ORIGINAL ARTICLE Middle Lutetian climate in the Paris Basin: implications for a marine hotspot of paleobiodiversity Damien Huyghe • Didier Merle • Franck Lartaud • Emilie Cheype • Laurent Emmanuel Received: 8 December 2011 / Accepted: 29 March 2012 / Published online: 18 April 2012 Ó Springer-Verlag 2012 Abstract The present study reports the evolution of environmental conditions and seawater temperatures during the establishment of a marine hotspot of paleobiodiversity that took place in the Paris Basin during the Lutetian. The stable isotope compositions (d 18 O and d 13 C) of three spe- cies of molluscs (two bivalves: Cubitostrea plicata and Venericardia imbricata, and one gastropod: Sigmesalia multisulcata) collected along the reference section of Grignon (Falunie `re) are used for paleoenvironmental and paleoclimatic reconstructions. Additional high-resolution analyses on one specimen of Haustator imbricatarius allow the documentation of seasonal changes for tempera- ture. The high-resolution profiles of the d 18 O signatures of S. multisulcata reveal that these gastropods mineralized their shell during the warm months of the year, as did V. imbricata, which probably had a short life span (less than 1 year). These two species thus only yield temperatures for the summer period, from 22 to 30 °C. The d 18 O of C. pli- cata shells indicate mean annual sea surface temperatures ranging between 15 and 23 °C during the Middle Lutetian, with minimal temperatures probably reflecting greater depth at the base of the section. The seasonal contrasts reconstructed in the upper part of the section, from the large gastropod H. imbricatarius, ranged between 18 and 30 °C. Comparison of the isotopic values of the species indicates that the d 13 C of the three taxa seems to be mostly influenced by ecological features, leading to differences between en- dobenthic (V. imbricata) and epibenthic species (C. pli- cata); or the food habits. The paleoclimatic reconstructions show that the Lutetian climate was relatively stable in the Paris Basin with long-term cooling of the mean annual sea- surface temperatures. Nevertheless, this study shows that despite a context of colder conditions compared to the Early Eocene, the climate provided a favorable context for the increase of marine biodiversity in the Paris Basin during the Middle Lutetian. Keywords Lutetian Paris Basin Molluscs Climate Stable isotopes Paleobiodiversity Introduction In the Paris Basin, the Lutetian is a stage corresponding to the greatest shallow-water marine biodiversity during the Cenozoic and even one of the highest past biodiversity Electronic supplementary material The online version of this article (doi:10.1007/s10347-012-0307-3) contains supplementary material, which is available to authorized users. D. Huyghe F. Lartaud E. Cheype L. Emmanuel UMR 7193, ISTeP, UPMC University of Paris 06, 75005 Paris, France D. Huyghe F. Lartaud E. Cheype L. Emmanuel UMR 7193, ISTeP, CNRS, 75005 Paris, France D. Huyghe (&) Laboratoire des Fluides Complexes et leurs Re ´servoirs, I.P.R.A., Universite ´ de Pau et des Pays de l’Adour, BP 1155, 64013 Pau Cedex, France e-mail: [email protected] D. Merle De ´partement Histoire de la Terre, UMR CNRS 7207, CR2P, CP 38, Muse ´um National d’Histoire Naturelle, 8, rue Buffon, 75005 Paris, France F. Lartaud CNRS FRE 3350, Lab. Ecoge ´ochimie des environnements benthiques (LECOB), Observatoire Oce ´anologique, UPMC University of Paris 06, 66650 Banyuls-sur-mer, France 123 Facies (2012) 58:587–604 DOI 10.1007/s10347-012-0307-3

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ORIGINAL ARTICLE
Middle Lutetian climate in the Paris Basin: implicationsfor a marine hotspot of paleobiodiversity
Damien Huyghe • Didier Merle • Franck Lartaud •
Emilie Cheype • Laurent Emmanuel
Received: 8 December 2011 / Accepted: 29 March 2012 / Published online: 18 April 2012! Springer-Verlag 2012
Abstract The present study reports the evolution ofenvironmental conditions and seawater temperatures during
the establishment of a marine hotspot of paleobiodiversity
that took place in the Paris Basin during the Lutetian. Thestable isotope compositions (d18O and d13C) of three spe-
cies of molluscs (two bivalves: Cubitostrea plicata and
Venericardia imbricata, and one gastropod: Sigmesaliamultisulcata) collected along the reference section of
Grignon (Faluniere) are used for paleoenvironmental and
paleoclimatic reconstructions. Additional high-resolutionanalyses on one specimen of Haustator imbricatarius
allow the documentation of seasonal changes for tempera-ture. The high-resolution profiles of the d18O signatures of
S. multisulcata reveal that these gastropods mineralized
their shell during the warm months of the year, as did V.imbricata, which probably had a short life span (less than
1 year). These two species thus only yield temperatures for
the summer period, from 22 to 30 "C. The d18O of C. pli-cata shells indicate mean annual sea surface temperatures
ranging between 15 and 23 "C during the Middle Lutetian,
with minimal temperatures probably reflecting greaterdepth at the base of the section. The seasonal contrasts
reconstructed in the upper part of the section, from the large
gastropod H. imbricatarius, ranged between 18 and 30 "C.Comparison of the isotopic values of the species indicates
that the d13C of the three taxa seems to be mostly influenced
by ecological features, leading to differences between en-dobenthic (V. imbricata) and epibenthic species (C. pli-cata); or the food habits. The paleoclimatic reconstructions
show that the Lutetian climate was relatively stable in theParis Basin with long-term cooling of the mean annual sea-
surface temperatures. Nevertheless, this study shows thatdespite a context of colder conditions compared to the Early
Eocene, the climate provided a favorable context for the
increase of marine biodiversity in the Paris Basin during theMiddle Lutetian.
Keywords Lutetian ! Paris Basin ! Molluscs ! Climate !Stable isotopes ! Paleobiodiversity
Introduction
In the Paris Basin, the Lutetian is a stage corresponding to
the greatest shallow-water marine biodiversity during the
Cenozoic and even one of the highest past biodiversity
Electronic supplementary material The online version of thisarticle (doi:10.1007/s10347-012-0307-3) contains supplementarymaterial, which is available to authorized users.
D. Huyghe ! F. Lartaud ! E. Cheype ! L. EmmanuelUMR 7193, ISTeP, UPMC University of Paris 06,75005 Paris, France
D. Huyghe ! F. Lartaud ! E. Cheype ! L. EmmanuelUMR 7193, ISTeP, CNRS, 75005 Paris, France
D. Huyghe (&)Laboratoire des Fluides Complexes et leurs Reservoirs, I.P.R.A.,Universite de Pau et des Pays de l’Adour, BP 1155,64013 Pau Cedex, Francee-mail: [email protected]
D. MerleDepartement Histoire de la Terre, UMR CNRS 7207, CR2P,CP 38, Museum National d’Histoire Naturelle, 8, rue Buffon,75005 Paris, France
F. LartaudCNRS FRE 3350, Lab. Ecogeochimie des environnementsbenthiques (LECOB), Observatoire Oceanologique, UPMCUniversity of Paris 06, 66650 Banyuls-sur-mer, France
123
Facies (2012) 58:587–604
DOI 10.1007/s10347-012-0307-3

known in the world (Lozouet 1997; Merle 2008). This
elevated biodiversity is documented by around 2,700marine species, which include 1,550 gastropod species and
540 bivalve species (Fig. 1; Pacaud and Le Renard 1995;
Merle 2008). In the American Gulf Coast, a significantincrease in species richness is also observed during the
Lutetian (Hansen 1988) and thus, the increase in mollusc
species richness observed in the Paris Basin cannot beregarded only as a local event. However, the species
richness of the Paris Basin is 5.5 times higher than that ofthe Gulf Coast (Merle 2008). It is difficult to determine if
biases resulting from the quality of the geological record
(Crampton et al. 2011; Zuschin et al. 2011; Smith andMcGowan 2011) or from the effort of sampling (Bouchet
et al. 2002; Kantor and Sisoev 2005) drive to strengthen
this increase in the Paris Basin and make that it is sospectacular, but from the Ypresian to the Bartonian, the
preservation of the shells displays the same quality and an
artefact resulting from greater preservation of fossils can beexcluded.
The deposition of a vast carbonate platform in the Paris
Basin with various biofacies (Gely 2008), a paleogeo-graphic situation located in border of the Atlantic Ocean
and northern domains and close to the Tethys Ocean rep-
resent favorable global conditions to the preservation of ahotspot (Merle 2008). However, such an increase in species
richness during the Lutetian is surprising because this
period corresponds to an interval of relative cooling(Fig. 1), with more and more evidence arguing for the
presence of continental ice-sheets in both hemispheres
(e.g., Pekar et al. 2005; Tripati et al. 2005, 2008). Addi-tionally, this stage is located between two periods of global
warming, corresponding to the Early Eocene climaticoptimum (EECO) between 54 and 49 Ma (Zachos et al.
2001), i.e., during the Ypresian and the Middle Eocene
climatic optimum (MECO), near 40 Ma, at the beginningof the Bartonian (Bohaty et al. 2009). Theoretically, these
two warm periods should have constituted a better climatic
context for the establishment of a ‘‘hotspot’’ of marinebiodiversity, due to higher seawater temperatures (Miller
et al. 1987; Lear et al. 2000; Zachos et al. 2001, 2008).
Although the Paleogene global climatic evolution isrelatively well documented in the oceanic realm (e.g.,
Shackleton and Kennett 1975; Miller et al. 1987; Zachos
et al. 2001, 2008; Bohaty et al. 2009), few studies deal withnear-shore environments (e.g., Tivollier and Letolle 1968;
Buchardt 1978; Purton and Brasier 1997, 1999; Kobashi
et al. 2001; Tripati et al. 2001; Kobashi and Grossman2003; Ivany et al. 2008). The oxygen isotopic composition
(d18O) of mollusc shells has been shown to be a reliable
proxy for changes in coastal seawater temperature andsalinity (e.g., Epstein et al. 1953; Anderson and Arthur
1983; Grossman and Ku 1986; Surge et al. 2001). Seawater
d18O (d18Ow) depends on the degree of freshwater influxand/or evaporation, often resulting in a positive correlation
between salinity and d18Ow (Pierre 1999; Schmidt 1999).
Such a dependency must be taken into account for thereconstruction of paleo-seawater temperatures using
ancient mollusc shells because some species can tolerate
considerable salinity fluctuations. It is therefore necessaryto integrate other paleontological, sedimentological, and/or
geochemical data when interpreting the d18O values as
paleotemperatures. In addition to the detection of climatechanges on thousand to million year time-scales, mollusc
carbonate shells can also record infra-annual temperature
variations (Kirby et al. 1998; Andreasson and Schmitz2000; Lartaud et al. 2010a; Titschack et al. 2010), pro-
viding supplementary information on the paleoclimatic
context. Eocene seasonal temperature investigations arehowever sparse (e.g., Andreasson and Schmitz 2000;
Kobashi et al. 2001).
The aim of this study is to document the paleoclimaticand the paleoenvironmental contexts during the Lutetian in
which the ‘‘hotspot’’ of paleobiodiversity is documented.
The outcrop of Faluniere at Grignon (Yvelines, France), inthe Paris Basin, was chosen as a key reference because of
Pal
eoce
neE
ocen
eO
ligo
-cen
e
Danian
Selandian
Thanetian
Ypresian
Lutetian
Bartonian
Priabonian
Rupelian30
35
40
45
50
55
60
65
Age
(M
a)
EECO
Oi1 glaciation
PETM
MECO
BivalvesGastropods
3 2 1 0 -1
0 4 8 12Ice-free temperatures (°C)
! O (‰ VPDB)18
0 400 800 1200 1600
Number of species
Maximummarinebiodiversity
Lutetiancooling
Fig. 1 Evolution of species richness of bivalves and gastropods inthe Paris Basin during the Paleogene (from Merle 2008) compared tothe evolution of the d18O of deep-sea benthic foraminifers (afterZachos et al. 2008). PETM refers to the Paleocene–Eocene Thermalmaximum, EECO to the Early Eocene climatic optimum, MECO tothe Middle Eocene climatic optimum and Oi1 glaciation to theglaciation at the Eocene–Oligocene boundary
588 Facies (2012) 58:587–604
123

