CAN A RIVER-RESERVOIR INTERFACE SERVE AS NURSERY HABITAT FOR RIVERINE FISHES? by Matthew R. Acre, B.S. A Thesis In Wildlife, Aquatic, and Wildlands Science Management Submitted to the Graduate Faculty Of Texas Tech University in Partial Fulfillment of The Requirements for The Degree of MASTER OF SCIENCE Approved Timothy B. Grabowski Chair of Committee Nathan G. Smith Allison Pease Mark Sheridan Dean of the Graduate School August, 2015

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

CAN A RIVER-RESERVOIR INTERFACE SERVE AS NURSERY HABITAT FOR
RIVERINE FISHES?
by
Matthew R. Acre, B.S.
A Thesis
In
Wildlife, Aquatic, and Wildlands Science Management
Submitted to the Graduate Faculty
Of Texas Tech University in
Partial Fulfillment of
The Requirements for
The Degree of
MASTER OF SCIENCE
Approved
Timothy B. Grabowski
Chair of Committee
Nathan G. Smith
Allison Pease
Mark Sheridan
Dean of the Graduate School
August, 2015

Copyright © 2015
Matthew R. Acre

Texas Tech University, Matthew Acre, August 2015
ii
Acknowledgements
This work was made possible by funding from Texas Parks and Wildlife
Department (contract #432957), Texas Tech University, and the Texas Cooperative
Fisheries and Wildlife Research Unit. This work was conducted under the auspices of the
TTU Animal Care and Use (AUP #13004-01). I would like to recognize Bob Betsill at
the Heart of The Hills Research Unit for his support. I would like to thank my committee
chair, primary investigator, and friend Tim Grabowski. His guidance and wisdom is
appreciated and always needed. Thank you as well to my other committee members, Nate
Smith and Allison Pease, who always made themselves available to my questions and
always had time to lend a helping hand. Thank you to all the students from the
Grabowski Lab and Pease Lab both past and present for your support, the collaborative
nature of the graduate students from the labs made a daunting task a little less so. To
name a few of those graduate students who I could not have done this without; Jessica
East, Julia Mueller, Jillian Groeschel, Wade Massure, and Jessica Pease. During the
course of this research I was fortunate to have a committed undergraduate student as my
field technician. Daniel Logue took it upon himself to be responsible for much more than
just field work and was an invaluable resource whom without this work does not get
completed. Thank you, Dan. I am also grateful for lab technicians who spent countless
hours under the scope to complete this research; thank you Max Berlin, Amanda Emert,
and Grant Kilcrease. Also thank you to the countless number of people who chipped in
invaluable advice on everything from logistics to net construction. Last, but most
certainly not least, thank you to my mother and father for their patience and support

Texas Tech University, Matthew Acre, August 2015
iii
throughout a strenuous two years, I could not have done this without you and could not
have kept my sanity without your frequent field visits.

Texas Tech University, Matthew Acre, August 2015
iv
Table of Contents
Acknowledgements.............................................................................................................ii
Abstract...............................................................................................................................v
List of Tables......................................................................................................................vi
List of Figures....................................................................................................................vii
Introduction.........................................................................................................................1
Methods...............................................................................................................................6
Data Analyses....................................................................................................................15
Results................................................................................................................................17
Discussion..........................................................................................................................31
Literature Cited..................................................................................................................38
Appendix............................................................................................................................44

Texas Tech University, Matthew Acre, August 2015
v
Abstract
Anthropogenic modifications to riverine systems have reduced access to
important off-channel nursery habitats, but the river-reservoir interface (RRI) may offer
surrogate nursery habitat. I sampled ichthyoplankton assemblages in off-channel and
main channel habitats of the Lake Livingston RRI and middle Trinity River to: 1)
compare species composition and abundance in these different habitat types, and 2)
evaluate the influence of abiotic characteristics on ichthyoplankton assemblages in these
habitats. Ichthyoplankton was sampled using light traps and paired push nets deployed
off jet-propelled kayaks during March—July in 2013 and February—June in 2014. A
total of 44,029 larval fishes were collected, representing 27 taxa. A few taxa were
dominant at all sites, such as Threadfin Shad Dorosoma petenense and Inland Silverside
Menidia beryllina; however, less common taxa such as moronids, sciaenids, and
centrarchids were captured more frequently in RRI habitats. Sites with frequent
connectivity to the main channel, in particular the RRI backwater habitats, had higher
taxonomic richness, diversity, and overall abundance than similar habitats lacking this
connectivity. Canonical correspondence analysis suggests that there are major divisions
between mesohabitats in the Trinity River being driven by physicochemical components
and species assemblage structure related to connectivity to the main channel of the river.
These differences suggest RRI habitat may act as a surrogate nursery for some species
when access to other off-channel habitat is limited.

Texas Tech University, Matthew Acre, August 2015
vi
List of Tables
1 Means of the physicochemical and habitat variables collected for the river
backwaters (RIVBW), river main channel (RIVMC), river-reservoir interface
backwaters (RRIBW), and river-reservoir interface main channel (RRIMC) in the
Trinity River and Lake Livingston from March—July in 2013 and February—June
in 2014.............................................................................................................................9
2 Canonical correlation analysis results from push net transects of fish assemblage
data collected from 12 sites in the Trinity River and Lake Livingston, Texas
March—July in 2013 and February—June in 2014......................................................29
3 Standardized canonical discriminant coefficients for variables used in canonical
correlation analysis of the Trinity River and Lake Livingston larval fish
assemblages from push net transects, March—July in 2013 and February—June in
2014...............................................................................................................................30
A.1 Mean CPUE and H’ for larval fishes captured in 2013 and 2014 from the Trinity
River, Texas by site.......................................................................................................44
A.2 Canonical correlation analysis coefficients for larval fishes and habitat variables
collected from the Trinity River, Texas March—July in 2013 and February—June
in 2014...........................................................................................................................45
A.3 Taxonomic richness in push net (PN) and light trap (LT) samples taken from
riverine backwaters (RIVBW), riverine main channel (RIVMC), river-reservoir
interface backwaters (RRIBW), and river-reservoir interface main channel
(RRIMC) from the Trinity River and Lake Livingston, Texas March—July in 2013
and February—June in 2014.........................................................................................46
A.4 Fish collected in the Trinity River, Texas March—July in 2013 and
February—June in 2014 by site and year. An ‘x’ signifies a species was captured at
that site and a blank space indicates that the species was not captured........................47

