Journal of Plant Physiology 165 (2008) 1610—1619 Metalliferous and non-metalliferous populations of Viola tricolor represent similar mode of antioxidative response Aneta Słomka a, , Marta Libik-Konieczny b , Elz ˙bieta Kuta a , Zbigniew Miszalski b a Department of Plant Cytology and Embryology, Jagiellonian University, 52 Grodzka Street, 31-044 Cracow, Poland b Department of Plant Physiology, Polish Academy of Sciences, 21 Niezapominajek Street, 30-239 Cracow, Poland Received 14 September 2007; received in revised form 12 November 2007; accepted 19 November 2007 KEYWORDS Antioxidative enzymes; Heavy metals; Hydrogen peroxide; Microevolution; Viola tricolor Summary Heavy metal-contaminated sites are excellent areas to examine the antioxidative machinery responsible for physiological adaptations of many plant species. Superoxide dismutase (SOD), guaiacol peroxide (GPX), ascorbate peroxide (APX), catalase (CAT) activity and hydrogen peroxide (H 2 O 2 ) content were analyzed in leaves and roots of Viola tricolor (Viola) from contaminated soils (‘Bukowno’, ‘Saturn’, ‘Warpie’ heaps), and non-contaminated soil (‘Zakopane meadow’) to examine the level of oxidative stress and antioxidative response. In leaves, six isoforms of SOD were recognized. Roots possessed two additional bands, named manganese superoxide dismutase (MnSOD)-like form (MnSODI) and Cu/ZnSOD-like form (Cu/ZnSODIV). The H 2 O 2 content in leaves ranged from 554 to 5 098 mmol H 2 O 2 /gf.w. and was negatively correlated with CAT activity. The non- contaminated population was characterized by the lowest CAT activity combined with the highest H 2 O 2 concentration. Two isoforms of CAT, CAT-1 and CAT-2, were recognized in leaves of plants from non-contaminated and contaminated sites, respectively. In roots of individuals from two heaps (‘Warpie’ and ‘Saturn’), two distinct bands for each CAT isoform were observed. A slower migrating band may be an aggregate, exhibiting CAT and MnSODs activities. Both peroxidases (APX and GPX) presented the same pattern of activity, depending on the organ, indicating that in leaves and roots APX and GPX were regulated in parallel. Differences in enzyme activities and H 2 O 2 content between plants from different contaminated sites were statistically significant, but were tightly maintained at a ARTICLE IN PRESS www.elsevier.de/jplph 0176-1617/$ - see front matter & 2007 Elsevier GmbH. All rights reserved. doi:10.1016/j.jplph.2007.11.004 Abbreviations: APX, ascorbate peroxide; CAT, catalase; Cu/ZnSOD, copper and zinc superoxide dismutase; FeSOD, iron superoxide dismutase; GPX, guaiacol peroxide; H 2 O 2 , hydrogen peroxide; MnSOD, manganese superoxide dismutase; ROS, reactive oxygen species. Corresponding author. Tel.: +4812 6631764; fax: +4812 4228107. E-mail address: [email protected] (A. Słomka).

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ARTICLE IN PRESS
Journal of Plant Physiology 165 (2008) 1610—1619
0176-1617/$ - sdoi:10.1016/j.
Abbreviationdismutase; GPX�CorrespondE-mail addr
www.elsevier.de/jplph
Metalliferous and non-metalliferous populationsof Viola tricolor represent similar mode ofantioxidative response
Aneta Słomkaa,�, Marta Libik-Koniecznyb, Elzbieta Kutaa,Zbigniew Miszalskib
aDepartment of Plant Cytology and Embryology, Jagiellonian University, 52 Grodzka Street, 31-044 Cracow, PolandbDepartment of Plant Physiology, Polish Academy of Sciences, 21 Niezapominajek Street, 30-239 Cracow, Poland
Received 14 September 2007; received in revised form 12 November 2007; accepted 19 November 2007
KEYWORDSAntioxidativeenzymes;Heavy metals;Hydrogen peroxide;Microevolution;Viola tricolor
ee front matter & 2007jplph.2007.11.004
s: APX, ascorbate perox, guaiacol peroxide; H2Oing author. Tel.: +48 12ess: [email protected]
SummaryHeavy metal-contaminated sites are excellent areas to examine the antioxidativemachinery responsible for physiological adaptations of many plant species.Superoxide dismutase (SOD), guaiacol peroxide (GPX), ascorbate peroxide (APX),catalase (CAT) activity and hydrogen peroxide (H2O2) content were analyzed inleaves and roots of Viola tricolor (Viola) from contaminated soils (‘Bukowno’,‘Saturn’, ‘Warpie’ heaps), and non-contaminated soil (‘Zakopane meadow’) toexamine the level of oxidative stress and antioxidative response.In leaves, six isoforms of SOD were recognized. Roots possessed two additionalbands, named manganese superoxide dismutase (MnSOD)-like form (MnSODI) andCu/ZnSOD-like form (Cu/ZnSODIV). The H2O2 content in leaves ranged from 554 to5 098 mmol H2O2/g f.w. and was negatively correlated with CAT activity. The non-contaminated population was characterized by the lowest CAT activity combinedwith the highest H2O2 concentration. Two isoforms of CAT, CAT-1 and CAT-2, wererecognized in leaves of plants from non-contaminated and contaminated sites,respectively. In roots of individuals from two heaps (‘Warpie’ and ‘Saturn’), twodistinct bands for each CAT isoform were observed. A slower migrating band may bean aggregate, exhibiting CATand MnSODs activities. Both peroxidases (APX and GPX)presented the same pattern of activity, depending on the organ, indicating that inleaves and roots APX and GPX were regulated in parallel.Differences in enzyme activities and H2O2 content between plants from differentcontaminated sites were statistically significant, but were tightly maintained at a
Elsevier GmbH. All rights reserved.
ide; CAT, catalase; Cu/ZnSOD, copper and zinc superoxide dismutase; FeSOD, iron superoxide
2, hydrogen peroxide; MnSOD, manganese superoxide dismutase; ROS, reactive oxygen species.6631764; fax: +48 12 4228107..edu.pl (A. Słomka).

