Provided for non-commercial research and educational use only. Not for reproduction, distribution or commercial use. This chapter was originally published in the book Advances in Agronomy, Vol. 110, published by Elsevier, and the attached cop y is provided by Elsevier for the author's benefit and for the benefit of the author's institution, for non-commercial research and educational use including without limitation use in instruction at your institution, sending it to specific colleagues who know you, and providing a copy to your institution’s administrator. All other uses, reproduction and distribution, including without limitation commercial reprints, selling or licensing copies or access, or posting on open internet sites, your personal or institution’s website or repository, are prohibited. For exceptions, permission may be sought for such use through Elsevier's permissions site at: http://www.elsevier.com/locate/permissionusematerial From: N. K. Fageria and A. Moreira, The Role of Mineral Nutrition on Root Growth of Crop Plants. In Donald L. Sparks, editor: Advances in Agronomy, Vol. 110, Burlington: Academic Press, 2011, pp. 251-331. ISBN: 978-0-12-385531-2 © Copyright 2011 Elsevier Inc. Academic Press.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 1/82
Provided for non-commercial research and educational use only.
Not for reproduction, distribution or commercial use.
This chapter was originally published in the book Advances in Agronomy, Vol. 110,
published by Elsevier, and the attached copy is provided by Elsevier for the author's
benefit and for the benefit of the author's institution, for non-commercial research and
educational use including without limitation use in instruction at your institution,sending it to specific colleagues who know you, and providing a copy to your
institution’s administrator.
All other uses, reproduction and distribution, including without limitation commercial
reprints, selling or licensing copies or access, or posting on open internet sites, your
personal or institution’s website or repository, are prohibited. For exceptions,
permission may be sought for such use through Elsevier's permissions site at:
http://www.elsevier.com/locate/permissionusematerial
From: N. K. Fageria and A. Moreira, The Role of Mineral Nutrition on Root Growth
of Crop Plants. In Donald L. Sparks, editor: Advances in Agronomy, Vol. 110,
Burlington: Academic Press, 2011, pp. 251-331.
ISBN: 978-0-12-385531-2
© Copyright 2011 Elsevier Inc.
Academic Press.

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 2/82
C H A P T E R F O U R
The Role of Mineral Nutrition
on Root Growth of Crop Plants
N. K. Fageria* and A. Moreira†
Contents
1. Introduction 252
2. Root-Induced Changes in the Rhizosphere 255
3. Root Systems of Cereals and Legumes 2564. Contribution of Root Systems to Total Plant Weight 260
5. Rooting Depth and Root Distribution 263
6. Root Growth as a Function of Plant Age 265
7. Root–Shoot Ratio 268
8. Root Growth Versus Crop Yield 270
9. Genotypic Variation in Root Growth 271
10. Root Oxidation Activity in Oxygen-Deficient Soils 274
11. Root Growth in Conservation Tillage Systems 276
12. Mineral Nutrition Versus Root Growth 27812.1. Nitrogen 279
12.2. Phosphorus 287
12.3. Potassium 299
12.4. Calcium 301
12.5. Magnesium 302
12.6. Sulfur 303
12.7. Micronutrients 304
13. Management Strategies for Maximizing Root Systems 312
13.1. Soil management 312
13.2. Plant management 31614. Conclusions 317
Acknowledgment 318
References 318
Abstract
Agriculture is going through a profound revolution worldwide due to increasing
world demand for food, higher costs of energy and other inputs, environmental
pollution problems, and instability of cropping systems. In this context,
Advances in Agronomy, Volume 110 # 2011 Elsevier Inc.
ISSN 0065-2113, DOI: 10.1016/B978-0-12-385531-2.00004-9 All rights reserved.
* Rice and Bean Research Center of Embrapa, Santo Antonio de Goias, GO, Brazil{ Western Amazon Research Center of Embrapa, Manaus, AM, Brazil
251

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 3/82
knowledge of factors that affect root development is fundamental to improving
nutrient cycling and uptake in soil–plant systems. Roots are important organs
that supply water, nutrients, hormones, and mechanical support (anchorage) to
crop plants and consequently affect economic yields. In addition, roots improve
soil organic matter (OM) by contributing to soil pools of organic carbon (C),
nitrogen (N), and microbial biomass. Root-derived soil C is retained and forms
more stable soil aggregates than shoot-derived soil C. Although roots normally
contribute only 10–20% of the total plant weight, a well-developed root system
is essential for healthy plant growth and development. Root growth of plants is
controlled genetically, but it is also influenced by environmental factors. Min-
eral nutrition is an important factor influencing the growth of plant roots, but
detailed information on nutritional effects is limited, primarily because roots
are half-hidden organs that are very difficult to separate from soil. As a result, it
is difficult to measure the effect of biotic and abiotic factors on root growthunder field conditions. Root growth is mainly measured in terms of root density,
length, and weight. Root dry weight is often better related to crop yields than is
root length or density. The response of root growth to chemical fertilization is
similar to that of shoot growth; however, the magnitude of the response may
differ. In nutrient-deficient soils, root weight often increases in a quadratic
manner with the addition of chemical fertilizers. Increasing nutrient supplies
in the soil may also decrease root length but increase root weight in a quadratic
fashion. Roots with adequate nutrient supplies may also have more root hairs
than nutrient-deficient roots. This may result in greater uptake of water and
nutrients by roots well supplied with essential plant nutrients, compared with
roots grown in nutrient-deficient soils. Under favorable conditions, a major part
of the root system is usually found in the top 20 cm of soil. Maximum root
growth is generally achieved at flowering in cereals and at pod-setting in
legumes. Genotypic variations are often found in the response of root growth
to nutrient applications, and the possibility of modifying root system response
to soil properties offers exciting prospects for future improvements in crop
yields. Rooting pattern in crop plants is under multi- or polygenic control, and
breeding programs can be used to improve root system properties for environ-
ments where drought is a problem. The use of crop species and cultivarstolerant to biotic and abiotic stresses, as well as the use of appropriate cultural
practices, can improve plant root system function under favorable and unfavor-
able environmental conditions.
1. Introduction
Roots are important plant organs. They absorb water and nutrientsfrom the soil and translocate them to plant tops (Merrill et al ., 1996, 2002;Sainju et al ., 2005a; Stone et al ., 2001). Roots also give mechanical supportto plants and supply hormones that affect many physiological and biochem-ical processes associated with growth and development. Roots exert control
252 N. K. Fageria and A. Moreira

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 4/82
over whole-plant growth and development by controlling the uptake of mineral nutrients (Zobel, 1986). Cytokinins produced in roots are translo-cated to shoots and participate in the control of leaf protein biosynthesis
(Oritani, 1995). In addition, cytokinins may control nitrogen assimilation inthe root itself. Oritani (1995) also reported that rice roots synthesize zeatinglucoside, a bound cytokinin, in addition to isopentenyladenine (IP),isopentenyl-adenosine (IPA), trans-zeatin riboside (tZR), and trans-zeatin(tZ), which are free cytokinins in zeatin-related compounds. Zobel (2005a)reported that root system dynamics are instrumental in the maintenance of biological and chemical equilibrium within the soil and modulate changesto soil quality. In addition, genotypes with inherently large root systemshave been associated with reduced lodging in cereals and legumes (Stoffella
and Kahn, 1986). Soil is knitted together by plant roots, which formcomplex and structurally diverse reinforcing structures. Near the soil sur-faces, fibrous plant roots hold soil aggregates together against the stresses of water and wind.
Vigorous root systems are needed for the development of healthy plantsand consequently, higher yields. Roots that are left in the soil after cropharvest improve soil organic matter (OM) content and contribute to thenitrogen cycle and microbial activity (Sainju et al ., 2005a). All these activ-ities improve soil structure, soil water holding capacity, water infiltrationinto the soil, as well as reduce soil bulk density and soil erosion, ultimatelyleading to greater soil productivity.
Processes that are largely controlled or directly influenced by roots andoften occur in the vicinity of the root surface are often referred to asrhizosphere processes (Cheng and Kuzyakov, 2005). These processes mayinclude root turnover, rhizodeposition, root respiration, and rhizospheremicrobial respiration that are a result of microbial utilization of rhizodepo-sits. Rhizosphere processes play an important role in the global C cycle.Terrestrial ecosystems are intimately connected to atmospheric carbondioxide levels through photosynthetic fixation of CO
2
, sequestration of CO2 in plant and soil biomass, and the subsequent release of C throughrespiration and decomposition of organic matter (Cheng and Kuzyakov,2005). Carbon cycling belowground is increasingly being recognized as oneof the most significant components of the ecosystem C fluxes and pools(Cheng and Kuzyakov, 2005; Jackson et al ., 1997; Zak and Pregitzer, 1998).
Roots improve soil aggregation, which controls biological and hydro-logic properties of the soil. A soil aggregate is a group of primary soilparticles that adhere to one another more strongly than to surroundingsoil particles (Follett et al ., 2009). Root materials remain mixed within thesoil as they decompose, providing a gum-like material that cements soilparticles into aggregates (Melillo and Gosz, 1983; Tresder et al ., 2005). Rootexudation occurs when organic acids either are leaked from the root or arereleased as a means of interacting with microbes in the rhizosphere. These
The Role of Mineral Nutrition on Root Growth of Crop Plants 253

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 5/82
exudates can influence microbial activity and the subsequent metabolism of root detritus. Mucilages released by the root cap and epidermis link partic-ulate organic residues with mineral fragments. Microbes in the rhizosphere
use plant mucilage as a substrate for growth and secrete their own mucilage,thereby producing mucigel, a mucilaginous material of mixed origin whichstabilizes finer aggregates. In addition, in many soils, fungal hyphae areimportant for stabilizing larger structural units (Goss and Kay, 2005).Finally, allocation of carbohydrates and other C-containing moleculesdirectly to mycorrhizal fungi forms another conduit of C into the soil asthese microbes secrete their own exudates and as their tissues senesce(Tresder et al ., 2005).
The amount of C and N supplied by roots can be significant for main-
taining or improving soil organic matter (Sainju et al ., 2005b). The organicinput from plant roots to the surrounding soil is the principal support of thebiological activity and abundance of organisms in the rhizosphere (Chenget al ., 1994; Kirchner et al ., 1993). As much as 7–43% of the total above-ground and belowground plant biomass can be contributed by roots (Kuoet al ., 1997a,b). Roots can supply from 400 to 1460 kg C ha1 during agrowing season (Kuo et al ., 1997a; Qian and Doran, 1996). Liang et al . (2002)reported that roots contributed as much as 12% of soil organic C, 31% of water soluble C, and 52% of microbial biomass C within a growing season.Roots may play a dominant role in soil C and N cycles (Gale et al ., 2000a;Puget and Drinkwater, 2001; Wedin and Tilman, 1990). Roots may haverelatively greater influence on soil organic C and N levels than the above-ground plant biomass (Boone, 1994; Haider et al ., 1993; Milchumas et al .,1985; Norby and Cortrufo, 1998; Sanchez et al ., 2002). Balesdent andBalabane (1996) reported that corn roots contributed 1.6 times more C tosoil organic C than did stover. Root-derived C is retained and forms morestable aggregates than does shoot-derived C (Gale et al ., 2000a,b).
Rhizodeposition, such as root exudates, mucilages, and sloughed cells,may be a significant source of soil organic C (Balesdent and Balabane, 1996;Buyanovsky et al ., 1986; Sainju et al ., 2005a). Helal and Sauerbeck (1987)estimated that the amount of C released from roots as rhizodeposit could bemore than 580 kg C ha1. This rhizodeposition increases microbial activityand influences N mineralization in the soil (Bakken, 1990; Texier and Biles,1990). Carbon contribution from corn root biomass and rhizodeposition tosoil organic C can be as much as 1.7–3.5 times greater than from stover (Allmaras et al ., 2004; Wilts et al ., 2004).
The environment is seldom optimum for extensive and effective rootgrowth. Canopy conditions that limit photosynthesis reduce shoot growthand limit assimilate translocation to the roots, thus reducing root growth(Miller, 1986). Root growth is under multi- or polygenic control and is alsoinfluenced by environmental factors, including soil temperature, soil mois-ture content, solar radiation and soil physical, chemical, and biological
254 N. K. Fageria and A. Moreira

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 6/82
properties (Fageria, 2002c, 2009; Klepper, 1992; Merrill et al ., 1996;Russell, 1977; Sainju et al ., 2005b; Zobel, 1991). Most of the root biomassof annual crops is located in the 0–20 cm soil depth. This may be associated
with greater organic matter, nutrients, aeration, and water availability in thetop soil layer compared to lower soil depths (Merrill et al ., 1996, 2002; Qinet al ., 2004; Sainju et al ., 2005a; Stone et al ., 2001).
Increased knowledge of root architecture and root development dynam-ics could help improve crop productivity in agroecosystems. Better under-standing of root architecture and growth dynamics of annual crops may leadto a more efficient use of applied nutrients and water. The study of plantroots is one of the most promising, but least explored, areas of researchrelated to plant growth. The aerial portions of plant species have received
greater attention and study, probably because of their conspicuousness andeasy access, while the subterranean portions have been neglected because of the difficulty of observing and sampling them and the disruption of rootsystems when they are removed from soil. Many crop root studies haverelied on soil cores and more recently on minirhizotron observations (Boxand Ramseur, 1993; Zobel, 2005b). The data collected using these methodsmay not be representative of the crop as a whole (Andren et al ., 1991;Hansson et al ., 1992; Hoad et al ., 2001; Parker et al ., 1991). In addition,information about annual field crop root growth dynamics as a function of environmental factors is scattered and often not readily accessible.
The primary objectives of this chapter are to review the latest advances inrelation to the role of mineral nutrition in the growth and development of roots of annual crops. To make the subject matter as practical as possible, mostof the discussion is supported by experimental results. Our approach shouldenhance understanding on the contribution of roots to total dry matter of crops, to assess the effects of root system size and form on overall crop growth,and to relate the effects of root growth on the environment. This informationmay be useful for agricultural scientists in the fields of plant nutrition, water use, breeding, and plant physiology who are interested in conductingresearch to manipulate plant root systems in favor of higher yields.
2. Root-Induced Changes in the Rhizosphere
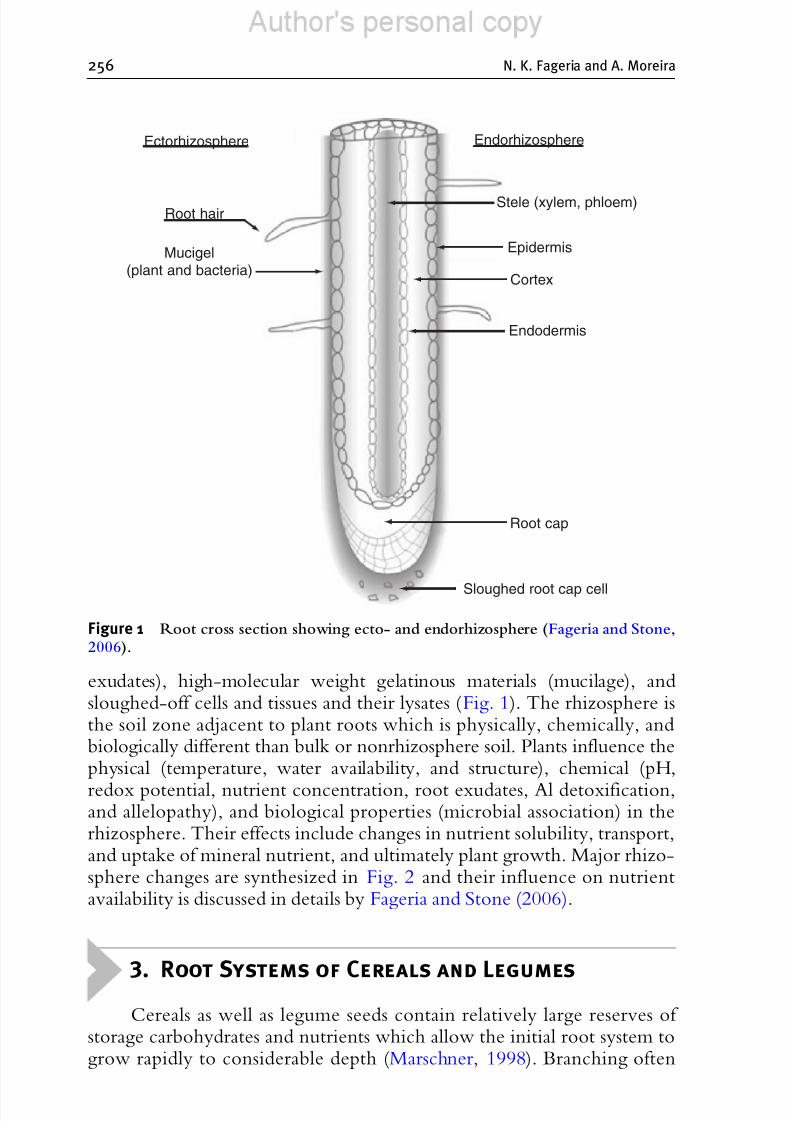
Pinton and Varannini (2001) suggested that the soil layer surroundingroots should be termed the ectorhizosphere and the root inside the layer colonized by microorganisms should be designated as endorhizosphere. Thetwo areas are separated by the root surface known as rhizoplane (Fig. 1).Growing roots release an appreciable amount of organic components intothe rhizosphere. Marschner (1995) reported that three major componentsreleased by roots are low-molecular weight organic compounds (free
The Role of Mineral Nutrition on Root Growth of Crop Plants 255
8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 7/82
exudates), high-molecular weight gelatinous materials (mucilage), andsloughed-off cells and tissues and their lysates (Fig. 1). The rhizosphere isthe soil zone adjacent to plant roots which is physically, chemically, andbiologically different than bulk or nonrhizosphere soil. Plants influence thephysical (temperature, water availability, and structure), chemical (pH,
redox potential, nutrient concentration, root exudates, Al detoxification,and allelopathy), and biological properties (microbial association) in therhizosphere. Their effects include changes in nutrient solubility, transport,and uptake of mineral nutrient, and ultimately plant growth. Major rhizo-sphere changes are synthesized in Fig. 2 and their influence on nutrientavailability is discussed in details by Fageria and Stone (2006).
3. Root Systems of Cereals and Legumes
Cereals as well as legume seeds contain relatively large reserves of storage carbohydrates and nutrients which allow the initial root system togrow rapidly to considerable depth (Marschner, 1998). Branching often
Endodermis
Cortex
Epidermis
Stele (xylem, phloem)
Root cap
Mucigel
(plant and bacteria)
Root hair
Ectorhizosphere Endorhizosphere
Sloughed root cap cell
Figure 1 Root cross section showing ecto- and endorhizosphere (Fageria and Stone,2006).
256 N. K. Fageria and A. Moreira

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 8/82
begins before the leaves have unfolded, with the result that the plantestablishes early contact with moist soil (Hoad et al ., 2001). Generally,roots are classified into four groups. These groups are the taproot, basalroots, lateral roots, and shootborn or adventitious roots (Zobel, 2005a).When plants produce secondary shoots (tillers) or shoot branches whichdevelop roots, these roots are commonly called adventitious roots. Toindicate the true origin of these adventitious roots, the term shootborn issometimes used (Zobel, 2005a). The primary function of the taproot, basalroots, and adventitious roots is to establish the most optimum frameworkfrom which to initiate small lateral roots to effect water and nutrient uptake(Zobel, 2005b). The taproot penetrates relatively deeply to ensure an
Physical changes
Chemical changes
Biological changes
Temperature
Water holding capacity
Nitrogen fixation
PGPR bacteriasMycorrhizal fungi
Harmful microorganisms
pHRedox potential
Release of organic compounds by rootsNutrient concentration and availability
AI detoxification
Allelopathy
Structure
Figure 2 Major physical, chemical, and biological changes in the rhizosphere (Fageriaand Stone, 2006).
The Role of Mineral Nutrition on Root Growth of Crop Plants 257

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 9/82
adequate supply of soil water, the basal roots spread out laterally to ensure astructure for lateral roots that take up P and other nutrients that are lessabundant in the lower levels of the soil profile (Zobel, 2005b), and to
provide a degree of lodging resistance to the plant as it matures and producesseed (Barlow, 1986; Stoffella et al ., 1979). For many grasses and other speciesin which root secondary thickening is not important, the shootborn rootstake over the role of the basal roots. The shootborn roots continue to buildthe framework with larger and larger conducting roots as the plant increasesin size (Zobel, 2005b). The basal and shootborn roots probably provide littledirect uptake of nutrients and water (St. Aubin et al ., 1986).
Besides length and weight, surface area is an important parameter of theroot system in crop plants. The form of root systems and their development
conditions greatly affect the surface area of roots. The surface area of rootshas a high positive correlation with the amount of nutrient absorption(Takenaga, 1995). Various studies show that 90–95% or more of the rootlength of an intact plant is made up of roots <0.6 mm in diameter (Zobel,2003, 2005b).
Monocots and dicots typically have different root system structures.Root systems of monocots are fibrous, whereas dicots often have taproots.The fibrous root systems of monocots consist of seminal, nodal, and lateralroots. Seminal roots develop from primordia within seeds and nodal rootsdevelop adventitiously from lower stem nodes. All adventitious roots of stem origin are called nodal roots to distinguish them from other adventi-tious roots that emerge from the mesocotyl or elsewhere on the plant.Nodal roots are identified by the node number from which they originate.Nodal roots may be functional or nonfunctional (Thomas and Kaspar,1997). Functional nodal roots are defined as roots that have emerged fromstem nodes, entered the soil, and developed lateral roots and/or root hairs.Nonfunctional nodal roots are defined as roots that have emerged fromaboveground stem nodes and have not entered the soil or produced lateralroots (Thomas and Kaspar, 1997).
Initial seminal or nodal roots develop laterals that are classed as roots of the first order, roots that develop from first-order roots are classed assecond-order roots, and additional roots that develop from these lateralsare classed as third-order roots, fourth-order roots, etc. ( Yamauchi et al .,1987a,b). Nodal roots are also known as adventitious, coronal, and/or crown roots. Roots of cereals such as rice include mesocotyl, radical(seminal), and nodal or adventitious roots ( Yoshida, 1981). Mesocotylroots emerge from the axis between the coleoptile node and the base of the radical, and they typically develop only when seeds are planted verydeep or are treated with chemicals ( Yoshida, 1981). Until adventitious rootsdevelop, seedlings must rely on roots which initiate on the subcoleoptileinternodes above the seed or seminal roots below the seed. Adventitiousroots are important to seedling establishment because they can conduct
258 N. K. Fageria and A. Moreira

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 10/82
more water than smaller diameter seminal roots. Adventitious rootsmay develop as early as 2 weeks after sowing. Seedling survival may increasewhen seeds are sown at greater soil depths, where greater soil water avail-
ability may increase adventitious root development (Fageria et al ., 2006).Tiller roots do not form in cereals until tillers have two to three leaves(Kleeper et al ., 1984), and until these roots have developed, parent culmsmust provide nutrients and water. Parent culms may also have to providehormonal control so essential for tiller survival. Delayed root production bytillers may explain why late tillers often do not survive (Kleeper et al ., 1984).Figure 3 shows the radical and adventitious root system of upland rice(Oryza sativa L.) (cereal) and Fig. 4 shows the tap root system of dry bean(Phaseolus vulgaris L.) (legume).
In addition to their morphological differences, roots of cereals andlegumes have different physiochemical properties. The surface of plantroots has a negative electric charge, mainly due to carboxyl groups in thepectin of the root cell walls. The density of this negative charge is defined asa cation exchange capacity (CEC; Takenaga, 1995). The CEC of cereals
3rd leaf
4th leaf
2nd leaf
1st leaf
1st tiller
Adventitious root
Radicle root
Figure 3 Root system of upland rice seedlings (Fageria, 2007).
The Role of Mineral Nutrition on Root Growth of Crop Plants 259

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 11/82
such as rice, barley, and corn is typically lower than the CEC of legumes likedry bean and broad bean. Roots with high CEC absorb more divalentcations like Ca2þ and Mg2þ than monovalent cations such as Kþ andNH4
þ. On the other hand, roots with lower CEC absorb more monovalentthan divalent cations. Hence, in grass–legume mixtures, legumes generallysuffer with Kþ deficiency due to large uptake of this element by grasses.Essau (1977), Fageria et al . (2006), Klepper (1992), Leskovarant and Stofella
(1995), O’Toole and Bland (1987), and Zobel (1991, 2005a,b) have dealtextensively with various types of monocotyledonous and dicotyledonousroots and root hairs, and their growth and morphology.
4. Contribution of Root Systems to TotalPlant Weight
Crops can accumulate photosynthetic products in their stems, leaves,grains, and roots, and the development of robust root systems is necessary toproduce good crop yields. A well-developed root system is needed toabsorb adequate amounts of water and nutrients, especially when plants
First trifoliate
Gem
Epicotyl
Primary leaf
Hypocotyl
Adventitious roots
Basal roots
Tap rootLateral roots
Figure 4 Root system of dry bean seedling (Fageria and Santos, 2008).
260 N. K. Fageria and A. Moreira

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 12/82
are under abiotic or biotic stress. The traditional view is that a large vigorousroot system, through avoidance of plant water deficits, is required for high
yields in water-limited environments (Ludlow and Muchow, 1990). In
addition, large root systems add more organic matter to the soil, creatingbetter environmental conditions for the growth and development of crops.The distribution of photosynthetic products in the root, shoot, and grain isdetermined genetically, but it also varies with environmental conditions(Fageria, 1992, 2009; Sainju et al ., 2005a). Because roots are so difficult toseparate from soils, little is known about the effects of soil and crop manage-ment practices on their growth and development (Sainju et al ., 2005b).
In most annual plant species, only 10–20% of mature plant biomassconsists of root tissue (Zobel, 1986). Similarly, Fageria (1989a) also reported
that the contribution of roots of food crops to the total plant weight variesfrom 10–20%, depending on species and cultivars within species. However,Sainju et al . (2005b) reported that C accumulation in cotton and sorghumroots ranged from 1% to 14%. Data in Table 1 show the contribution of theroot system of 20 upland rice genotypes grown at two N rates in a BrazilianOxisol. In this study, the nitrogen genotype interaction was significantbecause some genotypes were highly responsive to the N application whileothers were not. Thus, genotype selection is an important strategy for upland rice production in Brazilian Oxisols. In the control treatment, thecontribution of the root system to total plant weight varied from 12% to30%, with an average value of 22%. At the 300 mg N kg1 soil treatment,the contribution of root weight to total plant weight varied from 3% to21%, with an average value of 14%. The proportionally lower root dryweight at the higher N rate was associated with a significant increase in thegrain and shoot weight of rice genotypes with the addition of N fertilizer (Fageria and Baligar, 2005).
The contribution of root systems to total plant weights of tropicallegume cover crops under three P levels is shown in Table 2. There was asignificant influence of P rate, cover crop species and P species interac-tion was significant, indicating different responses of cover crops at differentP rates. At 0 mg P kg1 level, the contribution of roots to total plant weightvaried from 4.91% to 21.50%, with an average value of 12.98%. With100 mg P kg1, the root contribution to the total plant weight variedfrom 11.30% to 25.16%, with an average value of 18.93%. At the200 mg P kg1 P rate, the contribution of roots to the total plant weightvaried from 6.36% to 25.91%, with an average value of 14.79%. Overall,increase in root contribution to the total plant weight with the increase in Prate may be associated with the response of legumes to P fertilization.Significant differences have been reported among the crop species andgenotypes of the same species in the absorption and utilization of P(Epstein and Bloom, 2005; Fageria, 2009; Marschner, 1995). Tian et al .(1998) reported significant responses of legume cover crops grown on
The Role of Mineral Nutrition on Root Growth of Crop Plants 261

