Mobile hard substrata e An additional biodiversity source in a high latitude shallow subtidal system Piotr Balazy a, * , Piotr Kuklinski a, b a Marine Ecology Department, Institute of Oceanology, Polish Academy of Sciences, Powstancow Warszawy 55, Sopot 81-712, Poland b Department of Life Sciences, Natural History Museum, Cromwell Road, London SW7 5BD, United Kingdom article info Article history: Received 13 May 2012 Accepted 3 January 2013 Available online 17 January 2013 Keywords: mobile hard substrata epifaunal assemblages hermit crabs biodiversity polar waters Arctic abstract This study demonstrates the importance of a hard mobile substratum (hermit crab shells) for Arctic biodiversity. Based on previous observations from other geographic regions we hypothesized that this niche at high latitudes would support a higher biodiversity of epifauna than might be predicted from similar substrata. We test whether the hermit crab epifauna is specific to that substratum providing unique biodiversity components to the local community. From four study sites in Isfjorden (78 N), West Spitsbergen and two study sites in Northern Norway (69 N) we collected approximately 50 each of hermit crabs, gastropods and pebbles, of visually similar surface area using SCUBA diving. Hermit crab shells were colonized by a larger number of epifaunal species than either gastropods or pebbles, even when they were of a larger size. Among 87 taxa found on all the three substrata, 22 occurred only on hermit crab shells. Except for two study sites hermit crab shells also supported more individuals. This study shows that the contribution of shells carried by hermit crabs to high-latitude, shallow-subtidal diversity is higher than might be predicted by their surface area alone and that hermit crabs modify, maintain and create a unique habitat. This is the result of a number of factors interacting positively on the presence of epifauna including shell surface heterogeneity and the complex influence of the crab host. Ó 2013 Elsevier Ltd. All rights reserved. 1. Introduction Factors affecting biodiversity is one of the most frequent topics addressed in ecological studies. In recent years biodiversity related issues have received even more attention due to growing interest in global and regional environmental changes (Chapin et al., 2000; IPCC, 2001; Pereira et al., 2010; Dawson et al., 2011). Yet in com- parison to terrestrial ecosystems, we still know very little about the processes that control marine biodiversity patterns at different spatial and temporal scales (Heip, 2003; Worm et al., 2006; Tittensor et al., 2010). Although there has been much progress in this topic in the last decade, there are still many gaps in our knowledge especially in polar regions (Piepenburg, 2005; Costello et al., 2010; Bluhm et al., 2011; Appeltans et al., 2012). Hermit crabs and the communities of species they support are regarded as an important model system to study biodiversity pat- terns over a variety of scales (Williams and McDermot, 2004). This extensively studied group is abundant worldwide in shallow water environments (Hazlett, 1981; Lancaster, 1988, 1990). Epifaunal assemblages occurring on empty gastropod shells inhabited by hermit crabs can be very diverse (Jensen and Bender, 1973; Reiss et al., 2003, for a complete review see Williams and McDermot, 2004). The large numbers of species associated with hermit crab shells results from the fact that the mobile hard substratum ben- efits from a number of advantages for epifaunal taxa. These include mobility and stability of substratum preventing epifauna from be- ing buried in sediment and increased food supply to organisms colonizing shells by constant movement of the antennae and mouth parts of the hermit crab (Conover, 1975; Stachowitsch, 1980). There is much evidence that hermit crabs are ecosystem engineers (Stachowitsch, 1977; McLean, 1983; Jones et al., 1994, 1997; Gutierrez et al., 2003; Reiss et al., 2003; Bell, 2005) that efficiently modify, maintain and create habitat for other biota (Williams and McDermot, 2004). Several studies, from both tropical and temperate regions, show that hermit crabs support a higher biodiversity than other firm substrata of similar surface area (Conover, 1979; Karlson and Cariolou, 1982; Karlson and Shenk, 1983; Van Winkle et al., 2000; Bell, 2005; Bick, 2006). Whether this is a global pattern or just a local phenomenon is not known. This study investigates the possible influence of hard mobile substrata on local epifaunal diversity in a high latitudinal coastal system. Shallow Arctic seas are dominated by muddy bottoms and * Corresponding author. E-mail addresses: [email protected] (P. Balazy), [email protected] (P. Kuklinski). Contents lists available at SciVerse ScienceDirect Estuarine, Coastal and Shelf Science journal homepage: www.elsevier.com/locate/ecss 0272-7714/$ e see front matter Ó 2013 Elsevier Ltd. All rights reserved. http://dx.doi.org/10.1016/j.ecss.2013.01.004 Estuarine, Coastal and Shelf Science 119 (2013) 153e161

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

at SciVerse ScienceDirect
Estuarine, Coastal and Shelf Science 119 (2013) 153e161
Contents lists available
Estuarine, Coastal and Shelf Science
journal homepage: www.elsevier .com/locate/ecss
Mobile hard substrata e An additional biodiversity source in a highlatitude shallow subtidal system
Piotr Balazy a,*, Piotr Kuklinski a,b
aMarine Ecology Department, Institute of Oceanology, Polish Academy of Sciences, Powstancow Warszawy 55, Sopot 81-712, PolandbDepartment of Life Sciences, Natural History Museum, Cromwell Road, London SW7 5BD, United Kingdom
a r t i c l e i n f o
Article history:Received 13 May 2012Accepted 3 January 2013Available online 17 January 2013
Keywords:mobile hard substrataepifaunal assemblageshermit crabsbiodiversitypolar watersArctic
* Corresponding author.E-mail addresses: [email protected] (P.
(P. Kuklinski).
0272-7714/$ e see front matter � 2013 Elsevier Ltd.http://dx.doi.org/10.1016/j.ecss.2013.01.004
a b s t r a c t
This study demonstrates the importance of a hard mobile substratum (hermit crab shells) for Arcticbiodiversity. Based on previous observations from other geographic regions we hypothesized that thisniche at high latitudes would support a higher biodiversity of epifauna than might be predicted fromsimilar substrata. We test whether the hermit crab epifauna is specific to that substratum providingunique biodiversity components to the local community. From four study sites in Isfjorden (78�N), WestSpitsbergen and two study sites in Northern Norway (69�N) we collected approximately 50 each ofhermit crabs, gastropods and pebbles, of visually similar surface area using SCUBA diving. Hermit crabshells were colonized by a larger number of epifaunal species than either gastropods or pebbles, evenwhen they were of a larger size. Among 87 taxa found on all the three substrata, 22 occurred only onhermit crab shells. Except for two study sites hermit crab shells also supported more individuals. Thisstudy shows that the contribution of shells carried by hermit crabs to high-latitude, shallow-subtidaldiversity is higher than might be predicted by their surface area alone and that hermit crabs modify,maintain and create a unique habitat. This is the result of a number of factors interacting positively on thepresence of epifauna including shell surface heterogeneity and the complex influence of the crab host.
� 2013 Elsevier Ltd. All rights reserved.
1. Introduction
Factors affecting biodiversity is one of the most frequent topicsaddressed in ecological studies. In recent years biodiversity relatedissues have received evenmore attention due to growing interest inglobal and regional environmental changes (Chapin et al., 2000;IPCC, 2001; Pereira et al., 2010; Dawson et al., 2011). Yet in com-parison to terrestrial ecosystems, we still know very little about theprocesses that control marine biodiversity patterns at differentspatial and temporal scales (Heip, 2003; Worm et al., 2006;Tittensor et al., 2010). Although there has been much progress inthis topic in the last decade, there are still many gaps in ourknowledge especially in polar regions (Piepenburg, 2005; Costelloet al., 2010; Bluhm et al., 2011; Appeltans et al., 2012).
