Journal of General Virology (1998), 79, 1469–1477. Printed in Great Britain ................................................................................................................................................................................................................................................................................... MHC class I molecules are enriched in caveolae but do not enter with simian virus 40 Howard A. Anderson, 2 † Yuzhi Chen 3 ‡ and Leonard C. Norkin 1, 2, 3 Department of Microbiology 1 , Graduate Program in Molecular and Cellular Biology 2 , and Graduate Program in Neuroscience and Behavior 3 , University of Massachusetts Amherst, Amherst, MA 01003, USA Simian virus 40 (SV40) binds to MHC class I molecules anywhere on the cell surface and then enters through caveolae. The fate of class I mol- ecules after SV40 binding is not known. Sensitivity of 125 I-surface-labelled class I molecules to papain cleavage was used to distinguish internalized class I molecules from class I molecules remaining at the cell surface. Whereas the caveolae-enriched mem- brane microdomain was found to also be enriched for class I molecules, no internalized papain-re- sistant 125 I-surface-labelled class I molecules could be detected at any time in either control cells or in Introduction Simian virus 40 (SV40) initiates infection by binding to class I molecules encoded by the major histocompatibility complex (MHC) [human lymphocyte antigens (HLA) in humans] (Atwood & Norkin, 1989 ; Breau et al., 1992). MHC class I molecules are best known for their role in presenting endogenously synthesized antigenic peptide fragments for recognition at the cell surface by cytotoxic T lymphocytes. These molecules are heterodimers, composed of a 44 kDa transmembrane heavy chain and a noncovalently associated 12 kDa β # microglobulin light chain. Viruses that enter cells by endocytosis are generally considered to enter through clathrin-coated pits (Marsh & Helenius, 1989). In contrast, SV40 enters cells through caveolae (Anderson et al., 1996; Stang et al., 1997), which are flask- shaped plasma membrane invaginations, approximately 70 to 100 nm wide. Caveolae are distinguished from clathrin-coated Author for correspondence : Leonard C. Norkin. Fax ›1 413 545 1578. e-mail lnorkin!microbio.umass.edu † Present address : National Cancer Institute, Bethesda, MD 20892, USA. ‡ Present address : Department of Microbiology and Molecular Genetics, Harvard Medical School, Boston, MA 02115, USA. cells preadsorbed with saturating amounts of SV40. Instead, 125 I-surface-labelled class I molecules, as well as preadsorbed 125 I-labelled anti-class I anti- bodies, accumulated in the medium, coincident with the turnover of class I molecules at the cell surface. The class I heavy chains that accumulated in the medium were truncated and their release was specifically prevented by the metalloprotease in- hibitor 1,10-phenanthroline. Thus, whereas class I molecules mediate SV40 binding, they do not appear to mediate SV40 entry. pits in electron microscopy images by their size and shape. Also, caveolae contain a characteristic marker protein called caveolin (Rothberg et al., 1992). The regular functions of caveolae are not yet entirely clear, but they have been implicated in endocytosis, transcytosis, potocytosis and in- tracellular signalling (Montesano et al., 1982 ; Simionescu et al., 1982 ; Anderson, 1993 a, b ; Lisanti et al., 1994 a, b ; Schnitzer et al., 1995 a, 1996). In general, little is known about the fate of virus receptors following virus binding. We are interested here in the fate of MHC class I molecules following SV40 binding for several reasons. First, an earlier study reported that cross-linking class I molecules with antibodies against β # microglobulin induced the internalization of class I molecules through small non- coated plasma membrane invaginations that would now be recognized as caveolae (Huet et al., 1980). Second, the caveolae- mediated SV40 entry pathway delivers the virus to the endoplasmic reticulum (ER), rather than to the endosomal} lysosomal compartment, which is the usual target for endocytic traffic (Kartenbeck et al., 1989 ; Norkin & Anderson, 1996). Since MHC class I molecules acquire antigenic peptides in the ER (Cox et al., 1990), and since it is not clear whether class I molecules at the cell surface might recycle to the ER, we were interested in the possibility that SV40 might target the ER through its association with class I molecules. Several studies provided evidence that class I molecules are 0001-5435 # 1998 SGM BEGJ

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Journal of General Virology (1998), 79, 1469–1477. Printed in Great Britain. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
MHC class I molecules are enriched in caveolae but do notenter with simian virus 40
Howard A. Anderson,2† Yuzhi Chen3‡ and Leonard C. Norkin1, 2, 3
Department of Microbiology1, Graduate Program in Molecular and Cellular Biology2, and Graduate Program in Neuroscience andBehavior3, University of Massachusetts Amherst, Amherst, MA 01003, USA
Simian virus 40 (SV40) binds to MHC class Imolecules anywhere on the cell surface and thenenters through caveolae. The fate of class I mol-ecules after SV40 binding is not known. Sensitivityof 125I-surface-labelled class I molecules to papaincleavage was used to distinguish internalized class Imolecules from class I molecules remaining at thecell surface. Whereas the caveolae-enriched mem-brane microdomain was found to also be enrichedfor class I molecules, no internalized papain-re-sistant 125I-surface-labelled class I molecules couldbe detected at any time in either control cells or in
IntroductionSimian virus 40 (SV40) initiates infection by binding to
class I molecules encoded by the major histocompatibilitycomplex (MHC) [human lymphocyte antigens (HLA) inhumans] (Atwood & Norkin, 1989 ; Breau et al., 1992). MHCclass I molecules are best known for their role in presentingendogenously synthesized antigenic peptide fragments forrecognition at the cell surface by cytotoxic T lymphocytes.These molecules are heterodimers, composed of a 44 kDatransmembrane heavy chain and a noncovalently associated12 kDa β
#microglobulin light chain.
Viruses that enter cells by endocytosis are generallyconsidered to enter through clathrin-coated pits (Marsh &Helenius, 1989). In contrast, SV40 enters cells through caveolae(Anderson et al., 1996 ; Stang et al., 1997), which are flask-shaped plasma membrane invaginations, approximately 70 to100 nm wide. Caveolae are distinguished from clathrin-coated
Author for correspondence: Leonard C. Norkin.
Fax 1 413 545 1578. e-mail lnorkin!microbio.umass.edu
† Present address: National Cancer Institute, Bethesda, MD 20892,
USA.
‡ Present address: Department of Microbiology and Molecular
Genetics, Harvard Medical School, Boston, MA 02115, USA.
cells preadsorbed with saturating amounts of SV40.Instead, 125I-surface-labelled class I molecules, aswell as preadsorbed 125I-labelled anti-class I anti-bodies, accumulated in the medium, coincident withthe turnover of class I molecules at the cell surface.The class I heavy chains that accumulated in themedium were truncated and their release wasspecifically prevented by the metalloprotease in-hibitor 1,10-phenanthroline. Thus, whereas class Imolecules mediate SV40 binding, they do notappear to mediate SV40 entry.
pits in electron microscopy images by their size and shape.Also, caveolae contain a characteristic marker protein calledcaveolin (Rothberg et al., 1992). The regular functions ofcaveolae are not yet entirely clear, but they have beenimplicated in endocytosis, transcytosis, potocytosis and in-tracellular signalling (Montesano et al., 1982 ; Simionescu et al.,1982 ; Anderson, 1993a, b ; Lisanti et al., 1994a, b ; Schnitzer etal., 1995a, 1996).
In general, little is known about the fate of virus receptorsfollowing virus binding. We are interested here in the fate ofMHC class I molecules following SV40 binding for severalreasons. First, an earlier study reported that cross-linking classI molecules with antibodies against β
#microglobulin induced
the internalization of class I molecules through small non-coated plasma membrane invaginations that would now berecognized as caveolae (Huet et al., 1980). Second, the caveolae-mediated SV40 entry pathway delivers the virus to theendoplasmic reticulum (ER), rather than to the endosomal}lysosomal compartment, which is the usual target for endocytictraffic (Kartenbeck et al., 1989 ; Norkin & Anderson, 1996).Since MHC class I molecules acquire antigenic peptides in theER (Cox et al., 1990), and since it is not clear whether class Imolecules at the cell surface might recycle to the ER, we wereinterested in the possibility that SV40 might target the ERthrough its association with class I molecules.