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 13/82
Alfisols in Africa. Similarly, Ae et al . (1990) reported that pigeon pea as acover crop was more efficient in utilizing iron-bound P than several other cover crops. These authors also reported that this ability of pigeon pea wasattributed to root exudates, in particular, piscidic acid and its p-O -methylderivative, which release P from Fe–P by chelating Fe3þ. These results alsoshow that P fertilization also improves the root weight of cover crops,which may be beneficial in improving soil organic matter content and soilmicrobial activities.
Table 1 Contribution of upland rice roots in the total plant dry weight (%) asinfluenced by N rates
Genotype 0 mg N kg1 soil 300 mg N kg1 soil
BRA01506 18abcd 3g
BRA01596 18abcd 3g
BRA01600 23abcd 6fgBRA02535 25abcd 14cde
BRA02601 23abcd 19abcdBRA032033 20abcd 15bcde
BRA032039 14cd 21ab
BRA032048 12d 19abcBRA032051 23abcd 14cde
BRA042094 25abcd 12eBRA042156 24abcd 15cde
BRA042160 28abc 15cdeBRA052015 17abcd 13de
BRA052023 16bcd 21aBRA052033 22abcd 15bcde
BRA052034 30a 12eBRA052045 27abc 17abcde
BRA052053 29ab 11ef
BRS Primavera 28abc 12eBRS Sertaneja 26abc 14de
Average 22 14F -test
N rate (N) *Genotype (G) **
N G **CV(%) 17
Contribute of roots in total plant dry weight %ð Þ ¼ Root dry weight
Dry weight of root; shoot; and grain 100:
*,** Significant at the 5% and 1% probability levels, respectively. Means followed by the same letter inthe same column are not significant at the 5% probability level by Tukey’s test.
262 N. K. Fageria and A. Moreira

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 14/82
5. Rooting Depth and Root Distribution
Rooting depth and distribution are important traits for absorption of water and nutrients from the soil profile. Rooting depth, that is, themaximum depth that roots reach, is difficult to ascertain in the field(Hoad et al ., 2001). Hsiao et al . (2009) reported that roots typically reachmaximum depth about the time when the canopy begins to senesce under nonstress conditions. Gregory (1994) reported that rooting depth in cerealsincreases until anthesis. Hoad et al . (2001) reviewed the rooting depthliterature for cereals and concluded that individual roots of cereal cropscan reach a depth of over 2.0 m under favorable conditions. Soil compac-tion reduces rooting depth (Lipiec et al ., 1991; Unger and Kaspar, 1994).
Table 2 Contribution of tropical legume cover crops root dry weight (%) in the totalplant weight as influenced by phosphorus rates
Cover crop 0 mg P kg1 100 mg P kg1 200 mg P kg1
Crotalaria breviflora 20.77a 11.39ef 21.22abCrotalaria juncea L. 12.28abc 19.77abcdef 12.49bcdCrotalaria mucronata 4.91c 19.29abcdef 21.96abCrotalaria spectabilis Roth 11.66abc 24.72ab 9.26cdCrotalaria ochroleuca G. Don 5.18c 22.76abc 17.38abcCalopogonium mucunoides 11.66abc 12.16def 25.91aPueraria phaseoloides Roxb. 7.22bc 17.17abcdef 8.15cdCajanus cajan L. Millspaugh 21.50a 21.12abcde 6.36dCajanus cajan L. Millspaugh 16.90abc 21.80abcd 13.77bcd
Dolichos lablab L. 12.81abc 22.22abc 14.71bcdMucuna deeringiana (Bort) Merr. 19.70ab 15.08bcdef 13.36bcdMucuna aterrima (Piper & Tracy)
Holland14.17abc 25.16a 16.43abc
Mucuna cinereum L. 11.09abc 13.44cdef 16.94abcCanavalia ensiformis L. DC. 11.83abc 11.30f 9.12cd
Average 12.98 18.93 14.79F -test
P rate (P) **
Cover crops (C) **P C **
Contribute of roots in total plant dry weight %ð Þ ¼ Root dry weight
Dry weight of root and shoot 100:
** Significant at the 1% probability level. Means followed by the same letter in the same column are notsignificant at the 5% probability level by Tukey’s test.
The Role of Mineral Nutrition on Root Growth of Crop Plants 263

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 15/82
Martino and Shaykewich (1994) reported that the proportion of rootspenetrating the soil is inversely related to the soil penetration resistance.
Ueda (1936) observed that wheat cultivars with relatively great cold
resistance had roots that penetrated rapidly into deeper soil layer at earlystages of growth. Similarly, Sanders and Brown (1978) reported that differ-ences in the yields of indeterminate and determinate soybean varietiesmirrored the distribution of their root systems, although the dry weightsof their root systems were almost the same. Yamauchi et al . (1987b)compared the root system distributions of 13 species of cereals and reportedthat there was a significant difference among cereals in the distribution anddepth of penetration of root systems. Some of the cereals had “concen-trated” root systems and others had “scattered” type root systems.
Large amounts of organic matter and immobile nutrients are generallyfound in the upper soil layers. Hence, a major part of the roots of the mostcrops is concentrated in the upper 0–20 cm soil depth (Gregory, 1994).Application of nitrogen fertilizer to barley caused an accumulation of 90–97% of the root mass in the top 30 cm soil layer (Hansson andAndren, 1987). Similarly, Haberle et al. (1996) reported only a fewunbranched primary roots below a depth of 25 cm in fertilized wheat.Differences in root lengths, dry weights of roots at different soil depths,and the extent of rooting at the seedling stage were related to differences in
yield and the ability of wheat cultivars to escape drought (Hurd, 1974).Upland rice cultivars, which are more drought tolerant than lowlandcultivars, have deeper and more prolific rooting systems (Steponkus et al .,1980). When soil types did not restrict the rooting potential, deep rooting of bean cultivars was positively associated with seed yield, crop growth, cooler canopy temperature, and soil water extraction (Sponchiado et al ., 1989).
Because about 90% of the total NH4, P, and K uptake and root length of flooded rice cultivars occur within the surface 20 cm of soil, samplescollected for routine soil tests should be taken from the top 20 cm (Teoet al ., 1995). Lowland rice plants develop a surface mat of roots in theoxygenated zone near the soil surface soon after application of flood waters(University of Arkansas Cooperative Extension Service Rice Committee,1990).
Durieux et al . (1994) reported that more than half of the root length of maize was located in the surface 0–20 cm depth at all sampling times duringa season. Roots of the peanut ( Arachis hypogaea L.) cultivar Florunner penetrated to depths up to 280 cm when grown in a sandy soil, and themost extensive root growth occurred in the top 30 cm (Boote et al ., 1982).Sharratt and Cochran (1993) reported that 85% and 95% of the root mass of barley was located in interrows of the top 20 and 40 cm of soil, respectively.Welbank and Williams (1968) also found that nearly 80% of barley rootsoccupied the uppermost 15 cm of soil. A study conducted by Stone andPereira (1994a,b) of four common bean cultivars and three upland rice
264 N. K. Fageria and A. Moreira

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 16/82
cultivars to evaluate rooting depths in an Oxisol showed that 70% of theroots were concentrated in the top 20 cm layer and about 90% wereconcentrated in the top 40 cm soil depth of both crops.
The presence of these roots in surface soil layers may contribute to largeamounts of nutrients measured in the upper 20 cm of the soil. Using theClaassen–Barber model to predict nutrient uptake by maize grown in siltloam soil, >90% of K and P uptake occurred in the top 20 cm soil depth(Schenk and Barber, 1980). Silberbush and Barber (1984) reported thatabout 80% of P and 54% of K uptake by soybean was from 0 to 15 cmdepths.
Soybean cultivars differ in their rate of downward growth during specificshoot development stages and in their maximum rooting depth on specific.
Cultivars selected for rapid taproot elongation rates in a greenhouse trialwere found to have greater rooting depths in rhizotron and field trials thancultivars selected for slow taproot elongation (Kaspar et al ., 1978, 1984).
6. Root Growth as a Function of Plant Age
Root development varies with stages of plant growth and develop-
ment. The most rapid development of corn (Zea mays L.) roots occursduring the first 8 weeks after planting (Anderson, 1987). As corn plants age,growth of roots generally increases at slower rates than shoots (Baligar,1986). After silking, corn root length declines (Mengel and Barber, 1974).This decline in root length after silking presumably is due to the high Cdemand of grain resulting in enhanced translocation of C and N to grain,including some C and N that roots would normally obtain (Wiesler andHorst, 1993).
Peanut ( A. hypogaea L.) root length density and root weight densityincreased at each soil depth increment from planting to 80 days after planting (Ketring and Reid, 1993). These authors reported that roots hadpenetrated to depths of 120 cm 40–45 days after planting and spread laterallyto 46 cm in mid-furrow. The 0–15 cm depth increment had the highestmean root length density, which increased to a maximum of 2.1 cm cm3 at80 days after planting (Ketring and Reid, 1993). This meant that peanutroots were established both deeply and laterally in the soil profile early in thegrowing season. This would be advantageous in drought environments andhelpful for water management.
Sunflower (Helianthus annuus L.) rooting depth reached 1.88 m at thebeginning of disk flowering and 2.02 m at the completion of disk flowering( Jaffar et al ., 1993). In a review of depth development of roots with time for 55 crop species (Borg and Grimes, 1986), it was shown that maximumrooting depth for most crop species was generally achieved at physiological
The Role of Mineral Nutrition on Root Growth of Crop Plants 265
8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 17/82
maturity. Kaspar et al . (1984) noted that the rate of soybean (Glycine maxL. Merr.) root depth penetration reached a maximum during early flower-ing and declined during seed fill. However, some root growth was observed
throughout the reproductive stage until physiological maturity (Klepper andKaspar, 1994).Slaton et al . (1990) studied root growth dynamics of lowland rice and
found that maximum root growth rates were reached between activetillering and panicle initiation, and maximum root length was reached byearly booting. Beyrouty et al . (1987) noted that the most rapid rate of rootand shoot growth in flooded rice occurred before panicle initiation, whichcorresponds to the plant transition between vegetative and reproductivegrowth. Approximately 77% and 81% of total shoot and root biomass,
respectively, was achieved before panicle initiation. Following panicleinitiation, the length of roots and shoots increased only slightly until harvest(physiological maturity). Beyrouty et al . (1988) also reported that lowlandrice root growth was most rapid during vegetative growth, with maximumroot length occurring at panicle initiation. Root length either plateaued or declined during reproductive growth.
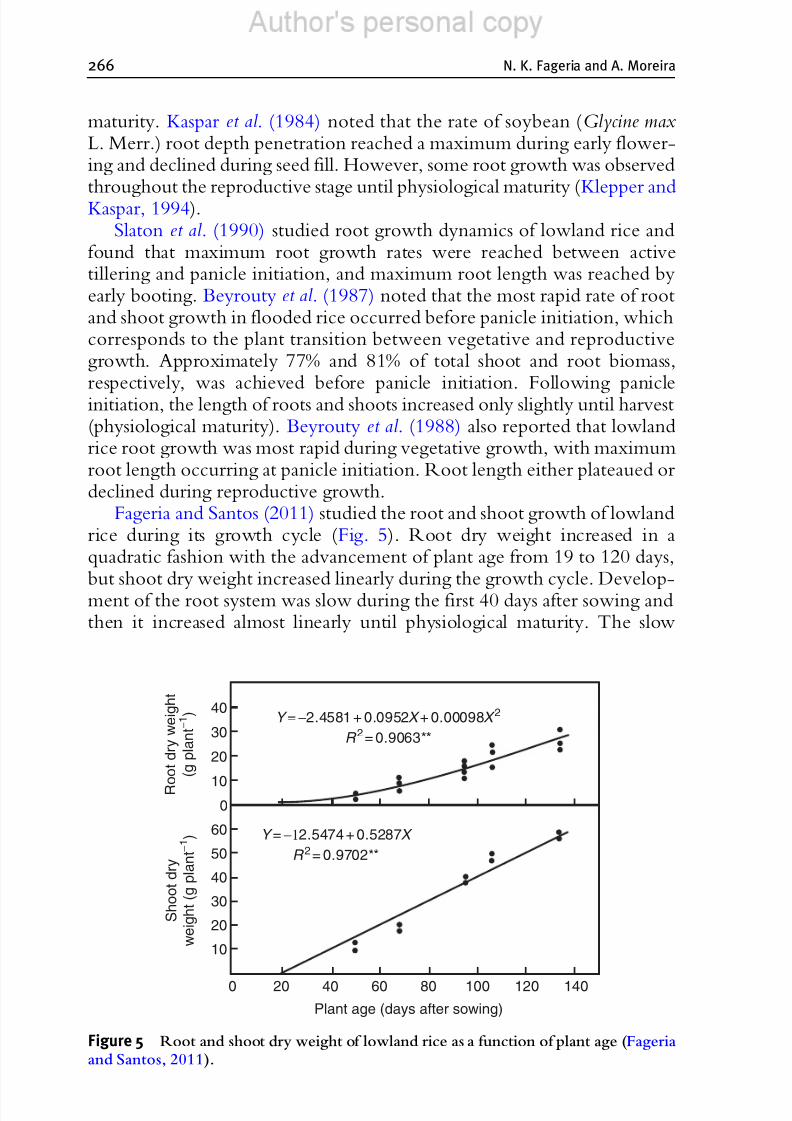
Fageria and Santos (2011) studied the root and shoot growth of lowlandrice during its growth cycle (Fig. 5). Root dry weight increased in aquadratic fashion with the advancement of plant age from 19 to 120 days,but shoot dry weight increased linearly during the growth cycle. Develop-ment of the root system was slow during the first 40 days after sowing andthen it increased almost linearly until physiological maturity. The slow
0
0
20
20
30
30
50
20
10
S h o o t d r y
w e i g h t ( g p l a n t − 1 )
R o o t d r y w e i g h t
( g p l a n t − 1 )
10
40
40
40Y =−2.4581 + 0.0952X + 0.00098X 2
Y =−12.5474+ 0.5287X
R 2= 0.9063**
R 2= 0.9702**
60
60
80
Plant age (days after sowing)
100 120 140
Figure 5 Root and shoot dry weight of lowland rice as a function of plant age (Fageriaand Santos, 2011).
266 N. K. Fageria and A. Moreira

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 18/82
increase in root dry weight early in the growth cycle may be associated withlow translocation of photosynthetic materials due to low leaf area (Fageria,2007). When there is a low amount of photosynthetic product, a major part
goes to the shoot, and very little is translocated to the roots (Fageria, 1992).Root growth generally parallels shoot growth in crop plants. When alarge amount of nutrients, especially N, is supplied to leaves from roots,photosynthesis remains high during maturation, which secures the supply of carbohydrates to roots. Hence, the activities of roots and shoots are mutuallydependent (Osaki et al ., 1997). Figure 6 shows maximum root length androot dry weight of dry bean during the growth cycle of a Brazilian cultivar BRS Valente under greenhouse conditions. Maximum root length was
0
10
20
0
0
0
2
4
10
20
20
10
30
40
20
Plant age (days after sowing)
Y = 0.0297EXP(0.1161X −0.00054X 2)
Y = 0.4461EXP(0.0787X −0.00041X 2)
Y = 0.0267EXP(0.0824X −0.00032X 2)
Y =6.0778+0.7862X −0.0049X 2)
R 2= 0.9216**
R 2
= 0.8545**
R 2= 0.8353**
R 2= 0.9430**
S h o o t d r y w e i g h t
( g p l a n t − 1 )
R o o t d r y w e i g
h t
( g p l a n t − 1 )
M a x i m u m
r o o t
l e n g t h ( c m )
N u m b e r o f
t r i f o l i a
t e s ( p l a n t − 1 )
40 60 80 100
Figure 6 Relationship between plant age and dry bean growth parameters (Fageriaand Santos, 2008).
The Role of Mineral Nutrition on Root Growth of Crop Plants 267

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 19/82
achieved at 80 days after sowing. However, root dry weight increasedquadratically with plant age, much like shoot growth. The youngestgrowing portions of the roots are most active in water and mineral nutrient
uptake; therefore, the root system must continually grow to provideadequate nutrition for the plant (Brown, 1984).
7. Root–Shoot Ratio
The partitioning of photoassimilate between roots and shoots hasfrequently been analyzed as a balance between root and shoot activity(Brouwer, 1966; Davidson, 1969a; Werf, 1996). Different plant species
may have different patterns for photosynthate transportation and allocationto shoot and root (Dyer et al ., 1991; Freckman et al ., 1991). There is aninterdependence of shoot and root for growth and development. The shootrelies on the root for water and nutrients, while the roots depend on theshoot for carbohydrates (Hoad et al ., 2001). The terms “shoot” and “root”are used here in a botanical sense and refer, respectively, to the entire aerialand subterranean portions of higher seed plants (Aung, 1974). In the earlypart of the twentieth century, shoot–root ratios were used rather extensivelyto characterize plant response to imposed nutritional changes. Root growth
is closely related to the whole plant growth. This relationship is called“allometry” or relative growth. Root dry weight is related to the total dryweight of a plant using the following equation ( Yoshida, 1981):
W R ¼ HW hT;
where W R is the root dry weight, W T is the total dry weight (shootdry weight þ root dry weight), and H and h are constants. The aboverelationship has been tested for different rice cultivars grown under variousenvironmental conditions, and can be expressed by the following equation( Yoshida, 1981):
W R ¼ 0:212W 0:936T :
When plants are small (substitute 1 for W T), W R/W T is 0.2; W R/W Tvalues approach 0.1 as plants grow larger (substitute 105 for W T). In other words, ratio of root dry weight to total dry weight ranges from 0.2 at theseedling stage to 0.1 at the reproductive stage (heading) for rice ( Yoshida,1981).
The above relationship between root and total dry weights gives anestimate of root mass that remains in soil if shoot weight is known. For example, when plants produce shoot dry weights of 3 Mg ha1 at heading,
268 N. K. Fageria and A. Moreira

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 20/82
root dry weights remaining in soil should be 330 kg ha1. Partitioning of dry matter in roots relative to shoots is high during the seedling stages of growth and steadily declines throughout development (Evans and Wardlaw,
1976). The shoot:root biomass ratio changes during ontogeny, generallybecoming high as the plant approaches flowering, and stabilizing after flowering (Zobel, 1986). Shoot-to-root ratios of common bean, rice,wheat, and cowpea increased as plants advanced in age (Fageria, 1992).Increases in shoot-to-root ratios indicate that shoots have a higher priorityfor photosynthate accumulation than roots. If shoot–root ratios decreasewith time, roots have preferential utilization of photosynthates under theexisting plant growth conditions. A relatively high conservation of photo-assimilate in shoots may increase the plant’s photosynthetic leaf area while
decreasing root biomass and the plant’s capacity for water and nutrientuptake (Werf, 1996).Environmental stresses increase the relative weights of roots compared to
shoots (Eghball and Maranville, 1993). Decrease in the availability of N, P,or water increased root–shoot ratios of perennial ryegrass (Lolium perenne L.)(Davidson, 1969b). Although deficiencies of many mineral elements influ-ence plant growth and root–shoot relationships, invariably water and Ndeficiency limit shoot growth the most. Root–shoot ratios of 28-day-oldmaize plants were 0.27, 0.15, and 0.18 at volumetric soil moisture contentsof 0.22, 0.27, and 0.32 m3 m3, respectively (Mackay and Barber, 1985).When plants are N-deficient, relatively more photosynthate is used by rootsas they develop greater length to aid the plant in obtaining more N. Ingeneral, when low nutrient levels do not reduce maize grain yield by morethan 20%, addition of N will reduce total root weights even though shootweights increase (Barber, 1995). Champigny and Talouizte (1981) reportedthat under N deprivation, translocation of photoassimilates from shoots toroots increased because of increased sink strength of roots compared toshoot sinks. In an experiment with 18-day-old wheat seedlings deprived of N for 7 days, soluble sugar contents in roots were higher than in thecorresponding roots of seedlings grown continuously with complete nutri-ent solutions (Talouizte et al ., 1984). Similarly, root–shoot ratios of maizeplants were higher when grown with low soil N compared to adequate N(Eghball and Maranville, 1993).
Soil salinity is another important soil chemical property that influencesshoot–root ratios. The depressing effect of salinity on root growth isgenerally less severe than its effect on shoot growth. Shalhevet et al .(1995) summarized the results of 10 experiments relating shoot and rootgrowth to salinity. In all the experiments, the root and shoot responses wereevaluated by measuring fresh or dry weights at the end of the experimentalperiods. All the 10 experiments produced either the same or stronger growth responses of shoots than roots because of the imposed osmoticpotential. However, Slaton and Beyrouty (1992) observed shoot–root ratios
The Role of Mineral Nutrition on Root Growth of Crop Plants 269

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 21/82
of rice remained constant as a result of a functional equilibrium in whichshoot growth was proportional to root growth.
Partitioning of photosynthates and their effects on dry matter distribu-
tion is influenced by several environmental factors such as low temperature,drought, and mineral nutrient deficiency (Wardlaw, 1990). The mineralnutrients P and N exerted pronounced influences on photosynthate and drymatter partitioning between shoots and roots (Costa et al ., 2002). Phospho-rus and N-deficient plants usually produce proportionately more dry matter to roots than shoots, compared with unstressed plants. This probably resultsfrom higher export rates of photosynthate to roots in deficient plants. Leaf expansion is highly sensitive to low tissue P concentrations, producinghigher concentrations of sucrose and starch in P-deficient leaves because
of reduced demand (Fredeen et al ., 1989). Thus, roots become morecompetitive for photosynthates than shoots, which leads to higher exportof carbohydrates to roots with correspondingly lower shoot–root ratios(Rufty et al ., 1993). Cakmak et al . (1994) reported that dry matter partition-ing between shoots and roots of common bean was affected differently bylow supplies of P, K, and Mg. Although total dry matter production wassomewhat similar in P-, K-, and Mg-deficient plants, K- and especially Mg-deficient plants had greater than normal shoot–root ratios, while P-deficientplants had smaller than normal shoot–root ratios (Cakmak et al ., 1994).Shoot–root dry weight ratios were 1.8 in P-deficient, 4.9 in control, 6.9 inK-deficient, and 10.2 in Mg-deficient plants. Upland rice usually has highroot/shoot weight ratios than lowland rice, an adaptation improving accessto soil water (Dingkuhn and Kropff, 1996).
8. Root Growth Versus Crop Yield
Roots are responsible for absorption of water and nutrients which areimportant resources affecting crop yields. In addition, roots improve soilorganic matter content and biological activity in the rhizosphere. Rootlength and root dry weight are standard root parameters that are measuredin many studies, largely because they are more easily determined than other root system properties (Gregory, 1994). Barber and Silberbush (1984)studied the relationship between root length and soybean yield and con-cluded that yield was significantly related to total root length at the R6 (fullseed) stage. These authors concluded that root growth is important indetermining the nutrient supply to the shoot which, in turn, affects crop
yield. Similarly, Thangaraj et al . (1990) reported that root length density of lowland rice at flowering was directly proportional to grain yield. Leon andSchwang (1992) used the grid intercept method (Newman, 1966) to evalu-ate differences in total root length between cultivars of oats and barley and
270 N. K. Fageria and A. Moreira
8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 22/82
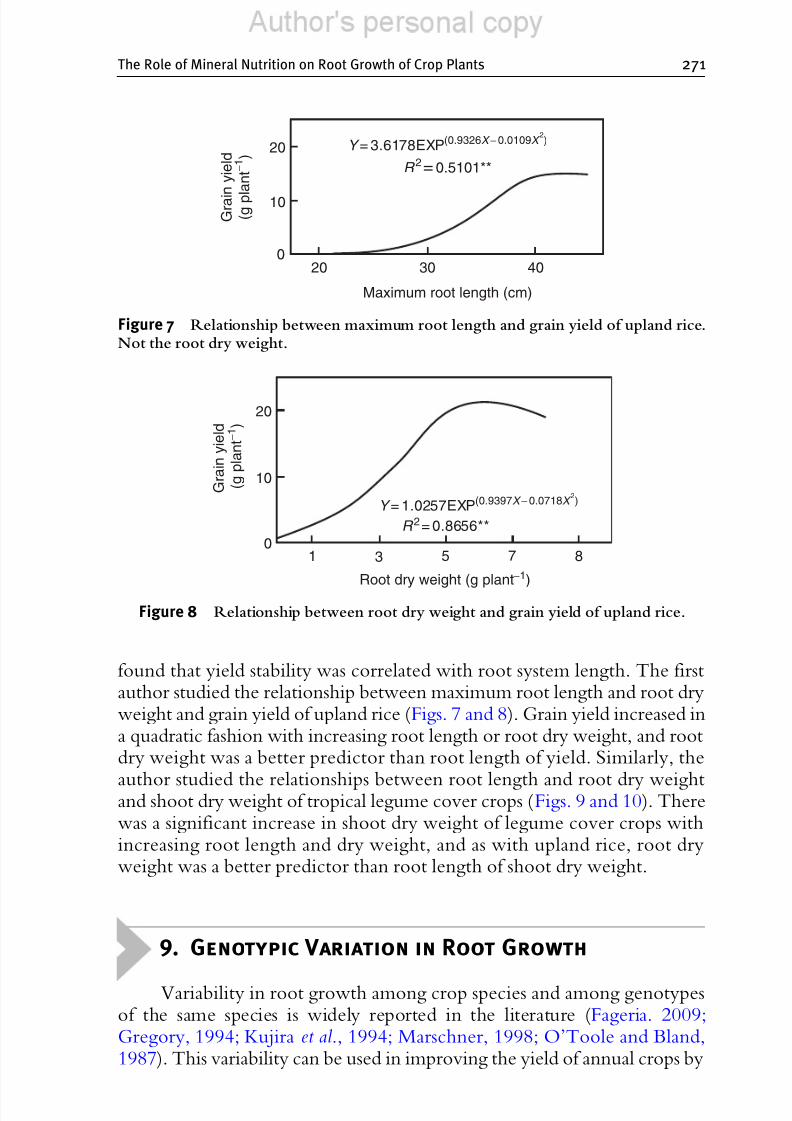
found that yield stability was correlated with root system length. The firstauthor studied the relationship between maximum root length and root dryweight and grain yield of upland rice (Figs. 7 and 8). Grain yield increased ina quadratic fashion with increasing root length or root dry weight, and rootdry weight was a better predictor than root length of yield. Similarly, the
author studied the relationships between root length and root dry weightand shoot dry weight of tropical legume cover crops (Figs. 9 and 10). Therewas a significant increase in shoot dry weight of legume cover crops withincreasing root length and dry weight, and as with upland rice, root dryweight was a better predictor than root length of shoot dry weight.
9. Genotypic Variation in Root Growth
Variability in root growth among crop species and among genotypesof the same species is widely reported in the literature (Fageria. 2009;Gregory, 1994; Kujira et al ., 1994; Marschner, 1998; O’Toole and Bland,1987). This variability can be used in improving the yield of annual crops by
200
10
20 Y = 3.6178EXP(0.9326X −0.0109X 2)
R 2= 0.5101**
Maximum root length (cm)
G r a i n y i e l d
( g p l a n t − 1 )
30 40
Figure 7 Relationship between maximum root length and grain yield of upland rice.Not the root dry weight.
Y = 1.0257EXP(0.9397X −0.0718X 2)
R 2= 0.8656**
87510
Root dry weight (g plant−1)
G r a i n y i e l d
( g p l a n t − 1 )
10
20
3
Figure 8 Relationship between root dry weight and grain yield of upland rice.
The Role of Mineral Nutrition on Root Growth of Crop Plants 271

