the balance between functionalspecialization and functional integration may rely upon the sorts of mechanism modelledin this paper The brain appears to adhere to two principles of functional organization namelysegregation of function within cortical areas and sub-areas and the integration of these areasin perceptual synthesis cognition and sensorimotor integration (Tononi et al 1994 Fristonet al 1995) The balance between preserving specialized dynamics unique to each arearequired by specialization and the coupling among these areas mediating functional integrationmay be achieved by the neuromodulatory mechanisms described above Significantlythe spontaneous variation in the numbers of degrees of freedom (for example whenlow-dimensional synchronization manifolds are shadowed) imparts a dynamic characteristicto this perspective that is precluded by linear mechanisms alone
Finally from a mechanistic perspective complexity and diversity of brain dynamics arerequired for selection of adaptive responses (Friston 2000b) There are many examples ofselective mechanisms operating within the brain All can be seen as performing an ascenton some objective function or fitness landscape These mechanisms require complex anditinerant dynamics to facilitate a thorough exploration of the landscape These landscapesmay range from those involved in perception (where the landscape is defined by predictionerror see eg Rao and Ballard (1998)) to those involved in value learning (where thelandscape is determined by the outcome of a behaviour eg in temporal difference models (seeeg Sutton and Barto 1990)) In short complexity is necessary to maintain an information-rich environment in which the brain can respond to sensory inputs An essential componentof this is the maintenance of diversity which is a prerequisite for selectionist mechanismsat many scales (Edelman 1993) Understanding the mechanistic basis of complex dynamicsas measured electrophysiologically may well depend on the sort of biologically plausiblemodels used in this paper This understanding may be important for both basic and clinicalneuroscience (eg Tass 2002)
The present study supplements and advances on other research in this field The studyof complex phase synchronization and clustering in neural oscillator networks similar tothe present model has previously been the subject of formal mathematical study Forexample Terman and Wang (1995) studied synchronization of blocks of neural oscillatorsthat were subject to perturbation by connected lsquostimulirsquo Somers and Kopell (1995) studiedthe emergence of both synchronization and travelling waves in networks of coupled neuraloscillators In the work of Terman and Wang the local dynamics were based on neurallsquocaricaturersquo models such as the FitzHughndashNagumo equations and are hence more abstract thanthe current study The approach of Somers and Kopell only required that the local dynamicsexhibit fast and relaxation phases and is hence more abstract still In both studies onlyperiodic dynamics are studied Complex patterns of chaos synchronization in neural arrayshave been characterized by nonlinear methods (Kaneko 1998) and their putative contributionto both learning (Nakajima and Ueda 1996) and perception (Zhao and Macau 2001 Breakspear2001) explored However these studies employed models such as the logistic equation andtent map which are quite abstract in comparison to the present study On the other handHansel and Sompolinsky (1996) studied the role of chaos synchronization in a model ofvisual perception in a cortical hypercolumn using a very similar approach to that adoptedhere but with the additional complexity that synaptic delays were incorporated However inthis and earlier work (Hansel and Sompolinsky 1992) only linear measures of dynamics andsynchrony (specifically the auto- and cross-correlation functions) were employed The analysisof chaotic synchrony in the study of Larter et al (1999) was based only upon observation of
730 M Breakspear et al
the Euclidean difference between variables from two nodes within a larger array Hence thesignificance of the present work is that it provides a quantitative nonlinear analysis of complexchaos synchronization in a biophysical model of cortical dynamics Such research is requiredin order to bridge the gap between quantitative nonlinear research into synchronization andcomputation in abstract models of neural arrays and qualitative or linear analysis in morebiophysically plausible models
Acknowledgments
The authors would like to thank Peter Robinson for assistance with the manuscript and R Larterfor helpful advice MB is a recipient of a NSW Institute of Psychiatry Research Grant anda University of Sydney SESQUI postdoctoral fellowship KJF is supported by the WellcomeTrust
References
Abarbanel H Rulkov N and Sushchik M 1996 Generalised synchronization of chaos the auxiliary system approachPhys Rev E 53 4528ndash35
Afraimovich V Verichev N and Rabinovich M 1986 Stochastic synchronization of oscillation in dissipative systemsRadiophys Quantum Electron 29 795ndash801
Breakspear M 2001 Perception of odours by a nonlinear model of the olfactory bulb Int J Neural Syst 11 101ndash24Breakspear M 2002 Nonlinear phase desynchronisation in human electroencephalographic data Human Brain Mapping
15 175ndash98Breakspear M and Terry J R 2002a Nonlinear interdependence in neural systems motivation theory and relevance
Int J Neurosci 112 1163ndash84Breakspear M and Terry J R 2002b Detection and description of nonlinear interdependence in normal multichannel
human EEG Clin Neurophysiol 113 735ndash53Breakspear M and Terry J R 2002c Topographic organization of nonlinear interdependence in multichannel human
EEG Neuroimage 16 822ndash35Destexhe A Rudolph M Fellous J M and Sejnowski T J 2001 Fluctuating synaptic conductances recreate in vivo-like
activity in neocortical neurons Neuroscience 107 13ndash24Douglas R J Mahowald M A and Martin K A 1996 Microarchitecture of neocortical columns Brain TheorymdashBiological
Basis and Computational Principles ed A Aertsen and V Braitenberg (Amsterdam Elsevier)Eckhorn R Bauer R Jordan W Brosch M Kruse W Munk M and Reitboeck H J 1988 Coherent oscillations a
mechanism of feature linking in the visual cortex Multiple electrode and correlation analyses in the cat BiolCybern 60 121ndash30
Edelman G M 1993 Neural Darwinism selection and reentrant signalling in higher brain function Neuron 10 115ndash25Engel A K Kreiter A K Konig P and Singer W 1991 Synchronization of oscillatory neuronal responses between
striate and extrastriate visual cortical areas of the cat Proc Natl Acad Sci USA 88 6048ndash52Friston K J 2000a The labile brain I Neuronal transients and nonlinear coupling Phil Trans R Soc B 355 215ndash36Friston K J 2000b The labile brain II Transients complexity and selection Phil Trans R Soc B 355 237ndash52Friston K J Tononi G Sporns O and Edelman G 1995 Characterising the complexity of neuronal interactions Human
Brain Mapping 3 302ndash14Fujisaka H and Yamada T 1983 Stability theory of synchronized motion in coupled-oscillator system Prog Theor
Phys 69 32ndash47Gatlin L 1972 Information Theory and the Living System (New York Columbia University Press)Gray C M Konig P Engel A and Singer W 1989 Oscillatory responses in cat visual cortex exhibit inter-columnar
synchronization which reflects global stimulus properties Science 338 334ndash7Gray C M and Singer W 1989 Stimulus specific neuronal oscillations in orientation columns of cat visual cortex Proc
Natl Acad Sci USA 86 1698ndash702Haig A R Gordon E Wright J J Meares R A and Bahramali H 2000 Synchronous cortical gamma-band activity in
task-relevant cognition Neuroreport 11 669ndash75Hansel D and Sompolinsky H 1992 Synchronization and computation in a chaotic neural network Phys Rev Lett 68
718ndash21
Synchronization in a model of neuronal dynamics 731
Hansel D and Sompolinsky H 1996 Chaos and synchrony in a model of a hypercolumn in visual cortex J ComputNeurosci 3 7ndash4
Heagy J Carroll T and Pecora L 1998 Desynchronization by periodic orbits Phys Rev E 57 R1253ndash6Hunt B Ott E and Yorke J 1997 Differentiable synchronization of chaos Phys Rev E 55 4029ndash34Josic K 1998 Invariant manifolds and synchronization of coupled dynamical systems Phys Rev Lett 80 3053ndash6Kandel E Schwartz J and Jessell T 2000 Principles of Neural Science (New York McGraw-Hill)Kaneko K 1998 On the strength of attractors in a high-dimensional system Physica D 124 322ndash44Kocarev L Parlitz U and Brown R 2000 Robust synchronization of chaotic systems Phys Rev E 61 3716ndash20Larter R Speelman B and Worth R M 1999 A coupled ordinary differential equation lattice model for the simulation
of epileptic seizures Chaos 9 795ndash804Lumer E Edelman G and Tononi G 1997a Neural dynamics in a model of the thalamocortical system I Layers loops
and the emergence of fast synchronous oscillations Cereb Cortex 7 207ndash27Lumer E Edelman G and Tononi G 1997b Neural dynamics in a model of the thalamocortical system II The role of
neural synchrony tested through perturbations of spike timing Cereb Cortex 7 228ndash36Magleby K 1987 Short-term modulation at synapses Synaptic Function ed G M Edelman W E Gall and
W M Cowan (New York Wiley)Maistrenko Y Maistrenko V Popovich A and Mosekilde E 1998 Transverse instability and riddled basins in a system
of two coupled logistic maps Phys Rev E 57 2713ndash24Morris C and Lecar H 1981 Voltage oscillations in the barnacle giant muscle fiber Biophys J 35 193ndash213Mountcastle V B 1997 The columnar organization of the neocortex Brain 120 701ndash22Mountcastle V B 1998 The Cerebral Cortex (Cambridge MA Harvard University Press)Nakajima H and Ueda Y 1996 Riddled basins of the optimal states in learning dynamical systems Physica D 99 35ndash44Nunez P L 1995 Neocortical Dynamics and Human Brain Rhythms (Oxford Oxford University Press)Osipov G V Pikovsky A S Rosenblum M G and Kurths J 1997 Phase synchronization effects in a lattice of nonidentical
Rossler oscillators Phys Rev E 55 2353ndash61Ott E and Sommerer J 1994 Blowout bifurcations the occurrence of riddled basins and onndashoff intermittency Phys