Texas Tech University, Matthew Acre, August 2015
vii
List of Figures
1 The Trinity River, located entirely in Texas, runs from Dallas-Ft. Worth metroplex
and discharges into the Galveston Bay near Houston, Texas..........................................8
2 Two-way ANOVA to analyze mean catch per unit effort (CPUE) and Shannon’s
Diveristy Index (H’) for light trap and push net collections in the Trinity River,
Texas which were collected every three weeks from March—July in 2013 and
February—June in 2014................................................................................................18
3 Shannon’s Diveristy Index (H’) for light trap and push net collections in the Trinity
River, Texas which were collected every three weeks from March—July in 2013
and February—June in 2014.........................................................................................19
4 Timing of species appearance in samples taken from riverine backwaters (RIVBW),
riverine main channel (RIVMC), river-reservoir interface backwaters (RRIBW),
and river-reservoir interface main channel (RRIMC) from the Trinity River and
Lake Livingston, Texas March—July in 2013 and February—June in 2014...............22
5 Larval fish diversity in the Trinity River, Texas March—July in 2013 and
February—June in 2014 by mesohabitat type; river-reservoir interface backwater
(RRIBW), river-reservoir interface main channel (RRIMC), riverine backwater
(RIVBW), riverine main channel (RIVMC) ................................................................23
6 Larval fish diversity in 10 backwater sites sampled in the upper middle (RIVBW…)
and the river-reservoir interface (RRIBW…) of the Trinity River, Texas from
March—July in 2013 and February—June in 2014......................................................25
7 Canonical correspondence analysis linking species composition collected in the river
backwaters (RIVBW), river main channel (RIVMC), river-reservoir interface main
channel (RRIMC), and river-reservoir interface backwaters (RRIBW) in
March—July in 2013 and February—June in 2014 with physicochemical variables
from the Trinity River and Lake Livingston, Texas......................................................28

Texas Tech University, Matthew Acre, August 2015
1
Chapter I
Introduction
Anthropogenic alterations of riverine systems, in particular dams and reservoirs,
have had negative consequences for the diversity and productivity of riverine fish
assemblages (Baxter 1977; Pringle et al. 2000; Poff and Zimmerman 2010). Dam and
reservoir construction is a widespread and repeated disturbance. It is not unusual for a
river basin to have multiple dams which effectively fragment the river longitudinally,
resulting in short, isolated river reaches (Ward and Stanford 1995). Furthermore the
operation of dams serves to alter the quantity and timing of water moving through the
river basin, resulting in highly altered flow regimes and physical habitat alterations (NRC
1992; Dynesius and Nilsso 1994; Naiman et al. 1995; Gido et al. 2013). Behavioral and
life-history adaptations to the natural flow regime are documented for many riverine
species including synchronizing life-cycle events (e.g. spawning and egg laying) to flood
pulsing (Lytle and Poff 2004). Fishes that use rising water levels in warmer months as
cues to induce spawning or egg deposition may receive false cues or may never deposit
as a result of regulated flow regimes (Lytle and Poff 2004). Miller et al. (1989) noted a
total of 27 species and 13 subspecies of North American fishes became extinct during the
past century. Fifteen of these species and subspecies were primarily fluvial species from
highly altered systems. Similarly, Anderson et al. (1995) found a marked decline in
relative fish abundance and diversity over a 30-year period after dam construction East
Texas rivers, particularly within fluvial-specialist and migrating taxa (e.g. catastomids,

Texas Tech University, Matthew Acre, August 2015
2
cyprinids, ictalurids, and percids) due to longitudinal fragmentation. Some reproductive
guilds, such as pelagic broadcast spawners, seem to be particularly susceptible to
longitudinal habitat fragmentation (Platania and Altenbach 1998; Perkin and Gido 2011).
The negative effects of longitudinal fragmentation in riverine systems are more
immediate than some of the other effects such as a reduction in connectivity to floodplain
habitats.
Altered flow regimes have dramatically diminished connections between rivers
and the off-channel habitats of their floodplains (Poff et al. 1997; Poff and Zimmerman
2010). Reductions in high flow events have also resulted in channelization and other
modifications to channel morphology which can exacerbate the already reduced
connectivity by necessitating higher flows to connect off-channel habitat (Ward and
Stanford 1995). While many fishes in temperate rivers are capable of completing their
life histories while confined to the main channel of the river (Galat and Zweimüller 2001;
Welcomme et al. 2006), there appear to be long-term negative consequences associated
with the loss of access to off-channel habitat due to altered flow regimes for a large
proportion of riverine fishes (Sammons and Macena 2009; Dutterer et al. 2012). For
example, Dutterer et al. (2012) found positive relationships between catch rates of age-0
Spotted Bass Micropterus punctulatus and Spotted Sucker Minytrema melanops and the
occurrence of discharge rates high enough to connect off-channel habitat in the
Apalachicola River, Florida. In years that discharge rates were lower and access to off-
channel habitat was not available, fewer age-0 fishes were caught, suggesting stronger

Texas Tech University, Matthew Acre, August 2015
3
year-classes in years with higher discharge rates. Weaker year-classes have been
associated with loss of connectivity to spawning and nursery habitats. Although nursery
habitats for some species of concern have yet to be identified, many are likely dependent
on floodplain habitat, at least partly, to complete their life histories (spawning and egg
deposition). For example, range-wide declines of Alligator Gar Atractosteus spatula have
been partly attributed to loss of access to off-channel spawning and nursery habitats
(Alfaro et al., 2008; Brinkman 2008; Inebnit 2009; Kluender 2011). Though little is
known about Alligator Gar in the earliest life-history stages, their spawning season
coincides with seasonal river pulsing that would connect floodplain habitats in less
modified systems (Simon and Wallus 1989). Additionally, their populations across the
southeastern U.S. have been in decline since the 1980s which coincides with a reduction
in connectivity to floodplain habitat (Etnier and Starnes 1993; Simon and Wallus 1989).
While there has been a general reduction in the connectivity of most altered rivers
with their floodplains (Ward and Stanford 1995), aging of the reservoirs in these altered
systems has resulted in the development of superficially similar nursery habitats at the
river-reservoir interface (RRI). The RRI is an ecotone where there is a gradual transition
from lotic conditions to lentic conditions, including a reduction in current velocity. Large
amounts of sediment are deposited in the RRI as a reservoir ages, building delta-like
formations consisting of off-channel backwater habitats, isolated coves, and drowned
creeks and tributaries (Palmieri et al. 2001). These RRI habitats are by their nature, low
relief, low elevation areas, and therefore require lower discharge rates to remain

Texas Tech University, Matthew Acre, August 2015
4
connected to the main channel. In the last 20 years much research has been done on adult
fish assemblages within the RRI (e.g., Carvalho et al.1998; Oliveira et al. 2004;
Kaemingk et al. 2007; Santos et al. 2010; Terra et al. 2010; Yang et al. 2012; Buckmeier
et al. 2013). These studies concluded that the RRI supports high abundance, high species
diversity, high taxonomic richness, and high proportions of fishes occurring in the river-
reservoir ecosystem, relative to river or reservoir segments. Fish assemblages in the RRI
also seem to be highly variable, with seasonal species turnover, suggesting the possibility
that many of the species may only be temporary residents as they move through the RRI
between riverine and reservoir habitats (Buckmeier et al. 2013). Little work has been
done to quantify use of the RRI as spawning or nursery habitats, particularly by
floodplain-dependent riverine species that require lateral connectivity to floodplain
habitat. The more frequent and consistent accessibility of RRI habitats and their
superficial similarity to off-channel floodplain habitats suggest they may have an
important long-term function in maintaining populations of fishes that rely on off-channel
habitats as nursery and spawning habitat.
Clarifying how the RRI is being used by fishes has important conservation
applications, as RRI habitats have been proposed as targets of management action to
preserve fish diversity within altered riverine systems (Buckmeier et al. 2013). The
Trinity River in eastern Texas is a highly altered riverine system that offers the
opportunity to compare the use of floodplain and RRI habitats as nurseries by the fish
assemblage. There are 35 dams within the Trinity River Basin, including Lake Livingston