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 23/82
incorporating vigorous root growth into desirable cultivars. Vigorous rootgrowth is especially important when nutrient and water stress are significant(Gregory, 1994). Ludlow and Muchow (1990), in their review of traitslikely to improve yields in water-limited environments, place a vigorousrooting system high in their list of properties to be sought.
O’Toole and Bland (1987) reviewed genotypic variation in root growthof annual crops and reported significant differences in rooting depths,maximum root length, and distribution pattern in the soil profile. Hurd(1974) and Yoshida and Hasegawa (1982) reported rooting depth differ-ences among genotypes of wheat (Triticum aestivum L.) and rice,
0
1
2
3
4
5
6
7
8 Y = 2.9808−0.2634X + 0.0076X 2
R 2= 0.8783**
10 20Maximum root length (cm)
S h o o t d r y w e i g h t ( g p l a n t − 1 )
30 40 50
Figure 9 Relationship between maximum root length and shoot dry weight of tropical legume cover crops. Values are averages of 14 tropical legume cover crops.
0
1
2
3
4
5
6
7
8 Y = 0.3422EXP(5.5187X −2.7205X 2)
R 2= 0.9712**
Root dry weight (g plant−1)
S h o o t d r y w e i g h t
( g p l a n t − 1 )
0.2 0.4 0.6 0.8 1.0 1.2
Figure 10 Relationship between root dry weight and shoot dry weight of tropicallegume cover crops. Values are averages of 14 tropical legume cover crops.
272 N. K. Fageria and A. Moreira

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 24/82
respectively. Similarly, Brown et al . (1987) and Pan et al . (1985) alsoreported differences in rooting depths of corn and barley (Hordeum vulgare L.), respectively. Similar variability exists in dicotyledonous species where
most work has been undertaken on legumes, including dry bean (Fageria,2002a; White and Castillo, 1989), soybean (Taylor et al ., 1978; Zobel, 2005a),chickpea (Cicer arietinum L.; Vincent and Gregory, 1986), peanut (Mathewset al ., 1988), and white clover (Trifolium repens L.; Caradus, 1990). Hamblin andTennant (1987) compared root growth and water uptake of wheat and lupin(Lupinus albus L.) and Gregory and Brown (1989) did similar work with barleyand chick pea. These authors concluded that there were differences betweenthe species in the root length necessary to extract water, and the rate of water extraction was greater in legumes than in cereals. Fageria (1991) studied root
dry weight of the Brazilian upland rice cultivars IAC 47 and the InternationalRice Research Institute (IRRI) cultivar IR 43 and concluded that root dryweight of Brazilian cultivars was almost double that of the IRRI cultivar during the entire growth cycle. Xiaoe et al . (1997) reported that hybrid ricehas a more vigorous root system, larger panicle, and more grains per paniclethan traditional rice cultivars.
The genotypic variability in the root growth of annual crops has beenused to identify superior genotypes for drought-prone environments(Gregory, 1994; Hurd et al ., 1972). Gregory and Brown (1989) reviewedthe role of root characters in moderating the effects of drought and con-cluded that roots may have a direct effect, by increasing the supply of water available to the crop, or an indirect effect by changing the rate at which thesupply becomes available. Where crops are grown on deep soils and water isstored throughout the whole soil profile, the depth of rooting has a major influence on the potential supply of water (Gregory, 1994). Rain mayreplenish the upper soil during the season, but later growth and grain fillingin many crops are accomplished during periods of low rainfall when soilmoisture stored deep in the profile must be utilized. Sponchiado et al . (1989)reported that in dry bean, drought avoidance results from root growth andsoil water extraction deep in the profile.
Atkinson (1990) reported significant variation in the speed of rootpenetration, specific root length, branching pattern, root density, totalroot mass, and root hair development of 25 spring barley cultivars. Infor-mation reported in the literature on old and new cereal cultivars indicatesthat more modern cultivars are more responsive than older cultivars to highnutrient availability (Haberle, 1993; Haberle et al ., 1995), although theytend to have a lower root fraction (Wahbi and Gregory, 1995). Rootphysiological characteristics also differ among cultivars and can affect pro-cesses like nutrient acquisition (Hoad et al ., 2001; Marschner, 1998). Therate of uptake of nutrient per unit root length depends on the nutrientavailability but also varies considerably among cultivars (Hoad et al ., 2001;Romer, 1985).
The Role of Mineral Nutrition on Root Growth of Crop Plants 273

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 25/82
10. Root Oxidation Activity in Oxygen-DeficientSoils
Roots require O2 for respiration, water and mineral absorption, andother metabolic activities. Brailsford et al . (1993) reported that roots asphyx-iate within a few hours or days at normal growing temperatures unless atleast a small amount of oxygen (0.1 kPa in a flowing gas phase) is available.The O2 used by roots is replaced largely by molecular diffusion between soilair and the aboveground atmosphere (Miller, 1986). Waterlogging, definedas the soil saturated or nearly saturated with water, significantly reducesoxygen availability to roots, causes the soil to become “reduced,” results in
the production of toxic compounds, and generally reduces root growth(Ellis, 1979; Hoad et al ., 2001). Cereals are generally more tolerant towaterlogging than legumes. Hoad et al . (2001) reported that cereal rootscan survive short periods of waterlogging without adverse effects becausethey can use the small quantities of oxygen dissolved in the soil water andare capable of anaerobic metabolism when oxygen demand exceeds supply.Oxygen requirements are higher for growing root tips than for the older parts of the root system. The effect of waterlogging is least at low tempera-tures when more oxygen can be dissolved in the water and biological
activity is depressed (Hoad et al ., 2001). The effect of waterlogging isreduced with the application of chemical fertilizers, especially K, whichcan improve the respiration capacity of roots (Fageria, 2009).
Most agricultural crops are mesophytes, which, for maximum growth,require an environment that is neither too wet nor too dry (Fageria, 1992).However, some plant species grow well under anaerobic conditions. It haslong been known that marsh plants or hydrophytes, such as rice, aregenetically adapted to grow in reduced soil environments (Horiguchi,1995). Adaptation to waterlogging in hydrophytes is the result of their
unique ability to translocate O2 from the shoot to the root system. Andoet al . (1983) reported that in rice oxygen absorbed from the atmosphere bythe shoots can be translocated to the root system and released into therhizosphere within 5 min. Generally, flooded rice roots have a reddishbrown color due to oxidation of the Fe ions by oxygen release from theroots and their deposition on the root surface (Fageria et al ., 2008a,b).
Flooding induces many changes in plant roots, of which formation of aerenchyma (large interconnected intercellular spaces) is an important adap-tive mechanism (Laan et al ., 1989). Some species like corn (Z. mays L.)
develop aerenchyma as a response to flooding or anoxia (Armstrong andDrew, 2002), a facultative development, while others develop them rou-tinely regardless of the environmental conditions (constitutive develop-ment; Barlow, 2002; Zobel, 2005a). The development of aerenchyma in
274 N. K. Fageria and A. Moreira

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 26/82
rice (O. sativa L.), and other species and crops growing in wet soils, allowsthe roots to grow into saturated soils and still extract nutrients (Zobel,2005b). In corn, the leaf bases also form aerenchyma when submerged,
while the enclosed stem bearing aerenchymatous nodal (adventitious) rootsmay elongate more rapidly ( Jackson, 1994). Aerenchyma formation in cornroots appears to be triggered by the loss of tonoplast integrity (Campbell andDrew, 1983), although cell wall disintegration is also involved at an earlystage, at least in rice ( Jackson, 1994; Webb and Jackson, 1986). Aerench-
ymatous roots are formed either by some cell wall separation and cellcollapse (lysigeny) or by cell separation without collapse (schizogeny).Figure 11 shows cross sections of rice and wheat roots, and lysigenous andschizogenous intercellular space. Both forms result in large longitudinal
channels in root cortices, and such structures enhance diffusion of atmo-spheric or photosynthetic oxygen from shoots to roots so that aerobicrespiration and growth can be maintained (Armstrong, 1979). Changes inroot morphology occur after flooding for both wetland and nonwetlandplant species. Flooding may also increase branching of roots, developmentof new adventitious roots, and superficial rooting (Laan et al ., 1989).
Cellular spaces that exist in roots facilitate oxygen diffusion; however,the amount of aeration varies greatly among plant species. In the case of certain marsh plants like rice, root cortical cells are arranged in columns, andwhen channels form, the spaces become large and continuous, facilitatingdiffusion. However, in the case of certain terrestrial plants, the cellular arrangement is oblique and the spaces formed are small (Horiguchi, 1995).There are marsh plants, however, that do not display columnar forms, whilesome terrestrial plants do (Horiguchi, 1995). In the case of rice, largecortical aerenchyma spaces develop schizogenously and lysigenously evenin well-aerated soils. Horiguchi (1995) reported that in rice and other graminaceous plants, aerenchymous cells are well developed in both thenodes and the internodes. The oxidizing capacity of roots of crop plants alsodepends on the soil fertility level. Higher soil fertility improves the oxida-tion activity of rice roots (Horiguchi, 1995). Application of silicon to rice
RiceWheatLysigenous
intercellular
space
Schizogenous
intercellular
space
A B
Figure 11 Cross section of a rice (A) and wheat (B) root showing lysigenous andschizogenous intercellular spaces (from Horiguchi, 1995).
The Role of Mineral Nutrition on Root Growth of Crop Plants 275

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 27/82
crops increases the diffusion of oxygen from the tops to the roots, strength-ening oxidation activity and the deposition of ferrous iron on root surfaces,and restricting the transport of excess iron to the top (Horiguchi, 1995).
Development of adventitious roots in some crops is related to planttolerance of soil waterlogging. Such roots developed by corn after oneday of flooding were short and porous enough to allow significant O2
movement from the shoots to the root meristems (Miller, 1986; Wenkertet al ., 1981). Despite this adventitious root formation and increased rootporosity, corn and most other crop plants soon perish under such conditions(Miller, 1986).
11. Root Growth in ConservationTillage Systems
Conservation tillage is defined as any tillage sequence, the object of which is to minimize or reduce loss of soil and water; operationally, a tillageor tillage and planting combination that leaves a 30% or greater cover of crop residue on the surface (Soil Science Society of America, 2008). Thebenefits of conservation tillage are reducing soil erosion, conserving soil
moisture, avoiding fluctuations of soil temperature in the arable soil depth,and reducing the costs of soil preparation. In addition, the use of conserva-tion tillage is being encouraged as part of a strategy to reduce C loss fromagricultural soils (Kern and Johnson, 1993). Decomposition rates are gener-ally slower in no-till than conventional tillage in which the decompositionof soil organic matter is promoted by the stirring of the soil and alterations inthe soil microclimate (Parton et al ., 1996). Holland and Coleman (1987)suggested that C retention is increased in no-till because the surface residueis primarily decomposed by fungi, which have higher assimilation efficiencythan the bacteria, which dominate the decomposition processes of residuemixed into the soil. Gale and Cambardella (2000) reported that there was aclear difference in the partitioning of surface residue and root-derived Cduring decomposition and imply that the beneficial effects on no-till on soilorganic C accrual are primarily due to the increased retention of root-derived C in the soil. Knowledge of how plant root systems grow under conservation tillage is important because this practice is widely adopted inmany countries around the world, most notably, in countries such as UnitedStates, Brazil, Argentina, Canada, and Australia (Bolliger et al ., 2006). Forty-five percent of the total cultivated land in Brazil is now estimated to bemanaged with conservation tillage, although in southern Brazil, this figure isreported to exceed 80% (Bolliger et al ., 2006).
Conservation tillage reduces soil erosion, conserves soil moisture, con-serves energy, increases soil organic matter content, and consequently, soil
276 N. K. Fageria and A. Moreira

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 28/82
quality. However, conservation tillage may compact surface soil horizons andmay lead to poor root growth. Adverse effects of soil compaction on cropproduction have been recognized for many years. Cato the Elder (234–149 B.C.)
wrote that the first principle of good crop husbandry is to plow well and thesecond principle is to plow again, presumably to provide a “mellow” (well-aerated) seedbed (Unger and Kaspar, 1994). Conservation tillage increasessoil bulk density (Martino and Shaykewich, 1994) which may inhibit rootgrowth in the upper part of the soil profile (Cannell, 1985; Lampurlaneset al ., 2001), reducing nutrient uptake and plant growth (Peterson et al .,1984). Qin et al . (2004) reported that wheat root length density, mean rootdiameter, and percentage of small-diameter roots were lower in no-till thanconventional tillage. Root diameter may be indicative of the effects of soil
strength on root growth and affects the utilization of nutrients in the soil.Sidiras et al . (2001) reported thicker barley roots under conventional tillagethan under no-till. In general, bulk densities that impede root growth are1.55 Mg m3 for clay loams, 1.65 Mg m3 for silt loams, 1.80 Mg m3 for sandy loams, and 1.85 Mg m3 for loamy fine sands (Miller, 1986).
Tillage-induced differences in the soil nutrient status may also have asignificant impact on root growth (Qin et al ., 2004). Conservation tillageoften results in the stratification of soil nutrients, especially of immobilenutrients like P (Crozier et al ., 1999; Holanda et al ., 1998; Logan et al .,1991). This produces greater soil fertility near the soil surface which, incontrast to the effects of compaction described above, causes an increase inroot length density near the soil surface under conservation tillage (Cannelland Hawes, 1994; Gregory, 1994). Frequently, root growth is greater from0 to 5 cm in conservation and no-tillage systems than in conventional tillagesystems (Chan and Mead, 1992; Rasmussen, 1991; Wulfsohn et al ., 1996).
Radial root swelling has been reported for lupins (Lupinus angustifoliusL.) grown in compacted soil (Atwell, 1989), for barley (H. vulgare L.) under mechanical impedance (Wilson et al ., 1977), and for mustard (Brassica sp.) indrying soil (Vartanian, 1981). Studies on root elongation of cotton as afunction of soil strength and soil water content showed that root elongationis more sensitive to soil strength than water content (Taylor and Ratliff,1969). Root volumes were less affected than length, indicating an increasein root diameter (Ball et al ., 1994). Chassot et al . (2001) reported thatconservation tillage decreases soil temperature, and this may be the mainreason for the poor growth of the roots and shoots of corn seedlingscompared with conventional tillage under temperate humid conditions.
Considering the many advantages of conservation tillage compared toconventional tillage, the effects of soil compaction produced by conserva-tion tillage can be minimized. Unger and Kaspar (1994) reported thatgrowing deep-rooted crops in rotation will help avoid or alleviate compac-tion, improving root distribution and increasing rooting depth. Theseauthors also reported that weather conditions and soil moisture can enhance
The Role of Mineral Nutrition on Root Growth of Crop Plants 277

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 29/82
or diminish the effects of compaction on root growth. Even if compactionlimits root growth, subsequent weather events may either enhance or diminish the effect of the root limitation on crop growth. The first author
has conducted field experiments on an upland rice-dry bean rotation usingconservation tillage on Brazilian Oxisols. The upland rice root system wasadversely affected, and rice yields were low (around 2000 kg ha1), abouthalf that expected in a field experiment with conventional soil preparationand favorable environmental conditions. However, dry bean yields weregreater than 3000 kg ha1 (Fageria, 2008; Fageria and Stone, 2004). Hence,selecting the appropriate crop is important.
12. Mineral Nutrition Versus Root Growth
There are 17 nutrients essential for plant growth and development andthese are carbon (C), hydrogen (H), oxygen (O), nitrogen (N), phosphorus(P), potassium (K), calcium (Ca), magnesium (Mg), sulfur (S), zinc (Zn),copper (Cu), manganese (Mn), iron (Fe), boron (B), molybdenum (Mo),chlorine (Cl), and nickel (Ni). Among these nutrients, plants take C, H, andO from air and soil water and the remaining from the soil solution. About
95% of the plant weight is C, H, and O, and the remaining 5% is the other 14 nutrients. The C, H, O, N, P, K, Ca, Mg, and S are required by plants inlarge amounts and for this reason are known as major or macronutrients.The remaining nutrients, Zn, Cu, Mn, Fe, B, Mo, Cl, and Ni, are classifiedas micronutrients because they are required in small amounts by plants(Fageria et al ., 2002). Among the micronutrients, Cl is also absorbed byplants in large amounts, but it is needed in only small amounts, and Cldeficiency is rarely observed in crop plants.
All the 17 nutrients are equally important for plant growth. If any of these nutrients is limiting in the growth medium, plant growth will bereduced. An example is given in Fig. 12, where at an adequate fertility level,the relative dry weight of dry bean was 100%. When essential nutrients wereomitted from the soil or were not applied, growth was reduced, but theamount of reduction depended on the nutrient. The impacts of deficiencieson growth were in the order of P > Ca > Mg > N ¼ K > S amongmacronutrients and B > Zn > Cu > Fe > Mn > Mo among micronu-trients. Similarly, the influence of N, P, and K on shoot and root growth of dry bean is shown in Fig. 13. Both shoot and root growth were significantlyreduced when N, P, or K were omitted from the growth medium.Figures 14 and 15 show root growth of wheat, upland rice, and corn withthe addition of N þ P þ K and with the omission of these nutrients fromthe growth medium. Root growth of these three crop species was decreasedin the absence of N, P, and K in the soil. A significant amount of variation
278 N. K. Fageria and A. Moreira

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 30/82
exists, both within and among crop species, in nutrient acquisition and use.This variability reflects differences in root morphology and mechanisms thateither aid or prevent ion movement into the root (Gabelman et al ., 1986).
12.1. NitrogenNitrogen is one of the most yield-limiting nutrients in crop production inmost agroecosystems. Nitrogen plays numerous key roles in plant biochem-istry, including being an essential constituent of enzymes, chlorophyll,
A F L
− N
− P
− K
− C a
− M
g
− S
− Z n
− F e
− C u
− B
− M o
− M n
0
20
40
60
R e l a t i v e s h o o t d r y w e i g h t ( % )
80
100
120
Figure 12 Relative shoot dry weight of dry bean as influenced by adequate fertilitylevel (AFL) and other nutrients were not applied or omitted from the Oxisol.
N + P + K −N0
20
40
60
80
100
120
R e l a t i v e d r y w e i g h
t ( % )
−P −K
Shoot Root
Figure 13 Relative dry weight of shoot and root of dry bean as influenced by N, P, andK fertilization.
The Role of Mineral Nutrition on Root Growth of Crop Plants 279

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 31/82
nucleic acids, storage proteins, cell walls, and a vast array of other cellular components (Harper, 1994). Consequently, a deficiency of N in crop plantsprofoundly influences plant growth, development, and yield. The recovery
Wheat
N+P+K −N −P −K
Upland
rice
Figure 14 Root growth of wheat at the top and upland rice at the bottom grown atN þ P þ K and N, P, and K levels.
Corn
N + P + K −N −P −K
Figure 15 Corn root growth at N þ P þ K and N, P, and K levels.
280 N. K. Fageria and A. Moreira

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 32/82
of applied N with chemical fertilizers is lower than 50% for most annualcrops. The low recovery of N is associated with loss of this elementby leaching, denitrification, volatilization, incorporation into soil micro-
organisms, and soil erosion (Fageria and Baligar, 2005). To improvethe efficiency of N uptake and use by crop plants, root systems play animportant role.
Root morphology is influenced by the amount of N fertilizer applied(Eghball et al ., 1993) and factors such as temperature (Feil et al ., 1991) andsoil mechanical impedance (Bengough and Mullins, 1990). Eghball et al .(1993) showed that N stress in corn reduced root branching. Similarly,Maizlish et al . (1980) showed greater root branching in corn with increasinglevels of applied fertilizer N. Costa et al . (2002) reported that greater root
length and root surface area were obtained at an N fertilizer rate of 128 kg N ha1 compared with either the absence of fertilizer N or thehigher rate of 255 kg N ha1. Nitrogen fertilizer improves root growth insoils having low-OM content (Gregory, 1994; Robinson et al ., 1994).Nitrogen fertilization may increase crop root growth by increasing soil Navailability (Garton and Widders, 1990; Weston and Zandstra, 1989). Sainjuet al . (2001) observed that tomato (Lycopersicon esculentum Mill.) root growthwas greater with hairy vetch and crimson clover cover crops and90 kg N ha1 than with no cover crops or N fertilization. Nitrogen alsoimproves production of lateral roots and root hairs, as well as increasingrooting depth and root length density deep in the profile (Hansson andAndren, 1987). Hoad et al . (2001) reported that surface application of nitrogen fertilizer increases root densities in the surface layers of the soil.
Nitrogen fertilization can increase root length and root surface area anddecrease root mass per unit area of corn (Anderson, 1987; Costa et al ., 2002).It is well known that roots tend to proliferate in nutrient-enriched soil zones(Drew et al ., 1973; Qin et al ., 2005). Russell (1977) refers to this as acompensatory response. The results of pot experiments showed that cornroots were longer and thinner in zones that were rich in N (Durieux et al .,1994; Zhang and Barber, 1992, 1993).
Root mass was less affected by N than root length, but the effect maydepend on the stage of maturity of the crop (Baligar et al ., 1998). Higher rates of application of N reduced root growth and depth of rooting in wheat(Comfort et al ., 1988) and reduced root:shoot ratio in rye (Brouwer, 1966).In corn, the primary root system was 16% thicker when NH4
þ –N wasapplied rather than NO3 –N. The NH4
þ –N treatment also increased thediameters of lateral and first- and second-order nodal roots (Anderson et al .,1991). Baligar et al . (1998) reported that relative dry weights of roots of rice,dry bean, corn, and soybean were reduced by 38%, 56%, 35%, and 11%,respectively, when N was omitted from a complete fertilizer. Nitrogendeficiency also reduces branching and root hairs in cereals and legumes(Baligar et al ., 1998).
The Role of Mineral Nutrition on Root Growth of Crop Plants 281
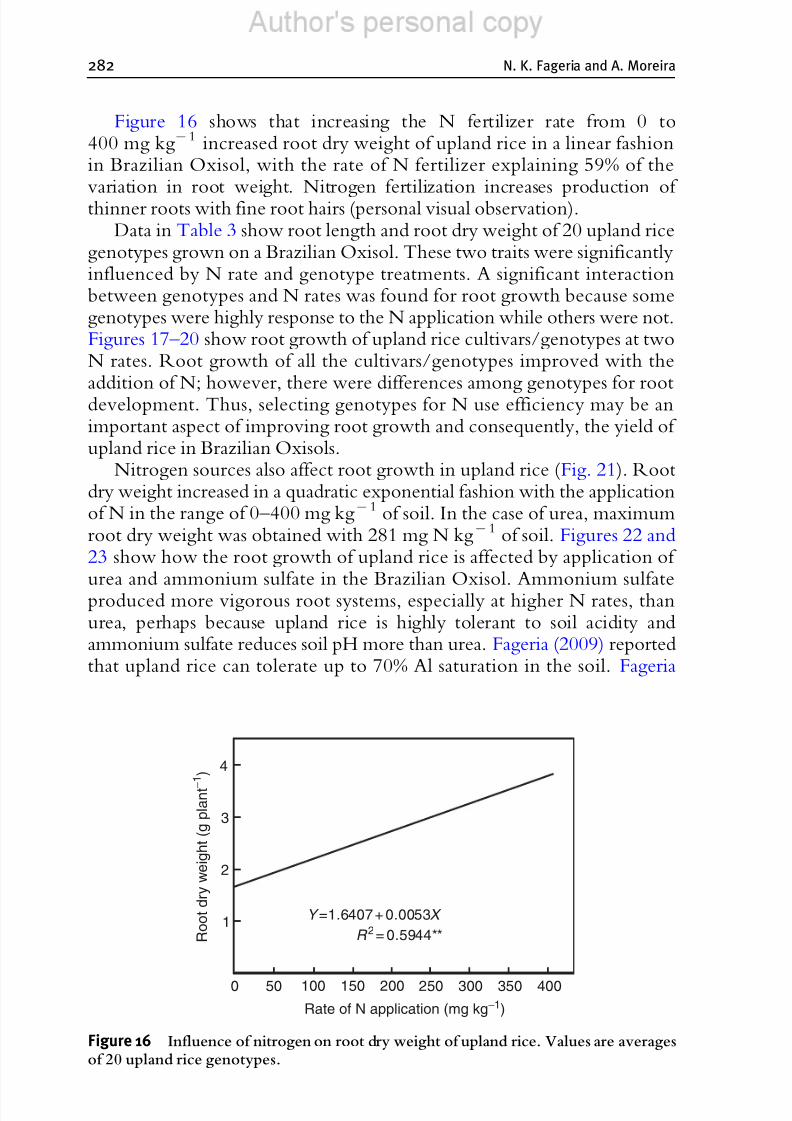
8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 33/82
Figure 16 shows that increasing the N fertilizer rate from 0 to400 mg kg1 increased root dry weight of upland rice in a linear fashionin Brazilian Oxisol, with the rate of N fertilizer explaining 59% of the
variation in root weight. Nitrogen fertilization increases production of thinner roots with fine root hairs (personal visual observation).Data in Table 3 show root length and root dry weight of 20 upland rice
genotypes grown on a Brazilian Oxisol. These two traits were significantlyinfluenced by N rate and genotype treatments. A significant interactionbetween genotypes and N rates was found for root growth because somegenotypes were highly response to the N application while others were not.Figures 17–20 show root growth of upland rice cultivars/genotypes at twoN rates. Root growth of all the cultivars/genotypes improved with the
addition of N; however, there were differences among genotypes for rootdevelopment. Thus, selecting genotypes for N use efficiency may be animportant aspect of improving root growth and consequently, the yield of upland rice in Brazilian Oxisols.
Nitrogen sources also affect root growth in upland rice (Fig. 21). Rootdry weight increased in a quadratic exponential fashion with the applicationof N in the range of 0–400 mg kg1 of soil. In the case of urea, maximumroot dry weight was obtained with 281 mg N kg1 of soil. Figures 22 and23 show how the root growth of upland rice is affected by application of urea and ammonium sulfate in the Brazilian Oxisol. Ammonium sulfateproduced more vigorous root systems, especially at higher N rates, thanurea, perhaps because upland rice is highly tolerant to soil acidity andammonium sulfate reduces soil pH more than urea. Fageria (2009) reportedthat upland rice can tolerate up to 70% Al saturation in the soil. Fageria
500
1
2
3
4
100 150 200
Y =1.6407+ 0.0053X
R 2= 0.5944**
Rate of N application (mg kg−1)
R o o t d r y w e i g h t ( g p l a n t −
1 )
250 300 350 400
Figure 16 Influence of nitrogen on root dry weight of upland rice. Values are averagesof 20 upland rice genotypes.
282 N. K. Fageria and A. Moreira