its very diversified and well-preserved faunal assemblage.
It contains more than 800 mollusc species and 190 fora-minifer species (Le Calvez and Le Renard 1980; Merle and
Courville 2008). The recent refreshment of the outcrop
allows sampling across the Middle Lutetian.The three most represented mollusc species along the
section were chosen to reconstruct the variations of envi-
ronmental conditions from their stable isotope composi-tions (d18O and d13C): two species of bivalves, Cubitostreaplicata (Solander in Brander, 1766) and Venericardiaimbricata (Gmelin, 1791), and one species of gastropod,
Sigmesalia multisulcata (Lamarck, 1804). Moreover, the
high-resolution analyses of the isotopic ratio of threeS. multisulcata gastropod shells, one Haustator imbrica-tarius (Lamarck, 1804), a large gastropod shell and one
C. plicata oyster shell, were performed to investigate theseasonal variations of temperatures.
Previous paleoclimatic work
Paleoenvironmental and paleoclimatic reconstructionsfrom stable isotopes of mollusc shells have already been
tried with specimens from the section of Grignon (Andre-
asson and Schmitz 1996). However, these samples camefrom an old collection from the Museum National d’His-
toire Naturelle (Paris) and their exact location in the
present section of Grignon is unknown. The two bivalves(Venericardia) and two gastropods (Haustator = Turritel-la) shells used by Andreasson and Schmitz (1996) show
that the mean annual temperatures (MAT) were about21 "C, with temperatures ranging between 14 "C in winter
and 28 "C in summer.
Klein et al. (1997), however, argued that ‘‘The stratig-raphy of the section at Grignon suggests that the fossils
could have secreted their shells in seawater diluted by fresh
water, which would give rise to erroneously high d18Oestimates of paleotemperature’’. This criticism flows from
the fact that Andreasson and Schmitz (1996) did not pro-
vide the precise location of the samples in Grignon section,which extends from Zone III to Zone IV of Abrard (1925).
In contrast, Klein et al. (1997) argued that the salinity
was probably under-estimated by Andreasson and Schmitz(1996), based on the presence of miliolids in the sediments,
which indicate hypersaline conditions. However, according
to Andreasson and Schmitz (1997), the presence of rotalidforaminifers such as Discorbis, Cibicides and Nonion,
whose modern equivalents are observed in normal marine
environments, does not indicate strong hypersalinity.The contradictions raised by these authors highlight the
difficulties of constraining the paleosalinities and of inter-
preting the stable isotope signature in mollusc shells aspaleotemperatures. This is why it is indispensable to
present isotopic values in a well-defined stratigraphic
context. A revision of the paleoenvironmental changes thatoccur in the section of Grignon, based on paleontological,
sedimentological, and geochemical proxies, will yield
more accurate constraints on the climatic reconstitutions.
Geological setting
The Paris Basin is an intracratonic basin, with reducedtectonic activity during the Middle Eocene (Cavelier and
Pomerol 1979; Brunet and Le Pichon 1982; Guillocheau
et al. 2000). The sediments from the ‘‘Faluniere de Gri-gnon’’ are correlated of the Middle Lutetian, according to
the identification of biozone NP15 (Aubry 1985). They
belong to the regional Zone III and the base of Zone IV ofAbrard (1925) and correspond to parasequences A6–A10
of Gely (1996). Recent paleogeographic reconstructions,
for parasequence A7 and the base of parasequence A8,placed Grignon on the southwestern border of the basin,
close to the sea-shore and the Bray anticline (Fig. 2; Gely
2008). During this period, the Paris Basin was connected tothe North Sea and the Atlantic Ocean via the English
Channel (Fig. 2; Pomerol 1973; Gely 2008).
Description of the section
As seen above with the criticism of Klein et al. (1997)against the results of Andreasson and Schmitz (1996), it is
essential to define the sedimentological and stratigraphic
context to reconstruct the paleoclimatic evolution. Thefamous site of Grignon was studied by several great nat-
uralists such as Lamarck, Cuvier, and Brongniart who were
interested in its remarkable fossil richness, but, paradoxi-cally, no complete section was described for this quarry.
Abrard (1925) and Le Calvez and Le Renard (1980)
described the sedimentary succession, but they only gave acomposite section, corresponding to different exposures
found in the park of Grignon. Figure 9 of Abrard (1925)
clearly indicates the place of these different exposures(Trou rouge, Faluniere, Route de la Maugere and Cotes-
aux-Buis). In contrast, Le Calvez and Le Renard (1980) did
not locate and describe the sedimentary succession pre-cisely and the base of their section (level A) is missing in
the quarry called ‘‘Faluniere’’. This well-known quarry was
rehabilitated in 2006, thanks to new field work and a sec-tion, 13 m in thickness, can be described. From the base to
the top, it includes seventeen stratigraphic intervals (Fig. 3)
defined by lithological and paleontological criteria. In theparagraphs below, we propose an upgrade description of
the section, in relation to the sequential framework estab-
lished by Gely (1996) for the Lutetian deposits of the ParisBasin.
Facies (2012) 58:587–604 589
123

Parasequence A6
Intervals 1–6 (2.60 m thick)—More or less consolidatedlimestone, slightly glauconitic and containing echinoids
(e.g., Maretia grignonensis, Echinolampas calvimontensisand Gitolampas issyaensis). The intervals can be correlated
to the ‘‘Calcaire dur avec poches sableuses a Echinides’’ of
Abrard (1925) and to level B of Le Calvez and Le Renard(1980).
Intervals 7–11 (3.70 m thick)—Glauconitic calcareous
sands with quartz and shell accumulations. The shells aregenerally decalcified, but we recognized: Glycymerispulvinata, Meroena semisulcata, Cardium (Orthocardi-um) subporulosum, Cubitostrea plicata, Fustiaria circi-nata, Athleta (Volutospina) spinosus, Haustatorimbricatarius, and Galeoda enodis. They can be corre-
lated to the level of ‘‘calcaire endurci a Ampullospirahybrida’’ of Abrard (1925) and level C of Le Calvez and
Le Renard (1980).
Parasequence A7
Campanile giganteum level (1.5 m thick) [=interval 12]—Glauconitic calcareous sands containing Campanilegiganteum and a very rich and famous assemblage of well-
preserved organisms, including molluscs, bryozoans, andforaminifers. It corresponds to a rich, 0.40-m-thick, accu-
mulation, whose top of is less dense and displays abundant
bioturbation (Ophiomorpha type). The Campanile gigant-eum level is easily correlated to level D of Le Calvez and
Le Renard (1980).
Parasequence A8
Interval 13 (2.3 m thick)—Calcareous sands with Orbito-lites complanatus and miliolids. The base of the level
displays a rich shell accumulation deposited as fills ofpluricentimetric channels. The thickness of this shell
accumulation varies from 0.2 m to 0.4 m. At Grignon, it is
the second famous assemblage of macro-organisms. Themid-part of the level contains some Terebratula bisinuataand clusters of Chama in life position. The first Avicular-ium lithocardium of the section appear at the top of thisinterval and are associated with many Seraphs sopitus.
Interval 13 can be correlated to the ‘‘Couche a Orbitolitescomplanatus’’, ‘‘couche a Terebratula bisinuata’’ and‘‘couche a Lithocardium aviculare’’ (=Avicularium litho-cardium) of Abrard (1925; p. 134) and to the levels E–F of
Le Calvez and Le Renard (1980).
Parasequence A9
Interval 14 (1.25 m thick)—Limestone with Orbitolites,
sea grass (Cymodoceites), corals; and internal molds of
Seraphs sopitus and Avicularium lithocardium. This levelcorresponds to level G of Le Calvez and Le Renard (1980)
and to the base of Banc Vert of Abrard (1925).
Interval 15 (1.10 m thick)—Clayey limestone withdecalcified shells and corals. This interval is not described
by Abrard (1925) and Le Calvez and Le Renard (1980), but
could be related to the Banc Vert.Interval 16 (0.1 m thick)—Limestone with internal
molds of Seraphs sopitus, Avicularium lithocardium and
0° 5° E5° W
50° N
45° N
ParisBasin
AquitaineBasin
Bay ofBiscay
FRANCE
ENGLAND
SPAIN
Paris
a
English Channel
study area
Grignon
ChampagneRiver
Lake ofMorancez
Lake of Provins
Mainland
Dep
th
10 to 20 m
5 to 10 m
> 5 m
Lake
Cliff
Foreshore
Coastal drift
Etampes Gulf
Houdan
Pontoise
Meulun
Compiègne
Château-Thierry
NoyonLaon
Reims
La Chapelle-en-Vexin
Paris
bNBRAY ANTICLINE
0 50 km
Fig. 2 a Large-scale paleogeographic reconstruction for the Middle Lutetian and location of the study area. b Paleogeographic map of thecentral Paris Basin during the Middle Lutetian (parasequences A7 and A8) and location of the site of Grignon (modified after Gely 2008)
590 Facies (2012) 58:587–604
123

Saxolucina saxorum. Like level 15, this interval is notdescribed by Abrard (1925) and Le Calvez and Le Renard
(1980), but could be related to the Banc Vert.
Parasequence A10
Interval 17 (0.1 m thick)—Limestone with internal molds
of Saxolucina saxorum and Batillaria. It corresponds to the
‘‘couches a Cerithes’’ of Abrard (1925).
Paleoecological and paleoenvironmental contexts
Although the paleontological content of Grignon (Falu-
niere) is well known and illustrates its exceptional species
richness, few studies concern the paleoecology of the
fossils found in this outcrop (Chaix 1979; Le Calvez 1970;Guelorget and Perthuizot 1983; Andreasson and Schmitz
1996). Additionally, these studies did not focus on the
paleoenvironmental variations through the entire sectionbut refer to punctual samples. According to Gely (1996),
the paleontological assemblage of parasequence A6 indi-cates infralittoral marine environments, with normal
salinity. Abundant Glycymeris at the top of parasequence 6
(level 7–11) could be compared to the Glycymeris com-munity found in the Rupelian of the Paris Basin (Lozouet
1997), which corresponds to the infralittoral biocoenosis
‘‘Sables Fin Bien Calibres’’ defined in the MediterraneanSea (Perez and Picard 1964). In reference to this modern
biocoenosis, the water depth was probably between 20 and
30 m (Guernet et al. in press). The fossil assemblage of
0
1
2
3
4
5
6
7
8
9
10
11
12
131617
15
14
13
12
11
10
9
8
7
6
5
4
321
Thickness(m)
A8
A9
A10
A7
A6
Mid
dle
Lute
tian
C. plicata
S. multisulcata
V. imbricata
H. imbricatarius
Glauconitic sandswith quartz and shell accumulations
Glauconitic sandswith quartz
Calcareous sandswith Orbitolites complanatus,miliolids and shell accumulations levels
Limestoneswith Orbitolites, sea grass, corals and internalmolds of molluscs
Clayey limestone with decalcifiedshells and corals
Limestones with internal moldsof Saxolucina saxorum and Batillaria
Campanile giganteum level
Limestoneswith glauconiteand echinids
Para-
se
quen
ce
Inter
val
Fig. 3 Sedimentologicalsuccession and parasequences(from Gely 1966) of the outcropof the Faluniere of Grignon. Thestratigraphic positions of thethree groups of molluscscollected for this study appearas grey squares (Cubitostreaplicata), dark diamonds(Sigmesalia multisulcata), andgrey triangles (Venericardiaimbricata). The open squares,diamonds, and circles representthe fossils analyzed for theinfra-annual profiles(Cubitostrea plicata, Sigmesaliamultisulcata, and Haustatorimbricatarius, respectively)
Facies (2012) 58:587–604 591
123

