UNIVERSIDADE ESTADUAL PAULISTA “JULIO DE MESQUITA FILHO” FACULDADE DE CIÊNCIAS AGRÁRIAS E VETERINÁRIAS CÂMPUS DE JABOTICABAL MODELOS DE AVALIAÇÃO GENÉTICA ALTERNATIVOS NA PRESENÇA DE DIFERENÇAS GENÉTICAS ENTRE AMBIENTES Roberto Carvalheiro Zootecnista Orientadora: Profa. Dra. Sandra Aidar de Queiroz Co-orientadores: Dr. Luiz Alberto Fries Dr. Flávio Schramm Schenkel Tese apresentada à Faculdade de Ciências Agrárias e Veterinárias, Câmpus de Jaboticabal - UNESP, para a obtenção do Título de Doutor em Zootecnia (Produção Animal). JABOTICABAL - SÃO PAULO - BRASIL 2004

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

UNIVERSIDADE ESTADUAL PAULISTA “JULIO DE MESQUITA FILHO”FACULDADE DE CIÊNCIAS AGRÁRIAS E VETERINÁRIAS
CÂMPUS DE JABOTICABAL
MODELOS DE AVALIAÇÃO GENÉTICA ALTERNATIVOS NAPRESENÇA DE DIFERENÇAS GENÉTICAS ENTRE
AMBIENTES
Roberto CarvalheiroZootecnista
Orientadora: Profa. Dra. Sandra Aidar de QueirozCo-orientadores: Dr. Luiz Alberto Fries
Dr. Flávio Schramm Schenkel
Tese apresentada à Faculdade deCiências Agrárias e Veterinárias, Câmpusde Jaboticabal - UNESP, para a obtençãodo Título de Doutor em Zootecnia(Produção Animal).
JABOTICABAL - SÃO PAULO - BRASIL
2004

ii
DADOS CURRICULARES DO AUTOR
Roberto Carvalheiro, filho de Manuel Henriques Carvalheiro e de Alda Gonçalves
Carvalheiro, nasceu em 27 de setembro de 1974 em São Paulo, SP. Em janeiro de
1998, graduou-se no curso de Zootecnia pela Universidade Federal de Lavras (UFLA),
onde foi bolsista do Programa Institucional de Bolsa de Iniciação Científica –
CNPq/PIBIC no período de agosto de 1995 a julho de 1997. Em março de 2000, obteve
o título de Mestre em Zootecnia pela Faculdade de Ciências Agrárias e Veterinárias,
Universidade Estadual Paulista “Julio de Mesquita Filho”, Câmpus de Jaboticabal
(FCAV–UNESP/Jaboticabal), onde foi bolsista da CAPES no período de março de 1998
a fevereiro de 2000. Em fevereiro de 2004, obteve o título de Doutor em Zootecnia pela
FCAV–UNESP/Jaboticabal, sendo bolsista do CNPq por seis meses, da FAPESP por
dois anos e seis meses, e da CAPES - do programa de doutorado com estágio no
exterior (PDEE) - por um ano. O estágio no exterior foi realizado no Centre for Genetic
Improvement of Livestock, Department of Animal and Poultry Science, University of
Guelph, Guelph, Ontario, Canada, no período de setembro de 2002 a agosto de 2003.

iii
Dedico
À minha família e à Liziane, por tudo que vocês são e representam para mim!
Agradeço
À Profa. Dra. Sandra Aidar de Queiroz, ao Dr. Luiz Alberto Fries e ao Dr. Flávio
Schramm Schenkel, pela amizade, orientação, ensinamentos, apoio, motivação, ...
Vocês foram fundamentais para o meu crescimento profissional e pessoal!
Aos professores do Departamento de Zootecnia e do Departamento de Ciências Exatas
da FCAV-UNESP/Jaboticabal, pelo agradável convívio e por toda a contribuição em
minha formação acadêmica.
À Coordenadora do Programa de Pós-Graduação em Zootecnia da FCAV-
UNESP/Jaboticabal, Profa. Dra. Telma Teresinha Berchielli Moreno, por todo o auxílio e
incentivo prestados ao longo do curso.
Aos membros da Comissão Examinadora, Prof. Dr. Danísio Prado Munari, Prof. Dr.
Elias Nunes Martins, Dr. Roberto Augusto de Almeida Torres Júnior, Dr. Maurício Mello
de Alencar, pelas contribuições valiosas. Foi uma honra contar com a participação de
vocês!
Ao CNPq, FAPESP e CAPES, pela concessão de recursos durante períodos distintos
do doutorado.
Aos pesquisadores do Centre for Genetic Improvement of Livestock, University of
Guelph, em especial ao Dr. S. P. Miller, pela oportunidade prestada para a realização
do doutorado “sanduíche”, e ao Dr. L. R. Schaeffer, pelos ensinamentos.

iv
Aos amigos do GenSys, por todo o auxílio, apoio e motivação prestados ao longo do
doutorado.
À todos os amigos da pós-graduação, em especial aos meus contemporâneos, Laila,
Rodrigo, Luciele e Vânia. Foi uma benção e um privilégio ter a oportunidade de
“caminhar” com vocês!
Por fim, gostaria de agradecer de forma especial aos amigos da família Schenkel -
Sandra, Mariana, Daniel e Flávio - e ao amigo Vanerlei, pela acolhida formidável e por
terem tornado muito agradável a minha estada no Canadá.

v
SUMÁRIO
Resumo ........................................................................................................ vii
Summary..................................................................................................... viii
Capítulo 1 - Considerações Gerais ............................................................. 1
1.1 – Problema em estudo ............................................................................. 1
1.2 – Justificativa para a condução do estudo ............................................... 3
1.3 – Objetivos do estudo .............................................................................. 4
1.4 – Estratégias de desenvolvimento do trabalho......................................... 4
Capítulo 2 - Propriedades empíricas das soluções das equações demodelo misto de Henderson para diferentes situações deassociação entre touros e grupos de contemporâneos...... 6
Resumo ................................................................................................ 6
2.1 – Introdução ............................................................................................. 7
2.2 – Material e Métodos................................................................................ 8
2.2.1 – Estrutura das populações simuladas ....................................... 8
2.2.2 – Modelo de simulação ............................................................ 11
2.2.3 – Análises estatísticas ............................................................. 11
2.2.4 – Critérios de comparação dos modelos .................................. 13
2.3 – Resultados e Discussão...................................................................... 14
2.3.1 – Dados simulados ................................................................... 14
2.3.2 – Soluções para o efeito de grupo de contemporâneos ........... 16
2.3.3 – Soluções para touros............................................................. 18
2.3.4 – Discussão geral ..................................................................... 23
2.4 – Conclusões ......................................................................................... 26

vi
Capítulo 3 - Modelos de avaliação genética alternativos em situaçõesde seleção parental............................................................... 27
Resumo .............................................................................................. 27
3.1 – Introdução ........................................................................................... 28
3.2 – Material e Métodos.............................................................................. 29
3.2.1 – Estrutura da população simulada .......................................... 30
3.2.2 – Modelo de simulação............................................................. 30
3.2.3 – Critérios de seleção .............................................................. 31
3.2.4 – Informações adicionais do processo de simulação ............... 32
3.2.5 – Modelos de avaliação genética ............................................ 32
3.2.6 – Critérios de comparação dos modelos .................................. 36
3.3 – Resultados e Discussão...................................................................... 37
3.3.1 – Viés........................................................................................ 37
3.3.2 – Médias dos VGVs.................................................................. 42
3.3.3 – Tendências genéticas............................................................ 44
3.3.4 – Discussão geral ..................................................................... 49
3.4 – Implicações ......................................................................................... 51
3.5 – Conclusões ......................................................................................... 52
Referências Bibliográficas ....................................................................... 53
Apêndice 1 ................................................................................................. 57
Apêndice 2 ................................................................................................. 59
Apêndice 3 ................................................................................................. 65
Apêndice 4 ................................................................................................. 69
Apêndice 5 ................................................................................................. 76

vii
MODELOS DE AVALIAÇÃO GENÉTICA ALTERNATIVOS NA PRESENÇA DEDIFERENÇAS GENÉTICAS ENTRE AMBIENTES
Resumo: Investigou-se, por meio de simulação de dados, propriedades
empíricas das soluções fornecidas pelo modelo misto de Henderson (MMH) e por
modelos de avaliação alternativos, sob diferentes situações de diferenças genéticas
entre grupos de contemporâneos (GCs). Num primeiro estudo, diferenças genéticas
entre GCs foram geradas pela distribuição não aleatória da progênie dos touros nos
GCs. Sob esta condição, as soluções do modelo MMH foram viesadas, o que reduziu a
habilidade de MMH em classificar apropriadamente os animais, bem como sua
eficiência em selecionar animais com mérito genético superior. Dentre os modelos
alternativos testados, o modelo que assumiu touro como efeito fixo e vaca como efeito
aleatório apresentou os melhores resultados quanto aos critérios estudados,
mostrando-se superior a MMH nas situações de diferenças genéticas entre GCs, e
similar na situação controle. No segundo estudo, diferenças genéticas entre GCs foram
geradas pelo processo de seleção parental. Foram conduzidos três tipos de seleção
parental: com base no BLUP; no fenótipo; e no valor genético verdadeiro (VGV). O
modelo MMH apresentou soluções viesadas quando houve seleção com base no VGV,
e o viés dos modelos alternativos ocorreu principalmente na situação de seleção
baseada no BLUP. O viés apresentado pelo modelo MMH ocorreu no sentido de
subestimar as soluções dos touros usados em gerações com animais de mérito
genético superior. O viés apresentado pelos modelos alternativos, com seleção
baseada no BLUP, ocorreu no sentido oposto. O viés apresentado nas diferentes
situações teve conseqüências sobre a habilidade de cada modelo em selecionar
animais de mérito genético superior e em estimar a tendência genética. Quanto à estas
propriedades, evidências foram observadas de que modelos alternativos podem
apresentar resultados superiores aos do modelo MMH, e que nem sempre a seleção,
ou mais explicitamente, as diferenças genéticas entre níveis dos efeitos ambientais,
deveriam ser ignoradas nas avaliações genéticas.
Palavras-chave: BLUP, efeito de seleção, modelo misto, predição viesada

viii
ALTERNATIVE GENETIC EVALUATION MODELS UNDER THE PRESENCE OFMEAN GENETIC DIFFERENCES BETWEEN LEVELS OF ENVIRONMENTAL
EFFECTS
Summary: Stochastic simulations were carried out to investigate empirical
proprieties of solutions from Henderson's Mixed Model (HMM) and other alternative
models, under the presence of mean genetic differences between levels of
contemporary group effects (CGs). In a first study, mean genetic differences were
caused by non-randomly use of sires across CGs. Under this situation, HMM solutions
were biased. The bias reduced the ability of HMM in properly ranking animals and its
efficiency in selecting superior genetic animals. The alternative model, that assumed sire
as fixed and dam as random effects, presented, in general, better empirical statistical
and genetic proprieties than the other studied models under the presence of mean
genetic differences, and it was similar to HMM in a control situation. In the second study,
mean genetic differences between CGs were generated as a result of parental selection.
Three types of parental selection were carried out: based on BLUP, phenotype and true
breeding value (TBV). MMH presented biased solutions when selection was based on
TBV, the bias being in the direction of underestimating superior genetic sires. The
biases in the alternative tested models occurred mainly for BLUP selection, where sires
with superior genetic merit tended to be over evaluated. Biases presented by the
different models for each situation affected models’ ability to estimate genetic trends and
to select better animals. Regarding to these proprieties, evidences were observed that
alternative models can provide better results than MMH, and that genetic mean
differences between levels of CG effects should not always be ignored in genetic
evaluations.
Keywords: biased estimation, BLUP, mixed model, selection effect

1
CAPÍTULO 1 – CONSIDERAÇÕES GERAIS
1.1 - Problema em estudo
Há interesse, em programas de melhoramento, na predição de valores genéticos
e na utilização destas predições, ou funções das mesmas, como critério de seleção dos
animais, para que progresso genético possa ser obtido.
Sob normalidade e na classe de funções invariantes a translação, o melhor
preditor linear não viesado (BLUP), desenvolvido por HENDERSON (1949) e com
propriedades descritas por HENDERSON et al. (1959) e por HENDERSON (1963),
apresenta-se como critério de seleção ótimo no sentido de maximizar a probabilidade
de classificação correta dos animais quanto ao mérito genético, assim como de
maximizar a média dos valores genéticos de um determinado número de animais
selecionados (HENDERSON, 1973; FERNANDO & GIANOLA, 1986).
Na derivação do BLUP, pressupõe-se que a esperança marginal dos valores
genéticos de todos os animais que estão sendo avaliados seja igual a zero (E(ai)=0).
Esta pressuposição, como discutido a seguir, é raramente atendida para dados de
campo de programas de melhoramento animal.
A violação da pressuposição E(ai)=0 é geralmente tratada na literatura como um
problema de efeito de seleção. Entretanto, cabe salientar que forças ou práticas de
manejo dos animais que atuam no sentido de gerar diferenças genéticas entre níveis
dos efeitos ambientais podem fazer com que a pressuposição E(ai)=0 não seja válida.
Na prática, diferenças genéticas entre ambientes podem ser geradas como resultado de
tomadas de decisão dos produtores (ex.: quais touros usar e como alocar sua progênie
em diferentes grupos de manejo). A seleção parental, por exemplo, se bem aplicada,
faz com que animais de gerações posteriores tenham valor genético esperado superior
ao da média dos valores genéticos da população dos animais que os geraram.
Diferenças genéticas também podem ser criadas a partir do uso não aleatório dos
touros em diferentes ambientes (ex.: touros com mérito genético superior sendo
utilizados nos rebanhos com melhores condições ambientais).

2
Como conseqüência da violação de E(ai)=0, as soluções das Equações de
Modelo Misto de Henderson (EMMH), que forneceriam sob condições ideais o BLUP
dos valores genéticos, não são garantidas de serem não viesadas (HARVEY, 1979;
FRIES & SCHENKEL, 1993). Por sua vez, o viés das soluções das EMMH pode fazer
com que animais com mérito genético inferior sejam selecionados para a reprodução,
podendo também afetar as estimativas e predições de tendências ambientais e
genéticas, bem como a proporção de animais jovens de determinado rebanho a receber
certificado de superioridade genética, e outras conseqüências práticas. Resultados
nesse sentido foram observados por HARVEY (1979), SIMIANER & WOLLNY (1989),
UGARTE et al. (1992), FRIES & SCHENKEL (1993), VISSCHER & GODDARD (1993),
TÄUBERT et al. (2002), entre outros.
HENDERSON (1973 e 1975) apresentou condições especiais em que as
soluções das EMMH regulares são não viesadas mesmo que a pressuposição E(ai)=0seja violada. Tais condições, como, por exemplo, informação de parentesco completa e
decisões tomadas exclusivamente com base em funções invariantes a translação ou em
variáveis não correlacionadas ao valor genético, dificilmente são atendidas em
conjuntos de dados reais oriundos de programas de melhoramento. Para casos em que
estas condições não são atendidas, HENDERSON (1975) propôs modificações nas
EMMH regulares para que o BLUP dos valores genéticos fosse obtido. As EMMH
modificadas requerem uma matriz para descrever o processo seletivo que, no geral,
não é trivial de ser construída. Além disso, não há garantia de que as EMMH
modificadas forneçam valores genéticos preditos que sejam um critério de seleção
ótimo no sentido de permitir a seleção de animais com a média dos valores genéticos
maximizada.
GIANOLA et al. (1988), GIANOLA et al. (1989) e IM et al. (1989) propuseram
modelos alternativos baseados em métodos de estimação por máxima verossimilhança.
Nestes trabalhos, são apresentados argumentos favoráveis ao uso de funções de
verossimilhança não condicionais ao processo de seleção (uL). Se uL for utilizada, o
processo de seleção é ignorável (as funções de verossimilhança resultariam nos
mesmos estimadores considerando ou não os processos de seleção) quando for

3
baseado apenas em informação contida nos dados para análise, ou nos dados e em
variáveis independentes a eles. Caso contrário, o conhecimento do processo de
seleção é necessário para que inferência correta seja feita sobre os parâmetros de
interesse. Se a função conjunta das variáveis aleatórias e das observações fenotípicas
for maximizada condicionalmente ao processo de seleção (cL), as condições para
ignorá-lo são, de acordo com IM et al. (1989), ainda mais restritas do que as
apresentadas por HENDERSON (1975). GIANOLA et al. (1988) derivaram as equações
resultantes da maximização de cL e descreveram as situações especiais em que os
processos de seleção poderiam ser ignorados, como, por exemplo, quando a seleção
ocorre com base em funções das observações fenotípicas que sejam invariantes a
translação.
A derivação do BLUP pressupondo E(ai)≠≠≠≠0 resulta em soluções para os efeitos
ambientais e genéticos sendo obtidas pelo método de quadrados mínimos ponderados
(FRIES & SCHENKEL, 1993; SCHENKEL, 1998), ou pelo método de quadrados
mínimos ordinários (OLS) se homogeneidade de variância residual for assumida. Neste
caso, os efeitos ambientais e genéticos seriam tratados como efeitos fixos e, no geral,
não haveria possibilidade de aplicação do modelo animal individual em virtude da
deficiência de posto da matriz de coeficientes. Uma alternativa seria a utilização de um
modelo touro e vaca, considerando-os como efeitos fixos. Entretanto, assim como nos
modelos de seleção propostos por HENDERSON (1975), não há garantia de que esse
modelo forneça valores genéticos preditos que sejam um critério de seleção ótimo no
sentido de maximizar a média dos valores genéticos dos animais selecionados.
1.2 - Justificativa para a condução do estudoComo não há, até o presente momento, uma solução estatística clara e definitiva
para tratar a não aleatoriedade de ocorrência dos níveis de efeito genético nos níveis
de efeito ambiental, isto é, para tratar forças que atuam no sentido de gerar diferenças
genéticas entre ambientes, torna-se importante a condução de estudos comparando
diferentes modelos e procurando aqueles que forneçam melhores critérios de seleção
sob estas condições.

4
1.3 - Objetivos do estudoOs principais objetivos do presente trabalho foram: (1) avaliar propriedades das
soluções fornecidas pelas Equações de Modelos Mistos de Henderson, sob diferentes
situações de diferenças genéticas entre ambientes; (2) comparar modelos alternativos
de avaliação genética, para cada situação estudada, com base nas propriedades das
soluções fornecidas pelos diferentes modelos; e (3) fornecer informações que possam
auxiliar os melhoristas na escolha de modelos mais apropriados para situações de
diferenças genéticas entre ambientes.
1.4 - Estratégias de desenvolvimento do trabalho1.4.1 - Como comparar os modelos?
Na dificuldade de comparar analiticamente diferentes modelos quanto às suas
propriedades estatísticas e genéticas, especialmente quando E(ai)=0 não é uma
pressuposição atendida, optou-se pela condução de estudos de simulação, seguindo
recomendações de HENDERSON (1989 e 1990).
1.4.2 - Que situações simular?
Na deficiência e impossibilidade de simular todas as situações práticas possíveis,
dois casos isolados foram escolhidos de forma a gerar diferenças genéticas entre
ambientes. Em um primeiro trabalho, investigaram-se as propriedades empíricas das
soluções fornecidas por diferentes modelos sob situações de associação não aleatória
entre touros e grupos de contemporâneos. Um segundo estudo foi conduzido para
comparar os modelos na presença de diferentes tipos de seleção parental.
1.4.3 - Que modelos comparar?
As EMMH foram aplicadas sob seu modelo usual, considerando os valores
genéticos como efeito aleatório e os efeitos de GC como fixos. Os modelos alternativos
utilizados foram baseados nas propostas de FRIES & SCHENKEL (1993) e de
SCHAEFFER (2003), de considerar os valores genéticos ou parte deles como efeitos
fixos. Optou-se pelo emprego de tais modelos porque: (1) há evidências na literatura de

5
que os mesmos possam ser mais apropriados para situações de diferenças genéticas
entre ambientes (HARVEY, 1979; FRIES & SCHENKEL, 1993; SCHAEFFER, 2003); (2)
são modelos que não requerem a descrição/conhecimento dos processos responsáveis
pela não aleatoriedade de ocorrência dos níveis de efeito genético nos níveis de efeito
ambiental; e (3) são modelos de fácil implementação computacional.

