MCP-1 deficiency causes altered inflammation with impaired skeletal muscle regeneration Paula K. Shireman,* ,†,‡,§,1 Vero ´ nica Contreras-Shannon, Oscar Ochoa, † Bijal P. Karia, † Joel E. Michalek, ¶ and Linda M. McManus §,#, ** *South Texas Veterans Health Care System, San Antonio, Texas, USA; and Departments of † Surgery, ‡ Medicine, Cellular & Structural Biology, ¶ Epidemiology and Biostatistics, # Periodontics, and **Pathology and § Sam and Ann Barshop Institute for Longevity and Aging Studies, University of Texas Health Science Center, San Antonio, Texas, USA Abstract: We examined the role of MCP-1, a po- tent chemotactic and activating factor for macro- phages, in perfusion, inflammation, and skeletal mus- cle regeneration postischemic injury. MCP-1/ or C57Bl/6J control mice [wild-type (WT)] underwent femoral artery excision (FAE). Muscles were col- lected for histology, assessment of tissue chemokines, and activity measurements of lactate dehydrogenase (LDH) and myeloperoxidase. In MCP-1/ mice, restoration of perfusion was delayed, and LDH and fiber size, indicators of muscle regeneration, were decreased. Altered inflammation was observed with increased neutrophil accumulation in MCP-1/ versus WT mice at Days 1 and 3 (P<0.003), whereas fewer macrophages were present in MCP-1/ mice at Day 3. As necrotic tissue was removed in WT mice, macrophages decreased (Day 7). In contrast, macro- phage accumulation in MCP-1/ was increased in association with residual necrotic tissue and impaired muscle regeneration. Consistent with altered inflam- mation, neutrophil chemotactic factors (keratino- cyte-derived chemokine and MIP-2) were increased at Day 1 post-FAE. The macrophage chemotactic factor MCP-5 was increased significantly in WT mice at Day 3 compared with MCP-1/ mice. However, at post-FAE Day 7, MCP-5 was elevated significantly in MCP-1/ mice versus WT mice. Addition of exogenous MCP-1 did not induce proliferation in murine myoblasts (C2C12 cells) in vitro. MCP-1 is essential for reperfusion and the successful comple- tion of normal skeletal muscle regeneration after ischemic tissue injury. Impaired muscle regeneration in MCP-1/ mice suggests an important role for macrophages and MCP-1 in tissue reparative processes. J. Leukoc. Biol. 81: 000 – 000; 2007. Key Words: CCL2 macrophage neutrophils chemokines isch- emia INTRODUCTION Acute and chronic tissue ischemia is a common complication of atherosclerosis, embolic disease, and traumatic injury. De- spite treatment, these conditions often result in lower extremity amputations. New and adjuvant therapies for limb salvage will probably derive from a better understanding of the mechanisms involved in postischemic collateral artery development and skeletal muscle regeneration. Macrophages and inflammation are important components in collateral artery formation [1] and skeletal muscle regeneration [2– 4] via biological responses that are incompletely under- stood. MCP-1 (also known as CCL2), through its receptor CCR2, is a chemokine with potent macrophage recruitment and activating functions [5, 6]. In wild-type (WT) mice, within 3 days after femoral artery excision (FAE), MCP-1 protein levels were elevated maximally in ischemic calf muscle in association with a robust macrophage infiltrate; however, in the thigh, where arteriogenesis occurs, MCP-1 did not increase [7]. Thus, MCP-1 is probably involved in the modulation of mac- rophage responses following ischemia. Previous studies have documented MCP-1 involvement in the restoration of perfusion after the induction of hind-limb ischemia [8 –11]; however, little is known about the role of MCP-1 in skeletal muscle regeneration following ischemic injury. Injured muscle secretes factors that are chemotactic for macrophages and other inflammatory cells [12], essential cells in the removal of damaged or necrotic tissue [13]. Further, macrophages enhance proliferation and delay differentiation [2] of satellite cells, the multipotent myogenic stem cells that are integral to regenerating damaged muscle [14]. Myogenic precursor cells can differentiate into skeletal mus- cle as well as transdifferentiate into adipocytes [15], osteo- blasts [16], and endothelial cells [17]. The role that inflamma- tion and perhaps MCP-1 plays in modulating the plasticity of myogenic stem cell growth and differentiation remains largely unknown but may be an important modifier in the skeletal muscle response to ischemia. Given the essential roles of macrophages in muscle regen- eration [2, 4] and collateral artery formation [1], we hypothe- 1 Correspondence: Departments of Surgery and Medicine and Sam and Ann Barshop Institute for Longevity and Aging Studies, University of Texas Health Science Center, MSC 7741, 7703 Floyd Curl Drive, San Antonio, TX 78229- 3900. E-mail: [email protected] Received May 23, 2006; revised October 31, 2006; accepted November 2, 2006. doi: 10.1189/jlb.0506356 0741-5400/07/0081-0001 © Society for Leukocyte Biology Journal of Leukocyte Biology Volume 81, March 2007 1 Uncorrected Version. Published on November 29, 2006 as DOI:10.1189/jlb.0506356 Copyright 2006 by The Society for Leukocyte Biology.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

MCP-1 deficiency causes altered inflammation with impairedskeletal muscle regeneration
Paula K. Shireman,*,†,‡,§,1 Veronica Contreras-Shannon,�� Oscar Ochoa,†
Bijal P. Karia,† Joel E. Michalek,¶ and Linda M. McManus§,#,***South Texas Veterans Health Care System, San Antonio, Texas, USA; and Departments of †Surgery, ‡Medicine,��Cellular & Structural Biology, ¶Epidemiology and Biostatistics, #Periodontics, and **Pathology and §Sam andAnn Barshop Institute for Longevity and Aging Studies, University of Texas Health Science Center,San Antonio, Texas, USA
Abstract: We examined the role of MCP-1, a po-tent chemotactic and activating factor for macro-phages, in perfusion, inflammation, and skeletal mus-cle regeneration postischemic injury. MCP-1�/� orC57Bl/6J control mice [wild-type (WT)] underwentfemoral artery excision (FAE). Muscles were col-lected for histology, assessment of tissue chemokines,and activity measurements of lactate dehydrogenase(LDH) and myeloperoxidase. In MCP-1�/� mice,restoration of perfusion was delayed, and LDH andfiber size, indicators of muscle regeneration, weredecreased. Altered inflammation was observed withincreased neutrophil accumulation in MCP-1�/�versus WT mice at Days 1 and 3 (P<0.003), whereasfewer macrophages were present in MCP-1�/� miceat Day 3. As necrotic tissue was removed in WT mice,macrophages decreased (Day 7). In contrast, macro-phage accumulation in MCP-1�/� was increased inassociation with residual necrotic tissue and impairedmuscle regeneration. Consistent with altered inflam-mation, neutrophil chemotactic factors (keratino-cyte-derived chemokine and MIP-2) were increasedat Day 1 post-FAE. The macrophage chemotacticfactor MCP-5 was increased significantly in WT miceat Day 3 compared with MCP-1�/� mice. However,at post-FAE Day 7, MCP-5 was elevated significantlyin MCP-1�/� mice versus WT mice. Addition ofexogenous MCP-1 did not induce proliferation inmurine myoblasts (C2C12 cells) in vitro. MCP-1 isessential for reperfusion and the successful comple-tion of normal skeletal muscle regeneration afterischemic tissue injury. Impaired muscle regenerationin MCP-1�/� mice suggests an important role formacrophages and MCP-1 in tissue reparativeprocesses. J. Leukoc. Biol. 81: 000–000; 2007.
Key Words: CCL2 � macrophage � neutrophils � chemokines � isch-emia
INTRODUCTION
Acute and chronic tissue ischemia is a common complicationof atherosclerosis, embolic disease, and traumatic injury. De-
spite treatment, these conditions often result in lower extremityamputations. New and adjuvant therapies for limb salvage willprobably derive from a better understanding of the mechanismsinvolved in postischemic collateral artery development andskeletal muscle regeneration.
Macrophages and inflammation are important components incollateral artery formation [1] and skeletal muscle regeneration[2–4] via biological responses that are incompletely under-stood. MCP-1 (also known as CCL2), through its receptorCCR2, is a chemokine with potent macrophage recruitmentand activating functions [5, 6]. In wild-type (WT) mice, within3 days after femoral artery excision (FAE), MCP-1 proteinlevels were elevated maximally in ischemic calf muscle inassociation with a robust macrophage infiltrate; however, in thethigh, where arteriogenesis occurs, MCP-1 did not increase [7].Thus, MCP-1 is probably involved in the modulation of mac-rophage responses following ischemia.
Previous studies have documented MCP-1 involvement inthe restoration of perfusion after the induction of hind-limbischemia [8–11]; however, little is known about the role ofMCP-1 in skeletal muscle regeneration following ischemicinjury. Injured muscle secretes factors that are chemotactic formacrophages and other inflammatory cells [12], essential cellsin the removal of damaged or necrotic tissue [13]. Further,macrophages enhance proliferation and delay differentiation[2] of satellite cells, the multipotent myogenic stem cells thatare integral to regenerating damaged muscle [14].
Myogenic precursor cells can differentiate into skeletal mus-cle as well as transdifferentiate into adipocytes [15], osteo-blasts [16], and endothelial cells [17]. The role that inflamma-tion and perhaps MCP-1 plays in modulating the plasticity ofmyogenic stem cell growth and differentiation remains largelyunknown but may be an important modifier in the skeletalmuscle response to ischemia.
Given the essential roles of macrophages in muscle regen-eration [2, 4] and collateral artery formation [1], we hypothe-
1 Correspondence: Departments of Surgery and Medicine and Sam and AnnBarshop Institute for Longevity and Aging Studies, University of Texas HealthScience Center, MSC 7741, 7703 Floyd Curl Drive, San Antonio, TX 78229-3900. E-mail: [email protected]
Received May 23, 2006; revised October 31, 2006; accepted November 2,2006.
doi: 10.1189/jlb.0506356
0741-5400/07/0081-0001 © Society for Leukocyte Biology Journal of Leukocyte Biology Volume 81, March 2007 1
Uncorrected Version. Published on November 29, 2006 as DOI:10.1189/jlb.0506356
Copyright 2006 by The Society for Leukocyte Biology.

