Full Terms & Conditions of access and use can be found at https://www.tandfonline.com/action/journalInformation?journalCode=lcss20 Communications in Soil Science and Plant Analysis ISSN: (Print) (Online) Journal homepage: https://www.tandfonline.com/loi/lcss20 Microbial Dynamics and Nutrient Mineralization in Soil Amended with Cacao Pod and Water Hyacinth Composts: Implication for Nitrogen Fixed by Soybean Cornelius Talade Atere , Michael Omofowa Osunde & Akin Olayinka To cite this article: Cornelius Talade Atere , Michael Omofowa Osunde & Akin Olayinka (2020) Microbial Dynamics and Nutrient Mineralization in Soil Amended with Cacao Pod and Water Hyacinth Composts: Implication for Nitrogen Fixed by Soybean, Communications in Soil Science and Plant Analysis, 51:19, 2466-2478, DOI: 10.1080/00103624.2020.1836202 To link to this article: https://doi.org/10.1080/00103624.2020.1836202 Published online: 03 Nov 2020. Submit your article to this journal Article views: 17 View related articles View Crossmark data

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Full Terms & Conditions of access and use can be found athttps://www.tandfonline.com/action/journalInformation?journalCode=lcss20
Communications in Soil Science and Plant Analysis
ISSN: (Print) (Online) Journal homepage: https://www.tandfonline.com/loi/lcss20
Microbial Dynamics and Nutrient Mineralization inSoil Amended with Cacao Pod and Water HyacinthComposts: Implication for Nitrogen Fixed bySoybean
Cornelius Talade Atere , Michael Omofowa Osunde & Akin Olayinka
To cite this article: Cornelius Talade Atere , Michael Omofowa Osunde & Akin Olayinka (2020)Microbial Dynamics and Nutrient Mineralization in Soil Amended with Cacao Pod and WaterHyacinth Composts: Implication for Nitrogen Fixed by Soybean, Communications in Soil Scienceand Plant Analysis, 51:19, 2466-2478, DOI: 10.1080/00103624.2020.1836202
To link to this article: https://doi.org/10.1080/00103624.2020.1836202
Published online: 03 Nov 2020.
Submit your article to this journal
Article views: 17
View related articles
View Crossmark data

Microbial Dynamics and Nutrient Mineralization in Soil Amended with Cacao Pod and Water Hyacinth Composts: Implication for Nitrogen Fixed by SoybeanCornelius Talade Atere , Michael Omofowa Osunde , and Akin Olayinka
Department of Soil Science and Land Resources Management, Obafemi Awolowo University, (OAU), Ife, Nigeria
ABSTRACTComposting organic wastes can be relatively cheap and environmentally- friendly sources of mineralizable nutrients for legumes. We amended 5 kg topsoil (0–15 cm) samples of an Ultisol (Lixisol) with 0, 2.5 and 5 t ha−1 each of cacao pod (CC) and water hyacinth (WC) composts with and without starter fertilizer (F) (25 kg N ha−1 as urea and 26 kg P ha−1 as single super phos-phate). The amended soil samples were used for 16-week laboratory incuba-tion experiment and two-consecutive 8-week plantings of soybean (Glycine max. L. Merril) in the screenhouse. Measurements were carried out on some indices of microbial activity, namely, microbial respiration, N and P mineralization as well as on nodulation and N2 fixed by soybean. The microbial respiration, and N and P mineralization increased with application rates of the composts, but generally declined with incubation duration. Applications of 5 t ha−1 each of CC and WC with or without F increased (p < .05) the microbial respiration (24–93%), N mineralized (21–35%), and P mineralized (32–180%) during the incubation period. The highest of these increments was, however, obtained with WC. Increases of 1162 and 981% (CC2F and WC2F) and 28% (CC2F) were also recorded in nodulation and the amount of N2-fixed, respectively. It was concluded that 5 t ha−1 water hyacinth or cacao pod composts with starter fertilizer is capable of enhan-cing soybean N2 fixation by serving as mineralizable nutrient sources prior to the onset of nodulation and N2 fixation.
ARTICLE HISTORY Received 2 April 2020 Accepted 15 May 2020
KEYWORDS N2 fixation; cacao pod; water hyacinth; compost; N and P mineralization
Introduction
Soil microorganisms play numerous and important functions in the soil ecosystem functioning. Their roles in soil include, but not limited to formation of organic matter from dead plant and animal residues through the process of decomposition, nutrient (C, N, P, and K) cycling, improvement of soil structure, suppression of invading pathogens and biological nitrogen fixation (BNF) (Aislabie and Deslippe 2013; Jacoby et al. 2017). Symbiotic nitrogen fixation which is mediated by groups of bacteria and actinomycetes (Hillel 2008; Mahmud et al. 2020) is especially advantageous for increased yields and reduction of chemical fertilizer applications in legume’s production.Soybeans is an important legume, with production of approximately 349 million tonnes globally (Food and Agriculture Organization of the United Nations Statistics Division (FAOSTAT) 2018), accounting for 50% of the world’s oil-seed production (OECD-FAO 2019). Soybean is rich in protein, and can serve as a cheaper alternative to cowpea, meat, eggs and pork (Adino, Wondifraw, and Addis 2018; Atere and Olayinka 2013). It generates 200 kg N ha−1 in above ground biomass each growing season with 58–68% of its nitrogen content resulting from symbiotic nitrogen fixation (Brookshire et al. 2019). By fixing its own nitrogen, and sparing some soil nitrate, soybean leaves positive balance of N in the soil,
CONTACT Cornelius Talade Atere [email protected] Department of Soil Science and Land Resources Management, Obafemi Awolowo University, (OAU), Ife, Nigeria
COMMUNICATIONS IN SOIL SCIENCE AND PLANT ANALYSIS 2020, VOL. 51, NO. 19, 2466–2478 https://doi.org/10.1080/00103624.2020.1836202
© 2020 Taylor & Francis Group, LLC

thereby replenishing the soil and increasing the yields of non N-fixers planted next to it (Hauggaard- Nielsen, Ambus, and Jensen 2001). Although the N-demand of soybean can be largely met through N2 fixation, external supply of nutrients through mineral and/or organic fertilizers may be required prior to the onset of N2 fixation, and to supply other nutrients for balanced plant nutrition. Mineral fertilizers are ready sources of plant nutrients for increased crop production (Timsina 2018). However, their applications by peasant farmers have been significantly reduced recently due to scarcity and high costs (Lin et al. 2019). Such adverse side effects on the soil as acidity and Aluminum toxicity have also discouraged the use of mineral fertilizers (Adesanwo et al. 2009). Comparatively, organic fertilizers are relatively cheaper and environmentally friendly, and has gained tremendous attention in recent times despite few limitations such as slow nutrient release and bulkiness. Several agricultural wastes have been converted to organic fertilizers among which are the water hyacinth (Eichhornia crassipes [Mart.] Solms) and cacao pods. Water hyacinth is a free-floating macrophyte belonging to the family Pontedericeae, Liliales that grows in fresh water. Water hyacinth has been recognized as one of the most harmful aquatic weeds in the world due to its negative effects on people’s livelihoods and waterways (McKeown and Bugyi 2016). It is of negative importance to the aquatic ecosystem due to its ability to impede traffic and cause a reduction in the oxygen concentration of the water (McKeown and Bugyi 2016). Numerous physical, chemical, and biological weed control methods adopted for its control have yielded little success (Abdelsabour 2010). However, there has been continued efforts from some researchers that water hyacinth can be converted to positive uses other than being a nuisance to both terrestrial and aquatic ecosystems (Abdelsabour 2010; Anjanabha and Kumar 2010). Thus, the weed is considered a valuable source of macronutrients such as phosphorus, nitrogen and potassium that are essential for plant nutrition (Atere and Olayinka 2013; Kuke and Hunde 2019). The biomass of this weed has been composted and used as organic fertilizer (Matsumura 2002) for crop production, especially grains. Another compostable waste is cacao pod husks. Cacao (Theobroma cacao L., family: Sterculiaceae) is a major cash crop in Nigeria and many tropical countries. Global annual production of dry cocoa beans exceeded 3.98 million tons in 2011–2012 (Fidelis and Rao 2017). Cacao pod husks is known to generate tons of waste, and the global burden of pod husk waste continues to increase well over 40 million tons (Fidelis and Rao 2017; Vriesmann, Teofilo, and Petkowicz 2012). Attempts have been made on the diversification of cacao pod husks to other uses like animal feeds (Alexander et al. 2008), local soaps production (Taiwo and Osinowo 2001), feedstocks like crude gums and pectins for food and pharmaceutical industries (Vriesmann, Teofilo, and Petkowicz 2012) and as organic fertilizer for increased crop production (Fidelis and Rao 2017). Although, these wastes are generated in large quantities in their respective regions of prevalence, their utilization as composts for crop growth remain very low. It is generally believed that small amount of N supply at the early stage of growth can boost the N2 fixing capability of legumes (Vieira et al. 2010) so as not to shift them to dependence on external N supply at the expense of symbiotically fixed nitrogen (Dent and Cocking 2017). Cacao pod and water hyacinth composts are rich in plant nutrients including N and P but, the microbially mediated pattern of nutrient release from these composts such that they are beneficial for improved N2 fixation process in legumes is poorly understood.The objectives of this study were, therefore, to (i) determine the dynamics of microbial respiration, and N and P mineralization in cacao pod- and water hyacinth-amended soils, and (ii) estimate the N2 fixed by soybean in the compost-amended soils.
