Chapter 32 Modulation of Early Inflammatory Reactions to Promote Engraftment and Function of Transplanted Pancreatic Islets in Autoimmune Diabetes Lorenzo Piemonti, Luca G. Guidotti, and Manuela Battaglia Abstract We acknowledge that successful long-term islet survival in the liver and immune tolerance to intrahepatic islet antigens are highly dependent upon the ini- tial inflammatory and priming events that occur at this site. Thromboembolic and necroinflammatory events occurring in the liver early after portal vein islet trans- plantation are thought to reduce the total islet mass by up to 75%. The magnitude of such loss represents a major factor necessitating the extremely large number of islets needed to achieve normoglycemia. A better understanding and control of these events – including their likely support to effector immune responses – are required if we are to develop ways to prevent them, improve intrahepatic islet engraftment, and achieve long-term tolerance. Keywords Type 1 diabetes · Pancreatic islet transplantation · Instant blood-mediated inflammatory reaction 32.1 Introduction Despite the substantial improvements in insulin therapy thanks to new commer- cially available drugs, and the adoption of intensive treatment regimens able to improve glycemic control, exogenous insulin administration cannot avoid the long- term complications of diabetes and the life expectancy of diabetic patients is still shorter compared to that of the general population [1, 2]. In principle, treatment of type 1 diabetes (T1D) and many cases of type 2 diabetes lies in the possibility of finding a β-cell mass replacement capable of performing two essential functions: assessing blood sugar levels and secreting appropriate levels of insulin in the vascular bed. Currently, the only available clinical therapy capable of L. Piemonti (B ) San Raffaele Diabetes Research Institute (HSR-DRI), Via Olgettina 60, 20132, Milano, Italy e-mail: [email protected] 725 M.S. Islam (ed.), The Islets of Langerhans, Advances in Experimental Medicine and Biology 654, DOI 10.1007/978-90-481-3271-3_32, C Springer Science+Business Media B.V. 2010

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Chapter 32Modulation of Early Inflammatory Reactionsto Promote Engraftment and Functionof Transplanted Pancreatic Isletsin Autoimmune Diabetes
Lorenzo Piemonti, Luca G. Guidotti, and Manuela Battaglia
Abstract We acknowledge that successful long-term islet survival in the liver andimmune tolerance to intrahepatic islet antigens are highly dependent upon the ini-tial inflammatory and priming events that occur at this site. Thromboembolic andnecroinflammatory events occurring in the liver early after portal vein islet trans-plantation are thought to reduce the total islet mass by up to 75%. The magnitudeof such loss represents a major factor necessitating the extremely large number ofislets needed to achieve normoglycemia. A better understanding and control of theseevents – including their likely support to effector immune responses – are requiredif we are to develop ways to prevent them, improve intrahepatic islet engraftment,and achieve long-term tolerance.
Keywords Type 1 diabetes · Pancreatic islet transplantation · Instantblood-mediated inflammatory reaction
32.1 Introduction
Despite the substantial improvements in insulin therapy thanks to new commer-cially available drugs, and the adoption of intensive treatment regimens able toimprove glycemic control, exogenous insulin administration cannot avoid the long-term complications of diabetes and the life expectancy of diabetic patients is stillshorter compared to that of the general population [1, 2].
In principle, treatment of type 1 diabetes (T1D) and many cases of type 2 diabeteslies in the possibility of finding a β-cell mass replacement capable of performing twoessential functions: assessing blood sugar levels and secreting appropriate levels ofinsulin in the vascular bed. Currently, the only available clinical therapy capable of
L. Piemonti (B)San Raffaele Diabetes Research Institute (HSR-DRI),Via Olgettina 60, 20132, Milano, Italye-mail: [email protected]
725M.S. Islam (ed.), The Islets of Langerhans, Advances in ExperimentalMedicine and Biology 654, DOI 10.1007/978-90-481-3271-3_32,C© Springer Science+Business Media B.V. 2010

726 L. Piemonti et al.
restoring β-cell mass in diabetic patients is the allogeneic/autologous transplantationof β-cells (i.e., somatic cell therapy with total pancreas, and Langherans’ islets orβ-cell transplantation).
Replacement of the whole gland reestablishes long-term normoglycemia, witha success rate of 80% [3], and it is especially successful in patients who undergosimultaneous pancreas and kidney transplantation. However, because of the risk ofsurgical complications, this procedure will never be a viable option for most T1Dpatients. The subjects offered this treatment are patients who have already developedmany of the secondary complications, including end-stage renal failure and have aquality of life that is adequate for undergoing such a difficult treatment. Since thebreakthrough made by Shapiro and colleagues [4], islet transplantation has emergedas an attractive alternative to whole pancreas transplantation. Despite advances inrecent years [5], allogeneic somatic therapy is still problematic.
A nonspecific immune response mediated predominantly by innate inflamma-tory processes related to mechanics and site, and preexisting and transplant-inducedauto- and allo-specific cellular immune responses (possibly promoted by the initialinflammation) play a major role in the loss of islets and islet function trans-planted in the liver. Although significantly improved by the implementation of theEdmonton protocol, our capacity of achieving long-lasting insulin independencein T1D patients undergoing portal vein islet transplantation remains limited [5–7].This indicates that the detrimental impact of innate and adaptive immune responsesis not fully contained by the Edmonton protocol-associated regimen of general-ized immunosuppression (i.e., induction with daclizumab [anti-IL-2Ralpha mAb]and maintenance with rapamycin [mTOR inhibitor] plus tacrolimus [calcineurininhibitor]).
Prolong intrahepatic islet survival by increasing the potency of such regimen isnot practicable, due to the likelihood of enhancing susceptibility to cancer and infec-tions, and the toxicity that some of these drugs may have toward kidney functionsand transplanted islets. Rather, it is intuitive that alternative strategies aimed at selec-tively inhibiting undesired islet-specific or nonspecific immune responses representan ideal step toward a better management (i.e., weaning/withdrawal of generalizedimmune suppression) and outcome (i.e., long-lasting insulin independence) of islettransplanted T1D patients.
32.2 Defining the Site for Islet Transplantation
The liver was suggested as an optimal site for islet transplantation by Lacy andcolleagues, by using a rat model of diabetes [8]. By the 1980s, successful trans-plantation of islet autografts was reported in humans by using infusion of cellsinto the patient’s liver through the portal venous circulation [9–11]. Subsequently,the publication of the first case of insulin independence in a diabetic patient afterislet infusion through the portal vein consecrated the liver as the site of choice forislet transplantation in humans [12]. Because of this early success, the subsequent

32 Modulation of Early Inflammatory Reactions 727
clinical experience of islet transplantation has been developed almost exclusivelyusing the intrahepatic infusion through the portal vein. However, in the last years, ithas becoming increasingly recognized that the liver may not be the optimal environ-ment as a recipient site for pancreatic islets, owing not only to immunological [13,14] but also to anatomical [15, 16] and physiological factors that likely contributeto the decline of islet mass after implantation [17–19].
The potential advantages of the intrahepatic islet transplantation include thelow risk of the procedure and the delivery of insulin directly to the liver. Froma clinical point of view, the process of intrahepatic infusion is currently consid-ered safe, although there is a low risk of portal vein thrombosis and elevated portalpressure, in addition to bleeding from the percutaneous hepatic puncture site [20–23]. Numerous investigators, however, have recently addressed these complications,using high doses of heparin in conjunction with sealing the parenchymal track withthrombogenic material [24, 25].
From a metabolic point of view the process of intrahepatic infusion is consideredoptimal due to the fact that insulin is delivered more physiologically after intraportaltransplantation [26]. In healthy individuals indeed, insulin is secreted by the pan-creas into the portal venous circulation to the liver. Thus, the physiological balancebetween hepatic and extrahepatic insulin exposure requires portal delivery of insulin[27–29], and chronically implanted intrahepatic islets were described to be capableof restoring a pattern of insulin secretion and clearance that closely reproduces thatof the native pancreas [26]. However, the argument that islets would work morephysiologically after intraportal transplantation has recently received little supportin the literature. It has clearly been demonstrated that intraportally transplanted isletsin experimental models respond to glucose stimulation only when perfused via thehepatic artery; no response is observed after challenge via the portal vein [30]. Thereare also reports on alterations in islet function after intraportal islet transplantation,such as a defective glucagon response to hypoglycemia [19, 31, 32] and a defectiveglucose-stimulated insulin release [33].
The potential disadvantages of the intrahepatic route of islet transplantationinclude: (i) induction of instant blood-mediated inflammatory reaction, thrombo-sis, and nonantigen-specific inflammation; (ii) delayed vascularization of islets; (iii)exposure to high level of immunosuppressive agents; and (iv) glucolipotoxicity.
Intrahepatic islet infusion in men is associated with an immediate blood-mediated inflammatory reaction, thrombosis, and hepatic tissue ischemia withelevated blood liver enzymes [34–45]. Loss of as many as 50–75% of islets duringengraftment in the liver [46] has been suggested to be a prime factor necessitatingthe very large number of islets needed to achieve normoglycemia [35]. Furthermore,the necessity for cannulation of the portal system to seed the islets leads to anincrease in the portal pressure proportional to the administered islet mass [47], thusrestricting the total mass that can be implanted. As a consequence, a highly purifiedsuspension of islets is needed to transplant sufficient cells to achieve insulin inde-pendence. Because the purity of the suspension is inversely proportional to the isletyield per donor [48], fewer islets can be isolated from the already scarce donor pool,further limiting broad clinical applicability of pancreatic islet transplantation. It is

