Kasetsart J. (Nat. Sci.) 46 : 346 - 362 (2012) 1 Center for Agricultural Biotechnology, Kasetsart University, Kamphaeng Saen Campus, Nakhon Pathom 73140, Thailand. 2 Center of Excellence on Agricultural Biotechnology (AG-BIO/PERDO-CHE), Bangkok 10900, Thailand. 3 Chachoengsao Rubber Research Center, Sanam Chai Khet, Chachoengsao 24160, Thailand. 4 Department of Plant Science, Faculty of Science, Mahidol University, Bangkok 10400, Thailand. 5 Institut de Recherche pour le Développement (IRD), UMR DIA-PC (UR118), 911 Avenue Agropolis, 34032 Montpellier, France. * Corresponding author, e-mail: [email protected] Received date : 06/12/11 Accepted date : 30/03/12 Mitochondrial/Cytosolic Acetyl CoA and Rubber Biosynthesis Genes Expression in Hevea brasiliensis Latex and Rubber Yield Somjintana Ruderman 1,2,3 , Panida Kongsawadworakul 4 , Unchera Viboonjun 4 Orarat Mongkolporn 1,2 and Hervé Chrestin 5,* ABSTRACT Expressions of the 22 genes involved in rubber biosynthesis, from the acetyl CoA genesis and the mevalonate (MVA) pathway to IPP polymerization, were analyzed by real-time polymerase chain reaction (PCR). The results revealed that the expressions of most genes were not related to the latex yield trait when compared among three high- and three low-yielding rubber clones. Only the genes HMGR3 and SRPP1 were significantly up-regulated in low-yielding clones. The higher expression of the SRPP1 gene in low-yielding clones suggested a possible increase in the number and higher rubber biosynthetic activity of the active small rubber particles, which may produce latex with higher viscosity, impairing the latex flow and limiting the latex yield. The up-regulation of the HMGR3 gene in low-yielding clones may reflect a higher amount or activity or both of Frey-Wyssling particles (plastids) in their latex. These plastids have been shown to be involved in oxidative processes that induced earlier laticifers plugging, which also impaired the latex flow and yield. In conclusion, a possible crossover between the cytosolic MVA and the plastidic 1-deoxy-D-xylulose-5-phosphate/2-C-methyl-D-erythritol-4-phosphate (DXP/ MEP) pathways is suggested, which might especially take place in the laticifers of the high-yielding clones, to fulfill the requirement in isopentenyl pyrophosphate for more active rubber biosynthesis. Keywords: acetyl CoA, 1-deoxy-D-xylulose-5-phosphate/2-C-methyl-D-erythritol-4-phosphate (DXP/ MEP) pathway, Hevea brasiliensis, mevalonate pathway, rubber biosynthesis Abbreviations: ACAT = Acetyl CoA acetyltransferase; ACL = ATP-citrate lyase; DMAPP = Dimethylallyl diphosphate; DPMDC = Disphosphomevalonate decarboxylase; DRC = Dry rubber content; DXP/MEP = 1-Deoxy-D-xylulose-5-phosphate/2-C-methyl-D-erythritol-4-phosphate; FWPs = Frey-Wyssling particles; HMGR = 3-Hydroxy 3-methylglutaryl CoA reductase; HMGS = 3-Hydroxy 3-methylglutaryl CoA synthase; IPP = Isopentenyl pyrophosphate; IPPI = Isopentenyl diphosphate isomerase; MVA = Mevalonate; MVK = Mevalonate kinase; PDCE = Pyruvate dehydrogenase complex; PMK = Phosphomevalonate kinase; PT = Prenyltransferase; RCB = Randomized complete block design; REF = Rubber elongation factor; rGE = Relative gene expression; SRPP = Small rubber particle protein.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Kasetsart J. (Nat. Sci.) 46 : 346 - 362 (2012)
1 Center for Agricultural Biotechnology, Kasetsart University, Kamphaeng Saen Campus, Nakhon Pathom 73140, Thailand.2 Center of Excellence on Agricultural Biotechnology (AG-BIO/PERDO-CHE), Bangkok 10900, Thailand.3 Chachoengsao Rubber Research Center, Sanam Chai Khet, Chachoengsao 24160, Thailand.4 Department of Plant Science, Faculty of Science, Mahidol University, Bangkok 10400, Thailand.5 Institut de Recherche pour le Développement (IRD), UMR DIA-PC (UR118), 911 Avenue Agropolis, 34032 Montpellier,
France. * Corresponding author, e-mail: [email protected]
Received date : 06/12/11 Accepted date : 30/03/12
Mitochondrial/Cytosolic Acetyl CoA and Rubber Biosynthesis Genes Expression in Hevea brasiliensis Latex and Rubber Yield
Somjintana Ruderman1,2,3, Panida Kongsawadworakul4, Unchera Viboonjun4 Orarat Mongkolporn1,2 and Hervé Chrestin5,*
ABSTRACT
Expressions of the 22 genes involved in rubber biosynthesis, from the acetyl CoA genesis and the mevalonate (MVA) pathway to IPP polymerization, were analyzed by real-time polymerase chain reaction (PCR). The results revealed that the expressions of most genes were not related to the latex yield trait when compared among three high- and three low-yielding rubber clones. Only the genes HMGR3 and SRPP1 were signifi cantly up-regulated in low-yielding clones. The higher expression of the SRPP1 gene in low-yielding clones suggested a possible increase in the number and higher rubber biosynthetic activity of the active small rubber particles, which may produce latex with higher viscosity, impairing the latex fl ow and limiting the latex yield. The up-regulation of the HMGR3 gene in low-yielding clones may refl ect a higher amount or activity or both of Frey-Wyssling particles (plastids) in their latex. These plastids have been shown to be involved in oxidative processes that induced earlier laticifers plugging, which also impaired the latex fl ow and yield. In conclusion, a possible crossover between the cytosolic MVA and the plastidic 1-deoxy-D-xylulose-5-phosphate/2-C-methyl-D-erythritol-4-phosphate (DXP/MEP) pathways is suggested, which might especially take place in the laticifers of the high-yielding clones, to fulfi ll the requirement in isopentenyl pyrophosphate for more active rubber biosynthesis.Keywords: acetyl CoA, 1-deoxy-D-xylulose-5-phosphate/2-C-methyl-D-erythritol-4-phosphate (DXP/
MEP) pathway, Hevea brasiliensis, mevalonate pathway, rubber biosynthesis
Abbreviations: ACAT = Acetyl CoA acetyltransferase; ACL = ATP-citrate lyase; DMAPP = Dimethylallyl diphosphate; DPMDC = Disphosphomevalonate decarboxylase; DRC = Dry rubber content; DXP/MEP = 1-Deoxy-D-xylulose-5-phosphate/2-C-methyl-D-erythritol-4-phosphate; FWPs = Frey-Wyssling particles; HMGR = 3-Hydroxy 3-methylglutaryl CoA reductase; HMGS = 3-Hydroxy 3-methylglutaryl CoA synthase; IPP = Isopentenyl pyrophosphate; IPPI = Isopentenyl diphosphate isomerase; MVA = Mevalonate; MVK = Mevalonate kinase; PDCE = Pyruvate dehydrogenase complex; PMK = Phosphomevalonate kinase; PT = Prenyltransferase; RCB = Randomized complete block design; REF = Rubber elongation factor; rGE = Relative gene expression; SRPP = Small rubber particle protein.

