Mitochondrial D-Loop instability in thyroid tumours is not a marker of malignancy Valdemar Ma ´ximo a , Jorge Lima a , Paula Soares a,b , Tiago Botelho a , Lı ´gia Gomes a , Manuel Sobrinho-Simo ˜es a,b,c, * a Institute of Molecular Pathology and Immunology of the University of Porto (IPATIMUP), Porto, Portugal b Department of Pathology, Medical Faculty of the University of Porto, Porto, Portugal c Department of Pathology, Hospital Sa ˜o Joa ˜o, Porto, Portugal Received 4 February 2005; received in revised form 20 May 2005; accepted 1 June 2005 Abstract Despite the numerous studies describing a high frequency of mitochondrial DNA (mtDNA) somatic mutations in many types of human primary tumors the mechanisms that generate such mutations and the role of mtDNA mutations in tumor development remain unclear. We present the results obtained in the study of mtDNA displacement-loop (D-Loop) region in a series of 66 thyroid tumors, and respective adjacent parenchyma, including benign (adenomas, nZ30) and malignant tumors (follicular carcinomas, nZ17 and papillary carcinomas, nZ19). Three repetitive regions were analyzed [two mononucleotide repetitive (D310 and D568) and one dinucleotide repetitive (D514)]. Thirty-two (48.5%) of the 66 tumors [15/30 (50.0%) adenomas, 8/17 (47.1%) follicular carcinomas and 9/19 (47.4%) papillary carcinomas] harbored somatic insertions in D-Loop repetitive regions. Twenty (30.3%) of the 66 tumors [12/30 (40%) adenomas, 3/17 (17.6%) follicular carcinomas and 5/19 (26.3%) papillary carcinomas] harbored somatic insertions at the D310 mononucleotide repeat. Three (4.6%) of the 66 tumors [1/30 (3.3%) adenomas and 2/17 (11.8%) follicular carcinomas] harbored somatic insertions at the D568 mononucleotide repeat. Fifteen (22.7%) of the 66 tumors [3/30 (10.0%) adenomas, 5/17 (29.4%) follicular carcinomas and 7/19 (36.8%) papillary carcinomas] harbored somatic insertions at the D514 dinucleotide repeat. Five (7.6%) of the 66 tumors [1/30 (3.3%) adenomas, 1/17 (5.9%) follicular carcinomas and 2/19 (10.5%) papillary carcinomas] harbored somatic insertions in more than one region, and in one of them (a carcinoma) alterations were detected in the three regions. We conclude that mutations in the mtDNA D-Loop region are frequent in benign and malignant thyroid tumors and cannot be considered a marker of malignancy. Our study shows, furthermore, two repetitive regions (D310 and D514) that appear to be susceptible to mutation in thyroid tumors. q 2005 Published by Elsevier B.V. on behalf of Mitochondria Research Society. Keywords: Thyroid tumors; mtDNA; D-Loop instability; Hu ¨rthle cell tumors; D310 1. Introduction The human mtDNA is a 16-kb circular, double stranded DNA that encodes 13 polypeptides of the mitochondrial respiratory chain (MRC), 22 transfer Mitochondrion xx (2005) 1–8 www.elsevier.com/locate/mito 1567-7249/$ - see front matter q 2005 Published by Elsevier B.V. on behalf of Mitochondria Research Society. doi:10.1016/j.mito.2005.06.003 * Corresponding author. Address: IPATIMUP, Rua Dr Roberto Frias s/n, 4200-465 Porto, Portugal. Tel.: C351 22 55 70 700; fax: C351 22 55 70 799. E-mail address: [email protected] (M. Sobrinho-Simo ˜es). DTD 5 ARTICLE IN PRESS

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

DTD 5 ARTICLE IN PRESS
Mitochondrial D-Loop instability in thyroid tumours
is not a marker of malignancy
Valdemar Maximoa, Jorge Limaa, Paula Soaresa,b, Tiago Botelhoa,
Lıgia Gomesa, Manuel Sobrinho-Simoesa,b,c,*
aInstitute of Molecular Pathology and Immunology of the University of Porto (IPATIMUP), Porto, PortugalbDepartment of Pathology, Medical Faculty of the University of Porto, Porto, Portugal
cDepartment of Pathology, Hospital Sao Joao, Porto, Portugal
Received 4 February 2005; received in revised form 20 May 2005; accepted 1 June 2005
Abstract
Despite the numerous studies describing a high frequency of mitochondrial DNA (mtDNA) somatic mutations in many types
of human primary tumors the mechanisms that generate such mutations and the role of mtDNA mutations in tumor development
remain unclear. We present the results obtained in the study of mtDNA displacement-loop (D-Loop) region in a series of 66
thyroid tumors, and respective adjacent parenchyma, including benign (adenomas, nZ30) and malignant tumors (follicular
carcinomas, nZ17 and papillary carcinomas, nZ19). Three repetitive regions were analyzed [two mononucleotide repetitive
(D310 and D568) and one dinucleotide repetitive (D514)]. Thirty-two (48.5%) of the 66 tumors [15/30 (50.0%) adenomas, 8/17
(47.1%) follicular carcinomas and 9/19 (47.4%) papillary carcinomas] harbored somatic insertions in D-Loop repetitive
regions. Twenty (30.3%) of the 66 tumors [12/30 (40%) adenomas, 3/17 (17.6%) follicular carcinomas and 5/19 (26.3%)
papillary carcinomas] harbored somatic insertions at the D310 mononucleotide repeat. Three (4.6%) of the 66 tumors [1/30
(3.3%) adenomas and 2/17 (11.8%) follicular carcinomas] harbored somatic insertions at the D568 mononucleotide repeat.