Several studies provided evidence that class I molecules are
0001-5435 # 1998 SGM BEGJ

H. A. Anderson, Y. Chen and L. C. NorkinH. A. Anderson, Y. Chen and L. C. Norkin
internalized by T cells (Capps et al., 1989 ; Dasgupta et al.,1988 ; Machy et al., 1987 ; Tse & Pernis, 1984 ; Vega &Strominger, 1989), and perhaps other cells as well (Huet et al.,1980 ; Reid & Watts, 1990). However, internalization was notobserved in other studies (Eichholtz et al., 1992 ; Neefjes et al.,1990, 1992). These discrepancies might be due to cell type-specific differences and perhaps to differences in experimentalapproach. Regardless, although we found that the caveolin-containing membrane microdomain is enriched for MHC classI molecules, we could not detect any spontaneous, or antibody-induced, or SV40-induced internalization of class I moleculeson CV-1 monkey kidney fibroblasts, which are standardpermissive host cells for SV40. Instead, class I moleculesappeared to be spontaneously released from the cell surface bythe action of a metalloprotease. These results imply theexistence of a putative coreceptor for SV40 that might benecessary for SV40 to enter and infect cells.
Methods+ Chemicals. The following chemicals were purchased from Sigma :1,10-phenanthroline, phenylmethylsulfonyl fluoride (PMSF), aprotinin,leupeptin, soybean trypsin inhibitor, papain and lactoperoxidase. A1±0 M stock of 1,10-phenanthroline was prepared before use in 95%ethanol. The protease inhibitors PMSF, aprotinin, leupeptin and soybeantrypsin inhibitor were prepared, and stored, as described by Harlow &Lane (1988). Papain was reconstituted to a concentration of 7±5 mg}ml inPBS (pH 7±2), and frozen at ®20 °C until use. Lactoperoxidase wasreconstituted with PBS to 268 units}ml (2±5 mg}ml), and stored at®20 °C until use.
+ Cells and virus. CV-1 African green monkey kidney fibroblastswere purchased from the ATCC and maintained in DMEM containing10% foetal bovine serum (Atlanta Biologicals). SV40 wild-type strain 776was triple plaque-purified and grown by infecting CV-1 cells at an m.o.i.of 1 p.f.u. per cell. Virus lysates, harvested at 3 days, were frozen andthawed three times, and cell debris was removed by centrifugation at15000 g. Virus titres were determined by plaque assay on CV-1 cells.
+ Antibodies. W6}32 hybridoma (Parham et al., 1979) was obtainedfrom the American Type Tissue Culture Association, and maintained inRPMI containing 10% FBS until cells reached maximum density.Hybridoma cells were removed by centrifugation at 300 g, and W6}32tissue culture supernatant was frozen until use. W6}32 hybridoma wasalso injected into mice for ascites production (Harlow & Lane, 1988).Rabbit antiserum specific for a peptide sequence on the cytoplasmic tailof human MHC class I heavy chain (Rougon et al., 1984), and denaturedHLA-B27 (Neefjes et al., 1986) were generous gifts from H. L. Ploegh.
+ Cell surface iodination. CV-1 cells were grown to confluency andwashed twice with PBS. Radiolabelling was performed as described byPhilips & Morrison (1970). Lactoperoxidase (90 µg}ml, 10 units}ml) in1 ml of PBS was added to each plate. Na"#&I (0±25 mCi ; ICN) was thenadded to each plate, followed by addition of 37 µl of H
#O
#(7 µM final
concentration), every minute, for 5 min. Cells were incubated for 5 min,and the iodination reaction was stopped by addition of 2 ml of stopbuffer (PBS, 1 mg}ml tyrosine). Cells were incubated for an additional5 min, and washed twice with PBS. After iodination, cell viability wasgreater than 90%, as judged by trypan blue dye exclusion. Cells did notdisplay any altered morphology.
+ Immunoprecipitations and SDS–PAGE. CV-1 cells were sol-ubilized with 500 µl of lysis buffer (50 mM Tris–HCl, 1 mM MgCl
#, 1%
NP40, 1 mM PMSF, 15 µg}ml aprotinin, 2 µg}ml leupeptin, 5 mM 1,10-phenanthroline, 10 µg}ml soybean trypsin inhibitor (pH 7±4) for 1 h onice. Insoluble material was removed by centrifugation at 15000 g for5 min. Protein concentrations were measured using the Bio-Rad proteinassay (Bradford, 1976). Extracts were precleared with 5 µl of normalrabbit serum, for 1 h on ice, followed by addition of 100 µl of formalin-fixed Staphylococcus aureus for 1 h on ice. S. aureus was pelleted bycentrifugation at 15000 g for 5 min. Extracts were usually preclearedthree times to remove proteins that bound nonspecifically to immuno-precipitation complexes. MHC class I proteins were immunoprecipitatedwith 100 µl of W6}32 tissue culture supernatant fluid or with 5 µl ofrabbit antiserum specific for the MHC class I cytoplasmic tail. Antibodieswere added to cell extracts for 1 h, followed by the addition of 100 µl ofS. aureus, for 1 h. To immunoprecipitate class I proteins using antiserumspecific for denatured class I proteins, samples were first boiled for10 min. Samples were cooled on ice, precleared, and class I proteins wereimmunoprecipitated with 5 µl of antiserum against denatured HLA-B27,for 1–3 h, followed by the addition of 100 µl of S. aureus. Allimmunoprecipitates were washed three times with 500 µl of lysis buffer,resuspended in 50 µl of sample buffer (63 mM Tris, 10% glycerol, 2%SDS, 40 mM DTT, 0±01% bromphenol blue), and boiled for 10 min.Samples were then resolved by SDS–PAGE. Gels were fixed with 25%methanol and 5% acetic acid, dried, and visualized by autoradiography.
+ Papain treatment to remove surface proteins. Cell surfaceproteins were labelled with "#&I. Cells were then washed and incubatedfor up to 3 h at 37 °C in DMEM containing 10% FBS. At the indicatedtimes cells were detached by gently scraping monolayers with a rubberpoliceman and then treated with papain (1 mg}ml) in 500 µl of PBScontaining 1 mM EDTA for 1 h on ice in glass sealed vials that werepulsed for 10 s with N
#gas. Viability of cells after this procedure was
routinely about 80% as judged by trypan blue dye exclusion. Cells werewashed three times with PBS to remove papain, and solubilized with lysisbuffer.
Papain was also used to remove "#&I-labelled W6}32 and "#&I-labelledtransferrin prebound to cells. W6}32 was purified from mouse ascitesusing a protein A affinity column (Harlow & Lane, 1988). Purified W6}32and transferrin were labelled with "#&I by the chloramine T method(Parham et al., 1979). "#&I-labelled W6}32 or "#&I-labelled transferrin werebound to cells on ice in PBS for 1 h, and cells were washed three times.Cells were cultured for various lengths of time at 37 °C in DMEMcontaining 10% FBS. Cells were then treated with papain as describedabove, washed, and the amount of radioactivity associated with cells afterpapain treatment was determined with a gamma counter.
+ Isolation of caveolin-enriched membrane. Caveolin-enrichedmembrane was isolated by the Triton X-100-based procedure aspreviously described (Anderson et al., 1996). Caveolin-enriched mem-brane was isolated by a detergent-free procedure, essentially as describedby Song et al. (1996). In brief, CV-1 cells were washed in ice-cold PBS andresuspended in 1 ml of lysis buffer containing 500 mM Na
#CO
$(pH 11)
and homogenized by pipetting up and down 14 times through a 1000 µlblue tip, followed by three cycles of sonication (10 s each). The lysate wasthen adjusted to 45% sucrose by adding 1 ml of 90% sucrose in MESbuffer (25 mM MES, pH 6±5, and 150 mM NaCl). The lysate was thentransferred to an ultracentrifuge tube and overlaid sequentially with 2 mlof 35% sucrose and 1 ml of 5% sucrose, each in MES buffer containing250 mM Na
#CO
$. The tube was centrifuged at 39000 r.p.m. for 14 h at
37 °C in a SW50.1 rotor. Caveolin-enriched membrane was isolated as aband at the 5}35% sucrose interface. It was then diluted 1 :3 in buffercontaining 1 :1 500 mM Na
#CO
$(pH 11) and MES buffer (pH 6±5) with
1 mM PMSF and pelleted by centrifugation at 14000 r.p.m. in amicrocentrifuge. The proteins in the pellet were solubilized in a buffer
BEHA
MHC class I molecules and SV40 entryMHC class I molecules and SV40 entry
containing 10 mM Tris, pH 8±0, 150 mM NaCl, 1% Triton X-100,60 mM n-octyl β--glucopyranoside, 1 mM PMSF, 10 µg}ml aprotinin,10 µg}ml soybean trypsin inhibitor, 1 mM sodium vanadate, and10 µg}ml leupeptin. Protein concentrations were determined using theBio-Rad protein assay (Bradford, 1976).