728 L. Piemonti et al.
known that islets are highly vascularized. Pancreatic islets comprise only approxi-mately 2% of the pancreas cell mass and yet consume up to 20% of the arterial bloodflow [49–51]. Blood vessels within pancreatic islets are of a greater density thanthose in the surrounding exocrine tissue and are lined with fenestrated endothelialcells. These specialized features are responsible for the greater partial pressure ofoxygen in islets as compared with acinar tissue and other organs, which is importantfor normal islet cell function and survival. Pancreatic islets lose this vascular supplyduring the isolation process [52]. In contrast to whole-organ transplantation, whereorgan perfusion is quickly reestablished by reconnection of arterial and venous ves-sels, the reestablishment of blood flow to transplanted islets requires several daysand involves angiogenesis and possibly vasculogenesis. Importantly, not only arethe islets avascular for several days following transplantation, but they are also lessvascularized and have a lower oxygen tension than islets in the pancreas when revas-cularization is complete [15, 53]. Revascularization of the islet graft is estimated torequire 7–14 days [54–57]. Vascular endothelial cells from both donor [58] andhost [52] stimulate angiogenesis to form intragraft blood vessels in 3–5 days post-transplant, and full blood circulation is reestablished within approximately 1 week.Decreased vascular density and low revascularization have been reported in trans-planted mouse [53] and human pancreatic islets [59] although studies have alsoshown normal vascularity [55].
The portal vein carries blood with oxygen tension slightly less than that of arte-rial blood, contains higher concentrations of substances from the gut that may betoxic to the islets, and the immunosuppressive agents (known to be toxic to isletcell function) are absorbed from the gut and thus their toxic effects might be mag-nified for islets bathed in portal blood. Recently, portal vein and peripheral bloodimmunosuppressant drug concentrations in islet transplant patients were measuredand higher levels of both sirolimus and tacrolimus in the portal blood circulationwere reported [18, 60]. These drugs may interfere with angiogenesis and may becytotoxic to β-cells at high local concentrations.
Islets are metabolically active and require access to oxygen, glucose, and othermetabolites in a hospitable environment at physiological pH, and to be free fromtoxic metabolites and oxygen free radicals. Chronic exposure of transplanted isletsto the liver would lead to high portal vein levels of nutrients and gut hormones; theresulting hypersecretion of undiluted insulin into surrounding hepatocytes wouldelicit a powerful lipogenic response, overloading the nearby hepatocytes with tri-acylglycerol. Islets would be chronically exposed to both a uniquely high lipidenvironment and a high glucose environment. This combination would result inglucolipotoxicity [61]. This hypothesis is supported by liver biopsy evidence [39,62, 63] and by the fact that fatty livers occur in 20% of subjects in association withgraft dysfunction [64]. The potential pathogenic consequences of the lipid excessare suggested by the demonstration that exposure of isolated human islets to fattyacids damages β-cells and directly or indirectly results in apoptosis [65].
The recognition of these problems has renewed the interest in the search foran alternative site for implantation such as the intramuscular site and the omentalpouch [66].

32 Modulation of Early Inflammatory Reactions 729
32.3 Main Biological Events Triggering Early Graft Failureof Transplanted Pancreatic Islets
Among the components concurring to the outcome of islet transplantation, theinstant blood-mediated inflammatory reaction (IBMIR) is considered a crucialevent associated with early loss of function of transplanted islets. The IBMIRis a thrombotic reaction occurring when purified human islets are incubated inABO-compatible blood. This reaction causes morphology disruption of those isletsentrapped within a thrombus [67, 68]. The IBMIR is a likely cause of both loss oftransplanted tissue and the intraportal thrombosis associated with clinical islet trans-plantation. The IBMIR is triggered by production of tissue factor (TF) and secretionby the endocrine cells of the islets of Langerhans in the islet preparation [42]. Afterinitial generation of thrombin, by TF-expressing islets, thrombin-activated plateletsstart to bind to the islet surface. Via the amplification loop involving factor XI andactivated platelets [69], more thrombin is formed, generating a fibrin capsule sur-rounding the islets. The IBMIR occurs in clinical islet transplantation as shown byan increase in concentrations of thrombin–antithrombin complex immediately afterislet infusion [42], even without clinical signs of intraportal thrombosis. Indeed,intraportal thrombosis can occur since the thrombus does not originate from thevessel wall but from the transplanted islets and it is therefore not occlusive. TheIBMIR culminates in the disruption of islet morphology by infiltrating leukocytes.Polymorphonuclear cells (PMN) were found to be the predominant cell type infil-trating the islets in vitro [67, 68]. PMN appeared already 15 minutes after incubationwith ABO-compatible blood, with massive infiltration occurring within 1 hour andpeaked at 2 hours. Macrophages were also found to infiltrate the islets, althoughthe number of infiltrated cells increased slightly over time [70]. B and T cellswere not detected at all in the islets during the whole period of incubation, sug-gesting that the specific immune response is not involvedin the early phase of theIBMIR. In line with this hypothesis, immunosuppressive therapy presently used inT1D patients undergoing islet transplantation does not affect IBMIR [71]. PMN andmacrophage recruitment and infiltration into transplanted islets is supposed to be acrucial event in their loss of function. It is worthwhile mentioning that islet β-cellsare exquisitely susceptible to oxidative stress because of their insufficient antiox-idant pool [72], a situation that points to a rapid and direct damage to the isletsby infiltrating PMN and macrophages. In addition, infiltrating PMN are supposedto be directly involved in monocyte and T-lymphocyte recruitment, since it wasdemonstrated that PMN release chemotactic factors for T cells and macrophages[73, 74].
The mechanism(s) by which the newly transplanted islets stimulate PMN andmonocyte recruitment are not completely understood. It was suggested that leuko-cyte infiltration is a result of complement activation [67, 68]. The anaphylatoxinsC3a and C5a, released upon activation of IBMIR cascade events, were supposed tobe responsible at least in part for leukocyte recruitment and infiltration into trans-planted islets. However, a consistent leukocyte infiltration into the islets was alsoobserved when both coagulation and complement activation were abrogated, clearly

730 L. Piemonti et al.
indicating that mechanisms other than complement activation are responsible forleukocyte recruitment [68]. In the same context, it was hypothesized that leukocyterecruitment could be elicited by proinflammatory mediators released by the islets[75, 76]. The mechanism responsible for PMN and monocyte/macrophage recruit-ment could be, respectively, CXCL8 and CCL2 production. Indeed, islets have beenshown to express CXCL8 and CCL2 [77, 75] and this event might trigger the inflam-matory reaction at the site of transplant and may play a relevant role in the clinicaloutcome of islet transplantation. It was demonstrated that primary cultures of pan-creatic islets expressed and secreted CCL2 and low production of CCL2 by theislets resulted as the most relevant factor for long-lasting insulin independence [75].In addition, a significant relationship between TF and CCL2 released in vitro bythe islets and plasma biochemical parameters of coagulation in patients after islettransplantation was demonstrated [36].
Overall, these results suggest that the infiltration pattern observed during theIBMIR resembles, at least in part, that detected in the reperfused organs. In thiscontext, PMN are attracted to the graft due to upregulation and release of agentsby the ischemia-induced alterations of the endothelial cells and parenchyma cellsof the transplanted organ. Similarly, PMN and subsequent mononuclear leukocyterecruitment to the islets could also be due to induced specific chemotactic mediatorsreleased from islets and/or infiltrating PMN themselves. The massive infiltration byPMN probably causes direct damage to the islets, not only by functionally impair-ing or reducing the mass of the implanted islets but probably also by amplifying thesubsequent immune responses [78].
Recently, the inflammatory reaction observed in the liver of the recipient afterislet transplantation has been considered as an additional component, other thanIBMIR, that could contribute, together with the subsequent immune responses, toearly graft failure. In this context, intrahepatic islet infusion in humans, as well asin experimental animal models, is associated with histopathologic changes againresembling at least in part the post-ischemic organ situation. Indeed, histologicalevaluation of livers following portal vein islet transplant showed the presence ofembolism, thrombosis, and abundant areas of liver necrosis around the transplantedislets [44, 45]. In experimental animal models of allogeneic islet transplantation,ischemia and necrosis of the liver reach a peak at 1 day after islet transplantation,are reduced by day 3, and are largely resolved by day 7 [79]. The interpretationof these observations is that islet transplant blocks the blood flow to the capillarybed resulting in acute necrosis of the surrounding liver tissue. This phenomenon isconsidered to be clinically relevant since no substantial dimensional differences insmall liver vessels and capillaries are evident in mice and men, suggesting that theportal size difference should not be relevant at the levels of single islet microen-vironment in terms of ischemia and reperfusion. Starting from day 2 after isletallotransplantation, leukocytes, mainly PMN, infiltrate the necrotic hepatic regions.Subsequently, a mononuclear leukocyte infiltrate colonizes islets, starting as peri-islets and becoming intra-islets, with progressive endocrine tissue destruction andloss of the insulin production [79] (Fig. 32.1). Again, infiltrated PMN, as demon-strated in post-ischemic situations, could have a crucial physiopathological role indirectly inducing tissue damage and islet loss of function as well as in orchestrating
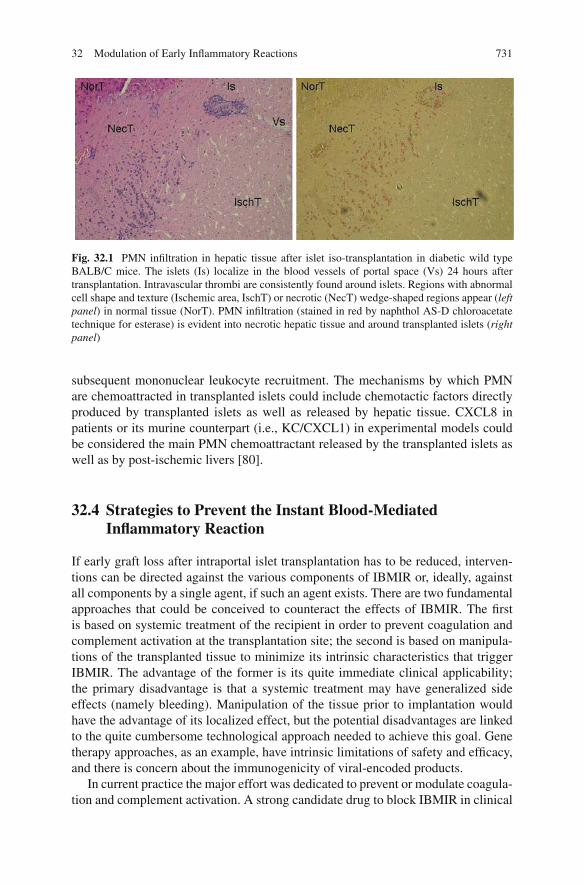
32 Modulation of Early Inflammatory Reactions 731
Fig. 32.1 PMN infiltration in hepatic tissue after islet iso-transplantation in diabetic wild typeBALB/C mice. The islets (Is) localize in the blood vessels of portal space (Vs) 24 hours aftertransplantation. Intravascular thrombi are consistently found around islets. Regions with abnormalcell shape and texture (Ischemic area, IschT) or necrotic (NecT) wedge-shaped regions appear (leftpanel) in normal tissue (NorT). PMN infiltration (stained in red by naphthol AS-D chloroacetatetechnique for esterase) is evident into necrotic hepatic tissue and around transplanted islets (rightpanel)
subsequent mononuclear leukocyte recruitment. The mechanisms by which PMNare chemoattracted in transplanted islets could include chemotactic factors directlyproduced by transplanted islets as well as released by hepatic tissue. CXCL8 inpatients or its murine counterpart (i.e., KC/CXCL1) in experimental models couldbe considered the main PMN chemoattractant released by the transplanted islets aswell as by post-ischemic livers [80].
32.4 Strategies to Prevent the Instant Blood-MediatedInflammatory Reaction
If early graft loss after intraportal islet transplantation has to be reduced, interven-tions can be directed against the various components of IBMIR or, ideally, againstall components by a single agent, if such an agent exists. There are two fundamentalapproaches that could be conceived to counteract the effects of IBMIR. The firstis based on systemic treatment of the recipient in order to prevent coagulation andcomplement activation at the transplantation site; the second is based on manipula-tions of the transplanted tissue to minimize its intrinsic characteristics that triggerIBMIR. The advantage of the former is its quite immediate clinical applicability;the primary disadvantage is that a systemic treatment may have generalized sideeffects (namely bleeding). Manipulation of the tissue prior to implantation wouldhave the advantage of its localized effect, but the potential disadvantages are linkedto the quite cumbersome technological approach needed to achieve this goal. Genetherapy approaches, as an example, have intrinsic limitations of safety and efficacy,and there is concern about the immunogenicity of viral-encoded products.
In current practice the major effort was dedicated to prevent or modulate coagula-tion and complement activation. A strong candidate drug to block IBMIR in clinical

