M ecanisme d’absorption intestinale des acides gras a longue chaı ˆne : r ^ ole emergent du CD36 L’apport excessif de lipides, associ e a un d es equilibre qualitatif (exc es des graisses satur ees et de cholest erol, rapport n-6/n- 3 trop elev e), contribue a l’augmenta- tion de la pr evalence de l’ob esit e et a l’apparition des maladies associ ees (ath eroscl erose, diab ete non insulino- d ependant) (Bray et al., 2004 ; de Wit et al., 2011). Parmi les diff erents organes impliqu es dans le maintien de l’hom eostasie lipidique, le tractus diges- tif est le moins etudi e. Ce constat est paradoxal puisque l’intestin conditionne la biodisponibilit e des lipides alimentai- res a l’organisme et contribue a la r egulation de la triglyc erid emie post- prandiale (Tomkin et Owens, 2012). Or une hypertriglyc erid emie postprandiale est souvent associ ee aux dyslipid emies, a l’ob esit e, a l’insulino-r esistance et est un facteur de risque des maladies cardio- vasculaires. Par ailleurs, l’intestin parti- cipe a la r egulation de la prise alimentaire en particulier par l’interm ediaire de la s ecr etion d’hormones intestinales (CCK, GLP-1, GLP2...GIP) (Steinert et Beglinger, 2011) et de l’apolipoprot eine AIV qui est constitutive des lipoprot eines intestinales (Tso et Liu, 2004). L’ensemble de ces fonctions fait de l’intestin un organe pertinent dans l’ etude de la mise en place de l’ob esit e et des maladies associ ees. Plus particu- li erement, les perturbations du m etabo- lisme lipidique intestinal peuvent retentir sur les capacit es d’absorption et sur la lip emie postprandiale et affecter plus largement le m etabolisme des autres lipoprot eines (Mora et al., 2008). En ce qui concerne l’absorption intesti- nale des lipides, les m ecanismes cellulai- res et mol eculaires ne sont pas encore totalement elucid es. Cela s’explique par le fait que l’intestin a longtemps et e consid er e comme une simple barri ere passive capable d’absorber de grandes quantit es de lipides alimentaires. Etant donn e les r^ oles fondamentaux des acides gras a longue chaı ˆne (AGLC) dans la cellule (fonctionnel, energ etique, m etabolique et de signalisation cellu- laire), on conc ¸oit que leur biodisponibilit e ne soit pas limitante. Le fait que l’excr e- tion f ecale des lipides soit de l’ordre de 5 % chez l’homme malgr e de grandes variations de l’apport lipidique et que la st eatorrh ee soit diagnostiqu ee d es que la perte exc ede 7 % souligne l’efficacit e de l’absorption intestinale des AGLC (Ross, 1993). Ces derni eres ann ees plusieurs lipid-binding proteins (LBP) ont et e identi- fi ees au niveau de l’ent erocyte montrant la complexit e du m ecanisme d’absorp- tion et sugg erant que des modifications de leur niveau d’expression intestinale Abstract: Mechanism of Intestinal Absorption of Long chain Fatty acid: Emerging role of CD36 Excessive lipid intake, associated with a qualitative imbalance, favors the development of obesity and associated diseases. From organs involved in the lipid homeostasis, the small intestine remains the most poorly known although it is responsible for the lipid bioavailability and largely contributes to the regulation of postprandial hypertriglycer- idemia. The mechanism of long chain fatty acid (LCFA) intestinal absorption is not totally elucidated. Over the two last decades, cloning of lipid binding proteins (LBP), proteins involved in trafficking and metabolic fate of LCFA in gut have provided new insights on cellular and molecular mechanisms involved in fat absorption. The synthesis of recent literature indicates that intestine is able to adapt its absorption capacity to the fat content of the diet. This adaptation takes place through a fat-coordinated induction of LBP and apolipoproteins. CD36 could operate as a lipid sensor responsible for a transducing signal related to the lipid content of the diet at the origin of this intestinal adaptation. This lipid-mediated metabolic response may lead to the formation of large chylomicrons rapidly degraded in the blood. All together, these new data indicate that this intestinal lipid sensing mechanism may be a therapeutic target for reducing the postprandial hypertriglyceridemia and associated cardiovascular risks. Key words: intestine, CD36, lipid-binding proteins, intestinal adaptation, sensing, lipid absorption, postprandial hypertriglyceridemia Thi Thu Trang TRAN Marjorie BUTTET V eronique TRAYNARD Philippe BESNARD H el ene POIRIER Isabelle NIOT Physiologie de la nutrition et toxicologie (Nutox), UMR U866 Inserm/universit e de Bourgogne/AgroSup Dijon 1 Esplanade Erasme, 21000 Dijon, France <[email protected]> Article rec ¸u le 1 er juin 2012 Accept e le 8 juin 2012 Pour citer cet article : Tran TTT., Buttet M, Traynard V, Besnard P, Poirier H, Niot I. M ecanisme d’absorption intestinale des acides gras a longue chaı ˆne : r^ ole emergent du CD36. OCL 2012 ; 19(4) : 200-208. doi : 10.1684/ocl.2012.0461 doi: 10.1684/ocl.2012.0461 200 OCL VOL. 19 N8 4 juillet-aou ˆ t 2012 DOSSIER : ABSORPTION INTESTINALE DES LIPIDES

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

M�ecanisme d’absorption intestinale des acides gras�a longue chaıne : role �emergent du CD36
L’apport excessif de lipides, associ�e �a und�es�equilibre qualitatif (exc�es des graissessatur�ees et de cholest�erol, rapport n-6/n-3 trop �elev�e), contribue �a l’augmenta-tion de la pr�evalence de l’ob�esit�e et �al’apparition des maladies associ�ees(ath�eroscl�erose, diab�ete non insulino-d�ependant) (Bray et al., 2004 ; de Witet al., 2011). Parmi les diff�erents organesimpliqu�es dans le maintien del’hom�eostasie lipidique, le tractus diges-tif est le moins �etudi�e. Ce constat estparadoxal puisque l’intestin conditionnela biodisponibilit�e des lipides alimentai-res �a l’organisme et contribue �a lar�egulation de la triglyc�erid�emie post-prandiale (Tomkin et Owens, 2012). Orune hypertriglyc�erid�emie postprandialeest souvent associ�ee aux dyslipid�emies, �al’ob�esit�e, �a l’insulino-r�esistance et est unfacteur de risque des maladies cardio-vasculaires. Par ailleurs, l’intestin parti-
cipe �a la r�egulation de la prise alimentaireen particulier par l’interm�ediaire dela s�ecr�etion d’hormones intestinales(CCK, GLP-1, GLP2. . .GIP) (Steinert etBeglinger, 2011) et de l’apolipoprot�eineAIV qui est constitutive des lipoprot�einesintestinales (Tso et Liu, 2004).L’ensemble de ces fonctions fait del’intestin un organe pertinent dansl’�etude de la mise en place de l’ob�esit�eet des maladies associ�ees. Plus particu-li�erement, les perturbations du m�etabo-lisme lipidique intestinal peuvent retentirsur les capacit�es d’absorption et sur lalip�emie postprandiale et affecter pluslargement le m�etabolisme des autreslipoprot�eines (Mora et al., 2008).
