The FASEB Journal • Research Communication Microenvironment drives the endothelial or neural fate of differentiating embryonic stem cells coexpressing neuropilin-1 and Flk-1 Anna Gualandris,* ,‡,1,2 Alessio Noghero,* ,‡,1 Massimo Geuna, †,‡ Marco Arese,* ,‡ Donatella Valdembri,* ,‡ Guido Serini,* ,‡ and Federico Bussolino* ,‡ *Division of Molecular Angiogenesis, † Division of Clinical and Experimental Cytometry, and ‡ Department of Oncological Sciences, Institute for Cancer Research and Treatment (IRCC), University of Turin School of Medicine, Candiolo, Torino, Italy ABSTRACT The observation that the architecture of the cardiovascular and nervous systems is drawn by common guidance cues and the closeness between neural progenitors and endothelial cells in the vascular niche strongly suggests the existence of links between endothelial and neural cell fates. We identified an embryonic stem cell-derived discrete, nonclonal cell population expressing the two vascular endothelial growth factor receptors neuropilin-1 (Nrp1) and Flk1 that differentiates in vitro toward endothelial or neural phenotypes depending on microenvironmental cues. When microinjected in the chick embryo, Nrp1 cells integrate within the host, developing vessels and brain, and acquire endothelial and neural markers, respec- tively. These results show that precursors of endothelial cells and precursors of neural cells arise from the same pool of differentiating embryonic stem cells and share the expression of Nrp1 and Flk1. These data reinforce the parallelism between vascular and nervous system at the level of cell fate and commitment and open new perspective in regenerative medicine of neurovascular diseases.—Gualandris, A., Noghero, A., Geuna, M., Arese, M., Valdembri, D., Serini, G., Bussolino, F. Microenvironment drives the endothelial or neural fate of differentiating embryonic stem cells coexpressing neuropilin-1 and Flk-1. FASEB J. 23, 68 –78 (2009) Key Words: cell commitment endothelial differentiation em- bryoid bodies During the past two decades, the parallelism be- tween the vascular and nervous system has become increasingly evident. By following similar pathways of migration in the developing body, vessels and nerves establish interdependent reciprocal relations (1). Among the attractive/repulsive molecules that are shared by nerves and vessels there are the semaphorins with their cognate receptors neuropilins (Nrp) and plexins (2). Besides being localized on axons and growth cones, where it modulates nerves trajectory during embryonic development (3), the glycoprotein Nrp1 is also expressed by human endothelial cells, where it mediates the activity of VEGF-A 165 and VEGF- A 121 (4, 5). Accordingly, knocking out and overexpress- ing Nrp1 in engineered mice both lead to lethality because of massive vascular defects (6, 7), whereas the specific loss of Nrp1 in endothelial cells decreases arterial marker expression in small arteries (8). The parallelism between vascular and nervous system is also mirrored at the level of cell commitment and differen- tiation. Even though the origins from different germ layers make the endothelial and neural lineages re- stricted to specific progenitors, some data show a plastic correlation between the endothelial and neural com- mitments. In vitro, in appropriate culture conditions, bone marrow and cord blood cells acquire a neural phenotype (9, 10), while neural stem cells (NSCs) acquire the endothelial phenotype (11). In vivo, bone marrow-derived cells enter the brain and differentiate into neural cells (12, 13), whereas murine NSCs can engraft into the hematopoietic system of irradiated hosts to produce blood cells (14). Furthermore, in the quail-chick chimera model, the avian cranial neuroec- toderm originates smooth muscle cells (15). Consider- ing that Nrp1 and VEGFR-2/Flk-1 are shared by mature neurons and endothelial cells, we wondered whether Nrp1 could be shared also by the respective precursors of both cell types, ultimately marking points of contacts between the two commitments. To achieve our goal, we used murine embryonic stem cells (ES cells) as a source of precursors of both neural and endothelial cells. ES cells differentiate efficiently in vitro to originate the embryoid bodies (EBs), three- dimensional differentiated cell masses that provide a large number of cells representing an early or primitive stage of development, which is otherwise difficult to 1 These authors contributed equally to this work. 2 Correspondence: Department of Oncological Sciences, Institute for Cancer Research and Treatment (IRCC), Strada Provinciale 142, km 3,95, 10060, Candiolo (TO), Italy. E-mail: [email protected] doi: 10.1096/fj.08-112847 68 0892-6638/09/0023-0068 © FASEB

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

The FASEB Journal • Research Communication
Microenvironment drives the endothelial or neural fateof differentiating embryonic stem cells coexpressingneuropilin-1 and Flk-1
Anna Gualandris,*,‡,1,2 Alessio Noghero,*,‡,1 Massimo Geuna,†,‡ Marco Arese,*,‡
Donatella Valdembri,*,‡ Guido Serini,*,‡ and Federico Bussolino*,‡
*Division of Molecular Angiogenesis, †Division of Clinical and Experimental Cytometry, and‡Department of Oncological Sciences, Institute for Cancer Research and Treatment (IRCC),University of Turin School of Medicine, Candiolo, Torino, Italy
ABSTRACT The observation that the architecture ofthe cardiovascular and nervous systems is drawn bycommon guidance cues and the closeness betweenneural progenitors and endothelial cells in the vascularniche strongly suggests the existence of links betweenendothelial and neural cell fates. We identified anembryonic stem cell-derived discrete, nonclonal cellpopulation expressing the two vascular endothelialgrowth factor receptors neuropilin-1 (Nrp1) and Flk1that differentiates in vitro toward endothelial or neuralphenotypes depending on microenvironmental cues.When microinjected in the chick embryo, Nrp1� cellsintegrate within the host, developing vessels and brain,and acquire endothelial and neural markers, respec-tively. These results show that precursors of endothelialcells and precursors of neural cells arise from the samepool of differentiating embryonic stem cells and sharethe expression of Nrp1 and Flk1. These data reinforcethe parallelism between vascular and nervous systemat the level of cell fate and commitment and open newperspective in regenerative medicine of neurovasculardiseases.—Gualandris, A., Noghero, A., Geuna, M.,Arese, M., Valdembri, D., Serini, G., Bussolino, F.Microenvironment drives the endothelial or neural fateof differentiating embryonic stem cells coexpressingneuropilin-1 and Flk-1. FASEB J. 23, 68–78 (2009)
Key Words: cell commitment � endothelial differentiation � em-bryoid bodies
During the past two decades, the parallelism be-tween the vascular and nervous system has becomeincreasingly evident. By following similar pathways ofmigration in the developing body, vessels and nervesestablish interdependent reciprocal relations (1).Among the attractive/repulsive molecules that areshared by nerves and vessels there are the semaphorinswith their cognate receptors neuropilins (Nrp) andplexins (2). Besides being localized on axons andgrowth cones, where it modulates nerves trajectoryduring embryonic development (3), the glycoproteinNrp1 is also expressed by human endothelial cells,
where it mediates the activity of VEGF-A165 and VEGF-A121 (4, 5). Accordingly, knocking out and overexpress-ing Nrp1 in engineered mice both lead to lethalitybecause of massive vascular defects (6, 7), whereas thespecific loss of Nrp1 in endothelial cells decreasesarterial marker expression in small arteries (8). Theparallelism between vascular and nervous system is alsomirrored at the level of cell commitment and differen-tiation. Even though the origins from different germlayers make the endothelial and neural lineages re-stricted to specific progenitors, some data show a plasticcorrelation between the endothelial and neural com-mitments. In vitro, in appropriate culture conditions,bone marrow and cord blood cells acquire a neuralphenotype (9, 10), while neural stem cells (NSCs)acquire the endothelial phenotype (11). In vivo, bonemarrow-derived cells enter the brain and differentiateinto neural cells (12, 13), whereas murine NSCs canengraft into the hematopoietic system of irradiatedhosts to produce blood cells (14). Furthermore, in thequail-chick chimera model, the avian cranial neuroec-toderm originates smooth muscle cells (15). Consider-ing that Nrp1 and VEGFR-2/Flk-1 are shared by matureneurons and endothelial cells, we wondered whetherNrp1 could be shared also by the respective precursorsof both cell types, ultimately marking points of contactsbetween the two commitments.
To achieve our goal, we used murine embryonic stemcells (ES cells) as a source of precursors of both neuraland endothelial cells. ES cells differentiate efficientlyin vitro to originate the embryoid bodies (EBs), three-dimensional differentiated cell masses that provide alarge number of cells representing an early or primitivestage of development, which is otherwise difficult to
1 These authors contributed equally to this work.2 Correspondence: Department of Oncological Sciences,
Institute for Cancer Research and Treatment (IRCC), StradaProvinciale 142, km 3,95, 10060, Candiolo (TO), Italy. E-mail:[email protected]
doi: 10.1096/fj.08-112847
68 0892-6638/09/0023-0068 © FASEB

access in an embryo (16). This model has been used inthe past to identify the hemangioblasts, the commonprecursors of both hematopoietic and endothelial cells(17), and an Flk1� cell population that could differen-tiate not only into hematopoietic and endothelial cells,but also into smooth muscle cells, skeletal muscle cells,and cardiomyocytes (18–20). Despite that no proofshave been reported of the existence of such progeni-tors in vivo and recent studies performed with mouseembryo and tetrachimeric blastocysts argued against it(21, 22), the Flk1� cells isolated from EBs still repre-sent an interesting tool to easily address the topic of cellcommitment and differentiation.
On the basis of these observations, we identified anES-derived population of nondefinitively committedNrp1� cells that retains the capacity to embrace theendothelial or the neural commitments. Such potenti-ality was proved in vitro by culturing the two purifiedNrp1�/Flk1� and Nrp1�/Flk1� populations in cell-type restrictive conditions, and, in vivo, by injecting thecells into different areas of the developing chickenembryos.
