UNCORRECTED PROOF Chapter 15 Microenvironmental Regulation of Ovarian Cancer Metastasis Maria V. Barbolina, Natalie M. Moss, Suzanne D. Westfall, Yueying Liu, Rebecca J. Burkhalter, Francoise Marga, Gabor Forgacs, Laurie G. Hudson, and M. Sharon Stack Introduction: Ovarian Carcinoma Metastasis Tumors arising from the ovarian surface epithelium (OSE) account for the vast majority of ovarian malignancies; however, the etiology of epithelial ovarian cancer (EOC) remains poorly understood, 1 and the analysis of early events in ovarian carcinogenesis is limited by the relative lack of early-stage tumors for study. The normal OSE is a single layer of mesodermally derived cells that exhibit the remarkable ability to transition between epithelial and fibroblastic phenotypes in response to microenvironmental cues. 2–4 Such phenotypic plas- ticity is usually limited to immature, regenerating, or neoplastic epithelium. Unlike most carcinomas that initially de-differentiate during neoplastic pro- gression, ovarian carcinomas undergo a mesenchymal-epithelial transition and acquire a more differentiated epithelial phenotype resulting in significant mor- phologic heterogeneity as tumors acquire increasingly complex differentiation reminiscent of the highly specialized epithelia of Mu¨llerian duct origin. 5,6 Differentiated primary ovarian tumors acquire morphologic characteristics of the fallopian tube (serous carcinoma), endometrium (endometrioid carcinoma), endocervix (mucinous carcinoma), and vagina (clear cell carcinoma). 5,6 More recently, classification of ovarian tumors into low-grade (type I) versus high- grade (type II) malignancies has been proposed based on presumed pathways leading to tumorigenesis, rather than histopathologic characteristics. 7 Low- grade carcinomas are more indolent, develop from a recognized precursor lesion, and are often confined to the ovary at diagnosis. In contrast, high- grade tumors are clinically aggressive at initial presentation, are not associated SPB-104209 15 March 30, 2009 Time: 10:17 Proof 1 M.S. Stack (*) Department of Pathology & Anatomical Sciences, University of Missouri School of Medicine, 1 Hospital Drive, M214E Medical Sciences Bldg., Columbia, MO 65212, USA e-mail: [email protected] M.S. Stack, D.A. Fishman (eds.), Ovarian Cancer, Cancer Treatment and Research 149, DOI 10.1007/978-0-387-98094-2_15, Ó Springer ScienceþBusiness Media, LLC 2009 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

UNCORRECTEDPROOF
Chapter 15
Microenvironmental Regulation of Ovarian
Cancer Metastasis
Maria V. Barbolina, Natalie M. Moss, Suzanne D. Westfall,
Yueying Liu, Rebecca J. Burkhalter, Francoise Marga, Gabor Forgacs,
Laurie G. Hudson, and M. Sharon Stack
Introduction: Ovarian Carcinoma Metastasis
Tumors arising from the ovarian surface epithelium (OSE) account for the vast
majority of ovarian malignancies; however, the etiology of epithelial ovarian
cancer (EOC) remains poorly understood,1 and the analysis of early events in
ovarian carcinogenesis is limited by the relative lack of early-stage tumors for
study. The normal OSE is a single layer of mesodermally derived cells that
exhibit the remarkable ability to transition between epithelial and fibroblastic
phenotypes in response to microenvironmental cues.2–4 Such phenotypic plas-
ticity is usually limited to immature, regenerating, or neoplastic epithelium.
Unlike most carcinomas that initially de-differentiate during neoplastic pro-
gression, ovarian carcinomas undergo a mesenchymal-epithelial transition and
acquire a more differentiated epithelial phenotype resulting in significant mor-
phologic heterogeneity as tumors acquire increasingly complex differentiation
reminiscent of the highly specialized epithelia of Mullerian duct origin.5,6
Differentiated primary ovarian tumors acquire morphologic characteristics of
the fallopian tube (serous carcinoma), endometrium (endometrioid carcinoma),
endocervix (mucinous carcinoma), and vagina (clear cell carcinoma).5,6 More
recently, classification of ovarian tumors into low-grade (type I) versus high-
grade (type II) malignancies has been proposed based on presumed pathways
leading to tumorigenesis, rather than histopathologic characteristics.7 Low-
grade carcinomas are more indolent, develop from a recognized precursor
lesion, and are often confined to the ovary at diagnosis. In contrast, high-
grade tumors are clinically aggressive at initial presentation, are not associated
SPB-104209 15 March 30, 2009 Time: 10:17 Proof 1
M.S. Stack (*)Department of Pathology & Anatomical Sciences, University of Missouri Schoolof Medicine, 1 Hospital Drive, M214E Medical Sciences Bldg., Columbia,MO 65212, USAe-mail: [email protected]
M.S. Stack, D.A. Fishman (eds.), Ovarian Cancer,Cancer Treatment and Research 149, DOI 10.1007/978-0-387-98094-2_15,� Springer ScienceþBusiness Media, LLC 2009
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44

UNCORRECTEDPROOF
with a morphologically recognizable precursor, metastasize early, and areassociated with poor clinical outcome. Molecular analyses have establisheddistinct molecular changes that distinguish type I and II tumors, supportingthe concept of two pathways of ovarian carcinogenesis.1
It is predicted that more than 15,500 women will die this year from complica-tions related to epithelial ovarian cancer metastasis.8 As 75% of women withEOC are initially diagnosed with previously disseminated intra-abdominaldisease, a more detailed understanding of the cellular and biological andbiophysical factors that promote successful metastatic dissemination clearlycan ultimately improve patient survival. In addition to genetic alterations thatpromote tumorigenesis, the contribution of the unique ovarian carcinomamicroenvironment to the development of metastatic disease is gaining increas-ing recognition. Unlike other solid tumors, hematogenous dissemination ofovarian cancer cells is uncommon. Instead, an early event in EOC metastaticdissemination is the exfoliation of cells from the primary ovarian tumor into theperitoneal cavity (Fig. 15.1A, B). Shed tumor cells are believed to block peri-toneal lymphatics9 and, together with expression of vascular endothelial growthfactor (VEGF), contribute to the build-up of peritoneal ascites.6,10 Individualtumor cells as well as multicellular aggregates (MCAs) or spheroids have beendetected in ovarian cancer ascites.11–13 Shed tumor cells interact with mesothe-lial cells lining the inner surface of the peritoneal cavity (Fig. 15.1C), whereuponcell-cell and cell-matrix adhesion molecules contribute to anchoring of tumorcells to establish secondary lesions.14,15 The dissemination of ovarian cancer islargely contained within the peritoneal cavity, establishing an unique micro-environmental niche composed of tumor cells, inflammatory components, anda host of soluble factors secreted by—or in response to—tumor cells. Theseinclude growth factors, bioactive lipids, proteolytic enzymes, extracellularmatrix components, and inflammatory mediators.16–18 The primary tumor aswell as both suspended and anchored metastatic cells maintain direct contactwith ascites, providing a mechanism for dynamic and reciprocal regulation ofthe tumor microenvironment.