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 34/82
(2009) also reported that rice growth was better in 10 mg L1 Al than with0 mg Al L1 in nutrient solution. Brazilian rice cultivars (lowland as well asupland) are highly tolerant to soil acidity (Fageria et al ., 2004). Another possible explanation is that ammonium sulfate has about 24% S, which mayimprove root growth if the extractable soil S level is lower than 10 mg kg1.
The timing of nitrogen application can also influence the root growth of crop plants (Table 4). The treatment T 3, which produced maximum grain
yield, also produced minimum root length; treatment T 2 which producedminimum grain yield produced maximum root length. There was a
Table 3 Root length and root dry weight of 20 upland rice genotypes as influencedby nitrogen fertilization
Genotype
Root length (cm) Root dry weight (g plant1)
0 mg N kg1 300 mg N kg1 0 mg N kg1 300 mg N kg1
BRA01506 34.67ab 21.00cd 0.92a 0.40f
BRA01596 30.00b 15.67d 0.87a 0.45f BRA01600 27.00ab 25.33bcd 1.14a 1.03ef
BRA02535 31.67ab 30.00abcd 1.33a 3.25abcdBRA02601 28.00b 32.67abc 1.11a 3.73ab
BRA032033 30.00a 28.00abcd 1.12a 2.41cdBRA032039 43.00a 34.67abc 1.24a 3.77ab
BRA032048 28.50b 32.00abc 1.05a 3.62abcBRA032051 30.67ab 35.67ab 1.31a 2.82bcd
BRA042094 30.33b 38.00ab 1.78a 2.31de
BRA042156 29.33b 33.00abc 1.19a 2.84bcdBRA042160 29.67b 32.50abc 1.51a 3.33abcd
BRA052015 31.00ab 35.00abc 1.67a 2.83bcdBRA052023 29.67b 26.50abcd 1.48a 4.14a
BRA052033 30.33b 31.33abc 1.56a 3.38abcdBRA052034 29.00b 37.67ab 1.78a 2.58bcd
BRA052045 28.67b 37.67ab 1.75a 3.66abc
BRA052053 29.00b 36.50ab 1.65a 2.72bcdBRS Primavera 29.33b 36.33ab 1.68a 2.49bcdBRS Sertaneja 30.00b 40.33a 1.45a 2.68bcd
Average 30.49 31.99 1.38 2.72F -test
N rate (N) NS *Genotype (G) ** **
N G ** **CV(%) 14.25 15.86
Source: Fageria (2011).*,**Significant at the 5% and 1% probability levels, respectively. Means followed by the same letter in
the same column are not significant at the 5% probability level by Tukey’s test.
The Role of Mineral Nutrition on Root Growth of Crop Plants 283

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 35/82
N0
BRS Sertaneja
N300
Figure 17 Root growth of cultivar BRS Sertaneja at 0 and 300 mg N kg1 soil(Fageria, 2011).
N0
BRS Primavera
N300
Figure 18 Root growth of cultivar BRS Primavera at 0 and 300 mg N kg1 soil(Fageria, 2011).

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 36/82
negative association between root length and grain yield (Y ¼ 31.2041 0.2718X , R 2 ¼ 0.7396**). Root dry weight was significantly related tograin yield (Y ¼ 7.6345 þ 5.8030X 0.4081X 2, R 2 ¼ 0.8747**;Fageria, 2011). Figure 24 shows root growth of upland rice under differentN timing treatments. Root dry weight was least (treatment T 2) when all Nfertilizer was applied at planting. Root and shoot yields were better intreatments T 3 and T 4, when N was applied later in the growth cycle(Fageria, 2011). Better root growth may be responsible for higher absorp-tion of nutrients and water in the T 3 and T 4 treatments which resulted inhigher grain and straw yields.
The depth of N placement can influence NO3 –N distribution in the soiland root growth of wheat (Sharma and Chaudhary, 1984). Root lengthdensity decreased abruptly below the 15 cm depth when N was surfaceapplied, whereas root length density decreased more gradually below 15 cmwhen N was placed at the 10 cm depth. Drew (1975) reported that barleyroot weight increased in the zone of nutrient localization and decreased inthe deficient zone. Murphy and Zaurov (1994) reported that N fertilizationat the 5, 10, and 15 cm soil depths produced greater root mass than Nfertilization at 0 cm soil depth or surface fertilization.
BRA052053
N0 N300
Figure 19 Root growth of genotype BRA052053 at 0 and 300 mg N kg1 soil(Fageria, 2011).
The Role of Mineral Nutrition on Root Growth of Crop Plants 285

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 37/82
BRA052045
N0 N300
Figure 20 Root growth of genotype BRA0522045 at 0 and 300 mg N kg1 soil(Fageria, 2011).
0
1
2
4
3
5
50 100
Nitrogen application rate (mg kg−1)
R o o t d r y w e i g h t ( g p l a n t − 1 )
150
CO(NH2)2 (Y ) = 1.1194+0.0135X −0.000024X 2
(NH4)2SO4 (Y ) = 1.9132EXP(0.00093X + 0.000031X 2)
R 2 = 0.5296**
R 2= 0.4122**
200 250 300 350 400
Figure 21 Relationship between nitrogen application rate by ammonium sulfate andurea and root dry weight of upland rice (Fageria et al. 2011).
286 N. K. Fageria and A. Moreira

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 38/82
The effects of two N rates on root length and root dry weight of sixtropical legume cover crops are showed in Table 5. Root length as well asroot dry weight were significantly increased with the addition of N fertil-izer. Similarly, root length as well as root dry weight were significantlydifferent among cover crop species. Root growth of four legume cover crops is shown in Figs. 25–28. In all cases, root growth was better at thehigher N rate compared to low N rate.
12.2. Phosphorus
Phosphorus is one of the most yield-limiting nutrients in tropical highlyweathered soils (Fageria, 2009; Fageria and Baligar, 2003, 2008). Thedeficiency of P in these soils may be related to the low natural level of thiselement in these soils as well as to the immobilization of P in these soils(Fageria and Baligar, 2008). Phosphate plays many roles in the physiologyand biochemistry of pants. It is a component of important compounds like
0 N 50 N 300 N
CO(NH2)2
Figure 22 Root growth of upland rice at 0 (left), 50 (center), and 300 (right)mg N kg1 soil supplied with urea.
The Role of Mineral Nutrition on Root Growth of Crop Plants 287

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 39/82
(NH4)2SO4
300 N0 N 400 N
Figure 23 Root growth of upland rice at 0 (left), 300 (center), and 400 (right)mg N kg1 soil with ammonium sulfate.
Table 4 Root length and root dry weight of upland rice as influenced by nitrogentiming treatments
Nitrogen timing treatmenta Root length (cm) Root dry weight (g plant1)
T 1 28.75b 4.21bcT 2 35.00a 2.56cT 3 25.75b 5.74abT 4 27.00b 7.70aT 5 28.25b 4.98b
Average 28.95 5.04F -test ** **
CV(%) 8.7 20.9
Source: Fageria (2011).** Significant at the 1% probability level. Means followed by the same letter in the same column are not
significant at the 5% probability level by the Tukey’s test.a T 1 (1/2 N applied at sowing þ 1/2 applied at panicle initiation), T 2 (total N applied at sowing),
T 3 (1/3 N applied at sowing þ 1/3 N applied at active tillering þ 1/3 N applied at the panicleinitiation), T 4 (1/2 N applied at initiation of tillering þ 1/2 N applied panicle initiation), and T 5 (2/3N applied at sowing þ 1/3 N applied at panicle initiation).
288 N. K. Fageria and A. Moreira

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 40/82
T4T3T2T1 T5
Figure 24 Root growth of upland rice at harvest under N timing treatments.T 1 (1/2 N applied at sowing þ 1/2 applied at panicle initiation), T 2 (total N appliedat sowing), T 3 (1/3 N applied at sowing þ 1/3 N applied at active tillering þ 1/3 Napplied at the panicle initiation), T 4 (1/2 N applied at initiation of tillering þ 1/2 Napplied panicle initiation), and T 5 (2/3 N applied at sowing þ 1/3 N applied at panicleinitiation). Source: Fageria (2011).
Table 5 Influence of nitrogen rate on root length and root dry weight of six tropicallegume cover crops
Cover crops
Root length (cm) Root dry weight (g plant1)
0 mg N kg1 100 mg N kg1 0 mg N kg1 100 mg N kg1
Showy
crotalaria
22 29 0.48 0.77
Calopogonio 28 34 0.30 0.46
Pueraria 23 24 0.16 0.15
Pigeon pea 30 34 0.73 0.48Lablab 27 32 0.74 1.72Gray mucuna
bean
28 31 2.01 1.84
Average 26b 31a 0.74b 0.90aF -test
N rate (N) ** **Cover crops
(C)
** **
N C NS **CV(%) 6.47 16.27
**, NS Significant at the 1% probability level and nonsignificant, respectively. Means in the same line for each growth parameter, followed by the same letter are not significantly different by Tukey’s testat the 5% probability level.
The Role of Mineral Nutrition on Root Growth of Crop Plants 289

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 41/82
0 N 100 N
Black mucuna bean
Figure 25 Root growth of black mucuna bean tropical legume cover crop at 0 (left)and 100 (right) mg N kg1 soil.
0 N100 N
Gray mucuna bean
Figure 26 Root growth of gray mucuna bean tropical legume cover crop at 0 (left)and 100 (right) mg N kg1 soil.
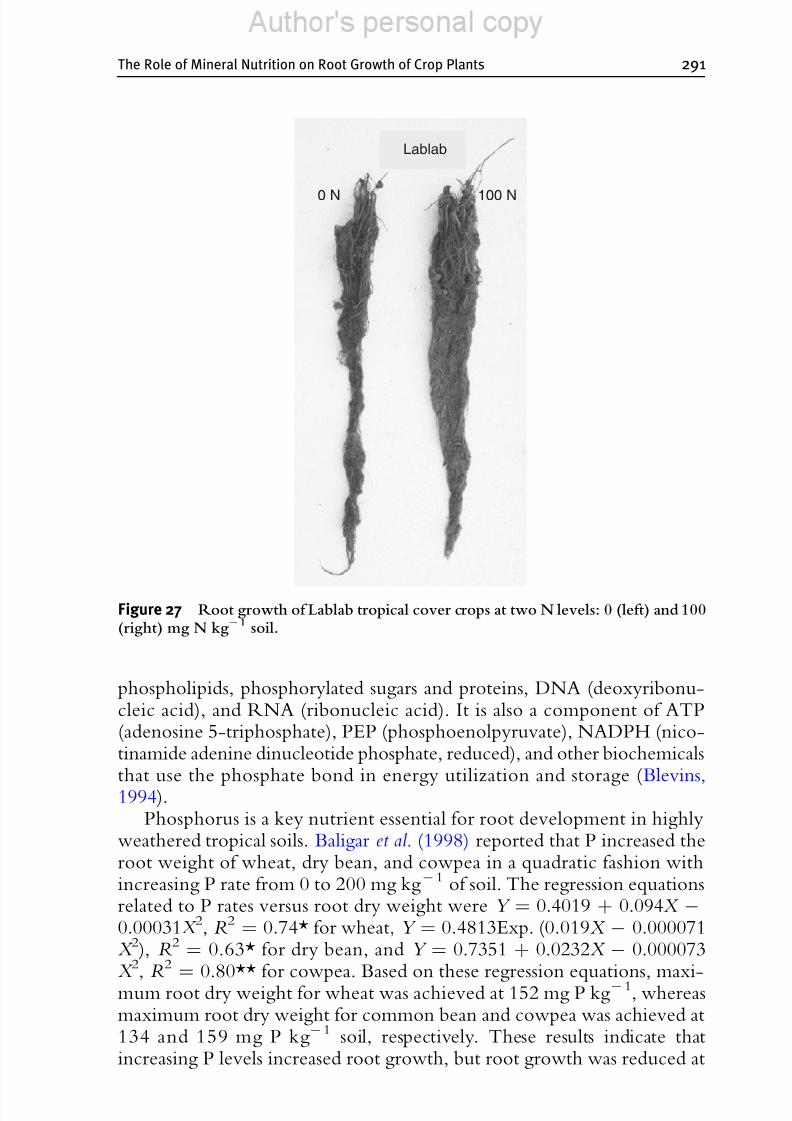
8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 42/82
phospholipids, phosphorylated sugars and proteins, DNA (deoxyribonu-cleic acid), and RNA (ribonucleic acid). It is also a component of ATP(adenosine 5-triphosphate), PEP (phosphoenolpyruvate), NADPH (nico-tinamide adenine dinucleotide phosphate, reduced), and other biochemicalsthat use the phosphate bond in energy utilization and storage (Blevins,1994).
Phosphorus is a key nutrient essential for root development in highlyweathered tropical soils. Baligar et al . (1998) reported that P increased theroot weight of wheat, dry bean, and cowpea in a quadratic fashion withincreasing P rate from 0 to 200 mg kg1 of soil. The regression equationsrelated to P rates versus root dry weight were Y ¼ 0.4019 þ 0.094X 0.00031X 2, R 2 ¼ 0.74* for wheat, Y ¼ 0.4813Exp. (0.019X 0.000071X 2), R 2 ¼ 0.63* for dry bean, and Y ¼ 0.7351 þ 0.0232X 0.000073X 2, R 2 ¼ 0.80** for cowpea. Based on these regression equations, maxi-mum root dry weight for wheat was achieved at 152 mg P kg1, whereasmaximum root dry weight for common bean and cowpea was achieved at134 and 159 mg P kg1 soil, respectively. These results indicate thatincreasing P levels increased root growth, but root growth was reduced at
0 N 100 N
Lablab
Figure 27 Root growth of Lablab tropical cover crops at two N levels: 0 (left) and 100(right) mg N kg1 soil.
The Role of Mineral Nutrition on Root Growth of Crop Plants 291
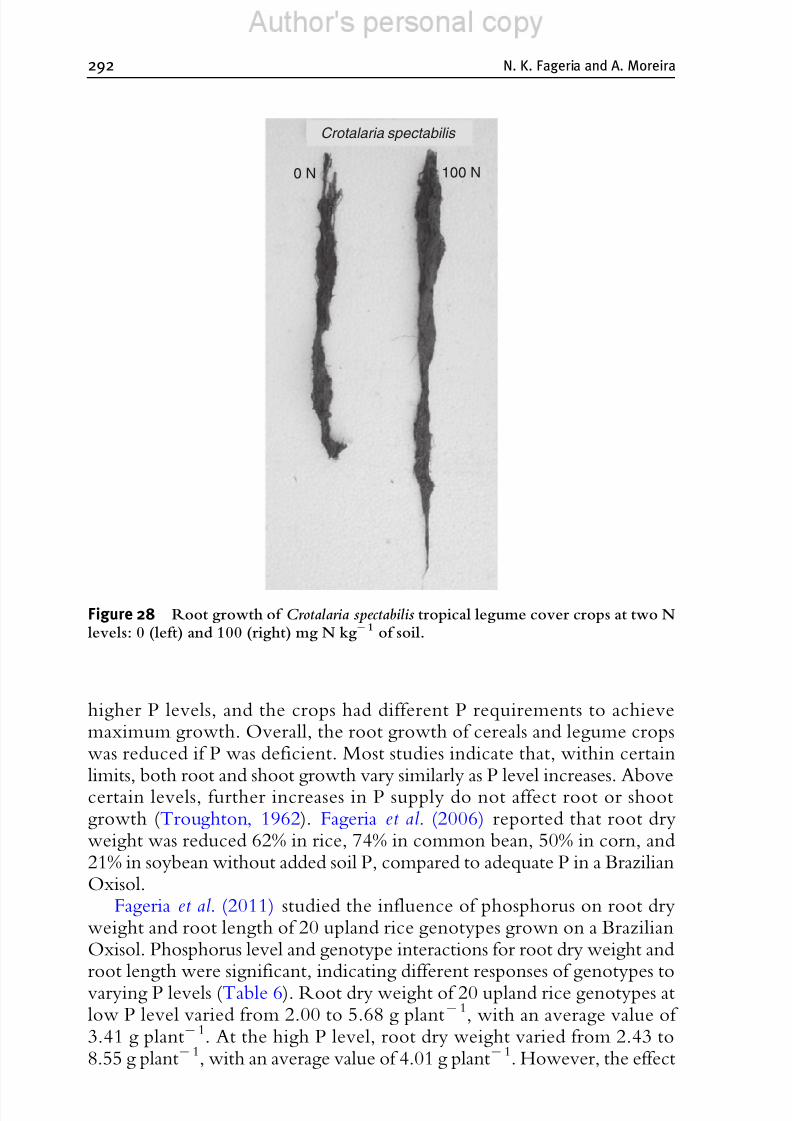
8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 43/82
higher P levels, and the crops had different P requirements to achievemaximum growth. Overall, the root growth of cereals and legume cropswas reduced if P was deficient. Most studies indicate that, within certainlimits, both root and shoot growth vary similarly as P level increases. Abovecertain levels, further increases in P supply do not affect root or shootgrowth (Troughton, 1962). Fageria et al . (2006) reported that root dryweight was reduced 62% in rice, 74% in common bean, 50% in corn, and21% in soybean without added soil P, compared to adequate P in a BrazilianOxisol.
Fageria et al . (2011) studied the influence of phosphorus on root dryweight and root length of 20 upland rice genotypes grown on a BrazilianOxisol. Phosphorus level and genotype interactions for root dry weight androot length were significant, indicating different responses of genotypes tovarying P levels (Table 6). Root dry weight of 20 upland rice genotypes atlow P level varied from 2.00 to 5.68 g plant1, with an average value of 3.41 g plant1. At the high P level, root dry weight varied from 2.43 to8.55 g plant1, with an average value of 4.01 g plant1. However, the effect
0 N 100 N
Crotalaria spectabilis
Figure 28 Root growth of Crotalaria spectabilis tropical legume cover crops at two Nlevels: 0 (left) and 100 (right) mg N kg1 of soil.
292 N. K. Fageria and A. Moreira

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 44/82
of P level on root system dry weight was not significant. Root length variedfrom 23.00 to 38.33 cm with an average value of 30.9 cm at low P level.At high P level, root length varied from 23.67 to 34.33 cm, with an averagevalue of 28.20 cm. There was a significant 10% decrease in root length at thehigh P level compared to the low P level. However, higher P level roots hadmore fine hairs compared to lower P level (personal observations).
Table 6 Root dry weight and root length of 20 upland rice genotypes as influencedby P levels
Genotype
Root dry weight (g plant1) Root length (cm)
Low P
(25 mg kg1)
High P
(200 mg kg1)
Low P
(25 mg kg1)
High P
(200 mg kg1)
BRA01506 3.92ab 3.22c 26.00ab 26.67a
BRA01596 2.78ab 2.73c 35.67ab 28.00aBRA01600 2.81ab 3.03c 36.00ab 29.00a
BRA02535 3.12ab 4.30c 28.67ab 33.33aBRA02601 4.42ab 3.20c 31.33ab 27.00a
BRA032033 3.70ab 3.62c 23.00b 29.67a
BRA032039 2.91ab 4.36c 27.67ab 27.00aBRA032048 3.96ab 3.91c 37.00a 33.33aBRA032051 2.00b 2.58c 36.00ab 30.67a
BRA042094 2.82ab 3.92c 30.00ab 27.33aBRA042156 2.50b 2.91c 29.00ab 27.00a
BRA042160 5.68a 8.32ab 32.67ab 33.00aBRA052015 3.91ab 2.98c 27.00ab 27.00a
BRA052023 4.69ab 8.55a 29.67ab 34.33aBRA052033 2.23b 2.43c 27.00ab 29.00a
BRA052034 3.18ab 3.99c 31.00ab 24.67a
BRA052045 3.07ab 3.08c 38.33a 24.67aBRA052053 2.57ab 3.87c 28.33ab 23.67a
BRSPrimavera
3.56ab 5.21bc 29.67ab 25.00a
BRSSertaneja
4.36ab 3.92c 34.00ab 23.67a
Average 3.41 4.01 30.9 28.20F -test
P level (P) NS *
Genotype(G)
** **
P G * **
Source: Fageria et al . (2011).*,**, NS Significant at the 5% and 1% probability level and nonsignificant, respectively. Means in the
same column followed by the same letter are not significantly different at the 5% probabilitylevel by the Tukey’s test.
The Role of Mineral Nutrition on Root Growth of Crop Plants 293

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 45/82
Figures 29–31 show root growth of upland rice under different P rates.Unlike the data cited above, root growth of all the cultivars/genotypesincreased with increasing P levels.
The first author studied the influence of P levels on the root growth of 14 tropical legume cover crops grown on a Brazilian Oxisol. Root dryweight was significantly influenced by P; crop species and P crop speciesinteractions were significant (Fig. 32). The significant P crop speciesinteraction indicates significant variation in shoot dry weight with thevariation in P levels. At the lowest P level (0 mg kg1), maximum rootdry weight of 0.77 g plant1 was produced by white jack bean (Canavaliaensiformis) and minimum root dry weight of 0.01 g plant1 was produced bycrotalaria (Crotalaria mucronata) and pueraria (Pueraria phaseoloides). At the
medium P level (100 mg P kg
1
), maximum root dry weight of 1.91 g plant1 was produced by black mucuna bean (Mucuna cinereum)and minimum root dry weight of 0.07 was produced by crotalaria (Crotalaria
0 P 50 P 175 P
Figure 29 Root growth of upland rice under 0 (left), 50 (center), and 175 (right)mg P kg1 soil (Fageria and Stone, 1999).
0 P 25 P 50 P 75 P 100 P 200 P
IAC 164
Figure 30 Root growth of upland rice cultivar IAC 164 at different P levels.
294 N. K. Fageria and A. Moreira

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 46/82
breviflora), with an average value of 0.63 g plant1. At the highest P level(200 mg P), maximum root dry weight of 1.42 g plant1 was produced bygray mucuna bean (Mucuna cinereum) and minimum root dry weight of 0.09 g plant1 was produced by calopogonio (Calopogonium mucunoides)and pueraria (Pueraria phaseoloides). The variation in root dry weight isgenetically controlled and also influenced by environmental variables, likethe supply of mineral nutrition (Baligar et al ., 2001; Caradus, 1990; Fageriaet al ., 2006).
Maximum root length of tropical legume cover crops varied from 15.5to 36 cm at the low P level, from 20.5 to 50.33 cm at the medium P leveland 18.33 to 52.33 cm at the high P level (Fig. 33). Overall, root length alsoincreased with increasing P level. Figures 34–36 show root growth of tropical legume cover crops as influenced by P levels. The improvementin root length by improved P nutrition has been reported by Fageria (2009)in various crop species. Barber (1995), Fageria et al . (2006), Marschner (1995), and Mengel et al . (2001) reported that mineral nutrition has tremen-dous effects on root growth, development, and function and, subsequently,the ability of roots to absorb and translocate nutrients. These authors further reported that mineral deficiencies induce considerable variations in thegrowth and morphology of roots and such variations are strongly influencedby plant species and genotypes.
200 P
25 P
BRA 01596
Figure 31 Root growth of upland rice genotype BRA01596 at two P levels.
The Role of Mineral Nutrition on Root Growth of Crop Plants 295

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 47/82
The influence of P fertilization on dry bean root dry weight was studiedby Fageria (1989b). Root dry weight of three bean genotypes increasedsignificantly in a quadratic fashion but differed from genotype to genotype(Table 7). The variability in root dry weight due to P fertilization was about
200mg P kg−1
1.6
1.4
1.2
1.0
0.8
0.6
0.4
0.2
0
2.0
1.81.6
1.4
1.2
1.0
0.8
0.8
0.6
0.6
0.4
0.4
0.2
0.2
0
0
C r o t a l a r i a b r e v i f l o r a
C r o t a l a r i a j u n c e a
C r o t a l a r i a m u c r o n a t a
C r o t a l a r i a s p e c t a b l i s
C r o t a l a r i a o c h r o l e u c a
C a l o p o g ô n i u m
m u c u n o i d e s
P u e r a r i a p h a s e o l o i d e s
C a j a n u s c a j a n
( m i x e d c o l o r )
C a j a n u s c a j a n ( b l a c k )
D o l i c h o s l a b l a b
M u c u n a d e e r i n g i a n a
M u c u n a a t e r r i m a
M u c u n a c i n e r e u m
C a n a v a l i a e n s i f o r m i s
100mg P kg−1
0 mg P kg−1
de de
cde
cde
b
a
a
b
b
bcbc bcd
df
f
efef
ef ff
c
de
de de de dee e
d
b
aa
bb
c
c
R o o t d r y w e i g h t ( g p l a n t − 1 )
b
de
cd cd
e e e
Figure 32 Root dry weight of 14 tropical legume cover crops as influenced by P rates.
296 N. K. Fageria and A. Moreira