ARTICLE IN PRESS
Antioxidative response of Viola tricolor from contaminated soils 1611
very similar level. Prolonged and permanent heavy metal stress evoked a very similarmode of antioxidative response in specimens of analyzed metalliferous populationsnot causing measurable oxidative stress. Thus, our results clearly indicate thatV. tricolor is a taxon well adapted to heavy metal-contaminated soils, and thatdifferences in enzyme activities and H2O2 content result from adjustment of plantsto a variety of conditions.& 2007 Elsevier GmbH. All rights reserved.
Introduction
In southern Poland, in a range of 80 km expandingfrom NW to SE, long-time mining and industrialhuman activity have created an anthropogenicallymodified landscape with ubiquitous heaps and pits.Ore mining has a long history in this region,beginning in the 13th century. Many smelters inthe vicinity of Olkusz that use primitive technologyno longer operate, while others are still working(Grodzinska and Szarek-Łukaszewska, 2002). Thecalamine waste heaps are spoils of different ages.The oldest heaps are more than 100 years old, andyounger heaps are approximately 10–20 years old.Fresh heaps are totally bare. Soils of heaps in theOlkusz area are rich in heavy metals, such as zinc,cadmium and lead, which are distributed inconcentrations exceeding standards by even a1000 times (Szarek-Łukaszewska and Niklinska,2002; Szarek-Łukaszewska et al., 2004). Extremeliving conditions, specifically a microclimatestrongly influenced by wind, has led to very poorseed germination and seedling survival on thewaste heaps (Wierzbicka and Rostanski, 2002). Onlygenotypes resistant to the enormous amounts ofheavy metals in the soil and to physiologicaldrought can survive in the area.
In soils enriched in metals due to geologicalstructures, such as serpentine soils (nickel, copper)or calamine soils (zinc, lead), long-term adapta-tions have created new plant genotypes. Incontrast, old and newly created calamine wasteheaps are colonized by species that have adaptedto these anthropogenically polluted areas in arelatively short time period. Adaptation to theseconditions can be a fast evolutionary process. Theselection of fully metal-tolerant ecotypes may berapid and evolve within some tens of years, or insome plant species, within a few years or even fromthe first contact with the new terrain by a randomlydistributed seed or mature plant already tolerantof the extreme living conditions (Ernst, 1999; Boneand Farres, 2001).
Taxa colonizing soils rich in heavy metals areclassified as either obligate or facultative metallo-phytes. In facultative metallophytes, interspecific
variation enables certain ecotypes to cope withharsh edaphic conditions and favors their adapta-tion to these soil types (Reznick and Ghalambor,2001).
Viola tricolor (heartsease or wild pansy) is amongthe species abundantly present on heaps in theOlkusz area. In the family Violaceae, V. tricolor andViola arvensis are recognized as facultative metal-lophytes, occurring both on contaminated and non-contaminated sites, whereas Viola lutea ssp.westfalica, Viola lutea ssp. calaminaria and Violaboashanensis are considered obligate metallo-phytes with restricted areas of distribution, exclu-sively on soils with enhanced heavy metalsconcentration (Prasad and de Oliviera Freitas,1999; Ernst et al., 2004; Siuta et al., 2005; Zhuanget al., 2005; Bansova et al., 2006; Hildebrandt etal., 2006). Several populations of V. tricolor on soilscontaminated with heavy metals have been de-scribed from Harz Mountains (Germany), fromBanska Stiavnica, Stare Hory (Slovakia) and fromthe ‘‘old’’ Boleslaw heap (Poland) (Dobrzanska,1955; Ernst et al., 2004; Bansova et al., 2006).Recently, two new populations were discovered onheavy metal-rich soils in Southern Poland, the‘Saturn’ heap in Olkusz, and in the vicinity of the‘Warpie’ grove near Chrzanow (Siuta, unpubl.data).
Previous studies on V. tricolor from the Boleslawheap concentrated on determination of the mycor-rhizal status of the species and the mineralcomposition of the rhizosphere (Pawlowska et al.,1996; Cabala et al., 2004). Despite the presenceof large amounts of Fe, Zn and Pb-sulfide inthe ground, plants appeared to be protectedagainst intensified absorption of metal ions bythe occurrence of an abundant (87.3% of rootscolonized) arbuscular myccorhizal fungus (Krupaand Piotrowska-Seget, 2003; Pawlowska and Charvat,2004).
Heavy metals are toxic to plants at a certainthreshold value depending on the species andthe heavy metal content. They affect the plasmamembrane (primarily through lipid peroxidation),DNA and RNA structure, protein expression,photosynthesis and respiration through generation

ARTICLE IN PRESS
A. Słomka et al.1612
of reactive oxygen species (ROS), blocking of func-tional groups in biomolecules, and/or displacementof essential metal ions from biomolecules (Mittler,2002).