Materials and methods
Site description
The study locations were the screenhouse and the Department of Soil Science & Land Resources Management’s Soil Microbiology Laboratory, Obafemi Awolowo University (OAU), Ile-Ife, Nigeria.. The area is within the rain forest zone and located between latitudes Latitude 7° 30’–7° 35’ N and Longitude 4° 30’–4° 35’ E. The altitude is about 200 meters above mean sea level with approximately eight months (March–October) of bimodal annual rainfall having its peaks in June and September.
COMMUNICATIONS IN SOIL SCIENCE AND PLANT ANALYSIS 2467
The dry season spans about four months, usually, November–February subject to some irregularity in the rainfall distribution pattern over the years (Ojetade et al. 2016). The mean annual precipitation is about 1400 mm. The soil properties were as described by Atere and Olayinka (2013, 2019) as follows: field moisture capacity (FMC) 22.30%, pH 4.40 (0.01 M CaCl2), 11.00 g organic C kg−1 soil, 5.80 g total N (TN) kg−1, 21.34 g available P kg−1, 0.21 cmol K kg−1, 2.10 cmol Ca kg−1, 3.21 cmol Mg kg−1, 0.28 cmol Na kg−1. The textural classification of the soil was clay, comprising 310 g kg−1 sand, 230 g kg−1
silt, and 460 g kg−1 clay.
Sample preparation
Top soil samples (0 to 15 cm) of Itagunmodi soil series (Ultisol/Lixisol) were obtained from an uncultivated plot close to the OAU’s main gate. The soil samples were bulked, air-dried, and passed through 2 mm sieve to remove coarse particles. The portion that passed through the 2 mm sieve was used for laboratory analyses, laboratory incubation and screenhouse soil culture experiments. To prepare the composts, water hyacinth (Eichhornia crassipes L.) and freshly-discarded cacao pods were obtained, chopped into smaller pieces and composted in jute bags over a period of 3 months with regular turning. Thereafter, the composted materials were compounded with and without N (0 and 25 kg ha−1 as urea) and P (0 and 26 kg ha−1 as SSP), mixed and further composted for 1 month to obtain organo-mineral fertilizers. The chemical properties of the cacao pod and the water hyacinth are shown in Table 1.
Analytical procedures
SoilThe following parameters were determined: soil particle size distribution using the hydrometer method (Bouyoucos 1962), but with 0.2 M sodium hydroxide (NaOH) solution used as the dispersing agent instead of 5% Calgon, water holding capacity using the core sampling method (Blake and Hartge1986), soil pH potentiometrically in a soil-solution ratio of 1:2 in 0.01 M CaCl2 using a glass electrode pH meter (Peech, Olsen, and Bolt 1953), available P using the Bray-1 method (Bray and Kurtz 1945) and read at 660 nm wavelength after the development of the molybdenum blue color, exchangeable cations extracted with 1 N ammonium acetate (NH4OAc) at pH 7 and the sodium (Na), calcium (Ca), and potassium (K) determined using flame photometer while magnesium (Mg) the ethylene-diaminetetraacetic acid (EDTA) titration method (Page, Miller, and Keeney 1982). The total N contents were determined using micro-Kjeldahl digestion and distillation process (Bremner and Mulvaney 1982).
Cacao pod and water hyacinthSome total elemental contents of the fresh and composted cacao pod and water hyacinth were determined by oven drying portions at 65°C, grinding with stainless steel mill and digesting using
Table 1. Chemical properties of cacao pod and water hyacinth.
Cacao Pod Water hyacinth
Fresh Composted Fresh Composted
C (%) 33.60 21.80 37.44 15.31N (%) 0.49 0.88 0.84 1.05P (%) 0.36 0.56 0.92 0.67K (%) 4.06 7.28 2.25 4.81Ca (%) 0.30 0.60 0.28 0.67Mg (%) 0.03 0.02 0.02 0.02Na (%) 0.24 0.31 1.14 0.48C:N 68.57 24.77 44.57 14.58
Atere and Olayinka (2013, 2019))
2468 C. T. ATERE ET AL.

concentrated H2SO4 and 30% H2O2. The total contents of cations were read: K, Na, and Ca with flame photometer and Mg with Atomic Absorption Spectrophotometer. Their organic carbon and total N contents were also determined using chromic acid digestion method of Allison (1965) and micro- Kjeldahl digestion and distillation process (Bremner and Mulvaney 1982), respectively.
Screenhouse soil culture experiment
The soil culture experiment was carried out in the screenhouse at the Obafemi Awolowo University, Ile-Ife, Nigeria. Five kilograms (5 kg) of soil was amended with water hyacinth or cacao pod compost with and without N and P. Thus, there were three levels (0, 2.5, and 5 t ha−1) each of the of the two composts, and two levels (addition or no addition) of inorganic fertilizer. as follows: Control, F, CC1, WC1, CC1F, WC1F, CC2, WC2, CC2F, WC2F. Where, Control = soil only; F = Inorganic N (25 kg ha−1) and P (26 kg ha−1); CC = cacao pod compost; WC = water hyacinth compost; 1 = 2.5 t ha−1; 2 = 5.0 t ha−1.
The two composts were treated individually and not combined as a treatment because emphasis was on their individual effects on the measured soil microbial indices and N2 fixation. Further, the materials were sorted from locations far apart; while cacao pod is abundant in the forestry upland zones, water hyacinth is more abundant on water bodies, especially in the riverine areas. Thus, either of them, depending on accessibility, can be used for compost.