Lett A 188 39ndash47Pecora L and Carroll T 1990 Synchronization in chaotic systems Phys Rev Lett 64 821ndash4Pikovsky A and Grassberger P 1991 Symmetry breaking bifurcation for coupled chaotic attractors J Phys A Math
Gen 24 4587ndash97Rao R P and Ballard D H 1998 Predictive coding in the visual cortex a functional interpretation of some extra-classical
receptive field effects Nature Neurosci 2 79ndash87Robinson P A Wright J J and Rennie C R 1998 Synchronous oscillations in the cerebral cortex Phys Rev E 57
4578ndash88Rodriguez E George N Lachauz J P Martinerie J Renault B and Varela F J 1999 Perceptionrsquos shadow long distance
synchronization of human brain activity Nature 397 430ndash3Rosenblum M Pikovsky A and Kurths J 1996 Phase synchronization of chaotic oscillators Phys Rev Lett 76 1804ndash7Rosenblum M Pikovsky A and Kurths J 1997 From phase to lag synchronization in coupled chaotic oscillators Phys
Rev Lett 78 4193ndash6Rulkov N and Sushchik M 1997 Robustness of synchronized chaotic oscillations Int J Bifurc Chaos 7 625ndash43Rulkov N Sushchik M Tsimring L and Abarbanel H 1995 Generalized synchronization of chaos in unidirectionally
coupled chaotic systems Phys Rev E 51 980ndash94Schiff S So P Chang T Burke R and Sauer T 1996 Detecting dynamical interdependence and generalized synchrony
through mutual prediction in a neural ensemble Phys Rev E 54 6708ndash24Silberstein R B 1995 Neuromodulations of neocortical dynamics Neocortical Dynamics and Human EEG Rhythms
ed P L Nunez (Oxford Oxford University Press)Singer W 1995 Putative functions of temporal correlations in neocortical processing Large-Scale Neuronal Theories
of the Brain ed C Koch and J Davis (London MIT Press)Somers D and Kopell N 1995 Waves and synchrony in networks of oscillators of relaxation and non-relaxation type
Physica D 89 169ndash83Sutton R S and Barto A G 1990 Time derivative models of Pavlovian reinforcement Learning and Computational
Neuroscience Foundations of Adaptive Networks ed M Gabriel and J Moore (Cambridge MA MIT Press) pp497ndash538
Szentagothai J 1983 The modular architectonic principle of neural centers Rev Physiol Biochem Pharmacol 9811ndash61
Tallon-Baudry C Betrand O and Fischer C 2001 Oscillatory synchrony between human extrastriate areas during visualshort-term memory maintenance J Neurosci 21 RC177(1ndash5)
Tass P A 2002 Desynchronization of brain rhythms with soft phase-resetting techniques Biol Cybern 87 102ndash15
732 M Breakspear et al
Tass P Rosenblum M Weule J Kurths J Pikovsky A Volkmann J Schnitzler A and Freund H 1998 Detection of nmphase locking from noisy data application to magnetoencephalography Phys Rev Lett 81 3291ndash4
Terman D and Wang D 1995 Global competition and local cooperation in a network of neural oscillators Physica D81 148ndash76
Terry J and Breakspear M 2003 An improved algorithm for the detection of dynamical interdependence in bivariatetime-series Biol Cybern 88 129ndash36
Tononi G Sporns O and Edelman G M 1994 A measure for brain complexity relating functional segregation andintegration in the nervous system Proc Natl Acad Sci USA 91 5033ndash7
von der Malsburg C 1985 Nervous structures with dynamical links Phys Chem 89 703ndash10Wang X 1991 Period-doublings to chaos in a simple neural network an analytic proof Complex Syst 5 425ndash41Wennekers T and Pasemann F 1996 Synchronous chaos in high dimensional modular neural networks Int J Bifurc
Chaos 6 2055ndash67Zaks M A Parks E H Rosenblum M G and Kurths J 1999 Alternative locking ratios in imperfect phase synchronization
Phys Rev Lett 82 4228ndash31Zhao L and Macau E N 2001 A network of dynamically coupled chaotic maps for scene segmentation IEEE Trans
Neural Netw 12 1375ndash85
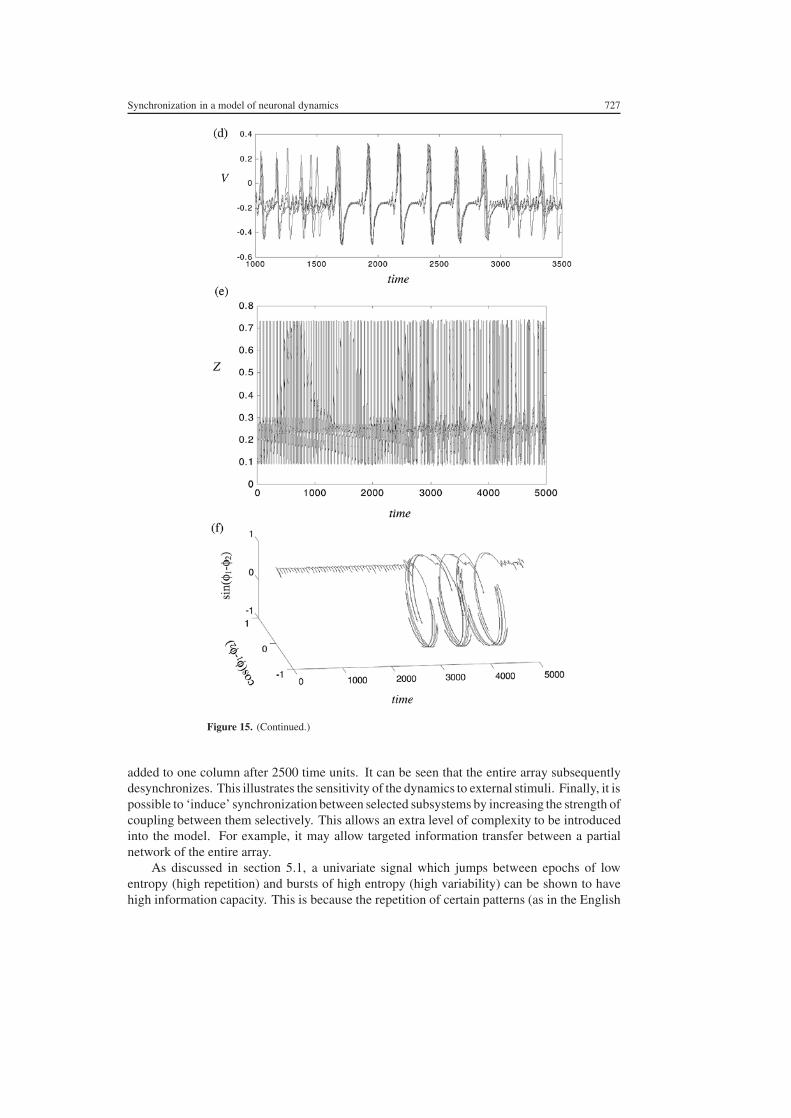
Synchronization in a model of neuronal dynamics 727
Figure 15 (Continued)
added to one column after 2500 time units It can be seen that the entire array subsequentlydesynchronizes This illustrates the sensitivity of the dynamics to external stimuli Finally it ispossible to lsquoinducersquo synchronization between selected subsystems by increasing the strength ofcoupling between them selectively This allows an extra level of complexity to be introducedinto the model For example it may allow targeted information transfer between a partialnetwork of the entire array
As discussed in section 51 a univariate signal which jumps between epochs of lowentropy (high repetition) and bursts of high entropy (high variability) can be shown to havehigh information capacity This is because the repetition of certain patterns (as in the English
728 M Breakspear et al
language) permits error detectability Bursts of high entropy act in the opposite directionto increase message variability With a multivariate signal in addition to optimizing theinformation capacity within each signal the overall information capacity can be increasedthrough the type of itinerant synchronization briefly illustrated in this section Synchronizationacross an entire array increases the message reliability (since a single lsquoerrant letterrsquo in onesignal can be easily detected) Conversely desynchronization into multiple clusters increasesmessage variability since each cluster will produce independent information Hence a neuralarray which switches dynamically between a range of cluster configurations can be seen asadaptive in the sense that high synchronous states can be expressed when error detectabilityis a priority (such as during quick and reliable execution of a familiar task) but the array canrapidly desynchronize if variability becomes salient (for example if the environment suddenlybecomes novel) A formal operationalized demonstration of this principle is to be the subjectof future work
6 Discussion
In this paper we present a dynamical model for activity in an array of weakly coupledlocal neural assemblies The model incorporates the fundamental principles governing cellmembrane potential as determined by voltage- and ligand-gated ion channels In this wayit allows a reasonable amount of physiological complexity to be incorporated includingsynaptic activation and receptor subtypeswithout recourse to overwhelming detail Numericalanalysis of this model reveals a transition from independent oscillations to stable chaoticsynchronization depending on the balance of local versus long-range connectivity andbiophysical heterogeneity These excitatory couplings are subject to ascending subcorticalneuromodulation arising principally from monoaminergic nuclei such as Locus Coeruleus(Silberstein 1995) Neuromodulation of extrinsic connections was incorporated into the modelvia a coupling parameter C The stability of the synchronization is determined by the valueof the largest transverse Lyapunov exponent which in turn depends on the coupling parameterC If the coupling is weakened with the result that this exponent becomes positive a blowoutbifurcation occurs and independent oscillations re-emerge In the vicinity of the transitionnon-linear desynchronizations with complex temporal and spatial properties are observed
It is pleasing that complex and itinerant behaviour reminiscent of real neuronaldynamics emerges through such a simple manipulation of a single dynamic control parameterFurthermore the ubiquitous projections of classical neuromodulatory systems in real brainssuggest a clear biological mechanism by which the brain could self-organize throughmoderating the complexity of its own dynamics There are many reasons that the complicateddynamics of the sort demonstrated in this paper may be neurobiologically important Wemention four
First the presence of multiple clusters of phase-synchronized subsystems as discussedabove may be a uniquely non-linear mechanism facilitating the lsquocarriagersquo and distinction ofmultiple concurrent dynamic cell assemblies (and hence cognitive processing) This may beimportant for perceptual grouping binding and transitions as suggested by others (eg von derMalsburg 1985 Gray and Singer 1989)
Second the facility to decouple and reconfigure dynamic cell assemblies througha neuromodulatory control parameter may be important for avoiding low-entropysynchronization manifolds In the absence of itinerancy synchronization would renderneuronal dynamics incapable of representing changes in sensory input or indeed cognitive stateFor example in the model it was observed that small perturbations to one column were ableto induce a global change in neuronal dynamics by disrupting a state of weak synchronization
Synchronization in a model of neuronal dynamics 729
Inevitably alternative dynamic configurations emerge epitomizing the importance of lsquolabilersquobrain dynamics (Friston 2000a)
Thirdly from a purely functionalist point of view the balance between functionalspecialization and functional integration may rely upon the sorts of mechanism modelledin this paper The brain appears to adhere to two principles of functional organization namelysegregation of function within cortical areas and sub-areas and the integration of these areasin perceptual synthesis cognition and sensorimotor integration (Tononi et al 1994 Fristonet al 1995) The balance between preserving specialized dynamics unique to each arearequired by specialization and the coupling among these areas mediating functional integrationmay be achieved by the neuromodulatory mechanisms described above Significantlythe spontaneous variation in the numbers of degrees of freedom (for example whenlow-dimensional synchronization manifolds are shadowed) imparts a dynamic characteristicto this perspective that is precluded by linear mechanisms alone
Finally from a mechanistic perspective complexity and diversity of brain dynamics arerequired for selection of adaptive responses (Friston 2000b) There are many examples ofselective mechanisms operating within the brain All can be seen as performing an ascenton some objective function or fitness landscape These mechanisms require complex anditinerant dynamics to facilitate a thorough exploration of the landscape These landscapesmay range from those involved in perception (where the landscape is defined by predictionerror see eg Rao and Ballard (1998)) to those involved in value learning (where thelandscape is determined by the outcome of a behaviour eg in temporal difference models (seeeg Sutton and Barto 1990)) In short complexity is necessary to maintain an information-rich environment in which the brain can respond to sensory inputs An essential componentof this is the maintenance of diversity which is a prerequisite for selectionist mechanismsat many scales (Edelman 1993) Understanding the mechanistic basis of complex dynamicsas measured electrophysiologically may well depend on the sort of biologically plausiblemodels used in this paper This understanding may be important for both basic and clinicalneuroscience (eg Tass 2002)