En ce qui concerne l’absorption intesti-nale des lipides, les m�ecanismes cellulai-res et mol�eculaires ne sont pas encoretotalement �elucid�es. Cela s’explique par
le fait que l’intestin a longtemps �et�econsid�er�e comme une simple barri�erepassive capable d’absorber de grandesquantit�es de lipides alimentaires. �Etantdonn�e les roles fondamentaux des acidesgras �a longue chaıne (AGLC) dansla cellule (fonctionnel, �energ�etique,m�etabolique et de signalisation cellu-laire), onconcoit que leur biodisponibilit�ene soit pas limitante. Le fait que l’excr�e-tion f�ecale des lipides soit de l’ordre de5 % chez l’homme malgr�e de grandesvariations de l’apport lipidique et que last�eatorrh�ee soit diagnostiqu�ee d�es que laperte exc�ede 7 % souligne l’efficacit�e del’absorption intestinale des AGLC (Ross,1993). Ces derni�eres ann�ees plusieurslipid-binding proteins (LBP) ont �et�e identi-fi�ees au niveau de l’ent�erocyte montrantla complexit�e du m�ecanisme d’absorp-tion et sugg�erant que des modificationsde leur niveau d’expression intestinale
Abstract: Mechanism of Intestinal Absorption of Long chain Fatty acid:Emerging role of CD36Excessive lipid intake, associated with a qualitative imbalance, favors the development ofobesity and associated diseases. From organs involved in the lipid homeostasis, the smallintestine remains the most poorly known although it is responsible for the lipidbioavailability and largely contributes to the regulation of postprandial hypertriglycer-idemia. Themechanism of long chain fatty acid (LCFA) intestinal absorption is not totallyelucidated. Over the two last decades, cloning of lipid binding proteins (LBP), proteinsinvolved in trafficking and metabolic fate of LCFA in gut have provided new insights oncellular and molecular mechanisms involved in fat absorption. The synthesis of recentliterature indicates that intestine is able to adapt its absorption capacity to the fatcontent of the diet. This adaptation takes place through a fat-coordinated induction ofLBP and apolipoproteins. CD36 could operate as a lipid sensor responsible for atransducing signal related to the lipid content of the diet at the origin of this intestinaladaptation. This lipid-mediated metabolic response may lead to the formation of largechylomicrons rapidly degraded in the blood. All together, these new data indicate thatthis intestinal lipid sensing mechanism may be a therapeutic target for reducing thepostprandial hypertriglyceridemia and associated cardiovascular risks.
Key words: intestine, CD36, lipid-binding proteins, intestinal adaptation, sensing, lipidabsorption, postprandial hypertriglyceridemia
Thi Thu Trang TRANMarjorie BUTTETV�eronique TRAYNARDPhilippe BESNARDH�el�ene POIRIERIsabelle NIOT
Physiologie de la nutrition et toxicologie
(Nutox),
UMR U866 Inserm/universit�e de
Bourgogne/AgroSup Dijon
1 Esplanade Erasme,
21000 Dijon, France
Article recu le 1er juin 2012
Accept�e le 8 juin 2012
Pour citer cet article : Tran TTT., Buttet M, Traynard V, Besnard P, Poirier H, Niot I. M�ecanisme d’absorption intestinale des acides gras �a longuechaıne : role �emergent du CD36. OCL 2012 ; 19(4) : 200-208. doi : 10.1684/ocl.2012.0461
doi:10.1684/o
cl.2012.0461
200 OCL VOL. 19 N8 4 juillet-aout 2012
DOSSIER : ABSORPTION INTESTINALE DES LIPIDES

retentissent sur l’efficacit�e de l’absorptionet la lip�emie postprandiale, et plusg�en�eralement sur l’hom�eostasie des lipi-des de l’organisme (Niot et al., 2009).C’est pourquoi, cette revue abordera lerole des LBP en particulier les membra-naires dans le m�ecanisme d’absorptiondes lipides et sa r�egulation.
Absorption intestinaledes lipides alimentaires
L’alimentation apporte de 30 �a 40 % del’�energie sous forme de lipides, princi-palement des triglyc�erides (TG). Du faitde leur caract�ere hydrophobe, l’absorp-tion des TG au niveau intestinal estun processus complexe comportantplusieurs �etapes (Niot et al., 2009)(figure 1) : le captage des AGLC, letrafic intracellulaire, la synth�ese et las�ecr�etion des chylomicrons.
�A l’arriv�ee dans la lumi�ere intestinale, leslipides alimentaires, principalement desTG sont hydrolys�es en monoglyc�eride(MG) et AGLC grace aux lipases gas-
triques et pancr�eatiques. Ces produitssont ensuite associ�es au cholest�erol, auxphospholipides, aux sels biliaires etforment des micelles mixtes (voir l’arti-cle pr�esent�e dans ce num�ero par Beaslaset al., 2012). Il est admis que lesAGLC sont absorb�es pr�ef�erentiellementau niveau j�ejunal (Mariadason et al.,2005) par les ent�erocytes hautementdiff�erenci�es se trouvant dans les deuxtiers sup�erieurs des villosit�es intestinales(Niot et al., 2009).
Le captage des acides gras�a longue chaıne
Les ent�erocytes sont hautementpolaris�es. La membrane apicale del’ent�erocyte sedistinguede lamembranebasolat�erale par l’existence de micro-villosit�es qui augmentent la surfaced’�echange. Ces microvillosit�es en plusdu glycocalix (r�eseau de glycoprot�eines)et du mucus participent �a la formationd’une couche d’eau non agit�ee dontl’�epaisseur peut aller jusqu’�a 500 mm etqui est caract�eris�ee par un pH acide du
fait de la pr�esence de pompes �a protonsau niveau de la membrane apicale(figure 1). Ce microenvironnement est�a l’originede ladissociationmicellairequilib�ere les AGLC proton�es �a proximit�eimm�ediate de la surface de la bordure enbrosse des ent�erocytes (Shiau et al.,1985) et facilite leur captagepardiffusionpassive (Niot et al., 2009). Ce constatpose la question du role physiologiquedes LBPmembranaires identifi�ees dans leprocessus d’absorption des AGLC : laFABPm, le FATP4 et CD36.
La FABPpm
La FABPpm est exprim�ee aussi bien dansla bordure en brosse que dans lesmembranes cellulaires lat�erales desent�erocytes en cours de diff�erenciationet diff�erenci�es (Stremmel et al., 1985b).Le fait que l’utilisation de l’anticorps anti-FABPpm sur des explants j�ejunauxr�eduise le captagedes acidesgrasmontrequ’elle pourrait jouer un role dans cette�etape membranaire (Stremmel, 1988).De plus, la FABPpm est aussi pr�esentedans la mitochondrie puisqu’elle estl’homologue de la mitochondrial aspar-tate-aminotransferase (mASpAT) quicatalyse les r�eactions de transaminationreliant le cycle de l’ur�ee au cycle de Krebs(Stump et al., 1993). Ces donn�eesmontrent que des �etudes plus approfon-dies sont n�ecessaires pour d�eterminer sila FABPpm a un role physiologique dansl’absorption intestinale des lipides.L’invalidation de ce g�ene n’a toujourspas �et�e r�ealis�ee �a ce jour.
La FATP4
La FATP4 est une prot�eine membranaireappartenant �a la famille des FATP quicomporte cinq FATP murines et six FATPhumaines et dont chaquemembre a uneexpression tissulaire sp�ecifique (Doege etStahl, 2006). Au niveau intestinal, laFATP4 est majoritairement exprim�ee auniveau des ent�erocytes hautementdiff�erenci�es du j�ejunum. Sa localisationent�erocytaire fait l’objet de discussionspuisqu’elle a �et�e initialement d�ecrite auniveau des membranes microvillositaires(Stahl et al., 1999), mais serait plusmajoritairement pr�esente dans ler�eticulum endoplasmique (RE) (Garcia-Martinez et al., 2005 ; Milger et al.,2006). Il a �et�e montr�e en utilisant desent�erocytes isol�es de souris d�epourvusdecouche d’eau non agit�ee, qu’il existe unecorr�elation entre l’efficacit�e de captagedes AGLC et l’expression de la FATP4
Lipides alimentairesTG AGLC-
2-MG
Sels biliaires etphospholipides
Couche d’eaunon agitée Diffusion
passive
AGLC
Captage
Micelles mixtes
FABPpmFATP4
CD36
Traficintracellulaire
AGLCACS
RE
Golgi
Synthèse et sécrétiondes lipoprotéines
TG, PL, EC
ApoB48
Lymphe SangChylomicron
ApoA-IV
ApoC-IIApoC-III
I-FABPL-FABP
PCTV
AGLC-SCoA
L-FABP
MTP
Figure 1. Principales �etapes de l’absorption intestinale des AGLC.
