Microphytobenthos vertical migratory photoresponse as characterised by light-response curves of surface biomass Jo~ ao Sero ˆdio * , Helena Coelho, So ´nia Vieira, So ´nia Cruz Departamento de Biologia, Universidade de Aveiro, Campus de Santiago, 3810-193 Aveiro, Portugal Received 30 November 2005; accepted 8 March 2006 Available online 22 May 2006 Abstract The migratory response of intertidal microphytobenthos to changes in irradiance was studied on undisturbed estuarine sediments. Two non- destructive optical techniques were used to trace variations in vivo of surface biomass: PAM fluorometry, for measuring the minimum fluores- cence level (F o ); and spectral reflectance analysis, for quantifying the normalized difference vegetation index (NDVI). Following the formation of a dense biofilm at the surface, replicated sediment samples were simultaneously exposed to six different irradiance levels, ranging from 50 to 1500 mmol m ÿ2 s ÿ1 , during a period of 120 min. The migratory photoresponse of the biofilms was characterised by constructing biomass vs. light curves (BLC), relating the accumulation of microalgal biomass after that period (estimated by F o or NDVI) to the irradiance level incident on the surface. BLCs allow characterising the main features of the migratory photoresponse of intact biofilms. Typical BLC showed a clear biphasic pattern, with an increase in microalgal accumulation under irradiances below 100 mmol m ÿ2 s ÿ1 , maximum values under 100e250 mmol m ÿ2 s ÿ1 , and a gradual decrease of surface biomass under higher irradiances, indicating a strong photophobic downward migra- tory response. Similar BLC patterns were obtained when measuring F o or NDVI. The construction of BLCs for biofilms from intertidal sites with distinctive sediment characteristics and diatom taxonomic composition allowed to detected significant differences in the migratory photores- ponse. Biofilms from a muddy sediment exhibited considerably larger amplitude in the migratory photoresponse than the biofilms from a sandy mud site, especially under high irradiances. The photophobic migratory response to high light was found to vary among diatom species, particularly in the case of the biofilms from the muddy sediments. Ó 2006 Elsevier Ltd. All rights reserved. Keywords: biomass; chlorophyll fluorescence; microphytobenthos; phototaxis; reflectance; vertical migrations 1. Introduction The migratory behaviour of sediment-inhabiting microalgae is a long known phenomenon, particularly well documented for estuarine intertidal microphytobenthos (Consalvey et al., 2004b). Many species of diatoms, euglenophytes and cyano- bacteria are capable of moving vertically within the upper layers of the sediment, in synchronization with dayenight and tidal cycles. These rhythmic movements are not limited to the photic zone of the sediment (Pinckney et al., 1994), and are known to be partially endogenously controlled (Round and Palmer, 1966; Sero ˆdio et al., 1997). In recent years, this phenomenon has been increasingly studied, largely because of its recognized ecological importance as a key controlling factor of short-term variability in community-level microphyto- benthic productivity (Pinckney and Zingmark, 1991; Sero ˆdio et al., 2001). The rhythmic vertical migration of large numbers of microalgae causes the periodic accumulation of large amounts of microalgal biomass in the photic zone of the sediment. By considerably affecting the fraction of solar irradi- ance that can be absorbed and used for photosynthesis by the biofilm, large fluctuations in depth-integrated production rates are thus caused (Sero ˆdio et al., 2001). Accordingly, this source of short-term variability has been formally included in the formulation of primary production models (Pinckney and Zingmark, 1993; Guarini et al., 2000; Sero ˆdio and Catarino, 2000). * Corresponding author. E-mail address: [email protected] (J. Sero ˆdio). 0272-7714/$ - see front matter Ó 2006 Elsevier Ltd. All rights reserved. doi:10.1016/j.ecss.2006.03.005 Estuarine, Coastal and Shelf Science 68 (2006) 547e556 www.elsevier.com/locate/ecss

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Estuarine, Coastal and Shelf Science 68 (2006) 547e556www.elsevier.com/locate/ecss
Microphytobenthos vertical migratory photoresponse as characterisedby light-response curves of surface biomass
Jo~ao Serodio*, Helena Coelho, Sonia Vieira, Sonia Cruz
Departamento de Biologia, Universidade de Aveiro, Campus de Santiago, 3810-193 Aveiro, Portugal
Received 30 November 2005; accepted 8 March 2006
Available online 22 May 2006
Abstract
The migratory response of intertidal microphytobenthos to changes in irradiance was studied on undisturbed estuarine sediments. Two non-destructive optical techniques were used to trace variations in vivo of surface biomass: PAM fluorometry, for measuring the minimum fluores-cence level (Fo); and spectral reflectance analysis, for quantifying the normalized difference vegetation index (NDVI). Following the formationof a dense biofilm at the surface, replicated sediment samples were simultaneously exposed to six different irradiance levels, ranging from 50 to1500 mmol m�2 s�1, during a period of 120 min. The migratory photoresponse of the biofilms was characterised by constructing biomass vs.light curves (BLC), relating the accumulation of microalgal biomass after that period (estimated by Fo or NDVI) to the irradiance level incidenton the surface. BLCs allow characterising the main features of the migratory photoresponse of intact biofilms. Typical BLC showed a clearbiphasic pattern, with an increase in microalgal accumulation under irradiances below 100 mmol m�2 s�1, maximum values under100e250 mmol m�2 s�1, and a gradual decrease of surface biomass under higher irradiances, indicating a strong photophobic downward migra-tory response. Similar BLC patterns were obtained when measuring Fo or NDVI. The construction of BLCs for biofilms from intertidal sites withdistinctive sediment characteristics and diatom taxonomic composition allowed to detected significant differences in the migratory photores-ponse. Biofilms from a muddy sediment exhibited considerably larger amplitude in the migratory photoresponse than the biofilms from a sandymud site, especially under high irradiances. The photophobic migratory response to high light was found to vary among diatom species,particularly in the case of the biofilms from the muddy sediments.� 2006 Elsevier Ltd. All rights reserved.
Keywords: biomass; chlorophyll fluorescence; microphytobenthos; phototaxis; reflectance; vertical migrations
1. Introduction
The migratory behaviour of sediment-inhabiting microalgaeis a long known phenomenon, particularly well documented forestuarine intertidal microphytobenthos (Consalvey et al.,2004b). Many species of diatoms, euglenophytes and cyano-bacteria are capable of moving vertically within the upperlayers of the sediment, in synchronization with dayenightand tidal cycles. These rhythmic movements are not limitedto the photic zone of the sediment (Pinckney et al., 1994),and are known to be partially endogenously controlled (Roundand Palmer, 1966; Serodio et al., 1997). In recent years, this
* Corresponding author.
E-mail address: [email protected] (J. Serodio).
0272-7714/$ - see front matter � 2006 Elsevier Ltd. All rights reserved.
doi:10.1016/j.ecss.2006.03.005
phenomenon has been increasingly studied, largely becauseof its recognized ecological importance as a key controllingfactor of short-term variability in community-level microphyto-benthic productivity (Pinckney and Zingmark, 1991; Serodioet al., 2001). The rhythmic vertical migration of large numbersof microalgae causes the periodic accumulation of largeamounts of microalgal biomass in the photic zone of thesediment. By considerably affecting the fraction of solar irradi-ance that can be absorbed and used for photosynthesis by thebiofilm, large fluctuations in depth-integrated production ratesare thus caused (Serodio et al., 2001). Accordingly, this sourceof short-term variability has been formally included in theformulation of primary production models (Pinckney andZingmark, 1993; Guarini et al., 2000; Serodio and Catarino,2000).