INTRODUCTION
Natural rubber is produced as rubber particles in the cytoplasm of laticifers, the specialized anastomozed cells located in the inner bark of the rubber tree (Hevea brasiliensis). The latex (laticifers cytoplasm) is simply collected by bark wounding or deliberate tapping. Hevea
brasiliensis natural rubber has long been reported to be synthesized through the cytosolic MVA pathway (Archer and Audley, 1967) as shown in Figure 1. Rubber biosynthesis includes three successive steps: 1) the generation of acetyl CoA; 2) the isoprenoid biosynthesis (MVA pathway) resulting in the production of the IPP molecule and its isomer, DMAPP; and 3) the polymerization of
Figure 1 Interconnections between the glycolysis, mevalonate and rubber biosynthesis pathways, through the mitochondrial pyruvate dehydrogenase complex (PDCE) and cytosolic ATP-citrate lyase (ACL) acetyl CoA biosynthesis. ATP = Adenosine triphosphate; CoA = Coenzyme A; Dimethylallyl-PP = Dimethylallyl diphosphate; DPMDC = Disphosphomevalonate decarboxylase; HMGCoA = 3-Hydroxy 3-methylglutaryl CoA; HMGR = 3-Hydroxy 3-methylglutaryl CoA reductase; HMGS = 3-Hydroxy 3-methylglutaryl CoA synthase; Isopentenyl-PP = Isopentenyl pyrophosphate; Mevalonate-5-P = Mevalonate-5-phosphate; Mevalonate-5-PP = Mevalonate-5-diphosphate; MK = Mevalonate kinase; PMK = Phosphomevalonate kinase; PTs = Prenyltransferases; REFs = Rubber elongation factors; SRPPs = Small rubber particle proteins
Kasetsart J. (Nat. Sci.) 46(3) 347

Kasetsart J. (Nat. Sci.) 46(3)348
IPP and DMAPP for initiation of the rubber chains and then of IPP into long cis-1,4-polyisoprene chains (rubber) by PT, assisted by the REF (Dennis and Light, 1989) and SRPP (Oh et al., 1999), which are located on the surface of the rubber particles. Another plant metabolic pathway for IPP synthesis, called DXP/MEP, which is located in the plastids (Rohmer et al., 1996; Lichtenthaler, 1999), has recently been evidenced through molecular studies in Hevea brasiliensis latex (Ko et al., 2003). Radiolabeling of intermediates of the cytosolic MVA pathway provided evidence that synthesized IPP did incorporate into natural rubber (Skilleter and Kekwick, 1971). Expression of the enzymes in the DXP/MEP pathway could be proven in laticifers of Hevea brasiliensis; however, using [1-13C] 1-deoxy-D-xylulose triacetate, an intermediate of the MEP pathway, no isotopic-labelled rubber molecules could be detected (Ko et al., 2003; Sando et al., 2009). This suggested that the IPP derived from the plastidial DXP/MEP pathway might not be involved in Hevea brasiliensis rubber biosynthesis, but rather in carotenoids and other small isoprenoids biosynthesis in the latex plastids, FWPs (Chow et al., 2012), at least under normal conditions. It is worth noting that whatever the pathway, IPP biosynthesis operates at an equivalent high energy and reducing power cost, since the generation of each IPP molecule consumes 3 ATP and 2 NADPH through both the MVA and DXP/MEP pathways, from mitochondrial citrate and the cytosolic pyruvate, respectively. Acetyl CoA, a metabolite connecting the catabolic and anabolic metabolisms, is a key intermediate in energy generation, especially through the initiation of the mitochondrial citric acid/Krebs cycle followed by oxidative phosphorylation. It is also involved in numerous anabolic pathways (Oliver et al., 2009), including Hevea brasiliensis rubber biosynthesis in the latex cytosol (Archer and Audley, 1967). Since acetyl CoA is not permeable to biological
membranes, its biogenesis occurs independently via various pathways in subcellular compartments, cytosol, mitochondria, plastids, nuclei and peroxisomes, where it is required (Oliver et al., 2009). In mitochondria, acetyl CoA is mostly synthesized from the glycolysis-generated cytosolic pyruvate through the mitochondrial multi-enzyme pyruvate dehydrogenase complex (PDCE). The PDCE is composed of three major enzymes that act sequentially (Perham and Packman, 1989)—pyruvate dehydrogenase (PDCE1) (EC 1.2.4.1), dihydro-lipoamide S-acetyltransferase (PDCE2) (EC 2.3.1.12) and dihydrolipoamide dehydrogenase (PDCE3) (EC 1.8.1.4). It is now well documented that most of the cytosolic acetyl CoA pool is synthesized from the mitochondrial citrate, which after its transportation into the cytosol by mitochondrial di(oxaloacetate)/tricarboxylate(citrate) translocators (Hanning et al., 1999), the citrate is split into oxaloacetate and acetyl CoA through the action of cytosolic ACL (EC 2.3.3.8) (Fatland et al., 2005) as shown in Figure 1. The cytosolic-released oxaloacetate can be re-imported into the mitochondria through the same di/tricarboxylate shuttle, resulting in a citrate-oxaloacetate Krebs-cycle short circuit (Fatland et al., 2005) and thus in a decrease in energy (ATP) availability. The latex yield is limited by two main intrinsic factors in the latex-producing tissues. The fi rst is the latex fl ow rate and its duration, which are infl uenced by: 1) the turgor pressure of the inner bark tissues (d’Auzac et al., 1989); 2) the latex viscosity which, as in all synthetic and natural colloids, depends on the percentage of dry matter (dry rubber content [DRC] of latex) (Van Gils, 1951; Cornish and Brichta, 2002) and mainly on the rubber particle size and particle size dispersion (PSD) (Greenwood et al., 1995; Blackley, 1997; Chu et al., 1998; Cornish and Brichta, 2002); and 3) the latex coagulation effi ciency (Brzozowska-Hanower et al., 1978; Gidrol et al., 1994; Kongsawadworakul and Chrestin, 2003;

Kasetsart J. (Nat. Sci.) 46(3) 349
Wititsuwannakul et al., 2008). The second intrinsic limiting factor is the ability of the latex cells to regenerate their whole lost cytoplasm, including rubber, between two consecutive tappings, which mainly depends on their metabolic orientation and activity (Jacob et al., 1989). As the MVA pathway has long been proven to be directly involved in Hevea brasiliensis rubber synthesis, the objective of this study aimed to investigate the expression of 22 selected genes involved in the acetyl CoA genesis and MVA pathways in relation to the clonal latex yield trait. Genes that might be found to be related to rubber yield may eventually be used as specifi c molecular markers of yield potential in the rubber tree breeding and selection programs.
MATERIALS AND METHODS
Plant materials Latex samples of three high- and three low-latex yielding clones planted in Thailand were used (Table 1). These rubber clones, aged 8.5 y had been planted in a small-scale clone trial at the Surat Thani Rubber Research Center (SRRC), Thailand in 1999. An RCB with three replications was used for the rubber planting layout: six rubber clones, eight trees per clone per replication and a row spacing of 2.75 × 3 m. Alternate day tapping without any bark stimulation treatment was started in 2006 when the trees were age 7 y.
Measurement of latex (rubber) yield and dry rubber content Both latex yield and DRC measurements were performed according to the standard method for small-scale trial data collection at SRRC (Sangsing et al., 1998). The latex yield of individual trees was measured only twice a month (once every 2 w). After stopping the latex fl ow, 5% formic acid was added into the latex cup and stirred to accelerate latex coagulation. The coagulated rubber (cup lump) of individual trees
was collected, air dried for 16 d and then weighed. The yield was calculated by subtracting 15% of the moisture content from the total air-dried weight (Equation 1): Yield = Air-dried weight × 0.85 (1)where the yield and total air-dried weight were expressed in grams of dry rubber per tree per tapping. To measure the DRC of each clone, three replications of 50 mL of pooled latex per clone per plot (eight trees) were collected in just one tapping before latex yield collection. Then, 2% formic acid was added to 8–10 g fresh latex to induce coagulation. The coagulated latex was fl atted into a small rubber sheet, dried in a hot air-oven at 70 °C for 16 h and weighed. The DRC was calculated using Equation 2: DRC = (dry rubber weight/fresh latex weight) × 100 (2) Latex collection and RNA extraction Trees that had been tapped for 1.5 y were selected based on their within-groups girth size homogeneity as well as their yield per tree per tapping per girth size (Table 1). An RCB design with six treatments (clones) and two replications was designed for latex collection. Each treatment consisted of three trees per replication. The latex sample was equally pooled from each tree (2 mL × three trees) and mixed in 6 mL of 2X “fi xation/extraction” buffer (50 mM Tris-HCl, 300 mM LiCl, 10 mM EDTA, 10% SDS, pH 9.0). The latex samples were immediately deep-frozen in liquid nitrogen and then stored at -80 °C. Total RNA was extracted using the latex LiCl precipitation method, as described by Pujade-Renaud et al. (1994).
cDNA synthesis After treating with DNase I (Ambion; USA), 1.5 μg of total RNA was used as the template for the fi rst-strand cDNA synthesis (SuperScript® III, First-Strand Synthesis System; Invitrogen;

Kasetsart J. (Nat. Sci.) 46(3)350Ta
ble
1 Va
lues
(mea
n ±
SE) f
or la
tex
sam
ples
of h
igh-
and
low
-yie
ldin
g cl
ones
from
the
smal
l-sca
le c
lone
tria
l at S
urat
Tha
ni R
ubbe
r Res
earc
h C
ente
r, Th
aila
nd.
