Mitosis in diatoms: rediscovering an old model for cell division Alessandra De Martino, 1 Alberto Amato, 1 and Chris Bowler 1,2 * 1 Laboratory of ‘Molecular Biology of Photosynthetic Organisms’, UMR 8186, Department of Biology, Ecole Normale Supe ´rieure, 46 rue d’Ulm, 75230 Paris cedex 05, France 2 Stazione Zoologica Anton Dohrn, Villa Comunale, I-80121 Naples, Italy Diatoms are important protists that generate one fifth of the oxygen produced annually on earth. These aquatic organisms likely derived from a secondary endosymbio- sis event, and they display peculiar genomic and struc- tural features that reflect their chimeric origin. Diatoms were one of the first models of cell division and these early studies revealed a range of interesting features including a unique acentriolar microtubule-organising centre. Unfortunately, almost nothing is known at the molecular level, in contrast to the advances in other experimental organisms. Recently the full genome sequences of two diatoms have been annotated and molecular tools have been developed. These resources offer new possibilities to re-investigate the mechanisms of cell division in diatoms by recruiting information from more intensively studied organisms. A renaissance of the topic is further justified by the current interest in diatoms as a source of biofuels and for understanding massive diatom proliferation events in response to environmental stimuli. Keywords: cell division; diatom; mitosis; MTOC; spindle Introduction Our understanding of cell division derives almost entirely from intensively studied model organisms. But the living world is wider and more diversified than these elite models and many organismal groups hide peculiarities that deserve to be studied and understood. Diatoms (class Bacillariophyceae) are one of the most species-rich algal groups and play a fundamental role in marine primary production and in biogeochemical cycles of several elements such as carbon, silicon, nitrogen and phosphorus. (1) They derive from a secondary endosymbiosis in which a photosynthetic eukaryote was engulfed by another eukaryote. (2) The guest and host genomes then merged yielding a chimeric genome. (3–5) The most well-known feature of diatoms is their highly ornamented external cell wall made of amorphous silica, (6) called the frustule, which is composed of two unequal valves, the epivalve fitting onto the hypovalve like a lid fits onto its box. They predominantly divide vegetatively, through mitosis and cytokinesis. During cytokinesis each daughter cell inherits one maternal valve as the epivalve, while the hypovalve is synthesised de novo within the mother cell. (7,8) Nuclear division and cytokinesis were studied in diatoms as early as 1896 by Lauterborn (9) in an extensive dissertation, which was then translated and summarised almost one century later. (10) For several diatoms, mitosis has been studied by light and transmission electron microscopy (TEM), but there are only a few reports from a molecular standpoint. The diatom genera for which mitosis has been most commonly studied belong to the order Pennales (pennate diatoms (7) ), in particular Surirella (Supporting Information Table S1). Many of these studies were innovative at that time and contained impressive microscopic observations, and they revealed a unique microtubule (MT)-organising centre (MTOC), called the MT centre (MC). The MC shows peculiar structure and behaviour during the cell cycle. In the late 1980s, Cande and coworkers worked intensively on spindle elongation in the centric diatom Stephanopyxis turris both in vivo and by isolating mitotic spindles and studying them in vitro using different approaches such as indirect immunofluorescence. (11–13) Only in the mid 1990s diatom mitosis was investigated molecularly, first in the pennate Cylindrotheca fusiformis, revealing the role of diatom spindle kinesin 1, a novel kinesin-related protein (KRP), in spindle elongation. (14–16) Now that whole genome sequences from diatoms are available (3,5) as well as high- throughput reverse genetics, (17–19) the molecular mechan- isms involved in diatom mitosis and the distinctive features characterising diatom biology (see below and (7) ) warrant re-investigation. Review article DOI 10.1002/bies.200900007 Abbreviations: Bb, basal body; CDK, cyclin-dependent kinase; EB1, end- binding protein 1; EM, electron microscopy; GCP, g-tubulin complex protein; g- tub, g-tubulin; KRP, kinesin-related protein; MC, microtubule centre; MT, microtubule; MTOC, microtubule-organising centre; NE, nuclear envelope; NEBD, NE breakdown; PC, polar complex; SPB, spindle polar body; TEM, transmission electron microscopy; YFP, yellow fluorescent protein. *Correspondence to: C. Bowler, Laboratory of ‘Molecular Biology of Photosynthetic Organisms’, UMR 8186, Department of Biology, Ecole Normale Supe ´rieure, 46 rue d’Ulm, 75230 Paris cedex 05, France. E-mail: [email protected] 874 BioEssays 31:874–884, ß 2009 Wiley Periodicals, Inc.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Review article DOI 10.1002/bies.200900007
Mitosis in diatoms: rediscoveringan old model for cell divisionAlessandra De Martino,1 Alberto Amato,1 and Chris Bowler1,2*
1 Laboratory of ‘Molecular Biology of Photosynthetic Organisms’, UMR 8186, Department of Biology, Ecole Normale Superieure,46 rue d’Ulm, 75230 Paris cedex 05, France2Stazione Zoologica Anton Dohrn, Villa Comunale, I-80121 Naples, Italy
Diatoms are important protists that generate one fifth ofthe oxygen produced annually on earth. These aquaticorganisms likely derived from a secondary endosymbio-sis event, and they display peculiar genomic and struc-tural features that reflect their chimeric origin. Diatomswere one of the first models of cell division and theseearly studies revealed a range of interesting featuresincluding a unique acentriolar microtubule-organisingcentre. Unfortunately, almost nothing is known at themolecular level, in contrast to the advances in otherexperimental organisms. Recently the full genomesequences of two diatoms have been annotated andmolecular tools have been developed. These resourcesoffer new possibilities to re-investigate the mechanismsof cell division in diatoms by recruiting informationfrom more intensively studied organisms. A renaissanceof the topic is further justified by the current interestin diatoms as a source of biofuels and for understandingmassive diatom proliferation events in response toenvironmental stimuli.
Keywords: cell division; diatom; mitosis; MTOC; spindle
Introduction
Our understanding of cell division derives almost entirely from
intensively studied model organisms. But the living world is
wider and more diversified than these elite models and many
organismal groups hide peculiarities that deserve to be
studied and understood. Diatoms (class Bacillariophyceae)
are one of the most species-rich algal groups and play a
fundamental role in marine primary production and in
biogeochemical cycles of several elements such as carbon,
silicon, nitrogen and phosphorus.(1) They derive from a
Abbreviations: Bb, basal body; CDK, cyclin-dependent kinase; EB1, end-
binding protein 1; EM, electron microscopy; GCP, g-tubulin complex protein; g-
tub, g-tubulin; KRP, kinesin-related protein; MC, microtubule centre; MT,
microtubule; MTOC, microtubule-organising centre; NE, nuclear envelope;
NEBD, NE breakdown; PC, polar complex; SPB, spindle polar body; TEM,
transmission electron microscopy; YFP, yellow fluorescent protein.
*Correspondence to: C. Bowler, Laboratory of ‘Molecular Biology of
Photosynthetic Organisms’, UMR 8186, Department of Biology, Ecole
Normale Superieure, 46 rue d’Ulm, 75230 Paris cedex 05, France.
E-mail: [email protected]
874
secondary endosymbiosis in which a photosynthetic
eukaryote was engulfed by another eukaryote.(2) The guest
and host genomes then merged yielding a chimeric
genome.(3–5)
The most well-known feature of diatoms is their highly
ornamented external cell wall made of amorphous silica,(6)
called the frustule, which is composed of two unequal
valves, the epivalve fitting onto the hypovalve like a lid fits
onto its box. They predominantly divide vegetatively, through
mitosis and cytokinesis. During cytokinesis each daughter
cell inherits one maternal valve as the epivalve, while
the hypovalve is synthesised de novo within the mother
cell.(7,8)
Nuclear division and cytokinesis were studied in diatoms
as early as 1896 by Lauterborn(9) in an extensive dissertation,
which was then translated and summarised almost one
century later.(10) For several diatoms, mitosis has been
studied by light and transmission electron microscopy (TEM),
but there are only a few reports from a molecular standpoint.
The diatom genera for which mitosis has been most
commonly studied belong to the order Pennales (pennate
diatoms(7)), in particular Surirella (Supporting Information
Table S1). Many of these studies were innovative at that time
and contained impressive microscopic observations, and they
revealed a unique microtubule (MT)-organising centre
(MTOC), called the MT centre (MC). The MC shows peculiar
structure and behaviour during the cell cycle.