The present study supplements and advances on other research in this field The studyof complex phase synchronization and clustering in neural oscillator networks similar tothe present model has previously been the subject of formal mathematical study Forexample Terman and Wang (1995) studied synchronization of blocks of neural oscillatorsthat were subject to perturbation by connected lsquostimulirsquo Somers and Kopell (1995) studiedthe emergence of both synchronization and travelling waves in networks of coupled neuraloscillators In the work of Terman and Wang the local dynamics were based on neurallsquocaricaturersquo models such as the FitzHughndashNagumo equations and are hence more abstract thanthe current study The approach of Somers and Kopell only required that the local dynamicsexhibit fast and relaxation phases and is hence more abstract still In both studies onlyperiodic dynamics are studied Complex patterns of chaos synchronization in neural arrayshave been characterized by nonlinear methods (Kaneko 1998) and their putative contributionto both learning (Nakajima and Ueda 1996) and perception (Zhao and Macau 2001 Breakspear2001) explored However these studies employed models such as the logistic equation andtent map which are quite abstract in comparison to the present study On the other handHansel and Sompolinsky (1996) studied the role of chaos synchronization in a model ofvisual perception in a cortical hypercolumn using a very similar approach to that adoptedhere but with the additional complexity that synaptic delays were incorporated However inthis and earlier work (Hansel and Sompolinsky 1992) only linear measures of dynamics andsynchrony (specifically the auto- and cross-correlation functions) were employed The analysisof chaotic synchrony in the study of Larter et al (1999) was based only upon observation of
730 M Breakspear et al
the Euclidean difference between variables from two nodes within a larger array Hence thesignificance of the present work is that it provides a quantitative nonlinear analysis of complexchaos synchronization in a biophysical model of cortical dynamics Such research is requiredin order to bridge the gap between quantitative nonlinear research into synchronization andcomputation in abstract models of neural arrays and qualitative or linear analysis in morebiophysically plausible models
Acknowledgments
The authors would like to thank Peter Robinson for assistance with the manuscript and R Larterfor helpful advice MB is a recipient of a NSW Institute of Psychiatry Research Grant anda University of Sydney SESQUI postdoctoral fellowship KJF is supported by the WellcomeTrust
References
Abarbanel H Rulkov N and Sushchik M 1996 Generalised synchronization of chaos the auxiliary system approachPhys Rev E 53 4528ndash35
Afraimovich V Verichev N and Rabinovich M 1986 Stochastic synchronization of oscillation in dissipative systemsRadiophys Quantum Electron 29 795ndash801
Breakspear M 2001 Perception of odours by a nonlinear model of the olfactory bulb Int J Neural Syst 11 101ndash24Breakspear M 2002 Nonlinear phase desynchronisation in human electroencephalographic data Human Brain Mapping
15 175ndash98Breakspear M and Terry J R 2002a Nonlinear interdependence in neural systems motivation theory and relevance
Int J Neurosci 112 1163ndash84Breakspear M and Terry J R 2002b Detection and description of nonlinear interdependence in normal multichannel
human EEG Clin Neurophysiol 113 735ndash53Breakspear M and Terry J R 2002c Topographic organization of nonlinear interdependence in multichannel human
EEG Neuroimage 16 822ndash35Destexhe A Rudolph M Fellous J M and Sejnowski T J 2001 Fluctuating synaptic conductances recreate in vivo-like
activity in neocortical neurons Neuroscience 107 13ndash24Douglas R J Mahowald M A and Martin K A 1996 Microarchitecture of neocortical columns Brain TheorymdashBiological
Basis and Computational Principles ed A Aertsen and V Braitenberg (Amsterdam Elsevier)Eckhorn R Bauer R Jordan W Brosch M Kruse W Munk M and Reitboeck H J 1988 Coherent oscillations a
mechanism of feature linking in the visual cortex Multiple electrode and correlation analyses in the cat BiolCybern 60 121ndash30
Edelman G M 1993 Neural Darwinism selection and reentrant signalling in higher brain function Neuron 10 115ndash25Engel A K Kreiter A K Konig P and Singer W 1991 Synchronization of oscillatory neuronal responses between
striate and extrastriate visual cortical areas of the cat Proc Natl Acad Sci USA 88 6048ndash52Friston K J 2000a The labile brain I Neuronal transients and nonlinear coupling Phil Trans R Soc B 355 215ndash36Friston K J 2000b The labile brain II Transients complexity and selection Phil Trans R Soc B 355 237ndash52Friston K J Tononi G Sporns O and Edelman G 1995 Characterising the complexity of neuronal interactions Human
Brain Mapping 3 302ndash14Fujisaka H and Yamada T 1983 Stability theory of synchronized motion in coupled-oscillator system Prog Theor
Phys 69 32ndash47Gatlin L 1972 Information Theory and the Living System (New York Columbia University Press)Gray C M Konig P Engel A and Singer W 1989 Oscillatory responses in cat visual cortex exhibit inter-columnar
synchronization which reflects global stimulus properties Science 338 334ndash7Gray C M and Singer W 1989 Stimulus specific neuronal oscillations in orientation columns of cat visual cortex Proc
Natl Acad Sci USA 86 1698ndash702Haig A R Gordon E Wright J J Meares R A and Bahramali H 2000 Synchronous cortical gamma-band activity in
task-relevant cognition Neuroreport 11 669ndash75Hansel D and Sompolinsky H 1992 Synchronization and computation in a chaotic neural network Phys Rev Lett 68
718ndash21
Synchronization in a model of neuronal dynamics 731
Hansel D and Sompolinsky H 1996 Chaos and synchrony in a model of a hypercolumn in visual cortex J ComputNeurosci 3 7ndash4
Heagy J Carroll T and Pecora L 1998 Desynchronization by periodic orbits Phys Rev E 57 R1253ndash6Hunt B Ott E and Yorke J 1997 Differentiable synchronization of chaos Phys Rev E 55 4029ndash34Josic K 1998 Invariant manifolds and synchronization of coupled dynamical systems Phys Rev Lett 80 3053ndash6Kandel E Schwartz J and Jessell T 2000 Principles of Neural Science (New York McGraw-Hill)Kaneko K 1998 On the strength of attractors in a high-dimensional system Physica D 124 322ndash44Kocarev L Parlitz U and Brown R 2000 Robust synchronization of chaotic systems Phys Rev E 61 3716ndash20Larter R Speelman B and Worth R M 1999 A coupled ordinary differential equation lattice model for the simulation
of epileptic seizures Chaos 9 795ndash804Lumer E Edelman G and Tononi G 1997a Neural dynamics in a model of the thalamocortical system I Layers loops
and the emergence of fast synchronous oscillations Cereb Cortex 7 207ndash27Lumer E Edelman G and Tononi G 1997b Neural dynamics in a model of the thalamocortical system II The role of
neural synchrony tested through perturbations of spike timing Cereb Cortex 7 228ndash36Magleby K 1987 Short-term modulation at synapses Synaptic Function ed G M Edelman W E Gall and
W M Cowan (New York Wiley)Maistrenko Y Maistrenko V Popovich A and Mosekilde E 1998 Transverse instability and riddled basins in a system
of two coupled logistic maps Phys Rev E 57 2713ndash24Morris C and Lecar H 1981 Voltage oscillations in the barnacle giant muscle fiber Biophys J 35 193ndash213Mountcastle V B 1997 The columnar organization of the neocortex Brain 120 701ndash22Mountcastle V B 1998 The Cerebral Cortex (Cambridge MA Harvard University Press)Nakajima H and Ueda Y 1996 Riddled basins of the optimal states in learning dynamical systems Physica D 99 35ndash44Nunez P L 1995 Neocortical Dynamics and Human Brain Rhythms (Oxford Oxford University Press)Osipov G V Pikovsky A S Rosenblum M G and Kurths J 1997 Phase synchronization effects in a lattice of nonidentical
Rossler oscillators Phys Rev E 55 2353ndash61Ott E and Sommerer J 1994 Blowout bifurcations the occurrence of riddled basins and onndashoff intermittency Phys
Lett A 188 39ndash47Pecora L and Carroll T 1990 Synchronization in chaotic systems Phys Rev Lett 64 821ndash4Pikovsky A and Grassberger P 1991 Symmetry breaking bifurcation for coupled chaotic attractors J Phys A Math
Gen 24 4587ndash97Rao R P and Ballard D H 1998 Predictive coding in the visual cortex a functional interpretation of some extra-classical
receptive field effects Nature Neurosci 2 79ndash87Robinson P A Wright J J and Rennie C R 1998 Synchronous oscillations in the cerebral cortex Phys Rev E 57
4578ndash88Rodriguez E George N Lachauz J P Martinerie J Renault B and Varela F J 1999 Perceptionrsquos shadow long distance
synchronization of human brain activity Nature 397 430ndash3Rosenblum M Pikovsky A and Kurths J 1996 Phase synchronization of chaotic oscillators Phys Rev Lett 76 1804ndash7Rosenblum M Pikovsky A and Kurths J 1997 From phase to lag synchronization in coupled chaotic oscillators Phys
Rev Lett 78 4193ndash6Rulkov N and Sushchik M 1997 Robustness of synchronized chaotic oscillations Int J Bifurc Chaos 7 625ndash43Rulkov N Sushchik M Tsimring L and Abarbanel H 1995 Generalized synchronization of chaos in unidirectionally
coupled chaotic systems Phys Rev E 51 980ndash94Schiff S So P Chang T Burke R and Sauer T 1996 Detecting dynamical interdependence and generalized synchrony
through mutual prediction in a neural ensemble Phys Rev E 54 6708ndash24Silberstein R B 1995 Neuromodulations of neocortical dynamics Neocortical Dynamics and Human EEG Rhythms
ed P L Nunez (Oxford Oxford University Press)Singer W 1995 Putative functions of temporal correlations in neocortical processing Large-Scale Neuronal Theories
of the Brain ed C Koch and J Davis (London MIT Press)Somers D and Kopell N 1995 Waves and synchrony in networks of oscillators of relaxation and non-relaxation type