The amended soils were each transferred into plastic pots (19 and 13 cm upper and lower inner diameters, and 21 cm height)-perforated at the bottom for free drainage and air movement), replicated thrice, arranged in a Completely Randomized Design (CRD). The soil moisture content at 70% of the FMC was monitored with moisture meter. Soil moisture content at 60–70% of the FMC is believed to be optimum for most plant and microbial growths (Ilstedt, Nordgren, and Malmer 2000) Three (3) soybean seeds of an early maturing Tropical Glycine max (TGX1448-ZE) were planted per pot and later thinned to 2 at two weeks after planting. The plants were allowed to grow for 8 weeks and terminated at 50% flowering. At harvesting, the shoots were cut at 2 cm above the soil surface and the leaves detached, leaving the petioles with the stems which were used to determine the nitrogen fixed. The roots were also carefully harvested in order to expose the root nodules. The fresh and dry weights, respectively, of the nodules, roots, and stems with petioles were taken. A repeated planting of soybean was done on the same soils, but without fresh addition of amendments. This was to evaluate the residual effects of the amendments.
Laboratory incubation
The laboratory incubation was carried out to determine the effects of the treatments on some soil microbial indices, namely, microbial respiration, and nitrogen (N) and phosphorus (P) mineralization as described below.
Determination of microbial respiration, N and P mineralizationTo determine microbial respiration, one hundred grams (100 g) portions of soil were amended with water hyacinth or cacao pod compost with and without starter N and P at the rates of 0, 2.5, and 5 t ha−1. There was a total of ten (10) treatments replicated three times, just as done for the screenhouse study, and arranged in a completely randomized design (CRD). The soil samples were moistened with distilled water to about 70% of its FMC and put in 500 ml glass jars. With the aid of lengths of thread, vials containing 10 ml of 1 M NaOH solutions were suspended into the jars and capped tightly. The CO2-C evolved was determined fortnightly using the double acid titration method (Anderson 1982). The unreacted NaOH was titrated with 0.5 N H2SO4 first in the presence of phenolphthalein indicator to a colorless end point, followed by the addition of methyl orange for further titration to determine the carbonate formed. The experiment lasted for 16 weeks.
The concentration of C was calculated using the following formula
COMMUNICATIONS IN SOIL SCIENCE AND PLANT ANALYSIS 2469

mgC=100g ¼Titre � N H2SO4 � 30 � 12
60 Where 30 is equivalent of CO3
= in mg; 12/60 is ratio of C to the mass of CO3=
For N and P mineralization, four hundred grams (400 g) portions of soil were weighed and incubated as described above and N and P mineralized were monitored fortnightly for 16 weeks. The amount of N mineralized was determined using the steam distillation technique of Bremner and Keeney (1966). Ten grams (10 g) portion of each sample was extracted with 30 ml of 1 M KCl by shaking the suspension for an hour and then filtering with Whatman no.2 (11 cm) filter paper. Twenty milliliters (20 ml) of the filtrate was steam-distilled into 10 ml of 4% H3BO3 indicator mixture after addition of 0.1 g MgO and Devarda’s alloy. The borate produced was later titrated against 0.05 N H2SO4. Available P on the other hand was determined using the Bray-1 technique (Bray and Kurtz 1945). Briefly, two (2) g of soil sample was weighed into 50 ml conical flask. 20 ml of solution A (0.025 NHCl + 0.03 N NH4F) was added and shaken for 5 minutes in a mechanical shaker. The suspension was then immediately filtered with 9 cm, Whatman No. 2 filter paper into funnel tube. A 3 ml. aliquot of filtrate was transferred into a colorimeter tube. Three (3) ml of solution B (0.87 N HCl, 0.38% ammonium molybdate, 0.5% H3BO3) was then added to the colorimeter tube and mixed. Solution C was prepared by dissolving 8 g of the dry powder (prepared by thoroughly mixing and grinding 2.5 g of 1-amino-2-sulfonic acid, 5.0 g of sodium sulfite (Na2SO3) and 146 g of sodium metabisulphite (Na2S2O5) to a fine powder) in 50 ml of warm distilled water, and leaving to stand overnight if possible. Five (5) drops of the solution C was then added to the solution in each colorimeter tube and mixed immediately. This is the reductant which forms the blue color with the phosphor-molybdate. The final solution is read on the colorimeter (already calibrated with standard P solutions) after 15–45 minutes of adding the P-C solution, at a wavelength of 660 nm. The absorbance of the samples is then plotted against the absorbance of P standards to obtain the concentrations of P in the samples.
Determination of nitrogen fixed
Hot water extraction (HWE) technique of tissueThe nitrogen fixed was determined using Hot Water Extraction (HWE) technique of ureide assay (Herridge, Bergersen, and People 1990; Herridge, O’Connell, and Donnelly 1988). The harvested plant tissues (stem + petiole) were used for this analysis. The fresh samples were weighed and put in labeled paper bags under natural air-dry condition for about 48–72 hours. They were later oven-dried at 65°C in a forced-air oven for 2 days and ground. About 0.5 g portion of the ground tissue were weighed into 100 ml beaker. Twenty-five (25) ml of distilled water was added to the weighed sample using measuring cylinder and placed on a hot plate already set to its boiling level. The content was allowed to boil for 1–2 minutes and filtered while still hot into 250 ml conical flask. The filtrate was poured quantitatively into a 50 ml volumetric flask and made up to volume with distilled water after cooling. The eluant was transferred into plastic vials with fitting caps and kept in a freezer until analysis for N-solutes, viz, ureide-N, and nitrate-N.
N solute analysesUreide N. One (1) mM standard of Ureide (allantoin) was prepared by adding 39.53 mg allantoin to 250 ml water and used in making concentrations of 0.0, 0.1, 0.2, 0.4, and 1.0 mM for a standard curve at 525 nm wavelength. A standard curve of the concentrations of allantoin within the range of 0–0.10 mM against the optical density was constructed. A 0.5 ml of 0.5 N NaOH was added to a test tube containing 2.5 ml of each of the concentrations, respectively, and placed in a boiling water bath for 10 minutes. The test tubes were later removed and placed on a bench and one ml of the mixture of 0.65 N HCl/0.33% phenylhydrazine was added and placed in the boiling water bath for exactly 2 minutes. The rack containing the test tubes was removed from the boiling water bath and
2470 C. T. ATERE ET AL.

immediately plunged into an ice bath (plastic cooler containing ice) for 15 minutes. After the rack of the tubes was removed from the ice bath, 2.5 ml of cold mixture of 10 N HCl/1.67% KFeCn was added and mixed thoroughly for even color development and better accuracy. A uniform red color with high ureide concentrations developed was left for 10 minutes and the optical density was read at 525 nm on a spectrophotometer immediately to avoid color fade after 15 minutes.
Nitrate-N. Nitrate standard (25 mM) was prepared by adding 635 mg KNO3 to 250 ml water and this was used to make concentrations (0.0, 1.25, 2.5, 5.0, 10.0, and 15.0 mM) for a standard curve. A standard curve of concentrations (within the range 0–15.0 mM) against optical density of nitrate was plotted. A small portion (0.05 ml) of each of the six concentrations was transferred into test tubes in duplicates with pipette. A small measure (0.20 ml) of salicylic acid was added and mixed thoroughly with each of the concentrations to obtain a clear solution and thereafter left to stand for 20 minutes. After 20 minutes, 4.75 ml of 2 N NaOH was added and left for another 10 minutes. The optical density was then read at 410 nm on a spectrometer.