MATERIALS AND METHODS
Cell lines
W4 mouse ES cells were kindly provided by Dr. Alexandra L.Joyner (Developmental Biology Program, Memorial Sloan-Kettering Cancer Center, New York, NY, USA) (23). R1 EScells expressing the enhanced yellow fluorescent protein(EYFP) were obtained from Dr. Andras Nagy (Samuel Lunen-feld Research Institute, Mt. Sinai Hospital, Toronto, ON,Canada) (24). Both ES cell lines were cultured as describedpreviously (25). Murine embryonic fibroblasts (MEFs) werepurchased from American Type Culture Collection (LGCStandards, Milan, Italy). OP9 and PA6 stromal cell lines,kindly provided by Dr. Timm Schroeder (Institute of CellResearch, Helmholtz Zentrum Munchen–German ResearchCenter for Environmental Health, Neuherberg, Germany),were cultured as described (26).
In vitro differentiation of ES cells
EBs were obtained by the “hanging drop” procedure, asdescribed previously (25). The age of the EBs was indicatedwith progressing numbers starting from the first day ofculture in the absence of leukemia inhibitory factor (LIF).For fluorescence immunostaining, the EBs were plated ontoLab-Tek II CC2 chamber slides (Nunc, Naperville, IL, USA).
Cell staining for FACS analysis and for cell sorting
EBs were dissociated into cell suspension by trypsin treatment(0.05%) for 1 min and by a mechanical dissociation througha 25-gauge needle. Cells were stained with 1 �g/106 cells ofgoat anti-Nrp1 antibody alone or in association with one ortwo of the following antibodies: anti-Flk1, CD31, ICAM2,CD133, PSA-NCAM, RC2, APC-anti-PDGFR�, APC-anti-CXCR4, APC-anti-Flk1, and PE-anti-Flk1. Antibody brandsand dilutions are given in Supplemental Table 1. The second-ary antibodies used were fluoresecin isothiocyanate (FITC) orAPC-conjugated-anti-goat to detect Nrp1, PE-conjugated-anti-
rat to detect Flk1 and CD133, and FITC-conjugated anti-mouse immunoglobulin G (IgG) or IgM to detect nestin,PSA-NCAM, and RC2 (Southern Biotech., Birmingham, AL,USA). To detect cytoplasmic antigens, the IntraPrep Perme-abilization Reagent was used (Beckman Coulter, Marseille,France) by following the manufacturer’s instructions. As acontrol of intracellular labeling, isotype IgGs were used (datanot shown).
To prepare cells for cell sorting, the secondary antibodiesused were PE-conjugated anti-rat and biotin-conjugated anti-goat followed by an AlexaFluor 647-PE-conjugated streptavi-din (Invitrogen, Carlsbad, CA, USA). Cell sorting was per-formed with Beckman Coulter Epics Altra. The accuracy ofthe sorting procedure was confirmed by analyzing represen-tative samples of the two sorted populations through a BectonDickinson FACS Calibur (Becton Dickinson, Franklin Lakes,NJ, USA) purposely reserved for routine analysis only. Suchanalysis revealed that cell populations were sorted at 95–97%of purity.
Culturing sorted cells
Once sorted, cells were seeded onto OP9 monolayers andcultured in EGM-2 medium composed by endothelial cellbasal medium-2 and the EGM-2 bullet kit (Bio Whittaker,Walkersville, MD, USA).
For the neural differentiation, sorted cells were seededonto laminin-1-coated coverslips and grown for the first 2 daysin a serum-free Dulbecco modified Eagle medium (DMEM)/F12 medium supplemented with N2 supplement (Invitro-gen), 1 �g/ml laminin-1 (Sigma, St. Louis, MO, USA), and 10ng/ml fibroblast growth factor (FGF-2) (R&D Systems, Min-neapolis, MN, USA). FGF-2 was withdrawn from the mediumfor the following 48 h after that medium was replaced withNeurobasal medium plus B27 supplement, 2% horse serum,50 ng/ml nerve growth factor (Roche, Mannheim, Germany),and 10 ng/ml brain-derived neurotrophic factor (BDNF;R&D Systems).
Single-cell deposition assay
YFP� ES cells sorted for Nrp1 and Flk1 expression wereseeded as a single cell per well into optical-bottom 96-wellplates (Nunc, Rochester, NY, USA) containing confluentmonolayers of PA6 feeders and differentiating W4 ES cells.YFP� clones were derived in N2/B27 medium (27) supple-mented with FGF2 20 ng/ml for the first 24 h. Medium waschanged every 2 days; after 8 days of coculture, clones werefixed and processed for immunostaining.
RNA extraction and reverse transcriptase-polymerase chainreaction (RT-PCR)
Total RNAs from the cocultures of YFP sorted cells and OP9cells or from OP9 cells alone were purified at the timesindicated using TriZOL reagent (Invitrogen) and RNase-freeDNase I (DNA-free; Ambion, Austin, TX, USA). RNAs werereverse transcribed using SuperscrptII cDNA synthesis kit(Invitrogen). Control reactions without reverse transcriptasewere performed for each RNA sample. PCR reactions werecarried out with 100 ng of first-strand cDNA using PlatinumTaq polymerase (Invitrogen) and optimized to allow semi-quantitative comparisons within the log phase of amplifica-tion. Comprehensive list of the primer sequences is given inSupplemental Table 2. Images of ethidium bromide-stainedgels were acquired with a Molecular Imager Chemidoc XRS(Bio-Rad, Hercules, CA, USA), and densitometric analysis wasperformed with Quantity One software (Bio-Rad); results
69ENDOTHELIAL AND NEURAL FATE IN ES-DERIVED LINEAGES
were normalized over the expression of the HPRT house-keeping gene and expressed as relative units.
Indirect immunofluorescence
The list of the primary and secondary antibodies used in theseexperiments is given in Supplemental Table 1. All the imageswere captured by using a Leica TCS SP2 AOBS confocalmicroscope and analyzed with Leica Confocal Software (LCS;Leica Microsystems, Wetzlar, Germany).
EBs and cultured sorted cells were fixed with PBS and 4%paraformaldehyde. Incubation with the primary antibodieswas carried out for 1 h at 37°C in a moist chamber. Frozenchicken sections were incubated in a moist chamber over-night at 4°C with primary antibodies; only when required,they were incubated with biotinylated sambucus nigra agglu-tinin (Vector Laboratories, Burlingame, CA, USA) and Alexa-Fluor-555-streptavidin (Invitrogen).
The immunofluorescence staining of the 96-well platescontaining the single-cell deposition assay was performedwith a mixture of the following primary antibodies: anti-greenfluorescent protein (GFP) with anti-RC2 and with anti-VE-cadherin. Alternatively, anti-O4, anti-� smooth muscle actin(�SMA), anti-E-cadherin, or anti-cytokeratin antibody wasused.
Immunoprecipitation and Western blot analysis
Cell lysates of W4 undifferentiated ES cells and of 7-day-oldEBs were obtained under cold conditions with 10 mM Tris/HCL pH 7.5, 150 mM NaCl, 5 mM EDTA, 1% Triton-X-100,0.1 mM ZnCl2 supplemented with protease inhibitors cocktail(Sigma). Protein content was measured by BCA assay (Pierce,Rockford, IL, USA). Nrp1 protein was immunoprecipitatedwith 1 �g of rabbit polyclonal anti-Nrp1 antibody (Santa Cruz
Biotechnology, Santa Cruz, CA, USA), and immunocom-plexes were loaded onto 7.5% SDS-PAGE and transferred toPVDF membrane (Millipore, Bedford, MA, USA). The filterwas decorated with the goat polyclonal anti-Nrp1 antibodyC19 (Santa Cruz). Immunocomplexes were then visualized byenhanced chemiluminescence (ECL) system (Pharmacia-Amersham Biotech., Little Chalfont, Buckinghamshire, UK).
Microinjection experiments on chick embryos
Purified Nrp1�/Flk1� or Nrp1�/Flk1� cells were microin-jected into the beating heart or into the mesencephalic cavityof HH19 chicken embryos by using a manual microinjector(CellTram Oil; Eppendorf, Hamburg, Germany). At differentdays after injection, the embryo vasculature was labeled withAlexaFluor 405-conjugated wheat germ agglutinin that wasinjected into an artery of the vitelline sac. Immediatelyafterward, embryos were sacrificed, fixed with zinc salineformalin (Bio-Optica, Milan, Italy) for 48 h at 4°C or with PBS4% paraformaldehyde for 24 h at 4°C and cryoprotected in15% sucrose for 8 h and then in 30% sucrose overnight at4°C. After embedding and freezing them in Killik compound(Bio-Optica), 12- to 20-�m sections were cut using a LeicaCM3050 S cryostat.
