Modulated Expression of Notchl During Thymocyte Development By R.P. Hasserjian, J.C. Aster, F. Davi, D.S. Weinberg, and J. Sklar The Notch gene family encodes transmembrane proteins that have been implicated in control of diverse cellular differ- entiation events in the fly, frog, and mouse. Mammalian Notchl is expressed at high levels in thymus and is mutated in a subset of human T-cell acute lymphoblastic neoplasms, suggesting a role in T-cell differentiation. To investigate the patterns of expression of NOTCHI protein in thymocytes of the developing and mature thymus, antibodies raised against NOTCHI were used to perform immunohistochemi- cal and flow cytometric analyses. Strong staining for NOTCHl within the fetal murine thymus was observed as early as 13.5 days postcoitum. By 17.5 days postcoitum, pref- erential staining of superficial cortical thymocytes was ob- HE DROSOPHILA gene Notch encodes a transmem- T brane receptor (NOTCH) that functions in a novel sig- nal transduction pathway.' Dysregulation of Notch activity during critical phases of development leads to hyperplasia and/or hypoplasia of diverse cell types, indicating a role in the control of programs of differentiation in a variety of Signaling by NOTCH appears to normally depend on binding of extracellular ligands, including DELTA and SERRATE, two additional transmembrane proteins.5 These and other observations have suggested that NOTCH coordi- nates differentiation among groups of equipotent progenitor cells that lie in close physical proximity. Mammals possess three genes, Notchl -3, that are highly homologous to Drosophila Notch, and at least one more distantly related gene, Overexpression of a constitu- tively active form of NOTCHl inhibits neural and myoid differentiation of murine cell lines, indicating that the ability of NOTCH-like proteins to inhibit certain types of differenti- ation has been conserved." Furthermore, analysis of mice containing homozygous disruptions in Notchl has shown that NOTCH 1 is essential for postimplantation development, as all embryos die by 11.5 days postcoitum (dpc)." These embryos display subtle abnormalities in somitogenesis and also show widespread cell death, particularly in regions of the central nervous system.''.'* Other studies have noted a relationship between increased NOTCHl expression and mi- From the Division of Molecular Oncology, Department of Pathol- ogy, Brigham and Women's Hospital, Harvard Medical School, Bos- ton, MA. Submitted September 20, 1995; accepted April 2, 1996. Supported in part by grants from the National Institutes of Health No. CA38621 and No. CO62450 (J.S.), and No. CA65889 (R.P.H.), a grant from the Massachusetts Division of the American Cancer Society (J.C.A.), and the Tata Fellowship (F.D.). Address reprint requests to J. Sklar, MD, PhD, Department of Pathology, Brigham & Women's Hospital, 75 Francis St, Boston, MA 02115. The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked "advertisement" in accordance with 18 U.S.C. section 1734 solely to indicate this fact. 0 1996 by The American Society of Hematology. 0006-4971/96/8803-00I 5$3.00/0 served, with weak staining of developing medulla. Flow cy- tometric analysis and immunohistochemical staining of flow-sorted cells confirmed that the highest levels of NOTCHI expression in adult murine thymus were present in immature cortical thymocytes (CD24high. CD4-CD8-). In contrast, NOTCHI expression was low or absent in more mature cortical thymocytes (CD241ow, CD4'CD8'), whereas intermediate levels of expression were observed in CD4TD8- and CD4-CD8' cells. These data indicate a dy- namic pattern of NOTCHI expression during T-cell differenti- ation and suggest that downregulation of NOTCHI may be required for maturation of cortical thymocytes. 0 1996 by The American Society of Hematology. totic activity within developing neural tissues of fly, fish, and Thus, NOTCHl might function by maintaining cells in a proliferative state, thereby inhibiting terminal dif- ferentiation. Notchl mRNA is widely expressed during murine and human development, but is found at highest levels in devel- oping thymus.I5 Human NOTCHI (also referred to as TAN- I), located on chromosome 9, is broken in human T-cell lymphoblastic neoplasms containing a recurrent (7; 9) (q34;q34.3) chromosomal transl~cation.'~ This rearrange- ment joins portions of NOTCHl and TCRP, and leads to overexpression of truncated polypeptides containing most or all of the cytoplasmic domain of NOTCH1.15 These data suggest that NOTCHl may function normally to control T- cell differentiation, possibly through an effect on the cell cycle. To gain insight into these possibilities, immunohisto- chemical and flow cytometric analyses were performed to correlate NOTCHl expression with normal thymocytic dif- ferentiation and proliferation. MATERIALS AND METHODS Animals. Embryos were obtained from timed-pregnant ICR mice (Taconic Farms Inc, Germantown, NY). Thymuses and peripheral blood (PB) were obtained from 4- to 7-week-old Balbk mice (Ta- conic Farms Inc). Isolation of thymocytes and PB mononuclear cells (PBMC). Freshly sacrificed mice were exsanguinated, and PBMC were sepa- rated in a Ficoll-Hypaque gradient (Pharmacia, Piscataway, NJ). Thymocytes or splenocytes were disaggregated by pressing the thy- muses or spleens through a fine steel mesh. PBMC, thymocytes, and splenocytes were washed and incubated in RPMI containing 20% heat-inactivated fetal calf serum (FCS) at 37" for 30-60 minutes before analysis. Antibodies. Immunophenotyping was performed with the fol- lowing antibodies: CD4-fluorescein (H129.19; GIBCO-BRL, Gaith- ersburg, MD), CDCTRI-COLOR (Caltag, South San Francisco, CA), CD8a-R-phycoerythrin (53-6.7; GIBCO-BRL), CD24-fluores- cein (CT-HSA; Caltag), CD25-R-phycoerythrin (3C7; GIBCO- BRL), and anti-PCNA (monoclonal mouse IgG; Oncogene Science, Cambridge, MA). Immunologic staining for NOTCHI was per- formed with xffinity-purified polyclonal rabbit antibodies against a cytoplasmic domain of human NOTCH1, termed T3 (human NOTCHl amino acids 1733-1877). synthesized as a glutathione-S- transferase (GST) fusion protein.16 Anti-GST antibodies purified from the same animals were used as a negative control. A second portion of human NOTCHI, termed TC, (amino acids 2278-2470) 970 Blood, VOl 88, NO 3 (August I), 1996: pp 970-976 For personal use only. on April 10, 2019. by guest www.bloodjournal.org From