The relative abundance of ureide-N (RU) in extracts of whole stem segments was calculated using the equation below:
RU %ð Þ ¼4 � ureide
4 � ureideð Þ þ nitrate½ �� 100
The standard curve relating proportion of N derived from N2 fixation (Ndfa%) to RU for soybean during vegetative and flowering stages was obtained using the equation below:
RU ¼ 10:7þ 0:50P þ 0:0034P2
Where P is the proportion of plant N from N2 fixation (%Ndfa), and RU is the relative abundance of ureide-N in stem extracts.
Statistical analysis
The data collected were subjected to ANOVA and means separated using the Duncan’s New Multiple Range Test (DNMRT) at 5% probability level.
Results
Microbial respiration in soils amended with cacao pod and water hyacinth composts during sixteen (16) weeks of laboratory incubation study
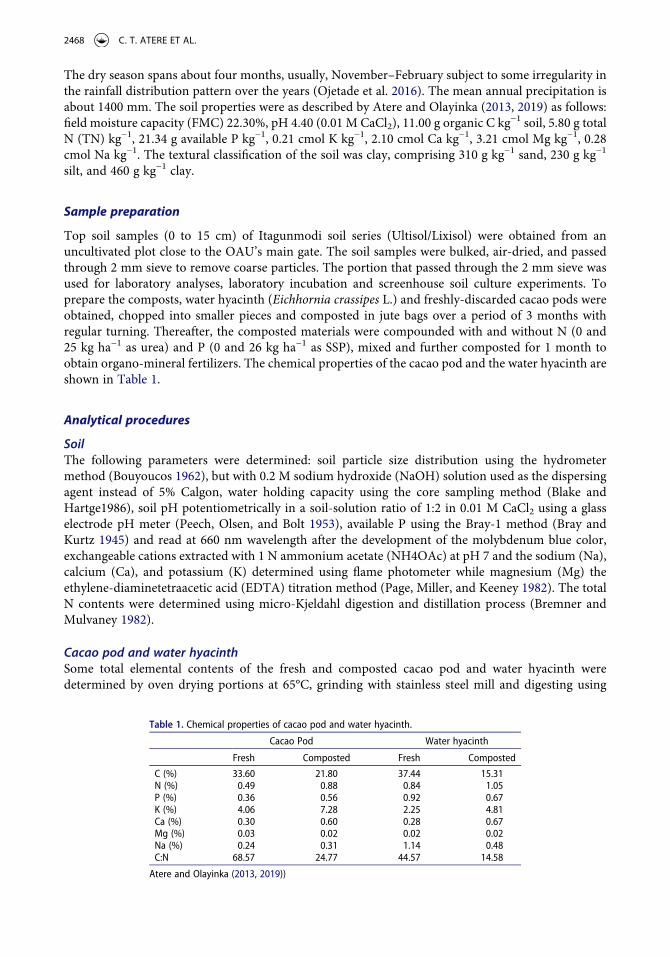
The highest CO2-C evolution from the amended soils during the incubation period was recorded in the 2nd week of incubation, indicating a flush of microbial activity (respiration) at this period. (Table 2). The CO2-C released in this 2nd week was 24–93% (p < .05) higher in CC1, CC2, CC2F, WC2, and WC2F than in the Control, which showed increase in microbial respiration with the increasing application rates of water-hyacinth and cacao-pod composts. . Except in the 10th and 16th weeks, only WC2F consistently resulted in higher C evolution (p < .05) than the Control throughout the incubation period. The CO2-C released from the sole fertilizer treatment was similar to that from the Control during the entire incubation period. It was also observed that with increasing duration of incubation, especially from the 8th week, the CO2-C released became increasingly similar among the treatments. The highest cumulative CO2-C evolved was obtained with WC2F and CC2, and the least with fertilizer and the Control (Figure 1).
COMMUNICATIONS IN SOIL SCIENCE AND PLANT ANALYSIS 2471
Nitrogen (N) and phosphorus (P) mineralization in soil samples amended with cacao pod and water hyacinth composts during sixteen (16) weeks of laboratory incubation study
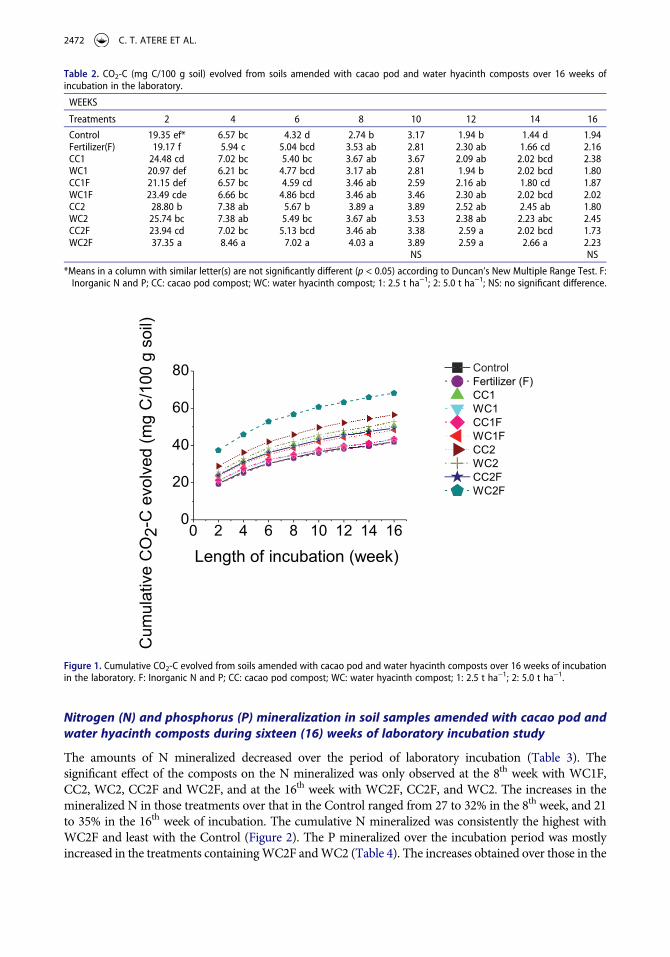
The amounts of N mineralized decreased over the period of laboratory incubation (Table 3). The significant effect of the composts on the N mineralized was only observed at the 8th week with WC1F, CC2, WC2, CC2F and WC2F, and at the 16th week with WC2F, CC2F, and WC2. The increases in the mineralized N in those treatments over that in the Control ranged from 27 to 32% in the 8th week, and 21 to 35% in the 16th week of incubation. The cumulative N mineralized was consistently the highest with WC2F and least with the Control (Figure 2). The P mineralized over the incubation period was mostly increased in the treatments containing WC2F and WC2 (Table 4). The increases obtained over those in the
Table 2. CO2-C (mg C/100 g soil) evolved from soils amended with cacao pod and water hyacinth composts over 16 weeks of incubation in the laboratory.