Plants exposed to high heavy metal concentra-tions are more susceptible to damage brought on byoverproduction of ROS, generated during oxidativestress, than plants growing under non-heavy metalconditions (Mithofer et al., 2004). H2O2, as themost stable molecule, plays also a signaling role inplants (Vranova et al., 2002). Normally, steady-state levels of O2
d� and H2O2 are sustained bytightly controlled activity of non-enzymatic anti-oxidants such as glutathione, carotenoids, vitaminsC, E and antioxidant enzymes e.g., superoxidasedismutase (SOD, EC 1.15.1.1), catalase (CAT, EC1.11.1.6), ascorbate peroxidase (APX, EC 1.11.1.11),guaiacol peroxidase (GPX, EC 1.11.1.7) (Mittler,2002). During oxidative bursts (rapid, non-controlROS production), the antioxidative system is usuallyoverwhelmed due to a large increase in oxidativeload. Nonetheless, smaller localized bursts of H2O2
production strengthen stress resistance. Overall,H2O2 and O2
d�, as second messengers in the plantcell, orchestrate the selective expression of manydefense genes owing to improved oxidative stresstolerance (Mittler et al., 2004).
For plants growing in soils with a relativelyconstant level of heavy metals, the balancemaintenance between ROS and the antioxidativesystem is crucial for survival and adaptation,although the ability of plants to increase antioxi-dative protection to combat the negative effect ofheavy metals is limited. Many studies have con-firmed that exposure to heavy metals usuallyresults in decreased activity of antioxidativeenzymes (Schutzendubel and Polle, 2002).
Although it is accepted that environmental stressfactors never act alone, experimental studies arenormally restricted to plant response to a specificfactor. It must be considered that stress usuallyacts as a complex of various interacting factors,and consequently, plants are forced to respond to aunique complex of growth conditions. In ourstudies, V. tricolor specimens were collecteddirectly from sites they naturally occupy andimmediately examined. The advantage of researchon plants growing in natural rather than experi-mental conditions is direct documentation ofmicroevolutionary processes, i.e., genotype leveladaptation and its plasticity and ecological status.
The aim of our studies was to examine acti-vity of selected antioxidant enzymes combinedwith concentration of hydrogen peroxide inV. tricolor individuals from contaminated and non-contaminated sites.
Materials and Methods
Plant material
V. tricolor L. (Violaceae) individuals were randomlycollected on sunny days during two summer seasons(June–September 2005 and 2006) from four populationslocalized in southern Poland. Three occurred on Zn–Pbspoil heaps: ‘Bukowno heap’ (BH), ‘Saturn heap’ (SH) and‘Warpie heap’ (WH), and the fourth on a meadow in theTatra Mountains area, near the village Zakopane (ZM).
Because heartsease, (V. tricolor), occurring on zincspoil heaps, is biennial, individuals in bloom wereexamined carefully to collect uniform plant material.
Samples were collected at mid-day and covered withdark plastic bags to avoid light exposure and photo-respiration, and were then immediately transported tothe laboratory. Young leaves similar in size and roots of�20 plants from each stand were washed carefully indistilled water and frozen at �80 1C for further analysis.
Site description
(A)
‘Bukowno heap’ – a small part of ‘‘Michalowska’’quarry. Initially, only galena (PbS) was exploited.Since the XIX century, calamine (ZnCO3 and ZnSiO3),and, before the First World War, sphalerite (ZnS)were the essential stocks. The mining activity in thisarea ended in 1924. In 1950, when a smelting workwas built, the old shafts and tunnels were buried(Krygier et al., 1971). The smelting facility is still inuse; however, ores are transported from a minelocated nearby. The mean concentration of zinc andlead in soil amounts to 6725 and 1769 ppm, respec-tively. The pH is �6.9 (Siuta, unpubl. data).(B)
‘Saturn heap’ – this site exhibits the oldest (800years) remnants of mining activity. The meanconcentrations in soil amount to 1866 ppm forzinc and 3026 ppm for lead. SH differs fromother sites in having enhanced concentrations ofcalcium and magnesium in the soil (Ca �60,250 ppm,Mg –33,858 ppm), which are approximately fourtimes higher than at other sites. The pH of the soilis slightly alkaline (7.4) (Siuta, unpubl. data).(C)
‘Warpie heap’ – this site is located in the old districtof Chrzanow city. The last documented marks of mi-ning activity come from the 18th century (Molenda,1963). The ores are still exploited in the Balin-Trzebionka mine located 12 km from the ‘Warpieheap’. The pH of the soil is the same as for thesoil from SH (7.4). The concentration of lead in thesoil is the highest among all the sites and reached5004 ppm.(D)
‘Zakopane meadow’ – this site is located at 838mabove sea level. The soil is richer in copper and ironcompared with others sites. The pH of the soil is 7.1(Siuta, unpubl. data). The population from Zakopanewas recognized as a control due to low zinc and leadconcentrations in the soil.
ARTICLE IN PRESS
Antioxidative response of Viola tricolor from contaminated soils 1613
Methods
Protein isolationTo isolate fractions of soluble proteins, we homoge-
nized plant material (1 g fresh weight) at 4 1C with amortar in 2.5mL homogenization buffer (17.9 g L�1
Tricine, 0.74 g L�1 MgSO4, 0.155 g L�1 DTT, 1.14 g L�1
EDTA, adjusted with 1M Tris to pH 8.0). Non-solublematerial was removed by centrifugation for 1min at12,000g.
Determination of protein contentThe protein concentration was determined according
to Bradford (1976) using the BioRad protein assay(Bio-Rad, Hercules, California) with BSA as a standard.Protein fractions were used immediately for measuringenzyme activity or stored at �80 1C until further use.
Analysis of superoxide dismutase (SOD) and catalase(CAT) activity by native PAGE
For determination of SOD and CAT activity, weanalyzed the fractions of soluble proteins isolated asdescribed above using native PAGE at 4 1C and 180 V andthe Laemmli (1970) buffer system without sodiumdodecyl sulfate.