732 L. Piemonti et al.
islet transplantation is the low molecular weight dextran sulfate (LMW-DS; MM5000), today available for clinical use, that inhibits both complement and coagula-tion activation [81, 82]. In in vitro studies, replacement of heparin by low molecularweight dextran sulfate blocked IBMIR to a greater extent. In in vivo rodent stud-ies, treatment of the recipient with dextran sulfate significantly prolonged survivalof intraportally transplanted islets [83]. Based on these evidences a clinical trial toassess the safety and effectiveness of LMW-SD on post transplant islet function inpeople with T1D is currently ongoing (ClinicalTrials.gov Identifier:NCT00790439).
Other anticoagulant or complement inhibitors have been shown to prevent isletdamage in vitro or in vivo: melagatran, a specific thrombin inhibitor [68]; nacyste-lyn, a derivative of N-acetylcysteine [84, 85]; activated protein C [38, 86]; sCR1,a complement inhibitor [87]; TP10, the soluble complement receptor 1 [88]; lowmolecular mass factor VIIa inhibitor [41]; nicotinamide [89]. In general, all thesestrategies aimed at inhibiting the IBMIR have shown only a modest benefit in a lim-ited series of in vivo studies in animal models. It is unlikely that an agent targetingonly one component of IBMIR would block all elements of the reaction (coagula-tion, complement activation, production of proinflammatory mediators); therefore,the effect of a single agent on the engraftment may be limited. The identificationof new strategies to reduce the detrimental effects of IBMIR should be one of theobjectives of the research in the next years. For example, PMN recruitment andinfiltration into transplanted islets is a key pathophysiological event responsible fordirect damage of islet functionality and, at least in part, of subsequent mononuclearleukocyte infiltration and related loss of insulin production. Among chemotacticmediators, CXCL8 in transplanted patients, and the murine counterpart in experi-mental transplants, is supposed to be a crucial mediator in PMN recruitment intotransplanted islets and thus represents, together with its receptors (CXCR1 andCXCR2), a primary therapeutic target to prevent early graft failure.
Manipulation of the tissue prior to implantation has the advantage of its local-ized effect. Surface heparinization of islets is an attractive alternative to solubleheparin. It provides a means to render biocompatible the islet surface when exposedto blood, thereby mimicking the protective characteristics conferred by heparan sul-fate on the endothelial cells lining the vascular wall. In addition to the effects onthe cascade systems and on the circulating cells, heparin coating reduces expo-sure of collagen and other extracellular matrix proteins on the islets that may beprothrombotic and trigger inflammation. On this basis it was demonstrated that mod-ification of pancreatic islets with surface-attached heparin or thrombomodulin canreduce the deleterious IBMIR associated with islet transplantation [90]. Similarly,since endothelial cells (EC) readily tolerate contact with blood, a conceivable strat-egy to overcome IBMIR would be to create composite islet-endothelial cell grafts.This approach was recently reported. Human islets were cocultured with primaryhuman aortic endothelial cells (HAEC) for 2–7 days to obtain 50–90% coverage[91]. Exposed to blood, HAEC-coated islets induced less activation of coagulationand complement compared to control islets with decreased platelet and leukocyteconsumption and less infiltration of CD11b+ cells in clots. After transplantationto athymic nude mice, composite islet-HAEC grafts stained positive for insulin and

32 Modulation of Early Inflammatory Reactions 733
PECAM-1 demonstrating the presence of both islets and HAEC within the islet graft7 weeks after transplantation. The refinement of this technique could allow introduc-tion of composite islet-EC grafts in clinical islet transplantation, using autologousEC expanded in vitro and kept frozen until allogeneic islets become available for thatspecific recipient. Similar results were reported using composite pig islet-human ECgraft [92]. More recently it was reported that addition of mesenchymal stem cellsto composite islets enhanced the capacity of EC to enclose the islets without com-promising the islet functionality. Moreover, the mesenchymal stem cells stimulatedEC sprout formation not only into the surrounding matrices but also into the isletswhere intra-islet capillary-like structures were formed [93].
32.5 Strategies for Cytoprotection and Revascularization
Methods that favor islet engraftment by modulating islet cell resistance to the nox-ious stimuli and/or the level of inflammation at the transplant site and/or the levelof vascularization may result in long-term insulin independence after transplanta-tion of a reduced number of islets. Induction of islet cytoprotection to reduce and/orprevent the negative effects of noxious stimuli may be achieved by multiple means,including preconditioning of the graft in culture prior transplantation and/or treat-ment of the recipients in the peri-transplant period. Several approaches have beenproposed toward this goal, including the use of a number of cytoprotective regimensvia pharmacological administration, gene therapy, gene silencing, and protein trans-duction domains. Potential candidate molecules that have been used in experimentalstudies include 17-β-estradiol, nicotinamide, metal protoporphyrins, glucagon-likepeptide-1 (GLP-1), which may be used during isolation, added in the culture media,or administered to the recipient as they may avoid or partially prevent the effectsof oxidative stress and proinflammatory cytokines early after transplant, thereforemaximizing islet engraftment.
An example of molecule useful for both pretreatment of isolated islets and recip-ient is α1-antitrypsin, a major protease inhibitor that inhibits the enzyme activityof neutrophil elastase and thrombin. In vitro, in the presence of α1-antitrypsin,mouse islets were protected against the effect of the cytokines, IL-1β, and IFN-γ, bymeans of greater viability, a 40% reduction in nitric oxide production, and greatlydiminished TNF-α production. Moreover, administration of human α1-antitrypsin torecipient mice improved islet survival [94–96] and would represent a safe approachin the clinic.
Similarly it was recently reported that methylprednisolone is efficient in reduc-ing the inflammatory status of human islets and thus has the potential to improvegraft function following islet transplantation [97]. As a consequence, some groupshave introduced glucocorticoid preconditioning of the islet preparation priortransplantation. In addition, in these centers all organ donors are given methyl-prednisolone prior procurement [97]. Moreover, antioxidant supplementation tothe islet culture medium for scavenging oxygen radicals helps human islets to

734 L. Piemonti et al.
reduce their inflammatory state, determined by reduction of cytokine and MCP-1expression [98].
Several factors may improve graft angiogenesis and vascularity. Vascularendothelial growth factor-A, a well-known angiogenic factor that is secreted byislets in response to hypoxia, appears to play a significant role in angiogenesis andimprovement of graft function [99, 100]. Additional growth factors, such as platelet-derived growth factor, epidermal growth factor, and fibroblast growth factor, arealso thought to promote graft revascularization [101]. Overexpression of moleculesknown to enhance revascularization such as VEGF has been attempted, but thesemolecules have yet to exhibit a significant impact on islet graft survival [102, 103].This is likely related to the fact that although vascular endothelial growth factorexpression will hasten the revascularization process, it cannot provide an immediatebenefit to the transplanted tissue.
Ex vivo gene transfer to isolated islets has been performed using several genecandidates including interleukin-1 receptor antagonist [104, 105], TNF-α antagonist[46, 106–108], heme oxygenase I [109, 110], insulin-like growth factor-I [111, 112],dominant negative protein kinase C[113], dominant negative MyD88 [114], nuclearfactor B [115], inhibitor of B repressor [116], heat shock protein 70 [113], man-ganese superoxide dismutase [118], and catalase [119] for immune, inflammation,and apoptosis protection.
Several different molecules that inhibit the generation of and/or damage mediatedby reactive oxygen species (ROS), including glutathione peroxidase, superoxide dis-mutase, and heme oxygenase-1 have tested [109, 110, 120, 121]. Although thesemolecules can individually protect islets during controlled in vitro challenges whereROS are specifically produced, the in vivo benefit of such an approach has onlybeen demonstrated when glutathione peroxidase and superoxide dismutase wereco-expressed in transgenic islet grafts [122].
Significant efforts have been also made to inhibit specific apoptotic triggers,either extrinsic (cFLIP and A20) or intrinsic (BCL-2 and BCL-XL). These proteinshave proven to be quite effective in enhancing β-cell survival in vitro. However,reproducing the protective effect using transplanted islets has been difficult andlargely unfruitful. Recently, investigations of downstream apoptosis inhibitors thatblock the final common pathway (i.e., X-linked inhibitor of apoptosis protein[XIAP]) have demonstrated promise in both human and rodent models of engraft-ment [123, 124]. The studies using XIAP strongly support the concept that inhibitionof apoptosis at the level of caspases promotes β-cell survival in islet transplantation,effectively preventing cell death triggered by extrinsic and intrinsic pathways at thesame time.
32.6 Modification of Transplant Site and Biomaterial-BasedStrategies to Improve Engraftment
A simple and attractive approach to improve islet transplantation engraftment is toconsider whether sites and techniques other than intraportal infusion into the liverare better adapted for islet implantation and survival. Although many implantation