WEEKS
Treatments 2 4 6 8 10 12 14 16
Control 19.35 ef* 6.57 bc 4.32 d 2.74 b 3.17 1.94 b 1.44 d 1.94Fertilizer(F) 19.17 f 5.94 c 5.04 bcd 3.53 ab 2.81 2.30 ab 1.66 cd 2.16CC1 24.48 cd 7.02 bc 5.40 bc 3.67 ab 3.67 2.09 ab 2.02 bcd 2.38WC1 20.97 def 6.21 bc 4.77 bcd 3.17 ab 2.81 1.94 b 2.02 bcd 1.80CC1F 21.15 def 6.57 bc 4.59 cd 3.46 ab 2.59 2.16 ab 1.80 cd 1.87WC1F 23.49 cde 6.66 bc 4.86 bcd 3.46 ab 3.46 2.30 ab 2.02 bcd 2.02CC2 28.80 b 7.38 ab 5.67 b 3.89 a 3.89 2.52 ab 2.45 ab 1.80WC2 25.74 bc 7.38 ab 5.49 bc 3.67 ab 3.53 2.38 ab 2.23 abc 2.45CC2F 23.94 cd 7.02 bc 5.13 bcd 3.46 ab 3.38 2.59 a 2.02 bcd 1.73WC2F 37.35 a 8.46 a 7.02 a 4.03 a 3.89
NS2.59 a 2.66 a 2.23
NS
*Means in a column with similar letter(s) are not significantly different (p < 0.05) according to Duncan’s New Multiple Range Test. F: Inorganic N and P; CC: cacao pod compost; WC: water hyacinth compost; 1: 2.5 t ha−1; 2: 5.0 t ha−1; NS: no significant difference.
0 2 4 6 8 10 12 14 160
20
40
60
80 ControlFertilizer (F)CC1WC1CC1FWC1FCC2WC2CC2FWC2F
evitalumu
CC
O2
)liosg
001/C
gm(
devloveC-
Length of incubation (week)
Figure 1. Cumulative CO2-C evolved from soils amended with cacao pod and water hyacinth composts over 16 weeks of incubation in the laboratory. F: Inorganic N and P; CC: cacao pod compost; WC: water hyacinth compost; 1: 2.5 t ha−1; 2: 5.0 t ha−1.
2472 C. T. ATERE ET AL.
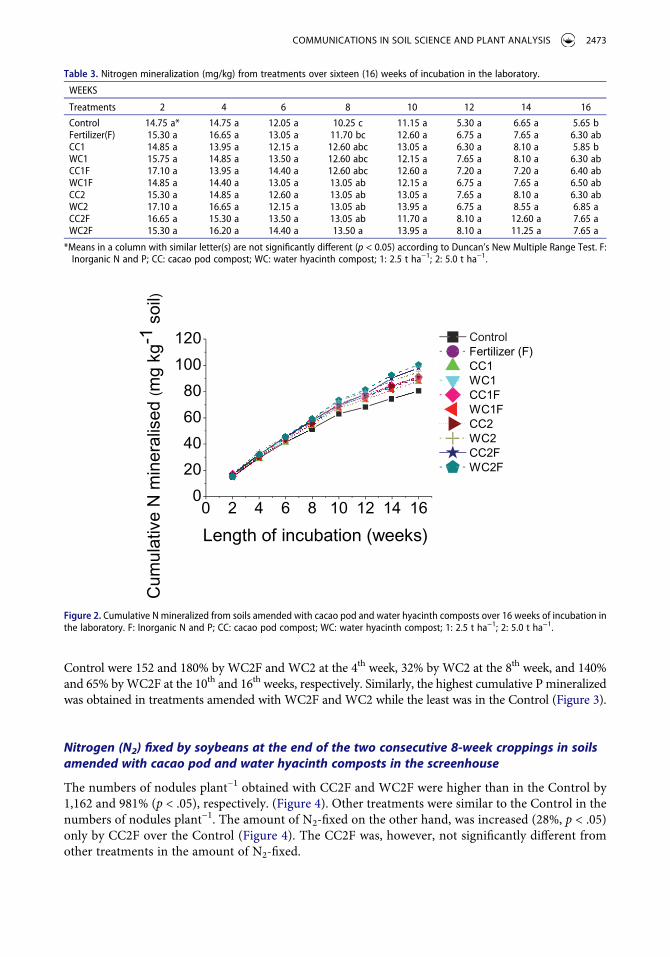
Control were 152 and 180% by WC2F and WC2 at the 4th week, 32% by WC2 at the 8th week, and 140% and 65% by WC2F at the 10th and 16th weeks, respectively. Similarly, the highest cumulative P mineralized was obtained in treatments amended with WC2F and WC2 while the least was in the Control (Figure 3).
Nitrogen (N2) fixed by soybeans at the end of the two consecutive 8-week croppings in soils amended with cacao pod and water hyacinth composts in the screenhouse
The numbers of nodules plant−1 obtained with CC2F and WC2F were higher than in the Control by 1,162 and 981% (p < .05), respectively. (Figure 4). Other treatments were similar to the Control in the numbers of nodules plant−1. The amount of N2-fixed on the other hand, was increased (28%, p < .05) only by CC2F over the Control (Figure 4). The CC2F was, however, not significantly different from other treatments in the amount of N2-fixed.
Table 3. Nitrogen mineralization (mg/kg) from treatments over sixteen (16) weeks of incubation in the laboratory.
WEEKS
Treatments 2 4 6 8 10 12 14 16
Control 14.75 a* 14.75 a 12.05 a 10.25 c 11.15 a 5.30 a 6.65 a 5.65 bFertilizer(F) 15.30 a 16.65 a 13.05 a 11.70 bc 12.60 a 6.75 a 7.65 a 6.30 abCC1 14.85 a 13.95 a 12.15 a 12.60 abc 13.05 a 6.30 a 8.10 a 5.85 bWC1 15.75 a 14.85 a 13.50 a 12.60 abc 12.15 a 7.65 a 8.10 a 6.30 abCC1F 17.10 a 13.95 a 14.40 a 12.60 abc 12.60 a 7.20 a 7.20 a 6.40 abWC1F 14.85 a 14.40 a 13.05 a 13.05 ab 12.15 a 6.75 a 7.65 a 6.50 abCC2 15.30 a 14.85 a 12.60 a 13.05 ab 13.05 a 7.65 a 8.10 a 6.30 abWC2 17.10 a 16.65 a 12.15 a 13.05 ab 13.95 a 6.75 a 8.55 a 6.85 aCC2F 16.65 a 15.30 a 13.50 a 13.05 ab 11.70 a 8.10 a 12.60 a 7.65 aWC2F 15.30 a 16.20 a 14.40 a 13.50 a 13.95 a 8.10 a 11.25 a 7.65 a
*Means in a column with similar letter(s) are not significantly different (p < 0.05) according to Duncan’s New Multiple Range Test. F: Inorganic N and P; CC: cacao pod compost; WC: water hyacinth compost; 1: 2.5 t ha−1; 2: 5.0 t ha−1.
0 2 4 6 8 10 12 14 160
20
40
60
80
100
120 ControlFertilizer (F)CC1WC1CC1FWC1FCC2WC2CC2FWC2F
desilarenim
Nevitalu
muC
(gk
gm
-1lios)
Length of incubation (weeks)
Figure 2. Cumulative N mineralized from soils amended with cacao pod and water hyacinth composts over 16 weeks of incubation in the laboratory. F: Inorganic N and P; CC: cacao pod compost; WC: water hyacinth compost; 1: 2.5 t ha−1; 2: 5.0 t ha−1.