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 48/82
52% in genotype Carioca, 35% in genotype CNF10, and 70% in genotypeCNF4856. Such information may contribute to the selection of cultivarsspecific to soil type and management systems, resulting in increasing yieldson the soils of variable P fertility (Fageria, 1989b).
70
60
50
40
30
20
10
0
c
cd
fdef
cde
C r o t a l a r i a b r e v i f l o r a
C r o t a l a r i a j u n c e a
C r o t a l a r i a m u c r o n a t a
C r o t a l a r i a s p e c t a b l i s
C r o t a l a r i a o c h r o l e u c a
C a l o p o g o n i u m
m u c u n o i d e s
P u e r a r i a p h a s e o l o i d e s
C a j a n u s c a j a n ( b l a c k )
C a j a n u s c a j a n
( m i x e d c o l o r )
D o l i c h o s l a b l a b
M u c u n a d e e r i n g i a n a
M u c u n a a t e r r i m a
M u c u n a c i n e r e u m
C a n a v a l i a e n s i f o r m i s
cdecd
cdcd cd
bcab
a
e
de dede
ab
defdef def
ef
cde cde cdebcd
bc
aa
b
cdcd
cd cdc c
ab
a
b ab
d
60
50
40
30
20
10
0
40
30
20
10
0
ab
200mg P kg−1
100mg P kg−1
M a x i m u m
r o o t l e n g t h ( c
m )
0 mg P kg−1
Figure 33 Maximum root length of 14 tropical legume cover crops at three P levels.
The Role of Mineral Nutrition on Root Growth of Crop Plants 297
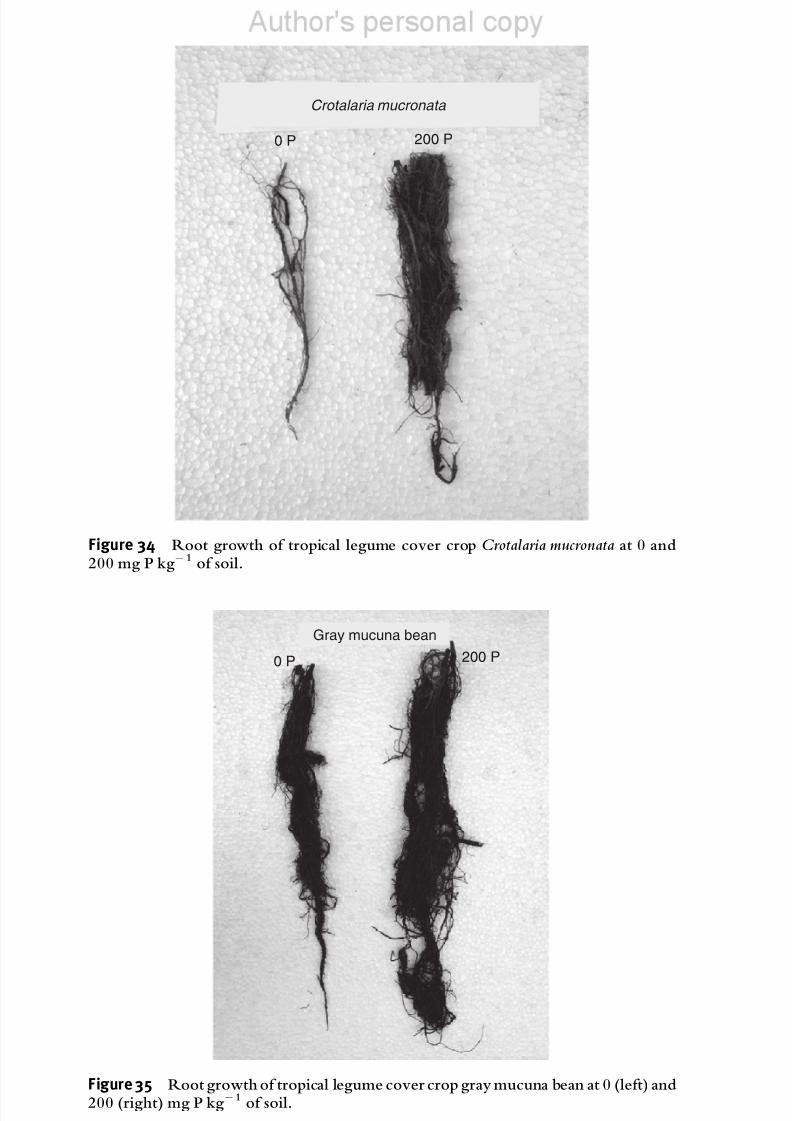
8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 49/82
0 P 200 P
Crotalaria mucronata
Figure 34 Root growth of tropical legume cover crop Crotalaria mucronata at 0 and200 mg P kg1 of soil.
Gray mucuna bean
0 P 200 P
Figure 35 Root growth of tropical legume cover crop gray mucuna bean at 0 (left) and200 (right) mg P kg1 of soil.

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 50/82
12.3. Potassium
Potassium plays an important role in the growth and development of plants,including the root system. Many enzymes are activated in plants by potas-sium, and it is also required for photosynthesis, transport of photosynthate,and protein synthesis (Blevins, 1994). Potassium plays a role in cell growthfollowing cell division by serving as a major component in cell turgor. It alsomaintains ionic balance and electrical neutrality in plants. Crops that pro-duce large quantities of protein per unit area of land require more K thanthose that produce less protein (Blevins, 1994). Potassium also plays animportant role in opening and closing the stomata.
Tennant (1976) reported that potassium deficiency stops root growthcompletely within 10–12 days of planting in wheat. Inadequate K reducesroot growth and consequently, crop yields (Baligar et al ., 1998). A defi-ciency of K in an Inceptisol reduced root growth by 23% in lowland rice, by30% in dry bean, by 12% in corn, and by 11% in soybean (Baligar et al .,
0 P 200 P
Crotalaria breviflora
Figure 36 Root growth of tropical legume cover crop Crotalaria breviflora at 0 and200 mg P kg1 of soil.
The Role of Mineral Nutrition on Root Growth of Crop Plants 299

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 51/82
1998). On an Oxisol, a 35% lower root dry mass was observed in 13 corngenotypes when K levels were 0 versus 200 mg kg1 of soil (Baligar et al .,1998). Data in Table 8 show that at 0 mg K kg1 of soil root dry weight of common bean (P. vulgaris L.) genotypes varied from 1.54 to 3.14 g per 3 plants, a variation of twofold. At the 200 mg K kg1 level, root dry weightvaried from 1.50 to 2.30 g per 3 plants, a variation of 1.5-fold. Similarly,maximum root length varied from 42 to 46 cm at low K level and 32–44 cmat higher K level. At the higher K level, there was a slight decrease in theroot length of all the genotypes, and the root weight of three genotypes alsodecreased at the higher K level. However, at the higher K level, there weremore root hairs than at the low K level (visual observations).
There is widespread evidence for genotype diversity in the root char-acteristics of many crops in response to the environment and increasinginterest in using this diversity to improve agricultural production andconsequently, nutrient-use efficiency (Barber, 1994; Gregory, 1994).
Mullins et al . (1994) studied K placement effects on the root growth of cotton grown on a fine sandy loam soil. Root density measurements takenin-row showed that root growth at depths >20 cm was improved with in-row subsoil additions of K. Tupper (1992) also observed increased cottontaproot length when K fertilizer was band-applied in the subsoils of
Table 7 Influence of phosphorus fertilization on the root growth of three dry beangenotypes at harvest
P rate (mg kg1)
Root dry weight (g per 2 plants)
Carioca CNF10 CNF4856
0 0.29 0.45 0.23
25 1.13 1.79 1.3850 1.66 1.28 1.83
75 1.31 1.29 1.52100 1.34 1.60 1.59
125 1.62 1.33 1.78150 1.31 1.75 1.82
175 1.18 1.29 1.48200 1.79 1.50 1.67Regression analysis
P rate versus Carioca (Y ) ¼ 0.6459 þ 0.0128X 0.000044X 2,R 2 ¼ 0.5179**
P rate versus CNF10 (Y ) ¼ 0.8704 þ 0.0109X 0.000042X 2,R 2 ¼ 0.3509*
P rate versus CNF4856 (Y ) ¼ 0.6145 þ 0.0193X 0.000075X 2,R 2 ¼ 0.6953**
Source: Adapted from Fageria (1989b).*,** Significant at the 5% and 1% probability levels, respectively.
300 N. K. Fageria and A. Moreira

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 52/82
Mississippi soils with low soil-test K. On the other hand, Hallmark and
Barber (1984) and Yibrin et al . (1993) reported that localized applications of K did not promote root growth. However, K has been shown to promotethe root growth of some vegetable crops (Zhao et al ., 1991). Fageria (1992)determined the root growth of rice grown in nutrient solution as well as inan Oxisols at the stress and nonstress levels of K. At the stress levels of K, riceroot growth was reduced compared with nonstress levels. Alfalfa herbage
yield, root weight, and root total nonstructural carbohydrates increasedwith increasing K fertilizer (Kitchen et al ., 1990).
12.4. Calcium
Absolute Ca deficiency is difficult to identify on plants grown in acidic soils(Kamprath and Foy, 1985). Most acidic soils contain adequate total Ca for most plants, and Ca-deficiency symptoms are rarely observed in the field.Only in highly leached, acidic, low-cation exchange soils (Oxisols andUltisols) would absolute deficiencies be likely to occur (Garrity et al .,1983). Levels of Ca required for essential growth functions are so low asto approach those of micronutrients. Hence, the major role of Ca in soilsand in plants is to exclude or detoxify other elements such as Al, Mn, andheavy metals that might otherwise become toxic (Garrity et al ., 1983).
Gonzalez-Erico et al . (1979) evaluated the maize response to deepincorporation of limestone on an Oxisol. They reported that incorporation
Table 8 Root dry weight and maximum root length of six common bean genotypes asinfluenced by potassium levels applied to a Brazilian Oxisol
Genotypes
0 mg K kg1 200 mg K kg1
Root dry weight
(g per 3 plants)
Maximum
root length
(cm)
Root dry weight
(g per 3 plants)
Maximum
root length
(cm)
Apore 1.54 45 1.67 32Perola 1.97 42 2.04 39
Ruda 1.94 44 2.30 35
IACCarioca
3.14 45 1.70 38
JaloPrecoce 2.24 42 1.67 36
Safira 1.77 46 1.50 44
Average 2.10 44 1.81 37
Source: Fageria et al . (2008a).Values were determined at physiological maturity.
The Role of Mineral Nutrition on Root Growth of Crop Plants 301
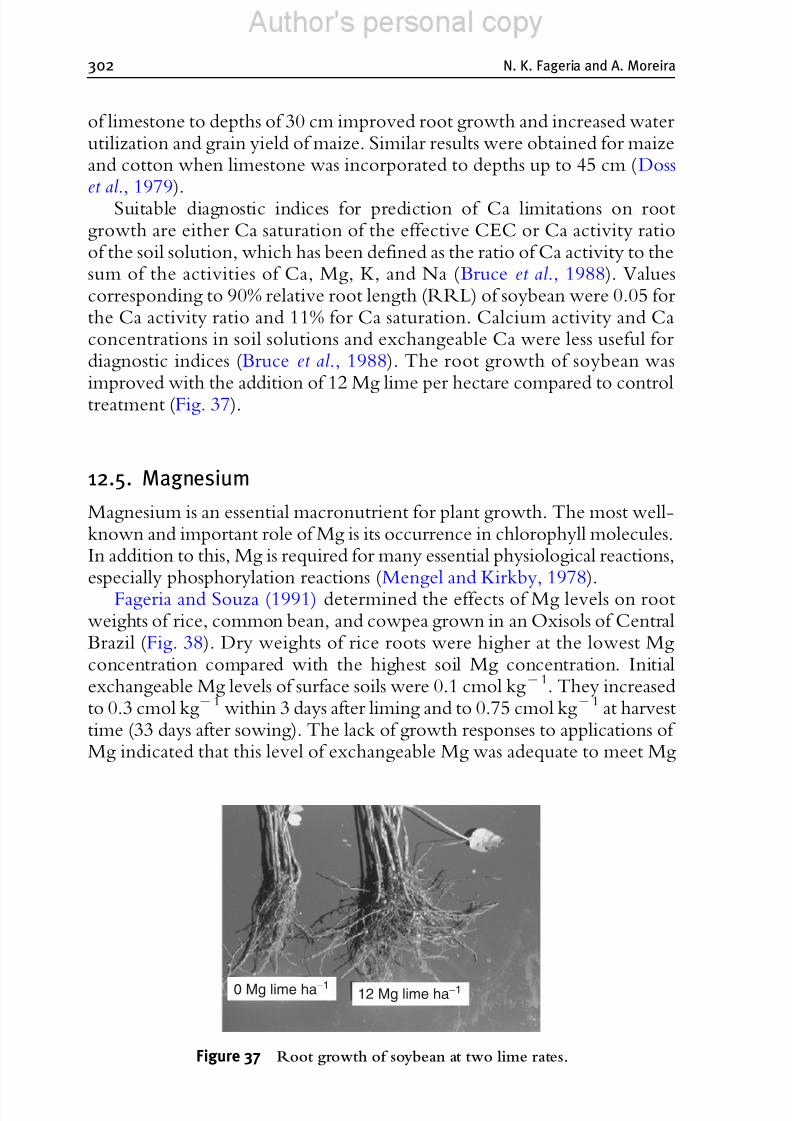
8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 53/82
of limestone to depths of 30 cm improved root growth and increased water utilization and grain yield of maize. Similar results were obtained for maizeand cotton when limestone was incorporated to depths up to 45 cm (Doss
et al ., 1979).Suitable diagnostic indices for prediction of Ca limitations on rootgrowth are either Ca saturation of the effective CEC or Ca activity ratioof the soil solution, which has been defined as the ratio of Ca activity to thesum of the activities of Ca, Mg, K, and Na (Bruce et al ., 1988). Valuescorresponding to 90% relative root length (RRL) of soybean were 0.05 for the Ca activity ratio and 11% for Ca saturation. Calcium activity and Caconcentrations in soil solutions and exchangeable Ca were less useful for diagnostic indices (Bruce et al ., 1988). The root growth of soybean was
improved with the addition of 12 Mg lime per hectare compared to controltreatment (Fig. 37).
12.5. Magnesium
Magnesium is an essential macronutrient for plant growth. The most well-known and important role of Mg is its occurrence in chlorophyll molecules.In addition to this, Mg is required for many essential physiological reactions,especially phosphorylation reactions (Mengel and Kirkby, 1978).
Fageria and Souza (1991) determined the effects of Mg levels on rootweights of rice, common bean, and cowpea grown in an Oxisols of CentralBrazil (Fig. 38). Dry weights of rice roots were higher at the lowest Mgconcentration compared with the highest soil Mg concentration. Initialexchangeable Mg levels of surface soils were 0.1 cmol kg1. They increasedto 0.3 cmol kg1 within 3 days after liming and to 0.75 cmol kg1 at harvesttime (33 days after sowing). The lack of growth responses to applications of Mg indicated that this level of exchangeable Mg was adequate to meet Mg
12 Mg lime ha−10 Mg lime ha−1
Figure 37 Root growth of soybean at two lime rates.
302 N. K. Fageria and A. Moreira

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 54/82
requirements of upland rice grown in this limed soil. Dry weight of roots of common bean increased with Mg application up to 3 cmol kg1 of soil.Similarly, significant responses of cowpea root growth to soil Mg levelswere observed, and maximum root weight was achieved at2.5 cmol Mg kg1 of soil.
12.6. Sulfur
Sulfur has long been recognized as an essential element for plant growth anddevelopment and classified as a macronutrient. Crop responses to appliedsulfur have been reported in a wide range of soils in many parts of the world(Fageria, 2009). Sulfur plays many important roles in the growth anddevelopment of plants. Fageria and Gheyi (1999) summarized importantfunctions of the sulfur in the plant. It is an important component of twoamino acids, cysteine and methionine, which are essential for proteinformation. Since animals cannot reduce sulfate, plants play a vital role in
0
Y = 2.6658exp(0.3007X −0.05983X 2)
Y = 2.4497exp(0.3009X −0.05047X 2)
Y =1.0752 −0.08852X
R 2= 0.80**
R 2= 0.88*
R 2= 0.85*
Upland rice
Common bean
Cowpea
Extractable Mg in soil (cmolc kg−1
)
R o o t d r y
w e i g h t ( g / 3 p l a n t s )
0
0
0
1
1
1
1
2
2
2
2
3
3
3
3
4
4
4
5 6 7
Figure 38 Root dry weights of upland rice, common bean, and cowpea grown withdifferent Mg levels in an Oxisol (adapted from Fageria and Souza, 1991).
The Role of Mineral Nutrition on Root Growth of Crop Plants 303

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 55/82
supplying essential S-containing amino acids to them. Sulfur plays animportant role in enzyme activation. It promotes nodule formation inlegumes. Sulfur is necessary in chlorophyll formation, although it is not a
constituent of chlorophyll. Maturity of seeds and fruits is delayed in theabsence of adequate sulfur. Sulfur is required by the plants in the formationsof nitrogenase. It increases the crude protein content of forages andimproves the quality of cereals for milling and baking. Sulfur increases theoil content of oilseed crops and increases winter hardiness in plants.It increases drought tolerance in plants, controls certain soil-borne diseases,and helps in the formation of glucosides that give characteristic odors andflavors to onion, garlic, and mustard. Sulfur is necessary for the formation of vitamins and synthesis of some hormones and glutathione, and it is involved
in oxidation–reduction reactions. Sulfur improves tolerance to heavy metaltoxicity in plants, and it is a component of sulfur contain sulfolipids. Organicsulfates may serve to enhance the water solubility of organic compounds,which may be important in dealing with salinity stress, and fertilization withsulfate decreases fungal diseases in many crops. Few studies have assessed theimpacts of sulfur on root growth and function; however, the effects of sulfur on root growth may be similar to those of N. Zhao et al. (2008) reportedthat S application increased the root number and root dry weight of soybeancompared to control treatment.
12.7. Micronutrients
Micronutrients have also been called minor or trace elements, indicatingthat their required concentrations in plant tissues are small compared to themacronutrients (Fageria et al ., 2002; Mortvedt, 2000). Based on physico-chemical properties, except B and Cl, the essential micronutrients aremetals. Even though micronutrients are required in small quantities byfield crops, their influence can be as great as that of macronutrients incrop production. Micronutrients are normally constituents of prostheticgroups that catalyze redox processes by electron transfer (such as withthe transition elements Cu, Fe, Mn, and Mo) and form enzyme–substratecomplexes by coupling enzymes with substrates (Fe and Zn) or enhanceenzyme reactions by influencing molecular configurations betweenenzyme and substrate (Zn) (Fageria et al ., 2002).
Micronutrient deficiencies in crop plants are widespread because of (i)increased micronutrient demands from intensive cropping practices andadaptation of high-yielding cultivars which may have higher micronutrientdemand, (ii) enhanced production of crops on marginal soils that containlow levels of essential micronutrients, (iii) increased use of high-analysisfertilizers with low amounts of micronutrients, (iv) decreased use of animalmanures, composts, and crop residues, (v) use of many soils that are inher-ently low in micronutrient reserves, (vi) use of liming in acid soils, and (vi)
304 N. K. Fageria and A. Moreira

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 56/82
involvement of natural and anthropogenic factors that limit adequatesupplies and create elemental imbalance (Fageria et al ., 2002).
12.7.1. ZincDeficiency of Zn in crop production is spread worldwide (Alloway, 2008).Graham (2008) reported that half of the world’s soils are intrinsicallydeficient in Zn. Zinc deficiency in annual crops is reported in Brazil(Fageria and Stone, 2008), Australia (Graham, 2008), India (Singh, 2008),China (Zou et al ., 2008), Turkey (Cakmak, 2008), Europe (Sinclair andEdwards, 2008), USA (Brown, 2008), and Africa (Waals and Laker, 2008).Micronutrient deficiencies are also a worldwide problem in human health(Welch, 2008). Zinc deficiency is the highest priority among micronutri-
ents for agriculture to address (Graham, 2008).In the Brazilian Cerrado region, Zn deficiency is very common inannual crops, especially upland rice and corn (Fageria, 2009). Figures 39and 40 show Zn-deficiency symptoms in upland rice and corn grown onBrazilian Oxisols. Data in Table 9 show that Zn application of up to120 mg kg1 improved the root growth of upland rice and wheat signifi-cantly. Similarly, Figs. 41 and 42 show improvement in the root growth of soybean and dry dean, respectively, with the addition of Zn in BrazilianOxisols.
12.7.2. BoronBoron deficiency is common for plants grown in arid, semiarid, and heavyrainfall areas in calcareous, sandy, light textured, acid, and low-OM soils(Gupta, 1993). Differences between B sufficiency and toxicity are narrow
Figure 39 Zinc deficiency in upland rice grown on Brazilian Oxisol.
The Role of Mineral Nutrition on Root Growth of Crop Plants 305

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 57/82
(Marschner, 1995), and soils supplied with high amounts of municipalcompost, sludge, and biosolids tend to accumulate high amounts of B,which may result in B toxicity. Boron is essential for pollen germinationand pollen tube growth in crop plants (Blevins, 1994). Boron requirements
Figure 40 Zinc deficiency in corn grown on Brazilian Oxisol.
Table 9 Root dry weight of upland rice and root length of corn as influenced by zincfertilization
Zn rate (mg kg1)
Upland rice root dry weight
(g per 4 plants)aWheat maximum root
length (cm)a
0 1.08 555 1.33 54
10 1.48 5220 1.55 55
40 1.73 5780 1.45 58
120 1.18 52R 2 0.75** 0.24*
Source: Adapted from Fageria (2002a).*,** Significant at the 5% and 1% probability levels, respectively.a Upland rice plants were harvested 6 weeks after sowing and corn plants were harvested 4 weeks after
sowing.
306 N. K. Fageria and A. Moreira

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 58/82
of dicots are generally higher than monocots (Fageria, 2000). Fageria (2000)reported that maximum root growth of upland rice can be achieved withthe application of 0.4 mg B kg1 soil, whereas maximum root growth of dry bean required 1.9 mg B kg1 of soil. Figure 43 shows that B require-ments for root growth varied among the crop species. Application of 24 mg B kg1 soil decreased the root dry weight of upland rice and cornin greenhouse studies. However, application of B at lower rates to the samesoil increased the root dry weight in dry bean, soybean, and wheat (Fig. 43).
0 5 120
Dry
bean
mg Zn kg−1
Figure 42 Root growth of dry bean at three Zn levels.
40
Soja
1050 80 mg Zn kg−1
Figure 41 Root growth of soybean at different Zn levels.
The Role of Mineral Nutrition on Root Growth of Crop Plants 307
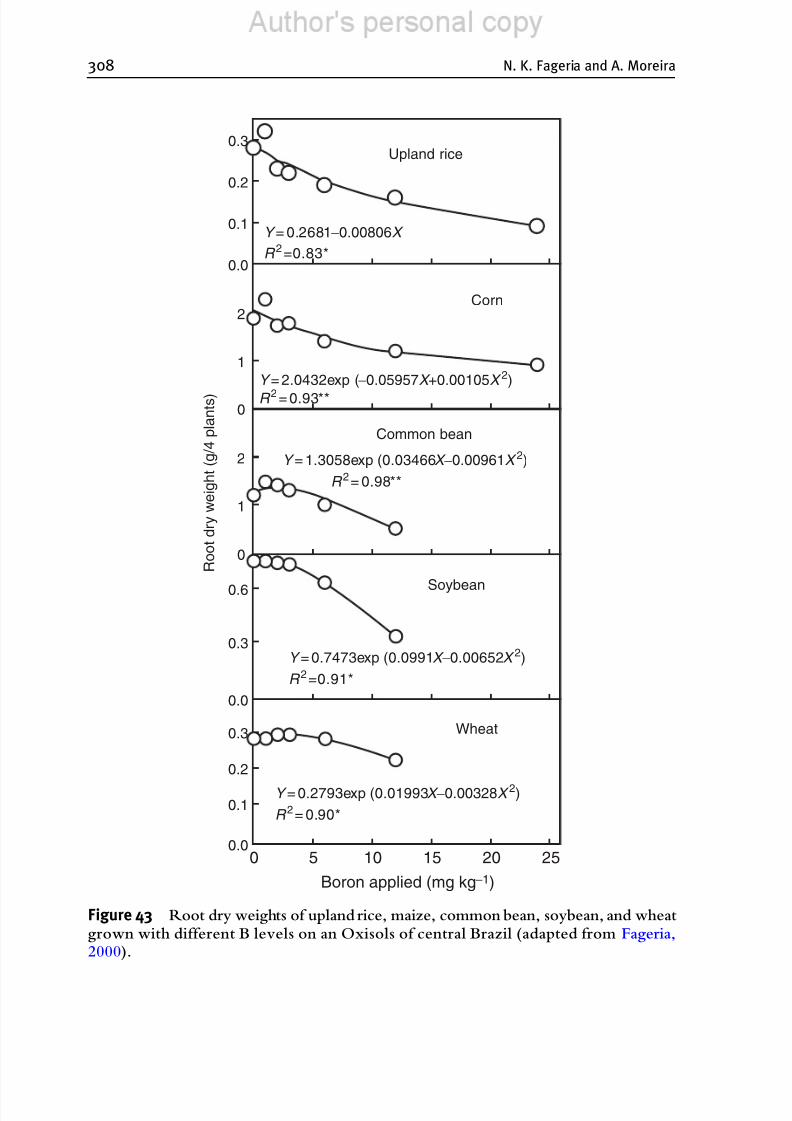
8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 59/82
0.3
Y = 0.2681−0.00806X
R 2=0.83*
Y = 2.0432exp (−0.05957X +0.00105X 2)
R 2 = 0.93**
Y = 1.3058exp (0.03466X −0.00961X 2)
R 2= 0.98**
R 2=0.91*
Y = 0.2793exp (0.01993X −0.00328X 2)
R 2= 0.90*
0.2
0.1
0.0
2
1
0
2
1
0
0.6
0.3
0.0
0.3
0.2
0.1
0.00 5 10 15 20 25
Boron applied (mg kg−1)
R o o t d r y w e i g h t ( g / 4 p l a n t s )
Upland rice
Corn
Common bean
Soybean
Wheat
Y = 0.7473exp (0.0991X −0.00652X 2)
Figure 43 Root dry weights of upland rice, maize, common bean, soybean, and wheatgrown with different B levels on an Oxisols of central Brazil (adapted from Fageria,2000).
308 N. K. Fageria and A. Moreira