+ Purification of whole plasma membrane. Plasma membranewas isolated using the colloidal silica technique, essentially as describedby Stolz & Jacobson (1991). In brief, CV-1 cells were washed with MES-buffered saline (MBS) containing 20 mM MES, pH 5±5, 135 mM NaCl,1 mM MgCl
#and 0±5 mM CaCl
#with protease and phosphotase
inhibitors. Cells were then coated with 1% colloidal silica (kindlyprovided by B. S. Jacobson) in MBS for 2–5 min. Cells were next washedwith MBS and then overcoated with 1 mg}ml polyacrylic acid (mol. mass50000) in MBS to balance the positive charges on the silica beads. Cellswere washed again in MBS and lysed by osmotic pressure in the lysisbuffer containing 2±5 mM imidazole, 1 mM MgCl
#and 0±5 mM CaCl
#with all inhibitors. Cells were then scraped off the flask and vortexedbriefly. The exposed surface of the cell membrane was pelleted at 900 gfor 5 min in a clinical centrifuge. The membrane pellet was cleaned bywashing gently in the lysis buffer and by pelleting through a 70%metrizamide cushion for 20 min at 20000 r.p.m. in an SW50.1 rotor. Themembrane pellet was then solubilized in a strong detergent and sonicatedthree times (5 s each). The proteins were precipitated in ice-cold acetonefor 20 min (1 part solubilized protein to 4 parts of acetone). Theprecipitated proteins were then solubilized as described for the caveolin-enriched fraction, and concentrations determined using the Bio-Radprotein assay (Bradford, 1976).
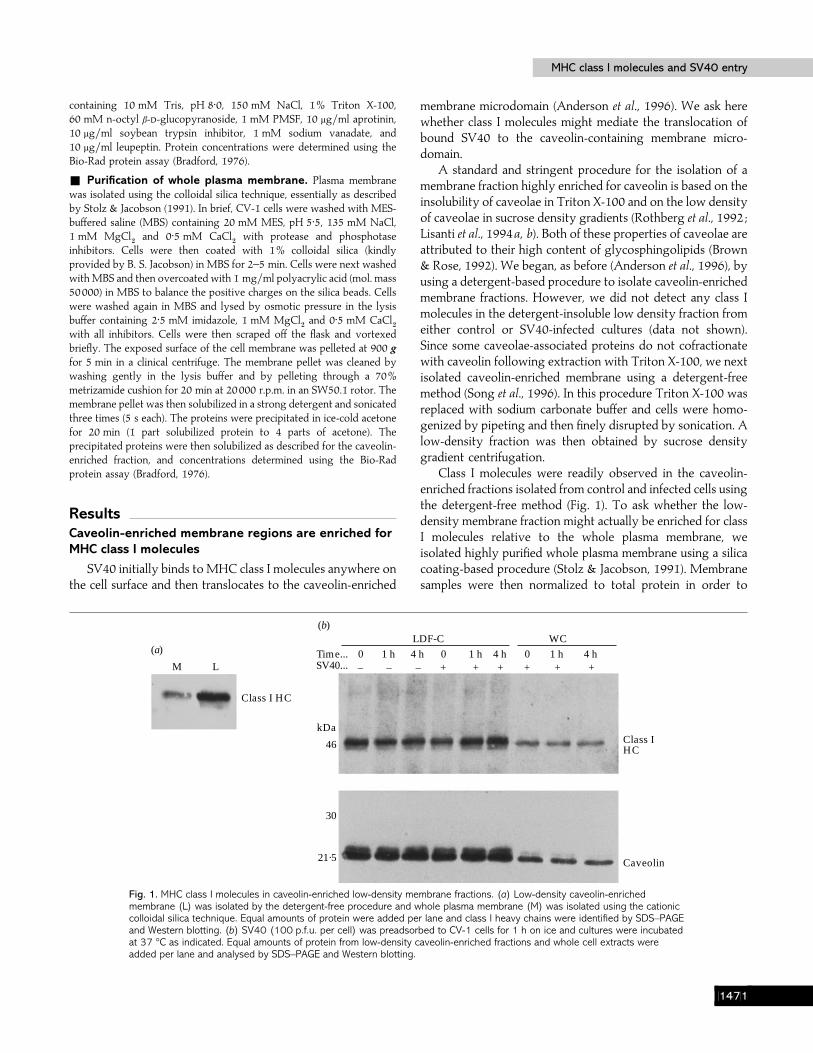
ResultsCaveolin-enriched membrane regions are enriched forMHC class I molecules
SV40 initially binds to MHC class I molecules anywhere onthe cell surface and then translocates to the caveolin-enriched
M L
Class I HC
(a)
+ + + + + +–––SV40...0 1 h 4 h 0 1 h 4 h 0 1 h 4 hTime...
LDF-C WC
46
kDa
21·5
30
Caveolin
Class IHC
(b)
Fig. 1. MHC class I molecules in caveolin-enriched low-density membrane fractions. (a) Low-density caveolin-enrichedmembrane (L) was isolated by the detergent-free procedure and whole plasma membrane (M) was isolated using the cationiccolloidal silica technique. Equal amounts of protein were added per lane and class I heavy chains were identified by SDS–PAGEand Western blotting. (b) SV40 (100 p.f.u. per cell) was preadsorbed to CV-1 cells for 1 h on ice and cultures were incubatedat 37 °C as indicated. Equal amounts of protein from low-density caveolin-enriched fractions and whole cell extracts wereadded per lane and analysed by SDS–PAGE and Western blotting.
membrane microdomain (Anderson et al., 1996). We ask herewhether class I molecules might mediate the translocation ofbound SV40 to the caveolin-containing membrane micro-domain.
A standard and stringent procedure for the isolation of amembrane fraction highly enriched for caveolin is based on theinsolubility of caveolae in Triton X-100 and on the low densityof caveolae in sucrose density gradients (Rothberg et al., 1992 ;Lisanti et al., 1994a, b). Both of these properties of caveolae areattributed to their high content of glycosphingolipids (Brown& Rose, 1992). We began, as before (Anderson et al., 1996), byusing a detergent-based procedure to isolate caveolin-enrichedmembrane fractions. However, we did not detect any class Imolecules in the detergent-insoluble low density fraction fromeither control or SV40-infected cultures (data not shown).Since some caveolae-associated proteins do not cofractionatewith caveolin following extraction with Triton X-100, we nextisolated caveolin-enriched membrane using a detergent-freemethod (Song et al., 1996). In this procedure Triton X-100 wasreplaced with sodium carbonate buffer and cells were homo-genized by pipeting and then finely disrupted by sonication. Alow-density fraction was then obtained by sucrose densitygradient centrifugation.
Class I molecules were readily observed in the caveolin-enriched fractions isolated from control and infected cells usingthe detergent-free method (Fig. 1). To ask whether the low-density membrane fraction might actually be enriched for classI molecules relative to the whole plasma membrane, weisolated highly purified whole plasma membrane using a silicacoating-based procedure (Stolz & Jacobson, 1991). Membranesamples were then normalized to total protein in order to
BEHB
H. A. Anderson, Y. Chen and L. C. NorkinH. A. Anderson, Y. Chen and L. C. Norkin
(a) (b)
kDa97
47
30
22
17
kDa97
47
30
22
17
0·0 0·25
0·75
1·5 2·0 3·0 5·0 9·0
Chase time (h)
0·0 0·75
1·5 2·0 3·00·0 0·0 0·0 0·75
1·5 2·0 3·0
SV40
PapainPapain
Chasetime (h)...
Heavychain
â2m
0·0 0·75
1·5 2·0 3·0 5·0 9·0Chasetime (h)...