548 J. Serodio et al. / Estuarine, Coastal and Shelf Science 68 (2006) 547e556
Benthic microalgal motility is known to be affected byvarious environmental factors, such as light intensity (Perkins,1960; Paterson, 1986; Kingston, 1999a), light spectrum (Cohnet al., 1999), sediment physical disturbance (Hopkins, 1966),water cover (Pinckney et al., 1994; Mitbavkar and Anil,2004), wave action (Kingston, 1999b), temperature (Cohnet al., 2003), and sub-surface nutrients (Kingston, 2002).Migratory responses to irradiance are particularly interestingdue to the large variability of this parameter under in situintertidal conditions and because it directly affects the func-tioning of the photosynthetic apparatus, triggering photopro-tective mechanisms, and causing photoinhibition (Serodioet al., 2005a). The influence of irradiance on the migratorybehaviour is also important for the modelling of primaryproductivity and for the characterisation of the phototacticresponses under high light, in the context of the often sug-gested hypothesis that the downward vertical migration mayrepresent a form of behavioural photoacclimation or photopro-tection (Admiraal, 1984; Kromkamp et al., 1998; Serodioet al., 2001). Most manipulative studies of benthic microalgalmotility as a response to environmental stimuli have been car-ried out on single-species microalgal populations grown inculture, under conditions markedly different from the naturalmicroenvironment inhabited by natural assemblages (Cohnand Disparti, 1994; Cohn et al., 1999, 2004). On undisturbedsediments, such investigations have been limited to eugleno-phytes (Kingston, 1999a). To our knowledge, no systematicstudies have been carried out explicitly on the migratoryphotoresponse of sediment-inhabiting diatoms while compris-ing natural biofilms.
This work addresses the characterisation of the migratory re-sponse of natural microphytobenthic biofilms to irradiance. Thephototactic response was studied by measuring the variationwith incident irradiance of the surface biomass, here definedas the biomass present in the photic zone of the sediment andthat is detectable by the optical techniques used in vivo tonon-destructively estimate it: pulse amplitude modulation(PAM) fluorometry (for measuring the minimum fluorescencelevel (Fo)) and spectral reflectance analysis (for measuringthe normalized difference vegetation index (NDVI)). Themigratory photoresponse of undisturbed microphytobenthoswas quantitatively characterised by constructing biomass vs.light curves, plots that relate the photoaccumulation of biomass(estimated by Fo or NDVI) to the irradiance level incident onthe sediment surface. This approach was applied to the compar-ative study of the phototactic response of microphytobenthicbiofilms growing on sediments of different granulometriesand with different species composition.
2. Materials and methods
2.1. Sampling
Undisturbed sediment samples were collected on two inter-tidal flats in the Ria de Aveiro, a mesotidal estuary of the westcoast of Portugal: Vista Alegre (VA; 40 � 350 N, 8 � 410 W), onthe west margin of the Canal de Ilhavo, and Gafanha da
Encarnac~ao (GE; 40 � 380 N, 8 � 440 W), on the east marginof the Canal de Mira. Sampling sites had different granulomet-ric characteristics. The VA sampling site composed of finemuddy sediments (97% particles below 63 mm) while theGE site composed of sandy mud (45.3% particles between63 and 125 mm; 42.7% below 63 mm). Sampling was carriedout from November 2004 to April 2005. Sediment sampleswere collected using plastic corers (3.6 cm diameter) andtaken to the laboratory where all the measurements were car-ried out. Samples were sectioned with minimum disturbanceinto plastic rings of 5 mm deep (same diameter of samplingcorers), and maintained in a Petri dish containing water col-lected at the sampling site. When necessary, slurries were pre-pared by resuspending repeatedly the top 2 mm of sediment infiltered site water using a syringe. Microalgal suspensionswere prepared by placing two pieces of lens tissue on the sur-face of illuminated sediment cores during daytime low tide.Microalgae were collected by resuspending the upper pieceof lens tissue in filtered site water (Eaton and Moss, 1966).
2.2. Chlorophyll fluorescence
Variable chlorophyll a fluorescence was measured usinga PAM fluorometer comprising a computer-operated PAM-Control Unit (Walz, Effeltrich, Germany) and a WATER-EDF-Universal emitter-detector unit (Gademann InstrumentsGmbH, Wurzburg, Germany). This instrument uses a modulatedblue light (LED-lamp peaking at 450 nm, half-bandwidth of20 nm; further details in Serodio et al., 2005b) as source formeasuring, actinic and saturating light, emitted at a frequencyof 18 Hz when measuring Fo. When using sediment samples(undisturbed samples or slurries), fluorescence was measuredusing a 6-mm diameter Fluid Light Guide fiberoptics bundlethat delivered the measuring and saturating light provided bythe fluorometer. Measurements were taken by positioning thefiberoptics perpendicularly to the sediment surface, at a fixeddistance of 1 mm. The relative position of the fiberoptics andthe sediment surface was controlled using a micromanipulator(MM33, Martzhauher, Germany). When measuring on microal-gal suspensions, the fiberoptics bundle was connected to a fluo-rescence cuvette (KS-101, Walz). Unless stated otherwise,samples were dark-adapted for 2 min before the fluorescencelevel (Fo) was measured. On each occasion, Fo was measuredin three separate, non-overlapping areas within each sample.
2.3. Spectral reflectance
Reflectance spectra were measured using a fiber optic spec-trometer (USB2000-VIS-NIR, grating #3, Ocean Optics,Duiven, The Netherlands). Reflectance was recorded overthe 350e1000 nm bandwidth with a sampling spectral resolu-tion of 0.38 nm, using a 400-mm diameter fiber optic (modelQP400-2-VIS/NIR-BX, Ocean Optics), positioned perpendic-ularly to the sediment surface at a fixed distance of 2.2 cm,set to determine a view field coincident with the area moni-tored by the PAM fiberoptics. Reflectance was determinedfrom the light spectrum reflected from the undisturbed

549J. Serodio et al. / Estuarine, Coastal and Shelf Science 68 (2006) 547e556
sediment sample, normalized to spectrum reflected from areference white panel. A reflectance spectrum measured in thedark was subtracted with both the sample and the whitereference spectra to account for the dark current noise ofthe spectrometer. Spectrum measurement was carried out byexposing the samples (and the reference panel) to a constant ir-radiance of 200 mmol m�2 s�1, provided by the same lightsource that was used to study the migratory photoresponses(see below). Three replicated spectra were measured on each oc-casion, and the mean spectrum was used for the subsequent cal-culations. Reflectance measurements were used to estimate themicrophytobenthos surface biomass by calculating the normal-ized difference vegetation index (NDVI; Rouse et al., 1973):
NDVI¼ R750�R675
R750þR675
ð1Þ
where R750 and R675 represent the reflectance measured at 750and 675 nm, respectively. R750 and R675 were calculated byaveraging the values obtained in the intervals of 749.62e750.38 nm and 674.62e675.38 nm, respectively.