Pericellular Adhesive Microenvironment
Ovarian tumor progression is accompanied by changes in the pericellularadhesive microenvironment that may reflect, in part, the unique phenotypicplasticity of the normal OSE. The OSE is composed of a single layer of simplesquamous, cuboidal, or columnar epithelium separated by a basement mem-brane from the underlying connective tissue layer, the tunica albuginea.19 OSEdisplays both epithelial and mesenchymal characteristics in situ, expressingepithelial markers such as keratin, desmosomes, and basement membrane aswell as mesenchymal markers including vimentin and interstitial collagens.2,20
Reversible modulation between epithelial and fibroblastic phenotypes occurs
SPB-104209 15 March 30, 2009 Time: 10:17 Proof 1
M.V. Barbolina et al.
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89

UNCORRECTEDPROOF
during postovulatory repair and includes alterations in cell shape, cell-cell and
cell-matrix contacts, and altered expression of differentiation markers.3,4 Simi-
larly, reversible modulation of cellular adhesive events likely plays a critical role
in remodeling of the OSE during tumor progression.Cell-matrix and cell-cell adhesion, mediated by integrins and cadherins,
respectively, are required for the maintenance of tissue integrity. Moreover,
modulation of the expression and/or function of adhesive proteins may con-
tribute to metastatic dissemination. The expression of integrins, heterodimeric
transmembrane receptors for extracellular matrix components, has not been
SPB-104209 15 March 30, 2009 Time: 10:17 Proof 1
Fig. 15.1 Schematic model of distinct adhesive microenvironments in epithelial ovarian cancer
(EOC) metastasis. (A) The primary tumor likely forms from malignant transformation ofovarian surface epithelial (OSE) cells.While the OSE expresses primarilyN-cadherin, primaryovarian tumors undergo a mesenchymal-epithelial transition and acquire expression of theepithelial cell-cell adhesion molecule E-cadherin. (B) Metastatic cells are shed from theprimary tumor as single cells and multicellular aggregates (MCAs). Formation of malignantascites is common, particularly with advanced-stage tumors. Tumor cells in ascites lackintegrin-matrix contacts, whereas cadherin-based adhesion is prevalent. (C) Peritoneal metas-tasis involves reestablishment of integrin-matrix contacts as single cells or adherent MCAsinteract with submesothelial types I and III collagens. Tumor cells proliferate to establishedanchored secondary lesions containing cell-matrix binding integrins such as a2b1 and a3b1 aswell as cell-cell binding cadherins, predominately N-cadherin. The role of this distinct adhe-sive milieu on cellular response to microenvironmental regulators including growth factors,cytokines, and bioactive lipids is under investigation
15 Microenvironmental Regulation of Ovarian Cancer Metastasis
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134

UNCORRECTEDPROOF
extensively evaluated in OSE; however several integrins including a2, a3, a5, av,b1, and b3 have been detected.21–23 Localized expression of a6b4-integrin on thebasal epithelial surface of OSE, which is lost in progression to EOC, has alsobeen reported.23,24 Adhesion mediated by b1-integrins may represent a poten-tial therapeutic target, as this integrin subunit has been implicated in binding ofindividual tumor cells and MCAs to peritoneal mesothelium or submesothelialmatrix and promoting MCA disaggregation11,12,25–31
In marked contrast with other epithelia that express E-cadherin, normalOSE epithelial cell-cell integrity is maintained by N-cadherin, and moderateN-cadherin staining of some EOC tumors has been reported (reviewed inHudson et al.14). The epithelial cell-cell adhesion molecule E-cadherin is largelyabsent in OSE yet becomes more abundant in primary differentiated EOC,suggesting a role in early events leading to cellular transformation.14 ReducedE-cadherin staining is found in late-stage carcinomas and ascites-derived tumorcells,32–34 whereasN-cadherin immunoreactivity is elevated in advanced tumorsand peritoneal metastases.14 This reacquisition of mesenchymal-type adhesivefeatures in advanced tumors may change the morphology of MCAs by redis-tribution (sorting) of E- and N-cadherin presenting cells, thereby modifying thebiomechanical properties of the aggregate (such as surface tension)35–37 andcontributing to intraperitoneal anchoring of metastatic lesions.
Reversible changes in the expression and/or function of cellular adhesiveproteins occur during ovarian tumor progression (Fig. 15.1). Initial dissemina-tion of cells from the primary tumor on the ovarian surface requires disruptionof both cadherin-mediated cell-cell contacts and integrin-matrix interactions.Tumor cells in ascites lack integrin-matrix contacts, yet cadherin-based adhe-sion is prevalent in MCAs. Integrin-matrix adhesion is reestablished duringperitoneal anchoring, and subsequent tumor cell proliferation generatescadherin-expressing metastatic colonies. The influence of this reversible mod-ulation in the pericellular adhesive milieu on tumor progression and metastaticcompetence is largely unexplored. Furthermore, the adhesive milieu may alsoalter the response of tumor cells to soluble and/or matrix-associated micro-environmental regulators such as growth factors, bioactive lipids, and ECMproteins.
Microenvironment of Ascitic Tumor Cells
The peritoneal cavity of normal women contains 5–20 ml of serous exudateincluding many plasma proteins and free-floating cells such as macrophages,desquamated mesothelial cells, lymphocytes, and mast cells.38 In women withovarian cancer, obstruction of peritoneal lymphatics together with enhancedvascular permeability often results in abdominal distension caused by accumu-lated malignant ascites fluid in the peritoneal cavity ranging from <500 ml to>2 L. Whereas women with early-stage malignancies (stage I and II) are often
SPB-104209 15 March 30, 2009 Time: 10:17 Proof 1
M.V. Barbolina et al.