Y
ield
Y
ield
Y
ield
(g
ram
s per
(g
ram
s per
tree
(g
ram
s per
tree
per
G
irth
Cut
leng
th
tre
e pe
r pe
r tap
ping
per
ta
ppin
g pe
r cm
cut
(c
m)
(cm
)
tapp
ing)
cm
girt
h)
leng
th)
Rep
licat
ion
I
RR
IT 2
51
HY
51
.29
± 1
.64
1.00
± 0
.07
1.79
± 0
.17
51.5
7 ±
2.08
28
.83
± 1.
89
-
CH
-35-
210
HY
46
.63
± 2
.05
0.88
± 0
.04
1.49
± 0
.06
52.7
3 ±
1.81
31
.33
± 0.
76
-
BPM
24
HY
44
.63
± 10
.31
0.85
± 0
.16
1.42
± 0
.27
51.9
3 ±
2.98
31
.33
± 1.
44
-
RR
IM60
0 LY
25
.02
± 6
.34
0.48
± 0
.11
0.88
± 0
.19
52.0
0 ±
1.13
28
.25
± 1.
06
-
CH
-35-
449
LY
19.8
6 ±
2.0
2 0.
34 ±
0.0
3 0.
60 ±
0.0
2 59
.00
± 3.
63
33.0
0 ±
3.28
-
C
H-3
5-O
P-20
19
LY
19.8
8 ±
1.0
3 0.
30 ±
0.0
1 0.
55 ±
0.0
4 66
.90
± 2.
71
36.0
0 ±
0.87
-
Rep
licat
ion
II
R
RIT
251
H
Y
59.2
6 ±
5.7
8 1.
04 ±
0.1
2 1.
97 ±
0.1
7 56
.97
± 1.
52
30.1
7 ±
1.76
-
C
H-3
5-21
0 H
Y
64.5
7 ±
9.1
7 1.
08 ±
0.1
0 1.
78 ±
0.0
7 59
.90
± 4.
60
36.1
7 ±
3.88
-
B
PM 2
4 H
Y
48.1
8 ±
8.3
9 1.
00 ±
0.0
0 1.
60 ±
0.2
6 53
.44
± 8.
39
30.1
7 ±
1.61
-
R
RIM
600
LY
28.6
2 ±
0.4
6 0.
48 ±
0.0
4 0.
91 ±
0.0
3 59
.45
± 6.
15
31.5
0 ±
1.41
-
C
H-3
5-44
9 LY
24
.42
± 1
.14
0.43
± 0
.03
0.72
± 0
.05
57.4
0 ±
6.61
33
.83
± 1.
53
-
CH
-35-
OP-
2019
LY
24
.96
± 7
.35
0.38
± 0
.09
0.66
± 0
.14
64.8
0 ±
6.66
37
.33
± 3.
25
-Av
erag
e
(fro
m 2
repl
icat
ions
)
RR
IT 2
51
HY
55
.28
± 5
.63
1.02
± 0
.03
1.88
± 0
.13
54.2
7 ±
3.82
29
.50
± 0.
94
35.9
C
H-3
5-21
0 H
Y
55.6
0 ±
12.6
9 0.
98 ±
0.1
4 1.
63 ±
0.2
1 56
.32
± 5.
07
33.7
5 ±
3.42
39
.5
BPM
24
HY
46
.41
± 2
.51
0.93
± 0
.10
1.51
± 0
.13
52.6
9 ±
1.07
30
.75
± 0.
82
33.7
R
RIM
600
LY
26.8
2 ±
2.5
5 0.
48 ±
0.0
0 0.
90 ±
0.0
2 55
.73
± 5.
27
29.8
8 ±
2.30
34
.7
CH
-35-
449
LY
22.1
4 ±
3.2
2 0.
38 ±
0.0
6 0.
66 ±
0.0
9 58
.20
± 1.
13
33.4
2 ±
0.59
36
.3
CH
-35-
OP-
2019
LY
22
.42
± 3
.59
0.34
± 0
.06
0.61
± 0
.08
65.8
5 ±
1.48
36
.67
± 0.
94
38.2
HY
= h
igh-
yiel
ding
clo
ne; L
Y =
low
-yie
ldin
g cl
one;
DR
C =
dry
rubb
er c
onte
nt.
C
lone
C
lass
ifi ca
tion
D
RC

Kasetsart J. (Nat. Sci.) 46(3) 351
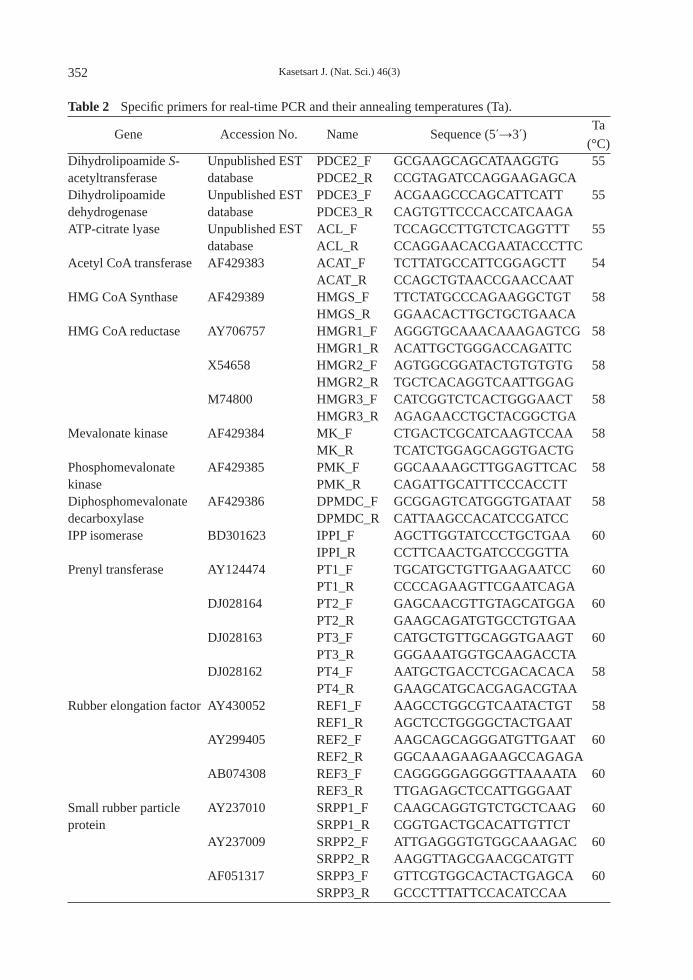
USA) in accordance with the manufacturer’s protocol. Primer design and verifi cation The 22 specific primer pairs were designed from either full-length cDNA sequences available in the NCBI database, or ESTs sequences from an unpublished database, using the Primer3 program. These primers were designed to have a Tm range of 54–60 °C and PCR product sizes of 150–250 bp (Table 2). The primers of genes that had more than one isoform were designed from the less conserved parts of the aligned sequences. The PCR products from every primer pair were validated by reverse transcription polymerase chain reaction and the amplifi ed fragments were isolated, purified, ligated to pGEM®-T Easy Vector (Promega) and transformed into E. coli DH5α. The plasmid DNA was sequenced by 1st BASE DNA Sequencing Service (Malaysia). After removing the vector sequences, the sequences were validated by homology searches against the GenBank database using BLASTN and BLASTX programs. Gene expression analysis by quantitative real-time PCR All quantitative real-time PCRs (qRT-PCRs) were performed in triplicate, using generated cDNA as a template and an ABI-7500 real-time PCR machine (Applied Biosystems; Foster City, CA, USA) as described by Tungngoen et al. (2009). Actin (a house keeping gene) was selected as a suitable reference gene. The percentage amplifi cation effi ciency (PE) of all genes, including the reference gene was verifi ed by qRT-PCR with serial dilutions of cDNA templates: 1/10, 1/25, 1/50, 1/100, 1/200 and 1/300. Standard curves for PE calculations were performed according to Livak and Schmittgen (2001). The acceptable PE should be in the range 0.8–1.2 (Yuan et al., 2006) for relative gene expression calculation using Equation 3: Relative gene expression = 2-ΔCt (3)
where ∆Ct = (Ct of target gene – Ct of reference gene)
Statistical analysis Normal distribution and the signifi cance of the relative gene expression were tested by one-way analysis of variance (ANOVA). Differences were accepted as highly signifi cant at P ≤ 0.01 and signifi cant at P ≤ 0.05. Mean comparisons were performed by Duncan’s test, at P ≤ 0.05. The correlation coeffi cient was calculated using Microsoft Excel 2003.