6
CAPÍTULO 2 - PROPRIEDADES EMPÍRICAS DAS SOLUÇÕES DAS EQUAÇÕES DEMODELO MISTO DE HENDERSON PARA DIFERENTES SITUAÇÕES DEASSOCIAÇÃO ENTRE TOUROS E GRUPOS DE CONTEMPORÂNEOS
Resumo: Simulações estocásticas foram conduzidas com o objetivo de
investigar propriedades empíricas das soluções das equações de modelo misto de
Henderson (EMMH) sob quatro diferentes situações (ST1 a ST4) de associação entre
touros e grupos de contemporâneos (GC). Os touros tiveram progênie distribuída nos
GCs independentemente de seus valores genéticos verdadeiros (VGV) na situação
ST1, e de acordo com seus VGV nas situações ST2 a ST4, em que diferenças
genéticas entre GCs foram geradas. ST2 foi caracterizada por ausência de correlação
entre os efeitos de touro e de GC. ST3 e ST4 apresentaram, respectivamente,
correlação positiva e negativa entre os efeitos de touro e de GC. As EMMH foram
aplicadas sob um modelo que incluiu os efeitos de GC como fixo, e touro e vaca como
aleatórios (M1). Três modelos alternativos foram usados (M2 a M4). GC, touro e vaca
foram assumidos como efeitos aleatórios no modelo M2, e como efeitos fixos em M3.
No modelo M4, consideraram-se os efeitos de GC e touro como fixos e vaca como
aleatório. Na situação em que não houve diferenças genéticas entre GCs (ST1), os
modelos M1, M2 e M4 apresentaram resultados semelhantes entre si quanto aos
critérios de comparação estudados, enquanto que M3 forneceu soluções com maior
quadrado médio do erro de predição em comparação com os demais modelos. Na
presença de diferenças genéticas entre GCs (ST2 a ST4), as soluções do M1 foram
viesadas, o que reduziu sua habilidade em classificar apropriadamente os animais, bem
como sua eficiência em selecionar touros geneticamente superiores. Dentre os modelos
alternativos testados, M4 apresentou os melhores resultados quanto aos critérios
estudados, mostrando-se superior a M1 nas situações ST2 a ST4, e similar na ST1.
Palavras-chave: avaliação genética, BLUP, correlação entre efeitos ambientais e
genéticos, predições viesadas

7
2.1 - Introdução
Em melhoramento animal, o modelo de avaliação genética usualmente aplicado,
proposto por HENDERSON (1963), é baseado na pressuposição de que a esperança
marginal dos valores genéticos de todos os animais que estão sendo avaliados seja
igual a zero (E(ai)=0).
Diferentes formas de seleção ou práticas que resultem na distribuição não
aleatória dos animais nos níveis de efeitos ambientais (ex.: grupos de contemporâneos
(GC)) podem gerar diferenças genéticas entre estes níveis e, consequentemente,
invalidar a pressuposição E(ai)=0. A seleção parental, por exemplo, se bem aplicada,
faz com que animais de gerações posteriores tenham valor genético esperado superior
ao da média dos valores genéticos da população dos animais que os geraram.
Diferenças genéticas também podem ser criadas a partir de associações não aleatórias
entre touros e GCs como, por exemplo, touros com mérito genético superior sendo
utilizados nos melhores ambientes.
Como conseqüência da violação de E(ai)=0, as soluções das Equações de
Modelo Misto de Henderson (EMMH), que forneceriam sob condições ideais o BLUP
dos valores genéticos, não são garantidas de serem não viesadas (HARVEY, 1979;
FRIES & SCHENKEL, 1993). HENDERSON (1973, 1975) apresentou condições
especiais em que as soluções das EMMH são não viesadas mesmo que a
pressuposição E(ai)=0 seja violada. Tais condições, como, por exemplo, informação de
parentesco completa e diferenças genéticas geradas com base em funções das
observações fenotípicas invariantes a translação, dificilmente são atendidas em
conjuntos de dados reais oriundos de programas de melhoramento.
O viés das soluções das EMMH pode fazer com que animais com mérito
genético inferior sejam selecionados para a reprodução, podendo também afetar as
estimativas e predições de tendências genéticas e ambientais, bem como a proporção
de animais jovens de determinado rebanho a receber certificado de superioridade
genética. Diante de tais conseqüências, torna-se importante o estudo de modelos de
avaliação genética alternativos e o fornecimento de informações que auxiliem os

8
melhoristas na escolha de modelos mais adequados ou menos sensíveis à violação da
pressuposição E(ai)=0.
Os objetivos do presente trabalho foram os de investigar, por meio de simulação
estocástica, propriedades empíricas das soluções das EMMH sob situações de
diferenças genéticas entre GCs causadas pela associação não aleatória entre os
efeitos de touro e de GC, e comparar modelos alternativos nestas situações.
2.2 - Material e Métodos2.2.1 - Estrutura das populações simuladas
A população base simulada consistiu de 250 machos (touros) e 2.500 fêmeas
(vacas), não aparentados, não selecionados e assumidos de serem amostrados
aleatoriamente de uma população conceitualmente infinita. Cada vaca foi acasalada
duas vezes, com touros escolhidos aleatoriamente, gerando dois produtos, um por
acasalamento. Apenas uma geração de progênie foi simulada. Os 5.000 produtos
gerados foram distribuídos em 200 grupos de contemporâneos (GC) de acordo com
quatro situações distintas (ST1 a ST4).
Para gerar as diferentes situações, cinco conjuntos de 40 GCs (sGC1 a sGC5) e
cinco conjuntos de 50 touros foram formados com base na sua identificação (ID) ou na
classificação dos valores verdadeiros de seus efeitos (rank), dependendo da situação
simulada (segue ilustração na Tabela 2.1, e no Apêndice 1). Os touros foram agrupados
conforme sua ID na ST1 e conforme seu rank, em ordem decrescente dos valores
genéticos verdadeiros, nas situações ST2 a ST4. Os GCs foram agrupados conforme
sua ID nas situações ST1 e ST2, e seu rank nas situações ST3 (ordem decrescente) e
ST4 (ordem crescente).
9
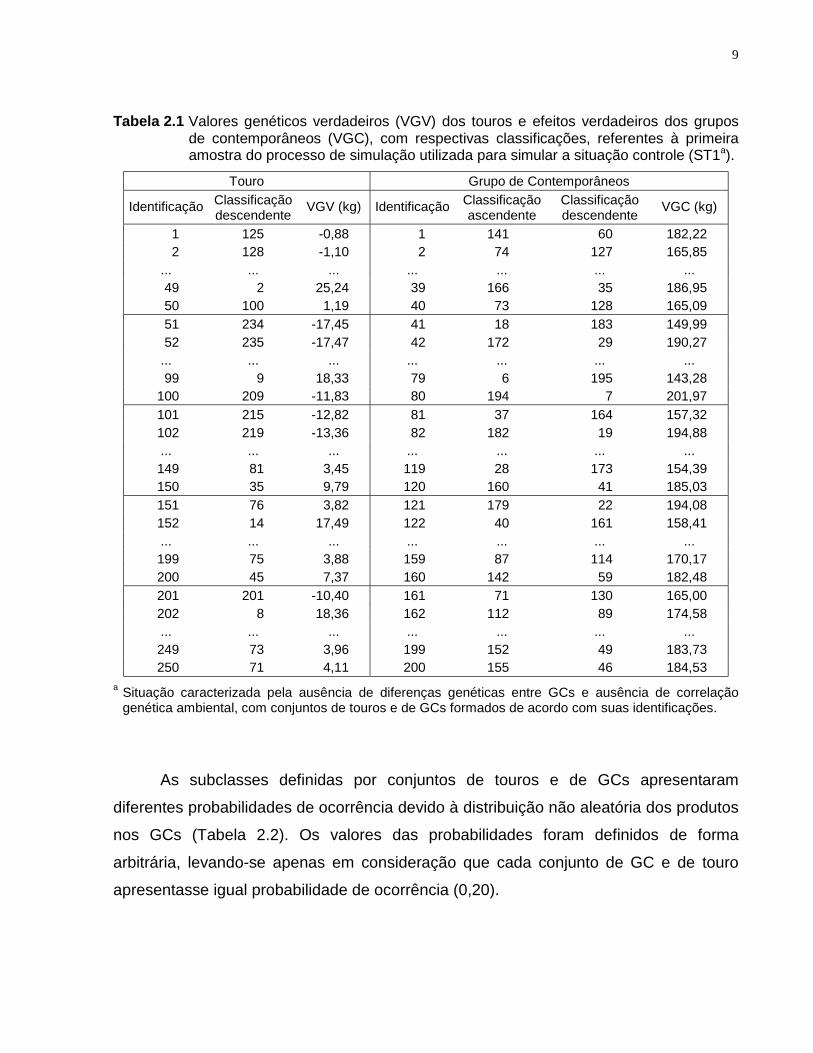
Tabela 2.1 Valores genéticos verdadeiros (VGV) dos touros e efeitos verdadeiros dos gruposde contemporâneos (VGC), com respectivas classificações, referentes à primeiraamostra do processo de simulação utilizada para simular a situação controle (ST1a).
Touro Grupo de Contemporâneos
Identificação Classificaçãodescendente VGV (kg) Identificação Classificação
ascendenteClassificaçãodescendente VGC (kg)
1 125 -0,88 1 141 60 182,222 128 -1,10 2 74 127 165,85
... ... ... ... ... ... ...49 2 25,24 39 166 35 186,9550 100 1,19 40 73 128 165,0951 234 -17,45 41 18 183 149,9952 235 -17,47 42 172 29 190,27
... ... ... ... ... ... ...99 9 18,33 79 6 195 143,28
100 209 -11,83 80 194 7 201,97101 215 -12,82 81 37 164 157,32102 219 -13,36 82 182 19 194,88... ... ... ... ... ... ...149 81 3,45 119 28 173 154,39150 35 9,79 120 160 41 185,03151 76 3,82 121 179 22 194,08152 14 17,49 122 40 161 158,41... ... ... ... ... ... ...199 75 3,88 159 87 114 170,17200 45 7,37 160 142 59 182,48201 201 -10,40 161 71 130 165,00202 8 18,36 162 112 89 174,58... ... ... ... ... ... ...249 73 3,96 199 152 49 183,73250 71 4,11 200 155 46 184,53
a Situação caracterizada pela ausência de diferenças genéticas entre GCs e ausência de correlaçãogenética ambiental, com conjuntos de touros e de GCs formados de acordo com suas identificações.
As subclasses definidas por conjuntos de touros e de GCs apresentaram
diferentes probabilidades de ocorrência devido à distribuição não aleatória dos produtos
nos GCs (Tabela 2.2). Os valores das probabilidades foram definidos de forma
arbitrária, levando-se apenas em consideração que cada conjunto de GC e de touro
apresentasse igual probabilidade de ocorrência (0,20).

10
Tabela 2.2 Freqüências (%) esperadas e observadas da ocorrência das subclasses definidaspor conjuntos de touros e conjuntos de grupos de contemporâneos (GCs).
Conjuntos Conjuntos de GCs a
de Touros b 1-40 41-80 81-120 121-160 161-2001-50 12 c (11,96 d) 6 (5,93) 2 (1,96) 0 (0,00) 0 (0,00)
51-100 6 (6,01) 8 (8,02) 4 (4,04) 2 (1,98) 0 (0,00)101-150 2 (2,00) 4 (3,98) 8 (8,14) 4 (4,05) 2 (2,00)151-200 0 (0,00) 2 (2,01) 4 (4,05) 8 (7,99) 6 (5,91)201-250 0 (0,00) 0 (0,00) 2 (1,98) 6 (6,10) 12 (11,89)
a Os conjuntos de GCs foram formados conforme o número de identificação do GC nas situações 1 (ST1)e 2 (ST2), de acordo com a classificação descendente dos efeitos verdadeiros de GC (VGC) na ST3, econforme a classificação ascendente de VGC na ST4.
b Os conjuntos de touros foram formados de acordo com o número de identificação do touro na ST1, econforme a classificação descendente de seus valores genéticos verdadeiros nas situações ST2 a ST4.
c Valores esperados, que foram utilizados no processo de simulação.d Valores observados (média das 50 repetições).
Na ST1, situação controle, diferenças genéticas entre GCs não são esperadas
visto que os conjuntos de touros foram formados de acordo com sua ID. Entretanto,
para as situações ST2 a ST4, diferenças genéticas entre GCs foram geradas pois os
touros foram agrupados conforme seu rank, de forma que, sGC1 e sGC2 tiveram maior
proporção de produtos dos melhores touros, sGC3 apresentou maior proporção de
produtos dos touros intermediários, enquanto que sGC4 e sGC5 apresentaram maior
proporção de produtos dos piores touros (Tabela 2.2).
ST2 foi caracterizada por ausência de correlação genética ambiental (rG,E), pois
os GCs foram agrupados com base em sua ID. Já, para as situações ST3 e ST4, rG,E
positivas e negativas foram geradas em decorrência da formação de conjuntos de GCs
baseados em seu rank nas ordens decrescente e crescente, respectivamente.
A conectabilidade dos GCs foi garantida pela imposição de restrições sobre o
valor mínimo para número de produtos por touro (5), produtos por GC (10), touros por
GC (3) e GC por touro (3), e também por meio da aplicação do programa MILC (FRIES,
1998), que considera como dado conectado aquele que estiver em um GC com mais de
10 laços genéticos com outros GCs e que tenha pelo menos três pais (touros e/ou

11
vacas) conectados. Novas replicações foram simuladas caso algum GC estivesse
desconectado.
Para cada situação, 50 repetições foram simuladas, sendo gerados, portanto,
200 conjuntos de dados. Variaram, a cada replicação, o acasalamento entre os animais
base, o número de filhos por touro, a alocação e o número de animais por GC.
2.2.2 - Modelo de simulaçãoA cada repetição, observações fenotípicas de uma característica hipotética, que
será denominada como ganho de peso pós-desmame (GPPD), foram simuladas, exceto
para os animais base, de acordo com o modelo: yij = bi + aij + eij , em que yij é o GPPD
do animal j no GC i, bi é o efeito do GC i, aij é o valor genético aditivo do animal j no GC
i e eij é o termo referente ao erro aleatório.
Valores para eij foram independentemente amostrados de uma distribuição
normal com média zero e variância de 180 kg2 (N(0;180)). Valores para bi foram
amostrados de uma N(172,5;300). Os valores genéticos dos animais base foram
gerados a partir de uma N(0;120), e os valores genéticos de seus produtos foram
gerados como: aij = ½( asj + adj) + mij, em que asj e adj são os valores genéticos aditivos
do touro e da vaca que geraram o animal j, e mij é o efeito da Segregação Mendeliana
do animal j, que foi amostrada de uma N(0;60). Os parâmetros usados no processo de
simulação reproduziram uma característica com herdabilidade igual a 0,4 e são
semelhantes aos estimados por CARVALHEIRO (2000) para a característica GPPD de
um conjunto de dados de bovinos da raça Nelore.
Os programas para simular a estrutura dos dados e as observações fenotípicas
foram desenvolvidos no software estatístico SAS (Statistical Analysis System) (SAS,
1998), e se encontram nos Apêndices 2 e 3, respectivamente.
2.2.3 - Análises Estatísticas
Para cada um dos 200 conjuntos de dados, análises foram conduzidas sob
quatro diferentes modelos, que podem ser descritos pela equação:
y = Xββββ + Zss + Zdd + e ,

12
em que: y é o vetor de GPPD, ββββ é o vetor de efeito de GC, s e d são, respectivamente,
os vetores de efeito genético aditivo de touro e de vaca, e é o vetor de resíduos, X, Zs e
Zd são as matrizes de incidência que relacionam os elementos de ββββ, s e d com os
elementos de y, respectivamente. O modelo de touro e vaca foi utilizado, pois o mesmo
eqüivale ao modelo animal para as situações simuladas no presente estudo, em que
animais base não apresentaram dados (observações fenotípicas) e animais com dados
não apresentaram progênie (HENDERSON, 1984).
No modelo 1 (M1), ββββ foi considerado como vetor de efeito fixo, e s e d como
vetores de efeitos aleatórios. As matrizes de esperança e (co)variância são dadas por:
E(y) = Xββββ, E(s) = E(d) = E(e) = 0, Var(y) = V = 222 '' endddsss IZZZZ σσσσσσσσσσσσ ++ , Var(s) =
2ssI σσσσ , Var(d) = 2
ddI σσσσ e Var(e) = 2enI σσσσ , em que: sI , dI e nI são matrizes identidade de
ordens iguais aos números de touros, de vacas e de observações, respectivamente, 2sσσσσ
e 2dσσσσ são as variâncias dos efeitos genéticos de touro e vaca, que são iguais a um
quarto da variância genética aditiva ( 2sσσσσ = 2
dσσσσ =1/4 2aσσσσ ), e 2
eσσσσ é a variância residual.
Matrizes de (co)variância com uma estrutura diagonal foram utilizadas para s e d, pois
os touros e as vacas eram animais não aparentados nas populações simuladas.
Soluções para ββββ, s, e d foram obtidas com base nas equações de modelos mistos de
Henderson (EMMH) (HENDERSON, 1963).
O modelo 2 (M2) diferiu do M1 com respeito a ββββ. No M2, ββββ foi considerado como
vetor de efeito aleatório. Portanto, para M2, E(y) = µµµµ, E(ββββ) = 0, Var(y) =2en
2ddd
2sss
2gc I'ZZ'ZZ'XX σσσσ++++σσσσ++++σσσσ++++σσσσ e Var(ββββ) = 2
gcgcI σσσσ , em que: µµµµ é o vetor da média
geral, gcI é a matriz de incidência de ordem igual ao número de GC, e 2gcσσσσ é a variância
devida ao efeito de GC. As matrizes de esperança e (co)variância dos demais efeitos
aleatórios são iguais àquelas definidas no M1. As soluções também foram obtidas
conforme as EMMH, mas com o valor da relação entre as variâncias residual e de GC
( 2eσσσσ / 2
gcσσσσ ) adicionado aos elementos da diagonal da matriz de coeficientes (lado
esquerdo do sistema de equações) correspondentes ao efeito de GC.

13
O modelo M2 foi aplicado para investigar as considerações de VAN VLECK
(1987), UGARTE et al. (1992), e VISSCHER & GODDARD (1993) de que benefícios
poderiam ocorrer ao assumir GC como efeito aleatório em situações similares à ST3
simulada no presente trabalho.
No modelo 3 (M3), ββββ, s, e d foram assumidos como vetores de efeitos fixos.
Assim, E(y) = Xββββ + Zss + Zdd, E(e) = 0, e Var(y) = Var(e) = 2enI σσσσ . As soluções para M3
foram obtidas por um procedimento em dois passos. No primeiro, soluções foram
obtidas pelo método de quadrados mínimos ordinários (OLS), impondo-se as restrições
de que a soma dos elementos de s e d fossem iguais a zero. Como segundo passo, as
soluções para s e d foram multiplicadas pelo fator de regressão: np/(np+ 2eσσσσ /(1/4 2
aσσσσ )),
sendo np o tamanho da progênie dos correspondentes elementos em s e d.
Para o quarto modelo empregado (M4), ββββ e s foram considerados como vetores
de efeitos fixos e d como vetor de efeito aleatório, com E(y) = Xββββ + Zss, E(d) = E(e) = 0,
Var(y) = 22' enddd IZZ σσσσσσσσ + , Var(d) = 2ddI σσσσ , e Var(e) = 2
enI σσσσ . As soluções foram obtidas
conforme equações similares as EMMH, mas sem adicionar a relação entre as
variâncias residual e de touro ( 2eσσσσ / 2
sσσσσ ) aos elementos da diagonal da matriz de
coeficientes correspondentes ao efeito de touro. A restrição de que a soma dos
elementos em s fosse igual a zero foi imposta, e as soluções para s foram regredidas
como segundo passo usando o mesmo fator de regressão definido para o M3.
Para todos os modelos, os valores genéticos esperados (VGE) dos touros e
vacas foram obtidos multiplicando-se as soluções para s e d por dois. Os componentes
de variância usados nas análises foram iguais àqueles empregados no processo de
simulação dos dados, ou seja, não foram estimados.
Os programas desenvolvidos para conduzir as avaliações genéticas, sob os
quatro diferentes modelos, encontram-se no Apêndice 4.
2.2.4 - Critérios de comparação dos modelosOs modelos foram comparados com relação ao viés e ao quadrado médio do
erro de predição (QME), que foram definidos por:

14
viésw = 1/q∑=
−q
iii ww
1
)ˆ( , e QMEw = 1/q∑=
−q
iii ww
1
2)ˆ( ,
em que: wi é o efeito verdadeiro de GC (VGC) ou efeito genético verdadeiro (VGV) de
touro ou de vaca, iw é o efeito estimado de GC (EGC) ou o VGE de touro ou de vaca,
e q é o número de GCs, touros ou vacas, respectivamente.
Correlações de Spearman entre VGV e VGE foram calculadas como uma
alternativa de avaliar a habilidade de cada modelo em classificar apropriadamente os
animais. Na tentativa de mensurar a eficiência de cada modelo em selecionar animais
de mérito genético superior, também foram calculadas as médias dos VGVs de touros
selecionados conforme os VGEs fornecidos pelos diferentes modelos. Para este último
critério, foram conduzidos testes-t pareados com um nível de significância de 5%.
Para todos os critérios descritos acima, os modelos foram comparados com base
na média aritmética dos valores que cada critério assumiu em cada uma das 50
repetições.
2.3 - Resultados e Discussão2.3.1 - Dados Simulados
Resultados referentes à estrutura das populações simuladas são apresentados
na Tabela 2.3. Em média, 9,2% das subclasses definidas por touro e GC foram
preenchidas, os touros apresentaram o mínimo de 8,6 e o máximo de 33,5 produtos, e
os GCs apresentaram o mínimo de 12,5 e o máximo de 40,3 animais.
Tabela 2.3 Informações referentes à estrutura das populações simuladas (médias das 50repetições).
Média Mínimo MáximoProdutos por touro 20,00 8,60 33,48Produtos por GC 25,00 12,54 40,28Touros por GC 22,94 11,54 36,08GCs por touro 18,35 8,12 30,04% de subclasses touro x GCpreenchidas 9,18 9,10 9,26

15
Na Tabela 2.4, são apresentadas as médias e variâncias dos valores genéticos
verdadeiros (VGV) dos produtos e dos efeitos verdadeiros de GC (VGC) para as
diferentes situações simuladas. Como desejado, os valores observados das médias e
variâncias gerais (última linha da tabela) foram próximos aos parâmetros utilizados no
processo de simulação. As médias de VGV por conjunto de GC foram próximas a zero
na situação ST1, e variaram de –5,52 a 5,36 kg nas situações ST2 a ST4. As variâncias
de VGV foram reduzidas nas situações ST2 a ST4, mas foram semelhantes entre os
conjuntos de GC, isto é, não houve heterogeneidade de variância genética. As médias
de VGC por conjunto de GC foram próximas a 172,5 kg em ST1 e ST2, e variaram de
148,37 a 197,27 kg em ST3 e ST4. Heterogeneidade de variância de VGC entre os
conjuntos de GC foi observada nas situações ST3 e ST4. Seguindo o processo de
simulação, ST1 foi caracterizada por ausência de diferenças genéticas entre GCs e de
correlação entre o VGV dos produtos e o VGC (rG,E); ST2, ST3 e ST4 apresentaram
diferenças genéticas entre GCs, e rG,E iguais a 0,00, 0,34, e –0,34, respectivamente (as
correlações entre o VGV dos touros e o VGC foram iguais a 0,00 para ST1 e ST2, 0,68
para ST3 e –0,68 para ST4).
Tabela 2.4 Médias (kg) e variânciasa (kg2) dos valores genéticos verdadeiros dos produtos(VGV) e dos efeitos verdadeiros dos grupos de contemporâneos (VGC), porconjunto de GC e situação simuladab (médias das 50 repetições).
Conjunto VGV VGCde GC ST1 ST2 a ST4 ST1 e ST2 ST3 ST41-40 -0,09 (120,58) 5,36 (102,06) 172,89 (292,50) 197,27 (68,16) 148,37 (64,65)41-80 -0,07 (119,85) 3,07 (105,01) 172,73 (303,08) 181,97 (9,66) 163,30 (9,13)81-120 -0,12 (118,71) -0,13 (105,79) 172,64 (305,74) 172,55 (7,24) 172,55 (7,24)121-160 -0,03 (119,52) -3,20 (104,73) 172,50 (315,48) 163,30 (9,13) 181,97 (9,66)161-200 -0,15 (119,60) -5,52 (101,73) 172,70 (318,77) 148,37 (64,65) 197,27 (68,16)
Geral -0,09 (119,78) -0,08 (119,60) 172,69 (307,44) 172,69 (307,44) 172,69 (307,44)a Valores entre parênteses.b ST1: ausência de diferenças genéticas entre GCs e de correlação entre os efeitos de touro e de GC.
ST2: diferenças genéticas entre GCs e ausência de correlação entre os efeitos de touro e de GC.ST3: diferenças genéticas entre GCs e correlação positiva entre os efeitos de touro e de GC.ST4: diferenças genéticas entre GCs e correlação negativa entre os efeitos de touro e de GC.

