QUANTITATIVE MODELING OF GENETIC CIRCUITS INTEGRATING TRANSCRIPTIONAL AND S RNA MEDIATED REGULATIONS Matteo Brilli INRIA -RHONE-ALPES LBBE UMR CNRS 5558 UNIV-LYON1 Trento November 27, 2012 (INRIA, CNRS, UNIV-LYON1) MODELING INTEGRATED GENETIC CIRCUITS NOVEMBER 27, 2012 1 / 31

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

QUANTITATIVE MODELING OF GENETIC CIRCUITS
INTEGRATING TRANSCRIPTIONAL AND SRNAMEDIATED REGULATIONS
Matteo Brilli
INRIA - RHONE-ALPESLBBE
UMR CNRS 5558UNIV-LYON1
Trento November 27, 2012
(INRIA, CNRS, UNIV-LYON1) MODELING INTEGRATED GENETIC CIRCUITS NOVEMBER 27, 2012 1 / 31

TOC
1 INTRODUCTION
2 EXAMPLES
3 BASICS OF MATHEMATICAL MODELING
4 NETWORK MOTIFS AND THEIR DYNAMICAL PROPERTIES
5 MODELING SRNA REGULATIONDynamical properties of sRNA-transcription integrated circuits
(INRIA, CNRS, UNIV-LYON1) MODELING INTEGRATED GENETIC CIRCUITS NOVEMBER 27, 2012 2 / 31

INTRODUCTION
INTRODUCTION
GENERAL FEATURES
1 sRNAs are today recognized as pivotal post-transcriptional regulators;
2 size ranges from 50 to a few hundreds nucleotides;
3 the majority modulate gene expression by direct base-pairing with targetmRNA;
4 regulation is predominantly negative;
5 increasing evidence of multiple targets per sRNA.
(INRIA, CNRS, UNIV-LYON1) MODELING INTEGRATED GENETIC CIRCUITS NOVEMBER 27, 2012 3 / 31

INTRODUCTION
WIDESPREAD OCCURRENCE
(INRIA, CNRS, UNIV-LYON1) MODELING INTEGRATED GENETIC CIRCUITS NOVEMBER 27, 2012 4 / 31

INTRODUCTION
FUNDAMENTAL ROLES IN PATHOGENESIS
FIGURE 1: Gopel2011a
(INRIA, CNRS, UNIV-LYON1) MODELING INTEGRATED GENETIC CIRCUITS NOVEMBER 27, 2012 5 / 31

INTRODUCTION
MAIN ROLES IN E. coli
FIGURE 2: Predictions from Modi2011.
A ROLE IN..Mainly stress andenvironmental relatedfunctions.
(INRIA, CNRS, UNIV-LYON1) MODELING INTEGRATED GENETIC CIRCUITS NOVEMBER 27, 2012 6 / 31

INTRODUCTION
FULLY INTEGRATED WITHIN THE GENE REGULATORY
NETWORK
FIGURE 3: sRNA are often regulated by specific transcription factors (TF) and often regulate TFsStorz2011
(INRIA, CNRS, UNIV-LYON1) MODELING INTEGRATED GENETIC CIRCUITS NOVEMBER 27, 2012 7 / 31

INTRODUCTION
MOST COMMON MECHANISMS OF ACTION
FIGURE 4: Waters2009
DIFFERENT MODES OF ACTION
1 Block translation (often bind theShine-Dalgarno) and increasedegradation;
2 Increase mRNA degradation;
3 Promote transcription termination;
4 Increase translation rate by removinginhibitory secondary structures;
5 Act in stoichiometric fashion(degraded with target);
6 Often in conjunction with Hfq.
(INRIA, CNRS, UNIV-LYON1) MODELING INTEGRATED GENETIC CIRCUITS NOVEMBER 27, 2012 8 / 31

INTRODUCTION
HFQ
FIGURE 5: Storz2011
HFQ
1 Interacts with both thesRNA and the targetmRNA;
2 Interacts with the RNAdegradosome;
3 Affects the translationand turnover rates ofspecific transcripts;
4 Distant homologues inArchaea and Eukaryotes.
(INRIA, CNRS, UNIV-LYON1) MODELING INTEGRATED GENETIC CIRCUITS NOVEMBER 27, 2012 9 / 31