RESULTS
Rubber yield and rubber biosynthesis genes correlation Taking into account the rubber clones tested, statistical analysis showed there was a negative but not signifi cant correlation between the clonal rubber yield and the latex DRC (Table 3). These results were in accordance with Vinod et al. (2000), who studied rubber yield in relation to various putatively related traits in 10 different high-, medium- and low-yielding clones that showed no signifi cant correlation between DRC and the total volume of latex collected, as well as with the dry rubber yield. In contrast, a negative correlation between rubber yield and DRC had been previously reported for some rubber clones (Paardekooper and Sookmark, 1969). Furthermore, the statistical analysis in the current study did not show any highly signifi cant relationships between the expression of the 22 studied genes putatively involved in the different steps of rubber biosynthesis and the average rubber yields. Indeed, three genes (ACAT, HMGR1 and IPPI) exhibited only not signifi cant positive correlations, while the other 19 genes exhibited only not signifi cant negative correlations (Table 3) with yield. These results suggested that there was no or a putative nonlinear relationship between the expression of these genes and the clonal rubber yield.
Kasetsart J. (Nat. Sci.) 46(3)352
Table 2 Specifi c primers for real-time PCR and their annealing temperatures (Ta). Ta (°C)Dihydrolipoamide S- Unpublished EST PDCE2_F GCGAAGCAGCATAAGGTG 55acetyltransferase database PDCE2_R CCGTAGATCCAGGAAGAGCA Dihydrolipoamide Unpublished EST PDCE3_F ACGAAGCCCAGCATTCATT 55dehydrogenase database PDCE3_R CAGTGTTCCCACCATCAAGA ATP-citrate lyase Unpublished EST ACL_F TCCAGCCTTGTCTCAGGTTT 55 database ACL_R CCAGGAACACGAATACCCTTC Acetyl CoA transferase AF429383 ACAT_F TCTTATGCCATTCGGAGCTT 54 ACAT_R CCAGCTGTAACCGAACCAAT HMG CoA Synthase AF429389 HMGS_F TTCTATGCCCAGAAGGCTGT 58 HMGS_R GGAACACTTGCTGCTGAACA HMG CoA reductase AY706757 HMGR1_F AGGGTGCAAACAAAGAGTCG 58 HMGR1_R ACATTGCTGGGACCAGATTC X54658 HMGR2_F AGTGGCGGATACTGTGTGTG 58 HMGR2_R TGCTCACAGGTCAATTGGAG M74800 HMGR3_F CATCGGTCTCACTGGGAACT 58 HMGR3_R AGAGAACCTGCTACGGCTGA Mevalonate kinase AF429384 MK_F CTGACTCGCATCAAGTCCAA 58 MK_R TCATCTGGAGCAGGTGACTG Phosphomevalonate AF429385 PMK_F GGCAAAAGCTTGGAGTTCAC 58kinase PMK_R CAGATTGCATTTCCCACCTT Diphosphomevalonate AF429386 DPMDC_F GCGGAGTCATGGGTGATAAT 58decarboxylase DPMDC_R CATTAAGCCACATCCGATCC IPP isomerase BD301623 IPPI_F AGCTTGGTATCCCTGCTGAA 60 IPPI_R CCTTCAACTGATCCCGGTTA Prenyl transferase AY124474 PT1_F TGCATGCTGTTGAAGAATCC 60 PT1_R CCCCAGAAGTTCGAATCAGA DJ028164 PT2_F GAGCAACGTTGTAGCATGGA 60 PT2_R GAAGCAGATGTGCCTGTGAA DJ028163 PT3_F CATGCTGTTGCAGGTGAAGT 60 PT3_R GGGAAATGGTGCAAGACCTA DJ028162 PT4_F AATGCTGACCTCGACACACA 58 PT4_R GAAGCATGCACGAGACGTAA Rubber elongation factor AY430052 REF1_F AAGCCTGGCGTCAATACTGT 58 REF1_R AGCTCCTGGGGCTACTGAAT AY299405 REF2_F AAGCAGCAGGGATGTTGAAT 60 REF2_R GGCAAAGAAGAAGCCAGAGA AB074308 REF3_F CAGGGGGAGGGGTTAAAATA 60 REF3_R TTGAGAGCTCCATTGGGAAT Small rubber particle AY237010 SRPP1_F CAAGCAGGTGTCTGCTCAAG 60protein SRPP1_R CGGTGACTGCACATTGTTCT AY237009 SRPP2_F ATTGAGGGTGTGGCAAAGAC 60 SRPP2_R AAGGTTAGCGAACGCATGTT AF051317 SRPP3_F GTTCGTGGCACTACTGAGCA 60 SRPP3_R GCCCTTTATTCCACATCCAA
Gene Accession No. Name Sequence (5 →3 )
Kasetsart J. (Nat. Sci.) 46(3) 353
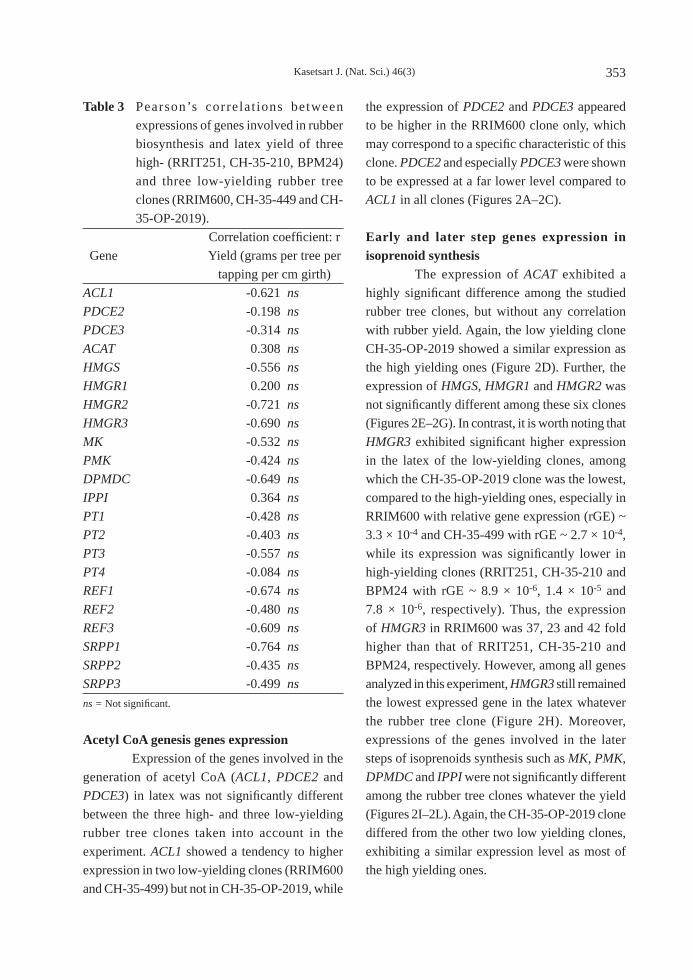
Acetyl CoA genesis genes expression Expression of the genes involved in the generation of acetyl CoA (ACL1, PDCE2 and PDCE3) in latex was not signifi cantly different between the three high- and three low-yielding rubber tree clones taken into account in the experiment. ACL1 showed a tendency to higher expression in two low-yielding clones (RRIM600 and CH-35-499) but not in CH-35-OP-2019, while
the expression of PDCE2 and PDCE3 appeared to be higher in the RRIM600 clone only, which may correspond to a specifi c characteristic of this clone. PDCE2 and especially PDCE3 were shown to be expressed at a far lower level compared to ACL1 in all clones (Figures 2A–2C).