SOD bands were visualized on 12% polyacrylamide gelsusing the activity staining procedure described byMiszalski et al. (1998). Isoforms of SOD were identifiedusing two inhibitors: 3mM KCN inhibiting Cu/ZnSOD and5mM H2O2 inhibiting Cu/ZnSOD and FeSOD.
CAT activity was visualized after electrophoresis on10% polyacrylamide gels according to the methoddescribed by Woodbury et al. (1971).
Spectrophotometric analysis of catalase (CAT),guaiacol peroxide (GPX) and ascorbate peroxidase(APX) activity
CAT activity was measured according to the methoddescribed by Aebi (1984). The disappearance of H2O2
[initial concentration: 0.04% (v/v) H2O2] in a phosphatebuffer (50mM KH2PO4, 50Mm Na2HPO4, pH 7.0) wasmonitored at 240 nm. The enzyme activity [I.U.] wasdefined as 1mmol of H2O2 decomposed per minute permilligram of total protein. For calculation, the absor-bance coefficient 43 LM�1 cm�1 was used.
GPX was determined according to Zheng and vanHuystee (1992). The reaction was run for 5min at 25 1C,in 1mL cuvette for 50 mL of purified extract in 300mMpotassium phosphate buffer, pH 6.1, in the presence of8.42mM guaiacol and 2.10mM H2O2. Increase of absor-bance was read at 470 nm and calculated with the use ofabsorbance coefficient 26,600 LM�1 cm�1.
APX was measured according to Nakana and Asada(1981). The reaction was run for 2min at 25 1C, in 1mLcuvette for 50 mL extract in 300mM KPO4, pH 7.0, in thepresence of 0.75mM ascorbate and 2mM H2O2. Decreaseof absorbance was read at 290 nm and calculated withthe use of absorbance coefficient 2800 LM�1 cm�1.
Spectrophotometric analysis of endogenous hydrogenperoxide (H2O2) concentration
The endogenous H2O2 level was investigated accordingto the modified method described previously by Brennanand Frenkel (1977). Hydrogen peroxide was extracted bythe homogenization of 0.5–1 g of tissue in 2mL of coldacetone. After centrifugation (5min; 12,000g) the pelletwas discarded, and 0.5mL of the extract was collected.Titanium reagent (50 mL of 20% titanium tetrachloride inconcentrated HCl, v/v) was added to 0.5mL of theextract, followed by the addition of 0.1mL of NH3 aq.(25%) to precipitate the peroxide–titanium complex.After 5min of centrifugation at 10,000g, the supernatantwas discarded and the precipitate was repeatedlywashed in 1mL of acetone and centrifuged for 5min at10,000g. The precipitate was solubilized in 1mL of 1MH2SO4, and brought to a final volume of 2mL. Theabsorbance of the obtained solution was read at 415 nmagainst a water blank. The concentration of peroxide inthe extract was determined by comparing the absorbanceagainst a standard curve representing the titanium–H2O2
complex, over the range from 0 to 300 mM. All H2O2
measurements were normalized to a fresh weight oftissue.
Statistical analysis
Statistical analyses were performed using theSTATISTICA 7.1 computer program. To determine indivi-dual treatment effects at the Pp0.05 level of prob-ability, we used a two-way ANOVA followed by a Duncan’smultiple range test.
Results
SOD activity was visualized on native polyacryla-mide gels after electrophoretic separation ofisolated proteins. Leaves and roots exhibitedactivity of several SOD isoforms (Figure 1A and B),which were recognized after selective stainingusing specific inhibitors (Figure 1C). In roots,in contrast to leaves, additional bands of manga-nese superoxide dismutase (MnSOD) (MnSODI)and Cu/ZnSOD (Cu/ZnSODIV) were recognized(Figure 1B). SOD activity was higher in roots thanin leaves (Figure 1A and B). In leaves of plantsgrowing on the WH, SOD activity was higher than inleaves collected from other sites, while in roots,activity of this enzyme was the lowest amonganalyzed root samples (Figure 1A and B).