32 Modulation of Early Inflammatory Reactions 735
sites have been proposed, few have found their way into the clinical setting [125].In one clinical investigation using infusion of islets under the kidney capsule, twoof three recipients showed autograft survival, measured by C-peptide secretion.However, the high transplant mass at this site relative to intraportal infusions doesnot justify the approach [126]. The intraperitoneal and omental pouch sites are alsoattractive for clinical islet cell transplantation [127] as they allow a large implan-tation volume and the concurrent use of transplant devices or capsules [128]. Acase report of autologous islet transplantation into the brachioradialis muscle of a7-year-old girl who had complete pancreatectomy for severe hereditary pancreati-tis described C-peptide levels maintained even 2 years after transplantation [66].The patient received conservative insulin therapy via a pump, but the maintainedfunction of the transplant was believed to contribute to good glycemic control andprevention of hypoglycemic events.
Applications for biomaterials in improving islet engraftment by immunoisola-tion of the transplanted tissue through semipermeable membranes are increasing[129–131]. Three different kinds of encapsulated systems can be used for the pur-pose of islet transplantation: (1) perfusion chambers directly connected to the bloodcirculation (intravascular macrocapsules), (2) diffusion chambers in the shape ofa tube or disk that can be implanted i.p. or s.c. (extravascular macrocapsules), or(3) the encapsulation of one or few islets in globular membranes (extravascularmicrocapsules). Intravascular macrocapsules are based on the principle of “dialy-sis cartridges” in which islets are seeded in the space between hollow fibers that areperfused with blood. The islets may be in a packed form or dispersed in a spacermatrix that prevents mutual adhesion and improves diffusional nutrient transport ofthe islets. These hollow fibers are enclosed within a larger tube, and the device isimplanted into the vessels of the host by vascular anastomoses. Biomaterial used forthe construction of these microcapillaries is polyacrylonitrile and polyvinylchlo-ride copolymer, a biocompatible matrix often used in spinal cord injury. Thesedevices permit close contact between the bloodstream and the islets, leading toefficient diffusional transport of metabolites. Encapsulation of islets in this devicehas been shown to induce normoglycemia in various animal models of diabetesincluding rats, dogs, and monkeys [132, 133]. The duration of this normoglycemiawas usually restricted to several hours and success of a somewhat longer dura-tion was exceptional. Blood clotting in the lumen of these small-diameter artificialcapillaries proved to be a major obstacle in spite of intense systemic anticoag-ulation, which is indicative of low biocompatibility of the implant material. Anincrease in the diameter of the capillaries led to increased flow rate of blood andreduced the risk of thromboembolism, but not without accompanying risks plusthe complications associated with vascular prosthetic surgery [134]. These con-siderations shifted the research focus toward extravascular macrocapsules for isletengraftment.
Extravascular macrocapsules are based on the same principles as intravascularones but have the advantage that biocompatibility issues do not pose a serious riskto the patient. These devices have been designed in both flat sheet membranous andhollow fiber formats [135]. They can be implanted into the peritoneal cavity, the sub-cutaneous tissue, or under the kidney capsule. Various biomaterials have been used

736 L. Piemonti et al.
to generate these devices including nitrocellulose acetate, 2-hydroxyethyl methacry-late (HEMA), acrylonitrile, polyacrylonitrile and polyvinylchloride copolymer,sodium methallylsulfonate, and alginate. Biocompatibility of these devices is seenin terms of fibrosis at the site of implantation and covering the device. Variousapproaches have been used to enhance the biocompatibility of these devices, includ-ing the use of hollow fiber geometry because it offers reduced surface area ofcontact with the host per islet. Use of a smooth outer surface and hydrogelsfurther improves biocompatibility of these devices by the absence of interfa-cial tension, thus reducing protein adsorption and cell adhesion. Hydrogels alsoprovide higher permeability for low molecular weight nutrients and metabolites.Hydrogel materials that have been used include alginate [136], agarose [137–139], polyurethane [140], chitosan–polyvinyl pyrrolidone hydrogels [141], cellulose[142], cross-linked hydrophilic poly(N,N-dimethyl acrylamide) with hydrophobicdi-, tri-, and octamethacrylate telechelic polyisobutylene [143], and a copolymerof acrylonitrile and sodium methallylsulfonate [144]. Other approaches to addressthe problem of biocompatibility of these devices include membrane coating withpoly-ethylene-oxide to reduce surface protein adsorption and surface hydrophobiza-tion with corona discharge. Surface fibrosis and biocompatibility remain the mostsignificant hurdles to the successful use of both macrocapsule and microcapsuledevices.
Extravascular microcapsules enclose one or a few islets and are implanted atextravascular sites for obvious reasons. Microencapsulation of islets offers sev-eral advantages over macroencapsulation: higher surface area per unit volume forbetter diffusive transfer of nutrients and metabolites, mechanical stability, ease inmanufacturing, and easy implanting procedures. Encapsulated islets have shownimproved graft function and survival compared with unencapsulated islets [145,146]. The long-term survival and function of islets microencapsulate, however, islimited. Primary impediments to the success of microencapsulation for islet trans-plantation include: (1) biocompatibility, (2) inadequate immunoprotection, and (3)hypoxia. Inadequate biocompatibility is recognized by the pericapsular overgrowthon microcapsules that consist of fibroblasts and macrophages [147, 148]. Moreover,since encapsulation precludes the ingrowth of blood vessels and islet revascular-ization is important for long-term islet function, use of encapsulated islets hasbeen complicated by cell death secondary to chronic hypoxia and/or decreasedaccessibility to nutrients and growth factors. Islets microencapsulated within analginate-poly-(L-lysine) membrane and an agarose hydrogel membrane have beeninvestigated for use as a bioartificial pancreas. Many groups have reported that along-term normoglycemia in a diabetic small animal, such as a mouse or a rat,can be realized by transplanting microencapsulated islets into its peritoneal cav-ity. However, in clinical settings, about 10 ml of islet suspension should be injectedthrough a catheter into the portal vein in the liver. The diameter of microencap-sulated islets was several times larger than that of islets, which could result inplugged vessels if infused into the portal vein. A report documented normoglycemiain a human patient with intraportal transplantation of microencapsulated islets for aperiod of 9 months [149].

32 Modulation of Early Inflammatory Reactions 737
An emerging strategy to improve engraftment avoiding the limit of encapsulationis the use of synthetic biocompatible microporous polymer scaffold [127, 150–153].The polymer scaffolds are porous and not intended to serve as an immune bar-rier. Rather, they were specifically designed to provide a solid support for isletsthat would allow cellular infiltration and formation of a vascular network withinthe transplant graft. Several basic requirements for cell transplantation on micro-porous scaffolds have been identified, including biocompatibility, a high surfacearea/volume ratio with sufficient mechanical integrity, and a suitable environmentfor new tissue formation that can integrate with the surrounding tissue [154, 155].Microporous scaffolds with a high surface area/volume ratio not only have sufficientsurface area to support cell adhesion but also can support nutrient transport by dif-fusion from surrounding tissue. Moreover, they can be fabricated from material thathas sufficient mechanical properties to resist collapse while maintaining an inter-connected pore structure that allows for cell infiltration from the surrounding tissue.This is important not only for integration of the engineered tissue with the host butalso for development of a vascular network throughout the tissue to supply the nec-essary metabolites once the transplanted cells are engrafted. Encouraging reportsindicate that a synthetic polymer scaffold can serve as a platform for islet transplan-tation and improves the function of extrahepatically transplanted islets comparedto islets transplanted without a scaffold. The scaffold may also be useful to deliverbioactive molecules to modify the microenvironment surrounding the transplantedislets and, thus, enhance islet survival and function.
Camouflaging the surface of islets instead of incorporating them in a membranebarrier or in a scaffold is another approach to improve engraftment. This processinvolves attachment of polymeric, hydrophilic chains to the islet surface to achievemolecular coating of the islets. Surface modification of islets by bioconjugation canovercome several potential problems with islet encapsulation. The diffusional bar-rier of less resistance and reduced thickness can be generated around the islets,compared with an encapsulation membrane. Furthermore, fine surface coating ofislets leads to reduced volume of tissue per equivalent islet, which makes transplant-ing islets into human subjects feasible via the portal venous route of administration[156]. Strategies for surface coating of islets essentially use linear hydrophilicpolymers such as PEG with an activated functional group and a mild conjugationreaction. Surface coating of rat islets with PEG was first reported by Panza et al.[157] and was subsequently shown to be cytoprotective for porcine islet xenotrans-plantation in diabetic SCID mice [158]. Panza and colleagues demonstrated that theviability of islets was not compromised upon PEGylation and that islets retainedthe in vitro insulin response to glucose stimulation activity. Xie et al. additionallydemonstrated protection in vitro against human antibody/complement-induced cyto-toxicity in coated porcine islets and in in vivo islet function in the diabetic SCIDmice model [158]. Xie et al. further introduced the concept of albumin shielding ofislets using a disuccinimidyl derivative that is attached on one end to the islets andon the other end to an albumin moiety [158]. This concept harbors the possibility ofmodifying encapsulation technology to literally “build” capsules on the islet surfaceinstead of “encapsulating” islets. Heterobifunctional PEGs can be conjugated to the

738 L. Piemonti et al.
islet surface followed by attachment of another moiety on the exposed end of thePEG chains, which can then be cross-linked to each other to result in a firm micro-capsule. Formation of a microcapsule in this manner will obviate many problemsassociated with the processing technology of microencapsulation discussed earlier[159–161].
Methods for immobilizing enzyme and glycosaminoglycans, such as urokinaseand heparin, to the surface of islets utilizing layer-by-layer approach were described[162]. The surface of islets was modified with a poly(ethylene glycol)-phospholipidconjugate bearing a biotin group (biotin-PEG-lipids, PEG MW: 5000). Biotin-PEG-lipids were anchored to the cell membranes of islets, and the PEG-lipid layer onthe islets was further covered by streptavidin and biotin—bovine serum albuminconjugate using a layer-by-layer method. The surface was further activated withoxidized dextran. Urokinase was anchored to the islets through Schiff base forma-tion. Heparin was anchored to the islets through polyion complex formation betweenanionic heparin and a cationic protamine coating on the islets. No practical islet vol-ume increase was observed after surface modification, and the modifications did notimpair insulin release in response to glucose stimulation. The anchored urokinaseretained high fibrinolytic activity, which could help to improve graft survival bypreventing thrombosis on the islet surface.
32.7 Concluding Remarks
Clinical islet transplantation is currently being explored as a treatment for personswith type 1 diabetes and hypoglycemia unawareness. Although ‘proof of princi-ple’ has been established in recent clinical studies, the procedure suffers from lowefficacy and a large number of islets are needed to obtain insulin independencein clinical islet transplantation, requiring two to four cadaveric pancreases. Isletengraftment remains an unresolved problem in humans and the possibility of havingclinically applicable solutions will be one of the determinants of success or failureof islet transplantation in the coming years.
References
1. Franco OH, Steyerberg EW, Hu, FB, Mackenbach J, Nusselder W. Associations of dia-betes mellitus with total life expectancy and life expectancy with and without cardiovasculardisease. Arch Intern Med 2007:167:1145–51.
2. Hu FB, Stampfer MJ, Solomon CG, Liu S, Willett WC, Speizer FE, Nathan DM, MansonJE. The impact of diabetes mellitus on mortality from all causes and coronary heart diseasein women: 20 years of follow-up. Arch Intern Med 2001;161:1717–23.
3. Robertson RP, Davis C, Larsen J, Stratta R, Sutherland DE. Pancreas and islet transplantationfor patients with diabetes. Diabetes care 2000;23:112–6.
4. Shapiro AM, Lakey JR, Ryan EA, Korbutt GS, Toth E, Warnock GL, Kneteman NM,Rajotte RV. Islet transplantation in seven patients with type 1 diabetes mellitus using aglucocorticoid-free immunosuppressive regimen. N Engl J Med 2000;343:230–8.