Hermit crabs and the communities of species they support areregarded as an important model system to study biodiversity pat-terns over a variety of scales (Williams and McDermot, 2004). Thisextensively studied group is abundant worldwide in shallow waterenvironments (Hazlett, 1981; Lancaster, 1988, 1990). Epifaunal
Balazy), [email protected]
All rights reserved.
assemblages occurring on empty gastropod shells inhabited byhermit crabs can be very diverse (Jensen and Bender, 1973; Reisset al., 2003, for a complete review see Williams and McDermot,2004). The large numbers of species associated with hermit crabshells results from the fact that the mobile hard substratum ben-efits from a number of advantages for epifaunal taxa. These includemobility and stability of substratum preventing epifauna from be-ing buried in sediment and increased food supply to organismscolonizing shells by constant movement of the antennae andmouth parts of the hermit crab (Conover, 1975; Stachowitsch,1980). There is much evidence that hermit crabs are ecosystemengineers (Stachowitsch, 1977; McLean, 1983; Jones et al., 1994,1997; Gutierrez et al., 2003; Reiss et al., 2003; Bell, 2005) thatefficiently modify, maintain and create habitat for other biota(Williams andMcDermot, 2004). Several studies, from both tropicaland temperate regions, show that hermit crabs support a higherbiodiversity than other firm substrata of similar surface area(Conover, 1979; Karlson and Cariolou, 1982; Karlson and Shenk,1983; Van Winkle et al., 2000; Bell, 2005; Bick, 2006). Whetherthis is a global pattern or just a local phenomenon is not known.
This study investigates the possible influence of hard mobilesubstrata on local epifaunal diversity in a high latitudinal coastalsystem. Shallow Arctic seas are dominated by muddy bottoms and

P. Balazy, P. Kuklinski / Estuarine, Coastal and Shelf Science 119 (2013) 153e161154
hard surfaces are locally rare, which often results in species-poorecosystems. The presence of even a small patch of hard surfacecan significantly increase local biodiversity (McLean, 1983; Daleet al., 1989; Kuklinski et al., 2006). Therefore, gastropod shellsused by hermit crabs are considered an important source of hardmobile surfaces in those regions (Kuklinski et al., 2008). Despitethis there are few studies describing shallow water Arctic marinecommunities even though this zone seems to be the most sus-ceptible to on-going global climate change (IPCC, 2007;Schiermeier, 2007; Beuchel and Gulliksen, 2008). The ice cap of theArctic Ocean is predicted to be seasonally free of ice in less than 30years (Wang and Overland, 2009). Increasing water temperaturealready has very large impact on this marine ecosystem. Newlyinvading species are appearing (Berge et al., 2005; Renaud et al.,2008) while truly Arctic ones are predicted to move polewards orvanish (Weslawski et al., 2010, 2011). Therefore, reliable up-to-dateinformation about Arctic diversity and ecology is needed.
At high latitudes we expected to observe differences in speciesnumber, species composition and their relative abundance be-tween hermit crab shells and other hard substrata of the samesurface area. In this study we test (1) whether hermit crab shellsprovide a unique substratum for benthic biodiversity as expressedby an increased number of species and relative abundance incomparison to similar substratum in terms of complexity and sur-face area. We compare epifaunal assemblage parameters of shellscarried by hermit crabs to similar shells of live gastropods from thesame locality. In addition to these two biotic groups of substratum
75°N75°N
70°N70°N
80°N80°N
S
T
Fig. 1. Study area with marked sampling sites (S e Isfjorde
we also analyze epifaunal assemblages of pebbles with similarsurface area to the shells. By comparing epifaunal assemblagesfrom these three substrata we test (2) to what degree they differ inspecies composition. We analyze (3) whether the hermit crabepifauna is specific to that substratum providing unique compo-nents to the local community. The study encompasses distantgeographic locations enabling us to understand (4) whether theseprocesses are local or relevant over broader scale. Lastly we hy-pothesize (5) that the hermit crab epifaunal assemblage will sup-port a higher proportion of species than either gastropod shells orpebbles at the soft bottom sites, where it is often the one of fewproviders of firm surface, in comparison to rocky bottoms wherethe species pool of typical hard bottom organisms is much higher.
2. Material and methods
2.1. Study area
The study was carried out in Northern Norway (69�N) and inWest Spitsbergen (Svalbard, 78�N) (Fig. 1). In Northern Norway twostudy sites (T1, T3) were selected north of Tromsø (Fig. 1). The fjordhere cuts deeply into the mainland and is sheltered from the opensea, however, water circulation and exchange is high due to strongtidal currents (Holte and Oug, 1996). The sea temperature usuallyvaries from 1 to 9 �C and salinities range from 31 to 34 (Oug, 1998).Water masses in the area are the transitional zone between borealand Arctic areas (Holte and Oug, 1996). The water rarely freezes,
KongsfjordenKongsfjorden
Isfjorden
Bellsund
10°E10°E
70°N70°N
S1S2
S3S4
T1T3
n, West Spitsbergen; T e Tromsø, Northern Norway).

P. Balazy, P. Kuklinski / Estuarine, Coastal and Shelf Science 119 (2013) 153e161 155
and then only in the shallow, innermost parts of the fjord. There isno terrestrial influence from glaciers. The bottom in this locationconsists of sand and gravel with an admixture of cobbles to boul-ders. Thick crusts of coralline red algae cover the substratum increvices and between boulders (Oug, 2001) while serpulid poly-chaete reefs dominate the macrobenthos. Three pagurids werefound in northern Norway: Pagurus pubescens Krøyer, 1838, Pagurusbernhardus (Linnaeus, 1758) and Anapagurus chiroacanthus (Lillje-borg, 1856) (Barnes et al., 2007).
Four study sites (S1eS4) were selected in Isfjorden, which is thelargest fjord on the west coast of the largest island of the SvalbardArchipelago e West Spitsbergen (Fig. 1). Isfjorden has an open,broad mouth without any shallows. Hydrological conditions insidethe fjord are the result of an interplay between warmer highly sa-line Atlantic and colder Arctic waters. There is some freshwaterdischarge and sedimentation mainly due to calving of surroundingglaciers but also to river runoff (Zajaczkowski and Wlodarska-Kowalczuk, 2007; Nilsen et al., 2008). The salinity in the subtidalvaries from 25 to 32, while the sea temperature range is fromminus1.8e4 �C. The sea surface is usually at least partially covered by iceduringwintermonths, but the study sites closer to themouth of thefjord have been ice-free for the last few years. Sites S1 and S2 arecharacterized by hard bedrock with a steep scree of pebbles, cob-bles and boulders. At these locations a rich epibenthic communityconsisting mostly of algae (dense concentrations of Laminaria spp.down to depth of w10 m), ascidians, barnacles, bryozoans, poly-chaetes and sponges has been reported (Barnes and Kuklinski,2005). On the other hand, sites S3 and S4 were characterized bymuddy sediments. Such locations in Isfjorden are dominated bya characteristic soft bottom fauna includingmostly polychaetes andbivalves (Wlodarska-Kowalczuk et al., 2007). Hermit crabs are fairlycommon at all localities reaching abundances up to fiveindividuals m�2 (Barnes et al., 2007). At Svalbard the hermit crabPagurus pubescens is at the absolute northernmost limit (Birula,1907; Heegaard, 1941). In the shallow waters of the archipelagoSandberg and McLaughlin (1988), Gulliksen and Svendsen (2004)reported another crab in the same family e Pagurus bernhardus.