COMMUNICATIONS IN SOIL SCIENCE AND PLANT ANALYSIS 2473
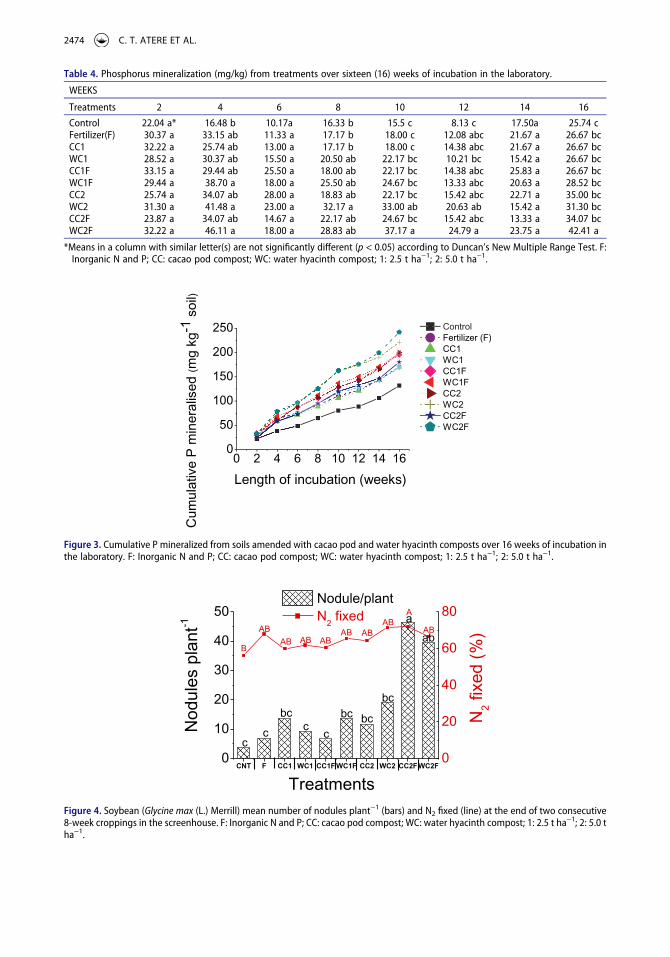
Table 4. Phosphorus mineralization (mg/kg) from treatments over sixteen (16) weeks of incubation in the laboratory.
WEEKS
Treatments 2 4 6 8 10 12 14 16
Control 22.04 a* 16.48 b 10.17a 16.33 b 15.5 c 8.13 c 17.50a 25.74 cFertilizer(F) 30.37 a 33.15 ab 11.33 a 17.17 b 18.00 c 12.08 abc 21.67 a 26.67 bcCC1 32.22 a 25.74 ab 13.00 a 17.17 b 18.00 c 14.38 abc 21.67 a 26.67 bcWC1 28.52 a 30.37 ab 15.50 a 20.50 ab 22.17 bc 10.21 bc 15.42 a 26.67 bcCC1F 33.15 a 29.44 ab 25.50 a 18.00 ab 22.17 bc 14.38 abc 25.83 a 26.67 bcWC1F 29.44 a 38.70 a 18.00 a 25.50 ab 24.67 bc 13.33 abc 20.63 a 28.52 bcCC2 25.74 a 34.07 ab 28.00 a 18.83 ab 22.17 bc 15.42 abc 22.71 a 35.00 bcWC2 31.30 a 41.48 a 23.00 a 32.17 a 33.00 ab 20.63 ab 15.42 a 31.30 bcCC2F 23.87 a 34.07 ab 14.67 a 22.17 ab 24.67 bc 15.42 abc 13.33 a 34.07 bcWC2F 32.22 a 46.11 a 18.00 a 28.83 ab 37.17 a 24.79 a 23.75 a 42.41 a
*Means in a column with similar letter(s) are not significantly different (p < 0.05) according to Duncan’s New Multiple Range Test. F: Inorganic N and P; CC: cacao pod compost; WC: water hyacinth compost; 1: 2.5 t ha−1; 2: 5.0 t ha−1.
0 2 4 6 8 10 12 14 160
50
100
150
200
250 ControlFertilizer (F)CC1WC1CC1FWC1FCC2WC2CC2FWC2F
desilarenim
Pevitalu
muC
(gk
gm
-1lios)
Length of incubation (weeks)
Figure 3. Cumulative P mineralized from soils amended with cacao pod and water hyacinth composts over 16 weeks of incubation in the laboratory. F: Inorganic N and P; CC: cacao pod compost; WC: water hyacinth compost; 1: 2.5 t ha−1; 2: 5.0 t ha−1.
CNT F CC1 WC1 CC1F WC1F CC2 WC2 CC2FWC2F0
10
20
30
40
50AB
AAB
ABABABABAB
AB
Bab
a
bc
bcbc
cc
bc
c
N2 fi
xed
(%)
Nodule/plant
tnalpseludo
N-1
Treatments
c0
20
40
60
80 N2 fixed
Figure 4. Soybean (Glycine max (L.) Merrill) mean number of nodules plant−1 (bars) and N2 fixed (line) at the end of two consecutive 8-week croppings in the screenhouse. F: Inorganic N and P; CC: cacao pod compost; WC: water hyacinth compost; 1: 2.5 t ha−1; 2: 5.0 t ha−1.
2474 C. T. ATERE ET AL.

Discussion
Dynamics of microbial respiration, and N and P mineralization in cacao pod- and water hyacinth-amended soils during laboratory incubation
The C flush observed at the second week of incubation could arise from the hitherto inactive microorganisms becoming more active with the addition of carbon substrates, and having overcome the lag time required to adjust to the new system (Miransari 2013). At this point also, the native soil organic matter could become more susceptible to microbial attack due to positive priming (Zhu et al. 2016), resulting in increased microbial respiration (Fageria 2012; Oladipo et al. 2010). The CO2-C released thereafter decreased up to the end of incubation as a result of decline in the food materials available for microbial growth. The increasing resistance of the remaining native and added organic matter to microbial decomposition with length of incubation when the more easily decomposable compounds were getting exhausted, could also contribute to the observed trend. In a similar study by Hossain et al. (2017), when organic materials were added to soil, the CO2 emission reached the peak at 5th week of incubation and then decreased with irregular fashion until 21st week. The earlier peak of CO2 emission (at the 2nd week) in our study could be attributed to the composting of the materials before adding them to soil, making them more easily attacked by microorganisms. The higher microbial activity indicated by the increased CO2-C release at higher application rate (5.0 t ha−1) of the composts was because of the richness of the organic amendments in carbon and basic nutrients such as N, P, K, Ca, and Mg (Table 1). Thus, the composts were readily available sources of cell C and energy to the microorganisms. Conversely, the treatment containing only fertilizer (inorganic N and P) was not different from the Control in term of C release due to the absence of organic materials which could serve as energy source for the microorganisms.
The increase in N and P mineralized mainly by 5.0 t ha−1 water hyacinth compost with and without inorganic N and P addition could be attributed to the richness of the compost material in N and P, and its lower C: N ratio than cacao pod compost. It was found that the addition to soil of organic amendments with C:N ratios less than 20:1 resulted in net mineralization of N for plant uptake (Masunga et al. 2015). Fortifying the composts with starter inorganic N and P also improved the compost performance by providing the immediate nutrients required by microorganisms, thereby aiding mineralization of the organic nutrients. This fortification was also going to improve plant nutrient uptake prior to release of nutrients from the composts. The significantly higher (p < .05) amounts of CO2-C evolved from these treatments having higher mineralized N and P (Table 2) was a further indication of available nutrients, especially, N and P (Zong et al. 2015). Microbial prolifera-tion is enhanced in the presence of nutrients (Gilani and Bahmanyar 2008) which favors mineraliza-tion of immobilized nutrients for plant uptake (Franzluebbers 2004). This high microbial activity, especially, as early as the second week of compost addition was expected to aid early release of nutrients for improved plant growth, .
