REVIEW MicroRNAs and Tumor Vasculature Normalization: Impact on Anti-Tumor Immune Response Agata Matejuk • Guillaume Collet • Mahdi Nadim • Catherine Grillon • Claudine Kieda Received: 20 July 2012 / Accepted: 15 January 2013 / Published online: 11 April 2013 Ó L. Hirszfeld Institute of Immunology and Experimental Therapy, Wroclaw, Poland 2013 Abstract Inefficient immune response is a major glitch during tumor growth and progression. Chaotic and leaky blood vessels created in the process of angiogenesis allow tumor cells to escape and extricate anti-cancer immunity. Proangiogenic characteristics of hypoxic tumor microen- vironment maintained by low oxygen tension attract endothelial progenitor cells, drive expansion of cancer stem cells, and deviantly differentiate monocyte descendants. Such cellular milieu further boosts immune tolerance and eventually appoint immunity for cancer advantage. Blood vessel normalization strategies that equilibrate oxygen levels within tumor and fix abnormal vasculature bring exciting promises to future anticancer therapies especially when combined with conventional chemotherapy. Recently, a new group of microRNAs (miRs) engaged in angiogenesis, called angiomiRs and hypoxamiRs, emerged as new therapeutic targets in cancer. Some of those miRs were found to efficiently regulate cancer immunity and their dysregulation efficiently programs aberrant angio- genesis and cancer metastasis. The present review highlights new findings in the field of miRs proficiency to normalize aberrant angiogenesis and to restore anti-tumor immune responses. Keywords MicroRNAs regulation Hypoxia Angiogenesis Cancer Vessels normalization Tumor immune response Introduction Vasculogenesis and angiogenesis are highly regulated processes. In physiological conditions they are abundant during fetus development but restricted to wound healing and menstrual cycles during adulthood. They become prominent in pathological conditions such as cancer, dia- betic retinopathy, macular degeneration, atherosclerosis, and arthritis. On the other hand, inefficient angiogensis is a concern during stroke and coronary arterial disease. Hypoxia plays critical role in initiation of molecular events directly leading to formation of new blood vessels. Hypoxia-inducible factor (HIF) family by its pivotal influence on a variety of genes including vascular endo- thelial growth factor (VEGF) is a key regulator of tumor angiogenesis (Gordan and Simon 2007; Gruber and Simon 2006). Hypoxia-dependent growth factors and pro-angio- genic genes activate previously quiescent endothelial cells, induce proinflamatory factors, degrade and remodel extracellular matrix, and increase cancer cell mobility and their dedifferentiation. All these events lead to progressive tumor angiogenesis and result in immature, leaky, and unevenly distributed vessels leading to inefficient engage- ment of anti-cancer immune response and poor accessibility of tumor cells to chemotherapeutic drugs. Additionally, persisting low oxygen levels in tumor mass render it unresponsive to radiotherapy (Jordan and Son- veaux 2012). Anti-angiogenic agents are now being approved and introduced in clinics for the prevention and treatment of A. Matejuk (&) G. Collet M. Nadim C. Grillon C. Kieda Centre de Biophysique Mole ´culaire, CNRS UPR 4301, rue Charles Sadron, 45071 Orle ´ans, France e-mail: [email protected] A. Matejuk Le StudiumÒ, Loire Valley Institute for Advanced Studies, 3D, avenue de la Recherche Scientifique, 45071 Orle ´ans, France M. Nadim Libragen–Induchem Company, 3, rue des Satellites, Bat. Canal Biotech, 31400 Toulouse, France Arch. Immunol. Ther. Exp. (2013) 61:285–299 DOI 10.1007/s00005-013-0231-4 123

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

REVIEW
MicroRNAs and Tumor Vasculature Normalization:Impact on Anti-Tumor Immune Response
Agata Matejuk • Guillaume Collet •
Mahdi Nadim • Catherine Grillon • Claudine Kieda
Received: 20 July 2012 / Accepted: 15 January 2013 / Published online: 11 April 2013
� L. Hirszfeld Institute of Immunology and Experimental Therapy, Wroclaw, Poland 2013
Abstract Inefficient immune response is a major glitch
during tumor growth and progression. Chaotic and leaky
blood vessels created in the process of angiogenesis allow
tumor cells to escape and extricate anti-cancer immunity.
Proangiogenic characteristics of hypoxic tumor microen-
vironment maintained by low oxygen tension attract
endothelial progenitor cells, drive expansion of cancer stem
cells, and deviantly differentiate monocyte descendants.
Such cellular milieu further boosts immune tolerance and
eventually appoint immunity for cancer advantage. Blood
vessel normalization strategies that equilibrate oxygen
levels within tumor and fix abnormal vasculature bring
exciting promises to future anticancer therapies especially
when combined with conventional chemotherapy.
Recently, a new group of microRNAs (miRs) engaged in
angiogenesis, called angiomiRs and hypoxamiRs, emerged
as new therapeutic targets in cancer. Some of those miRs
were found to efficiently regulate cancer immunity and
their dysregulation efficiently programs aberrant angio-
genesis and cancer metastasis. The present review
highlights new findings in the field of miRs proficiency to
normalize aberrant angiogenesis and to restore anti-tumor
immune responses.
Keywords MicroRNAs regulation � Hypoxia �Angiogenesis � Cancer � Vessels normalization �Tumor immune response
Introduction
Vasculogenesis and angiogenesis are highly regulated
processes. In physiological conditions they are abundant
during fetus development but restricted to wound healing
and menstrual cycles during adulthood. They become
prominent in pathological conditions such as cancer, dia-
betic retinopathy, macular degeneration, atherosclerosis,
and arthritis. On the other hand, inefficient angiogensis is a
concern during stroke and coronary arterial disease.
Hypoxia plays critical role in initiation of molecular events
directly leading to formation of new blood vessels.
Hypoxia-inducible factor (HIF) family by its pivotal
influence on a variety of genes including vascular endo-
thelial growth factor (VEGF) is a key regulator of tumor
angiogenesis (Gordan and Simon 2007; Gruber and Simon
2006). Hypoxia-dependent growth factors and pro-angio-
genic genes activate previously quiescent endothelial cells,
induce proinflamatory factors, degrade and remodel
extracellular matrix, and increase cancer cell mobility and
their dedifferentiation. All these events lead to progressive
tumor angiogenesis and result in immature, leaky, and
unevenly distributed vessels leading to inefficient engage-
ment of anti-cancer immune response and poor
accessibility of tumor cells to chemotherapeutic drugs.
Additionally, persisting low oxygen levels in tumor mass
render it unresponsive to radiotherapy (Jordan and Son-
veaux 2012).
Anti-angiogenic agents are now being approved and
introduced in clinics for the prevention and treatment of
A. Matejuk (&) � G. Collet � M. Nadim � C. Grillon � C. Kieda
Centre de Biophysique Moleculaire, CNRS UPR 4301, rue
Charles Sadron, 45071 Orleans, France
e-mail: [email protected]
A. Matejuk
Le Studium�, Loire Valley Institute for Advanced Studies, 3D,
avenue de la Recherche Scientifique, 45071 Orleans, France
M. Nadim
Libragen–Induchem Company, 3, rue des Satellites, Bat. Canal
Biotech, 31400 Toulouse, France
Arch. Immunol. Ther. Exp. (2013) 61:285–299
DOI 10.1007/s00005-013-0231-4
123

cancer. However, the benefits remain modest. As much as
these therapies bring exciting results majority of tumors
eventually become refractory. Monoclonal antibody-based
therapies are mostly directed towards the main key players
in angiogenesis: VEGF and its receptor VEGFR2. Indeed
they were found to efficiently target vasculature but the
effect was only transient (Carmeliet and Jain 2011a).
Especially anti-VEGF monotherapies failed (Carmeliet and
Jain 2011a; Cobleigh et al. 2003; Giantonio et al. 2007;
Jain et al. 2006) since anti-VEGF/VEGFR treatments boost
hypoxic state that further allow for selection of highly
resistant and aggressive stem-like cancer cells with pro-
gressively lower response to conventional therapies.
Accumulating data clearly show that cutting off the blood
flow to the tumor initiates atrocious compensatory mech-
anisms. In contrast, blood vessel normalization approaches
that fix the leaky, twisty blood vessels that feed tumors,
thereby improving the delivery of drugs are encouraging
and are under fast development (Carmeliet and Jain 2011b;
Jain 2009). Consequently, blood vessel normalization
concept covers and targets hypoxia, angiogenesis, cancer
stem cells differentiation, radio-, drug-response efficacy,
and reestablishment of anti-tumor immune response. Cur-
rent aims of anti-angiogenic therapy is to create a
normalization window where blood vessels regain their
typical structure with decreased permeability for better
drug penetration (Jain 2005; Tong et al. 2004; Wildiers
et al. 2003).
Recently, new opportunities for better understanding of
tumor biology came with discovery of microRNAs (miRs)
a new class of gene regulatory molecules. miRs are 19–25
single-stranded RNAs capable of targeting cleavage and/or
mRNA translational inhibition or inducing mRNA insta-
bility. Those non-coding RNAs control the expression level
of genes involved in development, proliferation, differen-
tiation, and apoptosis (He and Hannon 2004). Alterations in
miRs synthesis pathways and processing have important
consequences for both physiological and pathological
processes such as development and cancer (Winter et al.
2009). In fact, miRs have been shown to affect the hall-
marks of cancer, including angiogenesis and aberrant
immune response and their profile and level of expression
are changed in malignancies. Presently, more than 1400
human miRs are identified (Kozomara and Griffiths-Jones
2011) and the number is growing. A single miR can reg-
ulate a variable number of targets (Thomas et al. 2010).
Deeper understanding of the crosstalk between tumor and
immune cells in a language of angio-and/or hypoxa-miRs
should unravel miRs implications and potential new targets
for treatment and diagnostics. Recent review by Hartmann
and Thum (2011) describes in detail individual miRs
involved in vascular (dys)function. Caporali and Emanueli
(2011) present miRs that regulate angiogenesis. In this
review, we briefly discuss miRs that control tumor angi-
ogensis and have potential to target aberrant anti-cancer
immune responses.
Blood Vessel Normalization Paradigm
Vascular normalization hypothesis was first introduced by
Jain (2001). Tumor hypoxic environment boosts produc-
tion of VEGF and other proangiogenic factors and initiates
angiogenesis that results in abnormal blood vessel network.
Tumor vessels are highly irregular; the junctions among
endothelial cells and connective tissue cells such as peri-
vascular cells are loose, leading to leaky vascular structure
with multiple fenestrations (Fukumura et al. 2010; Nagy
et al. 2009). Pseudo-vessel formation is also linked to a
decrease in endothelial cell ability to adhere to each other.
Dysregulation of normal vessel maturation has catastrophic
consequences for any given anti-cancer therapy. Hyper-
permeable phenotype allows metastatic tumor cells to
escape and strongly prevents infiltration of anti-cancer
immunity and drug penetration to the tumor mass. It also
poorly delivers oxygen further enhancing hypoxic state
(Hockel and Vaupel 2001). Hypoxic condition induces
dedifferentiation of cells, epithelial-mesenchymal transi-
tion, increased cell migration and invasion, and high
resistance to chemo-, radio- (Teicher 1996), and immuno-
therapy (Noman et al. 2011). It also precludes cytotoxicity
of anti-cancer immunity (Ganss et al. 2004; Hamzah et al.
2008). In clinical applications, anti-VEGF monotherapy
that targets vascular recession by cutting off blood supply
was significantly unsuccessful. However, direct targeting
of VEGF as addition to standard chemotherapy showed
improvements in patient outcomes (Saltz et al. 2008;
Tebbutt et al. 2010). To explain this paradox Jain proposed
the ‘‘vascular normalization’’ hypothesis. It states that
cautious use of anti-angiogenic therapy creates a ‘‘nor-
malization window’’ that reverses aberrant angiogenesis,
leads to normalized vascular phenotype, and reduces vessel
permeability and hypoxia (Goel et al. 2011, 2012). Direct
or indirect anti-angiogenic therapy regulates equilibrium
between pro- and anti-angiogenic factors and controls
pathways crucial for maintenance of healthy angiogenesis.
Besides controlling VEGF levels, direct anti-angiogenic
therapies regulate angiopoietin-Tie2 pathway (Falcon et al.
2009), expression of placental growth factor (PlGF)
(Fischer et al. 2007; Van de Veire et al. 2010), endothelial
cell-specific integrins (Desgrosellier and Cheresh 2010;
Primo et al. 2010; Skuli et al. 2009), and the prolyl-4-
hydroxylase domain (PHD) tumor-oxygen sensor proteins
(Mazzone et al. 2009). The latter senses low oxygen ten-
sion and by predisposing to hypoxia generates abnormal
vessels. PHD2?/- mice displayed reduced metastasis due
286 Arch. Immunol. Ther. Exp. (2013) 61:285–299
123

