J Pineal Res 1998: 24rl79-192 Printed in the United States ofAmerica--all rights reserved. Copyright 0 Munksgaard. 1998 ISSN 0742-3098 Journal of Pineal Research Melatonin prevents apoptosis induced by 6-hydroxydopamine in neuronal cells: Implications for Parkinson’s disease Mayo JC, Sainz RM, Uria H, Antolin I, Esteban MM, Rodriguez C. Melatonin prevents apoptosis induced by 6-hydroxydopamine in neuronal cells: Implications for Parkinson’s disease. J. Pineal Res. 1998; 24: 179- 1 9 2 . 0 Munksgaard, Copenhagen Abstract: It was recently reported that low doses of 6-hydroxydopamine (6- OHDA) induce apoptosis of naive (undifferentiated) and neuronal (differentiated) PC 12 cells, and this system has been proposed as an adequate experimental model for the study of Parkinson’s disease. The mechanism by which this neurotoxin damages cells is via the production of free radicals. Given that the neurohormone melatonin has been reported 1) to be a highly effective endogenous free radical scavenger, 2) to increase the mRNA levels and the activity of several antioxidant enzymes, and 3) to inhibit apoptosis in other tissues, we have studied the ability of melatonin to prevent the programmed cell death induced by 6-OHDA in PC12 cells. We found that melatonin prevents the apoptosis caused by 6-OHDA in naive and neuronal PC12 cells as estimated by 1) cell viability assays, 2) counting of the number of apoptotic cells, and 3) analysis and quantification of DNA fragmentation. Exploration of the mechanisms used by melatonin to reduce programmed cell death revealed that this chemical mediator prevents the 6-OHDA induced reduction of mRNAs for several antioxidant enzymes. The possibility that melatonin utilized additional mechanisms to prevent apoptosis of these cells is also discussed. Since this endogenous agent has no known side effects and readily crosses the blood- brain-barrier, we consider melatonin to have a high clinical potential in the treatment of Parkinson’s disease and possibly other neurodegenerative diseases, although more research on the mechanisms is yet to be done. Introduction Exogenous neurotoxin damage has been proposed as the cause for neurodegenerative diseases (Meldrum and Garthwaite, 1990; Zuber and Alperovitch, 1991; Drachman and Lippa, 1992; Martin and Beal, 1992). In vivo models of Parkinson’s disease mainly use two neurotoxins: 6-hydroxydopamine (6-OHDA) and 1 -methyl-4- phenyltetrahydropyridine (MPTP) (Zigmond and Stricker, 1989). Both of these agents destroy cat- echolamine containing neurons although by differ- ent mechanisms: 6-OHDA toxicity is mediated by the production of oxygen species by nonenzymatic oxidation (Cohen and Heikkila, 1974), while MPTP inhibits the NADH-linked electron transport in mi- Juan Carlos Mayo,’ Rosa Maria Sainz,’ Higinio Uria,’ Isaac Antolin, Manuel M. Esteban? and Carmen Rodriguez’ ‘Departamento de Morfologia y Biologia Celular and ‘Departmento de biologia Funcional, Universidad de Oviedo, c/Julian Claveria s/n 33006 Oviedo, Spain Key words: melatonin - apoptosis - antioxidant enzymes mRNA - PC12 cells - 6-OHDA - Parkinson’s disease Address reprint requests to Or. Carmen Rodriguez, Departamento de Morfologia y Biologia Celular, Facultad de Medicina, c/Julian Claveria, 33006, Oviedo, Spain. E-mail: [email protected] Received November 4, 1997; accepted December 1, 1997. tochondria (Ramsay et al., 1986); 6-OHDA destroys selectively catecholaminergic neurons (Ichitany et al., 1991) as its injection into the substantia nigra induces degeneration of the nigro-striatal dopamine system, with reduction of dopamine in the striatum and in the substantia nigra (Michel and Hefty, 1990). These neurotoxins induce programmed cell death in PC12 cells (Walkinshaw and Waters, 1994; Desole et al., 1996). Apoptotic cells present typical morphological and biochemical markers very differ- ent from cells dying of necrosis, the other usual cel- lular death (Wyllie et al., 1980). Walkinshaw and Waters (1994) found that low doses of 6-OHDA cause apoptotic cell death of a small number of neu- rons, although repeated exposure can kill most of 179

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

J Pineal Res 1998: 24rl79-192 Printed in the United States ofAmerica--all rights reserved.
Copyright 0 Munksgaard. 1998
ISSN 0742-3098 Journal of Pineal Research
Melatonin prevents apoptosis induced by 6-hydroxydopamine in neuronal cells: Implications for Parkinson’s disease
Mayo JC, Sainz RM, Uria H, Antolin I, Esteban MM, Rodriguez C. Melatonin prevents apoptosis induced by 6-hydroxydopamine in neuronal cells: Implications for Parkinson’s disease. J. Pineal Res. 1998; 24: 179- 192 .0 Munksgaard, Copenhagen
Abstract: It was recently reported that low doses of 6-hydroxydopamine (6- OHDA) induce apoptosis of naive (undifferentiated) and neuronal (differentiated) PC 12 cells, and this system has been proposed as an adequate experimental model for the study of Parkinson’s disease. The mechanism by which this neurotoxin damages cells is via the production of free radicals. Given that the neurohormone melatonin has been reported 1) to be a highly effective endogenous free radical scavenger, 2) to increase the mRNA levels and the activity of several antioxidant enzymes, and 3 ) to inhibit apoptosis in other tissues, we have studied the ability of melatonin to prevent the programmed cell death induced by 6-OHDA in PC12 cells. We found that melatonin prevents the apoptosis caused by 6-OHDA in naive and neuronal PC12 cells as estimated by 1) cell viability assays, 2) counting of the number of apoptotic cells, and 3 ) analysis and quantification of DNA fragmentation. Exploration of the mechanisms used by melatonin to reduce programmed cell death revealed that this chemical mediator prevents the 6-OHDA induced reduction of mRNAs for several antioxidant enzymes. The possibility that melatonin utilized additional mechanisms to prevent apoptosis of these cells is also discussed. Since this endogenous agent has no known side effects and readily crosses the blood- brain-barrier, we consider melatonin to have a high clinical potential in the treatment of Parkinson’s disease and possibly other neurodegenerative diseases, although more research on the mechanisms is yet to be done.
Introduction
Exogenous neurotoxin damage has been proposed as the cause for neurodegenerative diseases (Meldrum and Garthwaite, 1990; Zuber and Alperovitch, 1991; Drachman and Lippa, 1992; Martin and Beal, 1992). In vivo models of Parkinson’s disease mainly use two neurotoxins: 6-hydroxydopamine (6-OHDA) and 1 -methyl-4- phenyltetrahydropyridine (MPTP) (Zigmond and Stricker, 1989). Both of these agents destroy cat- echolamine containing neurons although by differ- ent mechanisms: 6-OHDA toxicity is mediated by the production of oxygen species by nonenzymatic oxidation (Cohen and Heikkila, 1974), while MPTP inhibits the NADH-linked electron transport in mi-
Juan Carlos Mayo,’ Rosa Maria Sainz,’ Higinio Uria,’ Isaac Antolin, Manuel M. Esteban? and Carmen Rodriguez’ ‘Departamento de Morfologia y Biologia Celular and ‘Departmento de biologia Funcional, Universidad de Oviedo, c/Julian Claveria s/n 33006 Oviedo, Spain
Key words: melatonin - apoptosis - antioxidant enzymes mRNA - PC12 cells - 6-OHDA - Parkinson’s disease
Address reprint requests to Or. Carmen Rodriguez, Departamento de Morfologia y Biologia Celular, Facultad de Medicina, c/Julian Claveria, 33006, Oviedo, Spain. E-mail: [email protected]
Received November 4, 1997; accepted December 1, 1997.
tochondria (Ramsay et al., 1986); 6-OHDA destroys selectively catecholaminergic neurons (Ichitany et al., 1991) as its injection into the substantia nigra induces degeneration of the nigro-striatal dopamine system, with reduction of dopamine in the striatum and in the substantia nigra (Michel and Hefty, 1990).
These neurotoxins induce programmed cell death in PC12 cells (Walkinshaw and Waters, 1994; Desole et al., 1996). Apoptotic cells present typical morphological and biochemical markers very differ- ent from cells dying of necrosis, the other usual cel- lular death (Wyllie et al., 1980). Walkinshaw and Waters (1994) found that low doses of 6-OHDA cause apoptotic cell death of a small number of neu- rons, although repeated exposure can kill most of
179

Mayo et a].
the cells, demonstrating that they are not resistant to the toxin. On the other hand, high doses mas- sively kill cells by necrosis. When neurons die of necrosis, several die at the same time and the sur- rounding tissue shows signs of inflammation result- ing in early clinical symptoms. As Parkinson symptoms may require years to become apparent, when 80% of the cells in the substantia nigra are already lost, it is likely that recurrent apoptosis of small number of cells after long term exposure to very low levels of a neurotoxin may be the way in which the neurons die.
Melatonin easily passes the blood-brain barrier (Reiter, 1991) entering neurons and glial cells (Menendez-Pelaez and Reiter, 1993; Menendez- Pelaez et al., 1993). This chemical agent exerts a strong antioxidant activity and is a highly effective endogenous free radical scavenger (Tan et al., 1993; Pieri et al., 1994; Reiter, 1997). Additionally, it in- creases the mRNAs for antioxidant enzymes in sev- eral tissues (Antolin et al., 1996; Kotler et al., 1998). This agent protects DNA, proteins and lipids in cel- lular membranes against free radical attack (Pierrefiche et al., 1993; Tan et al., 1994; Abe et al., 1994; Melchiorri et al., 1995). Additionally, we have shown that melatonin can prevent physi- ological apoptosis of thymocytes, both in vivo and in vitro, due to age and dexametasone treat- ment (Sainz et al., 1995).
As free radicals have been suggested as a cause of Parkinson's disease (Gotz et al., 1990) and given that 6-OHDA induces apoptosis through free radi- cal production, we investigated if melatonin can prevent neuronal cell death produced by this neuro- toxin. The experimental model used in this study was the rat adrenal pheochromocytoma cell line PC 12. This cell line synthesizes dopamine and nor- epinephrine. The addition to the culture medium of nerve growth factor (NGF) interrupts cellular mul- tiplication and induces neurites similar to those of the sympathetic neurons in primary cell cultures; in this state they retain their capacity to synthesize dopamine (Greene and Tischler, 1976). These two properties together with neuronal apoptosis induced by 6-OHDA in these cells make the PC12 cell line a good model for the study of the Parkinson's dis- ease. In vivo experimental models for neurode- generative diseases are technically difficult to perform due to the long duration and slow devel- opment of these diseases. Also, in these models, apoptosis is difficult to demonstrate because of the small number of cells that die at any one time point and the normal appearing condition of surrounding tissue; this further justifies the use of in vitro mod- els in our study.