Texas Tech University, Matthew Acre, August 2015
5
which is one of the largest reservoirs in Texas. The RRI between the Trinity River and
Lake Livingston is approximately 34 km and contains numerous backwaters of varying
areas and connection sizes that are superficially similar to off-channel floodplain habitat.
The objectives of this study were to 1) quantify abundance and diversity of larval fishes
in the RRI relative to naturally-occurring off-channel habitats with reduced connectivity
to the main channel 2) examine whether abundance and species composition of the
ichthyoplankton assemblage varies between main channel habitats and off-channel
habitats in the RRI, and 3) evaluate the degree of influence physicochemical
characteristics of off-channel habitats in the RRI and riverine sites have on abundance
and species composition.

Texas Tech University, Matthew Acre, August 2015
6
Chapter II
Methods
Study Area
The Trinity River flows approximately 885 km from its headwaters north of the
Dallas-Fort Worth metroplex area southeast towards Houston, eventually emptying into
the Galveston Bay system (Figure 1). The Trinity River supplies a large proportion of the
water used by Dallas-Fort Worth and Houston metropolitan areas, and 34 impoundments
have been constructed in the Trinity River Basin since 1914 to meet the increasing water
demands of these rapidly growing cities (Perkin and Bonner 2011). Most of these
impoundments occur on the tributaries of the Trinity River, as the main channel of the
river is unobstructed within this study area except for an incomplete lock and dam. My
research focused on a portion of the middle Trinity River from Richland Chambers
Reservoir to the Lake Livingston RRI (Figure 1). The middle Trinity River is heavily
channelized with sandy-clay substrates and relatively high discharge (358-29,100 m3s-1).
Lake Livingston, constructed in 1969, meets the water demands of the four surrounding
counties which includes the city of Houston. It is the largest single-purpose reservoir in
Texas and has a large delta-like transition zone, or RRI (Figure 1; C), in its upper reaches
that is typical of reservoirs of similar age and size.
This study encompassed off-channel backwaters and riverine habitats from the
Trinity River near U.S. Highway 287 and FM 488, Big Lake Bottom Wildlife

Texas Tech University, Matthew Acre, August 2015
7
Management Area (Figure 1; A & B), and near Riverside, Texas off State Highway 19
(Figure 1; C). Each sampling location was classified as one of four mesohabitat types;
river main channel (RIVMC), river backwater (RIVBW), RRI main channel (RRIMC),
and RRI backwater (RRIBW), and each varied in physical size, complexity, and
physicochemical condition (Table 1).

Texas Tech University, Matthew Acre, August 2015
8
Figure 1 The Trinity River, located entirely in Texas, runs from Dallas-Ft. Worth metroplex and discharges
into the Galveston Bay near Houston, Texas. Inset A: Three river backwater sites and the river main channel
site. Inset B: One river backwater site. Inset C: Six river-reservoir interface backwater and one river-reservoir
interface main channel site. Sampling localities were sampled every three weeks from March—July in 2013
and February—June in 2014.

Texas Tech University, Matthew Acre, August 2015
9
Table 1 Means of the physicochemical and habitat variables collected for the river backwaters (RIVBW), river main channel (RIVMC), river-reservoir interface
backwaters (RRIBW), and river-reservoir interface main channel (RRIMC) in the Trinity River and Lake Livingston from March—July in 2013 and February—
June in 2014. Area and connection size were measured to determine the ratio of connection size relative to the area of a backwater (RCA). Turbidity (NTU;
nephelometric turbidity units), conductivity (mS/cm; millisiemens per centimeter), temperature (°C), and dissolved oxygen (milligrams/liter) were collected
during each sampling event and a mean is presented here to illustrate the variability in backwater habitats.
Site Area
(ha)
Mean
estimated
connection
size (m)
Perimeter
(m)
Connection
size/area
(RCA)
Mean
turbidity
(NTU)
Mean
cond.
(mS/cm)
Mean
temp.
(°C)
Mean
DO
(mg/L)
River RIVBW9 28.69 - 6750.0 - 19.58 0.28 23.95 8.27
RIVBW12 56.26 - 6307.3 - 32.45 0.33 21.37 10.10
RIVBW13 28.96 - 2484.5 - 12.46 0.26 22.29 16.86
RIVBW14 61.72 - 11988.8 - 9.88 0.72 16.91 7.76
RIVMC - - - - 85.80 0.71 23.03 9.54
RRI RRIBW2 82.02 56 12907.7 6.85E-05 38.40 0.52 23.68 8.73
RRIBW3 162.84 145 19676.2 8.91E-05 36.71 0.44 23.76 10.41
RRIBW4 4.52 12 1094.7 2.66E-04 32.01 0.50 24.05 10.67
RRIBW5 463.32 177 34013.9 3.82E-05 45.55 0.41 23.04 8.69
RRIBW6 44.26 11 5453.3 2.42E-05 66.98 0.45 21.94 9.45
RRIBW7 100.39 32 8045.0 3.22E-05 89.23 0.47 24.39 11.81
RRIMC - - - - 40.79 0.45 23.26 7.55

Texas Tech University, Matthew Acre, August 2015
10
River Sites
I sampled four river backwater sites and one river main channel site on the Trinity
River (Figure 1; A & B). Sites were selected to encompass variations in natural
backwater habitats. However, most are not accessible from the river as they do not
maintain a connection to the main channel. Therefore, I was limited to backwaters that
had public access or privately-owned backwaters to which I was given permission to
access. The river main channel site was accessed via public boat ramp at the US-287
highway crossing (Figure 1; A). One riverine backwater (RIVBW14) was only sampled a
total of four times; once in 2013 and three times in 2014 due to gear limitations in heavily
vegetated habitat.
RRI Sites
Six RRI backwater sites (Figure 1; C) were identified and sampled as well as one
RRI main channel site in the southernmost section of the study area. As there was large
variation among backwater habitats in the RRI (Table 1), I selected sites that accounted
for much of this variation. The main channel in the RRI (Figure 1; C) was not as clearly
defined as the main channel in the river sites; however, using the National Hydrography
Dataset (USGS), the thalweg was identified flowing into Lake Livingston and sampling
transects were established in this predefined RRI main channel habitat.