Fifteen (22.7%) of the 66 tumors [3/30 (10.0%) adenomas, 5/17 (29.4%) follicular carcinomas and 7/19 (36.8%) papillary
carcinomas] harbored somatic insertions at the D514 dinucleotide repeat. Five (7.6%) of the 66 tumors [1/30 (3.3%) adenomas,
1/17 (5.9%) follicular carcinomas and 2/19 (10.5%) papillary carcinomas] harbored somatic insertions in more than one region,
and in one of them (a carcinoma) alterations were detected in the three regions. We conclude that mutations in the mtDNA
D-Loop region are frequent in benign and malignant thyroid tumors and cannot be considered a marker of malignancy. Our
study shows, furthermore, two repetitive regions (D310 and D514) that appear to be susceptible to mutation in thyroid tumors.
q 2005 Published by Elsevier B.V. on behalf of Mitochondria Research Society.
Keywords: Thyroid tumors; mtDNA; D-Loop instability; Hurthle cell tumors; D310
1567-7249/$ - see front matter q 2005 Published by Elsevier B.V. on be
doi:10.1016/j.mito.2005.06.003
* Corresponding author. Address: IPATIMUP, Rua Dr Roberto
Frias s/n, 4200-465 Porto, Portugal. Tel.: C351 22 55 70 700; fax:
C351 22 55 70 799.
E-mail address: [email protected] (M. Sobrinho-Simoes).
1. Introduction
The human mtDNA is a 16-kb circular, double
stranded DNA that encodes 13 polypeptides of the
mitochondrial respiratory chain (MRC), 22 transfer
Mitochondrion xx (2005) 1–8
www.elsevier.com/locate/mito
half of Mitochondria Research Society.

V. Maximo et al. / Mitochondrion xx (2005) 1–82
DTD 5 ARTICLE IN PRESS
RNAs and 2 ribosomal RNAs required for protein
synthesis (Anderson et al., 1981; Andrews et al.,
1999). The mutation rate of mtDNA is about 10!20
times higher than that of the nuclear DNA (nDNA)
(Wallace et al., 1998; Brown and Wallace 1994). This
high mutation rate may be due to one or more of the
following factors: a high concentration of oxygen
radicals at the mitochondrial inner membrane, the
lack of efficient mtDNA repair mechanisms, and/or
the absence of DNA-coating proteins such as the
histones (Anderson et al., 1981; Wallace et al., 1998).
Due to this high mutation rate, mtDNA is likely a
hotspot for mutations in cancer as it is preferentially
modified by many carcinogens (Backer et al., 1980).
Alterations of mtDNA have been demonstrated in
various types of primary human cancers and include
large deletions, missense mutations, frameshift
mutations and small insertions/deletions in repetitive
regions (Sanchez-Cespedes et al., 2001; Maximo et
al., 2001, 2002; Habano et al., 1998, 2000; Richard et
al., 2000; Burgart et al., 1995; Polyak et al., 1998;
Fliss et al., 2000; Yeh et al., 2000; Lewis et al., 2000).
Despite the high number of studies on mtDNA
mutations in human primary neoplasms, the mechan-
isms that generate such mutations and the putative
role played by mtDNA mutations in tumor develop-
ment remain unclear.
In a previous study, Sanchez-Cespedes et al.
(2001) reported the identification of a specific and
highly polymorphic homopolymeric C stretch (D310),
located within the displacement-Loop (D-Loop) of the
mtDNA, as a mutational hotspot in human benign
and malignant primary tumors. Moreover, these
alterations were also present in 2 out of 14
preneoplastic lesions from head and neck tumors
(Sanchez-Cespedes et al., 2001). Most of the somatic
alterations found were deletion/insertions generating
D310 variants identical to constitutive polymorph-
isms described previously (Sanchez-Cespedes et al.,
2001; MITOMAP: A Human Mitochondrial Genome
Database, 2001).
In an attempt to clarify the putative pathogenic
meaning of somatic mitochondrial D-Loop instability
in thyroid tumors, we have studied three repetitive
regions of the D-Loop region in a series of 66 benign
and malignant thyroid tumors and adjacent thyroid
parenchyma by PCR/SSCP and automated
sequencing.
2. Material and methods
2.1. Material
Sixty-six thyroid tumors from 59 patients were
studied. In 7 patients there were two distinct lesions
that were separately studied. The 66 lesions were
classified according to Hedinger et al. (1988) and
Rosai et al. (1992) as follicular adenoma (nZ10),
Hurthle cell follicular adenoma (nZ20), follicular
carcinoma (nZ4), Hurthle cell follicular carcinoma
Polyak et al. (1998), papillary carcinoma (nZ12) and
Hurthle cell papillary carcinoma (nZ7). Samples
from 27 lesions were obtained at the time of surgery,
together with the corresponding normal adjacent
tissues; these samples were carefully dissected by
expert pathologists and snap frozen. In the remaining
33 cases, microdissected paraffin-embedded material
was used for the screening of mtDNA D-Loop
mutations, due to the absence of representative
tumor tissue in the frozen samples.