to decreased number of cancer cells from primary tumor in
blood stream (Mazzone et al. 2009). A recent study shows
that gene-targeting PHD2 improves tumor response to
chemotherapy and prevents side-toxicity (Leite de Oliveira
et al. 2012). Indirect inhibitors of angiogenesis target tumor
cell oncogenes such as HER-2, the PI3K-AKT-mTOR axis,
Ras and epidermal growth factor receptor, and include
endocrine therapies and metronomic chemotherapy (Goel
et al. 2012). The goal in ‘‘blood vessel normalization’’
strategy is to balance the action of pro-angiogenic factors
such as VEGF, basic fibroblast growth factor (bFGF), and
angiopoietin-2 (Ang-2) with action of anti-angiogenic
mediators such as thrombospondin-1 (TSP-1) and Ang-1
(Jain 2003; Relf et al. 1997). Ang-1 is an agonist of
endothelial Tie-2 receptor promoting blood vessel nor-
malization and Ang-2 is an antagonist of the same receptor
leading to increased angiogenesis. Ang-Tie-2 axis is crucial
in regulating both healthy and tumor-associated angio-
genesis (Huang et al. 2010). A recent approach that might
lead to blood vessel normalization is based on the regula-
tion of oxygen partial pressure within tumor mass.
Previously Kieda et al. (2006) and Sihn et al. (2007)
showed that myo-inositol trispyrophosphate (ITPP), a
ubiquitous carbohydrate and allosteric effector of hemo-
globin causing oxygen release, was able both in vitro and
in vivo to inhibit hypoxia-induced angiogenesis. Studies on
ITPP’s ability to normalize oxygen tension in tumor are
ongoing.
The Role of Blood Vessel Normalization Strategies
in Anti-Cancer Immune Responses
The ability to create metastasis and neovascularization
strongly depends on active interplay between tumor cells
and their microenvironment including extracellular matrix,
endothelial cells, and immune cells. Proper immune
responses achieved mainly by natural killer (NK) cells and
cytotoxic T cells are paramount in the eradication of can-
cer. As tumor growth progresses and hypoxic state
propagates, the development of immune tolerance and
activation of immune-related suppressor cells ascends.
Additionally, hypoxia surge resistance to cytotoxicity of
immune cells infiltrating tumor mass (Ganss et al. 2004).
Over the past few years there is a constant progression in
the development of new immunotherapies either with
active vaccinations or with adoptive cell transfers that
assure enhancement of cytotoxic anti-cancer immunity.
Vaccines targeting the neovasculature of tumors show
antitumor efficacy, inhibition of local suppression mecha-
nisms, and boosting anti-tumor immunity (Matejuk et al.
2011). However, the outcome of such therapies is still
modest. A few preclinical studies show increase of anti-
cancer immune responses when immunotherapy is com-
bined with anti-angiogenic approaches (Huang et al. 2002;
Li et al. 2006; Manning et al. 2007; Shrimali et al. 2010).
One of the possible mechanisms of such therapies is abo-
lition of immunosuppressive properties of VEGF.
Moreover, normalization of blood vessels can directly
improve infiltration of immune-competent cells. The role
of VEGF in immunosuppression is still not explained well.
It has been reported that VEGF upregulates adhesion
molecules on angiogenic vessels and eases adhesion and
rolling of NK cells (Melder et al. 1996). On the contrary,
Griffioen et al. (1996) showed that VEGF inhibits expres-
sion of leukocytes adhesion molecules and prevents
leukocyte tumor infiltration. In inducible nitric oxide syn-
thase (iNOS)-/- mice despite the increased level of
intratumoral VEGF, interleukin (IL)-12 induced a stronger
inhibition of blood vessel formation and recruitment of NK
cells (Bielawska-Pohl et al. 2010). Nitric oxide has been
shown to modulate endothelial interactions and expression
of junction molecules such as CD31 (Carreau et al. 2011).
In some studies VEGF has been shown to inhibit the
maturation and function of dendritic cells (Gabrilovich
et al. 1996; Osada et al. 2008). Additionally, VEGF was
proven to possess additive effect on immunosuppression in
tumor by attracting regulatory T cells (Tregs) and Gr1?
myeloid suppressor cells (Li et al. 2006; Shojaei et al.
2007). Besides tumor cells, myeloid cell that infiltrate
tumor site are main sources of VEGF. Stockmann et al.
(2008) by using mice lacking myeloid cell-derived VEGF
showed that myeloid cell-specific deletion of VEGF
resulted in vascular normalization, better oxygenation, and
better response to chemotherapeutic cytotoxicity. However,
unexpectedly, deletion of myeloid-cell VEGF-a resulted in
an accelerated tumor progression. Recently, Shrimali et al.
(2010) observed beneficial effect of anti-VEGF-a antibody
therapy combined with immunotherapy in melanoma
models by increased influx of antitumor cytotoxic T cells.
Vessel stability and normalization can also be obtained by
targeting Ang-Tie2 pathway. Ang-1 promotes maturation
of vessels and is produced by perivascular cells, whereas
Ang-2 produced by endothelial cells possesses proangio-
genic and vasculature destabilizing activity. Both growth
factors compete for Tie-2 receptor. The attempts to block
Ang-2 were successful in blood vessel normalization and
interfered with the proangiogenic, immunosupressive
activities of Tie-2 expressing monocytes (TEMs) (Coffelt
et al. 2010; Mazzieri et al. 2011). Abnormal vessel angi-
ogenesis is also mediated by PlGF. The histidine-rich
glycoprotein can downregulate PlGF and induce blood
vessel maturation, inhibition of hypoxia and switch from
pro-tumorogenic M2 macrophages to M1 with anti-tumor
activity (Huang et al. 2011b; Rolny et al. 2011). Thus,
regulation of abnormal tumor vessel phenotype has clear
Arch. Immunol. Ther. Exp. (2013) 61:285–299 287
123

effect on the restoration of a proper immune response in
tumor environment (Fig. 1).
Angiomirs are Regulators of Angiogenesis
and Endothelial Cell Functions
miRs that tightly regulate the process of endothelial cell
function and regulate the vascular integrity, especially
angiogenesis, are referred as angiomiRs (Wang and Olson
2009). Pioneering work on miRs involved in the process of
angiogenesis came from studies on two key miRs biosyn-
thesis enzymes: Dicer and Drosha. The first report came
from the studies on mice homozygous for hypomorphic
allele of Dicer. Homozygous embryos displayed a retarded
phenotype, no angiogenesis, and died during gestation
period (Yang et al. 2005). Silencing of Dicer and Drosha in
endothelial cells inhibited endothelial cell migration and
tube formation; however, silencing Dicer only has a pro-
found effect on angiogenesis in vivo (Yang et al. 2005).
Later, several other in vitro studies on human endothelial
cells, using Dicer and Drosha knockdowns, demonstrated
inhibition of angiogenic events like proliferation, migra-
tion, and capillary sprouting (Kuehbacher et al. 2007; Shilo
et al. 2008; Suarez et al. 2007, 2008). Several key angio-
genic mediators such as VEGFs, their receptors: VEGFR1
and VEGFR2 as well as Ang-2 receptor Tie-1, endothelial
NOS (eNOS), and IL-8, were strongly affected by Dicer
deletion (Suarez et al. 2007; Yang et al. 2005). In addition,
TSP-1, a potent anti-angiogenic factor was significantly up-
regulated (Kuehbacher et al. 2007). Interestingly, recently a
novel Dicer-independent miRs processing pathway has
been described (Cifuentes et al. 2010). Rescue for endo-
thelial cell proliferation and morphogenesis in Dicer null
mice could be achieved by overexpression of miR-17-92
cluster members, especially miR-18a (Suarez et al. 2008).
Of note, the miR-17-92 cluster was the first oncogenic miR
described in mammals (He et al. 2005). MiR-17-92 toge-
ther with miR-155 are proven to cause oncogenesis
(Mendell 2008; O’Connell et al. 2008). Mounting evidence
indicates that miRs are emerging as important modulators
of angiogenesis in normal and pathological conditions such
as cancer.
First 15 miRs in human umbilical vein endothelial cells
(HUVEC) were identified by Poliseno et al. (2006). Pres-
ently 28 miRs are known as expressed by cultured
endothelial cells (Chen and Gorski 2008; Fasanaro et al.
2008; Fish et al. 2008; Kuehbacher et al. 2007; Suarez et al.
Fig. 1 Schematic representation of complex anti-cancer approaches.
The illustration shows the result of action of combinatory treatment of
blood vessel normalization stratagies that might include also miRs
characteristic for physioxia state (tissue-dependent proper oxygen
partial pressure) and conventional therapy (chemotherapy and/or
radiotherapy). It leads to reduction of hypoxia, downregulation of
hypoxiamiRs, and reduction of aberrant immune response mostly
derived from monocyte lineage. Restoration of physioxia and proper
anti-cancer immune response is achieved
288 Arch. Immunol. Ther. Exp. (2013) 61:285–299
123