The H2O2 concentration in leaves was as much asseveral times higher than in roots (Figure 2A and B),excluding leaves from the WH, which showed asignificantly lower H2O2 level (Figure 2A) similar tothe concentration in roots (Figure 2B). In bothorgans, except for roots from the BH, the tendencyto decrease the level of H2O2 in plants from
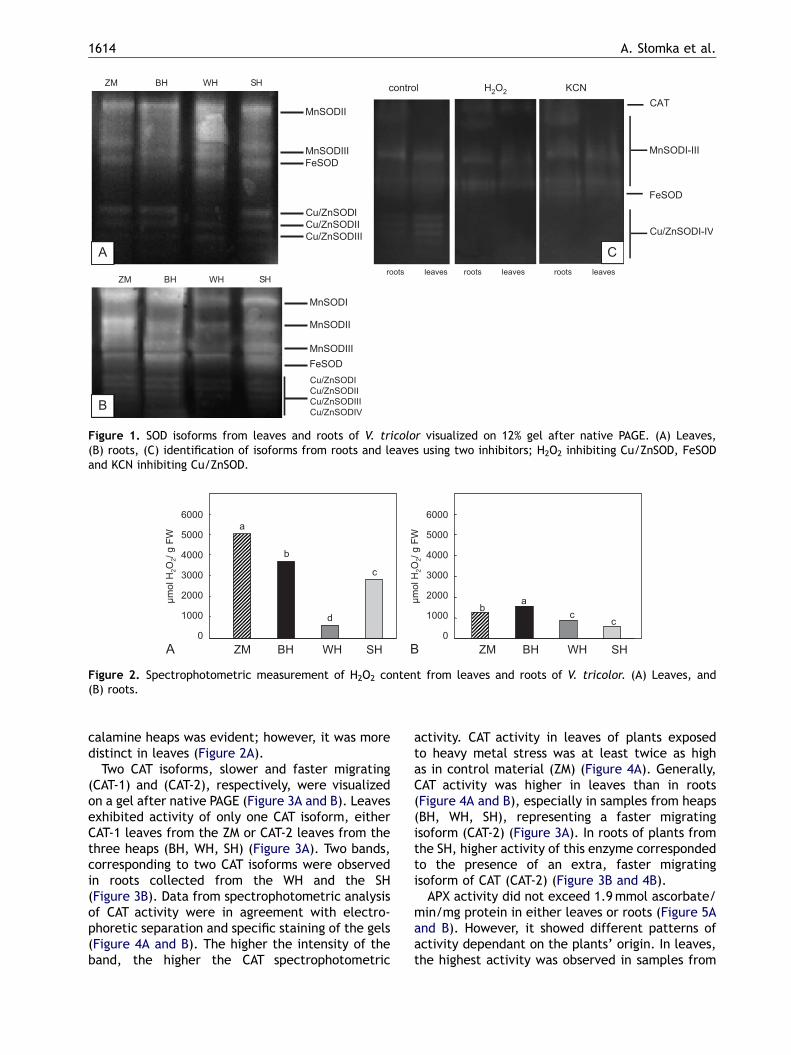
ARTICLE IN PRESS
Figure 1. SOD isoforms from leaves and roots of V. tricolor visualized on 12% gel after native PAGE. (A) Leaves,(B) roots, (C) identification of isoforms from roots and leaves using two inhibitors; H2O2 inhibiting Cu/ZnSOD, FeSODand KCN inhibiting Cu/ZnSOD.
Figure 2. Spectrophotometric measurement of H2O2 content from leaves and roots of V. tricolor. (A) Leaves, and(B) roots.
A. Słomka et al.1614
calamine heaps was evident; however, it was moredistinct in leaves (Figure 2A).
Two CAT isoforms, slower and faster migrating(CAT-1) and (CAT-2), respectively, were visualizedon a gel after native PAGE (Figure 3A and B). Leavesexhibited activity of only one CAT isoform, eitherCAT-1 leaves from the ZM or CAT-2 leaves from thethree heaps (BH, WH, SH) (Figure 3A). Two bands,corresponding to two CAT isoforms were observedin roots collected from the WH and the SH(Figure 3B). Data from spectrophotometric analysisof CAT activity were in agreement with electro-phoretic separation and specific staining of the gels(Figure 4A and B). The higher the intensity of theband, the higher the CAT spectrophotometric
activity. CAT activity in leaves of plants exposedto heavy metal stress was at least twice as highas in control material (ZM) (Figure 4A). Generally,CAT activity was higher in leaves than in roots(Figure 4A and B), especially in samples from heaps(BH, WH, SH), representing a faster migratingisoform (CAT-2) (Figure 3A). In roots of plants fromthe SH, higher activity of this enzyme correspondedto the presence of an extra, faster migratingisoform of CAT (CAT-2) (Figure 3B and 4B).
APX activity did not exceed 1.9mmol ascorbate/min/mg protein in either leaves or roots (Figure 5Aand B). However, it showed different patterns ofactivity dependant on the plants’ origin. In leaves,the highest activity was observed in samples from

ARTICLE IN PRESS
Antioxidative response of Viola tricolor from contaminated soils 1615
the ZM and the SH (Figure 5A), while in roots,activity decreased gradually and significantly, withthe highest activity in plants from the ZM and thelowest in plants from the SH (Figure 5B).
GPX activity exhibited differences betweenstudied samples, and was distinctly higher in roots
Figure 4. Spectrophotometric measurement of catalase acti(B) roots.
Figure 5. Spectrophotometric measurement of APX activit(B) roots.
Figure 3. Catalase isoforms from leaves and roots ofV. tricolor visualized on 10% gel after native PAGE.(A) Leaves, and (B) roots.
than in leaves (Figure 6A and B). In leaves, thehighest level of GPX activity was noted in plantsfrom the ZM and the SH (Figure 6A). In roots, it wasthe highest in the ZM and the lowest in the SH(Figure 6B). The patterns of GPX and APX activity,disposing H2O2, were similar and typical for eachorgan (compare Figures 5 and 6), which maysuggest that their activities are regulated inparallel.
Discussion
The analyses of antioxidative response of plantsoriginating from sites contaminated with heavymetals are thus far limited to algae, lichens andferns (Ratkevicius et al., 2003; Cao et al., 2004;Cuny et al., 2004). Data concerning floweringplants have focused mainly on cultivated plantssubjected to heavy metal treatment under labora-tory conditions (Chen et al., 2000; Kuo and Kao,2004; Qadir et al., 2004; Metwally et al., 2005;Tiryakioglu et al., 2006) and to hyperaccumulatorsof heavy metals (Boominathan and Doran, 2002;Wang et al., 2004; Wojcik et al., 2006). Our resultsprovide valuable information about the mode of
vity from leaves and roots of V. tricolor. (A) Leaves, and
y from leaves and roots of V. tricolor. (A) Leaves, and

ARTICLE IN PRESS
Figure 6. Spectrophotometric measurement of GPX activity from leaves and roots of V. tricolor. (A) Leaves, and(B) roots.