16
2.3.2 - Soluções para o efeito de grupo de contemporâneos
São apresentados, na Tabela 2.5, o viés e o QME das soluções para os efeitos
de GC (EGC) fornecidas pelos diferentes modelos. Observou-se que o modelo M1
apresentou EGC viesado para as situações em que houve diferenças genéticas entre
GCs (ST2 a ST4). GCs com animais superiores geneticamente tiveram seus efeitos
superestimados (e vice versa), de acordo com as considerações de HARVEY (1979) e
FRIES & SCHENKEL (1993), de que as soluções para os efeitos fixos fornecidas pelas
EMMH podem estar contaminadas por efeitos genéticos. É interessante notar que
quando todos os GCs foram considerados (viés geral) o viés não foi observado.
O viés e o QME das soluções de M1 foram os mesmos para as situações ST2,
ST3 e ST4, evidenciando que o problema está associado à existência de diferenças
genéticas entre GCs, e não em razão da covariância genética ambiental. Mesmo
quando rG,E foi igual a zero, a soluções das EMMH foram viesadas (soluções do M1 na
ST2).
Considerar GC como efeito aleatório (modelo M2) resultou em pequena redução
do viés em comparação com o modelo M1, na ST2. A redução do QME também foi
pequena, em ST1 e ST2. Estes resultados estão relacionados ao tamanho dos GCs,
que em média apresentaram 25 animais, e ao valor relativamente pequeno da razão
entre as variâncias residual e de GC (HENDERSON, 1973). Na situação ST3, em
concordância com UGARTE et al. (1992) e VISSCHER & GODDARD (1993), M2
apresentou melhores resultados do que o modelo M1 com relação ao viés e ao QME.
As soluções regredidas do efeito de GC, obtidas ao considerá-lo como efeito aleatório
(M2), foram menos contaminadas pelos valores genéticos em comparação com M1. A
mesma redução no viés não foi observada em ST4. Nesta situação, o viés e o QME
foram maiores no modelo M2 do que no M1, assim como observado por VISSCHER &
GODDARD (1993). Portanto, parece que a decisão de tratar o efeito de GC como fixo
ou aleatório não depende apenas do tamanho do GC e do valor da razão entre as
variâncias residual e de GC, mas também da direção de uma possível covariância
genética ambiental.
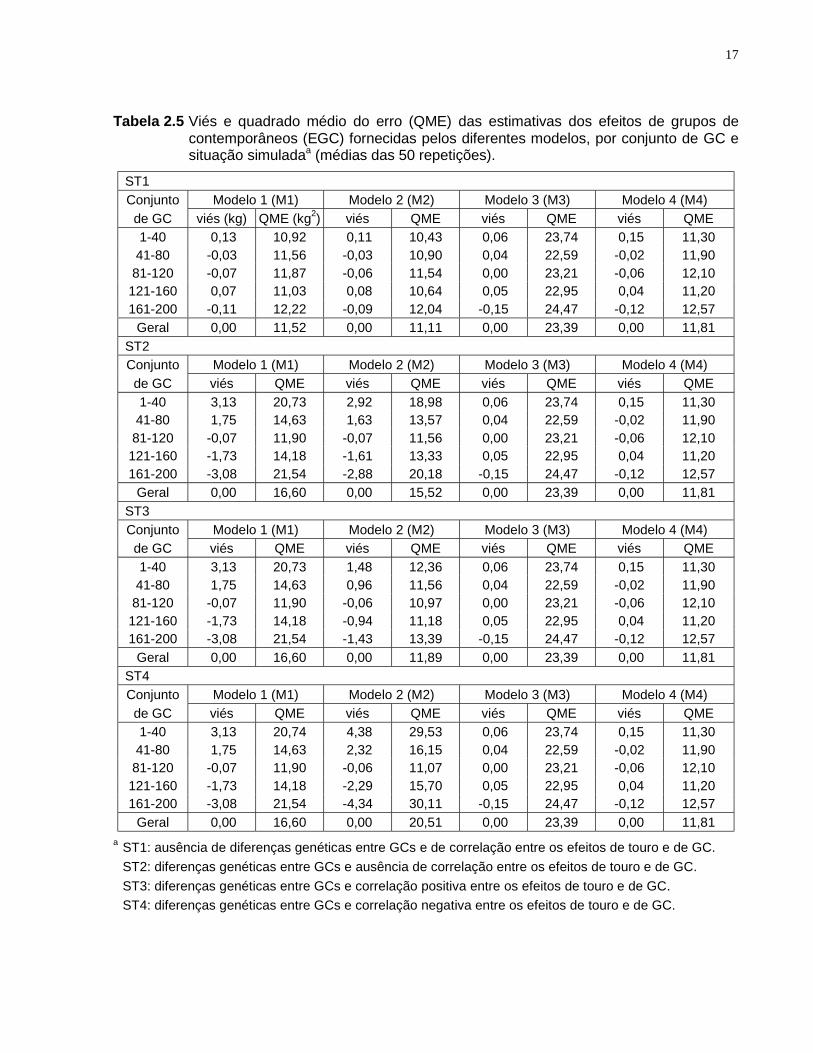
17
Tabela 2.5 Viés e quadrado médio do erro (QME) das estimativas dos efeitos de grupos decontemporâneos (EGC) fornecidas pelos diferentes modelos, por conjunto de GC esituação simuladaa (médias das 50 repetições).
ST1Conjunto Modelo 1 (M1) Modelo 2 (M2) Modelo 3 (M3) Modelo 4 (M4)de GC viés (kg) QME (kg2) viés QME viés QME viés QME1-40 0,13 10,92 0,11 10,43 0,06 23,74 0,15 11,3041-80 -0,03 11,56 -0,03 10,90 0,04 22,59 -0,02 11,9081-120 -0,07 11,87 -0,06 11,54 0,00 23,21 -0,06 12,10121-160 0,07 11,03 0,08 10,64 0,05 22,95 0,04 11,20161-200 -0,11 12,22 -0,09 12,04 -0,15 24,47 -0,12 12,57
Geral 0,00 11,52 0,00 11,11 0,00 23,39 0,00 11,81ST2Conjunto Modelo 1 (M1) Modelo 2 (M2) Modelo 3 (M3) Modelo 4 (M4)de GC viés QME viés QME viés QME viés QME1-40 3,13 20,73 2,92 18,98 0,06 23,74 0,15 11,3041-80 1,75 14,63 1,63 13,57 0,04 22,59 -0,02 11,9081-120 -0,07 11,90 -0,07 11,56 0,00 23,21 -0,06 12,10121-160 -1,73 14,18 -1,61 13,33 0,05 22,95 0,04 11,20161-200 -3,08 21,54 -2,88 20,18 -0,15 24,47 -0,12 12,57
Geral 0,00 16,60 0,00 15,52 0,00 23,39 0,00 11,81ST3Conjunto Modelo 1 (M1) Modelo 2 (M2) Modelo 3 (M3) Modelo 4 (M4)de GC viés QME viés QME viés QME viés QME1-40 3,13 20,73 1,48 12,36 0,06 23,74 0,15 11,3041-80 1,75 14,63 0,96 11,56 0,04 22,59 -0,02 11,9081-120 -0,07 11,90 -0,06 10,97 0,00 23,21 -0,06 12,10121-160 -1,73 14,18 -0,94 11,18 0,05 22,95 0,04 11,20161-200 -3,08 21,54 -1,43 13,39 -0,15 24,47 -0,12 12,57
Geral 0,00 16,60 0,00 11,89 0,00 23,39 0,00 11,81ST4Conjunto Modelo 1 (M1) Modelo 2 (M2) Modelo 3 (M3) Modelo 4 (M4)de GC viés QME viés QME viés QME viés QME1-40 3,13 20,74 4,38 29,53 0,06 23,74 0,15 11,3041-80 1,75 14,63 2,32 16,15 0,04 22,59 -0,02 11,9081-120 -0,07 11,90 -0,06 11,07 0,00 23,21 -0,06 12,10121-160 -1,73 14,18 -2,29 15,70 0,05 22,95 0,04 11,20161-200 -3,08 21,54 -4,34 30,11 -0,15 24,47 -0,12 12,57
Geral 0,00 16,60 0,00 20,51 0,00 23,39 0,00 11,81a ST1: ausência de diferenças genéticas entre GCs e de correlação entre os efeitos de touro e de GC.
ST2: diferenças genéticas entre GCs e ausência de correlação entre os efeitos de touro e de GC.ST3: diferenças genéticas entre GCs e correlação positiva entre os efeitos de touro e de GC.ST4: diferenças genéticas entre GCs e correlação negativa entre os efeitos de touro e de GC.

18
Seguindo as considerações de HARVEY (1979) e FRIES & SCHENKEL (1993),
as soluções do efeito de GC fornecidas pelo modelo M3 foram não viesadas em todas
as situações simuladas (ST1 a ST4). Entretanto, como esperado (HENDERSON, 1978),
M3 apresentou maior QME do que M1, em decorrência da maior variância do erro de
predição (VEP) (VEP = QME - viés2). Note que as soluções do M3 foram obtidas por
OLS como primeiro passo e que as vacas apresentaram apenas dois produtos, fator
este que contribuiu para os resultados observados. O VEP de soluções por OLS
também pode ser altamente influenciado pelo grau de desbalanceamento dos dados. O
viés e o QME do M3 foram idênticos para todas as situações (ST1 a ST4), isto é, seus
valores independeram da presença de diferenças genéticas entre GCs e de correlação
genética ambiental.
Similarmente ao modelo M3, os resultados do modelo M4 com respeito ao viés e
ao QME foram os mesmos para todas as situações simuladas. Além de não viesadas,
as soluções do modelo M4 apresentaram QME semelhante ao do modelo M1 na
situação ST1. Para casos em que as fêmeas apresentam número reduzido de progênie
e não são a principal causa da existência de diferenças genéticas entre GCs (como, por
exemplo, para espécies em que há pouca oportunidade para a seleção operar sobre as
fêmeas), o uso do modelo M4 parece ser um comprometimento entre remover o viés
sem que ocorra aumento expressivo do VEP. Com relação ao viés e ao QME, M4
apresentou melhores resultados do que o modelo M1 nas situações ST2 a ST4. Mesmo
quando a pressuposição usual requerida pelas EMMH para fornecer estimativas não
viesadas foi atendida (ST1), M4 não apresentou resultados inferiores a M1.
2.3.3 - Soluções para touros
A Tabela 2.6 apresenta os valores de viés e QME dos VGEs dos touros para
cada situação (ST1 a ST4) e modelo (M1 a M4). Correlações de Spearman (CORR)
entre VGV e VGE também são apresentadas. Foram obtidos resultados gerais e por
conjunto de touros.

19
Tabela 2.6 Viés (kg) e quadrado médio do erro (QME) (kg2) dos valores genéticos esperadosdos touros (VGE), e correlação de Spearman entre VGE e o valor genéticoverdadeiro (CORR), para cada modelo, conjunto de touros e situação simuladaa
(médias das 50 repetições).
ST1Conjunto Modelo 1 (M1) Modelo 2 (M2) Modelo 3 (M3) Modelo 4 (M4)de touros viés QME CORR viés QME CORR viés QME CORR viés QME CORR
1-50 0,06 39,72 0,79 0,07 39,75 0,79 -0,06 72,27 0,67 0,02 40,45 0,7951-100 0,12 41,20 0,78 0,13 41,13 0,78 0,20 69,12 0,68 0,12 41,60 0,78101-150 0,05 38,92 0,80 0,05 38,85 0,80 0,01 69,81 0,67 0,04 39,23 0,80151-200 0,08 39,35 0,80 0,08 39,23 0,80 0,01 67,17 0,68 0,10 39,42 0,80201-250 0,35 39,57 0,80 0,33 39,58 0,80 0,49 67,41 0,69 0,37 39,75 0,80
Geral 0,13 39,75 0,80 0,13 39,71 0,80 0,13 69,16 0,69 0,13 40,09 0,80ST2Conjunto Modelo 1 (M1) Modelo 2 (M2) Modelo 3 (M3) Modelo 4 (M4)de touros viés QME CORR viés QME CORR viés QME CORR viés QME CORR
1-50 -8,06 95,55 0,51 -7,84 92,18 0,51 -4,55 85,52 0,38 -4,46 53,45 0,5151-100 -3,59 40,53 0,21 -3,46 39,58 0,22 -1,41 58,99 0,18 -1,49 32,35 0,21101-150 0,00 26,51 0,22 0,00 26,46 0,22 -0,05 58,91 0,16 -0,02 28,92 0,22151-200 3,94 41,72 0,25 3,81 40,63 0,25 1,76 59,18 0,19 1,85 32,08 0,25201-250 8,38 99,73 0,51 8,16 96,12 0,51 4,92 83,76 0,40 4,80 54,72 0,51
Geral 0,13 60,81 0,65 0,13 59,00 0,66 0,13 69,27 0,69 0,13 40,30 0,80ST3Conjunto Modelo 1 (M1) Modelo 2 (M2) Modelo 3 (M3) Modelo 4 (M4)de touros viés QME CORR viés QME CORR viés QME CORR viés QME CORR
1-50 -8,05 95,55 0,51 -6,40 71,68 0,51 -4,55 85,50 0,38 -4,46 53,44 0,5151-100 -3,59 40,53 0,21 -2,60 34,17 0,21 -1,41 58,99 0,18 -1,49 32,35 0,21101-150 0,00 26,51 0,22 -0,01 26,32 0,22 -0,05 58,91 0,16 -0,02 28,92 0,22151-200 3,94 41,72 0,25 2,96 34,83 0,25 1,75 59,17 0,19 1,84 32,08 0,25201-250 8,38 99,73 0,51 6,72 74,72 0,51 4,91 83,74 0,40 4,80 54,71 0,51
Geral 0,13 60,81 0,65 0,13 48,35 0,74 0,13 69,26 0,69 0,13 40,30 0,80ST4Conjunto Modelo 1 (M1) Modelo 2 (M2) Modelo 3 (M3) Modelo 4 (M4)de touros viés QME CORR viés QME CORR viés QME CORR viés QME CORR
1-50 -8,06 95,57 0,51 -9,29 116,97 0,51 -4,55 85,54 0,38 -4,46 53,46 0,5151-100 -3,59 40,53 0,21 -4,33 46,53 0,21 -1,41 58,99 0,18 -1,49 32,36 0,21101-150 0,00 26,51 0,22 0,00 26,66 0,22 -0,05 58,91 0,16 -0,02 28,92 0,22151-200 3,94 41,72 0,25 4,67 48,08 0,25 1,76 59,18 0,19 1,85 32,08 0,25201-250 8,38 99,75 0,51 9,62 122,19 0,51 4,92 83,78 0,40 4,80 54,73 0,51
Geral 0,13 60,82 0,65 0,13 72,08 0,56 0,13 69,28 0,69 0,13 40,31 0,80a ST1: ausência de diferenças genéticas entre GCs e de correlação entre os efeitos de touro e de GC.
ST2: diferenças genéticas entre GCs e ausência de correlação entre os efeitos de touro e de GC.ST3: diferenças genéticas entre GCs e correlação positiva entre os efeitos de touro e de GC.ST4: diferenças genéticas entre GCs e correlação negativa entre os efeitos de touro e de GC.

20
Em ST1, os VGEs fornecidos por todos os modelos estudados foram não
viesados. O modelo M3 apresentou maiores QME e menores CORR. Os demais
modelos apresentaram QME e CORR semelhantes entre si.
Nas situações ST2 a ST4, todos os modelos apresentaram soluções viesadas.
O viés dos modelos M3 e M4 está relacionado ao uso do fator de regressão (como
segundo passo) para obtenção das soluções, fazendo com que os touros extremos
tivessem seus VGEs proporcionalmente mais regredidos em relação a zero do que os
touros intermediários. M1 apresentou viés adicional pois, como mencionado
anteriormente, as soluções para os efeitos de GC deste modelo foram contaminadas
por efeitos genéticos. Os touros superiores tiveram seus VGEs subestimados porque
parte de sua superioridade foi estimada como efeito de GC, sendo que o oposto
ocorreu para os touros inferiores.
Ao considerar GC como efeito aleatório (M2), funções dos efeitos de GC são
incorporadas nos VGEs (VAN VLECK, 1987). Assim como em VISSCHER &
GODDARD (1993), a conseqüência deste confundimento foi que os resultados de M2
foram, em comparação com M1, favoráveis (menor viés e QME e maior CORR) nas
situações ST2 e ST3 e desfavoráveis em ST4. Os valores das correlações genética
ambiental nas situações ST2, ST3 e ST4 explicam estes resultados. Na situação ST4,
por exemplo, em que os melhores touros estão associados aos piores GCs, as funções
dos efeitos de GC incorporadas nos VGEs, ao considerar GC como aleatório, fizeram
com que os VGEs dos melhores touros fossem subestimados de forma mais acentuada,
e vice versa.
Em todas as situações estudadas, o modelo M3 apresentou menores CORR por
conjunto de touros do que o modelo M1, em decorrência da maior VEP. Entretanto, é
interessante notar que, nas situações ST2 a ST4, M3 apresentou maior CORR geral do
que M1, em decorrência do confundimento entre os efeitos de touro e de GC nas
soluções fornecidas por M1. Semelhantemente aos resultados de HARVEY (1979), a
CORR geral do modelo M3 permaneceu a mesma para as diferentes situações,
enquanto que a CORR geral do modelo M1 foi maior em ST1 do que nas situações ST2
a ST4.

21
Diferenças genéticas entre GCs (ST2 a ST4) reduziram a habilidade de M4 em
classificar apropriadamente os animais por conjunto de touros, assim como para os
outros modelos, pois touros de mérito genético similar tenderam a ser comparados mais
freqüentemente entre eles. Contudo, a CORR geral do modelo M4 permaneceu
constante (0,8) para todas as situações, e foi superior àquelas obtidas pelos demais
modelos nas situações ST2 a ST4.
Na tentativa de mensurar a eficiência de cada modelo em predizer a
superioridade genética nas diferentes situações, foram calculadas as médias dos VGVs
de touros “top” selecionados conforme os VGEs fornecidos pelos diferentes modelos
(Tabela 2.7). Três intensidades de seleção foram aplicadas: 50% (top50), 20% (top20) e
2% (top2) dos melhores touros. Na última coluna da Tabela 2.7 são apresentadas as
médias dos VGVs dos touros “top” numa situação hipotética em que estes seriam
selecionados de acordo com o VGV, apenas para avaliar quão distante cada modelo
está desta situação ideal.
Para os modelos M1 e M2, as diferenças genéticas entre GCs tiveram,
proporcionalmente, maior impacto sobre o VGV médio de touros selecionados com
intensidade moderada (20%) e baixa (50%). No M1, o VGV médio dos top20 e top50 foi
menor para as situações ST2 a ST4 do que para a ST1 (p<0,05). O mesmo foi
observado para o modelo M2 (p<0,05). A habilidade de M3 e M4 em selecionar touros
não foi afetada pela situação simulada, isto é, os resultados de ST1 e ST2 a ST4 não
diferiram (p>0,05). Considerando as situações ST2 a ST4, o modelo M1 apresentou
maior VGV médio do que M3 para os top2, mas menor para os top50, e o modelo M4
apresentou maior VGV médio do que M1 para todas as intensidades de seleção
(p<0,05). Contrastando os modelos M4 e M2, M4 foi superior a M2 para os top50 e
top20 na situação ST3 e para todas as intensidades de seleção na situação ST4
(p<0,05).