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Modulated Expression of Notchl During Thymocyte Development
By R.P. Hasserjian, J.C. Aster, F. Davi, D.S. Weinberg, and J. Sklar
The Notch gene family encodes transmembrane proteins that have been implicated in control of diverse cellular differ- entiation events in the fly, frog, and mouse. Mammalian Notchl is expressed at high levels in thymus and is mutated in a subset of human T-cell acute lymphoblastic neoplasms, suggesting a role in T-cell differentiation. To investigate the patterns of expression of NOTCHI protein in thymocytes of the developing and mature thymus, antibodies raised against NOTCHI were used to perform immunohistochemi- cal and flow cytometric analyses. Strong staining for NOTCHl within the fetal murine thymus was observed as early as 13.5 days postcoitum. By 17.5 days postcoitum, pref- erential staining of superficial cortical thymocytes was ob-
HE DROSOPHILA gene Notch encodes a transmem- T brane receptor (NOTCH) that functions in a novel sig- nal transduction pathway.' Dysregulation of Notch activity during critical phases of development leads to hyperplasia and/or hypoplasia of diverse cell types, indicating a role in the control of programs of differentiation in a variety of
Signaling by NOTCH appears to normally depend on binding of extracellular ligands, including DELTA and SERRATE, two additional transmembrane proteins.5 These and other observations have suggested that NOTCH coordi- nates differentiation among groups of equipotent progenitor cells that lie in close physical proximity.
Mammals possess three genes, Notchl -3, that are highly homologous to Drosophila Notch, and at least one more distantly related gene, Overexpression of a constitu- tively active form of NOTCHl inhibits neural and myoid differentiation of murine cell lines, indicating that the ability of NOTCH-like proteins to inhibit certain types of differenti- ation has been conserved." Furthermore, analysis of mice containing homozygous disruptions in Notchl has shown that NOTCH 1 is essential for postimplantation development, as all embryos die by 11.5 days postcoitum (dpc)." These embryos display subtle abnormalities in somitogenesis and also show widespread cell death, particularly in regions of the central nervous system.''.'* Other studies have noted a relationship between increased NOTCHl expression and mi-
From the Division of Molecular Oncology, Department of Pathol- ogy, Brigham and Women's Hospital, Harvard Medical School, Bos- ton, MA.
Submitted September 20, 1995; accepted April 2, 1996. Supported in part by grants from the National Institutes of Health
No. CA38621 and No. CO62450 (J.S.) , and No. CA65889 (R.P.H.), a grant from the Massachusetts Division of the American Cancer Society (J.C.A.), and the Tata Fellowship (F.D.).
Address reprint requests to J . Sklar, MD, PhD, Department of Pathology, Brigham & Women's Hospital, 75 Francis St, Boston, MA 02115.
The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked "advertisement" in accordance with 18 U.S.C. section 1734 solely to indicate this fact. 0 1996 by The American Society of Hematology. 0006-4971/96/8803-00I 5$3.00/0
served, with weak staining of developing medulla. Flow cy- tometric analysis and immunohistochemical staining of flow-sorted cells confirmed that the highest levels of NOTCHI expression in adult murine thymus were present in immature cortical thymocytes (CD24high. CD4-CD8-). In contrast, NOTCHI expression was low or absent in more mature cortical thymocytes (CD241ow, CD4'CD8'), whereas intermediate levels of expression were observed in CD4TD8- and CD4-CD8' cells. These data indicate a dy- namic pattern of NOTCHI expression during T-cell differenti- ation and suggest that downregulation of NOTCHI may be required for maturation of cortical thymocytes. 0 1996 by The American Society of Hematology.
totic activity within developing neural tissues of fly, fish, and Thus, NOTCHl might function by maintaining cells in a proliferative state, thereby inhibiting terminal dif- ferentiation.
Notchl mRNA is widely expressed during murine and human development, but is found at highest levels in devel- oping thymus.I5 Human NOTCHI (also referred to as TAN- I ) , located on chromosome 9, is broken in human T-cell lymphoblastic neoplasms containing a recurrent (7; 9) (q34;q34.3) chromosomal transl~cation.'~ This rearrange- ment joins portions of NOTCHl and TCRP, and leads to overexpression of truncated polypeptides containing most or all of the cytoplasmic domain of NOTCH1.15 These data suggest that NOTCHl may function normally to control T- cell differentiation, possibly through an effect on the cell cycle. To gain insight into these possibilities, immunohisto- chemical and flow cytometric analyses were performed to correlate NOTCHl expression with normal thymocytic dif- ferentiation and proliferation.
MATERIALS AND METHODS
Animals. Embryos were obtained from timed-pregnant ICR mice (Taconic Farms Inc, Germantown, NY). Thymuses and peripheral blood (PB) were obtained from 4- to 7-week-old Balbk mice (Ta- conic Farms Inc).
Isolation of thymocytes and PB mononuclear cells (PBMC). Freshly sacrificed mice were exsanguinated, and PBMC were sepa- rated in a Ficoll-Hypaque gradient (Pharmacia, Piscataway, NJ). Thymocytes or splenocytes were disaggregated by pressing the thy- muses or spleens through a fine steel mesh. PBMC, thymocytes, and splenocytes were washed and incubated in RPMI containing 20% heat-inactivated fetal calf serum (FCS) at 37" for 30-60 minutes before analysis.