parasequence A7, does not differ strongly from the previ-
ous parasequence (Le Calvez and Le Renard 1980), butcontains Campanile giganteum. A significant facies change
is observed within parasequence A8, with calcareous bio-
clastic sand containing Orbitolites and miliolids and ahighly diverse fauna of molluscs, which indicates a prob-
able diminution of the paleodepth to less than 20 m
(Fig. 2). In parasequence A9, numerous algae (Genot2009) and phytophagous gastropods indicate sea grass
environments, while stenohaline forms such as corals,echinids, the bivalve Chama, and the brachiopod Tere-bratula suggest marine conditions and high water clarity.
Intervals 14–16 with Seraphs sopitus, Lithocardium avic-ularium, remains of Cymodoceites and branched corals, do
not indicate significant salinity changes. On the contrary, a
major salinity change occurs in interval 17 (parasequenceA10), with the appearance of the Batillaria assemblage
found in lagoonal environments in different localities of the
Lutetian from the Paris Basin (Abrard 1925; Gely 1996).To conclude, our analysis of the section shows that from
interval 1 to interval 16, the faunal associations observed
indicate marine conditions with deceasing water depth andnormal and stable salinity (near 35%), except at the top of
the section (interval 17), where the salinity became
unstable. Because of this change, we did not sampleinterval 17 (Fig. 3).
Materials and methods
Studied taxa
The three most abundant fossil species present throughout
the section at Grignon were chosen for the study of thepaleoenvironmental changes: Cubitostrea plicata (n = 39),
Venericardia imbricata (n = 13), and Sigmesalia multi-sulcata (n = 21) (Figs. 3, 4). Three additional shells ofV. imbricata, one C. plicata and one Haustator imbrica-tarius were collected to perform high-resolution stable
isotope analysis (Fig. 3).Cubitostrea plicata is a small oyster (\4 cm). Like
modern Ostreidae, C. plicata is considered to be an epi-
faunal filter-feeder living in shallow-water environments(Stenzel 1971). The stable isotope signature of oyster shells
is commonly used for paleoclimatic reconstructions (e.g.,
Kirby et al. 1998; Surge et al. 2003; Lartaud et al. 2010a;Titschack et al. 2010; Fan et al. 2011) and can thus yield
reliable values of past seasonal seawater temperatures.
C. plicata is the most abundant mollusc species found atGrignon and was sampled from the bottom (interval 3) to
the top (interval 15) of the section (Fig. 2).
The bivalve Venericardia imbricata has no modernequivalent, but morphological features indicate that this
species is closely related to the Recent Cardita ventricosa(Yonge 1969; Watters 1993; Andreasson and Schmitz1996). C. ventricosa is an endobiont suspension-feeder
living in shallow waters. Stable isotope analyses have
never been performed for Recent Cardita, but fossil Ven-ericardia specimens were used to reconstruct seawater
paleotemperatures (Seward 1978; Stevens and Vella 1981;
Purton and Brasier 1999; Ivany et al. 2004; Haveles andIvany 2010) and particularly at Grignon, where Andreasson
and Schmitz (1996) showed that V. imbricata mineralizedtheir shells mostly during the warmer months of the year,
with no or very slow shell growth during the winter period.
Thus, the d18O of these shells enables only the maximumseawater paleotemperatures of the year to be estimated.
V. imbricata shells have been collected from intervals 12 to
15 and one isolated specimen in interval 5 (Fig. 3).Haustator imbricatarius is a large gastropod (*10 cm
in height) that belongs to the family Turritellidae (Turri-
tellinae) and is closely related to the Recent genus Turri-tella. Ecological observations on this genus indicate that it
is a shallow burrowing ciliary suspension-feeder (Graham
1938; Yonge 1946; Davitashvili and Merklin 1968; Allmon1988, 2011; Waite and Strasser 2011). However, a detailed
study of the feeding behavior of the Australian turritelline
Gazameda gunnii by Carter (1980) found that this speciesis not only a ciliary suspension-feeder in the style of
T. communis, but also a deposit feeder. Analyses of gut and
fecal contents show that G. gunnii consumes a range ofmaterial including diatoms, sponge spicules, bryozoan and
crustacean skeletal fragments, and coarse siliceous sedi-
ment grains. Thus, turritellines should be regarded as notstrict, ciliary suspension feeder. Turritella shells are com-
monly used for paleoclimatic reconstructions (Andreasson
and Schmitz 1996, 2000; Tripati et al. 2009). The shell wassampled in interval 13, which is the only interval where
they are represented.
Sigmesalia multisulcata is a small gastropod (*3 cm inheight). The extinct genus Sigmesalia is closely related to
the living genus Mesalia (Squires and Saul 2007). There
are no ecological data on Mesalia but the genus belongs tothe family Turritellidae, such as Turritella and Haustator.
As with other turritellids, Sigmesalia is supposed to have
been a shallow-infaunal not strict ciliary suspension feeder(Davitashvili and Merklin 1968; Allmon 1988; Waite and
Strasser 2011), or possibly deposit feeder like some mod-
ern species of turritellids (Carick 1980). No isotopic workhas been performed on Sigmesalia species. However,
numerous studies have shown that gastropods precipitate
their shell in isotopic equilibrium with seawater, andthus can be used as an accurate archive of past tempera-
tures (Grossman and Ku 1986; Wefer and Berger 1991).
S. multisuculata shells were sampled from interval 11 tothe base of interval 14 (Fig. 3).
592 Facies (2012) 58:587–604
123

Preservation of the shells
The use of biogenic carbonates to reconstruct past envi-ronments and temperatures requires the estimation of the
influence of diagenesis on the shells to ensure that a pri-
mary isotopic signal is preserved. The preservation of thesamples has been tested using different methods according
to each type of shell structures.
In oyster shells, the hinge contains an ontogenetic recordof both the oysters’ growth and the environmental condi-
tions experienced throughout their life (Richardson et al.
1993; Kirby et al. 1998). The upper part of the hinge iscomposed of a single mineralogy (low magnesian calcite)
and a uniform microstructure (foliated calcite) (Carter
1980). This part is preferentially used for paleoclimaticstudies inferred from isotopic composition since it is
assumed to be more resistant to diagenetic alteration
(Lartaud et al. 2006). Moreover, the hinge area is com-monly the sole remaining part of the fragile shells of fossil
C. plicata found at Grignon. The primary foliated calcite of
the hinge section has been checked by cathodolumines-
cence. The preservation of shell growth patterns, i.e., reg-ular seasonal cycles, naturally present in living organisms
attests to the good preservation of the samples and the
preservation of a primary isotopic signal (Fig. 4; Lartaudet al. 2006, 2010b; Ivany and Runnegar 2010).
Venericardia imbricata, S. multisulcata and H. imbri-catarius have an aragonite primary mineralogy. X-raydiffraction was used to determine the mineralogy of fossil
shells. The preservation of the aragonitic mineralogy isgenerally considered to be a reliable indicator of a reduced
diagenetic overprint and of the preservation of the primary
isotopic signal, because it is an unstable polymorph ofcalcium carbonate in comparison to calcite (Al-Aasm and
Veizer 1986). Additionally, the preservation of residual
color patterns in the external layer of the shell precludesany aragonite to aragonite diagenetic transformation. Fol-
lowing the method of Merle et al. (2008), each shell is
observed under UV-light (Fig. 3), after NaHClO treatmentfor 24 h. The presence of color patterns, which exist
0.5 1 1.5 2 2.50
! OCL
! C
18
13
40
60
80
100
120
1
0
-1
-2
GROWTH
Lenght (mm)
a
b c
d
f g
h
e
1 mm
a
Fig. 4 Fossils from Grignon analyzed in this study. a Haustatorimbricatarius viewed under natural light; b, c Sigmesalia multisulcataviewed under natural light (b) and under UV light (c); d, e Venericar-dia imbricata under natural light (d) and under UV light (e); f, g view
of the external part of Cubitostrea plicata shell under natural light(f) and internal part of the shell (g) h cathodoluminescence view andcorresponding profile and d18O and d13C pattern of the micro-sampledhinge of Cubitostrea plicata. Scale bar 1 cm
Facies (2012) 58:587–604 593
123
naturally in the external layer of the mollusc shells, indi-
cates that fossils did not undergo strong diagenesis (Cazeet al. 2010; Caze et al. 2011).
Sampling and isotopic analysis
Before sampling, the shells are ultrasonically cleaned to
remove any residual sediment. Following the cleaning pro-tocol of Lartaud et al. (2010a), organic mater is removed by a
6 % hydrogen peroxide (H2O2) bath for 6 h and any carbon-ate-based superficial contamination is dissolved using 0.15 %
nitric acid (HNO3) for 20 min. Shells are rinsed in deminer-
alized water before sampling with a 0.5-mm drill bit.The sampling area for C. plicata shells corresponds to
the whole hinge (excluding altered or secondarily filled
areas) which is assumed to reflect both seasonal contrastsand several years of growth of the shell, leading to an
estimation of seawater temperatures close to, or just above,
the annual average throughout the life of the specimen
(Goodwin et al. 2003; Lartaud et al. 2010a). The whole
hinge of each oyster is drilled to reconstruct mean annualtemperatures and one measurement corresponds to one
oyster. The observation of the hinge under cathodolumi-
nescence allows avoiding altered areas or sediment-filledareas to be sampled (Fig. 4). One fossil was micro-sampled
and five samples were made on its hinge to obtain infra-
annual data (Figs. 4h, 6b).The samples for V. imbricata shells are performed on
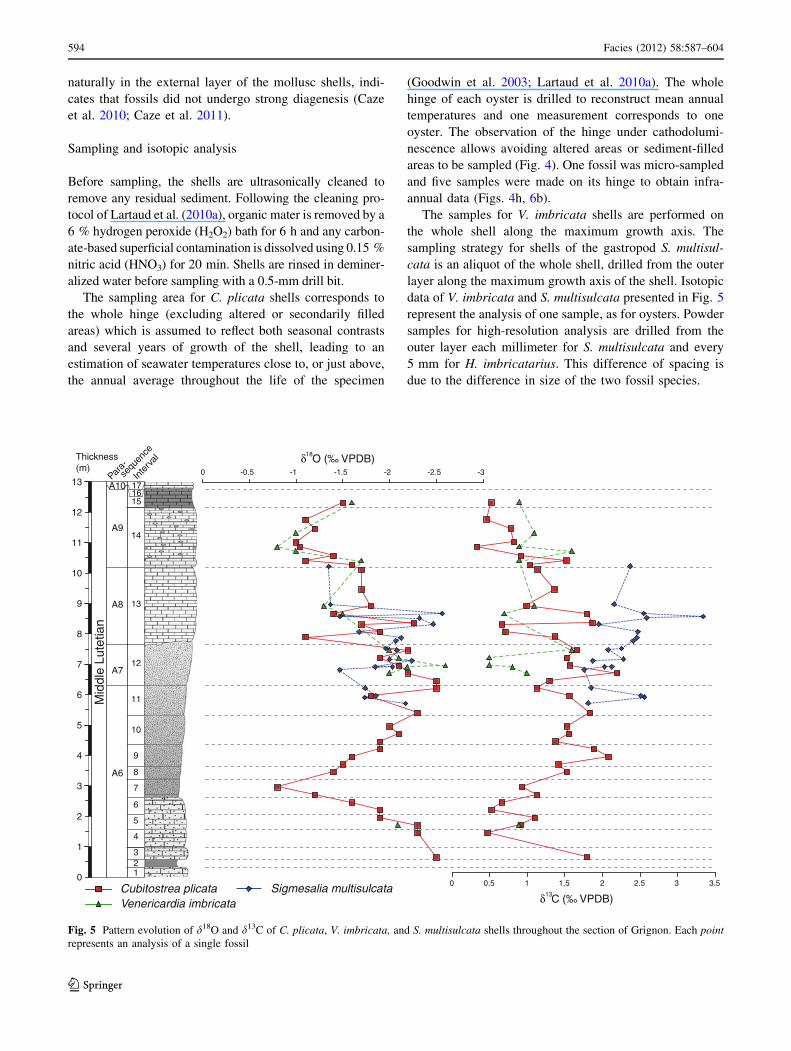
the whole shell along the maximum growth axis. Thesampling strategy for shells of the gastropod S. multisul-cata is an aliquot of the whole shell, drilled from the outer
layer along the maximum growth axis of the shell. Isotopicdata of V. imbricata and S. multisulcata presented in Fig. 5
represent the analysis of one sample, as for oysters. Powder
samples for high-resolution analysis are drilled from theouter layer each millimeter for S. multisulcata and every
5 mm for H. imbricatarius. This difference of spacing is
due to the difference in size of the two fossil species.
0
1
2
3
4
5
6
7
8
9
10
11
12
13
Thickness(m)
Para-
se
quen
ce
Inter
val
A8
A9
A10
A7
A6
Mid
dle
Lute
tian
Cubitostrea plicata Sigmesalia multisulcataVenericardia imbricata
14
13
12
11
10
9
8
7
6
5
4
321
1617
15
-3-2.5-2-1.5-1-0.50
! O (‰ VPDB)18
0 0.5 1 1.5 2 2.5 3 3.5
! C (‰ VPDB)13
Fig. 5 Pattern evolution of d18O and d13C of C. plicata, V. imbricata, and S. multisulcata shells throughout the section of Grignon. Each pointrepresents an analysis of a single fossil
594 Facies (2012) 58:587–604
123

