Modeling and Analysis of Radial Flow Mammalian Cell Culture John P. Tharakan and Pao C. Chau Department of AMES (Chemical Engineering), University of California, San Diego, La Jolla, California 92093 Accepted for publication September 2, 1986 A model for radial flow mammalian cell culture has been developed. Two situations, one with permeable hollow fibers and the other with a porous matrix in the annulus, were considered. The hollow fibers were modeled as con- tinuously distributed sinks. Numerical solutions are pre- sented for the complete model as well as limiting ana- lytical solutions. The analysis identified the importance of various kinetic, transport, and design dimensionless groups for maintenance of radial flow cell culture sys- tems under uniform conditions. The important design parameter was the depth of the bed and the important operating parameter was a modified Damkohler number, both of which should be maintained low for gradient free systems. Dispersion was included in the analysis but sub- strate consumption was relatively independent of dis- persion. Preliminary separation of a low- and high-mo- lecular-weight product was also modeled, and shown to be strongly dependent on the permeability of the fibers, as well as the aspect ratio and the magnitude of the trans- membrane flux. INTRODUCTION Process development for large-scale mammalian cell cultures is a topic of current interest.' An important step in understanding cell culture systems lies in the development of engineering models, enabling one to design and analyze culture configurations for the pro- duction of useful biochemicals. The present study is directed at understanding the transport, kinetic, and design parameters that play a significant role in the distribution of nutrient and prod- uct in radial flow mammalian cell culture bioreactors for the large-scale culture of mammalian cells.2 In the analysis, both hollow fiber and nonpermeable polymer matrix supports are investigated. Hollow fiber system^^,^ offer several advantages, which include high cell den- sities, reduced shear fields for shear sensitive cells, the ability to maintain a three-dimensional cell matrix, pre- liminary product separation, and decreased risks of contamination. Crossflow systems offer additional ad- vantages, both in flat bed5 and radial flow2 configura- tions, in the ability to minimize nutrient gradients and to enable oxygen transfer in the culture system.6 Biotechnology and Bioengineering, Vol. XXIX, Pp. 657-671 (1987) 0 1987 John Wiley & Sons, Inc. A radial flow catalytic reactor is a special case of a crossflow reactor. It was developed to handle large gas flow rates with minimal pressure drop.'** The first an- alytical work on radial flow catalytic reactors was con- ducted by Genkin and co-w~rkers,~ who studied the distribution of feed gas streams and radial velocity pro- files for centripetal and centrifugal flows. Their work was extended by Kaye.'O Isothermal fixed bed radial reactors were described by Hlavacek and Kublicek. I1 Dudukovic and Lamba12obtained analytical solutions for first-order conversions and used numerical solu- tions for reactions of order other than one. Ponzi and KayeI3 studied the effect of flow maldistribution on conversion and selectivity and this work was extended by Chang and Calo14who analyzed the effects of flow direction on conversion and confirmed asymptotic re- sults using numerical experiments. Recently, Yo0 and Dixon" imposed the effects of volume change, heat transfer, flow maldistribution, and complex kinetics in a numerical analysis. Radial flow is also employed in reverse osmosis where large volumes of liquid need to be processed. The anal- ysis of hollow fiber reverse osmosis units was studied by Gill and co-workers.1620 Both experimental and theoretical results were used to establish design pa- rameters and optimal operation conditions to maximize the efficiency of radial flow hollow fiber reverse os- mosis units. A common approach in analyzing radial catalytic reactors has been to assume an inlet profile and in- vestigate the effect of flow direction (centripetal or centrifugal) on conver~ion.'~ In addition, studies of both radial reverse osmosis units and radial catalytic reac- tors have usually assumed that convective effects pre- dominate over dispersion. Kinetics has also been lim- ited to the traditional cases of first or second order, and the aim in catalytic reactor design is to maximize conversion. The design of cell culture systems is very different. As a consequence of using low superficial velocities to minimize shear fields, the ratio of disper- sion and convection terms becomes close to unity and CCC 0006-3592/87/060657-15$04.00

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Modeling and Analysis of Radial Flow Mammalian Cell Culture
John P. Tharakan and Pao C. Chau Department of AMES (Chemical Engineering), University of California, San Diego, La Jolla, California 92093
Accepted for publication September 2, 1986
A model for radial flow mammalian cell culture has been developed. Two situations, one with permeable hollow fibers and the other with a porous matrix in the annulus, were considered. The hollow fibers were modeled as con- tinuously distributed sinks. Numerical solutions are pre- sented for the complete model as well as limiting ana- lytical solutions. The analysis identified the importance of various kinetic, transport, and design dimensionless groups for maintenance of radial flow cell culture sys- tems under uniform conditions. The important design parameter was the depth of the bed and the important operating parameter was a modified Damkohler number, both of which should be maintained low for gradient free systems. Dispersion was included in the analysis but sub- strate consumption was relatively independent of dis- persion. Preliminary separation of a low- and high-mo- lecular-weight product was also modeled, and shown to be strongly dependent on the permeability of the fibers, as well as the aspect ratio and the magnitude of the trans- membrane flux.
INTRODUCTION
Process development for large-scale mammalian cell cultures is a topic of current interest.' An important step in understanding cell culture systems lies in the development of engineering models, enabling one to design and analyze culture configurations for the pro- duction of useful biochemicals.
The present study is directed at understanding the transport, kinetic, and design parameters that play a significant role in the distribution of nutrient and prod- uct in radial flow mammalian cell culture bioreactors for the large-scale culture of mammalian cells.2 In the analysis, both hollow fiber and nonpermeable polymer matrix supports are investigated. Hollow fiber system^^,^ offer several advantages, which include high cell den- sities, reduced shear fields for shear sensitive cells, the ability to maintain a three-dimensional cell matrix, pre- liminary product separation, and decreased risks of contamination. Crossflow systems offer additional ad- vantages, both in flat bed5 and radial flow2 configura- tions, in the ability to minimize nutrient gradients and to enable oxygen transfer in the culture system.6
Biotechnology and Bioengineering, Vol. XXIX, Pp. 657-671 (1987) 0 1987 John Wiley & Sons, Inc.
A radial flow catalytic reactor is a special case of a crossflow reactor. It was developed to handle large gas flow rates with minimal pressure drop.'** The first an- alytical work on radial flow catalytic reactors was con- ducted by Genkin and co -w~rke r s ,~ who studied the distribution of feed gas streams and radial velocity pro- files for centripetal and centrifugal flows. Their work was extended by Kaye.'O Isothermal fixed bed radial reactors were described by Hlavacek and Kublicek. I 1
Dudukovic and Lamba12 obtained analytical solutions for first-order conversions and used numerical solu- tions for reactions of order other than one. Ponzi and KayeI3 studied the effect of flow maldistribution on conversion and selectivity and this work was extended by Chang and Calo14 who analyzed the effects of flow direction on conversion and confirmed asymptotic re- sults using numerical experiments. Recently, Yo0 and Dixon" imposed the effects of volume change, heat transfer, flow maldistribution, and complex kinetics in a numerical analysis.
Radial flow is also employed in reverse osmosis where large volumes of liquid need to be processed. The anal- ysis of hollow fiber reverse osmosis units was studied by Gill and co-workers.1620 Both experimental and theoretical results were used to establish design pa- rameters and optimal operation conditions to maximize the efficiency of radial flow hollow fiber reverse os- mosis units.
A common approach in analyzing radial catalytic reactors has been to assume an inlet profile and in- vestigate the effect of flow direction (centripetal or centrifugal) on conver~ion.'~ In addition, studies of both radial reverse osmosis units and radial catalytic reac- tors have usually assumed that convective effects pre- dominate over dispersion. Kinetics has also been lim- ited to the traditional cases of first or second order, and the aim in catalytic reactor design is to maximize conversion. The design of cell culture systems is very different. As a consequence of using low superficial velocities to minimize shear fields, the ratio of disper- sion and convection terms becomes close to unity and
CCC 0006-3592/87/060657- 15$04.00

both transport phenomena need to be considered in a complete analysis. Kinetics in biological systems are nonlinear, and the goal is to minimize gradients of nu- trient, which is the opposite of maximizing conversion, to provide a more uniform environment in v i m for cell propagation.