2007, 2008; Wurdinger et al. 2008). In vivo studies
revealed miR-126, -24 and -23a as specifically expressed
by microvascular endothelial cells, and predominant
expression of miR-145 by pericytes (Larsson et al. 2009).
In a recent study miR-145 was found to be expressed by
HUVEC and was able to control smooth muscle pheno-
types with potential to combat atherosclerosis
(Hergenreider et al. 2012). MiR-126 is the most habitually
expressed miR found in HUVEC and is characteristic of
endothelial cells from veins, arteries, and in brain and skin.
So far miR-126 seems to be a master regulator of vascular
integrity and angiogenesis that can inhibit tumor growth
and invasion. It positively regulates signaling downstream
of growth factors like VEGF, bFGF, and EGF (Fish et al.
2008; Kuhnert et al. 2008; Wang et al. 2008) thus inhib-
iting miR-126 might suppress cancer neovascularization.
Knockout of an endothelial cell-specific miR-126 in zeb-
rafish and mice caused leaky vessels, hemorrhaging, and
impaired neoangiogenesis (Fish et al. 2008; Kuhnert et al.
2008; Wang et al. 2008). Its importance in vascular
inflammation is evident by controlling leukocyte traffick-
ing to sites of inflammation and suppressing vascular cell
adhesion molecule-1, a molecule coordinating adhesion of
leukocytes to endothelium (Harris et al. 2008). Low levels
of miR-126 promoted adhesive interactions between leu-
kocytes and endothelium (Harris et al. 2008; van Solingen
et al. 2009) and its downregulation was observed in many
cancer cell lines (Crawford et al. 2008; Guo et al. 2008;
Tavazoie et al. 2008). Recently, miR-126 was reported to
increase endothelial VEGF signaling by pressure of blood
flow and the mechano-sensitive zinc finger transcription
factor klf2a (Nicoli et al. 2010). Besides miR-126, exam-
ples of other classic angio-miRs are miR-221/222, miR-23-
27, and miR-17-92 cluster (Wang and Olson 2009). miR-
221/222 modulate the angiogenic capacity of HUVEC.
They specifically target stem cell factor receptor and c-kit
and thus are able to inhibit angiogenesis by reducing
endothelial cell migration and proliferation (Li et al. 2009;
Matsui et al. 2004; Poliseno et al. 2006). A recent study in
zebrafish revealed the essential role for miR-221 in angi-
ogenesis with its critical role in tip cell formation and
migration (Nicoli et al. 2012). It has been shown that miR-
221/222 can regulate the level of eNOS involved in
endothelial cell migration, but the effect must be indirect
since they do not target 30-UTR of eNOS (Suarez et al.
2007). Minami et al. (2009) suggested that miR-221/222
are related to proliferative capacity of endothelial cells.
They showed that in patients with coronary artery disease
levels of miR-221/222 were higher which correlated with
lower number of endothelial progenitor cells (Minami et al.
2009). Of note, these miRs can also regulate p27(Kip1)
tumor suppressor promoting cancer cell proliferation (le
Sage et al. 2007). The crosstalk between miRs and
angiogenic factors is two-directional. The angiogenic fac-
tors produced by the tumor cells can change miR profile.
For example, miR-378 promotes angiogenesis and can-
cerogenesis in tumor models (Lee et al. 2007) by inhibiting
cell differentiation (Kahai et al. 2009; Lee et al. 2007) as
well as induction of VEGF expression (Hua et al. 2006).
MiR-378 competes with miR-125a for the same seed
region of VEGFs (Lee et al. 2007). Another angiomiR,
miR-296 expression increases during endothelial cells
co-stimulation with glioma cells and its inhibition reduces
vascularization of tumor xenografts (Wurdinger et al.
2008). miR-296 contributes to angiogenesis by targeting
the hepatocyte growth factor-regulated tyrosine kinase
substrate mRNA and reducing degradation of VEGFR2 and
PDGFRb (Hua et al. 2006; Wurdinger et al. 2008). The
secreted factor, TSP1 acting on angiogenesis inhibition is
targeted by the cluster of miRs-17 through -92 (Dews et al.
2006). miRs belonging to miR-17-92 family are known
oncogenes and are integrated components of the molecular
pathways that regulate tumor development and tumor
maintenance (Olive et al. 2010). All crucial events in
cancer development and progression such as uncontrolled
cell proliferation, inhibition of apoptosis, and tumor angi-
ogenesis are promoted by these miRs (Mendell 2008).
miR-17-92 cluster has potent tumor angiogenesis stimu-
lating activity. Inhibition of miR-17, -18a, and -20a
increased HUVEC sprouting in 3D spheroid model in vitro,
and in vivo increased number of blood vessels was
observed by blocking of miR-17 and miR-20a with a lesser
effect done by miR-18a and miR-19a (Doebele et al. 2010).
MiR-17/20 targets proangiogenic genes including Janus
kinase 1 (Doebele et al. 2010). Inhibition of miR-17/20
specifically augmented neovascularization of Matrigel
plugs but did not affect tumor angiogenesis indicating a
context-dependent regulation of angiogenesis by miR-17/
20 in vivo (Doebele et al. 2010). Moreover, miR-17-92
cluster belongs to hypoxamiRs that can affect HIF inde-
pendently of hypoxia (Loscalzo 2010). There is an intricate
and finely tuned circuit involving this family of miRs,
c-myc, and HIF-1a that may play a role in cancer cell
proliferation under normoxia (Taguchi et al. 2008). Inter-
estingly, recent findings depict one of miR-17-92 cluster
members, miR-92a, to be a negative regulator of new blood
vessel formation. AntagomiR that counteracted the effect
of miR-92a caused induction of blood vessel growth and
reduced damages caused by ischemia and myocardial
infarction in mouse model (Bonauer et al. 2009). MiR-21
and miR-31 possess pro-angiogenic function and are
upregulated in cancer promoting invasion and metastasis.
VEGF produced by tumor cells, crucial for angiogenesis, is
suppressed by several miRs including miR-15b, -16, -20a,
and -20b (Hua et al. 2006). Those miRs might be of special
interest in vessel normalization strategy since transfection
Arch. Immunol. Ther. Exp. (2013) 61:285–299 289
123

to the cells can specifically target VEGF. miR-16 nega-
tively regulates VEGF translation by binding to VEGF 30-UTR (Karaa et al. 2009). Recently Chamorro-Jorganes
et al. (2011), reported that miR-16-like family members
(possessing the same seed sequence), including miR-15,
-16 and -424 critically regulate expression of VEGF-a,
their receptor VEGFR2, and fibroblast growth factor
receptor-1 (FGFR1). miR-15 and miR-16, by targeting
Acvr2a, a ligand belonging to transforming growth factor
(TGF)-b superfamily, interfere with TGF-b signaling. They
are also negatively regulated by Wnt/b-catenin pathway
(Martello et al. 2007). Members of another family miR-23-
27 cluster, miR-27a, and miR-27b are highly expressed in
endothelial cells and inhibition of miR-27b markedly
reduced endothelial sprouting in vitro (Kuehbacher et al.
2007). This family has been also found to regulate angio-
genesis (Zhou et al. 2011). Recently Urbich et al. (2012)
showed that miR-27a/b promote angiogenesis by targeting
the angiogenesis inhibitor Semaphorin 6A, which controls
repulsion of neighboring endothelial cells. Another potent
regulator of angiogenesis is miR-130a that induces novel
vessel formation by down-regulating anti-angiogenic
homeobox genes GAX and HOXA5 (Chen and Gorski
2008).
HypoxamiRs are Potential Targets for Blood Vessel
Normalization Strategies
Decrease in the partial pressure of oxygen to reach a low
level, called hypoxia, initiates genetic changes directly
leading to angiogenic response. Other cellular functions
controlled by hypoxia are related to proliferation, apopto-
sis, and metabolism. Hypoxic state is a hallmark of cancer
environment. Tumor cells under hypoxia tend to be resis-
tant to therapies and constitute a poor prognosis. Persistent
hypoxia correlates with metastasis and reduced survival in
patients (Hockel et al. 1999). Hypoxia triggers the stabil-
ization of oxygen-dependent transcription factor HIF that
further initiates the synthesis of the main pro-angiogenic
factors VEGF-A, -B, and -C (Rocha 2007). This leads to
initiation of neoangiogenesis and creation of new patho-
logic blood vessels within tumor environment. Thus VEGF
like many other factors is secondary to the hypoxia and as
much as current anti-VEGF approaches are helpful, even-
tually are over-compensated by progressive and persistent
hypoxia. Novel approaches to anti-cancer therapy by
modulating the oxygen tension in tumor environment and
thus directly targeting hypoxia are under development and
might bring new clinical applications. Aiming hypoxia in
cancer by specific hypoxamiRs can provide another layer
of controlling aberrant angiogenesis and blood vessel
normalization. Application of hypoxamiRs can also
directly lead to regulation of immune response in tumor
since HIF-1a has been demonstrated to regulate survival
and function of immune cells in the inflammatory micro-
environment such as cancer (Cramer and Johnson 2003;
Cramer et al. 2003; Palazon et al. 2012). Many cells
including macrophages express HIF-1a as an adaptation
mechanism in low oxygen environment. In T cells HIF-1anegatively regulates activation (Thiel et al. 2007). miRs
that target HIF-1a belong to miR-17-92 family and have
been shown to induce Th1 responses in lung cancer
(Taguchi et al. 2008). The term ‘‘hypoxamir’’ was initially
defined by Loscalzo et al. (Chan and Loscalzo 2010;
Loscalzo 2010). He classified hypoxamirs into three sub-
classes. The first group consists of HIF-dependent
hypoxamiRs, the second group involves miRs that are
induced by hypoxia and in turn affect HIF-1a expression,
and the last group comprises miRs that regulate HIF-1a in
hypoxia-independent manner. We added yet another group
called ‘‘miRs repressed by hypoxia’’ (Table 1) since sev-
eral new reports show that hypoxia not only induces
expression of diverse miRs but is also able to suppress their
expression. In rapidly growing field of miRs we realize that
our classification is only temporarily and many more newly
discovered molecules are on the horizon. Some of miRs
belong to two different categories since depending on the
conditions they might be induced or repressed by hypoxia.
The following are the examples of classical miRs whose
expression is related to hypoxia and/or HIF: cluster 17-92,
miR-20b, -31, -107, -199a, -210, -373, -424, and -519c.
miR-17-92 targets HIF-1a and is induced by oncogene
c-MYC (Taguchi et al. 2008). Similar to 17-92 cluster,
miR-519c suppresses expression of HIF-1a independently
on hypoxia (Cha et al. 2010). As found in MCF-7 breast
cancer cells, miR-20b reduces VEGF expression by tar-
geting HIF-1a and STAT3 (Cascio et al. 2010; Hua et al.
2006). miR-31 was identified as a pro-hypoxamiR inducing
expression of HIF-1a by affecting factor-inhibiting HIF
(Liu et al. 2010). miR-107 inhibits expression of HIF-1bsubunit of HIF-1 in response to p53 (Yamakuchi et al.
2010). miR-199a was found to decrease expression of HIF-
1a in cardiomyocytes (Rane et al. 2009). So far, the key
master of miRs that are steadily induced by hypoxia is
miR-210. Inhibition of miR-210 repressed the formation of
capillary-like structures in hypoxic environment and
reduced migration of cells in response to VEGF; endo-
thelial expression of miR-210 was induced by HIF-1a(Ivan et al. 2008). In hypoxia, miR-210 is a critical regu-
lator of endothelial cell survival, migration, and
differentiation (Fasanaro et al. 2008). Several target genes
have been identified for miR-210 such as ephrin A3,
RAD52, FGFRL2 and many others involved in angiogen-
esis, tumor propagation, cell cycle regulation, and stem cell
generation (Crosby et al. 2009b; Fasanaro et al. 2008;
290 Arch. Immunol. Ther. Exp. (2013) 61:285–299
123

Pulkkinen et al. 2008; Tsuchiya et al. 2011). In vitro studies
show its strong up-regulation by hypoxia in cultured HU-
VEC, and in vivo in limb and brain ischemia (Jeyaseelan
et al. 2008; Pulkkinen et al. 2008). Another hypoxamiR,
miR-424, was shown to be specifically expressed by
endothelial cell in response to hypoxia (Ghosh et al. 2010).
VEGF-a, VEGFR2, and FGFR1 were identified as targets
of miR-424 in endothelial cells (Chamorro-Jorganes et al.
2011). Decreased miR-424 expression and increased levels
of MEK1 or cyclin E1 in senile hemangioma caused
abnormal cell proliferation in the tumor and abnormal
angiogenesis (Nakashima et al. 2010). With collaboration
of miR-155, -222 and -503, -424 regulates monocyte dif-
ferentiation (Forrest et al. 2010; Rosa et al. 2007).
Overexpression of miR-424 leads to maturation of the
monoblastic U937 cells and expression of monocyte/mac-
rophage characteristics. Another set of oncomiRs: miR-210
and miR-373 whose expression is dependent upon HIF has
been identified (Crosby et al. 2009a, b). miR-373 as onco-
gene was found to play an important role in colon cancer
induction via regulating expression of RAB22A gene
(Tanaka et al. 2011). In another study, miR-373 increased
the expression of metalloproteinase 9 by activation of the
Ras/Raf/MEK/Erk signaling pathway and nuclear factor
kappa-light-chain-enhancer of activated B cells (NF-jb)
(Liu and Wilson 2012). Opposite role for miR-373 as a
tumor suppressor was documented in estrogen receptor
negative breast cancer by targeting NF-jb and TGF-b sig-
naling pathways (Keklikoglou et al. 2012). From the breast
and colon cancer studies, a group of miRs regulated by low
oxygen level including miR-210, 26 and 181 has been
identified (Kulshreshtha et al. 2008). It becomes increas-
ingly clear that miRs are implicated in the regulation of
various aspects of hypoxia including angiogenesis and
appear as targets for novel therapeutic approaches in cancer.
The discovery of novel miRs (angiomiRs) that are able to
lead to physioxia state, a proper tissue-dependent oxygen
partial pressure, can be of special interest.
miRs that Regulate the Immune Responses
Pre-existing condition for tumor initiation is persisting
inflammation that eventually leads to tissue damage,
release of metalloproteinases, pro-inflammatory cytokines,
and chemokines. Unresolved inflammation activates
endothelial cells and angiogenesis, cell proliferation,
motility, and engagement of stem cells. Myeloid cells are
first to respond to local inflammation. Immune cells display
a relative plasticity and their developmental fate strongly
depends on microenvironmental stimuli. Myeloid cells
have potential to differentiate into monocytes and macro-
phages. The latter can further polarize into M1/M2
macrophages or DC1/DC2 dendritic cells depending on the
cellular and signaling milieu. Early immune response in
cancer is characterized by influx of these innate immune
effectors that also include neutrophils and NK cells.
Complex networks created by several immune-competent
cells such as dendritic cells, B cells, cytotoxic CD8? T,
CD4? T-helper and NK cells in combination with cyto-
kines, chemokines, and other immune mediators are
required for effective immune reactions against cancer.
Ineffective immune response to cancer and promotion of
M2 and DC2 phenotypes as well involvement of regulatory
T cells favor tumor development and expansion. Specific
regulation of immune function by miRs plays a particularly
important role in cancer immunology. In physiological
conditions, the immune system is cautiously regulated,
coordinated, and stabilized by multiple miRs (Baltimore
et al. 2008). There is a selective tissue expression of some
immune-related miRs. For example, miR-142 is found in
all hematopoietic tissues, miR-223 is almost solely
expressed in bone marrow, and miR-181 is typically pro-
duced in brain, lung, and thymus (Chen et al. 2004). miRs
constitute an important connection between the adaptive
and innate immune responses (O’Connell et al. 2010).
Immune cells can be recognized by the specific set of
miRs. For example, hematopoietic cells are characterized
by the following miRs: miR-142, -144, -150, -155, and -
223 (Landgraf et al. 2007). miR-150, -155, -223 as well as
miR-17-92 cluster and miR-181 actively participate in
maturation and differentiation of myeloid and lymphoid
lineages (Tsitsiou and Lindsay 2009). Different immune
cell types can express the same miRs; however, their
magnitude of expression can be altered, for example miR-
342 is 10-times more abundant in T cells than B cells
(Merkerova et al. 2008). miR-155 plays a crucial role in
development, maintenance, and function of immune sys-
tem (Rodriguez et al. 2007). It is a constituent of innate
response to a variety of inflammatory mediators (Sonkoly
et al. 2008; Tili et al. 2007) and is essential for proper T
cell responses. miR-155 as oncomiR directly links immu-
nity with oncogenesis and is dependent on MAPK
signaling (O’Connell et al. 2007). Disregulation of miR-
155 in hematopietic cells leads in mice to malignancy,
myeloproliferative disorder, and in human to acute myeloid
leukemia (Costinean et al. 2006; O’Connell et al. 2008).
miR-155 is induced by inflammatory mediators during
early macrophage responses (O’Connell et al. 2007). Dis-
ruption of gene encoding miR-155 in mice caused deficient
antigen presentation by dendritic cells, breakdown in
mounting memory, and T-cell-specific immune responses
(Rodriguez et al. 2007). It is a key regulator of adaptive
immunity and fate of Th1 vs Th2 cells (Thai et al. 2007).
Malfunction of miR-155 skews immune response towards
Th2 phenotype and production of IL-4, IL-5 and IL-10
Arch. Immunol. Ther. Exp. (2013) 61:285–299 291
123