The carbonate powders are reacted with 100 %
orthophosphoric acid at 50 "C, and oxygen isotope ratiosare measured using a VG MM903 mass spectrometer.
Isotopic data are reported in conventional delta (d)
notation relative to the Vienna Pee Dee Belemnite (%VPDB). The standard used for the analyses is an internal
standard calibrated on the NBS-19. Standard deviation
for both d18O and d13C is ±0.1 %. All results arereported in Appendix 1 in Electronic supplementary
material.
Paleotemperature calculation
Paleotemperatures can be reconstructed using the equation
of Anderson and Arthur (1983) for calcite shells (i.e.,
C. plicata):
T "C# $ % 16& 4:14 #d18Oc & d18Ow$ ' 0:13 #d18Oc
& d18Ow$2
and from the equation of Grossman and Ku (1986) for
aragonite shells (i.e., V. imbricata, S. multisulcata andH. imbricatarius):
T "C# $ % 20:6& 4:34 #d18Oa & d18Ow$
Estimating paleotemperatures requires control of thed18O of seawater (d18Ow), which is a function of salinity.
The reconstruction of past salinities throughout the
section at Grignon, according to the faunal assemblagesdescribed and to previous studies (Guelorget and
Perthuizot 1983; Le Calvez 1970; Andreasson and
Schmitz 1996; Guernet et al. in press), show normalmarine waters for the levels sampled for isotopic analyses,
leading to an estimated salinity that remained stable and
close to 35%. No correction of the latitudinal effect istaken into account because the latitudinal gradient was
reduced during Middle Eocene times (Greenwood and
Wing 1995; Andreasson and Schmitz 1996; Bijl et al.2009). The d18Ow is also a function of the glacial effect,
which corresponds to the amount of ice stored on the
continents. For full greenhouse periods, Shackleton andKennett (1975) estimated a global mean d18Ow of -1 %(SMOW). The first permanent glaciations of the Cenozoic
occurred at the Eocene—Oligocene boundary (*34 Ma;Zachos et al. 1996), but more and more studies have
shown that significant glaciations could have occurred
also before this period (Lear et al. 2000; Tripati et al.2005, 2008; Payros et al. 2009; Huyghe et al. 2012),
although this remains controversial (Burgess et al. 2008).
For this study, we consider d18Ow of -0.9 % in referenceto Lear et al. (2000), who estimated the global d18Ow
according to the comparison of the Mg/Ca ratio and thed18O of benthic foraminifers in the Pacific.
Results
Evolution of d18O and d13C of mollusc shells through
the Middle Lutetian
The isotopic values of the three species of fossils analyzed
throughout the section at Grignon are reported in Fig. 5. The
d18O profiles along the sedimentary succession display rel-atively large variations. The d18O values of C. plicatashells, which are present throughout the section, range
between -2.7 and ?0.5 %. Low values are observed at thebottom of the section (-2.5 %), followed by an increase
until interval 7 (-0.8 %), followed by a decrease into the
base of interval 12 (-2.5 %). From interval 12 to the middleof interval 13, the oxygen isotopic composition is more
variable (-1.1 to -2.5 %), but exhibits an important
increase until the middle of interval 14, where high values arerecorded (-1 %). The upper part of the section shows a
slight decrease of d18O until interval 15 (-1.5 %). The trend
followed by V. imbricata and S. multisulcata shells, whichoccur solely in the upper part of the section, is the same as that
of the oysters, ranging between -2.6 and -0.7 % for V. im-bricata and between -2.6 and -1.3 % for S. multisulcata.
The d13C of C. plicata shells exhibits great fluctuations,
with values ranging between ?0.3 and ?2.1 %. V. im-bricata shells record the same range of variation (?0.5 to?1. 6%). With very few exceptions, d13C values of
S. multisulcata shells are higher than those of the two otherspecies, ranging between ?1.1 and ?3.4 %. The evolution
of the d13C of oysters displays an initial decrease at the
base of the section, followed by a long increase from about?0.5 % in interval 4 to ?2.1 % in interval 5. In the upper
part of the section, i.e., from interval 11 to interval 15, the
d13C of C. plicata shells decreases slightly and the d13C ofV. imbricata shells remains relatively stable, whereas
S. multisulcata shells display a slight increase in d13C.
Comparison of d18O and d13C between species
Isotopic results presented in Fig. 5 highlight some shiftsbetween the different species of fossils. Stable isotope ratios of
the three taxa (C. plicata, V. imbricata and S. multisulcata)sampled along the section are compared to identify the causesof the shifts between them (Fig. 6), in the intervals where all of
them are present (Fig. 3). Because C. plicata has a calcitic
shell, in contrast to V. imbricata and S. multisulcata, which arearagonitic, we have corrected for the mineralogical effect
between calcite and aragonite. Synthetic aragonite exhibits
higher isotopic values than calcite, i.e., ?0.6 % for d18O(Tarutani et al. 1969) and ?1.7 % for d13C (Romanek et al.
1992). After this correction, C. plicata shells present signifi-
cantly higher values of d18O (between -1.4 and -2.5 %)and d13C (between ?0.6 and ?2.2 %) than S. multisulcata
Facies (2012) 58:587–604 595
123
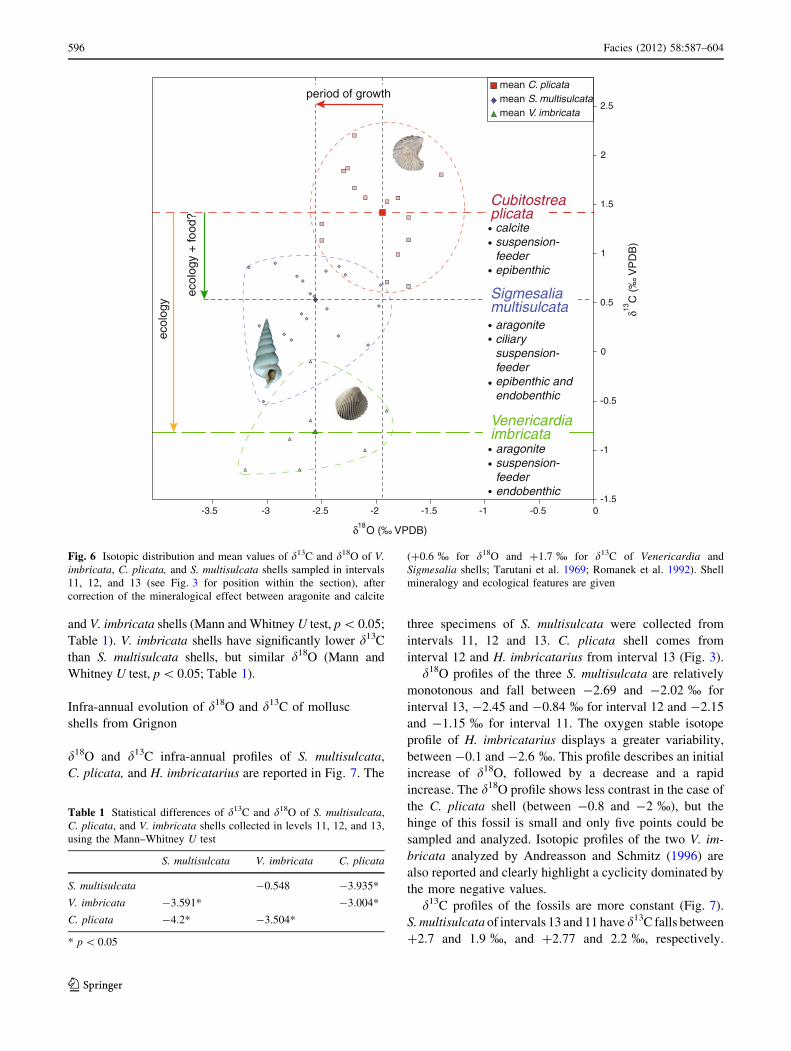
and V. imbricata shells (Mann and Whitney U test, p \ 0.05;
Table 1). V. imbricata shells have significantly lower d13C
than S. multisulcata shells, but similar d18O (Mann andWhitney U test, p \ 0.05; Table 1).
Infra-annual evolution of d18O and d13C of molluscshells from Grignon
d18O and d13C infra-annual profiles of S. multisulcata,C. plicata, and H. imbricatarius are reported in Fig. 7. The
three specimens of S. multisulcata were collected from
intervals 11, 12 and 13. C. plicata shell comes from
interval 12 and H. imbricatarius from interval 13 (Fig. 3).d18O profiles of the three S. multisulcata are relatively
monotonous and fall between -2.69 and -2.02 % for
interval 13, -2.45 and -0.84 % for interval 12 and -2.15and -1.15 % for interval 11. The oxygen stable isotope
profile of H. imbricatarius displays a greater variability,
between -0.1 and -2.6 %. This profile describes an initialincrease of d18O, followed by a decrease and a rapid
increase. The d18O profile shows less contrast in the case of
the C. plicata shell (between -0.8 and -2 %), but thehinge of this fossil is small and only five points could be
sampled and analyzed. Isotopic profiles of the two V. im-bricata analyzed by Andreasson and Schmitz (1996) arealso reported and clearly highlight a cyclicity dominated by
the more negative values.
d13C profiles of the fossils are more constant (Fig. 7).S. multisulcata of intervals 13 and 11 have d13C falls between
?2.7 and 1.9 %, and ?2.77 and 2.2 %, respectively.
! O (‰ VPDB)18
! C
(‰
VP
DB
)13
ecol
ogy
+ fo
od?
period of growth
Sigmesaliamultisulcata
Cubitostreaplicata
Venericardiaimbricata
-1.5
-1
-0.5
0
0.5
1
1.5
2
2.5
-3.5 -3 -2.5 -2 -1.5 -1 -0.5 0
mean C. plicatamean S. multisulcatamean V. imbricata
ecol
ogy
aragoniteciliary suspension-feederepibenthic and endobenthic
aragonitesuspension-feederendobenthic
calcitesuspension-feederepibenthic
Fig. 6 Isotopic distribution and mean values of d13C and d18O of V.imbricata, C. plicata, and S. multisulcata shells sampled in intervals11, 12, and 13 (see Fig. 3 for position within the section), aftercorrection of the mineralogical effect between aragonite and calcite
(?0.6 % for d18O and ?1.7 % for d13C of Venericardia andSigmesalia shells; Tarutani et al. 1969; Romanek et al. 1992). Shellmineralogy and ecological features are given
Table 1 Statistical differences of d13C and d18O of S. multisulcata,C. plicata, and V. imbricata shells collected in levels 11, 12, and 13,using the Mann–Whitney U test
S. multisulcata V. imbricata C. plicata
S. multisulcata -0.548 -3.935*
V. imbricata -3.591* -3.004*
C. plicata -4.2* -3.504*
* p \ 0.05
596 Facies (2012) 58:587–604
123