In the late 1980s, Cande and coworkers worked intensively
on spindle elongation in the centric diatom Stephanopyxis
turris both in vivo and by isolating mitotic spindles and
studying them in vitro using different approaches such as
indirect immunofluorescence.(11–13) Only in the mid 1990s
diatom mitosis was investigated molecularly, first in the
pennate Cylindrotheca fusiformis, revealing the role of
diatom spindle kinesin 1, a novel kinesin-related protein
(KRP), in spindle elongation.(14–16) Now that whole genome
sequences from diatoms are available(3,5) as well as high-
throughput reverse genetics,(17–19) the molecular mechan-
isms involved in diatom mitosis and the distinctive features
characterising diatom biology (see below and(7)) warrant
re-investigation.
BioEssays 31:874–884, � 2009 Wiley Periodicals, Inc.

A. De Martino, A. Amato and C. Bowler Review article
The microtubule centre of diatoms: anacentriolar MTOC with atypical featuresand behaviour during the cell cycle
The term MTOC was first defined by Pickett-Heaps(20) to
designate structures from which MTs emanate. MTOCs play a
crucial role in the division of eukaryotic cells. They nucleate
MTs and determine temporal and spatial distributions of the
interphase cytoplasmic MT arrays and the mitotic bipolar
spindle. Alterations of MTOC function lead to defects in
polarity, cytoplasmic organisation and fidelity of transmission
of genetic material during cell division. For instance, failure to
correctly regulate MTOC number or structure has dramatic
consequences in animal cells, leading to monopolar or
multipolar spindles, a characteristic of cancer cells.(21)
The MTOCs of yeast and slime mould look very different
from animal centrosomes (Fig. 1A–D) in that they are
multilayer structures lacking centrioles. In Saccharomyces
cerevisiae, the spindle polar body (SPB) is totally embedded
in the nuclear envelope (NE) throughout the cell cycle, and is
able to nucleate both nuclear and cytoplasmic MTs.(22) By
contrast, the SPB of the fission yeast Schizosaccharomyces
pombe(23) and the MTOC of Dictyostelium discoideum are
cytoplasmic, lying adjacent to the NE (Fig. 1A–D).
Centrioles likely derived from a basal body (Bb) that
nucleated MTs in flagella and cilia. It is thought that Bbs were
present in an ancestor, and during the evolution of eukaryotic
cells both structures (Bb and centrioles) were either
maintained or differentially lost, as in somatic cells of higher
plants.(24) Notably, angiosperms lack conspicuous MTOCs(25)
and MTs appear to be nucleated from several dispersed sites
organised as cortical parallel arrays encircling the inner
plasma membrane during interphase.
Bbs are still present in flagellated unicellular algae (e.g.
Chlamydomonas reinhardtii) and flagellated gametes of some
other algae (e.g. brown algae or centric diatoms), but have
been lost in pennate diatoms.(7) Centrics genetically differ
from pennates as much as humans from fish,(5) and such a big
distance is also reflected in their sexual cycle. Bbs in the
microgametes of centric diatoms are the only centriole-like
structures present among diatoms.
The diatom acentriolar MC has an ultrastructure different
from other MTOCs (Fig. 1). It diverges from those of animals
by the absence of centrioles and it diverges from yeast SBP
and slime mould MTOC because it has a spherical structure. It
also differs from the MTOC in brown algae that have an
animal-like centrosome consisting of a pair of centrioles and a
pericentriolar matrix.(26) This highlights the diversity of MTOC
structure and organisation even in phylogenetically closely
related organisms.
The first light microscopic descriptions of diatom MCs were
reported by Lauterborn at the end of the 19th century in the
pennate Surirella calcarata,(9) and this work represents one of
BioEssays 31:874–884, � 2009 Wiley Periodicals, Inc.
the first illustrations of mitosis and cytokinesis in a single living
cell. Later, TEM studies (Fig. 1, Supporting Information Table
S1) confirmed Lauterborn’s hypotheses.(9) The MC is clearly
visible by light microscopy in large pennate diatoms (several
species of the genera Surirella, Pinnularia and Nitzschia) and
in a few centric diatoms where it is said to be ‘less obvious’ to
observe (Supporting Information Table S1). It is always
closely associated with the interphase nucleus and with the
Golgi apparatus. In the centric diatom Lythodesmium the Bb is
involved in flagellum formation during sexual reproduction and
forms de novo during meiosis. For these reasons this
structure has been termed ‘paracentrosome’.(27) Centrics
may therefore produce Bbs de novo during flagellar formation
during sexual reproduction as in brown algae,(26) whereas in
pennates the centriolar structure has been totally lost.(28)
Besides its peculiar acentriolar ultrastructure, the diatom MC
also displays unusual behaviour during the cell cycle (Fig. 2
and 3). Animal centrosomes, yeast SPB and slime mould
MTOC are all present as single entities in the cell and they
duplicate before mitosis to constitute the spindle poles(29)
(Fig. 2). By contrast, the diatom MC (Fig. 2A) does not duplicate
before mitosis; rather a second structure, the polar complex
(PC), appears in premitotic cells (at early prophase in several
pennate diatoms) close to the MC. The PC is a cubic laminar
structure. At prophase, the PC splits into two parallel structures
known as the polar plates, which constitute the poles of the
mitotic spindle.(30) Both the MC and PC function as MT
nucleation centres but they act at different periods of the cell
cycle. Once the PC appears, the MC disintegrates and vice
versa. Lauterborn(9) already observed these as two indepen-
dent structures. The PC has been identified in several pennates
but the behaviour of MC/PC shows some variation among
diatoms (Supporting Information Table S1). It is thought that the
PC is produced by the MC(9,10) or forms very close to it,(31) but
almost nothing is known about their origin or whether the PC
derives from the MC. Ultrastructurally, the PC clearly differs from
the two centriolar poles of the mitotic spindle in animals
(Fig. 2B), while it is more similar to acentriolar yeast SBP
(Fig. 2C–D) and D. discoideum MTOC(22,32) (Fig. 2E). Further-
more, the duplication and splitting process of the PC recalls the
duplication and peeling off of the D. discoideum MTOC
(Fig. 2E). Again, the appearance of diatom MTOCs during
mitosis is very different from what is observed in higher plants,
which are characterised by broad spindle poles (not shown).
To conclude, the diatom MC is an MTOC model known for
many years with unusual and uncharacterised features that
merits being explored at the molecular level. The interconver-
sion of two spatially and temporally distinct structures during
the cell cycle is unique to the diatom MTOC, and the question
of whether the PC derives from MC components has not been
addressed at the molecular level. Why are there two spatially
distinct structures in diatoms, instead of a single entity that
duplicates as in the centrosome or SPB? An additional
875

Figure 1. Diatom MTOC ultrastructure in an interphase cell compared to other MTOC models. A: Electron micrograph of animal centrosome
(left) and schematic representation of the pair of centrioles surrounded by a pericentriolar matrix (PCM) (right).(29) Reprinted with permission from
� Pearson Education, Inc., publishing as Benjamin Cummings (Campbell N. A. and Reece J. B., Biology, � 2002). B–E: Electron micrographs of
acentriolar MTOC of yeast (SPB) (B and C), slime mould (D) and pennate diatom (E). The SPB in S. cerevisiae (B) is a disc-like structure
embedded in the NE connected to cytoplasmic MTs (cMTs) and nuclear MTs (nMTs). The half bridge (HB) is the site of the new SPB assembly.(22)
Reprinted with permission from the Annual Review of Cell and Developmental Biology, Volume 20, � 2004 by Annual Reviews, www.
annualreviews.org. The SPB of S. pombe (C) is a single dense layer beside the NE (arrow).(23) Reproduced with permission from Ding
et al., 1997, Mol Biol Cell, 8 1461–1479, � The Biochemical Society. D. discoideum MTOC (D) is a matchbox-shaped three-layered structure.(110)
Reprinted with permission from the Molecular Biology of the Cell, Volume 10, �1999 by the American Society of Cell Biology. Pennate diatom
(Surirella ovalis) MC (E) is an extranuclear structure, appearing as a well-defined dark granule from which MTs radiate around the interphase
nucleus (n) and in all directions.(30) Reprinted with permission from Rockefeller University Press, � Tippit and Pickett-Heaps, 1977. Originally
published in The Journal of Cell Biology. 73: 705–727.
Review article A. De Martino, A. Amato and C. Bowler
question emerges from old electron microscopy (EM) studies:
does the MC in centric and pennate diatoms differ in
molecular structure?