2.4. Fo, NDVI vs. sediment Chl a content
To test the possibility to use the fluorescence parameter Fo
and the reflectance-based index NDVI to trace changes in themicroalgal concentration in the upper layers of the sediment,Fo and NDVI were measured in vertically homogenous sedi-ment samples of increasing Chl a content, prepared by addingincreasing amounts of microalgae to sterilized sediment(Serodio et al., 1997). Microalgae used in these slurries werecollected by using lens tissue on sediment samples exposed tolow light (100e150 mmol m�2 s�1) for several hours in themorning after the day of collection. Fo was determined afterthe slurries were dark-adapted for 30 min. Chl a was extractedin 90% aqueous acetone and quantified spectrophotometricallyfollowing the method of Lorenzen (1967).
Because the fluorescence parameter Fo may be affected byphysiological effects determined by light exposure prior todark-adaptation, the relationship between Fo and Chl a was stud-ied in samples that were previously exposed to a range of differ-ent irradiance levels. To isolate the physiological response of Fo
and to avoid the confounding effects of migratory responses toincident irradiance, experiments were carried out using microal-gal suspensions. Microalgae were collected as described aboveand were resuspended in filtered site water. Suspensions wereexposed to 100, 500 and 1500 mmol m�2 s�1 during 90 min,after which they were dark-adapted for 2 min and Fo was deter-mined. The suspensions were filtered on GF/F filters and Chla was determined spectrophotometrically.
2.5. Migratory photoresponse: biomass vs. light curves
The migratory photoresponse of microphytobenthic assem-blages was studied by simultaneously exposing replicate sed-iment samples to different irradiance levels and by followingthe induced changes in surface biomass through variations in
Fo and NDVI. The results of these experiments were used toconstruct biomass vs. light curves, which by summarizingthe migratory response to irradiance allow to characterisethe phototactic responses of intact assemblages along a rangeof irradiances. Coinciding with the beginning of the low tide inthe sampling site, samples were exposed to 150 mmol m�2 s�1
for 1 h to induce the upward migration of motile microalgae(determined in previous tests). After this period, the sampleswere simultaneously exposed to a range of different irradi-ances for a period of 120 min. Each sample was exposed sep-arately to a constant irradiance level during this period. Every20 min, spectral reflectance was measured on each sample, af-ter which was darkened for 2 min before Fo was determined.NDVI and Fo were determined for three matching non-overlapping areas of each sample. Samples were exposed toactinic cold white light, provided by a halogen lamp (Quartz-line lamp DDL 150W, General Electric, USA) emitting lightwith a continuous colour spectrum similar to natural sunlight.Light was delivered to the sample by a fiberoptics bundle con-nected to a 5-cm diameter light ring (Standard Ringlight andVolpi Intralux 5000-1, Volpi, Switzerland) which provideda homogeneous shade-free light field. Samples were placedbelow the centre of the light ring, at a fixed vertical distancefrom it (ca. 2 cm). Fo and NDVI were measured by passingthe respective fiberoptics through the centre of the light ring.Photon irradiance incident on the sample surface was mea-sured using a PAR micro-sensor (Spherical MicroQuantumSensor, US-SQS/W, Walz). A total of six illumination systemswere used to separately and simultaneously expose each of sixsediment samples to each of the following irradiance levels:50, 100, 250, 500, 1000 and 1500 mmol m�2 s�1. One addi-tional sample was transferred to darkness and maintained inthe dark during the measuring period.
2.6. Species identification
After the construction of biomass vs. light curves, the upper1 mm of the sediment samples exposed to 100, 500 and1500 mmol m�2 s�1 was sectioned and diluted in a knownvolume of filtered sea water for determination of taxonomiccomposition. Sub-samples of the diluted slurries were fixedin 1% v/v formaldehyde and viewed under bright-field micro-scope for determination of the relative abundance of majortaxonomic groups (diatoms, euglenophytes and cyanobacte-ria), by counting a minimum of 400 cells on three replicatedsub-samples. Diatom identification was done on sub-samplesoxidized using nitric acid and potassium dichromate, anddigested in concentrated HCl.
3. Results
3.1. Fo, NDVI vs. sediment Chl a content
Both Fo and NDVI were found to vary linearly with thesediment Chl a content (Fig. 1). Highly significant correlationswere obtained between Chl a content and Fo (r¼ 0.965,p< 0.001) and NDVI (r¼ 0.938, p< 0.001). Linear
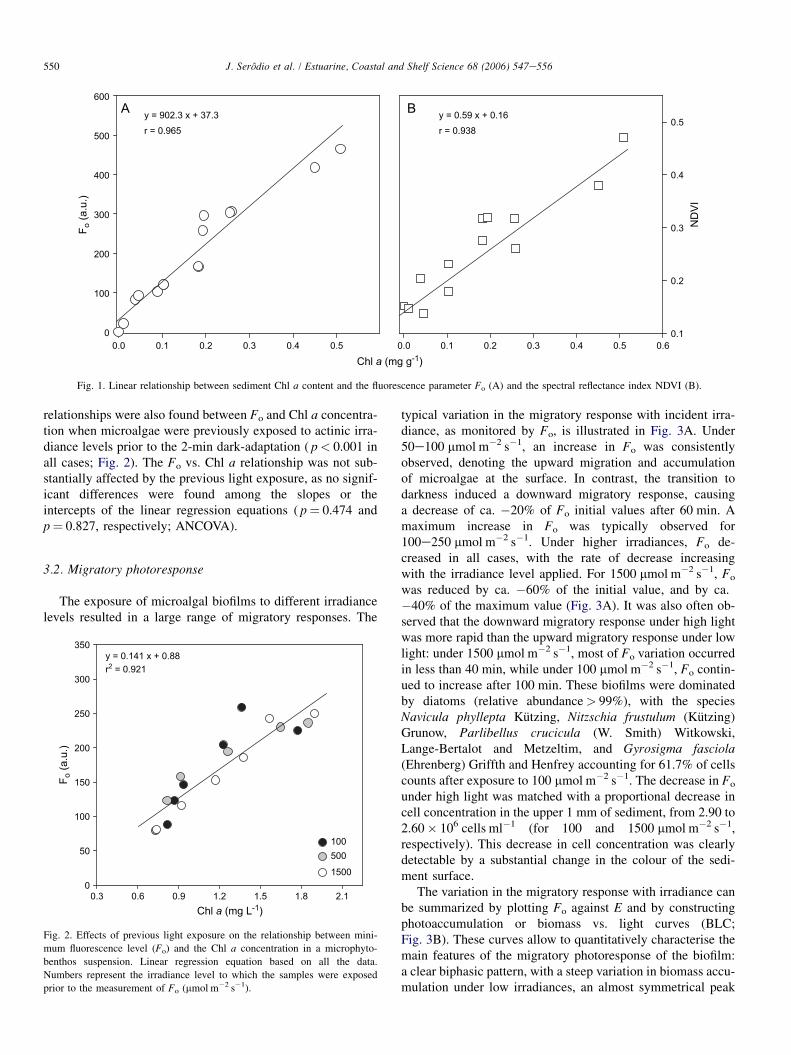
550 J. Serodio et al. / Estuarine, Coastal and Shelf Science 68 (2006) 547e556
Chl a (mg g-1)
F o (a
.u.)