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
UNCORRECTEDPROOF
free of ascites, the vast majority of women with advanced disease (stage III/IV)
produced >500 ml of ascites.39 In women with advanced ovarian cancer, the
presence of ascites is an independent adverse prognostic factor40 and is corre-
lated with both intraperitoneal and retroperitoneal tumor spread.41 Together,
these data indicate that presence of ascites is highly predictive of ovarian
malignancy in women with a pelvic mass.39
The complexity of ascites fluid is highlighted by a recent proteomic analysis
that identifiedmore than 200 proteins in the soluble fraction of ascites andmore
than 2500 in the combined soluble and cellular fractions.42 Soluble components
include the bioactive lipid lysophosphatidic acid (LPA), synthesized by both
platelets and activated mesothelial cells,43,44 cytokines including interleu-
kins17,45 and macrophage migration inhibitory factor,46 growth factors such
as EGF, VEGF, and HB-EGFAQ1 ,47–50 and newly synthesized as well as degraded
ECM proteins such as collagens types I and III.51–53 These soluble factors can
have dramatic consequences on tumor cell behavior and metastatic progres-
sion. For example, LPA can directly interact with tumor cell LPA receptors54 to
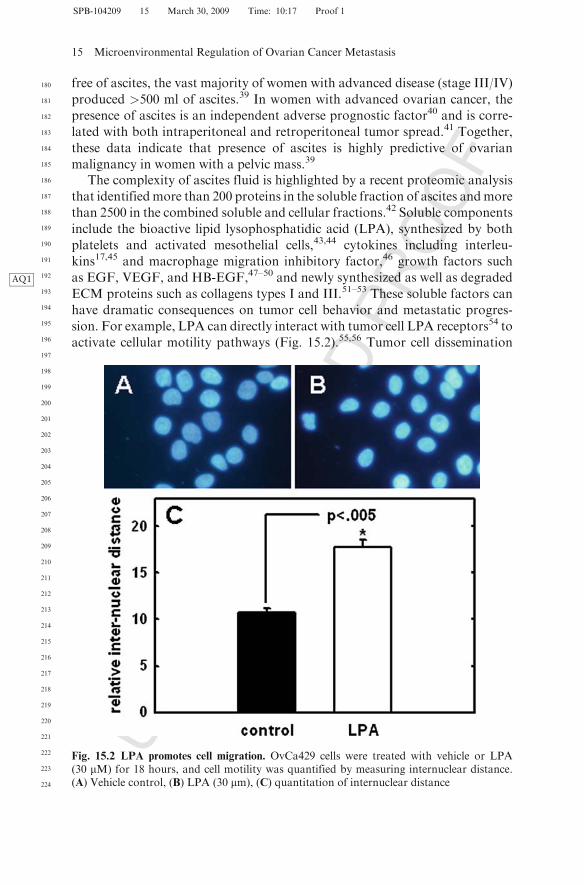
activate cellular motility pathways (Fig. 15.2).55,56 Tumor cell dissemination
SPB-104209 15 March 30, 2009 Time: 10:17 Proof 1
Fig. 15.2 LPA promotes cell migration. OvCa429 cells were treated with vehicle or LPA(30 mM) for 18 hours, and cell motility was quantified by measuring internuclear distance.(A) Vehicle control, (B) LPA (30 mm), (C) quantitation of internuclear distance
15 Microenvironmental Regulation of Ovarian Cancer Metastasis
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
223
224

UNCORRECTEDPROOF
may be promoted through other mechanisms such as regulating the expressionand/or activation of metastasis-associated proteinases in the matrix metallo-proteinase (MMP) and plasminogen activator families.54,55,57 Alternatively,LPA may also influence tumor progression through altered expression ofsecondary regulators of tumor cell behavior including interleukins andchemokines.45,58–60
Cellular components including activated mesothelial cells and inflammatorycells are also prevalent in ascites. Stromal cells in the tumor microenvironmentmay contribute to regulation of EOC metastasis, and recent studies support arole for inflammatory cells in promotion of intraperitoneal disease. Tumor celldissemination from the ovaries was found to correlate temporally withenhanced peritoneal inflammation61 and direct intraperitoneal injection oftumor cells also showed a prometastatic effect on inflammation. Depletion ofperitoneal macrophages reducedmetastasis, suggesting that inflammation facil-itates metastasis via amacrophage-mediatedmechanism.61 This is supported bydata demonstrating that co-culture of ovarian cancer cells with macrophagesled to dynamic changes in macrophage expression of IL-10, -12, and -6, TNF-a,and CSF-1AQ2 .47,62 Macrophage-secreted TNF-a increased tumor cell invasive-ness,47 providing an example of stromal-epithelial cross-talk in the control ofEOC metastasis via dynamic regulation of the cytokine network.
Does Cadherin Expression Contribute to Intraperitoneal Survival?
A unique feature of tumor cells in ascites is the ability to survive the lack of cell-matrix contact and proliferate as a floating tumor population (Fig. 15.1B). It isinteresting to speculate that these conditions favor the survival of a subpopula-tion composed of highly neoplastic cells.63 In support of this hypothesis, recentstudies have demonstrated that cells isolated from murine ascites proliferatemore readily and are more aggressive relative to the parental cells when re-injected in vivo.64 In gene expression profiling studies comparing cells culturedas monolayers, MCAs, or tumor xenografts, MCA expression profiles werefound to cluster with tumor xenografts rather than with monolayer cells.65
These data suggest that MCAs represent a more advanced stage of malignancyrelative to the parental primary tumor cells. A self-renewing population ofovarian cancer initiating cells that form self-renewing spheroids was recentlyisolated from solid primary ovarian tumors.66 When cultured under stem cellselective conditions, these MCAs displayed a 100,000-fold increase in tumor-igenicity, resulting in tumors that metastasized to the omentum and colon.These data support the hypothesis that the nonadherent MCA populationfound in human ovarian ascites is the primary source of intraperitoneal meta-static lesions.
Although survival of single cells and MCAs in ascites is likely influenced byready access to soluble growth factors,48,49 recent data support a role for
SPB-104209 15 March 30, 2009 Time: 10:17 Proof 1
M.V. Barbolina et al.
225
226
227
228
229
230
231
232
233
234
235
236
237
238
239
240
241
242
243
244
245
246
247
248
249
250
251
252
253
254
255
256
257
258
259
260
261
262
263
264
265
266
267
268
269

UNCORRECTEDPROOF
cadherin expression in MCA survival. MCAs in human ovarian carcinomatousascites are relatively heterogeneous, ranging in size from 30 to 200 mm.11,12 Assummarized above, primary ovarian carcinomas gain expression of the cell-celladhesion molecule E-cadherin relative to unaffected ovarian surface epithe-lium, as tumor cells undergo an initial mesenchymal-epithelial transition inearly carcinogenesis.14 It has been suggested that acquisition of E-cadherin–mediated adhesion early in tumor progression functions to suppress anoikis, atype of programmed cell death resulting from loss of integrin-based cell-matrixcontacts as cells are shed from the primary tumor into the peritoneal cavity.14