2.2. Protocol
At each site, in 2009 and 2010, 50 hermit crabs (Pagurus spp.)(except S4 where only 14 hermit crabs were found), 50 gastropodsand 50 pebbles of visually similar size were collected by SCUBAdivers from 5 m2 at approximately 15 m depth. Hermit crabs andgastropod shells were identified to species in order to compareshells originating from the same gastropod species. All organismslarger than 1 mm directly attached to the shell and pebble wereidentified to the lowest possible taxonomic level. Determination ofSpirorbidaewasmade according to Jirkov (2001). Due to the state ofthe samples (retrieved after preservation) not all morphologicalcharacters of the body could be recognized. Identifications werebased on the structure of the tube attached to the shell or pebble,therefore, some spirorbids were classified into morpho-groups:Spirorbis sp./Bushiella sp./Pilleolaria sp.; Spirorbidae juvenile; Spi-rorbidae undetermined. Number of individuals were counted andeach colonial organism was considered as a single individual.External shell surface and pebble area (SA) were estimated usingthe technique described by Bergey and Getty (2006). Shells andpebbles were carefully wrapped in a thin layer of stock aluminumfoil paying special attention to avoid overlapping any regions. Allexcess foil was trimmed off. The foil was then weighed (B) and theresults were plotted against aluminum foil pieces of known size(SA ¼ 0.0495 þ 413.59*B, R2 ¼ 0.948).
To investigate and compare richness values between three dif-ferent substrata (hermit crab shell, gastropod shell, pebble) species
accumulation curves (increase in number of epifaunal species withnumber of individuals/colonies counted) were plotted. Hermitcrabs might use a different number of shell species than the gas-tropods therefore additionally the same analysis was limited onlyto Buccinum sp. shells. This ensured full comparability of these twosubstrata and excluded the possible impact of different shell spe-cies composition. Because the sizes of hermit crab and gastropodshells, and pebbles were estimated underwater only visually ananalysis of variance (ANOVA) was used to test the statistical dif-ference in the surface areas of the three substrata. Post-hoc pair-wise comparisons were performed using a Tukey’s honestlysignificant difference (HSD) test for equal number of samples(Tukey, 1953).
The comparison of epifaunal species composition between dif-ferent substrata was calculated using zero-adjusted BrayeCurtissimilarity index (to improve interpretability of displays and sig-nificance of tests when working on denuded or entirely blanksamples) and PRIMER software package (Bray and Curtis, 1957;Clarke and Gorley, 2001; Clarke et al., 2006). In order to amelioratethe effects of differing sample areas, data were first standardized.Non-metric multidimensional scaling (nMDS) of square roottransformed data as well as presence/absence data was carried out(Clarke and Gorley, 2001). This provided a balanced view of theassemblage structure (square root transformation) and a compari-son of the species list (presence/absence data) occurring in sam-ples. One-way analyses of similarities (ANOSIM; Clarke and Green,1988) were used to test differences between the three studiedsubstrata. ANOSIM uses the statistic R, which is calculated usingaverage rank similarities among pairs of replicates (in this caseepifaunal species) within each of the groups (different years) minusthe average rank similarity of replicates between groups and itranges between �1 and 1. R ¼ 1 when all similarities within groupsare less than any similarity between groups, R> 0.75 when there islarge difference with the groups either well separated, R > 0.5when overlapping but clearly different, R < 0.25 when groups arebarely separable and R ¼ 0 when samples within and betweengroups are equally similar. R ¼ �1 means greater dissimilarityamong replicate units within samples than between samples, forexample when the community is species poor and individuals havea heavily clustered distribution, so that variability in a group isextreme. Alternatively, it may indicate an incorrect labeling ofsamples (Chapman and Underwood, 1999; Clarke and Gorley,2001).
3. Results
At all study sites there were differences in surface area betweenthe substrata despite visual similarities (ANOVA p < 0.001, Table 1).However, at no site did hermit crab shells have significantly largermean shell area than gastropod shells or pebbles. At stations S1, S2and T1 hermit crab and gastropod shells had similar surface area (inall cases Tukey HSD test p > 0.59). Pebbles were the largest sub-stratum at stations S1, S2, S3 and T1 while at stations S4 gastropodshells had the largest surface area (in all cases Tukey HSD testp < 0.025). At station T3 gastropod shells and pebbles had similarsurface on average (Tukey HSD test p ¼ 0.99).
The 264 specimens of hermit crabs collected at all sitesbelonged to two species: Pagurus pubescens Krøyer, 1838 andPagurus bernhardus (Linnaeus, 1758). At West Spitsbergen only P.pubescens was recorded. Hermit crabs used 24 species of gastro-pod shells (mostly Buccinum glaciale Linnaeus, 1758 and Buccinumundatum Linnaeus, 1758) with a total surface area of 5302 cm2. Intotal 14,404 individuals representing 78 epifaunal taxa were foundon the investigated hermit crab shells (11,571 individuals of 69taxa at West Spitsbergen, 2833 individuals of 45 taxa in Northern
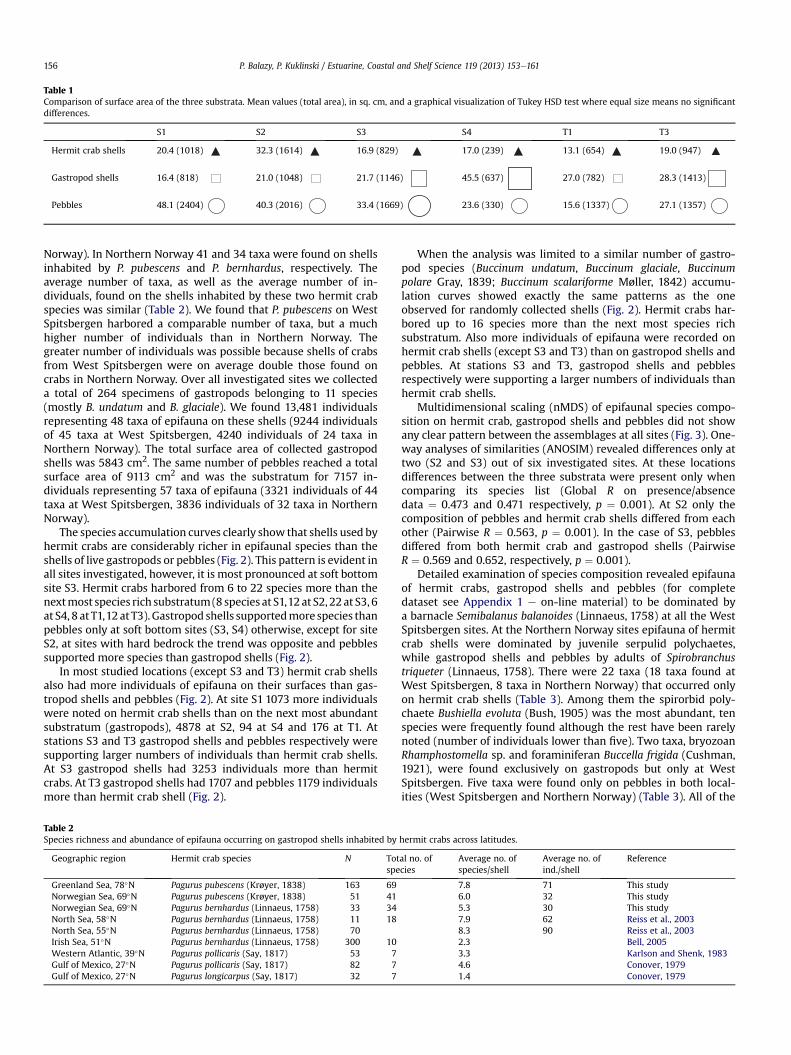
Table 1Comparison of surface area of the three substrata. Mean values (total area), in sq. cm, and a graphical visualization of Tukey HSD test where equal size means no significantdifferences.