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 60/82
12.7.3. CopperCopper deficiency is often observed on plants grown in soils inherently lowin Cu (coarse-textured and calcareous soils) and in soils high in OM, where
Cu complexes with organic substances (Fageria et al ., 2002). Higher-than-normal Cu supplies usually inhibit root growth more than shoot growth(Lexmond and Vorm, 1981). Use of Cu-containing fungicides and anthel-mintic compounds in agriculture has resulted in Cu toxicity in some plants,but naturally occurring Cu toxicity is relatively uncommon (Welch et al .,1991). Root dry weight of wheat and root length of dry bean were signifi-cantly increased by the application of copper fertilization (Table 10).
12.7.4. IronIron deficiency is a worldwide problem and occurs in numerous crops(Fageria et al ., 2002; Marschner, 1995). Iron deficiency occurs not becauseof Fe scarcity in soil or plants, but because various soil and plant factors affectFe availability, inhibit its absorption, or impair its metabolic use (Marschner,1995; Welch et al ., 1991). Plant species that commonly become Fe-deficientare peanut ( A. hypogaea L.), soybean (G. max L.), sorghum (Sorghum bicolor L. Moench), and upland rice (O. sativa L.). Iron deficiency reduces rootgrowth (Table 11). Iron toxicity (indicated by leaf bronzing) can be seriousfor production of crops in waterlogged soils. For wetland rice, Fe toxicity isthe second most severe yield-limiting nutrient disorder, and it has beenreported in South America, Asia, and Africa (Fageria et al ., 2008b). Irontoxicity decreases the root growth of lowland rice (Fageria et al ., 2008b);however, genotypic differences exist (Fig. 44).
Table 10 Root dry weight of wheat and root length of dry bean asinfluenced by copper fertilization
Cu rate (mg kg1) Wheat (g per 4 plants)a Dry bean (cm)a
0 0.53 252 0.60 30
4 0.50 278 0.48 28
16 0.47 28
32 0.47 2464 0.43 30
96 0.17 14R 2 0.88** 0.42**
Source: Adapted from Fageria (2002a).** Significant at the 1% probability levels.a Wheat and dry bean plants were harvested 5 weeks after sowing.
The Role of Mineral Nutrition on Root Growth of Crop Plants 309
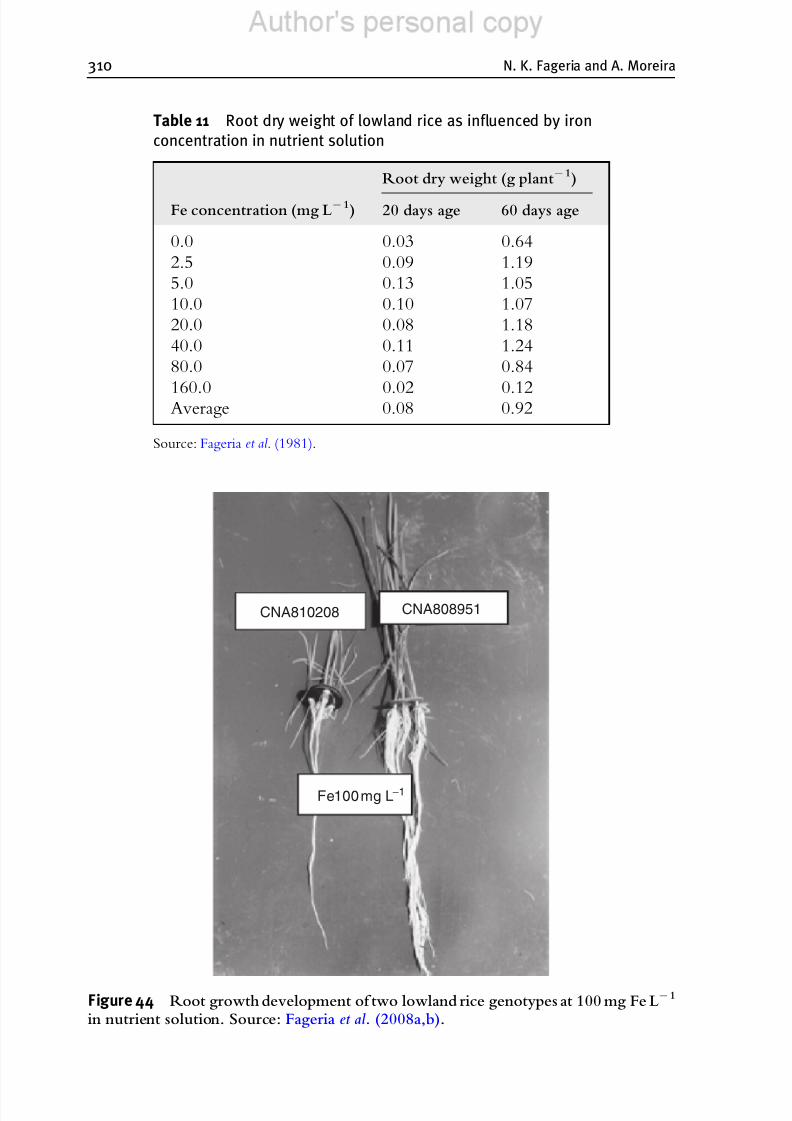
8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 61/82
CNA810208 CNA808951
Fe100mg L−1
Figure 44 Root growth development of two lowland rice genotypes at 100 mg Fe L 1
in nutrient solution. Source: Fageria et al. (2008a,b).
Table 11 Root dry weight of lowland rice as influenced by ironconcentration in nutrient solution
Fe concentration (mg L 1)
Root dry weight (g plant1)
20 days age 60 days age
0.0 0.03 0.64
2.5 0.09 1.195.0 0.13 1.05
10.0 0.10 1.0720.0 0.08 1.18
40.0 0.11 1.2480.0 0.07 0.84
160.0 0.02 0.12Average 0.08 0.92
Source: Fageria et al . (1981).
310 N. K. Fageria and A. Moreira

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 62/82
12.7.5. Other micronutrientsIn addition to zinc, boron, copper, and iron, the other micronutrients thatare essential for the growth of higher plants are manganese, molybdenum,
chlorine, and nickel. Manganese deficiency has been reported for plantsgrown in the coarse-textured and poorly drained coastal plain soils of theUnited States (Reuter et al ., 1988) and in the soils of Central America,Bolivia, and Brazil (Leon et al ., 1985). In Europe, Mn deficiency has beenreported on plants grown in peaty (England and Denmark), coarse-textured(Sweden and Denmark), coarse/fine textured (Netherlands), podzolic, andbrown forest (Scotland) soils (Welch et al ., 1991). Manganese deficiency hasalso been reported on plants grown in the semiarid regions of China, India,southeast and western Australia, Congo, Ivory Coast, Nigeria, and other
western African countries (Fageria et al ., 2002). Manganese toxicity on cropplants grown in many parts of the world has been reported to be moreimportant than Mn deficiency (Foy, 1984; Welch et al ., 1991).
Molybdenum is the least abundant of the micronutrients in the litho-sphere (Mortvedt, 2000), and soil concentrations range from 0.2 to5 mg kg1 (mean of 2 mg kg1). Mo deficiency usually occurs on plantsgrown in the broad areas of well-drained acid soils and in soils formed fromparent materials low in Mo. In Australia, Mo deficiency occurs on cropsgrown in soils derived from sedimentary rocks, basalts, and granites
(Anderson, 1970). Peaty, alkaline, and poorly drained soils commonlyhave high Mo. Iron oxides adsorb more Mo than Al oxides (Fageria et al .,2002), and clay mineralogy can affect Mo adsorption, in the order mont-morillonite > illite > kaolinite (Goldberg, 1993). Hydrous ferric oxides or ferric oxide molybdate complexes and insoluble ferric molybdates may formin well-aerated soils so that Mo solubility and availability to plants is low(Welch et al ., 1991). In poorly drained soils, the formation of soluble ferrousmolybdates or molybdites may lead to high Mo availability to plants. Plantsgrown in high Mo soils of the intermountain valleys of western UnitedStates have been reported to accumulate high Mo which has induced“molybdenosis” (Cu deficiency) in cattle (Welch et al ., 1991).
Chloride is essential to higher plants and is required for the water-splitting reactions in photosystem II (Kelley and Izawa, 1978). Nickel hasbeen shown to be essential for soybean (Eskew et al ., 1983), and it is knownto be a constituent of urease (Blevins, 1994; Klucas et al ., 1983). Since urea isa widely used fertilizer in crop plants worldwide, Ni nutrition could beimportant (Blevins, 1994). Information on the influence of these micronu-trients on the root growth of crop plants is not available, and this aspect isnot discussed here.
The Role of Mineral Nutrition on Root Growth of Crop Plants 311

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 63/82
13. Management Strategies for MaximizingRoot Systems
Root growth of crop plants can be improved by adopting manage-ment practices that modify soils to fit crops, as well as to modify the plants tofit the soil. Both of these strategies can be combined to get maximumeconomic results.
13.1. Soil management
Soil management practices that can improve root growth of crop plants
include liming acid soils, use of gypsum, maintenance of organic matter,use of adequate rates, sources and methods of fertilizer application, and deepplowing.
13.1.1. Liming acid soilsLiming is the most common and effective practice to reduce soil acidity.Lime requirements of crops grown on acid soils are determined by the qualityof liming material, status of soil fertility, crop species and cultivar withinspecies, crop management practices, and economic considerations. Soil pH,
base saturation, and aluminum saturation are important acidity indices thatare used to determine liming. Liming improves soil pH, Ca, and Mg contentsand reduces Al concentrations in the soil solution. In addition, limingimproves beneficial microbe populations in the soil. Furthermore, limingimproves P concentration in the soil solution by reducing P immobilizationby Fe and Al in acid soils (Fageria and Baligar, 2008). All these beneficialeffects of liming improve the root growth of crop plants. Nurlaeny et al .(1996) found that liming increased shoot dry weight, total root length, andmycorrhizal colonization of roots in soybean and corn grown on tropical acid
soils. Gonzalez (1976) reported that incorporation of lime to a 30 cm soildepth allowed the corn roots to penetrate and use stored water throughoutthe lime layer. Data in Table 12 show that liming increases soil pH andconsequently, the root dry weight of dry bean grown on a Brazilian Oxisol.Similarly, Figs. 45 and 46 show how the root growth of dry bean and soybeanis influenced by soil pH. The root growth of both the legumes was signifi-cantly influenced by increasing soil pH. Dry bean root growth was maximumat pH 5.9 and soybean produced vigorous root systems at pH 6.4.
13.1.2. Use of gypsumGypsum (CaSO4 2H2O) or phosphogypsum (e.g., byproducts of phospho-ric acid manufacturing processes) applications are used to leach Ca deeper into soil profiles where Ca can replace Al on cation exchange complexes.
312 N. K. Fageria and A. Moreira

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 64/82
Much of the Al displaced by Ca can be leached from the root zones. Thispractice works well in sandy soils or Oxisols with clay loam aggregateswhich behave hydrologically like sands (Foy, 1992). Poor root growth of crop plants has been frequently observed in highly weathered acid soils invarious countries (Alcordo and Rechcigl, 1993). The chemical factor iden-tified as most responsible for poor root growth is excess soluble Al (Alcordoand Rechcigl, 1993; Foy, 1992). Excess Al3þ has been reported to inhibitroot growth by binding to the PO4 portion of DNA in the root cell nuclei,reducing template activity and thus cell division (Matsumoto andMorimura, 1980). In legumes, it has been shown to impair the growth of root hairs and rhizobia, reducing root nodule initiation and function(Munns and Franco, 1982). Excess Al may also adversely affect the rootand overall plant growth in nonphytotoxic ways by competing with Ca and
Table 12 Influence of soil pH on root growth of dry bean grown ona Brazilian Oxisol
Soil pH in H2O Root dry weight (g per 4 plants)
4.1 1.90
4.7 3.58
5.3 4.675.9 5.40
6.6 4.737.0 3.80R 2 0.99**
Source: Adapted from Fageria (2002b).
** Significant at the 1% probability level.
Lowland soil
Dry
bean
pH 4.9 pH 5.9 pH 6.4 pH 6.7 pH 7.0
Figure 45 Root growth of dry bean grown on a Brazilian lowland soil (Inceptisol;Fageria and Stone, 1999).
The Role of Mineral Nutrition on Root Growth of Crop Plants 313

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 65/82
Mg for uptake by plants (Rengel and Robinson, 1989). The use of gypsum
can neutralize subsoil acidity by leaching CaSO4 and forming AlSO4þ
which is not toxic for root growth (Alva and Sumner, 1989). Ritcheyet al . (1980) reported that application of gypsum increased subsoil Ca andMg while decreasing Al and improving the root growth of corn in BrazilianOxisol.
13.1.3. Maintenance of adequate amounts of organic matterThe benefits of organic matter addition to soils include improving nutrientcycling and availability to plants through direct additions as well as throughmodification in soil physical and biological properties. The complementaryuse of organic manures and chemical fertilizers has proved to be the best soilfertility management strategy in the tropics (Fageria and Baligar, 2005).Enhanced soil organic matter increases soil aggregation and water-holdingcapacity, provides an additional source of nutrients, and reduces P fixation,toxicities of Al and Mn, and leaching of nutrients (Baligar and Fageria,1999). Build-up of organic matter through additions of crop and animalresidues increases the population and species diversity of microorganismsand their associated enzyme activities and respiration rates (Fageria, 2002c).The use of organic compost may result in a soil that has greater capacity toresist the spread of plant pathogenic organisms. The improvement in theoverall soil quality may produce more vigorous root systems and higher crop yields (Fageria, 2002c).
Lowland soil
pH 4.9 pH 5.9 pH 6.4 pH 6.7 pH 7.0
Soybean
Figure 46 Root growth of soybean in a Brazilian lowland soil (Inceptisol) at different pH.
314 N. K. Fageria and A. Moreira

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 66/82
13.1.4. Use of adequate nutrient rates, sources, and methods of application
Use of adequate rates and effective sources of nutrients are important
management practices to improve crop yield and plant root systems.In addition, appropriate methods of fertilizer application are also importantfor the development of vigorous root systems. Immobile nutrients like Pand K should be applied in bands to improve their uptake by plant roots.
13.1.5. Deep plowingDeep plowing improves soil conditions for root growth by breaking com-pacted layers that roots cannot readily penetrate. If water tables are near thesoil surfaces, drainage can also be useful. When depths to root-restricting
hardpans are relatively shallow (<0.25 m), chisel plowing can be effectivefor disrupting compacted layers.
13.1.6. Mulching, greater sowing depth, and sowing larger seedsRoot growth may be manipulated through cultural practices like mulching,which can affect soil temperatures. Warmer soil temperatures generallyproduce larger root systems. In addition, crop residues on soil surfacesdecrease soil evaporation and improve water-use efficiency. Greater sowingdepths may decrease seedling emergence rate, but it can also increase the
survival of emergent seedlings by increasing water availability. Larger seedsgenerally produce seedlings with more extensive root systems. Sowing goodquality seeds could also improve root systems.
13.1.7. Leaching salts from soil profilesLeaching may be required to prevent harmful accumulation of salts in theroot zone.. Steady-state leaching (L r ) requirements may be estimated(Reichman and Trooien, 1993) as
L r ¼D d
D a¼ ECa
ECd
;
where D d and D a are depths of drainage water and applied water, respec-tively, and ECa and ECd are electrical conductivity of applied water anddrainage water, respectively.
13.1.8. Integrated cropping systems and pest managementReduced tillage, N side-dressing, and early planting can be included inintegrated pest management programs with no risk of increasing potentialfor root damage from western maize rootworm (Roth et al ., 1995). Miltner et al . (1991) reported that cyst nematodes suppressed soybean root growthon susceptible cultivars, whereas root growth of tolerant cultivars was
The Role of Mineral Nutrition on Root Growth of Crop Plants 315

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 67/82
stimulated by the presence of soybean cyst nematodes. The use of tolerantor nutrient-use-efficient cultivars may be an important practice to improveroot growth in stress environments. Stress-tolerant genotypes are being
identified and bred worldwide to solve some of the most difficult problemsof soil fertility such as subsoil acidity, salinity, low plant availability of Fe incalcareous soils, and low P availability in acidic soils (Foy, 1992). Rhizobialspecies/strains differ markedly in tolerance to low pH, and toxicities of Aland Mn in tropical soils have been identified. The use of improved strainscan improve N2 fixation and the root growth of legumes grown in acidicsoils.
13.2. Plant management
Plant management is another important strategy in improving root systemsof crop plants. Adequate plant density and spacing can improve the rootgrowth of crop plants.
13.2.1. Genetic variabilityAn important plant management strategy is to exploit root system geneticvariability of crop species or cultivars within species. Plant genetic variabilitycan be defined as the heritable characters of a particular crop species or
cultivar that produce differences in the growth or production amongspecies, or cultivars of the same species, under favorable or unfavorablegrowth conditions (Fageria, 1989a). Cultivar differences in root size arequite common and have been related to differences in nutrient uptake(Baligar et al ., 1998; Caradus, 1990; Fageria et al ., 2006). Differencesbetween white clover (T. repens L) populations and cultivars in P uptakeper plant at low levels of P have been related to differences in root size andabsolute growth rate (Caradus and Snaydon, 1986). There is widespreadevidence for genotype diversity in the root characteristics of many crops inresponse to environment and increasing interest in using this diversity toimprove agricultural production and consequently, nutrient use efficiency(Barber, 1994; Gregory, 1994). Mineral deficiency and toxicity, mechanicalimpedance, moisture stress, oxygen stress, and temperature have tremen-dous effects on root growth, development, and function and, subsequently,the ability of roots to absorb and translocate nutrients (Arkin and Taylor,1981; Baligar et al ., 1998; Barber, 1995; Marschner, 1995; Mengel et al .,2001). Mineral deficiency induces considerable variations in growth andmorphology of roots, and such variations are strongly influenced by plantspecies and genotypes. Overall, the growth of the main axis is little affectedby nutrient deficiency, but growth of lateral branches and their elongationrates appear to be substantially reduced. Baligar et al . (1998) summarized theeffects of various essential elements as follows: nitrogen deficiency increasesroot hair length, increases or has no effect on root hair density, and reduces
316 N. K. Fageria and A. Moreira

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 68/82
branching. Phosphorus deficiency increases the overall growth of roots androot hair length, increases the number of second-order laterals, and either increases or does not affect root hair density. K and Ca deficiencies reduce
root growth; however, high Mg levels reduce the dry mass of roots. Theeffects of these nutrient stress factors on nutrient-use efficiency have notbeen well explored. Baligar et al . (1998) states that low pH reduces rootmass, length, and root hair formation. In alkaline soils, ammonium toxicitycauses severe root inhibition and in general, salinity leads to reduction in themass and length of roots and dieback of laterals.
14. Conclusions
The study of plant roots is one of the most promising, but leastexplored, areas of research related to mineral nutrition. Roots are poorlystudied primarily because of their physical location and growth habit.Understanding plant root growth is important for improving productivityof annual crops in agroecosystems. Root systems are important plant organsbecause they absorb water and nutrients and also provide mechanicalsupport to the plant. In addition, roots synthesize growth substances and
hormones such as cytokinins that are important in leaf function and possi-bly, grain development. In plant growth analysis, the role of roots isgenerally ignored due to the difficulty in getting accurate root growthdata under field condition. The principal structure of the root systemincludes four developmentally distinct classes of roots. These are taproots,basal roots, lateral roots, and shootborn roots. Current evidence suggeststhat the four root classes that make up the primary and secondary rootsystems are physiologically distinct from each other. Rooting depth andspreading capacity are important traits for uptake of water and nutrients.Root growth varies among plant species and cultivars within species and canbe modified by environmental factors. Genotypic differences in root growthamong crop species and genotypes of the same species under similar andvariable environmental conditions are now well demonstrated, and thepossibility of developing genotypes of desirable root systems to soil proper-ties offers exciting prospects for the future. Root number, maximum length,and root dry weight increase with increasing levels of macro- and micro-nutrients in the soil to a point beyond which root growth is suppressed.Plant roots together with their associated bacteria and fungi play an impor-tant role in the formation, maintenance, and turnover of soil aggregates.Currently, the techniques available to measure root systems are laboriousand time consuming, and this limits their use in plant physiological research.Hence, it is necessary to develop root measurement techniques that aresimple, cheap, and less time consuming.
The Role of Mineral Nutrition on Root Growth of Crop Plants 317

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 69/82
ACKNOWLEDGMENT
The authors thank Dr. Charles Allan Jones, Professor, Texas-A&M University, TexasAgriLife Research and Extension Service, Dallas, Texas for their peer review of the chapter
and for giving valuable suggestions for its improvement.
REFERENCES
Ae, N., Arihara, J., Okada, K., Yoshihara, T., and Johnson, C. (1990). Phosphorus uptake bypigeon pea and its role in cropping systems of the Indian subcontinent. Science 248,477–480.
Alcordo, I. S., and Rechcigl, J. E. (1993). Phosphogypsum in agriculture: A review. Adv.
Agron. 49, 55–118.Allmaras, R. R., Linden, D. R., and Clapp, C. E. (2004). Corn-residue transformations intoroot and shoot carbon as related to nitrogen, tillage, and stover management. Soil Sci. Soc.
Am. J. 68, 1366–1375.Alloway, B. J. (2008). Micronutrients and crop production: An introduction.
In “Micronutrient Deficiencies in Global Crop Production” (B. J. Alloway, Ed.),pp. 1–39. Springer, New York.
Alva, A. K., and Sumner, M. E. (1989). Alleviation of aluminum toxicity to soybeans byphosphogypsum or calcium sulfate in dilute nutrient solutions. Soil Sci. 147, 278–285.
Anderson, B. J. (1970). Tracer elements for sheep, pastures and fodder crops in Australia. J. Aust. Inst. Agric. Sci. 36, 15–29.
Anderson, E. L. (1987). Corn root growth and distribution as influenced by tillage andnitrogen fertilization. Agron. J. 79, 544–549.
Anderson, D. S., Teyker, R. H., and Rayburn, A. L. (1991). Nitrogen form effects on earlycorn root morphological and anatomical development. J. Plant Nutr. 14, 1255–1266.
Ando, T., Yoshida, S., and Niiyama, I. (1983). Nature of oxidizing power of rice roots. Plant Soil 72, 57–71.
Andren, O., Rajkai, K., and Katterer, T. (1991). A non-destructive technique for studies of root distribution in relation to soil-moisture. Agric. Ecosyst. Environ. 34, 269–278.
Arkin, G. F., and Taylor, H. M. (1981). Modifying the Root Environment to Reduce CropStressModifying the Root Environment to Reduce Crop Stress. ASAE, St. Joseph, MI.
Armstrong, J. (1979). Aeration in higher plants. In “Advances in Botanical Research”
(H. W. Woolhouse, Ed.), Vol. 7, pp. 226–332. Academic Press, London.Armstrong, W., and Drew, M. C. (2002). Root growth and metabolism under oxygen
deficiency. In “Plant Roots: The Hidden Half” (Y. Waisel, et al., Eds.), pp. 729–761.Marcel Dekker, New York.
Atkinson, D. (1990). Influence of root system morphology and development on the need for fertilizers and the efficiency of use. In “Crops as Enhancers of Nutrient Use”(V. C. Baligar and R. R. Duncan, Eds.), pp. 411–450. Academic Press, San Diego, CA.
Atwell, B. J. (1989). Physiological responses of lupin roots to soil compaction. In “Structuraland Functional Aspects of Transport in Roots” (B. C. Loughman, Ed.), pp. 251–255.Kluwer Academic Publishers, Dordrecht, The Netherlands.
Aung, L. H. (1974). Root–shoot relation-ships. In “The Plant Root and Its Environment”
(E. W. Carson, Ed.), pp. 29–61. University Press of Virginia, Charlottesville, VA.Bakken, L. R. (1990). Microbial growth and immobilization/mineralization of N in the
rhizosphere. Symbiosis 9, 37–41.Balesdent, J., and Balabane, M. (1996). Major contribution of roots to soil carbon storage
inferred from maize cultivated soils. Soil Biol. Biochem. 28, 1261–1263.
318 N. K. Fageria and A. Moreira

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 70/82
Baligar, V. C. (1986). Interrelationships between growth and nutrient uptake in alfalfa andcorn. J. Plant Nutr. 9, 1391–1404.
Baligar, V. C., and Fageria, N. K. (1999). Plant nutrient efficiency: Towards the secondparadigm. In “Soil Fertility, Soil Biology and Plant Nutrition Interrelationships”
(J. Q. Siqueira, F. M. S. Moreira, A. S. Lopes, L. R. G. Guilherme, V. Faquin,A. E. Furtini Neto, and J. G. Carvallo, Eds.), pp. 183–204. Brazilian Society of SoilScience and University of Lavras, Lavras, MG Brazil.
Baligar, V. C., Fageria, N. K., and Elrashidi, M. (1998). Toxicity and nutrient constraints onroot growth. HortScience 33, 960–965.
Baligar, V. C., Fageria, N. K., and He, Z. L. (2001). Nutrient use efficiency in plants.Commun. Soil Sci. Plant Anal. 32, 921–950.
Ball, R. A., Oosterhuis, D. M., and Mauromoustakos, A. (1994). Growth dynamics of thecotton plant during water-deficit stress. Agron. J. 86, 788–795.
Barber, S. A. (1994). Root growth and nutrient uptake. In “Physiology and Determinationof Crop Yield” (K. J. Boote, J. M. Bennett, T. R. Sinclair, and G. M. Paulsen, Eds.),
pp. 95–99. ASA, CSSA, and SSSA, Madison, WI.Barber, S. A. (1995). Soil Nutrient Bioavailability: A Mechanistic Approach.2nd edn.. John
Wiley & Sons, New York.Barber, S. A., and Silberbush, M. (1984). Plant root morphology and nutrient uptake.
In “Roots, Nutrient and Water Influx, and Plant Growth” (S. A. Barber andD. R. Bouldin, Eds.), pp. 65–85. ASA Special Publication No. 49, ASA, CSSA, andSSSA, Madison, WI. 65–87.
Barlow, P. W. (1986). Adventitious roots of whole plants: Their forms, functions andevolution. In “New Root Formation in Plants and Cuttings” (M. B. Jackson, Ed.),pp. 67–110. Martinus Nijhoff, Dordrecht, The Netherlands.
Barlow, P. W. (2002). Cellular pattering in root meristems: Its origins and significance.
In “Plant Roots: The Hidden Half” (Y. Waisel, et al., Eds.), pp. 49–82. Marcel Dekker,New York.
Bengough, A. G., and Mullins, C. E. (1990). Mechanical impedance to root growth:A review of experimental techniques and root growth responses. J. Soil Sci. 41, 341–358.
Beyrouty, C. A., Wells, B. R., Norman, R. J., Marvel, J. N., and Pillow, J. A. (1987).Characterization of rice roots using a minirhizotron technique. In “Minirhizotron Obser-vation Tubes: Methods and Applications for Measuring Rhizosphere Dynamics”(H. M. Taylor, Ed.), , ASA Special Publication No. 50, pp. 99–108. American Societyof Agronomy, Madison, WI.
Beyrouty, C. A., Wells, B. R., Norman, R. J., Marvel, J. N., and Pillow, J. A. (1988). Rootgrowth dynamics of a rice cultivar grown at two locations. Agron. J. 80, 1001–1004.
Blevins, D. G. (1994). Uptake, translocation, and function of essential mineral elements incrop plants. In “Physiology and Determination of Crop Yield” (G. A. Peterson, Ed.),pp. 259–275. ASA, CSSA, and SSSA, Madison, WI.
Bolliger, A., Magid, J., Amado, T. J. C., Neto, F. S., Ribeiro, M. F. S., Calegari, A.,Ralisch, R., and Neergaard, A. (2006). Taking stock of the Brazilian zero-till revolution:A review of landmark research and farmers practice. Adv. Agron. 91, 47–110.
Boone, R. D. (1994). Light-fraction soil organic matter: Origin and contribution to netnitrogen mineralization. Soil Biol. Biochem. 26, 1459–1468.
Boote, K. J., Stansell, J. R., Schubert, A. M., and Stone, J. F. (1982). Irrigation, water use,and water relationships. In “Peanut Science and Technology” (H. E. Pattee andC. T. Yong, Eds.), pp. 164–205. American Peanut Research & Education Society, Yoakum, TX.
Borg, H., and Grimes, D. W. (1986). Depth development of roots with time: An empiricaldescription. Trans. Am. Soc. Agric. Eng. 29, 194–197.
The Role of Mineral Nutrition on Root Growth of Crop Plants 319