sized that MCP-1 may have a vital function in skeletal muscleresponses to ischemia via recruitment and activation of mac-rophages. The present study was conducted to explore therelationship among the restoration of perfusion, inflammation,and skeletal muscle regeneration post-FAE in mice lackingMCP-1. Novel findings in this report include impairments inskeletal muscle regeneration and dysregulated inflammation inMCP-1�/� mice.
MATERIALS AND METHODS
Experimental animals
MCP-1�/� mice, on a C57Bl/6J background, were derived as describedpreviously [5] (original mice for the MCP-1�/� breeding colony were a kindgift from Barrett Rollins) and backcrossed to C57Bl/6J mice from Jackson Labs(Bar Harbor, ME) for six generations. MCP-1�/� mice, on the C57Bl/6Jbackground, were bred at the Audie Murphy Veterans Hospital (San Antonio,TX), and C57Bl/6J WT control mice were purchased from Jackson Labs. Malemice, 10–20 weeks old, were used in this study. All procedures complied withthe National Institutes of Health (NIH) Animal Care and Use Guidelines andwere approved by the Institutional Animal Care and Use Committee of theUniversity of Texas Health Science Center at San Antonio and at the SouthTexas Veterans Health Care System (San Antonio).
Mouse FAE model and laser doppler imaging(LDI)
Male mice [MCP-1�/� or C57Bl/6J (WT)] underwent FAE. Mice were anes-thetized with an i.p. injection of pentobarbital (2 mg; Abbott Laboratories,Chicago, IL) or 2–3% isoflurane with oxygen for FAE and LDI. The femoralartery was excised from the inguinal ligament to just proximal to the bifurcationof the popliteal and saphena arteries as described previously [18]. In allanimals, the femoral vein and nerve were preserved, and a sham surgery wasperformed on the left leg. LDI (Moor LDI, Moor Instruments, Wilmington, DE)was performed sequentially on 20 mice/strain; each animal was imaged at alltime-points, as described previously [18].
Preparation of tissue lysates
After right FAE and left sham surgery (Days 1, 3, 7, and 14; n�5/time-point),mice were killed humanely, and all of the muscles of the hind limb from theknee to the ankle (designated as “calf”) or from the inguinal ligament to theknee (designated as thigh) were removed, weighed, and used immediately inpreparation of tissue lysates, as described previously [7]. Similar samples werealso collected from animals not subjected to surgery and served as negativecontrols (baseline). Briefly, on ice, muscles were minced and mixed, and aweighed portion was homogenized (Tissumizer, Tekmar, Cincinnati, OH) on icefor 15 s in 1.5 ml lysate buffer, i.e., 50 mM Tris buffer (pH 7.4) containing 250mM NaCl, 1% Nonidet P-40 (Roche Applied Science, Indianapolis, IN), 50mM NaF, 2 mM Na3VO4, 5.5 mM EDTA, 1 mM 4-(2-aminoethyl) benzenesul-fonyl fluoride hydrochloride, 65 �M bestatin hydrochloride, 7 �M transepoxy-succinyl-L-leucylamido-(4-guanidino) butane, 11 �M leupeptin, 0.15 �Maprotinin, and 1 mM PMSF (from Sigma-Aldrich, St. Louis, MO, unlessotherwise specified). The tissue homogenate was then centrifuged immediately(4400 g, 5 min, 4°C), and the supernatant was removed and centrifuged further(2300 g, 5 min, 4°C). Aliquots of the final supernatant were stored at –80°C.
Measurement of tissue levels of MCP-5, MIP-2,MIP-1�, and keratinocyte-derived chemokine(KC) and enzymatic activity of lactatedehydrogenase (LDH) and myeloperoxidase(MPO)
Tissue lysates were thawed on ice for 15 min and centrifuged immediately(3300–16,100 g, 4°C, 10 min). Protein was determined by the Pierce BCA™protein assay (Pierce Biotechnology, Inc., Rockford, IL) using a microtiter
plate format; BSA from ICN Biomedicals Inc. (Costa Mesa, CA) in lysate bufferwas used as the standard. Absorption in all microtiter plate assays wasmonitored in a SpectraMax Plus plate reader (Molecular Devices, Sunnyvale,CA), and results were analyzed with SOFTmax PRO software (MolecularDevices).
MCP-5, MIP-2, MIP-1�, and KC levels in the tissue lysates were assessedby an ELISA by R&D Systems (Minneapolis, MN), according to the manufac-turer’s protocol with slight modifications. Standards and unknowns were di-luted in lysate buffer, and results were expressed as pg/mg protein.
Reaction rates (Vmax) for LDH and MPO were determined as describedpreviously [7]. LDH and MPO activities were normalized to mg proteinmeasured in each lysate. Data for LDH, MPO, muscle weight, and protein/weight for WT mice have been reported previously [7] and were included herefor purposes of comparison with results from MCP-1�/� mice. Data for WTand MCP-1�/� mice were obtained in parallel experiments.
Histologic studies
After right FAE and left sham surgery, mice were killed humanely after 1, 3,7, 14, 21, or 28 days, and hind-limb tissues were collected for histology; n �4–12 mice/time-point/strain. After the i.p. injection of heparin (250 U) andpentobarbital (3 mg), the abdominal aorta was cannulated and pressure-perfused as described previously [7]. Hind limbs were removed at the inguinalligament and placed in 10% neutral-buffered formalin (NBF) for 1–2 h.Complete cross-sections of each leg at the calf or thigh (including bone,muscle, and skin) were then obtained and decalcified prior to routine paraffinembedding. All paraffin-embedded specimens were sectioned (6 �m) andstained with H&E prior to light microscopic examination. In a separate groupof animals not pressure-perfused, tissues from the anterior and posteriorcompartments of the lower leg were removed en bloc, transected axiallythrough the mid-portion of the muscle belly, fixed in 10% NBF, and paraffin-embedded.
Immunohistochemical studies
Routine, indirect immunohistochemical procedures were used to localizemonocytes/macrophages (F4/80 and mac3) or neutrophils (Ly-6) in deparaf-finized sections. Nonspecific binding of antibodies was blocked by treatment ofsections (30 min) with 1% BSA in PBS. Primary antibodies (diluted in 1% BSAin PBS) were rat monoclonal antimouse F4/80 (Serotec, Inc., Raleigh, NC),antimouse mac3 (BD Biosciences PharMingen, San Diego, CA), and antimouseLy-6 (BD Biosciences PharMingen). Biotinylated secondary antibodies(mouse-absorbed rabbit antirat IgG) and streptavidin-HRP were obtained fromVector Laboratories (Burlingame, CA). After enzymatic development in dia-minobenzidine and hydrogen peroxide, sections were counterstained withhematoxylin.
Histology and immunohistochemistry images were captured with a NikonE600 Eclipse microscope (Nikon, Melville, NY), equipped with a high-reso-lution digital camera (Nikon DXM 1200), interfaced to a personal computerequipped with Act-1 (Nikon) networking software for image-capture and ar-chiving.
Histomorphometric measurement ofintermuscular fat and muscle fiber area
Morphometric analyses were performed on images captured with a NikonEclipse TE2000-U inverted microscope, equipped with a high-resolution dig-ital camera (Nikon DXM 1200F), interfaced with a personal computer,equipped with Metamorph (Nikon) software. Using the anterior compartmentspecimen removed en bloc at 21 days post-FAE, 2–6 �m cross-sections wereobtained through the mid-portion of the muscle and stained with H&E ortrichrome. Preliminary studies demonstrated no significant difference in fibercross-sectional area between H&E and trichrome-stained sections (data notshown). The 21-day time-point was chosen for morphometric analysis, as onlysmall foci of necrotic muscle were present occasionally post-FAE in bothmouse strains. The tibialis anterior (TA), the largest and most superficialmuscle in the anterior compartment, was identified and used for all histomor-phometric measurements. The TA was chosen, as it consistently underwentnecrosis and regeneration, and muscles in the posterior compartment, such asthe gastrocnemious muscle, exhibited various amounts of necrosis and regen-eration [7]. For TA specimen following FAE, three different microscopic
2 Journal of Leukocyte Biology Volume 81, March 2007 http://www.jleukbio.org

sections separated by at least 100 �m were used for morphometric analysis,and one microscopic section was used for sham surgery. Within each TAsection, eight nonoverlapping areas (20� magnification) were selected ran-domly and digitally captured; areas containing large blood vessels or fibrousbands between muscle bundles were excluded.
In each of the digitally captured images, intermuscular adipocytes wereoutlined manually and divided by the total image area to calculate the percentfat. The average percent fat area of eight images for each TA specimen wasthen determined. The average percent fat among replicate TA sections for eachanimal was calculated for subsequent use in comparisons of results for eachgroup of animals.
The average cross-sectional area (�m2) of individual muscle fibers for agiven animal was determined after manual outlining of individual muscle fibersin each of the digitally captured images for a given TA; fibers, which wereincluded only partially within the images, were excluded. In the TA of FAElimbs, only regenerated fibers (identified by the presence of a centrally locatednucleus [19]) were measured, whereas fibers with peripherally located nuclei(i.e., mature, nonregenerated fibers) were measured in the TA of the shamsurgery limbs.
Cell culture
Mouse C2C12 myoblasts [American Type Culture Collection (ATCC), Manas-sas, VA] were cultured in growth media (GM) containing DMEM (ATCC),supplemented with 10% FBS (Hyclone, Logan, UT) and 1% penicillin-strep-tomycin (Mediatech, Inc., Herndon, VA) at 37°C in a humidified atmosphereof 5% CO2 in air. Cells were allowed to reach 60% confluency before passage.