22
Tabela 2.7 Médias (kg) dos valores genéticos verdadeiros dos touros (VGV) selecionadosconforme as soluções de cada modelo, para diferentes intensidades de seleção esituações simuladasa (médias das 50 repetições).
N.º de touros Modelo 1 (M1) Modelo 2 (M2) Modelo 3 (M3) Modelo 4 (M4) VGVSelecionados ST1 ST2-ST4 ST1 ST2 ST3 ST4 ST1 ST2-ST4 ST1 ST2-ST4 ST1-ST4
125 (50%) 6,92 5,61 6,90 5,73 6,40 4,84 5,99 5,99 6,93 6,92 8,5250 (20%) 12,28 10,37 12,29 10,53 11,47 9,23 10,68 10,60 12,28 12,28 15,055 (2%) 20,73 20,64 20,68 20,59 21,11 20,26 18,01 18,73 20,94 21,29 25,84
a ST1: ausência de diferenças genéticas entre GCs e de correlação entre os efeitos de touro e de GC.ST2: diferenças genéticas entre GCs e ausência de correlação entre os efeitos de touro e de GC.ST3: diferenças genéticas entre GCs e correlação positiva entre os efeitos de touro e de GC.ST4: diferenças genéticas entre GCs e correlação negativa entre os efeitos de touro e de GC.
Conforme os resultados apresentados, é possível notar que a existência de
heterogeneidade de variância ambiental entre os conjuntos de GC nas situações ST3 e
ST4 não afetou as soluções dos modelos que assumiram GC como fixo (M1, M2 e M3),
pois estes apresentaram resultados iguais para as situações ST3 e ST4 em
comparação com a situação ST2 - caracterizada pela ausência de heterogeneidade de
variância ambiental.
As comparações para as soluções de vacas não foram apresentadas porque
todos os modelos tiveram resultados semelhantes independente da situação simulada,
como esperado, uma vez que as vacas não eram fonte da existência de diferenças
genéticas entre GCs.
Os VGEs dos produtos foram obtidos por retro-substituição (HENDERSON,
1984). Considerando o modelo M1 e as situações ST2 a ST4, os VGEs dos produtos
não foram tão afetados pelo viés quanto os VGEs dos touros (dados não
apresentados). Este resultado é explicado pela existência de viés em direção oposta
para as soluções de touro e de GC, que tenderam a se cancelar ao estimar o efeito da
Segregação Mendeliana.

23
2.3.4 - Discussão geral
Propriedades empíricas das soluções das equações de modelo misto de
Henderson foram investigadas para diferentes situações de associação entre touros e
GCs. As situações ST2 a ST4 são casos de “seleção do tipo L’u” de acordo com a
terminologia adotada por HENDERSON (1973), pois os touros tiveram seus produtos
alocados nos GCs com base em informações “externas” (conhecimento a priori do VGV
dos touros). Segundo o autor, se a associação entre touros e GCs ocorresse com base
no BLUP, em vez do VGV, as soluções do M1 seriam não viesadas.
Na prática, diferenças genéticas entre GCs podem ser geradas como resultado
de tomadas de decisão dos produtores (ex.: quais touros usar e como alocar sua
progênie em diferentes grupos de manejo). Normalmente, estas decisões não são
tomadas apenas com base no BLUP dos valores genéticos. HENDERSON (1975, 1990)
sugeriu que o estudo de situações de “seleção do tipo L’u” poderia ser útil para
representar casos reais comuns como, por exemplo, diferenças genéticas sendo
criadas com base em informações prévias ou em variáveis correlacionadas com o VGV
e não disponíveis ou utilizadas na análise. Nestas condições, o uso do modelo M1
resultou em soluções viesadas e, como conseqüência, em redução da habilidade em
classificar os animais apropriadamente.
HENDERSON (1973) mencionou que embora seja lógico considerar rebanho
como efeito aleatório, considerá-lo como fixo elimina o viés causado por diferenças no
mérito de rebanhos associados à determinados touros. Os resultados do presente
estudo não estão em desacordo com esta afirmação, pois HENDERSON (1973) não se
referiu à situações como a ST3 ou a ST4 simuladas no presente estudo, mas sim à
situação em que a média dos efeitos de GC difere entre os touros, mas não há
diferenças genéticas entre GCs.
Ao expressar o melhor preditor linear não viesado (BLUP) dos valores genéticos
(û) desenvolvido por HENDERSON (1949, 1963) na forma )ˆXy(V'GZu 1 ββββ−−−−==== −−−− , talvez
torna-se mais claro que o viés em ββββ pode afetar as soluções de û. Se ββββ é tratado como
fixo, HENDERSON (1973) mostrou que o valor esperado de û fornecido pelas EMMH é

24
invariante a ββββ , o que não significa necessariamente que û é livre de um possível viés
em ββββ .
Estimadores não viesados podem ser inadmissíveis sob o critério QME, isto é,
estimadores viesados com menor QME podem existir (GIANOLA, 1990). No entanto,
com relação as EMMH e as situações simuladas, o viés que afetou ββββ foi uma função
dos efeitos genéticos, o que reduziu a habilidade das EMMH em discriminar os
melhores animais, o que pode ser observado na Tabela 2.6 contrastando a CORR do
M1 na ST1 contra as demais situações (ST2 a ST4).
Viés nas soluções das EMMH também foi observado por HARVEY (1979),
SIMIANER & WOLLNY (1989), UGARTE et al. (1992), FRIES & SCHENKEL (1993),
VISSCHER & GODDARD (1993), TORRES JR. et al. (1997), e TÄUBERT et al. (2002).
SIMIANER & WOLLNY (1989) reportaram que a estratégia de tratar as filhas de
determinados touros em teste para que esses apresentassem melhores provas (VGE)
teve efeito oposto, ou seja, fez com que tivessem valores genéticos subestimados, pois
parte da superioridade genética dos touros foi estimada como efeito de tratamento.
TORRES JR. et al. (1997) simularam uma situação comparável a ST3 do
presente estudo e observaram que a presença de parentesco entre os touros reduziu,
mas não eliminou o viés das soluções fornecidas pelas EMMH.
O uso de grupos genéticos poderia ser uma alternativa para reduzir ou eliminar o
viés apresentado pelo modelo M1. Essa alternativa não foi testada no presente estudo
porque em conjuntos de dados reais dificilmente há oportunidade de definir grupos
genéticos de forma apropriada como em estudos de simulação. A inclusão de grupos
genéticos desnecessários no modelo pode levar à redução da acurácia de predição
(TORRES JR. et al., 1997).
Tratar GC como efeito aleatório (M2) parece não ser uma alternativa adequada
se há evidências da existência de covariância genética ambiental negativa. VISSCHER
& GODDARD (1993) discutiram algumas situações práticas em que os efeitos de touro
e de GC podem estar negativamente associados e salientaram que, normalmente, na
prática, a covariância entre touros e GC é meramente o resultado da escolha de

25
criadores por determinados touros, e que esta associação não deveria ser usada para
predizer a performance futura da progênie de determinado touro, uma vez que esse não
é um parâmetro populacional propriamente dito. Ao assumir GC como aleatório, em
situações de diferenças genéticas entre GCs, UGARTE et al. (1992) observaram que a
redução na VEP não acompanhou o aumento do viés para casos em que o GC era
relativamente pequeno e o valor da razão entre as variâncias residual e de GC era
baixo.
As soluções do primeiro passo dos modelos M3 e M4 podem ser vistas como
casos especiais dos modelos de seleção de Henderson para lidar com casos de
“seleção do tipo L’u” (HENDERSON, 1975), em que a matriz que descreve o processo
de seleção (L) foi considerada igual a uma matriz identidade para touros e vacas no
modelo M3, e igual a uma matriz identidade para os touros e a uma matriz nula para as
vacas no M4. Este procedimento adotado foi sugerido por HENDERSON (1973) caso L
fosse difícil de ser definida (como ocorre usualmente com dados de campo). Conforme
os resultados do presente estudo, o modelo M3 apresentou soluções não viesadas para
as diferentes situações simuladas às custas de alta variância do erro de predição das
estimativas (VEP), o que pode não ser vantajoso sob o aspecto de maximizar o
progresso genético. Por sua vez, o uso do modelo M4 parece ser um comprometimento
entre remover o viés sem que ocorra aumento expressivo da VEP, para situações em
que as fêmeas apresentam número reduzido de progênie e não são a principal causa
da existência de diferenças genéticas entre GCs.
É importante notar que a comparação entre métodos de estimação viesados e
não viesados pode ser altamente dependente do espaço paramétrico em estudo
(GIANOLA, 1990; SIMIANER, 1991), e generalizações com respeito aos resultados do
presente estudo devem ser feitas com precaução.
HENDERSON (1973, 1975) apresentou algumas situações como, por exemplo,
quando diferenças genéticas entre GCs são geradas com base em funções dos dados
invariantes a translação, em que as soluções de modelos semelhantes a M3 e M4 do
presente estudo poderiam ser viesadas, o que evidencia a importância da condução de
mais estudos nesta linha de pesquisa.

26
2.4 - ConclusõesEvidências empíricas mostraram que as soluções das equações regulares de
modelo misto de Henderson podem apresentar propriedades afetadas pela presença de
diferenças genéticas entre GCs, causadas pela associação não aleatória entre os
efeitos de touro e de GC (uma forma de “seleção do tipo L’u”), e que modelos
alternativos podem fornecer melhores resultados sob esta condição.
Os resultados sugerem que a fonte principal de um possível viés nas soluções
fornecidas pelas EMMH regulares está mais associada à presença de diferenças
genéticas entre GCs do que a existência de correlação entre os efeitos genéticos e
ambientais.
A busca de modelos que forneçam soluções com menor QME parece estar mais
associada à obtenção de critérios ótimos no sentido de permitir que a média do mérito
genético de um grupo de animais selecionados seja maximizada, do que a busca de
modelos que forneçam soluções não viesadas ou com variância do erro de predição
reduzida, isoladamente.

27
CAPÍTULO 3 - MODELOS DE AVALIAÇÃO GENÉTICA ALTERNATIVOS EMSITUAÇÕES DE SELEÇÃO PARENTAL
Resumo: Simulações estocásticas foram conduzidas com o objetivo de
investigar propriedades empíricas das equações de modelo misto de Henderson
utilizando um modelo animal (MMH_a) e de outros sete modelos de avaliação genética
alternativos, sob três tipos de seleção parental (com base no BLUP, no fenótipo e no
valor genético verdadeiro (VGV)) e duas situações de informação de parentesco
(completa e 10% da informação de parentesco referente ao touro perdida). Foram
simuladas 50 repetições de populações compostas por 12.000 animais distribuídos em
600 grupos de contemporâneos (GCs) e seis gerações. A cada geração houve
reposição de 50% dos machos e de 10% das fêmeas em reprodução. Os parâmetros
utilizados conferiram à característica hipotética simulada uma herdabilidade igual a 0,4.
Quanto ao viés das soluções dos diferentes modelos, para a situação de parentesco
completo, MMH_a apresentou soluções viesadas quando houve seleção com base no
VGV, e o viés dos modelos alternativos ocorreu principalmente, no geral, na situação de
seleção baseada no BLUP. O viés apresentado pelo modelo MMH_a ocorreu no sentido
de subestimar as soluções dos touros usados em gerações com animais de mérito
genético superior. O viés apresentado pelos modelos alternativos, com seleção
baseada no BLUP, ocorreu no sentido oposto. Informação de touro perdida fez com que
todos os modelos apresentassem viés no sentido de subestimar as soluções dos touros
usados em gerações com animais de mérito genético superior. O viés apresentado nas
diferentes situações teve conseqüências sobre a habilidade de cada modelo em
selecionar animais de mérito genético superior e em estimar a tendência genética.
Quanto à estas propriedades empíricas, evidências foram observadas de que modelos
alternativos podem apresentar resultados superiores aos do modelo MMH_a, e que
nem sempre a seleção, ou mais explicitamente, as diferenças genéticas entre níveis
dos efeitos ambientais, deveriam ser ignoradas.
Palavras-chave: avaliação genética, BLUP, efeito de seleção, predições viesadas

28
3.1 – Introdução
Há interesse, em programas de melhoramento, na predição de valores genéticos
e na utilização destas predições ou funções das mesmas como critério de seleção dos
animais. Sob normalidade e na classe de funções invariantes a translação, o melhor
preditor linear não viesado (BLUP), desenvolvido por HENDERSON (1949) e com
propriedades descritas por HENDERSON et al. (1959) e por HENDERSON (1963),
apresenta-se como critério de seleção ótimo no sentido de maximizar a probabilidade
de classificação correta dos animais quanto ao mérito genético, assim como de
maximizar a média dos valores genéticos de determinado número de animais
selecionados (HENDERSON, 1973; FERNANDO & GIANOLA, 1986).
Na derivação do BLUP, pressupõe-se que a esperança marginal dos valores
genéticos de todos os animais que estão sendo avaliados seja igual a zero (E(ai)=0).
Diferentes formas de seleção ou práticas que resultem na distribuição não aleatória dos
animais nos níveis de efeitos ambientais podem gerar diferenças genéticas entre estes
níveis e, consequentemente, invalidar a pressuposição E(ai)=0. A seleção parental, por
exemplo, se bem aplicada, faz com que animais de gerações posteriores tenham valor
genético esperado superior ao da média dos valores genéticos da população dos
animais que os geraram.
Como conseqüência da violação de E(ai)=0, as soluções das Equações de
Modelo Misto de Henderson (EMMH), que forneceriam sob condições ideais o BLUP
dos valores genéticos, não são garantidas de serem não viesadas (HARVEY, 1979;
FRIES & SCHENKEL, 1993). HENDERSON (1973 e 1975) apresentou condições
especiais em que as soluções das EMMH regulares são não viesadas mesmo que a
pressuposição E(ai)=0 seja violada. Tais condições, como, por exemplo, informação de
parentesco completa e seleção dos animais com base em funções invariantes a
translação, dificilmente são atendidas em conjuntos de dados reais oriundos de
programas de melhoramento. Para casos em que estas condições não são atendidas,
HENDERSON (1975) propôs modificações nas EMMH regulares para que o BLUP dos
valores genéticos fosse obtido. As EMMH modificadas requerem uma matriz para
descrever o processo seletivo que, no geral, não é trivial de ser construída.

29
FRIES & SCHENKEL (1993) e SCHAEFFER (2003) propuseram modelos
alternativos de avaliação genética que não requerem a pressuposição de ausência de
diferenças genéticas entre os níveis dos efeitos ambientais. Entretanto, assim como nos
modelos de seleção propostos por HENDERSON (1975), não há garantia de que os
valores genéticos preditos sejam um critério de seleção ótimo no sentido de permitir a
seleção de animais com a média dos valores genéticos maximizada.
Simulações conduzidas por CARVALHEIRO et al. (2004 – capítulo anterior)
evidenciaram que modelos alternativos baseados nas propostas de FRIES &
SCHENKEL (1993) e de SCHAEFFER (2003), de considerar os valores genéticos ou
parte deles como efeitos fixos, podem apresentar soluções com propriedades empíricas
superiores àquelas fornecidas pelas EMMH regulares, para casos em que a
pressuposição E(ai)=0 não é válida. O estudo de CARVALHEIRO et al. (2004– capítulo
anterior) se limitou, no entanto, a situações que envolviam apenas uma geração de
progênie e que apresentavam diferenças genéticas entre os níveis de efeitos
ambientais (grupos de contemporâneos (GC)) geradas a partir da associação entre os
valores genéticos dos touros e os GC.
No presente estudo de simulação, houve interesse em comparar modelos de
avaliação genética alternativos em situações de seleção parental, visto que esta é uma
prática comum e fundamental em programas de melhoramento e que não há uma
solução estatística definitiva para resolver os possíveis problemas de inferência
causados por ela.
3.2 - Material e Métodos
Na dificuldade de comparar analiticamente diferentes modelos quanto às suas
propriedades, especialmente sob violação da pressuposição E(ai)=0, optou-se pela
condução de um estudo de simulação, seguindo recomendações de HENDERSON
(1989 e 1990).

30
3.2.1 - Estrutura da população simuladaA população base (G0) simulada consistiu de 100 machos e 2.000 fêmeas, não
aparentados, não selecionados e assumidos de serem amostrados aleatoriamente de
uma população conceitualmente infinita. Cada fêmea foi acasalada aleatoriamente com
um dos machos e gerou um produto. Os 2.000 produtos gerados constituíram os
animais da geração G1 e foram distribuídos aleatoriamente em 100 grupos de
contemporâneos (GC), sendo estipulado que os animais dos primeiros 50 GCs eram
fêmeas e os demais eram machos.
Caracterizando o processo de seleção parental, 50 machos e 1.800 fêmeas da
G0 e 50 machos e 200 fêmeas da G1 foram selecionados, com base em critérios de
seleção descritos adiante, para gerar os animais da G2. Houve, portanto, sobreposição
de gerações. Este processo de reposição de 50% dos machos e de 10% das fêmeas
em reprodução se repetiu até que os animais da G6 fossem gerados. A cada período
de acasalamento, 2.000 produtos foram gerados e distribuídos aleatoriamente em 100
GCs, totalizando 12.000 animais em 600 GCs.
A conectabilidade entre os GCs foi garantida pela imposição de restrições sobre
o valor mínimo para número de produtos por reprodutor (5), produtos por GC (10),
reprodutor por GC (3) e GC por reprodutor (3), e também por meio da aplicação do
programa MILC (FRIES, 1998). Sempre uma nova repetição foi simulada caso algum
GC estivesse desconectado.
3.2.2 - Modelo de simulaçãoObservações fenotípicas foram simuladas, exceto para os animais da geração
G0, seguindo o modelo: yij = bi + aij + eij , sendo yij a observação fenotípica do animal j
no GC i, bi o efeito do GC i, aij o valor genético aditivo do animal j no GC i e eij o termo
referente ao erro aleatório associado a observação yij.
Valores para bi foram amostrados de uma distribuição uniforme com parâmetros
(valores de mínimo e máximo) iguais a 122,5 e 222,5 kg. Valores para eij foram
independentemente amostrados de uma distribuição normal com média zero e variância
de 180 kg2 (eij ~N(0;180)). Os valores genéticos dos animais base foram gerados a

31
partir de uma N(0;120), e os valores genéticos dos demais animais foram gerados como
aij = ½( asj + adj) + mij, em que asj e adj são os valores genéticos aditivos do macho e da
fêmea que geraram o animal j, e mij é o efeito da Segregação Mendeliana do animal j,
que foi amostrada de uma N(0;60).
Os parâmetros usados no processo de simulação reproduziram uma
característica com herdabilidade igual a 0,4, e são semelhantes aos estimados por
CARVALHEIRO (2000) para a característica ganho de peso pós-desmame de um
conjunto de dados de bovinos da raça Nelore.
3.2.3 - Critérios de seleção
Na prática, inúmeras são as fontes de informação que podem ser utilizadas como
critério de seleção dos animais e que, se correlacionadas com o valor genético, podem
gerar diferenças genéticas entre anos/gerações/períodos consecutivos de seleção. Na
deficiência e impossibilidade de simular todas as situações práticas possíveis, três
casos isolados foram escolhidos de forma a gerar diferenças genéticas. Os três critérios
de seleção parental empregados a cada período de acasalamento foram: (1) o BLUP
dos valores genéticos, obtido com o emprego das EMMH regulares a cada geração; (2)
as observações fenotípicas; e (3) os valores genéticos verdadeiros (VGV).
De acordo com a terminologia adotada por HENDERSON (1973 e 1975), as
situações de (1) a (3) são, respectivamente, casos de: (1) “seleção do tipo L’y” com
L’X=0, em que L é a matriz que descreve o processo de seleção e L’X=0 caracteriza a
condição de seleção invariante a translação ou, equivalentemente, invariante aos
valores assumidos pelos efeitos ambientais (ββββ) (no Apêndice 5, encontra-se uma prova
da equivalência entre “seleção do tipo L’y” com L’X=0 e a seleção com base no BLUP);
(2) “seleção do tipo L’y” com L’X≠≠≠≠0, isto é, seleção com base em funções de y
(observações fenotípicas) que não são invariantes a ββββ; e (3) “seleção do tipo L’u”, em
que u é o valor genético verdadeiro. Embora, na prática, não seja possível selecionar
os animais com base no valor genético verdadeiro, HENDERSON (1975 e 1990)
sugeriu que o estudo desta situação poderia ser útil para representar casos reais

32
comuns como, por exemplo, seleção com base em informações prévias ou em variáveis
correlacionadas com o VGV e não disponíveis ou não utilizadas na análise.
As fêmeas de descarte, 10% das fêmeas em reprodução, foram sempre
escolhidas aleatoriamente. As fêmeas de reposição foram selecionadas com base nos
critérios descritos anteriormente, de acordo com cada uma das três situações
simuladas. Os machos, tanto os de descarte quanto os de reposição, foram sempre
escolhidos com base nos critérios de seleção. Quando o critério de seleção adotado foi
o fenótipo, os machos da geração G0, por não apresentarem essa informação, foram
selecionados com base em uma variável aleatória com correlação de 0,4 ao VGV.
3.2.4 - Informações adicionais do processo de simulação
Com relação à informação de parentesco, duas situações foram simuladas: (1)
informação de parentesco completa; e (2) 10% dos animais da G2 a G6 (1.000 animais)
com informação de progenitor (pai) perdida. Houve interesse em simular esta segunda
situação, pois é comum, em conjuntos de dados reais, que parte dos animais não tenha
informação de parentesco conhecida devido, por exemplo, ao uso de lotes com mais de
um reprodutor.
Para cada uma das seis situações simuladas, compreendidas pela combinação
entre os três critérios de seleção e os dois tipos de informação de parentesco utilizados,
50 repetições foram geradas, resultando em um total de 300 conjuntos de dados.
Caracterizando a condição de efeito fixo, conforme a escola estatística
frequentista, os valores dos efeitos de GC (bi) gerados na primeira repetição foram
mantidos nas demais repetições.
3.2.5 - Modelos de avaliação genética
Os componentes de variância utilizados nas avaliações genéticas foram iguais
àqueles empregados no processo de simulação dos dados, ou seja, eles não foram
estimados. Portanto, o termo avaliação genética se refere apenas à estimação dos
efeitos de GC e à predição dos valores genéticos.