In the present work we induced apoptosis in
180
PC12 cells by the addition of 6-OHDA to the cul- ture medium. We demonstrate that melatonin pre- vents programmed cell death induced by these neurotoxin to naive and neuronal PC12 cells,
Materials and methods
Materials
PC 12 cells (ATCC ## I72 1 -CRL) were purchased from American type Culture Collection (ATCC). NGF (mNGF 7s) was purchased from Alomone Labs, Ltd. (Israel). Horse serum, fetal bovine serum, and rat-tail collagen were obtained from GIBCO BRL (Life Technologies, Spain). Terminal trans- ferase and the biotinilated d-UTP were purchased from Boehringer Mannheim S.A. (Spain). [aP32]d- CTP (3,000 Ci/mmol) was obtained from Amersham Iberica S.A. (Spain). All other reagents were from Sigma Chemical Co. (Spain). 6-OHDA was dis- solved in sterile distilled water to avoid early oxi- dation and diluted from a stock 100-fold to the desired concentration directly in the culture medium. Melatonin was dissolved in a small amount of etha- nol and diluted from a stock 100-fold to the desired concentration directly in the culture medium.
Experimental procedures
Cell cultures. Naive (undifferentiated and therefore proliferative) PC 12 cells between passages 20 and 40 were cultured on dishes previously coated with 0.1 mg/ml rat-tail collagen in RPMI 1640 medium supplemented with 10% heat-inactivated horse se- rum, 5% fetal bovine serum, 2 mM L-glutamine, 100 units/ml penicillin, 100 pdml streptomycin, and 0.25 pg/ml amphotericin (Greene and Tischler, 1976). Neuronal (differentiated and nonpro- liferative) PC 12 cells were obtained by treatment with NGF at a concentration of 100 ng/ml during 14 days in RPMI 1640 medium supplemented with 1 % heat-inactivated horse serum. The medium con- taining NGF was changed every 2 days. Cell viability determination. The M'TT reduction assay de- scribed by Hansen et al, (1 989) was followed. Briefly, 45,000 cells per well were seeded in 96-multiwell plates (Falcon Becton Dickinson, NJ, USA). Cells were left to attach overnight and after 3 hr preincubation with melatonin, 6-OHDA was added; 24 hr later, MTT from a stock solution (5 mdml in RPMI-1640) at a fmal concentration of 1 mdml was added. Plates were incubated 3 hr at 37°C. Lysing buffer (20% SDS, 50% N,N-dimethylformamide, pH 4.7) was added and cells were incubated overnight at 37°C. Samples were read on an ELISA reader at a wavelength of 570 nm. Val- ues from blank dishes containing only medium were

Melatonin reduces 6-OHDA neuronal apoptosis
subtracted from the values of the samples. Standard curves were built using serial two-fold dilutions of freshly plated cells. Viability was expressed as the per- centage of the controls.
For trypan blue exclusion assay, 1 ml of trypan blue solution (final concentration of 0.1 %) was added to each plate after removal of the medium. The dye was aspirated after 3 min and cells washed with saline. Cells excluding fsypan blue were counted in a hemocy- tometer. Viability was expressed as the percentage of the controls. Morphornetrical studies. A) For the morphometrical analysis, the naive PC12 cells were seeded at a den- sity of lo6 cells/ml and collected after a brief incu- bation with PBS-EDTA and scrapping of the dishes. After centrifugation at 500g, cells were fixed over- night in 3% glutaraldehyde in phosphate buffer 0.1 M, pH 7.4. Then, they were postfixed for 10 rnin at 4°C in Os04, dehydrated in acetone, and embedded in Spurr resin. Several semithin sections of 1 pm separated at least 20 ym were stained with 0.1% toluidine blue and observed under a Nikon Labophot optical microscope. Cells with apoptotic morphol- ogy (basophilia and perinuclear condensation of the chromatin) were counted at a final magnification of lOOx, in a minimal area of 2 mm2 for each group.
B) The detached neuronal PC12 cells on the su- pernatant were pelleted by centrifugation at 500g for 5 min, resuspended in a small volume of RPMI 1640 medium, and fixed in an Eppendorf tube with an equal volume of 4% paraformaldehyde. An equal volume of acridine orange (AO) solution ( I 0 yg/ml in phosphate-buffered saline) was added and a sample was mounted on a slide for fluorescence mi- croscope observation (Orthoplan Leitz). Cells brightly stained with condensed chromatin were considered apoptotic and counted. The results are presented as the average of two independent experiments. DNA nick end labeling of cells (TUNEL). This labeling is indicative of the number of 3'OH ends present in the nucleus. A terminal deoxynucleotidyl transferase (TdT)-mediated deoxyuridine triphosphate incorpo- ration to the sites of the DNA breaks and amplified by avidin-peroxidase was used (ABC, Vectostain). Given that apoptotic cells undergo a higher rate of DNA fragmentation than normal cells, they display a strong staining of their nuclei while the nuclei of the remainder of the cells are hardly recognizable. Cells were fixed overnight in phosphate-buffered 4% paraformaldehyde at 4"C, dehydrated in acetone and embedded in methacrylate (HistoresinTM, Reitcher-Jung, Germany). Sections of 3 pm were treated as described by Gavrieli et al. (1992). DNA gel electrophoresis. Detached cells were used for DNA electrophoresis. DNA was extracted follow- ing the method described by Colotta et al. (1992).
Cells were centrifuged at 500g for 5 rnin at 4°C. The pellet obtained was incubated in 400 p1 of the lysis buffer (0.2% Triton X-100, 10 mM Tris, and 1 mM EDTA) pH 8.0 for 20 min on ice and centrifuged for 20 rnin at 13,OOOg at 4°C. The supernatant, con- taining the low molecular weight DNA, was sepa- rated and incubated with RNase DNase-free (20 pgl ml) for 1 hr at 37°C. Low molecular weight DNA was extracted with phenol-chloroform and precipi- tated with 2.5 vol ethanol and 0.1 vol sodium ac- etate (3 M, pH 5.2) at -20°C overnight. After centrifugation at 13,OOOg to recover DNA, the pel- let was washed with ethanol (70%) and resuspended in TE (pH 8); low molecular weight DNA was elec- trophoresed in a 2% agarose gel containing 0.5 pg/ ml ethidium bromide at 4V/cm. DNA ladder was observed by UV light and photographed with a Polaroid camera. Quantification of DNA fragmentation. High and low mo- lecular weight DNA were separated as described above. High molecular weight DNA (pellet) was re- suspended in 400 p1 of the same lysis buffer and precipitated with 12.5% trichloroacetic acid (TCA). The supernatant (containing the low molecular weight DNA) was precipitated with 12.5% TCA for 18 hr. Both high and low molecular weight DNA were centrifuged at 13,OOOg for 5 min at 4"C, ex- tracted from the precipitates with 30 pl 1 M perchlo- ric acid and 30 p15 mM NaOH at 70°C for 20 min, and quantified accord Ing to the method described by Burton (1956) in an LLISA reader at 600 nm wave- length. The percentage of DNA fragmentation was calculated as follows: [(OD600 of supematant)/(OD 600 pellet + OD 600 supernatant)] x 100. RNA isolation and Northern analysis. Total RNA was ex- tracted according to Chomczynski and Sacchi ( I 987). Northern blot analysis was accomplished by standard procedures previously described (Menendez-Pelaez et al., 1991). After electro- phoresis in a 1% agarose gel, samples were trans- ferred to nylon filters (Amersham Life Sciences, Spain). Hybridization was carried out at 42°C overnight in a solution containing 50% for- mamide. The following probes were used for the DNA/RNA hybridization: a 1.4 kb EcoRI frag- ment from rat manganese-containing superoxide dismutase (Mn-SOD) cDNA clone, pSP65 (Ho and Crapo, 1987a); a 0.6 kb EcoRI fragment from rat copper-zinc-containing superoxide dismutase (Cu-Zn-SOD) cDNA clone, pUC13 (Ho and Crapo, 1987b); a 0.8 kb SalI fragment from the rat glutathione peroxidase (GSH-Px) cDNA clone, LK440 (Yoshimura et al., 1988); a 1.6 kb Hind III/ Eco RI fragment from the rat catalase cDNA clone, pTZCTL (Furuta et al., 1986); and a 2.1 kb BamHI fragment from human p-actin cDNA
181
Mayo et al.
20
clone, pHFBA-1 (Gunning et al., 1983) (used to normalize the rest of the mRNA values). Signals were quantified by densitometric scanning of the au- toradiograph y. Statistical analysis. Three plates per group were used in each experiment and the results shown are the average of three independent experiments unless otherwise indicated. Data are presented as the means f SEM. Significance was tested by one-way analy- sis of variance (ANOVA) followed by a Student Newman-Keuls multiple range test.