Antibodies. Immunophenotyping was performed with the fol- lowing antibodies: CD4-fluorescein (H129.19; GIBCO-BRL, Gaith- ersburg, MD), CDCTRI-COLOR (Caltag, South San Francisco, CA), CD8a-R-phycoerythrin (53-6.7; GIBCO-BRL), CD24-fluores- cein (CT-HSA; Caltag), CD25-R-phycoerythrin (3C7; GIBCO- BRL), and anti-PCNA (monoclonal mouse IgG; Oncogene Science, Cambridge, MA). Immunologic staining for NOTCHI was per- formed with xffinity-purified polyclonal rabbit antibodies against a cytoplasmic domain of human NOTCH1, termed T3 (human NOTCHl amino acids 1733-1877). synthesized as a glutathione-S- transferase (GST) fusion protein.16 Anti-GST antibodies purified from the same animals were used as a negative control. A second portion of human NOTCHI, termed TC, (amino acids 2278-2470)
970 Blood, VOl 88, NO 3 (August I), 1996: pp 970-976
For personal use only.on April 10, 2019. by guest www.bloodjournal.orgFrom

NOTCH7 IN THYMOCYTE DEVELOPMENT 97 1
was expressed from the vector pQE8 (Qiagen, Chatsworth, CA) as a 6xHis fusion protein in Escherichia coli strain mlS(pREP4) cells, purified on Ni2+-NTA resin as described in the package insert (Qia- gen), and used to immunize rabbits. The TC portion of NOTCHl shows no homology to NOTCH2 or NOTCH3.7.8 Anti-TC antibodies were affinity purified as previously described.16 Secondary antibodies used for immunoperoxidase studies were anti-rabbit IgG-peroxidase conjugate (Sigma, St Louis, MO), antimouse IgG-peroxidase conju- gate (Sigma), and peroxidase-antiperoxidase soluble complex (Sigma). Affinity-purified anti-T3 and anti-GST control antibodies were directly conjugated to fluorescein-EX succinimidyl ester (Mo- lecular Probes, Eugene, OR) according to the package insert. A11 anti-NOTCH1 and control antibodies for immunohistochemistry were used at a concentration of 2 pghnL.
Whole or sagitally bisected mouse em- bryos were fixed for 4 hours in 10% neutral buffered formalin, processed routinely, and embedded in paraffin. Similarly fixed, par- affin-embedded human thymuses from a 22-week gestation human fetus and human adult were obtained from the surgical pathology files of Brigham & Women’s Hospital. Sections (3 pm) were deparaf- finized and boiled in 5% urea for IO minutes to enhance the immuno- reactivity of antigens (the “antigen retrieval” proced~re).’~ Immuno- peroxidase staining was performed as previously described, using anti-T3, anti-GST, anti-TC, or anti-PCNA antibodies.’*
Immunoperoxidase staining for NOTCHl in jow-sorted cells. lo6 thymocytes were incubated with primary antibody in 100 pL phosphate-buffered saline with 2% FCS (PBSF) for 30 minutes on ice, washed twice with PBSF, and sorted on a FACS Vantage flow cytometer (Beckton Dickinson, San Jose, CA). Using CD4-fluores- cein and CD8-R-phycoerythrin antibodies, four fractions were col- lected in RPMI with 10% FCS by gating on the following popula- tions: CD4-CD8-, CD4+CD8’, CD4’CD8-, and CD4-CD8’. The purity of the populations was assessed by flow cytometnc analysis of each population following sorting, and was as follows: CD4-CD8-: 88%, CD4TD8’: 95%, CD4’CD8-. 96%, and CD4-CD8’: 70%. In a separate experiment, four populations were collected using CD24-fluorescein and CD25-R-phycoerythrin anti- bodies: CD24highCD25’, CD24highCD25-, CD241owCD25- and CD24-CD25-. The use of CD24 to define thymocyte subpopulations at different stages of maturation has been previously described.” CD25 (interleukin-2 receptor a chain) expression is an early event during T-cell development.” Cytocentnhge preparations were made following rinsing in phosphate-buffered saline (PBS). Slides were air-dried, fixed in 3% paraformaldehyde in PBS for 15 minutes, permeabilized in methanol at -20” for 1 minute, rinsed five times in PBS, and subjected to antigen retrieval as previously described. Slides were stained with anti-T3 or anti-GST antibody as described for paraffin sections. Individual cells were scored for immunoreac- tivity by two independent observers in a blinded fashion, after view- ing the cells at original magnification X 1,000 (see Table 1 ) . A total of 100 to 309 cells from each population were counted. Flow sorting was performed twice for CD4ICD8 and once for CD24ICD25.
Cells were stained with the antibodies CD4-TRI-COLOR and CD8-R-phycoer- ythrin, as previously described. The cells were then fixed in 1% paraformaldehyde in PBS for 10 minutes, permeabilized in 0.2% Triton-X-100 (J.T. Baker Inc, Phillipsburg, NJ) in PBS (PBST) for 3 minutes, and nnsed once in PBST. Cells were incubated with 0.1 pg anti-T3-fluorescein or anti-GST-fluorescein antibodies in PES with 2% heat-inactivated rabbit serum (PBSR) on ice for 30 minutes and rinsed twice in PBST. After fixation in 0.5% paraformaldehyde in PBS, cells were analyzed on a FACScan flow cytometer (Becton Dickinson), which is equipped with optical filters appropriate for simultaneous 3-COh analysis. Between 10,000 and 30,000 cells
Immunohistochemistry.