cytokines. miR-155 inhibits interferon (IFN)-c signaling in
CD4 cells and its over-expression promotes Th1 responses
(Banerjee et al. 2010). It also promotes the production of B
cell memory responses, high-affinity B cell clones that
undergo isotype-switching and produce high-affinity anti-
bodies, and its malfunction leads to plasma cell
transformation (Calame 2007). In contrast to miR-155,
-146a is a negative regulator of inflammation. miR-146a
knockout mice display autoimmunity and immunoprolif-
erative disorders (Boldin et al. 2011). miR-146a has been
found to be a NF-jb-dependent gene and plays an impor-
tant role in control of Toll-like receptor (TLR) and pro-
inflammatory cytokine signaling (Taganov et al. 2006).
NF-jb dysregulation in miR-146a deficient mice drives the
development of myeloid malignancies (Zhao et al. 2011).
Bone marrow-derived macrophages in miR-146a deficient
mice show increased cytokine production such as IL-1b,
IL-6 and TNF-a upon lipopolysaccharide stimulation
(Boldin et al. 2011). miR-146a is a negative regulator of
the IFN pathway targeting TLR and STAT-5 (Tang et al.
2009). miRs that target T cells include miR-181a and
members of miR-17-92 family. miR-181a controls T cells
positive and negative selection in thymus and its overex-
pression causes augmentation of sensitivity to peptide
antigens in mature T cells (Li et al. 2007). miR-17-92
expression in activated T cells promotes Th1 responses
such as increase production of IFN-c (Xiao et al. 2008).
Overexpression of this family renders T cells more sensi-
tive to T cell receptor antigenic stimulation (Xiao et al.
2008). miR-223 and miR-150 are selectively expressed by
B and T cells (Merkerova et al. 2008; Monticelli et al.
2005). miR-223 promotes differentiation of myeloid blast
cells in bone marrow (Fazi et al. 2005) and regulates
activation of granulocytes (Johnnidis et al. 2008). miR-222
and miR-339 are oncomiRs that directly influence tumor
immune responses by suppression of intracellular cell
adhesion molecule-1, thus decreasing the receptiveness of
cancer cells to cytotoxic T lymphocytes (Ueda et al. 2009).
Deletion of Dicer and Dgcr8 had an impact on the
expression level of natural-killer group 2, member D
Table 1 Overview of hypoxamiRs
Hypoxamirs induced by HIF Hypoxamirs that
affect HIF
miRNAs that affect HIF
independent of hypoxia
miRNAs repressed by
hypoxia
miR-10b (Haque et al. 2011) miR-20b (Cascio et al.
2010; Lei et al.
2009)
miR-17-92 cluster(Taguchi et al.
2008)
miR-17-3p miR-15b (Hua et al. 2006)
miR-155 (Bruning et al. 2011) miR-130 (Saito et al.
2011)
miR-17-5p miR-16 (Dejean et al. 2011;
Hua et al. 2006)
miR-210 (Chan and Loscalzo 2010; Crosby et al.
2009a, b; Fasanaro et al. 2008; Giannakakis et al.
2008)
miR-145 (Bussolati
et al. 2012; Xu et al.
2012)
miR-18a miR-20a (Hua et al. 2006)
miR-373 (Crosby et al. 2009a, b) miR-155 (Bruning
et al. 2011)
miR-19a miR-20b (Hua et al. 2006)
miR-199a (Kang et al.
2012)
miR-19b miR-34a (Du et al. 2012)
miR-424 (Ghosh et al.
2010)
miR-20a
(Kang
et al. 2012)
miR-101 (Cao et al. 2010)
miR-92a miR-135a (Gonsalves and
Kalra 2010)
miR-21 (Liu et al. 2011) miR-199a (Gonsalves and
Kalra 2010; Rane et al.
2009)
miR-22 (Yamakuchi et al. 2011) miR200b (Chan et al. 2011)
miR-31 (Liu et al. 2010) miR-378 (Fang et al. 2012)
miR-107 (Yamakuchi et al. 2010) miR-449a/b (Muth et al.
2010)
miR-130 (Kulshreshtha et al. 2007;
Saito et al. 2011)
miR-138 (Song et al. 2011)
miR-199a (Mizuno et al. 2012)
miR-519c (Cha et al. 2010)
The miRs classifed by Loscalzo (2010) are written in bold
292 Arch. Immunol. Ther. Exp. (2013) 61:285–299
123
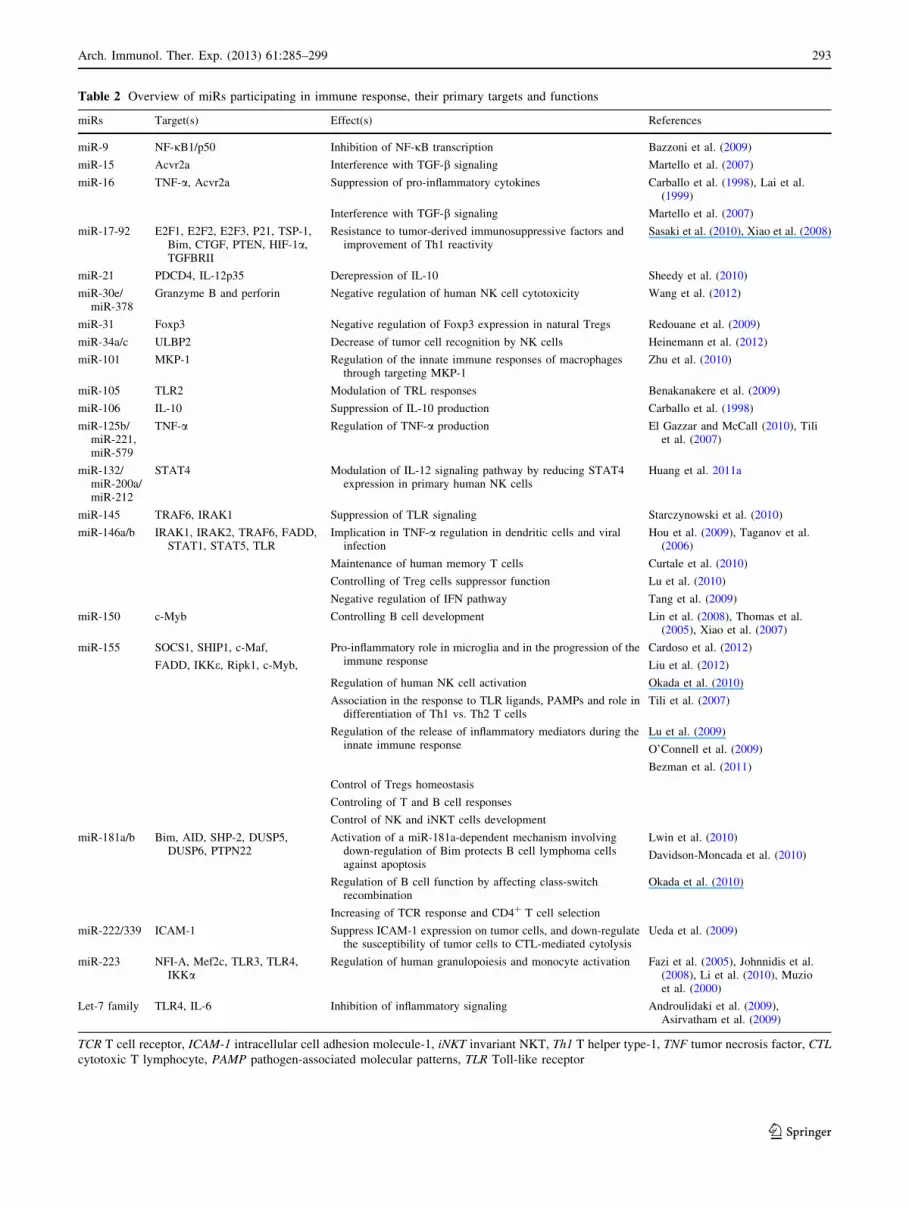
Table 2 Overview of miRs participating in immune response, their primary targets and functions
miRs Target(s) Effect(s) References
miR-9 NF-jB1/p50 Inhibition of NF-jB transcription Bazzoni et al. (2009)
miR-15 Acvr2a Interference with TGF-b signaling Martello et al. (2007)
miR-16 TNF-a, Acvr2a Suppression of pro-inflammatory cytokines Carballo et al. (1998), Lai et al.(1999)
Interference with TGF-b signaling Martello et al. (2007)
miR-17-92 E2F1, E2F2, E2F3, P21, TSP-1,Bim, CTGF, PTEN, HIF-1a,TGFBRII
Resistance to tumor-derived immunosuppressive factors andimprovement of Th1 reactivity
Sasaki et al. (2010), Xiao et al. (2008)
miR-21 PDCD4, IL-12p35 Derepression of IL-10 Sheedy et al. (2010)
miR-30e/miR-378
Granzyme B and perforin Negative regulation of human NK cell cytotoxicity Wang et al. (2012)
miR-31 Foxp3 Negative regulation of Foxp3 expression in natural Tregs Redouane et al. (2009)
miR-34a/c ULBP2 Decrease of tumor cell recognition by NK cells Heinemann et al. (2012)
miR-101 MKP-1 Regulation of the innate immune responses of macrophagesthrough targeting MKP-1
Zhu et al. (2010)
miR-105 TLR2 Modulation of TRL responses Benakanakere et al. (2009)
miR-106 IL-10 Suppression of IL-10 production Carballo et al. (1998)
miR-125b/miR-221,miR-579
TNF-a Regulation of TNF-a production El Gazzar and McCall (2010), Tiliet al. (2007)
miR-132/miR-200a/miR-212
STAT4 Modulation of IL-12 signaling pathway by reducing STAT4expression in primary human NK cells
Huang et al. 2011a
miR-145 TRAF6, IRAK1 Suppression of TLR signaling Starczynowski et al. (2010)
miR-146a/b IRAK1, IRAK2, TRAF6, FADD,STAT1, STAT5, TLR
Implication in TNF-a regulation in dendritic cells and viralinfection
Hou et al. (2009), Taganov et al.(2006)
Maintenance of human memory T cells Curtale et al. (2010)
Controlling of Treg cells suppressor function Lu et al. (2010)
Negative regulation of IFN pathway Tang et al. (2009)
miR-150 c-Myb Controlling B cell development Lin et al. (2008), Thomas et al.(2005), Xiao et al. (2007)
miR-155 SOCS1, SHIP1, c-Maf, Pro-inflammatory role in microglia and in the progression of theimmune response
Cardoso et al. (2012)
FADD, IKKe, Ripk1, c-Myb, Liu et al. (2012)
Regulation of human NK cell activation Okada et al. (2010)
Association in the response to TLR ligands, PAMPs and role indifferentiation of Th1 vs. Th2 T cells
Tili et al. (2007)
Regulation of the release of inflammatory mediators during theinnate immune response
Lu et al. (2009)
O’Connell et al. (2009)
Bezman et al. (2011)
Control of Tregs homeostasis
Controling of T and B cell responses
Control of NK and iNKT cells development
miR-181a/b Bim, AID, SHP-2, DUSP5,DUSP6, PTPN22
Activation of a miR-181a-dependent mechanism involvingdown-regulation of Bim protects B cell lymphoma cellsagainst apoptosis
Lwin et al. (2010)
Davidson-Moncada et al. (2010)
Regulation of B cell function by affecting class-switchrecombination
Okada et al. (2010)
Increasing of TCR response and CD4? T cell selection
miR-222/339 ICAM-1 Suppress ICAM-1 expression on tumor cells, and down-regulatethe susceptibility of tumor cells to CTL-mediated cytolysis
Ueda et al. (2009)
miR-223 NFI-A, Mef2c, TLR3, TLR4,IKKa
Regulation of human granulopoiesis and monocyte activation Fazi et al. (2005), Johnnidis et al.(2008), Li et al. (2010), Muzioet al. (2000)
Let-7 family TLR4, IL-6 Inhibition of inflammatory signaling Androulidaki et al. (2009),Asirvatham et al. (2009)
TCR T cell receptor, ICAM-1 intracellular cell adhesion molecule-1, iNKT invariant NKT, Th1 T helper type-1, TNF tumor necrosis factor, CTL
cytotoxic T lymphocyte, PAMP pathogen-associated molecular patterns, TLR Toll-like receptor
Arch. Immunol. Ther. Exp. (2013) 61:285–299 293
123