INTRODUCTION
HFQ
FIGURE 6: Chao2010
(INRIA, CNRS, UNIV-LYON1) MODELING INTEGRATED GENETIC CIRCUITS NOVEMBER 27, 2012 10 / 31

INTRODUCTION
SHORT SUMMARY FOR EUKARYOTES
FIGURE 7: RNA regulation in Eukaryotes Kim2005
• Different types ofsmall RNAs;
• Different helperproteins/proteincomplexes;
• Pre-processing;• Act catalytically.
(INRIA, CNRS, UNIV-LYON1) MODELING INTEGRATED GENETIC CIRCUITS NOVEMBER 27, 2012 11 / 31

EXAMPLES
RYHB AND IRON HOMEOSTASIS
FIGURE 8: Ferrous iron (Fe2+) is essential but itbecomes toxic in the presence of normal respiratoryby-products (H2O2):→ finely controlledhomeostasis; Salvail2012
UNDER IRON STARVATION...RyhB is a master regulator of ironhomestasis:
1 stimulates the degradationof ∼ 18 mRNAs encodingFe-proteins;
2 feedbacks on Fur;
3 promotes siderophoreproduction e.g. activatingshiA mRNA translation;
(INRIA, CNRS, UNIV-LYON1) MODELING INTEGRATED GENETIC CIRCUITS NOVEMBER 27, 2012 12 / 31

EXAMPLES
QRR AND QUORUM SENSING REGULATION IN Vibrio
Quorum-sensing: regulation of geneexpression in response to cell density;it allows to track population density,synchronize gene expression on apopulation-wide scale, and therebycarry out collective activities.
FIGURE 9: Fenley2011
DIFFERENT ARRANGEMENTS→ DIFFERENTPHENOTYPES
1 V. harveyi produces and monitors theconcentrations of 3 autoinducers (AI), V.cholerae produces and monitors 2 AIs;
2 AI-1 and AI-2 act additively in V. harvey, butredundantly in V. cholerae;
3 ∆luxU: always bright (density-independent)in V. harveyi but not in V. cholerae.
4 ∆ sensor kinases (e.g. cqsS and luxQ)changed the luminescence phenotype in V.harveyi but not in V. cholerae.
(INRIA, CNRS, UNIV-LYON1) MODELING INTEGRATED GENETIC CIRCUITS NOVEMBER 27, 2012 13 / 31
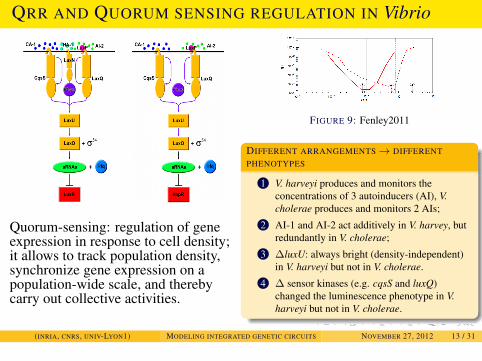
EXAMPLES
QRR AND QUORUM SENSING REGULATION IN Vibrio
Quorum-sensing: regulation of geneexpression in response to cell density;it allows to track population density,synchronize gene expression on apopulation-wide scale, and therebycarry out collective activities.
FIGURE 9: Fenley2011
DIFFERENT ARRANGEMENTS→ DIFFERENTPHENOTYPES
1 V. harveyi produces and monitors theconcentrations of 3 autoinducers (AI), V.cholerae produces and monitors 2 AIs;
2 AI-1 and AI-2 act additively in V. harvey, butredundantly in V. cholerae;
3 ∆luxU: always bright (density-independent)in V. harveyi but not in V. cholerae.
4 ∆ sensor kinases (e.g. cqsS and luxQ)changed the luminescence phenotype in V.harveyi but not in V. cholerae.
(INRIA, CNRS, UNIV-LYON1) MODELING INTEGRATED GENETIC CIRCUITS NOVEMBER 27, 2012 13 / 31