2.2. DNA extraction
DNA was extracted from microdissected frozen
and/or paraffin-embedded pathologic and normal
thyroid tissue pairs using the NucleoSpinw Tissue
Kit (Macherey-Nagel, Duren, Germany).
2.3. Screening of mtDNA D-Loop mutations
The screening of mtDNA D-Loop mutations was
performed by PCR/SSCP and automated sequencing.
Two independent fragments of 157 bp (between nt
267 and 463) and 185 bp (between nt 453 and 637),
using the following primers: the forward primer used
for the 157 bp fragment amplification was 5 0-
TCCACACAGACATCATAACA-3 0 and the reverse
was 5 0-AAAGTGCATACCGCCAAAAG-3 0; the for-
ward primer used for the 185 bp fragment amplifi-
cation was 5 0-TTTCCCCTCCCACTCCCATACT-3 0
and the reverse was 5 0-GTGATGTGAGCCCGTC-
TAAACA-3 0. PCR amplifications were performed in
a 25 ml volume containing 200 mM of each dNTP,
12.5 pmol of each of the forward and reverse primers,
50 mM KCl, 10 mM Tris–HCl (pH 9.0), 1.5 mM
MgCl2 and 1 U of Taq DNA polymerase (Amersham
Pharmacia Biotech). Cycling conditions were a single
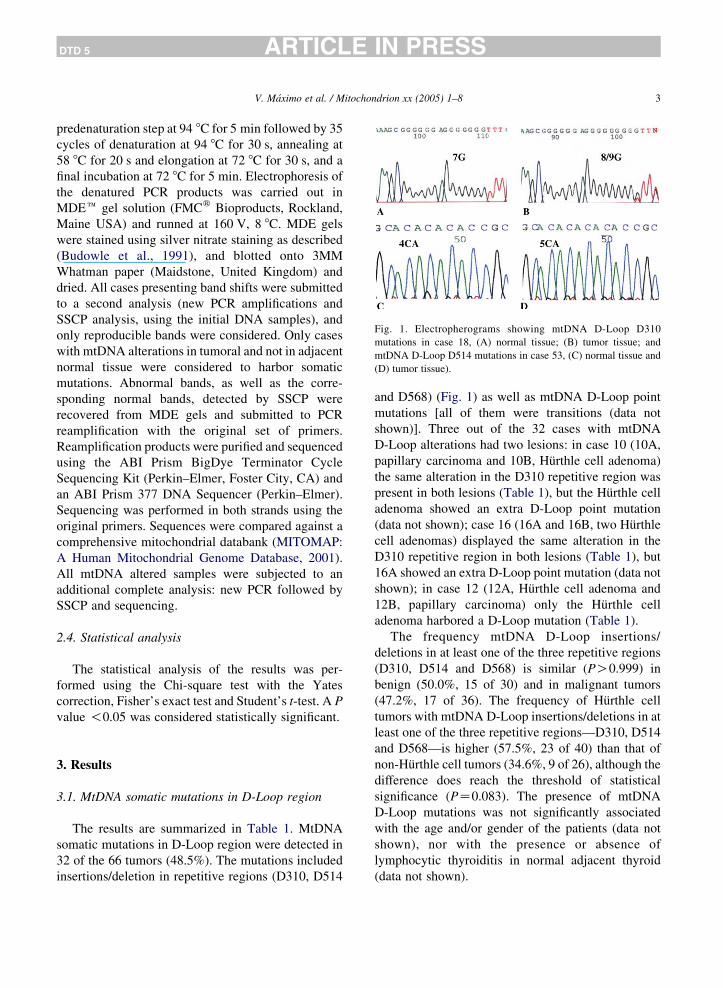
Fig. 1. Electropherograms showing mtDNA D-Loop D310
mutations in case 18, (A) normal tissue; (B) tumor tissue; and
mtDNA D-Loop D514 mutations in case 53, (C) normal tissue and
(D) tumor tissue).
V. Maximo et al. / Mitochondrion xx (2005) 1–8 3
DTD 5 ARTICLE IN PRESS
predenaturation step at 94 8C for 5 min followed by 35
cycles of denaturation at 94 8C for 30 s, annealing at
58 8C for 20 s and elongation at 72 8C for 30 s, and a
final incubation at 72 8C for 5 min. Electrophoresis of
the denatured PCR products was carried out in
MDEe gel solution (FMCw Bioproducts, Rockland,
Maine USA) and runned at 160 V, 8 8C. MDE gels
were stained using silver nitrate staining as described
(Budowle et al., 1991), and blotted onto 3MM
Whatman paper (Maidstone, United Kingdom) and
dried. All cases presenting band shifts were submitted
to a second analysis (new PCR amplifications and
SSCP analysis, using the initial DNA samples), and
only reproducible bands were considered. Only cases
with mtDNA alterations in tumoral and not in adjacent
normal tissue were considered to harbor somatic
mutations. Abnormal bands, as well as the corre-
sponding normal bands, detected by SSCP were
recovered from MDE gels and submitted to PCR
reamplification with the original set of primers.
Reamplification products were purified and sequenced
using the ABI Prism BigDye Terminator Cycle
Sequencing Kit (Perkin–Elmer, Foster City, CA) and
an ABI Prism 377 DNA Sequencer (Perkin–Elmer).