A. Słomka et al.1616
antioxidative defense in common plant specieswidespread on calamine soils.
It is known that SOD is the main antioxidantenzyme functioning as a superoxide radicalscavenger in living organisms, a first-line defenderagainst oxidative stress caused by ROS. V. tricolorproduces several SOD isoforms. Generally,roots showed higher SOD activity, expressed bythe presence of additional bands (MnSODI andCu/ZnSODIV) not detected in leaves. This is notsurprising because roots are directly affected byhigh heavy metal concentrations in the soil and arealso directly affected by drought.
SOD catalyzes a reaction producing hydrogenperoxide, the most stable molecule among all ROS,often measured as a marker of oxidative stress(Cheeseman, 2006). The concentration of thismolecule should thus correlate with SOD activity.This was confirmed in roots of V. tricolor from theZM and the BH. Higher H2O2 concentrations wereaccompanied by higher SOD activity, in contrast toresults obtained from leaves from the WH, wherelow H2O2 concentrations accompanied high SODactivity. These results might suggest that, in leavesfrom the WH, H2O2 is produced independently ofSOD function.
It is evident that the level of H2O2 differssignificantly between leaves and roots. Thesedifferences do not necessarily reveal signs ofmeasurable oxidative stress. It is known thatspecies well adapted to harsh conditions maintainstable levels of H2O2, owing to either low levelsof ROS production or high capabilities for deto-xification (Cheeseman, 2006). The substantialdifferences in H2O2 content (in mmol/mg f.w.)between leaves and roots of V. tricolor areconsistent with those found in other species(Dixit et al., 2001). In Hordeum vulgare, thegreater load of H2O2 was observed in leaves,although de facto roots were affected by heavymetals (Erdei et al., 2002).
The level of hydrogen peroxide in plant cells isunder the control of CAT and peroxidases, whichcan lower concentration if it is produced in excess.In flowering plants, there are three genes thatencode CAT isoforms. External factors stimulate thetranscription of these genes in a different way(Pereira et al., 2002). We observed only twoisoforms of CAT in leaves, CAT-1 in leaves fromthe ZM and CAT-2 in leaves from the three heaps.Roots of V. tricolor from the WH and SH possessedtwo distinct bands for each. Sandalio et al. (2001)found the presence of two bands of CAT visualizedin traditional native PAGE in leaves of Pisumsativum treated with cadmium and mercury.CAT is a homotetrameric protein and exhibits atendency to form aggregates (Sheptovitsky andBrudvig, 1996). In Mesembryanthemum crystal-linum leaves, two bands of CAT were also revealed(Niewiadomska and Miszalski, unpubl. data). Theauthors suggested that a slower migrating band isan aggregate exhibiting CATand MnSOD activity. Wecannot exclude the possibility that, in V. tricolorleaves, the CAT-1 band represents activity of bothCAT and MnSODII and/or MnSODIII. The fastermigrating band (CAT-2) may correspond to majorCAT activity and be responsible for H2O2 removal.Thus, in leaves of plants growing in heavy metals-contaminated sites, we observed a decrease ofH2O2 content. Similarly, the decrease of H2O2
concentration in roots from the WH and the SHcould be explained by the presence of CAT-2 inthese organs. The decrease of CAT activity mayresult not only from changes in the assembly ofenzyme subunits but also from the decrease of totalprotein content due to inhibition of enzymesynthesis or photoinactivation, or by a combinationof all three (Pereira et al., 2002).
APX and GPX are other enzymes involved inthe removal of H2O2. It should be noted thatboth peroxidases participate in controlling slightchanges in hydrogen peroxide concentration, which

ARTICLE IN PRESS
Antioxidative response of Viola tricolor from contaminated soils 1617
result from a metabolic pathway rather than fromoxidative stress. In contrast, high H2O2 content isusually removed by CAT. The highest activity ofboth peroxidases coupled with low CAT activity inroots and leaves from the ZM population suggestshigh metabolic activity for the plants. In rootsof V. tricolor, CATand peroxidases seem to functionconcurrently. Low CAT activity followed higher APXand GPX activity, and, inversely, high CAT activityresulted in lower total peroxidase activity. Thesame mode of regulation of H2O2 has beendescribed in the arsenic-hyperaccumulator Pterisvittata (Cao et al., 2004). Parallel regulation of APXand GPX activity (compare Figures 5 and 6)observed in V. tricolor is in accordance with dataof Wojcik et al. (2006), who observed complemen-tary activity of these enzymes under zinc stress inThlaspi caerulescens.
The ability of plants to increase antioxidativeprotection to combat the negative consequences ofheavy metal stress appears to be limited, as manystudies have shown that exposure to elevatedconcentrations of redox reactive metals result indecreased, and not in increased, activities ofantioxidative enzymes (Schutzendubel and Polle,2002). The conclusion that enzymatic activityincreases in lower heavy metal concentrationsand decreases when they become higher is gene-rally accepted (Cao et al., 2004). Similarly, theprolonged influence of heavy metals initiallyinduces an increase of enzyme activity, followedby a subsequent decrease (Qadir et al., 2004). Wemay assume that if V. tricolor colonized heaps foryears, the species might have developed andimproved physiological mechanisms that enable itto survive and propagate. Plants of V. tricolor fromall populations possessed a tightly regulated andbalanced state between activities of their antiox-idative enzymes and ROS. The prolonged andchronic heavy metal stress evoked a very similarmode of antioxidative response in all representa-tives of the four analyzed populations and did notcause measurable oxidative stress. In conclusion,plants of V. tricolor are well adapted to heavymetal-contaminated soils they occupy. Differencesin enzymes activities and H2O2 content are theresult of adjustment of the plants to variousconditions.