Physica D 89 169ndash83Sutton R S and Barto A G 1990 Time derivative models of Pavlovian reinforcement Learning and Computational
Neuroscience Foundations of Adaptive Networks ed M Gabriel and J Moore (Cambridge MA MIT Press) pp497ndash538
Szentagothai J 1983 The modular architectonic principle of neural centers Rev Physiol Biochem Pharmacol 9811ndash61
Tallon-Baudry C Betrand O and Fischer C 2001 Oscillatory synchrony between human extrastriate areas during visualshort-term memory maintenance J Neurosci 21 RC177(1ndash5)
Tass P A 2002 Desynchronization of brain rhythms with soft phase-resetting techniques Biol Cybern 87 102ndash15
732 M Breakspear et al
Tass P Rosenblum M Weule J Kurths J Pikovsky A Volkmann J Schnitzler A and Freund H 1998 Detection of nmphase locking from noisy data application to magnetoencephalography Phys Rev Lett 81 3291ndash4
Terman D and Wang D 1995 Global competition and local cooperation in a network of neural oscillators Physica D81 148ndash76
Terry J and Breakspear M 2003 An improved algorithm for the detection of dynamical interdependence in bivariatetime-series Biol Cybern 88 129ndash36
Tononi G Sporns O and Edelman G M 1994 A measure for brain complexity relating functional segregation andintegration in the nervous system Proc Natl Acad Sci USA 91 5033ndash7
von der Malsburg C 1985 Nervous structures with dynamical links Phys Chem 89 703ndash10Wang X 1991 Period-doublings to chaos in a simple neural network an analytic proof Complex Syst 5 425ndash41Wennekers T and Pasemann F 1996 Synchronous chaos in high dimensional modular neural networks Int J Bifurc
Chaos 6 2055ndash67Zaks M A Parks E H Rosenblum M G and Kurths J 1999 Alternative locking ratios in imperfect phase synchronization
Phys Rev Lett 82 4228ndash31Zhao L and Macau E N 2001 A network of dynamically coupled chaotic maps for scene segmentation IEEE Trans
Neural Netw 12 1375ndash85

728 M Breakspear et al
language) permits error detectability Bursts of high entropy act in the opposite directionto increase message variability With a multivariate signal in addition to optimizing theinformation capacity within each signal the overall information capacity can be increasedthrough the type of itinerant synchronization briefly illustrated in this section Synchronizationacross an entire array increases the message reliability (since a single lsquoerrant letterrsquo in onesignal can be easily detected) Conversely desynchronization into multiple clusters increasesmessage variability since each cluster will produce independent information Hence a neuralarray which switches dynamically between a range of cluster configurations can be seen asadaptive in the sense that high synchronous states can be expressed when error detectabilityis a priority (such as during quick and reliable execution of a familiar task) but the array canrapidly desynchronize if variability becomes salient (for example if the environment suddenlybecomes novel) A formal operationalized demonstration of this principle is to be the subjectof future work
6 Discussion
In this paper we present a dynamical model for activity in an array of weakly coupledlocal neural assemblies The model incorporates the fundamental principles governing cellmembrane potential as determined by voltage- and ligand-gated ion channels In this wayit allows a reasonable amount of physiological complexity to be incorporated includingsynaptic activation and receptor subtypeswithout recourse to overwhelming detail Numericalanalysis of this model reveals a transition from independent oscillations to stable chaoticsynchronization depending on the balance of local versus long-range connectivity andbiophysical heterogeneity These excitatory couplings are subject to ascending subcorticalneuromodulation arising principally from monoaminergic nuclei such as Locus Coeruleus(Silberstein 1995) Neuromodulation of extrinsic connections was incorporated into the modelvia a coupling parameter C The stability of the synchronization is determined by the valueof the largest transverse Lyapunov exponent which in turn depends on the coupling parameterC If the coupling is weakened with the result that this exponent becomes positive a blowoutbifurcation occurs and independent oscillations re-emerge In the vicinity of the transitionnon-linear desynchronizations with complex temporal and spatial properties are observed
It is pleasing that complex and itinerant behaviour reminiscent of real neuronaldynamics emerges through such a simple manipulation of a single dynamic control parameterFurthermore the ubiquitous projections of classical neuromodulatory systems in real brainssuggest a clear biological mechanism by which the brain could self-organize throughmoderating the complexity of its own dynamics There are many reasons that the complicateddynamics of the sort demonstrated in this paper may be neurobiologically important Wemention four
First the presence of multiple clusters of phase-synchronized subsystems as discussedabove may be a uniquely non-linear mechanism facilitating the lsquocarriagersquo and distinction ofmultiple concurrent dynamic cell assemblies (and hence cognitive processing) This may beimportant for perceptual grouping binding and transitions as suggested by others (eg von derMalsburg 1985 Gray and Singer 1989)
Second the facility to decouple and reconfigure dynamic cell assemblies througha neuromodulatory control parameter may be important for avoiding low-entropysynchronization manifolds In the absence of itinerancy synchronization would renderneuronal dynamics incapable of representing changes in sensory input or indeed cognitive stateFor example in the model it was observed that small perturbations to one column were ableto induce a global change in neuronal dynamics by disrupting a state of weak synchronization
Synchronization in a model of neuronal dynamics 729
Inevitably alternative dynamic configurations emerge epitomizing the importance of lsquolabilersquobrain dynamics (Friston 2000a)
Thirdly from a purely functionalist point of view the balance between functionalspecialization and functional integration may rely upon the sorts of mechanism modelledin this paper The brain appears to adhere to two principles of functional organization namelysegregation of function within cortical areas and sub-areas and the integration of these areasin perceptual synthesis cognition and sensorimotor integration (Tononi et al 1994 Fristonet al 1995) The balance between preserving specialized dynamics unique to each arearequired by specialization and the coupling among these areas mediating functional integrationmay be achieved by the neuromodulatory mechanisms described above Significantlythe spontaneous variation in the numbers of degrees of freedom (for example whenlow-dimensional synchronization manifolds are shadowed) imparts a dynamic characteristicto this perspective that is precluded by linear mechanisms alone
Finally from a mechanistic perspective complexity and diversity of brain dynamics arerequired for selection of adaptive responses (Friston 2000b) There are many examples ofselective mechanisms operating within the brain All can be seen as performing an ascenton some objective function or fitness landscape These mechanisms require complex anditinerant dynamics to facilitate a thorough exploration of the landscape These landscapesmay range from those involved in perception (where the landscape is defined by predictionerror see eg Rao and Ballard (1998)) to those involved in value learning (where thelandscape is determined by the outcome of a behaviour eg in temporal difference models (seeeg Sutton and Barto 1990)) In short complexity is necessary to maintain an information-rich environment in which the brain can respond to sensory inputs An essential componentof this is the maintenance of diversity which is a prerequisite for selectionist mechanismsat many scales (Edelman 1993) Understanding the mechanistic basis of complex dynamicsas measured electrophysiologically may well depend on the sort of biologically plausiblemodels used in this paper This understanding may be important for both basic and clinicalneuroscience (eg Tass 2002)
The present study supplements and advances on other research in this field The studyof complex phase synchronization and clustering in neural oscillator networks similar tothe present model has previously been the subject of formal mathematical study Forexample Terman and Wang (1995) studied synchronization of blocks of neural oscillatorsthat were subject to perturbation by connected lsquostimulirsquo Somers and Kopell (1995) studiedthe emergence of both synchronization and travelling waves in networks of coupled neuraloscillators In the work of Terman and Wang the local dynamics were based on neurallsquocaricaturersquo models such as the FitzHughndashNagumo equations and are hence more abstract thanthe current study The approach of Somers and Kopell only required that the local dynamicsexhibit fast and relaxation phases and is hence more abstract still In both studies onlyperiodic dynamics are studied Complex patterns of chaos synchronization in neural arrayshave been characterized by nonlinear methods (Kaneko 1998) and their putative contributionto both learning (Nakajima and Ueda 1996) and perception (Zhao and Macau 2001 Breakspear2001) explored However these studies employed models such as the logistic equation andtent map which are quite abstract in comparison to the present study On the other handHansel and Sompolinsky (1996) studied the role of chaos synchronization in a model ofvisual perception in a cortical hypercolumn using a very similar approach to that adoptedhere but with the additional complexity that synaptic delays were incorporated However inthis and earlier work (Hansel and Sompolinsky 1992) only linear measures of dynamics andsynchrony (specifically the auto- and cross-correlation functions) were employed The analysisof chaotic synchrony in the study of Larter et al (1999) was based only upon observation of
730 M Breakspear et al