The profile of the shells of interval 12 ranges between ?1.64
and ?2.83 % and slightly differs with two rapid decreases ofthe d13C values in a long monotonous trend. H. imbricatariusexhibits d13C values that increase during its life, withextreme values between ?1.5 and ?3.2 %. Concerning
C. plicata, its d13C values are more negative than those of the
two other species and range from -0.7 to ?0.4 %.
Inter-taxon comparison
Sclerochronology and temperature
Considering that few shells have undergone transport
given the taphonomic observations and shell preservation,
the specimens analyzed built their shells from the samewater body for each stratigraphic level. As no environ-
mental origin can explain the differences observed on the
d18O corrected for mineralogical effects (Fig. 6) betweenthe three taxa and within the same collection interval,
influence of a vital effect and/or of the period of miner-
alization can be suggested. Although it is often difficult toidentify a vital effect for fossil shells, Wefer and Berger
(1991) have shown that many molluscs mineralise their
shell in equilibrium with water. Thus, d18O changesbetween species rather reflect seasonal biomineralization
differences.
Specimens of the species H. imbricatarius build theirshell throughout the year and thus record seasonal varia-
tions of temperature (Andreasson and Schmitz 1996). The
cyclicity recorded in their isotopic profile reflects seasonalvariations with maximum d18O values corresponding to
winter and minimum values to summer temperatures.
d18O values of S. multisulcata are relatively stable,
except for the shell from interval 12 (Fig. 7). The seasonalrange of the S. multisulcata shell from the same level as
H. imbricatarius previously described (interval 13) islower, which suggests that this organism seems to construct
its shell not continuously during the year and/or minerali-
zation is restricted to less than 1 year. Mean d18O valuesfrom the three S. multisulcata shells are closely similar to
the lowest values measured for H. imbricatarius (around
-2.5 %), corresponding to summer temperatures. As aconsequence, S. multisulcata appears to record only
the warmer temperatures of the year, like V. imbricata(Andreasson and Schmitz 1996). However, contrary toV. imbricata, the profiles of S. multisulcata exhibit reduced
variation (Fig. 7). As no apparent cessation of shell growth
was recorded either in the isotopic signal or on the externalpart of the shell, in contrast to V. imbricata (Fig. 7d;
Andreasson and Schmitz 1996), S. multisulcata had a very
short period of growth (\1 year), which seems to haveoccurred from spring to summer.
Oysters are known to build their shell throughout the
year, with slight decrease in shell growth during the winterperiod (Kirby et al. 1998; Kirby 2000; Surge et al. 2001;
Lartaud et al. 2010b; Titschack et al. 2010). Consequently,
mean d18O values of oysters will provide mean or weakover-estimations of the mean annual seawater tempera-
tures. However, the intra-annual range might reflect sea-
sonal changes in temperature (Lartaud et al. 2010a;Titschack et al. 2010). The lower range observed in
C. plicata shells in comparison to the H. imbricatariusshell (Fig. 7) could be due to lower temporal resolution ofsampling in the hinge of such small oysters, which induce a
bias in the interpretation of the isotopic signal leading to
-3
-2.5
-2
-1.5
-1
0
-0.5
0 10 20 30 40 50
level 13
0 10 20 30 40 50 60
level 12
0 10 20 30 40 50 60
level 11
Distance from the apex (mm) Distance from the apex (mm) Distance from the apex (mm)
growth
a b
dc
! O
(‰
VP
DB
)18
4.5
4
3.5
3
2.5
1.5
2
! C (‰
VP
DB
)13
-3
-2.5
-2
-1.5
-1
0
-0.5! O
(‰
VP
DB
)18
level 12
Umbo length (mm)0 1 2 3
growth
1
0.5
0
-0.5
-1
-2
-1.5
! C (‰
VP
DB
)13
level 13
! O
(‰
VP
DB
)18
-3
-2.5
-2
-1.5
-1
-0.5
0
0.50 50 100 150 200 250
Distance from the apex (mm)
growth
! C (‰
VP
DB
)13
3.5
3
2.5
2
1.5
1
0.5
0
3.5
3
2.5
2
1.5
1
0.5
0
! O18
! C13
! O18
! C13
! O18
! C13
-3
-2.5
-2
-1.5
-1
-0.5
0
0.520 30 40 50
Umbo length (mm)
! O (‰
VP
DB
)18
! O18
! C13
Fig. 7 High-resolution oxygen stable isotope profiles. a Three shells of S. multisulcata; b one shell of C. plicata; c one shell of H. imbricatarius;d V. imbricata (from Andreasson and Schmitz 1996). The precise position within the section is given in Fig. 3
Facies (2012) 58:587–604 597
123

time averaging of the d18O values (Goodwin et al. 2003;
Lartaud et al. 2010a).As a consequence, it seems that H. imbricatarius is the
only fossil analyzed in this study able to document the
seasonal range of temperature, whereas d18O values ofC. plicata reflect mean annual seawater temperatures and
d18O values of V. imbricata and S. multisulcata correspond
to summer temperatures. However, it is important to con-sider that molluscs are able to change the period of min-
eralization of their shell according to variations inenvironmental parameters, such as the temperature (Jones
and Quitmyer 1996). Nevertheless, the shift in the d18O
values of C. plicata shells compared to the ones of V.imbricata and S. multisulcata shells appears to be relatively
constant, which suggests that the period of growth of the
three taxa is relatively constant.
Habitat and feeding influence
The carbon stable isotope values differ significantly
between the three species (Fig. 6; Table 1). Shell d13C
derives from both dissolved inorganic carbon (DIC) andorganic carbon sources (food), the latter being incorporated
into the shell via metabolic pathways (e.g., Killingley and
Berger 1979; McConnaughey 1989; Wefer and Berger1991; McConnaughey et al. 1997). Although some meta-
bolically induced shell isotopic changes—so-called vital
effects—can be strong according to some taxa (Wefer andBerger 1991) or to breeding conditions (Lartaud et al.
2010c), they remain relatively low for marine molluscs
(i.e., the metabolic carbon is typically less than 10 % in theshells of marine molluscs, see a review in McConnaughey
and Gillikin 2008). The d13C of marine mollusc shells is
thus closely related to changes in the environmental DICand the source of carbon derived from the food (see review
in Lartaud et al. 2010c).
Infra-annual isotopic profiles of S. multisulcata, C. pli-cata, and H. imbricatarius shells analyzed in this study and
of V. imbricata shells from Andreasson and Schmitz (1996)
exhibit very few variations, meaning that the DIC at Gri-gnon was relatively constant throughout the year. The only
exception concerns the S. multisulcata shell from interval
12 for which two rapid increases in d13C are observed. Thisevolution might be related to phytoplanktonic blooms
during the summer period. An increase in primary pro-
ductivity should increase influx of organic matter to thebenthos, which will increase benthic respiration. As a
consequence, benthic respiration will return 12C to the DIC,
thereby decreasing d13CDIC values (Purton and Brasier1997). However, large differences are observed in the
d13C values of species taken from the same level
(Fig. 6), including species with the same period of growth(S. multisulcata and V. imbricata), which cannot reflect
environmental DIC changes. Latal et al. (2006) have shown
that the d13C of bivalve and gastropod shells, includingturritellids, is primarily derived from the original habitat
and nutrition (suspension-feeding, deposit-feeding or car-
nivorous). Based on their living equivalents, C. plicata andV. imbricata are both considered to have been suspension-
feeders (Yonge 1969; Stenzel 1971; Watters 1993; An-
dreasson and Schmitz 1996), whereas S. multisulcata isprobably a ciliary suspension-feeder or deposit-feeder,
similar to turritellids (Davitashvili and Merklin 1968; Ca-rick, 1980; Allmon, 2011). Consumption of rich 12C
organic matter on the seafloor explains the more 13C
depleted d13C of S. multisulcata shells.Venericardia imbricata, which is a burrowing organism,
exhibits low d13C values that cannot be related to the feeding
pathways. Remineralization processes of organic matter couldbe important in the sediment, leading to a decrease of the pore
water d13CDIC, which leads to low d13C values in shells of
burrowing organisms (McConnaughey and Gillikin 2008;Lartaud et al. 2010d), whereas at the seafloor the contribution
of seawater-derived DIC prevails.
However, we cannot rule out the influence of otherinternal factors influencing the d13C. The proportion of the
contribution of metabolic CO2, even if not prevailing in
comparison with other sources of carbon, can change withontogeny and environment within the same taxon
(McConnaughey and Gillikin 2008). It could explain, for
example, why the d13C of S. multisulcata increases in theupper part of the section, whereas the one of C. plicata and
V. imbricata decreases.
Paleotemperature reconstructions
Conversion of d18O values into seawater temperatures
using paleotemperature equations is shown in Fig. 8 and
mean temperatures for each taxon in each level is reportedin Table 2. Mean annual seawater temperature (MAT)
variations are inferred from C. plicata shells and temper-
atures of the warmer months (TWM) are derived from thed18O of S. multisulcata and V. imbricata shells and are only
available for the upper part of the section.
An initial decrease in MAT is recorded from interval 1to interval 7, from 23 to 16 "C, followed by warming until
the base of interval 12 (23 "C). Then, a long decrease in
temperatures is observed up to the base of interval 14(*16 "C) and the interval finishes with a warming in
interval 15 (19 "C). Maximum TWM are recorded in
intervals 12 and 13 (30 "C), with a relative coolingin interval 14 (21 "C) before a final warming in interval 15
(25 "C). The offset between the MAT calculated from C.plicata shells and the TWM inferred from V. imbricata andS. multisulcata shells is constant.
598 Facies (2012) 58:587–604
123