Nitrogen (N2) fixed by soybeans in cacao pod- and water hyacinth-amended soils under screenhouse conditions
The results obtained from the current study indicated that the N2 fixing rhizobia could benefit from the additions of composts, especially when fortified with small amounts of inorganic nutrients. Applications of water hyacinth and cacao pod composts increased microbial activity by serving as their sources of energy and cell carbon. In return, the locked-up nutrients in the composts were mineralized for the plant’s uptake. The applied composts were rich in mineralizable plant nutrients, including N and P. Nitrogen is an essential component of chlorophyll and thus, plays key role in photosynthesis (Mehata 2018). Phosphorus on the other hand, is required for physiological growth, root development, protein synthesis, promotion of meristematic tissue and carbohydrate formation (Malhotra, Sharma, and Pandey 2018). Phosphorus had also been reported to be directly related to nodulation, N2 fixation, and specific nodule activity in legumes (Míguez-Montero, Valentine, and
COMMUNICATIONS IN SOIL SCIENCE AND PLANT ANALYSIS 2475

Pérez-Fernández 2019). Nodulation and N2 fixed were increased at higher application rate (5 t ha−1) of the water hyacinth and cacao pod composts with starter amounts of inorganic N and P. These rates corresponded with the points at which larger amounts of N and P were mineralized (Figures 2 & 3), indicating the nexus between organic-material input, microbial activity and N2 fixation by legumes.
Conclusion
This findings from the study showed that application of cacao pod and water hyacinth composts with and without starter N and P increased the soil microbial activity. The microbial activity was, however, much higher with water hyacinth than with cacao pod composts. The supply of the composts at 5 t ha−1 with starter amounts of N and P exhibited higher influence on soil microbial activity, and increased soybean nodulation and nitrogen fixation. It was concluded that stimulation in microbial activity resulting from addition to soil of 5 t ha−1 cacao pod or water hyacinth composts with starter N and P aided mineralization of nutrients for increased soybean nodulation and nitrogen fixation.
Declaration of interest statement
The authors declare that there is no conflict of interest.
ORCID
Cornelius Talade Atere http://orcid.org/0000-0002-4885-4051Michael Omofowa Osunde http://orcid.org/0000-0003-4934-2784Akin Olayinka http://orcid.org/0000-0003-3990-9599
References
Abdelsabour, M. F. 2010. Water hyacinth: Available and renewable resource. Electronic Journal of Environmental Agriculture and Food Chemistry 9 (11):1746–59.
Adesanwo, O. O., M. T. Adetunji, J. K. Adesanwo, O. A. Osiname, S. Diatta, and D. O. Torimiro. 2009. Evaluation of traditional soil fertility management practices for rice cultivation in Southwestern Nigeria. American-Eurasian Journal of Agronomy 2 (2):45–49.
Adino, S., Z. Wondifraw, and M. Addis. 2018. Replacement of Soybean grain with Cowpea grain (Vigna Unguiculata) as protein supplement in Sasso x Rir crossbred chicks diet. Poultry, Fisheries and Wildlife Sciences 188 (6):2–6.
Aislabie, J., and J. R. Deslippe. 2013. Soil Microbes and their Contribution to Soil Services. In Ecosystem services in New Zealand – Conditions and trends, ed. J. R. Dymond, 143–61. Lincoln, New Zealand: Manaaki Whenua Press.
Alexander, J., D. Benford, A. Cockburn, J. P. Cravedi, E. Dogliotti, and A. D. Dominico. 2008. Theobromine as undesirable substances in animal feed: Scientific opinion of the panel on contaminants in the food chain. European Food Safety Authority Journal 725:1–66.
Allison, L. E. 1965. Organic carbon. In Methods of soil analysis, part 2, chemical and microbiological properties, ed. C. A. Black, 1367–78. Madison: American Society of Agronomy.
Anderson, J. 1982. Soil respiration. In Methods of soil analysis, A. L. Page, R. H. Miller, and D. R. Keeney. ed., Vol. 9, 831–66. Madison, WI, Agron: American Society of Agronomy and Soil Science of America.
Anjanabha, B., and P. Kumar. 2010. Water hyacinth as a potential biofuel crop. Electronic Journal of Environmental, Agriculture and Food Chemistry 9 (1):112–22.
Atere, C. T., and A. Olayinka. 2013. Soil chemical properties and growth of maize (Zea mays l.) as affected by cocoa pod compost-based N and P fertilizer. Nigerian Journal of Soil Science 23 (1):83–93.
Atere, C. T., and A. Olayinka. 2019. Enhancing maize (Zea mays l.) growth and nutrient uptake via application of water hyacinth (Eichornia crassipes [Mart.] Solms) compost and inorganic nutrients. Nigerian Journal of Soil Science 29 (1):27–34.
Blake, G. R., and K. H. Hartge. 1986. Bulk density. In Methods of soil analysis, A. Klute, A. ed., part I: Physical and mineralogical methods‟, 2nd ed.,363–76. Madison, WI: ASA Monograph.
Bouyoucos, G. J. 1962. Hydrometer method improved for making particle size analysis of soils. Soil Science Society of America Proceedings 26:464–65.
2476 C. T. ATERE ET AL.

Bray, R. H., and L. T. Kurtz. 1945. Determination of total, organic, and available forms of phosphorus in soils. Soil Science 59:39–45.
Bremner, J. M., and C. S. Mulvaney. 1982. Nitrogen-Total. In Methods of Soil Analysis, A. L. Page and R. H. Miller. ed., 2nd ed ed., 595–624. Madison, WI: American Society of Agronomy. Agronomy Monograph 9, Part 2.
Bremner, J. M., and D. R. Keeney. 1966. Determination of Isotope-ratio analysis of different forms of Nitrogen in Soil. Soil Science Society of America Proceedings 30:577–82.
Brookshire, E. N. J., N. Wurzburger, B. Currey, D. N. L. Menge, M. P. Oatham, and C. Roberts. 2019. Symbiotic nitrogen fixation is sufficient to support net aboveground biomass accumulation in a humid tropical forest. Scientific Report 7571 (9):2–10.
Dent, D., and E. Cocking. 2017. Establishing symbiotic nitrogen fixation in cereals and other non-legume crops: The greener nitrogen revolution. Agric and Food Secur 6 (7):1–9. doi:10.1186/s40066-016-0084–2.
Fageria, N. K. 2012. Role of soil organic matter in maintaining sustainability of cropping systems. Communications in Soil Science and Plant Analysis 43 (16):2063–113.
Fidelis, C., and R. B. K. Rao. 2017. Enriched cocoa pod composts and their fertilizing effects on hybrid cocoa seedlings. International Journal of Recycling of Organic Waste in Agriculture 6:99–106. doi:10.1007/s40093-017-0156-8.
Food and Agriculture Organization of the United Nations Statistics Division (FAOSTAT). 2018. Accessed May 6, 2020. http://www.fao.org/faostat/en/#data/QC
Franzluebbers, A. J. 2004. Organic Residues, Decomposition. In Encyclopedia of Soils in the Environment, ed. J. L. Hatfield, D. S. Powlson, C. Rosenzweig, K. M. Scow, M. J. Singer, and D. L. Sparks, Vol. 4, 112–13. Amsterdam: Elsevier Academic Press.