0
100
200
300
400
500
600
0.0 0.1 0.2 0.3 0.4 0.5 0.0 0.1 0.2 0.3 0.4 0.5 0.6
ND
VI
0.1
0.2
0.3
0.4
0.5y = 902.3 x + 37.3 r = 0.965
y = 0.59 x + 0.16r = 0.938
A B
Fig. 1. Linear relationship between sediment Chl a content and the fluorescence parameter Fo (A) and the spectral reflectance index NDVI (B).
relationships were also found between Fo and Chl a concentra-tion when microalgae were previously exposed to actinic irra-diance levels prior to the 2-min dark-adaptation ( p< 0.001 inall cases; Fig. 2). The Fo vs. Chl a relationship was not sub-stantially affected by the previous light exposure, as no signif-icant differences were found among the slopes or theintercepts of the linear regression equations ( p¼ 0.474 andp¼ 0.827, respectively; ANCOVA).
3.2. Migratory photoresponse
The exposure of microalgal biofilms to different irradiancelevels resulted in a large range of migratory responses. The
Chl a (mg L-1)0.3 0.6 0.9 1.2 1.5 1.8 2.1
F o (a
.u.)
0
50
100
150
200
250
300
350
100500
1500
y = 0.141 x + 0.88 r2 = 0.921
Fig. 2. Effects of previous light exposure on the relationship between mini-
mum fluorescence level (Fo) and the Chl a concentration in a microphyto-
benthos suspension. Linear regression equation based on all the data.
Numbers represent the irradiance level to which the samples were exposed
prior to the measurement of Fo (mmol m�2 s�1).
typical variation in the migratory response with incident irra-diance, as monitored by Fo, is illustrated in Fig. 3A. Under50e100 mmol m�2 s�1, an increase in Fo was consistentlyobserved, denoting the upward migration and accumulationof microalgae at the surface. In contrast, the transition todarkness induced a downward migratory response, causinga decrease of ca. �20% of Fo initial values after 60 min. Amaximum increase in Fo was typically observed for100e250 mmol m�2 s�1. Under higher irradiances, Fo de-creased in all cases, with the rate of decrease increasingwith the irradiance level applied. For 1500 mmol m�2 s�1, Fo
was reduced by ca. �60% of the initial value, and by ca.�40% of the maximum value (Fig. 3A). It was also often ob-served that the downward migratory response under high lightwas more rapid than the upward migratory response under lowlight: under 1500 mmol m�2 s�1, most of Fo variation occurredin less than 40 min, while under 100 mmol m�2 s�1, Fo contin-ued to increase after 100 min. These biofilms were dominatedby diatoms (relative abundance> 99%), with the speciesNavicula phyllepta Kutzing, Nitzschia frustulum (Kutzing)Grunow, Parlibellus crucicula (W. Smith) Witkowski,Lange-Bertalot and Metzeltim, and Gyrosigma fasciola(Ehrenberg) Griffth and Henfrey accounting for 61.7% of cellscounts after exposure to 100 mmol m�2 s�1. The decrease in Fo
under high light was matched with a proportional decrease incell concentration in the upper 1 mm of sediment, from 2.90 to2.60� 106 cells ml�1 (for 100 and 1500 mmol m�2 s�1,respectively). This decrease in cell concentration was clearlydetectable by a substantial change in the colour of the sedi-ment surface.
The variation in the migratory response with irradiance canbe summarized by plotting Fo against E and by constructingphotoaccumulation or biomass vs. light curves (BLC;Fig. 3B). These curves allow to quantitatively characterise themain features of the migratory photoresponse of the biofilm:a clear biphasic pattern, with a steep variation in biomass accu-mulation under low irradiances, an almost symmetrical peak

551J. Serodio et al. / Estuarine, Coastal and Shelf Science 68 (2006) 547e556
Irradiance (µmol quanta m-2 s-1)0 250 500 750 1000 1250 1500
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.660 min120 min
Time (min)0 20 40 60 80 100 120 140
Fo (r
el. u
nits
)
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
100
50250
0500
1000
1500
Fo (r
el. u
nits
)
A B
Fig. 3. (A) Typical migratory light response of undisturbed microphytobenthos biofilms, as determined by measuring fluorescence parameter Fo on samples pre-
exposed to constant irradiance of 150 mmol m�2 s�1 for 120 min. Numbers represent the irradiance level prior to the measurement of Fo (mmol m�2 s�1). Fo values
normalized to initial values. (B) Photoaccumulation or biomass vs. light curves (BLC), constructed from the data in panel (A). Examples of BLCs obtained after 60
and 120 min of light exposure. Mean values of three measurements.
under E� 100 mmol m�2 s�1, and a continuous decrease underhigher irradiances, almost linear for E� 500 mmol m�2 s�1.Although some variability was found among days, the sameoverall pattern was consistently observed in all the experiments,which was usually formed and stabilized after 1 h of lightexposure (Fig. 3B). The biphasic pattern observed whenmeasuring Fo was confirmed to represent changes in microalgalbiomass accumulation at the surface as very similar resultswere obtained when measuring NDVI. Highly significantcorrelations ( p< 0.001) were obtained between Fo and NDVIfor all the BLC constructed. This general pattern of photores-ponse was observed in all the experiments that were carriedout during a period of several months, both when measuringFo and NDVI.
3.3. Application to the comparison of biofilms
The construction of BLCs for microphytobenthos from in-tertidal sites with distinctive sediment granulometry revealeda significant difference in the migratory photoresponse(Fig. 4). Although the same general pattern was found inboth cases, similar to the one described above (Fig. 3), the mi-croalgal biofilm from the site with finer sediment (VA) ex-hibited a larger amplitude in the migratory response tochanges in irradiance, with Fo varying by more than 120% be-tween maximum and minimum values (under 100 and1500 mmol m�2 s�1, respectively). In contrast, the migratoryresponse of the biofilm from the sandier sediment (GE) wasless accentuated, both under low and high irradiances, withthe maximum range of Fo variation being less than 40%.The difference in the migratory response of the two types ofbiofilms is particularly evident regarding the decrease of sur-face biomass under high light. While for VA, Fo was reducedby more than �20% (relatively to initial values) under500 mmol m�2 s�1 and by ca. �65% under 1500 mmol m�2 s�1,
in the case of GE, values below the initial value were foundonly under 1000 mmol m�2 s�1, and the maximum decreasereached less than �17%.
The two types of biofilms presented important differencesregarding species composition and relative abundance. Thebiofilms from the muddy site VA were dominated by epipelicdiatoms Navicula gregaria, Navicula phyllepta, Parlibelluscrucicula and Gyrosigma fasciola. The assemblages fromthe sandy mud site GE were dominated by diatoms Ach-nanthes delicatula Kutzing, Achnanthes minutissima Kutzingand N. gregaria Donkin. The relative abundance of eugleno-phytes (Euglena sp.) and cyanobacteria (Oscillatoria sp.,Merismopedia sp.) was in all cases <1%. Despite the differ-ence in species composition, in both types of biofilms the de-crease in Fo was followed by a parallel decrease inmicroalgal cell concentration in the upper 1 mm of sediment
0 250 500 750 1000 1250 1500
F o (r
el. u
nits
)
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
1.8VAGE
Irradiance (µmol quanta m-2 s-1)
Fig. 4. Biomass vs. light curves for microphytobenthos biofilms from two in-
tertidal sites with distinctive sediment characteristics (VA: mud; GE: sandy
mud). Mean values of three measurements. Vertical bars represent 1 standard
deviation.