Seasonal variations of paleotemperatures deduced from
the isotopic profile of H. imbricatarius shell at the base of
interval 13 range between 18 and 30 "C with MAT of23 "C. These temperatures are in good agreement with both
MAT and TWM calculated from the other species at this
level (Fig. 8). Additionally, these temperatures are close tothe estimations of Andreasson and Schmitz (1996, 2000)
based on the stable isotope analysis of H. imbricatariusshells at Grignon and in Southern England (28 "C insummer and 14 "C in winter).
Paleoclimatic and paleoecological implications
The long trend showing a decrease in MAT through theMiddle Lutetian at Grignon is consistent with the long-term
cooling well documented in the oceanic realm during the
same period (e.g., Zachos et al. 2001). However, greater
temperature variability is observed in our study, i.e., a
cooling and warming event occurs between interval 1 andinterval 11, characterized by D MAT of 8 "C. The strong
cooling observed in interval 7 could correspond to the
variation of 1–2 "C recorded in the d18O of deep-seabenthic foraminifers near 44 Ma (Zachos et al. 2008).
The reliability of the observed variation of the temper-
atures from the base to interval 7 is, however, questionableas it represents only sea surface temperature variations.
Indeed, the Lutetian is known to be a relatively stable
period concerning the climate (Zachos et al. 2008). It isthus conceivable to consider that the observed shift rather
reflects variation of the d18Ow. Nevertheless, the fossil
associations do not suggest any significant salinity change.If this were the case, it would be necessary to consider a
salinity of 40 % in interval 7, i.e., distinct hypersaline
0
1
2
3
4
5
6
7
8
9
10
11
12
13
Thickness(m)
A8
A9
A10
A7
A6
Mid
dle
Lute
tian
14
13
12
11
10
9
8
7
6
5
4
321
Para-
se
quen
ce
Inter
val
1617
15
WATER
DEPTH
VARIATION
deepening
shallowing
Cubitostrea plicataSigmesalia multisulcataVenericardia imbricataHaustator imbricatarius
Temperatures (°C)
Mean growthtemperatures
Temperatures ofthe warmer months
14 16 18 20 22 24 26 28 30 32
Fig. 8 Changes of the calculated mean annual temperatures (from C. plicata d18O), the temperature of the warmer months of the year (from S.multisulcata and V. imbricata d18O), and the seasonal temperature variations (from H. imbricatarius d18O) during the Middle Lutetian at Grignon
Facies (2012) 58:587–604 599
123

conditions, to obtain a temperature higher than 20 "C, i.e.,
consistent with the rest of the section. In the lower part of
the section, fossil associations and the transgressive trenddisagree with such conditions. On the contrary, the
observed tendency could rather reflect an increase of the
water depth. The lower part of the sedimentary successionis the interval for which deeper environments have been
reconstructed (*30 m; Gely 2008; Guernet et al. in press)and it is related to a major regional transgression in the
Paris Basin (Gely 1996), as attested by the rich content in
glauconite (Fig. 3). Variations of global d18O could also beconceivable considering that small ephemeral ice-sheets
could have existed during the Lutetian (Lear et al. 2000;
Pekar et al. 2005; Payros et al. 2009). However, even if thiswere the case, the amplitude of the observed shift is too
great to result only from variation of ice volume.
Concerning the cooling of the upper part of the section,considerations of salinity changes have to be discussed
(Andreasson and Schmitz 1996, 1997; Klein et al. 1997;
Klein and Fricke 1997). We have considered a meansalinity of 35 % for this interval according to fossil asso-
ciations, but it is possible that the reduction of water depth
combined with relatively warm conditions could haveinduced ephemeral variation of the salinity by increasing
evaporation, in particular at the base of level 14 (Fig. 8).
However, the observed decrease of the temperatures isprogressive and such a progressive salinity-related trend
appears unrealistic. Moreover, seasonal variations of the
salinity are excluded because all taxa, even with differentperiods of mineralization, follow the same trend from
interval 11 to interval 15, even if the variability of the
values recorded by the Sigmesalia is higher than for theoysters and the Venericardia. Nevertheless, the Lutetian is
a period known to have experienced global long-term
cooling (e.g., Zachos et al. 2008), which is consistent withthe tendency recorded in the isotopic composition of the
fossils analyzed at Grignon.
This study shows that despite a context of global relativecooling, the Lutetian remained a warm period in the Paris
Basin, with MAT * 20 "C (Table 2), maximum summertemperatures close to 30 "C, and a mean annual range of
temperatures (MART) of 12 "C. Given these climatic
features, the southern part of the modern MediterraneanSea, which is characterized by subtropical conditions with
MAT * 22 "C, warm summer temperatures (*30 "C),
and MART of about 10 "C, could be considered as amodern analogue of the Middle Lutetian climate in the
Paris Basin. Other areas have some similar characteristics
to the Lutetian Paris Basin like the Red Sea. However,although molluscan diversities of the modern Red Sea are
probably more comparable to the Lutetian of the Paris
Basin (Zuschin and Oliver 2003; Zuschin et al. 2009;Janssen et al. 2011) than those of the southern Mediterra-
nean, the Red Sea cannot be considered as a full modern
climatic analogue of the Paris Basin during the Lutetian.Indeed, mean annual temperatures are comparable, but the
seasonal range of temperature is lower in the modern Red-
Sea (6–8 "C) than in the Paris Basin (12–14 "C). Thehypothesis considering the southern Mediterranean Sea as a
modern climatic analogue of the Lutetian of the Paris Basin
is also supported by the assemblages of ostracod generafound at Grignon. They are similar to the ones found
nowadays in the southern Mediterranean Sea, implying
MAT comprised between 20 and 25 "C and seasonal cli-matic differences of about 10 "C (Guernet et al. in press).
In addition, the Lutetian paleofloristic assemblages indicate
subtropical conditions with subarid seasons close to thosefound in southern Asia today (de Franceschi 2009). In
contrast to Paleocene and Early Eocene periods, the Paris
Basin was thus not characterized by a humid tropical cli-mate during the Middle Lutetian (Pomerol 1973; Thiry
1989).
The paleoclimatic results presented here are of majorinterest regarding the establishment of the ‘‘hotspot’’ of
biodiversity in the Paris Basin during the Lutetian. This
basin was characterized by warm summers and relativelymild winters. It would seem therefore that the decrease in
temperatures since the EECO was not sufficient to exert a
negative effect on the biodiversity of molluscs as was thecase at the Eocene–Oligocene boundary (Prothero 1994;
Ivany et al. 2003). This lack of negative effect on the
paleobiodiversity is also suggested in the Eastern Atlanticby the migration of molluscs from the southwestern
Table 2 Mean temperatures of C. plicata, V. imbricata and S. mul-tisulcata for each stratigraphic interval in which they were analyzed
Stratigraphiclevel
Mean temperatures ("C)
Cubitostreaplicata
Venericardiaimbricata
Sigmesaliamultisulcata
15 20 25
14 17 23
13 19 24 27
12 22 28 27
11 22 26
10 21
9 19
8 19
7 16
6 20
5 21
4 22
3 23
2
1
600 Facies (2012) 58:587–604
123

Atlantic coast of France to the Paris Basin during the
Lutetian. For example, at Gan, in the southern AquitaineBasin (Fig. 2), the Late Ypresian assemblage (biozone NP
14a) shares 32 % of species (118 species) with the Lutetian
of the Paris Basin (Merle 1986), implying that these speciesmigrated northward during the Lutetian and that the
paleotemperatures were sufficiently high to allow this
biogeographic dispersion. A colonization of the Paris Basinby southern species suggests even that this basin was
probably not the richest one during the Lutetian. Thepaleogeography of the western Tethys was formed by many
island arcs and could represent the centre of a large hot-
spot, as it is the case with the Recent Indo-Pacific biota(Merle 2008). Considering this paleogeographic context,
the Lutetian of northern Italy is placed within the Tethysian
island arcs, and its rich mollusc fauna (de Gregorio 1880;Quaggiotto and Mellini 2008) represents a good point of
comparison with the Paris Basin.
Conclusions
Reconstructing paleoenvironmental conditions, especially
variations of paleotemperatures, requires very well con-
strained ecological characteristics of the studied fossils toobtain reliable interpretations. In this study, we have
demonstrated that features such as the growth period of
fossils needs to be known. Nevertheless, the association ofdata coming from various species of molluscs with dif-
ferent periods of growth can constitute a powerful tool to
estimate both the mean annual temperatures and the max-imum temperatures of the year. Taking advantage of the
particularities of the fossils sampled at Grignon, we have
reconstructed, from stable isotopic compositions of oystersshells that mean annual seawater surface-temperatures ranged
between 18 and 23 "C during the Middle Lutetian, whereas V.imbricata and S. multisulcata shells indicate that the temper-ature of the warmer months could have reached 30 "C. This
study also shows the necessity to combine sedimentological,
paleontological, and geochemical data to constrain preciselythe environmental parameters and in particular the d18O of
seawater and the paleodepth.
The temperatures estimated indicate that the Lutetianclimate in the Paris Basin was similar to that of the present-
day southern Mediterranean Sea. It appears that these
subtropical climatic conditions, even if colder than duringthe previous stages, did not constitute a negative factor for
the establishment of the hot spot of marine biodiversity.
Rather, it seems that this event should be related to thepaleogeographic setting of western Europe at this time,
which exhibited a configuration close to the one where the
modern hot spot is observed, i.e., in the Indo-Pacificdomain, with important insularity.
This study confirms that the stable isotope compositions
of molluscs from near-shore environments are reliablewitnesses of the paleoenvironmental and paleoclimatic
evolution, as long as it is combined with sedimentological
and paleontological data. It seems necessary to diversifypaleoclimatic studies in these kinds of environments to
improve past climate models for the beginning of the
Cenozoic.
Acknowledgments The authors would like to thank P. Loubry andC. Lemzaouda (MNHN, Paris), who have taken the photos under UVlight, and N. Labourdette for the stable isotopes analyses. This paperis a contribution to the PPF ‘‘MNHN Etat et structure phylogenetiquede la biodiversite actuelle et fossile’’ (director: Ph. Janvier). Thisstudy was funded by a PhD grant from the French Ministry ofResearch and Education to Damien Huyghe and by funds from UPMCand CNRS to UMR 7193. We would like to thank the editor F.T. Fursich, M. Zuschin and an anonymous reviewer for their con-structive comments, and Martin Pickford (College de France, Paris)for correcting the English.
References
Abrard R (1925) Le Lutetien du Bassin de Paris: Essai demonographie stratigraphique. Societe Francaise d’Imprimerie,Angers 388 p
Al-Aasm I, Veizer J (1986) Diagenetic stabilization of aragonite andlow-Mg calcite; II, Stable isotopes in rudists. J Sediment Petrol56:763–770
Allmon W (1988) Ecology of Recent turritelline gastropods (Pros-obranchia, Turritellidae): current knowledge and paleontologicalimplications. Palaios 3:259–284
Allmon W (2011) Natural history of turritelline gastropods (Ceri-thiodea: Turritellidae): a status report. Malacologia 54:159–202
Anderson TF, Arthur MA (1983) Stable isotopes of oxygen andcarbon and their applications to sedimentological and palaeoen-vironmental problems. In: Arthur MA, Anderson TF, Kaplan IR,Veizer J, Land LS (eds) Stable isotopes in sedimentary geology.Soc Econ Paleontol Miner Short Course, vol 10, pp 1–151
Andreasson FP, Schmitz B (1996) Winter and summer temperaturesof the early middle Eocene of France from Turritella d18Oprofiles. Geology 24:1067–1070
Andreasson FP, Schmitz B (1997) Winter and summer temperaturesof the early middle Eocene of France from Turritella d18Oprofiles: reply. Geology 25:958–960
Andreasson FP, Schmitz B (2000) Temperature seasonality in theearly middle Eocene North Atlantic region: evidence from stableisotope profiles of marine gastropod shells. Geol Soc Am Bull112:628–640
Aubry M-P (1985) Northwestern European Paleogene magnetostra-tigraphy: calcareous nannofossil evidence. Geology 13:198–202
Bijl PK, Schouten S, Sluijs A, Reichart GJ, Zachos JC, Brinkhuis H(2009) Early Palaeogene temperature evolution of the southwestPacific Ocean. Nature 461:776–779
Bohaty SM, Zachos JC, Florindo F, Delaney ML (2009) Coupledgreenhouse warming and deep-sea acidification in the middleEocene. Paleoceanography 24, doi:10.1029/2008PA001676
Bouchet P, Lozouet P, Maestrati P, Heros V (2002) Assessing themagnitude of species richness in tropical marine environments:exceptionally high number of molluscs at a New Caledonia site.Biol J Linn Soc 75:421–436
Facies (2012) 58:587–604 601
123