32 Modulation of Early Inflammatory Reactions 739
5. Shapiro AM, Ricordi C, Hering BJ, Auchincloss H, Lindblad R, Robertson RP, Secchi A,Brendel MD, Berney T, Brennan DC, et al. International trial of the Edmonton protocol forislet transplantation. N Engl J Med 2006;355:1318–30.
6. Ryan EA, Lakey JR, Rajotte RV, Korbutt GS, Kin T, Imes S, Rabinovitch A, Elliott JF, BigamD, Kneteman NM, et al. Clinical outcomes and insulin secretion after islet transplantationwith the Edmonton protocol. Diabetes 2001;50:710–9.
7. Ryan EA, Paty BW, Senior PA, Bigam D, Alfadhli E, Kneteman NM, Lakey JR, ShapiroAM. Five-year follow-up after clinical islet transplantation. Diabetes 2005;54:2060–9.
8. Kemp CB, Knight MJ, Scharp DW, Ballinger WF, Lacy PE. Effect of transplantation site onthe results of pancreatic islet isografts in diabetic rats. Diabetologia 1973;9:486–91.
9. Largiader F, Kolb E, Binswanger U. A long-term functioning human pancreatic isletallotransplant. Transplantation 1980;29:76–7.
10. Najarian JS, Sutherland DE, Baumgartner D, Burke B, Rynasiewicz JJ, Matas AJ, GoetzFC. Total or near total pancreatectomy and islet autotransplantation for treatment of chronicpancreatitis. Ann Surg 1980;192:526–42.
11. Sutherland DE, Matas AJ, Goetz FC, Najarian JS. Transplantation of dispersed pancreaticislet tissue in humans: autografts and allografts. Diabetes 29 Suppl 1980;1:31–44.
12. Scharp DW, Lacy PE, Santiago JV, McCullough CS, Weide LG, Falqui L, Marchetti P,Gingerich RL, Jaffe AS, Cryer PE, et al. Insulin independence after islet transplantationinto type I diabetic patient. Diabetes 1990;39:515–8.
13. Toyofuku A, Yasunami Y, Nabeyama K, Nakano M, Satoh M, Matsuoka N, Ono J,Nakayama T, Taniguchi M, Tanaka M, Ikeda S. Natural killer T-cells participate in rejectionof islet allografts in the liver of mice. Diabetes 2006;55:34–9.
14. Yasunami Y, Kojo S, Kitamura H, Toyofuku A, Satoh M, Nakano M, Nabeyama K,Nakamura Y, Matsuoka N, Ikeda S, et al. Valpha14 NK T cell-triggered IFN-gamma pro-duction by Gr-1+CD11b+ cells mediates early graft loss of syngeneic transplanted islets. JExp Med 2005;202:913–8.
15. Carlsson PO, Palm F, Andersson A, Liss P. Markedly decreased oxygen tension in trans-planted rat pancreatic islets irrespective of the implantation site. Diabetes 2001;50:489–95.
16. Korsgren O, Lundgren T, Felldin M, Foss A, Isaksson B, Permert J, Persson NH, Rafael E,Ryden M, Salmela K, et al. Optimising islet engraftment is critical for successful clinicalislet transplantation. Diabetologia 2008;51:227–32.
17. Al-Jazaeri A, Xu, BY, Yang H, Macneil D, Leventhal JR, Wright JR, Jr. Effect of glucosetoxicity on intraportal tilapia islet xenotransplantation in nude mice. Xenotransplantation2005;12:189–196.
18. Desai NM, Goss JA, Deng S, Wolf BA, Markmann E, Palanjian M, Shock AP, Feliciano S,Brunicardi FC, Barker CF, et al. Elevated portal vein drug levels of sirolimus and tacrolimusin islet transplant recipients: local immunosuppression or islet toxicity? Transplantation2003;76:1623–5.
19. Gupta V, Wahoff DC, Rooney DP, Poitout V, Sutherland DE, Kendall DM, Robertson RP.The defective glucagon response from transplanted intrahepatic pancreatic islets duringhypoglycemia is transplantation site-determined. Diabetes 1997;46:28–33.
20. Hafiz MM, Faradji RN, Froud T, Pileggi A, Baidal DA, Cure P, Ponte G, Poggioli R,Cornejo A, Messinger S, et al. Immunosuppression and procedure-related complicationsin 26 patients with type 1 diabetes mellitus receiving allogeneic islet cell transplantation.Transplantation 2005;80:1718–28.
21. Maleux G, Gillard P, Keymeulen B, Pipeleers D, Ling Z, Heye S, Thijs M, Mathieu C,Marchal G. Feasibility, safety, and efficacy of percutaneous transhepatic injection of beta-cell grafts. J Vasc Interv Radiol 2005;16:1693–7.
22. Owen RJ, Ryan EA, O’Kelly K, Lakey JR, McCarthy MC, Paty BW, Bigam DL, KnetemanNM, Korbutt GS, Rajotte RV, Shapiro AM. Percutaneous transhepatic pancreatic islet celltransplantation in type 1 diabetes mellitus: radiologic aspects. Radiology 2003;229:165–70.
23. Venturini M, Angeli E, Maffi P, Fiorina P, Bertuzzi F, Salvioni M, De Cobelli F, Socci C,Aldrighetti L, Losio C, et al. Technique, complications, therapeutic efficacy of percutaneous

740 L. Piemonti et al.
transplantation of human pancreatic islet cells in type 1 diabetes: the role of US. Radiology2005;234:617–24.
24. Neeman Z, Hirshberg B, Harlan D, Wood BJ. Radiologic aspects of islet cell transplantation.Curr Diabetes Rep 2006;6:310–5.
25. Villiger P, Ryan EA, Owen R, O’Kelly K, Oberholzer J, Al Saif F, Kin T, Wang H, LarsenI, Blitz SL, et al. Prevention of bleeding after islet transplantation: lessons learned from amultivariate analysis of 132 cases at a single institution. Am J Transplant 2005;5:2992–8.
26. Meier JJ, Hong-McAtee I, Galasso R, Veldhuis JD, Moran A, Hering BJ, Butler PC.Intrahepatic transplanted islets in humans secrete insulin in a coordinate pulsatile mannerdirectly into the liver. Diabetes 2006;55:2324–32.
27. Meier JJ, Veldhuis JD, Butler PC. Pulsatile insulin secretion dictates systemic insulindelivery by regulating hepatic insulin extraction in humans. Diabetes 2005;54:1649–56.
28. Polonsky KS, Given BD, Hirsch L, Shapiro ET, Tillil H, Beebe C, Galloway JA, Frank BH,Karrison T, Van Cauter E. Quantitative study of insulin secretion and clearance in normaland obese subjects. J Clin Invest 1988;81:435–41.
29. Waldhausl W, Bratusch-Marrain P, Gasic S, Korn A, Nowotny P. Insulin production ratefollowing glucose ingestion estimated by splanchnic C-peptide output in normal man.Diabetologia 1979;17:221–7.
30. Lau J, Jansson L, Carlsson PO. Islets transplanted intraportally into the liver are stimu-lated to insulin and glucagon release exclusively through the hepatic artery. Am J Transplant2006;6:967–75.
31. Kendall DM, Teuscher AU, Robertson RP. Defective glucagon secretion during sustainedhypoglycemia following successful islet allo- and autotransplantation in humans. Diabetes1997;46:23–7.
32. Zhou H, Zhang T, Bogdani M, Oseid E, Parazzoli S, Vantyghem MC, Harmon J, SluccaM, Robertson RP. Intrahepatic glucose flux as a mechanism for defective intrahepatic isletalpha-cell response to hypoglycemia. Diabetes 2008;57:1567–74.
33. Mattsson G, Jansson L, Nordin A, Andersson A, Carlsson PO. Evidence of functionalimpairment of syngeneically transplanted mouse pancreatic islets retrieved from the liver.Diabetes 2004;53:948–54.
34. Barshes NR, Lee TC, Goodpastor SE, Balkrishnan R, Schock AP, Mote A, Brunicardi FC,Alejandro R, Ricordi C, Goss JA. Transaminitis after pancreatic islet transplantation. J AmColl Surg 2005a;200:353–61.
35. Barshes NR, Wyllie S, Goss JA. Inflammation-mediated dysfunction and apoptosisin pancreatic islet transplantation: implications for intrahepatic grafts. J Leukoc Biol2005b;77:587–97.
36. Bertuzzi F, Marzorati S, Maffi P, Piemonti L, Melzi R, de Taddeo F, Valtolina V, D’AngeloA, di Carlo V, Bonifacio E, Secchi A. Tissue factor and CCL2/monocyte chemoattractantprotein-1 released by human islets affect islet engraftment in type 1 diabetic recipients. JClin Endocrinol Metabol 2004;89:5724–8.
37. Bhargava R, Senior PA, Ackerman TE, Ryan EA, Paty BW, Lakey JR, Shapiro AM.Prevalence of hepatic steatosis after islet transplantation and its relation to graft function.Diabetes 2004;53:1311–7.
38. Contreras JL, Eckstein C, Smyth CA, Bilbao G, Vilatoba M, Ringland SE, Young C,Thompson JA, Fernandez JA, Griffin JH, Eckhoff DE. Activated protein C preserves func-tional islet mass after intraportal transplantation: a novel link between endothelial cellactivation, thrombosis, inflammation, islet cell death. Diabetes 2004;53:2804–14.
39. Eckhard M, Lommel D, Hackstein N, Winter D, Ziegler A, Rau W, Choschzick M, BretzelRG, Brendel MD. Disseminated periportal fatty degeneration after allogeneic intraportalislet transplantation in a patient with type 1 diabetes mellitus: a case report. Transplant Proc2004;36:1111–6.
40. Hyon SH, Ceballos MC, Barbich M, Groppa R, Grosembacher L, Vieiro MM, Barcan L,Algranati S, Litwak L, Argibay PF. Effect of the embolization of completely unpurified islets