AGLC : Acide gras �a longue chaıne ; MG : monoglyc�erides ; TG : triglyc�erides ; PL : phospholipi-des ; EC : ester de cholest�erol ; LBPs : lipid binding protein ; CD36 : fatty acid transporter ;FATP4 : fatty acid transporter protein 4 ; I-FABP : intestinal fatty acid-binding protein ; L-FABP :liver fatty acid-binding protein ; ACS : Acy CoA synth�etase ; AGLC-SCoA : Acyl CoA ; ACBP :Acyl CoA-binding protein ; MTP : microsomal triglyceride transfer protein ; ApoB48 : apolipo-prot�eine B48 ; ApoA-IV : apolipoprot�eine A-IV ; RE : r�eticulum endoplasmique.
OCL VOL. 19 N8 4 juillet-aout 2012 201

(Gimeno et al., 2003 ; Stahl et al., 1999).Ces donn�ees indiquent que la FATP4participe au captage des AGLC au niveauent�erocytaire.Cependant, la structuredela prot�eine FATP4 essentiellement cyto-solique, avec une courte s�equence extra-cellulaire du cot�e N-terminal qui necontient pas de domaine de liaison auxacides gras (Gertow et al., 2004 ; Stahlet al., 2001), met en doute son role detransporteur des AGLC. En revanche, ilest maintenant d�emontr�e que la FATP4porte une activit�e Acyl-CoA synth�etasedes acides gras �a longues et tr�es longueschaınes (Hall et al., 2005). Ainsi, laFATP4 en activant les AGLC en Acyl-CoA participe au maintien d’un gradientfavorable �a leur entr�ee dans l’ent�erocytepar diffusionpassive (Milger et al., 2006).L’ensemble de ces donn�ees expliquepourquoi la d�el�etion de FATP4 au niveauintestinal chez la souris ne modifie pasl’efficacit�e d’absorption intestinale deslipides alimentaires (captage et excr�etionf�ecale). On observe cependant uneaccumulation l�eg�ere de TG au niveaudes ent�erocytes quand ces sourisd�eficientes en FATP4 sont nourries avecun r�egime hyperlipidique (Shim et al.,2009). Ce r�esultat souligne son role dansle m�etabolisme ent�erocytaire des TG quis’effectue principalement au niveau duRE (Milger et al., 2006). Le fait que ladiminution d’expression de la FATP4(Frochot et al., 2012) et que le poly-morphismeGly-209-Ser sur le g�ene de laFATP4 observ�e chez l’homme se tradui-sent par une r�eduction de l’hypertri-glyc�erid�emie (Gertow et al., 2004) est enaccord avec cette hypoth�ese.
CD36
Le CD36 est une glycoprot�eine multi-fonctionnelle capable de lier les AGLCsatur�es ou non, ionis�es, avec une affinit�ede l’ordre du nanomolaire (Ibrahimiet al., 1996) mais �egalement le col-lag�ene, les phospholipides anioniques,les LDL oxyd�ees, le Plasmodium falcipa-rum, la thrombospondine et la b-amy-loıde (Martin et al., 2011 ; Silverstein etFebbraio, 2009 ; Su et Abumrad, 2009).Le CD36 est localis�e principalement auniveau des membranes des bordures enbrosse des deux tiers sup�erieurs desvillosit�es dans la partie proximale del’intestin grele (Lobo et al., 2001 ; Poirieret al., 1996). L’expression intestinale duCD36 est �etroitement corr�el�ee avec lateneur en lipides du r�egime. Alors qu’unr�egime pauvre en lipides diminue son
expression (Sukhotnik et al., 2001), unr�egime hyperlipidique chronique l’induitfortement (Poirier et al., 1996). Cepen-dant, �a la diff�erence de l’adipocyte et dumyocyte, la contribution significativede CD36 dans le captage ent�erocytairedes AGLC est mise en doute. En effet,selon les auteurs, la d�el�etion du g�ene duCD36 diminue le captage des AGLC parles ent�erocytes isol�es d�epourvus decouche d’eau non agit�ee (Nassir et al.,2007), ou n’a pas d’effet (Drover et al.,2005). Nos travaux ont d�emontr�ela pertinence physiologique de cesderni�eres donn�ees puisque des ansesj�ejunales isol�ees in situ de souris sauvageset d�eficientes en CD36 qui conservent lacouched’eaunonagit�eeet ladynamiquede l’absorption,ont lamemeefficacit�edecaptage (Tran et al., 2011). Ces derniersr�esultats expliquent pourquoi l’invalida-tion du g�ene de CD36 chez la souris nese traduit pas in vivo par une modifica-tion de l’absorption nette des AGLCexcept�e pour des acides gras �a tr�eslongues chaınes rares dans l’alimentation(Drover et al., 2008 ; Nassir et al., 2007 ;Nauli et al., 2006). Cependant, le CD36participe bien au m�ecanisme d’absorp-tion des lipides alimentaires puisque sad�eficience r�eduit consid�erablement las�ecr�etion lymphatique des chylomicronsqui sont moins riches en TG et donc deplus petites tailles (Masuda et al., 2009 ;Nauli et al., 2006). Ces effets sontassoci�es �a une r�etention de TG au niveauent�erocytaire en r�egime hyperlipidiquechronique (Drover et al., 2005). Cesdonn�ees d�emontrent que le CD36 joueun role tr�es important dans la formationdes chylomicrons mais pas commetransporteur efficace des lipides.
En conclusion, l’existence de la couched’eau non agit�ee en plus des propri�et�esstructurelles et fonctionnelles des LBPmembranaires indique que la contribu-tion d’un transport facilit�e dans lecaptage intestinal des acides gras alimen-taires n’est probablement pas pr�epond�e-rante en p�eriode postprandiale. Enrevanche, la diffusion passive, syst�emede transport �a forte capacit�e mais �afaible affinit�e, est particuli�erement bienadapt�ee et �eviterait ainsi que le captagedes AGLCnedevienne un facteur limitantdans l’absorption intestinale des lipidesapr�es un repas (Chow et Hollander,1979).
Il est important de noter que l’efficacit�ede cette diffusion passive est condition-n�ee par le maintien d’un gradient de
concentration en AGLC entre les milieuxextra- et intracellulaire d�ependantde l’efficacit�e du m�etabolisme ent�ero-cytaire des AGLC. Cependant, pour desconcentrations plus faibles, en situationinterprandiale par exemple, la diffusionfacilit�ee pourrait s’av�erer plus efficace,du fait d’une plus forte affinit�e pourles AGLC.
Trafic intracellulairedes acides gras �a longue chaıne
Une fois dans le cytoplasme, les AGLCsont pris en charge par des fatty acid-binding proteins (I et L-FABP) dont lesfonctions respectives ne sont pas tota-lement �elucid�ees. Cependant, leur liai-son avec les AGLC en maintenant legradient de concentration favorise leurcaptage par diffusion passive (Niot et al.,2009). Quand des AGLC sont d�elivr�es aupole apical des ent�erocytes, ils sontpr�ef�erentiellement li�es �a la I-FABP,isoforme, uniquement exprim�ee danscet organe, qui requiert un m�ecanismecollisionnel avec les phospholipidesmembranaires pour transf�erer les AGLC.Ces deux propri�et�es sugg�erent que la I-FABP est impliqu�ee dans le transfertvectoriel des AGLC vers le RE, site de r�e-est�erification des acides gras (Corsicoet al., 1998). Puisque, l’�echange desAGLC pour la L-FABP s’effectue pardiffusion aqueuse, elle serait plutot �al’origine de la formation d’un poolintracellulaire d’AGLC. Par ailleurs, elleserait un partenaire des PPARs. Enfin auniveau intestinal, la L-FABP participe autransport intracellulaire des pr�echylomi-crons comme nous le d�ecrirons plustard. En accord avec cette derni�erehypoth�ese, l’invalidation de la L-FABPconduit chez la souris soumise �a unecharge en lipides �a une moindres�ecr�etion de TG associ�ee �a leur r�etentionent�erocytaire (Newberry et al., 2006).