the Euclidean difference between variables from two nodes within a larger array Hence thesignificance of the present work is that it provides a quantitative nonlinear analysis of complexchaos synchronization in a biophysical model of cortical dynamics Such research is requiredin order to bridge the gap between quantitative nonlinear research into synchronization andcomputation in abstract models of neural arrays and qualitative or linear analysis in morebiophysically plausible models
Acknowledgments
The authors would like to thank Peter Robinson for assistance with the manuscript and R Larterfor helpful advice MB is a recipient of a NSW Institute of Psychiatry Research Grant anda University of Sydney SESQUI postdoctoral fellowship KJF is supported by the WellcomeTrust
References
Abarbanel H Rulkov N and Sushchik M 1996 Generalised synchronization of chaos the auxiliary system approachPhys Rev E 53 4528ndash35
Afraimovich V Verichev N and Rabinovich M 1986 Stochastic synchronization of oscillation in dissipative systemsRadiophys Quantum Electron 29 795ndash801
Breakspear M 2001 Perception of odours by a nonlinear model of the olfactory bulb Int J Neural Syst 11 101ndash24Breakspear M 2002 Nonlinear phase desynchronisation in human electroencephalographic data Human Brain Mapping
15 175ndash98Breakspear M and Terry J R 2002a Nonlinear interdependence in neural systems motivation theory and relevance
Int J Neurosci 112 1163ndash84Breakspear M and Terry J R 2002b Detection and description of nonlinear interdependence in normal multichannel
human EEG Clin Neurophysiol 113 735ndash53Breakspear M and Terry J R 2002c Topographic organization of nonlinear interdependence in multichannel human
EEG Neuroimage 16 822ndash35Destexhe A Rudolph M Fellous J M and Sejnowski T J 2001 Fluctuating synaptic conductances recreate in vivo-like
activity in neocortical neurons Neuroscience 107 13ndash24Douglas R J Mahowald M A and Martin K A 1996 Microarchitecture of neocortical columns Brain TheorymdashBiological
Basis and Computational Principles ed A Aertsen and V Braitenberg (Amsterdam Elsevier)Eckhorn R Bauer R Jordan W Brosch M Kruse W Munk M and Reitboeck H J 1988 Coherent oscillations a
mechanism of feature linking in the visual cortex Multiple electrode and correlation analyses in the cat BiolCybern 60 121ndash30
Edelman G M 1993 Neural Darwinism selection and reentrant signalling in higher brain function Neuron 10 115ndash25Engel A K Kreiter A K Konig P and Singer W 1991 Synchronization of oscillatory neuronal responses between
striate and extrastriate visual cortical areas of the cat Proc Natl Acad Sci USA 88 6048ndash52Friston K J 2000a The labile brain I Neuronal transients and nonlinear coupling Phil Trans R Soc B 355 215ndash36Friston K J 2000b The labile brain II Transients complexity and selection Phil Trans R Soc B 355 237ndash52Friston K J Tononi G Sporns O and Edelman G 1995 Characterising the complexity of neuronal interactions Human
Brain Mapping 3 302ndash14Fujisaka H and Yamada T 1983 Stability theory of synchronized motion in coupled-oscillator system Prog Theor
Phys 69 32ndash47Gatlin L 1972 Information Theory and the Living System (New York Columbia University Press)Gray C M Konig P Engel A and Singer W 1989 Oscillatory responses in cat visual cortex exhibit inter-columnar
synchronization which reflects global stimulus properties Science 338 334ndash7Gray C M and Singer W 1989 Stimulus specific neuronal oscillations in orientation columns of cat visual cortex Proc
Natl Acad Sci USA 86 1698ndash702Haig A R Gordon E Wright J J Meares R A and Bahramali H 2000 Synchronous cortical gamma-band activity in
task-relevant cognition Neuroreport 11 669ndash75Hansel D and Sompolinsky H 1992 Synchronization and computation in a chaotic neural network Phys Rev Lett 68
718ndash21
Synchronization in a model of neuronal dynamics 731
Hansel D and Sompolinsky H 1996 Chaos and synchrony in a model of a hypercolumn in visual cortex J ComputNeurosci 3 7ndash4
Heagy J Carroll T and Pecora L 1998 Desynchronization by periodic orbits Phys Rev E 57 R1253ndash6Hunt B Ott E and Yorke J 1997 Differentiable synchronization of chaos Phys Rev E 55 4029ndash34Josic K 1998 Invariant manifolds and synchronization of coupled dynamical systems Phys Rev Lett 80 3053ndash6Kandel E Schwartz J and Jessell T 2000 Principles of Neural Science (New York McGraw-Hill)Kaneko K 1998 On the strength of attractors in a high-dimensional system Physica D 124 322ndash44Kocarev L Parlitz U and Brown R 2000 Robust synchronization of chaotic systems Phys Rev E 61 3716ndash20Larter R Speelman B and Worth R M 1999 A coupled ordinary differential equation lattice model for the simulation
of epileptic seizures Chaos 9 795ndash804Lumer E Edelman G and Tononi G 1997a Neural dynamics in a model of the thalamocortical system I Layers loops
and the emergence of fast synchronous oscillations Cereb Cortex 7 207ndash27Lumer E Edelman G and Tononi G 1997b Neural dynamics in a model of the thalamocortical system II The role of
neural synchrony tested through perturbations of spike timing Cereb Cortex 7 228ndash36Magleby K 1987 Short-term modulation at synapses Synaptic Function ed G M Edelman W E Gall and
W M Cowan (New York Wiley)Maistrenko Y Maistrenko V Popovich A and Mosekilde E 1998 Transverse instability and riddled basins in a system
of two coupled logistic maps Phys Rev E 57 2713ndash24Morris C and Lecar H 1981 Voltage oscillations in the barnacle giant muscle fiber Biophys J 35 193ndash213Mountcastle V B 1997 The columnar organization of the neocortex Brain 120 701ndash22Mountcastle V B 1998 The Cerebral Cortex (Cambridge MA Harvard University Press)Nakajima H and Ueda Y 1996 Riddled basins of the optimal states in learning dynamical systems Physica D 99 35ndash44Nunez P L 1995 Neocortical Dynamics and Human Brain Rhythms (Oxford Oxford University Press)Osipov G V Pikovsky A S Rosenblum M G and Kurths J 1997 Phase synchronization effects in a lattice of nonidentical
Rossler oscillators Phys Rev E 55 2353ndash61Ott E and Sommerer J 1994 Blowout bifurcations the occurrence of riddled basins and onndashoff intermittency Phys
Lett A 188 39ndash47Pecora L and Carroll T 1990 Synchronization in chaotic systems Phys Rev Lett 64 821ndash4Pikovsky A and Grassberger P 1991 Symmetry breaking bifurcation for coupled chaotic attractors J Phys A Math
Gen 24 4587ndash97Rao R P and Ballard D H 1998 Predictive coding in the visual cortex a functional interpretation of some extra-classical
receptive field effects Nature Neurosci 2 79ndash87Robinson P A Wright J J and Rennie C R 1998 Synchronous oscillations in the cerebral cortex Phys Rev E 57
4578ndash88Rodriguez E George N Lachauz J P Martinerie J Renault B and Varela F J 1999 Perceptionrsquos shadow long distance
synchronization of human brain activity Nature 397 430ndash3Rosenblum M Pikovsky A and Kurths J 1996 Phase synchronization of chaotic oscillators Phys Rev Lett 76 1804ndash7Rosenblum M Pikovsky A and Kurths J 1997 From phase to lag synchronization in coupled chaotic oscillators Phys
Rev Lett 78 4193ndash6Rulkov N and Sushchik M 1997 Robustness of synchronized chaotic oscillations Int J Bifurc Chaos 7 625ndash43Rulkov N Sushchik M Tsimring L and Abarbanel H 1995 Generalized synchronization of chaos in unidirectionally
coupled chaotic systems Phys Rev E 51 980ndash94Schiff S So P Chang T Burke R and Sauer T 1996 Detecting dynamical interdependence and generalized synchrony
through mutual prediction in a neural ensemble Phys Rev E 54 6708ndash24Silberstein R B 1995 Neuromodulations of neocortical dynamics Neocortical Dynamics and Human EEG Rhythms
ed P L Nunez (Oxford Oxford University Press)Singer W 1995 Putative functions of temporal correlations in neocortical processing Large-Scale Neuronal Theories
of the Brain ed C Koch and J Davis (London MIT Press)Somers D and Kopell N 1995 Waves and synchrony in networks of oscillators of relaxation and non-relaxation type
Physica D 89 169ndash83Sutton R S and Barto A G 1990 Time derivative models of Pavlovian reinforcement Learning and Computational
Neuroscience Foundations of Adaptive Networks ed M Gabriel and J Moore (Cambridge MA MIT Press) pp497ndash538
Szentagothai J 1983 The modular architectonic principle of neural centers Rev Physiol Biochem Pharmacol 9811ndash61
Tallon-Baudry C Betrand O and Fischer C 2001 Oscillatory synchrony between human extrastriate areas during visualshort-term memory maintenance J Neurosci 21 RC177(1ndash5)
Tass P A 2002 Desynchronization of brain rhythms with soft phase-resetting techniques Biol Cybern 87 102ndash15
732 M Breakspear et al
Tass P Rosenblum M Weule J Kurths J Pikovsky A Volkmann J Schnitzler A and Freund H 1998 Detection of nmphase locking from noisy data application to magnetoencephalography Phys Rev Lett 81 3291ndash4
Terman D and Wang D 1995 Global competition and local cooperation in a network of neural oscillators Physica D81 148ndash76
Terry J and Breakspear M 2003 An improved algorithm for the detection of dynamical interdependence in bivariatetime-series Biol Cybern 88 129ndash36
Tononi G Sporns O and Edelman G M 1994 A measure for brain complexity relating functional segregation andintegration in the nervous system Proc Natl Acad Sci USA 91 5033ndash7
von der Malsburg C 1985 Nervous structures with dynamical links Phys Chem 89 703ndash10Wang X 1991 Period-doublings to chaos in a simple neural network an analytic proof Complex Syst 5 425ndash41Wennekers T and Pasemann F 1996 Synchronous chaos in high dimensional modular neural networks Int J Bifurc
Chaos 6 2055ndash67Zaks M A Parks E H Rosenblum M G and Kurths J 1999 Alternative locking ratios in imperfect phase synchronization
Phys Rev Lett 82 4228ndash31Zhao L and Macau E N 2001 A network of dynamically coupled chaotic maps for scene segmentation IEEE Trans
Neural Netw 12 1375ndash85

Synchronization in a model of neuronal dynamics 729
Inevitably alternative dynamic configurations emerge epitomizing the importance of lsquolabilersquobrain dynamics (Friston 2000a)