RESULTS
Analysis of Nrp1 expression in differentiatingES cells
ES cells were chosen as an in vitro model to analyze therole played by Nrp1 during cell differentiation. After 5
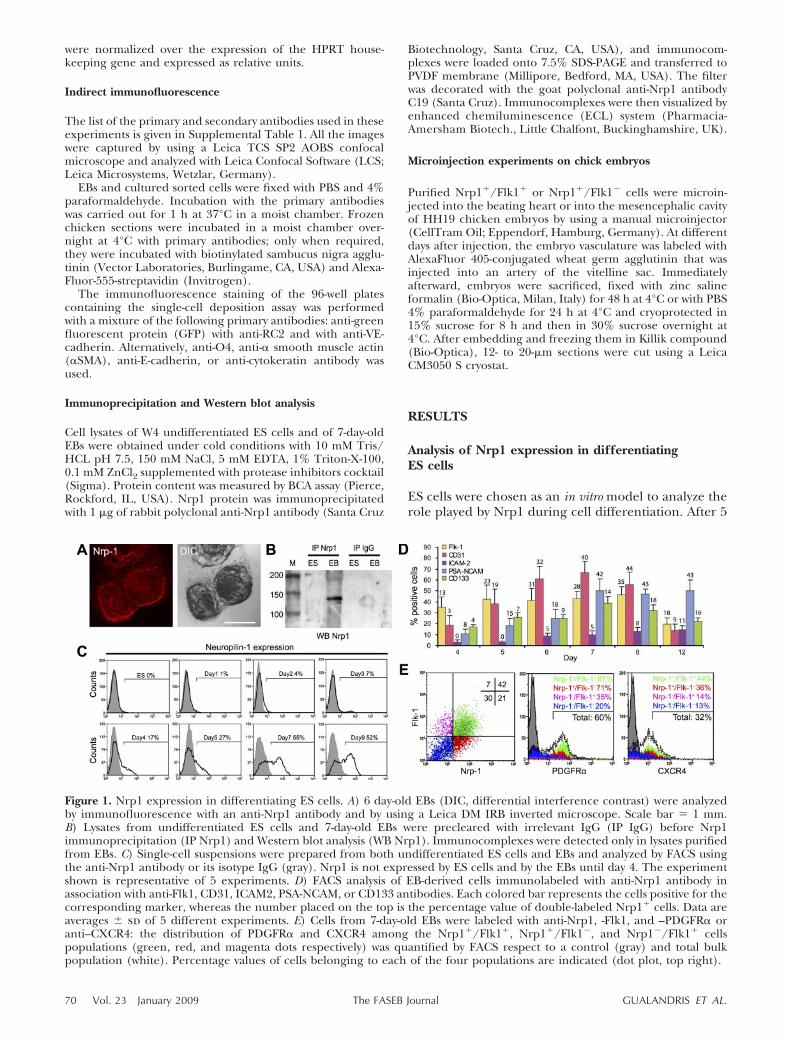
Figure 1. Nrp1 expression in differentiating ES cells. A) 6 day-old EBs (DIC, differential interference contrast) were analyzedby immunofluorescence with an anti-Nrp1 antibody and by using a Leica DM IRB inverted microscope. Scale bar � 1 mm.B) Lysates from undifferentiated ES cells and 7-day-old EBs were precleared with irrelevant IgG (IP IgG) before Nrp1immunoprecipitation (IP Nrp1) and Western blot analysis (WB Nrp1). Immunocomplexes were detected only in lysates purifiedfrom EBs. C) Single-cell suspensions were prepared from both undifferentiated ES cells and EBs and analyzed by FACS usingthe anti-Nrp1 antibody or its isotype IgG (gray). Nrp1 is not expressed by ES cells and by the EBs until day 4. The experimentshown is representative of 5 experiments. D) FACS analysis of EB-derived cells immunolabeled with anti-Nrp1 antibody inassociation with anti-Flk1, CD31, ICAM2, PSA-NCAM, or CD133 antibodies. Each colored bar represents the cells positive for thecorresponding marker, whereas the number placed on the top is the percentage value of double-labeled Nrp1� cells. Data areaverages � sd of 5 different experiments. E) Cells from 7-day-old EBs were labeled with anti-Nrp1, -Flk1, and –PDGFR� oranti–CXCR4: the distribution of PDGFR� and CXCR4 among the Nrp1�/Flk1�, Nrp1�/Flk1�, and Nrp1�/Flk1� cellspopulations (green, red, and magenta dots respectively) was quantified by FACS respect to a control (gray) and total bulkpopulation (white). Percentage values of cells belonging to each of the four populations are indicated (dot plot, top right).
70 Vol. 23 January 2009 GUALANDRIS ET AL.The FASEB Journal
days of growing ES cells as suspended aggregates inLIF-deprived medium, ES cells were allowed to attachto a substrate (day 5), where they formed EBs (Fig. 1A).To analyze Nrp1 expression in differentiating ES cells,6-day-old EBs were fixed and immunostained withanti-Nrp1 antibody. As shown in Fig. 1A, Nrp1 waswidely expressed in scattered areas throughout thegrowing EBs. Furthermore, Nrp1 protein was immuno-precipitated from lysates purified from 7-day-old EBsbut not from undifferentiated W4-ES cells (Fig. 1B).
To quantify Nrp1 expression in the EBs, differentiat-ing ES cells were analyzed by FACS. As shown in Fig. 1C,Nrp1 expression was absent in undifferentiated ES cellsand during the first 3 days of differentiation, it becamesignificant only at day 4 to finally stabilize at 45–50% ofthe total EB cells population in the next days.
Characterization of EB-derived Nrp1� cells
To characterize the phenotype of Nrp1� cells in theEBs, we performed a time-course FACS analysis of EBcells double-labeled for Nrp1 and for markers of eitherendothelial or neural cell precursors (Fig. 1D). Beingthe canonical marker of hemangioblasts, Flk1 is ex-pressed from day 4, which is the very first day ofappearance of Nrp1 expression. The amount of Nrp1�
cells within the Flk1 component progressively increasedover time to almost cover the totality of the Flk1expressing cells at day 12, time in which the Flk1�
endothelial cells are organized into mature vessels (Fig.1D, numbers above bars) (25). At these late time points,ICAM-2 expression and its Nrp1� fraction becamesignificant, whereas CD133 expression, which identifiesboth endothelial (28) and neural precursors (29),maintained a significant percentage of Nrp1-expressingcells throughout the entire EB growth. The polysialy-lated embryonic form of the neural molecule NCAM(PSA-NCAM) is characteristic of neural progenitors in
the forebrain (30), and its expression was shared almostentirely by Nrp1� cells at all the time points examined.CD31, which is expressed by undifferentiated ES cells(31), included a significant component of Nrp1� cellsat its peak of expression (day 7) to later stabilize at alower level at day 12.
Characterization of EB-derived Nrp1�/Flk1�
and Nrp1�/Flk1� cell populations
Because endothelial cells and neurons share the ex-pression of Nrp1 and VEGFR2/Flk1 (3, 4), we focusedon the cell populations defined by these two markers,the Nrp1�/Flk1�, Nrp1�/Flk1�, and Nrp1�/Flk1�
cell types (Fig. 1E, green, red, and magenta, respec-tively; and Supplemental Fig. 1). To better characterizethe potential of differentiation of these populations,the expression of the different germ layer markers wasevaluated by cytofluorimetry and immunofluorescence.As shown by Fig. 1E, in both Nrp1�/Flk1� and Nrp1�/Flk1� cell types, more than 70% of the cells express theparaxial mesoderm marker PDGFR� (32), whereas onlythe 44% of the Nrp1�/Flk1� cells and the 36% of theFlk1� counterpart express the definitive endodermmarker CXCR4 (33). Similarly, mesoderm orientationprevails on endoderm commitment also in those Flk1�
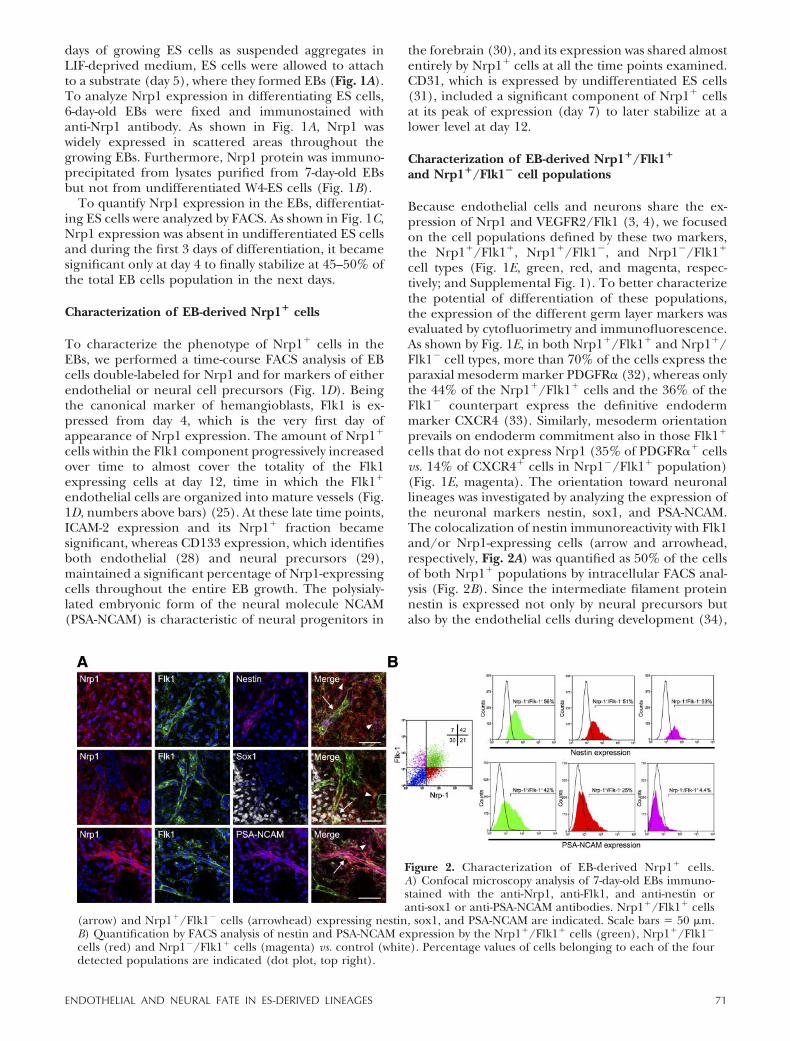
cells that do not express Nrp1 (35% of PDGFR�� cellsvs. 14% of CXCR4� cells in Nrp1�/Flk1� population)(Fig. 1E, magenta). The orientation toward neuronallineages was investigated by analyzing the expression ofthe neuronal markers nestin, sox1, and PSA-NCAM.The colocalization of nestin immunoreactivity with Flk1and/or Nrp1-expressing cells (arrow and arrowhead,respectively, Fig. 2A) was quantified as 50% of the cellsof both Nrp1� populations by intracellular FACS anal-ysis (Fig. 2B). Since the intermediate filament proteinnestin is expressed not only by neural precursors butalso by the endothelial cells during development (34),
Figure 2. Characterization of EB-derived Nrp1� cells.A) Confocal microscopy analysis of 7-day-old EBs immuno-stained with the anti-Nrp1, anti-Flk1, and anti-nestin oranti-sox1 or anti-PSA-NCAM antibodies. Nrp1�/Flk1� cells
(arrow) and Nrp1�/Flk1� cells (arrowhead) expressing nestin, sox1, and PSA-NCAM are indicated. Scale bars � 50 �m.B) Quantification by FACS analysis of nestin and PSA-NCAM expression by the Nrp1�/Flk1� cells (green), Nrp1�/Flk1�
cells (red) and Nrp1�/Flk1� cells (magenta) vs. control (white). Percentage values of cells belonging to each of the fourdetected populations are indicated (dot plot, top right).