Gilani, S. S., and M. A. Bahmanyar. 2008. Impact of organic Amendment with and without Mineral Fertilizers on soil microbial respiration. Journal of Applied Sciences 8 (4):642–47.
Hauggaard-Nielsen, H., P. Ambus, and E. S. Jensen. 2001. Temporal and spatial distribution of roots and competition for nitrogen in pea-barley intercrops – A field study employing 32P technique. Plant and Soil 236:63–74.
Herridge, D. F., F. J. Bergersen, and M. B. People. 1990. Measurement of nitrogen fixation by nodulated soybean calibrated by 15-N methods. Plant Physiology 93:495–503.
Herridge, D. F., P. O’Connell, and K. Donnelly. 1988. The Xylem Ureide Assay of Nitrogen fixation: Sampling Procedures and Sources of Error. Journal of Experimental Botany 39:12–22.
Hillel, D. 2008. Soil biodiversity. Soil in the Environment 163–74. doi:10.1016/C2009-0-00041-5.Hossain, M. B., M. M. Rahman, J. C. Biswas, M. U. Miah, S. Akhter, Maniruzzaman, A. K. Choudhury, F. Ahmed,
H. K. Shiragi, and N. Kalra. 2017. Carbon mineralization and carbon dioxide emission from organic matter added soil under different temperature regimes. International Journal of Recycling of Organic Waste in Agriculture 6:311–19. doi:10.1007/s40093-017-0179-1.
Ilstedt, U., A. Nordgren, and A. Malmer. 2000. Optimum soil water for soil respiration before and after amendment with glucose in humid tropical acrisols and a boreal mor layer. Soil Biology & Biochemistry 32 (11–12):1591–99.
Jacoby, R., M. Peukert, A. Succurro, A. Koprivova, and S. Kopriva. 2017. The role of soil microorganisms in plant mineral nutrition—current knowledge and future directions. Frontiers of Plant Science 8:1617. doi:10.3389/ fpls.2017.01617.
Kuke, A. C., and K. K. Hunde. 2019. Evaluation of water hyacinth (Eichornia crassipes) compost for its nutrient content at adami tulu agricultural research center. Innovative Techniques in Agriculture 3 (4):690–96.
Lin, W., M. Lin, H. Zhou, H. Wu, Z. Li, and W. Lin. 2019. The effects of Chemical and Organic Fertilizer Usage on Rhizosphere Soil in Tea Orchards. PLoS ONE 14 (5):1–16 e0217018. doi:10.1371/journal.pone.0217018.
Mahmud, K., S. Makaju, R. Ibrahim, and A. Missaoui. 2020. Current progress in nitrogen fixing plants and microbiome research. Plants 9 (97):1–17. doi:10.3390/plants9010097.
Malhotra, H., V. S. Sharma, and R. Pandey. 2018. Phosphorus nutrition: Plant growth in response to deficiency and excess. In Plant nutrients and abiotic stress tolerance, M. Hasanuzzaman, M. Fujita, H. Oku, K. Nahar, and B. Hawrylak-Nowak. ed.. Singapore: Springer. doi:10.1007/978-981-10-9044-8_7.
Masunga, R. H., V. N. Uzokwe, P. D. Mlay, I. Odeh, A. Singh, D. Buchan, and S. De Neve. 2015. Nitrogen mineralization dynamics of different valuable organic amendments commonly used in agriculture. Applied Soil Ecology 101:185–93.
Matsumura, Y. 2002. Evaluation of supercritical water gasification and biomethanation for wet biomass utilization in Japan. Energy Conversion and Management 43:1301–10.
McKeown, E., and G. Bugyi. 2016. Impact of water pollution on human health and environmental sustainability. Progress, and Proficiency in Sustainability (PPPS) Book Series, Hershey PA, USA, 346–65
Mehata, M. 2018. Measuring the influence of bedding on soil nitrogen losses and corn crop nitrogen characteristics for fall applied solid beef cattle manure in Eastern South Dakota. Electronic Theses and Dissertations 2437. https:// openprairie.sdstate.edu/etd/2437.
Míguez-Montero, M. A., A. Valentine, and M. A. Pérez-Fernández. 2019. Regulatory effect of phosphorus and nitrogen on nodulation and plant performance of leguminous shrubs. AoB PLANTS 11:plz047. doi:10.1093/aobpla/plz047.
Miransari, M. 2013. Soil microbes and the availability of soil nutrients. Acta Physiologiae Plantarum 35:3075–84.OECD-FAO. 2019. Oilseeds and oilseed products. In Agricultural outlook 2019–2028, 143–53. Chap 4. Paris: Food and
Agriculture Organization of the United Nations
COMMUNICATIONS IN SOIL SCIENCE AND PLANT ANALYSIS 2477

Ojetade, J. O., O. A. Fawole, O. M. Faturoti, and A. A. Amusan. 2016. Assessment of variability of soil properties under different vegetations in an Ultisol in Ile Area, Osun State, Nigeria. Ife Journal of Agriculture 28:1–12.
Oladipo, O. G., A. Olayinka, O. T. Aladesanmi, M. Sanni, A. J. Famurewa, and W. O. Siyanbola. 2010. Risk mitigation and policy implication for carbon dioxide (CO2) emission in organically-amended soils in Nigeria. African Journal of Environmental Science and Technology 4 (11):801–06.
Page, A. L., R. H. Miller, and D. R. Keeney. 1982. Methods of soil analysis, part 2. Chemical and microbiological properties. Agronomy No, 831–66. 92nd ed. Madison, Wisconsin, USA: American Society of Agronomy.
Peech, M., R. A. Olsen, and G. H. Bolt. 1953. The significance of potentiometric measurements involving liquid junction in clay and soil suspensions. Soil Science Society Am Proceedings 17:214–20.
Taiwo, O. E., and F. A. O. Osinowo. 2001. Evaluation of various agro-wastes for traditional black soap production. Bioresource Technology 79:95–97.
Timsina, J. 2018. Can organic sources of nutrients increase crop yield to meet global food demand? Agronomy 214 (8):1–20. doi:10.103390/agronomy8100214.
Vieira, R. F., I. C. Mendes, F. B. Reis-Junior, and M. Hungria. 2010. Symbiotic Nitrogen Fixation in Tropical Food Grain Legumes: Current Status. In Microbes for Legume Improvement, M. S. Khan, J. Musarrat, and A. Zaidi. ed.. , Microbes for Legume Improvement, 427–472, Vienna: Springer-Verlag/Wien. doi:10.1007/978-3-211-99753-6_18.
Vriesmann, L. C., R. F. Teofilo, and C. L. D. Petkowicz. 2012. Extraction and characterization of pectin from Cacao Pod Husks (Theobroma cacao L.) with citric acid. LWT 49 (1):108–16.
Zhu, Z., G. Zeng, T. Ge, Y. Hu, C. Tong, O. Shibistova, X. He, J. Wang, G. Guggenberger, and J. Wu. 2016. Fate of rice shoot and root residues, rhizodeposits, and microbeassimilated carbon in paddy soil — Part 1: Decomposition and priming effect. Biogeosciences 13:4481–89. doi:10.5194/bg-13-4481-2016.
Zong, N., J. Jiang, P. Shi, M. Song, Z. Shen, and X. Zhang. 2015. Soil Microbial Respiration with Climatic Factors in an Alpine Meadow. The Scientific World Journal 617471:11. doi:10.1155/2015/617471.
2478 C. T. ATERE ET AL.
Related Documents