(c)
kDa
97
47
30
22
Heavychain
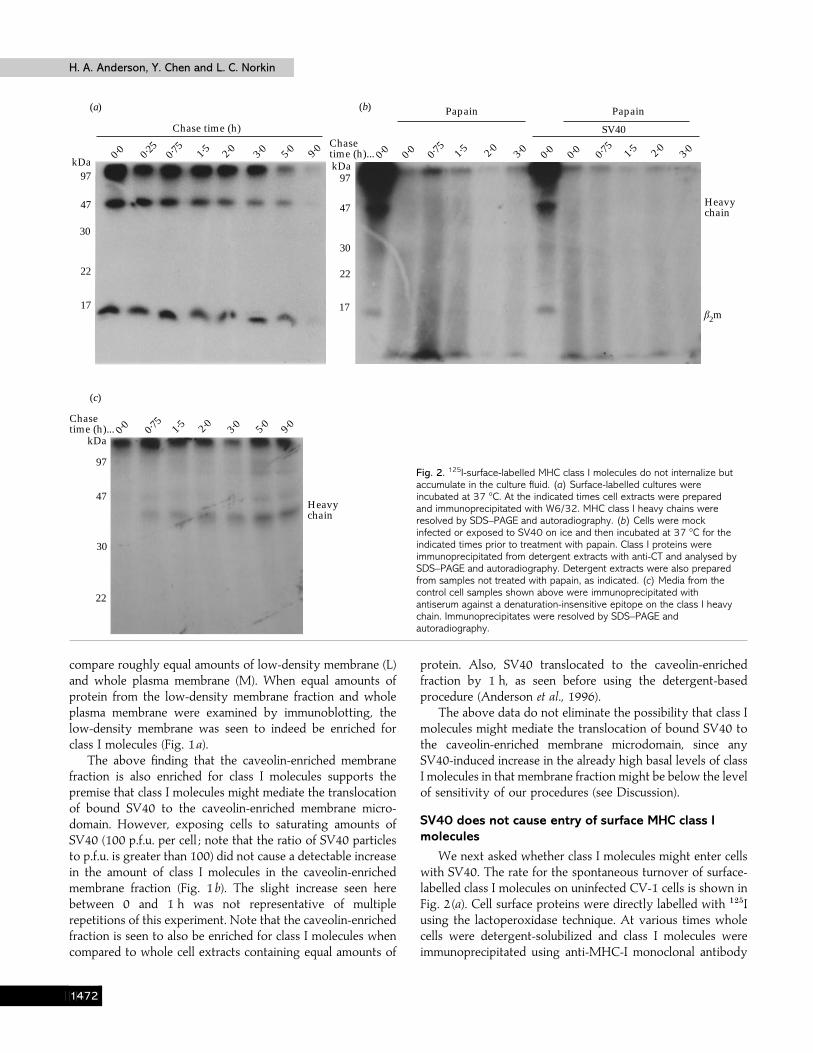
Fig. 2. 125I-surface-labelled MHC class I molecules do not internalize butaccumulate in the culture fluid. (a) Surface-labelled cultures wereincubated at 37 °C. At the indicated times cell extracts were preparedand immunoprecipitated with W6/32. MHC class I heavy chains wereresolved by SDS–PAGE and autoradiography. (b) Cells were mockinfected or exposed to SV40 on ice and then incubated at 37 °C for theindicated times prior to treatment with papain. Class I proteins wereimmunoprecipitated from detergent extracts with anti-CT and analysed bySDS–PAGE and autoradiography. Detergent extracts were also preparedfrom samples not treated with papain, as indicated. (c) Media from thecontrol cell samples shown above were immunoprecipitated withantiserum against a denaturation-insensitive epitope on the class I heavychain. Immunoprecipitates were resolved by SDS–PAGE andautoradiography.
compare roughly equal amounts of low-density membrane (L)and whole plasma membrane (M). When equal amounts ofprotein from the low-density membrane fraction and wholeplasma membrane were examined by immunoblotting, thelow-density membrane was seen to indeed be enriched forclass I molecules (Fig. 1a).
The above finding that the caveolin-enriched membranefraction is also enriched for class I molecules supports thepremise that class I molecules might mediate the translocationof bound SV40 to the caveolin-enriched membrane micro-domain. However, exposing cells to saturating amounts ofSV40 (100 p.f.u. per cell ; note that the ratio of SV40 particlesto p.f.u. is greater than 100) did not cause a detectable increasein the amount of class I molecules in the caveolin-enrichedmembrane fraction (Fig. 1b). The slight increase seen herebetween 0 and 1 h was not representative of multiplerepetitions of this experiment. Note that the caveolin-enrichedfraction is seen to also be enriched for class I molecules whencompared to whole cell extracts containing equal amounts of
protein. Also, SV40 translocated to the caveolin-enrichedfraction by 1 h, as seen before using the detergent-basedprocedure (Anderson et al., 1996).
The above data do not eliminate the possibility that class Imolecules might mediate the translocation of bound SV40 tothe caveolin-enriched membrane microdomain, since anySV40-induced increase in the already high basal levels of classI molecules in that membrane fraction might be below the levelof sensitivity of our procedures (see Discussion).
SV40 does not cause entry of surface MHC class Imolecules
We next asked whether class I molecules might enter cellswith SV40. The rate for the spontaneous turnover of surface-labelled class I molecules on uninfected CV-1 cells is shown inFig. 2 (a). Cell surface proteins were directly labelled with "#&Iusing the lactoperoxidase technique. At various times wholecells were detergent-solubilized and class I molecules wereimmunoprecipitated using anti-MHC-I monoclonal antibody
BEHC
MHC class I molecules and SV40 entryMHC class I molecules and SV40 entry
(a)
(b)CellPapain resistantMedia
150
100
50
00 0·75 1·5 3
Time at 37 °C (h)
CellPapain resistant
80
60
40
20
00 5 10
Time at 37 °C (min)
10–2
× c
.p.m
.
10–2
× c
.p.m
.
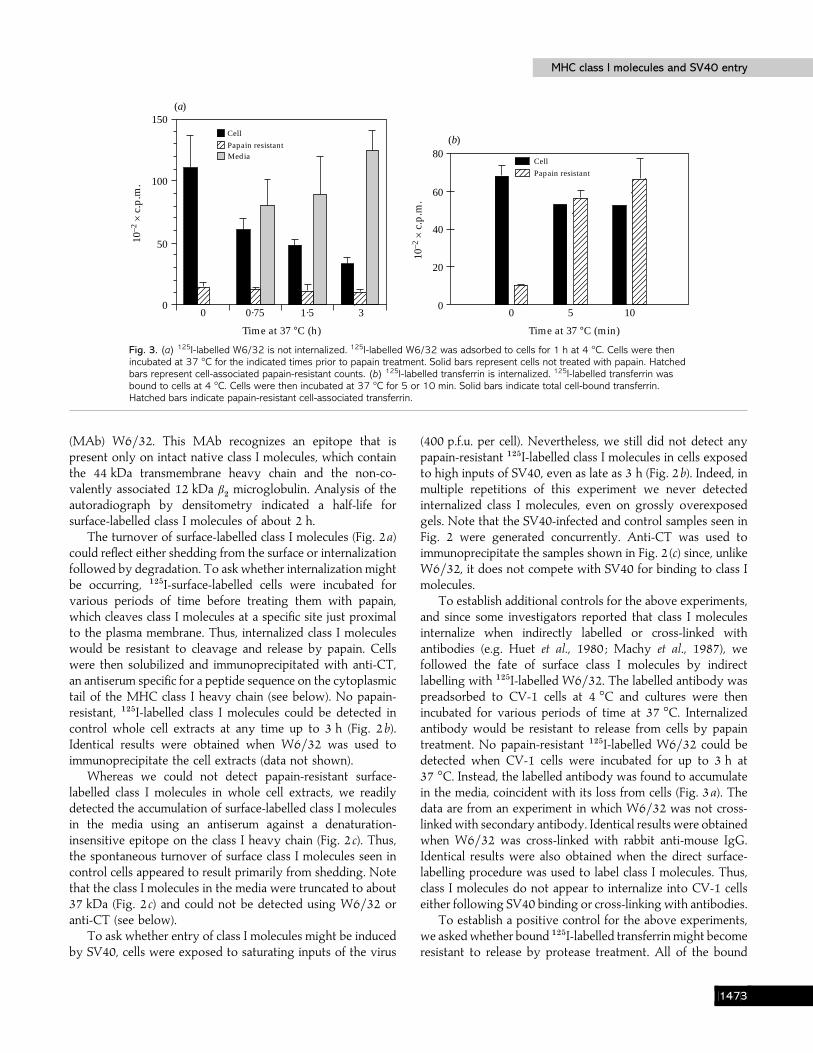
Fig. 3. (a) 125I-labelled W6/32 is not internalized. 125I-labelled W6/32 was adsorbed to cells for 1 h at 4 °C. Cells were thenincubated at 37 °C for the indicated times prior to papain treatment. Solid bars represent cells not treated with papain. Hatchedbars represent cell-associated papain-resistant counts. (b) 125I-labelled transferrin is internalized. 125I-labelled transferrin wasbound to cells at 4 °C. Cells were then incubated at 37 °C for 5 or 10 min. Solid bars indicate total cell-bound transferrin.Hatched bars indicate papain-resistant cell-associated transferrin.