Now that we benefit from two completed diatom gen-
omes,(3,5) molecular tools for reverse genetics,(17–19) and
time-lapse imaging of single cells overexpressing fluorescent
fusion proteins (paper in preparation), these questions can be
further re-examined. The centric (Thalassiosira pseudonana)
and pennate diatom (Phaeodactylum tricornutum) sequences
permit comparative studies to address the differences
876
between the two diatom groups. Moreover, the existence of
distinct MTOC structures acting at different moments of the
cell cycle offers several advantages for understanding the
dual function of MTOC, i.e. orchestration of cytoplasmic MTs
in interphase and mitotic spindle organisation. Furthermore,
since MTOCs are integrative structures for several regulatory
pathways during the cell cycle, e.g. cyclin/cyclin-dependent
kinases (CDKs) and other regulatory proteins control
centrosome and SPB duplication in animal cells, it will be
interesting to explore the function of these components in
BioEssays 31:874–884, � 2009 Wiley Periodicals, Inc.

Figure 2. The MC/PC cycle in diatoms compared to MTOC duplication in animals, yeasts and slime moulds. A: Electron micrographs (upper
panels; reprinted with permission from Rockefeller University Press, � Tippit and Pickett-Heaps, 1977; originally published in The Journal of Cell
Biology 73: 705–727) and schematic representation (lower panels) of the MC/PC cycle and spindle development in the diatom Surirella. At
preprophase, the PC appears close to, but separate from, the interphase MC and elongates. The central spindle arises before prophase from the
PC that splits into two and gives rise to the ‘polar plates’ (PP). A band of densely packed, parallel MTs, the central spindle, develops between the
two PP. Condensed chromosomes become visible, distributed as a ring around the spindle at metaphase. Red circles and rectangles, MC and PC,
respectively; green lines, MTs; light green full lines and dotted lines, NE and partially broken down NE, respectively; blue circles, chromosomes.
B: In animal cells, centrioles disengage at early G1 phase and duplication of each centriole occurs at S phase. Daughter centrioles reach
maturation at G2. Following NEBD at prometaphase, the two pairs of centrioles become the focused spindle poles at metaphase. Red cylinders,
centrioles; in grey, pericentriolar matrix (PCM); green lines, MTs; light green full lines and dotted lines, NE and broken down NE, respectively; blue
circles, chromosomes. C: In S. cerevisiae at S phase, the half bridge (HB) elongates and satellite material is deposited into a nascent SPB next to
the mother SPB. At S/G2 phase, the daughter SPB reaches maturation. At metaphase, the SPBs move to opposite sides of the NE, becoming the
spindle poles. In red, SPB and HB; green lines, MTs; light green full lines, NE; Blue circles, chromosomes.D: InS. pombe, a bridge-like structure is
formed in G2 phase and a new SPB is formed at the distal end of the bridge at preprophase. The NE invaginates and forms a fenestra into which
the two SPBs settle. At metaphase, the SPBs move to opposite sides of the NE, becoming the spindle poles. In red, SPB and bridge-like structure;
green lines, MTs; light green full lines and dotted lines, NE and fenestra, respectively; blue circles reorganisation, chromosomes. E: In
D. discoideum at prophase, the layered structures increase in size along their axes. At prometaphase, the corona and cytoplasmic MTs disperse,
then the middle layer of the three-layered core structure disappears and the two outer layers peel apart. The two outer layers become the spindle
poles. In red, MTOC; green lines, MTs; light green full lines, NE; blue circles, chromosomes; black dotted lines, corona. For each organism only
the most important steps during cell cycle are shown.
A. De Martino, A. Amato and C. Bowler Review article
BioEssays 31:874–884, � 2009 Wiley Periodicals, Inc. 877

Figure 3. Schematic representation of spindle formation, elongation and cytokinesis in diatoms compared to different model organisms.A: The
appearance of the PC (red rectangle) takes place at the beginning of preprophase close to the MC (red circle) outside the nucleus. The MC
disappears as the spindle elongates. By the end of prophase the spindle enters the nucleus but no complete NEBD occurs. At metaphase
chromosomes assemble around the spindle in a characteristic donut shape. At the end of anaphase B the cleavage furrow splits the fully
elongated spindle in two. B: In animals, centrioles (red rectangle) divide and the mitotic spindle elongates. At prometaphase NEBD occurs. At
metaphase chromosomes align between the poles of the spindle, then segregate. At telophase the cleavage furrow splits the spindle and two
daughter nuclei are reformed.C, D: In fission (C) and budding yeast (D), the SPB (red rectangle) divides and the spindle is generated between the
SPB pair sitting across the NE. No NEBD occurs. Cytokinesis takes place by fission in (C) and by budding in (D). E: In plants, MTs are nucleated
from numerous foci on the cortex in interphase. At preprophase foci condense at the centre to form the preprophase band. NEBD occurs.
Chromosomes align at metaphase. At telophase an outgrowing cell plate develops between the segregated chromosomes and splits the spindle.
In red, MTOCs; green lines, MTs; blue circles, chromosomes. Light green full, interrupted and dotted lines: NE, partially broken down NE and
broken down NE, respectively. Thin black lines, valves; thick black lines, cell wall, in grey, plasma membrane; black ring, contractile ring; violet
hemi-circles, polar cap.
Review article A. De Martino, A. Amato and C. Bowler
diatoms. We have recently identified several unique cyclin/
CDKs and other cell cycle regulators in P. tricornutum, and
diatoms contain a large number of diatom-specific cyclins.(5)
Hinting at functions of some molecularcomponents of the diatom MTOC
Although high-quality EM has been carried out on MC and PC
in diatoms, almost nothing is known about their composition at
the molecular level. These two structures have the same
function, namely MT nucleation, but they do not look the same
and they act at different times of the cell cycle, suggesting that
878
they could have different compositions and may be regulated
differently. From in silico analysis of the genomes of the
centric T. pseudonana(3) and the pennate P. tricornutum(5)
(http://genome.jgi-psf.org/Thaps3/Thaps3.home.html; http://
genome.jgi-psf.org/Phatr2/Phatr2.home.html), putative com-
ponents involved in the cell cycle regulation of MTOC and
the mitotic spindle can be identified (Supporting Information
Table S2).
Both the centrosome and SPB contain a g-tubulin (g-tub)
complex that serves as a template for polymerisation of
MTs(33) and establishes their polarity, with stable minus ends
facing the centrosome/SPB and plus ends facing outward.(34)
Although the g-tub complex and the nucleation mechanism
BioEssays 31:874–884, � 2009 Wiley Periodicals, Inc.
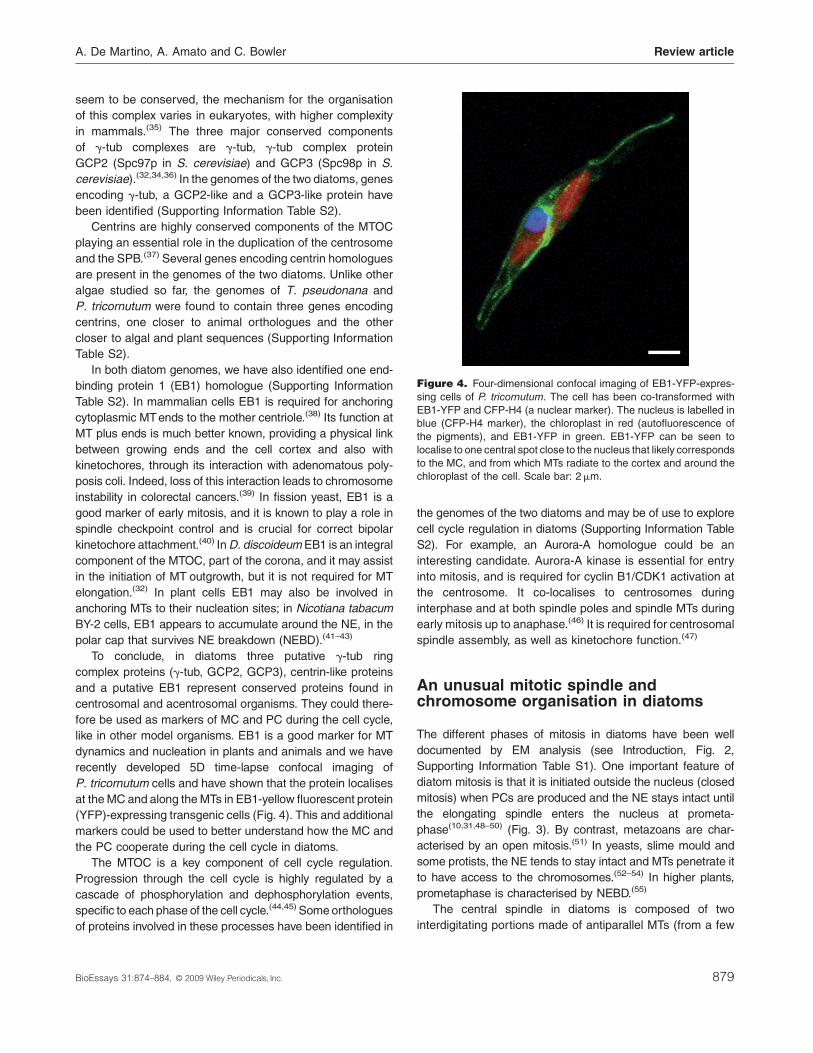
Figure 4. Four-dimensional confocal imaging of EB1-YFP-expres-
sing cells of P. tricornutum. The cell has been co-transformed with
EB1-YFP and CFP-H4 (a nuclear marker). The nucleus is labelled in
blue (CFP-H4 marker), the chloroplast in red (autofluorescence of
the pigments), and EB1-YFP in green. EB1-YFP can be seen to
localise to one central spot close to the nucleus that likely corresponds
to the MC, and from which MTs radiate to the cortex and around the
chloroplast of the cell. Scale bar: 2mm.