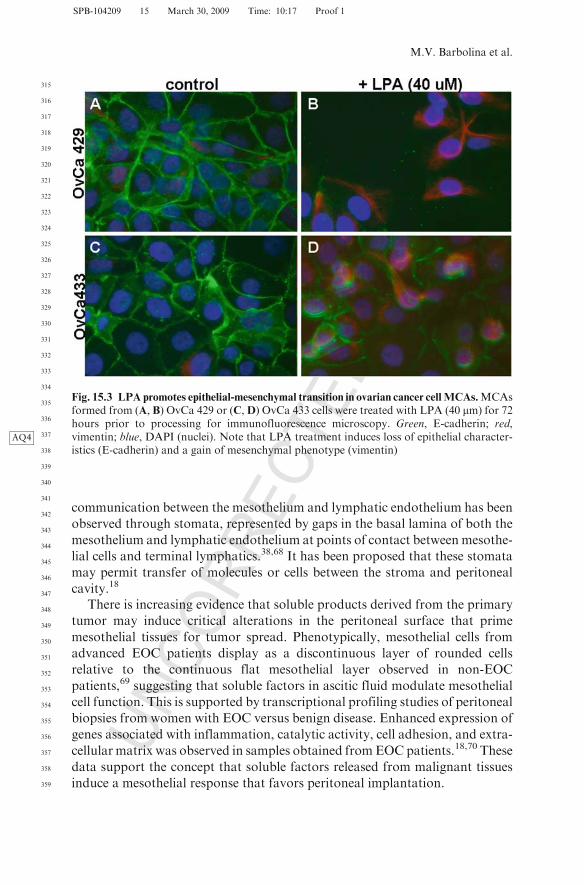
This is supported by data showing that E-cadherin engagement activates Aktvia PI3AQ3 -kinase signaling,66 and downregulation of E-cadherin leads to loss ofviability.66 Thus, proliferative signals downstream of E-cadherin activationmay contribute to ovarian cancer cell survival. Whereas reduced E-cadherinstaining is often found in late-stage carcinomas and ascites-derived tumor cells,N-cadherin staining persists in late-stage tumors and paired peritoneal metas-tases14; however, the role of N-cadherin in MCA survival has not beenexplored. Interestingly, N-cadherin expression levels of glioblastoma cells cul-tured as MCAs correlate with surface tension of the aggregate, indicative ofstronger aggregate cohesion.67 Furthermore, the effect of microenvironmentalregulators such as bioactive lipids, cytokines, growth factors, and matrix frag-ments on MCA survival and dissemination has not been extensively investi-gated. In this context, it is interesting to note that treatment of ovarian cancerMCA cultures with LPA induces expression of the mesenchymal marker vimen-tin (Fig. 15.3), suggesting that LPA may be a modulator of the late epithelial-mesenchymal transition (EMT) observed in progression to metastasis.14
Microenvironment of Peritoneal Metastatic Lesions
A major structural component of the ovarian cancer microenvironment is theperitoneum, a large membranous sheet that covers the abdominal organs andviscera. Involvement of the peritoneum or serosal surfaces is observed in at least80% of women with EOC18 and is a significant predictor of poor prognosis.Whereas women whose tumors lack peritoneal involvement have a 5-yearsurvival of 90%, survival is decreased to 20% over 5 years in women withmicroscopic or visible metastatic foci.18
The peritoneum is composed of a single layer of mesothelial cells withabundant long microvilli that form an extensive serous membrane withapproximately the same surface area as the skin.38 An associated basementmembrane overlays a stromal compartment composed predominately of aninterstitial (types I, III) collagen-based matrix. Elastic fibers, fibronectin, lami-nin, glycosaminoglycans, fibroblast-like cells, lymphatics, nerves, and capil-laries are also found here.68 Tight junctions and desmosomes connect peritonealmesothelial cells; however, intercellular spaces are also present.18,68 Direct
SPB-104209 15 March 30, 2009 Time: 10:17 Proof 1
15 Microenvironmental Regulation of Ovarian Cancer Metastasis
270
271
272
273
274
275
276
277
278
279
280
281
282
283
284
285
286
287
288
289
290
291
292
293
294
295
296
297
298
299
300
301
302
303
304
305
306
307
308
309
310
311
312
313
314
UNCORRECTEDPROOF
communication between the mesothelium and lymphatic endothelium has been
observed through stomata, represented by gaps in the basal lamina of both the
mesothelium and lymphatic endothelium at points of contact between mesothe-
lial cells and terminal lymphatics.38,68 It has been proposed that these stomata
may permit transfer of molecules or cells between the stroma and peritoneal
cavity.18
There is increasing evidence that soluble products derived from the primary
tumor may induce critical alterations in the peritoneal surface that prime
mesothelial tissues for tumor spread. Phenotypically, mesothelial cells from
advanced EOC patients display as a discontinuous layer of rounded cells
relative to the continuous flat mesothelial layer observed in non-EOC
patients,69 suggesting that soluble factors in ascitic fluid modulate mesothelial
cell function. This is supported by transcriptional profiling studies of peritoneal
biopsies from women with EOC versus benign disease. Enhanced expression of
genes associated with inflammation, catalytic activity, cell adhesion, and extra-
cellular matrix was observed in samples obtained fromEOC patients.18,70 These
data support the concept that soluble factors released from malignant tissues
induce a mesothelial response that favors peritoneal implantation.
SPB-104209 15 March 30, 2009 Time: 10:17 Proof 1
Fig. 15.3 LPA promotes epithelial-mesenchymal transition in ovarian cancer cellMCAs.MCAsformed from (A, B) OvCa 429 or (C,D) OvCa 433 cells were treated with LPA (40 mm) for 72hours prior to processing for immunofluorescence microscopy. Green, E-cadherin; red,vimentin; blue, DAPIAQ4 (nuclei). Note that LPA treatment induces loss of epithelial character-istics (E-cadherin) and a gain of mesenchymal phenotype (vimentin)
M.V. Barbolina et al.
315
316
317
318
319
320
321
322
323
324
325
326
327
328
329
330
331
332
333
334
335
336
337
338
339
340
341
342
343
344
345
346
347
348
349
350
351
352
353
354
355
356
357
358
359

UNCORRECTEDPROOF
Metastatic ovarian tumors arise as a consequence of CD44- and integrin-mediated intraperitoneal adhesion and localized invasion into theinterstitial collagen-rich submesothelial matrix to anchor secondary lesions(Fig. 15.1).11,12,15,27,28,71,72 After attachment of disseminated ovarian tumorcells to the peritoneal mesothelium, cells extend cytoplasmic processes throughthe junctional margins of neighboringmesothelial cells, inducing cellular retrac-tion and exposure of the underlying submesothelial ECM.73,74 Metastasizingovarian cancer cells encounter an interstitial collagen-rich environment, as thesubmesothelial matrix is composed primarily of types I and III col-lagen.51,68,73,74 Ovarian cancer cells adhere preferentially to interstitial collagesusing a2b1- and a3b1-integrins.27–29 Such affinity for interstitial collagens likelyreflects the phenotypic plasticity andmesenchymal origin of the ovarian surfaceepithelium.53,75,76 Matrix binding induces MCA dissociation, as multivalentcell-matrix contacts replace cell-cell adhesive interactions.11–13,77 b1-integrin–mediated submesothelial adhesion represents an important early event uniqueto ovarian cancer metastatic dissemination,12,13,27–29,30,77 and the resultingalterations in integrin signaling may contribute to metastatic success.