Synth�ese et s�ecr�etiondes chylomicrons
Les AGLC sont ensuite activ�es en Acyl-CoA par des Acyl-CoA synth�etases (ACS).Pendant la p�eriode postprandiale,les Acyl-CoA form�es sont r�e-est�erifi�espr�ef�erentiellement en TG avant d’etretransloqu�es dans le RE avec les phospho-lipides et les esters de cholest�erol par lamicrosomal triglyceride transfer protein(MTP).Ce transfert conduit�a la lipidationd’une apolipoprot�eine B48 (ApoB48),forme tronqu�ee de l’ApoB100, sp�eci-
202 OCL VOL. 19 N8 4 juillet-aout 2012

fique de l’intestin au moins chezl’homme et le hamster. L’assemblagedes pr�echylomicrons n�ecessite plusieurs�etapes (Iqbal et Hussain, 2009). En effet,l’association d’une ApoB48 avec unefaible quantit�e de lipides (TG et phos-pholipides) par la MTP constitue unelipoprot�eine primordiale qui fusionneavec des gouttelettes lipidiques conte-nant des TG et des esters de cholest�erol,form�ee ind�ependamment dans le REpar la MTP. La MTP est indispensable�aficience chez l’homme et la souris(par mutation ou transgen�ese) est �al’origine de l’abetalipoprot�ein�emie quise caract�erise par une malabsorptions�ev�ere des lipides caract�eris�ee par unest�eathorr�ee, une r�etention ent�erocytairede TG et l’absence de s�ecr�etion delipoprot�eines contenant l’ApoB48(tableau 1) (Berriot-Varoqueaux et al.,2001 ; Xie et al., 2007). Cette �etapeconstitue donc la premi�ere �etape limi-tante de la formation des chylomicrons.L’ApoAIV est aussi fondamentale auniveau de cette �etape puisqu’elle favori-serait la lipidation en stabilisant le pr�echy-lomicron et/ou l’ApoB48 (Mansbach etGorelick, 2007 ; Weinberg et al., 2012).
Ces pr�echylomicrons sont ensuite trans-port�es vers l’appareil de Golgi par desv�esicules issues du RE appel�ees prechylo-microns transfert vesicles (les PCTV)qui dufait de leur grande taille, sont retrouv�eesuniquement au niveau intestinal. Lespr�echylomicrons subissent dans l’appa-reil de Golgi des �etapes de maturation(glycosylation) et acqui�erent diff�erentesapolipoprot�eines comme les apolipo-prot�eines CII, CIII et A1 (ApoCII, ApoCIII,ApoA1).Ce transfert constitue la seconde�etape limitante de l’absorption deslipides. Il requiert la L-FABP pour initierla g�en�eration des PCTV au niveau du RE(Neeli et al., 2007) ou d’autres prot�einespour leur fusion avec les membranes del’appareil deGolgi commeGTPase Sar 1bou vesicule associatedmembraneprotein7(VAMP7). Les chylomicrons sont s�ecr�et�esdans la lymphe et rejoignent la circula-tion sanguine ou leurs TG sont d�egrad�espar la lipoproteine lipase (LPL).
L’activit�e de la LPL est consid�er�ee commele r�egulateurmajeur de la lip�emie. Or, enplus de sonniveaud’expression, l’activit�ede la LPL est r�egul�ee par le rapportApoCII/CIII port�e par la lipoprot�eine. Eneffet, l’ApoCII est un activateur puissantde la LPL alors que l’ApoC-III est uninhibiteur (Jong et al., 1999). Meme si le
niveau de ces apolipoprot�eines peut etremodul�e par �echange avec les lipoprot�ei-nes h�epatiques dans le sang, le niveauacquis par les chylomicrons dansl’ent�erocyte conditionne �egalementl’efficacit�e de la LPL (Olivecrona etBeisiegel, 1997). De plus, la clairancedes chylomicrons d�epend �egalement dunombre et de la taille des lipoprot�eines.En effet, l’activit�e de la LPL est plusefficace sur un petit nombre de groschylomicrons que sur grand nombre depetits chylomicrons (Martins et al.,1996 ; Xiang et al., 1999). Ce derniercrit�ere refl�ete finalement l’efficacit�e de lalipidation de l’ApoB48.
En conclusion, hormis la MTP, les LBP nesont pas indispensables au m�ecanismed’absorption (tableau 1). Cependant,elles semblent faciliter �a diff�erentsniveaux, la synth�ese et la s�ecr�etion deschylomicrons, en particulier en situationde challenge lipidique. Ce qui expliquepourquoi leur d�eficience n’alt�ere pasl’absorption globale des lipides (alt�era-tion des pertes f�ecales en lipides) maisplutot la triglyc�erid�emie postprandiale(Niot et al., 2009).
R�egulation del’absorption intestinalepar les lipidesalimentaires
La question qui se pose est de savoir sil’intestin poss�ede de mani�ere constitu-tive la capacit�e �a absorber de grandesquantit�es de lipides ou bien s’il existeune r�eponse m�etabolique adaptative enfonction de la quantit�e des lipidespr�esents dans le r�egime. Cette secondepossibilit�e sugg�ere qu’une d�er�egulationde cette adaptation puisse se traduirepar des pertes f�ecales en lipides et/oudes dyslipid�emies postprandiales et desr�etentions de TG ent�erocytaires. Il estadmis chez l’homme et la souris quel’augmentation de la teneur en lipides setraduit par une augmentation de la tailledes chylomicrons plutot que de leurnombre (Cartwright et Higgins, 1999 ;Karpe et al., 1997). Comme lemontre letableau 1, le fait que l’expression deplusieurs LBP (MTP, CD36, L-FABP) etapolipoprot�eines (ApoIV) soit r�egulablein vitro par les AGLC laisse supposer quel’efficacit�e de la formation des chylomi-crons puisse etre modul�ee par les lipidesalimentaires. Nos travaux ont confirm�ecette hypoth�ese, puisqu’un segment
intestinal isol�e in situ de souris nourriesavec un r�egime hyperlipidique chro-nique poss�ede une plus grande capacit�e�a absorber les AGLC (augmentation de lacapacit�e de captage et du m�etabolismelipidique sans r�etention de TG ent�ero-cytaire) que celui de souris nourriesavec un r�egime normolipidique (Petitet al., 2007). Ces modifications expli-quent pourquoi in vivo, un r�egimehyperlipidique chronique ne conduitpas �a l’augmentation des pertes f�ecalesen lipides. Cette adaptation intestinales’explique par deux ph�enom�enescompl�ementaires : l’induction de laprolif�eration intestinale et l’inductioncoordonn�ee de l’expression des g�enesimpliqu�es dans la formation des chylo-microns (CD36, FATP4, I et LFABP, MTP,ApoAIV et ApoCII) (Petit et al., 2007). Deplus, il est int�eressant de noter que cetter�egulation est r�eversible et d�ependantede la teneur en lipides du r�egime (deWitet al., 2011 ; Kondo et al., 2006 ; Petitet al., 2007). En effet, en utilisant unr�egime contenant diff�erentes quantit�esde lipides (10 %,20 %,30 %et45 %delipides en masse), de Wit et al., 2011 ontmontr�e que le nombre de g�enes r�egul�eset l’amplitude de leur induction sontcorr�el�es �a la quantit�e de lipides dur�egime.
On note que l’expression de l’ApoCIIIent�erocytaire estdiminu�eealorsquecellede l’ApoCII est induite en cas de r�egimehyperlipidique riche en AGLC insatur�es(Kondo et al., 2006 ; Petit et al., 2007).Par cons�equent, l’augmentation durapport ApoCII/ApoCIII et de la tailledes chylomicrons expliqueraient la plusfaiblehypertriglyc�erid�emiepostprandialeobserv�ee paradoxalement apr�es ce typede r�egime hyperlipidique chez le ron-gueur (Petit et al., 2007) et retrouv�eechez l’homme (Defoort et al., 2011).Cette adaptation intestinale est donc unfacteur �emergent de la r�egulation de latriglyc�erid�emie postprandiale.