A. De Martino, A. Amato and C. Bowler Review article
seem to be conserved, the mechanism for the organisation
of this complex varies in eukaryotes, with higher complexity
in mammals.(35) The three major conserved components
of g-tub complexes are g-tub, g-tub complex protein
GCP2 (Spc97p in S. cerevisiae) and GCP3 (Spc98p in S.
cerevisiae).(32,34,36) In the genomes of the two diatoms, genes
encoding g-tub, a GCP2-like and a GCP3-like protein have
been identified (Supporting Information Table S2).
Centrins are highly conserved components of the MTOC
playing an essential role in the duplication of the centrosome
and the SPB.(37) Several genes encoding centrin homologues
are present in the genomes of the two diatoms. Unlike other
algae studied so far, the genomes of T. pseudonana and
P. tricornutum were found to contain three genes encoding
centrins, one closer to animal orthologues and the other
closer to algal and plant sequences (Supporting Information
Table S2).
In both diatom genomes, we have also identified one end-
binding protein 1 (EB1) homologue (Supporting Information
Table S2). In mammalian cells EB1 is required for anchoring
cytoplasmic MTends to the mother centriole.(38) Its function at
MT plus ends is much better known, providing a physical link
between growing ends and the cell cortex and also with
kinetochores, through its interaction with adenomatous poly-
posis coli. Indeed, loss of this interaction leads to chromosome
instability in colorectal cancers.(39) In fission yeast, EB1 is a
good marker of early mitosis, and it is known to play a role in
spindle checkpoint control and is crucial for correct bipolar
kinetochore attachment.(40) InD. discoideum EB1 is an integral
component of the MTOC, part of the corona, and it may assist
in the initiation of MT outgrowth, but it is not required for MT
elongation.(32) In plant cells EB1 may also be involved in
anchoring MTs to their nucleation sites; in Nicotiana tabacum
BY-2 cells, EB1 appears to accumulate around the NE, in the
polar cap that survives NE breakdown (NEBD).(41–43)
To conclude, in diatoms three putative g-tub ring
complex proteins (g-tub, GCP2, GCP3), centrin-like proteins
and a putative EB1 represent conserved proteins found in
centrosomal and acentrosomal organisms. They could there-
fore be used as markers of MC and PC during the cell cycle,
like in other model organisms. EB1 is a good marker for MT
dynamics and nucleation in plants and animals and we have
recently developed 5D time-lapse confocal imaging of
P. tricornutum cells and have shown that the protein localises
at the MC and along the MTs in EB1-yellow fluorescent protein
(YFP)-expressing transgenic cells (Fig. 4). This and additional
markers could be used to better understand how the MC and
the PC cooperate during the cell cycle in diatoms.
The MTOC is a key component of cell cycle regulation.
Progression through the cell cycle is highly regulated by a
cascade of phosphorylation and dephosphorylation events,
specific to each phase of the cell cycle.(44,45) Some orthologues
of proteins involved in these processes have been identified in
BioEssays 31:874–884, � 2009 Wiley Periodicals, Inc.
the genomes of the two diatoms and may be of use to explore
cell cycle regulation in diatoms (Supporting Information Table
S2). For example, an Aurora-A homologue could be an
interesting candidate. Aurora-A kinase is essential for entry
into mitosis, and is required for cyclin B1/CDK1 activation at
the centrosome. It co-localises to centrosomes during
interphase and at both spindle poles and spindle MTs during
early mitosis up to anaphase.(46) It is required for centrosomal
spindle assembly, as well as kinetochore function.(47)
An unusual mitotic spindle andchromosome organisation in diatoms
The different phases of mitosis in diatoms have been well
documented by EM analysis (see Introduction, Fig. 2,
Supporting Information Table S1). One important feature of
diatom mitosis is that it is initiated outside the nucleus (closed
mitosis) when PCs are produced and the NE stays intact until
the elongating spindle enters the nucleus at prometa-
phase(10,31,48–50) (Fig. 3). By contrast, metazoans are char-
acterised by an open mitosis.(51) In yeasts, slime mould and
some protists, the NE tends to stay intact and MTs penetrate it
to have access to the chromosomes.(52–54) In higher plants,
prometaphase is characterised by NEBD.(55)
The central spindle in diatoms is composed of two
interdigitating portions made of antiparallel MTs (from a few
879

Review article A. De Martino, A. Amato and C. Bowler
units to hundreds), like in other organisms.(55,56) It is clearly
visible in EM, with a very well structured overlapping region
showing a unique paracrystalline arrangement, yielding the
conclusion that some bridge proteins might maintain the
structure(57–59) (Fig. 3A), as proposed for plant mitotic
spindles.(55) A set of motor proteins could act at the
overlapping region to pull apart the interdigitating MTs
during mitosis in diatoms (see below and(59)). At metaphase,
the chromosomes attached to the spindle have a unique
‘ring arrangement around the spindle equator’ that gives
the chromatin a characteristic ‘donut shape’(30,31) (Fig. 3A).
At this stage a dense matrix is visible in the spindles
of different diatoms (Fig. 3, Supporting Information
Table S1(30,48,57,60,61)). The so-called ‘collar material’ lies at
the chromosome-MT attachment site (see below).
Spindle elongation has been thoroughly studied in the
centric diatom S. turris,(11–13,50,59,62–64) and in the pennates
C. fusiformis(14–16,65,66) and Hantzschia amphioxys(67) using
different approaches. The spindle has been proposed to
elongate at anaphase B by different processes.(68) The
following evidence indicates that the main actor in spindle
elongation is the overlapping region: (i) biotinylated tubulin
incorporation studies showed that MT polymerisation occurs
mainly at the level of the overlapping region(11,64); (ii) a
reduction of the overlap was observed during mitosis (see
Fig. 3 in(59)); thus the overlapping region seems not to be a
fixed anchoring point for the two half-spindles but to play a
dynamic role as well as a structural one; (iii) in vitro studies on
isolated spindles showed that spindles elongate anyway even
though no cytoplasm surrounds the poles, and even after
chromatin digestion.(62,69) This means that the forces for
elongation do not reside in the PC or do not act on them but
are intrinsic to the spindle itself.
While the overlapping region is certainly a site for MT
polymerisation, it has been shown in vitro that isolated spindles
elongate even in the absence of tubulin (S. turris(11,50)) or in the
presence of MT polymerisation drugs,(62,70) and that anaphase
B movement is not affected in the presence of mitotic
inhibitors.(71) MT polymerisation can therefore be a permissive
feature but not a substantial one for spindle elongation itself.
More important is likely to be MT sliding as demonstrated
in vitro by Cande and coworkers.(12,13,68)
By coupling TEM micrographs with micro-cinematography,
it was shown that kinetochores in the pennate diatoms
Hantzschia and Nitzschia have a tri-plate structure, and
evidence based on EM observations was given to show that
they do not nucleate any MTs,(49) in contrast to what some
claimed at that time.(72) In diatoms the different MT types are
spatially separated, and the MTs involved in structural
maintenance of the central spindle do not attach to chromo-
somes and vice versa. After spindle entrance into the nucleus,
the chromosomes start oscillating back and forth to one or the
other pole possibly in order to orient their kinetochores to each
880
pole(49) (chromosome congression(73)); then they segregate. A
unique feature observed in Surirella(30) and Pinnularia(61) is an
ill-defined matrix, called the collar, permeating the central
spindle mid-way between the poles and chromosomes. In
these species no conventional kinetochore fibres were
observed (Supporting Information Table S1).