Integrin-Matrix Interaction in Peritoneal Metastasis
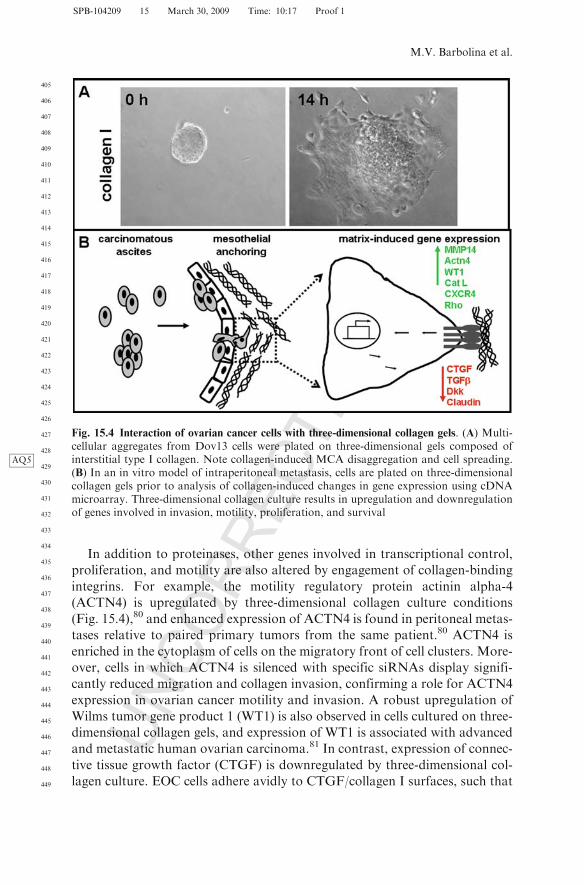
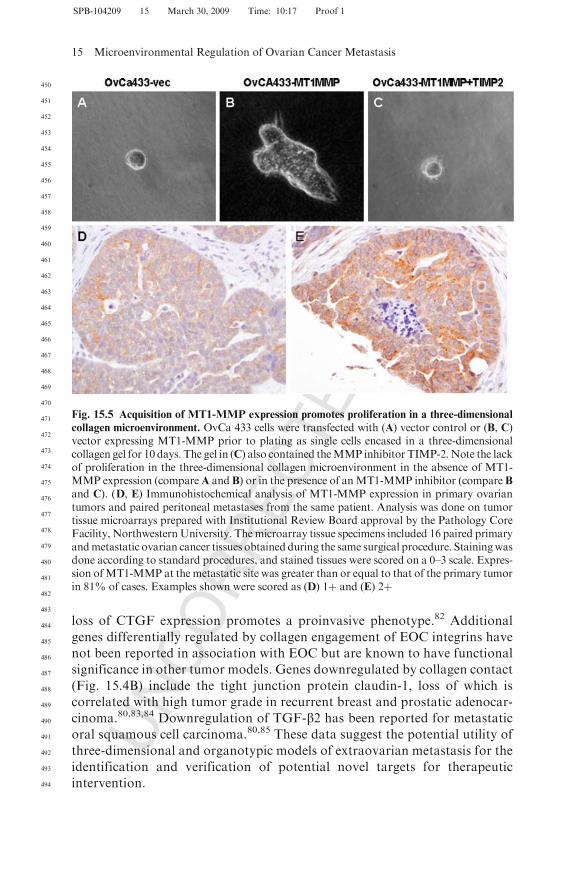
There is increasing evidence that the peritoneal microenvironment activelypromotes progression of metastatic ovarian carcinoma.15 Ovarian cancer cellsthat encounter this matrix barrier respond to de novo integrin signaling, result-ing in changes in gene expression that may ultimately potentiate metastasis(Fig. 15.4B). Using three-dimensional cell culture systems to model adhesiveevents in intraperitoneal metastasis, our laboratory has shown that aggregationof collagen-binding integrins activates integrin-mediated signaling via Srckinases to induce expression of the early growth response gene Egr-1.30 Theresulting Egr-1–mediated transcriptional activation of the membrane-type 1matrix metalloproteinase (MT1-MMP) promoter stimulates expression ofMT1-MMP, leading to enhanced collagen invasion.29,77 Acquisition of MT1-MMP collagenolytic activity may be key to metastatic success, as it was recentlyreported that MT1-MMP–driven collagenolysis is necessary to remove matrixbarriers to allow for the cytoskeletal reorganization necessary to drive a pro-liferative response.78 Indeed, the ability of ovarian cancer cells to survive long-term and proliferate in three-dimensional collagen gels is substantiallyenhanced by MT1-MMP expression (Fig. 15.5A–C), and MT1-MMP expres-sion is prevalent in peritoneal metastases from ovarian cancer patients(Fig. 15.5D, E). Additional metastasis-associated proteinases such as MMP-9are also induced by integrin-collagen contact.79 MMP-9 participates in E-cadherin ectodomain shedding, a process that likely contributes to disaggrega-tion of cells from the primary tumor or ascitic MCAs to facilitate subsequentintraperitoneal anchoring and metastatic dissemination.79
SPB-104209 15 March 30, 2009 Time: 10:17 Proof 1
15 Microenvironmental Regulation of Ovarian Cancer Metastasis
360
361
362
363
364
365
366
367
368
369
370
371
372
373
374
375
376
377
378
379
380
381
382
383
384
385
386
387
388
389
390
391
392
393
394
395
396
397
398
399
400
401
402
403
404
UNCORRECTEDPROOF
In addition to proteinases, other genes involved in transcriptional control,
proliferation, and motility are also altered by engagement of collagen-binding
integrins. For example, the motility regulatory protein actinin alpha-4
(ACTN4) is upregulated by three-dimensional collagen culture conditions
(Fig. 15.4),80 and enhanced expression of ACTN4 is found in peritoneal metas-
tases relative to paired primary tumors from the same patient.80 ACTN4 is
enriched in the cytoplasm of cells on the migratory front of cell clusters. More-
over, cells in which ACTN4 is silenced with specific siRNAs display signifi-
cantly reduced migration and collagen invasion, confirming a role for ACTN4
expression in ovarian cancer motility and invasion. A robust upregulation of
Wilms tumor gene product 1 (WT1) is also observed in cells cultured on three-
dimensional collagen gels, and expression of WT1 is associated with advanced
and metastatic human ovarian carcinoma.81 In contrast, expression of connec-
tive tissue growth factor (CTGF) is downregulated by three-dimensional col-
lagen culture. EOC cells adhere avidly to CTGF/collagen I surfaces, such that
SPB-104209 15 March 30, 2009 Time: 10:17 Proof 1
Fig. 15.4 Interaction of ovarian cancer cells with three-dimensional collagen gels. (A) Multi-cellular aggregates from Dov13 cells were plated on three-dimensional gels composed ofinterstitial type I collagenAQ5 . Note collagen-induced MCA disaggregation and cell spreading.(B) In an in vitro model of intraperitoneal metastasis, cells are plated on three-dimensionalcollagen gels prior to analysis of collagen-induced changes in gene expression using cDNAmicroarray. Three-dimensional collagen culture results in upregulation and downregulationof genes involved in invasion, motility, proliferation, and survival
M.V. Barbolina et al.
405
406
407
408
409
410
411
412
413
414
415
416
417
418
419
420
421
422
423
424
425
426
427
428
429
430
431
432
433
434
435
436
437
438
439
440
441
442
443
444
445
446
447
448
449
UNCORRECTEDPROOF
loss of CTGF expression promotes a proinvasive phenotype.82 Additional
genes differentially regulated by collagen engagement of EOC integrins have
not been reported in association with EOC but are known to have functional
significance in other tumor models. Genes downregulated by collagen contact
(Fig. 15.4B) include the tight junction protein claudin-1, loss of which is
correlated with high tumor grade in recurrent breast and prostatic adenocar-
cinoma.80,83,84 Downregulation of TGF-b2 has been reported for metastatic
oral squamous cell carcinoma.80,85 These data suggest the potential utility of
three-dimensional and organotypic models of extraovarian metastasis for the
identification and verification of potential novel targets for therapeutic
intervention.