De mani�ere int�eressante, au moins chezla souris, l’adaptation intestinale �a lateneur en lipides du r�egime contribue �ala r�egulation de l’hom�eostasie lipidique(de Wit et al., 2011 ; Kondo et al.,2006) puisqu’un d�efaut est associ�e aud�eveloppement de l’ob�esit�e (Kondoet al., 2006). Ces diff�erents constatsindiquent que la compr�ehension dum�ecanisme de l’adaptation du m�eta-bolisme ent�erocytaire �a la teneur enlipides du r�egime devrait permettred’envisager des actions th�erapeutiques
OCL VOL. 19 N8 4 juillet-aout 2012 203
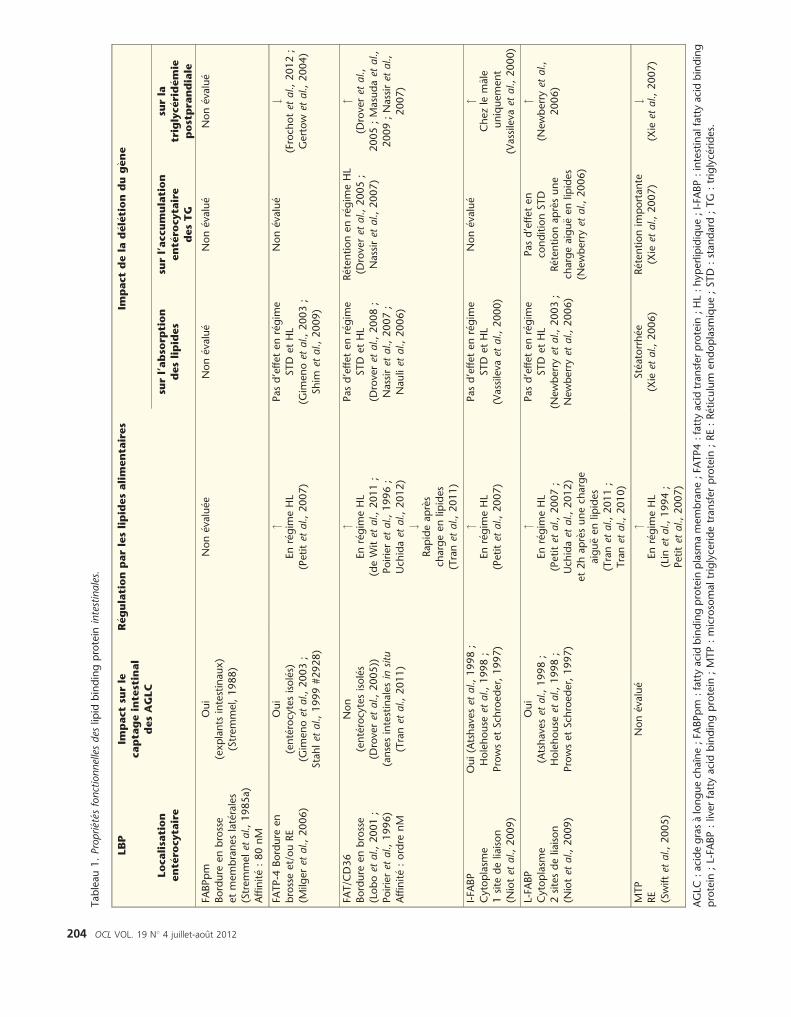
Tableau1.Propri� et� es
fonctionnellesdes
lipid
bindingprotein
intestinales.
LBP
Impactsu
rle
capta
geinte
stinal
desAGLC
R� egulationparleslipidesalimenta
ires
Impactdela
d� el� etion
dug� ene
Localisa
tion
ent� e
rocyta
ire
surl’abso
rption
deslipides
surl’accumulation
ent� e
rocyta
ire
desTG
surla
triglyc� erid� emie
postpra
ndiale
FABPpm
Bordure
enbrosse
etmembraneslat� erales
(Stremmelet
al.,1985a)
Affinit� e:80nM
Oui
(explants
intestinaux)
(Stremmel,1988)
Non� evalu� ee
Non� evalu� e
Non� evalu� e
Non� evalu� e
FATP-4
Bordure
en
brosseet/ouRE
(Milg
eret
al.,2006)
Oui
(ent� erocytesisol� es)
(Gim
enoet
al.,2003;
Stahlet
al.,1999#2928)
"Enr� egim
eHL
(Petitet
al.,2007)
Pasd’effetenr� egim
eSTD
etHL
(Gim
enoet
al.,2003;
Shim
etal.,2009)
Non� evalu� e
#(Froch
otet
al.,2012;
Gertow
etal.,2004)
FAT/C
D36
Bordure
enbrosse
(Loboet
al.,2001;
Poirieret
al.,1996)
Affinit� e:ordre
nM
Non
(ent� erocytesisol� es
(Droveret
al.,2005))
(ansesintestinalesin
situ
(Tranet
al.,2011)
"Enr� egim
eHL
(deWitet
al.,2011;
Poirieret
al.,1996;
Uch
idaet
al.,2012)
#Rapideapr� es
chargeenlip
ides
(Tranet
al.,2011)
Pasd’effetenr� egim
e
STD
etHL
(Droveret
al.,2008;
Nassiret
al.,2007;
Nau
liet
al.,2006)
R� etentionenr� egim
eHL
(Droveret
al.,2005;
Nassiret
al.,2007)
"(D
roveret
al.,
2005;Masudaet
al.,
2009;Nassiret
al.,
2007)
I-FA
BP
Cytoplasm
e1site
deliaison
(Niotet
al.,2009)
Oui(Atshaveset
al.,1998;
Holehouse
etal.,1998;
ProwsetSch
roeder,1997)
"Enr� egim
eHL
(Petitet
al.,2007)
Pasd’effetenr� egim
e
STD
etHL
(Vassileva
etal.,2000)
Non� evalu� e
"Chezle
male
uniquement
(Vassileva
etal.,2000)
L-FA
BP
Cytoplasm
e
2sitesdeliaison
(Niotet
al.,2009)
Oui
(Atshaveset
al.,1998;
Holehouse
etal.,1998;
ProwsetSch
roeder,1997)
"Enr� egim
eHL
(Petitet
al.,2007;
Uch
idaet
al.,2012)
et2hapr� esunech
arge
aigu€ eenlip
ides
(Tranet
al.,2011;
Tranet
al.,2010)
Pasd’effetenr� egim
eSTD
etHL
(Newberryet
al.,2003;
Newberryet
al.,2006)
Pasd’effeten
conditionSTD
R� etentionapr� esune
chargeaigu€ eenlip
ides
(Newberryet
al.,2006)
"(N
ewberryet
al.,
2006)
MTP
RE
(Swiftet
al.,2005)
Non� evalu� e
"Enr� egim
eHL
(Lin
etal.,1994;
Petitet
al.,2007)
St� eatorrh� ee
(Xie
etal.,2006)
R� etentionim
portante
(Xie
etal.,2007)
#(Xie
etal.,2007)
AGLC
:acidegras� alonguech
aıne;FA
BPpm
:fattyacidbindingprotein
plasm
amembrane;FA
TP4:fattyacidtransferprotein
;HL:hyperlipidique;I-FA
BP:intestinalfattyacidbinding
protein
;L-FA
BP:liverfattyacidbindingprotein
;MTP:microsomaltriglyceridetransferprotein
;RE:R� eticu
lum
endoplasm
ique;STD
:standard
;TG
:triglyc� erides.
204 OCL VOL. 19 N8 4 juillet-aout 2012

pour limiter la triglyc�erid�emie postpran-diale mais �egalement les maladiesm�etaboliques associ�ees.
Plus r�ecemment, des�etudesont rapport�eque l’adaptation g�enique observ�ee enr�egime chronique s’exercait �egalementsuite �a une seule charge en lipides(Tran et al., 2011 ; Uchida et al.,2012). Cette donn�ee d�emontre quel’adaptation ne r�esulte pas des effetsm�etaboliques secondaires au r�egimehyperlipidique chronique mais seraitdirectement d�ependante de la pr�esencedes lipides alimentaires dans la lumi�ereintestinale. En effet, un gavage avec del’huile richeenAGLCd�eclencheenmoinsde deux heures une induction du tauxd’ARNm et de la prot�eine L-FABP et dela MTP (Tran et al., 2011 ; Tran et al.,2010 ; Uchida et al., 2012). Pour exister,cette adaptation sugg�ere l’existence auniveau ent�erocytaire, d’un syst�eme ded�etection de la teneur en lipides durepas. Parmi les LBP, la r�egulation et lespropri�et�es fonctionnelles font du CD36un candidat pertinent.