In this study, we consider a radial flow bioreactor operating in a continuous perfusion mode and a sche- matic is shown in Figure 1 . The annular cell growth region may be occupied by a hollow fiber bundle or a porous polymer matrix support. The polymer matrix is considered to be nonpermeable. The analysis is de- veloped assuming the presence of semipermeable hol- low fibers with the special case of zero wall flux rep- resenting the polymer matrix. Cell mass is considered to be distributed uniformly in the shell space of the bioreactor. With hollow fiber systems, the fiber lumen is open at both ends to the drain ports, and the pressure in the lumen of the fibers may be maintained at ambient pressure. The liquid nutrient enters the inner annulus and can exit through either of two means. The fluid can diffuse across the fiber membrane wall and exit via the lumen of the fibers. Any residual fluid, mostly con- taining high molecular weight components rejected by the fibers, will exit via the outer annulus. In the con- tinuity equation, the flux of fluid through the fiber wall is modeled as a homogeneous sink.” In the analysis which follows, we shall also provide the limiting case in which the flux through the fiber wall is zero. This situation represents examples where the hollow fiber bundle is replaced by a porous polymer matrix that is useful in cell entrapment bioreactors.
In addition to substrate consumption, we also con- sider the formation of two products with very different molecular weights. One represents a metabolic waste product such as lactic acid, with a molecular weight around 100 D, while the other is a protein product with a molecular weight above lo4, and in the case of mono- clonal antibodies would be above 1.5 x lo5. Typically, the protein products are produced at very low con- centrations. For example, monoclonal antibodies are produced at a rate 1 pg/lo6 cells h in suspension cul- ture. Any initial purification step which can be per- formed in the bioreactor will facilitate downstream re- covery and purification. The objective of our work is
I L
CELL GROWTH REGION(SHELL SPACE) SHELL EFFLUENT
Figure 1. Schematic representation of radial flow hollow fiber bio- reactor and coordinate system. CFD is the central feed distributor.
to establish an engineering model for radial flow mam- malian cell culture and to evaluate the feasibility of effecting a preliminary separation of protein products.
MODEL DEVELOPMENT
We first state the assumptions and their physical justifications before presenting the balance equations.
1) The bioreactor is axisymmetric with no depen- dence on the angular coordinate.
2) With proper design of the system, especially the feed distributor,2 we can obtain an ideal inlet velocity profile, and the gradients in the radial direction can be much greater than those in the axial direction.
3) The annular bed depth is short and the superficial velocity is low. Thus the radial pressure drop in the annular region is negligible. 4) In typical mammalian cell culture systems, growth
rates are comparatively slow, with doubling times ranging from 20 to 50 h. With these slow growth rates, it is reasonable to assume pseudo-steady-state condi- tions for the cell culture model.
5) The annular cell growth region is considered to be a pseudohomogeneous region. The hollow fibers, which allows for a transmembrane flux to leave the bioreactor, are modeled as continuously distributed sinks. In limiting cases which the hollow fibers are replaced by a nonpermeable polymer supporting ma- trix, there is no transmembrane flux and the sink term is zero.
6) The rate of consumption of the limiting substrate and the formation of the products are described by pseudohomogeneous reaction terms in the annular cell growth region.
7) The individual mass fluxes of substrate and prod- ucts into the lumen of the fibers are related to the liquid flux velocity across the fiber membrane:
CLjU,, = k j (CSi - CLj) (1)
where c is concentration. The subscripts S and L refer to the shell and lumen, and the subscript i is either c , P I , or P2 which denote substrate and products 1 and 2, respectively. In cases which concentration polari- zation may become important, the mass transfer re- sistance across the gel layer can be incorporated into the mass transfer coefficient k j .
8) The velocity of the fluid through the fiber wall can be written as
U w ( r 7 z ) = A,bs(r) - PLG-,Z)I (2) and will be taken to be aconstant in the radial direction. The lumen pressure, p L , can be maintained constant with respect to the radial coordinate, and since we have assumed negligible pressure drop in the shell space, u,, is independent of radial position.
9) Simple unstructured kinetics is used for both cell mass and product formation. The consumption of lim-
658 BIOTECHNOLOGY AND BIOENGINEERING, VOL. 29, APRIL 1987

iting substrate in cell yield is described by the Monod model:
The analysis requires that only one limiting substrate be considered. This substrate is supplied through the inlet feed from the central feed distributor to the cell growth annulus. For practical purposes, either glucose or dissolved oxygen would be considered as the lim- iting substrate.
10) The Leudeking-Piret equation is used to model the formation of products,
R(P;,) = -- dx + bjX L / P i dt
where i = 1, 2 for the two products.
Model Equations
With the preceding assumptions, we may write the continuity equation and the mass balance equations for a substrate and two products in the annular cell growth region. The continuity equation in cylindrical coordinates without angular dependence is
- + - + “ = Q au, u, au ar r a,
where Q represents the sink term and is the volumetric flow rate entering the fiber lumen per unit reactor vol- ume. The mass balance for the substrate and the prod- ucts are
1 d(ru,cs) 1 d(ru,cs) (;) ar + (;) az = D, [$ + (;) l a % (r:)]
= DPi [$ + (;) l a (r?)]
+ QPiL + R(Pjs) (7)
where i = 1 , 2.
to both the inner, R , , and outer annulus, R2, viz.,
at r = R , u, = U,,
The Danckwert’s boundary conditions are applied
and 0 = U,,Pjs - Dpi - ( a::)
At the outlet,
_ - acs - 0 r = R2 (9) ar
- = O ; i = l , 2 apis ar
and
Equations (5)-(7) are integrated over the length of the reactor to give
+ - d r kL u,csdz = D,r dr
I d d r dr dr o
+ - - r - j L csdz] + r J L QcLdz
+ dr r kL u,Pis dz = Dpir {&(L.,r) dz
- -(O,r) dPiS + - - r - I L Pa dz} dz r dr dr o
+ r lL QPiLdz - r I 0 R(Pjs)dz L
(12)
where i = 1 , 2. These equations may be further simplified by real-
izing that u,(L,r) = u,(O,r) = 0, and that the gradients in the axial direction are assumed to be much smaller than those in the radial direction. For example, with the substrate,
dCS dCS -(L,r) = -(O,r) = 0 dz dz
In addition we define length averaged quantities L
CsU, = I u,cs dz (13) L o
- 1 ow = - /o’ u, dz
L
The total number of fibers per unit volume of reactor volume is
N ( 1 - €)
V s mr;L _ - -
THARAKAN AND CHAU: RADIAL FLOW OF MAMMALIAN CELL CULTURE 659

is the wall flux number; where V , is the volume of the annular shell space. The length averaged flow into the fiber can be written asl9
- 2(1 - E ) Q = O W ,
ro
The set of model equations can be rewritten in terms of these lengthwise average quantities. The model equations can further be put in dimensionless form. The velocity is rendered dimensionless by use of the inlet velocity, and the concentration of the substrate by the inlet substrate concentration. The product con- centrations are normalized by a reference concentra- tion based on the inlet substrate and appropriate yield coefficients. The radial coordinate is nondimension- alized by the depth of the annular shell space, p .
x = rlp; V = UJU,,; 8 = CslCso (18)
= PISIPrefl; = pZS/Prefl Y2/1
where Pres = Cs,YpI/c. After eliminating the lumen concentration with eqn.
(l), the system of equations in dimensionless form be- come
( x v ) = a I d x dx --
1 d28 1 d8 Pee (dx2 x dx)
(xve) = - - + -- + apce I d x dx --
and
I d 1 [d2 f l i + Ll] -- (XVfl;) = - - x dx Pep, dx2 x dx
+ Da, e + appifli + DaPjlx -
Y + O
where i = 1,2.
ditions in dimensionless form become:
atx = l /(u-l)
With the aspect ratio, a = R2/RI , the boundary con-
u = 1
1 de 1=8- - - . - Pec dx (22)
i = 1 , 2 1 dfl . 0 = fl. - -2. Pepi dx ’
and
at x = a/(a - 1) d8ldx = 0 (23) dflildx = 0; i = 1 , 2
where
f f = ( 2p(l - €))($)
is the Peclet number for species i; and
ki p. = - ‘ k + u
1 W’
is the fiber permeability number for species i. The sub- script i refers to substrate, c, or either of the two prod- ucts, PI and P2. The quantities Da and m are the mod- ified Damkohler numbers and are defined as follows:
Daclx = (”-.”) YX/CCSO (&) accounts for substrate consumption;
accounts for growth associated product maintenance;
accounts for nongrowth associated product mainte- nance:
accounts for growth associated product formation; and
Dapi = {x} {$-} (31) Prefl yi/l
accounts for nongrowth associated product formation. The Peclet number, pi, and the Damkohler numbers
will depend on the choice of limiting substrate used for the analysis. Table I provides ranges of typical values for the various coefficients in mammalian cell cultures. Kinetic parameters for both glucose and oxygen have been provided. It is important to note that the dimen- sionless analysis will hold for different substrates as long as the dimensionless groups are the same order of magnitude.