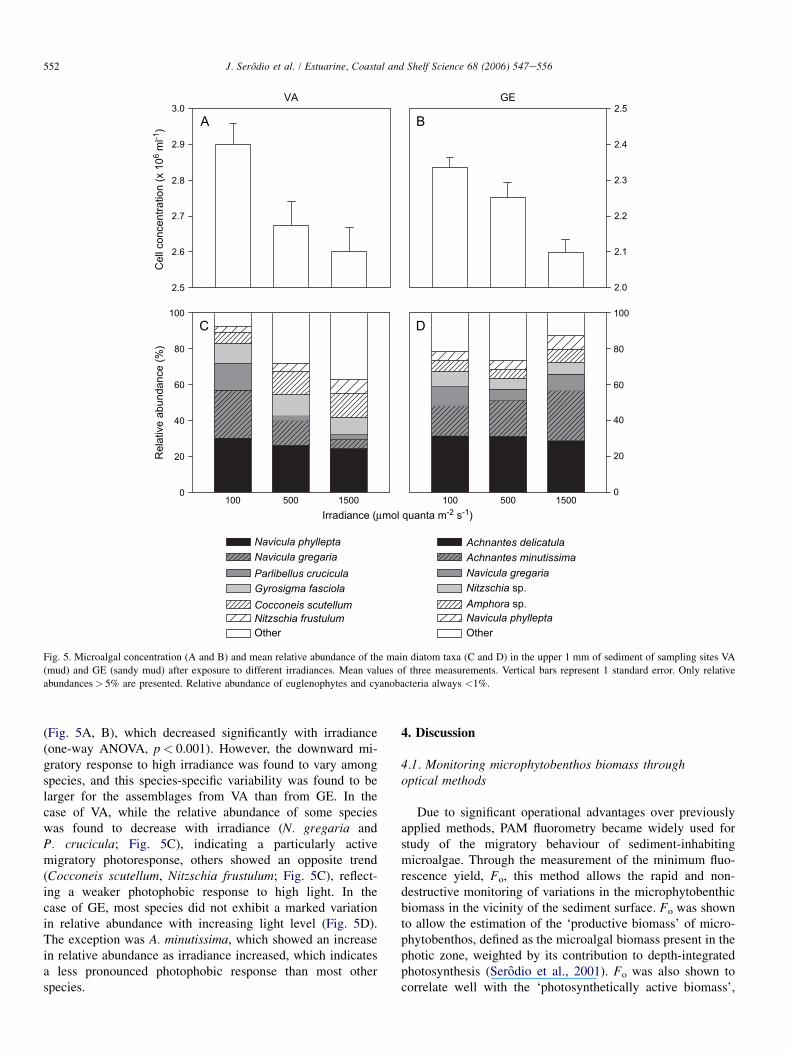
552 J. Serodio et al. / Estuarine, Coastal and Shelf Science 68 (2006) 547e556
100 500 1500
Rel
ativ
e ab
unda
nce
(%)
0
20
40
60
80
100
Navicula phyllepta
Navicula gregaria
Parlibellus crucicula
Gyrosigma fasciola
Cocconeis scutellum
Nitzschia frustulum
Other
VA GE
100 500 15000
20
40
60
80
100
Achnantes delicatula
Achnantes minutissima
Navicula gregaria
Nitzschia sp.Amphora sp.Navicula phyllepta
Other
Cel
l con
cent
ratio
n (x
106
ml-1
)
2.5
2.6
2.7
2.8
2.9
3.0
2.0
2.1
2.2
2.3
2.4
2.5A B
C D
Irradiance (µmol quanta m-2 s-1)
Fig. 5. Microalgal concentration (A and B) and mean relative abundance of the main diatom taxa (C and D) in the upper 1 mm of sediment of sampling sites VA
(mud) and GE (sandy mud) after exposure to different irradiances. Mean values of three measurements. Vertical bars represent 1 standard error. Only relative
abundances> 5% are presented. Relative abundance of euglenophytes and cyanobacteria always <1%.
(Fig. 5A, B), which decreased significantly with irradiance(one-way ANOVA, p< 0.001). However, the downward mi-gratory response to high irradiance was found to vary amongspecies, and this species-specific variability was found to belarger for the assemblages from VA than from GE. In thecase of VA, while the relative abundance of some specieswas found to decrease with irradiance (N. gregaria andP. crucicula; Fig. 5C), indicating a particularly activemigratory photoresponse, others showed an opposite trend(Cocconeis scutellum, Nitzschia frustulum; Fig. 5C), reflect-ing a weaker photophobic response to high light. In thecase of GE, most species did not exhibit a marked variationin relative abundance with increasing light level (Fig. 5D).The exception was A. minutissima, which showed an increasein relative abundance as irradiance increased, which indicatesa less pronounced photophobic response than most otherspecies.
4. Discussion
4.1. Monitoring microphytobenthos biomass throughoptical methods
Due to significant operational advantages over previouslyapplied methods, PAM fluorometry became widely used forstudy of the migratory behaviour of sediment-inhabitingmicroalgae. Through the measurement of the minimum fluo-rescence yield, Fo, this method allows the rapid and non-destructive monitoring of variations in the microphytobenthicbiomass in the vicinity of the sediment surface. Fo was shownto allow the estimation of the ‘productive biomass’ of micro-phytobenthos, defined as the microalgal biomass present in thephotic zone, weighted by its contribution to depth-integratedphotosynthesis (Serodio et al., 2001). Fo was also shown tocorrelate well with the ‘photosynthetically active biomass’,

553J. Serodio et al. / Estuarine, Coastal and Shelf Science 68 (2006) 547e556
defined simply as the microalgal biomass present in the photiczone of the sediment (Guarini et al., 2000; Honeywill et al.,2002).
Nevertheless, the use of Fo as a proxy for microphytoben-thos biomass has potentially important shortcomings. Becausethe fluorescence emission per unit Chl a depends on the phys-iological status of the microalgae, Fo has been preferred toother fluorescence parameters as it was shown to be the leastaffected by environmental factors (Serodio et al., 1997,2001; Consalvey et al., 2004a). However, the measurementof Fo requires the darkening of the sample which, in thecase of diatoms, should ideally be prolonged for a periodtime of tens of minutes (Ting and Owens, 1993). As the resultsof this study confirmed, the exposure to darkness, even duringthe early part of the light period, can induce a confoundingdecrease in surface biomass. As a compromise betweenminimizing the physiological effects of recent light historyand the artifactual migratory responses to darkness, Fo hasbeen measured by applying relatively short dark-adaptationperiods, such as 2 min (Serodio et al., 2005b), 5 min (Serodioet al., 2001) or 15 min (Honeywill et al., 2002; Consalveyet al., 2004a). The results of this study indicate that variationsin Fo, measured after only 2 min of dark-adaptation can bemostly attributed to changes in the Chl a content of the sedi-ment. Physiological effects causing the quenching of Fo underhigh light may be expected not to confound significantly thechanges in Fo resulting from variations in microalgal biomass,because its effects can be assumed as much smaller that thetypical changes in microalgal biomass associated to verticalmigrations.