Texas Tech University, Matthew Acre, August 2015
11
Sampling Methodology
I sampled larval fishes approximately every three weeks during March—July in
2013 and February—June in 2014 using both a modified push net design (Acre and
Grabowski in press) and quatrefoil light traps. I collected paired samples using two 0.75
x 0.50 x 1.6-m push nets mounted between two jet-propelled kayaks (Mokai
Manufacturing Inc., Newburgh, NY) with a 4.5 x 4.5 x 275-cm aluminum bar. Nets were
constructed using 1000-µm nylon mesh netting. Three 100-m transects were established
haphazardly where water depth was ≥ 0.75 m and obstructions were minimal. Transects
were surveyed each sampling event at a distance of ≤ 5 m to shore, with the exception of
one transect per site across the center of backwater in an attempt to cover most habitat
types available in a given backwater. All main channel transects were located as close to
shore as debris and depth allowed. During each push net sample, flow through each net
was recorded with a Model 2030R standard flowmeter (General Oceanics Inc., Miami,
FL) mounted across the mouth of each net. The duration of each sampling event and GPS
locations at the start and stop of each transect were recorded.
To supplement push net samples, I deployed modified quarter-foil light traps
(Floyd et al. 1984). Traps were constructed from high density polyethylene plastic
bottles, approximately 1750 cm3 in volume. A light source comprised of two amber-
colored waterproof LED lights was mounted internally. Three light traps were deployed
in close proximity to push net transects to encompass similar habitat and species
assemblages. Light traps were deployed for 1.0-6.0 hrs at a depth of 25 cm.

Texas Tech University, Matthew Acre, August 2015
12
A suite of physicochemical attributes was recorded at each sampling location.
Dissolved oxygen (mg L-1), conductivity (mS cm-1), and temperature (°C) were recorded
at each site using a YSI Pro2030 Processional Series unit (YSI incorporated, Yellow
Springs, Ohio). Turbidity (NTU) was recorded once at each site during every sampling
event with an Oakton turbidity meter (Oakton Instruments, Vernon Hills, Illinois). Water
temperature data were acquired from HOBO data loggers (Onset Computer Corporation,
Bourne, Massachusetts) placed at each site by Texas Parks and Wildlife Department.
Fish Processing and Identification
Upon capture, all larval fishes were euthanized using a ≥ 250 mg L-1 solution of
tricaine methanesulfonate (MS-222 Western Chemical Inc., Ferndale, Washington),
preserved in ExCell Plus (American MasterTech, Lodi, California), an ethanol-based
tissue fixative, and returned to the laboratory for processing. At the time of this study,
there was no freshwater fish larval fish identification key for the Trinity River Basin
specifically, nor for Texas generally. Therefore, larval fishes were identified to the lowest
taxon possible using Snyder and Seal (2008) and Wallus et al. (1990, 2004, 2006, 2006,
2008; Kay et al. 1994). Larval fishes were photographed against a 1.0 X 1.0 cm grid for
scale using an Infinity 1-2 microscopy camera (Lumenera Corporation, Ottawa, Ontario)
mounted on an Olympus SZX16 stereomicroscope (Olympus Corporation of the
Americas, Center Valley, Pennsylvania). A subsample was selected at random and
measured to the nearest 1.0 mm TL using ImageJ v 1.48 (Rasband 1997-2014). Larval

Texas Tech University, Matthew Acre, August 2015
13
fishes were stored in 1-mm glass vials filled with tissue fixative and deposited with the
Texas Natural History Collection in Austin, Texas (accn# 2015-14)
Discharge Data and Estimation of Lateral Connectivity
I estimated connection size and occurrence of backwaters to the main channel of
the Trinity River using a time series of satellite imagery from Google Earth (Google Inc.,
Mountain View, California) in the RRI of Lake Livingston. Images used in Google Earth
were captured on December 31, 2008 and June 4, 2010, from the Texas Orthoimagery
Program. A measurement of connection was taken for each date in the time series and the
mean was used (Table 1).
I used elevation of riverine backwaters, gauge height, and on-site observations (N.
Smith and D. Buckmeier, Texas Parks and Wildlife, personal communication) to
determine when riverine backwater connected to the main stem of the Trinity River. The
closest gage station to the riverine sites is approximately 31 km upstream (USGS gage
08062700 Trinity River at Trinidad, Texas). I used the bottom of channel elevation at
mean seal level at the Trinidad, TX gage (71.8 m) plus the gage height reading provided
by the USGS to estimate surface elevation at Trinidad, TX. I then subtracted the 5.6 m
which is the elevation difference between the Trinidad, TX gage and the spillway’s
confluence with the Trinity River (L. Byers, Richland Chambers Reservoir
Superintendent, personal communication). I then subtracted the elevation of the low-head

Texas Tech University, Matthew Acre, August 2015
14
dam in the tailrace of the Richland Chambers Reservoir. The resulting number, if
positive, indicated a connection between the Trinity River and at least one riverine
backwater site.

Texas Tech University, Matthew Acre, August 2015
15
Chapter III
Data Analyses
Relative Abundance (CPUE) and Shannon Index (H’)
All statistical analyses were performed using the program SAS v. 9.3 (SAS
Institute Inc., Cary, NC). Transect data was pooled by sampling event and catch per unit
effort (CPUE) and Shannon’s diversity index (Hʹ) were calculated for each sampling
event at each site, with light traps and push net data calculated separately. Catch per unit
effort for push net samples was calculated using the total number of fish captured per
transect and volume sampled per transect. Light trap CPUE was calculated using total
fish captured and time (minutes). Unless otherwise noted, push net data results are
presented as light trap data did not change the conclusions of this research. I used a
logarithmic transformation plus a constant to approximately meet the parametric
assumptions of normality and equality of variance (Guy and Brown 2007). I used a three-
way ANOVA with year (2013 and 2014), habitat (backwater and main channel), and type
(river and RRI) as the independent variables. A pairwise multiple comparison Tukey-
Kramer test was used for post-hoc comparisons.
Canonical Correspondence Analysis
Canonical correspondence analysis (CCA; Legendre and Legendre 1998) was
performed using only push net data to evaluate the influence of relevant environmental

Texas Tech University, Matthew Acre, August 2015
16
variables on assemblage composition across sample sites. This multivariate technique
merges multiple regression and ordination techniques and is commonly used to link
species abundance with environmental conditions at the time of sampling. Environmental
and habitat variables were assessed for normality in SAS prior to analyses with a
Kolmogorov-Smirnov test. A log transformation was performed on all data to
approximately meet the parametric assumptions of normality and equality of variance.
Once transformations were applied to all variables, species which were present in < 2%
of push net transects were removed from the CCA. A mean of zero and a standard
deviation of one (µ = 0, = 1) were used to standardize the transformed dataset.

Texas Tech University, Matthew Acre, August 2015
17
Chapter IV
Results
Relative Abundance and Species Diversity
I collected 44,029 larvae from the Trinity River and Lake Livingston RRI across
14 sampling events during 2013-2014. Overall, 82% of fishes were captured in the
backwaters of the RRI and 12% in the backwaters of the upper-middle Trinity River and
the remaining 6% were captured in the RRI and river main channels. RRI habitats
generally exhibited higher larval fish abundance and diversity than their riverine
counterparts for both push net transects and light traps (CPUE: F7,377 ≥ 2.26, P ≤ 0.03;
H’: F7,378 ≥ 10.67, P ≤ 0.001; Figure 2). There were no differences observed between
years for relative abundance of fishes, however, diversity between years were different
for both push net and light trap collections (F1,378 ≥ 5.95, P ≤ 0.02; Figure 3). This
relationship was driven mainly by the differences seen across the four mesohabitats
(F1,378 ≥ 3.79, P ≤ 0.05). While the two gear types exhibited similar spatial and temporal
patterns of larval abundance and taxonomic richness and diversity, push net transects
generally captured a larger number of larval fishes per sampling event and greater
taxonomic richness and diversity than light traps set at the same locations (F7,1231 ≥ 48.60,
P ≤ 0.001).