71ENDOTHELIAL AND NEURAL FATE IN ES-DERIVED LINEAGES

the more specific markers sox1 and PSA-NCAM werenext investigated. Sox1 is a SRY-related transcriptionfactor whose expression is characteristic of proliferat-ing neural precursors (35, 36). Nrp1�/Flk1� andNrp1�/Flk1� cells that expressed sox1 or PSA-NCAMwere detected by immunofluorescence in sparse areasof 7-day-old EBs (Fig. 2A, arrow and arrowhead). Theexpression of PSA-NCAM was quantified at 42% ofNrp1�/Flk1� cells and at 25% of Nrp1�/Flk1� cells byFACS analysis (Fig. 2B, green and red). Contrary towhat was observed with the PDGFR� molecule (Fig.1E), the lack of PSA-NCAM expression in the Nrp1�/Flk1� population suggested that neural orientation wasnot contemplated by the differentiation potential ofthose cells that did not carry Nrp1 (Fig. 2B, magenta).Similar results were obtained by analyzing the expres-sion of RC2, an antigen that identifies neural precur-sors at the stage of radial glia (37). The two Nrp1� celltypes were enriched in RC2, whereas the Nrp1� coun-terpart was not (Supplemental Fig. 2). To summarize,the molecular characterization of the populations de-fined by Nrp1 and Flk1 revealed an intriguing coexist-ence of markers of neural precursors with markers ofmesoderm/endothelial precursors, which make thefate of Nrp1� cells worth further investigation.
In vitro analysis of the potential of differentiationof Nrp1�/Flk1� and Nrp1�/Flk1� cells: endothelialand neural commitments
To better study the differentiation potential of Nrp1�
cells outside the EB environment, Nrp1�/Flk1� andNrp1�/Flk1� cells were isolated at 95% of purity from
7-day-old EBs by flow cytometry sorting and then cul-tured under different conditions. To support the en-dothelial differentiation in vitro, Nrp1�/Flk1� andNrp1�/Flk1� cells were seeded onto confluent mono-layers of OP9 stromal cells, described to be optimalfeeders for endothelial differentiation (38), and grownin the presence of EGM2 medium; the maturation intoendothelium was evaluated by analyzing the expressionof different endothelial markers by RT-PCR at differentdays (Fig. 3A; see also Supplemental Fig. 3A). Immedi-ately after sorting (day 0), the Nrp1�/Flk1� cellsexpressed all the endothelial genes tested at levels from2 to 8 times higher than those of the Nrp1�/Flk1�
cells, as shown by the quantification data plotted in Fig.3A. High expression of CD31 messenger at day 0 inboth cell types could reflect an inheritance from undif-ferentiated ES cells (31). During the next few days, thelevels of expression of the majority of the endothelialmarkers in the two cell populations became compara-ble. These last observations indicate that the coculturewith OP9 feeders could fill in the initial impairment ofthe Nrp1� cells sorted as Flk1�. Nevertheless, suchrescue was not complete or fast enough to allow theFlk1� cells to organize tubes on Matrigel, a basementmembrane matrix commonly used to evaluate theendothelial phenotype of cell cultures in vitro (Supple-mental Fig. 3B). Despite cell sorting procedures guar-anteeing 95% purity of the sorted cell populations, theFlk1 mRNA was amplified even in the Nrp1�/Flk1� cellsample at day 0. The possibility that the Nrp1�/Flk1�
cells expressed an intracellular inactive Flk1 proteinwas investigated by performing a FACS analysis on EBsat the same differentiation time of cell sorting (day 7),
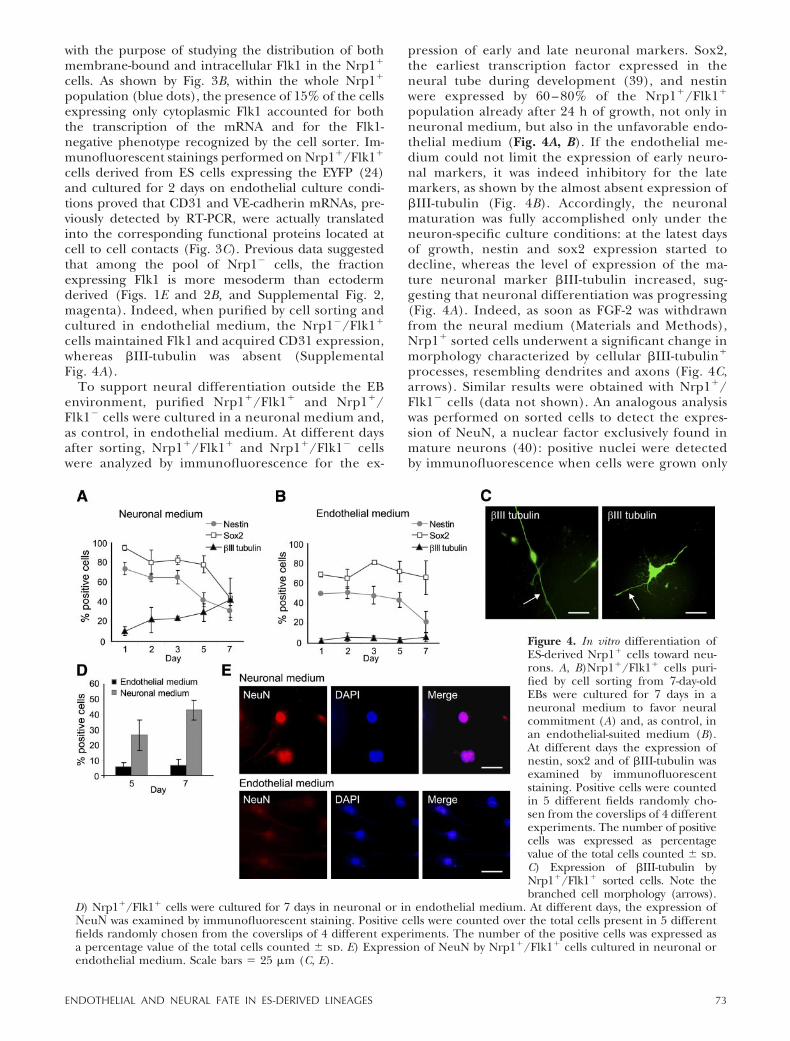
Figure 3. In vitro differentiation of ES-derivedNrp1� cells toward endothelium. A) Nrp1�/Flk1�
and Nrp1�/Flk1� cells purified by cell sorting from7-day-old EBs were cultured onto OP9 feeders for 4days. At different time points, the expression ofFlk1, CD31, Flt1, Endoglin, Tie1, Tie2, and VE-cadherin was evaluated by semiquantitative RT-PCRand normalized to the signal given by the house-keeping gene HPRT. After an initial impairment ofthe Nrp1�/Flk1� cells vs. Nrp1�/Flk1�, the levelsof expression of all the endothelial markers, withthe exception of Tie1, equalized over time. B) After
labeling of cells purified from 7-day-old EBs for the external Nrp1 (FITC) and the external Flk1 (PE), cells were fixed,permeabilized, and incubated with the antibody against cytoplasmic Flk1 (APC). The distribution of both externalmembrane-bound and intracellular cytoplasmic Flk1 was then analyzed by FACS specifically on the Nrp1� cells (blue dots).As indicated (second dot plot, top right) 15% of the Nrp1� cells expressed only cytoplasmic Flk1. C) Immunofluorescentstaining of Nrp1�/Flk1� cells derived from YFP�-ES cells revealed the presence of CD31 and VE-cadherin proteins in themembranes at cell-to-cell contacts. Scale bar � 30 �m.
72 Vol. 23 January 2009 GUALANDRIS ET AL.The FASEB Journal
with the purpose of studying the distribution of bothmembrane-bound and intracellular Flk1 in the Nrp1�
cells. As shown by Fig. 3B, within the whole Nrp1�
population (blue dots), the presence of 15% of the cellsexpressing only cytoplasmic Flk1 accounted for boththe transcription of the mRNA and for the Flk1-negative phenotype recognized by the cell sorter. Im-munofluorescent stainings performed on Nrp1�/Flk1�
cells derived from ES cells expressing the EYFP (24)and cultured for 2 days on endothelial culture condi-tions proved that CD31 and VE-cadherin mRNAs, pre-viously detected by RT-PCR, were actually translatedinto the corresponding functional proteins located atcell to cell contacts (Fig. 3C). Previous data suggestedthat among the pool of Nrp1� cells, the fractionexpressing Flk1 is more mesoderm than ectodermderived (Figs. 1E and 2B, and Supplemental Fig. 2,magenta). Indeed, when purified by cell sorting andcultured in endothelial medium, the Nrp1�/Flk1�
cells maintained Flk1 and acquired CD31 expression,whereas �III-tubulin was absent (SupplementalFig. 4A).