Early and later step genes expression in isoprenoid synthesis The expression of ACAT exhibited a highly signifi cant difference among the studied rubber tree clones, but without any correlation with rubber yield. Again, the low yielding clone CH-35-OP-2019 showed a similar expression as the high yielding ones (Figure 2D). Further, the expression of HMGS, HMGR1 and HMGR2 was not signifi cantly different among these six clones (Figures 2E–2G). In contrast, it is worth noting that HMGR3 exhibited signifi cant higher expression in the latex of the low-yielding clones, among which the CH-35-OP-2019 clone was the lowest, compared to the high-yielding ones, especially in RRIM600 with relative gene expression (rGE) ~ 3.3 × 10-4 and CH-35-499 with rGE ~ 2.7 × 10-4, while its expression was signifi cantly lower in high-yielding clones (RRIT251, CH-35-210 and BPM24 with rGE ~ 8.9 × 10-6, 1.4 × 10-5 and 7.8 × 10-6, respectively). Thus, the expression of HMGR3 in RRIM600 was 37, 23 and 42 fold higher than that of RRIT251, CH-35-210 and BPM24, respectively. However, among all genes analyzed in this experiment, HMGR3 still remained the lowest expressed gene in the latex whatever the rubber tree clone (Figure 2H). Moreover, expressions of the genes involved in the later steps of isoprenoids synthesis such as MK, PMK, DPMDC and IPPI were not signifi cantly different among the rubber tree clones whatever the yield (Figures 2I–2L). Again, the CH-35-OP-2019 clone differed from the other two low yielding clones, exhibiting a similar expression level as most of the high yielding ones.
Table 3 Pearson’s correlat ions between expressions of genes involved in rubber biosynthesis and latex yield of three high- (RRIT251, CH-35-210, BPM24) and three low-yielding rubber tree clones (RRIM600, CH-35-449 and CH-35-OP-2019).
Correlation coeffi cient: r Gene Yield (grams per tree per tapping per cm girth)ACL1 -0.621 nsPDCE2 -0.198 nsPDCE3 -0.314 nsACAT 0.308 nsHMGS -0.556 nsHMGR1 0.200 nsHMGR2 -0.721 nsHMGR3 -0.690 nsMK -0.532 nsPMK -0.424 nsDPMDC -0.649 nsIPPI 0.364 nsPT1 -0.428 nsPT2 -0.403 nsPT3 -0.557 nsPT4 -0.084 nsREF1 -0.674 nsREF2 -0.480 nsREF3 -0.609 nsSRPP1 -0.764 nsSRPP2 -0.435 nsSRPP3 -0.499 nsns = Not signifi cant.

Kasetsart J. (Nat. Sci.) 46(3)354
Figure 2 Expressions of A) ACL1; B) PDCE2; and C) PDCE3 genes involved in acetyl CoA genesis; D) ACAT; E) HMGS; F) HMGR1; and G) HMGR2 involved in the early steps of isoprenoids synthesis; H) plastidic HMGR3 probably involved in rubber biosynthesis; I) MK; J) PMK; K) DPMDC; and L) IPPI involved in the later steps of isoprenoids synthesis. ns = Not signifi cant, ** = Highly signifi cant (P ≤ 0.01). Different lower case letters indicate values that are signifi cantly different (One-way analysis of variance; Duncan’s test, P ≤ 0.05). Each bar graph is a mean of four independent experiments, with error bars representing the standard error; n = 4.
Kasetsart J. (Nat. Sci.) 46(3) 355
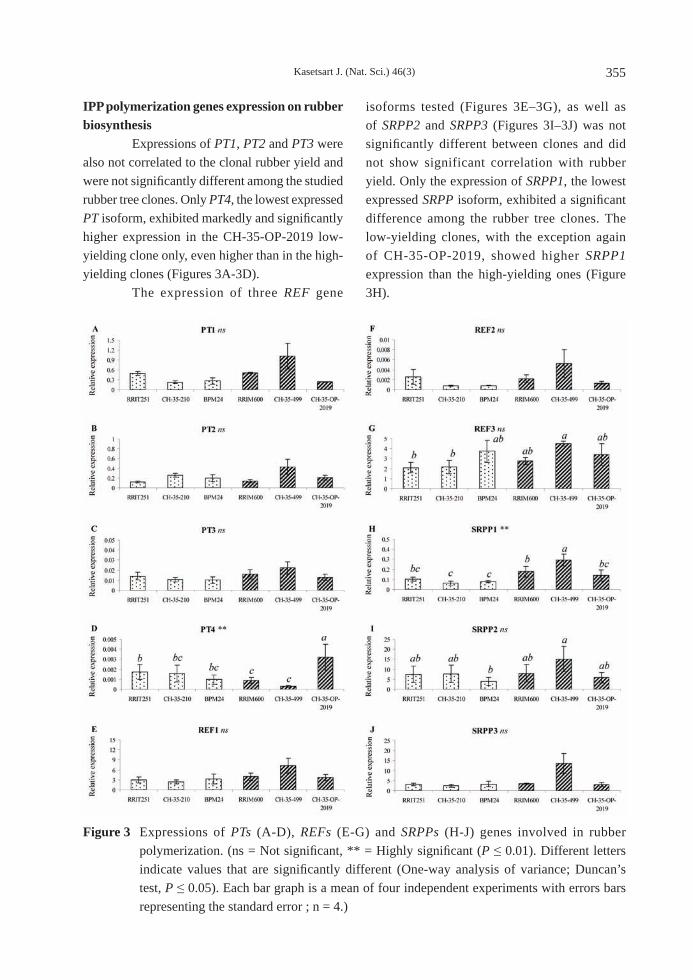
Figure 3 Expressions of PTs (A-D), REFs (E-G) and SRPPs (H-J) genes involved in rubber polymerization. (ns = Not signifi cant, ** = Highly signifi cant (P ≤ 0.01). Different letters indicate values that are signifi cantly different (One-way analysis of variance; Duncan’s test, P ≤ 0.05). Each bar graph is a mean of four independent experiments with errors bars representing the standard error ; n = 4.)
IPP polymerization genes expression on rubber biosynthesis Expressions of PT1, PT2 and PT3 were also not correlated to the clonal rubber yield and were not signifi cantly different among the studied rubber tree clones. Only PT4, the lowest expressed PT isoform, exhibited markedly and signifi cantly higher expression in the CH-35-OP-2019 low-yielding clone only, even higher than in the high-yielding clones (Figures 3A-3D). The expression of three REF gene
isoforms tested (Figures 3E–3G), as well as of SRPP2 and SRPP3 (Figures 3I–3J) was not signifi cantly different between clones and did not show significant correlation with rubber yield. Only the expression of SRPP1, the lowest expressed SRPP isoform, exhibited a signifi cant difference among the rubber tree clones. The low-yielding clones, with the exception again of CH-35-OP-2019, showed higher SRPP1 expression than the high-yielding ones (Figure 3H).