Cell proliferation
Cell proliferation was measured by the incorporation of BrdU using the cellproliferation ELISA BrdU colorimetric kit (Roche, Mannheim, Germany).C2C12 cells were seeded in 96-well plates (Falcon, Becton Dickinson, Frank-lin Lakes, NJ) in GM. After 24 h, the GM was removed, cells were washed withPBS, and the proliferation assay was performed under two different cultureconditions: 4000 cells/well were grown in serum-free media (SFM); DMEM,supplemented with 5 �g/ml transferrin (Sigma-Aldrich) and 0.2 �g/ml sele-nium (Sigma-Aldrich) or 2000 cells/well, were grown in DMEM supplementedwith 2% FBS. Cytokines or FBS were added after 2 h for SFM and after 24 hfor 2% FBS. The cytokines and FBS additions were as follows: as the negativecontrol, 1% BSA in PBS, the murine cytokine fibroblast growth factor 2 (FGF2)or MCP-1 (both from R&D Systems) in 1% BSA in PBS at 1, 10, or 100 ng/mland 20% FBS as the positive control. Cells were incubated with cytokines orFBS for 24 h or 48 h, followed by 2 h incubation with BrdU. After removingthe culture media, cells were fixed, and the plates were processed per manu-facturer protocol. After a 10-min incubation, absorbance was measured at 370nm using a SpectraMax Plus plate reader. Results for each experiment wereexpressed as fold change compared with the negative control (1% BSA in PBS),and the fold changes for three independent experiments were then averaged.
Data analysis
SAS software, Version 9.1 (SAS, Cary, NC), was used for all statisticalanalyses. Results were expressed as mean � SEM, and statistical significancewas set at the level of 5%.
Sequentially derived LDI ratios were analyzed by two different methods.First, a Bonferroni corrected multiple comparisons procedure was used todetermine if LDI ratios within a given group had returned to preoperative(baseline) levels. Data collected preoperatively were subtracted from each ofthe other time-points to produce a dataset of comparable LDI ratio differences.Second, an ANOVA incorporating repeated measures across time was usedwith a Bonferroni correction for multiple comparisons to determine if therewere differences between MCP-1�/� and WT mice at each time-point. Withinanimal, time correlations and heterogeneity of the variances across time weremodeled using the autoregressive order one-covariance structure. Animal wasconsidered a random effect.
Weight, protein/weight, chemokine levels, and enzyme activity were ana-lyzed by two different methods. First, a Dunnett-corrected multiple compari-sons procedure, using a two-way ANOVA of least-square means to determinewhether significant differences existed, was used to compare each time-point tobaseline values for each mouse strain. Second, a Hochberg or Bonferroni-
corrected two-way ANOVA was used to determine if there were differencesbetween MCP-1�/� and WT mice at each time-point. The MPO and chemo-kine data were analyzed in a similar manner, except a log transformation wasapplied to the data prior to analysis to adjust for unequal variances. For lysatesamples with chemokines below the level of detection in the ELISA (MIP-2,7.8; MIP-1�, 4.7 and 15.625 pg/ml for MCP-5 and KC), a value of lowestdetectable level/square root of 2 pg/ml was assigned to these samples [20], andthis value was corrected for the protein in each specimen.
Histomorphometry data for percent fat and fiber cross-sectional area wereanalyzed as follows. First, a repeated measures linear model was used todetermine that there were no significant interactions among the three differentlevels of the TA muscle. Replicate data for all images and levels were thenaveraged to generate a single number for each limb for each mouse—FAE orsham limb for fiber cross-sectional area and FAE-only for percent fat. Pairedt-tests were used to assess the significance of mean differences in fibercross-sectional area between FAE versus sham limbs within each strain.One-way analyses of variance were used to assess the significance of meandifferences between MCP-1�/� and WT mice for FAE (fiber cross-sectionalarea and percent fat) or sham (fiber cross-sectional area) limbs.
Prior to analysis of C2C12 proliferation data, the raw OD was background-corrected by subtracting the mean OD from blank wells and then divided bythe mean OD from the negative control wells. ANOVA was used to contrasttreatment and negative control OD means. The experiment-wide significancelevel was fixed at 5% using Dunnett’s method for repeated two-sided compar-isons with the control.
RESULTS
Restoration of perfusion post-FAE
Before FAE, perfusion was equivalent in the right and left legsof WT and MCP-1�/� mice. Immediately after FAE, perfu-sion decreased to �15% of that in the contralateral leg in bothgroups and remained unchanged through Day 3 (Fig. 1). InWT mice, perfusion increased by Day 7 and improved steadilythereafter. In contrast, there was a delay in the restoration ofperfusion in the MCP-1�/� mice and a significant decrease inthe perfusion ratio at Day 7 (P�0.01) as compared with WTanimals. In MCP-1�/� and WT mice, there was a significantdifference at all post-FAE time-points (P�0.0001) as com-
Fig. 1. Restoration of perfusion in MCP-1�/� and WT mice post-FAE. LDIperfusion ratios for MCP-1�/� and WT mice were measured sequentially foreach mouse at all time-points. For both groups, there were significant differ-ences at all time-points compared with pre-excision LDI measurements(P�0.001). Results represent the mean � SEM; n � 20 mice/strain. #, P �0.01, between MCP-1�/� and WT groups.
Shireman et al. Impaired muscle regeneration in MCP-1–/– mice 3
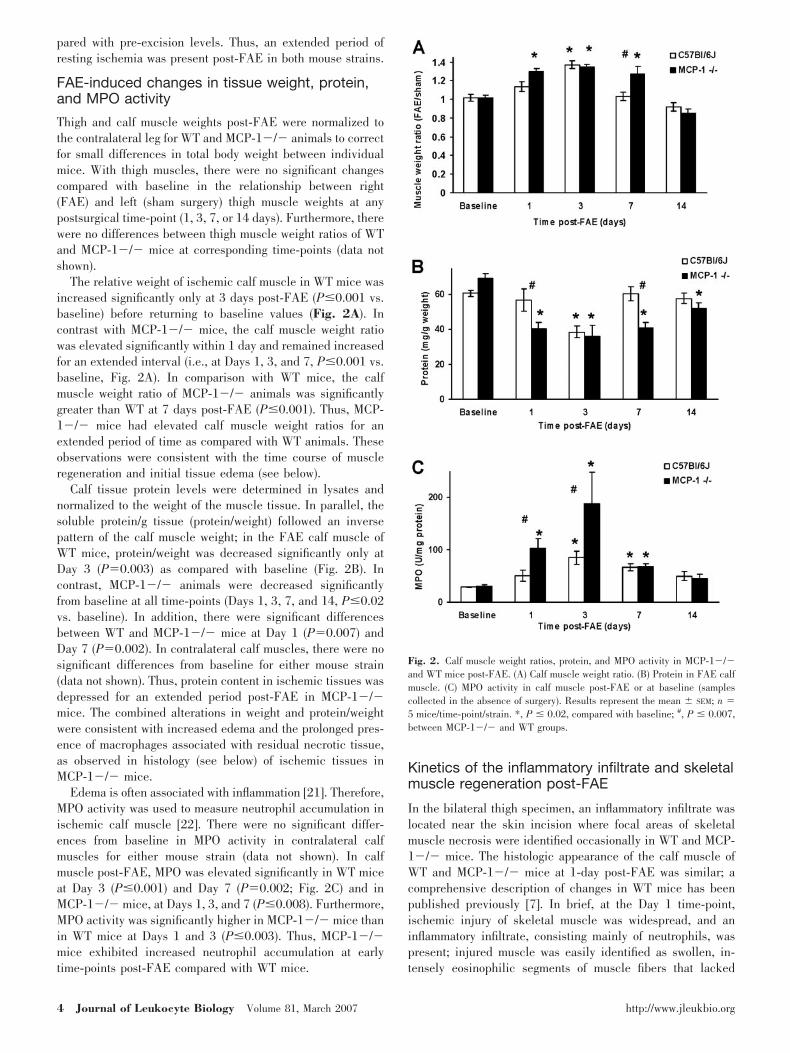
pared with pre-excision levels. Thus, an extended period ofresting ischemia was present post-FAE in both mouse strains.
FAE-induced changes in tissue weight, protein,and MPO activity
Thigh and calf muscle weights post-FAE were normalized tothe contralateral leg for WT and MCP-1�/� animals to correctfor small differences in total body weight between individualmice. With thigh muscles, there were no significant changescompared with baseline in the relationship between right(FAE) and left (sham surgery) thigh muscle weights at anypostsurgical time-point (1, 3, 7, or 14 days). Furthermore, therewere no differences between thigh muscle weight ratios of WTand MCP-1�/� mice at corresponding time-points (data notshown).
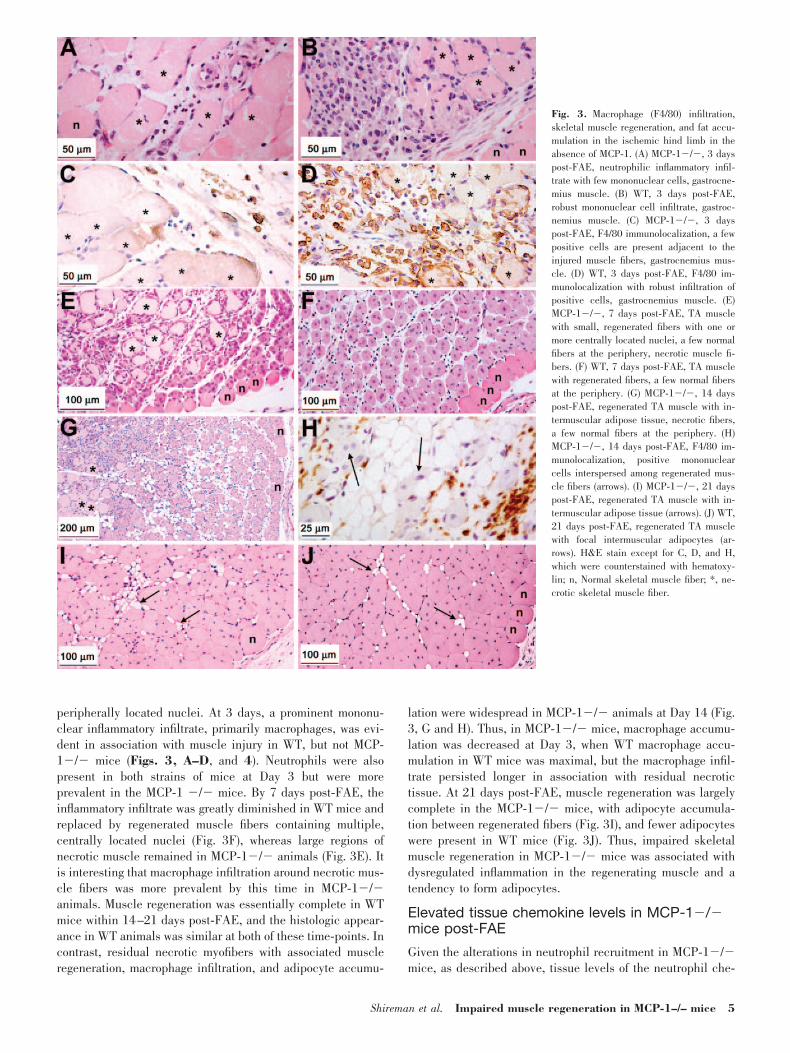
The relative weight of ischemic calf muscle in WT mice wasincreased significantly only at 3 days post-FAE (P�0.001 vs.baseline) before returning to baseline values (Fig. 2A). Incontrast with MCP-1�/� mice, the calf muscle weight ratiowas elevated significantly within 1 day and remained increasedfor an extended interval (i.e., at Days 1, 3, and 7, P�0.001 vs.baseline, Fig. 2A). In comparison with WT mice, the calfmuscle weight ratio of MCP-1�/� animals was significantlygreater than WT at 7 days post-FAE (P�0.001). Thus, MCP-1�/� mice had elevated calf muscle weight ratios for anextended period of time as compared with WT animals. Theseobservations were consistent with the time course of muscleregeneration and initial tissue edema (see below).