The continuity equation (19) provides an analytical solution for the velocity profile, V, which is substituted in the mass balances (20) and (21). Together with the boundary conditions (22) and (23), these equations con- stitute a boundary value problem. Since our goal is to avoid severe nutrient gradients, the shooting method with Gear’s numerical integration is adequate to obtain numerical solutions under circumstances relevant to mammalian cell culture systems. Analytical solutions are available for several limiting cases.
660 BIOTECHNOLOGY AND BIOENGINEERING, VOL. 29, APRIL 1987

Table I. Values of the kinetic and transport coeficients used in evaluating the range of dimensionless groups.
Magnitude Coefficient of Range Unit Reference
1 .o-2.0 1.0-4.5 0.1-0.2 O( 10 -2)
q 1 0 - 3 ) q 1 0 - 3 )
q10-6) O(10-1) q10-2)
0.3 5.0 2.0
O( 10-2) o( 10- 3)
10-1-10-4 o(10-6) I@-108 W . 3 ) O(1.0) 10s- 107 108-10’0 I 0s- 107 O( 1 .O) 0.3 3.0
qlo-2)
pg MAB/106 cells hr mg/mL (glucose) mM Oz/L cm2/s (oxygen) cm2/s (glucose) cm2/s (lactate) cm2/s (Ig) mg/mL (glucose) mM/L (oxygen) mg lactic acid/mL cm cm cm cm cm/s cm/s cells/mL mg lactic acid/mg glucose pg MABlmg glucose cells/mg glucose cells/mM 0, cells/mg lactic acid pg MAB/mg lactic acid dimensionless cm 1 h - 1
25 2 1
22” 22” 22” 22” 2
21b 2 2 2
26 26
2 19 2 2
2 21b
2
2 2
-
-
2,21 ~~~ ~
a Estimated from the correlation provided in ref. 22. Calculated based on raw data in ref. 21.
Limiting Cases for Substrate Profile
The solutions of the limiting cases are provided here. Only substrate concentration profiles are presented.
1) No wall flux (a = 0). The model equations reduce to a pseudohomogeneous reactor in radial coordinates. Plug flow is used in cases which the Peclet number is large.
la) Plug flow and zero-order kinetics with respect to the limiting substrate, 8 & y. The solution to this set of simplified equations is straightforward and is given by
The parameters are the aspect ratio, a and the zero- order Damkohler number for the substrate, D~G,.
lb) Plug flow and first-order kinetics with respect to the limiting substrate, y 9 8. The solution is
The parameters of importance are the first-order Damkohler number, Da,,, the aspect ratio, a, and the dimensionless Monod constant, y. It should be noted
that the reaction rate term is formulated such that the limiting zero and first order cases have different ap- parent rate parameters, and the effect of increasing y is equivalent to inhibition on the substrate consump- tion. Thus the solution of increasing y will not be bound within the zero and first order limits.
lc) Radial dispersion with zero-order dependence on substrate consumption. The solution of the substrate profile is
aA + I - 1
Pec(a - 1) +
Pec(a - 1)
aA+’ 1 + 1 -- 2 ( ~ - 1)’ (a - 1)*(1 -A) (34)
where A = 1 - Pec/(a - l), and Pec is the Peclet num- ber for the substrate.
Id) Radial dispersion with first-order dependence on substrate consumption. The solution to this case can be obtained analytically in terms of modified Bessel functions. The complete solution is
e = x q c l I y ( x ~ ) + C~IL~(XV‘Z)I (35)
THARAKAN AND CHAU: RADIAL FLOW OF MAMMALIAN CELL CULTURE 66 1

where Y is a constant for any one set of parameters and is equal to 2Pec/(a- l), and B is also a constant given by (PecDacl)/y. The two constants C1 and C2 can be evaluated from the boundary conditions and are given as:
] (37) c1 = [c23 [ Iw- ' (avE/a - 1)
I - " + ' ( U v E / a - 1)
2) Permeable fiber wall (a # 0). The sink term in the continuity equation is finite and simple analytical solutions are available only with negligible radial dis- persion (i.e., plug flow conditions).
2a) Plug flow with zero-order dependence on sub- strate consumption. With the inclusion of the wall flux, two additional parameters, a and p, are required. The solution is obtained through the use of an integrating factor:
O = 2(1 -P)+a a-1
2b) Plug flow with first-order dependence on sub- strate consumption. The solution is
e = [(i-u)($) - 1.0
+ ?(('>I ( 2 - 4 + DaJcry) (39) 2 a -1
RESULTS AND DISCUSSION
The results are first presented to illustrate conditions which help to reduce nutrient gradients and ensure a relatively uniform enviroment in the cell culture re- gion. The influence of each dimensionless groups is discussed individually. The solutions for both with (a # 0) and without (a = 0) transmembrane flux in the annular region are provided. The product dis- tributions are then presented to assess the feasibility of removing metabolic waste and improving the selec-
tivity of the high molecular weight product in the shell effluent. Typical magnitudes of various transport and kinetic constants in mammalian cell culture are given in Table I.
Modified Damkohler Number, Dac,x and Monod Constant, y
The modified substrate Damkohler number, Daclx, is a relative measure of the rates of substrate con- sumption to that of bulk convective transport through the cell growth region. At the time of inoculation, the cell mass loading is low, and the magnitude of Daclx is generally very small. For a longer term cell culture, the value of Daclx can increase by two orders of mag- nitude, depending on the number of cell doublings achieved.
The Damkohler number is strongly dependent on the choice of limiting substrate. From the results of Tolbert and Feder,2' a yield coefficient was estimated for oxy- gen uptake by hepatoma cells. The numbers for oxy- gen uptake rate provided by Glacken and co-workers' and numbers from our data were also used to estimate a range for the cell yield based on oxygen. Our own data2 were used to estimate appropriate values for glu- cose based cell yield.
Due to the low solubility of oxygen in culture media at 37°C (0.2mMIL'), the initial achievable concentra- tion is limited by this saturation value. However, the cell yield coefficient per mg glucose is much less than the cell yield/mM oxygen. Thus, for the same inlet superficial velocity, the Damkohler number based on oxygen is usually only an order of magnitude higher than that for glucose. To avoid nutrient limitations, the Damkohler number can be reduced by increasing the crossflow superficial flow rate. For example, with hep- atoma cells, and utilizing calculations based on the data of Tolbert and Feder,2' a Damkohler number of 0.1 for oxygen is possible with a superficial velocity as low as
cm/s and a cell loading as a high as lo8 cells/mL. With this same superficial velocity and cell loading, the Damkohler number for glucose is 0.01. Thus, if oxygen is suspected to be limiting as in the case of a porous polymer matrix in the annulus, the superficial velocity can be increased to reduce its effect.