protein activating receptor on NK cells, and leads to
apoptosis of peripheral NK cells (Sullivan et al. 2012), key
players in direct lysis of cancer cells. Multiple miRs
coordinate the function of NK cells; however, miR-21
seems to be a key regulator of NK cell survival (Bezman
et al. 2010). Recently, the major role in NK development
and maturation has been attributed to miR-150, which was
found to differentially control the development of NK and
invariant NKT cell lineages by targeting c-Myb (Bezman
et al. 2011). Additionally, miR-378 and miR-30e were
proved to suppress human NK cell cytotoxicity (Wang
et al. 2012). To delineate the role of miRs in the biology of
Tregs that effectively inhibit antitumor responses and
interfere with antitumor therapies, mouse models with
depleted Dicer in Foxp3 Treg lineage were developed
(Liston et al. 2008; Zhou et al. 2008). Dicer deletion
resulted in lymphoproliferative autoimmune syndrome and
loss of Treg suppression activity in vivo (Cobb et al. 2006;
Zhou et al. 2008). There is a strong overlap in miR
expression between Tregs and activated T cells with the
exception of miR-223 and miR-146 (Cobb et al. 2006). A
few Treg-specific miRs are directly regulated by Foxp3;
one of them is miR-155 that is important in response of
Tregs to IL-2 by targeting SOCS1 and as an oncogene is
constitutively and abundantly expressed in Tregs (Lu and
Rudensky 2009; Lu et al. 2009; Marson et al. 2007; Zheng
et al. 2007). Table 2 provides the overview on miRs par-
ticipating in immune response that might relate to tumor-
suppressive or tumor-promoting activities for which the
specific targets are known.
Concluding Remarks
Continuous investigation of miRs as epigenetic agents can
reveal multiple mechanisms to avoid early events impor-
tant in aberrant angiogenesis and immunological
surveillance. Besides, as miRs are differentially expressed
in tumorogenesis, and are markedly tissue specific, they
can serve as new biomarkers. Finding similar miR profiles
regulating angiogenesis as well as developmental stages
and function of immune cells in tumor microenvironment
can offer another level of our understanding of the com-
plexity of tumor paradigm. It would be of particular
interest to recognize set of miRs that beneficially acts to
reverse aberrant angiogenesis and improve anti-cancer
immunity. One of such targets could be members of miR-
16-like family that specifically target VEGFs, as well as
their receptors and possess the ability to interfere with
TGF-b signaling. Moreover, one of them, miR-424, a
potent hypoxamiR that modulates HIF1-a, has been proven
to play a key role in immunity. Improving anti-tumor
immune responses by engineering miRs such as miR-17-
92, -155, -181a with properties for T cell-based immuno-
therapies represent another possible approaches. The vessel
normalization strategy based on application of miRs that
might have significant impact on the function of anti-tumor
immunity should efficiently eradicate aberrant immune
responses caused by tumor-associated macrophages,
TEMs, N2-effector neutrophils, and Tregs and increase
participation and cytotoxicity of NKs and CD8 T cells. As
much as this novel strategies bring hope we still need to be
aware of potential pitfalls such as probable distinct func-
tions of particular miRs in different cellular scenery or
interference and redundancy with physiological pathways.
Nevertheless, a lot of optimism exists with the future
clinical applications for miRs. Tissue-restricted expression
profile and abundance of circulating miRs in body fluids
such as blood, serum, plasma and saliva reflecting physi-
ological and pathological conditions make them good
candidates as non-invasive biomarkers. miR signature is
directly correlated to prediction of patients survival, out-
come for therapeutic responses, and can be used
successfully in molecular diagnostics for array of diseases
such as cancer and cardiovascular malfunctions. Such
approaches are being currently carried out. Moreover,
some miRs can act as oncogenes or tumor suppressors; thus
their activity modulation may regulate targeted cellular
behaviors in cancer treatments. Our continuous study on
miRs and their correlation with molecular and cellular
targets with disease phenotypes will illuminate new bio-
logical pathways and disease mechanisms. Gain- and loss-
of-function manipulations using oligonucleotide-based
inhibitors or miRs decoys/mimics open novel opportunities
for therapuetic interventions.
A new generation of anti-cancer drugs based on com-
binatory therapy built on set of specific miRs, blood vessel
normalization strategies, and conventional chemotherapy
that evidently will inhibit multiple cancer targets is eagerly
awaited.
References
Androulidaki A, Iliopoulos D, Arranz A et al (2009) The kinase Akt1
controls macrophage response to lipopolysaccharide by regulat-
ing microRNAs. Immunity 31:220–231
Asirvatham AJ, Magner WJ, Tomasi TB (2009) miRNA regulation of
cytokine genes. Cytokine 45:58–69
Baltimore D, Boldin MP, O’Connell RM et al (2008) MicroRNAs:
new regulators of immune cell development and function. Nat
Immunol 9:839–845
Banerjee A, Schambach F, DeJong CS et al (2010) Micro-RNA-155
inhibits IFN-gamma signaling in CD4? T cells. Eur J Immunol
40:225–231
Bazzoni F, Rossato M, Fabbri M et al (2009) Induction and regulatory
function of miR-9 in human monocytes and neutrophils exposed
to proinflammatory signals. Proc Natl Acad Sci USA
106:5282–5287
294 Arch. Immunol. Ther. Exp. (2013) 61:285–299
123

Benakanakere MR, Li Q, Eskan MA et al (2009) Modulation of TLR2
protein expression by miR-105 in human oral keratinocytes.
J Biol Chem 284:23107–23115
Bezman NA, Cedars E, Steiner DF et al (2010) Distinct requirements
of microRNAs in NK cell activation, survival, and function.
J Immunol 185:3835–3846
Bezman NA, Chakraborty T, Bender T et al (2011) miR-150 regulates the
development of NK and iNKT cells. J Exp Med 208:2717–2731
Bielawska-Pohl A, Blesson S, Benlalam H et al (2010) The anti-
angiogenic activity of IL-12 is increased in iNOS-/- mice and
involves NK cells. J Mol Med 88:775–784
Boldin MP, Taganov KD, Rao DS et al (2011) miR-146a is a
significant brake on autoimmunity, myeloproliferation, and
cancer in mice. J Exp Med 208:1189–1201
Bonauer A, Carmona G, Iwasaki M et al (2009) MicroRNA-92a
controls angiogenesis and functional recovery of ischemic
tissues in mice. Science 324:1710–1713
Bruning U, Cerone L, Neufeld Z et al (2011) MicroRNA-155
promotes resolution of hypoxia-inducible factor 1alpha activity
during prolonged hypoxia. Mol Cell Biol 31:4087–4096
Bussolati B, Moggio A, Collino F et al (2012) Hypoxia modulates the
undifferentiated phenotype of human renal inner medullary
CD133? progenitors through Oct4/miR-145 balance. Am J
Physiol Renal Physiol 302:F116–F128
Calame K (2007) MicroRNA-155 function in B cells. Immunity
27:825–827
Cao P, Deng Z, Wan M et al (2010) MicroRNA-101 negatively
regulates Ezh2 and its expression is modulated by androgen
receptor and HIF-1alpha/HIF-1beta. Mol Cancer 9:108
Caporali A, Emanueli C (2011) MicroRNA regulation in angiogen-
esis. Vascul Pharmacol 55:79–86
Carballo E, Lai WS, Blackshear PJ (1998) Feedback inhibition of
macrophage tumor necrosis factor-alpha production by triste-
traprolin. Science 281:1001–1005
Cardoso AL, Guedes JR, Pereira de Almeida L et al (2012) miR-155
modulates microglia-mediated immune response by down-reg-
ulating SOCS-1 and promoting cytokine and nitric oxide
production. Immunology 135:73–88
Carmeliet P, Jain RK (2011a) Molecular mechanisms and clinical
applications of angiogenesis. Nature 473:298–307
Carmeliet P, Jain RK (2011b) Principles and mechanisms of vessel
normalization for cancer and other angiogenic diseases. Nat Rev
Drug Discov 10:417–427
Carreau A, Kieda C, Grillon C (2011) Nitric oxide modulates the
expression of endothelial cell adhesion molecules involved in
angiogenesis and leukocyte recruitment. Exp Cell Res 317:
29–41
Cascio S, D’Andrea A, Ferla R et al (2010) miR-20b modulates
VEGF expression by targeting HIF-1 alpha and STAT3 in MCF-
7 breast cancer cells. J Cell Physiol 224:242–249
Cha ST, Chen PS, Johansson G et al (2010) MicroRNA-519c
suppresses hypoxia-inducible factor-1alpha expression and
tumor angiogenesis. Cancer Res 70:2675–2685
Chamorro-Jorganes A, Araldi E, Penalva LO et al (2011) MicroRNA-
16 and microRNA-424 regulate cell-autonomous angiogenic
functions in endothelial cells via targeting vascular endothelial
growth factor receptor-2 and fibroblast growth factor receptor-1.
Arterioscler Thromb Vasc Biol 31:2595–2606
Chan SY, Loscalzo J (2010) MicroRNA-210: a unique and pleiotropic
hypoxamir. Cell Cycle 9:1072–1083
Chan YC, Khanna S, Roy S et al (2011) miR-200b targets Ets-1 and is
down-regulated by hypoxia to induce angiogenic response of
endothelial cells. J Biol Chem 286:2047–2056
Chen Y, Gorski DH (2008) Regulation of angiogenesis through a
microRNA (miR-130a) that down-regulates antiangiogenic
homeobox genes GAX and HOXA5. Blood 111:1217–1226
Chen CZ, Li L, Lodish HF et al (2004) MicroRNAs modulate
hematopoietic lineage differentiation. Science 303:83–86
Cifuentes D, Xue H, Taylor DW et al (2010) A novel miRNA
processing pathway independent of Dicer requires Argonaute2
catalytic activity. Science 328:1694–1698
Cobb BS, Hertweck A, Smith J et al (2006) A role for Dicer in
immune regulation. J Exp Med 203:2519–2527
Cobleigh MA, Langmuir VK, Sledge GW et al (2003) A phase I/II
dose-escalation trial of bevacizumab in previously treated
metastatic breast cancer. Semin Oncol 30:117–124
Coffelt SB, Tal AO, Scholz A et al (2010) Angiopoietin-2 regulates
gene expression in TIE2-expressing monocytes and augments
their inherent proangiogenic functions. Cancer Res 70:
5270–5280
Costinean S, Zanesi N, Pekarsky Y et al (2006) Pre-B cell
proliferation and lymphoblastic leukemia/high-grade lymphoma
in E(mu)-miR155 transgenic mice. Proc Natl Acad Sci USA
103:7024–7029
Cramer T, Johnson RS (2003) A novel role for the hypoxia inducible
transcription factor HIF-1alpha: critical regulation of inflamma-
tory cell function. Cell Cycle 2:192–193
Cramer T, Yamanishi Y, Clausen BE et al (2003) HIF-1alpha is
essential for myeloid cell-mediated inflammation. Cell 112:
645–657
Crawford M, Brawner E, Batte K et al (2008) MicroRNA-126 inhibits
invasion in non-small cell lung carcinoma cell lines. Biochem
Biophys Res Commun 373:607–612
Crosby ME, Devlin CM, Glazer PM et al (2009a) Emerging roles of
microRNAs in the molecular responses to hypoxia. Curr Pharm
Des 15:3861–3866
Crosby ME, Kulshreshtha R, Ivan M et al (2009b) MicroRNA
regulation of DNA repair gene expression in hypoxic stress.
Cancer Res 69:1221–1229
Curtale G, Citarella F, Carissimi C et al (2010) An emerging player in
the adaptive immune response: microRNA-146a is a modulator
of IL-2 expression and activation-induced cell death in T
lymphocytes. Blood 115:265–273
Davidson-Moncada J, Papavasiliou FN, Tam W (2010) MicroRNAs
of the immune system: roles in inflammation and cancer. Ann N
Y Acad Sci 1183:183–194
Dejean E, Renalier MH, Foisseau M et al (2011) Hypoxia-microR-
NA-16 downregulation induces VEGF expression in anaplastic
lymphoma kinase (ALK)-positive anaplastic large-cell lympho-
mas. Leukemia 25:1882–1890
Desgrosellier JS, Cheresh DA (2010) Integrins in cancer: biological
implications and therapeutic opportunities. Nat Rev Cancer
10:9–22
Dews M, Homayouni A, Yu D et al (2006) Augmentation of tumor
angiogenesis by a Myc-activated microRNA cluster. Nat Genet
38:1060–1065
Doebele C, Bonauer A, Fischer A et al (2010) Members of the
microRNA-17-92 cluster exhibit a cell-intrinsic antiangiogenic
function in endothelial cells. Blood 115:4944–4950
Du R, Sun W, Xia L et al (2012) Hypoxia-induced down-regulation of
microRNA-34a promotes EMT by targeting the Notch signaling
pathway in tubular epithelial cells. PLoS ONE 7:e30771
El Gazzar M, McCall CE (2010) MicroRNAs distinguish translational
from transcriptional silencing during endotoxin tolerance. J Biol
Chem 285:20940–20951
Falcon BL, Hashizume H, Koumoutsakos P et al (2009) Contrasting
actions of selective inhibitors of angiopoietin-1 and angiopoie-
tin-2 on the normalization of tumor blood vessels. Am J Pathol
175:2159–2170
Fang J, Song XW, Tian J et al (2012) Overexpression of microRNA-
378 attenuates ischemia-induced apoptosis by inhibiting caspase-
3 expression in cardiac myocytes. Apoptosis 17:410–423
Arch. Immunol. Ther. Exp. (2013) 61:285–299 295
123