Calf tissue protein levels were determined in lysates andnormalized to the weight of the muscle tissue. In parallel, thesoluble protein/g tissue (protein/weight) followed an inversepattern of the calf muscle weight; in the FAE calf muscle ofWT mice, protein/weight was decreased significantly only atDay 3 (P�0.003) as compared with baseline (Fig. 2B). Incontrast, MCP-1�/� animals were decreased significantlyfrom baseline at all time-points (Days 1, 3, 7, and 14, P�0.02vs. baseline). In addition, there were significant differencesbetween WT and MCP-1�/� mice at Day 1 (P�0.007) andDay 7 (P�0.002). In contralateral calf muscles, there were nosignificant differences from baseline for either mouse strain(data not shown). Thus, protein content in ischemic tissues wasdepressed for an extended period post-FAE in MCP-1�/�mice. The combined alterations in weight and protein/weightwere consistent with increased edema and the prolonged pres-ence of macrophages associated with residual necrotic tissue,as observed in histology (see below) of ischemic tissues inMCP-1�/� mice.
Edema is often associated with inflammation [21]. Therefore,MPO activity was used to measure neutrophil accumulation inischemic calf muscle [22]. There were no significant differ-ences from baseline in MPO activity in contralateral calfmuscles for either mouse strain (data not shown). In calfmuscle post-FAE, MPO was elevated significantly in WT miceat Day 3 (P�0.001) and Day 7 (P�0.002; Fig. 2C) and inMCP-1�/� mice, at Days 1, 3, and 7 (P�0.008). Furthermore,MPO activity was significantly higher in MCP-1�/� mice thanin WT mice at Days 1 and 3 (P�0.003). Thus, MCP-1�/�mice exhibited increased neutrophil accumulation at earlytime-points post-FAE compared with WT mice.
Kinetics of the inflammatory infiltrate and skeletalmuscle regeneration post-FAE
In the bilateral thigh specimen, an inflammatory infiltrate waslocated near the skin incision where focal areas of skeletalmuscle necrosis were identified occasionally in WT and MCP-1�/� mice. The histologic appearance of the calf muscle ofWT and MCP-1�/� mice at 1-day post-FAE was similar; acomprehensive description of changes in WT mice has beenpublished previously [7]. In brief, at the Day 1 time-point,ischemic injury of skeletal muscle was widespread, and aninflammatory infiltrate, consisting mainly of neutrophils, waspresent; injured muscle was easily identified as swollen, in-tensely eosinophilic segments of muscle fibers that lacked
Fig. 2. Calf muscle weight ratios, protein, and MPO activity in MCP-1�/�and WT mice post-FAE. (A) Calf muscle weight ratio. (B) Protein in FAE calfmuscle. (C) MPO activity in calf muscle post-FAE or at baseline (samplescollected in the absence of surgery). Results represent the mean � SEM; n �5 mice/time-point/strain. *, P � 0.02, compared with baseline; #, P � 0.007,between MCP-1�/� and WT groups.
4 Journal of Leukocyte Biology Volume 81, March 2007 http://www.jleukbio.org
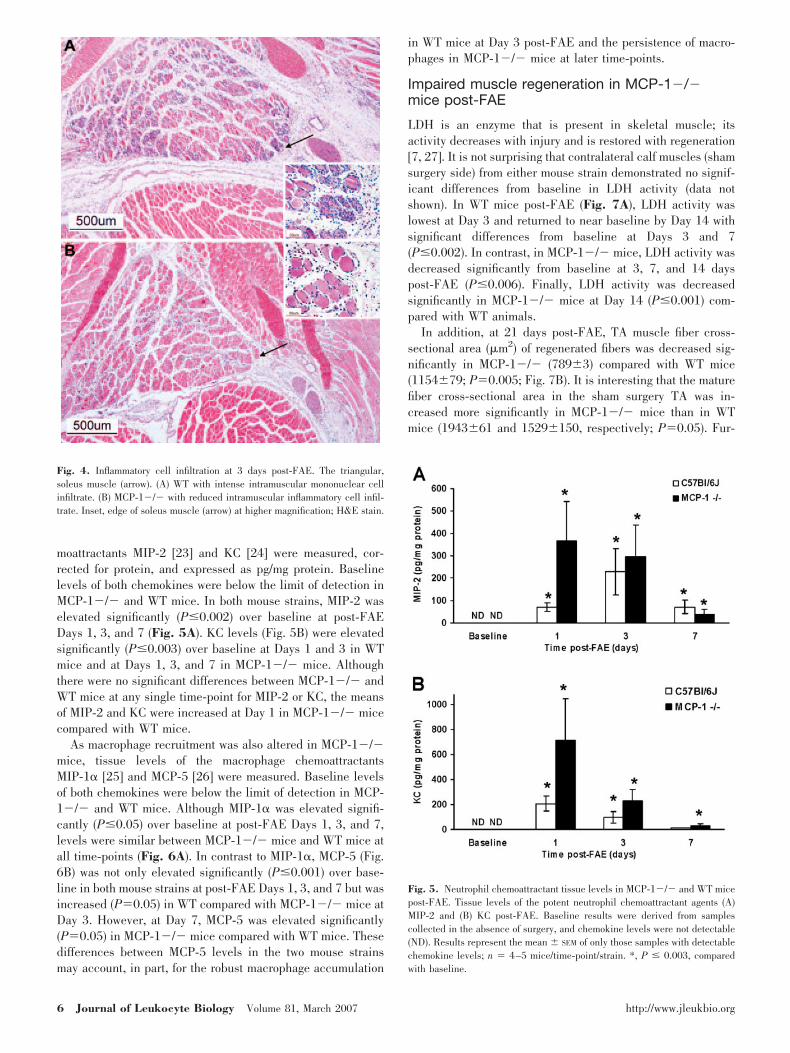
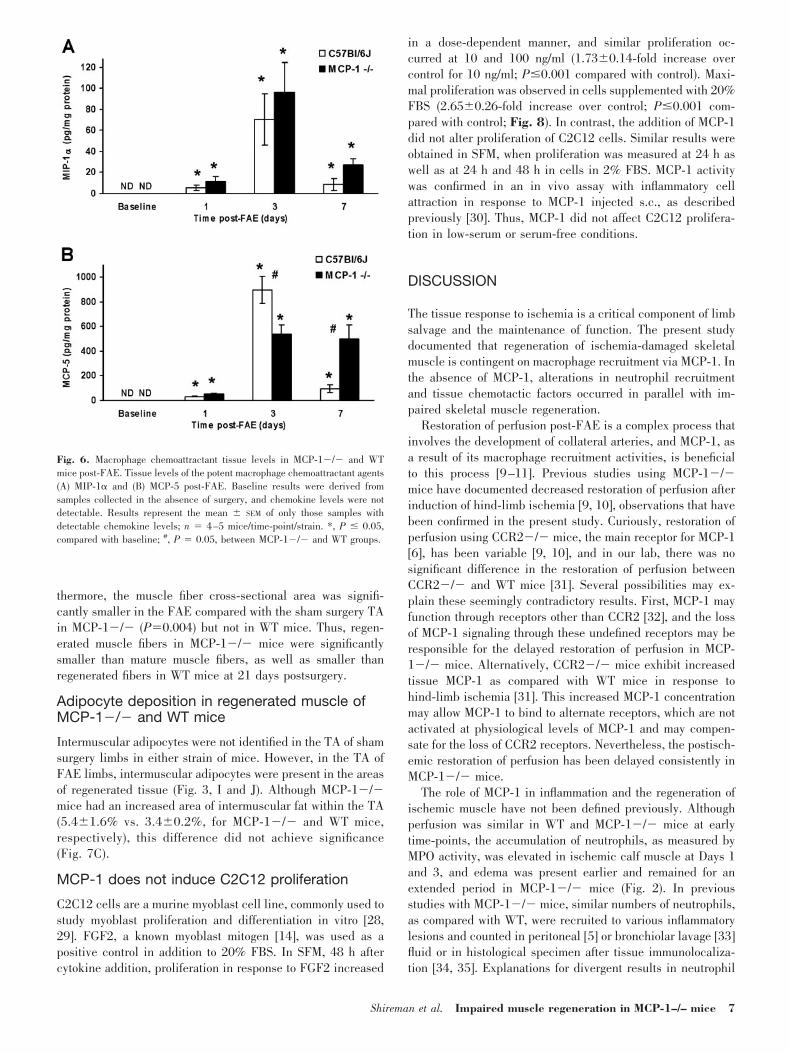
peripherally located nuclei. At 3 days, a prominent mononu-clear inflammatory infiltrate, primarily macrophages, was evi-dent in association with muscle injury in WT, but not MCP-1�/� mice (Figs. 3, A–D, and 4). Neutrophils were alsopresent in both strains of mice at Day 3 but were moreprevalent in the MCP-1 �/� mice. By 7 days post-FAE, theinflammatory infiltrate was greatly diminished in WT mice andreplaced by regenerated muscle fibers containing multiple,centrally located nuclei (Fig. 3F), whereas large regions ofnecrotic muscle remained in MCP-1�/� animals (Fig. 3E). Itis interesting that macrophage infiltration around necrotic mus-cle fibers was more prevalent by this time in MCP-1�/�animals. Muscle regeneration was essentially complete in WTmice within 14–21 days post-FAE, and the histologic appear-ance in WT animals was similar at both of these time-points. Incontrast, residual necrotic myofibers with associated muscleregeneration, macrophage infiltration, and adipocyte accumu-
lation were widespread in MCP-1�/� animals at Day 14 (Fig.3, G and H). Thus, in MCP-1�/� mice, macrophage accumu-lation was decreased at Day 3, when WT macrophage accu-mulation in WT mice was maximal, but the macrophage infil-trate persisted longer in association with residual necrotictissue. At 21 days post-FAE, muscle regeneration was largelycomplete in the MCP-1�/� mice, with adipocyte accumula-tion between regenerated fibers (Fig. 3I), and fewer adipocyteswere present in WT mice (Fig. 3J). Thus, impaired skeletalmuscle regeneration in MCP-1�/� mice was associated withdysregulated inflammation in the regenerating muscle and atendency to form adipocytes.