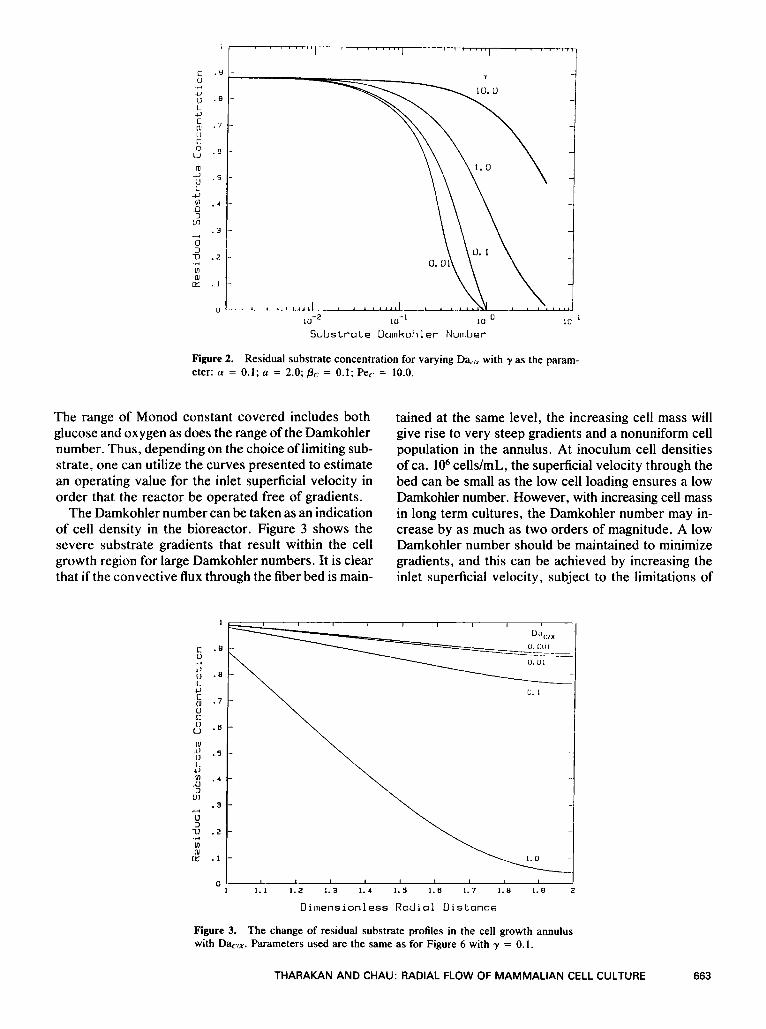
The effect of the Monod constant y can be taken as a measure of the inhibition to substrate utilization. The change of residual substrate concentration with the Damkohler number is shown in Figure 2 for increasing values of y. As y increases, the utilization of substrate decreases. At low Damkohler numbers corresponding to inoculum level cell concentrations, the bioreactor is kinetic rate limited and there is only a gradual de- crease in residual substrate concentration with in- crease in Damkohler number. As the Damkohler num- bers increases, the system becomes transport limited and the amount of residual substrate rapidly decreases.
662 BIOTECHNOLOGY AND BIOENGINEERING, VOL. 29, APRIL 1987
I I I
10-2 10-1 ID 10
Subst ru te Uaiiikohler Nuiiiber
Figure 2. Residual substrate concentration for varying Da,,, with y as the param- eter: a = 0.1; a = 2.0; Pc = 0.1; Pec = 10.0.
The range of Monod constant covered includes both glucose and oxygen as does the range of the Damkohler number. Thus, depending on the choice of limiting sub- strate, one can utilize the curves presented to estimate an operating value for the inlet superficial velocity in order that the reactor be operated free of gradients.
The Damkohler number can be taken as an indication of cell density in the bioreactor. Figure 3 shows the severe substrate gradients that result within the cell growth region for large Damkohler numbers. It is clear that if the convective flux through the fiber bed is main-
tained at the same level, the increasing cell mass will give rise to very steep gradients and a nonuniform cell population in the annulus. At inoculum cell densities of ca. lo6 cells/mL, the superficial velocity through the bed can be small as the low cell loading ensures a low Damkohler number. However, with increasing cell mass in long term cultures, the Damkohler number may in- crease by as much as two orders of magnitude. A low Damkohler number should be maintained to minimize gradients, and this can be achieved by increasing the inlet superficial velocity, subject to the limitations of
1 I I 1 I I I I I I 1 1 .1 1.2 1.3 1.4 1.5 1.6 1.7 1.8 1 .E 2
Dimens ion1 ess Radi a1 Ui stance
Figure 3. The change of residual substrate profiles in the cell growth annulus with Da,=,x. Parameters used are the same as for Figure 6 with y = 0.1.
THARAKAN AND CHAU: RADIAL FLOW OF MAMMALIAN CELL CULTURE 663
I. 4 1 ln . 4 .:
Ul
0 .3
.-I
1; .2 ln IU
LL . l
0
- , F i r s t o r d e r wi th diopers ion
- \<\L - ';\. ..
-
-- -_____ - -___ - /'\, --- --_ F i r e t o r d e r w i t l iout t l i eim-s 1 or1 --. L -
I I I I I --- -- ----__ .--.
I 2 3 4 5 6 I
Aspect R o L i o
1 I I I I I I I
c .. * .I I 1 I.
.).I II (U
c U
0
' I J
ln
LL
.,.a
I I - 1 rst order I
Asliect Ratio
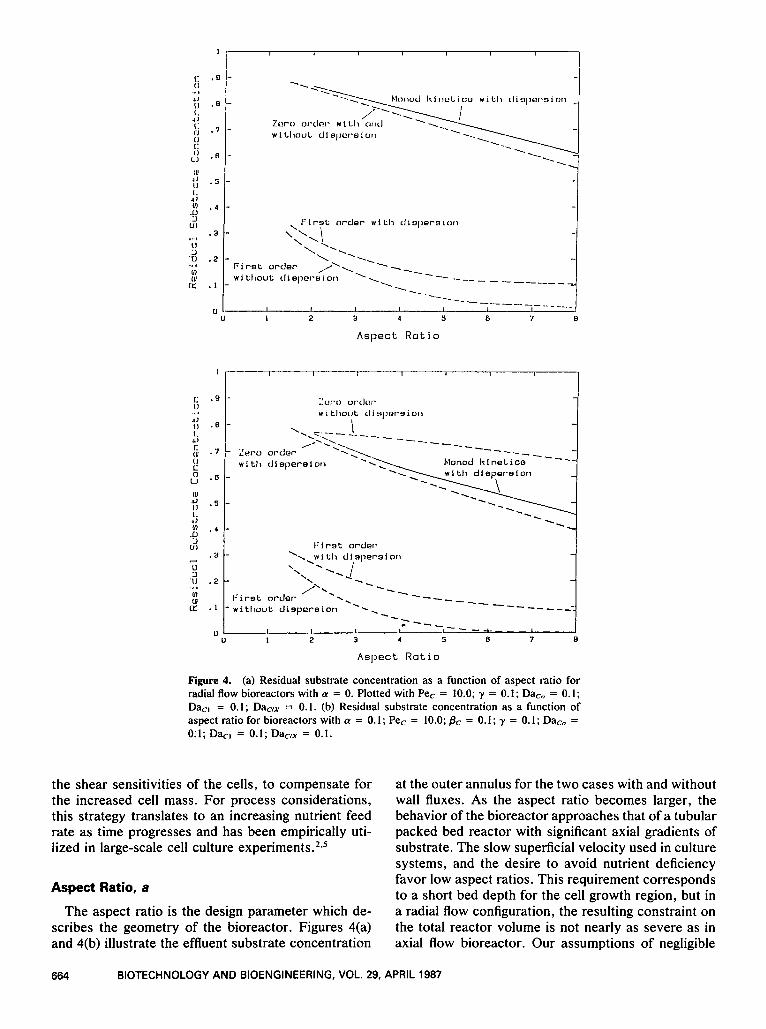
Figure 4. (a) Residual substrate concentration as a function of aspect ratio for radial flow bioreactors with a = 0. Plotted with Pe, = 10.0; y = 0.1; Da, , = 0.1; Dacl = 0.1; D&tX = 0.1. (b) Residual substrate concentration as a function of aspect ratio for bioreactors with a = 0. I ; Pec = 10.0; pc = 0.1 ; y = 0. I ; Da,,, = O i l ; D k l = 0.1; DacIx = 0.1.
the shear sensitivities of the cells, to compensate for the increased cell mass. For process considerations, this strategy translates to an increasing nutrient feed rate as time progresses and has been empirically uti- lized in large-scale cell culture experiment^.^,^
Aspect Ratio, a
The aspect ratio is the design parameter which de- scribes the geometry of the bioreactor. Figures 4(a) and 4(b) illustrate the effluent substrate concentration
at the outer annulus for the two cases with and without wall fluxes. As the aspect ratio becomes larger, the behavior of the bioreactor approaches that of a tubular packed bed reactor with significant axial gradients of substrate. The slow superficial velocity used in culture systems, and the desire to avoid nutrient deficiency favor low aspect ratios. This requirement corresponds to a short bed depth for the cell growth region, but in a radial flow configuration, the resulting constraint on the total reactor volume is not nearly as severe as in axial flow bioreactor. Our assumptions of negligible
664 BIOTECHNOLOGY AND BIOENGINEERING, VOL. 29, APRIL 1987
radial pressure drop and constant wall velocity, v,, also constrain the model to low aspect ratios. From the figures, 60% of the substrate still remains in the annular cell growth region for aspect ratios up to 6 with or without a wall flux. At larger aspect ratios, signifi- cant gradients of nutrients will develop and likely lead to nonuniform cell mass densities. The actual criterion depends on the cellular threshold of the particular lim- iting substrate.