To conclude, diatoms display several advantages for
studying the mitotic spindle: (i) some species are very large
and mitosis is rapid, enabling mitotic activity to be seen in living
cells with great clarity; (ii) they create a conspicuous central
spindle to which chromosomes attach; thus, unlike conven-
tional spindles, it is conceptually easy to see what structure
chromosomes attach to and move over during mitosis, and (iii)
their spindles are well organised, allowing their structure to be
analysed and correlated with their behaviour.
Glimpsing molecular features andregulation of diatom spindle andkinetochores
Several homologues of animal mitotic checkpoint and spindle-
associated regulatory proteins can be identified in the two
diatom genomes (Supporting Information Table S2). One
homologue of the kinetochore protein ZW10, which binds
the centromere and the kinetochore and is required for
chromosome segregation and for correct metaphase check-
point control, is present in P. tricornutum and T. pseudonana.
We have also identified several chromosome-associated
kinesins (chromokinesin) and several extra-spindle pole-like
proteins important for correct chromosome segregation
(Supporting Information Table S2). One is homologous to
the MAP65_ASE1 protein family, a putative MT-associated
protein that is essential for anaphase spindle elongation.
In animal cells there is a direct link between cell cycle
regulatory proteins and the assembly of the mitotic spindle,
which requires a protein complex that cross-links MTs
of opposite polarity.(74) A KRP called diatom spindle
kinesin 1(15,16) has been identified and characterised in
C. fusiformis as a motor protein involved in MT sliding.
Orthologues are present in both P. tricornutum and
T. pseudonana and could be good candidates to study the
spindle elongation process. However, in most of the previous
reports the presence of dynein was claimed instead of
KRP,(62) based on phosphorylation studies of vanadate-
treated in vitro isolated spindles.(12) Several dyneins are
encoded in the genomes of both diatoms.(3,4)
Kinetochores are multi-protein complexes that assemble
on centromeres and have three main functions: binding of
MTs, coordinating chromosome movements, and activating
spindle checkpoints. Centromeric regions vary in different
eukaryotes, from the 125-bp point centromeres in the budding
yeast(75) and the approximately 5-Mb human ones(76) to the
BioEssays 31:874–884, � 2009 Wiley Periodicals, Inc.

A. De Martino, A. Amato and C. Bowler Review article
holocentromeres in the nematode Caenorhabditis ele-
gans.(77) Albeit different in size and arrangement, centromeric
organisation is commonly associated with rapidly evolving
satellite sequences.(78) But the spontaneous birth of new
centromeres (neocentromeres) supports the idea that
centromeres are epigenetically specified and maintained,
generating a modification in chromatin structure(79) that is
inherited via meiotic drive.(80)
Despite low conservation of centromeric regions, centro-
mere or kinetochore proteins are highly conserved (at least
between metazoans and yeasts(81)). Centromeres likely carry
out the same function throughout eukaryotes, as do
kinetochores.(82) Among kinetochore proteins there is the
centromere-specific histone H3 variant, further referred to as
CENH3, according to Dawe and Henikoff.(83) CENH3
produces modified nucleosomes that line up on the chromo-
somes facing the pole.(84) This is where such nucleosomes
interact with the foundation kinetochore proteins(78) that in
turn recruit proteins involved in MTattachment and checkpoint
controls.(78) Dynamic binding of the MT to the kinetochores is
achieved through the motor proteins dynein and dynactin as
well as the KRP MCAK and CENP-E in vertebrates(85) and
several protein complexes such as the NDC complex (Ndc80/
Hec1-Nuf2-Spc24/25(86–88)), almost all of which are present in
diatoms (Supporting Information Table S2).
Kinetochores also play a substantial role in regulating the
progression from metaphase to anaphase. If the chromo-
somes are not properly aligned before segregation, aneuploid
daughter cells could be produced.(73) The spindle checkpoint
function is accomplished by a complicated orchestration of
different proteins(89) that sense free kinetochores and
transduce them into signals that inhibit the anaphase-
promoting complex/cyclosome.(89) A centromere-specific
histone H3 variant is present in P. tricornutum (Supporting
Information Table S2) but not in T. pseudonana, possibly
corroborating the hypothesis of a low level of similarity
of centromeric proteins even among diatoms (as hypothe-
sised for other groups(90)), but contrasting with what is
found in fungi, where all CENH3 proteins are orthologous.(91)
Notwithstanding the centrality of kinetochore function and
the molecular aspects underlying centromere organisation,
kinetochore construction, kinetochore-MT attachment and
spindle checkpoint activation are unclear in diatoms and to
our knowledge no major advances have been made since the
1980s.
Cytokinesis: a combination of plantand animal features
Diatom cytokinesis is a mixture of animal (cleavage furrow)
and plant (synthesis of new cell wall) features (Fig. 3). As has
been elegantly shown by Pickett-Heaps and Pickett-Heaps by
BioEssays 31:874–884, � 2009 Wiley Periodicals, Inc.
micro-cinematography (‘Diatoms: Life in glass houses’; http://
www.cytographics.com/pg-dia.htm), ‘cytokinesis occurs by
furrowing’(92) and, as in animal cells,(43) the two newly formed
daughter cells are split. ‘Cells undergoing cytokinesis . . . show
an [actin] band encircling the cell at the cleavage furrow’.(93)
This finding supports the hypothesis that an actin contractile
ring-like structure is involved in diatom cytokinesis like in
animals and yeasts.(94) An actin band is also involved in
diatom valve morphogenesis(93,95) surrounding the growing
edges of the silica deposition vesicle, a specialised organelle.
The silica deposition vesicle is generally positioned in the
middle of the furrow and grows outwards, and is often
adjacent to the newly ‘reforming’ MC (see Supporting
Information Table S1) and the Golgi apparatus.(93,96)
Recent studies of conserved protein families have
revealed a common basis for the mechanism of cytokinesis
in plants and animals.(43) A structure deriving from the spindle
midzone sits in the middle of the contractile ring in animals
(midbody) promoting the trafficking of vesicles towards the
ingressing furrow. In plants it is the phragmoplast, an MT/actin
structure that has the function of transporting Golgi-derived
vesicles to the site of cell plate assembly.(97) By contrast,
S. pombe divides by septation. At the end of mitosis, an
equatorial MTOC (distinct to SPB), called the EMTOC, forms
mid-way through anaphase, recruiting g-tub to the equatorial
F-actin ring before it constricts to cleave the cell into two.(98)
It seems that in diatoms actin plays a double role in cell
division: first during furrowing (as in animals), and later
on during cell wall deposition (as in plants), again highlighting
the chimeric nature of diatom regulatory processes and
cytological features.(3,5)
In animal cells the position of the mitotic spindle determines
the position of the furrow at anaphase,(99) while division zone
determination happens far before anaphase in plants. In plants
the preprophase band, a transitory MT structure, marks the
future localisation of the cell plate.(97) Among the different
models for cytokinesis, the budding and fission yeasts
represent rather unique examples. S. cerevisiae divides by
budding; i.e. a new cell is produced by the constriction of a
contractile ring. In fission yeast the contemporary deposition of
cell wall material and the action of a contractile ring-like
structure separate the two daughter cells by fission. Yeasts
also diverge from other organisms in that determination of the
division plane occurs far before mitosis, at the G1/S boundary
in S. cerevisiae and in G2 in S. pombe.(99) In the centric diatom
S. turris, the location of the plane of division is cortically
determined before the mitotic spindle is formed and it is totally
independent of the position of the nucleus. This evidence has
been proven by short-term spindle displacement experi-
ments.(92) Although cytokinesis in brown algae has been
relatively well studied (centripetal with the development of a
plasma membrane furrow),(26) we have not compared it with
diatoms because, in spite of being phylogenetically related to
881

Review article A. De Martino, A. Amato and C. Bowler
them, they show divergent features such as a centriolar mitotic
apparatus similar to animals,(26) have a different cell wall, and
are multicellular.
Conclusions
At the end of the 19th century, diatoms represented
one of the best models for scientists interested in
cell division. Lauterborn,(9) and subsequently Pickett-
Heaps,(10,20,28,30,31,48,49,57,58,60,61,71,93,95,96,100–104)
Cande(11–16,50,55,59,62–70,92,105) and collaborators have been
the pioneers of cell division studies in diatoms. Their beautiful
studies represent high-quality optical and EM descriptions of
nuclear division, spindle development and chromosome
segregation. They revealed intriguing features unique to
diatoms, such as (i) two independent organiser centres, the
MC and the PC, each with a role during interphase and
mitosis, respectively; (ii) a special ring distribution of the
chromosomes around the mitotic spindle at metaphase and a
paracrystalline organisation of the mitotic MT arrays; (iii) the
apparent absence of conventional kinetochores and centro-
somes, and (iv) cytokinesis with characteristics specific for
animals (cleavage furrow), plants (cell wall neosynthesis) and
diatoms (silica valve neosynthesis).