S1 S2 S3 S4 T1 T3
Hermit crab shells 20.4 (1018) 32.3 (1614) 16.9 (829) 17.0 (239) 13.1 (654) 19.0 (947)
Gastropod shells 16.4 (818) 21.0 (1048) 21.7 (1146) 45.5 (637) 27.0 (782) 28.3 (1413)
Pebbles 48.1 (2404) 40.3 (2016) 33.4 (1669) 23.6 (330) 15.6 (1337) 27.1 (1357)
P. Balazy, P. Kuklinski / Estuarine, Coastal and Shelf Science 119 (2013) 153e161156
Norway). In Northern Norway 41 and 34 taxa were found on shellsinhabited by P. pubescens and P. bernhardus, respectively. Theaverage number of taxa, as well as the average number of in-dividuals, found on the shells inhabited by these two hermit crabspecies was similar (Table 2). We found that P. pubescens on WestSpitsbergen harbored a comparable number of taxa, but a muchhigher number of individuals than in Northern Norway. Thegreater number of individuals was possible because shells of crabsfrom West Spitsbergen were on average double those found oncrabs in Northern Norway. Over all investigated sites we collecteda total of 264 specimens of gastropods belonging to 11 species(mostly B. undatum and B. glaciale). We found 13,481 individualsrepresenting 48 taxa of epifauna on these shells (9244 individualsof 45 taxa at West Spitsbergen, 4240 individuals of 24 taxa inNorthern Norway). The total surface area of collected gastropodshells was 5843 cm2. The same number of pebbles reached a totalsurface area of 9113 cm2 and was the substratum for 7157 in-dividuals representing 57 taxa of epifauna (3321 individuals of 44taxa at West Spitsbergen, 3836 individuals of 32 taxa in NorthernNorway).
The species accumulation curves clearly show that shells used byhermit crabs are considerably richer in epifaunal species than theshells of live gastropods or pebbles (Fig. 2). This pattern is evident inall sites investigated, however, it is most pronounced at soft bottomsite S3. Hermit crabs harbored from 6 to 22 species more than thenextmost species rich substratum(8 species at S1,12at S2, 22at S3, 6at S4, 8 at T1,12 at T3). Gastropod shells supportedmore species thanpebbles only at soft bottom sites (S3, S4) otherwise, except for siteS2, at sites with hard bedrock the trend was opposite and pebblessupported more species than gastropod shells (Fig. 2).
In most studied locations (except S3 and T3) hermit crab shellsalso had more individuals of epifauna on their surfaces than gas-tropod shells and pebbles (Fig. 2). At site S1 1073 more individualswere noted on hermit crab shells than on the next most abundantsubstratum (gastropods), 4878 at S2, 94 at S4 and 176 at T1. Atstations S3 and T3 gastropod shells and pebbles respectively weresupporting larger numbers of individuals than hermit crab shells.At S3 gastropod shells had 3253 individuals more than hermitcrabs. At T3 gastropod shells had 1707 and pebbles 1179 individualsmore than hermit crab shell (Fig. 2).
Table 2Species richness and abundance of epifauna occurring on gastropod shells inhabited by
Geographic region Hermit crab species N Totspe
Greenland Sea, 78�N Pagurus pubescens (Krøyer, 1838) 163 69Norwegian Sea, 69�N Pagurus pubescens (Krøyer, 1838) 51 41Norwegian Sea, 69�N Pagurus bernhardus (Linnaeus, 1758) 33 34North Sea, 58�N Pagurus bernhardus (Linnaeus, 1758) 11 18North Sea, 55�N Pagurus bernhardus (Linnaeus, 1758) 70Irish Sea, 51�N Pagurus bernhardus (Linnaeus, 1758) 300 10Western Atlantic, 39�N Pagurus pollicaris (Say, 1817) 53 7Gulf of Mexico, 27�N Pagurus pollicaris (Say, 1817) 82 7Gulf of Mexico, 27�N Pagurus longicarpus (Say, 1817) 32 7
When the analysis was limited to a similar number of gastro-pod species (Buccinum undatum, Buccinum glaciale, Buccinumpolare Gray, 1839; Buccinum scalariforme Møller, 1842) accumu-lation curves showed exactly the same patterns as the oneobserved for randomly collected shells (Fig. 2). Hermit crabs har-bored up to 16 species more than the next most species richsubstratum. Also more individuals of epifauna were recorded onhermit crab shells (except S3 and T3) than on gastropod shells andpebbles. At stations S3 and T3, gastropod shells and pebblesrespectively were supporting a larger numbers of individuals thanhermit crab shells.
Multidimensional scaling (nMDS) of epifaunal species compo-sition on hermit crab, gastropod shells and pebbles did not showany clear pattern between the assemblages at all sites (Fig. 3). One-way analyses of similarities (ANOSIM) revealed differences only attwo (S2 and S3) out of six investigated sites. At these locationsdifferences between the three substrata were present only whencomparing its species list (Global R on presence/absencedata ¼ 0.473 and 0.471 respectively, p ¼ 0.001). At S2 only thecomposition of pebbles and hermit crab shells differed from eachother (Pairwise R ¼ 0.563, p ¼ 0.001). In the case of S3, pebblesdiffered from both hermit crab and gastropod shells (PairwiseR ¼ 0.569 and 0.652, respectively, p ¼ 0.001).
Detailed examination of species composition revealed epifaunaof hermit crabs, gastropod shells and pebbles (for completedataset see Appendix 1 e on-line material) to be dominated bya barnacle Semibalanus balanoides (Linnaeus, 1758) at all the WestSpitsbergen sites. At the Northern Norway sites epifauna of hermitcrab shells were dominated by juvenile serpulid polychaetes,while gastropod shells and pebbles by adults of Spirobranchustriqueter (Linnaeus, 1758). There were 22 taxa (18 taxa found atWest Spitsbergen, 8 taxa in Northern Norway) that occurred onlyon hermit crab shells (Table 3). Among them the spirorbid poly-chaete Bushiella evoluta (Bush, 1905) was the most abundant, tenspecies were frequently found although the rest have been rarelynoted (number of individuals lower than five). Two taxa, bryozoanRhamphostomella sp. and foraminiferan Buccella frigida (Cushman,1921), were found exclusively on gastropods but only at WestSpitsbergen. Five taxa were found only on pebbles in both local-ities (West Spitsbergen and Northern Norway) (Table 3). All of the
hermit crabs across latitudes.
al no. ofcies
Average no. ofspecies/shell
Average no. ofind./shell
Reference
7.8 71 This study6.0 32 This study5.3 30 This study7.9 62 Reiss et al., 20038.3 90 Reiss et al., 20032.3 Bell, 20053.3 Karlson and Shenk, 19834.6 Conover, 19791.4 Conover, 1979

200 400 600 800 1000 1200 1400 1600 1800 2000 2200 2400 2600 28000
5
10
15
20
25
30
35
40
45
50
Spec
ies
S11000 2000 3000 4000 5000 6000 7000
0
5
10
15
20
25
30
35
40
45
50
S2
1000 2000 3000 4000 5000 60000
5
10
15
20
25
30
35
40
Spec
ies
S3100 200 300 400 500
0
2
4
6
8
10
12
14
16
18
20
S4
500 1000 1500 2000 2500 3000 3500 40000
5
10
15
20
25
30
35
40
T3100 200 300 400 500 600 700
0
5
10
15
20
25
30
35
Spec
ies
T1
Fig. 2. Species accumulation curves for epifauna found on hermit crab shell (triangle), live gastropod (square), pebble (circle) at different sites (S1, S2, S3, S4 e Isfjorden, WestSpitsbergen; T1, T3 e Tromsø, Northern Norway). In each case the number of samples of the three substrata was equal (N ¼ 50) except site S4 (N ¼ 14).