Brunet MF, Le Pichon X (1982) Subsidence of the Paris Basin.J Geophys Res 87:8547–8560
Buchardt B (1978) Oxygen isotope palaeotemperatures from theTertiary period in the North Sea area. Nature 275:121–123
Burgess CE, Pearson PN, Lear CH, Morgans HEG, Handley L,Pancost RD, Schouten S (2008) Middle Eocene climate cyclicityin the southern Pacific: implications for global ice volume.Geology 36:651–654
Carick N (1980) Aspects of the biology of molluscs on the continentalshelf of Sydney, NSW with particular reference to the populationbiology of Gazameda gunnii (Reeve, 1949). Thesis. Universityof Sydney, Australia 175 p
Carter JG (1980) Guide to bivalve shell microstructures. In: RhoadsDC, Lutz RA (eds) Skeletal growth of aquatic organisms.Plenum Press, New York, p 142
Cavelier C, Pomerol C (1979) Chronologie et interpretation desevenements tectoniques cenozoıques dans le Bassin de Paris.Bull Soc Geol Fr 21(7):33–48
Caze B, Merle D, Pacaud J-M, Saint Martin J-P (2010) Firstsystematic study using the variability of the residual colourpatterns: the case of the Paleogene Seraphsidae (Mollusca,Gastropoda, Stromboidea). Geodiversitas 32:417–477
Caze B, Merle D, Le Meur M, Pacaud J-M, Ledon D, Saint Martin J-P(2011) Taxonomic implications of the residual color patterns ofampullinid gastropods and their contribution to the discrimina-tion of naticids. Acta Paleont Polon 56:353–371
Chaix C (1979) Enseignements d’ordre paleoecologique tires del’etude des Turbinolia et Sphenotrochus (Scleractiniaires) duBassin de Paris. Repartition stratigraphique et geographique deces deux genres. Bull Inf Geol Bass Paris 16:35–37
Crampton JS, Foote M, Cooper RA, Beu AG, Peters SE (2011) Thefossil record and spatial structuring of environments andbiodiversity in the Cenozoic of New Zealand. In: McGowanAJ, Smith AB (eds) Comparing the geological and fossil records:implications for biodiversity studies. Geological Society ofLondon Special Publications, vol 358, pp 105–122, doi:10.1144/SP358.8
Davitashvili LS, Merklin RL (1968) Handbook on the ecology ofmarine Gastropoda. Akademi Nauk Gruzinskoi S.S.R. TbilisiInstitute of Paleobiology, Tbilissi
de Franceschi D (2009) Les grandes lignes de la paleobiodiversite auLutetien: les paleoflores. Geochronique 109:1–23
de Gregorio A (1880) Fauna di San Giovanni Ilarione. Montaina,Palermo 106 p
Epstein S, Buchsbaum R, Lowenstam HA, Urey HC (1953) Revisedcarbonate-water isotopic temperature scale. Geol Soc Am Bull64:1315–1326
Fan C, Koeniger P, Wang H, Frechen M (2011) Ligamentalincrements of the mid-Holocene Pacific oyster Crassostreagigas are reliable independent proxies for seasonality in thewestern Bohai Sea, China. Palaeogeogr Palaeoclimatol Palaeo-ecol 299:437–448
Gely J-P (1996) Le Lutetien du Bassin Parisien : de l’analysesequentielle haute resolution a la reconstitution paleogeograph-ique. Bull Inf Geol Bass Paris 34:3–27
Gely J-P (2008) La stratigraphie et la paleogeographie du Lutetien enFrance. In: Merle D (ed) Stratotype Lutetien. MNHN/BRGM,Paris/Orleans, pp 182–227
Genot P (2009) Cenozoic dasycladales. A photo-atlas of Lutetianspecies from French Cenozoic basins. Notebooks GeologicalSpecial Papers, vol 1, 180 p
Goodwin DH, Schone BR, Dettman DL (2003) Resolution andfidelity of oxygen isotopes as paleotemperature proxies inbivalve mollusk shells: models and observations. Palaios18:110–125
Graham A (1938) On a ciliary process of food-collecting in thegastropod Turritella communis Risso. Proc Zool Soc Lond108:543–563
Greenwood DR, Wing SL (1995) Eocene continental climates andlatitudinal temperature gradients. Geology 23:1044–1048
Grossman EL, Ku TL (1986) Oxygen and carbon isotope fraction-ation in biogenic aragonite: temperature effects. Chem Geol59:59–74
Guelorget O, Perthuizot J-P (1983) Le domaine paralique: expres-sions geologiques, biologiques et economique du confinement.Travaux du laboratoire de geologie de l’ENS 16:1–136
Guernet C, Huyghe D, Lartaud F, Merle D, Emmanuel L, Gely J-P,Michel F, Pilet O (in press) La faune d’Ostracodes de la falunierede Grignon (Lutetien du Bassin de Paris): implications strati-graphiques et paleogeographiques. Geodiversitas
Guillocheau F, Robin C, Allemand P, Bourquin S, Brault N, DromartG, Friedenberg R, Garcia J-P, Gaulierg J-M, Gaumet F,Grosdoya B, Hanot F, Le Stratj P, Mettraux M, Nalpas T,Prijaca C, Rigolleta C, Serrano O, Grandjean G (2000) Meso-Cenozoic geodynamic evolution of the Paris Basin: 3D strati-graphic constraints. Geodin Acta 13:189–246
Hansen TA (1988) Early Tertiary radiation of marine molluscs andthe long term effects of the Cretaceous Tertiary extinction.Paleobiology 14:37–51
Haveles AW, Ivany LC (2010) Rapid growth explains large size ofmolluscs in the Eocene Gosport Sand, United States Gulf Coast.Palaios 25:550–564
Huyghe D, Castelltort S, Mouthereau F, Serra-Kiel J, FilleaudeauP-Y, Emmanuel L, Berthier B, Renard M (2012) Large scalefacies change in the middle-Eocene South-Pyrenean forelandbasin: the role of tectonics and prelude to Cenozoic ice-ages.Sediment Geol 253–254:25–46
Ivany LC, Runnegar B (2010) Early Permian seasonality from bivalved18O and implications for the oxygen isotopic composition ofseawater. Geology 38:1027–1030
Ivany LC, Nesbitt EA, Prothero DR (2003) The marine Eocene–Oligocene transition: a synthesis. In: Prothero DR, Ivany LC,Nesbitt EA (eds) From Greenhouse to Icehouse: the marineEocene–Oligocene transition. Columbia University Press, NewYork, pp 522–534
Ivany LC, Wilkinson BH, Lohman KC, Johnson ER, McElroy BJ,Cohen GJ (2004) Intra-annual isotopic variation in Venericardiabivalves: implications for early Eocene temperatures, seasonal-ity, and salinity on the U.S. Gulf Coast J Sediment Res 74:7–19
Ivany LC, Lohmann KC, Hasiuk F, Blacke DB, Glass A, AronsonRB, Moody RM (2008) Eocene climate record of a high southernlatitude continental shelf: Seymour Island, Antarctica. Geol SocAm Bull 120:659–678
Janssen R, Zuschin M, Baal C (2011) Gastropods and their habitatsfrom the northern Red Sea (Egypt: Safaga) Part 2: Caenogas-tropoda: Sorbeoconcha and Littorinimorpha. Ann Naturhist MusWien, Ser A 113:373–509
Jones D, Quitmyer IR (1996) Marking time with bivalve shells:oxygen isotopes and season of annual increment formation.Palaios 11:340–346
Kantor Y, Sisoev AV (2005) A preliminary analysis of biodiversity ofmolluscs of Russia. Ruthenica 14:107–118
Killingley JS, Berger WH (1979) Stable isotopes in a mollusk shell:detection of upwelling events. Science 205:186–188
Kirby MX (2000) Paleoecological differences between Tertiary andQuaternary Crassostrea oysters as revealed by stable isotopesclerochronology. Palaios 15:132–141
Kirby MX, Soniat TM, Spero HJ (1998) Stable isotope sclerochro-nology of Pleistocene and recent oyster shells (Crassostreavirginica). Palaios 13:560–569
602 Facies (2012) 58:587–604
123