To support neural differentiation outside the EBenvironment, purified Nrp1�/Flk1� and Nrp1�/Flk1� cells were cultured in a neuronal medium and,as control, in endothelial medium. At different daysafter sorting, Nrp1�/Flk1� and Nrp1�/Flk1� cellswere analyzed by immunofluorescence for the ex-
pression of early and late neuronal markers. Sox2,the earliest transcription factor expressed in theneural tube during development (39), and nestinwere expressed by 60 – 80% of the Nrp1�/Flk1�
population already after 24 h of growth, not only inneuronal medium, but also in the unfavorable endo-thelial medium (Fig. 4A, B). If the endothelial me-dium could not limit the expression of early neuro-nal markers, it was indeed inhibitory for the latemarkers, as shown by the almost absent expression of�III-tubulin (Fig. 4B). Accordingly, the neuronalmaturation was fully accomplished only under theneuron-specific culture conditions: at the latest daysof growth, nestin and sox2 expression started todecline, whereas the level of expression of the ma-ture neuronal marker �III-tubulin increased, sug-gesting that neuronal differentiation was progressing(Fig. 4A). Indeed, as soon as FGF-2 was withdrawnfrom the neural medium (Materials and Methods),Nrp1� sorted cells underwent a significant change inmorphology characterized by cellular �III-tubulin�
processes, resembling dendrites and axons (Fig. 4C,arrows). Similar results were obtained with Nrp1�/Flk1� cells (data not shown). An analogous analysiswas performed on sorted cells to detect the expres-sion of NeuN, a nuclear factor exclusively found inmature neurons (40): positive nuclei were detectedby immunofluorescence when cells were grown only
Figure 4. In vitro differentiation ofES-derived Nrp1� cells toward neu-rons. A, B)Nrp1�/Flk1� cells puri-fied by cell sorting from 7-day-oldEBs were cultured for 7 days in aneuronal medium to favor neuralcommitment (A) and, as control, inan endothelial-suited medium (B).At different days the expression ofnestin, sox2 and of �III-tubulin wasexamined by immunofluorescentstaining. Positive cells were countedin 5 different fields randomly cho-sen from the coverslips of 4 differentexperiments. The number of positivecells was expressed as percentagevalue of the total cells counted � sd.C) Expression of �III-tubulin byNrp1�/Flk1� sorted cells. Note thebranched cell morphology (arrows).
D) Nrp1�/Flk1� cells were cultured for 7 days in neuronal or in endothelial medium. At different days, the expression of�eu� was examined by immunofluorescent staining. Positive cells were counted over the total cells present in 5 differentfields randomly chosen from the coverslips of 4 different experiments. The number of the positive cells was expressed asa percentage value of the total cells counted � sd. E) Expression of NeuN by Nrp1�/Flk1� cells cultured in neuronal orendothelial medium. Scale bars � 25 �m (C, E).
73ENDOTHELIAL AND NEURAL FATE IN ES-DERIVED LINEAGES
in neuronal medium (Fig. 4E). The results of thequantitative analysis are shown in Fig. 4D. Similarresults were obtained with Nrp1�/Flk1� cells (datanot shown). Contrary to the Nrp1� cell types, theNrp1�/Flk1� cells could not survive the restrictionsof the neural medium, thus confirming that thepresence of Flk1 in the absence of Nrp1 gave thesecells a mesoderm-restricted fate. All together, thesedata demonstrate that in response to the appropriateexogenous microenvironment, the bulk of ES-de-rived Nrp1� cell population gives rise to endothelialor neural cells.
Neuropilin-1 and Flk1 identify a discrete,nonclonal population with neurovasculardifferentiative potential
To analyze the capacity of single YFP� Nrp1�/Flk1�
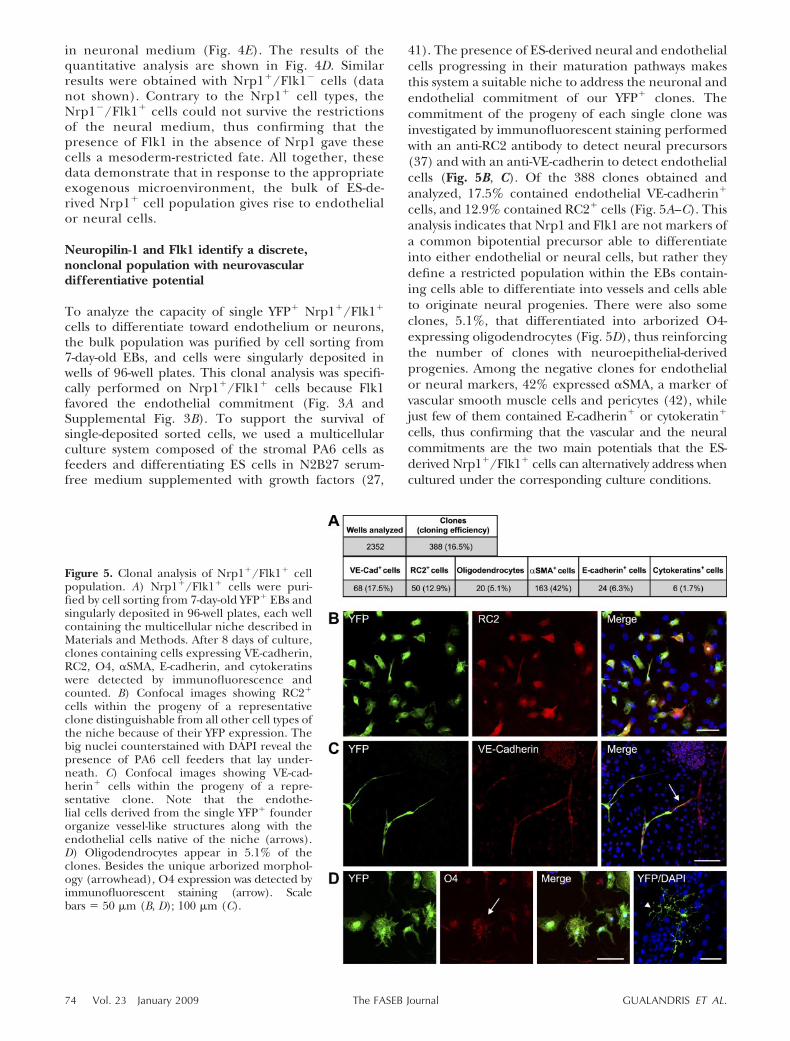
cells to differentiate toward endothelium or neurons,the bulk population was purified by cell sorting from7-day-old EBs, and cells were singularly deposited inwells of 96-well plates. This clonal analysis was specifi-cally performed on Nrp1�/Flk1� cells because Flk1favored the endothelial commitment (Fig. 3A andSupplemental Fig. 3B). To support the survival ofsingle-deposited sorted cells, we used a multicellularculture system composed of the stromal PA6 cells asfeeders and differentiating ES cells in N2B27 serum-free medium supplemented with growth factors (27,
41). The presence of ES-derived neural and endothelialcells progressing in their maturation pathways makesthis system a suitable niche to address the neuronal andendothelial commitment of our YFP� clones. Thecommitment of the progeny of each single clone wasinvestigated by immunofluorescent staining performedwith an anti-RC2 antibody to detect neural precursors(37) and with an anti-VE-cadherin to detect endothelialcells (Fig. 5B, C). Of the 388 clones obtained andanalyzed, 17.5% contained endothelial VE-cadherin�
cells, and 12.9% contained RC2� cells (Fig. 5A–C). Thisanalysis indicates that Nrp1 and Flk1 are not markers ofa common bipotential precursor able to differentiateinto either endothelial or neural cells, but rather theydefine a restricted population within the EBs contain-ing cells able to differentiate into vessels and cells ableto originate neural progenies. There were also someclones, 5.1%, that differentiated into arborized O4-expressing oligodendrocytes (Fig. 5D), thus reinforcingthe number of clones with neuroepithelial-derivedprogenies. Among the negative clones for endothelialor neural markers, 42% expressed �SMA, a marker ofvascular smooth muscle cells and pericytes (42), whilejust few of them contained E-cadherin� or cytokeratin�
cells, thus confirming that the vascular and the neuralcommitments are the two main potentials that the ES-derived Nrp1�/Flk1� cells can alternatively address whencultured under the corresponding culture conditions.
Figure 5. Clonal analysis of Nrp1�/Flk1� cellpopulation. A) Nrp1�/Flk1� cells were puri-fied by cell sorting from 7-day-old YFP� EBs andsingularly deposited in 96-well plates, each wellcontaining the multicellular niche described inMaterials and Methods. After 8 days of culture,clones containing cells expressing VE-cadherin,RC2, O4, �SMA, E-cadherin, and cytokeratinswere detected by immunofluorescence andcounted. B) Confocal images showing RC2�
cells within the progeny of a representativeclone distinguishable from all other cell types ofthe niche because of their YFP expression. Thebig nuclei counterstained with DAPI reveal thepresence of PA6 cell feeders that lay under-neath. C) Confocal images showing VE-cad-herin� cells within the progeny of a repre-sentative clone. Note that the endothe-lial cells derived from the single YFP� founderorganize vessel-like structures along with theendothelial cells native of the niche (arrows).D) Oligodendrocytes appear in 5.1% of theclones. Besides the unique arborized morphol-ogy (arrowhead), O4 expression was detected byimmunofluorescent staining (arrow). Scalebars � 50 �m (B, D); 100 �m (C).