Indices based on spectral reflectance, like NDVI, have beenincreasingly used as an alternative optical method for non-invasively monitoring changes in microphytobenthos biomass(Paterson et al., 1998; Decho et al., 2003; Carrere et al., 2004;Murphy et al., 2004). The use of NDVI avoids most problemsassociated to the use of Fo, as it is not affected by the physi-ological status of the microalgae and thus its determinationdoes not require dark-adaptation of the sample. A possiblecause for discrepancies between Fo and NDVI-based estimatesof surface Chl a content is the different depth interval belowthe sediment surface which is monitored by two types ofmethods. This depth interval is determined by the optical prop-erties of the sediment, which control the penetration ofdownwelling measuring light (of the PAM fluorometer) andof ambient light, and the attenuation of upwelling fluorescenceand reflected light (Serodio et al., 1997, 2001; Kromkampet al., 1998). In this study, the estimation of the depth intervalcorrespondent to reflectance signals was not attempted. How-ever, because reflectance was measured under a constant irra-diance level, the vertical zone monitored by NDVI and Fo maybe expected to be proportional. Accordingly, similar migratoryresponses were obtained using the two methods.
4.2. Biomass vs. light curves
Microphytobenthos are probably unique among aquaticphotoautotrophs in having the capacity of adjusting
photosynthetic activity not only through physiological regula-tion of the use of absorbed light energy by the photosyntheticapparatus, but also through active control of light absorption,by behaviourally exploiting environmental light gradients. Al-though the exposure to light is controlled at the individuallevel, by the movement of cells within the vertical light gradi-ent, its effects are expressed at the community level throughvariations in the biofilm biomass present in the photic zone.As photosynthesis vs. light (PeI ) curves are used tocharacterise the physiological response to changes in absorbedlight energy, the variable migratory response to changes in am-bient irradiance can be adequately described by biomass vs.light curves. BLCs characterise the migratory photoresponseof the whole biofilm, thus providing a way to compare thebehavioural photoresponse of microalgal assemblages alongtime, site characteristics, or species composition. Results onthe migratory response to light of microphytobenthic assem-blages were first reported in the form of BLCs by Kingston(1999a), summarizing the results of manipulative experimentson the phototactic response of Euglena proxima populationsunder in situ conditions. In this present study, to avoid thepossibly confounding effects of temperature on the migratoryphotoresponse (Cohn et al., 2003), unavoidable whenmeasurements are carried out in situ, cold light sources wereused to ensure the characterisation of the response to incidentirradiance independently of temperature. Nevertheless, itcannot be excluded that temperature, directly associated tovariations in incident solar irradiance, may also play a roleas an environmental clue in the determination of the migratorypattern of natural assemblages in situ.
4.3. Migratory photoresponse and behaviouralphotoprotection
The construction of BLC on intact microphytobenthos sam-ples revealed a consistent biphasic photoaccumulation patternthat resulted from the upward migration under low to moderateirradiances (50e250 mmol m�2 s�1) and the downward mi-gration under very low (<50 mmol m�2 s�1) and high(>500 mmol m�2 s�1) irradiances. The downward migratoryresponse to low light or darkness is likely to result from thefact that microalgae take the decrease in ambient light as anenvironmental clue indicating sunset or incoming high tide.Of particular interest is the downward migratory response un-der high light, consistently found in this study. This photopho-bic behaviour has often been hypothesized as a form of‘behavioural photoprotection’, through which motile microal-gae avoid exposure to potentially damaging irradiance levels(Admiraal, 1984; Kromkamp et al., 1998; Perkins et al.,2001; Serodio et al., 2001; Underwood, 2002). This eventualphotoprotective ability would be functionally equivalent tothe well-known phenomenon of chloroplast avoidance move-ments in higher plants (Haupt and Scheuerlein, 1990) whicheffectively decreases photodamage caused by high light(Kasahara et al., 2002). Although the downward migrationof sediment-inhabiting microalgae under high light is fre-quently mentioned in the literature, only a few studies have

554 J. Serodio et al. / Estuarine, Coastal and Shelf Science 68 (2006) 547e556
shown experimental evidence of this behaviour, either directly,through manipulation of light levels (Perkins, 1960; Kingston,1999a) or indirectly, through the analysis of the light responseof depth-integrated estimates of photosystem II electron trans-port rates (Perkins et al., 2001). Nevertheless, the migratoryphotoresponse behaviour of diatoms has been described in de-tailed from studies carried out on unialgal cultures (Cohnet al., 1999, 2003, 2004). The biphasic response to light re-ported in this study is in close agreement with the results ob-tained with the diatom Cruticula cuspidata, that has shown theoccurrence of step-down photophobic migratory responses un-der 50e100 mmol m�2 s�1 and step-up photophobic responsesunder irradiances above 500 mmol m�2 s�1 (Cohn et al., 2004).
While the decrease in Fo or NDVI observed when samplesare transferred from low to high irradiance is a clear indicationof a photophobic migratory reaction to high light, the availabledata do not allow to conclude on the photoprotective role ofthis behaviour. Nevertheless, it is interesting to note that theirradiance levels under which the photophobic response is trig-gered (100e250 mmol m�2 s�1) generally coincide with levelsfound, on microphytobenthic assemblages from the same in-tertidal site, to induce the operation of reversible photoprotec-tive nonphotochemical quenching (NPQ) mechanisms(Serodio et al., 2005a). NPQ processes operate by reducingthe transfer of absorbed light energy to the reaction centersand, while providing an effective protection against excessivelight, result in the lowering of the rate of photosynthesis(Muller et al., 2001). This suggests that the negative phototaxisunder high light may in fact reflect a way to regulate the lightlevel to which the cells are exposed to, in order to minimizethe triggering of physiological photoprotective mechanismsand to maintain the capacity for high photosynthetic rates.The peak in biomass accumulation observed under low lightmay simply reflect the fact that such irradiance levels aretoo low to cause photodamages and to induce physiologicalor behavioural photoprotective responses. Under suchfavourable light conditions, it may be expected the continuousaccumulation of microalgae at the surface, only limited byspace availability. This would explain the fact that under100 mmol m�2 s�1, Fo and NDVI increased continuously fora longer period than under other irradiance levels. A steepdecrease in surface biomass under high light was also foundby Kingston (1999a) on microphytobenthos dominated byEuglena proxima. However, a peak in biomass accumulation un-der intermediate light levels was not found, which may explainedby the absence of data for the range 30e300 mmol m�2 s�1.Thus, the undetected presence of a photoaccumulation patternsimilar to the one found in this study cannot be excluded.