Klein RT, Fricke HC, Purton L, Brasier M, Andreasson FP, SchmitzB (1997) Winter and summer temperatures of the early middleEocene of France from Turitella d18O profiles: comments andreply. Geology 25:957–958
Kobashi T, Grossman EL (2003) The oxygen isotopic record ofseasonality in Conus shells and is application to understandinglate middle Eocene (38 Ma) climate. Paleontol Res 7:343–355
Kobashi T, Grossman EL, Yancey TE, Dockery DT (2001) Reeval-uation of conflicting Eocene tropical temperature estimates:Molluskan oxygen evidence for warm low latitudes. Geology29:983–986
Lartaud F, Langlet D, de Rafelis M, Emmanuel L, Renard M (2006)Mise en evidence de rythmicite saisonniere dans la coquille deshuıtres fossiles Crassostrea aginensis Tournouer, 1914 (Aquita-nien) et Ostrea bellovacina Lamarck, 1806 (Thanetien). Ap-proche par cathodoluminescence et par sclerochronologie.Geobios 39:845–852
Lartaud F, Emmanuel L, de Rafelis M, Ropert M, Labourdette N,Richardson CA, Renard M (2010a) A latitudinal gradient ofseasonal temperature variation recorded in oyster shells from thecoastal waters of France and the Netherlands. Facies 56:13–25
Lartaud F, de Rafelis M, Ropert M, Emmanuel L, Geairon P, RenardM (2010b) Mn labelling of living oysters: artificial and naturalcathodoluminescence analysis as a tool for age and growth ratedetermination of C. gigas (Thunberg, 1793) shells. Aquaculture300:206–217
Lartaud F, Emmanuel L, de Rafelis M, Pouvreau S, Renard M(2010c) Influence of food supply on the d13C signature ofmollusc shells: implications for palaeoenvironmental reconsti-tutions. Geo-Mar Lett 30:23–34
Lartaud F, de Rafelis M, Olivier G, Krylova E, Dyment J, IldefonseB, Thibaud R, Gente P, Hoise E, Meistertzheim A-L, Fouquet Y,Gaill F, Le Bris N (2010d) Fossil clams from a serpentinite-hosted sedimented vent field near the active smoker complexRainbow, MAR, 36"130N: insight into the biogeography of ventfauna. Geochem Geophys Geosyst, vol 11. doi:10.1029/2010GC003079
Latal C, Piller WE, Harzhauser M (2006) Shifts in oxygen and carbonisotope signals in marine molluscs from the Central Paratethys(Europe) around the Lower/Middle Miocene transition. Palaeo-geogr Palaeoclimatol Palaeoecol 231:347–360
Le Calvez Y (1970) Contribution a l’etude des foraminiferespaleogenes du Bassin de Paris. Cahiers Paleontol CNRS, Paris
Le Calvez Y, Le Renard J (1980) Paleontologie de deux celebresgisements eocenes (Lutetien et Auversien) des environs de Paris.Compte rendu de l’excursion B-22. In: 26th Int Geol Cong,Paris. Bull Inf Geol Bass Paris, n" hors ser, pp 1–8
Lear CH, Elderfield H, Wilson PA (2000) Cenozoic deep-seatemperatures and global ice volumes from Mg/Ca in benthicforaminiferal calcite. Science 287:269–272
Lozouet P (1997) Le domaine atlantique europeen au Cenozoıquemoyen: diversite et evolution des Gasteropodes. PhD Thesis,Museum national d’Histoire naturelle, Paris, 309 p
McConnaughey TA (1989) 13C and 18O isotopic disequilibrium inbiological carbonates: II. In vitro simulation of kinetic isotopeeffects. Geochim Cosmochim Acta 53:163–171
McConnaughey TA, Gillikin DP (2008) Carbon isotopes in molluskshell carbonates. Geo-Mar Lett 28:287–299. doi:10.1007/s00367-008-0116-4
McConnaughey TA, Burdett J, Whelan JF, Paull CK (1997) Carbonisotopes in biological carbonates: respiration and photosynthesis.Geochim Cosmochim Acta 61:611–622
Merle D (1986) Contribution a l’etude paleontologique du gisementcuisien de Gan (Pyrenees-Atlantiques): systematique, evolutionet paleoecologie. Memoire de l’EPHE, (Dijon) 394 p
Merle D (2008) Le Lutetien du bassin de Paris: un exemple de point-chaud de la paleobiodiversite. In: Merle D (ed) StratotypeLutetien. MNHN/BRGM, Paris/Orleans, pp 174–181
Merle D, Courville P (2008) Les sites remarquables. In: Merle D (ed)Stratotype Lutetien. MNHN/BRGM, Paris/Orleans, pp 64–75
Merle D, Pacaud J-M, Kriloff A, Loubry P (2008) Le Lutetien duBassin de Paris: un exemple de point chaud de la paleobiodiv-ersite. Les motifs colores residuels des coquilles lutetiennes dubassin de Paris. In: Merle D (ed) Stratotype Lutetien. BRGM/MNHN, Orleans/Paris, pp 182–227
Miller KG, Fairbanks RA, Mountain GS (1987) Tertiary oxygenisotope synthesis, sea-level history and continental marginerosion. Paleoceanography 2:1–19
Pacaud J-M, Le Renard J (1995) Revision des mollusques paleogenesdu Bassin de Paris. IV—Liste systematique reactualisee. Coss-manniana 3:151–187
Payros A, Tosquella J, Bernaola G, Dinares-Turell J, Orue-EtxebarriaX, Pujalte V (2009) Filling the North European Early/MiddleEocene (Ypresian/Lutetian) boundary gap: insights from thePyrenean continental to deep-marine record. Palaeogeogr Palae-oclimatol Palaeoecol 280:313–332
Pekar SF, Hucks A, Fuller M, Li S (2005) Glacio-eustatic changes inthe early and middle Eocene (51–42 Ma): shallow-waterstratigraphy from ODP Leg 189 Site 1171 (South Tasman Rise)and deep-sea d18O records. Geol Soc Am Bull 117:1081–1093
Perez JM, Picard J (1964) Nouveau manuel de bionomie benthique dela Mer Mediterranee. Recueil des Travaux de la Station marined’Endoume 31:3–137
Pierre C (1999) The oxygen and carbon isotope distribution in theMediterranean water masses. Mar Geol 153:41–55
Pomerol C (1973) Ere Cenozoıque. Doin, ParisProthero DR (1994) The Eocene-Oligocene transition. Paradise lost.
Columbia University Press, New YorkPurton L, Brasier M (1997) Gastropod carbonate d18O and d13C
values record strong seasonal productivity and stratificationshifts during the late Eocene in England. Geology 25:871–874
Purton LMA, Brasier MD (1999) Life span and habitat insights fromd18O and d13C data from Nummulites and Venericardia,Hampshire basin, UK. Geology 27:711–714
Quaggiotto E, Mellini A (2008) Catalogo aggiornato dei molluschifossili eocenici di San Giovanni Ilarione (Verona–Italia setten-trionale) Prima parte: Mollusca, Gastropoda. Studi e Ricerche,Ass Amici Mus, Museo Civico ‘‘G. Zannato’’ 15:41–58
Richardson CA, Collis SA, Ekaratne K, Dare P, Key D (1993) Theage determination and growth rate of the European flat oyster,Ostrea edulis in British waters determined from acetate peels ofumbo growth lines. ICES J Mar Sci 50:493–500
Romanek CS, Grossman EL, Morse JW (1992) Carbon isotopicfractionation in synthetic aragonite and calcite: effects oftemperature and precipitation rate. Geochim Cosmochim Acta56:419–430
Schmidt GA (1999) Forward modeling of carbonate proxy data fromplanktonic Foraminifera using oxygen isotope tracers in a globalocean model. Paleoceanography 14:482–498
Seward D (1978) Palaeosalinities and palaeotemperatures fromcarbon and oxygen isotopes of carbonate shells in threeQuaternary formations, Wanganui Basin, New Zealand. Palae-ogeogr Palaeoclimatol Palaeoecol 23:47–55
Shackleton NJ, Kennett JP (1975) Paleotemperature history of theCenozoic and the initiation of Antarctic glaciation: oxygen andcarbon isotope analysis in DSDP site 277, 279 and 28. InitialRep Deep Sea Drill Proj 29:743–755
Smith AB, McGowan AJ (2011) The ties linking rock and fossilrecords and why they are important for paleobiodiversity studies.Geol Soc Lond Spec Publ 358:1–7
Facies (2012) 58:587–604 603
123

Squires RL, Saul LR (2007) Paleocene pareorine turritellid gastro-pods from the Pacific slope of North America. The Nautilus121:1–16
Stenzel HB (1971) Oysters. In: Moore RC (ed) Treatise in inverte-brate paleontology, Mollusca 6, Bivalvia (3). Geological Societyof America/Boulder, University of Kansas Press/Lawrence
Stevens KF, Vella P (1981) Palaeoclimatic interpretation of stableisotope ratios in molluscan fossils from middle Pleistocenemarine strata, Wanganui, New Zealand. Palaeogeogr Palaeocli-matol Palaeoecol 34:257–265
Surge DM, Lohmann KC, Dettman DL (2001) Controls on isotopicchemistry of the American oyster, Crassostrea virginica:implications for growth patterns. Palaeogeogr PalaeoclimatolPalaeoecol 172:283–296
Surge DM, Lohmann KC, Goodfriend GA (2003) Reconstructingestuarine conditions: oyster shells as recorders of environmentalchange, Southwest Florida. Estuar Coast Shelf Sci 57:737–756
Tarutani T, Clayton RN, Mayeda TK (1969) The effect of polymor-phism and magnesium substitution on oxygen isotope fraction-ation between calcium carbonate and water. GeochimCosmochim Acta 33:987–996
Thiry M (1989) Geochemical evolution and paleoenvironments of theEocene continental deposits in the Paris Basin. PalaeogeogrPalaeoclimatol Palaeoecol 70:153–163
Titschack J, Zuschin M, Spotl C, Baal C (2010) The giant oysterHyotissa hyotis from the northern Red Sea as a decadal-scalearchive for seasonal environmental fluctuations in coral reefhabitats. Coral Reefs 29:1061–1075
Tivollier J, Letolle R (1968) Resultat et interpretation d’analysesisotopiques de faunes malacologiques du Tertiaire parisien. BurRech Geol Min Mem 58:347–358
Tripati AK, Zachos J, Marincovich L, Bice K (2001) Late PaleoceneArctic coastal inferred from molluscan stable and radiogenicisotope ratios. Palaeogeogr Palaeoclimatol Palaeoecol170:101–113
Tripati AK, Backman J, Elderfield H, Ferretti P (2005) Eocene bipolarglaciation associated with global carbon cycle changes. Nature436:341–346
Tripati AK, Eagle RA, Morton A, Dowdeswell JA, Atkinson KL,Bahe Y, Dawber CF, Khadun E, Shaw RMH, Shorttle O,Thanabalasundaram L (2008) Evidence for glaciation in the
Northern Hemisphere back to 44 Ma from ice-rafted debris inthe Greenland Sea. Earth Planet Sci Lett 265:112–122
Tripati AK, Allmon WD, Sampson DE (2009) Possible evidence for alarge decrease in seawater strontium/calcium ratios and stron-tium concentrations during the Cenozoic. Earth Planet Sci Lett282:122–130
Waite R, Strasser A (2011) A comparison of Recent and fossil large,high-spired gastropods and their environments: the NopparatThara tidal Xat in Krabi, South Thailand, versus the SwissKimmeridgian carbonate platform. Facies 57:223–248
Watters GT (1993) Some aspects of the functional morphology of theshell of infaunal bivalves (Mollusca). Malacologia 35:315–342
Wefer G, Berger WH (1991) Isotope paleontology: growth andcomposition of extant calcareous species. Mar Geol 100:207–248
Yonge CM (1946) On the habits of Turritella communis Risso. J MarBiol Assoc 21:687–704
Yonge CM (1969) Functional morphology and evolution within theCarditacea (Bivalvia). Malacol Soc Lond Proc 38:493–527
Zachos JC, Quinn TM, Salamy K (1996) High resolution (104 yr)deep-sea foraminiferal stable isotope records of the Eocene-Oligocene climate transition. Paleoceanography 11:251–266
Zachos JC, Pagani M, Sloan L, Thomas E, Billups K (2001) Trends,rhythms, and aberrations in global climate 65 Ma to Present.Science 292:686–693
Zachos JC, Dickens GR, Zeebe RE (2008) An early Cenozoicperspective on greenhouse warming and carbon-cycle dynamics.Nature 451:279–283
Zuschin M, Oliver PG (2003) Bivalves and bivalve habitats in thenorthern Red Sea. The Northern Bay of Safaga (Red Sea, Egypt):an actuopalaeontological approach. VI. Bivalvia. Naturhistoris-ches Museum, Wien
Zuschin M, Janssen R, Baal C (2009) Gastropods and their habitatsfrom the northern Red Sea (Egypt, Safaga). Part 1: Patellogas-tropoda, Vetigastropoda and Cycloneritimorpha. Ann NaturhMus Wien Ser A 111:73–158
Zuschin M, Harzhauzer M, Mandic O (2011) Disentangling palae-odiversity signal from a biased sedimentary record: an examplefrom Early to Middle Miocene of Central Paratethys Sea. In:McGowan AJ, Smith AB (eds) Comparing the geological andfossil records: Implications for biodiversity studies. GeologicalSociety of London Special Publications, vol 358, pp 123–139
604 Facies (2012) 58:587–604
123
Related Documents