Flow cytometric analysis of NOTCHl expression.
were counted in each experiment. Data were stored in list mode and analyzed using FACScan Research software (Becton Dickinson). NOTCH1 expression was measured by fluorescein fluorescence in- tensity in gated thymocyte populations
RESULTS
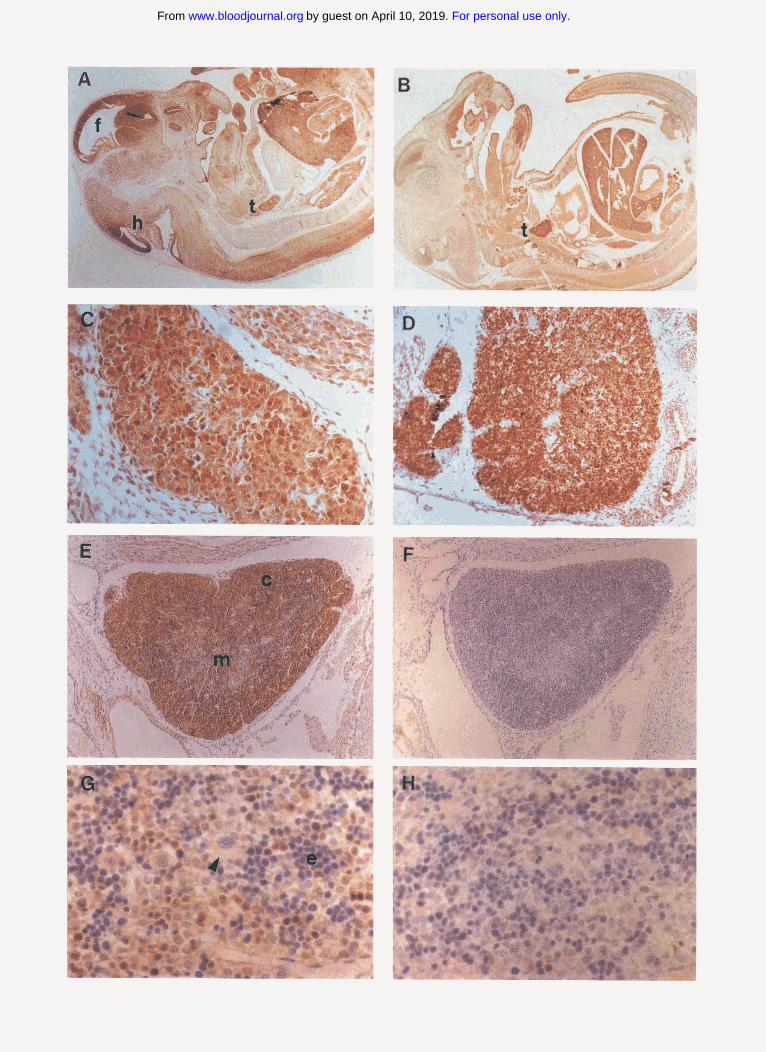
Immunohistochemical analysis of NOTCHI in embryos during murine development. The TC region of NOTCHl was selected as an immunogen because the amino acid se- quence of this region shows no homology to the correspond- ing regions of NOTCH2 and NOTCH3. Antibodies against the T3 region of human NOTCHl (anti-T3) recognize both human and murine NOTCHl in cell extracts.16 The staining patterns in murine embryonic tissue sections for anti-T3 and anti-TC were similar, and correlated well with previous anal- yses of Notchl mRNA by in situ h y b r i d i z a t i ~ n . ~ ~ ’ ~ . ~ ’ There- fore, cross-reactivity of either of these two antibodies with other related proteins, including NOTCH2 and NOTCH3, appears to be In addition to thymus (Fig 1A and lB, and described later), strong staining was also observed in penventricular neural tissue at 10.5 to 14.5 dpc, dorsal root ganglia at 1 1.5 to 18.5 dpc, and developing hair follicles at 16.5 to 18.5 dpc. Weaker staining was also seen in numer- ous other tissues, including testicular germ cells, mesenchy- mal cells, gut epithelium, and nephrons. Scattered mononu- clear cells (MNC) of uncertain lineage within the liver were also stained, whereas maturing erythroid and megakaryo- cytic elements within the liver were negative (Fig 1G). Sub- cellular localization of NOTCHl was difficult to determine in tissue sections. However, in those tissues with the most intense staining, such as spinal ganglia and early thymus, immunoreactivity appeared to be predominantly cyto- plasmic; no definite nuclear staining was observed in any embryonic tissue.
Immunohistochemical analysis of NOTCHl during thymic development. Within the early developing thymus (13.5 to 15.5 dpc, Fig 1C and ID), most cells were immunoreactive for NOTCHI, with a subset of cells showing particularly intense cytoplasmic staining (best seen in Fig IC). With further thymic maturation (18.5 dpc), strongest staining was observed in the thymic cortex, although weak staining per- sisted within the developing medulla (Fig 1E). No significant staining was seen with control anti-GST antibody (Fig 1 F and lH), or with anti-T3 antibody blocked with a 100-fold molar excess of T3 antigen (data not shown).