P. Balazy, P. Kuklinski / Estuarine, Coastal and Shelf Science 119 (2013) 153e161 157
taxa found on gastropods and pebbles were very rare and themajority of them were recorded only once and there was nevermore than 3 individuals.
4. Discussion
This study clearly shows that hermit crab shells provide a uniquesubstratum for epifauna. Species richness was much higher onhermit crab shells than on the shells of similar size and heteroge-neity of live gastropods or pebbles of even larger surface area. Thesame pattern was exhibited at all studied localities. If we take intoaccount that similar observations on a smaller scalewere also notedin other geographical regions, including western Atlantic (Conover,
1979; Karlson and Cariolou, 1982; Karlson and Shenk, 1983), IrishSea (Bell, 2005), Adriatic Sea (Stachowitsch, 1980), and Mediterra-nean Sea (Bick, 2006) it appears to be a widely recorded phenome-non. The differences in number of taxa and individuals associatedwith hermit crabs between geographic regions probably representsdisparities in sampling efforts of the hermit crab biocenoses ratherthan real differences in diversity patterns (Table 2, Williams andMcDermot, 2004). Our study supports these findings as the aver-age number of species associated with Pagurus pubescens shells inWest Spitsbergen andNorthernNorwaywas comparable. The largernumber of total species and average number of individuals inspecimens from West Spitsbergen results from over three timesa larger set of samples and a twice larger shell area, respectively.

2D Stress: 0.21 2D Stress: 0.19
S2S1
2D Stress: 0.08
S3
2D Stress: 0.09
S4
2D Stress: 0.18
T1
2D Stress: 0.15
T3
Fig. 3. nMDS plots based on zero-adjusted BrayeCurtis similarities of standardized abundances in samples on hermit crab (triangle), live gastropod shells (square) and pebbles(circle) at different sites (S1, S2, S3, S4 e Isfjorden, West Spitsbergen; T1, T3 e Tromsø, Northern Norway).
P. Balazy, P. Kuklinski / Estuarine, Coastal and Shelf Science 119 (2013) 153e161158
Epifaunal organisms living on the surface of hermit crab shellsbenefit frommany positive features of this substratum, which otherhabitats do not possess (Fernandez-Leborans, 2010). The mobilityof the crab seems to be one of the most important factors drivingincreased epifaunal biodiversity on their shells and it implies manyother positive parameters including enhanced food supply or pro-tection from predation. The constant movement of hermit crabsprevents epifauna from becoming buried in sediment (Conover,1975; Stachowitsch, 1977). Movement or shell elevation above thesediment seems to play an extremely important role especially insoft-bottom sites (e.g. sites S3 and S4) where high sedimentation ofmineral particles caused by melting glaciers is observed (pers. ob-servations). This was most evident at the soft bottom site S3 wherethe highest relative number of species on hermit crab shells wasrecorded. The fact that it was not shown at the other soft bottomsite S4 is most probably caused by low sampling effort as only 14hermit crabs were collected. The higher species richness recordedon gastropod shells compared with pebbles recorded at those twosoft bottom sites additionally supports the importance of substra-tum mobility. Gastropods and hermit crabs are capable of moving
on the surface of the soft bottom avoiding burial while pebbleswere often partly buried by sediment preventing colonization. Inplaces with hard bedrock (sites S1, S2, T1, T3) the pebbles weresupporting a higher biodiversity in comparison to gastropod shells.This is probably attributed to longer exposure of this substratum tobiotic colonization as pebbles at these sites can be exposed on thebottom for tens, if not hundreds of years while gastropod life spanshardly ever exceed twenty years.
Both hermit crab shells and shells of live gastropods providestable substrata not susceptible to water movement (Stachowitsch,1980; Kuklinski et al., 2008). They are likely to maintain their ver-tical position as they are supported by their host, which is able holdto the sea floor by use of its appendages or foot. It is believed thatowing to these characteristics, the probability of destruction ofassemblages occurring on hermit crab and gastropod shells is muchlower. In contrast, pebbles, influenced by wave action, can bemoved around, which can destroy epifauna. Such factors result inlonger periods of undisturbed growth and development of epifaunaon hermit crab and gastropod shells than on pebbles of similar size(Conover, 1975; Williams and McDermot, 2004). Even when the

Table 3List of taxa found only on given substratum. Rare taxa (number of individuals lower than five) are shown in gray. S e Isfjorden, West Spitsbergen; T e Tromsø, NorthernNorway.
Phylum Hermit crab shells Site Gastropod shells Site Rocks Site
Annelida Bushiella evoluta (Bush, 1905) S TChitinopomoa serrula (Stimpson, 1854) SSerpulidae undet. S
Arthropoda Balanus balanus (Linnaeus, 1758) S T Crustacea undet. TBalanus crenatus (Bruguiére, 1789) S
Bryozoa Callopora lineata (Linnaeus, 1767) S T Rhamphostomella sp. S Membraniporella nitida (Johnston, 1838) TCallopora smitti Kluge, 1946 S Porella alba (Nordgaard, 1906) SCribrilina cryptooecium Norman, 1903 S TCribrilina sp. TDendrobeania murrayana (Bean, in Johnston, 1847) SDendrobeania sp. SElectra pilosa (Linnaeus, 1767) TOncousoecia sp. SOshurkovia littoralis (Hastings, 1944) TRaymondcia rigida (Lorenz, 1886) SReussia sp. SSchizoporella pachystega (Kluge, 1929) SSeptentriopora karasi (Kuklinski and Taylor, 2006) SStomachetosella producta (Packard, 1863) STegella armifera (Hincks, 1880) SBryozoa ancestrulla S
Echinodermata Asteroidea undet. TForaminifera Buccella frigida (Cushman, 1921) SMollusca Bivalvia undet. TPorifera Porifera undet. T
P. Balazy, P. Kuklinski / Estuarine, Coastal and Shelf Science 119 (2013) 153e161 159
crab changes the shell there is large probability that anotherspecimenwill inhabit it shortly afterward since gastropod shells aregenerally a limiting resource in most hermit crab communities(Kellogg, 1976; Williams and McDermot, 2004; Barnes et al., 2007).
In addition, by using a mobile substratum, hermit crab associ-ates avoid slow-moving predators (Conover, 1979), such as sea ur-chins, which are common in the Arctic, and are able to crossunsuitable habitats (e.g. fine sediment patches) (Bell, 2009).Another potential advantage of life on the hermit crab shell is theaccess to increased food supplies provided by host excreta andmovement of its mouth parts and antennae (Heath, 1976;Stachowitsch, 1977; Conover, 1979 and references therein).This constant movement also supplies well-oxygenated water(Stachowitsch, 1980).