These observations invoke many exciting questions that
will need to be clarified by molecular approaches, such as the
precise and specific roles of MC and PC, their specific
structures and interconversion during the cell cycle, and the
mode of chromosome attachment and segregation on the
mitotic spindle. Although it is disappointing that these
pioneering works have been ‘frozen’ for the past 20 years
because of the lack of molecular technologies, molecular
tools and genomic data are now in place for cell division in
diatoms to be re-examined. In addition, diatoms offer several
advantages: (i) they are single cells easily observed in the
microscope and with easily identifiable mitotic structures; (ii)
the cells are easy to grow in the laboratory, and some species
are very large and mitosis is rapid; (iii) molecular tools for
reverse genetics have been developed; (iv) two completed
genome sequences are available, one from a pennate diatom
and one from a centric diatom, and (v) time-lapse imaging of
single cells expressing fluorescent fusion proteins has been
developed in P. tricornutum.
Several authors have recently reviewed and underlined the
diversity of structures involved in MT and mitotic spindle
organisation in eukaryotes, focusing on the animal and green
lineages.(43) Because of the chimeric and specific features of
cell division in diatoms, it will be interesting to add them to
such comparative analyses. Information about the origin and
evolution of the cellular structures involved in cell division in
eukaryotes may also aid our understanding of proliferative
diseases such as cancer.
882
Moreover some diatom species (mainly belonging to the
pennate genus Pseudo-nitzschia) can produce the neurotoxin
domoic acid(106) that accumulates in shellfish and can pose a
risk for human health.(107) These and other algal bloom
phenomena are examples of diatom cell proliferation in
response to specific environmental cues, and so it is of
interest to understand how external stimuli can provoke such
a dramatic activation of cell division. It is all the more urgent to
address such questions given current interest in large-scale
iron fertilisation of the southern oceans for carbon seques-
tration.(108) In these scenarios, iron enrichment is intended to
provoke massive diatom blooms which will remove CO2 from
the atmosphere through photosynthesis and export it to the
ocean interior.
Finally, diatoms are receiving attention as a source of algal
biofuels because of their high lipid content.(109) Such
approaches will require improvements of cell growth and
division in commercial settings and so an improved under-
standing of cell division will facilitate the development of
targeted strategies to optimise large-scale culture. To
summarise, we believe the new tools now available to explore
diatom biology can lead to a renaissance of studies of cell
division, thus vindicating the previous studies from the last two
centuries.
References
1. Litchman, E., Klausmeier, C. A., Miller, J. R., Schofield, O. M. and
Falkowski, P. G., Multi-nutrient, multi-group model of present and future
oceanic phytoplankton communities. Biogeosciences 2006. 3: 585–606.
2. Delwiche, C. F., Tracing the thread of plastid diversity through the
tapestry of life. Am Nat 1999. 154: S164–S177.
3. Armbrust, E. V., Berges, J. A., Bowler, C., Green, B. R., Martinez, D.,
et al. The genome of the diatom Thalassiosira pseudonana: ecology,
evolution, and metabolism. Science 2004. 306: 79–86.
4. Montsant, A., Allen, A. E., Coesel, S., De Martino, A., Falciatore, A.,
et al. Identification and comparative genomic analysis of signaling and
regulatory components in the diatom Thalassiosira pseudonana.
J Phycol 2007. 43: 585–604.
5. Bowler, C., Allen, A. E., Badger, J. H., Grimwood, J., Jabbari, K., et al.
The Phaeodactylum genome reveals the dynamic evolutionary history of
diatom genomes. Nature 2008. 456: 239–244.
6. Hoagland, K. D., Rosowski, J. R., Gretz, M. R. and Roemer, S. C.,
Diatom extracellular polymeric substances – function, fine-structure,
chemistry, and physiology. J Phycol 1993. 29: 537–566.
7. Round, F. E., Crawford, R. M. and Mann, D. G., The diatoms. Biology
abd morphology of the genera. Cambridge, UK, Cambridge University
Press, 1990. p 747.
8. Zurzolo, C. and Bowler, C., Exploring bioinorganic pattern formation in
diatoms. A story of polarized trafficking. Plant Physiol 2001. 127: 1339–
1345.
9. Lauterborn, R., Untersuchugen uber Bau, Kernteilung und Bewegung
der Diatomeen. Leipzig, Verlag von Willelm Engelmann, 1896.
10. Pickett-Heaps, J. D., Schmid, A.-M. M. and Tippit, D. H., Cell division in
diatoms. Protoplasma 1984. 120: 132–154.
11. Masuda, H. and Cande, W. Z., The role of tubulin polymerization during
spindle elongation in vitro. Cell 1987. 49: 193–202.
12. Wordeman, L., Masuda, H. and Cande, W. Z., Distribution of a thiopho-
sphorylated spindle midzone antigen during spindle reactivation in vitro.
J Cell Sci 1989. 93: 279–285.
BioEssays 31:874–884, � 2009 Wiley Periodicals, Inc.

A. De Martino, A. Amato and C. Bowler Review article
13. Wordeman, L., Davis, F. M., Rao, P. N. andCande,W. Z.,Distribution of
phosphorilated spindle-associated proteins in the diatom Stephanopyxis
turris. Cell Motil Cytoskeleton 1989. 12: 33–41.
14. Wein, H., Brady, B. and Cande, W. Z., Isolating the plant mitotic
apparatus: A procedure for isolating spindles from the diatom Cylin-
drotheca fusiformis. Method Cell Biol 1995. 50: 177–187.
15. Wein, H., Foss, M., Brady, B. and Cande, W. Z., DSK1, a novel kinesin-
related protein from the diatom Cylindrotheca fusiformis that is involved
in anaphase spindle elongation. J Cell Biol 1996. 133: 595–604.
16. Wein, H., Bass, H.W. and Cande,W. Z., DSK1, a kinesin-related protein
involved in anaphase spindle elongation, is a component of a mitotic
spindle matrix. Cell Motil Cytoskeleton 1998. 41: 214–224.
17. Falciatore, A., Casotti, R., Leblanc, C., Abrescia, C. and Bowler, C.,
Transformation of nonselectable reporter genes in marine diatoms. Mar
Biotechnol 1999. 1: 239–251.
18. Falciatore, A. and Bowler, C., Revealing the molecular secrets of
marine diatoms. Annu Rev Plant Biol 2002. 53: 109–130.
19. Siaut, M., Heijde, M., Mangogna, M., Montsant, A., Coesel, S., et al.
Molecular toolbox for studying diatom biology in Phaeodactylum tricor-
nutum. Gene 2007. 406: 23–35.
20. Pickett-Heaps, J. D., The evolution of the mitotic apparatus, an attempt
at comparative ultrastructural cytology in dividing cell plants. Cytobios
1969. 1: 257–280.
21. Bettencourt-Dias, M. and Glover, D. M., Centrosome biogenesis and
function: centrosomics brings new understanding. Nat Rev Mol Cell Biol
2007. 8: 451–463.
22. Jaspersen, S. L. and Winey, M., The budding yeast spindle pole body:
Structure, duplication, and function. Annu Rev Cell Dev Biol 2004. 20:
1–28.
23. McIntosh, J. R. and O’Toole, E. T., Life cycles of yeast spindle pole
bodies: Getting microtubules into a closed nucleus. Biol Cell 1999. 91:
305–312.
24. Beisson, J. and Wright, M., Basal body/centriole assembly and con-
tinuity. Curr Opin Cell Biol 2003. 15: 96–104.
25. Wasteneys, G. O., Microtubule organization in the green kingdom:
chaos or self-order? J Cell Sci 2002. 115: 1345–1354.
26. Katsaros, C., Karyophyllis, D. and Galatis, B., Cytoskeleton and
Morphogenesis in Brown Algae. Ann Bot 2006. 97: 679–693.
27. Manton, I., Kowallik, K. and von Stosch, V. A., Observations on the fine
structure and development of the spindle at mitosis and meiosis in a
marine centric diatom (Lithodesmium undulatum). IV. Second meiotic
division and conclusion. J Cell Sci 1970. 7: 407–445.
28. Pickett-Heaps, J. D., Postmitotic cellular reorganization in the diatom
Cymatopleura solea - the role of microtubules and the microtubule
center. Cell Motil Cytoskeleton 1991. 18: 279–292.
29. Azimzadeh, J. and Bornens, M., Structure and duplication of the
centrosome. J Cell Sci 2007. 120: 2139–2142.