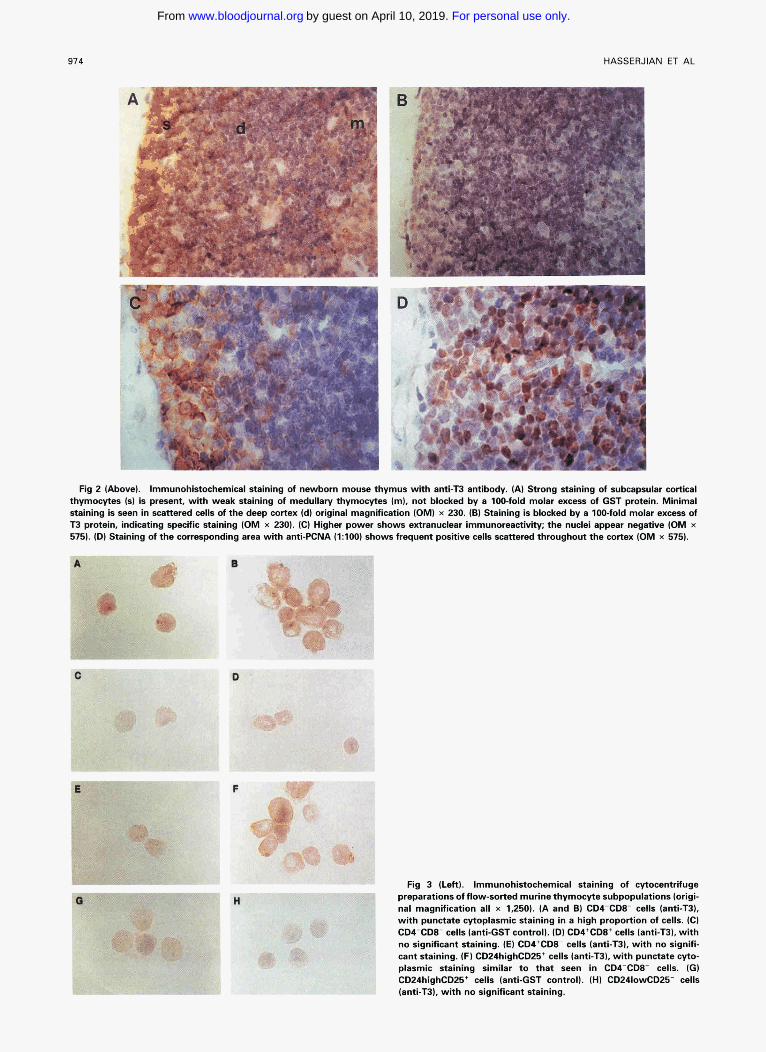
In neonatal, 4-week, and 7-week thymuses, strong staining was confined to a population of relatively large cells in the superficial cortex, with the deep cortex being negative and the medulla showing weak equjvocal staining (Fig 2A). A similar pattern of staining was observed in human fetal and adult thymus (data not shown). N o significant staining was seen with control antibody (anti-GST, not shown) or with antLT3 antibody blocked with a 100-fold molar excess of T3 antigen (Fig 2B). Staining for NOTCHl (Fig 2C) showed only partial correlation with the pattern of PCNA expression (Fig 2D), which was present in a high fraction of cells in the superficial and deep cortex and scattered cells throughout the medulla. Similarly, only partial correlation of NOTCHl
For personal use only.on April 10, 2019. by guest www.bloodjournal.orgFrom

972
and PCNA staining was observed in other embryonic tissues. including brain, skin, and mesenchyme (data not shown).
Analysis of NOTCHI in murine thymocyte subsets. To more precisely define the relationship between NOTCHl expression and thymocytic differentiation, studies were per- formed on subpopulations of thymocytes separated by FACS. In immature cortical thymocyte subpopulations (CD24highCD25', CD24highCD25-, and CD4-CD8-), im- munohistochemistry revealed a fine punctate pattern of extra- nuclear NOTCHl staining in a high proportion of cells (Fig 3A, 3B, and 3F). Minimal or no staining was seen in the other, more mature populations of thymocytes (CD4'CD8', CD4'CD8-, CD4-CD8', CD241owCD25 , and CD24.~ CD25-l (Fig 3D, 3E, and 3H). No staining of any population was seen with anti-GST control antibody (Figs 3C and 3G). The proportion of positively stained cells in each population was scored in a blinded fashion (Table l).
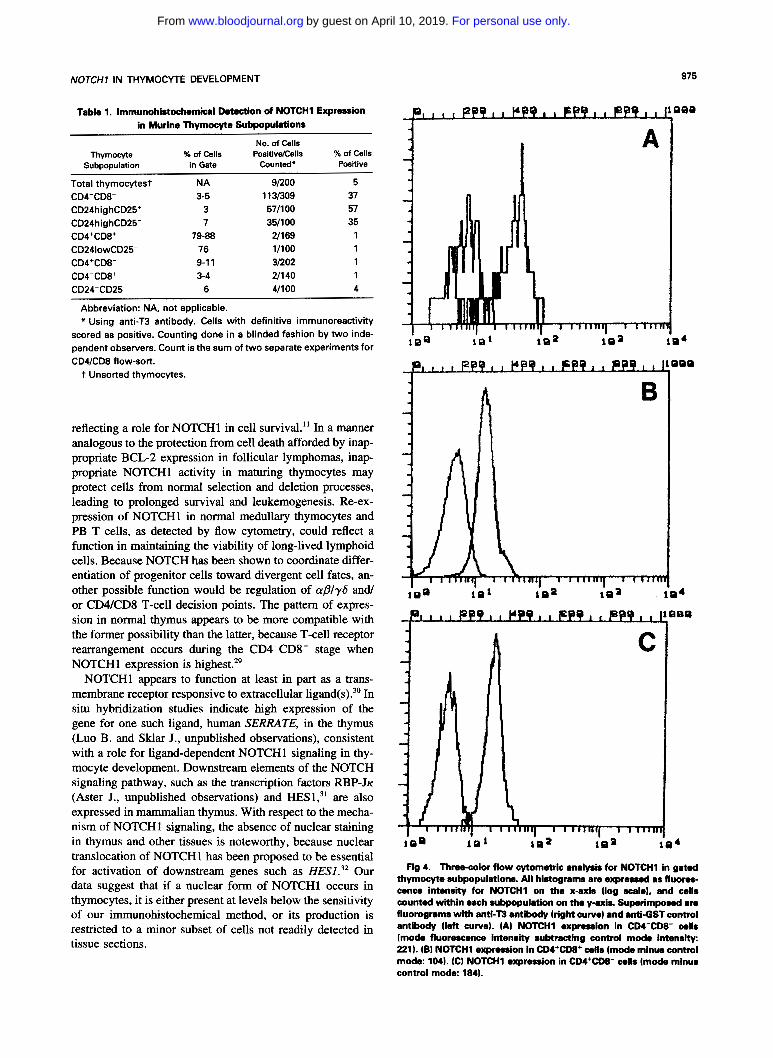
Because weak staining of thymic medulla was observed in tissue sections, the absence of immunohistochemical staining of isolated CD4+CD8- and CD4-CD8' thymocytes could have been due to either NOTCHI expression on other medullary cells (such as macrophages or epithelial cells) or differences in the sensitivity of immunohistochemistry car- ried out on sections versus on cells isolated by flow cytome- try. To address this issue and to further characterize NOTCHl expression in thymocyte subsets, three color FACS analysis was performed. Among the subsets, the strongest NOTCHl fluorescence intensity observed was in the immature cortical thymocyte populations (CD4- CD8-, Fig 4A). In contrast, the fluorescence intensity for NOTCH 1 in mature cortical thymocytes (CD4'CD8', Fig 4B) was much weaker, whereas intermediate fluorescence intensity was detected in CD4'CD8- (Fig 4C) and CD4-CD8' cells (data not shown). FACS analysis of murine PB and splenic CD4'CD8- and CD4-CD8' cells showed a fluorescence intensity for NOTCHl comparable to that observed in C D 4 T D 8 - and CD4-CD8+ thymocytes (data not shown). Thus, using three color FACS analysis, NOTCHl was de- tected in mature thymocytes (CD4'CD8-, CD4-CD8'), al- though at a lower level than in immature cortical thymocytes (CD4-CDK). The reason for the discrepancy between these results and those obtained by immunohistochemistry on flow-sorted CD4'CDK and CD4-CD8' cells is uncertain, but may be explained by lower sensitivity for detecting anti- gen in cytocentrifuge preparations compared with FACS analysis. In summary, our studies show that NOTCHhigh, NOTCHlow, and NOTCHintermediate immunophenotypes coincide with CD4-CD8-, CD4'CD8', and CD4/CD8 sin- gle positive stages of thymocyte maturation.