Sequencing was performed in both strands using the
original primers. Sequences were compared against a
comprehensive mitochondrial databank (MITOMAP:
A Human Mitochondrial Genome Database, 2001).
All mtDNA altered samples were subjected to an
additional complete analysis: new PCR followed by
SSCP and sequencing.
2.4. Statistical analysis
The statistical analysis of the results was per-
formed using the Chi-square test with the Yates
correction, Fisher’s exact test and Student’s t-test. A P
value !0.05 was considered statistically significant.
3. Results
3.1. MtDNA somatic mutations in D-Loop region
The results are summarized in Table 1. MtDNA
somatic mutations in D-Loop region were detected in
32 of the 66 tumors (48.5%). The mutations included
insertions/deletion in repetitive regions (D310, D514
and D568) (Fig. 1) as well as mtDNA D-Loop point
mutations [all of them were transitions (data not
shown)]. Three out of the 32 cases with mtDNA
D-Loop alterations had two lesions: in case 10 (10A,
papillary carcinoma and 10B, Hurthle cell adenoma)
the same alteration in the D310 repetitive region was
present in both lesions (Table 1), but the Hurthle cell
adenoma showed an extra D-Loop point mutation
(data not shown); case 16 (16A and 16B, two Hurthle
cell adenomas) displayed the same alteration in the
D310 repetitive region in both lesions (Table 1), but
16A showed an extra D-Loop point mutation (data not
shown); in case 12 (12A, Hurthle cell adenoma and
12B, papillary carcinoma) only the Hurthle cell
adenoma harbored a D-Loop mutation (Table 1).
The frequency mtDNA D-Loop insertions/
deletions in at least one of the three repetitive regions
(D310, D514 and D568) is similar (PO0.999) in
benign (50.0%, 15 of 30) and in malignant tumors
(47.2%, 17 of 36). The frequency of Hurthle cell
tumors with mtDNA D-Loop insertions/deletions in at
least one of the three repetitive regions—D310, D514
and D568—is higher (57.5%, 23 of 40) than that of
non-Hurthle cell tumors (34.6%, 9 of 26), although the
difference does reach the threshold of statistical
significance (PZ0.083). The presence of mtDNA
D-Loop mutations was not significantly associated
with the age and/or gender of the patients (data not
shown), nor with the presence or absence of
lymphocytic thyroiditis in normal adjacent thyroid
(data not shown).

Table 1
Summary of mtDNA D-Loop alterations in 66 thyroid tumors
Case Age Diagnosis mtDNA D-Loop alterations
D310 D514 D568
1 57 PC – CA7/CA4 –
2 51 HCFC C6/C6,7 – –
3 32 HCFC C6/C8,9 CA5/CA6 C6/C8
4 39 HCFC – – C6/C8–10
5 33 HCFC – CA5/CA7 –
6 76 HCFC – CA5/CA6 –
7 62 HCPC C7/C8 CA5/CA6 –
9 49 HCA C6/C8 – –
10A 63 PC C7/C8 – –
10B 63 HCA C7/C8 – –
12A 45 HCA C6/C8 – –
14 34 HCA C7/C9,10 – –
15 59 HCA C7/C8 – –
16A 67 HCA C7/C8 – –
16B 67 HCA C7/C8,9 – –
17 57 HCA C7/C8 – –
18 38 HCA C7/C8,9 – C6/C7
19 54 HCFC – CA5/CA6 –
21 8 HCPC – CA4/CA4–6 –
22 46 HCPC C7/C8 CA5/CA6 –
27 48 HCFC – CA5/CA6 –
28 42 A C7/C9,10 – –
32 32 HCPC C7/C9,10 – –
39 53 PC – CA5/CA8 –
40 77 HCA – CA5/CA6,7 –
41 40 FC C7/C8–10 – –
46 67 A – CA7/CA6 –
51 54 PC C7/C8–12 CA5/CA6,7 –
53 37 A – CA4/CA5 –
54 56 HCA C7/C8 – –
57 34 HCA C7/C8,9 – –
60 56 PC – CA5/CA6 –
V. Maximo et al. / Mitochondrion xx (2005) 1–84
DTD 5 ARTICLE IN PRESS
3.2. D310
Twenty (30.3%) of the 66 tumors [12/30 (40%)
adenomas, 5/19 (26.3%) papillary carcinomas and
3/17 (17.6%) follicular carcinomas] harbored somatic
insertions at the polymorphic homopolymeric C
stretch (D310). The mutations were insertions that
varied from 1 to 5 bp (Tables 1 and 2). The prevalence
of D310 alterations was higher, though not statisti-
cally significant (PZ0.178), in adenomas (40%, 12/
30) than in carcinomas (22.2%, 8/36; follicular
carcinoma K17.6%, 3 of 17: papillary carcinoma
K26.3%, 5 of 19). There were 11 tumors with 2 bp or
more insertions (7 adenomas, 2 follicular carcinomas
and 2 papillary carcinomas) (Tables 1 and 2). Five out
the 20 (25.0%) D310 alterations were outside the
usual polymorphic length range (between 7C and 9C)
and included unusual variants (10C, 11C and 12C).
Four of the 59 (6.1%) patients presented constitu-
tionally a deletion of 1 bp (6C).