The first order reaction curves represent nutrient starvation conditions which we seek to avoid and thus are not particularly useful in cell culture design. From the typical kinetic constants listed in Table I, y is usu- ally an order of magnitude less than the substrate con- centration, and thus the kinetics of substrate con- sumption approaches zero order. This will be true as long as the nutrient level in the cell growth region is above the cellular threshold value. Thus, the zero- order reaction with radial dispersion provides the clos- est approximation to the full numerical solution with Monod kinetics. With the flow rates used in cell cul- ture, convection and dispersion phenomena are of the same order of magnitude. Both sets of plots also in- dicate that the radial dispersion term cannot be omit- ted. The result agrees with the conventional chemical reaction engineering analysis that backmixing de- creases reactant conversion for positive order kinetics. In cell culture systems, the reactant is the chosen lim- iting substrate and dispersion effects now help to en- sure a more uniform nutrient profile in the bioreactor.
Comparison of Figures 4(a) and 4(b) shows that the behavior of the bioreactor with and without transmem- brane flux are similar. With hollow fibers [Fig. 4(b)],
the residual substrate concentration is always lower than that in the absence of transmembrane flux, even though the rates of substrate consumption are the same. This is an obvious consequence of the wall flux. While the fibers are designed to remove low molecular weight metabolic waste, the transmembrane flux will also carry along nutrient components which are usually of similar low molecular weight. To better utilize the nutrient, the cell culture should be operated as a continuous flow recycle reactor with a fairly high recycle ratio.2
Peclet Number, Pec
The Peclet number compares the relative signifi- cance of radial dispersion and bulk convection. With cell culture systems, the inlet superficial velocity is of the order c d s to minimize any possible shear damage to the mammalian cells. This velocity may be varied, depending on the choice of limiting substrate as well as the cell mass loading. The value for the dispersion coefficient estimated in Table I is calculated based on the effective diffusivity of the component in tissue adjusted using the correlations of Eidsath and co-workers.22 In this work, the effective dispersion coefficient is strongly dependent on the flow rate through the porous matrix and thus on the Peclet number. Since the flow rate through the bed is known, the Peclet number for each component may be calculated and the correlation of Eidsath and co-workers may be utilized to estimate the effective dispersion coefficient using the diffusion coefficient of the component through tis- sue.23
The magnitudes of convective and dispersive trans-
0 I I
Peclet Nuiiiber 10-1 10 O 10 ’ LO
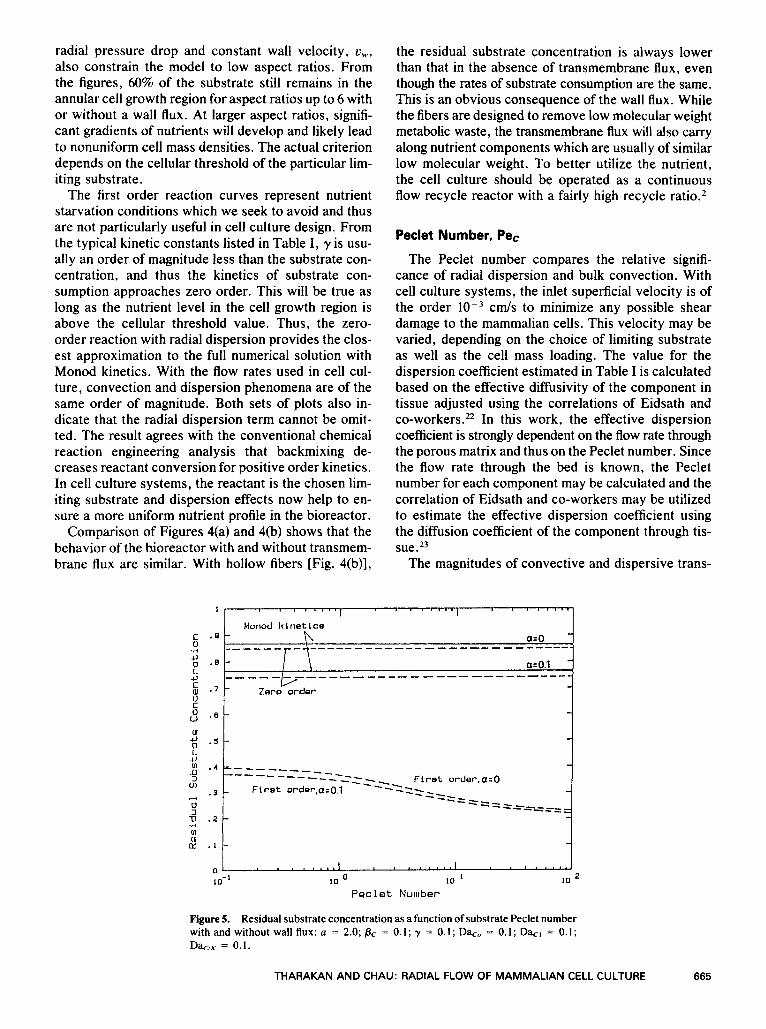
Figure 5. Residual substrate concentration as a function of substrate Peclet number with and without wall flux: a = 2.0; BC = 0.1; y = 0.1; Da,,, = 0.1; Da,=, = 0.1; D+,x = 0.1.
THARAKAN AND CHAU: RADIAL FLOW OF MAMMALIAN CELL CULTURE 665

port are similar and the Peclet number is relatively small. Typical values range from 1-10 for low-molec- ular-weight components to 10-100 for high-molecular- weight components. These values are in agreement with the work of Wei and R ~ s s ~ ~ who have shown that convection is the dominant transport mode for large molecules. Peclet numbers used in radial flow catalytic reactorsI2 or reverse osmosis unitsI9 are, on the other hand, of the order of 500 or greater.I6 The analysis provided by these workers would not be applicable for mammalian cell culture systems.
Figure 5 illustrates the residual substrate concentra- tion with increasing Pe,. The behavior of the system is very different for zero- and first-order kinetic ap- proximations. With zero-order kinetics, the Peclet number does not have any effect on the residual sub- strate concentration. This is in agreement with the no- tion that dispersion should not affect zero-order con- sumption of substrate. The solution for Monod kinetics indicates a finite albeit small decrease in the residual substrate concentration as the Peclet number in- creases. When a = 0, the residual substrate concen- tration is maintained at a higher level as a consequence of the absence of transmembrane flux. The approach to zero-order solution again is the result of a relatively high substrate concentration in the cell growth region.
membrane pressure drop. The membrane permeability depends on the molecular weight exclusion limit for the fiber,6 and decreases with increasing molecular weight cutoff. The operating transmembrane pressure drop increases with the bulk convective flow through the fiber bed. Thus, a can be varied by appropriate choices of hollow fibers and inlet velocity. The wall flux number takes on the limiting value of unity when all fluid exits via the fibers, and is zero when the hollow fiber bed is replaced by a nonpermeable porous matrix. The wall flux number can be viewed as both a design and operating parameter. The design consideration in- volves the choice of hollow fiber permeability, a de- cision that can be made on the basis of the product of interest and the molecular weight exclusion limit de- sired for the particular application. As an operating parameter, a suitable preliminary choice of inlet feed velocity can be made subject to the limitations of cell shear damage.