HASSERJIAN ET AL
DISCUSSION
These data show a dynamic pattern of NOTCHl expres- sion during normal thymocyte differentiation. Although some studies have found the highest levels of Notch mRNA expression in areas of increased mitotic activity within the central nervous systems of developing fish and prolif- eration and NOTCHl expression show only partial positive correlation in normal thymus (see Fig 2). For example, al- though CD4+CD8' thymocytes have a relatively high prolif- erative index as compared with mature C D 4 T D 8 - and CD4- CD8' cells,22 this subpopulation shows the lowest lev- els of NOTCHl expression by FACS analysis. These obser- vations argue against a simple role for NOTCHl in mainte- nance of progenitor cells in cell cycle. This conclusion is compatible with recent studies showing that NOTCH func- tion in the developing retina does not correlate with the cell cycle status of retinal progenitor
In contrast, the marked decline in NOTCHl expression observed during progression of thymocytes from the CD4-CD8 to the CD4'CD8' stage is compatible with a role in the control of early stages of thymocyte differentia- tion. Other events that appear to be required for cortical thymocyte maturation include signal transduction via a trans- membrane pre-TCR complex, LCK, and the IL-2 receptor, and possibly the action of cytokines such as tumor necrosis factor-a and IL-~cu.'~"' That NOTCHl may also be involved in coordinating this process is made plausible by its observed capacity to inhibit or modulate the differentiation of other types of progenitor cells. For instance, NOTCH I can inhibit neural and myoid differentiation when constitutively ac- tive.'" If NOTCHl has a similar function in the thymus, downregulation of NOTCHI may be required for normal differentiation of cortical thymocytes. This hypothesis is supported by recent studies showing that constitutively ac- tive forms of NOTCHl overexpressed in murine bone mar- row progenitor cells are potent inducers of T-cell lympho- blastic neoplasms, which immunophenotypically resemble cortical stages of thymocyte Constitutive expression of activated NOTCHl may block normal differ- entiation of cortical thymocytes, thereby contributing to the development of T-cell neoplasia. Additional studies with constitutively active Notchl constructs in transgenic models may serve to more precisely define the effects of increased NOTCHl activity on cortical thymocyte differentiation.
Alternatively, NOTCHl downregulation at the common thymocyte stage (CD4'CD8') may be necessary for positive and negative selection. Homozygous disruption of Notchl in mice leads to widespread cell death at l1 .S dpc, possibly
b
Fig 1. Immunohistochemical staining of mouse embryos. (A) Whole embryo, 13.5 dpc (anti-T3). NOTCHl staining is widespread in mesenchy- mal tissues, forebrain ( f ) and hindbrain (h) periventricular regions, and thymus (t) (original magnification (OM) x 13). (B) Whole embryo. 15.5 dpc (anti-TB). Strong staining is maintained in the thymus (t), whereas staining in other areas has decreased (OM x 7.5). (C) Thymus, 13.5 dpc (anti-T3). Staining is present diffusely, with scattered intensely staining cells (OM x 230). (D) Thymus, 15.5 dpc lanti-T3) (OM x 120). (E) Thymus, day 18.5 pc (antLT3). with strong staining of cortical thymocytes (c) and weaker staining in the medulla (m) (OM x SO). (F) Thymus, 18.5 dpc (anti-GST control, OM x SO). (G) Liver, 18.5 dpc (anti-TB). NOTCHl staining is present in scattered MNC of uncertain lineage, whereas megakaryocytes (arrowhead) and maturing erythroid elements (e) are negative (OM x 260). (HI Liver, 18.5 dpc (anti-GST control, OM x 260).
For personal use only.on April 10, 2019. by guest www.bloodjournal.orgFrom
I
-a
:.
For personal use only.on April 10, 2019. by guest www.bloodjournal.orgFrom
974
Fig 2 (Above). Immunohistochemical staining of newborn mouse thymus with anti-T3 antibody. (A) Strong staining of subcapsular cortical thymocytes 1s) is present, with weak staining of medullary thymocytes (m), not blocked by a 100-fold molar excess of GST protein. Minimal staining is seen in scattered cells of the deep cortex Id) original magnification (OM) x 230. (6) Staining is blocked by a 100-fold molar exmss of T3 protein, indicating opecific Staining (OM x 230). (C) Higher power shows extranudear immunoreactivity; the nuclei appear nwgative (OM x 575). (D) Staining of the wmesponding area with anti-PCNA (1:lOO) shows frequent positive cells scattered throughout the cortex (OM x 575).