Shells inhabited by hermit crabs are shells of previously livegastropods. Therefore, their exposure to colonization is longer andtheir epifauna could be a later and thus more diverse successionalstage than the epifauna of younger live gastropods. In addition itwas shown that marine invertebrate larvae prefer to settle onrough, irregular surfaces (Crisp and Ryland, 1960). Settling studiesperformed on a common hermit crab epibiont hydrozoan Hydrac-tinia echinata (Fleming, 1828) have shown that its larvae (planulae)preferred to settle in the pits and grooves of species with sculpturedshells rather than on species with smoother shells (Teitelbaum,1966). This shows that older shells with uneven surfaces andprobably without the organic periostracum layer present in livegastropods, probably increases settled larval survival (Dvoretskyand Dvoretsky, 2008). Therefore, higher species richness on her-mit crab shells is also enhanced by its heterogeneous surfaceespecially in comparison to pebbles, which are often smooth. In oursamples, the number of shell species among hermit crabs washigher than within live gastropods leading probably to higheroverall substratum heterogeneity of the former. This may havecontributed to the higher number of epifaunal species observed onhermit crab rather than gastropod shells. However, when wereduced hermit crab shells to the same species found in the livecollected group (only Buccinum sp. shells e mostly Buccinum gla-ciale and Buccinum undatum) exactly the same patterns at all sites
were recorded with hermit crabs harboring the most species richassemblages. This clearly shows that heterogeneity of the shells isnot the only factor leading to the highest species richness on hermitcrab shells.
At most of the study sites, hermit crabs were also supportingmore individuals of epifauna. However, at S3 and T3 numericalabundance of epifauna on gastropod shells (and pebbles in thecase of T3) was higher than on the shells used by hermit crabs.This was the result of one species occurring on gastropod shells/pebbles in large numbers. At S3 (West Spitsbergen) it was thebarnacle Semibalanus balanoides while at T3 (Northern Norway) itwas the serpulid polychaete Spirobranchus triqueter. Both speciesare known to produce large numbers of larvae which dominate inthe water column in late spring e early summer as well juvenileindividuals later during the season on all firm bottom substrata(Kuklinski et al., in press). The composition of epifaunal assem-blages at the earliest successional stage for both gastropod shellsand pebbles often seems to be a function of larval abundanceduring the time of given substratum exposure (Osman, 1977).Most likely our samples were taken during the settlement periodof barnacles and serpulids resulting in large numbers of theseopportunistic species on gastropods and pebbles. Furthermore, itis known that the settlement of marine invertebrate larvae isinfluenced by the presence of other co-occurring organisms(Grosberg, 1981). Semibalanus balanoides and some sedentarypolychaete species exhibit gregarious behavior during settlementin response to the occurrence of individuals of the same species(Knight-Jones, 1951; Crisp, 1961; Scheltema et al., 1981). On theother hand, a number of epifaunal species inevitably limit the areaavailable for colonization of others and space can be a limitingfactor for assemblage development (Menge and Sutherland, 1976and references therein,; Kuklinski et al., 2008). Therefore, spatialinteractions such as competition and overgrowth may havea direct effect on the final structure and the composition of theencrusting communities. Old, established assemblages on hermitcrab shells most probably prevent larvae from settling in suchlarge numbers, which is not the case with gastropods and pebblesthat have a less diverse assemblage structure.

P. Balazy, P. Kuklinski / Estuarine, Coastal and Shelf Science 119 (2013) 153e161160
Multidimensional scaling (nMDS) indicates that epifaunal as-semblages occurring on hermit crabs do not differ significantlyfrom those on gastropod shells and pebbles regardless of the bot-tom type (soft vs. hard bedrock). Dominance structure between thethree substrata was also similar suggesting comparable nature ofthese three firm habitats. Themain dominants arewell known fromthe literature. For example, barnacles and the tubeworm Circeisarmoricana Saint-Joseph, 1894 were described as one of the mostcommon components of fouling communities of hermit crabs(Reiss et al., 2003; Williams and McDermot, 2004). However,detailed analysis of the species composition on shells of hermitcrabs, gastropods and pebbles revealed that from 87 taxa found onall three substrata there were 22 that were present only on hermitcrab shells (Table 3). According to Williams and McDermot (2004and references therein) from at least 550 invertebrate speciesassociated with hermit crabs 114 are obligate commensals includ-ing bryozoans, cnidarians, polychaetes and cirripedes. However, inour study most epifauna are not obligate associates of hermit crabs,assuming that some of the taxonomical determination which wereto genus level only (juveniles, damaged individuals) do not includespecies specific to this substratum. Only the spirorbid Bushiellaevoluta is found attached solely to the inner side of gastropod shellsused by hermit crabs (Jirkov, 2001, personal observations). Other-wise a detailed literature search failed to identify any species foundduring this investigation that is specific to hermit crab shells.Therefore, epifaunal substratum specificity can be ruled out asa factor responsible for the high species richness observed onhermit crab shells. Gastropod shells and pebbles also lacked anyspecies specific to these two substrata. All epifaunal species werevery rare and nearly half of them represented mobile fauna such asjuvenile Asteroidea, Crustacea and Bivalvia also present in thesurrounding area. The remaining two bryozoan and one Foramin-ifera species (Table 3) are also known to live on other surfaces(Kuklinski and Barnes, 2005).
5. Conclusions
This study together with similar findings from other geographicregions show that hermit crabs shells are an example of hard mo-bile substrata that support higher biodiversity than would be pre-dicted by their surface area alone, not only at high latitudes but alsoworldwide. Gastropod shells inhabited by hermit crabs provide anexceptional substratum which increases epifaunal species numberand, in most of the cases, also relative abundance in comparison tosubstrata of similar complexity and surface area, regardless ofbottom type. However, a stronger impact on epifauna of this mobilehard substratum is more evident in soft bottom areas where theysupport higher proportion of species than gastropod shells andpebbles in comparison to rocky bottoms. In soft bottom areas,hermit crab shells are often one of few firm surfaces available andgastropods and pebbles are buried in the sediment limiting epi-faunal development thus leading to observed differences in bio-diversity values among these substrata. The observed pattern ofhighest diversity of epifauna on hermit crab shell is not caused bysymbiosis as, with one exception, species specific to these shellswere not found. It is the result of number of factors interactingpositively on the presence of epifauna including shell surface het-erogeneity and the complex influence of its host - the hermit crab.
Acknowledgments
The authors wish to thank: Jakub Beszczynski for his under-water assistance, Dominika Wojcieszek, Maria Wlodarska-Kowalczuk, Suzanne Williams, John Taylor and two anonymousreviewers for comments leading to a much-improved manuscript.
This study has been completed thanks to a grants from the PolishMinistry of Science and Higher Education (396/N-EOL-ENC/2009/0)and from the Polish National Science Centre (No 2011/01/N/NZ8/04493).
Appendix A. Supplementary data
Supplementary data related to this article can be found at http://dx.doi.org/10.1016/j.ecss.2013.01.004.
References
Appeltans, W., Ahyong, S.T., Anderson, G., Angel, M.V., Artois, T., Bailly, N.,Bamber, R., et al., 2012. The magnitude of global marine species diversity.Current Biology 22, 1e14.
Barnes, D.K.A., Kuklinski, P., 2005. Low colonization on artificial substrata in arcticSpitsbergen. Polar Biology 29, 65e69.
Barnes, D.K.A., Kuklinski, P., Wlodarska-Kowalczuk, M., 2007. Richness, abun-dance and shell use of subarctic and arctic hermit crabs. Marine Biology 152,1133e1142.