Thirdly from a purely functionalist point of view the balance between functionalspecialization and functional integration may rely upon the sorts of mechanism modelledin this paper The brain appears to adhere to two principles of functional organization namelysegregation of function within cortical areas and sub-areas and the integration of these areasin perceptual synthesis cognition and sensorimotor integration (Tononi et al 1994 Fristonet al 1995) The balance between preserving specialized dynamics unique to each arearequired by specialization and the coupling among these areas mediating functional integrationmay be achieved by the neuromodulatory mechanisms described above Significantlythe spontaneous variation in the numbers of degrees of freedom (for example whenlow-dimensional synchronization manifolds are shadowed) imparts a dynamic characteristicto this perspective that is precluded by linear mechanisms alone
Finally from a mechanistic perspective complexity and diversity of brain dynamics arerequired for selection of adaptive responses (Friston 2000b) There are many examples ofselective mechanisms operating within the brain All can be seen as performing an ascenton some objective function or fitness landscape These mechanisms require complex anditinerant dynamics to facilitate a thorough exploration of the landscape These landscapesmay range from those involved in perception (where the landscape is defined by predictionerror see eg Rao and Ballard (1998)) to those involved in value learning (where thelandscape is determined by the outcome of a behaviour eg in temporal difference models (seeeg Sutton and Barto 1990)) In short complexity is necessary to maintain an information-rich environment in which the brain can respond to sensory inputs An essential componentof this is the maintenance of diversity which is a prerequisite for selectionist mechanismsat many scales (Edelman 1993) Understanding the mechanistic basis of complex dynamicsas measured electrophysiologically may well depend on the sort of biologically plausiblemodels used in this paper This understanding may be important for both basic and clinicalneuroscience (eg Tass 2002)
The present study supplements and advances on other research in this field The studyof complex phase synchronization and clustering in neural oscillator networks similar tothe present model has previously been the subject of formal mathematical study Forexample Terman and Wang (1995) studied synchronization of blocks of neural oscillatorsthat were subject to perturbation by connected lsquostimulirsquo Somers and Kopell (1995) studiedthe emergence of both synchronization and travelling waves in networks of coupled neuraloscillators In the work of Terman and Wang the local dynamics were based on neurallsquocaricaturersquo models such as the FitzHughndashNagumo equations and are hence more abstract thanthe current study The approach of Somers and Kopell only required that the local dynamicsexhibit fast and relaxation phases and is hence more abstract still In both studies onlyperiodic dynamics are studied Complex patterns of chaos synchronization in neural arrayshave been characterized by nonlinear methods (Kaneko 1998) and their putative contributionto both learning (Nakajima and Ueda 1996) and perception (Zhao and Macau 2001 Breakspear2001) explored However these studies employed models such as the logistic equation andtent map which are quite abstract in comparison to the present study On the other handHansel and Sompolinsky (1996) studied the role of chaos synchronization in a model ofvisual perception in a cortical hypercolumn using a very similar approach to that adoptedhere but with the additional complexity that synaptic delays were incorporated However inthis and earlier work (Hansel and Sompolinsky 1992) only linear measures of dynamics andsynchrony (specifically the auto- and cross-correlation functions) were employed The analysisof chaotic synchrony in the study of Larter et al (1999) was based only upon observation of
730 M Breakspear et al
the Euclidean difference between variables from two nodes within a larger array Hence thesignificance of the present work is that it provides a quantitative nonlinear analysis of complexchaos synchronization in a biophysical model of cortical dynamics Such research is requiredin order to bridge the gap between quantitative nonlinear research into synchronization andcomputation in abstract models of neural arrays and qualitative or linear analysis in morebiophysically plausible models
Acknowledgments
The authors would like to thank Peter Robinson for assistance with the manuscript and R Larterfor helpful advice MB is a recipient of a NSW Institute of Psychiatry Research Grant anda University of Sydney SESQUI postdoctoral fellowship KJF is supported by the WellcomeTrust
References
Abarbanel H Rulkov N and Sushchik M 1996 Generalised synchronization of chaos the auxiliary system approachPhys Rev E 53 4528ndash35
Afraimovich V Verichev N and Rabinovich M 1986 Stochastic synchronization of oscillation in dissipative systemsRadiophys Quantum Electron 29 795ndash801
Breakspear M 2001 Perception of odours by a nonlinear model of the olfactory bulb Int J Neural Syst 11 101ndash24Breakspear M 2002 Nonlinear phase desynchronisation in human electroencephalographic data Human Brain Mapping
15 175ndash98Breakspear M and Terry J R 2002a Nonlinear interdependence in neural systems motivation theory and relevance
Int J Neurosci 112 1163ndash84Breakspear M and Terry J R 2002b Detection and description of nonlinear interdependence in normal multichannel
human EEG Clin Neurophysiol 113 735ndash53Breakspear M and Terry J R 2002c Topographic organization of nonlinear interdependence in multichannel human
EEG Neuroimage 16 822ndash35Destexhe A Rudolph M Fellous J M and Sejnowski T J 2001 Fluctuating synaptic conductances recreate in vivo-like
activity in neocortical neurons Neuroscience 107 13ndash24Douglas R J Mahowald M A and Martin K A 1996 Microarchitecture of neocortical columns Brain TheorymdashBiological
Basis and Computational Principles ed A Aertsen and V Braitenberg (Amsterdam Elsevier)Eckhorn R Bauer R Jordan W Brosch M Kruse W Munk M and Reitboeck H J 1988 Coherent oscillations a
mechanism of feature linking in the visual cortex Multiple electrode and correlation analyses in the cat BiolCybern 60 121ndash30
Edelman G M 1993 Neural Darwinism selection and reentrant signalling in higher brain function Neuron 10 115ndash25Engel A K Kreiter A K Konig P and Singer W 1991 Synchronization of oscillatory neuronal responses between
striate and extrastriate visual cortical areas of the cat Proc Natl Acad Sci USA 88 6048ndash52Friston K J 2000a The labile brain I Neuronal transients and nonlinear coupling Phil Trans R Soc B 355 215ndash36Friston K J 2000b The labile brain II Transients complexity and selection Phil Trans R Soc B 355 237ndash52Friston K J Tononi G Sporns O and Edelman G 1995 Characterising the complexity of neuronal interactions Human
Brain Mapping 3 302ndash14Fujisaka H and Yamada T 1983 Stability theory of synchronized motion in coupled-oscillator system Prog Theor
Phys 69 32ndash47Gatlin L 1972 Information Theory and the Living System (New York Columbia University Press)Gray C M Konig P Engel A and Singer W 1989 Oscillatory responses in cat visual cortex exhibit inter-columnar
synchronization which reflects global stimulus properties Science 338 334ndash7Gray C M and Singer W 1989 Stimulus specific neuronal oscillations in orientation columns of cat visual cortex Proc
Natl Acad Sci USA 86 1698ndash702Haig A R Gordon E Wright J J Meares R A and Bahramali H 2000 Synchronous cortical gamma-band activity in
task-relevant cognition Neuroreport 11 669ndash75Hansel D and Sompolinsky H 1992 Synchronization and computation in a chaotic neural network Phys Rev Lett 68
718ndash21
Synchronization in a model of neuronal dynamics 731
Hansel D and Sompolinsky H 1996 Chaos and synchrony in a model of a hypercolumn in visual cortex J ComputNeurosci 3 7ndash4
Heagy J Carroll T and Pecora L 1998 Desynchronization by periodic orbits Phys Rev E 57 R1253ndash6Hunt B Ott E and Yorke J 1997 Differentiable synchronization of chaos Phys Rev E 55 4029ndash34Josic K 1998 Invariant manifolds and synchronization of coupled dynamical systems Phys Rev Lett 80 3053ndash6Kandel E Schwartz J and Jessell T 2000 Principles of Neural Science (New York McGraw-Hill)Kaneko K 1998 On the strength of attractors in a high-dimensional system Physica D 124 322ndash44Kocarev L Parlitz U and Brown R 2000 Robust synchronization of chaotic systems Phys Rev E 61 3716ndash20Larter R Speelman B and Worth R M 1999 A coupled ordinary differential equation lattice model for the simulation
of epileptic seizures Chaos 9 795ndash804Lumer E Edelman G and Tononi G 1997a Neural dynamics in a model of the thalamocortical system I Layers loops
and the emergence of fast synchronous oscillations Cereb Cortex 7 207ndash27Lumer E Edelman G and Tononi G 1997b Neural dynamics in a model of the thalamocortical system II The role of
neural synchrony tested through perturbations of spike timing Cereb Cortex 7 228ndash36Magleby K 1987 Short-term modulation at synapses Synaptic Function ed G M Edelman W E Gall and
W M Cowan (New York Wiley)Maistrenko Y Maistrenko V Popovich A and Mosekilde E 1998 Transverse instability and riddled basins in a system
of two coupled logistic maps Phys Rev E 57 2713ndash24Morris C and Lecar H 1981 Voltage oscillations in the barnacle giant muscle fiber Biophys J 35 193ndash213Mountcastle V B 1997 The columnar organization of the neocortex Brain 120 701ndash22Mountcastle V B 1998 The Cerebral Cortex (Cambridge MA Harvard University Press)Nakajima H and Ueda Y 1996 Riddled basins of the optimal states in learning dynamical systems Physica D 99 35ndash44Nunez P L 1995 Neocortical Dynamics and Human Brain Rhythms (Oxford Oxford University Press)Osipov G V Pikovsky A S Rosenblum M G and Kurths J 1997 Phase synchronization effects in a lattice of nonidentical
Rossler oscillators Phys Rev E 55 2353ndash61Ott E and Sommerer J 1994 Blowout bifurcations the occurrence of riddled basins and onndashoff intermittency Phys
Lett A 188 39ndash47Pecora L and Carroll T 1990 Synchronization in chaotic systems Phys Rev Lett 64 821ndash4Pikovsky A and Grassberger P 1991 Symmetry breaking bifurcation for coupled chaotic attractors J Phys A Math
Gen 24 4587ndash97Rao R P and Ballard D H 1998 Predictive coding in the visual cortex a functional interpretation of some extra-classical