Texas Tech University, Matthew Acre, August 2015
18
Figure 2 Two-way ANOVA to analyze mean (± SE) catch per unit effort (CPUE) and Shannon’s Diveristy Index
(H’) for light trap and push net collections in the Trinity River, Texas which were collected every three weeks from
March—July in 2013 and February—June in 2014. The letters above each bar represents the mean seperation
between mesohabitat type; river backwater (RIV BW), river-reservoir interface backwater (RRI BW), river main
channel (RIV MC), and river-reservoir interface main channel (RRI MC). If the letter above the bar is the same then
the mesohabitats were not statistically different for that gear type. Shaded bars represent the riverine sites.

Texas Tech University, Matthew Acre, August 2015
19
Figure 3 Mean (±SE) Shannon’s Diveristy Index (H’) for light trap and push net collections in the Trinity River, Texas which were collected every three
weeks from March—July in 2013 and February—June in 2014

Texas Tech University, Matthew Acre, August 2015
20
Species Chronology and Richness
A total of 27 taxa were represented in the larval fish samples consisting of 25 that
were identified to the species level, one to genus, and one family-level group that likely
consisted of multiple genera and species. Percids were easily identified to the genus
level; however, a few species of Percina present in the Trinity River have no reliable
keys for larval identification. It is likely that collections of this genus are all Bigscale
Logperch Percina macrolepida, however, historical accounts exist of two other Percina
species (Hendrickson and Cohen 2012) in the Trinity River. Cyprinids were
unidentifiable past the family level due to a lack of larval keys for the species present in
the Trinity River Basin and therefore were treated as a single taxa for analysis.
Regardless of site, taxonomic richness was greater for push nets samples than light traps
samples (F23,1215 = 128.25, P ≤ 0.001). Push nets captured all 27 taxa represented in this
study and nine were unique to the push net samples.
Taxonomic richness varied both spatially and temporally during the study. Inland
Silverside Menidia beryllina and Threadfin Shad Dorosoma petenense were captured at
all sites and tended to be among the first species caught each spring. These two species
were collected throughout the course of each sampling season (Figure 4). Blue Catfish
Ictalurus furcatus, Channel Catfish Ictalurus punctatus, and White Bass Morone
chrysops were the only other species to be captured from all habitat types. However,
these species were only captured sporadically (Figure 4). Centrarchids, such as sunfishes
Lepomis spp., crappies Pomoxis spp., and Largemouth Bass were not encountered in the

Texas Tech University, Matthew Acre, August 2015
21
main channel of the river and tended to appear in samples in late spring and early summer
(Figure 4). Spotted Gar Lepisosteus oculatus was the only gar species encountered and
was represented by a single individual taken from the river main channel in 2014.
Catostomids, such as Smallmouth Buffalo and Spotted Sucker, were absent from riverine
main channel and backwater samples in 2013 (Figure 4), but were taken from one
riverine backwater in 2014. Smallmouth Buffalo was collected in both backwater and
main channel RRI habitats in both years; however, the species was captured in greater
numbers in the riverine backwaters (Figure 4).
The timing of species appearances in the samples tended to be later in 2013
relative to that in 2014 (Figure 4) and may have been related to the differences in
temperature and flow regimes between the two years (Figure 5b, d). In general, species
that were collected in both the riverine and RRI sites were collected earlier and for a
longer period than in the riverine sites. Species in the riverine main channel tended to be
first encountered later in the sampling season than the same species in riverine
backwaters or the RRI (Figure 4) and there were only a few taxa that were captured from
the riverine main channel on consecutive sampling events, such as cyprinids, Blue
Catfish, Channel Catfish, and Inland Silverside. A similar pattern was not noted in the
RRI where species were first encountered at the same time for main channel and
backwater sites (Figure 4).

Texas Tech University, Matthew Acre, August 2015
22
Figure 4 Timing of species appearance in samples taken from riverine backwaters (RIVBW), riverine main channel (RIVMC), river-reservoir interface
backwaters (RRIBW), and river-reservoir interface main channel (RRIMC) from the Trinity River and Lake Livingston, Texas March—July in 2013 and
February—June in 2014. A solid black bar indicates a species was captured during a sampling event. If a species was captured on non-consecutive
sampling events the time gap between sampling events was colored grey, indicating that the species was presumed to be present during that time. All
blank bars indicate that a species was not captured
Jun Jun
Spotted Gar
Threadfin Shad
Cyprinidae
Smallmouth Buffallo*
Spotted Sucker*
Blue Catfish
Channel Catfish
Tadpole Madtom*
Flathead Catfish*
Inland Silverside
Sheepshead Minnow*
White Bass
Redbreast Sunfish
Green Sunfish
Warmouth
Orangespotted Sunfish
Bluegill
Longear Sunfish
Sunfush species
Largemouth Bass*
White Crappie
Black Crappie*
Bluntnose Darter
Harlequin Darter*
Bigscale Logperch
Percina spp .
Freshwater Drum*
Spotted Gar
Threadfin Shad
Cyprinidae
Smallmouth Buffallo*
Spotted Sucker*
Blue Catfish
Channel Catfish
Tadpole Madtom*
Flathead Catfish*
Inland Silverside
Sheepshead Minnow*
White Bass
Redbreast Sunfish
Green Sunfish
Warmouth
Orangespotted Sunfish
Bluegill
Longear Sunfish
Sunfush species
Largemouth Bass*
White Crappie
Black Crappie*
Bluntnose Darter
Harlequin Darter*
Bigscale Logperch
Percina spp .
Freshwater Drum*
May
RIVMC
2013 2014
RIVBW
RRIBW
2013
RRIMC
2014
Mar Apr May Jun Jul Feb Mar Apr May Mar Apr May Jun Jul Feb Mar Apr

Texas Tech University, Matthew Acre, August 2015
23
Figure 5 Larval fish diversity in the Trinity River, Texas March—July in 2013 and February—June in 2014 by
mesohabitat type; river-reservoir interface backwater (RRIBW), river-reservoir interface main channel
(RRIMC), riverine backwater (RIVBW), riverine main channel (RIVMC). Temperature and flow data
downloaded from USGS gage station (USGS Gage 08065350 Trinity River near Crockett, Texas) roughly half
way between the riverine sites and the RRI sites and was chosen to be representative of the study area as a
whole. The vertical dashed line indicates when river discharge was high enough to potentially connect riverine
backwaters. Black dots on panels B and D represent sampling events.