(MAb) W6}32. This MAb recognizes an epitope that ispresent only on intact native class I molecules, which containthe 44 kDa transmembrane heavy chain and the non-co-valently associated 12 kDa β
#microglobulin. Analysis of the
autoradiograph by densitometry indicated a half-life forsurface-labelled class I molecules of about 2 h.
The turnover of surface-labelled class I molecules (Fig. 2a)could reflect either shedding from the surface or internalizationfollowed by degradation. To ask whether internalization mightbe occurring, "#&I-surface-labelled cells were incubated forvarious periods of time before treating them with papain,which cleaves class I molecules at a specific site just proximalto the plasma membrane. Thus, internalized class I moleculeswould be resistant to cleavage and release by papain. Cellswere then solubilized and immunoprecipitated with anti-CT,an antiserum specific for a peptide sequence on the cytoplasmictail of the MHC class I heavy chain (see below). No papain-resistant, "#&I-labelled class I molecules could be detected incontrol whole cell extracts at any time up to 3 h (Fig. 2b).Identical results were obtained when W6}32 was used toimmunoprecipitate the cell extracts (data not shown).
Whereas we could not detect papain-resistant surface-labelled class I molecules in whole cell extracts, we readilydetected the accumulation of surface-labelled class I moleculesin the media using an antiserum against a denaturation-insensitive epitope on the class I heavy chain (Fig. 2 c). Thus,the spontaneous turnover of surface class I molecules seen incontrol cells appeared to result primarily from shedding. Notethat the class I molecules in the media were truncated to about37 kDa (Fig. 2 c) and could not be detected using W6}32 oranti-CT (see below).
To ask whether entry of class I molecules might be inducedby SV40, cells were exposed to saturating inputs of the virus
(400 p.f.u. per cell). Nevertheless, we still did not detect anypapain-resistant "#&I-labelled class I molecules in cells exposedto high inputs of SV40, even as late as 3 h (Fig. 2b). Indeed, inmultiple repetitions of this experiment we never detectedinternalized class I molecules, even on grossly overexposedgels. Note that the SV40-infected and control samples seen inFig. 2 were generated concurrently. Anti-CT was used toimmunoprecipitate the samples shown in Fig. 2 (c) since, unlikeW6}32, it does not compete with SV40 for binding to class Imolecules.
To establish additional controls for the above experiments,and since some investigators reported that class I moleculesinternalize when indirectly labelled or cross-linked withantibodies (e.g. Huet et al., 1980 ; Machy et al., 1987), wefollowed the fate of surface class I molecules by indirectlabelling with "#&I-labelled W6}32. The labelled antibody waspreadsorbed to CV-1 cells at 4 °C and cultures were thenincubated for various periods of time at 37 °C. Internalizedantibody would be resistant to release from cells by papaintreatment. No papain-resistant "#&I-labelled W6}32 could bedetected when CV-1 cells were incubated for up to 3 h at37 °C. Instead, the labelled antibody was found to accumulatein the media, coincident with its loss from cells (Fig. 3a). Thedata are from an experiment in which W6}32 was not cross-linked with secondary antibody. Identical results were obtainedwhen W6}32 was cross-linked with rabbit anti-mouse IgG.Identical results were also obtained when the direct surface-labelling procedure was used to label class I molecules. Thus,class I molecules do not appear to internalize into CV-1 cellseither following SV40 binding or cross-linking with antibodies.
To establish a positive control for the above experiments,we askedwhether bound "#&I-labelled transferrin might becomeresistant to release by protease treatment. All of the bound
BEHD
H. A. Anderson, Y. Chen and L. C. NorkinH. A. Anderson, Y. Chen and L. C. Norkin
––0
––2
–+2
+–2
++2
+–0
+–2
++2Time (h)...
Phen...FBS...
Anti-cytoplasmic tail W6/32
(a)
––
––
–+
+–
++
+–
+–
++Phen...
FBS...
Anti-cytoplasmic tail W6/32(b)
100
80
60
40
20
0
% o
f MH
C c
lass
I pr
otei
n he
avy
chai
nim
mun
opre
cipi
tate
d b
y an
tibo
die
s
0 2 2 2 2 0 2 2
Time at 37 °C (h)
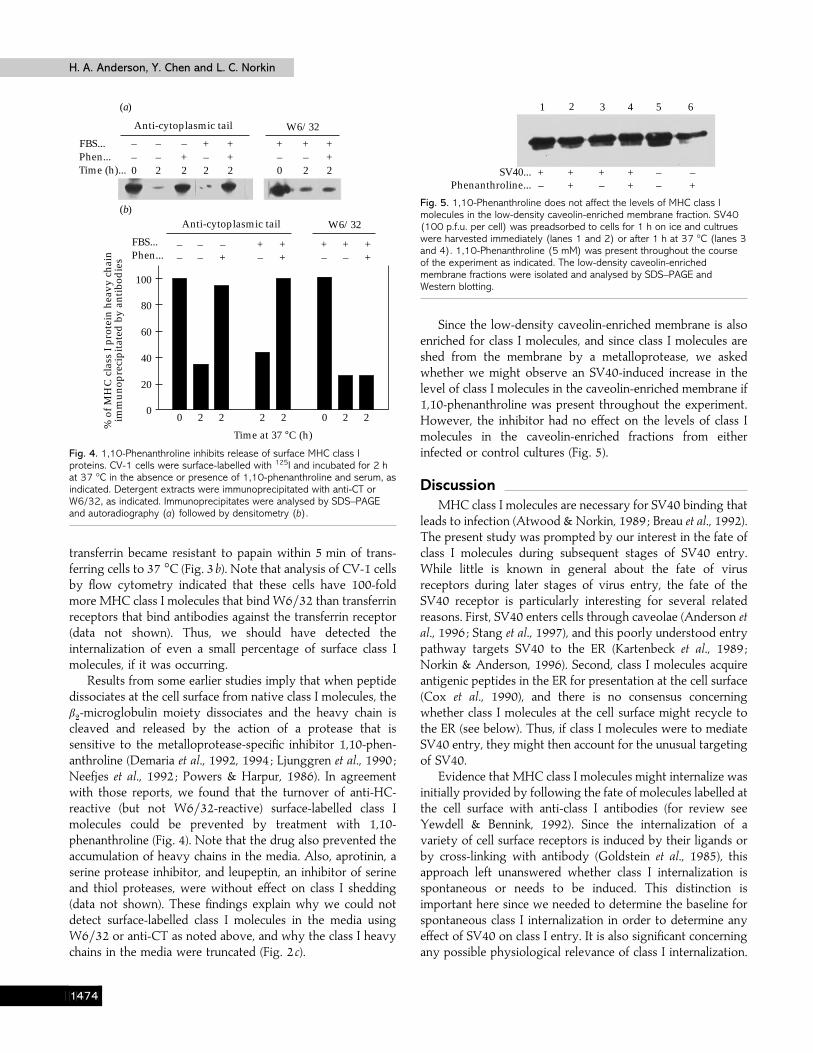
Fig. 4. 1,10-Phenanthroline inhibits release of surface MHC class Iproteins. CV-1 cells were surface-labelled with 125I and incubated for 2 hat 37 °C in the absence or presence of 1,10-phenanthroline and serum, asindicated. Detergent extracts were immunoprecipitated with anti-CT orW6/32, as indicated. Immunoprecipitates were analysed by SDS–PAGEand autoradiography (a) followed by densitometry (b).
transferrin became resistant to papain within 5 min of trans-ferring cells to 37 °C (Fig. 3b). Note that analysis of CV-1 cellsby flow cytometry indicated that these cells have 100-foldmore MHC class I molecules that bind W6}32 than transferrinreceptors that bind antibodies against the transferrin receptor(data not shown). Thus, we should have detected theinternalization of even a small percentage of surface class Imolecules, if it was occurring.