Fig 3 (Left). Immunohistochemical staining of cytocentrifuge preparations of flow-sorted murine thymocyte subpopulations (origi- nal magnification all x 1,250). (A and B) CD4-CD8- cells (antLT3). with punctate cytoplasmic staining in a high proportion of cells. (C) CDCCDE- cells (anti-GST control]. (D) CDI+CDE+ cells (anti-T3), with no significant staining. (E) CD4+CD8- cells lanti-T31, with no signifi- cant staining. (F) CD24highCD25+ cells lanti-T3), with punctate cyto- plasmic staining similar to that seen in CD4-CDE- cells. (G) CD24highCD25+ cells (anti-GST control). (H) CD241owCD25- cells
, . ..... . ,. ,A ...- I lanti-T3), with no significant staining.
For personal use only.on April 10, 2019. by guest www.bloodjournal.orgFrom
NOTCH7 IN THYMOCYTE DEVELOPMENT
Table 1. Immunohistochemical Detection of NOTCH1 Expression in Murine Thymocyte Subpopulations
Thymocyte % of Cells No. of Cells
PositivdCells % of Cells in Gate Counted* Positive Subpopulation
Total thymocytest NA 91200 5 CD4-CD8- 3-5 1 13B09 37
CD24highCD25' 3 571100 57
CD24highCD25- 7 351100 35 CD4+CD8+ 79-88 U169 1 CD241owCD25- 76 11100 1 CD4+CD8- 9-1 1 31202 1 CD4-CD8' 3-4 211 40 1 CD24-CD25- 6 41100 4
Abbreviation: NA, not applicable. * Using anti-T3 antibody. Cells with definitive immunoreactivity
scored as positive. Counting done in a blinded fashion by two inde- pendent observers. Count is the sum of two separate experiments for CD4lCD8 flow-sort.
t Unsorted thymocytes.
reflecting a role for NOTCHl in cell survival." In a manner analogous to the protection from cell death afforded by inap- propriate BCL-2 expression in follicular lymphomas, inap- propriate NOTCHl activity in maturing thymocytes may protect cells from normal selection and deletion processes, leading to prolonged survival and leukemogenesis. Re-ex- pression of NOTCHl in normal medullary thymocytes and PB T cells, as detected by flow cytometry, could reflect a function in maintaining the viability of long-lived lymphoid cells. Because NOTCH has been shown to coordinate differ- entiation of progenitor cells toward divergent cell fates, an- other possible function would be regulation of cu/3ly6 and or CD4lCD8 T-cell decision points. The pattern of expres- sion in normal thymus appears to be more compatible with the former possibility than the latter, because T-cell receptor rearrangement occurs during the CD4-CD8- stage when NOTCHl expression is highest."
NOTCHl appears to function at least in part as a trans- membrane receptor responsive to extracellular ligand(^).^' In situ hybridization studies indicate high expression of the gene for one such ligand, human SERRATE, in the thymus (Luo B. and Sklar J., unpublished observations), consistent with a role for ligand-dependent NOTCHl signaling in thy- mocyte development. Downstream elements of the NOTCH signaling pathway, such as the transcription factors RBP-JK (Aster J., unpublished observations) and HES1;l are also expressed in mammalian thymus. With respect to the mecha- nism of NOTCH1 signaling, the absence of nuclear staining in thymus and other tissues is noteworthy, because nuclear translocation of NOTCHl has been proposed to be essential for activation of downstream genes such as HES1.32 Our data suggest that if a nuclear form of NOTCHl occurs in thymocytes, it is either present at levels below the sensitivity of our immunohistochemical method, or its production is restricted to a minor subset of cells not readily detected in tissue sections.
h
Fig 4. Three-color flow cytometric analysis for NOTCHl in gated thymocyte rubpopulations. All histograms are expressed as fluom- cence intensity for NOTCHl on the x-axis (log scala), and cells counted within each subpopulation on the y-axis. Superimposed are fluorograms with anh"T3 antibody (right curve) and anti-GST control antibody (left curve). (A) NOTCHl expression in CDI-CD8- cells (mode fluorescence intensity subtracting control modo intensity: 221). IBI NOTCHl expression In CD4+CD8+ cells (mode minus control mode: 1041. (C) NOTCH1 expression in CD4+CD8- cells (mode minus control mode: 1841.
For personal use only.on April 10, 2019. by guest www.bloodjournal.orgFrom

976 HASSERJIAN ET AL
ACKNOWLEDGMENT
The authors thank Peter Lopez of the Dana-Farber Cancer Institute Core. Flow Cytometry Facility and Agatha Roma of the Brigham & Women’s HematologylFlow Cytometry Lab for assistance with FACS analysis.