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 71/82
Box, J. E., and Ramseur, E. L. (1993). Minirhizotron wheat root data: Comparisons to soilcore root data. Agron. J. 85, 1058–1060.
Brailsford, R. W., Voesenek, L. A. C. J., Blom, C. W. P. M., Smith, A. R., Hall, M. A., and Jackson, M. B. (1993). Enhanced ethylene production by primary roots of Zea mays L. in
response to sub-ambient partial pressures of oxygen. Plant Cell Environ. 16, 1071–1080.Brouwer, R. (1966). Root growth of grasses and cereals. In “The Growth of Cereals and
Grasses” (F. L. Milthorpe and D. J. Irvins, Eds.), pp. 153–166. Butterworth, London.Brown, R. H. (1984). Growth of the green plant. In “Physiological Basis of Crop Growth
and Development” (M. B. Tesar, Ed.), pp. 153–174. American Society of Agronomy andCrop Science Society of America, Madison, WI.
Brown, P. H. (2008). Micronutrient use in agriculture in the United States of America:Current practices, trends and constraints. In “Micronutrient Deficiencies in Global CropProduction” (B. J. Alloway, Ed.), pp. 267–286. Springer, New York.
Brown, S. C., Keating, J. D. H., Gregory, P. J., and Cooper, P. J. M. (1987). Effects of fertilizer, variety and location on barley production under rainfed conditions in northern
Syria. I. Root and shoot growth. Field Crops Res. 16, 53–66.Bruce, R. C., Warrell, L. A., Edwards, D. G., and Bell, L. C. (1988). Effects of aluminum
and calcium in the soil solution of acid soils on root elongation of Glycine max cv. Forrest. Aust. J. Agric. Res. 39, 319–338.
Buyanovsky, G. A., Wagner, G. H., and Gantzer, C. J. (1986). Soil respiration in a wheatecosystem. Soil Sci. Soc. Am. J. 50, 338–344.
Cakmak, I. (2008). Zinc deficiency in wheat in Turkey. In “Micronutrient Deficiencies inGlobal Crop Production” (B. J. Alloway, Ed.), pp. 181–200. Springer, New York.
Cakmak, I., Hengeler, C., and Marschner, H. (1994). Changes in phloem export of sucrosein leaves in response to phosphorus, potassium and magnesium deficiency in bean plants.
J. Exp. Bot. 45, 1251–1257.
Campbell, R., and Drew, M. C. (1983). Electron microscopy of gas space (aerenchyma)formation in adventitious roots of Zea mays L subjected to oxygen shortage. Planta 157,350–357.
Cannell, R. Q. (1985). Reduced tillage in North-West Europe: A review. Soil Tillage Res. 5,129–177.
Cannell, R. Q., and Hawes, J. D. (1994). Trends in tillage practices in relation to sustainablecrop production with special reference to temperate climates. Soil Tillage Res. 30,245–282.
Caradus, J. R. (1990). The structure and function of white clover root systems. Adv. Agron.43, 1–46.
Caradus, J. R., and Snaydon, R. W. (1986). Plant factors influencing phosphorus uptake bywhite clover from solution culture. I. Populations differences. Plant Soil 93, 153–163.
Champigny, M. L., and Talouizte, A. (1981). Photosynthate distribution and metabolic fatein relation to nitrogen metabolism in wheat seedlings. In “Photosynthesis: IV. Regulationof Carbon Metabolism” (G. Akoyunoglou, Ed.), pp. 645–652. Balban InternationalScience Services, Philadelphia, PA.
Chan, K. Y., and Mead, J. A. (1992). Tillage-induced differences in the growth anddistribution of wheat-roots. Aust. J. Agric. Res. 43, 19–28.
Chassot, A., Stamp, P., and Richner, W. (2001). Root distribution and morphology of maize seedlings as affected by tillage and fertilizer placement. Plant Soil 231, 123–135.
Cheng, W., and Kuzyakov, Y. (2005). Root effects on soil organic matter decomposition.In “Roots and Soil Management: Interactions Between Roots and the Soil” (R. W. Zobeland S. F. Wright, Eds.), pp. 119–143. ASA, CSSA, and SSSA, Madison, WI.
Cheng, W., Coleman, D. C., Carroll, C. R., and Hoffman, C. A. (1994). Investigatingshort-term carbon flows in the rhizospheres of different plant species, using isotopictrapping. Agron. J. 86, 782–788.
320 N. K. Fageria and A. Moreira

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 72/82
Comfort, S. D., Malzer, G. L., and Busch, R. H. (1988). Nitrogen fertilization of springwheat genotypes: Influence on root growth and soil water depletion. Agron. J. 80,114–120.
Costa, C., Dwyer, L. M., Zhou, X., Dutilleul, P., Hamel, C., Reid, L. M., and Smith, D. L.
(2002). Root morphology of contrasting maize genotypes. Agron. J. 94, 96–101.Crozier, C. R., Naderman, G. C., Tucker, M. R., and Sugg, R. E. (1999). Nutrient and pH
stratification with conventional and no-till management. Commun. Soil Sci. Plant Anal.30, 65–74.
Davidson, R. L. (1969a). Effect of root/leaf temperature differentials on root/shoot ratios inpasture grasses and clover. Ann. Bot. 33, 561–569.
Davidson, R. L. (1969b). Effects of soil nutrients and moisture on root/shoot ratios in Lolium perenne L. and Trifolium repens L. Ann. Bot. (London) 33, 571–577.
Dingkuhn, M., and Kropff, M. (1996). Rice. In “Photoassimilate Distribution in Plants andCrops” (E. Zamski and A. A. Schaffer, Eds.), pp. 519–547. Marcel Dekker, New York.
Doss, B. D., Dumas, W. T., and Lund, Z. F. (1979). Depth of lime incorporation for
correction of subsoil acidity. Agron. J. 71, 541–544.Drew, M. C. (1975). Comparison of the effects of localized supply of phosphate, nitrate,
ammonium, and potassium on the growth of the seminal root system and shoot of barley.New Phytol. 75, 470–490.
Drew, M. C., Saker, L. R., and Ashley, T. W. (1973). Nutrient supply and the growth of theseminal root system in barley. J. Exp. Bot. 24, 1189–1202.
Durieux, R. P., Kamprath, E. J., Jackson, W. A., and Moll, R. H. (1994). Root distributionof corn: The effect of nitrogen fertilization. Agron. J. 86, 958–962.
Dyer, M. I., Acra, M. A., Wang, G. M., Coleman, D. C., Freckman, D. W.,McNaughton, S. J., and Strain, B. R. (1991). Source-sink carbon relations in twoPanicum coloratum in response to herbivory. Ecology 72, 1472–1483.
Eghball, B., and Maranville, J. W. (1993). Root development and nitrogen influx of corngenotypes grown under combined drought and nitrogen stresses. Agron. J. 85, 147–152.
Eghball, B., Settimi, J. R., Maranville, J. W., and Parkhurst, A. M. (1993). Fractal analysis for morphological description of corn roots under nitrogen stress. Agron. J. 85, 287–289.
Ellis, F. B. (1979). Roots and their function in the soil. “The Yield of Cereals”, pp. 3–5.Cereal Unit National Agricultural Centre, Stoneleigh, UK.
Epstein, E., and Bloom, A. J. (2005). Mineral Nutrition of Plants: Principles and Perspecti-ves.2nd edn.. Sinauer Associations, Sunderland, MA.
Eskew, D. L., Welch, R. M., and Cary, E. E. (1983). Nickel: An essential micronutrients for legumes and possibly all higher plants. Science 222, 621–623.
Essau, K. (1977). Anatomy of Seed Plants.2nd edn.. John Wiley & Sons, New York.Evans, L. T., and Wardlaw, L. F. (1976). Aspects of the comparative physiology of grain
yield in cereals. Adv. Agron. 28, 301–359.Fageria, N. K. (1989a). Tropical Soils and Physiological Aspects of Crops. EMBRAPA,
Brasilia.Fageria, N. K. (1989b). Effect of phosphorus on growth, yield and nutrient accumulation in
the common bean. Trop. Agric. 66, 249–255.Fageria, N. K. (1991). Response of rice cultivars to phosphorus fertilization on a dark red
latosol of central Brazil. Rev. Bras. Cienc. Solo 15, 63–67.Fageria, N. K. (1992). Maximizing Crop Yields. Marcel Dekker, New York.Fageria, N. K. (2000). Adequate and toxic levels of boron for rice, common bean, corn,
soybean and wheat production in cerrado soil. Braz. J. Agric. Eng. Environ. 4, 57–62.Fageria, N. K. (2002a). Micronutrients influence on root growth of upland rice, common
bean, corn, wheat, and soybean. J. Plant Nutr. 25, 613–622.Fageria, N. K. (2002b). Nutrient management for sustainable dry bean production in the
tropics. Commun. Soil Sci. Plant Anal. 33, 1537–1575.
The Role of Mineral Nutrition on Root Growth of Crop Plants 321

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 73/82
Fageria, N. K. (2002c). Soil quality vs. environmentally based agriculture. Commun. Soil Sci.Plant Anal. 33, 2301–2329.
Fageria, N. K. (2007). Yield physiology of rice. J. Plant Nutr. 30, 843–879.Fageria, N. K. (2008). Optimum soil acidity indices for dry bean production on an Oxisol in
no-tillage system. Commun. Soil Sci. Plant Anal. 39, 845–857.Fageria, N. K. (2009). The Use of Nutrients in Crop Plants. CRC Press, Boca Raton, FL.Fageria, N. K. (2011). Root growth of upland rice genotypes as influenced by nitrogen
fertilization. J. Plant Nutr. 34, (in press).Fageria, N. K., and Baligar, V. C. (2003). Fertility management of tropical acid soils for
sustainable crop production. In “Handbook of Soil Acidity” (Z. Rengel, Ed.),pp. 359–385. Marcel Dekker, New York.
Fageria, N. K., and Baligar, V. C. (2005). Enhancing nitrogen use efficiency in crop plants. Adv. Agron. 88, 97–185.
Fageria, N. K., and Baligar, V. C. (2008). Ameliorating soil acidity of tropical Oxisols byliming for sustainable crop production. Adv. Agron. 99, 345–399.
Fageria, N. K., and Gheyi, H. R. (1999). Efficient Crop Production Federal University of Paraiba, Campina Grande, Brazil.
Fageria, N. K., and Moreira, A. (2011). Yield and yield components of upland rice asinfluenced by nitrogen sources. J. Plant Nutr. 34(in press).
Fageria, N. K., and Santos, A. B. (2008). Yield physiology of dry bean. J. Plant Nutr. 31,983–1004.
Fageria, N. K., and Santos, A. B. (2011). Lowland rice growth and development and nutrientuptake during growth cycle. J. Plant Nutr. 34(in press).
Fageria, N. K., and Souza, C. M. R. (1991). Upland rice, common bean, and cowpearesponse to magnesium application on an oxisols. Commun. Soil Sci. Plant Anal. 22,1805–1816.
Fageria, N. K., and Stone, L. F. (1999). Acidity management of cerrado and varzea soils of Brazil. Santo Antonio de Goias, Brazil.
Fageria, N. K., and Stone, L. F. (2004). Yield of common bean in no-tillage system withapplication of lime and zinc. Pesqui. Agropecu. Bras. 39, 73–78.
Fageria, N. K., and Stone, L. F. (2006). Physical, chemical, and biological changes in therhizosphere and nutrient availability. J. Plant Nutr. 29, 1327–1356.
Fageria, N. K., and Stone, L. F. (2008). Micronutrient deficiency problems in SouthAmerica. In “Micronutrient Deficiencies in Global Crop Production” (B. J. Alloway,Ed.), pp. 245–266. Springer, New York.
Fageria, N. K., Barbosa Filho, M. P., and Carvalho, J. R. P. (1981). Influence of iron ongrowth and absorption of P, K, Ca and Mg by rice plants in nutrient solution. Pesqui.
Agropecu. Bras. 16, 483–488.Fageria, N. K., Baligar, V. C., and Clark, R. B. (2002). Micronutrients in crop production.
Adv. Agron. 77, 185–268.Fageria, N. K., Castro, E. M., and Baligar, V. C. (2004). Response of upland rice genotypes
to soil acidity. In “The Red Soils of China: Their Nature, Management, and Utilization”(M. J. Wilson, Z. He, and X. Yang, Eds.), Kluwer Academic Publishers, Dordrecht,The Netherlands.
Fageria, N. K., Baligar, V. C., and Clark, R. B. (2006). Physiology of Crop Production.The Howarth Press, New York.
Fageria, N. K., Baligar, V. C., and Li, Y. C. (2008a). The role of nutrient efficient plants inimproving crop yields in the twenty first century. J. Plant Nutr. 31, 1121–1157.
Fageria, N. K., Santos, A. B., Barbosa Filho, M. P., and Guimaraes, C. M. (2008b). Irontoxicity in lowland rice. J. Plant Nutr. 31, 1676–1697.
Fageria, N. K., Moraes, O. P., and Vasconcelos, M. J. (2011). Yield and yield components of upland rice as influenced by nitrogen sources. J. Plant Nutr. 34(in press).
322 N. K. Fageria and A. Moreira

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 74/82
Feil, B., Thirapon, R., Geisler, G., and Stamp, P. (1991). The impact of temperature onseedling root traits of European and tropical corn (Zea mays L.) cultivars. J. Agron. 85,287–289.
Follett, R. F., Varvel, G. E., Kimble, J. M., and Vogel, K. P. (2009). No-till corn after
bromegrass: Effect on soil carbon and soil aggregates. Agron. J. 101, 261–268.Foy, C. D. (1984). Physiological effects of hydrogen, aluminum and manganese toxicities in
acid soils. In “Soil Acidity and Liming” (F. Adams, Ed.), 2nd edn., pp. 57–97. AmericanSociety of Agronomy, Madison, WI.
Foy, C. D. (1992). Soil chemical factors limiting plant root growth. Adv. Soil Sci. 19,97–149.
Freckman, D. W., Barker, K. R., Coleman, D. C., Acra, M. A., Dyer, M. I., Strain, B. R.,and McNaughton, S. J. (1991). The use of the 11C technique to measure plant responseto herbivorous soil nematodes. Funct. Ecol. 5, 810–818.
Fredeen, A. L., Rao, I. M., and Terry, N. (1989). Influence of phosphorus nutrition ongrowth and carbon partitioning in Glycine max. Plant Physiol. 89, 225–230.
Gabelman, W. H., Gerloff, G. C., Schettini, T., and Coltman, R. (1986). Genetic variabilityin root systems associated with nutrient acquisition and use. HortScience 21, 971–973.
Gale, W. J., and Cambardella, C. A. (2000). Carbon dynamics of surface residue and rootderived organic matter under simulated no-till. Soil Sci. Soc. Am. J. 64, 190–195.
Gale, W. J., Cambardell, C. A., and Bailey, T. B. (2000a). Root-derived carbon and theformation and stabilization of aggregates. Soil Sci. Soc. Am. J. 64, 201–207.
Gale, W. J., Cambardell, C. A., and Bailey, T. B. (2000b). Surface residue and root-derivedcarbon in stable and unstable aggregates. Soil Sci. Soc. Am. J. 64, 196–201.
Garrity, D. P., Sullivan, C. Y., and Watts, D. G. (1983). Moisture deficits and grain sorghumperformance: Drought stress conditions. Agron. J. 75, 997–1004.
Garton, R. W., and Widders, I. E. (1990). N and P preconditioning of small plug seedlings
influences growth and yield of processing tomatoes. HortScience 25, 655–657.Goldberg, S. (1993). Chemistry and mineralogy of boron in soils. In “Boron and Its Role in
Crop Production” (U. C. Gupta, Ed.), pp. 3–44. CRC Press, Boca Raton, FL.Gonzalez, E. E. (1976).Effect of depth of lime incorporation on the growth of corn in
Oxisols of central Brazil. Ph.D. Thesis, North Carolina State University, Raleigh, NC,University of Microfilms. Ann Arbor, MI (Diss. Abstract 37:5474).
Gonzalez-Erico, E., Kamprath, E. J., Nederman, G. C., and Soares, W. V. (1979). Effect of depth of lime incorporation on the growth of corn on an Oxisols of central Brazil. Soil Sci. Soc. Am. J. 43, 1155–1158.
Goss, M. J., and Kay, B. D. (2005). Soil aggregation. In “Roots and Soil Management:Interactions Between Roots and the Soil” (R. W. Zobel and S. F. Wright, Eds.),pp. 163–180. ASA, CSSA, and SSSA, Madison, WI.
Graham, R. D. (2008). Micronutrient deficiencies in crops and their global significance.In “Micronutrient Deficiencies in Global Crop Production” (B. J. Alloway, Ed.),pp. 41–61. Springer, New York.
Gregory, P. J. (1994). Root growth and activity. In “Physiology and Determination of Crop Yield” (G. A. Peterson, Ed.), pp. 65–93. ASA, CSSA, and SSSA, Madison, WI.
Gregory, P. J., and Brown, S. C. (1989). Root growth, water use and yield of crops in dryenvironments: What characteristics are desirable? Asp. Appl. Biol. 22, 235–243.
Gupta, U. C. (1993). Factors affecting boron uptake by plants. In “Boron and Its Role inCrop Production” (U. C. Gupta, Ed.), pp. 87–123. CRC Press, Boca Raton, FL.
Haberle, J. (1993). The comparison of root-system, nutrient, and water-uptake of two oldand present Czech winter wheat cultivars under different nitrogen fertilization. RostlinnaVyroba 39, 851–863.
Haberle, J., Svoboda, P., and Blaha, L. (1995). The comparison of shoot and root productionin old and new cultivars of winter cereals. Rostlinna Vyroba 41, 511–516.
The Role of Mineral Nutrition on Root Growth of Crop Plants 323

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 75/82
Haberle, J., Svoboda, P., and Ruzek, P. (1996). Root length of winter wheat and thecontent of mineral nitrogen in soil profile. Rostlinna Vyroba 42, 193–197.
Haider, K., Moiser, A., and Heinemeyer, O. (1993). Impact of growing crop plants on N-utilization from fertilizer, crop residues, and on organic N and C-mineralization. Soil Sci.
1, 183–191.Hallmark, W. B., and Barber, S. A. (1984). Root growth and morphology, nutrient uptake,
and nutrient status of early growth of soybeans as affected by soil P and K. Agron. J. 76,209–212.
Hamblin, A. P., and Tennant, A. P. (1987). Root length density and water uptake in cerealsand grain legumes; How well are they correlated? Aust. J. Agric. Res. 38, 513–527.
Hansson, A. C., and Andren, O. (1987). Root dynamics in barley, Lucerne, and meadowfescue investigated with a minirhizotron technique. Plant Soil 103, 33–38.
Hansson, A. C., Steen, E., and Andren, O. (1992). Root-growth of daily irrigated andfertilized barley investigation with ingrowth cores, soil cores, and minirhizotrons. Swed.
J. Agric. Res. 22, 141–152.
Harper, J. E. (1994). Nitrogen metabolism. In “Physiology and Determination of Crop Yield” (G. A. Peterson, Ed.), pp. 285–302. ASA, CSSA, and SSSA, Madison, WI.
Helal, H. M., and Sauerbeck, D. (1987). Direct and indirect influences of plant root onorganic matter and phosphorus turnover in soil. In “Soil Organic Matter Dynamics andSoil Productivity” (J. H. Couley, Ed.), INTECOl Bulletin 15, pp. 49–58. InternationalAssociation for Ecology, Athens, GA.
Hoad, S. P., Russell, G., Lucas, M. E., and Bingham, I. J. (2001). The management of wheat,barley, and oat root systems. Adv. Agron. 74, 193–246.
Holanda, F. S. R., Mengel, D. B., Paula, M. B., Carvalho, J. G., and Bertoni, J. C. (1998).Influence of crop rotations and tillage systems on phosphorus and potassium stratificationand root distribution in the soil profile. Commun. Soil Sci. Plant Anal. 29, 2383–2394.
Holland, E. A., and Coleman, D. C. (1987). Litter placement effects on microbial andorganic matter dynamics in an agro ecosystem. Ecology 68, 425–433.
Horiguchi, T. (1995). Rhizosphere and root oxidation activity. In “Science of the RicePlant” (T. Matsuo, K. Kumazawa, R. Ishii, K. Ishihara, and H. Hirata, Eds.), Vol. 2,pp. 221–248. Food and Agriculture Policy Research Center, Tokyo.
Hsiao, T., Steduto, L. H. P., Rojas-Lara, B., Raes, D., and Fereres, E. (2009). AquaCrop— The FAO crop model to stimulate yield response to water: III. Parameterization andtesting for maize. Agron. J. 101, 448–459.
Hurd, E. A. (1974). Phenotype and drought tolerance in wheat. Agric. Meteorol. 14, 39–55.Hurd, E. A., Townley-Smith, T. F., Patterson, L. A., and Owen, C. H. (1972). Techniques
used in producing Wascana wheat. Can. J. Plant Sci. 52, 689–691. Jackson, M. B. (1994). Root to shoot communication in flooded plants: Involvement of
abscisic acid, ethylene, and 1-aminocyclopropane-1-carboxylic acid. Agron. J. 86,775–782.
Jackson, R. B., Mooney, H. A., and Schulze, E. D. (1997). A global budget for fine rootbiomass, surface area, and nutrient contents. Proc. Natl. Acad. Sci. USA 94, 7362–7366.
Jaffar, M. N., Stone, L. R., and Goodwin, D. E. (1993). Rooting depth and dry matter development of sunflower. Agron. J. 85, 281–286.
Kamprath, E. J., and Foy, C. D. (1985). Lime–fertilizer–plant interactions in acid soils.In “Fertilizer Technology and Use” (O. P. Engelstad, Ed.), 3rd edn.. pp. 91–151. SoilScience Society of America, Madison, WI.
Kaspar, T. C., Stanley, C. D., and Taylor, H. M. (1978). Soybean root growth during thereproductive stages of development. Agron. J. 70, 1105–1107.
Kaspar, T. C., Taylor, H. M., and Shibles, R. M. (1984). Taproot elongation rates of soybean cultivars in the glasshouse and their relation to field rooting depth. Crop Sci.24, 916–920.
324 N. K. Fageria and A. Moreira

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 76/82
Kelley, P. M., and Izawa, S. (1978). The role of chloride ion in photosystem II. I. Effects of chloride ion on photosystem II electron transport and on hydroxylamine inhibition.Biochim. Biophys. Acta 502, 148–210.
Kern, J. S., and Johnson, M. G. (1993). Conservation tillage impacts on national soil and
atmospheric carbon levels. Soil Sci. Soc. Am. J. 57, 200–210.Ketring, D. L., and Reid, J. L. (1993). Growth and peanut roots under field conditions.
Agron. J. 85, 80–85.Kirchner, M. J., Wollum, A. G., and King, L. D. (1993). Soil microbial populations and
activities in reduced chemical input agro ecosystems. Soil Sci. Soc. Am. J. 57, 1289–1295.Kitchen, N. R., Buchholz, D. D., and Nelson, C. J. (1990). Potassium fertilizer and potato
leafhopper effects on alfalfa growth. Agron. J. 82, 1069–1074.Kleeper, B., Belford, R. K., and Rickman, R. W. (1984). Root and shoot development in
winter wheat. Agron. J. 76, 117–122.Klepper, B. (1992). Development and growth of crop root systems. Adv. Soil Sci. 19, 1–25.Klepper, B., and Kaspar, T. C. (1994). Rhizotrons: Their development and use in agricul-
tural research. Agron. J. 86, 745–753.Klucas, R. V., Hanus, F. J., Russell, S. A., and Evans, H. J. (1983). Nickel: A micronutrient
element for hydrogen-dependent growth of Rhizobium japonicum and for expression of urease activity in soybean leaves. Proc. Natl. Acad. Sci. USA 80, 2253–2257.
Kujira, Y., Grove, J. H., and Ronzelli, P. (1994). Varietal differences of root systems inwinter-wheat seedlings. Jpn. J. Crop Sci. 63, 524–530.
Kuo, S., Sainju, U. M., and Jellum, E. J. (1997a). Winter cover crop effects on soil organiccarbon and carbohydrate. Soil Sci. Soc. Am. J. 61, 145–152.
Kuo, S., Sainju, U. M., and Jellum, E. J. (1997b). Winter cover cropping influence onnitrogen in soil. Soil Sci. Soc. Am. J. 61, 1392–1399.
Laan, P., Benevoets, M. J., Lythe, S., Armstrong, W., and Blom, C. W. P. M. (1989). Root
morphology and aerenchyma formation as indicators of the flood-tolerance of Rumexspecies. J. Ecol. 77, 693–703.
Lampurlanes, J., Angas, P., and Cantero-Martines, C. (2001). Root growth, soil water content and yield of barley under different tillage systems on two soils in semiaridconditions. Field Crops Res. 69, 27–40.
Leon, J., and Schwang, K. U. (1992). Description and application of a screening method todetermine root morphology traits of cereal cultivars. Z. Acker Pflanzenbau 169, 128–134.
Leon, L. A., Lopez, A. S., and Vlek, P. L. G. (1985). Micronutrient problems in tropicalLatin America. Fertil. Res. 7, 95–129.
Leskovarant, D. I., and Stofella, P. J. (1995). Vegetable seedling root systems: Morphology,development, and importance. HortScience 30, 1153–1159.
Lexmond, T. M., and Vorm, P. D. J. (1981). The effect of pH on copper toxicity tohydroponically grown maize. Neth. J. Agric. Sci. 29, 217–238.
Liang, B. C., Wang, X. L., and Ma, B. L. (2002). Maize root-induced change in soil organiccarbon pools. Soil Sci. Soc. Am. J. 66, 845–847.
Lipiec, J., Hakansson, I., Tarkiewicz, S., and Kossowski, J. (1991). Soil physical propertiesand growth of spring barley as related to the degree of compactness of two soils. Soil Tillage Res. 19, 307–317.
Logan, T. J., Lal, R., and Dick, W. A. (1991). Tillage systems and soil properties in NorthAmerica. Soil Tillage Res. 20, 241–270.
Ludlow, M. M., and Muchow, R. C. (1990). A critical evaluation of traits for improvingcrop yields in water-limited environments. Adv. Agron 43, 107–153.
Mackay, A. D., and Barber, S. A. (1985). Effect of nitrogen on root growth of two corngenotypes in the field. Agron. J. 78, 659–703.
Maizlish, N. A., Fritton, D. D., and Kendall, W. A. (1980). Root morphology and earlydevelopment of maize at varying levels of nitrogen. Agron. J. 72, 25–30.
The Role of Mineral Nutrition on Root Growth of Crop Plants 325