74 Vol. 23 January 2009 GUALANDRIS ET AL.The FASEB Journal
In vivo analysis of the potential of differentiationof Nrp1�/Flk1� and Nrp1�/Flk1� cells: endothelialand neural commitments
Even with the limits of an in vitro culture system, thedata shown so far represented a supporting indica-tion of the capacity of the ES-derived Nrp1� cells todifferentiate into endothelium or neurons depend-ing on the appropriate stimuli. We next wonderedwhether a more physiological environment, such as aliving embryo, could allow the Nrp1� cells to betteraccomplish their fate. For that reason, Nrp1�/Flk1�
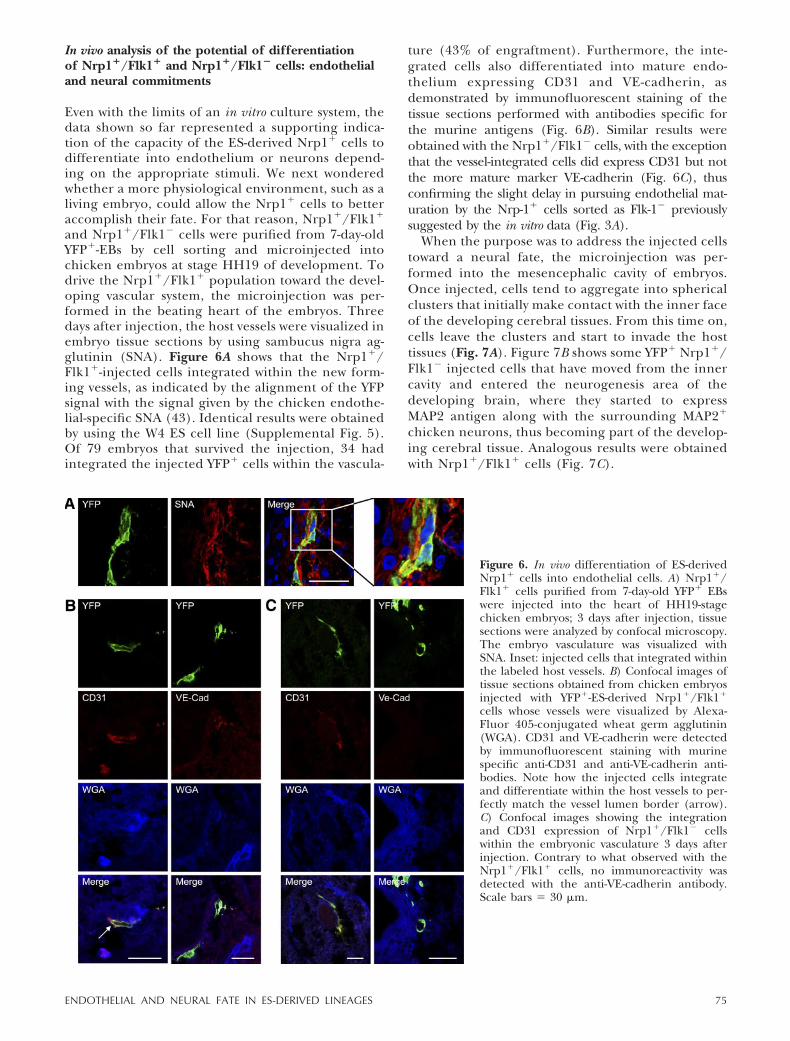
and Nrp1�/Flk1� cells were purified from 7-day-oldYFP�-EBs by cell sorting and microinjected intochicken embryos at stage HH19 of development. Todrive the Nrp1�/Flk1� population toward the devel-oping vascular system, the microinjection was per-formed in the beating heart of the embryos. Threedays after injection, the host vessels were visualized inembryo tissue sections by using sambucus nigra ag-glutinin (SNA). Figure 6A shows that the Nrp1�/Flk1�-injected cells integrated within the new form-ing vessels, as indicated by the alignment of the YFPsignal with the signal given by the chicken endothe-lial-specific SNA (43). Identical results were obtainedby using the W4 ES cell line (Supplemental Fig. 5).Of 79 embryos that survived the injection, 34 hadintegrated the injected YFP� cells within the vascula-
ture (43% of engraftment). Furthermore, the inte-grated cells also differentiated into mature endo-thelium expressing CD31 and VE-cadherin, asdemonstrated by immunofluorescent staining of thetissue sections performed with antibodies specific forthe murine antigens (Fig. 6B). Similar results wereobtained with the Nrp1�/Flk1� cells, with the exceptionthat the vessel-integrated cells did express CD31 but notthe more mature marker VE-cadherin (Fig. 6C), thusconfirming the slight delay in pursuing endothelial mat-uration by the Nrp-1� cells sorted as Flk-1� previouslysuggested by the in vitro data (Fig. 3A).
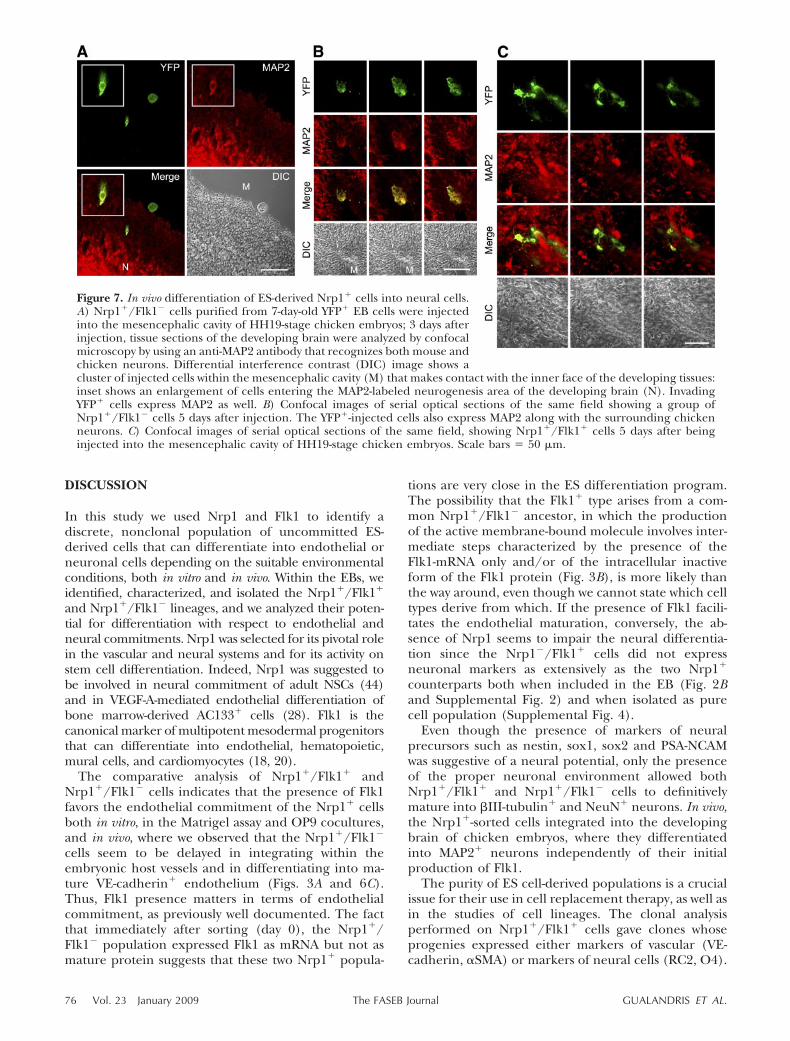
When the purpose was to address the injected cellstoward a neural fate, the microinjection was per-formed into the mesencephalic cavity of embryos.Once injected, cells tend to aggregate into sphericalclusters that initially make contact with the inner faceof the developing cerebral tissues. From this time on,cells leave the clusters and start to invade the hosttissues (Fig. 7A). Figure 7B shows some YFP� Nrp1�/Flk1� injected cells that have moved from the innercavity and entered the neurogenesis area of thedeveloping brain, where they started to expressMAP2 antigen along with the surrounding MAP2�
chicken neurons, thus becoming part of the develop-ing cerebral tissue. Analogous results were obtainedwith Nrp1�/Flk1� cells (Fig. 7C).
Figure 6. In vivo differentiation of ES-derivedNrp1� cells into endothelial cells. A) Nrp1�/Flk1� cells purified from 7-day-old YFP� EBswere injected into the heart of HH19-stagechicken embryos; 3 days after injection, tissuesections were analyzed by confocal microscopy.The embryo vasculature was visualized withSNA. Inset: injected cells that integrated withinthe labeled host vessels. B) Confocal images oftissue sections obtained from chicken embryosinjected with YFP�-ES-derived Nrp1�/Flk1�
cells whose vessels were visualized by Alexa-Fluor 405-conjugated wheat germ agglutinin(WGA). CD31 and VE-cadherin were detectedby immunofluorescent staining with murinespecific anti-CD31 and anti-VE-cadherin anti-bodies. Note how the injected cells integrateand differentiate within the host vessels to per-fectly match the vessel lumen border (arrow).C) Confocal images showing the integrationand CD31 expression of Nrp1�/Flk1� cellswithin the embryonic vasculature 3 days afterinjection. Contrary to what observed with theNrp1�/Flk1� cells, no immunoreactivity wasdetected with the anti-VE-cadherin antibody.Scale bars � 30 �m.
75ENDOTHELIAL AND NEURAL FATE IN ES-DERIVED LINEAGES
DISCUSSION
In this study we used Nrp1 and Flk1 to identify adiscrete, nonclonal population of uncommitted ES-derived cells that can differentiate into endothelial orneuronal cells depending on the suitable environmentalconditions, both in vitro and in vivo. Within the EBs, weidentified, characterized, and isolated the Nrp1�/Flk1�
and Nrp1�/Flk1� lineages, and we analyzed their poten-tial for differentiation with respect to endothelial andneural commitments. Nrp1 was selected for its pivotal rolein the vascular and neural systems and for its activity onstem cell differentiation. Indeed, Nrp1 was suggested tobe involved in neural commitment of adult NSCs (44)and in VEGF-A-mediated endothelial differentiation ofbone marrow-derived AC133� cells (28). Flk1 is thecanonical marker of multipotent mesodermal progenitorsthat can differentiate into endothelial, hematopoietic,mural cells, and cardiomyocytes (18, 20).