3.3. D514
Somatic mutations in the dinucleotide (CA) repeat
region D514 were detected in 15 cases (22.7%). The
prevalence of mutations was significantly lower
(PZ0.038) in adenomas (10.0%, 3/30) than in
carcinomas (33.3%, 12/36; follicular carcinoma
K29.4%; 5 of 17: papillary carcinoma K36.8%; 7
of 19). The type of alterations (number of CA repeats)

Table 2
Somatic insertions (1-bp or higher) in the D310 repeat in 66 thyroid tumors
Histotype No. of alterations (total) 1 bp O1 bp
Adenoma (nZ30) 12 (40.0%) 5 (16.7%) 7 (23.3%)
Carcinoma (nZ36) 8 (22.2%) 4 (11.1%) 4 (11.1%)
Follicular carcinoma (nZ17) 3 (17.6%) 1 (5.8%) 2 (11.8%)
Papillary carcinoma (nZ19) 5 (26.3%) 3 (15.8%) 2 (10.5%)
PZ0.178 (Adenomas vs. carcinomas)
V. Maximo et al. / Mitochondrion xx (2005) 1–8 5
DTD 5 ARTICLE IN PRESS
in the three groups of tumors is summarized in
Tables 1 and 3. The alterations we have found vary
from deletion of 3 CA to addition of 3 CA. Two of the
59 patients presented constitutionally an insertion of
4 bp (2CA) and another two presented constitutionally
a deletion of 2 bp (1CA).
3.4. D568
There were alterations in the mononucleotide (C)
repeat region D568 in 3 out of the 66 (4.5%): one
adenoma and two follicular carcinomas. The adenoma
showed an insertion of 2 bp (2C), one follicular
carcinoma showed a mixture of mtDNA molecules
with a range varying from 8C to 10C in the D568
stretch—addition of 2 bp to 4 bp (C) and the other
follicular carcinoma presented an insertion of 1 bp
(1C) (Table 1). All the 59 patients presented
constitutionally 6C in the D568 region.
3.5. D310 vs. D514 vs. D568
The frequency of alterations/instability was higher
in D310 (30.3%, 20/66) and D514 (22.7%, 15/66) than
in D568 (4.6%, 3/66) repetitive region. Alteration-
s/instability in at least one of the 3 repetitive regions
(D310, D514 and D568) were found in 32/66 (48.5%)
tumors [15/30 (50.0%) adenomas; 9/19 (47.4%)
papillary carcinomas and 8/17 (47.1%) follicular
carcinomas]. Five of the 20 cases with instability in
Table 3
Somatic insertions/deletions (1-CA repeat or higher) in the dinucleotide D
Histotype No. of alterations (total) 1
Adenoma (nZ30) 3 (10.0%) 2
Carcinoma (nZ36) 12 (33.3%) 7
Follicular carcinoma (nZ17) 5 (29.4%) 4
Papillary carcinoma (nZ19) 7 (36.8%) 3
PZ0.038 (Adenomas vs. carcinomas)
D310 region harbored also instability in one (four
cases) or both (one case) of the other repetitive
regions (D514 and D568): four cases presented
alterations in D310 and D514 regions and two cases
presented alterations in D310 and D568 regions; one
follicular carcinoma had alterations in the 3 repetitive
regions (Table 1).
4. Discussion
We detected mtDNA D-Loop somatic mutations in
almost fifty percent of the tumors, both benign and
malignant, thus supporting the assumption that these
alterations are early and common events in thyroid
carcinogenesis.
The mutations detected in our series were either
point mutation (transitions) which are the type of
mutations that may be expected as a consequence of
ROS damage (Backer et al., 1980; Beckman and
Ames, 1997; Cadet et al., 1997) or variations of
repetitive elements in monorepetitive (C) and
direpetitive (CA) D-Loop regions. Most of the
alterations we detected in D310, D514 and D568
regions were in the polymorphic length range
described previously (MITOMAP: A Human Mito-
chondrial Genome Database, 2001; Hauswirth and
Clayton, 1985; Cavelier et al., 2000; Wallace and
Mitochondrial, 1994), thus suggesting that most
mtDNA D-Loop variations detected by us in thyroid
514 repeat region in 66 thyroid tumors
CA repeat O1CA repeat
(6.7%) 1 (3.3%)
(19.4%) 5 (13.9%)
(23.5%) 1 (5.9%)
(15.8%) 4 (21.1%)

V. Maximo et al. / Mitochondrion xx (2005) 1–86
DTD 5 ARTICLE IN PRESS
tumors are unlikely to lead to functional impairment
of the mitochondria. Such alterations result probably
from the high ROS production in tumor cells and/or
from the high replication rate of the neoplastic cells.
Five of the 20 D310 alterations were outside the
usual polymorphic length range (between 7C and 9C)
and included unusual variants (10C, 11C and 12C).
Some of the observed D310 variants harbor large
C-tract insertions that are likely to interfere with the
initiation of mtDNA replication (Xu and Clayton,
1995; Lee and Clayton, 1998; Hauswirth and Clayton,
1985). These findings suggest that, at variance with
the aforementioned common D-Loop variations, these
unusual alterations may lead to the functional
impairment of the mitochondria. It remains to be
proved that these alterations are originated during
tumor development and may play a role in tumorigen-
esis (promoting the growth advantage of tumor cells).