Results from some earlier studies imply that when peptidedissociates at the cell surface from native class I molecules, theβ#-microglobulin moiety dissociates and the heavy chain is
cleaved and released by the action of a protease that issensitive to the metalloprotease-specific inhibitor 1,10-phen-anthroline (Demaria et al., 1992, 1994 ; Ljunggren et al., 1990 ;Neefjes et al., 1992 ; Powers & Harpur, 1986). In agreementwith those reports, we found that the turnover of anti-HC-reactive (but not W6}32-reactive) surface-labelled class Imolecules could be prevented by treatment with 1,10-phenanthroline (Fig. 4). Note that the drug also prevented theaccumulation of heavy chains in the media. Also, aprotinin, aserine protease inhibitor, and leupeptin, an inhibitor of serineand thiol proteases, were without effect on class I shedding(data not shown). These findings explain why we could notdetect surface-labelled class I molecules in the media usingW6}32 or anti-CT as noted above, and why the class I heavychains in the media were truncated (Fig. 2 c).
1 2 3 4 5 6
+–
++
+–
++
––
–+Phenanthroline...
SV40...
Fig. 5. 1,10-Phenanthroline does not affect the levels of MHC class Imolecules in the low-density caveolin-enriched membrane fraction. SV40(100 p.f.u. per cell) was preadsorbed to cells for 1 h on ice and cultrueswere harvested immediately (lanes 1 and 2) or after 1 h at 37 °C (lanes 3and 4). 1,10-Phenanthroline (5 mM) was present throughout the courseof the experiment as indicated. The low-density caveolin-enrichedmembrane fractions were isolated and analysed by SDS–PAGE andWestern blotting.
Since the low-density caveolin-enriched membrane is alsoenriched for class I molecules, and since class I molecules areshed from the membrane by a metalloprotease, we askedwhether we might observe an SV40-induced increase in thelevel of class I molecules in the caveolin-enriched membrane if1,10-phenanthroline was present throughout the experiment.However, the inhibitor had no effect on the levels of class Imolecules in the caveolin-enriched fractions from eitherinfected or control cultures (Fig. 5).
DiscussionMHC class I molecules are necessary for SV40 binding that
leads to infection (Atwood & Norkin, 1989 ; Breau et al., 1992).The present study was prompted by our interest in the fate ofclass I molecules during subsequent stages of SV40 entry.While little is known in general about the fate of virusreceptors during later stages of virus entry, the fate of theSV40 receptor is particularly interesting for several relatedreasons. First, SV40 enters cells through caveolae (Anderson etal., 1996 ; Stang et al., 1997), and this poorly understood entrypathway targets SV40 to the ER (Kartenbeck et al., 1989 ;Norkin & Anderson, 1996). Second, class I molecules acquireantigenic peptides in the ER for presentation at the cell surface(Cox et al., 1990), and there is no consensus concerningwhether class I molecules at the cell surface might recycle tothe ER (see below). Thus, if class I molecules were to mediateSV40 entry, they might then account for the unusual targetingof SV40.
Evidence that MHC class I molecules might internalize wasinitially provided by following the fate of molecules labelled atthe cell surface with anti-class I antibodies (for review seeYewdell & Bennink, 1992). Since the internalization of avariety of cell surface receptors is induced by their ligands orby cross-linking with antibody (Goldstein et al., 1985), thisapproach left unanswered whether class I internalization isspontaneous or needs to be induced. This distinction isimportant here since we needed to determine the baseline forspontaneous class I internalization in order to determine anyeffect of SV40 on class I entry. It is also significant concerningany possible physiological relevance of class I internalization.
BEHE

MHC class I molecules and SV40 entryMHC class I molecules and SV40 entry
In subsequent studies, approaches to measuring class Iinternalization were developed that were independent ofcross-linking with antibodies. For example, it was possible tofollow the fate of class I molecules that were radiolabelled atthe cell surface (Reid & Watts, 1990 ; Vega & Strominger,1989). Internalization of class I molecules has also beenfollowed by measuring the uptake of fluorescent β
#micro-
globulin (Hochman et al., 1991). Results from studies in whichseveral variations of these approaches were used have notprovided a consensus regarding whether class I molecules areinternalized constitutively, inducibly, or even at all (AbdelMotal et al., 1993 ; Aragnol et al., 1986 ; Capps et al., 1989 ;Eicholtz et al., 1992 ; Hochman et al., 1991 ; Huet et al., 1980 ;Machy et al., 1987 ; Neefjes et al., 1992 ; Reid & Watts, 1990 ;Tse & Pernis, 1984 ; Vega & Strominger, 1989).
Since these studies involved a variety of cell types, cell-type-specific effects might be responsible for some of thedissimilar findings. Differences in experimental methodologymight account for others. For example, it remains to be shownthat traffic of labelled antibody reagents reflects traffic of classI molecules. Indeed, most of the studies reporting theinternalization of class I molecules used reagents (e.g. anti-bodies) labelled with isotopes or fluorescent dyes to track classI molecules. It was assumed that these reagents remainedconjugated to the class I molecules and reflected theirmovements. In contrast, the few reports which did not findevidence for internalization (Eicholtz et al., 1992 ; Neefjes et al.,1990, 1992) used a more direct surface-labelling biochemicalapproach. Nevertheless, other studies in which a direct surface-labelling approach was used did report evidence for internal-ization of class I molecules (Reid & Watts, 1990 ; Vega &Strominger, 1989).
Since different experimental methodologies might lead todifferent conclusions, we followed the fate of surface class Imolecules on CV-1 cells using labelled antibodies as well as bydirect surface-labelling. Thus, it is noteworthy that we did notsee any evidence that surface class I molecules might internalizeusing either of these standard and sensitive procedures.
Our direct surface-labelling procedure depended on iso-lating "#&I-surface-labelled class I molecules from cell extractsby immunoprecipitation. Thus, some internalized class Imolecules might have gone undetected if they were degradedsoon after entry. However, this seems unlikely in the case ofclass I molecules that might have entered in association withSV40, since entering SV40 is largely targeted to the ER, ratherthan to the endosomal}lysosomal compartment (Kartenbeck etal., 1989 ; Norkin & Anderson, 1996). Furthermore, degra-dation is not an issue in experiments where surface class Imolecules were indirectly labelled with "#&I-labelled W6}32.
To establish a positive control for the above experiments,we followed the internalization of "#&I-labelled transferrin. Incontrast to our findings using labelled W6}32, preadsorbedtransferrin completely internalized within 5 min of pre-adsorption. Furthermore, analysis by flow cytometry showed
that CV-1 cells have 100-fold more class I molecules that bindW6}32 than transferrin receptors that bind antibodies againstthe transferrin receptor. Together, these results suggest thatwe should have detected internalized surface-labelled class Imolecules, even if only a small percentage of the surface-labelled molecules had internalized.
Since we found no evidence for the internalization of classI molecules coincident with SV40 entry, despite usingsaturating amounts of SV40 and the highly sensitive directsurface-labelling procedure to detect internalized class Imolecules, we conclude that class I molecules do not deliverSV40 into the cell, nor can they account for the targeting ofSV40 to the ER. Instead, our results imply the existence of aputative coreceptor for SV40 that might mediate its entry. Inthis regard, other viruses have also been found to use distinctcell surface factors to mediate the series of steps between initialvirus binding and entry. Examples include nonenvelopedviruses such as adenoviruses (Wickham et al., 1993), andenveloped viruses such as herpesviruses (McClain & Fuller,1994) and human immunodeficiency virus (Deng et al., 1996 ;Dragic et al., 1996 ; reviewed Norkin, 1995).
It is interesting to consider where in the SV40 entrypathway the virus might dissociate from class I molecules andassociate with the putative coreceptor. In this regard, SV40initially binds to class I molecules that are on flat regions of theplasma membrane. The virus later becomes associated withcaveolae. This was indicated by both cell fractionation(Anderson et al., 1996) and ultrastructure (Stang et al., 1997).The virus then enters in tight-fitting vesicles (Kartenbeck et al.,1989) that may derive from caveolae.