Acknowledgments
The authors would like to express their gratitudeto Professor Hermann Bothe from the University ofCologne for stimulating discussion and valuablecomments. The authors are in debt to Dr. Grazyna
Szarek-Łukaszewska for information concerningsites of plant origin and help in analyses of heavymetal content in the soil.
This work was supported by the JagiellonianUniversity research grant No. DBN–414/CRBW/XVII-19/2005.
References
Aebi H. Catalase in vitro. Methods Enzymol 1984;105:121–6.
Bansova V, Horak O, Ciamporova M, Nadubinska M,Lichtscheidl I. The vegetation of metalliferous andnon-metalliferous grassland in two former mineregions in central Slovakia. Biol Bratislava 2006;61(3):1–7.
Bone E, Farres A. Trends and rates of microevolution inplants. Genetica 2001;112–113:165–82.
Boominathan R, Doran PM. Ni-induced oxidative stress inroots of the Ni hyperaccumulator, Alyssum bertolonii.New Phytol 2002;156:205–15.
Bradford MM. A rapid and sensitive method for quantifi-cation of microgram quantities of protein utilizing theprinciple of protein–dye binding. Anal Biochem 1976;72:248–54.
Brennan T, Frenkel C. Involvement of hydrogen peroxidein the regulation of senescence in pear. Plant Physiol1977;59:411–6.
Cabala J, Teper E, Teper L, Małkowski E, Rostanski A.Mineral composition in rhizosphere of plants grown inthe vicinity of Zn–Pb ore flotation tailings Pond.Preliminary study. Acta Biol Cracov Bot 2004;46:65–74.
Cao X, Ma LQ, Tu C. Antioxidative responses toarsenic in the arsenic-hyperaccumulator Chinesebrake fern (Pteris vittata L.). Environ Pollut 2004;128:317–25.
Cheeseman JM. Hydrogen peroxide concentrations inleaves under natural conditions. J Exp Bot 2006;57(10):2435–44.
Chen L-M, Lin ChCh, Kao ChH. Copper toxicity in riceseedlings: changes in antioxidative enzyme activities,H2O2 level, and cell wall peroxidase activity in roots.Bot Bull Acad Sin 2000;41:99–103.
Cuny D, van Haluwyn Ch, Shirali P, Zerimech F, Jerome L,Haguenoer JM. Cellular impact of metal trace ele-ments in terricolous lichen Diploschistes muscorum(Scop.) R. Sant. – identification of oxidative stressbiomarkers. Water Air Soil Pollut 2004;152:55–69.
Dixit V, Pandey V, Shyam R. Differential antioxidativeresponse to cadmium in roots and leaves of pea(Pisum sativum L. cv. Azad). J Exp Bot 2001;52(358):1101–9.
Dobrzanska J. Flora and ecological studies on calamineflora in the district of Boleslaw and Olkusz. Acta SocBot Pol 1955;24(2):357–407.
Erdei S, Heged +us A, Hauptmann G, Szalai J, Horvath G.Heavy metal induced physiological changes in theantioxidative response system. Proceedings of the 7th

ARTICLE IN PRESS
A. Słomka et al.1618
Hungarian congress on plant physiology. Acta BiolSzegediensis 2002;46(3–4):89–90.
Ernst WHO. Evolution of plants on soils antropoge-nically contaminated by heavy metals. In: VanRaamsdonk LWD, den Nijs JCM, editors. Plant evolu-tion in man-made habitats. Proceedings of the VIIthsymposium IOPB. Amsterdam: Hugo de Vries Labora-tory; 1999.
Ernst WHO, Knolle F, Kratz S, Schung E. Aspects ofecotoxicology of heavy metals in the Harz region – aguided excursion. Landbauforsch V +olkenrode 2004;54:53–71.
Grodzinska K, Szarek-Łukaszewska G. Hałdy cynkowo-ołowiowe w okolicach Olkusza – przeszłosc, teraz-niejszosc i przyszłosc. Kosmos 2002;51:127–38.
Hildebrandt U, Hoef-Emden K, Backhausen S, Bothe H,Bozek M, Siuta A, et al. The rare endemic zinc violetsof Central Europe originate from Viola lutea Huds.Plant Syst Evol 2006;257(3–4):205–22.
Krupa P, Piotrowska-Seget Z. Positive aspects of interac-tion between plants and mycorrhizal fungi originatingfrom soils polluted with cadmium. Polish J Exp Stud2003;12(6):723–6.
Krygier E, Molenda D, Saładziak A. Katalog zabytkowbudownictwa przemysłowego w Polsce. Powiat Olkusz– wojewodztwo krakowskie. Vol. 3(4), part 1. Zabytkigornicze. Wrocław, Warszawa, Krakow, Gdansk: ZakładNarodowy im. Ossolinskich. Wydawnictwo PAN; 1971.
Kuo MCh, Kao ChH. Antoxidant enzyme activities areupregulated in response to cadmium in sensitive, butnot in tolerant, rice (Oryza sativa L.) seedlings. BotBull Acad Sin 2004;45:291–9.
Laemmli UK. Cleavage of structural proteins during theassembly of the head phase of bacteriophage T4.Nature 1970;227:680–5.
Metwally A, Safronova VI, Belimov AA, Dietz KJ.Genotypic variation of the response to cadmiumtoxicity in Pisum sativum L. J Exp Bot 2005;56(409):167–78.
Miszalski Z, Slesak I, Niewiadomska E, Baczek-Kwinta R,Luttge U, Ratajczak R. Subcellular localizationof superoxide dismutase isoforms from leaves inthe C3-CAM intermediate halophyte Mesembrynathe-mum crystallinum L. Plant Cell Environ 1998;21:169–79.