Elevated tissue chemokine levels in MCP-1�/�mice post-FAE
Given the alterations in neutrophil recruitment in MCP-1�/�mice, as described above, tissue levels of the neutrophil che-
Fig. 3. Macrophage (F4/80) infiltration,skeletal muscle regeneration, and fat accu-mulation in the ischemic hind limb in theabsence of MCP-1. (A) MCP-1�/�, 3 dayspost-FAE, neutrophilic inflammatory infil-trate with few mononuclear cells, gastrocne-mius muscle. (B) WT, 3 days post-FAE,robust mononuclear cell infiltrate, gastroc-nemius muscle. (C) MCP-1�/�, 3 dayspost-FAE, F4/80 immunolocalization, a fewpositive cells are present adjacent to theinjured muscle fibers, gastrocnemius mus-cle. (D) WT, 3 days post-FAE, F4/80 im-munolocalization with robust infiltration ofpositive cells, gastrocnemius muscle. (E)MCP-1�/�, 7 days post-FAE, TA musclewith small, regenerated fibers with one ormore centrally located nuclei, a few normalfibers at the periphery, necrotic muscle fi-bers. (F) WT, 7 days post-FAE, TA musclewith regenerated fibers, a few normal fibersat the periphery. (G) MCP-1�/�, 14 dayspost-FAE, regenerated TA muscle with in-termuscular adipose tissue, necrotic fibers,a few normal fibers at the periphery. (H)MCP-1�/�, 14 days post-FAE, F4/80 im-munolocalization, positive mononuclearcells interspersed among regenerated mus-cle fibers (arrows). (I) MCP-1�/�, 21 dayspost-FAE, regenerated TA muscle with in-termuscular adipose tissue (arrows). (J) WT,21 days post-FAE, regenerated TA musclewith focal intermuscular adipocytes (ar-rows). H&E stain except for C, D, and H,which were counterstained with hematoxy-lin; n, Normal skeletal muscle fiber; *, ne-crotic skeletal muscle fiber.
Shireman et al. Impaired muscle regeneration in MCP-1–/– mice 5
moattractants MIP-2 [23] and KC [24] were measured, cor-rected for protein, and expressed as pg/mg protein. Baselinelevels of both chemokines were below the limit of detection inMCP-1�/� and WT mice. In both mouse strains, MIP-2 waselevated significantly (P�0.002) over baseline at post-FAEDays 1, 3, and 7 (Fig. 5A). KC levels (Fig. 5B) were elevatedsignificantly (P�0.003) over baseline at Days 1 and 3 in WTmice and at Days 1, 3, and 7 in MCP-1�/� mice. Althoughthere were no significant differences between MCP-1�/� andWT mice at any single time-point for MIP-2 or KC, the meansof MIP-2 and KC were increased at Day 1 in MCP-1�/� micecompared with WT mice.
As macrophage recruitment was also altered in MCP-1�/�mice, tissue levels of the macrophage chemoattractantsMIP-1� [25] and MCP-5 [26] were measured. Baseline levelsof both chemokines were below the limit of detection in MCP-1�/� and WT mice. Although MIP-1� was elevated signifi-cantly (P�0.05) over baseline at post-FAE Days 1, 3, and 7,levels were similar between MCP-1�/� mice and WT mice atall time-points (Fig. 6A). In contrast to MIP-1�, MCP-5 (Fig.6B) was not only elevated significantly (P�0.001) over base-line in both mouse strains at post-FAE Days 1, 3, and 7 but wasincreased (P�0.05) in WT compared with MCP-1�/� mice atDay 3. However, at Day 7, MCP-5 was elevated significantly(P�0.05) in MCP-1�/� mice compared with WT mice. Thesedifferences between MCP-5 levels in the two mouse strainsmay account, in part, for the robust macrophage accumulation
in WT mice at Day 3 post-FAE and the persistence of macro-phages in MCP-1�/� mice at later time-points.
Impaired muscle regeneration in MCP-1�/�mice post-FAE
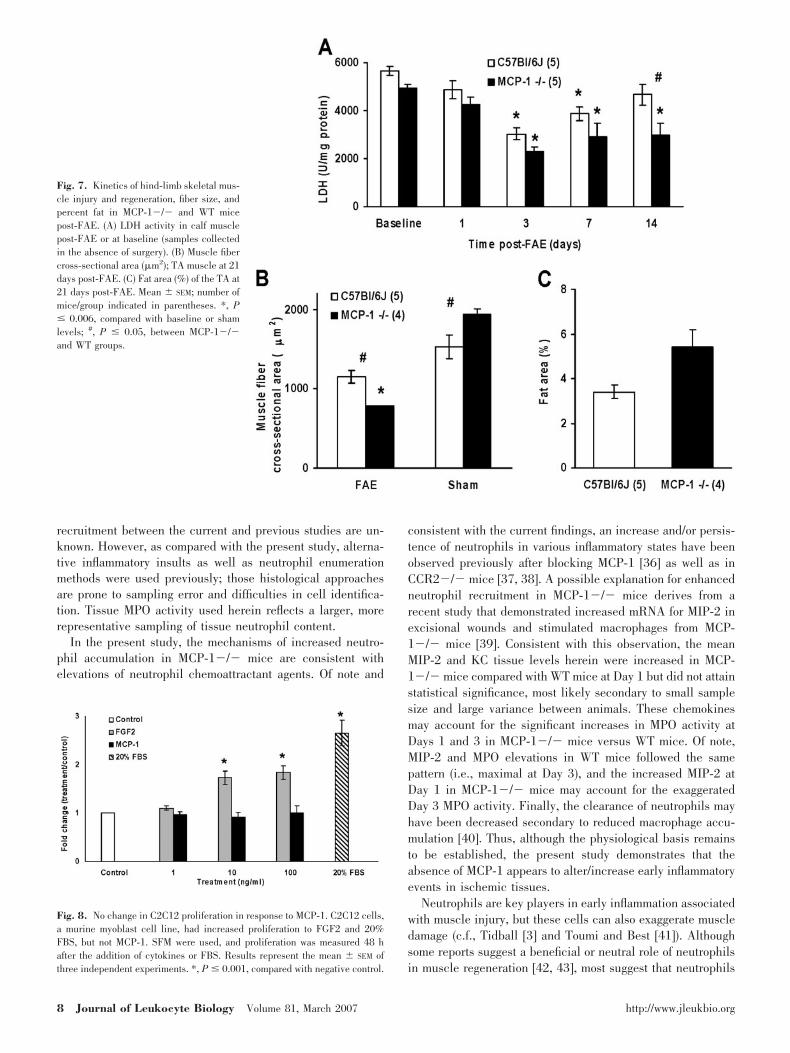
LDH is an enzyme that is present in skeletal muscle; itsactivity decreases with injury and is restored with regeneration[7, 27]. It is not surprising that contralateral calf muscles (shamsurgery side) from either mouse strain demonstrated no signif-icant differences from baseline in LDH activity (data notshown). In WT mice post-FAE (Fig. 7A), LDH activity waslowest at Day 3 and returned to near baseline by Day 14 withsignificant differences from baseline at Days 3 and 7(P�0.002). In contrast, in MCP-1�/� mice, LDH activity wasdecreased significantly from baseline at 3, 7, and 14 dayspost-FAE (P�0.006). Finally, LDH activity was decreasedsignificantly in MCP-1�/� mice at Day 14 (P�0.001) com-pared with WT animals.
In addition, at 21 days post-FAE, TA muscle fiber cross-sectional area (�m2) of regenerated fibers was decreased sig-nificantly in MCP-1�/� (789�3) compared with WT mice(1154�79; P�0.005; Fig. 7B). It is interesting that the maturefiber cross-sectional area in the sham surgery TA was in-creased more significantly in MCP-1�/� mice than in WTmice (1943�61 and 1529�150, respectively; P�0.05). Fur-
Fig. 5. Neutrophil chemoattractant tissue levels in MCP-1�/� and WT micepost-FAE. Tissue levels of the potent neutrophil chemoattractant agents (A)MIP-2 and (B) KC post-FAE. Baseline results were derived from samplescollected in the absence of surgery, and chemokine levels were not detectable(ND). Results represent the mean � SEM of only those samples with detectablechemokine levels; n � 4–5 mice/time-point/strain. *, P � 0.003, comparedwith baseline.
Fig. 4. Inflammatory cell infiltration at 3 days post-FAE. The triangular,soleus muscle (arrow). (A) WT with intense intramuscular mononuclear cellinfiltrate. (B) MCP-1�/� with reduced intramuscular inflammatory cell infil-trate. Inset, edge of soleus muscle (arrow) at higher magnification; H&E stain.
6 Journal of Leukocyte Biology Volume 81, March 2007 http://www.jleukbio.org
thermore, the muscle fiber cross-sectional area was signifi-cantly smaller in the FAE compared with the sham surgery TAin MCP-1�/� (P�0.004) but not in WT mice. Thus, regen-erated muscle fibers in MCP-1�/� mice were significantlysmaller than mature muscle fibers, as well as smaller thanregenerated fibers in WT mice at 21 days postsurgery.