Fasanaro P, D’Alessandra Y, Di Stefano V et al (2008) MicroRNA-
210 modulates endothelial cell response to hypoxia and inhibits
the receptor tyrosine kinase ligand Ephrin-A3. J Biol Chem
283:15878–15883
Fazi F, Rosa A, Fatica A et al (2005) A minicircuitry comprised of
microRNA-223 and transcription factors NFI-A and C/EBPalpha
regulates human granulopoiesis. Cell 123:819–831
Fischer C, Jonckx B, Mazzone M et al (2007) Anti-PlGF inhibits
growth of VEGF(R)-inhibitor-resistant tumors without affecting
healthy vessels. Cell 131:463–475
Fish JE, Santoro MM, Morton SU et al (2008) miR-126 regulates
angiogenic signaling and vascular integrity. Dev Cell
15:272–284
Forrest AR, Kanamori-Katayama M, Tomaru Y et al (2010) Induction
of microRNAs, mir-155, mir-222, mir-424 and mir-503, pro-
motes monocytic differentiation through combinatorial
regulation. Leukemia 24:460–466
Fukumura D, Duda DG, Munn LL et al (2010) Tumor microvascu-
lature and microenvironment: novel insights through intravital
imaging in pre-clinical models. Microcirculation 17:206–225
Gabrilovich DI, Chen HL, Girgis KR et al (1996) Production of
vascular endothelial growth factor by human tumors inhibits the
functional maturation of dendritic cells. Nat Med 2:1096–1103
Ganss R, Arnold B, Hammerling GJ (2004) Mini-review: overcoming
tumor-intrinsic resistance to immune effector function. Eur J
Immunol 34:2635–2641
Ghosh G, Subramanian IV, Adhikari N et al (2010) Hypoxia-induced
microRNA-424 expression in human endothelial cells regulates
HIF-alpha isoforms and promotes angiogenesis. J Clin Invest
120:4141–4154
Giannakakis A, Sandaltzopoulos R, Greshock J et al (2008) miR-210
links hypoxia with cell cycle regulation and is deleted in human
epithelial ovarian cancer. Cancer Biol Ther 7:255–264
Giantonio BJ, Catalano PJ, Meropol NJ et al (2007) Bevacizumab in
combination with oxaliplatin, fluorouracil, and leucovorin
(FOLFOX4) for previously treated metastatic colorectal cancer:
results from the Eastern Cooperative Oncology Group Study
E3200. J Clin Oncol 25:1539–1544
Goel S, Duda DG, Xu L et al (2011) Normalization of the vasculature
for treatment of cancer and other diseases. Physiol Rev 91:
1071–1121
Goel S, Wong AH, Jain RK (2012) Vascular normalization as a
therapeutic strategy for malignant and nonmalignant disease.
Cold Spring Harb Perspect Med 2:a006486
Gonsalves CS, Kalra VK (2010) Hypoxia-mediated expression of
5-lipoxygenase-activating protein involves HIF-1alpha and NF-
kappaB and microRNAs 135a and 199a–5p. J Immunol
184:3878–3888
Gordan JD, Simon MC (2007) Hypoxia-inducible factors: central
regulators of the tumor phenotype. Curr Opin Genet Dev 17:
71–77
Griffioen AW, Damen CA, Blijham GH et al (1996) Tumor
angiogenesis is accompanied by a decreased inflammatory
response of tumor-associated endothelium. Blood 88:667–673
Gruber M, Simon MC (2006) Hypoxia-inducible factors, hypoxia, and
tumor angiogenesis. Curr Opin Hematol 13:169–174
Guo C, Sah JF, Beard L et al (2008) The noncoding RNA, miR-126,
suppresses the growth of neoplastic cells by targeting phospha-
tidylinositol 3-kinase signaling and is frequently lost in colon
cancers. Genes Chromosomes Cancer 47:939–946
Hamzah J, Jugold M, Kiessling F et al (2008) Vascular normalization
in Rgs5-deficient tumours promotes immune destruction. Nature
453:410–414
Haque I, Banerjee S, Mehta S et al (2011) Cysteine-rich 61-connec-
tive tissue growth factor-nephroblastoma-overexpressed 5
(CCN5)/Wnt-1-induced signaling protein-2 (WISP-2) regulates
microRNA-10b via hypoxia-inducible factor-1alpha-TWIST
signaling networks in human breast cancer cells. J Biol Chem
286:43475–43485
Harris TA, Yamakuchi M, Ferlito M et al (2008) MicroRNA-126
regulates endothelial expression of vascular cell adhesion
molecule 1. Proc Natl Acad Sci USA 105:1516–1521
Hartmann D, Thum T (2011) MicroRNAs and vascular (dys)function.
Vascul Pharmacol 55:92–105
He L, Hannon GJ (2004) MicroRNAs: small RNA’s with a big role in
gene regulation. Nat Rev Genet 5:522–531
He L, Thomson JM, Hemann MT et al (2005) A microRNA
polycistron as a potential human oncogene. Nature 9:828–833
Heinemann A, Zhao F, Pechlivanis S et al (2012) Tumor suppressive
microRNAs miR-34a/c control cancer cell expression of ULBP2,
a stress-induced ligand of the natural killer cell receptor
NKG2D. Cancer Res 72:460–471
Hergenreider E, Heydt S, Treguer K et al (2012) Atheroprotective
communication between endothelial cells and smooth muscle
cells through miRNAs. Nat Cell Biol 14:249–256
Hockel M, Vaupel P (2001) Tumor hypoxia: definitions and current
clinical, biologic, and molecular aspects. J Natl Cancer Inst
93:266–276
Hockel M, Schlenger K, Hockel S et al (1999) Hypoxic cervical
cancers with low apoptotic index are highly aggressive. Cancer
Res 59:4525–4528
Hou J, Wang P, Lin L et al (2009) MicroRNA-146a feedback inhibits
RIG-I-dependent Type I IFN production in macrophages by
targeting TRAF6, IRAK1, and IRAK2. J Immunol 183:
2150–2158
Hua Z, Lv Q, Ye W et al (2006) MiRNA-directed regulation of VEGF
and other angiogenic factors under hypoxia. PLoS ONE 1:e116
Huang X, Wong MK, Yi H et al (2002) Combined therapy of local
and metastatic 4T1 breast tumor in mice using SU6668, an
inhibitor of angiogenic receptor tyrosine kinases, and the
immunostimulator B7.2-IgG fusion protein. Cancer Res
62:5727–5735
Huang H, Bhat A, Woodnutt G et al (2010) Targeting the ANGPT-
TIE2 pathway in malignancy. Nat Rev Cancer 10:575–585
Huang Y, Lei Y, Zhang H et al (2011a) MicroRNA regulation of
STAT4 protein expression: rapid and sensitive modulation of
IL-12 signaling in human natural killer cells. Blood 118:
6793–6802
Huang Y, Snuderl M, Jain RK (2011b) Polarization of tumor-
associated macrophages: a novel strategy for vascular normal-
ization and antitumor immunity. Cancer Cell 19:1–2
Ivan M, Harris AL, Martelli F et al (2008) Hypoxia response and
microRNAs: no longer two separate worlds. J Cell Mol Med
12:1426–1431
Jain RK (2001) Normalizing tumor vasculature with anti-angiogenic
therapy: a new paradigm for combination therapy. Nat Med
7:987–989
Jain RK (2003) Molecular regulation of vessel maturation. Nat Med
9:685–693
Jain RK (2005) Normalization of tumor vasculature: an emerging
concept in antiangiogenic therapy. Science 307:58–62
Jain RK (2009) A new target for tumor therapy. N Engl J Med
360:2669–2671
Jain RK, Duda DG, Clark JW et al (2006) Lessons from phase III
clinical trials on anti-VEGF therapy for cancer. Nat Clin Pract
Oncol 3:24–40
Jeyaseelan K, Lim KY, Armugam A (2008) MicroRNA expression in
the blood and brain of rats subjected to transient focal ischemia
by middle cerebral artery occlusion. Stroke 39:959–966
Johnnidis JB, Harris MH, Wheeler RT et al (2008) Regulation of
progenitor cell proliferation and granulocyte function by micr-
oRNA-223. Nature 451:1125–1129
296 Arch. Immunol. Ther. Exp. (2013) 61:285–299
123