The present findings do not eliminate the possibility thatclass I molecules might mediate the translocation of boundSV40 to caveolae. Consistent with this possibility, we foundthat the caveolin-enriched membrane fraction is also enrichedfor class I molecules relative to whole plasma membrane. Tothe best of our knowledge this finding is presently unique.Furthermore, Stang et al. (1997) recently reported that cross-linking class I molecules with W6}32 causes them to cluster incaveolae, as shown by immunoelectron microscopy. However,we could not detect any SV40- or W6}32-induced increase inthe level of class I molecules in the caveolin-enrichedmembranefraction. A possible explanation is that any SV40- or antibody-induced increase in the level of class I molecules in thecaveolin-enriched fraction might have been below the level ofdetection by our procedure, since that fraction is already highlyenriched for class I molecules.
We recently reported that SV40 entry is promoted by anintracellular signal that SV40 induces from the cell surface(Dangoria et al., 1996). More recently, we found that signaltransmission by SV40 requires the functional integrity of thelow-density membrane microdomain that surrounds thecaveolae (Y. Chen & L. C. Norkin, unpublished results).Furthermore, electron microscopy of detergent-insoluble low-density complexes showed that SV40 particles accumulated at
BEHF

H. A. Anderson, Y. Chen and L. C. NorkinH. A. Anderson, Y. Chen and L. C. Norkin
the ‘mouths ’ of the caveolae when signalling was blocked,whereas the virus was found within the caveolae whensignalling was unobstructed (Y. Chen & L. C. Norkin, un-published results). Note that the annular regions about thecaveolae, as well as the caveolae per se, are enriched for avariety of signal transducing molecules (Schnitzer et al., 1995b).
It may be of interest that whereas W6}32 did not induceinternalization of class I molecules, this MAb (as well as otheranti-class I MAbs) did induce a transmembrane signal in CV-1cells. However, the antibody-induced signal led to theactivation of mitogen-activated protein kinase (MAPK),whereas the SV40-induced signal did not (Dangoria et al.,1996). This finding suggests that the SV40 signal might betransmitted through the putative coreceptor, rather thanthrough class I molecules.
The following model is consistent with the above findings.First, SV40 binds to class I molecules anywhere on the cellsurface. Then, in association with class I molecules SV40translocates to the annuli surrounding the caveolae. Withinthat microdomain SV40 dissociates from class I molecules andassociates with the putative coreceptor. The SV40-inducedsignal is then transmitted, promoting virus entry into thecaveolae, followed by later stages of entry into the cell.
Although we detected no internalization of either directlyor indirectly labelled class I molecules into CV-1 cells, wereadily detected the accumulation of truncated directly labelledclass I heavy chains and "#&I-labelled W6}32 in the media. Ourfindings are entirely consistent with earlier reports (Demaria etal., 1992, 1994 ; Ljunggren et al., 1990 ; Neefjes et al., 1992),which implied that surface class I molecules are cleaved by asurface metalloprotease activity after the spontaneous releaseof peptide and the β
#microglobulin moiety. This instability of
empty class I heterodimers and their release from the cellsurface would prevent the rebinding of peptide that mightsensitize bystander cells to lysis by cytotoxic T cells (e.g.Demaria et al., 1994 ; Neefjes et al., 1992 ; Ortiz-Navarret &Hammerling, 1991 ; Rock et al., 1990).
We are grateful to Hidde L. Ploegh for antisera, to Bruce S. Jacobsonfor advice on membrane purification, and to Maryanne Wells for expertpreparation of the manuscript. This work was supported by NationalInstitutes of Health Grant CA50532, National Science Foundation GrantMCB-9219207, a Faculty Research grant from the University ofMassachusetts and a HATCH award from the USDA.
ReferencesAbdel Motal, U. M., Zhou, X., Joki, A., Siddiqi, A. R., Srinivasa, B. R.,Stenvall, K., Dahmen, J. & Jondal, M. (1993). Major histocompatibilitycomplex class I-binding peptides are recycled to the cell surface afterinternalization. European Journal of Immunology 23, 3224–3229.
Anderson, H. A., Chen, Y. & Norkin, L. C. (1996). Bound simian virus 40translocates to caveolin-enriched membrane domains, and its entry isinhibited by drugs that selectively disrupt caveolae. Molecular Biology ofthe Cell 7, 1825–1834.
Anderson, R. G. W. (1993a). Caveolae : where incoming and outgoingmessengers meet. Proceedings of the National Academy of Sciences, USA 90,10909–10913.
Anderson, R. G. W. (1993b). Potocytosis of small molecules and ions bycaveolae. Trends in Cell Biology 3, 69–72.
Aragnol, D., Malissen, B., Schiff, C., Piron, M. A. & Leserman, L. D.(1986). Endocytosis of class I molecules studied using B cell-B lymphomaand B cell-T lymphoma hybrids. Journal of Immunology 137, 3347–3353.
Atwood, W. J. & Norkin, L. C. (1989). Class I major histocompatibilityproteins as cell surface receptors for simian virus 40. Journal of Virology63, 4474–4477.
Bradford, M. M. (1976). A rapid and sensitive method for thequantitation of microgram quantities of protein utilizing the principle ofprotein-dye binding. Analytical Biochemistry 72, 248–254.
Breau, W. C., Atwood, W. J. & Norkin, L. C. (1992). Class I majorhistocompatibility proteins are an essential component of the simian virus40 receptor. Journal of Virology 66, 2037–2045.
Brown, D. A. & Rose, J. K. (1992). Sorting of GPI-anchored proteins toglycolipid-enriched membrane subdomains during transport to the apicalcell surface. Cell 68, 533–544.
Capps, G. G., VanKampen, M., Ward, C. L. & Zuniga, M. C. (1989).Endocytosis of the class I major histocompatibility antigen via a phorbolmyristate acetate-inducible pathway is a cell-specific phenomenon andrequires the cytoplasmic domain. Journal of Cell Biology 108, 1317–1329.
Cox, J. H., Yewdell, J. W., Eisenlohr, L. C., Johnson, P. R. & Bennink, J.R. (1990). Antigen presentation requires transport of MHC class Imolecules from the endoplasmic reticulum. Science 247, 715–718.
Dangoria, N. S., Breau, W. C., Anderson, H. A., Cishek, D. M. & Norkin,L. C. (1996). Extracellular simian virus 40 induces an ERK}MAPK-independent signaling pathway that activates primary response genesand promotes virus entry. Journal of General Virology 77, 2173–2182.
Dasgupta, J. S., Watkins, S., Slayter, H. & Yunis, E. J. (1988). Receptorlike nature of class I HLA: endocytosis via coated pits. Journal ofImmunology 141, 2577–2580.
Demaria, S., Schwab, R. & Bushkin, Y. (1992). The origin and fate ofβ#m-free MHC class I molecules induced on activated T cells. Cellular
Immunology 142, 103–113.
Demaria, S., Schwab, R., Gottesman, S. R. S. & Bushkin, Y. (1994).Soluble β
#-microglobulin-free class I heavy chains are released from the
surface of activated and leukemia cells by a metalloprotease. Journal ofBiological Chemistry 269, 6689–6694.
Deng, H., Liu, R., Ellmeier, W., Choe, S., Unutmaz, D., Buckhart, M., DiMarzio, P., Marmon, S., Sutton, R. E., Hill, C. M., Davis, C. B., Peiper,S. C., Schall, T. J., Littman, D. R. & Landau, N. R. (1996). Identificationof a major co-receptor for primary isolates of HIV-1. Nature 281,661–666.
Dragic, T., Litwin, V., Allaway, G. P., Martin, S. R., Huang, Y.,Nagashima, K. A., Cayanan, C., Maddon, P. J., Koup, R. A., Moore, J. P.& Paxton, W. A. (1996). HIV entry into CD4+ cells is mediated by thechemokine receptor CC-CKR-5. Nature 281, 667–673.
Eichholtz, T., Vossebeld, P., van Overveld, M. & Ploegh, H. (1992).Activation of protein kinase C accelerates internalization of transferrinreceptor but not of major histocompatibility complex class I, independentof their phosphorylation status. Journal of Biological Chemistry 267,22490–22495.
Goldstein, J. L., Brown, M. S., Anderson, R. G. W., Russell, D. W. W. &Schneider, W. J. (1985). Receptor mediated endocytosis : conceptsemerging from the LDL receptor system. Annual Review of Cell Biology 1,1–39.
BEHG

MHC class I molecules and SV40 entryMHC class I molecules and SV40 entry
Harlow, E. & Lane, D. (1988). Antibodies : A Laboratory Manual. ColdSpring Harbor, NY: Cold Spring Harbor Laboratory.