The residual substrate concentration in the cell growth region as a function of a is shown in Figure 6. Phys- ically, as a increases, a larger amount of substrate is transported into the fiber lumen together with other low-molecular-weight species, and the residual sub- strate in the outer annulus of the cell growth region decreases. The same result is true for any metabolic waste that is lower in molecular weight than the cutoff limit of the fibers. High-molecular-weight products, however, will be concentrated in the shell space as they are rejected by the fibers. Thus, while a high transmembrane flux helps to remove toxic metabolic waste, the large wall flux also leads to a more signifi- cant nutrient gradient in the cell growth region. This
Wall Flux Number, (Y
The wall flux number is the ratio of transmembrane flux across the fiber wall to the bulk convective flux through the annular fiber bed. The transmembrane flux depends on the membrane permeability and the trans-
c .Q c i -
-
-
-
- Zero order with dispersion
1 without dispersion t F 1
0 , 1 I I I I 1-
o . I . 2 .3 . 4 . 5 .B . 7 . a . a I
Wall F l u x Number
Figure 6. Residual substrate concentration as a function of the wall flux number, a: a = 2.0; Pec = 10.0; pc = 0.1; y = 0.1; Dac,, = 0.1; Dacl = 0.1; = 0.1.
666 BIOTECHNOLOGY AND BIOENGINEERING, VOL. 29, APRIL 1987
1
0
J (I)
U 3
v) (II tY . I - -
.3 - 4
: . 2 - -
0 I 1 I I I I I 1
I 1.1 1.2 1.3 1 . 4 1.5 1.8 1.7 1.8 1.e 2
Dii i rensionless Radi a 1 D i s t a n c e
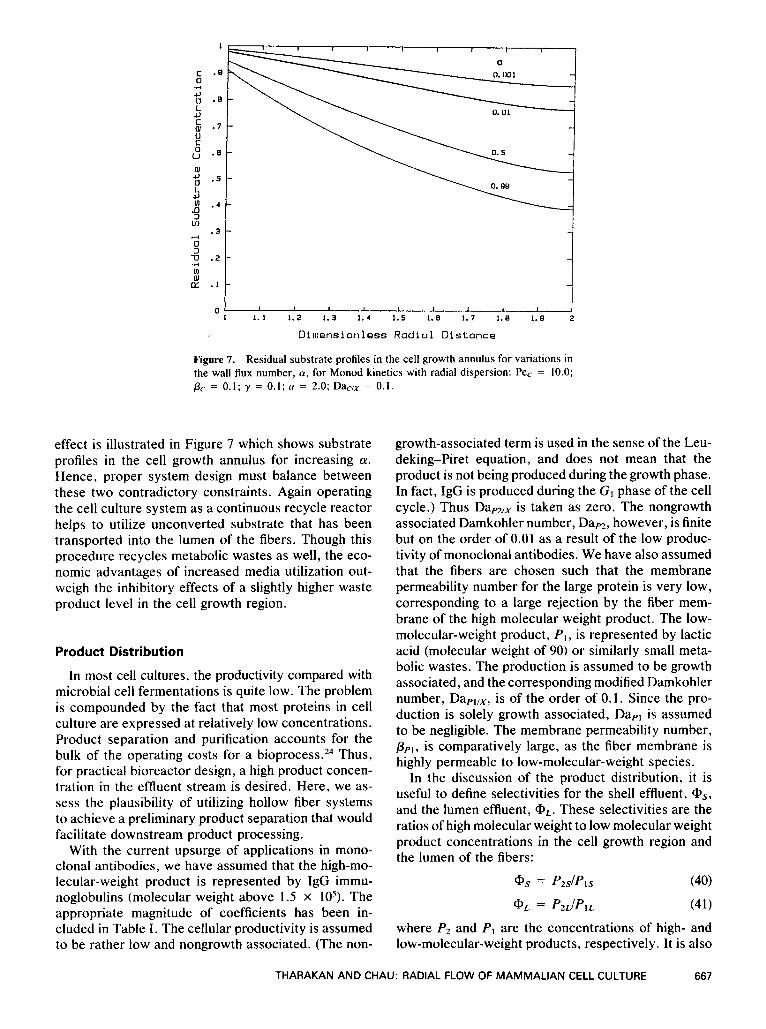
Figure 7. Residual substrate profiles in the cell growth annulus for variations in the wall flux number, 0, for Monod kinetics with radial dispersion: Pe, = 10.0; pc = 0.1; y = 0.1; u = 2.0; D+/x = 0.1.
0 ' I 1 I I I I 1.1 1.2 1.3 1 . 4 1.5 1.8 1.7 1.8 1.e 2
Dii i rensionless Radi a 1 D i s t a n c e
Figure 7. Residual substrate profiles in the cell growth annulus for variations in the wall flux number, 0, for Monod kinetics with radial dispersion: Pe, = 10.0; pc = 0.1; y = 0.1; u = 2.0; D+/x = 0.1.
effect is illustrated in Figure 7 which shows substrate profiles in the cell growth annulus for increasing a. Hence, proper system design must balance between these two contradictory constraints. Again operating the cell culture system as a continuous recycle reactor helps to utilize unconverted substrate that has been transported into the lumen of the fibers. Though this procedure recycles metabolic wastes as well, the eco- nomic advantages of increased media utilization out- weigh the inhibitory effects of a slightly higher waste product level in the cell growth region.
Product Distribution
In most cell cultures, the productivity compared with microbial cell fermentations is quite low. The problem is compounded by the fact that most proteins in cell culture are expressed at relatively low concentrations. Product separation and purification accounts for the bulk of the operating costs for a b i o p r o ~ e s s . ~ ~ Thus, for practical bioreactor design, a high product concen- tration in the effluent stream is desired. Here, we as- sess the plausibility of utilizing hollow fiber systems to achieve a preliminary product separation that would facilitate downstream product processing.
With the current upsurge of applications in mono- clonal antibodies, we have assumed that the high-mo- lecular-weight product is represented by IgG immu- noglobulins (molecular weight above 1.5 x lo5). The appropriate magnitude of coefficients has been in- cluded in Table I. The cellular productivity is assumed to be rather low and nongrowth associated. (The non-
growth-associated term is used in the sense of the Leu- deking-Piret equation, and does not mean that the product is not being produced during the growth phase. In fact, IgG is produced during the GI phase of the cell cycle.) Thus is taken as zero. The nongrowth associated Damkohler number, Darn, however, is finite but on the order of 0.01 as a result of the low produc- tivity of monoclonal antibodies. We have also assumed that the fibers are chosen such that the membrane permeability number for the large protein is very low, corresponding to a large rejection by the fiber mem- brane of the high molecular weight product. The low- molecular-weight product, P I , is represented by lactic acid (molecular weight of 90) or similarly small meta- bolic wastes. The production is assumed to be growth associated, and the corresponding modified Damkohler number, DaPIIX, is of the order of 0.1. Since the pro- duction is solely growth associated, Dapl is assumed to be negligible. The membrane permeability number, p p I , is comparatively large, as the fiber membrane is highly permeable to low-molecular-weight species.