33
Para cada um dos 300 conjuntos de dados, avaliações genéticas foram
conduzidas sob sete modelos diferentes, que podem ser, em geral, descritos pela
equação:
y = Xββββ + Zss + Zdd + m + e ,
em que: y é o vetor de observações fenotípicas, ββββ é o vetor de efeitos de GC, s e d são,
respectivamente, os vetores de efeito de progenitores e progenitoras (que serão
mencionados como efeitos de touro e vaca), m é o vetor referente aos efeitos de
Segregação Mendeliana, e é o vetor de erros aleatórios, X, Zs e Zd são as matrizes de
incidência que relacionam os elementos de ββββ, s e d com os elementos de y,
respectivamente. As pressuposições, e esperanças e (co)variâncias das variáveis
aleatórias de cada modelo são descritas a seguir.
3.2.5.1 - Modelo Misto de Henderson – Modelo Animal individual (MMH_a)
Modelo predominantemente aplicado em programas de melhoramento animal,
em que ββββ é considerado como vetor de efeitos fixos; Za = Zss + Zdd + m, sendo a ovetor aleatório de efeito genético aditivo de cada animal e Z a matriz que associa os
elementos de a com os de y. As matrizes de esperança e (co)variâncias são dadas por:
E(y) = Xββββ, E(a) = E(e) = 0, Var(y) = V = 2en
2a I'ZAZ σσσσ++++σσσσ , Var(a) = 2
aAσσσσ e Var(e) = 2enI σσσσ ,
em que A é a matriz de numeradores do coeficiente de parentesco genético aditivo
(WRIGHT, 1922), nI é uma matriz identidade de ordem n (número de animais em y),2aσσσσ é a variância genética aditiva na população base e 2
eσσσσ é a variância residual.
Soluções para ββββ e a foram obtidas com o emprego das EMMH regulares
(HENDERSON, 1963).
3.2.5.2 - Modelo Alternativo 1 – Modelo Touro e Vaca (MA1_tv)
A derivação do BLUP pressupondo E(a)≠0 resulta em soluções para ββββ e a sendo
obtidas pelo método de quadrados mínimos ponderados (FRIES & SCHENKEL, 1993;
SCHENKEL, 1998) ou pelo método de quadrados mínimos ordinários (OLS) se
homogeneidade de variância residual for assumida. Neste caso, ββββ e a seriam tratados

34
como efeitos fixos e, no geral, não haveria possibilidade de aplicação do modelo animal
devido à deficiência de posto da matriz de coeficientes. Uma alternativa, que foi
empregada no presente estudo, seria a utilização de um modelo touro e vaca,
considerando-os como efeitos fixos, e somando os vetores m e e para compor o vetor
aleatório de resíduos (εεεε). Neste modelo, assumiu-se: E(y) = Xββββ + Zss + Zdd, E(εεεε) = 0, e
Var(y) = Var(εεεε) = 2nI εεεεσσσσ . As soluções para ββββ, s, e d foram obtidas por OLS, impondo-se as
restrições de que a soma dos elementos de s e d fossem iguais a zero.
Os valores genéticos preditos (â) dos touros e vacas foram obtidos multiplicando-
se s e d por dois, e para os produtos (animais sem progênie), utilizou-se a expressão
â= s+ d+m , sendo:
com k=0,5 2aσσσσ /(0,5 2
aσσσσ + 2eσσσσ ) se a informação de parentesco fosse completa, e
k=0,75 2aσσσσ /(0,75 2
aσσσσ + 2eσσσσ ) se a informação de touro fosse perdida.
3.2.5.3 - Modelo Alternativo 2 – Modelo Touro e Vaca (MA2_tv)
Como as fêmeas apresentam, normalmente, número pequeno de descendentes,
considerá-las como efeito fixo pode resultar em soluções com elevado erro de predição
(CARVALHEIRO et al., 2004– capítulo anterior). Assim, um outro modelo alternativo
investigado considerou ββββ e s como vetores de efeitos fixos e d como vetor de efeito
aleatório, com E(y) = Xββββ + Zss, E(d) = E(εεεε) = 0, Var(y) = 2n
2ddd I'ZZ εεεεσσσσ++++σσσσ , Var(d) = 2
ddI σσσσ ,
e Var(εεεε) = 2nI εεεεσσσσ . As soluções foram obtidas conforme equações semelhantes às de OLS,
porém adicionando o termo ( 2εεεεσσσσ / 2
dσσσσ ) aos elementos da diagonal da matriz de
coeficientes correspondentes ao efeito de vaca. A restrição de que a soma dos
elementos em s seja igual a zero foi imposta. Os valores genéticos preditos (â) foram
obtidos como em MA1_tv.
Além de procurar não obter soluções com elevado erro de predição, o uso de
vaca como efeito aleatório também se justificaria pelo fato de, no geral, haver baixa
, k*)dZsZˆXy(m ds −−−−−−−−ββββ−−−−====

35
pressão de seleção sobre as fêmeas em condições reais, o que faz com que elas não
sejam a principal causa da existência de diferenças genéticas.
Conforme salientado por FRIES (1999), as soluções para s e d de um modelo
touro e vaca levam em consideração apenas a informação da progênie, ignorando a
informação própria e a dos pais. Com isso, as principais fontes de informação para
animais com tamanho reduzido de progênie e com informação própria não são
utilizadas na predição de seus valores genéticos. Procurando aproximar os dois
modelos alternativos descritos anteriormente (MA1_tv e MA2_tv) a um modelo animal,
para que as diferentes fontes de informação fossem utilizadas na predição dos valores
genéticos, quatro outros “modelos” alternativos (não são modelos em si, mas o foram
denominados como tal) foram empregados e são descritos a seguir.
3.2.5.4 - Modelo Alternativo 3 – Modelo “Animal” (MA3_a)
Os valores genéticos preditos (â) foram obtidos por: â= 2aAσσσσ Z’V-1(y- *ˆXββββ ), sendo
*ββββ as soluções para os efeitos de GC fornecidas pelo modelo MA1_tv.
A motivação para o uso deste modelo foi que, ao contrário das soluções
fornecidas pelas EMMH regulares para os efeitos de GC, os valores de *ββββ não são
correlacionados ou possivelmente contaminados pelos valores genéticos verdadeiros
(Cov( *ββββ ,a)=0) (FRIES & SCHENKEL, 1993).
3.3.5.5 - Modelo Alternativo 4 – Modelo “Animal” (MA4_a)
Os valores genéticos preditos (â) foram obtidos por: â= 2aAσσσσ Z’V-1(y- oˆXββββ ), sendo
oββββ as soluções para os efeitos de GC fornecidas pelo modelo MA2_tv.
3.2.5.6 - Modelo Alternativo 5 – Combinando Soluções (MA5_c)
Conforme descrito em VANRADEN & WIGGANS (1991), o â fornecido pelas
EMMH regulares pode ser decomposto em: â = w1(informação própria) +
w2(informação dos pais) + w3(informação da progênie), sendo w1=n/(n+2k+0,5pk),

36
w2=2k/(n+2k+0,5pk) e w3=0,5pk/(n+2k+0,5pk), em que n é o número de observações
fenotípicas do animal, k= 2eσσσσ / 2
aσσσσ e p é o tamanho da progênie. SCHAEFFER (2003)
propôs o uso destes fatores de ponderação para combinar as duas soluções que um
animal com informação própria e da progênie pode apresentar quando o modelo touro e
vaca é empregado. Este procedimento foi adotado considerando as soluções â, s e dfornecidas pelo modelo MA1_tv. Assim, os valores genéticos preditos dos animais com
progênie, combinando as diferentes fontes de informação (âc) foram obtidos por:
âc=(((w1+w2)â)+(w3 s ))/(w1+w2+w3), caso o animal fosse touro (exceto touros da G0),
e âc=(((w1+w2)â)+(w3 d))/(w1+w2+w3), caso o animal fosse vaca (exceto vacas da
G0).
3.2.5.7 - Modelo Alternativo 6 – Combinando Soluções (MA6_c)
Similar ao procedimento descrito anteriormente (MA5_c), porém utilizando as
soluções â, s e d do modelo MA2_tv, para obter âc.
3.2.6 - Critérios de comparação dos modelos
Os modelos foram comparados com relação ao viés das soluções, definido por:
viésw = 1/q∑∑∑∑====
−−−−q
1iii )ww( , em que wi é o efeito verdadeiro de GC (VGC) ou valor genético
verdadeiro (VGV), iw é o efeito estimado de GC (EGC) ou o valor genético predito
(VGE), e q é o número de níveis do efeito em consideração.
Na tentativa de mensurar a eficiência de cada modelo em predizer a
superioridade genética, também foram calculadas as médias dos VGVs de animais
selecionados sob diferentes pressões de seleção e com base nas soluções fornecidas
pelos diferentes modelos. Para este critério de comparação, testes-t pareados, com um
nível de significância de 5%, foram conduzidos contrastando todos os modelos, um por
vez, com o modelo MMH_a.
Os modelos também foram avaliados quanto à capacidade de predição da
tendência genética, calculando-se a média dos valores genéticos preditos, por geração.

37
Todos os critérios de comparação foram tomados com base na média dos
valores das 50 repetições.
3.3 - Resultados e Discussão
O problema de violação da pressuposição E(ai)=0 é geralmente tratado na
literatura como um problema de efeito de seleção. Entretanto, cabe salientar que forças
ou práticas de manejo dos animais que geram diferenças genéticas entre níveis dos
efeitos ambientais, entre GCs, por exemplo, estarão atuando no sentido de invalidar a
pressuposição E(ai)=0. Com isso, enfatiza-se que o principal foco dos resultados e
discussões a seguir deveria ser dado sobre as conseqüências da existência de
diferenças genéticas entre GCs e não da seleção em si.
3.3.1 - ViésO viés das estimativas de efeito de GC (EGC), por geração, encontra-se na
Tabela 3.1. Foi apresentado apenas o viés referente aos três primeiros modelos
descritos no material e métodos, pois para os demais “modelos” os efeitos de GC não
foram estimados.
Quando houve seleção com base no BLUP e informação de parentesco
completa, apenas as soluções das EMMH regulares (modelo MMH_a) não
apresentaram viés. Esse resultado empírico está de acordo com as considerações de
HENDERSON (1973) de que as EMMH regulares acomodariam diferenças genéticas
entre níveis de efeitos ambientais sendo geradas seguindo seleção com base em
funções lineares das observações fenotípicas e que sejam invariantes a translação,
como é o caso do BLUP.
Considerando ainda a situação de seleção com base no BLUP e informação de
parentesco completa, é possível notar, na Tabela 3.1, que os modelos touro e vaca
apresentaram EGCs viesados. O viés ocorreu no sentido de subestimar EGCs de
gerações com animais de mérito genético superior. Embora o modelo MA1_tv não
requeira em sua derivação a pressuposição de ausência de diferenças genéticas entre
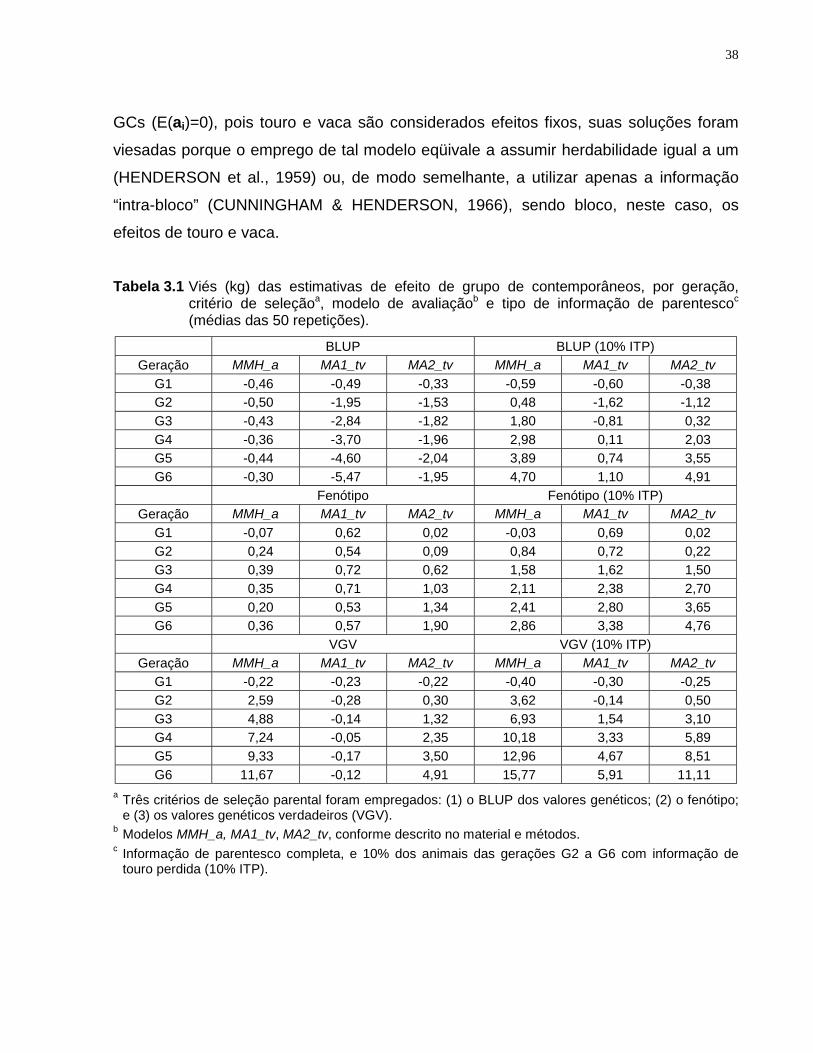
38
GCs (E(ai)=0), pois touro e vaca são considerados efeitos fixos, suas soluções foram
viesadas porque o emprego de tal modelo eqüivale a assumir herdabilidade igual a um
(HENDERSON et al., 1959) ou, de modo semelhante, a utilizar apenas a informação
“intra-bloco” (CUNNINGHAM & HENDERSON, 1966), sendo bloco, neste caso, os
efeitos de touro e vaca.
Tabela 3.1 Viés (kg) das estimativas de efeito de grupo de contemporâneos, por geração,critério de seleçãoa, modelo de avaliaçãob e tipo de informação de parentescoc
(médias das 50 repetições).
BLUP BLUP (10% ITP)Geração MMH_a MA1_tv MA2_tv MMH_a MA1_tv MA2_tv
G1 -0,46 -0,49 -0,33 -0,59 -0,60 -0,38G2 -0,50 -1,95 -1,53 0,48 -1,62 -1,12G3 -0,43 -2,84 -1,82 1,80 -0,81 0,32G4 -0,36 -3,70 -1,96 2,98 0,11 2,03G5 -0,44 -4,60 -2,04 3,89 0,74 3,55G6 -0,30 -5,47 -1,95 4,70 1,10 4,91
Fenótipo Fenótipo (10% ITP)Geração MMH_a MA1_tv MA2_tv MMH_a MA1_tv MA2_tv
G1 -0,07 0,62 0,02 -0,03 0,69 0,02G2 0,24 0,54 0,09 0,84 0,72 0,22G3 0,39 0,72 0,62 1,58 1,62 1,50G4 0,35 0,71 1,03 2,11 2,38 2,70G5 0,20 0,53 1,34 2,41 2,80 3,65G6 0,36 0,57 1,90 2,86 3,38 4,76
VGV VGV (10% ITP)Geração MMH_a MA1_tv MA2_tv MMH_a MA1_tv MA2_tv
G1 -0,22 -0,23 -0,22 -0,40 -0,30 -0,25G2 2,59 -0,28 0,30 3,62 -0,14 0,50G3 4,88 -0,14 1,32 6,93 1,54 3,10G4 7,24 -0,05 2,35 10,18 3,33 5,89G5 9,33 -0,17 3,50 12,96 4,67 8,51G6 11,67 -0,12 4,91 15,77 5,91 11,11
a Três critérios de seleção parental foram empregados: (1) o BLUP dos valores genéticos; (2) o fenótipo;e (3) os valores genéticos verdadeiros (VGV).
b Modelos MMH_a, MA1_tv, MA2_tv, conforme descrito no material e métodos.c Informação de parentesco completa, e 10% dos animais das gerações G2 a G6 com informação de
touro perdida (10% ITP).

39
Assim como em SCHENKEL et al. (2002), as soluções para GC do modelo
MMH_a não foram afetadas por seleção fenotípica, com informação de parentesco
completa. Esse resultado inesperado (HENDERSON ,1975; FERNANDO & GIANOLA,
1990) poderia, de acordo com SCHENKEL et al. (2002), estar em parte associado ao
fato de diferenças genéticas expressivas entre GCs de uma mesma geração não serem
esperadas para o processo de simulação utilizado, uma vez que, a cada geração, os
animais foram distribuídos aleatoriamente entre os GCs, e também pelo fato dos GCs
apresentarem, em média, número razoável de animais, permitindo assim estimação
acurada de seus efeitos.
Dentre os modelos touro e vaca empregados, apenas o modelo que os
considerou como efeitos fixos (MA1_tv) apresentou EGCs não viesados para a situação
de seleção fenotípica com informação de parentesco completa. O viés nos EGCs do
modelo MA2_tv está possivelmente associado ao uso de uma matriz diagonal para
descrever o parentesco entre as vacas.
Seleção do tipo L’u (com base no VGV) fez com que os modelos que
considerassem os efeitos genéticos ou parte deles como aleatórios, apresentassem
EGCs viesados. Mesmo quando uma matriz de parentesco correta e completa foi
definida, o modelo que assumiu o valor genético aditivo como efeito aleatório (modelo
MMH_a) não foi capaz de acomodar as diferenças genéticas entre GCs de diferentes
gerações, ao contrário do ocorrido para os demais tipos de seleção estudados.
Considerar touro e vaca como fixos (modelo MA1_tv) fez com que os EGCs fossem não
viesados. Resultados semelhantes foram observados por CARVALHEIRO et al. (2004–
capítulo anterior), porém estudando situações de diferenças genéticas entre GCs de
uma mesma geração.
Informação de touro perdida (10%ITP) fez com que todos os modelos
apresentassem EGCs viesados no sentido de superestimar o efeito de GCs de
gerações com animais de mérito genético superior. Mesmo os modelos MMH_a e
MA1_tv, que não haviam sido afetados por seleção com base no BLUP e no fenótipo e
com base no fenótipo e no VGV, respectivamente, apresentaram viés. É interessante
notar que considerando os modelos MA1_tv e MA2_tv e seleção com base no BLUP, o
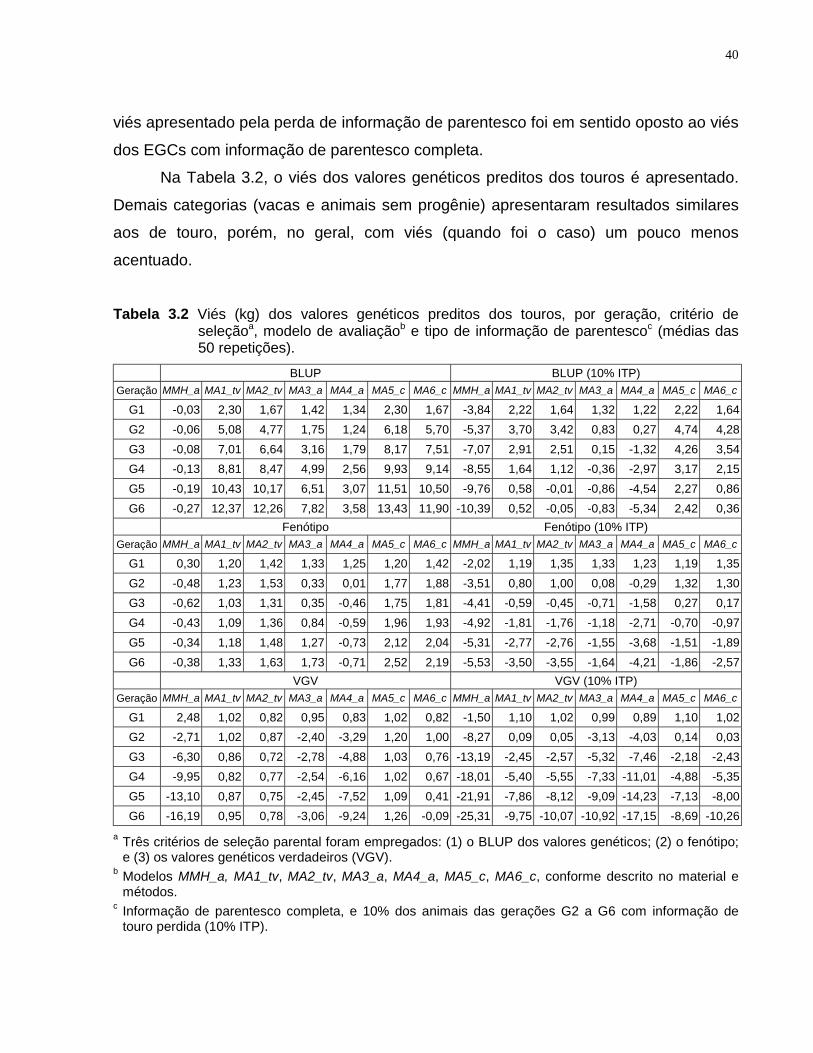
40
viés apresentado pela perda de informação de parentesco foi em sentido oposto ao viés
dos EGCs com informação de parentesco completa.
Na Tabela 3.2, o viés dos valores genéticos preditos dos touros é apresentado.
Demais categorias (vacas e animais sem progênie) apresentaram resultados similares
aos de touro, porém, no geral, com viés (quando foi o caso) um pouco menos
acentuado.
Tabela 3.2 Viés (kg) dos valores genéticos preditos dos touros, por geração, critério deseleçãoa, modelo de avaliaçãob e tipo de informação de parentescoc (médias das50 repetições).
BLUP BLUP (10% ITP)Geração MMH_a MA1_tv MA2_tv MA3_a MA4_a MA5_c MA6_c MMH_a MA1_tv MA2_tv MA3_a MA4_a MA5_c MA6_c
G1 -0,03 2,30 1,67 1,42 1,34 2,30 1,67 -3,84 2,22 1,64 1,32 1,22 2,22 1,64G2 -0,06 5,08 4,77 1,75 1,24 6,18 5,70 -5,37 3,70 3,42 0,83 0,27 4,74 4,28G3 -0,08 7,01 6,64 3,16 1,79 8,17 7,51 -7,07 2,91 2,51 0,15 -1,32 4,26 3,54G4 -0,13 8,81 8,47 4,99 2,56 9,93 9,14 -8,55 1,64 1,12 -0,36 -2,97 3,17 2,15G5 -0,19 10,43 10,17 6,51 3,07 11,51 10,50 -9,76 0,58 -0,01 -0,86 -4,54 2,27 0,86G6 -0,27 12,37 12,26 7,82 3,58 13,43 11,90 -10,39 0,52 -0,05 -0,83 -5,34 2,42 0,36
Fenótipo Fenótipo (10% ITP)Geração MMH_a MA1_tv MA2_tv MA3_a MA4_a MA5_c MA6_c MMH_a MA1_tv MA2_tv MA3_a MA4_a MA5_c MA6_c
G1 0,30 1,20 1,42 1,33 1,25 1,20 1,42 -2,02 1,19 1,35 1,33 1,23 1,19 1,35G2 -0,48 1,23 1,53 0,33 0,01 1,77 1,88 -3,51 0,80 1,00 0,08 -0,29 1,32 1,30G3 -0,62 1,03 1,31 0,35 -0,46 1,75 1,81 -4,41 -0,59 -0,45 -0,71 -1,58 0,27 0,17G4 -0,43 1,09 1,36 0,84 -0,59 1,96 1,93 -4,92 -1,81 -1,76 -1,18 -2,71 -0,70 -0,97G5 -0,34 1,18 1,48 1,27 -0,73 2,12 2,04 -5,31 -2,77 -2,76 -1,55 -3,68 -1,51 -1,89G6 -0,38 1,33 1,63 1,73 -0,71 2,52 2,19 -5,53 -3,50 -3,55 -1,64 -4,21 -1,86 -2,57
VGV VGV (10% ITP)Geração MMH_a MA1_tv MA2_tv MA3_a MA4_a MA5_c MA6_c MMH_a MA1_tv MA2_tv MA3_a MA4_a MA5_c MA6_c
G1 2,48 1,02 0,82 0,95 0,83 1,02 0,82 -1,50 1,10 1,02 0,99 0,89 1,10 1,02G2 -2,71 1,02 0,87 -2,40 -3,29 1,20 1,00 -8,27 0,09 0,05 -3,13 -4,03 0,14 0,03G3 -6,30 0,86 0,72 -2,78 -4,88 1,03 0,76 -13,19 -2,45 -2,57 -5,32 -7,46 -2,18 -2,43G4 -9,95 0,82 0,77 -2,54 -6,16 1,02 0,67 -18,01 -5,40 -5,55 -7,33 -11,01 -4,88 -5,35G5 -13,10 0,87 0,75 -2,45 -7,52 1,09 0,41 -21,91 -7,86 -8,12 -9,09 -14,23 -7,13 -8,00G6 -16,19 0,95 0,78 -3,06 -9,24 1,26 -0,09 -25,31 -9,75 -10,07 -10,92 -17,15 -8,69 -10,26
a Três critérios de seleção parental foram empregados: (1) o BLUP dos valores genéticos; (2) o fenótipo;e (3) os valores genéticos verdadeiros (VGV).
b Modelos MMH_a, MA1_tv, MA2_tv, MA3_a, MA4_a, MA5_c, MA6_c, conforme descrito no material emétodos.
c Informação de parentesco completa, e 10% dos animais das gerações G2 a G6 com informação detouro perdida (10% ITP).