Interestingly, the frequency of somatic mutations
in D310 region is higher, though not significantly
(PZ0.178), in adenomas than in carcinomas. This
finding fits with Sanchez-Cespedes et al. (2001)
description of D310 alterations in pre-neoplastic
lesions of head and neck tumors and indicates that
these alterations occur early in human carcinogenesis
(Sanchez-Cespedes et al., 2001). More recently,
Miyazono et al. (2002) verified that mtDNA D-Loop
mutations occur early and frequently in adenocarci-
nomas arising from Barrett’s esophagus.
Our results confirm that D310 is a major target
for mutations in human primary tumors
(Sanchez-Cespedes et al., 2001). The rate of D310
alterations (30.3%) lies in between the values
described by Sanchez-Cespedes et al. (2001) (0% in
ovarian and prostate tumors and 62.5% in gastric
tumors). The assumption that D310 was a hotspot was
indirectly confirmed by a very low rate of mutations/
instability in the other polymorphic homopolymeric C
stretch (D568).
The high prevalence (22.7%) of somatic mutations
in the dinucleotide D514 region identifies this stretch
as another mutational hotspot. The significantly
higher rate of D514 alterations in carcinomas than
in adenomas supports the assumption that such
alterations may be related to tumor progression, but
the study of a larger series of benign and malignant
tumors of the thyroid and other organs is necessary
before drawing any definitive conclusion. MtDNA
alterations, in other regions besides D-Loop, seem to
play also a role in tumor development. In previous
studies, our group (Maximo et al., 2002) and others
(Yeh et al., 2000), verified that mtDNA mutations/
variants, namely mtDNA complex I genes somatic
missense mutations, as well as mtDNA complex I
genes variants, are more common in human thyroid
malignant tumors, suggesting that mtDNA mutations
may play a role in tumor progression (Maximo et al.,
2002; Yeh et al., 2000). Recently, Petros et al. (2005)
and Shidara et al. (2005) demonstrated that mtDNA
mutations confer a positive contribution to the
promotion of cancer via the prevention from apoptosis
(Petros et al., 2005; Shidara et al., 2005). Taken
together, the aforementioned data suggest that
mtDNA mutations may play a role in tumor
progression. Our group has verified, furthermore,
that mutations in nuclear genes involved in the MRC
can also play a role in the causation and/or
development of neoplasms, namely Hurthle cell
tumors of the thyroid (Maximo et al., 2005).
The coexistence of deletions/insertions at two or
even three repetitive regions of the D-Loop in the
same tumors, reinforces the concept of mitochondrial
microsatellite instability (mtMSI) as advanced by
(Habano et al., 1998; Habano et al., 2000). The term
mtMSI was introduced by (Habano et al., 1998, 2000),
in a study on colorectal tumors, to describe alterations
(small insertions/deletions) in repetitive regions of
mtDNA D-Loop in analogy with the nuclear
microsatellite instability (nMSI). Hypermutable status
(instability) in the nuclear genome has been observed
in certain types of cancers, in which microsatellite
sequences are preferentially affected due to slippage
during the replication process (Aaltonen et al.,1993;
Ionov et al., 1993; Thibodeau et al., 1993). These
replication error phenotypes are related to functional
loss of mismatch repair (MMR) genes, including
hMSH2, hMLH1, hPMS1 and hPMS2 genes (Parsons
et al. 1993; Boyer et al., 1995). In the mitochondrial
genome, the MMR system has been found only in
yeast strains, in which MSH1 and MSH2 are
separately involved in mitochondrial and nuclear
DNA repair systems, respectively (Reenan and
Kolodner, 1992). No MSH1 homologue has been
found in mammalian cells and, therefore, it remains
uncertain whether or not an MMR system plays a role
in the maintenance of mammalian mitochondrial

V. Maximo et al. / Mitochondrion xx (2005) 1–8 7
DTD 5 ARTICLE IN PRESS
genome. (Habano et al., 1998, 2000) suggested that
this mtDNA instability was related with nuclear
microsatellite instability (nMSI), but we (Maximo
et al., 2001) and others (Richard et al., 2000) found
that these of alterations were not related to each other,
at least in thyroid (Maximo et al., 2001) and breast
(Richard et al., 2000) tumors. It seems likely that
oxidative damage plays a role in the acquisition of
concurrent alterations in several repetitive regions of
the D-Loop (Backer et al., 1980; Beckman and Ames,
1997; Cadet et al., 1997), but further studies are
necessary to clarify this point.
Regardless of the mechanism(s) that lead(s) to
mtDNA instability it is surprising the different
frequencies of instability in the different repetitive
regions studied, namely those that are similar, such as
D310 and D568 mononucleotide repetitive regions.
The different frequencies of alterations in the three
distinct mtDNA D-Loop regions may reflect the
importance of each region in mitochondrial biogen-
esis and/or tumor transformation, or differences on the
intrinsic susceptibility for mutations.
The different rates of D310 detected by Sanchez-
Cespedes et al. (2001) and Miyazono et al. (2002) in
different tumor types suggest the existence of
alternative mechanisms for the generation of some
D310 alterations, such as the rate of acquired
mutations during tumor development, the number of
mitochondria per cell, or the intrinsic differences in
the number of cell divisions prior to tumor formation
in different tissues. The results of the present study
and the findings of Sanchez-Cespedes et al. (2001)
support the assumption that mtDNA somatic
mutations may play different roles in different tumor
types. It remains to be proved, however, the
pathogenic role of mtDNA D-Loop instability in
human cancer, namely in thyroid tumors.