EXAMPLES
QRR AND QUORUM SENSING REGULATION IN Vibrioharvey
FIGURE 10: Input-output relation for the WT and mutated genetic circuits of
quorum-sensing in V. harvey. [Teng2011].
Different strains with one or moreregulatory feedback destroyed and singlecell fluorescence measurements as afunction of AI-1 and AI-2 concentrations.
RESULTS
1 Feedback into LuxN allows V.harvey to actively adjust its relativesensitivity to AI signals as cellstransition from low to high celldensities;
2 The other feedback loops controlthe input and output dynamicranges and the noise in the circuit.
(INRIA, CNRS, UNIV-LYON1) MODELING INTEGRATED GENETIC CIRCUITS NOVEMBER 27, 2012 14 / 31

EXAMPLES
VIRULENCE REGULATION IN Clostridium perfringens
FIGURE 11: [Frandi2010]
HERE COMES THE QUESTION...How to study the different regulatory schemes?
(INRIA, CNRS, UNIV-LYON1) MODELING INTEGRATED GENETIC CIRCUITS NOVEMBER 27, 2012 15 / 31

EXAMPLES
VIRULENCE REGULATION IN Clostridium perfringens
FIGURE 11: [Frandi2010]
HERE COMES THE QUESTION...How to study the different regulatory schemes?
(INRIA, CNRS, UNIV-LYON1) MODELING INTEGRATED GENETIC CIRCUITS NOVEMBER 27, 2012 15 / 31

EXAMPLES
COMPARATIVE SYSTEMS BIOLOGY
Sinorhizobium meliloti
Caulobacter crescentus
Rhodobacter sphaeroides
CckA-RD
CckA-HK~P
CckA-RD~P
CckA-HK
CtrA
ChpT~P
CtrA~P
ChpT
CpdR~PCpdR
proteolysis(ClpX)
Phos
phor
elay
DivK~P
DivK
DivJ
DivJ~P
PleC
PleC + Pi
CckA-RD
CckA-HK~P
CckA-RD~P
CckA-HK
CtrA
ChpT~P
CtrA~P
ChpT
CpdR~PCpdR
proteolysis(ClpX)
Phos
phor
elay
CtrA GcrA DivK~P
DivK~P
DivK
DivJ
DivJ~P
PleC
PleC + Pi
CckA-RD
CckA-HK~P
CckA-RD~P
CckA-HK
CtrA
ChpT~P
CtrA~P
ChpT
CpdR~PCpdR
proteolysis(ClpX)
Phos
phor
elay
CtrA GcrA DivK~P
CtrA GcrA
Modeled circuit Dynamics
FIGURE 12: Studying the phenotypes of different regulatory circuits indifferent organisms.
1 Reconstruct circuit indifferent species;
2 Build correspondingmathematical models;
3 Compare dynamicalbehaviors;
4 Similarly: mutate thesame circuits andexplore how itsproperties change.
(INRIA, CNRS, UNIV-LYON1) MODELING INTEGRATED GENETIC CIRCUITS NOVEMBER 27, 2012 16 / 31

BASICS OF MATHEMATICAL MODELING
MODELING GENE REGULATORY NETWORKS
TF
target
Cons-tu-ve synthesis
Regulated synthesis Degrada-on and dilu-on
A) Promoter activity:
• Positive regulation: A = h+(TF, θ, n) = TFnθn+TFn ;
• Negative regulation: A = h−(TF, θ, n) = 1− h+(TF, θ, n);• Combinatorial regulation: e.g.
for an AND logic: AAND =N∏
i=1
Ai;
for OR logic: AOR =N∑
i=1
Ai .
B) Protein degradation rate γ;
C) Dilution µ.
1 2 3 4 5
0
0.2
0.4
0.6
0.8
1
x
y=
xn
xn+
!n
Positive Hill
1 2 3 4 5
0
0.2
0.4
0.6
0.8
1
x
y=
!n
xn+
!n
Negative Hill
n=1n=2n=3n=4n=5n=6
(INRIA, CNRS, UNIV-LYON1) MODELING INTEGRATED GENETIC CIRCUITS NOVEMBER 27, 2012 17 / 31