41
Com informação de parentesco completa, os VGEs dos touros fornecidos pelo
modelo MMH_a foram não viesados para as situações de seleção com base no BLUP e
no fenótipo e viesados quando houve seleção com base no VGV. Para esta última
situação, como mencionado anteriormente, as soluções para os efeitos de GC do
modelo MMH_a foram contaminadas por efeitos genéticos. Com isso, os touros
superiores tiveram seus VGEs subestimados porque parte de sua superioridade foi
estimada como efeito de GC, sendo que o oposto ocorreu para os touros com mérito
genético inferior.
Os modelos touro e vaca (MA1_tv e MA2_tv), por considerarem touro como
efeito fixo, apresentaram, considerando informação de parentesco completa, soluções
não viesadas nos casos de seleção fenotípica e baseada no VGV, e VGEs viesados
para a situação de seleção com base no BLUP. O viés ocorreu no sentido de
superestimar os VGEs dos touros usados em gerações com mérito genético superior.
Os modelos MA5_c e MA6_c, por utilizarem as soluções dos modelos MA1_tv e
MA2_tv, respectivamente, para compor os VGEs dos touros, apresentaram
comportamento similar.
O procedimento de utilizar, como segundo passo, uma matriz de parentesco
correta e completa para descrever a covariância entre os valores genéticos fez com que
as soluções dos modelos MA3_a e MA4_a fossem menos viesadas do que aquelas
fornecidas pelos demais modelos alternativos (MA1_tv, MA2_tv, MA5_c e MA6_c), na
situação de informação de parentesco completa e seleção baseada no BLUP. Viés foi
observado, especialmente para o modelo MA4_a, quando houve seleção com base no
VGV, situação esta em que os outros modelos alternativos praticamente não
apresentaram viés. Esse resultado decorre do modelo MA4_a ter utilizado como
primeiro passo, no pré-ajuste das observações fenotípicas, as soluções dos efeitos de
GC fornecidas pelo modelo MA2_tv, soluções estas que apresentaram viés.
Assim como ocorreu com os EGCs, informação de touro perdida (10% ITP) fez
com que todos os modelos apresentassem VGEs viesados, para todas as situações
simuladas. O viés ocorreu no sentido de subestimar os VGEs dos touros usados em
gerações com animais de mérito genético superior. Na situação de seleção com base

42
no BLUP e 10% ITP, o viés dos modelos alternativos ocorreu no sentido oposto ao
observado na situação de parentesco completa, fazendo com que o viés dos VGEs
desses modelos, exceto MA4_a, fosse menos acentuado.
3.3.2 – Médias dos VGVsNa Tabela 3.3, são apresentadas as médias dos valores genéticos verdadeiros
dos 35 touros (10%) melhor classificados conforme os VGEs de cada modelo. Os
resultados mostraram que mesmo na situação ideal para o modelo MMH_a, isto é,
seleção com base no BLUP e informação de parentesco completa, as médias dos
modelos alternativos MA3_a e MA4_a não foram inferiores à do modelo MMH_a. Na
mesma situação, demais modelos alternativos, possivelmente por apresentarem VGEs
com viés relativamente acentuado, apresentaram médias inferiores à do modelo
MMH_a. Comportamento semelhante foi observado na situação de seleção fenotípica e
informação de parentesco completa, com MA3_a e MA4_a apresentando valores muito
próximos à MMH_a, e demais modelos com médias um pouco inferiores.
Tabela 3.3 Média (kg) dos valores genéticos verdadeiros dos touros 10% superiores,selecionados conforme soluções dos diferentes modelos de avaliação. Dados decada modelo são apresentados por critério de seleção e tipo de informação deparentesco, e são referentes à média dos valores das 50 repetições.
Modelo BLUP BLUP(10% ITP) Fenótipo Fenótipo
(10% ITP) VGV VGV(10% ITP)
MMH_a * 39,82 39,71 29,17 28,93 47,10 46,22MA1_tv 39,05 38,66 27,57 26,80 47,05NS 45,12MA2_tv 39,50 39,20 28,23 27,35 47,30> 45,44MA3_a 39,72NS 39,59 29,00 28,81NS 48,22> 47,61>
MA4_a 39,74NS 39,50 29,08NS 28,59 47,82> 46,78>
MA5_c 39,20 38,99 28,06 27,48 47,67> 46,43>
MA6_c 39,66 39,32 28,61 27,88 47,63> 46,31NS
* Testes-t pareados, com nível de significância de 5%, foram conduzidos contrastando todos os modelos,um por vez, com o modelo MMH_a. NS indica os contrastes que foram não significativos (p>0,05); >
refere-se à valores maiores do que os fornecidos pelo modelo MMH_a (p<0,05); ausência desobrescrito se refere à valores menores do que os fornecidos pelo modelo MMH_a (p<0,05).

43
Na situação de diferenças genéticas geradas com base no VGV, assim como
observado por CARVALHEIRO et al. (2004– capítulo anterior), modelos alternativos que
consideraram os efeitos genéticos ou parte deles como fixos permitiram, no geral, a
seleção de touros com mérito genético superior ao do modelo MMH_a.
Também foi possível observar na Tabela 3.3 que a informação de touro perdida
está associada à redução da habilidade em selecionar touros geneticamente
superiores, para todos os modelos e situações estudadas, mesmo que 10% ITP tenha
ocasionado redução do viés dos VGEs fornecidos pelos modelos MA1_tv, MA2_tv,
MA3_a, MA5_c e MA6_c na situação de seleção com base no BLUP.
Para as vacas 10% melhor classificadas, o modelo MMH_a apresentou médias
dos VGVs superiores aos demais modelos em todas as situações (p<0,05), embora os
modelos MA3_a e MA4_a tenham apresentado resultados muito próximos ao MMH_a
(Tabela 3.4).
Tabela 3.4 Média (kg) dos valores genéticos verdadeiros das vacas 10% superiores,selecionadas conforme soluções dos diferentes modelos de avaliação. Dados decada modelo são apresentados por critério de seleção e tipo de informação deparentesco, e são referentes à média dos valores das 50 repetições.
Modelo BLUP BLUP(10% ITP) Fenótipo Fenótipo
(10% ITP) VGV VGV(10% ITP)
MMH_a * 29,73 29,31 20,12 19,84 37,17 35,78MA1_tv 23,08 23,04 14,82 14,79 31,23 31,18MA2_tv 18,02 17,97 13,60 13,56 22,57 22,50MA3_a 29,29 28,89 19,53 19,38 36,46 35,46MA4_a 29,30 28,80 19,61 19,44 36,09 34,89MA5_c 21,58 20,79 14,01 13,87 27,07 26,19MA6_c 27,61 26,64 18,24 18,06 33,72 31,99
* Testes-t pareados, com nível de significância de 5%, foram conduzidos contrastando todos os modelos,um por vez, com o modelo MMH_a. Ausência de sobrescrito ao lado dos valores de média, refere-se àvalores menores do que os fornecidos pelo modelo MMH_a (p<0,05).
Pelos resultados da Tabela 3.4 se verificou que, conforme salientado por FRIES
(1999), a seleção das vacas foi seriamente comprometida ao utilizar modelo touro e
vaca (MA1_tv e MA2_tv), pois as principais fontes de informação (própria e dos pais)

44
não foram utilizadas no cálculo dos VGEs das vacas. A inferioridade do modelo MA2_tv
em relação ao modelo MA1_tv, quanto à média das vacas selecionadas, possivelmente
está associada ao uso, em MA2_tv, de uma matriz diagonal para descrever o
parentesco entre as vacas. Em CARVALHEIRO et al. (2004– capítulo anterior), MA2_tv
se mostrou superior à MA1_tv, quanto às propriedades empíricas estudadas, porém
naquele caso não havia realmente parentesco entre as vacas da população simulada.
Utilizar a proposta de SCHAEFFER (2003), de combinar as diferentes fontes de
informação, teve resultados favoráveis (maiores médias dos VGVs) para o modelo que
assumiu vaca como efeito aleatório (MA6_c) e desfavoráveis ao assumir vaca como fixo
(MA5_c). Mais investigações devem ser conduzidas para identificar porque o modelo
MA5_c apresentou médias dos VGVs das vacas inferiores às do modelo MA1_tv.
Outras intensidades foram aplicadas, selecionando 2%, 20% e 50% dos touros e
vacas melhores classificados, e resultados semelhantes aos descritos acima foram
observados (dados não apresentados).
3.3.3 – Tendências genéticasA seguir, o termo tendência genética será utilizado se referindo à media dos
valores genéticos preditos por geração. No presente estudo, tendências genéticas não
foram estimadas com a aplicação de modelos de regressão.
Os resultados de tendências genéticas apresentados nas Figuras 3.1 a 3.3
seguem os resultados referentes ao viés das soluções de cada modelo nas diferentes
situações. As tendências dos modelos MA5_c e MA6_c não foram apresentadas, pois
elas foram muito próximas as tendências dos modelos MA1_tv e MA2_tv,
respectivamente. A linha representada por VGV é referente a tendência verdadeira.
Considerando informação de parentesco completa, os modelos que assumiram
touro e vaca como efeitos fixos (MA1_tv, MA3_a e MA5_c) superestimaram o ganho
genético quando houve seleção com base no BLUP, e estimaram a tendência de forma
apropriada para as demais situações, exceto o modelo MA3_a que subestimou o ganho
na seleção com base no VGV, pois os VGEs dos touros e, consequentemente, dos
produtos, apresentaram um pouco de viés nesta situação. Informação de touro perdida

45
fez com que os modelos MA1_tv, MA3_a e MA5_c subestimassem o ganho em todas
as situações, assim como ocorrido com os demais modelos.
Modelos que consideraram touro como fixo e vaca como efeito aleatório
(MA2_tv, MA4_a e MA6_c), apenas não subestimaram o ganho genético para a
situação de seleção com base no BLUP e informação de parentesco completa.
Seguindo os resultados de viés, observou-se que o modelo MMH_a foi adequado
para estimar o ganho genético nas situações de seleção com base no BLUP e no
fenótipo e informação de parentesco completa. Quando diferenças genéticas com base
no VGV foram geradas, o modelo MMH_a foi o que mais subestimou a tendência
genética.
A combinação de seleção com base no BLUP e informação de parentesco
perdida fez com que as tendências genéticas estimadas pelos modelos MA1_tv, MA3_a
e MA5_c fossem mais próximas da tendência verdadeira do que aquela estimada pelo
modelo MMH_a.

46
Figura 3.1 Tendências genéticas para os diferentes modelos, com e sem informação de touroperdida (10% ITP), na situação de seleção com base no BLUP.
Critério de Seleção: BLUP
23,5
21,2
18,0
15,4
0,0
5,0
10,0
15,0
20,0
25,0
30,0
G1 G2 G3 G4 G5 G6Geração
Méd
ia d
os v
alor
es g
enét
icos
(kg)
TBVMMH_aMA3_aMA4_aMMH_a (10%MPI)MA3_a (10%MPI)MA4_a (10%MPI)
Critério de Seleção: BLUP
21,2
26,1
22,8
19,5
15,9
0,0
5,0
10,0
15,0
20,0
25,0
30,0
G1 G2 G3 G4 G5 G6Geração
Méd
ia d
os v
alor
es g
enét
icos
(kg)
TBVMA1_tvMA2_tvMA1_tv (10%MPI)MA2_tv (10%MPI)

47
Figura 3.2 Tendências genéticas para os diferentes modelos, com e sem informação de touroperdida (10% ITP), na situação de seleção fenotípica.
Critério de Seleção: Fenótipo
12,011,4
10,09,1
7,7
0,0
2,0
4,0
6,0
8,0
10,0
12,0
14,0
G1 G2 G3 G4 G5 G6Geração
Méd
ia d
os v
alor
es g
enét
icos
(kg)
TBVMMH_aMA3_aMA4_aMMH_a (10%MPI)MA3_a (10%MPI)MA4_a (10%MPI)
Critério de Seleção: Fenótipo
12,0
10,19,3
7,2
0,0
2,0
4,0
6,0
8,0
10,0
12,0
14,0
G1 G2 G3 G4 G5 G6Geração
Méd
ia d
os v
alor
es g
enét
icos
(kg)
TBVMA1_tvMA2_tvMA1_tv (10%MPI)MA2_tv (10%MPI)

48
Figura 3.3 Tendências genéticas para os diferentes modelos, com e sem informação de touroperdida (10% ITP), na situação de seleção com base no valor genético verdadeiro(VGV).
Critério de Seleção: TBV
28,5
16,6
23,9
20,3
12,3
18,8
15,2
0,0
5,0
10,0
15,0
20,0
25,0
30,0
G1 G2 G3 G4 G5 G6Geração
Méd
ia d
os v
alor
es g
enét
icos
(kg)
TBVMMH_aMA3_aMA4_aMMH_a (10%MPI)MA3_a (10%MPI)MA4_a (10%MPI)
Critério de Seleção: TBV
28,5
23,322,3
17,2
0,0
5,0
10,0
15,0
20,0
25,0
30,0
G1 G2 G3 G4 G5 G6Geração
Méd
ia d
os v
alor
es g
enét
icos
(kg)
TBVMA1_tvMA2_tvMA1_tv (10%MPI)MA2_tv (10%MPI)

49
3.3.4 – Discussão geralOs resultados empíricos do presente estudo de simulação, em concordância com
aqueles apresentados por CARVALHEIRO et al. (2004– capítulo anterior), evidenciaram
que o modelo de avaliação genética predominantemente aplicado em programas de
melhoramento (MMH_a) não é capaz de acomodar apropriadamente diferenças
genéticas entre GCs causadas por “seleção do tipo L’u”, mesmo que uma matriz de
parentesco correta e completa seja utilizada.
Na prática, não é possível conduzir “seleção do tipo L’u” ou tomar decisões, que
gerem diferenças genéticas entre GCs, com base no valor genético verdadeiro dos
animais. Entretanto, como descrito anteriormente, HENDERSON (1975 e 1990) sugeriu
que o estudo desta situação poderia ser útil para representar casos reais comuns como,
por exemplo, seleção com base em informações prévias ou em variáveis
correlacionadas com o VGV e não disponíveis ou não utilizadas na análise.
HENDERSON (1990) ainda empregou modelos de avaliação para lidar com “seleção do
tipo L’u” em casos de associação entre os efeitos de touro e de GC, população base
selecionada e grupos genéticos na avaliação de touros, ou seja, casos em que as
diferenças genéticas existentes não são geradas exclusivamente com base no BLUP
dos valores genéticos ou em outra função linear das observações fenotípicas que seja
invariante a translação.
HARVEY (1979), BURNSIDE & MEYER (1988), SIMIANER & WOLLNY (1989),
UGARTE et al. (1992), VISSCHER & GODDARD (1993), TORRES JR. et al. (1997),
TÄUBERT et al. (2002), entre outros, também observaram viés nas soluções do modelo
MMH_a em situações de diferenças genéticas entre níveis dos efeitos ambientais
causadas por variantes do processo de “seleção do tipo L’u”. No geral, os diferentes
autores observaram que, como no presente trabalho, as soluções para os efeitos
ambientais foram contaminadas por efeitos genéticos e, consequentemente, os animais
superiores tiveram seus VGEs subestimados porque parte de sua superioridade foi
estimada como efeito ambiental, sendo que o oposto ocorreu para os animais de mérito
genético inferior.

50
Dentre os modelos alternativos empregados, os mais promissores foram os
modelos MA3_a e MA4_a, que apresentaram certa vantagem em relação ao modelo
MMH_a na situação de “seleção do tipo L’u”, quanto aos critérios de comparação
estudados, e, nos demais tipos de seleção, apresentaram habilidade semelhante ao
modelo MMH_a em selecionar animais de mérito genético superior.
Sob o contexto Bayesiano, o uso dos modelos alternativos MA3_a e MA4_a não
seria adequado no sentido de não considerar a incerteza envolvida nas estimativas dos
efeitos de GC ao estimar os valores genéticos (SORENSEN & GIANOLA, 2002). As
conseqüências de uma estimação em dois passos, conforme empregada nos modelos
MA3_a e MA4_a, poderiam ser expressivas caso a quantidade de informação existente
para estimar os parâmetros de interesse secundário (EGCs, no caso) fosse limitada, o
que não ocorreu no presente estudo, em que os GCs apresentaram 20 animais, em
média.
GIANOLA et al. (1988), GIANOLA et al. (1989) e IM et al. (1989) propuseram
modelos alternativos baseados em métodos de estimação por máxima verossimilhança.
Nestes trabalhos, são apresentados argumentos favoráveis ao uso de funções de
verossimilhança não condicionais ao processo de seleção (uL). Se uL for utilizada, o
processo de seleção é ignorável quando for baseado apenas em informação contida
nos dados para análise, ou nos dados e em variáveis independentes a eles. Caso
contrário, o conhecimento do processo de seleção é necessário para que inferência
correta seja feita sobre os parâmetros de interesse. Se a função conjunta das variáveis
aleatórias e das observações fenotípicas for maximizada condicionalmente ao processo
de seleção (cL), as condições para ignorar o processo de seleção são, de acordo com
IM et al. (1989), ainda mais restritas do que as apresentadas por HENDERSON (1975).
As três situações de seleção simuladas no presente trabalho seriam ignoráveis se uL
fosse utilizada, ou seja, as funções de verossimilhança resultariam nos mesmos
estimadores considerando ou não os processos de seleção. Caso a opção fosse por
empregar cL, a seleção fenotípica não seria ignorável, por ser dependente dos efeitos
de GC (GIANOLA et al., 1988).