The construction of BLCs on microphytobenthos fromsediments of distinct granulometry revealed a clear differenceregarding the migratory photoresponse. The different speciescomposition of the two assemblages suggests that the variationin the photoresponse pattern may be associated to a distinct ca-pacity for behaviourally regulating light exposure. In fact, thebiofilms from fine the muddy sediments (VA) were dominatedby epipelic diatoms, for which longer and faster vertical mi-grations are expected. Also, the assemblages from sandier
sediments (GE) were composed to a large extent by diatomsof the genera Achnanthes and Amphora, known to includenon-migratory, epipsamic species which live attached to thelarger sediment grains (Round, 1979; Admiraal, 1984). The re-sults indicate that the light-induced migratory behaviour mayhave a higher relative importance as a photoregulatory andphotoprotective strategy in the case of epipelic-dominatedbiofilms. And that in the case of microalgal assemblagesgrowing on coarser sediments, the regulation of light absorp-tion and photoprotection may have to rely to a larger extenton physiological processes. BLCs could be used togetherwith measurements of NPQ to ascertain the relative impor-tance of behavioural and physiological photoprotection in epi-pelic and epipsamic-dominated microphytobenthic biofilms.
In this study, the migratory photoresponse of microphyto-benthos was characterised on the basis of variations in surfacebiomass. However, the occurrence of changes in species com-position in the photic zone cannot be excluded as there is wideevidence that overall migratory behaviour of the whole biofilmis in fact composed of different, species-specific behaviourpatterns (Palmer and Round, 1965; Round and Palmer, 1966;Round, 1979; Paterson, 1986; Perkins et al., 2002; Underwoodet al., 2005). Considerable diversity in the migratory photores-ponse may be expected as different species may have differentmigratory capacities or may respond diversely to changes inlight. Diatom motility has been shown to vary with species(Bertrand, 1990), type of mucilage produced while moving(Bellinger et al., 2005) and the presence of other species(Cohn et al., 2003). Furthermore, the replacement of cells inthe photic zone may also have occurred (‘micromigration’;Kromkamp et al., 1998), particularly during prolonged periodsunder high light. Because the photic depth (<200 mm; Serodioet al., 1997; Kromkamp et al., 1998) is certainly much smallerthan relatively large thickness of the sediment sections (1 mm)that were used for species identification and counting, it isconceivable that more important changes in species composi-tion occurring in this depth interval may have passed largelyundetected.
4.4. Implications for the modelling of primaryproductivity
Although BLCs relate quantitatively the surface biomasswith incident irradiance, they cannot be used directly topredict the variation in surface biomass from irradiance datameasured during daytime emersion periods. First, because bio-mass vs. light curves were constructed only after the biofilmwas formed at the sediment surface (induced by the exposureto low light), the relationship between biomass and irradiancethus established may not represent the pattern of microalgaeaccumulation following the hourly variation in solar light in-tensity. Secondly, the biomass measurements were madeover relatively short periods, therefore not allowing for thebuild-up of factors that in situ have cumulative effects on pho-tosynthesis and may enhance photodamage, like high pH, highO2 concentrations, or carbon depletion. Nevertheless, the ob-tained results have potentially important implications for the

555J. Serodio et al. / Estuarine, Coastal and Shelf Science 68 (2006) 547e556
modelling of short-term (intraday) variability of the photosyn-thetic rate of microphytobenthos and for the estimation of pri-mary productivity budgets for estuarine intertidal areas.Recent models acknowledge the importance of migratoryrhythms for the quantification of the instantaneous rates ofdepth-integrated photosynthesis, by modelling it as a functionof productive biomass, either implicitly (Pinckney and Zing-mark, 1993) or explicitly (Guarini et al., 2000; Serodio andCatarino, 2000). The hourly variability of productive biomassis in turn modelled as a function of solar and tidal cycles, re-sulting in the prediction of maximum biomass values duringthe middle of the day, under maximum irradiances (Pinckneyand Zingmark, 1991; Guarini et al., 2000; Serodio and Cata-rino, 2000). The strong and consistent photophobic responseunder high irradiances found in this study indicates that the as-sumptions regarding the hourly pattern of productive biomassshould be revised as they probably represent an oversimplifica-tion of the real situation.
Acknowledgements
We thank Bruno Jesus for his help in the use of the spec-trometer and Vanda Brotas for commenting on the manuscript.This work was supported by projects POCTI/MAR/15318/99and POCI/BIA-BDE/61977/2004, funded by Fundac~ao paraa Ciencia e a Tecnologia. We thank three anonymousreviewers for critical comments on the manuscript.
References
Admiraal, W., 1984. The ecology of estuarine sediment-inhabiting diatoms.
Progress in Phycological Research 3, 269e323.
Bellinger, B.J., Abdullahi, A.S., Gretz, M.R., Underwood, G.J.C., 2005.
Biofilm polymers: relationship between carbohydrate biopolymers from
estuarine mudflats and unialgal cultures of benthic diatoms. Aquatic
Microbial Ecology 38, 169e180.
Bertrand, J., 1990. La vitesse de deplacement des diatomees. Diatom Research
5, 223e239.
Carrere, V., Spilmont, N., Davoult, D., 2004. Comparison of simple techniques
for estimating chlorophyll a concentration in the intertidal zone using high
spectral-resolution field-spectrometer data. Marine Ecology Progress
Series 274, 31e40.
Cohn, S.A., Bahena, M., Davis, J.T., Ragland, R.L., Rauschenberg, C.D.,
Smith, B.J., 2004. Characterisation of the diatom photophobic response
to high irradiance. Diatom Research 19, 167e179.
Cohn, S.A., Disparti, N.C., 1994. Environmental factors influencing diatom
cell motility. Journal of Phycology 30, 818e828.
Cohn, S.A., Farrell, J.F., Munro, J.D., Ragland, R.L., Weitzell, R.E.,
Wibisono, B.L., 2003. The effect of temperature and mixed species com-
position on diatom motility and adhesion. Diatom Research 18, 225e243.
Cohn, S.A., Spurck, T.P., Pickett-Heaps, J.D., 1999. High energy irradiation at
the leading tip of moving diatoms causes a rapid change of cell direction.
Diatom Research 14, 193e206.
Consalvey, M., Jesus, B., Perkins, R.G., Brotas, V., Underwood, G.J.C.,
Paterson, D.M., 2004a. Monitoring migration and measuring biomass in
benthic biofilms: the effects of dark/far-red adaptation and vertical migra-
tion on fluorescence measurements. Photosynthesis Research 81, 91e101.
Consalvey, M., Paterson, D.M., Underwood, G.J.C., 2004b. The ups and
downs of life in a benthic biofilm: migration of benthic diatoms. Diatom
Research 19, 181e202.
Decho, A., Kawaguchi, T., Allison, M., Louchard, E., Reid, R., 2003. Sedi-
ment properties influencing upwelling spectral reflectance signatures:
The ‘‘biofilm gel effect’’. Limnology and Oceanography 48, 431e443.
Eaton, J., Moss, B., 1966. The estimation of numbers and pigment content of
epipelic algal populations. Limnology and Oceanography 11, 584e595.
Guarini, J.M., Blanchard, G.F., Gros, P., Gouleau, D., Bacher, C., 2000. Dy-
namic model of the short-term variability of microphytobenthic biomass
on temperate intertidal mudflats. Marine Ecology Progress Series 195,
291e303.
Haupt, W., Scheuerlein, R., 1990. Chloroplast movement. Plant, Cell and
Environment 13, 595e614.
Honeywill, C., Paterson, D.M., Hagerthey, S.E., 2002. Determination of
microphytobenthic biomass using pulse-amplitude modulated minimum
fluorescence. European Journal of Phycology 37, 485e492.