Kasetsart J. (Nat. Sci.) 46(3)356
DISCUSSION
Acetyl CoA genesis genes expression PDCE2 and PDCE3 are involved in the conversion of the glycolysis-derived cytosolic pyruvate into mitochondrial acetyl CoA, the common source of eukaryotic cell energy (ATP) production through the Krebs cycle. It has been reported that high latex metabolism traits are linked to high latex yield traits (Eschbach et al., 1986). Chrestin and Bangratz (1988) also reported the positive signifi cant correlations between latex ATP level, energy charge and the clonal latex yield. However, the current study found that the expression of these PDCEs was not signifi cantly correlated with the clonal rubber yield. Apart from its major role in energy production, acetyl CoA is an important molecule used in many biochemical reactions of cell metabolism. Thus, in Hevea brasiliensis laticifers, acetyl CoA is not only used for energy production and biosynthesis of rubber—the end product of secondary metabolism—but also for the entire range of the metabolic activities necessary for the synthesis/regeneration of the vital components of the cell cytoplasm (such as fatty acids, steroids, small isoprenoids and membrane structures among others), which fl ow out by tapping, especially in the case of tapped tissues. Even though PDCE2 and PDCE3 were expressed at a relatively low level in the laticifers mitochondria, it may suggest that in the Hevea brasiliensis laticifers, acetyl CoA biosynthesis/availability might not be directly limited by the expression of these two genes but regulated at other levels, including the competition with other acetyl CoA-dependent metabolic pathways. ACL is the cytosolic enzyme that catalyzes a cleavage of mitochondrial citrate, which is exported into the cytosol, to yield cytosolic acetyl CoA, a precursor of rubber and other phytochemical biosyntheses through the MVA pathway. Thus, the acetyl CoA derived from the activity of cytosolic ACL competes
with the ATP production at the starting point of the mitochondrial Krebs cycle. Although the expression of ACL1 was not signifi cantly different among high- and low-yielding rubber clones, this gene showed a tendency to be more highly expressed in two low-yielding clones (RRIM600 and CH-35-499) but not in the CH-35-OP-2019 clone compared with the high-yielding ones, which were quite homogeneous with regard to the expression of this gene. This may indicate that a higher expression of ACL1 might be potentially linked to low latex yield. However, ACL enzymatic activity, if mainly regulated at its gene transcription level, should lead to higher cytosolic acetyl CoA availability and thus to better rubber synthesis effi ciency, through the cytosolic MVA pathway.
Isoprenoids biosynthesis (mevalonate pathway) genes expression Most of the genes in the MVA pathway did not exhibit signifi cant differences in their expression levels among the high- and low-yielding rubber clones, except for ACAT and HMGR3. The expression of ACAT was signifi cantly different among the rubber clones, but not between high- and low-yielding groups. Similarly, Chuenwarin et al. (2010) reported that the expression of ACAT was not signifi cantly different between the low- (PB217) and high- (PB260) latex yielding clones. This can be explained by the fact that ACAT is involved not only in rubber biosynthesis but also in several metabolic pathways; thus, its expression level may not be directly linked to the rubber yield. Therefore, the difference in expression of ACAT in different clones may be only linked to some other clonal characteristics. The study also found that ACAT, MK, PMK, DPMDC and IPPI expressed at lower levels compared to HMGR1 (Figures 2D, 2I–2L). This confi rmed a previous report by Northern blot analysis (Ko et al., 2003) that expressions of MK, DPMDC and IPPI were lower compared to HMGR1, suggesting that these

Kasetsart J. (Nat. Sci.) 46(3) 357
genes supposed to be directly involved in rubber biosynthesis may be regulated at a low but not a limiting level in the laticifers. HMGR3 was significantly higher expressed in the low-yielding clones tested. However, subcellular localization prediction of three Hevea brasiliensis HMGR genes (GenBank accession no. X54659, X54658 and M74800), using PSORT programs (Nakai and Horton, 1999), indicated that HMGR3 encoded the most probable plastidic protein. Contrary to HMGR1 and HMGR2, which were much more expressed in the latex, HMGR3 was reported to be equally expressed in Hevea brasiliensis leaves and laticifers, and at a far lower level than the other two isoforms, suggesting that HMGR3 should probably not be involved in rubber synthesis (Chye et al., 1992). Thus, in the Hevea brasiliensis laticifers, the HMGR3 enzyme is most probably localized in the latex Frey-Wyssling particles (FWPs), which are known to synthesize and accumulate carotenoids and lipids, as well as a set of many other small isoprenoids (de Faÿ et al., 1989). The highly negative correlation between HMGR3 expression and the latex yield may be linked to a higher number and/or activity of FWPs in the latex of low-yielding clones. The FWPs are plastids containing latex coagulation factors, such as o-diphenol oxidases, which have been reported to play a role in the latex oxidative coagulation processes associated with the cessation of the latex fl ow (Coupé et al., 1972; Brzozowska-Hanower et al., 1978). Therefore, the high expression of HMGR3, if directly regulating the corresponding enzyme activity, might be linked to the earlier laticifers plugging, leading to a shortening of the latex fl ow and consequently to the lower latex yield. Besides playing a role in latex coagulation process, the FWPs were more recently suggested to be involved in supplying IPP for isoprenoids and eventually rubber biosynthesis through the tubular thread-like reticulum (Yoonram et al., 2008). Thus, the up-regulation of HMGR3 in low-yielding
clones may possibly indicate an increase in the global FWP activity, including the DXP/MEP pathway, which has been proposed to be involved in IPP synthesis and possibly in the intensifi cation of rubber biosynthesis (higher latex viscosity) and latex coagulation. Both activities could probably impair the latex fl ow and yield.
Isoprenoid polymerization genes expressions Genes involved in the polymerization of IPP into rubber molecules, such as PTs, REFs, and SRPPs, were abundantly expressed (high rGE levels) in all rubber clones. This confi rmed a previous report (Ko et al., 2003) that REF and SRPP were the most abundantly expressed genes in the latex from regularly tapped trees of the RRIM600 rubber clone. REF was present in large amounts at the surface of the large rubber particles in all laticifer layers (Sando et al., 2009). Contrary to a previous report (Priya et al., 2007), showing the differential expression of REF in the latex between some high- and low-yielding clones, results from the current study showed no correlation of REF expression in different yielding clones. Although REF had previously been claimed to be involved in rubber chains elongation (Dennis and Light, 1989), currently, its real role in rubber chain synthesis is rather controversial. SRPP1 expression only exhibited some negative correlation with the clonal rubber yield. SRPP1 proteins are found at the surface of the enzymatically active small rubber particles only (Rojruthai et al., 2010), especially in the young mature functional laticifers in or nearby the conducting phloem (Sando et al., 2009). Thus, it can be considered as a marker of small rubber particles. The involvement of SRPP from Hevea brasiliensis (Oh et al., 1999) or SRPP-like proteins from other rubber producing plant species such as Parthenium argentatum and Taraksakum kok, in the elongation of rubber chains has been confi rmed in vitro as well as in transformed bacterial strains (Kim et al., 2004). The higher expression of this

Kasetsart J. (Nat. Sci.) 46(3)358
gene may also indicate the higher effi ciency of the rubber biosynthesis and the higher number/longer chain of rubber particles that may cause an obstruction and impairment of the latex fl ow, thus limiting the latex yield. It is worth noting in this discussion that the expressions of most genes in the low-yielding clone CH-35-OP-2019 were similar to those in the high-yielding ones. At the same age, this low-yielding clone had a much bigger trunk size and cut length compared to the other fi ve clones (Table 1). Even though the current study did not verify the following hypothesis, it could be suggested that this clone may be characterized by faster trunk bark growth with thinner conducting phloem. This may be associated with a lower density of functional laticifers and a lower number of rings as well as with a lower number and density of feeding transversal rays per unit of bark volume. Such abnormalities can lead to a low yield especially due to anatomical and morphological reasons (Hénon and Nicolas, 1989; Garcia et al., 2002), rather than because of some disfunction in their laticifer physiology.