51
No caso de métodos de inferência baseados em distribuições amostrais,
HENDERSON (1990) mencionou que não estava certo se MMH_a ou um modelo
alternativo deveria ser adotado para casos em que MMH_a apresentasse soluções
viesadas. Os resultados do presente trabalho evidenciaram que modelos alternativos
podem apresentar propriedades superiores às do modelo MMH_a, e que nem sempre a
seleção, ou mais explicitamente, as diferenças genéticas entre níveis dos efeitos
ambientais, deveriam ser ignoradas.
O uso de grupos genéticos poderia ser uma alternativa para reduzir ou eliminar o
viés apresentado pelo modelo MMH_a na situação de “seleção do tipo L’u”, ou para o
caso de informação de parentesco perdida. Essa alternativa não foi testada no presente
estudo porque em conjuntos de dados reais dificilmente há oportunidade de definir
grupos genéticos de forma apropriada como em estudos de simulação. A inclusão de
grupos genéticos desnecessários no modelo pode acarretar em redução da acurácia de
predição (TORRES JR. et al., 1997), ou ainda resultar em problemas de estimabilidade
SULLIVAN (2002).
3.4 – Implicações
Os resultados encontrados indicam que progressos genéticos podem ser obtidos
com a aplicação dos diferentes modelos testados e que, de acordo com os resultados
referentes as médias dos valores genéticos verdadeiros dos touros classificados como
10% superiores, os progressos genéticos esperados seriam semelhantes.
Diferenças marcantes entre os modelos de avaliação ocorreram quanto a
habilidade em selecionar vacas de mérito genético superior. Neste caso, o progresso
genético poderia ser comprometido, e aconselha-se a adoção de modelos que não
usem apenas a informação da progênie para predizer o mérito genético das vacas.
Os valores de viés apresentados pelos diferentes modelos, nas diferentes
situações, evidenciam que a proporção de animais jovens de determinado rebanho a
receber certificado de superioridade genética poderia ser afetada, dependendo do
modelo adotado. Modelos que assumem o efeito de touro como aleatório, podem
favorecer a certificação de animais provenientes de rebanhos com mérito genético

52
inferior, caso as diferenças genéticas entre os rebanhos não possam ser explicadas
pelas informações contidas nos dados.
3.5 – Conclusões
Evidências foram encontradas de que modelos alternativos que consideram os
efeitos genéticos ou parte deles como efeito fixo podem apresentar propriedades
superiores às do modelo predominantemente aplicado em programas de melhoramento
(MMH_a), especialmente os modelos MA3_a e MA4_a.
Para casos em que as diferenças genéticas não são geradas seguindo decisões
tomadas unicamente com base no BLUP dos valores genéticos, touros com maior
proporção de filhos nos GCs/rebanhos/programas com animais de mérito genético
superior podem ter seu mérito genético subestimado se o modelo MMH_a for aplicado,
e tendências genéticas podem ser subestimadas.
Na prática, diferentes critérios/forças/decisões podem atuar no sentido de gerar
diferenças genéticas entre GCs e, possivelmente, todos os modelos testados
forneceriam soluções viesadas. Porém, com uma diferença marcante: modelos que
assumem os efeitos genéticos como aleatórios apresentariam viés no sentido de
subestimar o efeito de touros com maior proporção de filhos nos GCs/rebanhos com
animais de mérito genético superior, para casos em que não houvesse informação
completa e correta contida nos dados das forças que atuaram sobre a esperança dos
valores genéticos; e modelos que assumem os efeitos genéticos como fixos
apresentariam viés no sentido oposto, na presença de diferenças genéticas (entre GCs
de diferentes gerações) geradas com base no BLUP.

53
REFERÊNCIAS BIBLIOGRÁFICAS
BURNSIDE, E. B.; MEYER, K. Potential impact of bovine somatotropin on dairy sire
evaluation. J. Dairy Sci., v.71, p.2210-2219. 1988.
CARVALHEIRO, R. Flexibilizando a matriz R na predição de valores genéticos.
Dissertação (Mestrado em Zootecnia), Faculdade de Ciências Agrárias e
Veterinárias, Universidade Estadual Paulista, Câmpus de Jaboticabal, 2000. 107p.
CARVALHEIRO, R.; SCHENKEL, F. S.; FRIES, L. A.; QUEIROZ, S. A.; MILLER, S. P.
Empirical statistical and genetic proprieties of solutions from Henderson's mixed
model equations when sires are non-randomly used across contemporary groups. J.Anim. Breed. Genet. 2004 (a ser submetido).
CUNNINGHAM, E. P.; HENDERSON, C. R. Analytical techniques for incomplete block
experiments. Biometrics, v.22, p.829-842. 1966.
FERNANDO, R. L.; GIANOLA, D. Optimal properties of the conditional mean as a
selection criterion. Theor. Appl. Genet., v.72, p.822-825. 1986.
FERNANDO, R. L.; GIANOLA, D. Statistical inferences in populations undergoing
selection and non-random mating. In: Advances in Statistical Methods for GeneticImprovement of Livestock (ed. D. Gianola, and K. Hammond), Springer-Verlag,
Berlin. p.437-453. 1990.
FRIES, L. A. Connectability in beef cattle genetic evaluation: the heuristic approach
used in MILC.FOR. In: WORLD CONGRESS ON GENETICS APPLIED TO
LIVESTOCK PRODUCTION, 6., 1998, Armidale, Australia, Proceedings... v.27,
p.449-450.
FRIES, L. A. Selection amenable alternative models. In: INTERNATIONAL
SYMPOSIUM ON ANIMAL BREEDING AND GENETICS, 1999, Viçosa, MG, Brasil.
p.35-77.

54
FRIES, L. A.; SCHENKEL, F. S. Estimation and prediction under a selection model. In:
REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 30., 1993, Rio
de Janeiro, Brasil. Anais dos Simpósios... p.1-20.
GIANOLA, D. Can BLUP and REML be improved upon? In: WORLD CONGRESS ON
GENETICS APPLIED TO LIVESTOCK PRODUCTION, 4., 1990, Proceedings...
v.13, p.445-449.
GIANOLA, D.; FERNANDO, R. L.; IM, S.; FOULLEY, J. L. Likelihood estimation of
quantitative genetic parameters when selection occurs: models and problems.
Genome, v.31, p.768-777. 1989.
GIANOLA, D.; IM, S.; FERNANDO, R. L. Prediction of breeding value under
Henderson's selection model: A revisitation. J. Dairy Sci., v.71, p.2790-2798. 1988.
HARVEY, W.R. Accuracy of simplified regressed least-squares (SRLS) vs BLUP
methods for ranking sires. Indian J. Anim. Genet. Breed., v.1, p.7-9. 1979.
HENDERSON, C. R. Estimates of changes in herd environment. J. Dairy Sci., v.32,
p.706. 1949.
HENDERSON, C. R. Selection index and expected genetic advance. In: STATISTICAL
GENETICS AND PLANT BREEDING. Natl. Acad. Sci. Nat. Res. Council. Publ.,
p.141-163, 1963.
HENDERSON, C. R. Sire evaluation and genetics trends. In: ANIMAL BREEDING AND
GENETIC SYMPOSIUM IN HONOR OF DR. JAY L. LUSH, ASAS, 1973, Illinois,
USA, Proceedings... p.10-41.
HENDERSON, C. R. Best linear unbiased estimation and prediction under a selection
model. Biometrics, v.31, p.423-447. 1975.
HENDERSON, C. R. Undesirable properties of regressed least squares prediction of
breeding values. J. Dairy Sci., v.61, p.114-120. 1978.
HENDERSON, C. R. Applications of linear models in animal breeding. University of
Guelph, Ontario, Canada. 1984. 462 p.

55
HENDERSON, C. R. Comment. In: IM et al. (1989). Likelihood inferences in animal
breeding under selection: a missing-data theory view point. Genet. Sel. Evol., v.21,
p.409-410. 1989.
HENDERSON, C. R. Accounting for selection and mating biases in genetic evaluations.
In: Advances in Statistical Methods for Genetic Improvement of Livestock (ed.
D. Gianola, and K. Hammond), Springer-Verlag, Berlin. p.413-436. 1990.
HENDERSON, C. R.; KEMPTHORNE, O.; SEARLE, S. R.; VON KROSIGK, C. M. The
estimation of environmental and genetic trends from records subject to culling.
Biometrics, v.15, p.192-218. 1959.
IM, S.; FERNANDO, R. L.; GIANOLA, D. Likelihood inferences in animal breeding under
selection: a missing-data theory view point. Genet. Sel. Evol., v.21, p.399-414. 1989.
SAS Institute Inc. SAS/STAT, 1998. User guide. Cary: SAS Institute Inc., USA.
SCHAEFFER, L. R. Effects of non-random mating. In: QUANTITATIVE GENETICS
AND ANIMAL MODELS, Lesson 17, Winter Course. University of Guelph, Ontario,
Canada. 2003. 7p.
SCHENKEL, F. S. Studies on effects of parental selection on estimation of geneticparameters and breeding values of metric traits. Ph.D. Thesis. University of
Guelph, Ontario. 1998. 191p.
SCHENKEL, F. S.; SCHAEFFER, L. R.; BOETTCHER, P. J. Comparison between
estimation of breeding values and fixed effects using Bayesian and empirical BLUP
estimation under selection on parents and missing pedigree information. Genet. Sel.Evol., v.34, p.41-59. 2002.
SIMIANER, H. Prospects for third generation methods of genetic evaluation. In:
ANNUAL MEETING OF THE EUROPEAN ASSOCIATION FOR ANIMAL
PRODUCTION, 42., 1991, Germany, Proceedings... 13p.
SIMIANER, H.; WOLLNY, C. Impact of the potential use of bovine somatotropin on the
efficiency of a conventional dairy cattle breeding scheme. Livest. Prod. Sci., v.22,
p.31. 1989.

56
SORENSEN, D. A.; GIANOLA, D. Likelihood, Bayesian, and MCMC methods inquantitative genetics. Springer. 2002. 740p.
SORENSEN, D. A.; KENNEDY, B. W. Estimation of genetic variances from unselected
and selected populations. J. Anim. Sci., v.59, p.1213-1223. 1984.
SULLIVAN, P. Genetic evaluation strategies for multiple traits and countries. Ph.D.
Thesis. University of Guelph, Ontario. 2002. 152p.
TÄUBERT, H.; SWALVE, H. H.; SIMIANER, H. Treatment of management groups in
dairy cattle evaluations. In: WORLD CONGRESS ON GENETICS APPLIED TO
LIVESTOCK PRODUCTION, 7., 2002, Montpellier, France, Proceedings...
Communication 20-12.
TORRES JR., R. A. A.; BRACCINI NETO, J.; SILVA, M. A.; LOPES, P. S. Avaliação de
metodologias de predição de valor genético por meio de simulação. Rev. Bras.Zootec., v.26, p.1142-1146. 1997.
UGARTE, E.; ALENDA, R.; CARABAÑO, M. J. Fixed or random contemporary groups in
genetic evaluations. J. Dairy Sci., v.75, p.269-278. 1992.
VAN DER WERF, J. H. J. Models to estimate genetic parameters in cross breddairy cattle populations under selection. Ph. D. Thesis, Department of Animal
Breeding, Wageningen Agricultural University, Netherlands. 1990.
VAN VLECK, L. D. Contemporary groups for genetic evaluations. J. Dairy Sci., v.70,
p.2456-2464. 1987.
VANRADEN, P. M.; WIGGANS, G. R. Derivation, calculation and use of national animal
model information. J. Dairy Sci., v.74, p.2737-2746. 1991.
VISSCHER, P. M.; GODDARD, M. E. Fixed and random contemporary groups. J. DairySci., v.76, p.1444-1454. 1993.
WRIGHT, S. Coefficients of inbreeding and relationship. American Naturalist, v.56,
p.330-338. 1922.

57
Apêndice 1: Freqüências esperadas (%) da ocorrência das subclasses definidas por
conjuntos de touros e conjuntos de grupos de contemporâneos (GCs), para as
diferentes situação simuladas.
! Situação 1 – Os conjuntos de touros e de GCs foram formados conforme o número de
identificação (ID), resultando em uma situação caracterizada pela ausência de diferênças
genéticas entre conjuntos de GCs, e ausência de correlação genética ambiental.
! Situação 2 – Os conjuntos de touros foram formados conforme sua classificação (rank), em
ordem decrescente dos valores genéticos verdadeiros, e os conjuntos de GCs foram
formados conforme o número de identificação (ID), resultando em uma situação
caracterizada pela presença de diferênças genéticas entre conjuntos de GCs, e ausência de
correlação genética ambiental.
GCTouro 1-40 41-80 81-120 121-160 161-2001-50 12 6 2 0 0
51-100 6 8 4 2 0101-150 2 4 8 4 2151-200 0 2 4 8 6201-250 0 0 2 6 12
IDID
GCTouro 1-40 41-80 81-120 121-160 161-2001-50 12 6 2 0 0
51-100 6 8 4 2 0101-150 2 4 8 4 2151-200 0 2 4 8 6201-250 0 0 2 6 12
IDrank
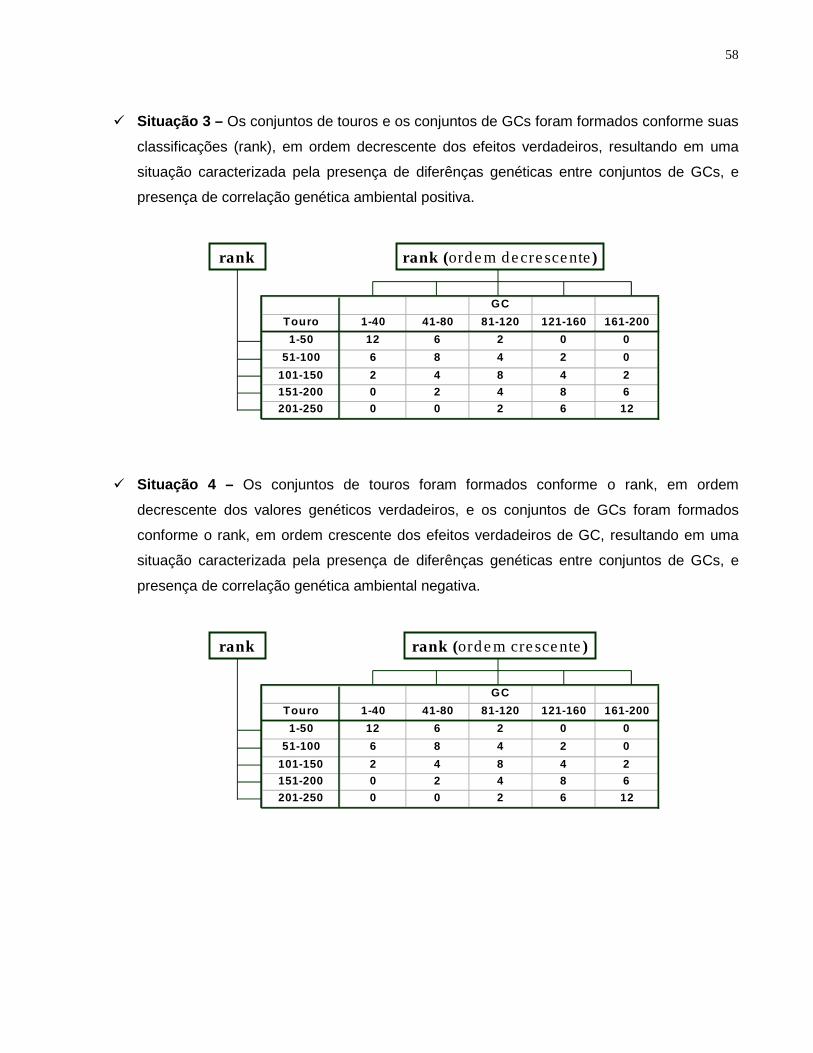
58
! Situação 3 – Os conjuntos de touros e os conjuntos de GCs foram formados conforme suas
classificações (rank), em ordem decrescente dos efeitos verdadeiros, resultando em uma
situação caracterizada pela presença de diferênças genéticas entre conjuntos de GCs, e
presença de correlação genética ambiental positiva.
! Situação 4 – Os conjuntos de touros foram formados conforme o rank, em ordem
decrescente dos valores genéticos verdadeiros, e os conjuntos de GCs foram formados
conforme o rank, em ordem crescente dos efeitos verdadeiros de GC, resultando em uma
situação caracterizada pela presença de diferênças genéticas entre conjuntos de GCs, e
presença de correlação genética ambiental negativa.
GCTouro 1-40 41-80 81-120 121-160 161-2001-50 12 6 2 0 0
51-100 6 8 4 2 0101-150 2 4 8 4 2151-200 0 2 4 8 6201-250 0 0 2 6 12
rank (ordem decrescente)rank
GCTouro 1-40 41-80 81-120 121-160 161-2001-50 12 6 2 0 0
51-100 6 8 4 2 0101-150 2 4 8 4 2151-200 0 2 4 8 6201-250 0 0 2 6 12
rank (ordem crescente)rank

59
Apêndice 2: Programa de simulação desenvolvido em SAS para gerar a estrutura dos
dados simulados no primeiro estudo, conforme descrito no Capítulo 2, item 2.2.1.
/*************************************//* RC *//* *//* doutorado JAN/2003 *//* *//* gerando estrutura dos dados *//* *//*************************************/
proc iml;
seed=27903441;
%MACRO CRIAR; /* comando MACRO usado para gerar repetições */%DO J=1 %TO 50; /* e gravar arq. saída com diferentes nomes */
n1=5000; /* n1=nº total de observações (animais) */n2=250; /* n2=nº de touros */n3=200; /* n3=nº de GC´s */
t=j(n1,1,0); /* t=touro (n2 touros gerando n1 animais) */gc=j(n1,1,0); /* n1 animais distribuídos em n3 GC´s */tgc=j(n2,n3,0); /* tgc subclasses touro*gc */ntgc=j(n3,1,0); /* ntgc=nº de touros por GC */ngct=j(n2,1,0); /* ngct=nº de GCs por touro */nt=j(n2,1,0); /* nt=nº de filhos por touro */ngc=j(n3,1,0); /* ngc=nº de animais por GC */
vaca=j(n1,1,0);an=j(n1,1,0);
/* "Restrições": mínimo máximo *//* (1) filhos por touro n4 n5 *//* (2) animais por GC n6 n7 *//* (3) touro por GC n8 n9 *//* (4) GC por touro n10 n11 */
n4=5;n5=50;n6=10;n7=100;n8=3;n9=50;n10=3;n11=40;

60
/* Iniciando Processo de amostragem (pseudo)aleatória *//* usando distribuição uniforme para amostrar touro e GC */
R=1;DO WHILE(R=1);
REINI:do i= 1 to n1;
an[i]=20000+i; if i<=(n1/2) then vaca[i]=10000+i; else vaca[i]=10000+i-(n1/2);
INIT: t[i]=int(ranuni(seed)*n2)+1;
if t[i]<=50 then rt=1; if t[i]>50 then if t[i]<=100 then rt=2; if t[i]>100 then if t[i]<=150 then rt=3; if t[i]>150 then if t[i]<=200 then rt=4; if t[i]>200 then rt=5;
prob1=0; rng1=ranuni(seed); if rng1<=.6 then prob1=1; if rng1>.6 then if rng1<=.9 then prob1=2; if rng1>.9 then prob1=3;
prob2=0; rng2=ranuni(seed); if rng2<=.3 then prob2=1; if rng2>.3 then if rng2<=.7 then prob2=2; if rng2>.7 then if rng2<=.9 then prob2=3; if rng2>.9 then prob2=4;
prob3=0; rng3=ranuni(seed); if rng3<=.1 then prob3=1; if rng3>.1 then if rng3<=.3 then prob3=2; if rng3>.3 then if rng3<=.7 then prob3=3; if rng3>.7 then if rng3<=.9 then prob3=4; if rng3>.9 then prob3=5;
if rt=1 then if prob1=1 then gc[i]=int(ranuni(seed)*20)+1; if rt=1 then if prob1=2 then gc[i]=int(ranuni(seed)*20)+41; if rt=1 then if prob1=3 then gc[i]=int(ranuni(seed)*20)+81;
if rt=2 then if prob2=1 then gc[i]=int(ranuni(seed)*20)+1; if rt=2 then if prob2=2 then gc[i]=int(ranuni(seed)*20)+41; if rt=2 then if prob2=3 then gc[i]=int(ranuni(seed)*20)+81; if rt=2 then if prob2=4 then gc[i]=int(ranuni(seed)*20)+121;

61
if rt=3 then if prob3=1 then gc[i]=int(ranuni(seed)*20)+1; if rt=3 then if prob3=2 then gc[i]=int(ranuni(seed)*20)+41; if rt=3 then if prob3=3 then gc[i]=int(ranuni(seed)*20)+81; if rt=3 then if prob3=4 then gc[i]=int(ranuni(seed)*20)+121; if rt=3 then if prob3=5 then gc[i]=int(ranuni(seed)*20)+161;
if rt=4 then if prob2=4 then gc[i]=int(ranuni(seed)*20)+41; if rt=4 then if prob2=3 then gc[i]=int(ranuni(seed)*20)+81; if rt=4 then if prob2=2 then gc[i]=int(ranuni(seed)*20)+121; if rt=4 then if prob2=1 then gc[i]=int(ranuni(seed)*20)+161;
if rt=5 then if prob1=3 then gc[i]=int(ranuni(seed)*20)+81; if rt=5 then if prob1=2 then gc[i]=int(ranuni(seed)*20)+121; if rt=5 then if prob1=1 then gc[i]=int(ranuni(seed)*20)+161;
resto=(mod(i,2)); if resto^=0 then gc[i]=gc[i]+20;
nt[t[i]]=nt[t[i]]+1; if nt[t[i]]>n5 then do; /* máximo de filhos por touro */ nt[t[i]]=nt[t[i]]-1; go to INIT; end;
ngc[gc[i]]=ngc[gc[i]]+1; if ngc[gc[i]]>n7 then do; /* máximo de animais por GC */ nt[t[i]]=nt[t[i]]-1; ngc[gc[i]]=ngc[gc[i]]-1; go to INIT; end;
tgc[t[i],gc[i]]=1; ntgc[gc[i]]=0; ngct[t[i]]=0;
do l=1 to n2; ntgc[gc[i]]=ntgc[gc[i]]+tgc[l,gc[i]]; end; if ntgc[gc[i]]>n9 then do; /* máximo de touros por GC */ nt[t[i]]=nt[t[i]]-1; ngc[gc[i]]=ngc[gc[i]]-1; tgc[t[i],gc[i]]=0; go to INIT; end;
do l=1 to n3; ngct[t[i]]=ngct[t[i]]+tgc[t[i],l]; end; if ngct[t[i]]>n11 then do; /* máximo de GCs por touro */ nt[t[i]]=nt[t[i]]-1; ngc[gc[i]]=ngc[gc[i]]-1; tgc[t[i],gc[i]]=0; go to INIT; end;

62
end;
restr1=0;restr2=0;restr3=0;restr4=0; /* Verificando Restrições */ /* */do m=1 to n2; /* */ if nt[m]<n4 then restr1=1; /* mínimo de filhos por touro */end; /* */ /* */
do m=1 to n3; /* */ if ngc[m]<n6 then restr2=1; /* mínimo de animais por GC */end; /* */ /* */do m=1 to n3; /* */ if ntgc[m]<n8 then restr3=1; /* mínimo de touros por GC */end; /* */ /* */do m=1 to n2; /* */ if ngct[m]<n10 then restr4=1; /* mínimo de GCs por touro */end; /* */
/* Iniciando avaliação de conectabilidade das subclasses (touro*GC) *//* de acordo com Schaeffer (1983 - p.39). *//* Se nsub=soma, então os dados estão conectados, *//* senão, REINIciar amostragem. */
nsub=0;obs=0;soma=sum(tgc);
do i1=1 to n2; do j1=1 to n3;
if obs=0 then do; if tgc[i1,j1]=1 then do; tgc[i1,j1]=0; nsub=nsub+1; obs=obs+1; CONT: do i2=1 to n3; if tgc[i1,i2]=1 then tgc[i1,i2]=2; end; do j2=1 to n2; if tgc[j2,j1]=1 then tgc[j2,j1]=2; end;
end; end;