- -.A-. Naive + MEL 10-7
t
Results
Melatonin promotes survival of PG12 cells after treatment with 6-OHDA
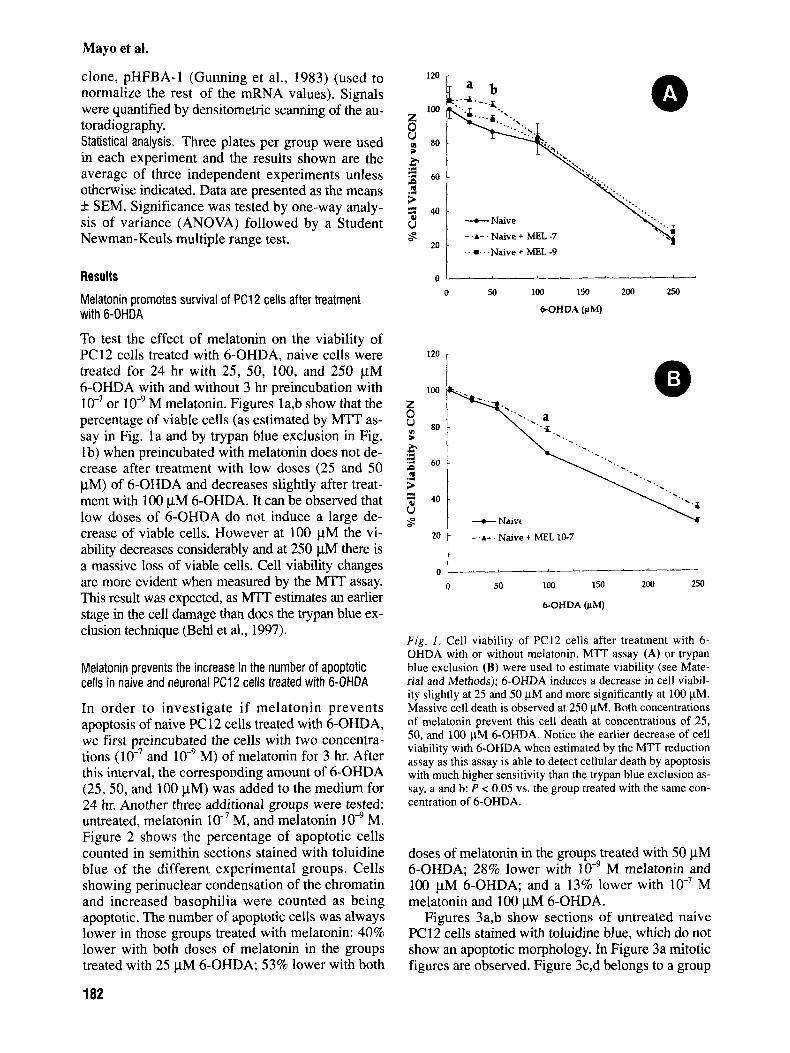
To test the effect of melatonin on the viability of PC12 cells treated with 6-OHDA, naive cells were treated for 24 hr with 25, 50, 100, and 250 pM 6-OHDA with and without 3 hr preincubation with
M melatonin. Figures la,b show that the percentage of viable cells (as estimated by MTT as- say in Fig. l a and by trypan blue exclusion in Fig. lb) when preincubated with melatonin does not de- crease after treatment with low doses (25 and 50 pM) of 6-OHDA and decreases slightly after treat- ment with 100 pM 6-OHDA. It can be observed that low doses of 6-OHDA do not induce a large de- crease of viable cells. However at 100 pM the vi- ability decreases considerably and at 250 pM there is a massive loss of viable cells. Cell viability changes are more evident when measured by the M?T assay. This result was expected, as MTT estimates an earlier stage in the cell damage than does the trypan blue ex- clusion technique (Behl et al., 1997).
or
Melatonin prevents the increase in the number of apoptotic cells in naive and neuronal PC12 cells treated with 6-OHDA
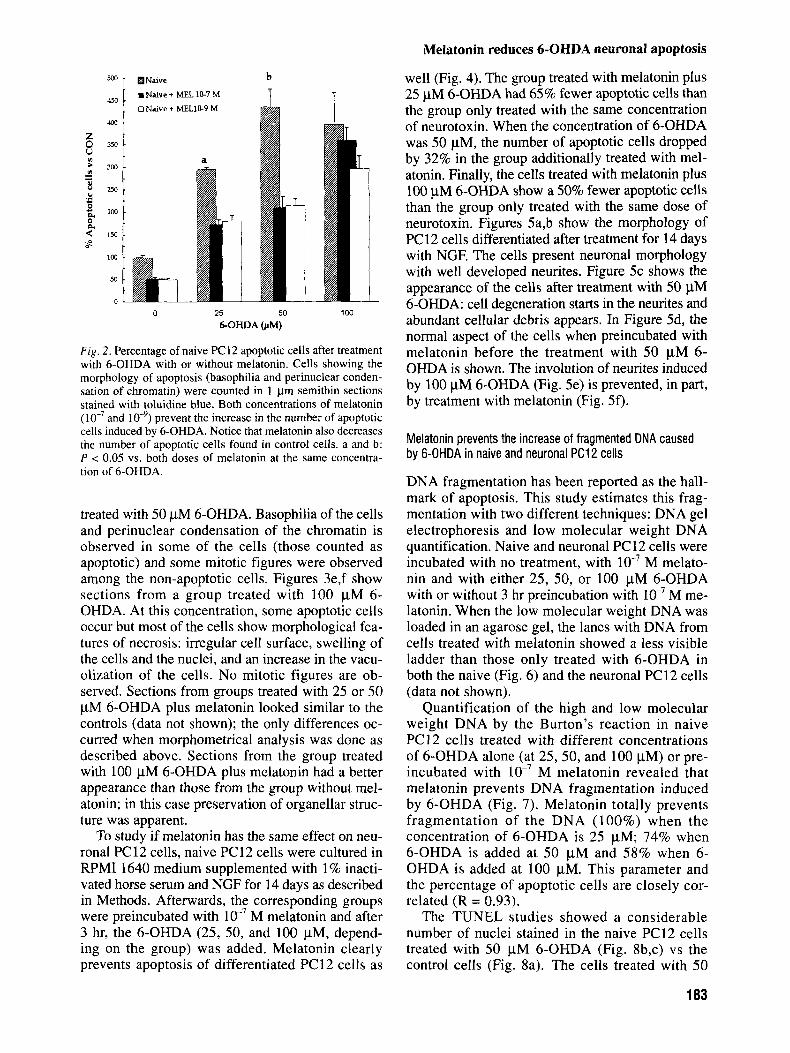
In order to investigate if melatonin prevents apoptosis of naive PC12 cells treated with 6-OHDA, we first preincubated the cells with two concentra- tions (lo-’ and M) of melatonin for 3 hr. After this interval, the corresponding amount of 6-OHDA (25,50, and 100 pM) was added to the medium for 24 hr. Another three additional groups were tested: untreated, melatonin M, and melatonin lo-’ M. Figure 2 shows the percentage of apoptotic cells counted in semithin sections stained with toluidine blue of the different experimental groups. Cells showing perinuclear condensation of the chromatin and increased basophilia were counted as being apoptotic. The number of apoptotic cells was always lower in those groups treated with melatonin: 40% lower with both doses of melatonin in the groups treated with 25 pM 6-OHDA; 53% lower with both
120 r
100 oz 2 80
3 40 U I
20
0
--c Naive - .A- Naive + mL -7 .. .m.. .Naive + MF,L -9
0 50 100 150 200 250
W H D A (JIM)
120 r
100 z 8 v)
LI - 40 s
F i g . 1. Cell viability of PC12 cells after treatment with 6- OHDA with or without melatonin. MTT assay (A) or trypan blue exclusion (B) were used to estimate viability (see Mate- rial and Methods); 6-OHDA induces a decrease in cell viabil- ity slightly at 25 and 50 FM and more significantly at 100 WM. Massive cell death is observed at 250 pM. Both concentrations of melatonin prevent this cell death at concentrations of 25, SO, and 100 FM 6-OHDA. Notice the earlier decrease of cell viability with 6-OHDA when estimated by the MTT reduction assay as this assay is able to detect cellular death by apoptosis with much higher sensitivity than the trypan blue exclusion as- say. a and b: P < 0.05 vs. the group treated with the same con- centration of 6-OHDA.
doses of melatonin in the groups treated with 50 pM 6-OHDA; 28% lower with M melatonin and 100 pM 6-OHDA; and a 13% lower with lo-’ M melatonin and 100 pM 6-OHDA.
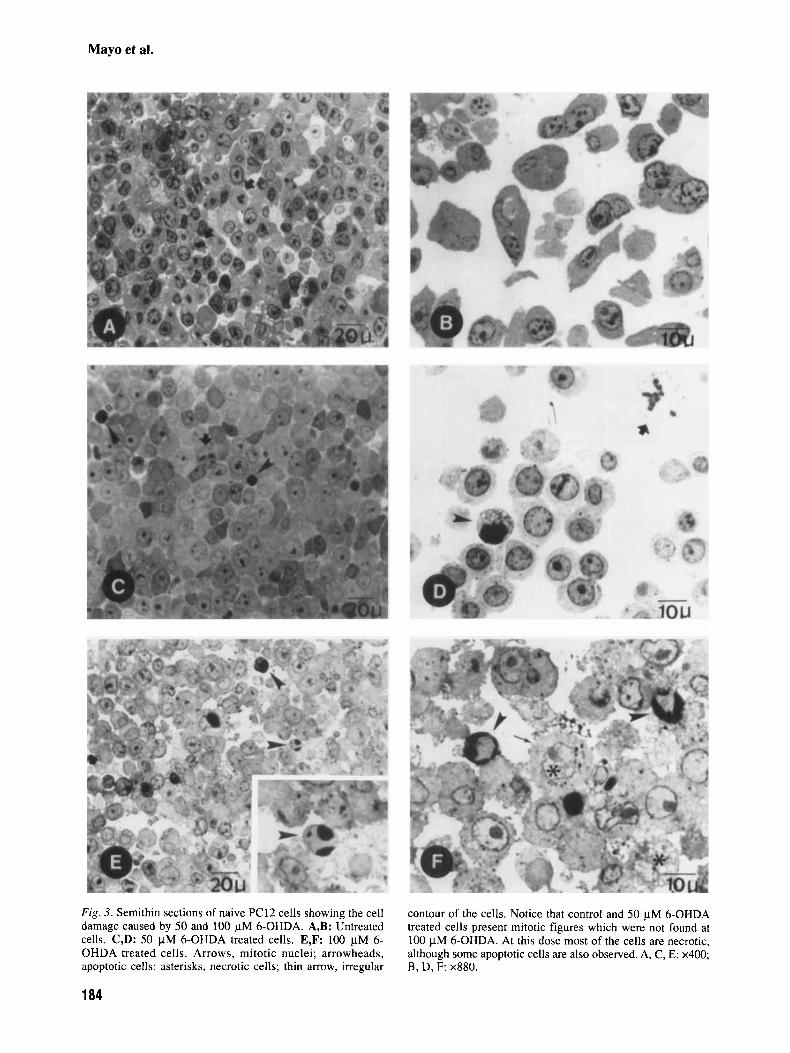
Figures 3a,b show sections of untreated naive PC12 cells stained with toluidine blue, which do not show an apoptotic morphology. In Figure 3a mitotic figures are observed. Figure 3c,d belongs to a group
182
Melatonin reduces 6-OHDA neuronal apoptosis
Naive
m Naive + MEL 1C-7 M 0 Naive t MELlC-9 M
a
0 25
b
50 100 6-OHDA (JIM)
Fig . 2 . Percentage of naive PC12 apoptotic cells after treatment with 6-OHDA with or without melatonin. Cells showing the morphology of apoptosis (basophilia and perinuclear conden- sation of chromatin) were counted in 1 pm semithin sections stained with toluidine blue. Both concentrations of melatonin
prevent the increase in the number of apoptotic cells induced by 6-OHDA. Notice that melatonin also decreases the number of apoptotic cells found in control cells. a and b: P < 0.05 vs. both doses of melatonin at the same concentra- tion of 6-OHDA.
and
treated with 50 pM 6-OHDA. Basophilia of the cells and perinuclear condensation of the chromatin is observed in some of the cells (those counted as apoptotic) and some mitotic figures were observed among the non-apoptotic cells. Figures 3e,f show sections from a group treated with 100 pM 6- OHDA. At this concentration, some apoptotic cells occur but most of the cells show morphological fea- tures of necrosis: irregular cell surface, swelling of the cells and the nuclei, and an increase in the vacu- olization of the cells. No mitotic figures are ob- served. Sections from groups treated with 25 or 50 pM 6-OHDA plus melatonin looked similar to the controls (data not shown); the only differences oc- curred when morphometrical analysis was done as described above. Sections from the group treated with 100 pM 6-OHDA plus melatonin had a better appearance than those from the group without mel- atonin; in this case preservation of organellar struc- ture was apparent.