CONCLUSION
No signifi cant correlation between the clonal rubber yield and the DRC was found, taking into account groups of mature tapped trees from three high- and three low-yielding clones of the same age. Further, expression of most genes involved in the MVA-dependent rubber biosynthesis (acetyl CoA genesis, MVA pathway and IPP polymerization) also did not show any significant correlation with the clonal rubber yield. Within this global cytosolic rubber biosynthesis pathway, ACL1 and SRPP1 only showed a tendency to negative correlation with rubber yield traits. The very highly significant negative correlation between HMGR3 gene expression and
the clonal yield was probably not directly linked to rubber biosynthesis, since HMGR3 is most probably expressed in the FWPs (latex plastids), while this gene is not involved in the DXP/MEP pathway. All these results lead to the following non-mutually exclusive hypotheses, suggestions and conclusions: • Because of no correlation between clonal latex DRC and yield, rubber biosynthesis per se may not be the main limiting factor in the latex yield at least for the six rubber clones tested. • The effi ciency of rubber biosynthesis may not be regulated mainly at the studied genes expression level due to no signifi cant correlation between most of the rubber biosynthesis genes expression levels and clonal yield. • Assuming that the activity of the corresponding enzymes may be regulated mainly at their gene expression level, the negative correlation between ACL1 and SRPP1 expression and clonal yield may be explained by a different clonal rubber particle size distribution. Latex from the low yielding clones would be characterized by a higher number and activity of the SRPs, leading, at similar DRC, to higher latex viscosity and less latex fl ow. This hypothesis should be verifi ed by characterization of the rubber particle size distribution in the latex of the six studied clones—for example, by the laser diffraction technique (Kongsawadworakul and Chrestin, 2003)—in parallel with the characterization of their latex initial fl ow rate and duration. • In the current study, only the genes involved in the MVA-dependent rubber biosynthesis were taken into consideration, since up to now, this has been the only single pathway that has been proven (through the use of isotope-labeled precursors) to be functional in the Hevea brasiliensis laticifer rubber biosynthesis (Skilleter and Kekwick, 1971). However, incorporation of IPP derived from the DXP/MEP pathway into natural

Kasetsart J. (Nat. Sci.) 46(3) 359
rubber cannot be excluded. Indeed, crossover of both pathways under certain conditions (for example, depletion of the MVA pathway derived-IPP to fulfi l the cytosolic IPP demand, which may be the case for very high yielding rubber trees) could be demonstrated in Arabidopsis thaliana and tobacco Bright Yellow-2 cells (Kasahara et al., 2002; Hemmerlin et al., 2003), and has been recently proposed for Hevea brasiliensis (Chow et al., 2012). Expression of plastidic DXP/MEP pathway genes in the latex in relation to the rubber clonal yield will be the next matter for study. • Up-regulation of the plastidic HMGR3 may indicate a higher number of FWPs or their higher activity in the latex of low-yielding clones or both. This may be not in agreement with the above hypothesis of the involvement of the plastidic DXP/MEP pathway in rubber synthesis in high-yielding clones. On the contrary, the higher number or activity of FWPs may, through probable higher phenol oxidase activity (Brzozowska-Hanower et al., 1978), contribute to faster latex coagulation and lower yield in low-yielding clones. This should be verifi ed by the measurement of the total latex pellet phenol oxidase activity and the carotenoids content in the latex of the low- and high-yielding clones.
ACKNOWLEDGEMENT
The authors would like to express their warm thanks to the Surat Thani Rubber Research Center, Department of Agriculture, Thailand, for providing planting material, the Department of Plant Science, Faculty of Science, Mahidol University, Thailand for providing laboratory facilities and the Institut de Recherche pour le Développement (IRD) and the Institut Français du Caoutchouc (IFC), France for providing chemicals. This research was also supported by the Center for Agricultural Biotechnology, Kasetsart University, Kamphaeng Saen Campus, Nakhon Pathom and the Center of Excellence on
Agricultural Biotechnology (AG-BIO/PERDO-CHE).
LITERATURE CITED
Archer, B.L. and B.G. Audley. 1967. Biosynthesis of rubber. Adv. Enzymol. Relat. Areas. Mol. Biol. 29: 221–257.
d’Auzac, J., J.L. Jacob and H. Chrestin. 1989. Physiology of the latex fl ow, pp. 219–285. In J. d'Auzac, J.L. Jacob and H. Chrestin, (eds). Physiology of Rubber Tree Latex. CRC Press Inc. Boca Raton, FL, USA.
Blackley, D.C. 1997. Polymer Latices: Science and Technology Types of Lattices. 2nd ed. Chapman & Hall. London, UK. 612 pp.
Brzozowska-Hanower, J., P. Hanower and C. Lioret. 1978. Étude du mècanisme de la coagulation du latex d'Hevea brasiliensis (Kunth) Müll. Arg. II. systémes enzymatiques impliqués dans le processus. Physiol. Vég. 16: 231–254. [in French]
Chow, K.S., M.N. Mat-Isa, A. Bahari, A.K. Ghazali, H. Alias, Z. Mohd-Zainuddin, C.C. Hoh and K.L. Wan. 2012. Metabolic routes affecting rubber biosynthesis in Hevea brasiliensis latex. J. Exp. Bot. 63(5): 1863–1871.
Chrestin, H. and J. Bangratz. 1988. The adenine nucleotides pool and energy charge in the Hevea latex cells: Clonal characteristics, relations with yield and sensitivity towards bark-dryness. Effects of yield stimulation with Ethrel, an ethylene releaser, pp. 547–562. In Proceedings of International Conference of Hevea Phisiology and Breeding. October 1988. Paris, France.
Chu, F., J. Guillot and A. Guyot. 1998. Rheology of concentrated multi-sized poly (St/BA/MMA) latexes. Colloid Polym. Sci. 276: 305–312.
C h u e n w a r i n , P. , U . Vi b o o n j u n , P. Kongsawadworakul, H. Chrestin and J. Narangajavana. 2010. Expressions of genes

Kasetsart J. (Nat. Sci.) 46(3)360
involved in isoprene biosynthesis pathways in the latex from Hevea brasiliensis high and low yielding trees. In Proceedings of the 16th Asian Agricultural Symposium and the 1st International Symposium on Agricultural Technology on Sufficiency Agriculture. King Mongkut’s Institute of Technology. Ladkrabang, Bangkok, Thailand.
Chye, M.L., C.T. Tan and N.H. Chua. 1992. Three genes encode 3-hydroxy-3-methylglutaryl-coenzyme A reductase in Hevea brasiliensis: hmg1 and hmg3 are differentially expressed. Plant Mol. Biol. 19: 473–484.
Cornish, K. and J.L. Brichta. 2002. Some rheological properties of latex from Parthenium argentatum Gray compared with latex from Hevea brasiliensis and Ficus elastica. J. Polym. Environ. 10: 13–18.
Coupé, M., S. Pujarniscle and J. d'Auzac. 1972. Compartmentation de diverses oxydo-réductases (peroxydase, o-diphenol-oxydase et malate déshydrogénase) dans le latex d' Hevea brasiliensis (Kunth). Müll Arg. Physiol. Vég. 10: 459–464. [in French]
Dennis, M.S. and D.R. Light. 1989. Rubber elongation factor from Hevea brasiliensis. J. Biol. Chem. 264: 18608–18617.
Eschbach, J.M., J. Tupy and R. Lacrotte. 1986. Photosynthate allocation and productivity of latex vessels in Hevea brasiliensis. Bio. Plantarum 28: 321–328.
Fatland, B.L., B.J. Nikolau and E.S. Wurtele. 2005. Reverse genetic characterization of cytosolic acetyl CoA generation by ATP-citrate lyase in Arabidopsis. Plant Cell 17: 182–203.
de Faÿ, E., C.H. Hébant and J.L. Jacob. 1989. Cytology and cytochemistry of the laticiferous system, pp.15–27. In J. d'Auzac, J.L. Jacob and H. Chrestin, (eds). Physiology of Rubber Tree Latex. CRC Press Inc. Boca Raton, FL, USA.
Garcia, D., V. Le Guen, C.R. Reis Mattos, P. de
Souza Conçalves and A. Clément-Demange. 2002. Relationships between yield and some structural traits of the laticiferous system in Hevea clones resistant to South American leaf blight. Crop Breed Appl. Biotech. 2(2): 307–318.
Gidrol, X., H. Chrestin, H.L. Ta and A. Kush. 1994. Hevein, a lectin-like protein from Hevea brasiliensis (rubber tree) is involved in the coagulation of latex. J. Biol. Chem. 269: 9278–9283.