Hochman, J., Jiang, H, Matyus, L., Edidin, M. & Pernis, B. (1991).Endocytosis and dissociation of class I MHC molecules with fluorescentβ-2 microglobulin. Journal of Immunology 146, 1862–1867.
Huet, C., Ash, J. F. & Singer, S. J. (1980). The antibody-inducedclustering and endocytosis of HLA antigens on cultured humanfibroblasts. Cell 21, 429–438.
Kartenbeck, J., Stukenbrok, H. & Helenius, A. (1989). Endocytosis ofsimian virus 40 into the endoplasmic reticulum. Journal of Cell Biology109, 2721–2729.
Lisanti, M. P., Scherer, P. E., Tang, Z. & Sargiacomo, M. (1994a).Caveolae, caveolin, and caveolin-rich membrane domains : a signalinghypothesis. Trends in Cell Biology 4, 231–235.
Lisanti, M. P., Scherer, P. E., Vidugiriene, J., Tang, Z. L., Hermanowski-Vosatka, A., Tu, Y. H., Cook, R. F. & Sargiacomo, M. (1994b).Characterization of caveolin-rich membrane domains isolated from anendothelial-rich source : implications for human disease. Journal of CellBiology 126, 111–126.
Ljunggren, H. G., Stam, J. N., O> hlen, C., Neefjes, J. J., Ha$ glund, P.,Heemels, M. T., Bastin, J., Schumacher, T. N. M., Townsend, A., Karre,K. & Ploegh, H. L. (1990). Empty class I molecules come out in the cold.Nature 346, 476–480.
McClain, D. & Fuller, A. O. (1994). Cell-specific kinetics and efficiency ofherpes simplex type 1 entry are determined by two distinct phases ofattachment. Virology 19, 690–702.
Machy, P., Truneh, A., Gennaro, D. & Hoffstein, S. (1987). Majorhistocompatibility complex class I molecules internalized via coated pitsin T lymphocytes. Nature 328, 724–726.
Marsh, M. & Helenius, A. (1989). Virus entry into animal cells. Advancesin Virus Research 36, 107–151.
Montesano, R., Roth, J., Robert, A. & Orci, L. (1982). Non-coatedmembrane invaginations are involved in binding and internalization oftetanus toxins. Nature 296, 651–653.
Neefjes, J. J., Breur-Vriesendorp, B. S., Van Seventer, G. A., Ivanyi, P.& Ploegh, H. L. (1986). An improved biochemical method for theanalysis of HLA class I antigens. Definition of new HLA-class I subtypes.Human Immunology 16, 169–181.
Neefjes, J. J., Stollarz, V., Peters, P. J., Geuze, H. J. & Ploegh, H. L.(1990). The biosynthetic pathway of MHC class II but not MHC classI intersects the endocytic route. Cell 61, 171–183.
Neefjes, J. J., Smit, L., Gehrmann, M. & Ploegh, H. L. (1992). The fateof the three subunits of major histocompatibility complex class Imolecules. European Journal of Immunology 22, 1609–1614.
Norkin, L. C. (1995). Virus receptors : implications for pathogenesis andthe design of antiviral agents. Clinical Microbiology Reviews 8, 293–315.
Norkin, L. C. & Anderson, H. A. (1996). Multiple stages of virus–receptor interactions as shown by simian virus 40. Advances inExperimental Medicine and Biology 408, 159–167.
Ortiz-Navarret, V. & Ha$ mmerling, G. J. (1991). Surface appearance andinstability of empty H-2 class I molecules under physiologic conditions.Proceeding of the National Academy of Sciences, USA 88, 3594–3597.
Parham, P., Bainstable, C. & Bodner, W. F. (1979). Use of monoclonalantibody (W6}32) in structural studies of HLA-A, B, C antigens. Journalof Immunology 123, 342–349.
Philips, D. R. & Morrison, M. (1970). The arrangement of proteins in thehuman erythrocyte membrane. Biochemical and Biophysical ResearchCommunications 40, 284–289.
Powers, J. & Harper, J. (1986). Inhibitors of metalloproteases. InProteinase Inhibitors, pp. 219–300. Edited by A. Barrett & G. Salveson.Amsterdam: Elsevier.
Reid, P. A. & Watts, C. (1990). Cycling of cell-surface MHC glyco-proteins through primaquine sensitive intracellular compartments. Nature346, 655–657.
Rock, K. L., Rothstein, L. E., Gamble, S. R. & Benacerraf, B. (1990).Reassociation with β
#-microglobulin is necessary for Kb class I major
histocompatibility complex binding of exogenous peptides. Proceedings ofthe National Academy of Sciences, USA 87, 7517–7521.
Rothberg, K. G., Heuser, J. E., Donzell, W. C., Ying, Y.-S., Glenney, J.R. & Anderson, R. G. W. (1992). Caveolin, a protein component ofcaveolae membrane coats. Cell 68, 673–682.
Rougon, G., Ceard, B., Van Rietscholen, J., Jordan, B. & Barbet, J.(1984). Induction with a synthetic peptide of antibodies to HLA class IC-terminal intracytoplasmic region. Molecular Immunology 21, 461–468.
Schnitzer, J. E., Liu, J. & Oh, P. (1995a). Endothelial caveolae have themolecular transport machinery for vesicle budding, docking, and fusionincluding VAMP, NSF, SNAP, annexins, and GTPases. Journal of BiologicalChemistry 270, 14399–14404.
Schnitzer, J. E., McIntosh, D. P., Dvorak, A. M., Liu, J. & Oh, P.(1995b). Separation of caveolae from associated microdomains of GPI-anchored proteins. Science 269, 1435–1439.
Schnitzer, J. E., Oh, P. & McIntosh, D. P. (1996). Role of GTP hydrolsisin fission of caveolae directly from plasma membranes. Science 274,239–242.
Simionescu, M., Simionescu, N. & Palade, G. E. (1982). Differentiatedmicrodomains on the luminal surface of capillary endothelium: dis-tribution of lectin receptors. Journal of Cell Biology 94, 406–413.
Song, K. S., Li, S., Okamoto, T., Quilliam, L. A., Sargiacomo, M. &Lisanti, M. P. (1996). Co-purification and direct interaction of ras withcaveolin, an integral membrane protein of caveolae microdomains.Detergent-free purification of caveolae membranes. Journal of BiologicalChemistry 271, 9690–9697.
Stang, E., Kartenbeck, J. & Parton, R. G. (1997). Major histo-compatibility complex class I molecules mediate association of SV40 withcaveolae. Molecular Biology of the Cell 8, 47–57.
Stolz, D. B. & Jacobson, B. S. (1991). Macro- and microvascularendothelial cells in vitro ; maintenance of biochemical heterogeneitydespite loss of ultrastructural characteristics. In Vitro Cell and De-velopmental Biology 27A, 169–181.
Tse, D. B. & Pernis, B. (1984). Spontaneous internalization of class Imajor histocompatibility complex molecules in T lymphoid cells. Journalof Experimental Medicine 159, 193–207.
Vega, M. A. & Strominger, J. L. (1989). Constitutive endocytosis ofHLA class I antigens requires a specific portion of the intracytoplasmictail that shares structural features with other endocytosed molecules.Proceedings of the National Academy of Sciences, USA 86, 2688–2692.
Wickham, T., Mathins, P., Cheresh, P. & Nimerow, G. (1993). IntegrinsAvB
$and AvB
&promote adenovirus internalization but not virus
attachment. Cell 73, 309–314.
Yewdell, J. W. & Bennink, J. R. (1992). Cell biology of antigenprocessing and presentation to MHC class I restricted T lymphocytes.Advances in Immunology 52, 1–124.
Received 12 January 1998; Accepted 27 January 1998
BEHH

H. A. Anderson, Y. Chen and L. C. NorkinH. A. Anderson, Y. Chen and L. C. Norkin
BEHI
Related Documents



![MANUAL DE USUARIO MÁQUINAS DE HIELO...MANUAL DE USUARIO [AUTOCONTENIDAS Y REMOTAS ] MHC-230/506MA - MHC-235/517MA - MHC-280/625MA - MHC-320/706MA MHC-500/1109MAR - MHC-680/1466MAR](https://static.cupdf.com/doc/110x72/5e93db5530a5a625c35ecff2/manual-de-usuario-mquinas-de-hielo-manual-de-usuario-autocontenidas-y-remotas.jpg)