To study if melatonin has the same effect on neu- ronal PC12 cells, naive PC12 cells were cultured in RPMI 1640 medium supplemented with 1% inacti- vated horse serum and NGF for 14 days as described in Methods. Afterwards, the corresponding groups were preincubated with M melatonin and after 3 hr, the 6-OHDA (25, 50, and 100 pM, depend- ing on the group) was added. Melatonin clearly prevents apoptosis of differentiated PC12 cells as
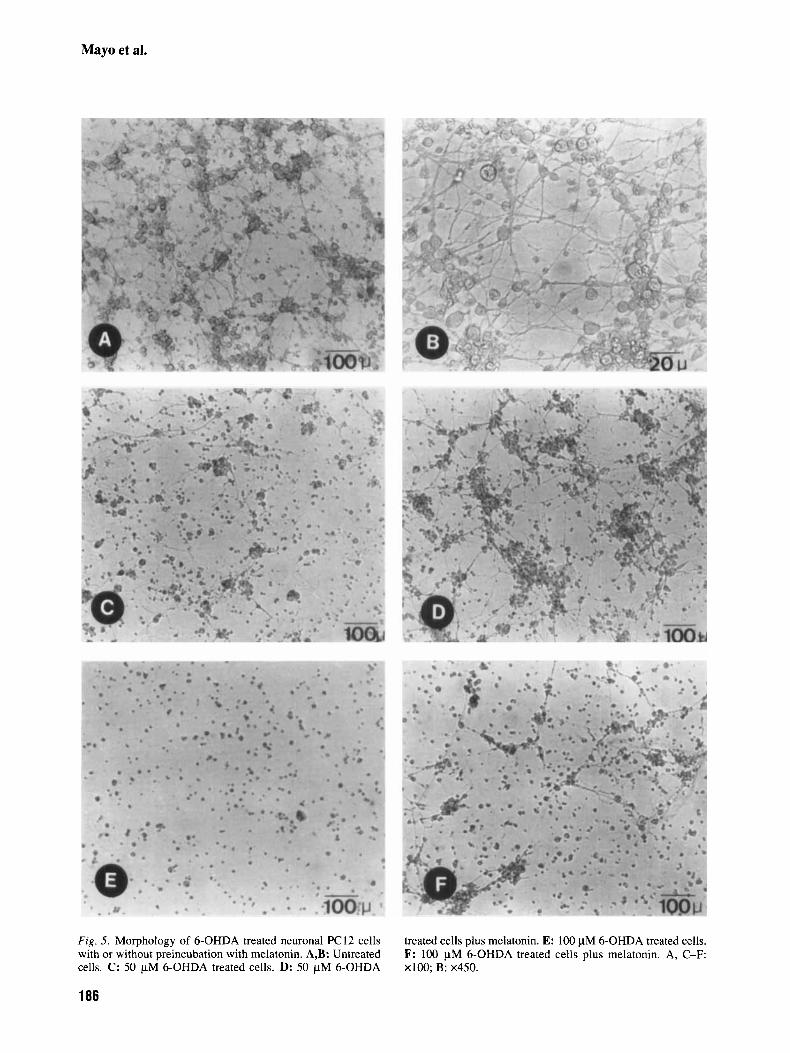
well (Fig. 4). The group treated with melatonin plus 25 pM 6-OHDA had 65% fewer apoptotic cells than the group only treated with the same concentration of neurotoxin. When the concentration of 6-OHDA was 50 pM, the number of apoptotic cells dropped by 32% in the group additionally treated with mel- atonin. Finally, the cells treated with melatonin plus 100 pM 6-OHDA show a 50% fewer apoptotic cells than the group only treated with the same dose of neurotoxin. Figures 5a,b show the morphology of PC12 cells differentiated after treatment for 14 days with NGF. The cells present neuronal morphology with well developed neurites. Figure 5c shows the appearance of the cells after treatment with 50 yM 6-OHDA: cell degeneration starts in the neurites and abundant cellular debris appears. In Figure 5d, the normal aspect of the cells when preincubated with melatonin before the treatment with 50 yM 6- OHDA is shown. The involution of neurites induced by 100 pM 6-OHDA (Fig. 5e) is prevented, in part, by treatment with melatonin (Fig. 5 0 .
Melatonin prevents the increase of fragmented DNA caused by 6-OHDA in naive and neuronal PC12 cells
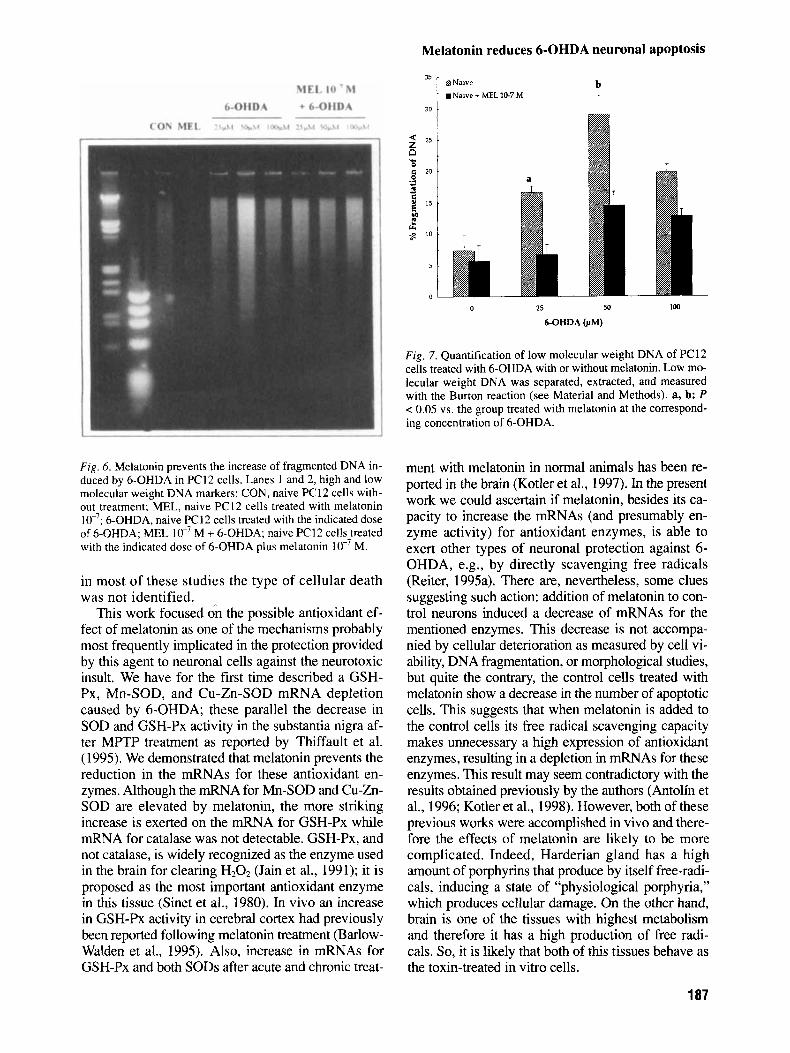
DNA fragmentation has been reported as the hall- mark of apoptosis. This study estimates this frag- mentation with two different techniques: DNA gel electrophoresis and low molecular weight DNA quantification. Naive and neuronal PC12 cells were incubated with no treatment, with M melato- nin and with either 25, 50, or 100 pM 6-OHDA with or without 3 hr preincubation with lo-’ M me- latonin. When the low molecular weight DNA was loaded in an agarose gel, the lanes with DNA from cells treated with melatonin showed a less visible ladder than those only treated with 6-OHDA in both the naive (Fig. 6) and the neuronal PC12 cells (data not shown).
Quantification of the high and low molecular weight DNA by the Burton’s reaction in naive PC 12 cells treated with different concentrations of 6-OHDA alone (at 25,50, and 100 pM) or pre- incubated with M melatonin revealed that melatonin prevents DNA fragmentation induced by 6-OHDA (Fig. 7). Melatonin totally prevents fragmentation of the DNA (100%) when the concentration of 6-OHDA is 25 pM; 74% when 6-OHDA is added at 50 pM and 58% when 6- OHDA is added at 100 pM. This parameter and the percentage of apoptotic cells are closely cor- related (R = 0.93).
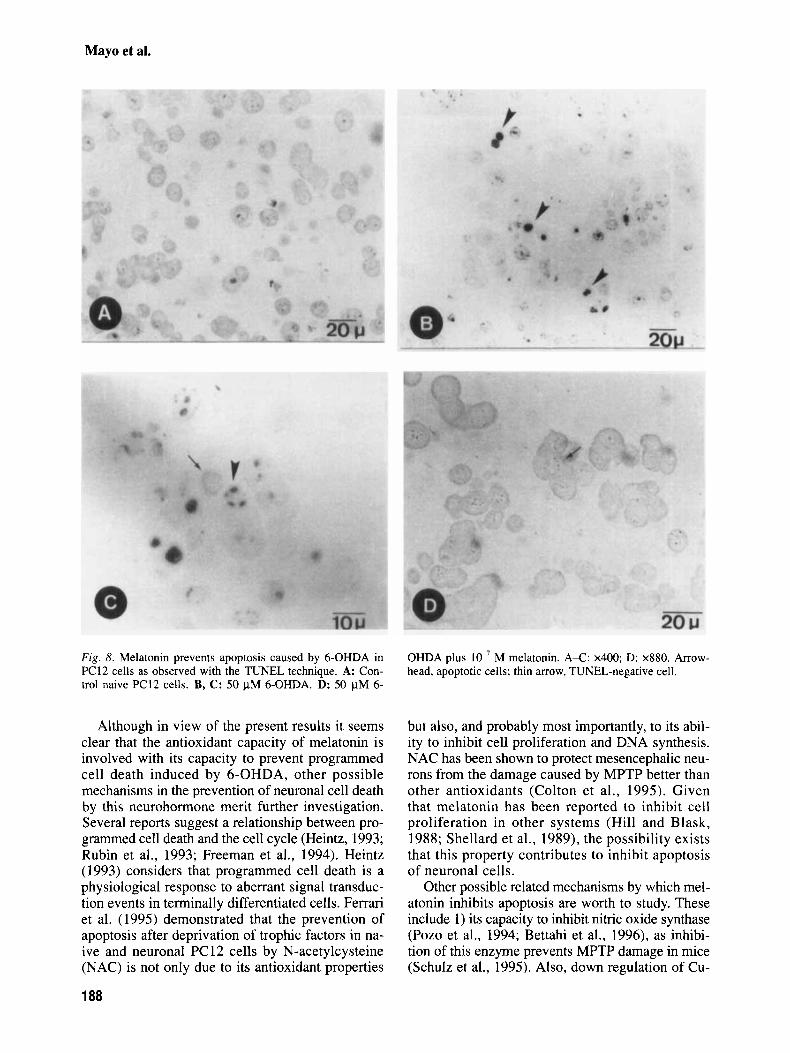
The TUNEL studies showed a considerable number of nuclei stained in the naive PC12 cells treated with 50 pM 6-OHDA (Fig. 8b,c) vs the control cells (Fig. 8a). The cells treated with 50
183
Mayo et al.