30. Tippit, D. H. and Pickett-Heaps, J. D., Mitosis in pennate diatom
Surirella ovalis. J Cell Biol 1977. 73: 705–727.
31. Pickett-Heaps, J. D., McDonald, K. L. and Tippit, D. H., Cell division in
pennate diatom Diatoma vulgare. Protoplasma 1975. 86: 205–242.
32. Graf, R., Daunderer, C. and Schulz, I., Molecular and functional ana-
lysis of the Dictyostelium centrosome. Int Rev Cytol 2004. 241: 155–202.
33. Vogel, J. M., Stearns, T., Rieder, C. L. and Palazzo, R. E.,Centrosomes
isolated from Spisula solidissima oocytes contain rings and an unusual
stoichiometric ratio of alpha/beta tubulin. J Cell Biol 1997. 137: 193–202.
34. Moritz, M., Braunfeld, M. B., Sedat, J. W., Alberts, B. and Agard, D. A.,
Microtubule nucleation by gamma-tubulin-containing rings in the centro-
some. Nature 1995. 378: 638–640.
35. Murata, T., Tanahashi, T., Nishiyama, T., Yamaguchi, K. and Hasebe,
M., How do plants organize microtubules without a centrosome? J Integr
Plant Biol 2007. 49: 1154–1163.
36. Seltzer, V., Janski, N., Canaday, J., Herzog, E., Erhardt, M., et al.
Arabidopsis GCP2 and GCP3 are part of a soluble gamma-tubulin
complex and have nuclear envelope targeting domains. Plant J 2007.
52: 322–331.
37. Bornens, M. and Azimzadeh, J., Origin and evolution of the centro-
some. In: Jekely, G. editor. Eukaryotic membranes and cytoskeleton:
origins and evolution. Berlin, Springer-Verlag Berlin, 2007. pp. 119–129.
BioEssays 31:874–884, � 2009 Wiley Periodicals, Inc.
38. Louie, R. K., Bahmanyar, S., Siemers, K. A., Votin, V., Chang, P., et al.
Adenomatous polyposis coli and EB1 localize in close proximity of the
mother centriole and EB1 is a functional component of centrosomes.
J Cell Sci 2004. 117: 1117–1128.
39. Fodde, R., Smits, R. and Clevers, H., APC, signal transduction and
genetic instability in colorectal cancer. Nat Rev Cancer 2001. 1: 55–67.
40. Asakawa, K. and Toda, T., Cooperation of EB1-Mal3 and the Bub1
spindle checkpoint. Cell Cycle 2006. 5: 27–30.
41. Chan, J., Calder, G. M., Doonan, J. H. and Lloyd, C. W., EB1 reveals
mobile microtubule nucleation sites in Arabidopsis. Nat Cell Biol 2003. 5:
967–971.
42. Piehl, M., Tulu, U. S., Wadsworth, P. and Cassimeris, L., Centrosome
maturation: Measurement of microtubule nucleation throughout the cell
cycle by using GFP-tagged EB1. Proc Natl Acad Sci U S A 2004. 101:
1584–1588.
43. Lloyd, C. and Chan, J., Not so divided: the common basis of plant and
animal cell division. Nat Rev Mol Cell Biol 2006. 7: 147–152.
44. Kramer, A., Lukas, J. and Bartek, J., Checking out the centrosome. Cell
Cycle 2004. 3: 1390–1393.
45. Kramer, J. and Hawley, R. S., The spindle-associated transmembrane
protein Axs identifies a new family of transmembrane proteins in eukar-
yotes. Cell Cycle 2003. 2: 174–176.
46. Hirota, T., Kunitoku, N., Sasayama, T., Marumoto, T., Zhang, D. W.,
et al. Aurora-A and an interacting activator, the LIM protein Ajuba, are
required for mitotic commitment in human cells. Cell 2003. 114: 585–598.
47. Barr, A. R. and Gergely, F., Aurora-A: the maker and breaker of spindle
poles. J Cell Sci 2007. 120: 2987–2996.
48. Tippit, D. H., Schulz, D. and Pickett-Heaps, J. D., Analysis of distribu-
tion of spindle microtubules in diatom Fragilaria. J Cell Biol 1978. 79:
737–763.
49. Tippit, D. H., Pickett-Heaps, J. D. and Leslie, R., Cell division in two
large pennate diatoms, Hantzschia and Nitzschia. III. A new proposal for
kinetochore function during prometaphase. J Cell Biol 1980. 86:
402–416.
50. McDonald, K. L., Pfister, K., Masuda, H., Wordeman, L., Staiger, C.,
et al. Comparison of spindle elongation in vivo and in vitro in Stepha-
nopyxis turris. J Cell Sci 1986. 5: 205–227.
51. Anderson, D. J. and Hetzer, M. W., The life cycle of the metazoan
nuclear envelope. Curr Opin Cell Biol 2008. 20: 386–392.
52. Lippincott-Schwartz, J., Cell biology – Ripping up the nuclear envel-
ope. Nature 2002. 416: 31–32.
53. Ribeiro, K. C., Pereira-Neves, A. and Benchimol, M., The mitotic
spindle and associated membranes in the closed mitosis of trichomo-
nads. Biol Cell 2002. 94: 157–172.
54. Heywood, P., Ultrastructure of mitosis in chloromonadophycean alga
Vacuolaria virescens. J Cell Sci 1978. 31: 37–51.
55. Baskin, T. I. and Cande, W. Z., The structure and function of the mitotic
spindle in flowering plants. Annu Rev Plant Physiol Plant Mol Biol 1990.
41: 277–315.
56. Karsenti, E. and Vernos, I., Cell cycle – the mitotic spindle: a self-made
machine. Science 2001. 294: 543–547.
57. Pickett-Heaps, J. D. and Tippit, D. H., Diatom spindle in perspective.
Cell 1978. 14: 455–467.
58. McDonald, K. L., Pickett-Heaps, J. D., McIntosh, J. R. and Tippit,
D. H., Mechanism of anaphase spindle elongation in Diatoma vulgare.
J Cell Biol 1977. 74: 377–388.
59. McDonald, K. L. and Cande, W. Z., Diatoms, the mechanism of ana-
phase spindle elongation. In: Coleman, A. W., Goff, L. J. and Stein-Taylor,
J. R. editors. Algae as experimental systems. New York, Alan R. Liss, Inc.,
1989. p. 3–18.
60. Pickett-Heaps, J. D., Tippit, D. H. and Andreozzi, J. A., Cell division in
pennate diatom Pinnularia. I. Early stages in mitosis. Biol Cell 1978. 33:
71–78.
61. Pickett-Heaps, J. D., Tippit, D. H. and Andreozzi, J. A., Cell division in
pennate diatom Pinnularia. II. Later stages in mitosis. Biol Cell 1978. 33:
79–84.
62. Cande, W. Z. and McDonald, K. L., In vitro reactivation of anaphase
spindle elongation using isolated diatom spindles. Nature 1985. 316:
168–170.
883

Review article A. De Martino, A. Amato and C. Bowler
63. Wordeman, L. and Cande, W. Z., Reactivation of spindle elongation
in vitro is correlated with the phosphorylation of a 205-kd spindle-
associated protein. Cell 1987. 50: 535–543.
64. Masuda, H., McDonald, K. L. and Cande, W. Z., The mechanism of
anaphase spindle elongation – uncoupling of tubulin incorporation and
microtubule sliding during in vitro spindle reactivation. J Cell Biol 1988.
107: 623–633.
65. Hogan, C. J., Wein, H., Wordeman, L., Scholey, J. M., Sawin, K. E.,
et al. Inhibition of anaphase spindle elongation in vitro by a peptide
antibody that recognizes kinesin motor domain. Proc Natl Acad Sci U S A
1993. 90: 6611–6615.
66. Hogan, C. J., Stephens, L., Shimizu, T. and Cande, W. Z., Physiolo-
gical evidence for involvement of a Kinesin-related protein during ana-
phase spindle elongation in diatom central spindles. J Cell Biol 1992.
119: 1277–1286.
67. Leslie, R. and Cande, W. Z., Spindle microtubule dynamics following
ultraviolet-microbeam irradiations of mitotic diatoms. Cell 1984. 36: 717–
727.
68. Cande, W. Z. and Hogan, C. J., The mechanism of anaphase spindle
elongation. Bioessays 1989. 11: 5–9.
69. McDonald, K. L., Pfister, K., Masuda, H., Wordeman, L., Staiger, C.,
et al. Comparison of spindle elongation in vivo and in vitro in Stepha-
nopyxis turris. J Cell Sci 1986. 5: 205–227.