receptive field effects Nature Neurosci 2 79ndash87Robinson P A Wright J J and Rennie C R 1998 Synchronous oscillations in the cerebral cortex Phys Rev E 57
4578ndash88Rodriguez E George N Lachauz J P Martinerie J Renault B and Varela F J 1999 Perceptionrsquos shadow long distance
synchronization of human brain activity Nature 397 430ndash3Rosenblum M Pikovsky A and Kurths J 1996 Phase synchronization of chaotic oscillators Phys Rev Lett 76 1804ndash7Rosenblum M Pikovsky A and Kurths J 1997 From phase to lag synchronization in coupled chaotic oscillators Phys
Rev Lett 78 4193ndash6Rulkov N and Sushchik M 1997 Robustness of synchronized chaotic oscillations Int J Bifurc Chaos 7 625ndash43Rulkov N Sushchik M Tsimring L and Abarbanel H 1995 Generalized synchronization of chaos in unidirectionally
coupled chaotic systems Phys Rev E 51 980ndash94Schiff S So P Chang T Burke R and Sauer T 1996 Detecting dynamical interdependence and generalized synchrony
through mutual prediction in a neural ensemble Phys Rev E 54 6708ndash24Silberstein R B 1995 Neuromodulations of neocortical dynamics Neocortical Dynamics and Human EEG Rhythms
ed P L Nunez (Oxford Oxford University Press)Singer W 1995 Putative functions of temporal correlations in neocortical processing Large-Scale Neuronal Theories
of the Brain ed C Koch and J Davis (London MIT Press)Somers D and Kopell N 1995 Waves and synchrony in networks of oscillators of relaxation and non-relaxation type
Physica D 89 169ndash83Sutton R S and Barto A G 1990 Time derivative models of Pavlovian reinforcement Learning and Computational
Neuroscience Foundations of Adaptive Networks ed M Gabriel and J Moore (Cambridge MA MIT Press) pp497ndash538
Szentagothai J 1983 The modular architectonic principle of neural centers Rev Physiol Biochem Pharmacol 9811ndash61
Tallon-Baudry C Betrand O and Fischer C 2001 Oscillatory synchrony between human extrastriate areas during visualshort-term memory maintenance J Neurosci 21 RC177(1ndash5)
Tass P A 2002 Desynchronization of brain rhythms with soft phase-resetting techniques Biol Cybern 87 102ndash15
732 M Breakspear et al
Tass P Rosenblum M Weule J Kurths J Pikovsky A Volkmann J Schnitzler A and Freund H 1998 Detection of nmphase locking from noisy data application to magnetoencephalography Phys Rev Lett 81 3291ndash4
Terman D and Wang D 1995 Global competition and local cooperation in a network of neural oscillators Physica D81 148ndash76
Terry J and Breakspear M 2003 An improved algorithm for the detection of dynamical interdependence in bivariatetime-series Biol Cybern 88 129ndash36
Tononi G Sporns O and Edelman G M 1994 A measure for brain complexity relating functional segregation andintegration in the nervous system Proc Natl Acad Sci USA 91 5033ndash7
von der Malsburg C 1985 Nervous structures with dynamical links Phys Chem 89 703ndash10Wang X 1991 Period-doublings to chaos in a simple neural network an analytic proof Complex Syst 5 425ndash41Wennekers T and Pasemann F 1996 Synchronous chaos in high dimensional modular neural networks Int J Bifurc
Chaos 6 2055ndash67Zaks M A Parks E H Rosenblum M G and Kurths J 1999 Alternative locking ratios in imperfect phase synchronization
Phys Rev Lett 82 4228ndash31Zhao L and Macau E N 2001 A network of dynamically coupled chaotic maps for scene segmentation IEEE Trans
Neural Netw 12 1375ndash85

730 M Breakspear et al
the Euclidean difference between variables from two nodes within a larger array Hence thesignificance of the present work is that it provides a quantitative nonlinear analysis of complexchaos synchronization in a biophysical model of cortical dynamics Such research is requiredin order to bridge the gap between quantitative nonlinear research into synchronization andcomputation in abstract models of neural arrays and qualitative or linear analysis in morebiophysically plausible models
Acknowledgments
The authors would like to thank Peter Robinson for assistance with the manuscript and R Larterfor helpful advice MB is a recipient of a NSW Institute of Psychiatry Research Grant anda University of Sydney SESQUI postdoctoral fellowship KJF is supported by the WellcomeTrust
References
Abarbanel H Rulkov N and Sushchik M 1996 Generalised synchronization of chaos the auxiliary system approachPhys Rev E 53 4528ndash35
Afraimovich V Verichev N and Rabinovich M 1986 Stochastic synchronization of oscillation in dissipative systemsRadiophys Quantum Electron 29 795ndash801
Breakspear M 2001 Perception of odours by a nonlinear model of the olfactory bulb Int J Neural Syst 11 101ndash24Breakspear M 2002 Nonlinear phase desynchronisation in human electroencephalographic data Human Brain Mapping
15 175ndash98Breakspear M and Terry J R 2002a Nonlinear interdependence in neural systems motivation theory and relevance
Int J Neurosci 112 1163ndash84Breakspear M and Terry J R 2002b Detection and description of nonlinear interdependence in normal multichannel
human EEG Clin Neurophysiol 113 735ndash53Breakspear M and Terry J R 2002c Topographic organization of nonlinear interdependence in multichannel human
EEG Neuroimage 16 822ndash35Destexhe A Rudolph M Fellous J M and Sejnowski T J 2001 Fluctuating synaptic conductances recreate in vivo-like
activity in neocortical neurons Neuroscience 107 13ndash24Douglas R J Mahowald M A and Martin K A 1996 Microarchitecture of neocortical columns Brain TheorymdashBiological
Basis and Computational Principles ed A Aertsen and V Braitenberg (Amsterdam Elsevier)Eckhorn R Bauer R Jordan W Brosch M Kruse W Munk M and Reitboeck H J 1988 Coherent oscillations a
mechanism of feature linking in the visual cortex Multiple electrode and correlation analyses in the cat BiolCybern 60 121ndash30
Edelman G M 1993 Neural Darwinism selection and reentrant signalling in higher brain function Neuron 10 115ndash25Engel A K Kreiter A K Konig P and Singer W 1991 Synchronization of oscillatory neuronal responses between
striate and extrastriate visual cortical areas of the cat Proc Natl Acad Sci USA 88 6048ndash52Friston K J 2000a The labile brain I Neuronal transients and nonlinear coupling Phil Trans R Soc B 355 215ndash36Friston K J 2000b The labile brain II Transients complexity and selection Phil Trans R Soc B 355 237ndash52Friston K J Tononi G Sporns O and Edelman G 1995 Characterising the complexity of neuronal interactions Human
Brain Mapping 3 302ndash14Fujisaka H and Yamada T 1983 Stability theory of synchronized motion in coupled-oscillator system Prog Theor
Phys 69 32ndash47Gatlin L 1972 Information Theory and the Living System (New York Columbia University Press)Gray C M Konig P Engel A and Singer W 1989 Oscillatory responses in cat visual cortex exhibit inter-columnar
synchronization which reflects global stimulus properties Science 338 334ndash7Gray C M and Singer W 1989 Stimulus specific neuronal oscillations in orientation columns of cat visual cortex Proc
Natl Acad Sci USA 86 1698ndash702Haig A R Gordon E Wright J J Meares R A and Bahramali H 2000 Synchronous cortical gamma-band activity in
task-relevant cognition Neuroreport 11 669ndash75Hansel D and Sompolinsky H 1992 Synchronization and computation in a chaotic neural network Phys Rev Lett 68
718ndash21
Synchronization in a model of neuronal dynamics 731
Hansel D and Sompolinsky H 1996 Chaos and synchrony in a model of a hypercolumn in visual cortex J ComputNeurosci 3 7ndash4
Heagy J Carroll T and Pecora L 1998 Desynchronization by periodic orbits Phys Rev E 57 R1253ndash6Hunt B Ott E and Yorke J 1997 Differentiable synchronization of chaos Phys Rev E 55 4029ndash34Josic K 1998 Invariant manifolds and synchronization of coupled dynamical systems Phys Rev Lett 80 3053ndash6Kandel E Schwartz J and Jessell T 2000 Principles of Neural Science (New York McGraw-Hill)Kaneko K 1998 On the strength of attractors in a high-dimensional system Physica D 124 322ndash44Kocarev L Parlitz U and Brown R 2000 Robust synchronization of chaotic systems Phys Rev E 61 3716ndash20Larter R Speelman B and Worth R M 1999 A coupled ordinary differential equation lattice model for the simulation
of epileptic seizures Chaos 9 795ndash804Lumer E Edelman G and Tononi G 1997a Neural dynamics in a model of the thalamocortical system I Layers loops
and the emergence of fast synchronous oscillations Cereb Cortex 7 207ndash27Lumer E Edelman G and Tononi G 1997b Neural dynamics in a model of the thalamocortical system II The role of
neural synchrony tested through perturbations of spike timing Cereb Cortex 7 228ndash36Magleby K 1987 Short-term modulation at synapses Synaptic Function ed G M Edelman W E Gall and
W M Cowan (New York Wiley)Maistrenko Y Maistrenko V Popovich A and Mosekilde E 1998 Transverse instability and riddled basins in a system
of two coupled logistic maps Phys Rev E 57 2713ndash24Morris C and Lecar H 1981 Voltage oscillations in the barnacle giant muscle fiber Biophys J 35 193ndash213Mountcastle V B 1997 The columnar organization of the neocortex Brain 120 701ndash22Mountcastle V B 1998 The Cerebral Cortex (Cambridge MA Harvard University Press)Nakajima H and Ueda Y 1996 Riddled basins of the optimal states in learning dynamical systems Physica D 99 35ndash44Nunez P L 1995 Neocortical Dynamics and Human Brain Rhythms (Oxford Oxford University Press)Osipov G V Pikovsky A S Rosenblum M G and Kurths J 1997 Phase synchronization effects in a lattice of nonidentical
Rossler oscillators Phys Rev E 55 2353ndash61Ott E and Sommerer J 1994 Blowout bifurcations the occurrence of riddled basins and onndashoff intermittency Phys
Lett A 188 39ndash47Pecora L and Carroll T 1990 Synchronization in chaotic systems Phys Rev Lett 64 821ndash4Pikovsky A and Grassberger P 1991 Symmetry breaking bifurcation for coupled chaotic attractors J Phys A Math
Gen 24 4587ndash97Rao R P and Ballard D H 1998 Predictive coding in the visual cortex a functional interpretation of some extra-classical
receptive field effects Nature Neurosci 2 79ndash87Robinson P A Wright J J and Rennie C R 1998 Synchronous oscillations in the cerebral cortex Phys Rev E 57
4578ndash88Rodriguez E George N Lachauz J P Martinerie J Renault B and Varela F J 1999 Perceptionrsquos shadow long distance
synchronization of human brain activity Nature 397 430ndash3Rosenblum M Pikovsky A and Kurths J 1996 Phase synchronization of chaotic oscillators Phys Rev Lett 76 1804ndash7Rosenblum M Pikovsky A and Kurths J 1997 From phase to lag synchronization in coupled chaotic oscillators Phys