63
if tgc[i1,j1]=2 then do; tgc[i1,j1]=0; nsub=nsub+1; go to CONT; end;
if i1=n2 then if j1=n3 then do; do i3=1 to n2; do j3=1 to n3; if tgc[i3,j3]=2 then do; i1=1; j1=1; end; end; end; end;
end;end;
do o=1 to n2; /* Zerando contadores */ nt[o]=0; /* para a próxima repetição */end; /* ou para REINIciar se */do p=1 to n3; /* restrições de mínimo não */ ngc[p]=0; /* foram atendidas (restr=1) */end; /* ou ainda se dados não */do q=1 to n2; /* estão conectados */ do s=1 to n3; tgc[q,s]=0; end;end;
print restr1 restr2 restr3 restr4 soma nsub;
if restr1=1 then go to REINI;if restr2=1 then go to REINI;if restr3=1 then go to REINI;if restr4=1 then go to REINI;if nsub^=soma then go to REINI;
R=R+1;END;
CREATE ESQ&J VAR{an t vaca gc};APPEND;CLOSE ESQ&J;
%END;%MEND CRIAR;%CRIAR;
quit;run;

64
%MACRO GRAVAR;%DO J=1 %TO 50;
data a;set esq&J;file "c:\roberto\s030331\dados&J";put an 1-6 t 8-13 vaca 15-20 gc 22-24;run;
%END;%MEND GRAVAR;%GRAVAR;

65
Apêndice 3: Programa de simulação desenvolvido em SAS para gerar os parâmetros
do modelo e as observações fenotípicas das simulações referentes ao primeiro estudo,
conforme descrito no Capítulo 2, item 2.2.2.
/********************************************************************** DOUTORADO (FEV/2003)
Programa para gerar Y (GPP), nas diferentes situações de associação touro_x_GC.
(1) var(a)=120; var(e) = 180; var(p)=300 kg^2 ; h^2=0,4; (2) E(egc)=172,5 ; var(egc)=300; (3) GPP=vga+egc+e**********************************************************************/
data clast;do i=1 to 250; touro=i; output;end;drop i;
%MACRO CRIAR;%DO J=1 %TO 50;
/* ST2: diferenças genéticas entre GCs (r(GC,touro)=0) */
data a;infile "c:\roberto\s030331\dados&J";input an touro vaca gc;proc sort;by touro;run;
data a2; /* TBV – touros */do i=1 to 250; vgat=rannor(0)*10.954; output;end;proc sort;by descending vgat;run;
data b;merge clast a2;drop i;proc sort;by touro;run;

66
data c;merge a b;by touro;proc sort;by vaca;run;
data d; /* TBV – vacas */do vaca=10001 to 12500; vgav=rannor(0)*10.954; output;end;
data e;merge c d;by vaca;
data f;set e; /* TBV – produtos */sm=rannor(0)*7.746;vga=((vgat+vgav)/2)+sm;proc sort;by gc;run;
data g; /* TGC */do gc=1 to 200; egc=rannor(0)*17.3205+172.5; output;end;
data h;merge f g;by gc;
data i;set h;e=rannor(0)*13.416;GPP=vga+egc+e;proc sort;by an;run;
data j;set i;file "c:\roberto\s030331\dat2r&J";put an 1-6 touro 8-13 vaca 15-20 gc 22-24 gpp 26-32 .3;run;
data k;set i;file "c:\roberto\s030331\cor2r&J";put an 1-6 vga 8-15 .4 touro 17-22 vgat 24-31 .4 vaca 33-38 vgav 40-47 .4 gc 49-51 egc 53-60 .4 e 62-69 .4 sm 71-78 .4;run;

67
/* ST1: ausência de diferenças genéticas entre GCs */
data l;set a2;rename i=touro;proc sort;by touro;run;
data l3;merge a l;by touro;proc sort;by an;run;
data l6;set k;keep an vgav egc e sm;
data l9;merge l3 l6;by an;vga=((vgat+vgav)/2)+sm;GPP=vga+egc+e;
data l11;set l9;file "c:\roberto\s030331\dat1r&J";put an 1-6 touro 8-13 vaca 15-20 gc 22-24 gpp 26-32 .3;run;
data l11;set l9;file "c:\roberto\s030331\cor1r&J";put an 1-6 vga 8-15 .4 touro 17-22 vgat 24-31 .4 vaca 33-38 vgav 40-47 .4 gc 49-51 egc 53-60 .4 e 62-69 .4 sm 71-78 .4;run;
/* ST3: diferenças genéticas entre GCs (r(GC,touro)>0) */
data m;do i=1 to 200; gc=i; output;end;drop i;
data m3;set g;rename gc=i;proc sort;by descending egc;run;

68
data m6;merge m m3;drop i;
data m9;set i;keep an vga touro vgat vaca vgav gc e sm;proc sort;by gc;run;
data m11;merge m6 m9;by gc;GPP=vga+egc+e;proc sort;by an;run;
data m13;set m11;file "c:\roberto\s030331\dat3r&J";put an 1-6 touro 8-13 vaca 15-20 gc 22-24 gpp 26-32 .3;run;
data m13;set m11;file "c:\roberto\s030331\cor3r&J";put an 1-6 vga 8-15 .4 touro 17-22 vgat 24-31 .4 vaca 33-38 vgav 40-47 .4 gc 49-51 egc 53-60 .4 e 62-69 .4 sm 71-78 .4;run;
%END;%MEND CRIAR;%CRIAR;

69
Apêndice 4: Programa desenvolvido em linguagem FORTRAN para conduzir
avaliações genéticas, conforme modelos descritos no Capítulo 2, item 2.2.3.
C RC - FEV/2003CCC M1 - (gc fixo e touro e vaca aleatorios)C M2 - (gc, touro e vaca aleatorios)C M3 - (gc, touro e vaca fixos)C M4 - (gc e touro fixos e vaca aleatorio)CCC h2=0.4 (rvep=8; regd=0.125)CC PARAMETER (nobs=20000,nivel=1000,rvep=8.0,regd=0.125)
INTEGER KA(nobs),KT(nobs),KV(nobs),KG(nobs) INTEGER SIRE(nivel),DAM(nobs),GC(nivel) INTEGER ISIRE(nobs),IDAM(nobs),IGC(nobs) INTEGER WT1(nobs),WT2(nobs),WV1(nobs),WV2(nobs), * WG1(nobs),WG2(nobs) INTEGER NT,NV,NG,IREG INTEGER BT,BV,BG REAL Y(nobs),MU,MUGF REAL VT(nivel),AT(nivel),VV(nobs),AV(nobs),VG(nivel),AG(nivel) INTEGER nIT(nivel),nIV(nobs),nIG(nivel) REAL LIM,DIFT,DIFT2,DIFV,DIFV2,DIFG,DIFG2
CC formato de entrada dos dados:C AN SIRE DAM GC GPPC 911 FORMAT(I6,1X,I6,1X,I6,1X,I3,1X,F7.3)CC obs: iSIRE, iDAM e iGC sao a "ID original"C
911 FORMAT(I6,1X,I6,1X,I6,1X,I3,1X,F7.3) 912 FORMAT(I6,1X,I6,1X,F12.7) 913 FORMAT(I6,1X,I6,1X,F12.7) 914 FORMAT(I6,1X,I6,1X,F12.7) 915 FORMAT(I6,1X,F12.7,1X,F12.7) 7891 FORMAT(T3,I5,T13,F10.5,T25,F10.5,T37,F10.5)
OPEN(11,FILE='dados',STATUS='OLD') OPEN(12,FILE='gcHtv',STATUS='UNKNOWN') OPEN(13,FILE='toHtv',STATUS='UNKNOWN') OPEN(14,FILE='vaHtv',STATUS='UNKNOWN') OPEN(15,FILE='anHtv',STATUS='UNKNOWN')

70
C Lendo arquivo de dadosC I=1 MU=0.0 10 CONTINUE READ(11,911,END=12)KA(I),ISIRE(I),IDAM(I),IGC(I),Y(I) MU=MU+(Y(I)-MU)/I I=I+1 GOTO 10 12 CONTINUE IREG=I-1 WRITE(*,*)' IREG=',IREG WRITE(*,*)' Media Geral=',mu
CC (i)Gerando nova ID para SIRE, DAM e GC de acordo com a ordem queC aparecem no arquivo de dadosC obs: KT, KV e KG sao a "ID sequencial"C (ii)Obtendo numero de touros, vacas e GC presentes noC arquivo de dados (NT,NV,NG)C
NT=0 NV=0 NG=0
DO 14 I=1,IREG WT1(I)=0 WT2(I)=0 WV1(I)=0 WV2(I)=0 WG1(I)=0 WG2(I)=0 14 CONTINUE
DO 16 I=1,IREG
IF(ISIRE(I).NE.0)THEN
DO 18 J=1,NT IF(WT1(J).EQ.ISIRE(I))THEN KT(I)=WT2(J) GOTO 20 ENDIF 18 CONTINUE NT=NT+1 WT1(NT)=ISIRE(I) WT2(NT)=NT KT(I)=NT
20 CONTINUE
ENDIF IF(IDAM(I).NE.0)THEN

71
DO 22 J=1,NV IF(WV1(J).EQ.IDAM(I))THEN KV(I)=WV2(J) GOTO 24 ENDIF 22 CONTINUE NV=NV+1 WV1(NV)=IDAM(I) WV2(NV)=NV KV(I)=NV
24 CONTINUE
ENDIF
DO 26 J=1,NG IF(WG1(J).EQ.IGC(I))THEN KG(I)=WG2(J) GOTO 16 ENDIF 26 CONTINUE NG=NG+1 WG1(NG)=IGC(I) WG2(NG)=NG KG(I)=NG
16 CONTINUE
DO 28 I=1,IREG IF(ISIRE(I).NE.0)SIRE(KT(I))=ISIRE(I) IF(IDAM(I).NE.0)DAM(KV(I))=IDAM(I) GC(KG(I))=IGC(I) 28 CONTINUE
DO 34 I=1,NG VG(I)=0.0 AG(I)=0.0 nIG(I)=0 34 CONTINUE WRITE(*,*)NG,' Grupos Contemporaneos'
DO 36 I=1,NT VT(I)=0.0 AT(I)=0.0 nIT(I)=0 36 CONTINUE WRITE(*,*)NT,' Touros'C DO 38 I=1,NV VV(I)=0.0 AV(I)=0.0 nIV(I)=0 38 CONTINUE WRITE(*,*)NV,' Vacas'

72
C INICIA PROCESSO ITERATIVOC
ITR=0 LIM=0.0005 MUGF=MU 50 CONTINUE IVOLTA=0 DIFT=0.0001 DIFV=0.0001 DIFG=0.0001 ITR=ITR+1 IF(ITR.EQ.501)GOTO 9000
CC TOUROSC
DO 54 J=1,NT AT(J)=0.0 nIT(J)=0 54 CONTINUE
DO 60 I=1,IREG IF(ISIRE(I).NE.0)THEN A1=Y(I)-MUGF-VG(KG(I))-VV(KV(I)) AT(KT(I))=AT(KT(I))+A1 nIT(KT(I))=nIT(KT(I))+1 ENDIF 60 CONTINUEC bt=0 SOMAT=0.0
C se touro eh fixo, usar AT(J)=AT(J)/nIT(J) e impor restricao AT(J)=AT(J)-CTcC se touro eh aleatorio, usar AT(J)=AT(J)*nIT(J)/(nIT(J)+rvep)C
DO 70 J=1,NT IF(nIT(J).NE.0)THEN AT(J)=AT(J)/nIT(J)ccc AT(J)=AT(J)*nIT(J)/(nIT(J)+rvep) SOMAT=SOMAT+AT(J) bt=bt+1 ENDIF 70 CONTINUE
CT=SOMAT/BT MUGF=MUGF+CT DO 80 J=1,NT IF(nIT(J).NE.0)THEN AT(J)=AT(J)-CT DIF=AT(J)-VT(J)

73
DIFT2=ABS(DIF) IF(DIFT2.GT.DIFT)DIFT=DIFT2 VT(J)=AT(J) ENDIF 80 CONTINUE IF(DIFT.GT.LIM)IVOLTA=1
CC GCSC DO 85 J=1,NG AG(J)=0.0 nIG(J)=0 85 CONTINUE
DO 90 I=1,IREG
IF(ISIRE(I).NE.0.AND.IDAM(I).NE.0) * A1=Y(I)-MUGF-VT(KT(I))-VV(KV(I)) AG(KG(I))=AG(KG(I))+A1 nIG(KG(I))=nIG(KG(I))+1
90 CONTINUE
C se GC eh fixo, usar AG(J)=AG(J)/nIG(J) e impor restricao AG(J)=AG(J)-CORGcC se GC eh aleatorio, usar AG(J)=AG(J)*nIG(J)/(nIG(J)*0.8)C bg=0 SOMAG=0.0 DO 95 J=1,NG IF(nIG(J).NE.0)THEN AG(J)=AG(J)/nIG(J)ccc AG(J)=AG(J)*nIG(J)/(nIG(J)*0.8) SOMAG=SOMAG+AG(J) bg=bg+1 ENDIF 95 CONTINUE CORG=SOMAG/BG MUGF=MUGF+CORG DO 100 J=1,NG IF(nIG(J).NE.0)THEN AG(J)=AG(J)-CORG DIF=AG(J)-VG(J) DIFG2=ABS(DIF) IF(DIFG2.GT.DIFG)DIFG=DIFG2 VG(J)=AG(J) ENDIF 100 CONTINUE IF(DIFG.GT.LIM)IVOLTA=1

74
CC VACASC
DO 150 J=1,NV AV(J)=0.0 nIV(J)=0 150 CONTINUE
DO 160 I=1,IREG IF(IDAM(I).NE.0)THEN A1=Y(I)-MUGF-VG(KG(I))-VT(KT(I)) AV(KV(I))=AV(KV(I))+A1 nIV(KV(I))=nIV(KV(I))+1 ENDIF 160 CONTINUE
C se vaca eh fixo, usar AV(J)=AV(J)/nIV(J) e impor restricao AV(J)=AV(J)-CORVcC se vaca eh aleatorio, usar AV(J)=AV(J)*nIV(J)/(nIV(J)+rvep)C bv=0 SOMAV=0.0 DO 170 J=1,NV IF(nIV(J).NE.0)THEN AV(J)=AV(J)/nIV(J)ccc AV(J)=AV(J)*nIV(J)/(nIV(J)+rvep) SOMAV=SOMAV+AV(J) bv=bv+1 ENDIF 170 CONTINUE
CORV=SOMAV/BV MUGF=MUGF+CORV DO 180 J=1,NV IF(nIV(J).NE.0)THEN AV(J)=AV(J)-CORV DIF=AV(J)-VV(J) DIFV2=ABS(DIF) IF(DIFV2.GT.DIFV)DIFV=DIFV2 VV(J)=AV(J) ENDIF 180 CONTINUE IF(DIFV.GT.LIM)IVOLTA=1C imit=mod(itr,10) if(itr.lt.6.or.imit.eq.0) then WRITE(*,7891) ITR,DIFT,DIFG,DIFV endif
IF(IVOLTA.EQ.1) GOTO 50CC WRITE(*,*) ' I T E R A C O E S T E R M I N A R A M ! ! !'

75
c WRITE(*,*)' GRAVANDO DP.G ...' DO 512 J=1,NG 512 WRITE(12,912)GC(J),nIG(J),VG(J)c WRITE(*,*)' GRAVANDO DP.T ...' DO 513 J=1,NT 513 WRITE(13,913)SIRE(J),nIT(J),VT(J)c WRITE(*,*)' GRAVANDO DP.V ...' DO 514 J=1,NV 514 WRITE(14,914)DAM(J),nIV(J),VV(J)c WRITE(*,*)' GRAVANDO DP.A ...' DO 515 I=1,IREG IF(ISIRE(I).NE.0.AND.IDAM(I).NE.0)THEN D=Y(I)-MUGF-VG(KG(I))-VT(KT(I))-VV(KV(I)) SM=D*regd DEPA=0.5*(VT(KT(I))+VV(KV(I)))+SM ENDIF
WRITE(15,915)KA(I),SM,DEPA 515 CONTINUECC GOTO 9001 9000 WRITE(*,*)' NAO HOUVE CONVERGENCIA APOS 500 ITERACOES !!! ' 9001 continueC STOP END

76
Apêndice 5: Desdobramento das equações de modelos mistos de Henderson
modificadas para a obtenção do BLUE/BLUP sob situações de “seleção do tipo L’y”
com L’X=0, e seleção com base no BLUP.
Baseado nos resultados de PEARSON (1903), HENDERSON (1975) derivou as
seguintes equações para a obtenção do BLUE/BLUP sob situações de seleção:
====
++++−−−−−−−−++++
−−−−
−−−−
−−−−
−−−−−−−−−−−−−−−−−−−−
−−−−−−−−−−−−−−−−−−−−
−−−−−−−−−−−−
yR'ByR'ZyR'X
tub
BR'BBR'BG'BZR'BXR'B
BGBR'ZGZR'ZXR'ZBR'XZR'XXR'X
1e
1
1
u1
ue1
e1
u1
e1
e
u1
e1111
e111
. (1)
Seleção com base em y:
Se a seleção for baseada nas observações fenotípicas, HENDERSON (1975)
demonstrou que as equações (1) se reduzem a:
====
++++
−−−−
−−−−
−−−−
−−−−−−−−−−−−
−−−−−−−−
yR'LyR'ZyR'X
tub
VL'L0X'L0GZR'ZXR'ZL'XZR'XXR'X
1
1
1
111
11
. (2)
Para que as equações (2) sejam equivalentes as EMMH regulares, a condição
L’X=0 deve ser atendida, isto é, a matriz que descreve o processo de seleção (L’) deve
ser ortogonal à matriz X. Esta condição ocorrerá quando as decisões baseadas em yforem tomadas dentro dos diferentes níveis dos efeitos fixos/ambientais, o que
raramente ocorre na prática, sobretudo na existência de covariáveis afetando y (FRIES,
1999; SCHAEFFER, 1999).
Seleção com base no BLUP:
Se o critério de seleção for o BLUP, então: Bu=Cov(u,BLUP) e
Be=Cov(e,BLUP). A seguir, as equações (1) são desdobradas considerando o BLUP

77
como critério de seleção. O objetivo desde desdobramento é averiguar se (1) se
reduziria às EMMH regulares caso a seleção fosse praticada com base no BLUP. Para
que isso ocorra, temos que provar que:
i) X’R-1Be=0ii) Z’R-1Be-G-1Bu=0
No 3º e 4º passos descritos a seguir, i) e ii) serão, respectivamente,
demonstrados.
1º passo: desdobrar Bu.
Bu = Cov(u,BLUP) = Cov(u,GZ’V-1(y-Xb ))
= Cov(u,GZ’V-1y-GZ’V-1Xb ) = Cov(u,GZ’V-1y-GZ’V-1X(X’V-1X)-1X’V-1y) = Cov(u,y)V-1ZG – Cov(u,y)V-1X(X’V-1X)-1X’V-1ZG = GZ’V-1ZG – GZ’V-1X(X’V-1X)-1X’V-1ZG
2º passo: desdobrar Be.
Be = Cov(e,BLUP) = Cov(y-Xb-Zu,BLUP) = Cov(y,BLUP) – Cov(Zu,BLUP) = Cov(y,GZ’V-1y- GZ’V-1X(X’V-1X)-1X’V-1y) – Z Cov(u,BLUP) = Cov(y,y)V-1ZG- Cov(y,y)V-1X(X’V-1X)-1X’V-1ZG) – Z Cov(u,BLUP) = ZG- X(X’V-1X)-1X’V-1ZG – ZGZ’V-1ZG + ZGZ’V-1X(X’V-1X)-1X’V-1ZG = ZG- X(X’V-1X)-1X’V-1ZG – (I-RV-1)ZG + (I-RV-1)X(X’V-1X)-1X’V-1ZG = ZG- X(X’V-1X)-1X’V-1ZG – ZG + RV-1ZG + X(X’V-1X)-1X’V-1ZG
- RV-1X(X’V-1X)-1X’V-1ZG = RV-1ZG - RV-1X(X’V-1X)-1X’V-1ZG

78
3º passo: desdobrar X’R-1Be.
X’R-1Be = X’R-1(RV-1ZG – RV-1 X(X’V-1X)-1X’V-1ZG) = X’V-1ZG – X’V-1X(X’V-1X)-1X’V-1ZG = X’V-1ZG – X’V-1ZG = 0 (c.q.d.)
4º passo: desdobrar Z’R-1Be-G-1Bu.
Z’R-1Be-G-1Bu = Z’R-1RV-1ZG – Z’R-1RV-1X(X’V-1X)-1X’V-1ZG – G-1GZ’V-1ZG + G-1GZ’V-1X(X’V-1X)-1X’V-1ZG
= Z’V-1ZG – Z’V-1X(X’V-1X)-1X’V-1ZG – Z’V-1ZG + Z’V-1X(X’V-1X)-1X’V-1ZG = 0 (c.q.d.)
Seguindo os resultados do 3º e 4º passos, as equações (1) se reduzem a:
====
++++++++
−−−−
−−−−
−−−−
−−−−−−−−
−−−−−−−−−−−−
−−−−−−−−
yR'ByR'ZyR'X
tub
BR'BBR'B00
0GZR'ZXR'Z0ZR'XXR'X
1e
1
1
u1
ue1
e
111
11
.
Related Documents