D�etection ent�erocytairedes lipides alimentaires :role �emergent du CD36
Au contraire de la MTP et de la L-FABP,nous avons observ�e que la prot�eineCD36 intestinale �etait, quant �a elle,diminu�ee une heure apr�es un gavageavec de l’huile chez les rongeurs (Tranet al., 2011). Des �etudes d’immuno-histochimie ont d�emontr�e que la dispari-tion partielle du CD36 des membranesmicrovillositaires (environ 75 %) s’exerceuniquement apr�es un repas contenantdes lipides (3 % en masse). Cette dis-parition membranaire s’explique parune d�egradation par le prot�easome,lipide-d�ependante cons�ecutive �a la poly-ubiquitination du CD36 (Tran et al.,2011). Ce ph�enom�ene rapide constitueun argument suppl�ementaire montrantque le CD36 n’est pas un transporteurefficace des lipides pendant leur phasede digestion. En revanche, ce type der�egulation est souvent observ�e pour lesr�ecepteurs associ�es �a une signalisationqui correspond �a la d�esensibilisation de lacellule vis-�a-vis du ligand. Or, il a d�ej�a �et�erapport�e que la prot�eine CD36 peut�egalement jouer un role de r�ecepteurvis-�a-vis de plusieurs de ses ligands(Martin et al., 2011). C’est le cas, auniveau des macrophages pour les LDLoxyd�es, au niveau des cellules endo-
th�eliales pour les thrombospondines(Silverstein et Febbraio, 2009) et enfinau niveau des cellules r�eceptrices gusta-tives des bourgeons du gout pour lesAGLC (El-Yassimi et al., 2008 ; Laugeretteet al., 2005).Dans tous les cas, les signauxintracellulaires d�eclench�es par les ligandsdu CD36 impliquent le recrutement etl’activation des Src kinases et/ou desmitogen-activated protein kinases (MAPKs)(Silverstein et Febbraio, 2009).
En accord avec cesdonn�ees, nos r�esultatsobtenus ex vivo sur des segments intesti-naux de souris ont d�emontr�e que lesAGLC d�eclenchent apr�es cinq minutesde contact, l’activation des extracellularsignal regulated kinase 1/2 (ERK1/2)suivie par une induction du tauxd’ApoB48 (apr�es dix minutes) et de laMTP (apr�es vingt minutes). On retrouve
apr�es vingt minutes la d�egradation duCD36 concomitante avec une diminu-tion du niveau de phosphorylation desERK1/2 correspondant sans doute auph�enom�ene de d�esensibilisation (Tranet al., 2011). Cette r�egulation estd�ependante du CD36 puisqu’elle estabsente dans les segments intestinauxprovenant d’animaux d�eficients enCD36 (Tran et al., 2011). La phosphory-lation des ERK1/2 a d�ej�a �et�e rapport�eecomme �etant impliqu�ee dans la forma-tion et la s�ecr�etion des lipoprot�einesh�epatiques et intestinales (Federico et al.,2006 ; Tsai et al., 2007). Ces r�egulationsex vivo ont une pertinence physiologiquepuisque chez les souris d�eficientes enCD36, l’induction des prot�eines MTPet L-FABP observ�ee deux heures apr�esun gavage est abolie (donn�ees nonpr�esent�ees).
CD361
3
2
5
4
AGLC
Dégradationprotéasomale
Formation des groschylomicrons
Ubiquitination
Ub
Cascade designalisation
ApoB48
L-FABP
MTP
ERK1/2 P
Figure 2. Role du CD36 dans l’absorption intestinale des lipides.1) Liaison des acides gras �alongue chaıne (AGLC) issus de la digestion des triglyc�erides alimentaires avec le CD36. 2)Activation des ERK1/2. 3) et 4) Le CD36 via l’activation des ERK1/2 d�eclenche l’induction del’ApoB48, de la MTP et de la L-FABP, trois prot�eines qui optimisent la synth�ese deschylomicrons. 5) Les AGLC d�eclenchent une d�egradation du CD36 par la voie ubiquitine-prot�easome : d�esensibilisation.
OCL VOL. 19 N8 4 juillet-aout 2012 205

En conclusion, au d�ebut de la phased’absorption intestinale, l’interactionentre CD36 et les AGLC provenantde l’hydrolyse des TG alimentairesd�eclencherait une activation du m�eta-bolisme intestinal conduisant �a laformation de chylomicrons riches enTG donc de grande taille efficacementd�egrad�es dans le sang (figure 2). Ce rolede lipido-r�ecepteur pour le CD36 intes-tinal expliquerait pourquoi les animauxet les patients d�eficients en CD36s�ecr�etent de petits chylomicrons �al’origine d’une hypertriglyc�erid�emiepostprandiale (Drover et al., 2005 ;Masuda et al., 2009). CD36 est doncla premi�ere �etape d’un m�ecanismequi conduit �a une adaptation post-prandiale du m�etabolisme ent�erocytairedes lipides permettant de prendre encharge de grandes quantit�es de lipidesalimentaires.
Conclusionet perspectives
Cette revue montre que des avanc�eesimportantes ont �et�e r�ealis�ees sur lesdiff�erentes �etapes de l’absorption deslipides par la d�ecouverte progressive desfonctions des diff�erentes LBP. Cepen-dant, le role physiologique des LBPmembranaires est encore ambigumemesi les fonctions �emergentes d�emontrentque ce ne sont pas des transporteursefficaces des AGLC en p�eriode d’absorp-tion des lipides. La recherche du rolephysiologique des LBP membranairesau niveau intestinal n�ecessite la priseen compte de la sp�ecificit�e intestinale(couche d’eau non agit�ee, micellesmixtes, pr�esence de grandes quantit�esde lipides par intermittence, etc.).
Au niveau du m�ecanisme d’absorption,l’�etape de lipidation de l’ApoB48 estla moins connue et pourtant elle estcritique pour la formation de chylomi-crons bien d�egrad�es dans la circulationsanguine (taille).
Cette revuemet enexergueque l’intestinest capable d’adapter ses capacit�esd’absorption et la qualit�e des lipoprot�ei-nes s�ecr�et�ees (taille et ApoC) �a la teneuren lipides du repas. Cette adaptationsemble d�ependante de la d�etection desproduits de digestion des lipides alimen-taires par l’interm�ediaire du CD36. Cesyst�eme de sensing semble faire inter-venir un autre r�ecepteur de la memefamille que CD36, le SRB1 (voir l’articlepr�esent�e dans ce num�ero par Beaslas
et al., 2012). Ainsi une perturbation dece syst�eme de sensing au niveau desr�ecepteurs ou de leur signalisationest susceptible d’alt�erer les capacit�esd’absorption et la lip�emie postprandiale.
Finalement, ce sensing ent�erocytairepeut affecter la biodisponibilit�e de tousles nutriments ou mol�ecules lipophilesv�ehicul�espar les chylomicrons (vitaminesA, D, E, TG, cholest�erol, m�edicaments).Ces derni�eres d�ecouvertes ouvrent denouvelles perspectives th�erapeutiquesou nutritionnelles pour moduler lescapacit�es d’absorption et limiter l’hyper-triglyc�erid�emie postprandiale.
Financements : Agence Francaise dela Recherche : programme SENSOFAT.
REFERENCES
Atshaves BP, Foxworth WB, Frolov A, et al.
Cellular differentiation and I-FABP proteinexpression modulate fatty acid uptake and
diffusion. Am J Physiol 1998 ; 274 : C633-644.
Beaslas O, Cueille C, Delers F, Chateau D,Chambaz J, Rousset M, Carri�ere V. D�etection
luminale des micelles lipidiques. OCL 2012 ;
19. (Sous presse) doi : 10.1684/ocl.2012.