Acknowledgements
This study was partially supported by a Ph.D.
Grant (SFRH/BD/8425/2002 - JL) and by a Post-Doc
Grant (SFRH/BPD/14594/2003 - VM) from the
Portuguese Science and Technology Foundation
(FCT) and by further funding from the same source
(Project - POCT/41055/NSE/2001).
References
Aaltonen, L.A., Peltomaki, P., Leach, F.S., Sistonen, P.,
Pylkkanen, L., Mecklin, J.P., Jarvinen, H., Powell, S.M.,
Jen, J., Hamilton, S.R., Petersen, G.M., Kinzler, K.W.,
Vogelstein, B., de la Chapelle, A., 1993. Clues to the
pathogenesis of familial colorectal cancer. Science 260,
812–816.
Anderson, S., Bankier, A.T., Barrell, B.G., de Bruijn, M.H.,
Coulson, A.R., Drouin, J., Eperon, I.C., Nierlich, D.P.,
Roe, B.A., Sanger, F., Schreier, P.H., Smith, A.J., Staden, R.,
Young, I.G., 1981. Sequence and organization of the human
mitochondrial genome. Nature 290, 457–465.
Andrews, R.M., Kubacka, I., Chinnery, P.F., Lightowlers, R.N.,
Turnbull, D.M., Howell, N., 1999. Reanalysis and revision of
the Cambridge reference sequence for human mitochondrial
DNA. Nat. Genet. 23, 147.
Backer, J.M., Weinstein, I.B., 1980. Mitochondrial DNA is a major
cellular target for a dihydrodiol-epoxide derivative of benzo[a]
pyrene. Science 209, 297–299.
Beckman, K.B., Ames, B.N., 1997. Oxidative decay of DNA.
J. Biol. Chem. 272, 19633–19636.
Boyer, J.C., Umar, A., Risinger, J.I., Lipford, J.R., Kane, M.,
Yin, S., Barrett, J.C., Kolodner, R.D., Kunkel, T.A., 1995.
Microsatellite instability, mismatch repair deficiency, and
genetic defects in human cancer cell lines. Cancer Res. 55,
6063–6070.
Brown, M.D., Wallace, D.C., 1994. Molecular basis of mitochon-
drial DNA disease. J. Bioenerg. Biomembr. 26, 273–289.
Budowle, B., Chakraborty, R., Giusti, A.M., Eisenberg, A.J.,
Allen, R.C., 1991. Analysis of the VNTR locus D1S80 by the
PCR followed by high-resolution PAGE. Am. J. Hum. Genet.
48, 137–144.
Burgart, L.J., Zheng, J., Shu, Q., Strickler, J.G., Shibata, D., 1995.
Somatic mitochondrial mutation in gastric cancer. Am.
J. Pathol. 147, 1105–1111.
Cadet, J., Berger, M., Douki, T., Ravanat, J.L., 1997. Oxidative
damage to DNA: formation, measurement, and biological
significance. Rev. Physiol. Biochem. Pharmacol. 131, 1–87.
Cavelier, L., Jazin, E., Jalonen, P., Gyllensten, U., 2000. MtDNA
substitution rate and segregation of heteroplasmy in coding and
noncoding regions. Hum. Genet. 107, 45–50.
Fliss, M.S., Usadel, H., Caballero, O.L., Wu, L., Buta, M.R.,
Eleff, S.M., Jen, J., Sidransky, D., 2000. Facile detection of
mitochondrial DNA mutations in tumors and bodily fluids.
Science 287, 2017–2019.
Habano, W., Nakamura, S., Sugai, T., 1998. Microsatellite
instability in the mitochondrial DNA of colorectal carcinomas:
evidence for mismatch repair systems in mitochondrial genome.
Oncogene 17, 1931–1937.
Habano, W., Sugai, T., Nakamura, S.I., Uesugi, N., Yoshida, T.,
Sasou, S., 2000. Microsatellite instability and mutation of
mitochondrial and nuclear DNA in gastric carcinoma. Gastro-
enterology 118, 835–841.
Hauswirth, W.W., Clayton, D.A., 1985. Length heterogeneity of a
conserved displacement-loop sequence in human mitochondrial
DNA. Nucleic Acids Res. 13, 8093–80104.

V. Maximo et al. / Mitochondrion xx (2005) 1–88
DTD 5 ARTICLE IN PRESS
Hedinger, C.E., Williams, E.D., Sobin, L.H., 1988. Histological
typing of thyroid tumors. In: Hedinger, C.E. (Ed.), International
Histological Classification of Tumors, vol. 11. Springer, Berlin.
Ionov, Y., Peinado, M.A., Malkhosyan, S., Shibata, D.,
Perucho, M., 1993. Ubiquitous somatic mutations in simple
repeated sequences reveal a new mechanism for colonic
carcinogenesis. Nature 363, 558–561.
Lee, D.Y., Clayton, D.A., 1998. Initiation of mitochondrial DNA
replication by transcription and R-loop processing. J. Biol.
Chem. 46, 30614–30621.
Lewis, P.D., Baxter, P., Paul Griffiths, A., Parry, J.M.,
Skibinski, D.O., 2000. Detection of damage to the mitochon-
drial genome in the oncocytic cells of Warthin’s tumor.