NETWORK MOTIFS AND THEIR DYNAMICAL PROPERTIES
NETWORK MOTIFS
FIGURE 13: Feed Forward Loops [Shoval2010].
SUMMARY ON NETWORK MOTIFS
• Transcription regulation andsignaling networks are composedof recurring patterns called networkmotifs;
• Network motifs are much moreabundant in biological networksthan would be expected by theirrandomized versions;
• The same small set of networkmotifs has been found frombacteria to plants to humans;
(INRIA, CNRS, UNIV-LYON1) MODELING INTEGRATED GENETIC CIRCUITS NOVEMBER 27, 2012 18 / 31

NETWORK MOTIFS AND THEIR DYNAMICAL PROPERTIES
MODELING THE FEEDFORWARD LOOP
-1-1
I-1
SYSTEM OF EQUATIONS FOR THE FFLC-1 AND
dYdt
= By︸︷︷︸Basal
+
Regulated synthesis︷ ︸︸ ︷κY
Xn
θnXY + Xn
− (γ + µ)︸ ︷︷ ︸Degr. and dil.
Y (1)
dZdt
= Bz + κZXn
θnXZ + Xn
Yn
θnYZ + Yn︸ ︷︷ ︸
AND logic
−(γ + µ)Z. (2)
Let’s put:α = γ + µ.
SYSTEM OF EQUATIONS FOR THE FFLI-1 ANDdYdt
= By + κYXn
θnXY + Xn
− αY (3)
dZdt
= Bz + κZXn
θnXZ + Xn
θnYZ
θnYZ + Yn
− αZ. (4)
(INRIA, CNRS, UNIV-LYON1) MODELING INTEGRATED GENETIC CIRCUITS NOVEMBER 27, 2012 19 / 31

NETWORK MOTIFS AND THEIR DYNAMICAL PROPERTIES
NETWORK MOTIFS HAVE SPECIFIC DYNAMICAL
PROPERTIES
0 5 10 15 20 25 30 35 400
0.2
0.4
0.6
0.8
1
Z(Y
)
FFLC-1 Vs Simple regulation
9.5 10 10.5 11 11.5 120
0.2
0.4
0.6
0.8
1
0 5 10 15 20 25 30 35 400
0.2
0.4
0.6
0.8
1
time
Z(Y
)
FFLI-1 Vs Simple regulation
9.5 10 10.5 11 11.5 120
0.2
0.4
0.6
0.8
1
time
Z FFLC−1Y FFLC−1Z SimpleY Simple
Z FFLI−1Y FFLI−1Z SimpleY Simple
FIGURE 14: [Shen-Orr2002,Mangan2003,Mangan2003a]
FFLC-1-1-1
• Sign-sensitive delay;• Persistence detector;• Noise reduction;
FFLI-1I-1
• Sign-sensitiveaccelerator;
• other incoherent FFL aregood pulsers;
(INRIA, CNRS, UNIV-LYON1) MODELING INTEGRATED GENETIC CIRCUITS NOVEMBER 27, 2012 20 / 31

MODELING SRNA REGULATION
INTRODUCING SRNA-MEDIATED REGULATION
FIGURE 15:
• sRNAs act stoichiometrically;• sRNAs affect mRNA
stability/translation;• most regulations are negative;
EQUATIONS OF THE SYSTEM
ds
dt= Ss −
Degr. and dil.︷︸︸︷αs − kms︸︷︷︸
Degr.complex
(5)
dm
dt= Sm − αm− kms. (6)
Note that this is the basic modeling framework for all works published on sRNA regulation e.g. from [Levine2007] on.
(INRIA, CNRS, UNIV-LYON1) MODELING INTEGRATED GENETIC CIRCUITS NOVEMBER 27, 2012 21 / 31

MODELING SRNA REGULATION
BUT...
THIS IS AN APPROXIMATIONThe [m] and [s] concentrations are in the same order of magnitude→ the complex should not be neglected. We can overcome this limitationby using a saturation function telling which is the complexed fraction of the total form at steady state ( dx
dt = 0):
YA =AB
Atot=
Atot − Afree
Atot. (7)
ms = mtot − mfree, (8)
stot = sfree + ms, (9)
andms =
mfree · sfree
Kd, (10)
where Kd is the dissociation constant for the complex formation; the free quantities are unknown, the tot are known. After some math we getthe final form of the saturation function:
Ym =mtot + stot + Kd −
√(stot − mtot + Kd)
2 + 4Kdmtot
2mtot(11)
Using this approach we can simply model the control by the sRNA in the following way:
dmtot
dt= Sm −
degr. free form︷ ︸︸ ︷γ · (mtot − Ym · mtot)−
degr. complex︷ ︸︸ ︷γ2 · Ym · mtot −µ · mtot. (12)
(INRIA, CNRS, UNIV-LYON1) MODELING INTEGRATED GENETIC CIRCUITS NOVEMBER 27, 2012 22 / 31