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 77/82
Marschner, H. (1995). Mineral Nutrition of Higher Plants.2nd edn.. Academic Press,New York.
Marschner, H. (1998). Role of root growth, arbuscular mycorrhiza, and root exudates for the efficiency in nutrient acquisition. Field Crops. Res. 56, 203–207.
Martino, D. L., and Shaykewich, C. F. (1994). Root penetration profiles of wheat andbarley as affected by soil penetration resistance in field conditions. Can. J. Soil Sci. 74,193–200.
Mathews, R. B., Harris, D., Nageswara Rao, R. C., Williamds, J. H., and Wadia, K. D. R.(1988). The physiological basis for yield differences between four genotypes of ground-nut ( Arachis hypogaea) in response to drought. I. Dry matter production and water use.Exp. Agric. 24, 191–202.
Matsumoto, H., and Morimura, S. (1980). Repressed template activity of chromatin of pearoots treated by aluminum. Plant Cell Physiol. 21, 951–959.
Melillo, J. M., and Gosz, J. R. (1983). Interactions of biogeochemical cycles in forestecosystems. In “The Major Biogeochemical Cycles and Their Interactions” (B. Bolin
and R. B. Cook, Eds.), pp. 177–220. John Wiley & Sons, Chichester, England.Mengel, D. B., and Barber, S. A. (1974). Development and distribution of the corn root
system under field conditions. Agron. J. 68, 341–344.Mengel, K., and Kirkby, E. A. (1978). Principles of Plant Nutrition International Potash
Institute, Bern, Switzerland.Mengel, K., Kirkbay, E. A., Kosegarten, H., and Appel, T. (2001). Principles of Plant
Nutrition.5th edn.. Kluwer Academic Publishers, Dordrecht, The Netherlands.Merrill, S. D., Black, A. L., and Bauer, A. (1996). Conservation tillage affects root growth of
dryland spring wheat under drought. Soil Sci. Soc. Am. J. 60, 575–583.Merrill, S. D., Tanaka, D. L., and Hanson, J. D. (2002). Root length growth of eight crop
species in haplustoll soils. Soil Sci. Soc. Am. J. 66, 913–923.
Milchumas, D. G., Lauenroth, W. K., Singh, J. S., and Cole, C. V. (1985). Root turnover and production by 14C dilution: Implications of carbon portioning in plants. Plant Soil 88, 353–365.
Miller, D. E. (1986). Root systems in relation to stress tolerance. HortScience 21, 963–970.Miltner, E. D., Karnok, K. J., and Hussey, R. S. (1991). Root response of tolerant and
intolerant soybean cyst nematodes. Agron. J. 83, 571–576.Mortvedt, J. J. (2000). Bioavailability of micronutrients. In “Hand Book of Soil Science”
(M. E. Sumner, Ed.), pp. D71–D88. CRC Press, Boca Raton, FL.Mullins, G. L., Reeves, D. W., Burmester, C. H., and Bryant, H. H. (1994). In-row
subsoiling and potassium placement on root growth and potassium content of cotton. Agron. J. 86, 136–139.
Munns, D. N., and Franco, A. A. (1982). Soil constraints to legume production. In “BiologicalNitrogen Fixation for Tropical Agriculture” (P. H. Graham and S. C. Harris, Eds.), pp. 135– 152. Centro Internacional de Agricultura Tropical, Cali, Colombia.
Murphy, J. A., and Zaurov, E. (1994). Shoot and root growth response of perennial ryegrassto fertilizer placement depth. Agron. J. 86, 828–832.
Newman, E. I. (1966). A method for estimating the total length of root in a sample. J. Appl.Ecol. 3, 139–145.
Norby, R. J., and Cortrufo, M. F. (1998). A question of litter quality. Nature 206, 17–18.Nurlaeny, N., Marschner, H., and George, E. (1996). Effects of liming and mycorrhizal
colonization on soil phosphate depletion and phosphate uptake by maize ( Zea mays L.)and soybean (Glycine max L.) grown in two tropical acid soils. Plant Soil 181,275–285.
O’Toole, J. C., and Bland, W. L. (1987). Genotypic variation in crop plant root systems. Adv. Agron. 41, 91–145.
326 N. K. Fageria and A. Moreira

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 78/82
Oritani, T. (1995). Cytokinins. In “Science of the Rice Plant” (T. Matsuo, K. Kumazawa,R. Ishii, K. Ishihara, and H. Hirata, Eds.), Vol. 2, pp. 189–198. Food and AgriculturePolicy Research Center, Tokyo.
Osaki, M., Shinano, T., Matsumoto, M., Zheng, T., and Tadano, T. (1997). A root-shoot
interaction hypothesis for high productivity of field crops. In “Plant Nutrition for Sustainable Food Production and Environment” (T. Ando, K. Fujita, T. Mae,H. Matsumoto, S. Mori, and J. Sekiya, Eds.), pp. 669–674. Kluwer Academic Publishers,Dordrecht, The Netherlands.
Pan, W. L., Jackson, W. A., and Moll, R. H. (1985). Nitrate uptake and portioning by corn(Zea mays L.) root systems and associated morphological differences among genotypesand stages of root development. J. Exp. Bot. 36, 1341–1351.
Parker, C. J., Carr, M. K. V., Jarvis, N. J., Puplampu, B. O., and Lee, V. H. (1991). Anevaluation of the minirhizotron technique for estimating root distribution in potatoes.
J. Agric. Sci. 116, 341–350.Parton, W. J., Ojima, D. S., and Schimel, D. S. (1996). Models to evaluate soil organic
matter storage and dynamics. In “Structure and Organic Matter Storage in Soils”(M. R. Carter and B. A. Stewart, Eds.), pp. 421–448. CRC Press, Boca Raton, FL.
Peterson, C. M., Klepper, B., Pumphrey, F. V., and Rickman, R. W. (1984). Restrictedrooting decreases tillering and growth of winter wheat. Agron. J. 76, 861–863.
Pinton, R., and Varannini, Z. (2001). The rhizosphere as a site of biochemical interactionsamong soil components, plants and microorganisms. In “The Rhizosphere: Biochemicaland Organic Substances at the Soil–Plant Interface” (R. Pinto, Z. Varanini, andP. Nannipieri, Eds.), pp. 1–17. Marcel Dekker, New York.
Puget, P., and Drinkwater, L. E. (2001). Short-term dynamics of root- and shoot-derivedcarbon from a leguminous green manure. Soil Sci. Soc. Am. J. 65, 771–779.
Qian, J. H., and Doran, J. W. (1996). Available carbon released from crop roots during
growth as determined by carbon-13 natural abundance. Soil Sci. Soc. Am. J. 60, 828–831.Qin, R., Stamp, P., and Richner, W. (2004). Impact of tillage on root systems of winter
wheat. Agron. J. 96, 1523–1530.Qin, R., Stamp, P., and Richner, W. (2005). Impact of tillage and banded starter fertilizer on
maize root growth in the top 25 centimeters of the soil. Agron. J. 97, 674–683.Rasmussen, J. K. (1991). Reduced soil tillage and Italian ryegrass as catch crop. II. Soil bulk
density, root development and soil chemistry. Tidsskr. Planteavl. 95, 139–154.Reichman, G. A., and Trooien, T. P. (1993). Corn yield and salinity response to irrigation
on slowly permeable subsoil. Soil Sci. Soc. Am. J. 57, 1549–1554.Rengel, Z., and Robinson, D. L. (1989). Aluminum effects on growth and micronutrient
uptake by annual ryegrass. Agron. J. 81, 208–215.Reuter, D. J., Alston, A. M., and McFarland, J. D. (1988). Occurrence and correction of
manganese deficiency in plants. In “Manganese in Soils and Plants” (R. D. Graham,R. J. Hannam, and N. C. Uren, Eds.), pp. 205–224. Kluwer Academic Publisher,Dordrecht, The Netherlands.
Ritchey, K. D., Souza, D. M. G., Lobato, E., and Correa, O. (1980). Calcium leaching toincrease rooting depth in Brazilian savannah Oxisol. Agron. J. 72, 40–44.
Robinson, D., Linehan, D. J., and Gordon, D. C. (1994). Capture of nitrate from soil bywheat in relation to root length, nitrogen inflow, and availability. New Phytol. 128,297–305.
Romer, W. (1985). The influence of shoot and root-growth on the uptake of phosphorus incereals. Bodenkultur 36, 203–211.
Roth, G. W., Calvin, D. D., and Lweloff, S. M. (1995). Tillage, nitrogen timing, andplanting date effects on western corn rootworm injury to corn. Agron. J. 87, 189–193.
Rufty, T. W. Jr., Israel, D. W., Volk, R. J., Qui, J., and Sa, T. (1993). Phosphate regulationof nitrate assimilation in soybean. J. Exp. Bot. 44, 879–891.
The Role of Mineral Nutrition on Root Growth of Crop Plants 327

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 79/82
Russell, R. S. (1977). Plant Root Systems. McGraw-Hill, New York.Sainju, U. M., Singh, B. P., and Whitehead, W. F. (2001). Comparison of the effects of
cover crops and nitrogen fertilization on tomato yield, root growth, and soil properties.Sci. Hortic. 91, 201–214.
Sainju, U. M., Singh, B. P., and Whitehead, W. F. (2005a). Tillage, cover crops, andnitrogen fertilization effects on cotton and sorghum root biomass, carbon, and nitrogen.
Agron. J. 97, 1279–1290.Sainju, U. M., Whitehead, W. F., and Singh, B. P. (2005b). Carbon accumulation in cotton,
sorghum, and underlying soil as influenced by tillage, cover crops, and nitrogen fertiliza-tion. Plant Soil 273, 219–234.
Sanchez, J. E., Paul, E. A., Wilson, T. C., Smeenk, J., and Harwood, R. R. (2002). Corn rooteffects on the nitrogen supplying capacity of a conditioned soil. Agron. J. 94, 391–396.
Sanders, J. L., and Brown, D. A. (1978). A new fiber-optic technique for measuring rootgrowth of soybean under field conditions. Agron. J. 70, 1073–1076.
Schenk, M. K., and Barber, S. A. (1980). Potassium and phosphorus uptake by corn
genotype grown in the field as influenced by root characteristics. Plant Soil 54, 65–75.Shalhevet, J., Huck, M. G., and Schroeder, B. P. (1995). Root and shoot growth responses
to salinity in maize and soybean. Agron. J. 87, 512–516.Sharma, B. R., and Chaudhary, T. N. (1984). Nitrogen uptake by wheat in relation to water
regime and depth of N placement in a loamy sand soil: Measurement and modelsimulation. J. Soil Sci. 35, 141–148.
Sharratt, B. S., and Cochran, V. L. (1993). Skip-row and equidistant-row barley withnitrogen placement; Yield, nitrogen uptake and root density. Agron. J. 85, 246–250.
Sidiras, N., Bilalis, D., and Vavoulidou, E. (2001). Effects of tillage and fertilization on someselected physical properties of soil (0–30 cm depth) and on the root grown dynamic of winter barley (Hordeum vulgare cv. Niki). J. Agron. Crop Sci. 187, 167–176.
Silberbush, M., and Barber, S. A. (1984). Phosphorus and potassium uptake of field grownsoybeans predicted by a simulation model. Soil Sci. Soc. Am. J. 48, 592–596.
Sinclair, A. H., and Edwards, A. C. (2008). Micronutrient deficiency problems in agricul-tural crops in Europe. In “Micronutrient Deficiencies in Global Crop Production”(B. J. Alloway, Ed.), pp. 225–266. Springer, New York.
Singh, M. V. (2008). Micronutrient deficiencies in crop and soils in India. In “MicronutrientDeficiencies in Global Crop Production” (B. J. Alloway, Ed.), pp. 93–123. Springer,New York.
Slaton, N. A., and Beyrouty, C. A. (1992). Rice shoot response to root confinement within amembrane. Agron. J. 84, 50–53.
Slaton, N. A., Beyrouty, C. A., Wells, B. R., Norman, R. J., and Gbur, E. E. (1990). Rootgrowth and distribution of two short-season rice genotypes. Plant Soil 126, 269–278.
Soil Science Society of America. (2008). Glossary of Soil Science Terms. ASA, Madison, WI.Sponchiado, B. N., White, J. W., Castillo, J. A., and Jones, P. G. (1989). Root growth of
four common bean cultivars in relation to drought tolerance in environments withcontrasting soil types. Exp. Agric. 25, 249–257.
St. Aubin, G., Canny, M. J., and McCully, M. E. (1986). Living vessel elements in the latemetaxylem of sheathed maize roots. Ann. Bot. 58, 577–588.
Steponkus, P. J., Cutler, J. M., and O’Toole, J. C. (1980). Adaptation to water stress in rice.In “Adaptation of Plants to Water and High Temperature Stress” (N. C. Turner andP. J. Kramer, Eds.), pp. 401–418. John Wiley & Sons, New York.
Stoffella, P. J., and Kahn, B. A. (1986). Root system effects on lodging of vegetable crops.HortScience 21, 960–963.
Stoffella, P. J., Sandsted, R. F., Zobel, R. W., and Hynes, W. L. (1979). Root characteristicsof some black beans. I. Relationship of root size to lodging and seed yield. Crop Sci. 19,823–826.
328 N. K. Fageria and A. Moreira

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 80/82
Stone, L. F., and Pereira, A. L. (1994a). Rice-common bean rotation under sprinkler irrigation: Effects of row spacing, fertilization and cultivar on growth, root developmentand water consumption of common bean. Pesqui. Agropecu. Bras. 29, 939–954.
Stone, L. F., and Pereira, A. L. (1994b). Rice-common bean rotation under sprinkler
irrigation: Effects of row spacing, fertilization and cultivar on growth, root developmentand water consumption of rice. Pesqui. Agropecu. Bras. 29, 1577–1592.
Stone, L. R., Goodrum, D. E., Jaafar, M. N., and Khan, A. H. (2001). Rooting front andwater depletion depths in grain sorghum and sunflower. Agron. J. 93, 1105–1110.
Takenaga, H. (1995). Internal factors in relation to nutrient absorption. In “Science of theRice Plant” (T. Matsuo, K. Kumazawa, R. Ishii, K. Ishihara, and H. Hirata, Eds.), Vol. 2,pp. 294–309. Food and Agriculture Policy Research Center, Tokyo.
Talouizte, A. M., Champigny, M. L., Bismuth, E., and Moyse, A. (1984). Root carbohy-drate metabolism associated with nitrate assimilation in wheat previously deprived of nitrogen. Physiol. Veg. 22, 19–27.
Taylor, H. M., and Ratliff, L. F. (1969). Root elongation rates of cotton and peanuts as a
function of soil strength and soil water content. Soil Sci. 108, 113–119.Taylor, H. M., Burnett, E., and Booth, G. D. (1978). Taproot elongation rates of soybeans.
Z. Acker Pflanzenbau 146, 33–39.Tennant, D. (1976). Root growth of wheat. I. Early patterns of multiplication and extension
of wheat roots including effects of leaves of nitrogen, phosphorus, and potassium. Aust. J. Agric. Res. 27, 183–196.
Teo, Y. H., Beyrouty, C. A., and Gbur, E. E. (1995). Evaluation of a model to predictnutrient uptake by field grown rice. Agron. J. 87, 7–12.
Texier, M., and Biles, G. (1990). The role of rhizosphere on C and N cycles in a plant–soilsystem. Symbiosis 9, 117–123.
Thangaraj, M., O’Tolle, J. C., and De Datta, S. K. (1990). Root response to water stress in
rainfed lowland rice. Exp. Agric. 26, 287–296.Thomas, A. L., and Kaspar, T. C. (1997). Maize nodal root response to time of soil ridging.
Agron. J. 89, 195–200.Tian, G., Carsky, R. J., and Kang, B. T. (1998). Differential phosphorus responses of
leguminous cover crops on soils with variable history. J. Plant Nutr. 21, 1641–1653.Tresder, K. K., Morris, S. J., and Allen, M. F. (2005). The contribution of root exudates,
symbionts, and detritus to carbon sequestration in the soil. In “Roots and Soil Manage-ment: Interactions Between Roots and the Soil” (R. W. Zobel and S. F. Wright, Eds.),pp. 145–162. ASA, CSSA, and SSSA, Madison, WI.
Troughton, A. (1962). The roots of temperate cereals (wheat, barley, oats and rye). Mimeo-graphed Publication No. 2 Commonwealth Bureau of Pasture and Field Crops, Hurley,Berkshire.
Tupper, G. R. (1992). Technologies to solve K deficiency: Deep placement. In “1992Proceedings of the Beltwide Cotton Production Research Conference,” pp. 73–76.National Cotton Council of America, Memphis, TN.
Ueda, S. (1936). On the development of root system of wheat and barley in sandy soil. Proc.Crop Sci. Soc. Jpn. 8, 140–143.
Unger, P. W., and Kaspar, T. C. (1994). Soil compaction and root growth: A review. Agron. J. 86, 759–766.
University of Arkansas Cooperative Extension Service Rice Committee. (1990). RiceProduction Handbook. University of Arkansas Cooperative Extension Service Miscella-neous Publication No. 192 University of Arkansas, Little Rock, AR.
Vartanian, N. (1981). Some aspects of structural and functional modifications induced bydrought in root systems. Plant Soil 63, 83–92.
Vincent, C. D., and Gregory, P. J. (1986). Differences in the growth and development of chickpea seedling roots (Cicer arietinum). Exp. Agric. 22, 233–242.
The Role of Mineral Nutrition on Root Growth of Crop Plants 329

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 81/82
Waals, J. H. V., and Laker, M. C. (2008). Micronutrient deficiencies in crops in Africa withemphasis on southern Africa. In “Micronutrient Deficiencies in Global Crop Produc-tion” (B. J. Alloway, Ed.), pp. 201–224. Springer, New York.
Wahbi, A., and Gregory, P. J. (1995). Growth and development of young roots of barley
(Hordeum vulgare L.) genotype. Ann. Bot. 75, 533–539.Wardlaw, I. A. (1990). The control of carbon partitioning in plants. New Phytol. 116,
341–381.Webb, J., and Jackson, M. B. (1986). A transmission and cryo-scanning electron microscope
study of the formation of aerenchyma (cortical gas filled space) in adventitious roots of rice (Oryza sativa L.) J . Exp. Bot. 37, 832–841.
Wedin, D. A., and Tilman, D. (1990). Species effects on nitrogen cycling: A test withperennial grasses. Oecologia 84, 433–441.
Welbank, P. J., and Williams, E. D. (1968). Root growth of a barley crop estimated bysampling with portable powered soil coring equipment. J. Appl. Ecol. 5, 447.
Welch, R. M. (2008). Linkages between trace elements in food crops and human health.
In “Micronutrient Deficiencies in Global Crop Production” (B. J. Alloway, Ed.),pp. 287–317. Springer, New York.
Welch, R. M., Allaway, W. H., House, W. A., and Kubota, J. (1991). Geographicdistribution of trace element problems. In “Micronutrients in Agriculture”(J. J. Mortvedt, F. R. Cox, L. M. Shuman, and R. M. Welch, Eds.), 2nd edn., pp. 31–57.Soil Science Society of America, Madison, WI.
Wenkert, W., Fausey, N. R., and Watters, H. D. (1981). Flooding Responses in Zea mays L.Plant Soil 62, 351–366.
Werf, A. V. D. (1996). Growth analysis and photoassimilate partitioning. In “PhotoassimilateDistribution in Plants and Crops” (E. Zamski and A. A. Schaffer, Eds.), pp. 1–20. MarcelDekker, New York.
Weston, L. A., and Zandstra, B. H. (1989). Transplant age and N and P nutrition effects ongrowth and yield of tomatoes. HortScience 24, 88–90.
White, J. W., and Castillo, J. A. (1989). Relative effect of root and shoot genotypes on yieldof common bean under drought stress. Crop Sci. 29, 360–362.
Wiesler, F., and Horst, W. J. (1993). Differences among maize cultivars in the utilization of soil nitrate and the related losses of nitrate through leaching. Plant Soil 151, 193–203.
Wilson, A. J., Robards, A. W., and Goss, M. J. (1977). Effects of mechanical impedance onroot growth in barley, Hordeum vulgare L.: II. Effects on cell development in seminalroots. J. Exp. Bot. 28, 1216–1227.
Wilts, A. A., Reicosky, D. C., Allmaras, R. R., and Clapp, C. E. (2004). Long-term cornresidues effects; Harvest alternatives, soil carbon turnover, and root-derived carbon. Soil Sci. Soc. Am. J. 68, 131342–131351.
Wulfsohn, D., Gu, Y., Wulfsohn, A., and Mojlaj, E. G. (1996). Statistical analysis of wheatroot growth patterns under conventional and no-till systems. Soil Tillage Res. 38, 1–16.
Xiaoe, Y., Romheld, V., Marschner, H., Baligar, V. C., and Martens, D. C. (1997). Shootphotosynthesis and root growth of hybrid and conventional rice cultivars as affected by Nand K levels in the root zone. Pedosphere 7, 35–42.
Yamauchi, A., Kono, Y., and Tatsumi, J. (1987a). Qualitative analysis of root systemstructure of upland rice and maize. Jpn. J. Crop Sci. 56, 608–617.
Yamauchi, A., Kono, Y., and Tatsumi, J. (1987b). Comparison of root system structures of 13 species of cereals. Jpn. J. Crop Sci. 56, 618–631.
Yibrin, H., Johnson, J. W., and Eckert, D. J. (1993). No-till corn production as affected bymulch, potassium placement, and soil exchangeable potassium. Agron. J. 85, 639–644.
Yoshida, S. (1981). Fundamentals of Rice Crop Science. IRRI, Los Banos, Philippines.
330 N. K. Fageria and A. Moreira

8/16/2019 Mineral Nutrition on Root Growth
http://slidepdf.com/reader/full/mineral-nutrition-on-root-growth 82/82
Yoshida, S., and Hasegawa, S. (1982). The rice root system: Its development and function.In “Drought Resistance of Crops with Emphasis on Rice” (IRRI, Ed.), pp. 97–114.IRRI, Los Banos, Philippines.
Zak, D. R., and Pregitzer, K. S. (1998). Integration of ecophysiological and biogeochemical
approaches to ecosystem dynamics. In “Success, Limitations, and Frontiers in EcosystemScience” (M. L. Pace and P. M. Groffman, Eds.), pp. 372–403. Springer, New York.
Zhang, J., and Barber, S. A. (1992). Maize root distribution between phosphorus fertilizedand unfertilized soil. Soil Sci. Soc. Am. J. 56, 819–822.
Zhang, J., and Barber, S. A. (1993). Corn root distribution between ammonium fertilizedand unfertilized soil. Commun. Soil Sci. Plant Anal. 24, 411–419.
Zhao, Z. R., Li, G. R., and Hung, G. Q. (1991). Promotive effect of potassium onadventitious root formation in some plants. Plant Sci. 79, 47–50.
Zhao, Y., Xiao, X., Bi, D., and Hu, F. (2008). Effects of sulfur fertilization on soybean rootand leaf traits, and soil microbial activity. J. Plant Nutr. 31, 473–483.
Zobel, R. W. (1986). Rhizogenetics (root genetics) of vegetable crops. HortScience 21,
956–959.Zobel, R. W. (1991). Root growth and development. In “The Rhizosphere and Plant
Growth” (D. L. Keister and P. B. Cregan, Eds.), pp. 61–71. Kluwer Academic Publishers,Dordrecht, The Netherlands.
Zobel, R. W. (2003). Sensitivity analysis of computer based diameter measurement fromdigital images. Crop Sci. 43, 583–591.
Zobel, R. W. (2005a). Primary and secondary root systems. In “Roots and Soil Manage-ment: Interactions Between Roots and the Soil” (R. W. Zobel and S. F. Wright, Eds.),pp. 3–14. ASA, CSSA, and SSSA, Madison, WI.
Zobel, R. W. (2005b). Tertiary root systems. In “Roots and Soil Management: InteractionsBetween Roots and the Soil” (R. W. Zobel and S. F. Wright, Eds.), pp. 35–56. ASA,
CSSA, and SSSA, Madison, WI.Zou, C., Gao, X., Shi, R., Fan, X., and Zhang, F. (2008). Micronutrient deficiencies in crop
production in China. In “Micronutrient Deficiencies in Global Crop Production”(B. J. Alloway, Ed.), pp. 127–148. Springer, New York.
The Role of Mineral Nutrition on Root Growth of Crop Plants 331
Related Documents