Greenwood, R., P.F. Luckham and T. Gregory. 1995. The effect of particle size on the layer thickness of a stabilizing polymer adsorbed onto two different classes of polymer latex, as determined from rheological experiments. Colloid Surface 98: 117–125.
Hanning, I., K. Baumgarten, K. Schott and H.W. Heldt. 1999. Oxaloacetate transport into plant mitochondria. Plant Physiol. 119(3): 1025–1031.
Hemmerlin, A., J-F. Hoeffler, O. Meyer, D. Tritsch, I. Kagan, C. Grosdemange-Billiard, M. Rohmer and T.J. Bach. 2003. Cross-talk between the cytosolic mevalonate and the plastidial methylerythritol phosphate pathways in tobacco BY-2 cells. J. Biol. Chem. 278: 26666–26676.
Hénon, J.M. and D. Nicolas. 1989. Relation between anatonomical characteritics of the laticifer system and latex yield: Search for early selection criteria, pp. 31–55. In J. d'Auzac, J.L. Jacob and H. Chrestin, (eds.). Physiology of Rubber Tree Latex. CRC Press Inc. Boca Raton, FL, USA.
Jacob, J.L., J.C. Prévôt, D. Roussel, R. Lacrotte, E. Serres, J. d’Auzac, J.M. Eschbach and H. Omont. 1989. Yield-limiting factors, latex physiological parameters, latex diagnosis and clonal typology, pp. 345–382. In J. d'Auzac, J.L. Jacob and H. Chrestin, (eds.). Physiology of Rubber Tree Latex. CRC Press Inc. Boca Raton, FL, USA.

Kasetsart J. (Nat. Sci.) 46(3) 361
Kasahara, H., A. Hanada, T. Kuzuyama, M. Takagi, Y. Kamiya and S. Yamaguchi. 2002. Contribution of the mevalonate and methylerythritol phosphate pathways to the biosynthesis of gibberellins in Arabidopsis. J. Biol. Chem. 277: 45188–45194.
Kim, J., S.B. Ryu, Y.S. Kwak and H. Kang. 2004. A novel cDNA from Parthenium argentatum Gray enhances the rubber biosynthetic activity in vitro. J. Exp. Bot. 55: 377–385.
Ko, J-H., K-S. Chow and K-H. Han. 2003. Transcriptome analysis reveals novel features of the molecular events occurring in the laticifers of Hevea brasiliensis (para rubber tree). Plant Mol. Biol. 53(4): 479–492.
Kongsawadworakul, P. and H. Chrestin. 2003. Laser diffraction: A new tool for identifi cation and studies of physiological effectors involved in aggregation/coagulation of the rubber particles from Hevea latex. Plant Cell Physiol. 44: 707–717.
Lichtenthaler, H.K. 1999. The 1-deoxy-D-xylulose-5-phosphate pathway of isoprenoid biosynthesis in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 50: 47–65.
Livak, K.J. and T.D. Schmittgen. 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 25: 402–408.
Nakai, K. and P. Horton. 1999. PSORT: A program for detecting sorting signals in proteins and predicting their subcellular localization. Trends Biochem. Sci. 24(1): 34–36.
Oh, S.K., H. Kang, D.H. Shin, J. Yang, K.S. Chow, H.Y. Yeang, B. Wagner, H. Breiteneder and K.H. Han. 1999. Isolation, characterization and functional analysis of a novel cDNA clone encoding a small rubber particle protein from Hevea brasiliensis. J. Biol. Chem. 274: 17132–17138.
Oliver, D.J., B.J. Nikolau and E.S. Wurtele. 2009. Acetyl CoA - Life at the metabolic nexus. Plant Sci. 176: 597–601.
Paardekooper, E.C. and S. Sookmark. 1969. Diurnal variations in latex yield and dry rubber content and relation to the saturation deficit of the air. J. Rubber Res. Inst. Malaya. 21: 341–347.
Perham, R.N. and L.C. Packman. 1989. 2-Oxo acid dehydrogenase multienzyme complexes: domains, dynamics and design. Ann. N. Y. Acad. Sci. 573:1–20.
Priya, P., P. Venkatachalam and A. Thulaseedharan. 2007. Differential expression pattern of rubber elongation factor (REF) mRNA transcripts from high and low yielding clones of rubber tree (Hevea brasiliensis Muell. Arg.). Plant Cell Rep. 26: 1833–1838.
Pujade-Renaud, V., A. Clement, C. Perrot-Rechenmann, J.C. Prévôt, H. Chrestin, J.L. Jacob and J. Guern. 1994. Ethylene-induced increase in glutamine synthetase activity and mRNA levels in Hevea brasiliensis latex cells. Plant Physiol. 105: 127–132.
Rohmer, M., M. Seemann, S. Horbach, S. Bringer-Meyer and H. Sahms. 1996. Glyceraldehyde-3-phosphate and pyruvate as precursors of isoprenic units in an alternative non-mevalonate pathway for terpenoid biosynthesis. J. Am. Chem. Soc. 118(1): 2564–2566.
Rojruthai, P., J.T. Sakdapipanich, S. Takahashi, L. Hyegin, M. Noike, T. Koyama and Y. Tanaka. 2010. In vitro synthesis of high molecular weight rubber by Hevea small rubber particles. J. Biosci. Bioeng. 109(2): 107–114.
Sando, T., T. Hayashi, T. Takeda, Y. Akiyama, Y. Nakazawa, E. Fukusaki and A. Kobayashi. 2009. Histochemical study of detailed laticifer structure and rubber biosynthesis-related protein localization in Hevea brasiliensis using spectral confocal laser scanning microscopy. Planta 230: 215–225.
Sangsing, K., R. Danskulphol, A. Rojsujitr and S. Suapan. 1998. Further proof clone trial. Annual Report. Vol. 2. Rubber Research Institute of Thailand. Bangkok, Thailand.

Kasetsart J. (Nat. Sci.) 46(3)362
Skilleter, D.N. and R.G.O. Kekwick. 1971. The enzymes forming isopentenyl pyrophosphate from 5-phosphomevalonate (mevalonate 5-phosphate) in the latex of Hevea brasiliensis. Biochem. J. 124: 407–417.
Tungngoen, K., P. Kongsawadworakul, U. Viboonjun, M. Katsuhara, N. Brunel, S. Sakr, J. Narangajavana and H. Chrestin. 2009. Involvement of HbPIP2;1 and HbTIP1;1 aquaporins in ethylene stimulation of latex yield through regulation of water exchanges between inner liber and latex cells in Hevea brasiliensis. Plant Physiol. 151: 843–856.
Van Gils, G.E. 1951. Studies on the latex viscosity. I: infl uence of the dry rubber content. Arch. Rubbercult. 28: 61–66.
Vinod, K.K., J. Pothen, D. Chaudhuri, P.M. Priyadarshan, T. Eappen, M. Varghese, D. Mandal, A.C. Sharma, T. Kumar Pal, A.S. Devakumar and A.K. Krisnakumar. 2000. Variation and trend of yield and related traits of Hevea brasiliensis Müell. Agr. in Tripura. Indian J. Nat. Rubber Res. 13(1-2): 69–78.
Wititsuwannakul, R., P. Pasitkul, K. Kanokwiroon and D.Wititsuwannakul. 2008. A role for a Hevea latex lectin-like protein in mediating rubber particle aggregation and latex coagulation. Phytochemistry 69(2): 339–347.
Yoonram, K., S. Takahashi, A. Rattanapittayaporn, T. Koyama, D. Wititsuwannakul and R. Wititsuwannakul. 2008. cDNA, from Hevea brasiliensis latex, encoding 1-deoxy-D-xylulose-5-phosphate reductoisomerase. Plant Sci. 175: 694–700.
Yuan, J.S., A. Reed, F. Chen and C.N. Stewart. 2006. Statistical analysis of real-time PCR data. BMC Bioinformatics 7: 85. doi:10.1186/1471-2105-7-85.
Related Documents