MODELING SRNA REGULATIONDYNAMICAL PROPERTIES OF SRNA-TRANSCRIPTION INTEGRATED
CIRCUITS
THRESHOLD LINEAR RESPONSE
0 0.2 0.4 0.6 0.8 1 1.2 1.4 1.6 1.8 20
1
2
3
4
5
6
7
mRNA synthesis rate.
[m
RN
A]S
S
plus: Levine et al., 2007 modelcircles: Our modificated model
other parameters are:m=0.2
s=0.1k=3.5354e 01Kd=0.01
A THRESHOLD LINEARRESPONSE:
• When αm � αs,translatable targetmRNA is very small;
• When αm � αs, thetarget mRNA starts to beavailable for translation.
(INRIA, CNRS, UNIV-LYON1) MODELING INTEGRATED GENETIC CIRCUITS NOVEMBER 27, 2012 23 / 31

MODELING SRNA REGULATIONDYNAMICAL PROPERTIES OF SRNA-TRANSCRIPTION INTEGRATED
CIRCUITS
THRESHOLD LINEAR RESPONSE
0.2 0.4 0.6 0.8 10
1
2
3
4
5
6
7
8
m
X SS
=0.5
=1.0
=4.0=2.0
FIGURE 16:
0 0.2 0.4 0.6 0.8 10
0.05
0.1
0.15
0.2
m SS / µ m SS
m
FIGURE 17:
A THRESHOLD LINEAR RESPONSE:
• The threshold depends only onthe two transcription rates;
• The smoothness of the transitiondepends on the degradation rateof the complex (half-life given bylog2γ
h);
• Around the crossover regiondifferences in steady state levelsof the target are much moredependent on the interactionstrength Kd .
• This gives an easy way forordering the expression ofdifferent genes by tuning αm andKd .
(INRIA, CNRS, UNIV-LYON1) MODELING INTEGRATED GENETIC CIRCUITS NOVEMBER 27, 2012 24 / 31

MODELING SRNA REGULATIONDYNAMICAL PROPERTIES OF SRNA-TRANSCRIPTION INTEGRATED
CIRCUITS
SRNA CAN PROVIDE A SWITCH-LIKE BEHAVIOR
10−2 10−1 100 10110−6
10−5
10−4
10−3
10−2
10−1
100
_s / _m
[ta
rge
t
mR
NA
]
Ultrasensitivity
a=10a=100a=1000a=10000
FIGURE 18: Ultrasensitive switch Mitarai2009.
SLIGHTLY DIFFERENT...
but comparable model from Mitarai2009:
ds
dt= α− s− γsm (13)
dm
dt= 1−
m
τ− γsm. (14)
where: α = αsαm
the relative transcription rate of s withrespect to that of m. γ = δαmτs quantifies theinactivation of the mRNA via sRNA:mRNA complexformation, and τ = τm
τsis the ratio of the mRNA
lifetime to the sRNA lifetime.
(INRIA, CNRS, UNIV-LYON1) MODELING INTEGRATED GENETIC CIRCUITS NOVEMBER 27, 2012 25 / 31

MODELING SRNA REGULATIONDYNAMICAL PROPERTIES OF SRNA-TRANSCRIPTION INTEGRATED
CIRCUITS
TARGET PRIORITIZATION
0 0.5 1 1.50
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
sRNA synthesis rate
norm
alized
steady
state
[mR
NA
]
sRNA mediated regulation
0 0.5 1 1.50
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
Repressor synthesis rate
norm
alized
steady
state
[mR
NA
]
Transcriptional repression
PRIORITIZATION
• Multistep switch-like degradationupon changing the rate ofsynthesis of the sRNA;
• Separation of the level betweenthe (k + 1)th mRNA and the kth
mRNA becomes clearerincreasing the difference in thedegradation rate.
• Separation between mRNAlevels following transcriptionalregulation is not as sharp as withsRNA regulation;
• prioritization through sRNA ismuch more effective especiallyfor small changes in the sRNAsynthesis rate.
(INRIA, CNRS, UNIV-LYON1) MODELING INTEGRATED GENETIC CIRCUITS NOVEMBER 27, 2012 26 / 31