32 Modulation of Early Inflammatory Reactions 741
on portal vein pressure and hepatic biochemistry in clinical practice. Cell Transplantation2004;13:61–65.
41. Johansson H, Lukinius A, Moberg L, Lundgren T, Berne C, Foss A, Felldin M, Kallen R,Salmela K, Tibell A, et al. Tissue factor produced by the endocrine cells of the islets ofLangerhans is associated with a negative outcome of clinical islet transplantation. Diabetes2005a;54:1755–62.
42. Moberg L, Johansson H, Lukinius A, Berne C, Foss A, Kallen R, Ostraat O, Salmela K,Tibell A, Tufveson G, et al. Production of tissue factor by pancreatic islet cells as a trigger ofdetrimental thrombotic reactions in clinical islet transplantation. Lancet 2002;360:2039–45.
43. Rafael E, Ryan EA, Paty BW, Oberholzer J, Imes S, Senior P, McDonald C, Lakey JR,Shapiro AM. Changes in liver enzymes after clinical islet transplantation. Transplantation2003;76:1280–4.
44. Sudo T, Hiyama E, Murakami Y, Yokoyama Y, Takesue Y, Sueda T. Hepatic regenerationpromotes engraftment of intraportally transplanted islet cells. Surgery 2005;137:612–9.
45. Yin D, Ding JW, Shen J, Ma, L, Hara M, Chong AS. Liver ischemia contributes to early isletfailure following intraportal transplantation: benefits of liver ischemic-preconditioning. AmJ Transplant 2006;6:60–8.
46. Contreras JL, Bilbao G, Smyth CA, Jiang XL, Eckhoff DE, Jenkins SM, Thomas FT, CurielDT, Thomas JM. Cytoprotection of pancreatic islets before and soon after transplantation bygene transfer of the anti-apoptotic Bcl-2 gene. Transplantation 2001;71:1015–23.
47. Casey JJ, Lakey JR, Ryan EA, Paty BW, Owen R, O’Kelly K, Nanji S, Rajotte RV,Korbutt GS, Bigam D, et al. Portal venous pressure changes after sequential clinical islettransplantation. Transplantation 2002;74:913–5.
48. Morrison CP, Wemyss-Holden SA, Dennison AR, Maddern GJ. Islet yield remains aproblem in islet autotransplantation. Arch Surg 2002;137:80–3.
49. Jansson L. Dissociation between pancreatic islet blood flow and insulin release in the rat.Acta physiologica Scandinavica 1985;124:223–28.
50. Jansson L. Glucose stimulation of pancreatic islet blood flow by redistribution of the bloodflow within the whole pancreatic gland. Pancreas 1988;3:409–12.
51. Jansson L. The regulation of pancreatic islet blood flow. Diabetes/Metabol Rev1994;10:407–16.
52. Menger MD, Yamauchi J, Vollmar B. Revascularization and microcirculation of freelygrafted islets of Langerhans. World J Surg 2001;25:509–15.
53. Mattsson G, Jansson L, Carlsson PO. Decreased vascular density in mouse pancreatic isletsafter transplantation. Diabetes 2002;51:1362–6.
54. Andersson A, Korsgren O, Jansson L. Intraportally transplanted pancreatic islets revascular-ized from hepatic arterial system. Diabetes 1989;38 Suppl 1:192–5.
55. Ballian N, Brunicardi FC. Islet vasculature as a regulator of endocrine pancreas function.World J Surg 2007;31:705–14.
56. Hart TK, Pino RM. Pseudoislet vascularization. Induction of diaphragm-fenestratedendothelia from the hepatic sinusoids. Laboratory Investigation; A J Tech Methods Pathol1986;54:304–13.
57. Jansson L, Carlsson PO. Graft vascular function after transplantation of pancreatic islets.Diabetologia 2002;45:749–63.
58. Brissova M, Fowler M, Wiebe P, Shostak A, Shiota M, Radhika A, Lin PC, GannonM, Powers AC. Intraislet endothelial cells contribute to revascularization of transplantedpancreatic islets. Diabetes 2004;53:1318–25.
59. Carlsson PO, Palm F, Mattsson G. Low revascularization of experimentally transplantedhuman pancreatic islets. J Clin Endocrinol Metabol 2002;87:5418–23.
60. Shapiro AM, Gallant HL, Hao EG, Lakey JR, McCready T, Rajotte RV, Yatscoff RW,Kneteman NM. The portal immunosuppressive storm: relevance to islet transplantation?Therapeutic Drug Monitor 2005;27:35–7.

742 L. Piemonti et al.
61. Lee Y, Ravazzola M, Park BH, Bashmakov YK, Orci L, Unger RH. Metabolic mechanismsof failure of intraportally transplanted pancreatic beta-cells in rats: role of lipotoxicity andprevention by leptin. Diabetes 2007;56:2295–301.
62. Hirshberg B, Mog S, Patterson N, Leconte J, Harlan DM. Histopathological study ofintrahepatic islets transplanted in the nonhuman primate model using edmonton protocolimmunosuppression. J Clin Endocrinol Metabol 2002;87:5424–9.
63. Robertson RP, Harmon JS. Pancreatic islet beta-cell and oxidative stress: the importance ofglutathione peroxidase. FEBS Lett 2007;581:3743–8.
64. Senior PA, Shapiro AM, Ackerman TE, Ryan EA, Paty BW, Bhargava R. Magneticresonance-defined perinephric edema after clinical islet transplantation: a benign findingassociated with mild renal impairment. Transplantation 2004;78:945–8.
65. Lupi R, Dotta F, Marselli L, Del Guerra S, Masini M, Santangelo C, Patane G, Boggi U, PiroS, Anello M, et al. Prolonged exposure to free fatty acids has cytostatic and pro-apoptoticeffects on human pancreatic islets: evidence that beta-cell death is caspase mediated,partially dependent on ceramide pathway, Bcl-2 regulated. Diabetes 2002;51:1437–42.
66. Rafael E, Tibell A, Ryden M, Lundgren T, Savendahl L, Borgstrom B, Arnelo U, IsakssonB, Nilsson B, Korsgren O, Permert J. Intramuscular autotransplantation of pancreatic isletsin a 7-year-old child: a 2-year follow-up. Am J Transplant 2008;8:458–62.
67. Bennet W, Sundberg B, Groth CG, Brendel MD, Brandhorst D, Brandhorst H, BretzelRG, Elgue G, Larsson R, Nilsson B, Korsgren O. Incompatibility between human bloodand isolated islets of Langerhans: a finding with implications for clinical intraportal islettransplantation? Diabetes 1999;48:1907–14.
68. Ozmen L, Ekdahl KN, Elgue G, Larsson R, Korsgren O, Nilsson B. Inhibition of throm-bin abrogates the instant blood-mediated inflammatory reaction triggered by isolatedhuman islets: possible application of the thrombin inhibitor melagatran in clinical islettransplantation. Diabetes 2002;51:1779–84.
69. Colman RW, Scott CF. When and where is factor XI activated by thrombin? Blood1996;87:2089.
70. Moberg L, Korsgren O, Nilsson B. Neutrophilic granulocytes are the predominant cell typeinfiltrating pancreatic islets in contact with ABO-compatible blood. Clin Exp Immunol2005;142:125–31.
71. Moberg L. The role of the innate immunity in islet transplantation. Ups J Med Sci2005;110:17–55.
72. Tiedge M, Lortz S, Drinkgern J, Lenzen S. Relation between antioxidant enzymegene expression and antioxidative defense status of insulin-producing cells. Diabetes1997;46:1733–42.
73. Chertov O, Yang D, Howard OM, Oppenheim JJ. Leukocyte granule proteins mobilize innatehost defenses and adaptive immune responses. Immunol Rev 2000;177:68–78.
74. Scapini P, Lapinet-Vera JA, Gasperini S, Calzetti F, Bazzoni F, Cassatella MA. Theneutrophil as a cellular source of chemokines. Immunol Rev 2000;177:195–203.
75. Piemonti L, Leone BE, Nano R, Saccani A, Monti P, Maffi P, Bianchi G, Sica A, Peri G,Melzi R, et al. Human pancreatic islets produce and secrete MCP-1/CCL2: relevance inhuman islet transplantation. Diabetes 2002;51:55–65.
76. Sordi V, Malosio ML, Marchesi F, Mercalli A, Melzi R, Giordano T, Belmonte N, FerrariG, Leone BE, Bertuzzi F, et al. Bone marrow mesenchymal stem cells express a restrictedset of functionally active chemokine receptors capable of promoting migration to pancreaticislets. Blood 2005;106:419–27.
77. Johansson U, Olsson A, Gabrielsson S, Nilsson B, Korsgren O. Inflammatory mediatorsexpressed in human islets of Langerhans: implications for islet transplantation. BiochemBiophys Res Commun 2003;308:474–9.
78. Badet L, Titus T, Metzen E, Handa A, McShane P, Chang LW, Giangrande P, Gray DW. Theinteraction between primate blood and mouse islets induces accelerated clotting with isletdestruction. Xenotransplantation 2002;9:91–6.