Jordan BF, Sonveaux P (2012) Targeting tumor perfusion and
oxygenation to improve the outcome of anticancer therapy.
Frontiers Pharmacol 3:94
Kahai S, Lee SC, Lee DY et al (2009) MicroRNA miR-378 regulates
nephronectin expression modulating osteoblast differentiation by
targeting GalNT-7. PLoS ONE 4:e7535
Kang SG, Lee WH, Lee YH et al (2012) Hypoxia-inducible factor-
1alpha inhibition by a pyrrolopyrazine metabolite of oltipraz as a
consequence of microRNAs 199a–5p and 20a induction. Carci-
nogenesis 33:661–669
Karaa ZS, Iacovoni JS, Bastide A et al (2009) The VEGF IRESes are
differentially susceptible to translation inhibition by miR-16.
RNA 15:249–254
Keklikoglou I, Koerner C, Schmidt C et al (2012) MicroRNA-520/
373 family functions as a tumor suppressor in estrogen receptor
negative breast cancer by targeting NF-kappaB and TGF-beta
signaling pathways. Oncogene 31:4150–4163
Kieda C, Greferath R, Crola da Silva C et al (2006) Suppression of
hypoxia-induced HIF-1alpha and of angiogenesis in endothelial
cells by myo-inositol trispyrophosphate-treated erythrocytes.
Proc Natl Acad Sci USA 103:15576–15581
Kozomara A, Griffiths-Jones S (2011) miRBase: integrating microR-
NA annotation and deep-sequencing data. Nucleic Acids Res
39:D152–D157
Kuehbacher A, Urbich C, Zeiher AM et al (2007) Role of Dicer and
Drosha for endothelial microRNA expression and angiogenesis.
Circ Res 101:59–68
Kuhnert F, Mancuso MR, Hampton J et al (2008) Attribution of
vascular phenotypes of the murine Egfl7 locus to the microRNA
miR-126. Development 135:3989–3993
Kulshreshtha R, Ferracin M, Wojcik SE et al (2007) A microRNA
signature of hypoxia. Mol Cell Biol 27:1859–1867
Kulshreshtha R, Davuluri RV, Calin GA et al (2008) A microRNA
component of the hypoxic response. Cell Death Differ 15:
667–671
Lai WS, Carballo E, Strum JR et al (1999) Evidence that tristetrapr-
olin binds to AU-rich elements and promotes the deadenylation
and destabilization of tumor necrosis factor alpha mRNA. Mol
Cell Biol 19:4311–4323
Landgraf P, Rusu M, Sheridan R et al (2007) A mammalian
microRNA expression atlas based on small RNA library
sequencing. Cell 129:1401–1414
Larsson E, Fredlund Fuchs P, Heldin J et al (2009) Discovery of
microvascular miRNAs using public gene expression data: miR-
145 is expressed in pericytes and is a regulator of Fli1. Genome
Med 1:108
le Sage C, Nagel R, Egan DA et al (2007) Regulation of the
p27(Kip1) tumor suppressor by miR-221 and miR-222 promotes
cancer cell proliferation. EMBO J 26:3699–3708
Lee DY, Deng Z, Wang CH et al (2007) MicroRNA-378 promotes
cell survival, tumor growth, and angiogenesis by targeting SuFu
and Fus-1 expression. Proc Natl Acad Sci USA 104:
20350–20355
Lei Z, Li B, Yang Z et al (2009) Regulation of HIF-1alpha and VEGF
by miR-20b tunes tumor cells to adapt to the alteration of oxygen
concentration. PLoS ONE 4:e7629
Leite de Oliveira R, Deschoemaeker S, Henze AT et al (2012) Gene-
targeting of phd2 improves tumor response to chemotherapy and
prevents side-toxicity. Cancer Cell 22:263–277
Li B, Lalani AS, Harding TC et al (2006) Vascular endothelial growth
factor blockade reduces intratumoral regulatory T cells and
enhances the efficacy of a GM-CSF-secreting cancer immuno-
therapy. Clin Cancer Res 12:6808–6816
Li QJ, Chau J, Ebert PJ et al (2007) miR-181a is an intrinsic
modulator of T cell sensitivity and selection. Cell 129:147–161
Li Y, Song YH, Li F et al (2009) MicroRNA-221 regulates high
glucose-induced endothelial dysfunction. Biochem Biophys Res
Commun 381:81–83
Li T, Morgan MJ, Choksi S et al (2010) MicroRNAs modulate the
noncanonical transcription factor NF-kappaB pathway by regu-
lating expression of the kinase IKKalpha during macrophage
differentiation. Nat Immunol 11:799–805
Lin YC, Kuo MW, Yu J et al (2008) c-Myb is an evolutionary
conserved miR-150 target and miR-150/c-Myb interaction is
important for embryonic development. Mol Biol Evol 25:
2189–2198
Liston A, Lu LF, O’Carroll D et al (2008) Dicer-dependent
microRNA pathway safeguards regulatory T cell function.
J Exp Med 205:1993–2004
Liu P, Wilson MJ (2012) miR-520c and miR-373 upregulate MMP9
expression by targeting mTOR and SIRT1, and activate the Ras/
Raf/MEK/Erk signaling pathway and NF-kappaB factor in
human fibrosarcoma cells. J Cell Physiol 227:867–876
Liu CJ, Tsai MM, Hung PS et al (2010) miR-31 ablates expression of
the HIF regulatory factor FIH to activate the HIF pathway in
head and neck carcinoma. Cancer Res 70:1635–1644
Liu LZ, Li C, Chen Q et al (2011) MiR-21 induced angiogenesis
through AKT and ERK activation and HIF-1alpha expression.
PLoS ONE 6:e19139
Liu X, Wang Y, Sun Q et al (2012) Identification of microRNA
transcriptome involved in human natural killer cell activation.
Immunol Lett 143:208–217
Loscalzo J (2010) The cellular response to hypoxia: tuning the system
with microRNAs. J Clin Invest 120:3815–3817
Lu LF, Rudensky A (2009) Molecular orchestration of differentiation
and function of regulatory T cells. Genes Dev 23:1270–1282
Lu LF, Thai TH, Calado DP et al (2009) Foxp3-dependent
microRNA155 confers competitive fitness to regulatory T cells
by targeting SOCS1 protein. Immunity 30:80–91
Lu LF, Boldin MP, Chaudhry A et al (2010) Function of miR-146a in
controlling Treg cell-mediated regulation of Th1 responses. Cell
142:914–929
Lwin T, Lin J, Choi YS et al (2010) Follicular dendritic cell-
dependent drug resistance of non-Hodgkin lymphoma involves
cell adhesion-mediated Bim down-regulation through induction
of microRNA-181a. Blood 116:5228–5236
Manning EA, Ullman JG, Leatherman JM et al (2007) A vascular
endothelial growth factor receptor-2 inhibitor enhances antitu-
mor immunity through an immune-based mechanism. Clin
Cancer Res 13:3951–3959
Marson A, Kretschmer K, Frampton GM et al (2007) Foxp3
occupancy and regulation of key target genes during T-cell
stimulation. Nature 445:931–935
Martello G, Zacchigna L, Inui M et al (2007) MicroRNA control of
Nodal signalling. Nature 449:183–188
Matejuk A, Leng Q, Chou ST et al (2011) Vaccines targeting the
neovasculature of tumors. Vasc Cell 3:7
Matsui J, Wakabayashi T, Asada M et al (2004) Stem cell factor/c-kit
signaling promotes the survival, migration, and capillary tube
formation of human umbilical vein endothelial cells. J Biol
Chem 279:18600–18607
Mazzieri R, Pucci F, Moi D et al (2011) Targeting the ANG2/TIE2
axis inhibits tumor growth and metastasis by impairing angio-
genesis and disabling rebounds of proangiogenic myeloid cells.
Cancer Cell 19:512–526
Mazzone M, Dettori D, Leite de Oliveira R et al (2009) Heterozygous
deficiency of PHD2 restores tumor oxygenation and inhibits
metastasis via endothelial normalization. Cell 136:839–851
Melder RJ, Koenig GC, Witwer BP et al (1996) During angiogenesis,
vascular endothelial growth factor and basic fibroblast growth
Arch. Immunol. Ther. Exp. (2013) 61:285–299 297
123

factor regulate natural killer cell adhesion to tumor endothelium.
Nat Med 2:992–997
Mendell JT (2008) miRiad roles for the miR-17-92 cluster in
development and disease. Cell 133:217–222
Merkerova M, Belickova M, Bruchova H (2008) Differential
expression of microRNAs in hematopoietic cell lineages. Eur J
Haematol 81:304–310
Minami Y, Satoh M, Maesawa C et al (2009) Effect of atorvastatin on
microRNA 221/222 expression in endothelial progenitor cells
obtained from patients with coronary artery disease. Eur J Clin
Invest 39:359–367
Mizuno S, Bogaard HJ, Gomez-Arroyo J et al (2012) MicroRNA-
199a-5p is associated with hypoxia inducible factor-1alpha
expression in the lung from COPD patients. Chest 142:663–672
Monticelli S, Ansel KM, Xiao C et al (2005) MicroRNA profiling of
the murine hematopoietic system. Genome Biol 6:R71
Muth M, Theophile K, Hussein K et al (2010) Hypoxia-induced
down-regulation of microRNA-449a/b impairs control over
targeted SERPINE1 (PAI-1) mRNA - a mechanism involved in
SERPINE1 (PAI-1) overexpression. J Transl Med 8:33
Muzio M, Bosisio D, Polentarutti N et al (2000) Differential
expression and regulation of toll-like receptors (TLR) in human
leukocytes: selective expression of TLR3 in dendritic cells.
J Immunol 164:5998–6004
Nagy JA, Chang SH, Dvorak AM et al (2009) Why are tumour blood
vessels abnormal and why is it important to know? Br J Cancer
100:865–869
Nakashima T, Jinnin M, Etoh T et al (2010) Down-regulation of mir-
424 contributes to the abnormal angiogenesis via MEK1 and
cyclin E1 in senile hemangioma: its implications to therapy.
PLoS ONE 5:e14334
Nicoli S, Standley C, Walker P et al (2010) MicroRNA-mediated
integration of haemodynamics and Vegf signalling during
angiogenesis. Nature 464:1196–1200
Nicoli S, Knyphausen CP, Zhu LJ et al (2012) miR-221 is required for
endothelial tip cell behaviors during vascular development. Dev
Cell 22:418–429
Noman MZ, Janji B, Kaminska B et al (2011) Blocking hypoxia-
induced autophagy in tumors restores cytotoxic T-cell activity
and promotes regression. Cancer Res 71:5976–5986
O’Connell RM, Taganov KD, Boldin MP et al (2007) MicroRNA-155
is induced during the macrophage inflammatory response. Proc
Natl Acad Sci USA 104:1604–1609
O’Connell RM, Rao DS, Chaudhuri AA et al (2008) Sustained
expression of microRNA-155 in hematopoietic stem cells causes
a myeloproliferative disorder. J Exp Med 205:585–594
O’Connell RM, Chaudhuri AA, Rao DS et al (2009) Inositol
phosphatase SHIP1 is a primary target of miR-155. Proc Natl
Acad Sci USA 106:7113–7118
O’Connell RM, Rao DS, Chaudhuri AA et al (2010) Physiological
and pathological roles for microRNAs in the immune system.
Nat Rev Immunol 10:111–122
Okada H, Kohanbash G, Lotze MT (2010) MicroRNAs in immune
regulation–opportunities for cancer immunotherapy. Int J Bio-
chem Cell Biol 42:1256–1261
Olive V, Jiang I, He L (2010) mir-17-92, a cluster of miRNAs in the
midst of the cancer network. Int J Biochem Cell Biol 42:
1348–1354
Osada T, Chong G, Tansik R et al (2008) The effect of anti-VEGF
therapy on immature myeloid cell and dendritic cells in cancer
patients. Cancer Immunol Immunother 57:1115–1124
Palazon A, Aragones J, Morales-Kastresana A et al (2012) Molecular
pathways: hypoxia response in immune cells fighting or
promoting cancer. Clin Cancer Res 18:1207–1213
Poliseno L, Tuccoli A, Mariani L et al (2006) MicroRNAs modulate
the angiogenic properties of HUVECs. Blood 108:3068–3071
Primo L, Seano G, Roca C et al (2010) Increased expression of alpha6
integrin in endothelial cells unveils a proangiogenic role for
basement membrane. Cancer Res 70:5759–5769
Pulkkinen K, Malm T, Turunen M et al (2008) Hypoxia induces
microRNA miR-210 in vitro and in vivo ephrin-A3 and neuronal
pentraxin 1 are potentially regulated by miR-210. FEBS Lett
582:2397–2401
Rane S, He M, Sayed D et al (2009) Downregulation of miR-199a
derepresses hypoxia-inducible factor-1alpha and Sirtuin 1 and
recapitulates hypoxia preconditioning in cardiac myocytes. Circ
Res 104:879–886
Redouane R, Hussein FK, Nabil EZ et al (2009) Human natural Treg
microRNA signature: Role of microRNA-31 and microRNA-21
in FOXp3 expression. Eur J Immunol 39:1–11
Relf M, LeJeune S, Scott PA, Fox S et al (1997) Expression of the
angiogenic factors vascular endothelial cell growth factor, acidic
and basic fibroblast growth factor, tumor growth factor beta-1,
platelet-derived endothelial cell growth factor, placenta growth
factor, and pleiotrophin in human primary breast cancer and its
relation to angiogenesis. Cancer Res 57:963–969
Rocha S (2007) Gene regulation under low oxygen: holding your
breath for transcription. Trends Biochem Sci 32:389–397
Rodriguez A, Vigorito E, Clare S et al (2007) Requirement of bic/
microRNA-155 for normal immune function. Science 316:608–611
Rolny C, Mazzone M, Tugues S et al (2011) HRG inhibits tumor
growth and metastasis by inducing macrophage polarization and
vessel normalization through downregulation of PlGF. Cancer
Cell 19:31–44
Rosa A, Ballarino M, Sorrentino A et al (2007) The interplay between
the master transcription factor PU.1 and miR-424 regulates
human monocyte/macrophage differentiation. Proc Natl Acad
Sci USA 104:19849–19854
Saito K, Kondo E, Matsushita M (2011) MicroRNA 130 family
regulates the hypoxia response signal through the P-body protein
DDX6. Nucleic Acids Res 39:6086–6099
Saltz LB, Clarke S, Diaz-Rubio E et al (2008) Bevacizumab in
combination with oxaliplatin-based chemotherapy as first-line
therapy in metastatic colorectal cancer: a randomized phase III
study. J Clin Oncol 26:2013–2019
Sasaki K, Kohanbash G, Hoji A et al (2010) miR-17-92 expression in
differentiated T cells - implications for cancer immunotherapy.
J Transl Med 8:17
Sheedy FJ, Palsson-McDermott E, Hennessy EJ et al (2010) Negative
regulation of TLR4 via targeting of the proinflammatory tumor
suppressor PDCD4 by the microRNA miR-21. Nat Immunol
11:141–147
Shilo S, Roy S, Khanna S et al (2008) Evidence for the involvement
of miRNA in redox regulated angiogenic response of human
microvascular endothelial cells. Arterioscler Thromb Vasc Biol
28:471–477
Shojaei F, Wu X, Malik AK et al (2007) Tumor refractoriness to anti-
VEGF treatment is mediated by CD11b? Gr1? myeloid cells.
Nat Biotechnol 25:911–920
Shrimali RK, Yu Z, Theoret MR et al (2010) Antiangiogenic agents
can increase lymphocyte infiltration into tumor and enhance the
effectiveness of adoptive immunotherapy of cancer. Cancer Res
70:6171–6180
Sihn G, Walter T, Klein JC et al (2007) Anti-angiogenic properties of
myo-inositol trispyrophosphate in ovo and growth reduction of
implanted glioma. FEBS Lett 581:962–966
Skuli N, Monferran S, Delmas C et al (2009) Alphavbeta3/alphavb-
eta5 integrins-FAK-RhoB: a novel pathway for hypoxia
regulation in glioblastoma. Cancer Res 69:3308–3316
Song T, Zhang X, Wang C et al (2011) MiR-138 suppresses expression of
hypoxia-inducible factor 1alpha (HIF-1alpha) in clear cell renal cell
carcinoma 786-O cells. Asian Pac J Cancer Prev 12:1307–1311
298 Arch. Immunol. Ther. Exp. (2013) 61:285–299
123