Adipocyte deposition in regenerated muscle ofMCP-1�/� and WT mice
Intermuscular adipocytes were not identified in the TA of shamsurgery limbs in either strain of mice. However, in the TA ofFAE limbs, intermuscular adipocytes were present in the areasof regenerated tissue (Fig. 3, I and J). Although MCP-1�/�mice had an increased area of intermuscular fat within the TA(5.4�1.6% vs. 3.4�0.2%, for MCP-1�/� and WT mice,respectively), this difference did not achieve significance(Fig. 7C).
MCP-1 does not induce C2C12 proliferation
C2C12 cells are a murine myoblast cell line, commonly used tostudy myoblast proliferation and differentiation in vitro [28,29]. FGF2, a known myoblast mitogen [14], was used as apositive control in addition to 20% FBS. In SFM, 48 h aftercytokine addition, proliferation in response to FGF2 increased
in a dose-dependent manner, and similar proliferation oc-curred at 10 and 100 ng/ml (1.73�0.14-fold increase overcontrol for 10 ng/ml; P�0.001 compared with control). Maxi-mal proliferation was observed in cells supplemented with 20%FBS (2.65�0.26-fold increase over control; P�0.001 com-pared with control; Fig. 8). In contrast, the addition of MCP-1did not alter proliferation of C2C12 cells. Similar results wereobtained in SFM, when proliferation was measured at 24 h aswell as at 24 h and 48 h in cells in 2% FBS. MCP-1 activitywas confirmed in an in vivo assay with inflammatory cellattraction in response to MCP-1 injected s.c., as describedpreviously [30]. Thus, MCP-1 did not affect C2C12 prolifera-tion in low-serum or serum-free conditions.
DISCUSSION
The tissue response to ischemia is a critical component of limbsalvage and the maintenance of function. The present studydocumented that regeneration of ischemia-damaged skeletalmuscle is contingent on macrophage recruitment via MCP-1. Inthe absence of MCP-1, alterations in neutrophil recruitmentand tissue chemotactic factors occurred in parallel with im-paired skeletal muscle regeneration.
Restoration of perfusion post-FAE is a complex process thatinvolves the development of collateral arteries, and MCP-1, asa result of its macrophage recruitment activities, is beneficialto this process [9–11]. Previous studies using MCP-1�/�mice have documented decreased restoration of perfusion afterinduction of hind-limb ischemia [9, 10], observations that havebeen confirmed in the present study. Curiously, restoration ofperfusion using CCR2�/� mice, the main receptor for MCP-1[6], has been variable [9, 10], and in our lab, there was nosignificant difference in the restoration of perfusion betweenCCR2�/� and WT mice [31]. Several possibilities may ex-plain these seemingly contradictory results. First, MCP-1 mayfunction through receptors other than CCR2 [32], and the lossof MCP-1 signaling through these undefined receptors may beresponsible for the delayed restoration of perfusion in MCP-1�/� mice. Alternatively, CCR2�/� mice exhibit increasedtissue MCP-1 as compared with WT mice in response tohind-limb ischemia [31]. This increased MCP-1 concentrationmay allow MCP-1 to bind to alternate receptors, which are notactivated at physiological levels of MCP-1 and may compen-sate for the loss of CCR2 receptors. Nevertheless, the postisch-emic restoration of perfusion has been delayed consistently inMCP-1�/� mice.
The role of MCP-1 in inflammation and the regeneration ofischemic muscle have not been defined previously. Althoughperfusion was similar in WT and MCP-1�/� mice at earlytime-points, the accumulation of neutrophils, as measured byMPO activity, was elevated in ischemic calf muscle at Days 1and 3, and edema was present earlier and remained for anextended period in MCP-1�/� mice (Fig. 2). In previousstudies with MCP-1�/� mice, similar numbers of neutrophils,as compared with WT, were recruited to various inflammatorylesions and counted in peritoneal [5] or bronchiolar lavage [33]fluid or in histological specimen after tissue immunolocaliza-tion [34, 35]. Explanations for divergent results in neutrophil
Fig. 6. Macrophage chemoattractant tissue levels in MCP-1�/� and WTmice post-FAE. Tissue levels of the potent macrophage chemoattractant agents(A) MIP-1� and (B) MCP-5 post-FAE. Baseline results were derived fromsamples collected in the absence of surgery, and chemokine levels were notdetectable. Results represent the mean � SEM of only those samples withdetectable chemokine levels; n � 4–5 mice/time-point/strain. *, P � 0.05,compared with baseline; #, P � 0.05, between MCP-1�/� and WT groups.
Shireman et al. Impaired muscle regeneration in MCP-1–/– mice 7
recruitment between the current and previous studies are un-known. However, as compared with the present study, alterna-tive inflammatory insults as well as neutrophil enumerationmethods were used previously; those histological approachesare prone to sampling error and difficulties in cell identifica-tion. Tissue MPO activity used herein reflects a larger, morerepresentative sampling of tissue neutrophil content.
In the present study, the mechanisms of increased neutro-phil accumulation in MCP-1�/� mice are consistent withelevations of neutrophil chemoattractant agents. Of note and
consistent with the current findings, an increase and/or persis-tence of neutrophils in various inflammatory states have beenobserved previously after blocking MCP-1 [36] as well as inCCR2�/� mice [37, 38]. A possible explanation for enhancedneutrophil recruitment in MCP-1�/� mice derives from arecent study that demonstrated increased mRNA for MIP-2 inexcisional wounds and stimulated macrophages from MCP-1�/� mice [39]. Consistent with this observation, the meanMIP-2 and KC tissue levels herein were increased in MCP-1�/� mice compared with WT mice at Day 1 but did not attainstatistical significance, most likely secondary to small samplesize and large variance between animals. These chemokinesmay account for the significant increases in MPO activity atDays 1 and 3 in MCP-1�/� mice versus WT mice. Of note,MIP-2 and MPO elevations in WT mice followed the samepattern (i.e., maximal at Day 3), and the increased MIP-2 atDay 1 in MCP-1�/� mice may account for the exaggeratedDay 3 MPO activity. Finally, the clearance of neutrophils mayhave been decreased secondary to reduced macrophage accu-mulation [40]. Thus, although the physiological basis remainsto be established, the present study demonstrates that theabsence of MCP-1 appears to alter/increase early inflammatoryevents in ischemic tissues.
Neutrophils are key players in early inflammation associatedwith muscle injury, but these cells can also exaggerate muscledamage (c.f., Tidball [3] and Toumi and Best [41]). Althoughsome reports suggest a beneficial or neutral role of neutrophilsin muscle regeneration [42, 43], most suggest that neutrophils
Fig. 7. Kinetics of hind-limb skeletal mus-cle injury and regeneration, fiber size, andpercent fat in MCP-1�/� and WT micepost-FAE. (A) LDH activity in calf musclepost-FAE or at baseline (samples collectedin the absence of surgery). (B) Muscle fibercross-sectional area (�m2); TA muscle at 21days post-FAE. (C) Fat area (%) of the TA at21 days post-FAE. Mean � SEM; number ofmice/group indicated in parentheses. *, P� 0.006, compared with baseline or shamlevels; #, P � 0.05, between MCP-1�/�and WT groups.
Fig. 8. No change in C2C12 proliferation in response to MCP-1. C2C12 cells,a murine myoblast cell line, had increased proliferation to FGF2 and 20%FBS, but not MCP-1. SFM were used, and proliferation was measured 48 hafter the addition of cytokines or FBS. Results represent the mean � SEM ofthree independent experiments. *, P � 0.001, compared with negative control.
8 Journal of Leukocyte Biology Volume 81, March 2007 http://www.jleukbio.org

increase muscle injury via generation of oxidative stress [3, 41,44]. In fact, MPO may be a major factor in neutrophil-mediateddamage; i.e., MPO�/� mice had less muscle membrane injuryinduced by mechanical loading than WT mice despite similarlevels of inflammation [44]. However, in the current study, aslight but not significant decrease in LDH activity at Day 3 inMCP-1�/� compared with WT mice does not support thelikelihood that there was increased tissue damage as a result ofincreased neutrophil accumulation. Nevertheless, altered skel-etal muscle regeneration in the absence of MCP-1 may, in part,be attributable to increased MPO accumulation at early time-points and is the basis for ongoing studies.
MCP-1 may affect the interactions of neutrophils and mac-rophages. As MCP-1 has been immunolocalized to vascularendothelial cells and macrophages within ischemic hind-limbmuscles [7], the absence of endothelial MCP-1 likely alters thebalance of neutrophils and monocytes transmigrating acrossthe endothelium. In this regard, previous reports have noted aninterdependence between monocyte and neutrophil trafficking[45] as well as a need for macrophages to replace neutrophilsto facilitate the resolution of inflammation [46]. Furthermore,MCP-1 enhances the ability of macrophages to phagocytoseapoptotic neutrophils [40], potentially removing harmful neu-trophilic enzymes, which could increase muscle damage. Thus,the absence of MCP-1 could alter neutrophil interactions withendothelial cells and macrophages.
Macrophages are important in muscle regeneration [2–4],and MCP-1 is a potent chemotactic factor for macrophagerecruitment [6]. In the current study, it was not surprising thatmacrophages were decreased in ischemic muscle tissue, asdefects in macrophage recruitment have been reported previ-ously in MCP-1�/� mice [5, 35, 47]. Normally, macrophagespredominate during skeletal muscle regeneration and are es-sential for removal of necrotic tissue [13]. Macrophages alsoproduce a vast array of growth factors and enzymes that influ-ence many aspects of the muscle regenerative process, includ-ing angiogenesis and the chemotaxis, proliferation, and differ-entiation of myoblasts [48]. Unless the damaged muscle fiberbecomes invaded by macrophages, it remains arrested in thestage of intrinsic degeneration [4, 49]. Further support for amacrophage role in muscle regeneration derives from inhibi-tion of P-selectin, an adhesion molecule important in the earlyevents of leukocyte extravasation, which resulted in fewerinflammatory cells in injured tissue as well as impaired muscleregeneration [50] and angiogenesis [51]. Thus, defective mac-rophage recruitment and/or function likely contributed to im-paired muscle regeneration in MCP-1�/� mice in the currentstudy.