In the discussion of the product distribution, it is useful to define selectivities for the shell effluent, QS,
and the lumen effluent, aL. These selectivities are the ratios of high molecular weight to low molecular weight product concentrations in the cell growth region and the lumen of the fibers:
@s = P2S/PIS (40)
@ L = P2LIPIL (41)
where P2 and P I are the concentrations of high- and low-molecular-weight products, respectively. It is also
THARAKAN AND CHAU: RADIAL FLOW OF MAMMALIAN CELL CULTURE 667
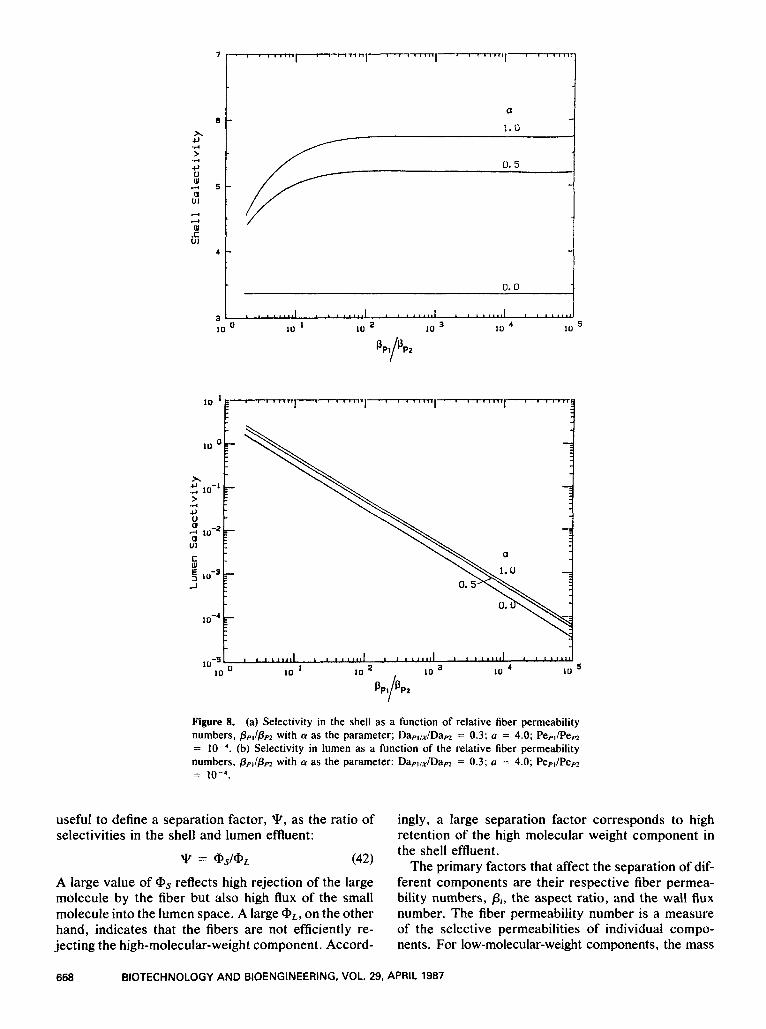
I ' ' "1--""""1'"""' ' "
L 4
Figure 8. (a) Selectivity in the shell as a function of relative fiber permeability numbers, pp,/ppz with a as the parameter; Dapl,JDapz = 0.3; a = 4.0; Pepl/Pep2 = (b) Selectivity in lumen as a function of the relative fiber permeability numbers, pp,/pp2 with a as the parameter: Dapllx/Dapz = 0.3; a = 4.0; Pep,/Pep2 = 1 0 - 4 .
useful to define a separation factor, q, as the ratio of selectivities in the shell and lumen effluent:
ingly, a large separation factor corresponds to high retention of the high molecular weight component in the shell effluent.
(42) The primary factors that affect the separation of dif- A large value of @s reflects high rejection of the large molecule by the fiber but also high flux of the small molecule into the lumen space. A large aL, on the other hand, indicates that the fibers are not efficiently re- jecting the high-molecular-weight component. Accord-
ferent components are their respective fiber permea- bility numbers, p i , the aspect ratio, and the wall flux number. The fiber permeability number is a measure of the selective permeabilities of individual compo- nents. For low-molecular-weight components, the mass
668 BIOTECHNOLOGY AND BIOENGINEERING, VOL. 29, APRIL 1987

transfer coefficient is relatively high and p p I will be much larger than pm. Numerically, p p I is close to unity and ppz is around 0.001.
With larger aspect ratios, the depth of the bed in- creases and there is a corresponding increase in the fiber surface area available for product separation. A larger aspect ratio would favor more efficient fraction- ation of the different molecular weight products. How- ever, as was discussed earlier, large aspect ratios lead to significant nutrient gradients that will be detrimental to uniform cell growth and viability. Thus, it is nec- essary to compromise between these two opposing constraints.
An increasing wall flux number corresponds to a larger transmembrane flux into the lumen. This is de- sirable for product separation, as the higher membrane transport rate will carry more low-molecular-weight components into the lumen, thus increasing the selec- tivity of the high-molecular-weight product in the shell space.
Figure 8(a) shows the shell effluent selectivity as a function of the ratio of fiber permeability numbers, ppI/ppz, with the wall flux number as a parameter. An increasing ratio corresponds to more efficient rejection of the high molecular weight component by the fibers. Thus the shell side selectivity increases. The selectiv- ity increases by twofold as the situation changes from no wall flux to that of a high wall flux number. The selectivity reaches an asymptote as the separation is limited by the total amount of fiber surface area avail- able. The corresponding lumen effluent selectivity is shown in Figure 8(b). As expected, the lumen selec- tivity decreases readily with the ratio p p I / p m as more
low-molecular-weight product is transported across the fiber membrane.
Figure 9 shows the change of shell effluent selectivity as a function of wall flux number. The parameter used is the aspect ratio. The result supports Figure 10 that a larger aspect ratio can improve the selectivity twofold in the shell effluent. With zero wall flux, the shell ef- fluent selectivity is the same regardless of the aspect ratio. This is an expected result since with no lumen effluent and the same production rates, the ratio of high- and low-molecular-weight products must be the same.
Figure 10 plots the separation factor as a function of the ratio of fiber membrane mass transfer coeffi- cients for the two molecules. When the ratio of the two mass transfer coefficients become unity, the sep- aration factor also becomes one and the distribution of the two components in the shell and lumen are iden- tical. The separation factor increases as the mass trans- fer coefficient of the low-molecular-weight product be- comes larger comparatively. As the mass transfer coefficients begin to differ by a factor of a 1000, ap- proaching complete rejection of the high-molecular- weight species and complete passage of the low-mo- lecular-weight species, the separation factor increases more significantly.
It should be mentioned that the efficiency of the separation depends strongly on the ability of the fibers to maintain their selective permeability as culture age progresses. With time, a gel layer of high-molecular- weight components will likely build up on the fiber surface and reduce the wall permeability for the low- molecular-weight components. In accordance with
1 I 1 I.-
o . I . 2 .3 . 4 .s . e .7 . a . a I
Wall F l u x Number
3
Figure 9. Shell selectivity as a function of wall flux number, a. with the aspect ratio as the parameter: Pep,/Pen = ppI/pn = 104; Dapl,x/Dan = 0.3.
THARAKAN AND CHAU: RADIAL FLOW OF MAMMALIAN CELL CULTURE 669
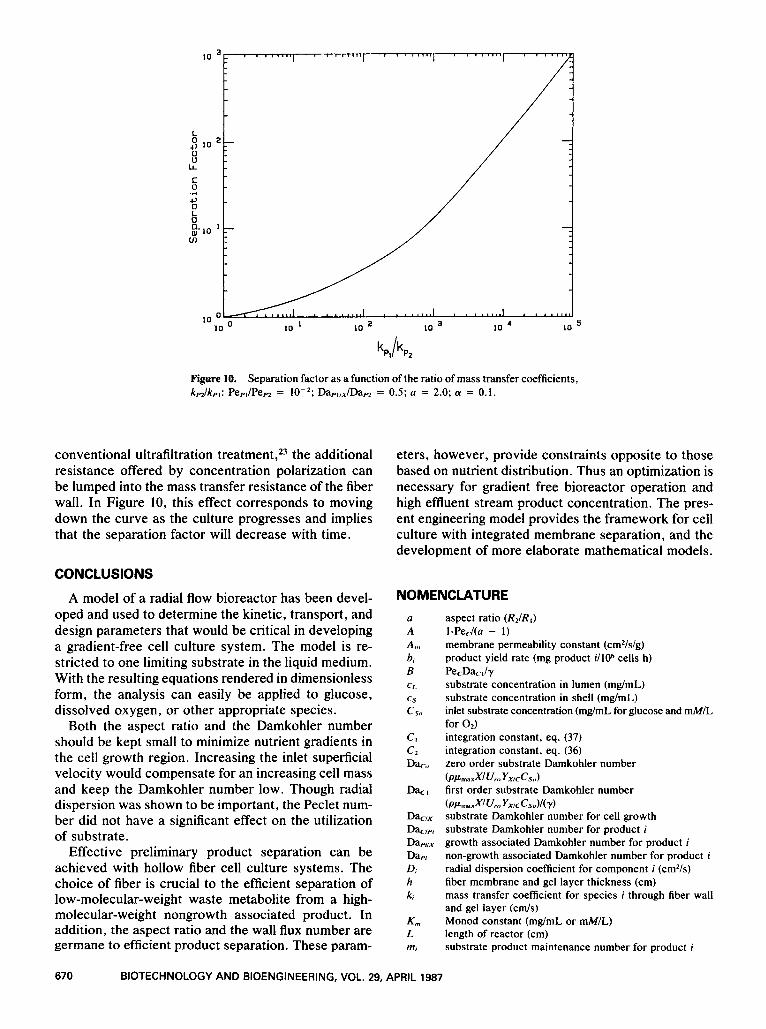
Figure 10. Separation factor as a function of the ratio of mass transfer coefficients, kp2/kpI: Pepl/Pe, = W2; Dapl/x/Dap2 = 0.5; a = 2.0; a = 0.1.
conventional ultrafiltration treatment,23 the additional resistance offered by concentration polarization can be lumped into the mass transfer resistance of the fiber wall. In Figure 10, this effect corresponds to moving down the curve as the culture progresses and implies that the separation factor will decrease with time.