Sonkoly E, Stahle M, Pivarcsi A (2008) MicroRNAs and immunity:
novel players in the regulation of normal immune function and
inflammation. Semin Cancer Biol 18:131–140
Starczynowski DT, Kuchenbauer F, Argiropoulos B et al (2010)
Identification of miR-145 and miR-146a as mediators of the 5q-
syndrome phenotype. Nat Med 16:49–58
Stockmann C, Doedens A, Weidemann A et al (2008) Deletion of
vascular endothelial growth factor in myeloid cells accelerates
tumorigenesis. Nature 456:814–818
Suarez Y, Fernandez-Hernando C, Pober JS et al (2007) Dicer
dependent microRNAs regulate gene expression and functions in
human endothelial cells. Circ Res 100:1164–1173
Suarez Y, Fernandez-Hernando C, Yu J et al (2008) Dicer-dependent
endothelial microRNAs are necessary for postnatal angiogenesis.
Proc Natl Acad Sci USA 105:14082–14087
Sullivan RP, Leong JW, Schneider SE et al (2012) MicroRNA-
deficient NK cells exhibit decreased survival but enhanced
function. J Immunol 188:3019–3030
Taganov KD, Boldin MP, Chang KJ et al (2006) NF-kappaB-
dependent induction of microRNA miR-146, an inhibitor
targeted to signaling proteins of innate immune responses. Proc
Natl Acad Sci USA 103:12481–12486
Taguchi A, Yanagisawa K, Tanaka M et al (2008) Identification of
hypoxia-inducible factor-1 alpha as a novel target for miR-17-92
microRNA cluster. Cancer Res 68:5540–5545
Tanaka T, Arai M, Wu S et al (2011) Epigenetic silencing of
microRNA-373 plays an important role in regulating cell
proliferation in colon cancer. Oncol Rep 26:1329–1335
Tang Y, Luo X, Cui H et al (2009) MicroRNA-146A contributes to
abnormal activation of the type I interferon pathway in human lupus
by targeting the key signaling proteins. Arthr Rheum 60:1065–1075
Tavazoie SF, Alarcon C, Oskarsson T et al (2008) Endogenous human
microRNAs that suppress breast cancer metastasis. Nature
451:147–152
Tebbutt NC, Wilson K, Gebski VJ et al (2010) Capecitabine,
bevacizumab, and mitomycin in first-line treatment of metastatic
colorectal cancer: results of the Australasian Gastrointestinal
Trials Group Randomized Phase III MAX Study. J Clin Oncol
28:3191–3198
Teicher BA (1996) A systems approach to cancer therapy. (Antionco-
genics ? standard cytotoxics–[mechanism(s) of interaction).
Cancer Metastasis Rev 15:247–272
Thai TH, Calado DP, Casola S et al (2007) Regulation of the germinal
center response by microRNA-155. Science 316:604–608
Thiel M, Caldwell CC, Kreth S et al (2007) Targeted deletion of HIF-
1alpha gene in T cells prevents their inhibition in hypoxic
inflamed tissues and improves septic mice survival. PLoS ONE
2:e853
Thomas MD, Kremer CS, Ravichandran KS et al (2005) c-Myb is
critical for B cell development and maintenance of follicular B
cells. Immunity 23:275–286
Thomas M, Lieberman J, Lal A (2010) Desperately seeking
microRNA targets. Nat Struct Mol Biol 17:1169–1174
Tili E, Michaille JJ, Cimino A et al (2007) Modulation of miR-155
and miR-125b levels following lipopolysaccharide/TNF-alpha
stimulation and their possible roles in regulating the response to
endotoxin shock. J Immunol 179:5082–5089
Tong RT, Boucher Y, Kozin SV et al (2004) Vascular normalization
by vascular endothelial growth factor receptor 2 blockade
induces a pressure gradient across the vasculature and improves
drug penetration in tumors. Cancer Res 64:3731–3736
Tsitsiou E, Lindsay MA (2009) microRNAs and the immune
response. Curr Opin Pharmacol 9:514–520
Tsuchiya S, Fujiwara T, Sato F et al (2011) MicroRNA-210 regulates
cancer cell proliferation through targeting fibroblast growth
factor receptor-like 1 (FGFRL1). J Biol Chem 286:420–428
Ueda R, Kohanbash G, Sasaki K et al (2009) Dicer-regulated
microRNAs 222 and 339 promote resistance of cancer cells to
cytotoxic T-lymphocytes by down-regulation of ICAM-1. Proc
Natl Acad Sci USA 106:10746–10751
Urbich C, Kaluza D, Fromel T et al (2012) MicroRNA-27a/b controls
endothelial cell repulsion and angiogenesis by targeting sem-
aphorin 6A. Blood 119:1607–1616
Van de Veire S, Stalmans I, Heindryckx F et al (2010) Further
pharmacological and genetic evidence for the efficacy of PlGF
inhibition in cancer and eye disease. Cell 141:178–190
van Solingen C, Seghers L, Bijkerk R et al (2009) Antagomir-
mediated silencing of endothelial cell specific microRNA-126
impairs ischemia-induced angiogenesis. J Cell Mol Med
13:1577–1585
Wang S, Olson EN (2009) AngiomiRs–key regulators of angiogen-
esis. Curr Opin Genet Dev 19:205–211
Wang S, Aurora AB, Johnson BA et al (2008) The endothelial-
specific microRNA miR-126 governs vascular integrity and
angiogenesis. Dev Cell 15:261–271
Wang P, Gu Y, Zhang Q et al (2012) Identification of resting and type
I IFN-activated human NK cell miRNomes reveals microRNA-
378 and microRNA-30e as negative regulators of NK cell
cytotoxicity. J Immunol 189:211–221
Wildiers H, Guetens G, De Boeck G et al (2003) Effect of
antivascular endothelial growth factor treatment on the intratu-
moral uptake of CPT-11. Br J Cancer 88:1979–1986
Winter J, Jung S, Keller S et al (2009) Many roads to maturity:
microRNA biogenesis pathways and their regulation. Nat Cell
Biol 11:228–234
Wurdinger T, Tannous BA, Saydam O et al (2008) miR-296 regulates
growth factor receptor overexpression in angiogenic endothelial
cells. Cancer Cell 14:382–393
Xiao C, Calado DP, Galler G et al (2007) MiR-150 controls B cell
differentiation by targeting the transcription factor c-Myb. Cell
131:146–159
Xiao C, Srinivasan L, Calado DP et al (2008) Lymphoproliferative
disease and autoimmunity in mice with increased miR-17-92
expression in lymphocytes. Nat Immunol 9:405–414
Xu Q, Liu LZ, Qian X et al (2012) MiR-145 directly targets p70S6K1
in cancer cells to inhibit tumor growth and angiogenesis. Nucleic
Acids Res 40:761–774
Yamakuchi M, Lotterman CD, Bao C et al (2010) P53-induced
microRNA-107 inhibits HIF-1 and tumor angiogenesis. Proc
Natl Acad Sci USA 107:6334–6339
Yamakuchi M, Yagi S, Ito T et al (2011) MicroRNA-22 regulates
hypoxia signaling in colon cancer cells. PLoS ONE 6:e20291
Yang WJ, Yang DD, Na S et al (2005) Dicer is required for
embryonic angiogenesis during mouse development. J Biol
Chem 280:9330–9335
Zhao JL, Rao DS, Boldin MP et al (2011) NF-kappaB dysregulation
in microRNA-146a-deficient mice drives the development of
myeloid malignancies. Proc Natl Acad Sci USA 108:9184–9189
Zheng Y, Josefowicz SZ, Kas A et al (2007) Genome-wide analysis of
Foxp3 target genes in developing and mature regulatory T cells.
Nature 445:936–940
Zhou X, Jeker LT, Fife BT et al (2008) Selective miRNA disruption
in T reg cells leads to uncontrolled autoimmunity. J Exp Med
205:1983–1991
Zhou Q, Gallagher R, Ufret-Vincenty R et al (2011) Regulation of
angiogenesis and choroidal neovascularization by members of
microRNA-23*27*24 clusters. Proc Natl Acad Sci USA
108:8287–8292
Zhu QY, Liu Q, Chen JX et al (2010) MicroRNA-101 targets MAPK
phosphatase-1 to regulate the activation of MAPKs in macro-
phages. J Immunol 185:7435–7442
Arch. Immunol. Ther. Exp. (2013) 61:285–299 299
123
Related Documents





![Review Article - Hindawi Publishing Corporationdownloads.hindawi.com/journals/jo/2012/193436.pdftherapy [22]. Furthermore, antiangiogenesis can normalize tumor vasculature and decrease](https://static.cupdf.com/doc/110x72/5f4d5f35bd4e976d402d7ac5/review-article-hindawi-publishing-therapy-22-furthermore-antiangiogenesis.jpg)


![arXiv:0801.0654v1 [q-bio.TO] 4 Jan 2008 · 2019-03-21 · arXiv:0801.0654v1 [q-bio.TO] 4 Jan 2008 Hot spot formation in tumor vasculature during tumor growth in an arterio-venous-network](https://static.cupdf.com/doc/110x72/5e6f32e6d0f2c50bca1bb0c1/arxiv08010654v1-q-bioto-4-jan-2008-2019-03-21-arxiv08010654v1-q-bioto.jpg)