32 Modulation of Early Inflammatory Reactions 743
79. Melzi R, Sanvito F, Mercalli A, Andralojc K, Bonifacio E, Piemonti L. Intrahepatic islettransplant in the mouse: functional and morphological characterization. Cell Transplant2008;17:1361–70.
80. Cavalieri B, Mosca M, Ramadori P, Perrelli MG, De Simone L, Colotta F, Bertini R, PoliG, Cutrin JC. Neutrophil recruitment in the reperfused-injured rat liver was effectivelyattenuated by repertaxin a novel allosteric noncompetitive inhibitor of CXCL8 recep-tors: a therapeutic approach for the treatment of post-ischemic hepatic syndromes. Int JImmunopathol Pharmacol 2005;18:475–86.
81. Johansson H, Goto M, Dufrane D, Siegbahn A, Elgue G, Gianello P, Korsgren O, Nilsson B.Low molecular weight dextran sulfate: a strong candidate drug to block IBMIR in clinicalislet transplantation. Am J Transplant 2006;6:305–12.
82. Spirig R, Gajanayake T, Korsgren O, Nilsson B, Rieben R. Low molecular weight dextransulfate as complement inhibitor and cytoprotectant in solid organ and islet transplantation.Mol Immunol 2008;45:4084–94.
83. Goto M, Johansson H, Maeda A, Elgue G, Korsgren O, Nilsson B. Low-molecular weightdextran sulfate abrogates the instant blood-mediated inflammatory reaction induced by adultporcine islets both in vitro and in vivo. Transplant Proc 2004;36:1186–7.
84. Beuneu C, Vosters O, Ling Z, Pipeleers D, Pradier O, Goldman M, Verhasselt V. N-Acetylcysteine derivative inhibits procoagulant activity of human islet cells. Diabetologia2007;50:343–7.
85. Vosters O, Beuneu C, Goldman M, Verhasselt V. N-acetylcysteine derivative inhibitsCD40-dependent proinflammatory properties of human pancreatic duct cells. Pancreas2008;36:363–8.
86. Stabler CL, Sun XL, Cui W, Wilson JT, Haller CA, Chaikof EL. Surface re-engineering of pancreatic islets with recombinant azido-thrombomodulin. BioconjugateChem 2007;18:1713–5.
87. Bennet W, Sundberg B, Lundgren T, Tibell A, Groth CG, Richards A, White DJ, Elgue G,Larsson R, Nilsson B, Korsgren O. Damage to porcine islets of Langerhans after exposure tohuman blood in vitro, or after intraportal transplantation to cynomolgus monkeys: protectiveeffects of sCR1 and heparin. Transplantation 2000;69:711–9.
88. Lundgren T, Bennet W, Tibell A, Soderlund J, Sundberg B, Song Z, Elgue G, Harrison R,Richards A, White DJ, et al. Soluble complement receptor 1 (TP10) preserves adult porcineislet morphology after intraportal transplantation into cynomolgus monkeys. Transplant Proc2001:33:725.
89. Moberg L, Olsson A, Berne C, Felldin M, Foss A, Kallen R, Salmela K, Tibell A,Tufveson G, Nilsson B, Korsgren O. Nicotinamide inhibits tissue factor expression in iso-lated human pancreatic islets: implications for clinical islet transplantation. Transplantation2003;76:1285–8.
90. Cabric S, Sanchez J, Lundgren T, Foss A, Felldin M, Kallen R, Salmela K, Tibell A,Tufveson G, Larsson R, et al. Islet surface heparinization prevents the instant blood-mediatedinflammatory reaction in islet transplantation. Diabetes 2007;56:2008–15.
91. Johansson U, Elgue G, Nilsson B, Korsgren O. Composite islet-endothelial cell grafts: anovel approach to counteract innate immunity in islet transplantation. Am J Transplant2005b;5:2632–.
92. Kim HI, Yu, JE, Lee SY, Sul AY, Jang MS, Rashid MA, Park SG, Kim SJ, Park CG, KimJH, Park KS. The effect of composite pig islet-human endothelial cell grafts on the instantblood-mediated inflammatory reaction. Cell Transplant 2009;18:31–7.
93. Johansson U, Rasmusson I, Niclou SP, Forslund N, Gustavsson L, Nilsson B, Korsgren O,Magnusson PU. Formation of composite endothelial cell-mesenchymal stem cell islets: anovel approach to promote islet revascularization. Diabetes 2008;57:2393–401.
94. Lewis EC, Mizrahi M, Toledano M, Defelice N, Wright JL, Churg A, Shapiro L, DinarelloCA. alpha1-Antitrypsin monotherapy induces immune tolerance during islet allografttransplantation in mice. Proc Natl Acad Sci USA 2008;105:16236–41.

744 L. Piemonti et al.
95. Weir GC, Koulamnda M. Control of inflammation with alpha1-antitrypsin: a potentialtreatment for islet transplantation and new-onset type 1 diabetes. Curr Diabetes Rep2009;9:100–2.
96. Zhang B, Lu, Y, Campbell-Thompson M, Spencer T, Wasserfall C, Atkinson M, Song S.Alpha1-antitrypsin protects beta-cells from apoptosis. Diabetes 2007;56:1316–23.
97. Lund T, Fosby B, Korsgren O, Scholz H, Foss A. Glucocorticoids reduce pro-inflammatorycytokines and tissue factor in vitro and improve function of transplanted human islets invivo. Transplant Int 2008;21:669–78.
98. Marzorati S, Antonioli B, Nano R, Maffi P, Piemonti L, Giliola C, Secchi A, Lakey JR,Bertuzzi F. Culture medium modulates proinflammatory conditions of human pancreaticislets before transplantation. Am J Transplant 2006;6:2791–5.
99. Brissova M, Shostak A, Shiota M, Wiebe PO, Poffenberger G, Kantz J, Chen Z, Carr C,Jerome WG, Chen J, et al. Pancreatic islet production of vascular endothelial growth fac-tor – a is essential for islet vascularization, revascularization, function. Diabetes 2006;55:2974–85.
100. Zhang N, Richter A, Suriawinata J, Harbaran S, Altomonte J, Cong L, Zhang H, Song K,Meseck M, Bromberg J, Dong H. Elevated vascular endothelial growth factor production inislets improves islet graft vascularization. Diabetes 2004;53:963–70.
101. Stagner JI, Samols E. Induction of angiogenesis by growth factors: relevance to pancreaticislet transplantation. EXS 1992;61:381–5.
102. Cheng K, Fraga D, Zhang C, Kotb M, Gaber AO, Guntaka RV, Mahato RI. Adenovirus-based vascular endothelial growth factor gene delivery to human pancreatic islets. GeneTher 2004;11:1105–16.
103. Mahato RI, Henry J, Narang AS, Sabek O, Fraga D, Kotb M, Gaber AO. Cationic lipid andpolymer-based gene delivery to human pancreatic islets. Mol Ther 2003;7:89–100.
104. Giannoukakis N, Rudert WA, Ghivizzani SC, Gambotto A, Ricordi C, Trucco M, RobbinsPD. Adenoviral gene transfer of the interleukin-1 receptor antagonist protein to human isletsprevents IL-1beta-induced beta-cell impairment and activation of islet cell apoptosis in vitro.Diabetes 1999;48:1730–6.
105. Gysemans C, Stoffels K, Giulietti A, Overbergh L, Waer M, Lannoo M, Feige U, Mathieu C.Prevention of primary non-function of islet xenografts in autoimmune diabetic NOD miceby anti-inflammatory agents. Diabetologia 2003;46:1115–23.
106. Dobson T, Fraga D, Saba C, Bryer-Ash M, Gaber AO, Gerling IC. Human pancreatic isletstransfected to produce an inhibitor of TNF are protected against destruction by humanleukocytes. Cell Transplant 2000;9:857–65.
107. Grey ST, Arvelo MB, Hasenkamp W, Bach FH, Ferran C. A20 inhibits cytokine-inducedapoptosis and nuclear factor kappaB-dependent gene activation in islets. J Exp Medi1999;190:1135–46.
108. Rabinovitch A, Suarez-Pinzon W, Strynadka K, Ju, Q, Edelstein D, Brownlee M, KorbuttGS, Rajotte RV. Transfection of human pancreatic islets with an anti-apoptotic gene (bcl-2)protects beta-cells from cytokine-induced destruction. Diabetes 1999;48:1223–9.
109. Pileggi A, Molano RD, Berney T, Cattan P, Vizzardelli C, Oliver R, Fraker C, Ricordi C,Pastori RL, Bach FH, Inverardi L. Heme oxygenase-1 induction in islet cells results inprotection from apoptosis and improved in vivo function after transplantation. Diabetes2001;50:1983–91.
110. Tobiasch E, Gunther L, Bach FH. Heme oxygenase-1 protects pancreatic beta cells fromapoptosis caused by various stimuli. J Investig Med 2001;49:566–71.
111. George M, Ayuso E, Casellas A, Costa C, Devedjian JC, Bosch F. Beta cell expression ofIGF-I leads to recovery from type 1 diabetes. J Clin Invest 2002a;109:1153–63.
112. Giannoukakis N, Mi, Z, Rudert WA, Gambotto A, Trucco M, Robbins P. Prevention of betacell dysfunction and apoptosis activation in human islets by adenoviral gene transfer of theinsulin-like growth factor I. Gene Ther 2000a;7: 2015–22.