Macrophage recruitment was delayed in the MCP-1�/�mice, but macrophages persisted longer as compared with theWT mice. Persistence of macrophages could potentially derivefrom the extended presence of necrotic tissue in the MCP-1�/� mice. Macrophages are critical to the removal of ne-crotic tissue [13] and injured muscle fibers release factors thatare chemotactic for macrophages [12]. In fact, chronic muscleinjury, as observed in muscular dystrophy, can result inchronic macrophage infiltration, which can actually increasemuscle damage and fibrosis [52]. Other mechanisms that maycontribute to the persistence of macrophages include alter-
ations in the production of macrophage chemoattractants in theMCP-1�/� mice. Support for this possibility derives from thesignificantly elevated MCP-5 levels at Day 7 in MCP-1�/�mice and may provide a mechanism for the prolonged presenceof macrophages in the absence of MCP-1. It is interesting thatMIP-1�, another potent macrophage chemoattractant, was sim-ilarly elevated in both mouse strains, suggesting that the de-layed and prolonged accumulation of macrophages in MCP-1�/� mice was regulated by specific chemokines rather thana generalized increase in chemotactic factors.
Another important aspect of skeletal muscle regeneration isthe activation of satellite cells, the multipotent adult stem cellsthat reside in skeletal muscle [14]. Satellite cell proliferation isrequired to repair/replace injured myofibers. Normally, satel-lite cells are quiescent and are physically distinct from themyofiber, as they reside in indentations between the sarco-lemma and the basal lamina. In response to muscle damage,satellite cells become activated, proliferate, and express myo-genic markers (now termed myoblasts) [14]. Myoblasts ulti-mately fuse to existing muscle or fuse together to form newmyofibers during regeneration of damaged skeletal muscle(reviewed in Hawke and Garry [14]). It is interesting thatmacrophage-conditioned media increase myoblast proliferationand chemotaxis [12, 53–55].
Although the cellular mechanisms responsible for the de-layed skeletal muscle regeneration in MCP-1 animals remainto be established, it is conceivable that MCP-1 has directeffects upon satellite cells and/or myoblasts to stimulate theirproliferation and/or chemotaxis. Support for this possibility isprovided by observations that CCR2 [56] and MCP-1 [57] arelocalized to satellite cells in vivo, and MCP-1 is synthesized bymyoblasts in tissue culture [57]. MCP-1 is chemotactic forsatellite cells [57] and could influence muscle regeneration byinducing satellite cells from neighboring regions to migrate intoareas of muscle injury. Nevertheless, exogenous MCP-1 did notenhance in vitro proliferation of C2C12 myoblasts in the cur-rent study (Fig. 6). Additional experiments to examine themitogenic and chemotactic effect of MCP-1 on isolated satellitecells and/or myoblasts are necessary to address this importantquestion.
Impaired muscle regeneration in MCP-1�/� mice was dem-onstrated in the current study by decreased tissue LDH anddiminished muscle fiber size as compared with WT mice (Fig.7). Although the increased adipocyte accumulation in MCP-1�/� mice was not significant, it followed an interesting trendin comparison with CCR2�/� mice, which do exhibit in-creased adipocyte deposition in regenerated muscle followingischemic [31] and freeze [58] injury compared with WT mice.It is interesting that myogenic progenitor cells can transdiffer-entiate into adipocytes [15, 59]. Thus, although MCP-1 defi-ciency may contribute to adipocyte accumulation in injuredmuscle, other ligands of CCR2 may play a more important role.Whether impaired muscle regeneration was a direct effect ofMCP-1 on satellite cells, an indirect effect (i.e., altered inflam-mation) or a combination of both factors cannot be answered bythe present study.
In conclusion, the absence of MCP-1 during ischemic injuryof skeletal muscle was associated with a modest delay in therestoration of perfusion, altered inflammation, altered chemo-
Shireman et al. Impaired muscle regeneration in MCP-1–/– mice 9

kine levels, and impaired muscle regeneration. In combination,these findings demonstrate that MCP-1 is essential for resto-ration of perfusion and the successful completion of normalskeletal muscle regeneration following ischemic injury, prob-ably via macrophage recruitment and activation.
ACKNOWLEDGMENTS
This work is supported, in part, by the San Antonio AreaFoundation, Veterans Administration Merit Review, TheAmerican Heart Association, Texas Affiliate (0365123Y), andNIH (HL070158, HL074236, HL04776, and AR052610). Weacknowledge the technical assistance of Richard Castillo, Jef-ferey Jimenez, and Didier Nuno in the completion of thesestudies. A portion of the statistical analyses was graciouslyprovided by John Cornell and Gary Chisholm.
REFERENCES
1. Deindl, E., Schaper, W. (2005) The art of arteriogenesis. Cell Biochem.Biophys. 43, 1–16.
2. Merly, F., Lescaudron, L., Rouaud, T., Crossin, F., Gardahaut, M. F.(1999) Macrophages enhance muscle satellite cell proliferation and delaytheir differentiation. Muscle Nerve 22, 724–732.
3. Tidball, J. G. (2005) Inflammatory processes in muscle injury and repair.Am. J. Physiol. Regul. Integr. Comp. Physiol. 288, R345–R353.
4. Lescaudron, L., Peltekian, E., Fontaine-Perus, J., Paulin, D., Zampieri,M., Garcia, L., Parrish, E. (1999) Blood borne macrophages are essentialfor the triggering of muscle regeneration following muscle transplant.Neuromuscul. Disord. 9, 72–80.
5. Lu, B., Rutledge, B. J., Gu, L., Fiorillo, J., Lukacs, N. W., Kunkel, S. L.,North, R., Gerard, C., Rollins, B. J. (1998) Abnormalities in monocyterecruitment and cytokine expression in monocyte chemoattractant protein1-deficient mice. J. Exp. Med. 187, 601–608.
6. Charo, I. F., Taubman, M. B. (2004) Chemokines in the pathogenesis ofvascular disease. Circ. Res. 95, 858–866.
7. Shireman, P. K., Contreras-Shannon, V., Reyes-Reyna, S. M., Robinson,S. C., McManus, L. M. (2006) MCP-1 parallels inflammatory and regen-erative responses in ischemic muscle. J. Surg. Res. 134, 145–157.
8. Niiyama, H., Kai, H., Yamamoto, T., Shimada, T., Sasaki, K., Murohara,T., Egashira, K., Imaizumi, T. (2004) Roles of endogenous monocytechemoattractant protein-1 in ischemia-induced neovascularization. J. Am.Coll. Cardiol. 44, 661–666.
9. Waeckel, L., Mallat, Z., Potteaux, S., Combadiere, C., Clergue, M., Duriez,M., Bao, L., Gerard, C., Rollins, B. J., Tedgui, A., Levy, B. I., Silvestre,J. S. (2005) Impairment in postischemic neovascularization in mice lack-ing the CXC chemokine receptor 3. Circ. Res. 96, 576–582.
10. Voskuil, M., Hoefer, I. E., van Royen, N., Hua, J., de Graaf, S., Bode, C.,Buschmann, I. R., Piek, J. J. (2004) Abnormal monocyte recruitment andcollateral artery formation in monocyte chemoattractant protein-1-defi-cient mice. Vasc. Med. 9, 287–292.
11. Ito, W. D., Arras, M., Winkler, B., Scholz, D., Schaper, J., Schaper, W.(1997) Monocyte chemotactic protein-1 increases collateral and periph-eral conductance after femoral artery occlusion. Circ. Res. 80, 829–837.
12. Robertson, T. A., Maley, M. A., Grounds, M. D., Papadimitriou, J. M.(1993) The role of macrophages in skeletal muscle regeneration withparticular reference to chemotaxis. Exp. Cell Res. 207, 321–331.
13. Tidball, J. G. (1995) Inflammatory cell response to acute muscle injury.Med. Sci. Sports Exerc. 27, 1022–1032.
14. Hawke, T. J., Garry, D. J. (2001) Myogenic satellite cells: physiology tomolecular biology. J. Appl. Physiol. 91, 534–551.
15. Teboul, L., Gaillard, D., Staccini, L., Inadera, H., Amri, E. Z., Grimaldi,P. A. (1995) Thiazolidinediones and fatty acids convert myogenic cellsinto adipose-like cells. J. Biol. Chem. 270, 28183–28187.
16. Sun, J. S., Wu, S. Y., Lin, F. H. (2005) The role of muscle-derived stemcells in bone tissue engineering. Biomaterials 26, 3953–3960.
17. Steffel, J., Wernig, M., Knauf, U., Kumar, S., Wiestler, O. D., Wernig, A.,Brustle, O. (2003) Migration and differentiation of myogenic precursors
following transplantation into the developing rat brain. Stem Cells 21,181–189.
18. Shireman, P. K., Quinones, M. P. (2005) Differential necrosis despitesimilar perfusion in mouse strains after ischemia. J. Surg. Res. 129,242–250.
19. Charge, S. B., Rudnicki, M. A. (2004) Cellular and molecular regulationof muscle regeneration. Physiol. Rev. 84, 209–238.
20. Hornung, R. W., Reed, D. R. (1990) Estimation of average concentrationin the presence of nondetectable values. Appl. Occup. Environ. Hyg. 5,46–51.
21. Ueno, A., Oh-ishi, S. (2002) Critical roles for bradykinin and prostanoidsin acute inflammatory reactions: a search using experimental animalmodels. Curr. Drug Targets Inflamm. Allergy 1, 363–376.
22. Bradley, P. P., Priebat, D. A., Christensen, R. D., Rothstein, G. (1982)Measurement of cutaneous inflammation: estimation of neutrophil contentwith an enzyme marker. J. Invest. Dermatol. 78, 206–209.
23. Jerva, L. F., Sullivan, G., Lolis, E. (1997) Functional and receptor bindingcharacterization of recombinant murine macrophage inflammatory protein2: sequence analysis and mutagenesis identify receptor binding epitopes.Protein Sci. 6, 1643–1652.
24. Bozic, C. R., Kolakowski Jr., L. F., Gerard, N. P., Garcia-Rodriguez, C.,von Uexkull-Guldenband, C., Conklyn, M. J., Breslow, R., Showell, H. J.,Gerard, C. (1995) Expression and biologic characterization of the murinechemokine KC. J. Immunol. 154, 6048–6057.
25. Menten, P., Wuyts, A., Van Damme, J. (2002) Macrophage inflammatoryprotein-1. Cytokine Growth Factor Rev. 13, 455–481.