Rev Lett 78 4193ndash6Rulkov N and Sushchik M 1997 Robustness of synchronized chaotic oscillations Int J Bifurc Chaos 7 625ndash43Rulkov N Sushchik M Tsimring L and Abarbanel H 1995 Generalized synchronization of chaos in unidirectionally
coupled chaotic systems Phys Rev E 51 980ndash94Schiff S So P Chang T Burke R and Sauer T 1996 Detecting dynamical interdependence and generalized synchrony
through mutual prediction in a neural ensemble Phys Rev E 54 6708ndash24Silberstein R B 1995 Neuromodulations of neocortical dynamics Neocortical Dynamics and Human EEG Rhythms
ed P L Nunez (Oxford Oxford University Press)Singer W 1995 Putative functions of temporal correlations in neocortical processing Large-Scale Neuronal Theories
of the Brain ed C Koch and J Davis (London MIT Press)Somers D and Kopell N 1995 Waves and synchrony in networks of oscillators of relaxation and non-relaxation type
Physica D 89 169ndash83Sutton R S and Barto A G 1990 Time derivative models of Pavlovian reinforcement Learning and Computational
Neuroscience Foundations of Adaptive Networks ed M Gabriel and J Moore (Cambridge MA MIT Press) pp497ndash538
Szentagothai J 1983 The modular architectonic principle of neural centers Rev Physiol Biochem Pharmacol 9811ndash61
Tallon-Baudry C Betrand O and Fischer C 2001 Oscillatory synchrony between human extrastriate areas during visualshort-term memory maintenance J Neurosci 21 RC177(1ndash5)
Tass P A 2002 Desynchronization of brain rhythms with soft phase-resetting techniques Biol Cybern 87 102ndash15
732 M Breakspear et al
Tass P Rosenblum M Weule J Kurths J Pikovsky A Volkmann J Schnitzler A and Freund H 1998 Detection of nmphase locking from noisy data application to magnetoencephalography Phys Rev Lett 81 3291ndash4
Terman D and Wang D 1995 Global competition and local cooperation in a network of neural oscillators Physica D81 148ndash76
Terry J and Breakspear M 2003 An improved algorithm for the detection of dynamical interdependence in bivariatetime-series Biol Cybern 88 129ndash36
Tononi G Sporns O and Edelman G M 1994 A measure for brain complexity relating functional segregation andintegration in the nervous system Proc Natl Acad Sci USA 91 5033ndash7
von der Malsburg C 1985 Nervous structures with dynamical links Phys Chem 89 703ndash10Wang X 1991 Period-doublings to chaos in a simple neural network an analytic proof Complex Syst 5 425ndash41Wennekers T and Pasemann F 1996 Synchronous chaos in high dimensional modular neural networks Int J Bifurc
Chaos 6 2055ndash67Zaks M A Parks E H Rosenblum M G and Kurths J 1999 Alternative locking ratios in imperfect phase synchronization
Phys Rev Lett 82 4228ndash31Zhao L and Macau E N 2001 A network of dynamically coupled chaotic maps for scene segmentation IEEE Trans
Neural Netw 12 1375ndash85

Synchronization in a model of neuronal dynamics 731
Hansel D and Sompolinsky H 1996 Chaos and synchrony in a model of a hypercolumn in visual cortex J ComputNeurosci 3 7ndash4
Heagy J Carroll T and Pecora L 1998 Desynchronization by periodic orbits Phys Rev E 57 R1253ndash6Hunt B Ott E and Yorke J 1997 Differentiable synchronization of chaos Phys Rev E 55 4029ndash34Josic K 1998 Invariant manifolds and synchronization of coupled dynamical systems Phys Rev Lett 80 3053ndash6Kandel E Schwartz J and Jessell T 2000 Principles of Neural Science (New York McGraw-Hill)Kaneko K 1998 On the strength of attractors in a high-dimensional system Physica D 124 322ndash44Kocarev L Parlitz U and Brown R 2000 Robust synchronization of chaotic systems Phys Rev E 61 3716ndash20Larter R Speelman B and Worth R M 1999 A coupled ordinary differential equation lattice model for the simulation
of epileptic seizures Chaos 9 795ndash804Lumer E Edelman G and Tononi G 1997a Neural dynamics in a model of the thalamocortical system I Layers loops
and the emergence of fast synchronous oscillations Cereb Cortex 7 207ndash27Lumer E Edelman G and Tononi G 1997b Neural dynamics in a model of the thalamocortical system II The role of
neural synchrony tested through perturbations of spike timing Cereb Cortex 7 228ndash36Magleby K 1987 Short-term modulation at synapses Synaptic Function ed G M Edelman W E Gall and
W M Cowan (New York Wiley)Maistrenko Y Maistrenko V Popovich A and Mosekilde E 1998 Transverse instability and riddled basins in a system
of two coupled logistic maps Phys Rev E 57 2713ndash24Morris C and Lecar H 1981 Voltage oscillations in the barnacle giant muscle fiber Biophys J 35 193ndash213Mountcastle V B 1997 The columnar organization of the neocortex Brain 120 701ndash22Mountcastle V B 1998 The Cerebral Cortex (Cambridge MA Harvard University Press)Nakajima H and Ueda Y 1996 Riddled basins of the optimal states in learning dynamical systems Physica D 99 35ndash44Nunez P L 1995 Neocortical Dynamics and Human Brain Rhythms (Oxford Oxford University Press)Osipov G V Pikovsky A S Rosenblum M G and Kurths J 1997 Phase synchronization effects in a lattice of nonidentical
Rossler oscillators Phys Rev E 55 2353ndash61Ott E and Sommerer J 1994 Blowout bifurcations the occurrence of riddled basins and onndashoff intermittency Phys
Lett A 188 39ndash47Pecora L and Carroll T 1990 Synchronization in chaotic systems Phys Rev Lett 64 821ndash4Pikovsky A and Grassberger P 1991 Symmetry breaking bifurcation for coupled chaotic attractors J Phys A Math
Gen 24 4587ndash97Rao R P and Ballard D H 1998 Predictive coding in the visual cortex a functional interpretation of some extra-classical
receptive field effects Nature Neurosci 2 79ndash87Robinson P A Wright J J and Rennie C R 1998 Synchronous oscillations in the cerebral cortex Phys Rev E 57
4578ndash88Rodriguez E George N Lachauz J P Martinerie J Renault B and Varela F J 1999 Perceptionrsquos shadow long distance
synchronization of human brain activity Nature 397 430ndash3Rosenblum M Pikovsky A and Kurths J 1996 Phase synchronization of chaotic oscillators Phys Rev Lett 76 1804ndash7Rosenblum M Pikovsky A and Kurths J 1997 From phase to lag synchronization in coupled chaotic oscillators Phys
Rev Lett 78 4193ndash6Rulkov N and Sushchik M 1997 Robustness of synchronized chaotic oscillations Int J Bifurc Chaos 7 625ndash43Rulkov N Sushchik M Tsimring L and Abarbanel H 1995 Generalized synchronization of chaos in unidirectionally
coupled chaotic systems Phys Rev E 51 980ndash94Schiff S So P Chang T Burke R and Sauer T 1996 Detecting dynamical interdependence and generalized synchrony
through mutual prediction in a neural ensemble Phys Rev E 54 6708ndash24Silberstein R B 1995 Neuromodulations of neocortical dynamics Neocortical Dynamics and Human EEG Rhythms
ed P L Nunez (Oxford Oxford University Press)Singer W 1995 Putative functions of temporal correlations in neocortical processing Large-Scale Neuronal Theories
of the Brain ed C Koch and J Davis (London MIT Press)Somers D and Kopell N 1995 Waves and synchrony in networks of oscillators of relaxation and non-relaxation type
Physica D 89 169ndash83Sutton R S and Barto A G 1990 Time derivative models of Pavlovian reinforcement Learning and Computational
Neuroscience Foundations of Adaptive Networks ed M Gabriel and J Moore (Cambridge MA MIT Press) pp497ndash538
Szentagothai J 1983 The modular architectonic principle of neural centers Rev Physiol Biochem Pharmacol 9811ndash61
Tallon-Baudry C Betrand O and Fischer C 2001 Oscillatory synchrony between human extrastriate areas during visualshort-term memory maintenance J Neurosci 21 RC177(1ndash5)
Tass P A 2002 Desynchronization of brain rhythms with soft phase-resetting techniques Biol Cybern 87 102ndash15
732 M Breakspear et al
Tass P Rosenblum M Weule J Kurths J Pikovsky A Volkmann J Schnitzler A and Freund H 1998 Detection of nmphase locking from noisy data application to magnetoencephalography Phys Rev Lett 81 3291ndash4
Terman D and Wang D 1995 Global competition and local cooperation in a network of neural oscillators Physica D81 148ndash76
Terry J and Breakspear M 2003 An improved algorithm for the detection of dynamical interdependence in bivariatetime-series Biol Cybern 88 129ndash36
Tononi G Sporns O and Edelman G M 1994 A measure for brain complexity relating functional segregation andintegration in the nervous system Proc Natl Acad Sci USA 91 5033ndash7
von der Malsburg C 1985 Nervous structures with dynamical links Phys Chem 89 703ndash10Wang X 1991 Period-doublings to chaos in a simple neural network an analytic proof Complex Syst 5 425ndash41Wennekers T and Pasemann F 1996 Synchronous chaos in high dimensional modular neural networks Int J Bifurc
Chaos 6 2055ndash67Zaks M A Parks E H Rosenblum M G and Kurths J 1999 Alternative locking ratios in imperfect phase synchronization
Phys Rev Lett 82 4228ndash31Zhao L and Macau E N 2001 A network of dynamically coupled chaotic maps for scene segmentation IEEE Trans
Neural Netw 12 1375ndash85

732 M Breakspear et al
Tass P Rosenblum M Weule J Kurths J Pikovsky A Volkmann J Schnitzler A and Freund H 1998 Detection of nmphase locking from noisy data application to magnetoencephalography Phys Rev Lett 81 3291ndash4
Terman D and Wang D 1995 Global competition and local cooperation in a network of neural oscillators Physica D81 148ndash76
Terry J and Breakspear M 2003 An improved algorithm for the detection of dynamical interdependence in bivariatetime-series Biol Cybern 88 129ndash36
Tononi G Sporns O and Edelman G M 1994 A measure for brain complexity relating functional segregation andintegration in the nervous system Proc Natl Acad Sci USA 91 5033ndash7
von der Malsburg C 1985 Nervous structures with dynamical links Phys Chem 89 703ndash10Wang X 1991 Period-doublings to chaos in a simple neural network an analytic proof Complex Syst 5 425ndash41Wennekers T and Pasemann F 1996 Synchronous chaos in high dimensional modular neural networks Int J Bifurc
Chaos 6 2055ndash67Zaks M A Parks E H Rosenblum M G and Kurths J 1999 Alternative locking ratios in imperfect phase synchronization
Phys Rev Lett 82 4228ndash31Zhao L and Macau E N 2001 A network of dynamically coupled chaotic maps for scene segmentation IEEE Trans
Neural Netw 12 1375ndash85
Related Documents