SPB-104209 15 March 30, 2009 Time: 10:17 Proof 1
Fig. 15.5 Acquisition of MT1-MMP expression promotes proliferation in a three-dimensional
collagen microenvironment. OvCa 433 cells were transfected with (A) vector control or (B, C)vector expressing MT1-MMP prior to plating as single cells encased in a three-dimensionalcollagen gel for 10 days. The gel in (C) also contained theMMP inhibitor TIMP-2.Note the lackof proliferation in the three-dimensional collagen microenvironment in the absence of MT1-MMP expression (compareA and B) or in the presence of anMT1-MMP inhibitor (compare Band C). (D, E) Immunohistochemical analysis of MT1-MMP expression in primary ovariantumors and paired peritoneal metastases from the same patient. Analysis was done on tumortissue microarrays prepared with Institutional Review Board approval by the Pathology CoreFacility, NorthwesternUniversity. Themicroarray tissue specimens included 16 paired primaryandmetastatic ovarian cancer tissues obtained during the same surgical procedure. Stainingwasdone according to standard procedures, and stained tissues were scored on a 0–3 scale. Expres-sion ofMT1-MMP at the metastatic site was greater than or equal to that of the primary tumorin 81% of cases. Examples shown were scored as (D) 1þ and (E) 2þ
15 Microenvironmental Regulation of Ovarian Cancer Metastasis
450
451
452
453
454
455
456
457
458
459
460
461
462
463
464
465
466
467
468
469
470
471
472
473
474
475
476
477
478
479
480
481
482
483
484
485
486
487
488
489
490
491
492
493
494

UNCORRECTEDPROOF
Conclusion
An early event in ovarian cancer metastasis is shedding of cells from the primarytumor into the peritoneal cavity as anchorage-independent MCAs that survive insuspension (ascites) and are exposed to specific microenvironmental cues thatpromote metastatic implantation into the submesothelial matrix to anchor sec-ondary metastatic lesions throughout the peritoneal cavity. This is distinct frommost other solid tumors that metastasize hematogenously and presents a distinctset of therapeutic challenges. In most carcinomas, metastasis is associated with adysregulated adhesion phenotype, typically characterized by a loss of epithelial(E)-cadherin. In contrast, EOC is unusual because early-stage disease reflects aninitial gain of epithelial characteristics (mesenchymal-epithelial transition, orMET), including E-cadherin expression. It is becoming apparent that MET andreestablishment of epithelial characteristics is important to later stages of metas-tasis; however, the MET of ovarian cancer occurs early in disease progression,suggesting that this event confers a competitive advantage. Most cells that losetheir contacts to the extracellular matrix undergo apoptosis (anoikis) and die,however EOC cells survive in suspension (ascites). It is generally assumed that thecell-cell contacts (cadherins) that hold MCAs together also help to ensure tumorcell survival. The role of additional soluble, cellular, and matrix factors in theovarian tumor microenvironment in promoting MCA survival, reversion to amesenchymal phenotype (EMT), and ultimate metastatic success is currentlyunder active investigation. A molecular-level understanding of how tumor cellsmetastasize is necessary for the development of novel therapies to inhibit intraper-itoneal spread and thereby improve the long-term survival of thousands of womenwith EOC. Development of models that accurately reflect the metastatic compe-tence of EOC presents a remarkable scientific challenge, because the EOC meta-static microenvironment involves a novel shedding of MCAs into a cavity asanchorage-independent, chemotherapy-resistant spheroids. In this unique niche,it is currently unknown what regulates the transition from free-floating MCA toanchoredmetastatic lesion. Understanding this transition will enable novel meansof targeting intraperitoneal therapies to appropriate multicellular populations.
Acknowledgments This work was supported by grants from the 2005–2006 and 2007–2008Penny Severns Breast, Cervical and Ovarian Cancer Fund, Illinois Department of PublicHealth (M.V.B.), the Ovarian Cancer Research Foundation Program of Excellence (M.V.B.),National Science Foundation NSF-0526854 (G.F.), and National Institutes of Health/National Cancer Institute Research Grants RO1CA86984 (M.S.S.), RO1CA086984-S1(N.M.), and RO1CA109545 (M.S.S. and L.G.H.).
References
1. Landen CN, Birrer MJ, Sood AK. Early events in the pathogenesis of epithelial ovariancancer. J Clin Oncol. 2008;26:995–1005.
2. Wong AST, Auersperg N. Normal ovarian surface epithelium. Cancer Treat Res.2002;107:161–183.
SPB-104209 15 March 30, 2009 Time: 10:17 Proof 1
M.V. Barbolina et al.
495
496
497
498
499
500
501
502
503
504
505
506
507
508
509
510
511
512
513
514
515
516
517
518
519
520
521
522
523
524
525
526
527
528
529
530
531
532
533
534
535
536
537
538
539

UNCORRECTEDPROOF
3. Darai E, Scoazec JY, Walker-Combrouze F, et al. Expression of cadherins in benign,borderline andmalignant ovarian epithelial tumors: a clinicopathologic study of 60 cases.Hum Pathol. 1997;28:922–928.
4. Alper O, DeSantis ML, Stromberg K, et al. Anti-sense suppression of epidermal growthfactor receptor expression alters cellular proliferation, cell adhesion and tumorigenicity inovarian cancer cells. Int J Cancer. 2000;88:566–574.
5. Auersperg N, Maines-Bandiera S, Dyck HG, et al. Characterization of cultured humanovarian surface epithelial cells: phenotypic plasticity and premalignant changes. LabInvest. 1994;71:510–518.
6. Naora H. The heterogeneity of epithelial ovarian cancers: reconciling old and newparadigms. Expert Rev Mol Med. 2007;9:1–12.
7. Shih EM, Kurman RJ. Ovarian tumorigenesis: a proposed model based on morphologi-cal and molecular genetic analysis. Am J Pathol. 2004;164:1511–1518.
8. ACS Cancer Facts & Figures. Available at: www.cancer.org.9. Hoskins WJ. Prospective on ovarian cancer; why prevent? J Cell Biochem Suppl.
1995;23:189–199.10. Ghosh S, Wu Y, Stack MS. Ovarian cancer-associated proteinases. Cancer Treat Res.
2002;107:331–354.11. Burleson KM, Hansen LK, Skubitz APN. Ovarian carcinoma spheroids disaggregate on
type I collagen and invade live human mesothelial cell monolayers. Clin Exp Metastasis.2004;21:685–697.
12. Burleson KM, Casey RC, Skubitz KM, et al. Ovarian carcinoma ascites spheroids adhereto extracellular matrix components and mesothelial cell monolayers. Gynecol Oncol.2004;93:170–181.
13. BurlesonKM, BoenteMP, Pambuccian SE, et al. Disaggregation and invasion of ovariancarcinoma ascites spheroids. J Transl Med. 2006;4:1–16.
14. Hudson LG, Zeineldin R, StackMS. Phenotypic plasticity of neoplastic ovarian epithelium:unique cadherin profiles in tumor progression. Clin Exp Metastasis. 2008;25:L643–L655.
15. KennyHA,Krausz T, Yamada SD, et al. Use of a novel 3D culture model to elucidate therole of mesothelial cells, fibroblasts and extracellular matrices on adhesion and invasionof ovarian cancer cells to the omentum. Int J Cancer. 2007;121:1463–1472.
16. Offner FA, Obrist P, Stadlmann S, et al. IL6 secretion by human peritoneal mesothelialand ovarian cancer cells. Cytokine. 1995;7:542–547.
17. Mustea A, Pirvulescu C, Konsgen D, et al. Decreased IL1 RA concentration in ascites isassociated with a significant improvement in overall survival in ovarian cancer. Cytokine.2008;42:77–84.
18. FreedmanRS,DeaversM, Liu J, et al. Peritoneal inflammation – amicroenvironment forepithelial ovarian cancer. J Transl Med. 2004;2:23–33.
19. Papadaki L, Beilby JOW. The fine structure of the surface epithelium of the human ovary.J Cell Sci. 1971;8:445–465.
20. Czernobilsky B, Moll R, Levy M, et al. Co-expression of cytokeratin and vimentinfilaments in mesothelial, granulosa and rete ovarii cells of the human ovary. Eur J CellBiol. 1985;37:175–190.
21. Kruk PA, Uitto VJ, Firth JD, et al. Reciprocal interactions between human ovariansurface epithelial cells and adjacent extracellular matrix. Exp Cell Res. 1994;215:97–108.
22. Carreiras F, Denoux Y, Staedel C, et al. Expression and localization of alphav integrinsand their ligand vitronectin in normal ovarian epithelium and in ovarian carcinoma.Gynecol Oncol. 1996;62:260–267.
23. Skubitz AP, Bast RC, Wayner EA, et al. Expression of alpha 6 and beta 4 integrins inserous ovarian carcinoma correlates with expression of the basement membrane proteinlaminin. Am J Pathol. 1996;148:1445–1461.
24. Bridges JE, Englefield P, Boyd IE, et al. Expression of integrin adhesion molecules innormal ovary and epithelial ovarian tumors. Int J Gynecol Cancer. 1995;5:187–192.