26. Sarafi, M. N., Garcia-Zepeda, E. A., MacLean, J. A., Charo, I. F., Luster,A. D. (1997) Murine monocyte chemoattractant protein (MCP)-5: a novelCC chemokine that is a structural and functional homologue of humanMCP-1. J. Exp. Med. 185, 99–109.
27. Fink, E., Fortin, D., Serrurier, B., Ventura-Clapier, R., Bigard, A. X.(2003) Recovery of contractile and metabolic phenotypes in regeneratingslow muscle after notexin-induced or crush injury. J. Muscle Res. CellMotil. 24, 421–429.
28. Koleva, M., Kappler, R., Vogler, M., Herwig, A., Fulda, S., Hahn, H.(2005) Pleiotropic effects of sonic hedgehog on muscle satellite cells. Cell.Mol. Life Sci. 62, 1863–1870.
29. Epstein, J. A., Lam, P., Jepeal, L., Maas, R. L., Shapiro, D. N. (1995) Pax3inhibits myogenic differentiation of cultured myoblast cells. J. Biol. Chem.270, 11719–11722.
30. Bleul, C. C., Fuhlbrigge, R. C., Casasnovas, J. M., Aiuti, A., Springer,T. A. (1996) A highly efficacious lymphocyte chemoattractant, stromalcell-derived factor 1 (SDF-1). J. Exp. Med. 184, 1101–1109.
31. Contreras-Shannon, V., Ochoa, O., Reyes-Reyna, S. M., Sun, D.,Michalek, J. M., Kuziel, W. A., McManus, L. M., Shireman, P. K. (2006)Fat accumulation with altered inflammation and regeneration in skeletalmuscle of CCR2 �/� mice following ischemic injury. Am. J. Physiol. CellPhysiol., Epub ahead of print.
32. Schecter, A. D., Berman, A. B., Yi, L., Ma, H., Daly, C. M., Soejima, K.,Rollins, B. J., Charo, I. F., Taubman, M. B. (2004) MCP-1-dependentsignaling in CCR2(�/�) aortic smooth muscle cells. J. Leukoc. Biol. 75,1079–1085.
33. Koth, L. L., Rodriguez, M. W., Bernstein, X. L., Chan, S., Huang, X.,Charo, I. F., Rollins, B. J., Erle, D. J. (2004) Aspergillus antigen inducesrobust Th2 cytokine production, inflammation, airway hyperreactivity andfibrosis in the absence of MCP-1 or CCR2. Respir. Res. 5, 12.
34. Kim, W. J., Chereshnev, I., Gazdoiu, M., Fallon, J. T., Rollins, B. J.,Taubman, M. B. (2003) MCP-1 deficiency is associated with reducedintimal hyperplasia after arterial injury. Biochem. Biophys. Res. Commun.310, 936–942.
35. Dewald, O., Zymek, P., Winkelmann, K., Koerting, A., Ren, G., Abou-Khamis, T., Michael, L. H., Rollins, B. J., Entman, M. L., Frangogiannis,N. G. (2005) CCL2/monocyte chemoattractant protein-1 regulates inflam-matory responses critical to healing myocardial infarcts. Circ. Res. 96,881–889.
36. Li, P., Garcia, G. E., Xia, Y., Wu, W., Gersch, C., Park, P. W., Truong, L.,Wilson, C. B., Johnson, R., Feng, L. (2005) Blocking of monocyte che-moattractant protein-1 during tubulointerstitial nephritis resulted in de-layed neutrophil clearance. Am. J. Pathol. 167, 637–649.
37. Jinnouchi, K., Terasaki, Y., Fujiyama, S., Tomita, K., Kuziel, W. A.,Maeda, N., Takahashi, K., Takeya, M. (2003) Impaired hepatic granulomaformation in mice deficient in C-C chemokine receptor 2. J. Pathol. 200,406–416.
38. Kuziel, W. A., Morgan, S. J., Dawson, T. C., Griffin, S., Smithies, O., Ley,K., Maeda, N. (1997) Severe reduction in leukocyte adhesion and mono-cyte extravasation in mice deficient in CC chemokine receptor 2. Proc.Natl. Acad. Sci. USA 94, 12053–12058.
10 Journal of Leukocyte Biology Volume 81, March 2007 http://www.jleukbio.org

39. Ferreira, A. M., Rollins, B. J., Faunce, D. E., Burns, A. L., Zhu, X.,Dipietro, L. A. (2005) The effect of MCP-1 depletion on chemokine andchemokine-related gene expression: evidence for a complex network inacute inflammation. Cytokine 30, 64–71.
40. Amano, H., Morimoto, K., Senba, M., Wang, H., Ishida, Y., Kumatori, A.,Yoshimine, H., Oishi, K., Mukaida, N., Nagatake, T. (2004) Essentialcontribution of monocyte chemoattractant protein-1/C-C chemokine li-gand-2 to resolution and repair processes in acute bacterial pneumonia.J. Immunol. 172, 398–409.
41. Toumi, H., Best, T. M. (2003) The inflammatory response: friend or enemyfor muscle injury? Br. J. Sports Med. 37, 284–286.
42. Teixeira, C. F., Zamuner, S. R., Zuliani, J. P., Fernandes, C. M., Cruz-Hofling, M. A., Fernandes, I., Chaves, F., Gutierrez, J. M. (2003) Neutro-phils do not contribute to local tissue damage, but play a key role inskeletal muscle regeneration, in mice injected with Bothrops asper snakevenom. Muscle Nerve 28, 449–459.
43. Teixeira, C. F., Chaves, F., Zamuner, S. R., Fernandes, C. M., Zuliani,J. P., Cruz-Hofling, M. A., Fernandes, I., Gutierrez, J. M. (2005) Effects ofneutrophil depletion in the local pathological alterations and muscleregeneration in mice injected with Bothrops jararaca snake venom. Int. J.Exp. Pathol. 86, 107–115.
44. Nguyen, H. X., Lusis, A. J., Tidball, J. G. (2005) Null mutation ofmyeloperoxidase in mice prevents mechanical activation of neutrophillysis of muscle cell membranes in vitro and in vivo. J. Physiol. 565,403–413.
45. Maus, U., von Grote, K., Kuziel, W. A., Mack, M., Miller, E. J., Cihak, J.,Stangassinger, M., Maus, R., Schlondorff, D., Seeger, W., Lohmeyer, J.(2002) The role of CC chemokine receptor 2 in alveolar monocyte andneutrophil immigration in intact mice. Am. J. Respir. Crit. Care Med. 166,268–273.
46. DiTirro, J., Rhoades, E. R., Roberts, A. D., Burke, J. M., Mukasa, A.,Cooper, A. M., Frank, A. A., Born, W. K., Orme, I. M. (1998) Disruptionof the cellular inflammatory response to Listeria monocytogenes infectionin mice with disruptions in targeted genes. Infect. Immun. 66, 2284–2289.
47. Huang, D. R., Wang, J., Kivisakk, P., Rollins, B. J., Ransohoff, R. M.(2001) Absence of monocyte chemoattractant protein 1 in mice leads todecreased local macrophage recruitment and antigen-specific T helpercell type 1 immune response in experimental autoimmune encephalomy-elitis. J. Exp. Med. 193, 713–726.
48. Grounds, M. D. (1998) Age-associated changes in the response of skeletalmuscle cells to exercise and regeneration. Ann. N. Y. Acad. Sci. 854,78–91.
49. Carlson, B. M., Faulkner, J. A. (1983) The regeneration of skeletal musclefibers following injury: a review. Med. Sci. Sports Exerc. 15, 187–198.
50. Baker, W., Schneider, B. A., Kulkarni, A., Sloan, G., Schaub, R., Sypek,J., Cannon, J. G. (2004) P-selectin inhibition suppresses muscle regener-ation following injury. J. Leukoc. Biol. 76, 352–358.
51. Egami, K., Murohara, T., Aoki, M., Matsuishi, T. (2006) Ischemia-inducedangiogenesis: role of inflammatory response mediated by P-selectin.J. Leukoc. Biol. 79, 971–976.
52. Gosselin, L. E., McCormick, K. M. (2004) Targeting the immune system toimprove ventilatory function in muscular dystrophy. Med. Sci. SportsExerc. 36, 44–51.
53. Cantini, M., Carraro, U. (1995) Macrophage-released factor stimulatesselectively myogenic cells in primary muscle culture. J. Neuropathol. Exp.Neurol. 54, 121–128.
54. Cantini, M., Massimino, M. L., Bruson, A., Catani, C., Dalla Libera, L.,Carraro, U. (1994) Macrophages regulate proliferation and differentiationof satellite cells. Biochem. Biophys. Res. Commun. 202, 1688–1696.
55. Massimino, M. L., Rapizzi, E., Cantini, M., Libera, L. D., Mazzoleni, F.,Arslan, P., Carraro, U. (1997) ED2 macrophages increase selectivelymyoblast proliferation in muscle cultures. Biochem. Biophys. Res. Com-mun. 235, 754–759.
56. Bartoli, C., Civatte, M., Pellissier, J. F., Figarella-Branger, D. (2001)CCR2A and CCR2B, the two isoforms of the monocyte chemoattractantprotein-1 receptor, are up-regulated and expressed by different cell sub-sets in idiopathic inflammatory myopathies. Acta Neuropathol. (Berl.)102, 385–392.
57. Chazaud, B., Sonnet, C., Lafuste, P., Bassez, G., Rimaniol, A. C., Poron,F., Authier, F. J., Dreyfus, P. A., Gherardi, R. K. (2003) Satellite cellsattract monocytes and use macrophages as a support to escape apoptosisand enhance muscle growth. J. Cell Biol. 163, 1133–1143.
58. Warren, G. L., Hulderman, T., Mishra, D., Gao, X., Millecchia, L.,O’Farrell, L., Kuziel, W. A., Simeonova, P. P. (2005) Chemokine receptorCCR2 involvement in skeletal muscle regeneration. FASEB J. 19, 413–415.
59. Li, W. C., Yu, W. Y., Quinlan, J. M., Burke, Z. D., Tosh, D. (2005) Themolecular basis of transdifferentiation. J. Cell. Mol. Med. 9, 569–582.
Shireman et al. Impaired muscle regeneration in MCP-1–/– mice 11
Related Documents