CONCLUSIONS
A model of a radial flow bioreactor has been devel- oped and used to determine the kinetic, transport, and design parameters that would be critical in developing a gradient-free cell culture system. The model is re- stricted to one limiting substrate in the liquid medium. With the resulting equations rendered in dimensionless form, the analysis can easily be applied to glucose, dissolved oxygen, or other appropriate species.
Both the aspect ratio and the Damkohler number should be kept small to minimize nutrient gradients in the cell growth region. Increasing the inlet superficial velocity would compensate for an increasing cell mass and keep the Damkohler number low. Though radial dispersion was shown to be important, the Peclet num- ber did not have a significant effect on the utilization of substrate.
Effective preliminary product separation can be achieved with hollow fiber cell culture systems. The choice of fiber is crucial to the efficient separation of low-molecular-weight waste metabolite from a high- molecular-weight nongrowth associated product. In addition, the aspect ratio and the wall flux number are germane to efficient product separation. These param-
eters, however, provide constraints opposite to those based on nutrient distribution. Thus an optimization is necessary for gradient free bioreactor operation and high effluent stream product concentration. The pres- ent engineering model provides the framework for cell culture with integrated membrane separation, and the development of more elaborate mathematical models.
NOMENCLATURE aspect ratio (R2/RI) l-Pec/(u - 1) membrane permeability constant (cm2/s/g) product yield rate (mg product i / lW cells h) PecDacl/y substrate concentration in lumen (mg/mL) substrate concentration in shell (mg/mL) inlet substrate concentration (mg/mL for glucose and mM/L for 02) integration constant, eq. (37) integration constant, eq. (36) zero order substrate Damkohler number @PmaxXIUr0 YX/CCS,J first order substrate Damkohler number @Prn.JI u.0 YX/CCS<>)4 Y) substrate Damkohler number for cell growth substrate Damkohler number for product i growth associated Damkohler number for product i non-growth associated Damkohler number for product i radial dispersion coefficient for component i (cm2/s) fiber membrane and gel layer thickness (cm) mass transfer coefficient for species i through fiber wall and gel layer (cm/s) Monod constant (mg/mL or mM/L) length of reactor (cm) substrate product maintenance number for product i
670 BIOTECHNOLOGY AND BIOENGINEERING, VOL. 29, APRIL 1987

number of fibers lumen side pressure (dyn/cm2) shell side pressure (dyn/cm*) shell side product concentration for i (mg/mL) lumen side product concentration for i (mg/mL) Peclet number for species i reference product concentration (Cs,,YPl,,) (mg/mL) volumetric flow rate into fiber lumen per unit volume of shell (s- ’) radial coordinate (cm) outside fiber radius (cm) inner radius of annulus (cm) outer radius of annulus (cm) substrate consumption rate (mg/mL h) product information rate (mg/mL h) time (s or h) radial superficial velocity ( c d s ) axial velocity (cds) inlet radial velocity (cmls) dimensionless radial velocity, u,/U,” velocity through fiber wall (cds) shell volume (cm’) dimensionless radial coordinate, rlp cell number density (cell/mL) reference yield coefficient (mg product i/mg substrate) relative product yield coefficient (mg product ilmg prod- uct I ) substrate yield coefficient (celllmg glucose or celllmM 0 2 )
product i yield coefficient (celumg product i) axial coordinate (cm)
Greek letters a wall flux number p, fiber permeability number for species i y dimensionless Monod constant, K,,JCsn E fiber bed void fraction Qs shell side product selectivity QL lumen side product selectivity Q, u 2Pe& - 1) p 9 separation factor 9 dimensionless substrate concentration, cs/Cso pmax maximum specific growth rate (h-I)
dimensionless product i concentration, P,slP,fl Y,/ l
fiber bed depth, Rz - R , (cm)
Subscripts C substrate 1 component i L fiber lumen space P I product 1 Pz product 2
r radial component S Z axial component
cell growth region, identical to shell space
References
1. M. W. Glacken, R. J . Fleischaker, and A. J. Sinskey, Ann. N Y
2. J. Tharakan and P. C. Chau, Biotechnol. Bioeng., 28,435 (1986). 3. R. A. Knazek, P. M. Gullino, P. 0. Kohler, and R. L. Dedrick,
4. P. M. Gullino and R. A. Knazek, Methods Enzymol., 58, 178
5 . K. Ku, M. J. Kuo, J. Delente, B. S. Wildi, and J. Feder, Bio-
6. J. Tharakan and P. C. Chau, Biotechnol. Bioeng., to appear. 7. K. Johansen, Chem. Eng. World, 5 , 8 (1970). 8. E. J . Newson, 2nd Intern. Symp. on Chem. Recat. Eng., Am-
9. V . S. Genkin, V. V. Dil’man, and S. P. Sergeev, Int . Chem.
10. L. A. Kaye, AIChE Annual Meeting, paper No. 12E, Miami
1 1 . V. Hlavacek and M. Kublicek, Chem. Eng. Sci., 27, 177 (1972). 12. M. P. Dudukovic and H. S. Lamba, AIChE Annual Meeting,
13. P. R. Ponzi and L. A. Kaye, AIChE J., 25, 100 ( 1979). 14. H. C. Chang and J. M. Calo, ACS Symp. Ser., 168, 305 (1981). 15. C. S. Yo0 and A. G. Dixon, “Modelling and Simulation of a
Radial Flow Fixed-Bed Reactor,” AIChE Annual Meeting, Chi- cago, IL, 1985.
Acad. Sci., 413, 355 (1983).
Science, 178, 65 (1972).
( 1979).
technol. Bioeng., 23, 79 (1981).
sterdam, (May 1972).
Eng., 13, 24 (1973).
Beach, FL, 1978.
paper No. 576, Boston, MA, 1975.
16. W. N. Gill and B. N. Bansal, AlChE J., 19, 823 (1973). 17. M. S. Dandavati, M. R. Doshi, and W. N. Gill. Chem. Eng.
18. V . N. Kabadi, M. R. Doshi, and W. N. Gill, Chem. Eng. Com-
19. M. Soltanieh and W. N. Gill, Chem. Eng. Commun., 18, 311
20. M. Soltanieh and W. N. Gill, Desalination, 49, 57 (1984). 21. R. W. Tolbert and J. Feder, Ann. Rep. Ferm. Proc.. 6,49 (1983). 22. A. Eidsath, R. G. Carbonell, S. Whitaker, and L. R. Herrmann,
23. J. Wei and M. Russ, J. Theor. Biol., 66, 775 (1977). 24. E. Flaschel, C. Wandrey, and M. R. Kula, Adv. Biochem. Eng.
Biotechnol., 26, 73 (1983). 25. S. B. Karkare, P. G. Phillips, D. H. Burke, and R. C. Dean,
“Continuous Production of Monoclonal Antibodies by Che- mostatic and Immobilized Hybridoma Culture,” ACS Annual Meeting, Philadelphia, PA, 1984.
26. Amicon Corporation, Technical Publication No. 494, Danvers, MA, 1984.
Sci., 30, 877 (1975).
mun., 3, 339 (1979).
(1982).
Chem. Eng. Sci., 38, 1803 (1983).
THARAKAN AND CHAU: RADIAL FLOW OF MAMMALIAN CELL CULTURE 67 1
Related Documents