70. Cande, W. Z. and McDonald, K. L., Physiological and ultrastructural
analysis of elongating mitotic spindles reactivated in vitro. J Cell Biol
1986. 103: 593–604.
71. Spurck, T. P. and Pickett-Heaps, J. D., The effects of diazepam on
mitosis and the microtubule cytoskeleton. I. Observations on the diatoms
Hantzschia amphioxys and Surirella robusta. J Cell Sci 1994. 107: 2643–
2651.
72. La Fountain, J. R. and Davidson, L. A., Analysis of spindle ultrastruc-
ture during prometaphase and metaphase of micronuclear division in
Tetrahymena. Chromosoma 1979. 75: 293–308.
73. Vogt, E., Kirsch-Volders, M., Parry, J. and Eichenlaub-Ritter, U.,
Spindle formation, chromosome segregation and the spindle checkpoint
in mammalian oocytes and susceptibility to meiotic error. Mutat Res,
Genet Toxicol 2008. 651: 14–29.
74. Mishima, M., Kaitna, S. and Glotzer, M., Central spindle assembly and
cytokinesis require a kinesin-like protein/RhoGAP complex with micro-
tubule bundling activity. Dev Cell 2002. 2: 41–54.
75. Clarke, L., Centromeres: proteins, protein complexes, and repeated
domains at centromeres of simple eukaryotes. Curr Opin Genet Dev
1998. 8: 212–218.
76. Murphy, T. D. and Karpen, G. H., Centromeres take flight: alpha satellite
and the quest for the human centromere. Cell 1998. 93: 317–320.
77. Dernburg, A. F., Here, there, and everywhere: kinetochore function on
holocentric chromosomes. J Cell Biol 2001. 153: F33–F38.
78. Amor, D. J., Kalitsis, P., Sumer, H. and Choo, K. H. A., Building the
centromere: from foundation proteins to 3D organization. Trends Cell Biol
2004. 14: 359–368.
79. Black, B. E., Foltz, D. R., Chakravarthy, S., Luger, K., Woods, V. L.,
et al. Structural determinants for generating centromeric chromatin.
Nature 2004. 430: 578–582.
80. Henikoff, S., Ahmad, K. and Malik, H. S., The centromere paradox:
stable inheritance with rapidly evolving DNA. Science 2001. 293: 1098–
1102.
81. Mellone, B. G. and Allshire, R. C., Stretching it: putting the CEN(P-A) in
centromere. Curr Opin Genet Dev 2003. 13: 191–198.
82. Kitagawa, K. and Hieter, P., Evolutionary conservation between budding
yeast and human kinetochores. Nat Rev Mol Cell Biol 2001. 2: 678–687.
83. Dawe, R. K. and Henikoff, S., Centromeres put epigenetics in the
driver’s seat. Trends Biochem Sci 2006. 31: 662–669.
84. Palmer, D. K., Oday, K., Trong, H. L., Charbonneau, H. and Margolis,
R. L., Purification of the centromere-specific protein CENP-A and
demonstration that it is a distinctive histone. Proc Natl Acad Sci U S A
1991. 88: 3734–3738.
85. Echeverri, C. J., Paschal, B. M., Vaughan, K. T. and Vallee, R. B.,
Molecular characterization of the 50-kD subunit of dynactin reveals
function for the complex in chromosome alignment and spindle organi-
zation during mitosis. J Cell Biol 1996. 132: 617–633.
884
86. Wei, R. R., Sorger, P. K. and Harrison, S. C., Molecular organization of
the Ndc80 complex, an essential kinetochore component. Proc Natl
Acad Sci U S A 2005. 102: 5363–5367.
87. Maiato, H., DeLuca, J. G., Salmon, E. D. and Earnshaw, W. C., The
dynamic kinetochore-microtubule interface. J Cell Sci 2004. 117: 5461–
5477.
88. DeLuca, J. G., Dong, Y. M., Hergert, P., Strauss, J., Hickey, J. M., et al.
Hec1 and Nuf2 are core components of the kinetochore outer plate
essential for organizing microtubule attachment sites. Mol Biol Cell 2005.
16: 519–531.
89. Kops, G. J. P. L., The kinetochore and spindle checkpoint in mammals.
Front Biosci 2008. 13: 3603–3620.
90. Malik, H. S. and Henikoff, S., Phylogenomics of the nucleosome. Nat
Struct Biol 2003. 10: 882–891.
91. Baker, R. E. and Rogers, K., Phylogenetic analysis of fungal centromere
H3 proteins. Genetics 2006. 174: 1481–1492.
92. Wordeman, L., McDonald, K. L. and Cande, W. Z., The distribution of
cytoplasmic microtubules throughout the cell cycle of the centric diatom
Stephanopixys turris: the role in nuclear migration and positioning the
mitotic spindle during cytokinesis. J Cell Biol 1986. 102: 1688–1698.
93. van de Meene, A. M. L. and Pickett-Heaps, J. D., Valve morphogenesis
in the centric diatom Rhizosolenia setigera (Bacillariophyceae, Cen-
trales) and its taxonomic implications. Eur J Phycol 2004. 39: 93–104.
94. Wang, H., Oliferenko, S. and Balasubramanian, M. K., Cytokinesis:
relative alignment of the cell division apparatus and the mitotic spindle.
Curr Opin Cell Biol 2003. 15: 82–87.
95. van de Meene, A. M. L. and Pickett-Heaps, J. D., Valve morphogenesis
in the centric diatom Proboscia alata Sundstrom. J Phycol 2002. 38: 351–
363.
96. Pickett-Heaps, J. D., Cell division and morphogenesis of the centric
diatom Chaetoceros decipiens (Bacillariophyceae) II. Electron micro-
scopy and a new paradigm for tip growth. J Phycol 1998. 34: 995–1004.
97. Van Damme, D. and Geelen, D., Demarcation of the cortical division
zone in dividing plant cells. Cell Biol Int 2008. 32: 178–187.
98. Heitz, M. J., Petersen, J., Valovin, S. and Hagan, I. M., MTOC formation
during mitotic exit in fission yeast. J Cell Sci 2001. 114: 4521–4532.
99. Glotzer, M., Animal cell cytokinesis. Annu Rev Cell Dev Biol 2001. 17:
351–386.
100. Pickett-Heaps, J. D., Tippit, D. H., Cohn, S. A. and Spurck, T. P.,
Microtubule dynamics in the spindle - theoretical aspects of assembly
disassembly reactions in vivo. J Theor Biol 1986. 1118: 153–169.
101. Pickett-Heaps, J. D., Tippit, D. H. and Leslie, R., Light and electron
microscopic observations on cell division in two large pennate diatoms,
Hantzschia and Nitzschia. I. Mitosis in vivo. Eur J Cell Biol 1980. 21: 1–11.
102. Pickett-Heaps, J. D., Tippit, D. H. and Leslie, R., Light and electron
microscopic observations on cell division in two large pennate diatoms,
Hantzschia and Nitzschia. II. Ultrastructure. Eur J Cell Biol 1980. 21: 12–
27.
103. Soranno, T. and Pickett-Heaps, J., Directionally controlled spindle
disassembly after mitosis in the diatom Pinnularia. Eur J Cell Biol
1982. 26: 234–243.
104. Tippit, D. H., McDonald, K. L. and Pickett-Heaps, J. D., Cell division in
centric diatom Melosira varians. Cytobiologie 1975. 12: 52–73.
105. Leslie, R. and Cande,W. Z., Ultraviolet microbeam irradiations of mitotic
diatoms - investigation of spindle elongation. J Cell Biol 1983. 96: 548–
561.
106. Mos, L., Domoic acid: a fascinating marine toxin. Environ Toxicol
Pharmacol 2001. 9: 79–85.
107. Bates, S. S., Bird, C. J., Defreitas, A. S. W., Foxall, R., Gilgan, M., et al.
Pennate diatom Nitzschia pungens as the primary source of domoic
acid, a toxin in shellfish from Eastern Prince Edward Island, Canada. Can
J Fish Aquat Sci 1989. 46: 1203–1215.
108. Buesseler, K. O., Andrews, J. E., Pike, S. M. and Charette, M. A., The
effects of iron fertilization on carbon sequestration in the Southern
Ocean. Science 2004. 304: 414–417.
109. Chisti, Y., Biodiesel from microalgae beats bioethanol. Trends Biotech-
nol 2008. 26: 126–131.
110. Ueda, M., Schliwa, M. and Euteneuer, U., Unusual centrosome cycle in
Dictyostelium: Correlation of dynamic behavior and structural changes.
Mol Biol Cell 1999. 10: 151–160.
BioEssays 31:874–884, � 2009 Wiley Periodicals, Inc.
Related Documents