The comparative analysis of Nrp1�/Flk1� andNrp1�/Flk1� cells indicates that the presence of Flk1favors the endothelial commitment of the Nrp1� cellsboth in vitro, in the Matrigel assay and OP9 cocultures,and in vivo, where we observed that the Nrp1�/Flk1�
cells seem to be delayed in integrating within theembryonic host vessels and in differentiating into ma-ture VE-cadherin� endothelium (Figs. 3A and 6C).Thus, Flk1 presence matters in terms of endothelialcommitment, as previously well documented. The factthat immediately after sorting (day 0), the Nrp1�/Flk1� population expressed Flk1 as mRNA but not asmature protein suggests that these two Nrp1� popula-
tions are very close in the ES differentiation program.The possibility that the Flk1� type arises from a com-mon Nrp1�/Flk1� ancestor, in which the productionof the active membrane-bound molecule involves inter-mediate steps characterized by the presence of theFlk1-mRNA only and/or of the intracellular inactiveform of the Flk1 protein (Fig. 3B), is more likely thanthe way around, even though we cannot state which celltypes derive from which. If the presence of Flk1 facili-tates the endothelial maturation, conversely, the ab-sence of Nrp1 seems to impair the neural differentia-tion since the Nrp1�/Flk1� cells did not expressneuronal markers as extensively as the two Nrp1�
counterparts both when included in the EB (Fig. 2Band Supplemental Fig. 2) and when isolated as purecell population (Supplemental Fig. 4).
Even though the presence of markers of neuralprecursors such as nestin, sox1, sox2 and PSA-NCAMwas suggestive of a neural potential, only the presenceof the proper neuronal environment allowed bothNrp1�/Flk1� and Nrp1�/Flk1� cells to definitivelymature into �III-tubulin� and NeuN� neurons. In vivo,the Nrp1�-sorted cells integrated into the developingbrain of chicken embryos, where they differentiatedinto MAP2� neurons independently of their initialproduction of Flk1.
The purity of ES cell-derived populations is a crucialissue for their use in cell replacement therapy, as well asin the studies of cell lineages. The clonal analysisperformed on Nrp1�/Flk1� cells gave clones whoseprogenies expressed either markers of vascular (VE-cadherin, �SMA) or markers of neural cells (RC2, O4).
Figure 7. In vivo differentiation of ES-derived Nrp1� cells into neural cells.A) Nrp1�/Flk1� cells purified from 7-day-old YFP� EB cells were injectedinto the mesencephalic cavity of HH19-stage chicken embryos; 3 days afterinjection, tissue sections of the developing brain were analyzed by confocalmicroscopy by using an anti-MAP2 antibody that recognizes both mouse andchicken neurons. Differential interference contrast (DIC) image shows acluster of injected cells within the mesencephalic cavity (M) that makes contact with the inner face of the developing tissues:inset shows an enlargement of cells entering the MAP2-labeled neurogenesis area of the developing brain (N). InvadingYFP� cells express MAP2 as well. B) Confocal images of serial optical sections of the same field showing a group ofNrp1�/Flk1� cells 5 days after injection. The YFP�-injected cells also express MAP2 along with the surrounding chickenneurons. C) Confocal images of serial optical sections of the same field, showing Nrp1�/Flk1� cells 5 days after beinginjected into the mesencephalic cavity of HH19-stage chicken embryos. Scale bars � 50 �m.
76 Vol. 23 January 2009 GUALANDRIS ET AL.The FASEB Journal

The fact that endothelial and neural cells were neverlocated together within the progeny of the same clonedemonstrates that Nrp1 and Flk1 define a populationof cells containing precursors restricted to the vascularfate and precursors restricted to the neural fate, thusexcluding the existence of a bipotential common pre-cursor. The topic of the bipotentiality or multipoten-tiality of progenitor cells has recently become con-troversial. Despite the numerous indirect evidencesupporting the existence of the hemangioblast (17, 20,45), recent in vivo studies argue against its bipotential-ity. Cell-lineage tracing of cells from the primitivestreak to the yolk sac failed to reveal a commonhematopoietic and endothelial progenitor (21). More-over, the analysis of tetrachimeras derived from differ-ent ES cell lines stably expressing separate fluorescenttracers showed that each blood island has contributionsfrom multiple clonal progenitors (22). Our results aremore in line with these studies, especially consideringthat a putative neuron-endothelial precursor, by tres-passing the ectoderm-mesoderm boundaries, wouldrepresent a concept of bipotent progenitor more chal-lenging than the hemangioblast. In this respect, ourdata are clear: the progeny of a single Nrp1�/Flk1� cellis only endothelial or neural. The idea of a bipotentprogenitor within our Nrp1�/Flk1� cells was justifiedby the model used, the ES cells, and by the presence ofFlk1. By using the same model with alternative proto-cols, different groups showed that ES-derived Flk1�
cells could differentiate into multiple mesoderm-de-rived lineages (18, 20, 46). Regarding these data, theunique presence of Nrp1 along with other neuralmarkers gives more challenges to our model. It wasproposed that Flk1 marks a common mesodermalprecursor that segregates to successive subsets of Flk1-expressing or nonexpressing cells whose fate is thendetermined by coexpression of lineage-specific tran-scription factors (47). Hence, our Nrp1�/Flk1� popu-lation could represent a transient intermediate pool ofcells, in which the committed progenitors arise, thanksto the loss or acquisition of specific molecules. Thepossibility to purify this pool of cells by using Nrp1 andFlk1 along with the encouraging in vivo transplantationdata opens new perspectives for the treatment of disor-ders characterized by defects of both the neural andvascular systems, including stroke, diabetic neuropathy,and Alzheimer’s disease (48).
This work was supported by Istituto Superiore di Sanita’(Programma Nazionale sulle Cellule Staminali, VI Pro-gramma nazionale AIDS), Associazione Italiana per la Ricercasul Cancro, Regione Piemonte (Progetto di Ricerca sanitariaFinalizzata 2006, Ricerca Scientifica 2004, grants A150 andD10), 6FP European Community (LSHM-CT-2003–503254),Fondazione Cassa di Risparmio di Torino and Ministero dellaSalute (Programma di Ricerca Finalizzata 2006 and Pro-gramma Straordinario di Ricerca Oncologica 2006), Minis-tero dell’Universita’ (PRIN2007), and Telethon, ItalyGGP04127. We thank Dr. Alexandra L. Joyner (Developmen-tal Biology Program, Memorial Sloan-Kettering Cancer Cen-ter, New York, NY, USA) for the gift of W4 129SvEv ES cells,Dr. Andras Nagy (Samuel Lunenfeld Research Institute, Mt.
Sinai Hospital, Toronto, ON, Canada) and Anna-KaterinaHadjantonakis (Developmental Biology Program, MemorialSloan-Kettering Cancer Center, New York, NY, USA) for R1EYFP-expressing ES cells, Dr. Timm Schroeder (Institute ofCell Research, Helmholtz Zentrum Munchen–German Re-search Center for Environmental Health, Neuherberg, Ger-many) for OP9 and PA6 cells and helpful suggestions, L.Primo for suggestions and comments, and Dr. S. Geuna forassistance with confocal microscopy. We thank Famarco andSusatrasporti S.p.A. for white Leghorn chicken eggs, and theDevelopmental Studies Hybridoma Bank of the University ofIowa for antibodies.