Fig. 3. Semithin sections of naive PC12 cells showing the cell damage caused by 50 and 100 pM 6-OHDA. A,B: Untreated cells. C,D: 50 pM 6-OHDA treated cells. E,F: 100 pM 6- OHDA treated cells. Arrows, mitotic nuclei; arrowheads, apoptotic cells; asterisks, necrotic cells; thin arrow, irregular
contour of the cells. Notice that control and 50 pM 6-OHDA treated cells present mitotic figures which were not found at 100 pM 6-OHDA. At this dose most of the cells are necrotic, although some apoptotic cells are also observed. A, C, E: x400; B, D, F: x880.
184
Melatonin reduces 6-OHDA neuronal apoptosis
C Discussion
50
2 40 - 8 B
P 30
ZL
d g 20
10
0
0 25 M 100 COHDA (pM)
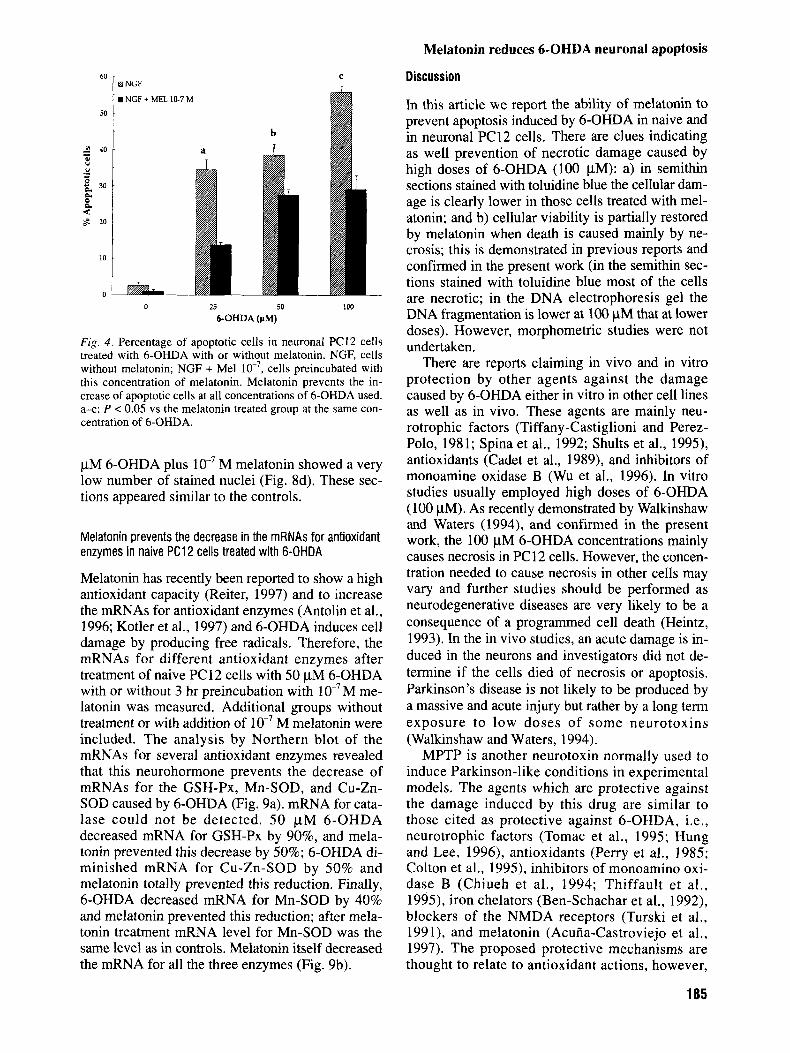
Fig. 4. Percentage of apoptotic cells in neuronal PC12 cells treated with 6-OHDA with or without melatonin. NGF, cells without melatonin; NGF + Me1 cells preincubated with this concentration of melatonin. Melatonin prevents the in- crease of apoptotic cells at all concentrations of 6-OHDA used. a-c: P < 0.05 vs the melatonin treated group at the same con- centration of 6-OHDA.
pM 6-OHDA plus M melatonin showed a very low number of stained nuclei (Fig. 8d). These sec- tions appeared similar to the controls.
Melatonin prevents the decrease in the mRNAs for antioxidant enzymes in naive PC12 cells treated with 6-OHDA
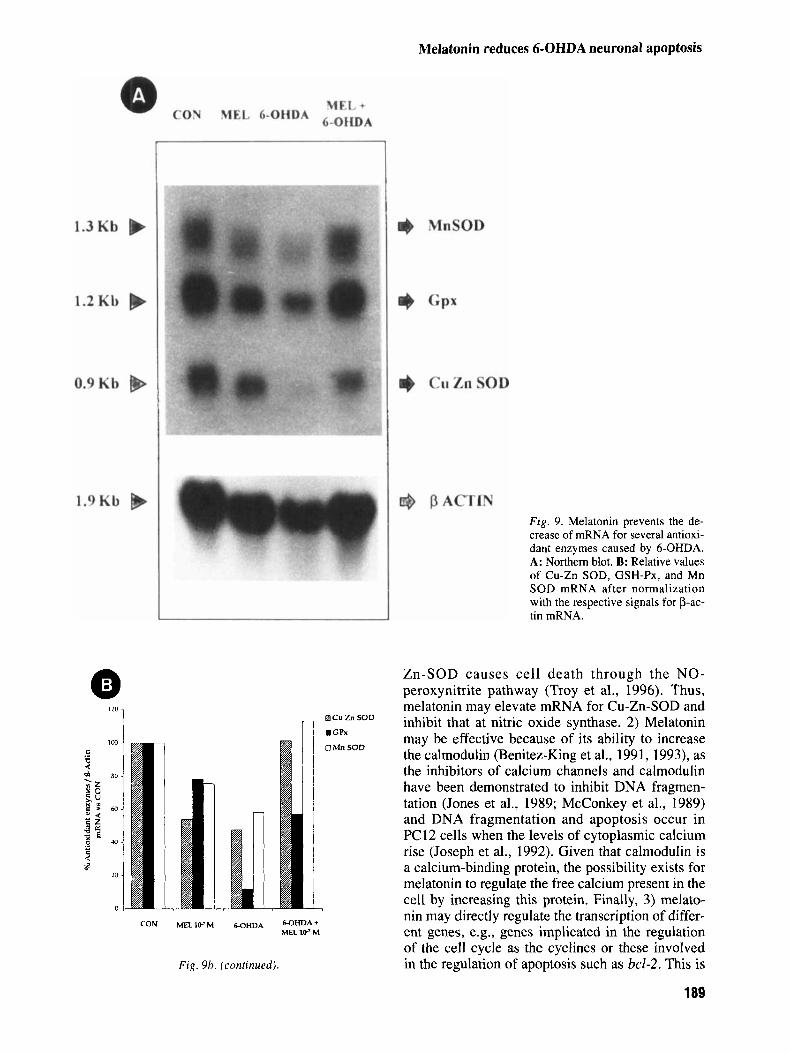
Melatonin has recently been reported to show a high antioxidant capacity (Reiter, 1997) and to increase the mRNAs for antioxidant enzymes (Antolin et al., 1996; Kotler et al., 1997) and 6-OHDA induces cell damage by producing free radicals. Therefore, the mRNAs for different antioxidant enzymes after treatment of naive PC12 cells with 50 pM 6-OHDA with or without 3 hr preincubation with M me- latonin was measured. Additional groups without treatment or with addition of M melatonin were included. The analysis by Northern blot of the mRNAs for several antioxidant enzymes revealed that this neurohormone prevents the decrease of mRNAs for the GSH-Px, Mn-SOD, and Cu-Zn- SOD caused by 6-OHDA (Fig. 9a). mRNA for cata- lase could not be detected. 50 p M 6-OHDA decreased mRNA for GSH-Px by 90%, and mela- tonin prevented this decrease by 50%; 6-OHDA di- minished mRNA for Cu-Zn-SOD by 50% and melatonin totally prevented this reduction. Finally, 6-OHDA decreased mRNA for Mn-SOD by 40% and melatonin prevented this reduction; after mela- tonin treatment mRNA level for Mn-SOD was the same level as in controls. Melatonin itself decreased the mRNA for all the three enzymes (Fig. 9b).
In this article we report the ability of melatonin to prevent apoptosis induced by 6-OHDA in naive and in neuronal PC12 cells. There are clues indicating as well prevention of necrotic damage caused by high doses of 6-OHDA (100 pM): a) in semithin sections stained with toluidine blue the cellular dam- age is clearly lower in those cells treated with mel- atonin; and b) cellular viability is partially restored by melatonin when death is caused mainly by ne- crosis; this is demonstrated in previous reports and confirmed in the present work (in the semithin sec- tions stained with toluidine blue most of the cells are necrotic; in the DNA electrophoresis gel the DNA fragmentation is lower at 100 pM that at lower doses). However, morphometric studies were not undertaken.