Mithofer A, Schulze B, Boland W. Biotic and heavy metalstress response in plants: evidence from commonsignals. FEBS Lett 2004;566:1–5.
Mittler R. Oxidative stress, antioxidants and stresstolerance. Trends Plant Sci 2002;7(9):405–10.
Mittler R, Vanderauwera S, Gollery M, van Breusegem F.Reactive oxygen gene network of plants. Trends PlantSci 2004;9(10):490–8.
Molenda D. Gornictwo kruszcowe na terenie złoz slasko-krakowskich do połowy XVI w, vol. 15. Wrocław,Warszawa, Krakow: Zakład Narodowy im. Ossolinskich.Wydawnictwo PAN; 1963.
Nakana Y, Asada K. Hydrogen peroxide is scavenged byascorbate-specific peroxidase in spinach chloroplasts.Plant Cell Physiol 1981;22:867–80.
Pawlowska TE, Charvat I. Heavy-metal stress and devel-opmental patterns of arbuscular mycorrhizal fungi.Appl Environ Microbiol 2004;70(11):6643–9.
Pawlowska TE, Błaszkowski J, R +uhling (A. The mycorrhizalstatus of plant colonizing a calamine spoil mound insouthern Poland. Mycorrhiza 1996;6:499–505.
Pereira GJG, Molina SMG, Lea PJ, Azevedo RA. Activity ofantioxidant enzymes in response to cadmium inCrotalaria juncea. Plant and Soil 2002;239:123–32.
Prasad MNV, de Oliviera Freitas HM. Feasible biotechno-logical and bioremediation strategies for serpentinesoils and mine spoils. Electron J Plant Biotechnol1999;2(1):20–34.
Qadir S, Qureshi MI, Javed S, Abdin MZ. Genotypicvariation in phytoremediation potential of Brassicajuncea cultivars exposed to Cd stress. Plant Sci 2004;167:1171–81.
Ratkevicius N, Correa JA, Moenne A. Copper accumula-tion, synthesis of ascorbate and activation of ascor-bate peroxide in Enteromorpha compressa (L.)Grev. (Chlorophyta) from heavy metal-enriched en-vironments in northern Chile. Plant Cell Environ 2003;26:1599–608.
Reznick DN, Ghalambor CK. The population ecology ofcontemporary adaptations: what empirical studiesreveal about the conditions that promote adaptiveevolution. Genetica 2001;112-113:183–98.
Sandalio LM, Dalurzo HC, Gomez M, Romero-Puertas MC,del Rio LA. Cadmium-induced changes in the growthand oxidative metabolism of pea plants. J Exp Bot2001;52(364):2115–26.
Schutzendubel A, Polle A. Plant responses to abioticstress: heavy metal-induced oxidative stress andprotection by mycorrhization. J Exp Bot 2002;53(372):1351–65.
Sheptovitsky YG, Brudvig GW. Isolation and characteriza-tion of spinach photosystem II membrane-associatedcatalase and polyphenol oxidase. Biochemistry 1996;35:16255–63.
Siuta A, Bozek M, Jedrzejczyk M, Rostanski A, Kuta E. Isthe blue zinc violet (Viola guestphalica Nauenb.)a taxon of hybrid origin? – Evidence from embryology.Acta Biol Cracov Bot 2005;47(1):237–45.
Szarek-Łukaszewska G, Niklinska M. Concentrationsof alkaline and heavy metals in Biscutella laevigataL. and Plantago lancoelata L. growing on calaminespoils (S. Poland). Acta Biol Cracov Bot 2002;44:29–38.
Szarek-Łukaszewska G, Słysz A, Wierzbicka M. Responseof Armeria maritima (Mill.) Willd. to Cd, Zn and Pb.Acta Biol Cracov Bot 2004;46:19–24.
Tiryakioglu M, Eker S, Ozkutlu F, Husted S, Cakmak I.Antioxidant defense system and cadmium uptake inbarley genotypes differing in cadmium tolerance.J Trace Elem Med Biol 2006;20:181–9.
Vranova E, Inze D, van Breusegem F. Signal transductionduring oxidative stress. J Exp Bot 2002;53(372):1227–36.
Wang H, Shan X-q, Wen B, Zhang S, Wang Z-j. Responsesof antioxidative enzymes to accumulation of copper ina copper hyperaccumulator of Commoelina communis.Arch Environ Contam Toxicol 2004;47:185–92.

ARTICLE IN PRESS
Antioxidative response of Viola tricolor from contaminated soils 1619
Wierzbicka M, Rostanski A. Microevolutionary changes inecotypes of calamine waste heap vegetation nearOlkusz, Poland: a review. Acta Biol Cracov Bot 2002;44:7–19.
Woodbury W, Spencer AK, Stahman MA. An improvedprocedure using ferricyanide for detecting catalaseisozymes. Anal Biochem 1971;44:301–5.
Wojcik M, Skorzynska-Polit E, Tukiendorf A. Organic acidsaccumulation and antioxidative enzyme activities in
Thlaspi caerulescens under Zn and Cd stress. J PlantGrowth Regul 2006;48:145–55.
Zheng X, van Huystee RB. Peroxidase-regulated elonga-tion of segments from peanuts hypocotyls. Plant Sci1992;81:47–56.
Zhuang P, Ye ZH, Lan CY, Xie ZW, Shu WS. Chemicallyassisted phytoextraction of heavy metal contaminatedsoils using three plant species. Plant Soil 2005;276:153–62.
Related Documents