REFERENCES
1. Mukouyama, Y. S., Shin, D., Britsch, S., Taniguchi, M., andAnderson, D. J. (2002) Sensory nerves determine the pattern ofarterial differentiation and blood vessel branching in the skin.Cell 109, 693–705
2. Bussolino, F., Valdembri, D., Caccavari, F., and Serini, G. (2006)Semaphoring vascular morphogenesis. Endothelium 13, 81–91
3. Fujisawa, H., Takagi, S., and Hirata, T. (1995) Growth-associatedexpression of a membrane protein, neuropilin, in Xenopus opticnerve fibers. Dev. Neurosci. 17, 343–349
4. Soker, S., Miao, H. Q., Nomi, M., Takashima, S., and Klagsbrun,M. (2002) VEGF165 mediates formation of complexes contain-ing VEGFR-2 and neuropilin-1 that enhance VEGF165-receptorbinding. J. Cell. Biochem. 85, 357–368
5. Shraga-Heled, N., Kessler, O., Prahst, C., Kroll, J., Augustin, H.,and Neufeld, G. (2007) Neuropilin-1 and neuropilin-2 enhanceVEGF121 stimulated signal transduction by the VEGFR-2 recep-tor. FASEB J. 21, 915–926
6. Kawasaki, T., Kitsukawa, T., Bekku, Y., Matsuda, Y., Sanbo, M.,Yagi, T., and Fujisawa, H. (1999) A requirement for neuropilin-1in embryonic vessel formation. Development 126, 4895–4902
7. Kitsukawa, T., Shimono, A., Kawakami, A., Kondoh, H., andFujisawa, H. (1995) Overexpression of a membrane protein,neuropilin, in chimeric mice causes anomalies in the cardiovas-cular system, nervous system and limbs. Development 121, 4309–4318
8. Mukouyama, Y. S., Gerber, H. P., Ferrara, N., Gu, C., andAnderson, D. J. (2005) Peripheral nerve-derived VEGF pro-motes arterial differentiation via neuropilin 1-mediated positivefeedback. Development 132, 941–952
9. Goolsby, J., Marty, M. C., Heletz, D., Chiappelli, J., Tashko, G.,Yarnell, D., Fishman, P. S., Dhib-Jalbut, S., Bever, C. T., Jr.,Pessac, B., and Trisler, D. (2003) Hematopoietic progenitorsexpress neural genes. Proc. Natl. Acad. Sci. U. S. A. 100, 14926–14931
10. Buzanska, L., Machaj, E. K., Zablocka, B., Pojda, Z., andDomanska-Janik, K. (2002) Human cord blood-derived cellsattain neuronal and glial features in vitro. J. Cell Sci. 115,2131–2138
11. Wurmser, A. E., Nakashima, K., Summers, R. G., Toni, N.,D’Amour, K. A., Lie, D. C., and Gage, F. H. (2004) Cellfusion-independent differentiation of neural stem cells to theendothelial lineage. Nature 430, 350–356
12. Mezey, E., Chandross, K. J., Harta, G., Maki, R. A., andMcKercher, S. R. (2000) Turning blood into brain: cells bearingneuronal antigens generated in vivo from bone marrow. Science290, 1779–1782
13. Mezey, E., Key, S., Vogelsang, G., Szalayova, I., Lange, G. D., andCrain, B. (2003) Transplanted bone marrow generates new neu-rons in human brains. Proc. Natl. Acad. Sci. U. S. A. 100, 1364–1369
14. Bjornson, C. R., Rietze, R. L., Reynolds, B. A., Magli, M. C., andVescovi, A. L. (1999) Turning brain into blood: a hematopoieticfate adopted by adult neural stem cells in vivo. Science 283,534–537
15. Korn, J., Christ, B., and Kurz, H. (2002) Neuroectodermalorigin of brain pericytes and vascular smooth muscle cells.J. Comp. Neurol. 442, 78–88
16. Keller, G. (2005) Embryonic stem cell differentiation: emer-gence of a new era in biology and medicine. Genes Dev. 19,1129–1155
77ENDOTHELIAL AND NEURAL FATE IN ES-DERIVED LINEAGES

17. Choi, K., Kennedy, M., Kazarov, A., Papadimitriou, J. C., andKeller, G. (1998) A common precursor for hematopoietic andendothelial cells. Development 125, 725–732
18. Yamashita, J., Itoh, H., Hirashima, M., Ogawa, M., Nishikawa, S.,Yurugi, T., Naito, M., Nakao, K., and Nishikawa, S. (2000)Flk1-positive cells derived from embryonic stem cells serve asvascular progenitors. Nature 408, 92–96
19. Motoike, T., Markham, D. W., Rossant, J., and Sato, T. N. (2003)Evidence for novel fate of Flk1� progenitor: contribution tomuscle lineage. Genesis 35, 153–159
20. Kattman, S. J., Huber, T. L., and Keller, G. M. (2006) Multipo-tent flk-1� cardiovascular progenitor cells give rise to thecardiomyocyte, endothelial, and vascular smooth muscle lin-eages. Dev. Cell 11, 723–732
21. Kinder, S. J., Loebel, D. A., and Tam, P. P. (2001) Allocationand early differentiation of cardiovascular progenitors in themouse embryo. Trends Cardiovasc. Med. 11, 177–184
22. Ueno, H., and Weissman, I. L. (2006) Clonal analysis of mousedevelopment reveals a polyclonal origin for yolk sac bloodislands. Dev. Cell. 11, 519–533
23. Auerbach, W., Dunmore, J. H., Fairchild-Huntress, V., Fang, Q.,Auerbach, A. B., Huszar, D., and Joyner, A. L. (2000) Establish-ment and chimera analysis of 129/SvEv- and C57BL/6-derivedmouse embryonic stem cell lines. BioTechniques 29, 1024–1028,1030, 1032
24. Hadjantonakis, A. K., Macmaster, S., and Nagy, A. (2002)Embryonic stem cells and mice expressing different GFP vari-ants for multiple non-invasive reporter usage within a singleanimal. BMC Biotechnol. 2, 11
25. Gualandris, A., Annes, J. P., Arese, M., Noguera, I., Jurukovski,V., and Rifkin, D. B. (2000) The latent transforming growthfactor-beta-binding protein-1 promotes in vitro differentiationof embryonic stem cells into endothelium. Mol. Biol. Cell 11,4295–4308
26. Schroeder, T., Meier-Stiegen, F., Schwanbeck, R., Eilken, H.,Nishikawa, S., Hasler, R., Schreiber, S., Bornkamm, G. W.,Nishikawa, S., and Just, U. (2006) Activated Notch1 altersdifferentiation of embryonic stem cells into mesodermal celllineages at multiple stages of development. Mech. Dev. 123,570–579
27. Ying, Q. L., and Smith, A. G. (2003) Defined conditions forneural commitment and differentiation. Methods Enzymol. 365,327–341
28. Fons, P., Herault, J. P., Delesque, N., Tuyaret, J., Bono, F., andHerbert, J. M. (2004) VEGF-R2 and neuropilin-1 are involved inVEGF-A-induced differentiation of human bone marrow pro-genitor cells. J. Cell. Physiol. 200, 351–359
29. Lee, A., Kessler, J. D., Read, T. A., Kaiser, C., Corbeil, D.,Huttner, W. B., Johnson, J. E., and Wechsler-Reya, R. J. (2005)Isolation of neural stem cells from the postnatal cerebellum.Nat. Neurosci. 8, 723–729
30. Gago, N., Avellana-Adalid, V., Evercooren, A. B., and Schu-macher, M. (2003) Control of cell survival and proliferation ofpostnatal PSA-NCAM(�) progenitors. Mol. Cell. Neurosci. 22,162–178
31. Li, Z. J., Wang, Z. Z., Zheng, Y. Z., Xu, B., Yang, R. C., Scadden,D. T., and Han, Z. C. (2005) Kinetic expression of plateletendothelial cell adhesion molecule-1 (PECAM-1/CD31) duringembryonic stem cell differentiation. J. Cell Biochem. 95, 559–570
32. Nishikawa, S. I., Nishikawa, S., Hirashima, M., Matsuyoshi, N.,and Kodama, H. (1998) Progressive lineage analysis by cellsorting and culture identifies FLK1�VE-cadherin� cells at a
diverging point of endothelial and hemopoietic lineages. Devel-opment 125, 1747–1757
33. Yasunaga, M., Tada, S., Torikai-Nishikawa, S., Nakano, Y.,Okada, M., Jakt, L. M., Nishikawa, S., Chiba, T., Era, T., andNishikawa, S. (2005) Induction and monitoring of definitiveand visceral endoderm differentiation of mouse ES cells. Nat.Biotechnol. 23, 1542–1550
34. Wiese, C., Rolletschek, A., Kania, G., Blyszczuk, P., Tarasov,K. V., Tarasova, Y., Wersto, R. P., Boheler, K. R., and Wobus,A. M. (2004) Nestin expression–a property of multi-lineageprogenitor cells? Cell. Mol. Life. Sci. 61, 2510–2522
35. Bylund, M., Andersson, E., Novitch, B. G., and Muhr, J. (2003)Vertebrate neurogenesis is counteracted by Sox1–3 activity. Nat.Neurosci. 6, 1162–1168
36. Kiefer, J. C. (2007) Back to basics: Sox genes. Dev. Dyn. 236,2356–2366
37. Anthony, T. E., Klein, C., Fishell, G., and Heintz, N. (2004)Radial glia serve as neuronal progenitors in all regions of thecentral nervous system. Neuron 41, 881–890
38. Hirashima, M., Kataoka, H., Nishikawa, S., Matsuyoshi, N., andNishikawa, S. (1999) Maturation of embryonic stem cells intoendothelial cells in an in vitro model of vasculogenesis. Blood 93,1253–1263
39. Zappone, M. V., Galli, R., Catena, R., Meani, N., De Biasi, S.,Mattei, E., Tiveron, C., Vescovi, A. L., Lovell-Badge, R., Otto-lenghi, S., and Nicolis, S. K. (2000) Sox2 regulatory sequencesdirect expression of a (beta)-geo transgene to telencephalicneural stem cells and precursors of the mouse embryo, revealingregionalization of gene expression in CNS stem cells. Develop-ment 127, 2367–2382
40. Sarnat, H. B., Nochlin, D., and Born, D. E. (1998) Neuronalnuclear antigen (NeuN): a marker of neuronal maturation inearly human fetal nervous system. Brain Dev. 20, 88–94
41. Kawasaki, H., Mizuseki, K., Nishikawa, S., Kaneko, S., Kuwana,Y., Nakanishi, S., Nishikawa, S. I., and Sasai, Y. (2000) Inductionof midbrain dopaminergic neurons from ES cells by stromalcell-derived inducing activity. Neuron 28, 31–40
42. Betsholtz, C., Lindblom, P., and Gerhardt, H. (2005) Role ofpericytes in vascular morphogenesis. EXS 94, 115–125
43. Nanka, O., Peumans, W. J., Van Damme, E. J., Pfuller, U.,Valasek, P., Halata, Z., Schumacher, U., and Grim, M. (2001)Lectin histochemistry of microvascular endothelium in chickand quail musculature. Anat. Embryol. (Berl.) 204, 407–411
44. Maurer, M. H., Tripps, W. K., Feldmann, R. E., Jr., andKuschinsky, W. (2003) Expression of vascular endothelialgrowth factor and its receptors in rat neural stem cells. Neurosci.Lett. 344, 165–168
45. Huber, T. L., Kouskoff, V., Fehling, H. J., Palis, J., and Keller, G.(2004) Haemangioblast commitment is initiated in the primi-tive streak of the mouse embryo. Nature 432, 625–630
46. Wu, S. M., Fujiwara, Y., Cibulsky, S. M., Clapham, D. E., Lien,C. L., Schultheiss, T. M., and Orkin, S. H. (2006) Developmentalorigin of a bipotential myocardial and smooth muscle cellprecursor in the mammalian heart. Cell 127, 1137–1150
47. Ema, M., and Rossant, J. (2003) Cell fate decisions in early bloodvessel formation. Trends Cardiovasc. Med. 13, 254–259
48. Murry, C. E., and Keller, G. (2008) Differentiation of embryonicstem cells to clinically relevant populations: lessons from embry-onic development. Cell 132, 661–680
Received for publication May 13, 2008.Accepted for publication August 14, 2008.
78 Vol. 23 January 2009 GUALANDRIS ET AL.The FASEB Journal
Related Documents