0448.
Berriot-Varoqueaux N, Dannoura AH,
Moreau A, et al. Apolipoprotein B48 glyco-
sylation in abetalipoproteinemia and Ander-
son’s disease. Gastroenterology 2001 ; 121 :1101-8.
Bray GA, Paeratakul S, Popkin BM. Dietary fat
and obesity: a review of animal, clinical and
epidemiological studies. Physiol Behav 2004 ;83 : 549-55.
Cartwright IJ, Higgins JA. Increased dietary
triacylglycerol markedly enhances the abilityof isolated rabbit enterocytes to secrete
chylomicrons: an effect related to dietary
fatty acid composition. J Lipid Res 1999 ; 40 :
1858-66.
Chow SL, Hollander D. Linoleic acid absorp-
tion in the unanesthetized rat: mechanism of
transport and influence of luminal factors on
absorption. Lipids 1979 ; 14 : 378-85.
Corsico B, Cistola DP, Frieden C, Storch J. The
helical domain of intestinal fatty acid binding
protein is critical for collisional transfer of fattyacids to phospholipid membranes. Proc Natl
Acad Sci U S A 1998 ; 95 : 12174-8.
de Wit NJ, Boekschoten MV, Bachmair EM,
et al.Dose-dependent effects of dietary fat ondevelopment of obesity in relation to intesti-
nal differential gene expression in C57BL/6J
mice. PLoS One 2011 ; 6 : e19145.
Defoort C, Vincent-Baudry S, LaironD. Effectsof 3-month Mediterranean-type diet on
postprandial TAG and apolipoprotein B48 in
the Medi-RIVAGE cohort. Public Health Nutr
2011 ; 14 : 2302-8.
Doege H, Stahl A. Protein-mediated fattyacid uptake: novel insights from in vivo
models. Physiology (Bethesda) 2006 ; 21 :
259-68.
Drover VA, Ajmal M, Nassir F, et al. CD36deficiency impairs intestinal lipid secretion
and clearance of chylomicrons from the
blood. J Clin Invest 2005 ; 115 : 1290-7.
Drover VA, Nguyen DV, Bastie CC, et al.CD36 mediates both cellular uptake of very
long chain fatty acids and their intestinal
absorption in mice. J Biol Chem 2008 ; 283 :13108-15.
El-Yassimi A, Hichami A, Besnard P,
Khan NA. Linoleic acid induces calcium
signaling, Src kinase phosphorylation, andneurotransmitter release in mouse CD36-
positive gustatory cells. J Biol Chem 2008 ;
283 : 12949-5.
Federico LM, Naples M, Taylor D, Adeli K.Intestinal insulin resistance and aberrant
production of apolipoprotein B48 lipopro-
teins in an animal model of insulin resistanceand metabolic dyslipidemia: evidence for
activation of protein tyrosine phosphatase-
1B, extracellular signal-related kinase, and
sterol regulatory element-binding protein-1cin the fructose-fed hamster intestine. Diabetes
2006 ; 55 : 1316-26.
Frochot V, Alqub M, Cattin AL, et al. The
transcription factor HNF-4alpha: a key factorof the intestinal uptake of fatty acids in
mouse. Am J Physiol Gastrointest Liver Physiol
2012b ; 302 : G1253-63.
Garcia-Martinez C, Marotta M, Moore-Carrasco R, et al. Impact on fatty acid
metabolism and differential localization of
FATP1 and FAT/CD36 proteins delivered incultured humanmuscle cells. Am J Physiol Cell
Physiol 2005 ; 288 : C1264-1272.
Gertow K, Bellanda M, Eriksson P, et al.
Genetic and structural evaluation of fatty acidtransport protein-4 in relation to markers
of the insulin resistance syndrome. J Clin
Endocrinol Metab 2004 ; 89 : 392-9.
Gimeno RE, Hirsch DJ, Punreddy S, et al.Targeted deletion of fatty acid transport
protein-4 results in early embryonic lethality.
J Biol Chem 2003 ; 278 : 49512-6.
Hall AM, Wiczer BM, Herrmann T, Stremmel
W, Bernlohr DA. Enzymatic properties of
purified murine fatty acid transport protein4 and analysis of acyl-CoA synthetase activi-
ties in tissues from FATP4 null mice. J Biol
Chem 2005 ; 280 : 11948-54.
Holehouse EL, Liu ML, Aponte GW. Oleic
acid distribution in small intestinal epithelialcells expressing intestinal-fatty acid binding
protein. Biochim Biophys Acta 1998 ; 1390 :
52-64.
206 OCL VOL. 19 N8 4 juillet-aout 2012

Ibrahimi A, Sfeir Z, Magharaie H, Amri EZ,
Grimaldi P, Abumrad NA. Expression of the
CD36 homolog (FAT) in fibroblast cells:effects on fatty acid transport. Proc Natl Acad
Sci U S A 1996 ; 93 : 2646-51.
Iqbal J, Hussain MM. Intestinal lipid absorp-
tion. Am J Physiol Endocrinol Metab 2009 ;296 : E1183-1194.
Jong MC, Hofker MH, Havekes LM. Role of
ApoCs in lipoprotein metabolism: functionaldifferences between ApoC1, ApoC2, and
ApoC3. Arterioscler Thromb Vasc Biol 1999 ;
19 : 472-84.
Karpe F, Olivecrona T, Hamsten A, Hultin M.
Chylomicron/chylomicron remnant turnover
in humans: evidence for margination ofchylomicrons and poor conversion of larger
to smaller chylomicron remnants. J Lipid Res
1997 ; 38 : 949-61.
Kondo H, Minegishi Y, Komine YV, et al.
Differential regulation of intestinal lipid meta-bolism-related genes in obesity-resistant A/J vs
obesity-prone C57BL/6J mice. Am J Physiol
Endocrinol Metab 2006 ; 291 : E1092-1099.
Laugerette F, Passilly-Degrace P, Patris B,
et al. CD36 involvement in orosensory detec-
tion of dietary lipids, spontaneous fat pre-ference, and digestive secretions. J Clin Invest
2005 ; 115 : 3177-84.
Lin MC, Arbeeny C, Bergquist K, Kienzle B,
Gordon DA, Wetterau JR. Cloning and
regulation of hamster microsomal triglyce-
ride transfer protein. The regulation isindependent from that of other hepatic
and intestinal proteins which participate in
the transport of fatty acids and triglycerides.
J Biol Chem 1994 ; 269 : 29138-45.
Lobo MV, Huerta L, Ruiz-Velasco N, et al.
Localization of the lipid receptors CD36 and
CLA-1/SR-BI in the human gastrointestinaltract: towards the identification of receptors
mediating the intestinal absorption of
dietary lipids. J Histochem Cytochem 2001 ;49 : 1253-60.
Mansbach 2nd CM, Gorelick F. Development
and physiological regulation of intestinallipid absorption. II. Dietary lipid absorption,
complex lipid synthesis, and the intracellular
packaging and secretion of chylomicrons. Am
J Physiol Gastrointest Liver Physiol 2007 ; 293 :G645-650.
Mariadason JM, Nicholas C, L’Italien KE, et al.
Gene expression profiling of intestinal epi-thelial cell maturation along the crypt-
villus axis. Gastroenterology 2005 ; 128 :
1081-8.
Martin C, Chevrot M, Poirier H, Passilly-
Degrace P, Niot I, Besnard P. CD36 as a lipid
sensor. Physiol Behav 2011 ; 105 : 36-42.
Martins IJ, Mortimer BC, Miller J, Redgrave
TG. Effects of particle size and number on the
plasma clearance of chylomicrons and
remnants. J Lipid Res 1996 ; 37 : 2696-705.
Masuda D, Hirano K, Oku H, et al.Chylomicron remnants are increased in the
postprandial state in CD36 deficiency. J Lipid
Res 2009 ; 50 : 999-1011.
Milger K, Herrmann T, Becker C, et al.Cellular uptake of fatty acids driven by the
ER-localized acyl-CoA synthetase FATP4. J Cell
Sci 2006 ; 119 : 4678-88.