SPB-104209 15 March 30, 2009 Time: 10:17 Proof 1
15 Microenvironmental Regulation of Ovarian Cancer Metastasis
540
541
542
543
544
545
546
547
548
549
550
551
552
553
554
555
556
557
558
559
560
561
562
563
564
565
566
567
568
569
570
571
572
573
574
575
576
577
578
579
580
581
582
583
584

UNCORRECTEDPROOF
25. Shield K, Riley C, Quinn MA, et al. Alpha2beta1 integrin affects metastatic potential ofovarian carcinoma spheroids by supporting disaggregation and proteolysis. J Carcinog.2007;14:6–11.
26. Sawada K, Mitra AK, Radjabi AR, et al. Loss of E-cadherin promotes ovarian cancermetastasis via alpha5 integrin, which is a therapeutic target. Cancer Res. 2008;68:2329–2339.
27. Moser TL, Pizzo SV, Bafetti LM, et al. Evidence for preferential adhesion of ovarianepithelial carcinoma cells to type I collagen mediated by the alpha2beta1 integrin. IntJ Cancer. 1996;67:695–701.
28. FishmanDA, Kearns AM, Chilikuri K, et al. Metastatic dissemination of human ovarianepithelial carcinoma is promoted by a2b1 integrin–mediated interaction with typeI collagen. Invasion Metastasis. 1998;18:15–26.
29. Ellerbroek SM, Fishman DA, Kearns AS, et al. Ovarian carcinoma regulation of matrixmetalloproteinase-2 and membrane type 1 matrix metalloproteinase through beta1 integ-rin. Cancer Res. 1999;59:1635–1641.
30. BarbolinaMV, Adley BP, Ariztia EV, et al.Microenvironmental regulation of membranetype 1 matrix metalloproteinase activity in ovarian carcinoma cells via collagen-inducedEGR1 expression. J Biol Chem. 2007;282:4924–4931.
31. Davidson B, Goldberg I, Reich R, et al. AlphaV and beta1-integrin subunits are com-monly expressed in malignant effusions from ovarian carcinoma patients. Gynecol Oncol.2003;90:248–257.
32. DaviesBR,WorsleySD,PonderBA.ExpressionofE-cadherin, alpha-catenin andbeta-cateninin normal ovarian surface epithelium and ovarian cancers.Histopathology. 1998;32:69–80.
33. Ho EY, Choi Y, Chae SW, et al. Immunohistochemical study of the expression ofadhesion molecules in ovarian serous neoplasms. Pathol Int. 2006;56:62–70.
34. Voutilainen KA, Anttila MA, Sillanpaa SM, et al. Prognostic significance of E-cadherin–catenin complex in epithelial ovarian cancer. J Clin Pathol. 2006;59:460–467.
35. ForgacsG, Foty RA, Shafrir Y, et al. Viscoelastic properties of living embryonic tissues: aquantitative study. Biophys J. 1998;74:2227–2234.
36. FotyRA, Pfleger Cm, Forgacs G, et al. Surface tensions of embryonic tissues predict theirmutual envelopment behavior. Development. 1996;122:1611–1620.
37. Duguay D, Foty RA, SteinbergMS. Cadherin-mediated cell adhesion and tissue segrega-tion: qualitative and quantitative determinants. Dev Biol. 2003;253:309–323.
38. diZerega GS, Rodgers KE. The Peritoneum. New York: Springer-Verlag; 1992.39. Shen-Gunther J, Mannel RS. Ascites as a predictor of ovarian malignancy. Gynecol
Oncol. 2002;87:77–83.40. Rahaman J, Cohen CJ. Impact of ascites on survival in advanced ovarian cancer. Proc
Am Soc Clin Oncol. 2002;21:837.41. Ayhan A, Gultekin M, Taskiran C, et al. Ascites and epithelial ovarian cancers: a
reappraisal with respect to different aspects. Int J Gynecol Cancer. 2006;17:68–75.42. Gortzak-Uzan L, Ignatchenko A, Evangelou AI, et al. A proteome resource of ovarian
cancer ascites: integrated proteomic and bioinformatics analyses to identify putativebiomarkers. J Proteome Res. 2008;7:339–351.
43. Xu Y, Gaudette DC, Boynton JD, et al. Characterization of an ovarian cancer activatingfactor in ascites from ovarian cancer patients. Clin Cancer Res. 1995;1:1223–1232.
44. Xu Y, Shen Z, Wiper DW, et al. Lysophosphatidic acid as a potential biomarker forovarian and other gynecologic cancers. JAMA. 1998;280:719–723.
45. Fang X, Yu S, Bast RC, et al. Mechanisms for lysophosphatidic acid-induced cytokineproduction in ovarian cancer cells. J Biol Chem. 2004;279:9653–9661.
46. Hagemann T,Wilson J, Burke F, et al. Ovarian cancer cells polarize macrophages towarda tumor-associated phenotype. J Immunol. 2006;176:5023–5032.
47. Reddy P, Liu L, Ren C, et al. Formation of E-cadherin mediated cell–cell adhesionactivates AKT andMAPKvia PI3 kinase and ligand-independent activation of epidermalgrowth factor receptor in ovarian cancer cells. Mol Endocrinol. 2005;19:2564–2578.
SPB-104209 15 March 30, 2009 Time: 10:17 Proof 1
M.V. Barbolina et al.
585
586
587
588
589
590
591
592
593
594
595
596
597
598
599
600
601
602
603
604
605
606
607
608
609
610
611
612
613
614
615
616
617
618
619
620
621
622
623
624
625
626
627
628
629

UNCORRECTEDPROOF
48. Hudson LG,AQ6 Zeineldin R, SilberbergM, et al. Activated epidermal growth factor receptorin ovarian cancer. Cancer Treat Res. 2009.
49. Lafky JM, Wilken JA, Baron AT, et al. Clinical implications of the ErbB/epidermalgrowth factor (EGF) receptor family and its ligands in ovarian cancer. Biochim BiophysActa. 2008;1785:232–265.
50. Said N, Socha MJ, Olearczyk JJ, et al. Normalization of the ovarian cancer microenvir-onment by SPARC. Mol Cancer Res. 2007;5:1015–1030.
51. Santala M, Risteli J, Risteli L, et al. Synthesis and breakdown of fibrillar collagens:concomitant phenomena in ovarian cancer. Br J Cancer. 1998;77:1825–1831.
52. Abdel Axia MT, Abdel Aziz Wassef M, Kamel M, et al. Clinical evaluation of serumaminoterminal propeptide of type III procollagen as tumor marker in gynecologic malig-nancies. Tumori. 1993;79:219–223.
53. Zhu GG, Risteli J, Puistola U, et al. Progressive ovarian carcinoma induces synthesis oftype I and type III procollagens in the tumor tissue and peritoneal cavity. Cancer Res.1993;53:5028–5032.
54. Mills GB, Eder A, Fang X, et al. Critical role of lysophospholipids in the pathophysiol-ogy, diagnosis, and management of ovarian cancer. Cancer Res Treat. 2002;107:259–284.
55. Do TV, Symowicz JC, Berman DM, et al. Lysophosphatidic acid down-regulates stressfibers and up-regulates pro-matrix metalloproteinase-2 activation in ovarian cancer cells.Mol Cancer Res. 2007;5:121–131.
56. Symowicz J, Adley BP, Woo MM, et al. Cyclooxygenase-2 functions as a downstreammediator of lysophosphatidic acid to promote aggressive behavior in ovarian carcinomacells. Cancer Res. 2005;65:2234–2242.