Texas Tech University, Matthew Acre, August 2015
24
Larval fish taxonomic richness varied during the spring of 2013 and 2014, but
was consistently higher in the backwaters relative to adjacent main channel habitats
(Figure 5). Taxonomic richness was positively correlated with water temperatures (r ≥
0.53, P ≤ 0.001), but did not exhibit a strong relationship with discharge on the day of
sampling (r ≤ -0.18, P = 0.18). Water temperatures were comparable both years, but
water temperatures at the start of 2014 sampling were cooler and experienced a rapid
drop in early March which appears to coincide with increased species richness in the
riverine backwaters in 2014 compared to 2013 (Figure 5). Taxonomic richness was
greater in the RRI backwaters than any other mesohabitat (F3,50 = 6.26, P ≤ 0.001).
Larval fish taxonomic richness in the backwaters of the river and the RRI peaked prior to
the first flood pulse of 2014. These two hydrologic peaks had the potential to connect at
least one backwater in the riverine portion of the study area and are delineated on the
graphs as a vertical dashed line. A third connection to the riverine backwaters was
identified, however, no sampling occurred after this pulse. Main channel sites tended to
peak in richness post-flood in both years with the exception of the river main channel in
2014. In the riverine backwaters a single sampling event in May of 2014 had a higher
taxonomic richness relative to the other events and was being driven by RIVBW12
(Figure 6c).

Texas Tech University, Matthew Acre, August 2015
25
Figure 6 Larval fish diversity in 10 backwater sites sampled in the upper middle (RIVBW…) and the river-reservoir
interface (RRIBW…) of the Trinity River, Texas from March—July in 2013 and February—June in 2014.
Temperature data in the riverine backwaters are daily averages from all four sites. Temperature data for RRI
backwaters are daily averages from six RRI backwaters. The vertical dashed line indicates when river discharge
was high enough to potentially connect riverine backwaters.

Texas Tech University, Matthew Acre, August 2015
26
Taxonomic richness was relatively consistent for most of the spring both years
among RRI backwaters, but highly variable in riverine backwaters (Figure 6). The high
taxonomic richness values in riverine backwaters during 2014 were primarily due to one
site, RIVBW12 located in the tailrace of Richland Chambers Reservoir (Figure 6). This
backwater is the only site of the riverine habitats to receive connection during the 2013-
2014 sampling seasons (Figure 5). The RRI backwaters were relatively similar in terms
or taxonomic richness and experienced similar changes in taxonomic richness and
composition through time. Backwater sites in 2013 experienced a peak diversity in early
May and a secondary peak later in the year around July. This secondary peak is due to a
shift in species composition from catastomids, percids, White Bass, and Largemouth Bass
early in the sampling year to Lepomis spp. and Channel Catfish.
Canonical Correspondence Analysis
Canonical correspondence analysis indicated that there were three major divisions
amongst sites based on larval fish assemblage structure and physicochemical
characteristics: the RRI (backwaters and main channel), riverine backwaters, and riverine
main channel (Figure 7). The sample sites were primarily separated along the first
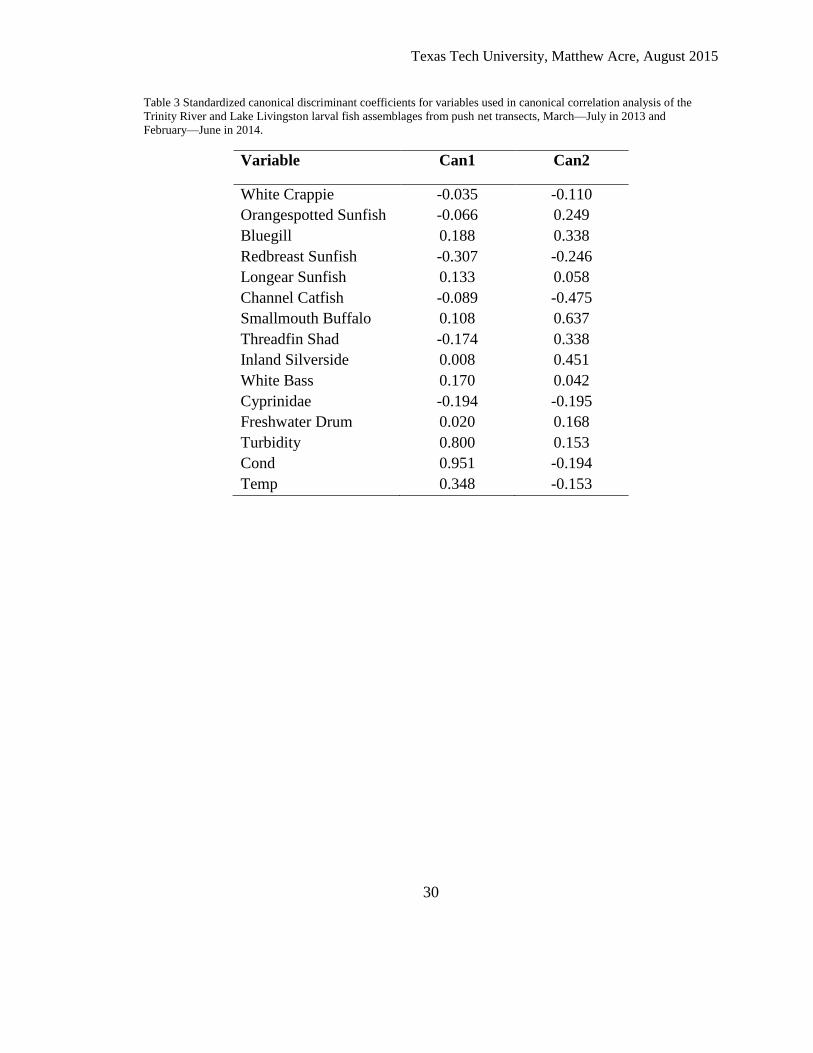
canonical axis (Table 3) which was strongly associated with turbidity and conductivity
and, to a lesser extent, temperature (Table 4). The first canonical axis had the greatest
explanatory power (Table 3) and further analysis of results from the first canonical
dimension suggest there was a difference in the scores along the first canonical

Texas Tech University, Matthew Acre, August 2015
27
dimension among sites (F11,110 = 35.94, P ≤ 0.001), indicating that the strongest driver to
these groupings is their differences in physicochemical properties and to a lesser extent
the composition of their larval fish assemblages (Table 4). In addition to the RRI main
channel, one of the riverine backwater sites (RIVBW14) grouped closely with the RRI
backwaters. Mean conductivity in RIVBW14 was greater than any other sampling
locality, but the site had lower turbidity and temperature than all other sites (Table 1).
However, this site was underrepresented in my samples as it was only sampled early in
the 2013 and 2014 seasons.
The second canonical axis was dominated by the relative abundance of various
fish species (Table 4). However, this axis explained only about 11.8% of the variance and
was not significant (Table 4) despite the habitat types exhibiting different mean scores
(F12,109 = 148.17, P ≤ 0.001). Abundance and occurrences of species varied across sites
and samples. The riverine habitats, particularly the main channel, exhibited relatively
depauperate larval fish assemblages compared to the RRI habitats (Figure 4); however,
the relationships between sites are primarily driven by conductivity, turbidity and to a
lesser extent temperature.