REFERENCES 1. Artavanis-Tsakonas S, Matsuno K, Fortini ME: Notch signal-
ing. Science 268:225, 1995 2. Fortini M E , Rebay I, Caron L, Artavanis-Tsakonas S: An acti-
vated Notch receptor blocks cell-fate commitment in the developing Drosophila eye. Nature 365:555, 1993
3. Corbin V, Michelson AM, Abmayr SM, Nee1 V, Alcamo E, Maniatis T, Young MW: A role for the Drosophila neurogenic genes in mesoderm differentiation. Cell 67:311, 1991
4. Rebay I, Fehon RG, Artavanis-Tsakonas S: Specific truncations of Drosophila Notch define dominant activated and dominant nega- tive forms of the receptor. Cell 74:319, 1993
5. Rebay I, Fleming M, Fehon RG, Cherbas L, Cherbas P, Arta- vanis-Tsakonas S: Specific EGF repeats of Notch mediate interac- tions with Delta and Serrate: Implications for Notch as a multifunc- tional receptor. Cell 67:687, 1991
6. Lardelli M, Lendahl U: Motch A and Motch B-two mouse Notch homologues coexpressed in a wide variety of tissues. Exp Cell Res 204:364, 1993
7. Lardelli M, Dahlstrand J, Lendahl U: The novel Notch homo- logue mouse Notch 3 lacks specific epidermal growth factor-repeats and is expressed in proliferating neuroepithelium. Mech Dev 46: 123, 1993
8. Weinmaster G, Roberts VJ, Lemke C: Notch2: A second mam- malian Notch gene. Development 116:931, 1992
9. Jhappan C, Gallahan D, Stahle C, Chu E, Smith GH, Merlino G, Callahan R: Expression of an activated Notch-related int-3 transgene interferes with cell differentiation and induces neoplastic transformation in mammary and salivary glands. Genes Dev 6:345, 1992
10. Nye JS, Kopan R, Axel R: An activated Notch suppresses neurogenesis and myogenesis, but not gliogenesis in mammalian cells. Development 120:2421, 1994
11. Swiatek PJ, Lindsell CE, del Amo FF, Weinmaster G, Gridley T: Notchl is essential for postimplantation development in mice. Genes Dev 8:707, 1994
12. Conlon RA, Reaume AG, Rossant J: Notchl is required for the coordinate segmentation of somites. Development 121:1533, 1995
13. Bierkamp C, Campos-Ortega JA: A zebrafish homologue of the Drosophila neurogenic gene Notch and its pattern of transcription during early embryogenesis. Mech Dev 43:87, 1993
14. Weinmaster G, Roberts VJ, Lemke C: A homolog of Dro- sophila Notch expressed during mammalian development. Develop- ment 113:119, 1991
15. Ellisen LW, Bird J, West DC, Soreng AL, Reynolds TC, Smith SD, Sklar J: TAN-l, the human homolog of the Drosophila
Notch gene, is broken by chromosomal translocations in T lympho- blastic neoplasms. Cell 66649, 1991
16. Aster J, Pear W, Hasserjian R, Erba H, Davi F, Luo B, Scott M, Baltimore D, Sklar J: Functional analysis of the TAN-l gene, a human homologue of Drosophila Notch. Cold Spring Harb Symp Quant Biol 59:125, 1994
17. Shan-Rong S, Chaiwan B, Young L, Cote RJ, Taylor C: Antigen retrieval technique utilizing citrate buffer or urea solution for immunohistochemical demonstration of androgen receptor in for- malin-fixed paraffin sections. J Histochem Cytochem 41:1599, 1993
18. Pinkus GS, O’Conner EM, Etheridge CL, Corson JM: Opti- mal immunoreactivity of keratin proteins in formalin-fixed, paraffin- embedded tissue requires preliminary trypsinization. J Histochem Cytochem 33:465, 1985
19. Hough MR, Takei F, Humphries RK, Kay R: Defective devel- opment of thymocytes overexpressing the costimulatory molecule, heat-stable antigen. J Exp Med 179:177, 1994
20. Godfrey DI, Kennedy J, Suda T, Zlotnik A: A developmental pathway involving four phenotypically and functionally distinct sub- sets of CD3-CD4-CD8-triple-negative adult mouse thymocytes de- fined by CD44 and CD25 expression. J Immunol 150:4244, 1993
21. Kopan R, Weintraub H: Mouse Notch: expression in hair follicles correlates with cell fate determination. J Cell Biol 121:631, 1993
22. Egerton M, Scollay R, Shortman K: Kinetics of mature T- cell development in the thymus. Proc Natl Acad Sci USA 87:2579, 1990
23. Corsky RI, Rapaport DH, Harris WH: Xotch inhibits cell differentiation in the Xenopus retina. Neuron 14:487, 1995
24. Austin CP, Feldman DE, Ida JA, Cepko CL: Vertebrate retinal ganglion cells are selected from competent progenitors by the action of Notch. Development 121:3637, 1995
25. Spits H, Lanier LL, Phillips JH: Development of human T and natural killer cells. Blood 85:2654, 1995
26. Zhiiiga-Pfliicker JC, Jiang D, Lenardo MJ: Requirement for TNF-a and IL-la in fetal thymocyte commitment and differentia- tion. Science 268:1906, 1995
27. Palmer DB, Hayday A, Owen MJ: Is TCRP expression an essential event in early thymocyte development? Immunol Today 14:460, 1993
28. Pear WS, Aster JC, Scott ML, Hasserjian RP, Soffer B, Sklar J, Baltimore D: Exclusive development of T-cell neoplasms in mice transplanted with bone marrow expressing activated Notch alleles. J Exp Med (in press)
29. Fowlkes BJ, Pardoll DM: Molecular and cellular events of T cell development. Adv Immunol44:273, 1989
30. Lindsell CE, Shawber CJ, Boulter J, Weinmaster G: Jagged: A mammalian ligand that activates Notchl. Cell 80:909, 1995
31. Sasai Y, Kageyama R, Tagawa Y, Shigemoto R, Shigetada N: Two mammalian helix-loop-helix factors structurally related to Drosophila hairy and Enhancer of split. Genes Dev 6:2620, 1992
32. Janiault S, Brou C, Logeat F, Schroeter E, Kopan R, Israel A: Signaling downstream of activated mammalian Notch. Nature 377:355, 1995
For personal use only.on April 10, 2019. by guest www.bloodjournal.orgFrom

1996 88: 970-976
RP Hasserjian, JC Aster, F Davi, DS Weinberg and J Sklar Modulated expression of notch1 during thymocyte development
http://www.bloodjournal.org/content/88/3/970.full.htmlUpdated information and services can be found at:
Articles on similar topics can be found in the following Blood collections
http://www.bloodjournal.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://www.bloodjournal.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://www.bloodjournal.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
Copyright 2011 by The American Society of Hematology; all rights reserved.Society of Hematology, 2021 L St, NW, Suite 900, Washington DC 20036.Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by the American
For personal use only.on April 10, 2019. by guest www.bloodjournal.orgFrom
Related Documents