Mora S, Rifai N, Buring JE, Ridker PM.Fasting compared with nonfasting lipids
and apolipoproteins for predicting incident
cardiovascular events. Circulation 2008 ;118 : 993-1001.
Nassir F, Wilson B, Han X, Gross RW,
Abumrad NA. CD36 is important for fattyacid and cholesterol uptake by the proximal
but not distal intestine. J Biol Chem 2007 ;
282 : 19493-501.
Nauli AM, Nassir F, Zheng S, et al. CD36 is
important for chylomicron formation and
secretion and may mediate cholesterol
uptake in the proximal intestine. Gastroente-rology 2006 ; 131 : 1197-207.
Neeli I, Siddiqi SA, Siddiqi S, et al. Liver fatty
acid-binding protein initiates budding of pre-chylomicron transport vesicles from intestinal
endoplasmic reticulum. J Biol Chem 2007 ;
282 : 17974-8.
Newberry EP, Xie Y, Kennedy S, et al.
Decreased hepatic triglyceride accumula-
tion and altered fatty acid uptake in micewith deletion of the liver fatty acid-binding
protein gene. J Biol Chem 2003 ; 278 : 51664-
72.
Newberry EP, Xie Y, Kennedy SM, Luo J,
Davidson NO. Protection against Western
diet-induced obesity and hepatic steatosis in
liver fatty acid-binding protein knockoutmice. Hepatology 2006 ; 44 : 1191-205.
Niot I, Poirier H, Tran TT, Besnard P. Intestinal
absorption of long-chain fatty acids: evidenceand uncertainties. Prog Lipid Res 2009 ; 48 :
101-15.
Olivecrona G, Beisiegel U. Lipid bindingof apolipoprotein CII is required for stimula-
tion of lipoprotein lipase activity against
apolipoprotein CII-deficient chylomicrons.Arterioscler Thromb Vasc Biol 1997 ; 17 :
1545-9.
Petit V, Arnould L, Martin P, et al. Chronichigh-fat diet affects intestinal fat absorption
and postprandial triglyceride levels in the
mouse. J Lipid Res 2007 ; 48 : 278-87.
Poirier H, Degrace P, Niot I, Bernard A,
Besnard P. Localization and regulation of the
putative membrane fatty-acid transporter
(FAT) in the small intestine. Comparison withfatty acid-binding proteins (FABP). Eur J
Biochem 1996 ; 238 : 368-73.
Prows DR, Schroeder F. Metallothionein-IIA
promoter induction alters rat intestinal fatty
acid binding protein expression, fatty aciduptake, and lipid metabolism in transfected
L-cells. Arch Biochem Biophys 1997 ; 340 :
135-43.
Ross AC. Overview of retinoid metabolism.J Nutr 1993 ; 123 : 346-50.
Shiau YF, Fernandez P, Jackson MJ,
McMonagle S. Mechanisms maintaining a
low-pH microclimate in the intestine. Am JPhysiol 1985 ; 248 : G608-617.
Shim J, Moulson CL, Newberry EP, et al. Fatty
acid transport protein 4 is dispensable forintestinal lipid absorption in mice. J Lipid Res
2009 ; 50 : 491-500.
Silverstein RL, Febbraio M. CD36, a scavenger
receptor involved in immunity, metabolism,angiogenesis, and behavior. Sci Signal 2009 ;
2 : re3.
Stahl A, Gimeno RE, Tartaglia LA, Lodish HF.
Fatty acid transport proteins: a current viewof a growing family. Trends Endocrinol Metab
2001 ; 12 : 266-73.
Stahl A, Hirsch DJ, Gimeno RE, et al. Identi-fication of the major intestinal fatty acid
transport protein.Mol Cell 1999 ; 4 : 299-308.
Steinert RE, Beglinger C. Nutrient sensing in
the gut: interactions between chemosensorycells, visceral afferents and the secretion of
satiation peptides. Physiol Behav 2011 ; 105 :
62-70.
Stremmel W. Uptake of fatty acids by jejunalmucosal cells is mediated by a fatty acid
binding membrane protein. J Clin Invest
1988 ; 82 : 2001-10.
Stremmel W, Lotz G, Strohmeyer G, Berk PD.Identification, isolation, and partial characte-
rization of a fatty acid binding protein from
rat jejunal microvillous membranes. J ClinInvest 1985a ; 75 : 1068-76.
Stremmel W, Strohmeyer G, Borchard F,
Kochwa S, Berk PD. Isolation and partial
characterization of a fatty acid-binding pro-tein in rat liver plasma membranes. Procee-
dings of the National Academy of Sciences USA
1985b ; 82 : 4-8.
Stump DD, Zhou SL, Berk PD. Comparison ofplasma membrane FABP and mitochondrial
isoform of aspartate aminotransferase from
rat liver. Am J Physiol 1993 ; 265 : G894-902.
Su X, AbumradNA. Cellular fatty acid uptake:a pathway under construction. Trends Endo-
crinol Metab 2009 ; 20 : 72-7.
Sukhotnik I, Gork AS, Chen M, Drongowski
RA, Coran AG, Harmon CM. Effect of low fat
diet on lipid absorption and fatty-acidtransport following bowel resection. Pediatr
Surg Int 2001 ; 17 : 259-64.
Swift LL, Jovanovska A, Kakkad B, Ong DE.
Microsomal triglyceride transfer protein
OCL VOL. 19 N8 4 juillet-aout 2012 207

expression in mouse intestine. Histochem Cell
Biol 2005 ; 123 : 475-82.
Tomkin GH, Owens D. The chylomicron:
relationship to atherosclerosis. Int J Vasc Med
2012 ; 2012 : 784536.
Tran TT, Poirier H, Clement L, et al. Luminal
lipid regulates CD36 levels and downstreamsignaling to stimulate chylomicron synthesis.
J Biol Chem 2011 ; 286 : 25201-10.
Tran TTT, Poirier H, Clement L, et al. CD36Displays Features of a Lipid-Sensor Involved
in Chylomicron Processing in the Rodent
Small Intestine. Atheroscler Suppl 2010 ; 11 :
82-3.
Tsai J, Qiu W, Kohen-Avramoglu R, Adeli K.
MEK-ERK inhibition corrects the defect in
VLDL assembly in HepG2 cells: potential roleof ERK in VLDL-ApoB100 particle assembly.
Arterioscler Thromb Vasc Biol 2007 ; 27 :
211-8.
Tso P, Liu M. Apolipoprotein A-IV, food
intake, and obesity. Physiol Behav 2004 ;83 : 631-43.
Uchida A, Whitsitt MC, Eustaquio T, et al.
Reduced triglyceride secretion in response to
an acute dietary fat challenge in obesecompared to lean mice. Front Physiol 2012 ;
3 : 26.
Vassileva G, Huwyler L, Poirier K, Agellon LB,
Toth MJ. The intestinal fatty acid bindingprotein is not essential for dietary fat
absorption in mice. FASEB J 2000 ; 14 :
2040-6.
Weinberg RB, Gallagher JW, Fabritius MA,
Shelness GS. ApoA-IV modulates the secre-
tory trafficking of apoB and the size of
triglyceride-rich lipoproteins. J Lipid Res 2012 ;
53 : 736-43.
Xiang SQ, Cianflone K, Kalant D, Sniderman
AD. Differential binding of triglyceride-rich
lipoproteins to lipoprotein lipase. J Lipid Res
1999 ; 40 : 1655-63.
Xie Y, Luo J, Kennedy S, Davidson NO.
Conditional intestinal lipotoxicity in Apobec-
1-/- Mttp-IKO mice: a survival advantage
for mammalian intestinal apolipoprotein BmRNA editing. J Biol Chem 2007 ; 282 :
33043-51.
Xie Y, Newberry EP, Young SG, et al.Compensatory increase in hepatic lipogene-
sis in mice with conditional intestine-specific
Mttp deficiency. J Biol Chem 2006 ; 281 :
4075-86.
208 OCL VOL. 19 N8 4 juillet-aout 2012
Related Documents