J. Pathol. 191, 274–281.
Maximo, V., Soares, P., Seruca, R., Rocha, A.S., Castro, P.,
Sobrinho-Simoes, M., 2001. Microsatellite instability, mito-
chondrial DNA large deletions, and mitochondrial DNA
mutations in gastric carcinoma. Genes Chromosomes. Cancer
32, 136–143.
Maximo, V., Soares, P., Lima, J., Cameselle-Teijeiro, J., Sobrinho-
Simoes, M., 2002. Mitochondrial DNA somatic mutations
(point mutations and large deletions) and mitochondrial DNA
variants in human thyroid pathology. A study with emphasis on
Hurthle cell tumors. Am. J. Pathol. 160, 1857–1865.
Maximo, V., Botelho, T., Capela, J., Soares, P., Lima, J.,
Taveira, A., Amaro, T., Barbosa, A.P., Preto, A.,
Harach, H.R., Williams, D., Sobrinho-Simoes, M., 2005.
Somatic and germline mutation in GRIM-19, a dual function
gene involved in mitochondrial metabolism and cell death, is
linked to mitochondrion-rich (Hurthle cell) tumours of the
thyroid. Br. J. Cancer 92, 1892–1898.
MITOMAP: A Human Mitochondrial Genome Database, 2001.
Center for Molecular Medicine, Emory University, Atlanta, GA,
USA. http://www.gen.emory.edu/mitomap.html.
Miyazono, F., Schneider, P.M., Metzger, R., Warnecke-Eberz, U.,
Baldus, S.E., Dienes, H.P., Aikou, T., Hoelscher, A.H., 2002.
Mutations in the mitochondrial DNA D-Loop region occur
frequently in adenocarcinoma in Barrett’s esophagus. Oncogene
21, 3780–3783.
Parsons, R., Li, G.M., Longley, M.J., Fang, W.H.,
Papadopoulos, N., Jen, J., de la Chapelle, A., Kinzler, K.W.,
Vogelstein, B., Modrich, P., 1993. Hypermutability and
mismatch repair deficiency in RERC tumor cells. Cell 75,
1227–1236.
Petros, J.A., Baumann, A.K., Ruiz-Pesini, E., Amin, M.B.,
Sun, C.Q., Hall, J., Lim, S., Issa, M.M., Flanders, W.D.,
Hosseini, S.H., Marshall, F.F., Wallace, D.C., 2005. mtDNA
mutations increase tumorigenicity in prostate cancer. Proc. Natl
Acad Sci. USA 102, 719–724.
Polyak, K., Li, Y., Zhu, H., Lengauer, C., Willson, J.K.,
Markowitz, S.D., Trush, M.A., Kinzler, K.W., Vogelstein, B.,
1998. Somatic mutations of the mitochondrial genome in human
colorectal tumors. Nat. Genet. 20, 291–293.
Reenan, R.A., Kolodner, R.D., 1992. Isolation and characterization
of two Saccharomyces cerevisiae genes encoding homologs of
the bacterial HexA and MutS mismatch repair proteins.
Genetics 132, 963–973.
Richard, S.M., Bailliet, G., Paez, G.L., Bianchi, M.S., Peltomaki, P.,
Bianchi, N.O., 2000. Nuclear and mitochondrial genome
instability in human breast cancer. Cancer Res. 60, 4231–4237.
Rosai, J., Carcangiu, M.L., DeLellis, R.A., 1992. Tumors of the
thyroid gland. In Atlas of Tumor Pathology 3rd series. Armed
Forces Institute of Pathology, Washington.
Sanchez-Cespedes, M., Parrella, P., Nomoto, S., Cohen, D.,
Xiao, Y., Esteller, M., Jeronimo, C., Jordan, R.C., Nicol, T.,
Koch, W.M., Schoenberg, M., Mazzarelli, P., Fazio, V.M.,
Sidransky, D., 2001. Identification of a mononucleotide repeat
as a major target for mitochondrial DNA alterations in human
tumors. Cancer Res. 61, 7015–7019.
Shidara, Y., Yamagata, K., Kanamori, T., Nakano, K., Kwong, J.Q.,
Manfredi, G., Oda, H., Ohta, S., 2005. Positive contribution of
pathogenic mutations in the mitochondrial genome to the
promotion of cancer by prevention from apoptosis. Cancer Res.
65, 1655–1663.
Thibodeau, S.N., Bren, G., Schaid, D., 1993. Microsatellite
instability in cancer of the proximal colon. Science 260,
816–819.
Wallace, D.C., 1994. Mitochondrial DNA sequence variation in
human evolution and disease. Proc. Natl Acad Sci. USA 91,
8739–8746.
Wallace, D.C., Brown, M.D., Melov, S., Graham, B., Lott, M.,
1998. Mitochondrial biology, degenerative diseases and aging.
Biofactors 7, 187–190.
Xu, B., Clayton, D.A., 1995. A persistent RNA–DNA hybrid is
formed during transcription at a phylogenetically conserved
mitochondrial DNA sequence. Mol. Cell. Biol. 15, 580–589.
Yeh, J.J., Lunetta, K.L., van Orsouw, N.J., Moore, F.D.,
Mutter, G.L., Vijg, J., Dahia, P.L., Eng, C., 2000. Somatic
mitochondrial DNA (mtDNA) mutations in papillary thyroid
carcinomas and differential mtDNA sequence variants in cases
with thyroid tumors. Oncogene 19, 2060–2066.
Related Documents