Texas Tech University, Matthew Acre, August 2015
28
Figure 7 Canonical correspondence analysis linking species composition collected in the river backwaters
(RIVBW), river main channel (RIVMC), river-reservoir interface main channel (RRIMC), and river-reservoir
interface backwaters (RRIBW) in March—July in 2013 and February—June in 2014 with physicochemical
variables from the Trinity River and Lake Livingston, Texas. Species that occurred in at least 2% of samples
were used for analyses. The length of each vector indicates the strength of correlation between that variable
and species composition. As the length of the line increases as does the strength of correlation. The direction
of the arrows on axes indicate when a particular species, in relation to species composition and relative
abundance among sites, is most likely to be captured. The Can_1 axis explained 86.6% of the variance with an
eigenvalue of 1.99 and the Can_2 axis described 11.8% of the variance with an eigenvalue of 0.27.

Texas Tech University, Matthew Acre, August 2015
29
Table 2 Canonical correlation analysis results from push net transects of fish assemblage data collected from 12 sites in
the Trinity River and Lake Livingston, Texas March—July in 2013 and February—June in 2014.
Can Canonical
correlation
Variance
explained F df 1 df 2 P
1 0.816 0.866 4.04 45 310 <
0.0001
2 0.462 0.118 1.11 28 210 0.33
Texas Tech University, Matthew Acre, August 2015
30
Table 3 Standardized canonical discriminant coefficients for variables used in canonical correlation analysis of the
Trinity River and Lake Livingston larval fish assemblages from push net transects, March—July in 2013 and
February—June in 2014.
Variable Can1 Can2
White Crappie -0.035 -0.110
Orangespotted Sunfish -0.066 0.249
Bluegill 0.188 0.338
Redbreast Sunfish -0.307 -0.246
Longear Sunfish 0.133 0.058
Channel Catfish -0.089 -0.475
Smallmouth Buffalo 0.108 0.637
Threadfin Shad -0.174 0.338
Inland Silverside 0.008 0.451
White Bass 0.170 0.042
Cyprinidae -0.194 -0.195
Freshwater Drum 0.020 0.168
Turbidity 0.800 0.153
Cond 0.951 -0.194
Temp 0.348 -0.153

Texas Tech University, Matthew Acre, August 2015
31
Chapter V
Discussion
The RRI may contain the most diverse and productive habitats in a river-reservoir
system in terms of adult fish assemblages (Carvalho et al.1998; Oliveira et al. 2004;
Kaemingk et al. 2007; Santos et al. 2010; Terra et al. 2010; Yang et al. 2012; Buckmeier
et al. 2013). My results demonstrate a similar pattern exists for larval fishes. Larval fish
diversity, richness, and relative abundance were greater in the both the RRI backwaters
and RRI main channel than in their counterparts in the riverine habitats. Furthermore,
RRI habitats, both main channel and backwater, were distinct from riverine backwaters
and the riverine main channel in terms of larval fish assemblage structure and
physicochemical conditions. Overall, RRI habitats may serve as an important surrogate
nursery habitat for riverine fishes, particularly for floodplain-dependent species, when
there is a lack of connectivity between the main river channel and floodplain habitats.
Relative Abundance
Lateral connectivity between the river channel and off-channel floodplain habitats
is important in maintaining species abundance and diversity of riverine fishes (Junk et al.
1989; Ward et al. 1999). For larval fishes, floodplain habitats typically have a higher
density of prey species, lower current velocities, and lower predation risk than habitats in
the river channel (Balcombe et al. 2005). However, anthropogenic modifications to river

Texas Tech University, Matthew Acre, August 2015
32
basins, such as dams, reduce the frequency, duration, and predictability of lateral
connectivity to floodplain habitats (Ward and Stanford 1995; Ward 1998; Bunn and
Arthington 2002) and thus have the potential to alter fish recruitment processes and
assemblage structure. In this particular study, riverine backwaters from the Trinity River
were largely isolated from the main channel with the exception of one backwater
(RIVBW12) and this is the only documented connection to floodplain habitats of the sites
sampled. Riverine backwaters in the Trinity River had depressed larval abundances
relative to those in the RRI and the difference in the accessibility of the habitats to the
main river channel is potentially an important factor driving the observed differences in
assemblage structure (Gibbons and Gee 1972; Werner and Hall 1976; Werneret al. 1977;
Tallman and Gee 1982; Schlosser and Toth 1984). Connectivity of lateral floodplain to
the main channel aids in the transport of nutrients essential in maintaining stable fish
populations and can be a limiting factor to floodplain productivity in times of extended
isolation (Junk et al. 1989). In contrast to riverine backwaters, RRI backwaters remained
connected to the main channel throughout the course of this study which likely aided in
nutrient transfer between main channel and backwaters and may have contributed to the
greater larval abundances collected. Overall, our estimates of the relative abundances of
the species encountered were consistent with other surveys for adult fishes in RRI
habitats (Oliveira et al. 2004; Santos et al. 2010; Terra et al. 2010).
The RRI backwaters vary greatly in the size of their connection to the main
channel of the RRI relative to their area (RCA; Table 1), which may be an abiotic factor

Texas Tech University, Matthew Acre, August 2015
33
contributing to the high abundance of larval fishes within them. There are several
potential explanations for why connection size is an important in determining the
abundance and diversity of the larval fish assemblage. The larvae of many marine fishes
can select their habitats once they have developed beyond the post-yolk sac larvae stage
(Leis 2006). Post-yolk sac larvae in freshwater systems may exhibit similar behavioral
patterns and the RCA value may be influencing an increase in the abundance of most
larvae captured during this study. Therefore, it is possible that a larger connection size
may allow more larvae to find a backwater while the smaller area increases my ability to
detect it. Conversely, the smaller connection size decreases the chances larvae will move
into a backwater, and the increased area allows for greater diffusion and lowers detection
probability. Additionally, most larval fish have limited locomotive function and the
method leading their distribution in large rivers at the broad scale should be dominated by
passive means (Bertolo et al. 2012). If larvae are mostly dispersing under passive means
the larger connection size would enable a greater abundance of fishes to be swept into a
backwater in the RRI. It is also possible that assemblages are comprised of permanent
adult residents and many generations live out the entirety of their lives in the RRI
backwaters. However, it is more likely that the RCA value has more of an effect for adult
fishes that are moving throughout the main channel searching for spawning grounds and
locations for egg deposition than yolk-sac larvae actively seeking out backwaters.