There are reports claiming in vivo and in vitro protection by other agents against the damage caused by 6-OHDA either in vitro in other cell lines as well as in vivo. These agents are mainly neu- rotrophic factors (Tiffany-Castiglioni and Perez- Polo, 1981; Spina et al., 1992; Shults et al., 1995), antioxidants (Cadet et al., 1989), and inhibitors of monoamine oxidase B (Wu et al., 1996). In vitro studies usually employed high doses of 6-OHDA (100 pM). As recently demonstrated by Walkinshaw and Waters (1994), and confirmed in the present work, the 100 pM 6-OHDA concentrations mainly causes necrosis in PC12 cells. However, the concen- tration needed to cause necrosis in other cells may vary and further studies should be performed as neurodegenerative diseases are very likely to be a consequence of a programmed cell death (Heintz, 1993). In the in vivo studies, an acute damage is in- duced in the neurons and investigators did not de- termine if the cells died of necrosis or apoptosis. Parkinson’s disease is not likely to be produced by a massive and acute injury but rather by a long term exposure to low doses of some neurotoxins (Walkinshaw and Waters, 1994).
MPTP is another neurotoxin normally used to induce Parkinson-like conditions in experimental models. The agents which are protective against the damage induced by this drug are similar to those cited as protective against 6-OHDA, i.e., neurotrophic factors (Tomac et al., 1995; Hung and Lee, 1996), antioxidants (Perry et al., 1985; Colton et al., 1995), inhibitors of monoamino oxi- dase B (Chiueh et al., 1994; Thiffault et al., 1995), iron chelators (Ben-Schachar et al., 1992), blockers of the NMDA receptors (Turski et al., 1991), and melatonin (Acuiia-Castroviejo et al., 1997). The proposed protective mechanisms are thought to relate to antioxidant actions, however,
185
Mayo et al.
Fig. 5. Morphology of 6-OHDA treated neuronal PC12 cells with or without preincubation with melatonin. A,B: Untreated cells. C: 50 pM 6-OHDA treated cells. D: 50 pM 6-OHDA
treated cells plus melatonin. E: 100 pM 6-OHDA treated cells. F: 100 pM 6-OHDA treated cells plus melatonin. A, C-F: ~ 1 0 0 ; B: x450.
186
Melatonin reduces 6-OHDA neuronal apoptosis
a Naive b Naive + MEL 1&7 M
Fig. 6. Melatonin prevents the increase of fragmented DNA in- duced by 6-OHDA in PC12 cells. Lanes 1 and 2, high and low molecular weight DNA markers; CON, naive PC12 cells with- out treatment; MEL, naive PC12 cells treated with melatonin lo-’; 6-OHDA, naive PC12 cells treated with the indicated dose of 6-OHDA; MEL lo-’ M + 6-OHDA; naive PC12 cells treated with the indicated dose of 6-OHDA plus melatonin lo-’ M.
in most of these studies the type of cellular death was not identified.
This work focused on the possible antioxidant ef- fect of melatonin as one of the mechanisms probably most frequently implicated in the protection provided by this agent to neuronal cells against the neurotoxic insult. We have for the first time described a GSH- Px, Mn-SOD, and Cu-Zn-SOD mRNA depletion caused by 6-OHDA; these parallel the decrease in SOD and GSH-Px activity in the substantia nigra af- ter MPTP treatment as reported by Thiffault et al. (1995). We demonstrated that melatonin prevents the reduction in the mRNAs for these antioxidant en- zymes. Although the mRNA for Mn-SOD and Cu-Zn- SOD are elevated by melatonin, the more striking increase is exerted on the mRNA for GSH-Px while mRNA for catalase was not detectable. GSH-Px, and not catalase, is widely recognized as the enzyme used in the brain for clearing H202 (Jain et al., 1991); it is proposed as the most important antioxidant enzyme in this tissue (Sinet et al., 1980). In vivo an increase in GSH-Px activity in cerebral cortex had previously been reported following melatonin treatment (Barlow- Walden et al., 1995). Also, increase in mRNAs for GSH-Px and both SODS after acute and chronic treat-
0 25 50 1w COHDA (pM)
Fig. 7. Quantification of low molecular weight DNA of PCl2 cells treated with 6-OHDA with or without melatonin. Low mo- lecular weight DNA was separated, extracted, and measured with the Burton reaction (see Material and Methods). a, b: P < 0.05 vs. the group treated with melatonin at the correspond- ing concentration of 6-OHDA.
ment with melatonin in normal animals has been re- ported in the brain (Kotler et al., 1997). In the present work we could ascertain if melatonin, besides its ca- pacity to increase the mRNAs (and presumably en- zyme activity) for antioxidant enzymes, is able to exert other types of neuronal protection against 6- OHDA, e.g., by directly scavenging free radicals (Reiter, 1995a). There are, nevertheless, some clues suggesting such action: addition of melatonin to con- trol neurons induced a decrease of mRNAs for the mentioned enzymes. This decrease is not accompa- nied by cellular deterioration as measured by cell vi- ability, DNA fragmentation, or morphological studies, but quite the contrary, the control cells treated with melatonin show a decrease in the number of apoptotic cells. This suggests that when melatonin is added to the control cells its free radical scavenging capacity makes unnecessary a high expression of antioxidant enzymes, resulting in a depletion in mRNAs for these enzymes. This result may seem contradictory with the results obtained previously by the authors (Antolin et al., 1996; Kotler et al., 1998). However, both of these previous works were accomplished in vivo and there- fore the effects of melatonin are likely to be more complicated. Indeed, Harderian gland has a high amount of porphyrins that produce by itself free-radi- cals, inducing a state of “physiological porphyria,” which produces cellular damage. On the other hand, brain is one of the tissues with highest metabolism and therefore it has a high production of free radi- cals. So, it is likely that both of this tissues behave as the toxin-treated in vitro cells.
187
Mayo et al.
Fig. 8. Melatonin prevents apoptosis caused by 6-OHDA in PC12 cells as observed with the TUNEL technique. A: Con- trol naive PC12 cells. B, C: 50 pM 6-OHDA. D: 50 FM 6-
OHDA plus lo-’ M melatonin. A-C: x400; D: x880. Arrow- head, apoptotic cells; thin arrow, TUNEL-negative cell.
Although in view of the present results it seems clear that the antioxidant capacity of melatonin is involved with its capacity to prevent programmed cell death induced by 6-OHDA, other possible mechanisms in the prevention of neuronal cell death by this neurohormone merit further investigation. Several reports suggest a relationship between pro- grammed cell death and the cell cycle (Heintz, 1993; Rubin et al., 1993; Freeman et al., 1994). Heintz (1993) considers that programmed cell death is a physiological response to aberrant signal transduc- tion events in terminally differentiated cells. Ferrari et al. (1995) demonstrated that the prevention of apoptosis after deprivation of trophic factors in na- ive and neuronal PC 12 cells by N-acetylcysteine (NAC) is not only due to its antioxidant properties
188
but also, and probably most importantly, to its abil- ity to inhibit cell proliferation and DNA synthesis. NAC has been shown to protect mesencephalic neu- rons from the damage caused by MPTP better than other antioxidants (Colton et al., 1995). Given that melatonin has been reported to inhibit cell proliferation in other systems (Hill and Blask, 1988; Shellard et al., 1989), the possibility exists that this property contributes to inhibit apoptosis of neuronal cells.
Other possible related mechanisms by which mel- atonin inhibits apoptosis are worth to study. These include 1) its capacity to inhibit nitric oxide synthase (Pozo et al., 1994; Bettahi et al., 1996), as inhibi- tion of this enzyme prevents MPTP damage in mice (Schulz et al., 1995). Also, down regulation of Cu-
Melatonin reduces 6-OHDA neuronal apoptosis
CON
Ei Cu Zn SOD n .GPx
M R I V M WHDA MHD*+ MR. 1 c M
Fig. 9b. (continued).
F ig . 9. Melatonin prevents the de- crease of mRNA for several antioxi- dant enzymes caused by 6-OHDA. A: Northern blot. B: Relative values of Cu-Zn SOD, GSH-Px, and Mn SOD mRNA after normalization with the respective signals for p-ac- tin mRNA.
Zn-SOD causes cell death through the NO- peroxynitrite pathway (Troy et al., 1996). Thus, melatonin may elevate mRNA for Cu-Zn-SOD and inhibit that at nitric oxide synthase. 2) Melatonin may be effective because of its ability to increase the calmodulin (Benitez-King et al., 1991, 1993), as the inhibitors of calcium channels and calmodulin have been demonstrated to inhibit DNA fragmen- tation (Jones et al., 1989; McConkey et al., 1989) and DNA fragmentation and apoptosis occur in PC12 cells when the levels of cytoplasmic calcium rise (Joseph et al., 1992). Given that calmodulin is a calcium-binding protein, the possibility exists for melatonin to regulate the free calcium present in the cell by increasing this protein. Finally, 3) melato- nin may directly regulate the transcription of differ- ent genes, e.g., genes implicated in the regulation of the cell cycle as the cyclines or these involved in the regulation of apoptosis such as bcl-2. This is
189

Mayo et al.
possible since melatonin is able to activate the nuclear receptor RZRp, which belongs to the nuclear hormone receptor family (Carlberg and Wiesenberg, 1995; Wiesenberg et al., 1995).
The brain is the most oxygen demanding tissue and therefore produces high levels of free radicals (Sokoloff, 1960); as a result, this tissue is likely to suffer damage from these agents (see Reiter, 1995a for review). The mentioned neuronal protectors act in different compartments of the cell, and all but the neurotrophic factors are exogenous. Melatonin is a potent endogenous antioxidant that is both lipid (Costa et al., 1995) and water soluble (Shida et al., 1994), able to act in all the compartments of the cell, giving protection to DNA (Tan et al., 1994), possi- bly to proteins (Abe et al., 1994) and to lipids (Melchiorri et al., 1995). In the present work we demonstrated that it provides protection to neuronal cells against 6-OHDA and it inhibits apoptosis caused by low doses of this neurotoxin. The mecha- nism by melatonin prevents programmed cell death caused by 6-OHDA is likely via antioxidant capac- ity. Considering that melatonin reduces neural dam- age induced by MPTP (Acuiia-Castroviejo et al., 1997), and given that cell death caused by low doses of 6-OHDA has been proposed as a model for Parkinson’s disease (Walkinshaw and Waters, 1994), the fact that melatonin truly prevents programmed cell death produced by this neurotoxin together with its negligible side effects opens the possibility of the use of this agent in the clinical setting against Parkinson’s disease. Additionally, its potential use in the treatment of other neurodegenerative diseases could also be considered since free radicals seem to be implicated in the origin of these diseases, as well as aging itself (Gotz et al., 1990; Jesberger and Richardson, 1991; Stadtman, 1992; Loo et al., 1993). In this context, melatonin has been recently shown to prevent neuronal death caused by a vari- ety of neurotoxins used as experimental models of neurodegenerative diseases (Lezoualc’h et al., 1996; Pappolla et al., 1997). Of additional interest is that melatonin, which possesses substantial antioxidant activity, decreases significantly with age (Reiter, 1992; Reiter et al., 1996) coincident with the in- crease in a variety of neurodegenerative conditions (Reiter, 1995b; Reiter et al., 1996).