Bell, J.J., 2005. Influence of occupant microhabitat on the composition of encrustingcommunities on gastropod shells. Marine Biology 147, 653e661.
Bell, J.J., 2009. Hitching a ride on a hermit crabs home: movement of gastropodshells inhabited by hermit crabs. Estuarine, Coastal and Shelf Science 85,173e178.
Berge, J., Johnsen, G., Nilsen, F., Gulliksen, B., Slagstad, D., 2005. Ocean temperatureoscillations enable reappearance of blue mussels Mytilus edulis in Svalbard aftera 1000 year absence. Marine Ecology Progress Series 303, 167e175.
Bergey, E., Getty, G.M., 2006. A review of methods for measuring the surface area ofstream substrates. Hydrobiologia 556, 7e16.
Beuchel, F., Gulliksen, B., 2008. Temporal patterns of benthic community develop-ment in an Arctic fjord (Kongsfjorden, Svalbard): results of a 24-year manipu-lation study. Polar Biology 31, 913e924.
Bick, A., 2006. Polychaete communities associated with gastropod shells inhabitedby the hermit crabs Clibanarius erythropus and Calcinus tubularis from Ibiza,Mediterranean Sea. Journal of the Marine Biological Association of the UnitedKingdom 86, 83.
Birula, A., 1907. Zoologische Ergebnisse der Russischen Expedition nach Spitzber-gen. Crustacea-Decapoda. Annuaire du Musee Zoologique de l’Academie d.Sciences de St. Petersbourg 11, 1e68.
Bluhm, B.A., Gradinger, R., Hopcroft, R.R., 2011. Editorial e Arctic Ocean diversity:synthesis. Marine Biodiversity 41, 1e4.
Bray, J.R., Curtis, J.T., 1957. An ordination of the upland forest communities ofsouthern Wisconsin. Ecological Monographs 27, 325e349.
Chapin, F.S., Zavaleta, E.S., Eviner, V.T., Naylor, R.L., Vitousek, P.M., Reynolds, H.M.,Hooper, D.U., Lavorel, S., Sala, O.E., Hobbie, S.E., Mack, M.C., Diaz, S., 2000.Consequences of changing biodiversity. Nature 405, 234e242.
Chapman, M.G., Underwood, A.J., 1999. Ecological patterns in multivariate assem-blages: information and interpretation of negative values in ANOSIM tests.Marine Ecology Progress Series 180, 257e265.
Clarke, K.R., Gorley, R.N., 2001. PRIMER v6: User Manual/Tutorial. PRIMER - E,Plymouth.
Clarke, K.R., Green, R.H., 1988. Statistical design and analysis for a ‘biological effects’study. Marine Ecology Progress Series 46, 213e226.
Clarke, K.R., Somerfield, P., Chapman, M., 2006. On resemblance measures forecological studies, including taxonomic dissimilarities and a zero-adjustedBrayeCurtis coefficient for denuded assemblages. Journal of ExperimentalMarine Biology and Ecology 330, 55e80.
Conover, M.R., 1975. Prevention of shell burial as a benefit hermit crabs provide totheir symbionts (Decapoda, Paguridea). Crustaceana 29, 311e313.
Conover, M.R., 1979. Effect of gastropod shell characteristics and hermit crabs onshell Epifauna. Journal of Experimental Marine Biology and Ecology 40, 81e94.
Costello, M.J., Coll, M., Danovaro, R., Halpin, P., Ojaveer, H., Miloslavich, P., 2010.A census of marine biodiversity knowledge, resources, and future challenges.PloS One 5, e12110. 1e15.
Crisp, D.J., 1961. Territorial behaviour in barnacle settlement. Journal of Exper-imental Biology 38, 429e446.
Crisp, D.J., Ryland, J.S., 1960. Influence of filming and of surface texture on thesettlement of marine organisms. Nature 185, 119.
Dale, J.E., Aitken, A.E., Gilbert, R., Risk, M.J., 1989. Macrofauna of Canadian arcticfjords. Marine Geology 85, 331e358.
Dawson, T.P., Jackson, S.T., House, J.I., Prentice, I.C., Mace, G.M., 2011. Beyond pre-dictions: biodiversity conservation in a changing climate. Science 332, 53e58.
Dvoretsky, A.G., Dvoretsky, V.G., 2008. Epifauna associated with the northern stonecrab Lithodes maia in the Barents Sea. Polar Biology 31, 1149e1152.
Fernandez-Leborans, G., 2010. Epibiosis in Crustacea: an overview. Crustaceana 83,549e640.
Grosberg, R.K., 1981. Competitive ability influences habitat choice in marine in-vertebrates. Nature 290, 700e702.
Gulliksen, B., Svendsen, E., 2004. Svalbard and Life in Polar Oceans. Kom Forlag,160 pp.

P. Balazy, P. Kuklinski / Estuarine, Coastal and Shelf Science 119 (2013) 153e161 161
Gutierrez, J.L., Jones, C.G., Strayer, D.L., Iribarne, O.O., 2003. Mollusks as ecosystemengineers: the role of shell production in aquatic habitats. Oikos 1, 79e90.
Hazlett, B.A., 1981. The behavioral ecology of hermit crabs. Annual Review ofEcology, Evolution, and Systematics 12, 1e22.
Heath, D.J., 1976. The distribution and orientation of epizoic barnacles on crabs.Zoological Journal of the Linnean Society 59, 59e67.
Heegaard, P.E., 1941. The zoology of East Greenland: decapod crustaceans. MeddGroe 121, 1e72.
Heip, C., 2003. What structures marine biodiversity and why does it vary? In:Wefer, G., Lamy, F., Mantoura, F. (Eds.), Marine Science Frontiers for Europe.Springer-Verlag, Berlin, Heidelberg, New York, Tokyo, pp. 251e264.
Holte, B., Oug, E., 1996. Soft-bottom macrofauna and responses to organic enrich-ment in the subarctic waters of Tromsø, Northern Norway. Journal of SeaResearch 36, 227e237.
IPCC, 2001. Climate Change 2001: Synthesis Report. A Contribution of WorkingGroups I, II, and III to the Third Assessment Report of the IntergovernmentalPanel on Climate Change. Cambridge University Press, Cambridge, and NewYork, 398 pp.
IPCC, 2007. Climate Change 2007: Synthesis Report. Contribution of WorkingGroups I, II and III to the Fourth Assessment Report of the IntergovernmentalPanel on Climate Change. IPCC, Geneva, 104 pp.
Jensen, K., Bender, K., 1973. Invertebrates associated with snail shells inhabited byPagurus bernhardus (L.) (Decapoda). Ophelia 10, 185e192.
Jirkov, I.A., 2001. Polychaeta of the Arctic Ocean. Yanus-K, Moscow, 632 pp.Jones, C.G., Lawton, J.H., Shachak, M., 1994. Organisms as ecosystem engineers.
Oikos 69, 373e386.Jones, C.G., Lawton, J.H., Shachak, M., 1997. Positive and negative effects of organ-
isms as physical ecosystem engineers. Ecology 78, 1946e1957.Karlson, R., Cariolou, M., 1982. Hermit crab shell colonization by Crepidula convexa
Say. Journal of Experimental Marine Biology and Ecology 65, 1e10.Karlson, R.H., Shenk, M.A., 1983. Epifaunal abundance, association, and overgrowth
patterns on large hermit crab shells. Journal of Experimental Marine Biologyand Ecology 70, 55e64.