57. Fishman DA, Liu, Y, Ellerbroek SM, et al. Lysophosphatidic acid promotes matrixmetalloproteinase (MMP) activation and MMP-dependent invasion in ovarian cancercells. Cancer Res. 2001;61:3194–3199.
58. Graves LE, Ariztia EV, Navari JR, et al. Proinvasive properties of ovarian cancer ascites-derived membrane vesicles. Cancer Res. 2004;64:7045–7049.
59. Lee Z, Swaby RF, Liang Y, et al. Lysophosphatidic acid is a major regulator of growth-regulated oncogene alpha in ovarian cancer. Cancer Res. 2006;66:2740–2748.
60. Said NA, Najwer I, Socha MJ, et al. SPARC inhibits LPA-mediated mesothelial-ovariancancer cell crosstalk. Neoplasia. 2007;9:23–35.
61. Robinson-Smith TM, Isaacsohn I, Mercer CA, et al. Macrophages mediateinflammation-enhanced metastasis of ovarian tumors in mice. Cancer Res. 2007;67:5708–5716.
62. Hageman T, Robinson SC, Thompson RG, et al. Ovarian cancer cell-derived migrationinhibitory factor enhances tumor growth, progression, and angiogenesis. Mol CancerTher. 2007;6:1993–2002.
63. Kassis J, Lkominek J, Kohn EC. Tumor microenvironment: what can effusions teach us?Diagn Cytopathol. 2004;33:316–319.
64. Greenaway J, Moorehead R, Shaw P, et al. Epithelial–stromal interaction increases cellproliferation, survival and tumorigenicity in a mouse model of human epithelial ovariancancer. Gynecol Oncol. 2008;108:385–394.
65. Zietarska M, Maugard CM, Filali-Mouhim A, et al. Molecular description of a 3D invitro model for the study of epithelial ovarian cancer (EOC). Mol Carcinog.2007;46:872–885.
66. Zhang S, Balch C, ChanMW, et al. Identification and characterization of ovarian cancerinitiating cells from primary human tumors. Cancer Res. 2008;68:4311–4320.
67. Hegedus B, Marga F, Jakab K, et al. The interplay of cell–cell and cell–matrix interac-tions in the invasive properties of brain tumors. Biophys J. 2006;91:2708–2720.
68. Battifora H, McCaughey WTE. Tumors of the serosal membranes. In: Atlas of TumorPathology, Third Series, Fascicle 15. Washington, DC: Armed Forces Institute of Patho-logy; 1994:1–117.
SPB-104209 15 March 30, 2009 Time: 10:17 Proof 1
15 Microenvironmental Regulation of Ovarian Cancer Metastasis
630
631
632
633
634
635
636
637
638
639
640
641
642
643
644
645
646
647
648
649
650
651
652
653
654
655
656
657
658
659
660
661
662
663
664
665
666
667
668
669
670
671
672
673
674

UNCORRECTEDPROOF
69. Zhang XY, Pettengell R, Nasiri N, et al. Characteristics and growth patterns of humanperitoneal mesothelial cells: comparison between advanced epithelial ovarian cancer andnon-ovarian cancer sources. J Soc Gynecol Investig. 1999;6:333–340.
70. Wang E, Ngalame Y, Panelli MC, et al. Peritoneal and subperitoneal stroma mayfacilitate regional spread of ovarian cancer. Clin Cancer Res. 11:113–122.
71. Cannistra SA, Kansas GS, Niloff J. et al. Binding of ovarian cancer cells to peritonealmesothelium in vitro is partly mediated by CD44H. Cancer Res. 1993;53:3830–3838.
72. Ahmed N, Riley C, Rice G, et al. Role of integrin receptors for fibronectin, collagen andlaminin in the regulation of ovarian carcinoma functions in response to a matrix micro-environment. Clin Exp Metastasis. 2005;22:391–402.
73. Niedbala MJ, Crickard K, Bernacki RJ. In vitro degradation of extracellular matrix byhuman ovarian carcinoma cells. Clin Exp Metastasis. 1987;5:181–197.
74. Sawada M, Shii J, Akedo H, et al. An experimental model for ovarian tumor invasion ofcultured mesothelial cell monolayer. Lab Invest. 1994;70:333–338.
75. Harvey W, Amlot, P. Colalgen production by human mesothelial cells in vitro. J Pathol.1983;139:337–347.
76. Stylianou E, Jenner LA, DaviesM, et al. Isolation, culture and characterization of humanperitoneal mesothelial cells. Kidney Int. 1990;37:1563–1570.
77. Ellerbroek SM, Wu YI, Overall CM, et al. Functional interplay between type I collagenand cell surface matrix metalloproteinase activity. J Biol Chem. 2001;276:24833–24842.
78. Hotary KB, Allen ED, Brooks PC, et al. Membrane type 1 matrix metalloproteinaseusurps tumor growth control imposed by the three-dimensional extracellular matrix.Cell.2003;114:33–45.
79. Symowicz J, Adley BP, Gleason KJ, et al. Engagement of collagen-binding integrinspromotes matrix metalloproteinase-9-dependent E-cadherin ectodomain shedding inovarian carcinoma cells. Cancer Res. 2007;67:2030–2039.
80. Barbolina M, Adley, BP, Kelly, et al. Motility-related actinin alpha-4 is associated withadvanced and metastatic ovarian carcinoma. Lab Invest. 2008;88:602–614.
81. Barbolina MV, Adley BP, Shea LD, et al. Wilms tumor gene protein 1 is associated withovarian cancer metastasis and modulates cell invasion. Cancer. 2008;112:1632–1641.
82. BarbolinaAQ7 MV, Adley BP, Kelly DL, et al. Downregulation of connective tissue growthfactor by three-dimensional matrix enhances ovarian carcinoma cell invasion. IntJ Cancer. 2008 (in press).
83. Langer S, Singer CF, Hudelist G, et al. Jun and Fos family protein expression in humanbreast cancer: correlation of protein expression and clinicopathological parameters. EurJ Gynaecol Oncol. 2006;27:345–352.
84. Zhai Y, Hotary KB, Nan B, et al. Expression of membrane type 1 matrix metalloprotei-nase is associated with cervical carcinoma progression and invasion. Cancer Res.2005;65:6543–6550.
85. Schaller G, Fuchs I, Ebert A, et al. The clinical importance of keratin 18 in breast cancer.Zentralbl Gynakol. 1999;121:126–130.
SPB-104209 15 March 30, 2009 Time: 10:17 Proof 1
M.V. Barbolina et al.
675
676
677
678
679
680
681
682
683
684
685
686
687
688
689
690
691
692
693
694
695
696
697
698
699
700
701
702
703
704
705
706
707
708
709
710
711
712
713
714
715
716
717
718
719

UNCORRECTEDPROOF
Chapter 15
Query No. Line No. Query
AQ1 192 Define acronyms EGF and HB-EGF.AQ2 242 Define acronym CSF-1.AQ3 280 Define acronyms Akt amd PI3.AQ4 337 Define acronym DAPI.AQ5 428 Check edited phase ‘‘composed of interstitial type I
collagen’’.AQ6 630 In Ref. 48, provide volume number and page numbers for
the article.AQ7 700 Please update Ref. 82.
SPB-104209 15 March 30, 2009 Time: 10:17 Proof 1
720
721
722
723
724
725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
763
764
Related Documents