Acknowledgments
This work was supported by grants from FICYT PB MAS 94- 12 (C.R.) and the University of Oviedo NP-97-18 (C.R.). H.U. thanks FICYT for his postgraduate fellowship. LA. was sup- ported by a postdoctoral fellowship from FICYT. We thank Dr. Y. S. Ho (Institute of Chemical Toxicology, Wayne State Uni- versity, Detroit, MI) for providing GSHP-x, Mn-SOD, and Cu- Zn SOD cDNA and Dr. T. Osumi (Laboratory of Cell and
Molecular Biology, Department of Life Science, Hyogo, Japan) for catalase cDNA.
References
ABE, M., R.J. REITER, P.H. ORHII, M. HARA, B. POEGGELER, L.C. MANCHESTER, L. BARLOW-WALDEN (1994) Inhibitory effect of melatonin on cataract formation in newborn rats: Evidence of an antioxidative role for melatonin. J. Pineal Res. 17:94-100.
AcuP~A-CASTROVIEIO, D., A. COTO-MONTES, G. MONTI, G.G. ORTIZ, R.J. REITER (1997) Melatonin is protective against MPTP-induced striatal and hippocampal lesions. Life Sci.
ANTOLIN, I., C. RODRIGUEZ, R.M. SAINZ, J.C. MAYO, H. UR~A, M. KOTLER, M.J. RODR~GUEZ-COLUNGA, D. TOLIVIA, A. MENENDEZ-PELAEZ (1 996) Neurohormone melatonin prevents cell damage: Effect on gene expression for antioxidant en- zymes. FASEB J. 105382-890.
BARLOW-WALDEN, L., R.J. REITER, M. ABE, M.I. PABLOS, A. MENENDEZ-PELAEZ, L.D. CHEN, B. POEGGELER (1995) Melato- nin stimulates brain glutathione peroxidase activity. Neurochem. Int. 26:497-502.
F. HOLSBOER (1 997) Glucocorticoids enhance oxidative stress- induced cell death in hipocampal neurons in vitro. Endocri- nology 138:lOl-106.
BEN-SACHAR, D., G. ESHEL, P. RIEDERER, M.B. YOUDIM (1992) Role of iron and iron chelation in dopaminergic-induced neurodegeneration: Implication for Parkinson’s disease. Ann. Neurol. 32:S105-110.
BENITEZ-KING, G., L. HUERTA DELGADILLO, F. ANTON TAY (1991) Melatonin modifies calmodulin levels in MDCK and N1E- 115 cell lines and inhibits phosphodiesterase activity in vitro. Brain Res. 557:289-292.
BENITEZ-KING, G., L. HUERTA DELGADILLO, F. ANTON TAY (1993) Binding of tritium melatonin to calmodulin. Life Sci.
BETTAHI, I., D. Pozo, C. OSUNA, R.J. REITER, D. ACURA- CASTROVIEJO, J.M. GUERRERO (1 996) Melatonin reduces nitric oxide synthase in rat hypothalamus. J. Pineal Res. 20:205-210.
BURTON, K. (1956) A study of the conditions and mechanism of the diphenylamine reaction for the colorimetric estimation of deoxyribonucleic acid. Biochem. J. 62:3 15-323.
CADET, J.L., M. KATZ, V. JAKSON-LEWIS, S. FAHN (1989) Vita- min E attenuates the toxic effects of intrastriatal injection of 6-hydroxydopamine (6-OHDA) in rats: behavioral and bio- chemical evidence. Brain Res. 476:1&15.
CARLBERG, C., I. WEISSENBERG (1995) The orphan receptor fam- ily RZR/ROR, melatonin and lipoxygenase: An unexpected relationship. J. Pineal Res. 18:121-128.
CHIUEH, C.C., S.J. HUANG, D.L. MURPHY (1994) Suppression of hydroxyl radical formation by M A 0 inhibitors: A novel possible neuroprotective mechanism in dopaminergic neuro- toxicity. J. Neural Transm. 41P: 189-196.
CHONCZYNSKI, P., N. SACCHI (1987) Single-step method of RNA isolation by a guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 162:156-159.
COHEN, G., R. HEILLILA (1974) The generation of hydrogen per- oxide, superoxide radical and hydroxyl radical by 6- hydroxydopamine, dialuric acid and related cytotoxic agents. J. Biol. Chem. 250:2447-2452.
COLOTTA, F., N. POLENTARUTTI, M. SIRONI, A. MANTOVANI (1992) Expression and involvement of c-fos and c-jun proto- oncogenes in programmed cell death induced by growth fac- tor deprivation in lymphoid cell lines. J. Biol. Chem.
60: 23-29.
BEHL, c., F. LEZOUALC’H, T. TRAPP, M. WIDMANN, T. SKUTELLA,
53 :20 1-207.
267: 18278-18283.
190

Melatonin reduces 6-OHDA neuronal apoptosis
JESBERBER, J.A., J.S. RICHARDSON (1991) Oxygen free radicals and brain dysfunction. Int J. Neurosci. 57: 1-17.
JONES, D.P., D.J. MCKONKEY, P. NICOTERA, S. ORRENIUS (1989) Calcium activated DNA fragmentation in rat liver nuclei. J. Biol. Chem. 264:6398-6403.
JOSEPH, R., W. LI, E. HAN (1992) Neuronal death, cytoplasmic calcium and internucleosomal DNA fragmentation: evidence for DNA fragments being released from cells. Mol. Brain Res.
KOTLER, M., C. RODRIGUEZ, R.M. SAINZ, I. ANTOLIN, A. MENENDEZ-PELAEZ (1998) Melatonin increases mRNA for an- tioxidant enzymes in brain cortex. J. Pineal Res. in press.
LEZOUALC’H, F. SKUTELLA, M. WIDMANN, C. BEHL (1996) Me- latonin prevents oxidative stress induced cell death in hippoc- ampal cells. NeuroReport 7:2071-2077.
Loo, D.T., A. COPANI, C.J. PIKE, E.R. WHITTEMORE, A.J. WALENCEWICZ, C.W. COTMAN (1993) Apoptosis is induced by P-amyloid in cultured central nervous system neurons. Proc. Natl. Acad. Sci. U.S.A. 90:795 1-7955.
MARTIN, J.B., M.F. BEAL (1992) Huntingtons disease and neu- rotoxins. Ann. N.Y. Acad. Sci. 648:169-175.
MCCONKEY, D.J., P. NICOTERA, P. HARTZELL, G. BELLOMO, A.H. WYLLIE, S. ORRENIUS (1989) Glucocorticoids activate a sui- cide process in thymocyte through an elevation of cytosolic Ca2+ concentration. Arch. Biochem. Biophys. 269:365-370.
MELCHORRI, D., R.J. REITER, A.M. ATIIA, M. HARA, G. NISTICO (1 995) Potent protective effect of melatonin on in vivo paraquat- induced oxidative damage in rats. Life Sci. 56:83-89.
MELDRUM, B., J. GARTHWAITE (1990) Excitatory amino acid neu- rotoxicity and neurodegenerative disease. Trends Pharmacol. Sci. 11:379-387.
MENENDEZ-PELAEZ, A., R.J. REITER (1993) Distribution of me- latonin in mammalian tissues: the relative importance of nuclear versus cytosolic localization. J. Pineal Res. 1559-69.
MENENDEZ-PELAEZ, A., C. RODRIGUEZ, P. DOMINGUEZ (1991) 5- aminolevulinate synthase mRNA levels in the Harderian gland of Syrian hamsters: Correlation with porphyrin concentrations and regulation by androgens and melatonin. Mol. Cell. Endocrinol. 80: 177-182.
MENENDEZ-PELAEZ, A,, B. POEGGELER, R.J. REITER, L. BARLOW- WALDEN, M. PABLOS, D.X. TAN (1993) Nuclear localization of melatonin in different mammalian tissues: Immunocy- tochemical and radioimmunoassay evidence. J. Cell. Biochem.
MICHEL, P.P., F. HEFTY (1990) Toxicity of 6-hydroxidopamine and dopamine for dopaminergic neurons in culture. J. Neurosci. Res. 26:428-435.
PAPPOLLA, M., M. Sos, R.A. OMAR, R.J. BICK, D.L.M. HICKSON- BICK, R.J. REITER, S. EFTHIMIOPOULOS, N.K. ROBAKIS (1997) Melatonin prevents death of neuroblastoma cells exposed to Alzheimer’s arnyloid peptide. J. Neurosci. 17:1683-1690.
PERRY, T.L., V.W. YONG, R.M. CLAVIER, K. JONES, J.M. WRIGHT, J.G. FOULKS, R.A. WALL (1985) Partial protection from the dopaminergic neurotoxin N-methyl-4-phenyl-l,2,3,6- tetrahydropyridine by four different antioxidants in the mouse. Neurosci. Lett. 60:109-114.
PIERI, C., M. MARRA, F. MORONI, R. DECCHIONI, F. MARCHESELLI (1994) Melatonin: A peroxyl radical scavenger more effec- tive than vitamin E. Life Sci. 55:271-276.
PIERREFICHE G., G. TOPALL, G. COURBAIN, I. HENRIET, H. LABORIT (1993) Antioxidant activity of melatonin in mice. Res. Commun. Chem. Pathol. Pharmacol. 80:211-223.
POZO, D., R.J. REITER, J.R. CALVO, J.M. GUERRERO (1994) Physi- ological concentrations of melatonin inhibit nitric oxide syn- thase in rat cerebellum. Life Sci. 55:455460.
RAMSAY, R.R., J. DADGAR, A. TREVOR, T.P. SINGER (1986) En- ergy-driven uptake of N-methyl-4-phenylpyridine by brain
17~70-76.
5 3 : 373-382.
191
COLTON, C.A., F. PAGAN, J. SNELL, J.S. COLTON, A. CUMMINS, D.L. GILBERT (1995) Protection from oxidation enhances the survival of cultured mesencephalic neurons. Exp. Neurol. 13254-61.
COSTA, E.J., R.H. LOPES, M.T. LAMY-FREUND (1995) Permeability of pure lipid bilayers to melatonin. J. Pineal. Res., 19:123-126.