MODELING SRNA REGULATIONDYNAMICAL PROPERTIES OF SRNA-TRANSCRIPTION INTEGRATED
CIRCUITS
BISTABILITY
sRNA
TF
THE MODEL
dx
dt=
Translation︷︸︸︷γy − δx︸︷︷︸
Degr.
(15)
dy
dt=
Synthesis︷ ︸︸ ︷λSy(x, p)−αy− σyz (16)
dz
dt= µSz(x, p)− βz− σyz. (17)
Where: x=protein TF, y=mRNA TF, z=sRNA. [Liu2011].
• Bistability: the capacity to achieve two alternative internal states in response to differentstimuli;
• ubiquitous in cellular systems;
• bistability is generated by regulatory interactions;
• fundamental biological significance e.g. cell differentiation, cell fate decision, adaptiveresponse to environmental stimuli, regulation of cell cycle oscillations and so on.
• switches involving ncRNA have been recently studied experimentally[Bumgarner2009,Iliopulos2009] and theoretically [Zhdanov2009].
(INRIA, CNRS, UNIV-LYON1) MODELING INTEGRATED GENETIC CIRCUITS NOVEMBER 27, 2012 27 / 31

MODELING SRNA REGULATIONDYNAMICAL PROPERTIES OF SRNA-TRANSCRIPTION INTEGRATED
CIRCUITS
BISTABILITY
THE MODEL
dx
dt=
Translation︷︸︸︷γy − δx︸︷︷︸
Degr.
(18)
dy
dt=
Synthesis︷ ︸︸ ︷λSy(x, p)−αy− σyz (19)
dz
dt= µSz(x, p)− βz− σyz. (20)
Where: x=protein TF, y=mRNA TF, z=sRNA.[Liu2011].
• bistability in this case only for intermediate association rates between sRNA and mRNA;
• In the monostable regimen lower degradation rates correspond to higher protein level and vice versa. On the converse, when theassociation rate is between A and B (the saddle points) the opposite can be true, depending on the initial conditions.
• the noise inherent in biological systems may induce switching between the two stable states.
(INRIA, CNRS, UNIV-LYON1) MODELING INTEGRATED GENETIC CIRCUITS NOVEMBER 27, 2012 28 / 31

MODELING SRNA REGULATIONDYNAMICAL PROPERTIES OF SRNA-TRANSCRIPTION INTEGRATED
CIRCUITS
CONCLUSIONS
• sRNAs provide an efficient regulatory mechanism;• integration of transcriptional and sRNA mediated regulations generates a
wide variety of interesting dynamical behaviors, such as threshold linearresponse, prioritization of targets and even bistability (and oscillations);
• modeling genetic circuits may provide information on both functionalityand evolution of genetic circuits.
(INRIA, CNRS, UNIV-LYON1) MODELING INTEGRATED GENETIC CIRCUITS NOVEMBER 27, 2012 29 / 31

MODELING SRNA REGULATIONDYNAMICAL PROPERTIES OF SRNA-TRANSCRIPTION INTEGRATED
CIRCUITS
COLLABORATORS AND ACKNOWLEDGEMENTS
A SPECIAL THANKS TO
Equipe Baobab (MF Sagot),LBBE, INRIA; Daniel Kahn(LBBE, INRA).
AND FOR YOUR ATTENTION,Thank you!
contact me: [email protected]
By the way...looking for a dog? 6 Labrador Chocolate available, contact me!
(INRIA, CNRS, UNIV-LYON1) MODELING INTEGRATED GENETIC CIRCUITS NOVEMBER 27, 2012 30 / 31

APPENDIX BIBLIOGRAPHY
(INRIA, CNRS, UNIV-LYON1) MODELING INTEGRATED GENETIC CIRCUITS NOVEMBER 27, 2012 31 / 31
Related Documents