32 Modulation of Early Inflammatory Reactions 745
113. Carpenter L, Cordery D, Biden TJ. Protein kinase Cdelta activation by interleukin-1betastabilizes inducible nitric-oxide synthase mRNA in pancreatic beta-cells. J Biol Chem2001;276:5368–74.
114. Dupraz P, Cottet S, Hamburger F, Dolci W, Felley-Bosco E, Thorens B. Dominant neg-ative MyD88 proteins inhibit interleukin-1beta/interferon-gamma -mediated induction ofnuclear factor kappa B-dependent nitrite production and apoptosis in beta cells. J Biol Chem2000;275:37672–8.
115. Wei JF, Zheng SS. NF-kappa B in allograft rejection. Hepatobiliary Pancreat Dis Int2003;2:180–3.
116. Giannoukakis N, Rudert WA, Trucco M, Robbins PD. Protection of human islets from theeffects of interleukin-1beta by adenoviral gene transfer of an Ikappa B repressor. J BiolChem 2000b;275:36509–13.
117. Burkart V, Liu H, Bellmann K, Wissing D, Jaattela M, Cavallo MG, Pozzilli P, Briviba K,Kolb H. Natural resistance of human beta cells toward nitric oxide is mediated by heat shockprotein 70. J Biol Chem 2000;275:19521–28.
118. Hohmeier HE, Thigpen A, Tran VV, Davis R, Newgard CB. Stable expression of manganesesuperoxide dismutase (MnSOD) in insulinoma cells prevents IL-1beta- induced cytotoxicityand reduces nitric oxide production. J Clin Invest 1998;101:1811–20.
119. Benhamou PY, Moriscot C, Richard MJ, Beatrix O, Badet L, Pattou F, Kerr-Conte J,Chroboczek J, Lemarchand P, Halimi S. Adenovirus-mediated catalase gene transfer reducesoxidant stress in human, porcine and rat pancreatic islets. Diabetologia 1998;41:1093–100.
120. Lepore DA, Shinkel TA, Fisicaro N, Mysore TB, Johnson LE, d’Apice AJ, Cowan PJ.Enhanced expression of glutathione peroxidase protects islet beta cells from hypoxia-reoxygenation. Xenotransplantation 2004;11:53–9.
121. Moriscot C, Pattou F, Kerr-Conte J, Richard MJ, Lemarchand P, Benhamou PY. Contributionof adenoviral-mediated superoxide dismutase gene transfer to the reduction in nitricoxide-induced cytotoxicity on human islets and INS-1 insulin-secreting cells. Diabetologia2000;43:625–31.
122. Mysore TB, Shinkel TA, Collins J, Salvaris EJ, Fisicaro N, Murray-Segal LJ, Johnson LE,Lepore DA, Walters SN, Stokes R, et al. Overexpression of glutathione peroxidase with twoisoforms of superoxide dismutase protects mouse islets from oxidative injury and improvesislet graft function. Diabetes 2005;54:2109–16.
123. Emamaullee JA, Rajotte RV, Liston P, Korneluk RG, Lakey JR, Shapiro AM, Elliott JF.XIAP overexpression in human islets prevents early posttransplant apoptosis and reducesthe islet mass needed to treat diabetes. Diabetes 2005;54:2541–8.
124. Plesner A, Liston P, Tan R, Korneluk RG, Verchere CB. The X-linked inhibitor of apoptosisprotein enhances survival of murine islet allografts. Diabetes 2005;54:2533–40.
125. Merani S, Toso C, Emamaullee J, Shapiro AM. Optimal implantation site for pancreatic islettransplantation. Br J Surg 2008;95:1449–61.
126. Jindal RM, Sidner RA, McDaniel HB, Johnson MS, Fineberg SE. Intraportal vs kidneysubcapsular site for human pancreatic islet transplantation. Transplant Proc 1998;30:398–9.
127. Berman DM, O’Neil JJ, Coffey LC, Chaffanjon PC, Kenyon NM, Ruiz P, Jr, Pileggi A,Ricordi C, Kenyon NS. Long-term survival of nonhuman primate islets implanted in anomental pouch on a biodegradable scaffold. Am J Transplant 2009;9:91–104.
128. Calafiore R, Basta G, Luca G, Lemmi A, Montanucci MP, Calabrese G, Racanicchi L,Mancuso F, Brunetti P. Microencapsulated pancreatic islet allografts into nonimmunosup-pressed patients with type 1 diabetes: first two cases. Diabetes Care 2006;29:137–8.
129. Fort A, Fort N, Ricordi C, Stabler CL. Biohybrid devices and encapsulation technologies forengineering a bioartificial pancreas. Cell Transplant 2008;17:997–1003.
130. Kizilel S, Garfinkel M, Opara E. The bioartificial pancreas: progress and challenges.Diabetes Technol Therapeutics 2005;7:968–85.
131. Kobayashi N. Bioartificial pancreas for the treatment of diabetes. Cell Transplant2008;17:11–7.

746 L. Piemonti et al.
132. Maki T, Lodge JP, Carretta M, Ohzato H, Borland KM, Sullivan SJ, Staruk J, Muller TE,Solomon BA, Chick WL, et al. Treatment of severe diabetes mellitus for more than one yearusing a vascularized hybrid artificial pancreas. Transplantation 1993;55:713–7; discussion717–8.
133. Sun AM, Parisius W, Healy GM, Vacek I, Macmorine HG. The use, in diabetic rats and mon-keys, of artificial capillary units containing cultured islets of Langerhans (artificial endocrinepancreas). Diabetes 1977;26:1136–9.
134. de Vos P, van Hoogmoed CG, van Zanten J, Netter S, Strubbe JH, Busscher HJ. Long-termbiocompatibility, chemistry, function of microencapsulated pancreatic islets. Biomaterials2003;24:305–12.
135. Zekorn T, Horcher A, Siebers U, Federlin K, Bretzel RG. Islet transplantation inimmunoseparating membranes for treatment of insulin-dependent diabetes mellitus. ExpClin Endocrinol Diabetes 103 Suppl 1995;2:136–9.
136. Simpson NE, Khokhlova N, Oca-Cossio JA, McFarlane SS, Simpson CP, ConstantinidisI. Effects of growth regulation on conditionally-transformed alginate-entrapped insulinsecreting cell lines in vitro. Biomaterials 2005;26:4633–41.
137. Balamurugan AN, Gu, Y, Tabata Y, Miyamoto M, Cui W, Hori H, Satake A, Nagata N,Wang W, Inoue K. Bioartificial pancreas transplantation at prevascularized intermuscularspace: effect of angiogenesis induction on islet survival. Pancreas 2003;26:279–85.
138. Wang T, Lacik I, Brissova M, Anilkumar AV, Prokop A, Hunkeler D, Green R, Shahrokhi K,Powers AC. An encapsulation system for the immunoisolation of pancreatic islets. NatureBiotechnol 1997;15:358–62.
139. Xu B, Iwata H, Miyamoto M, Balamurugan AN, Murakami Y, Cui W, Imamura M, InoueK. Functional comparison of the single-layer agarose microbeads and the developed three-layer agarose microbeads as the bioartificial pancreas: an in vitro study. Cell Transplant2001;10:403–8.
140. George S, Nair PD, Risbud MV, Bhonde RR. Nonporous polyurethane membranes as isletimmunoisolation matrices – biocompatibility studies. J Biomater Appl 2002b;16:327–40.
141. Risbud M, Hardikar A, Bhonde R. Chitosan-polyvinyl pyrrolidone hydrogels as candi-date for islet immunoisolation: in vitro biocompatibility evaluation. Cell Transplant 2000;9:25–31.
142. Risbud MV, Bhonde RR. Suitability of cellulose molecular dialysis membrane for bioartifi-cial pancreas: in vitro biocompatibility studies. J Biomed Mater Res 2001;54:436–44.
143. Isayeva IS, Kasibhatla BT, Rosenthal KS, Kennedy JP. Characterization and performanceof membranes designed for macroencapsulation/implantation of pancreatic islet cells.Biomaterials 2003;24:3483–91.
144. De Vos P, De Haan B, Van Schilfgaarde R. Effect of the alginate composition on thebiocompatibility of alginate-polylysine microcapsules. Biomaterials 1997;18:273–8.
145. Schneider S, Feilen PJ, Brunnenmeier F, Minnemann T, Zimmermann H, ZimmermannU, Weber MM. Long-term graft function of adult rat and human islets encapsulated innovel alginate-based microcapsules after transplantation in immunocompetent diabetic mice.Diabetes 2005;54:687–93.
146. Sun Y, Ma, X, Zhou D, Vacek I, Sun AM. Normalization of diabetes in spontaneouslydiabetic cynomologus monkeys by xenografts of microencapsulated porcine islets withoutimmunosuppression. J Clin Invest 1996;98:1417–22.
147. de Groot M, Schuurs TA, van Schilfgaarde R. Causes of limited survival of microencapsu-lated pancreatic islet grafts. J Surg Res 2004;121:141–50.
148. Safley SA, Kapp LM, Tucker-Burden C, Hering B, Kapp JA, Weber CJ. Inhibition of cellularimmune responses to encapsulated porcine islet xenografts by simultaneous blockade of twodifferent costimulatory pathways. Transplantation 2005;79:409–18.
149. Soon-Shiong P, Feldman E, Nelson R, Heintz R, Yao Q, Yao Z, Zheng T, MeridethN, Skjak-Braek G, Espevik T, et al. Long-term reversal of diabetes by the injection ofimmunoprotected islets. Proc Natl Acad Sci USA 1993;90:5843–7.

32 Modulation of Early Inflammatory Reactions 747
150. Blomeier H, Zhang X, Rives C, Brissova M, Hughes E, Baker M, Powers AC, Kaufman DB,Shea LD, Lowe WL, Jr. Polymer scaffolds as synthetic microenvironments for extrahepaticislet transplantation. Transplantation 2006;82:452–9.
151. Dufour JM, Rajotte RV, Zimmerman M, Rezania A, Kin T, Dixon DE, Korbutt GS.Development of an ectopic site for islet transplantation, using biodegradable scaffolds.Tissue Eng 2005;11:1323–31.
152. Kin T, O’Neil JJ, Pawlick R, Korbutt GS, Shapiro AM, Lakey JR. The use of anapproved biodegradable polymer scaffold as a solid support system for improvement of isletengraftment. Artificial Organs 2008;32:990–3.
153. Salvay DM, Rives CB, Zhang X, Chen F, Kaufman DB, Lowe WL, Jr, Shea LD. Extracellularmatrix protein-coated scaffolds promote the reversal of diabetes after extrahepatic islettransplantation. Transplantation 2008;85:1456–64.
154. Langer R, Vacanti JP. Tissue engineering. Science New York, NY 1993;260:920–6.155. Putnam AJ, Mooney DJ. Tissue engineering using synthetic extracellular matrices. Nat Med
1996;2:824–6.156. Lee DY, Yang K, Lee S, Chae SY, Kim KW, Lee MK, Han DJ, Byun Y. Optimization of
monomethoxy-polyethylene glycol grafting on the pancreatic islet capsules. J Biomed MaterRes 2002;62:372–77.
157. Panza JL, Wagner WR, Rilo HL, Rao RH, Beckman EJ, Russell AJ. Treatment of ratpancreatic islets with reactive PEG. Biomaterials 2000;21:1155–64.
158. Xie D, Smyth CA, Eckstein C, Bilbao G, Mays J, Eckhoff DE, Contreras JL.Cytoprotection of PEG-modified adult porcine pancreatic islets for improved xenotransplan-tation. Biomaterials 2005;26:403–12.
159. Krol S, del Guerra S, Grupillo M, Diaspro A, Gliozzi A, Marchetti P. Multilayer nanoen-capsulation. New approach for immune protection of human pancreatic islets. Nano Lett2006;6:1933–9.
160. Weber LM, Cheung CY, Anseth KS. Multifunctional pancreatic islet encapsulation barriersachieved via multilayer PEG hydrogels. Cell Transplant 2008;16:1049–57.
161. Wilson JT, Cui W, Chaikof EL. Layer-by-layer assembly of a conformal nanothin PEGcoating for intraportal islet transplantation. Nano Lett 2008;8:1940–8.
162. Teramura Y, Iwata H. Islets surface modification prevents blood-mediated inflammatoryresponses. Bioconjugate Chem 2008;19:1389–95.
Related Documents