DESOLE, M.S., L. SCIOLA, M.R. DELOGU, S. SIRCANA, R. MIGHELI (1996) Manganese and l-methy1-4-(2’-ethylphenyl)-l,2,3,6- tetrahydropyridine induce apoptosis of PC 12 cells. Neurosci. Lett. 209:193-196.
DRACHMAN, D.A. AND C.F. LIPPA (1992) The etiology of Alzheimers disease: the pathogenesis of dementia. Ann. N. Y. Acad. Sci. 648:176-186.
FERRARI, G., C.Y.I. YAN, L. GREENE (1 995) N-acetylcysteine (D- and L-stereoisomers) prevents apoptotic death of neuronal cells. J. Neurosci. 15:2857-2866.
FREEMAN, R.F., S. ESTUS, E.M. JOHNSON JR. (1994) Analysis of cell-related gene expression in postmitotic neurons: selective induction of Cyclin D l during programmed cell death. Neu- ron 12:343-355.
FURUTA, S., H. HAYASHI, M. HIJIKATA, S. MIYAZAWA, T. OSUMI AND T. HASHIMOTO ( 1 986) Complete nucleotide sequence of cDNA and deduced amino acid sequence of rat liver catalase. Proc. Natl. Acad. Sci. U.S.A. 83:313-317.
GAVRIELI, Y., Y. SHERMAN, S.A. BEN-SASSON (1992) Identifica- tion of programmed cell death in situ via specific labeling of nuclear DNA fragmentation. J. Cell. Biol. 119:493-501.
GOTZ, M.D., A. FREYBERG, P. RIEDERER (1990) Oxidative stress: A role in the pathogenesis of Parkinson’s disease. J. Neural Transm. 29:24 1-249.
GREENE, L., AS. TISCHLER (1976) Establishment of a noradr- energic clonal line of rat adrenal pheochromocytoma cells which respond to nerve growth factor. Proc. Natl. Acad. Sci. US .A. 73:2424-2428.
GUNNING, P., P. PONTE, H. OKAYAMA, J. ENGEL, H. BLAU, L. KEDES (1 983) Isolation and characterization of full-length cDNA for human alpha, beta and gamma actin have an amino- terminal cysteine that is subsequently removed. Mol. Cell Biol. 3:787-795.
HANSEN, M.B., S.E. NIELSEN, K. BERG (1989) Re-examination and further development of a precise and rapid dye method for measuring cell growth/cell kill. J. Immunol. Methods
HEINTZ, N. (1993) Cell death and the cell cycle: a relationship between transformation and degeneration? Trends Biochem. Sci. 18:157-159.
HILL, B.T., D. BLASK (1988) Effects of the pineal hormone me- latonin on the proliferation and morphological characteristics of human breast cancer cells (MCF7) in culture. Cancer Res. 48:6121-6126.
Ho, Y.S., J.D. CRAPO (1987a) Nucleotide sequencing of cDNA coding for rat manganese-containing superoxide dismutase. Nucleic Acid Res. 15:10070.
Ho, Y.S., J.D. CRAPO (1987b) cDNA and deduced amino acid sequence of rat copper-zinc-containing superoxide dismutase. Nucleic Acid Res. 15:6746.
HUNG, H-C., E.H.Y. LEE (1996) The mesolimbic dopaminer- gic pathway is more resistant than the nigrostriatal dopamin- ergic pathway to MPTP and MPP toxicity: Role of BDNF gene expression. Mol Brain Res. 41:16-26.
ICHITANY, Y., H. OKAMURA, Y. MATSUMOTO, I. NAGATSU, Y. IVATA (1991) Degeneration of the nigral dopamine neurons after 6-OHDA injection into the rat striatum. Brain Res. 549: 350-353.
JAIN, A., J. MARTENSSON, E. STOLE, P.A.M. AULD, A, MEISTER (1 99 1) Glutathione deficiency leads to mitochondria1 damage in brain. Proc. Natl. Acad. Sci. U S A . 88:1913-1917.
119~203-210.

Mayo et al.
mitochondria mediates the neurotoxicity of MFTP. Life Sci.
REITER, R.J. (1991) Pineal melatonin: cell biology of its syn- thesis and of its physiological interactions. Endocr. Res. 12: 15 1-180.
REITER, R.J. (1992) The aging pineal gland and its physiologi- cal consequences. BioEssays 14:169-175.
REITER, R.J. (1995a) Oxidative processes and antioxidative de- fense mechanisms in the aging brain. FASEB J. 9526-533.
REITER, R.J. (1995b) Oxygen radical detoxification processes during aging: The functional importance of melatonin. Ag- ing Clin. Exp. Res. 7:34&351.
REITER, R.J. (1997) Antioxidant actions of melatonin. Adv. Pharmacol. 38:103-117.
REITER, R.J., M.I. PABLOS, T.T. AGAPITO, J.M. GUERRERO ( 1996) Melatonin in the context of the free radical theory of aging. Ann. N.Y. Acad. Sci. 286:362-378.
RUBIN, L.L., K.L. PHILPOTT, S.F. BROOKS (1993) The cell cycle and cell death. Cum Biol. 3:391-394.
SAINZ, R.M., J.C. MAYO, H. URIA, M. KOTLER, I. ANTOL~N, C. RODRfGUEZ, A. MENENDEZ-PELAEZ (1995) The pineal neurohor- mone melatonin prevents in vivo and in vitro apoptosis of thy- mocytes. J. Pineal Res. 19:178-188.
SCHULZ, J.B., R.T. MATTEWS, M.M. MUQIT, S.E. BROWNE, M.F. BEAL (1995) Inhibition of neuronal nitric oxide synthase by 7-nitroindazole protects against MFTP-induced neurotoxicity in mice. J. Neurochem. 64:936-939.
SHELLARD, S.A., R.D. WHELAN, B.T. HIL (1989) Growth in- hibitory and citotoxic effect of melatonin and its metabo- lites on human tumor cell lines in vitro. Br. J. Cancer 60:
SHIDA, C.S., A.M.L. CASTRUCCI, M.T. LAMY-FREUND (1994) High melatonin solubility in aqueous medium. J. Pineal Res.
SHULTS, C., T. KIMBER, A. ALTAR (1995) BDNF attenuates the effects of intrastriatal injection of 6-hydroxydopamine. NeuroReport 6: 1109-1 1 12.
oxide production in rat brain in vivo. J. Neurochem. 34:1421-1428.
SOKOLOFF, L. (1996) The metabolism of the central nervous sys- tem in vivo. In: Handbook of Physiology-Neurophysiology. Field, J., H.W. Magoun, V.E. Hall, eds. American Physiologi- cal Society, Washington, Vol3, pp. 1843-1864.
SPINA, M.B., S.P. SQUINTO, J. MILLER, R.M. LINDSAY, c. HYMAN (1992) Brain-derived neurotrophic factor protects dopamine neurons against 6-hydroxydopamine and N-methys-4- phenylpyridinium ion toxicity: involvement of the glutathione system. J. Neurochem. 59:99-106.
STADTMAN, E.R. (1992) Protein oxidation and aging. Science 257: 1220-1224.
39:581-588.
288-295.
16: 198-20 1 .
SINET, P.M., R.E. HEIKKILA, G. COHEN (1980) Hydrogen per-
TAN, D.X., L.D. CHEN, B. POEGGELER, L.C. MANCHESTER, R.J. REITER (1993) Melatonin: A potent endogenous hydroxyl radi- cal scavenger. Endocr. J. 157-60.
TAN, D.X., R.J. REITER, L.D. CHEN, B. POEGGELER, L.C. MANCHESTER, L. BARLOW-WALDEN (1994) Both physiological and pharmacological levels of melatonin reduce DNA adduct formation induced by the carcinogen safrole. Carcinogenesis 15:215-218.
fect of MPTP and L-deprenyl on antioxidant enzymes and lipid peroxidation levels in mouse brain. J. Neurochem.
THIFFAULT, c., N. AUMONT, R. QUIRION, J. POIRIER (1995) Ef-
65:2725-2733. TIFFANY-CASTIGLIONI, E., J.R. ~ R E Z - ~ O L O (198 I ) Stimulation Of
resistance to 6-hydroxidopamine in a human neuroblastoma cell line by nerve growth factor. Neurosci. Lett. 26:157-161.
B.J. HOFFER, L. OLSON (1995) Protection and repair of the nigrostriatal dopaminergic system by GDNF in vivo. Nature 373:335-339.
SHELANSKI (1 996) Downregulation of Cu/Zn superoxide dismutase leads to cell death via the nitric oxide-peroxinitrite pathway. J. Neurosci. 16:253-261.
TURSKI, L., K. BRESSLER, K.J. RETTIG, P.A. LOSCHMANN, H. Wachtel(l991) Protection of substantia nigra from MPP neu- rotoxicity by N-methyl-D-aspartate antagonists. Nature 349:414-418.
WALKINSHAW, G., C.M. WATERS (1994) Neurotoxin-induced cell death in neuronal PC12 cells is mediated by induction of apoptosis. Neuroscience 63:975-987.
WIESENBERG, I., M. MISSBACH, J.-P. KAHLEN, M. SCHRADER, C. CARLBERG (1995) Transcriptional activation of the nuclear re- ceptor RZRa by the pineal gland hormone melatonin and identification of CGP 52608 as a synthetic ligand. Nucleic Acids Res. 23:327-333.
Wu, R.M., D.L. MURPHY, C.C. CHIUEH (1996) Suppression of hydroxyl radical formation and protection of nigral neurons by I-deprenyl (selegiline). Ann. N.Y. Acad. Sci. 786:379-390.
WYLLiE, A.H., J.F.R. KERR, A.R. CURRIE (1980) Cell death: the significance of apoptosis. Int. Rev. Cytol. 58:25 1-301.
(1988) Determination of nucleotide sequence of cDNA cod- ing rat glutathione peroxidase and diminished of the mRNA in selenium deficient rat liver. Biochem Biophys Res Commun 154:1024-1028.
ZIGMOND, M.J., E.M. STRICKER (1989) Animal models of Par- kinsonism using selective neurotoxins: clinical and basic im- plications. Int. Rev. Neurobiol. 3 1 : 1-79.
ZUBER, M., A. ALPEROVITCH (1991) Maladie de Parkinson et facteurs environnementaux. Rev. Epidemiol. Sante Publique,
TOMAC, A., E. LINDQVIST, L.F. H. LIN, S.O. OGREN, D. YOUNG,
TROY, M., D. DEROSSI, A. PROCHIANTZ, L.A. GREENE, L.
YOSHIMURA, S., S. TAKEKOSHI, K. WATANABE, Y. FUJII-KURIYAMA
39:373-387.
192
Related Documents