Kellogg, C.W., 1976. Gastropod shells: a potentially limiting resource for hermitcrabs. Journal of Experimental Marine Biology and Ecology 22, 101e111.
Knight-Jones, E.W., 1951. Gregariousness and some other aspects of the settlingbehaviour of Sipirorbis. Journal of the Marine Biological Association of theUnited Kingdom 30, 201e222.
Kuklinski, P., Barnes, D.K.A., 2005. Microhabitat diversity of Svalbard Bryozoa.Journal of Natural History 39, 539e554.
Kuklinski, P., Gulliksen, B., Lonne, O.J., Weslawski, J.M., 2006. Substratum as a struc-turing influence on assemblages of Arctic bryozoans. Polar Biology 29, 652e661.
Kuklinski, P., Barnes, D.K.A., Wlodarska-Kowalczuk, M., 2008. Gastropod shells,hermit crabs and arctic bryozoan richness. In: Hageman, S.J., Key Jr., M.M.J.,Winston, J.E. (Eds.), Bryozoan Studies 2007. Proceedings of the 14th Interna-tional Bryozoology Association Conference. Virginia Museum of Natural HistorySpecial Publication No. 15, Martinsville, pp. 93e100.
Kuklinski, P., Berge, J., McFadden, L., Dmoch, K., Zajaczkowski, M., Nygard, H.,Piwosz, K., Tatarek, A. Seasonality of occurrence and recruitment of Arcticmarine benthic invertebrate larvae in relation to environmental variables. PolarBiology, in press.
Lancaster, I., 1988. Pagurus bernhardus (L.) e an introduction to the natural historyof hermit crabs. Field Studies 7, 189e238.
Lancaster, I., 1990. Reproduction and life history strategy of the hermit crab Pagurusbernhardus. Journal of the Marine Biological Association of the United Kingdom70, 129e142.
McLean, R., 1983. Gastropod shells: a dynamic resource that helps shape benthic com-munity structure. Journal of Experimental Marine Biology and Ecology 69, 151e174.
Menge, B.A., Sutherland, J.P., 1976. Species diversity gradients: synthesis of the rolesof predation, competition, and temporal heterogeneity. American Naturalist110, 351e369.
Nilsen, F., Cottier, F., Skogseth, R., Mattsson, S., 2008. Fjord e shelf exchangescontrolled by ice and brine production: the interannual variation of Atlanticwater in Isfjorden, Svalbard. Continental Shelf Research 28, 1838e1853.
Osman, R.W., 1977. The establishment and development of a marine epifaunalcommunity. Ecological Monographs 47, 37e63.
Oug, E., 1998. Relating species patterns and environmental variables by canonicalordination: an analysis of soft-bottom macrofauna in the region of Tromsø,northern Norway. Marine Environmental Research 45, 29e45.
Oug, E., 2001. Polychaetes in intertidal rocky and sedimentary habitats in the regionof Tromsø, Northern Norway. Sarsia 86, 75e83.
Pereira, H.M., Leadley, P.W., Proenca, V., Alkemade, R., Scharlemann, J.P.W., Fer-nandez-Manjarrés, J.F., Araújo, M.B., et al., 2010. Scenarios for global bio-diversity in the 21st century. Science 330, 1496e1501.
Piepenburg, D., 2005. Recent research on Arctic benthos: common notions need tobe revised. Polar Biology 28, 733e755.
Reiss, H., Knauper, S., Kroncke, I., 2003. Invertebrate associationswith gastropod shellsinhabited by Pagurus bernhardus (Paguridae) secondary hard substrate increasingbiodiversity in North Sea soft-bottom communities. Sarsia 88, 404e415.
Renaud, P.E., Carroll, M.L., Ambrose Jr., W.G., 2008. Effects of global warming onArctic sea-floor communities and its consequences for higher trophic levels. In:Duarte, C. (Ed.), Impacts of Global Warming on Polar Ecosystems. FBBVA Press,Bilbao, pp. 139e177.
Sandberg, L., McLaughlin, P.A., 1988. Crustacea, Decapoda, Paguridea. Marine in-vertebrates of Scandinavia No. 10. Scandinavian University Press, Oslo, 113 pp.
Scheltema, R.S., Williams, I.P., Loudon, C., 1981. Gregarious settlement by the larvaeof Hydroides. Marine Ecology Progress Series 5, 69e74.
Schiermeier, Q., 2007. The new face of the Arctic. Nature 446, 133e135.Stachowitsch, M., 1977. The hermit crab micro biocenosis e the role of mobile
secondary hard bottom elements in a north Adriatic benthic community. In:Keegan, B.F. (Ed.), Biology of Benthic Organisms. Proceedings of the EuropeanSymposium in Marine Biology. Pergamon Press, Oxford, pp. 549e558.
Stachowitsch, M., 1980. The epibiotic and endolithic species associated with thegastropod shells inhabited by the hermit crabs Paguristes oculatus and Paguruscuanensis. Marine Ecology 1, 73e101.
Teitelbaum, M., 1966. Behavior and Settling Mechanism of Planulae of Hydractiniaechinata, vol. 131. Biological Bulletin, Marine Biological Laboratory, Woods Hole,pp. 410e411.
Tittensor, D.P., Mora, C., Jetz, W., Lotze, H.K., Ricard, D., Berghe, E.V., Worm, B., 2010.Global patterns and predictors of marine biodiversity across taxa. Nature 466,1098e1101.
Tukey, J.W., 1953. Some selected quick and easy methods of statistical analysis.Transactions of the New York Academy of Sciences Series II 16, 88e97.
Wang, M., Overland, J.E., 2009. A sea ice free summer Arctic within 30 years?Geophysical Research Letters 36, L07502.
Weslawski, J.M., Wiktor Jr., J., Kotwicki, L., 2010. Increase in biodiversity in the arcticrocky littoral, Sorkappland, Svalbard, after 20 years of climate warming. MarineBiodiversity 40, 123e130.
Weslawski, J.M., Kendall, M., Wlodarska-Kowalczuk, M., Iken, K., Kedra, M.,Legezynska, J., Sejr, M.K., 2011. Climate change effects on Arctic fjord and coastalmacrobenthic diversity e observations and predictions. Marine Biodiversity 41,71e85.
Williams, J.D., McDermot, J.J., 2004. Hermit crab biocoenoses: a worldwide reviewof the diversity and natural history of hermit crab associates. Journal ofExperimental Marine Biology and Ecology 305, 1e128.
Van Winkle, D.H., Longnecker, K., Blackstone, N.W., 2000. The effects of hermitcrabs on hydractiniid hydroids. Marine Ecology 21, 55e67.
Wlodarska-Kowalczuk, M., Szymfeling, M., Zajaczkowski, M., 2007. Dynamic sedi-mentary environments of an Arctic glacier-fed river estuary (Adventfjorden,Svalbard). II. Meio- and macrobenthic fauna. Estuarine, Coastal and Shelf Sci-ence 74, 274e284.
Worm, B., Barbier, E.B., Beaumont, N., Duffy, J.E., Folke, C., Halpern, B.S.,Jackson, J.B.C., et al., 2006. Impacts of biodiversity loss on ocean ecosystemservices. Science 314, 787e790.
Zajaczkowski, M., Wlodarska-Kowalczuk, M., 2007. Dynamic sedimentary envi-ronments of an Arctic glacier-fed river estuary (Adventfjorden, Svalbard). I.Flux, deposition and dynamics of the sediment. Estuarine, Coastal and ShelfScience 74, 285e296.
Related Documents