Hopkins, J.T., 1966. The role of water in the behaviour of an estuarine mudflat
diatom. Journal of the Marine Biology Association of the United Kingdom
46, 617e626.
Kasahara, M., Kagawa, T., Iokawa, K., Suetsugu, N., Miyao, M., Wada, M.,
2002. Chloroplast avoidance movement reduces photodamage in plants.
Nature 420, 829e832.
Kingston, M.B., 1999a. Effect of light on vertical migration and photosynthe-
sis of Euglena proxima (Euglenophyta). Journal of Phycology 35,
245e253.
Kingston, M.B., 1999b. Wave effects on the vertical migration of two benthic
microalgae: Hantzschia virgata var. intermedia and Euglena proxima. Es-
tuaries 22, 81e91.
Kingston, M.B., 2002. Effect of subsurface nutrient supplies on the vertical
migration of Euglena proxima (Euglenophyta). Journal of Phycology 38,
872e880.
Kromkamp, J., Barranguet, C., Peene, J., 1998. Determination of microphyto-
benthos PSII quantum efficiency and photosynthetic activity by means of
variable chlorophyll fluorescence. Marine Ecology Progress Series 162,
45e55.
Lorenzen, C.J., 1967. Determination of chlorophyll and pheo-pigments: spec-
trophotometric equations. Limnology and Oceanography 12, 343e346.
Mitbavkar, S., Anil, A., 2004. Vertical migration rhythms of benthic diatoms
in a tropical intertidal sand flat: influence of irradiance and tides. Marine
Biology 145, 9e20.
Muller, P., Li, X.-P., Niyogi, K., 2001. Non-photochemical quenching. A re-
sponse to excess light energy. Plant Physiology 125, 1558e1566.
Murphy, R.J., Tolhurst, T.J., Chapman, M.G., Underwood, A.J., 2004.
Estimation of surface chlorophyll on an exposed mudflat using digital
colour-infrared (CIR) photography. Estuarine, Coastal and Shelf Science
59, 625e638.
Palmer, J.D., Round, F.E., 1965. Persistent, vertical-migration rhythms in
benthic microflora. I. The effect of light and temperature on the rhythmic
behaviour of Euglena obtusa. Journal of the Marine Biology Association
of the United Kingdom 45, 567e582.
Paterson, D.M., 1986. The migratory behaviour of diatom assemblages in a lab-
oratory tidal micro-ecosystem examined by low temperature scanning
electron microscopy. Diatom Research 1, 227e239.
Paterson, D.M., Wiltshire, K.H., Miles, A., Blackburn, J., Davidson, I.,
Yates, M.G., McGrorty, S., Eastwood, J., 1998. Microbiological mediation
of spectral reflectance from intertidal cohesive sediments. Limnology and
Oceanography 43, 1207e1221.
Perkins, E.J., 1960. The diurnal rhythm of the littoral diatoms of the River
Eden estuary. Journal of Ecology 48, 725e728.
Perkins, R.G., Oxborough, K., Hanlon, A.R.M., Underwood, G.J.C.,
Baker, N.R., 2002. Can fluorescence be used to estimate the rate of photo-
synthetic electron transport within microphytobenthic biofilms? Marine
Ecology Progress Series 228, 47e56.
Perkins, R.G., Underwood, G.J.C., Brotas, V., Snow, G.C., Jesus, B.,
Ribeiro, L., 2001. Responses of microphytobenthos to light: primary pro-
duction and carbohydrate allocation over an emersion period. Marine Ecol-
ogy Progress Series 223, 101e112.
Pinckney, J., Zingmark, R.G., 1991. Effects of tidal stage and sun angles on
intertidal benthic microalgal productivity. Marine Ecology Progress Series
76, 81e89.

556 J. Serodio et al. / Estuarine, Coastal and Shelf Science 68 (2006) 547e556
Pinckney, J., Zingmark, R., 1993. Modeling the annual production of the inter-
tidal benthic microalgae in estuarine ecosystems. Journal of Phycology 29,
396e407.
Pinckney, J., Piceno, Y., Lovell, C.R., 1994. Short-term changes in the vertical
distribution of benthic microalgal biomass in intertidal muddy sediments.
Diatom Research 9, 143e153.
Round, F.E., 1979. A diatom assemblage living below the surface of intertidal
sand flats. Marine Biology 54, 219e223.
Round, F.E., Palmer, J.D., 1966. Persistent, vertical-migration rhythms in ben-
thic microflora. II. Field and laboratory studies on diatoms from the banks
of the river Avon. Journal of the Marine Biology Association of the United
Kingdom 46, 191e214.
Rouse, J.W., Haas, R.H., Schell, J.A., Deering, D.W., 1973. Monitoring vege-
tation systems in the great plains with ERTS. In: Third Earth Resources
Technology Satellite Symposium, NASA SP-351, vol. 1. NASA, Washing-
ton, DC, pp. 309e317.
Serodio, J., Catarino, F., 2000. Modelling the primary productivity of intertidal
microphytobenthos: time scales of variability and effects of migratory
rhythms. Marine Ecology Progress Series 192, 13e30.
Serodio, J., Marques da Silva, J., Catarino, F., 1997. Nondestructive tracing of
migratory rhythms of intertidal benthic microalgae using in vivo chloro-
phyll a fluorescence. Journal of Phycology 33, 542e553.
Serodio, J., Marques da Silva, J., Catarino, F., 2001. Use of in vivo chlorophyll
a fluorescence to quantify short-term variations in the productive biomass of
intertidal microphytobenthos. Marine Ecology Progress Series 218, 45e61.
Serodio, J., Cruz, S., Vieira, S., Brotas, V., 2005a. Non-photochemical quench-
ing of chlorophyll fluorescence and operation of xanthophyll cycle in mi-
crophytobenthos. Journal of Experimental Marine Biology and Ecology
326, 157e169.
Serodio, J., Vieira, S., Cruz, S., Barroso, F., 2005b. Short-term variability in
the photosynthetic activity of microphytobenthos as detected by measuring
rapid light curves using variable fluorescence. Marine Biology 146,
903e914.
Ting, C.S., Owens, T.G., 1993. Photochemical and nonphotochemical fluores-
cence quenching processes in the diatom Phaeodactylum tricornutum.
Plant Physiology 101, 1323e1330.
Underwood, G.J.C., 2002. Adaptations of tropical marine microphytobenthic
assemblages along a gradient of light and nutrient availability in Suva La-
goon, Fiji. European Journal of Phycology 37, 449e462.
Underwood, G.J.C., Perkins, R.G., Consalvey, M., Hanlon, A.R.M.,
Oxborough, K., Baker, N.R., Paterson, D.M., 2005. Patterns in microphy-
tobenthic primary productivity: species-specific variation in migratory
rhythms and photosynthesis in mixed-species biofilms. Limnology and
Oceanography 50, 755e767.
Related Documents




![Characterised Control Valves with Actuators - Belimo15.04.2011].pdf · Characterised Control Valves with Actuators Version 5.1. ... Select the Characterised Control Valve according](https://static.cupdf.com/doc/110x72/5a9df4b67f8b9a29228b9d94/characterised-control-valves-with-actuators-15042011pdfcharacterised-control.jpg)