Texas Tech University, Matthew Acre, August 2015
34
Species Diversity and Richness
A total of 78 native species and six non-native fishes have been reported from the
Trinity River Basin (Hendrickson and Cohen 2012), of which at least 27 were captured in
this study. The cyprinid family is of particular interest, because this family potentially
represents 21 of the species reported from the Trinity River Basin and could alter our
analysis significantly if parsed to the species level. Cyprinids may have had several
independent occurrences based on their sporadic appearances in collection. This may
indicate multiple species at different times using these mesohabitats which would
corroborate similar finding in the RRI of the Colorado River, Texas (Buckmeier et al.
2013). However, with the current identifications, diversity of larval fishes collected were
greater in the RRI habitats compared to their counterparts in the riverine sites.
Taxonomic richness varied amongst the sites and was higher in the backwaters
relative to the adjacent main channel habitats. Taxonomic richness was also positively
correlated with temperature, but did not exhibit a clear relationship with discharge.
Richness and diversity patterns observed in the riverine backwaters seem to be
disproportionally driven by this high taxonomic richness and diversity in the tailrace of
Richard Chambers Dam (RIVBW12). This site is unique compared to the other riverine
backwaters because it maintained a flow of approximately 1 m3 s-1 in the duration of the
study (L. Byers personal communication.) and is impounded by a low-head dam before it
enters into the Trinity River. Fish populations in this site are likely isolated for much
shorter periods of time than fishes in the other riverine backwaters. It is also possible that

Texas Tech University, Matthew Acre, August 2015
35
fish passage over the low-head dam could have occurred three times, during the sampling
periods, from 2013-2014 contributing to the diversity and richness seen in this particular
riverine backwater which appears to be functioning, in terms of larval fish assemblage,
similarly to RRI habitats.
Previous research has shown that the RRI may be the most species rich and
diverse ecotone of the river-reservoir ecosystem (Kaemingk et al. 2007; Betsill 2012,
Buckmeier et al. 2013), and my results supports these findings. However, the appearance
of Spotted Sucker and the increased abundance of Smallmouth Buffalo in the riverine
habitats suggests the RRI may not be a substitute for riverine backwaters for all riverine
fishes particularly species with more specific spawning requirements. Spotted Sucker
spawn in rock and gravel riffles above pools and have semi-buoyant eggs (Kay et al.
1994) which may also contribute to a low detection rate as my gear was designed to
sample the top 0.5-m of the water column. Smallmouth buffalo have similar spawning
requirements but typically deposit their eggs over freshly inundated floodplains which in
turn may contribute to their higher detection rate relative to Spotted Sucker. Additionally,
even common species of gar in the Trinity River, such as Spotted Gar, were not captured
with the exception of a single specimen from the main channel of the Trinity River. Gar,
in their early life history stages, are attached to plants and detritus and then likely move
into shallow habitats (Alfaro et al. 2008) where our push nets cannot sample.
Year Effect

Texas Tech University, Matthew Acre, August 2015
36
The timing of species appearance in samples was delayed for most species in
2014 relative to 2013. There was also a reduction in abundance observed across most
sites in 2014 which coincides with a precipitous drop in water temperatures in early
March. Reduced abundance of dominant larval ichthyofauna, such as Threadfin Shad, can
decrease competition for resources and result in increased survival rates for less common
ichthyofauna (Byström and Garcia-Berthou 1999; Welker et al. 1994). It is possible that
the later appearance of Threadfin Shad in 2014 reduced competition and contributed to
the first time appearance of Spotted Sucker and the increased abundance of Smallmouth
Buffalo in the riverine backwaters. Both species were captured mostly in RIVBW12, a
completely artificial spillway, and a single Spotted Sucker specimen captured in a large
backwater in the RRI during the same sampling event. The spillway (RIVBW12)
maintains a flow into the main channel of the Trinity throughout the year, though adult
fish are unlikely to make the passage over the low-head dam at the base into the
backwater during lower flows. However, the connection does allow for nutrient transfer
and displayed similar physicochemical characteristics as RRI backwaters (Table 1).
Conclusion and Management Implications
Overall, the backwaters in the RRI appear to be acting as nursery habitat for many
fishes of the Trinity River in times when naturally-occurring backwaters lack
connectivity. Whether RRI species assemblages are comprised mostly of permanent
residents or if these assemblages are dynamic groups with individuals and taxa coming

Texas Tech University, Matthew Acre, August 2015
37
and going in different stages of their life-history cycle is unclear. It is apparent, however,
that the RRI may be important for diversity, richness and abundance for many fishes, but
there was a distinct lack of floodplain-dependent species, particularly Alligator Gar and
Spotted Sucker. The absence of these floodplain-dependent species may indicate that the
RRI may not be a substitute for natural connections to floodplain habitats. A more
targeted effort for specific obligate-floodplain species to determine if it was bias in our
sampling methodology given their early life-history strategies or that these species only
exist in low numbers is needed to determine if the obligate floodplain-species are using
RRI habitats or if there is no legitimate substitute for natural flood pulsing that connects
floodplain habitats. While it may be clear that the RRI is a more productive system than
its riverine counterparts during drier years, an extended study is needed to elucidate the
interactions of less common species and those fishes of conservation need. For example,
the RRI backwaters share connectivity to the main channel and thus share the same
potential pool of species, but I did not observe the same densities, relative abundance, or
assemblage structure across all sites. Research directed at connectivity size, duration, and
distance to main channel would be helpful in the management of riverine fish. My data
suggest that the RRI habitats are important for maintaining abundance of fish populations
but may not be the solution for diversity. Though the RRI may be important in drought
years it is unlikely to be a perfect substitute for natural flood pulsing.

Texas Tech University, Matthew Acre, August 2015
38
Literature Cited
Acre, M. R., and T. B. Grabowski. In Press. Deployment of paired push nets from jet-
propelled kayaks to sample ichthyoplankton. North American Journal of Fisheries
Management.
Alfaro, R., C. González, and A. Ferrara. 2008. Gar biology and culture: status and
prospects. Aquaculture Research 39(7):748–763.
Anderson, A. A., C. Hubbs, K. O. Winemiller, and R. J. Edwards. 1995. Texas freshwater
fish assemblages following three decades of environmental change. The
Southwestern Naturalist 40(3):314-321.
Balcombe, S., S. Bunn, F. McKenzie‐Smith, and P. Davies. 2005. Variability of fish diets
between dry and flood periods in an arid zone floodplain river. Journal of Fish
Biology 67(6):1552–1567.
Baxter, R. M. 1977. Environmental Effects of Dams and Impoundments. Annual Review
of Ecology and Systematics 8:255-283
Betsill, R. K. 2012. Aquatic Ecosystems Research and Coordination. Texas Parks and
Wildlife Department, USFWS Federal Assistance Grant T-53-1 Final Report,
Austin.
Bertolo, A., F. Blanchet, P. Magnan, P. Brodeur, M. Mingelbier, and P. Legendre. 2012.
Inferring Processes from Spatial Patterns: The Role of Directional and Non–
Directional Forces in Shaping Fish Larvae Distribution in a Freshwater Lake
System. PLoS ONE 7(11).
Brinkman, E. L. 2008. Contributions to the life history of alligator gar, Atractosteus
spatula, in Oklahoma. Master’s Thesis. Oklahoma State University, Stillwater,
Oklahoma.
Buckmeier, D. L., N. G. Smith, B. P. Fleming, and K. A. Bodine. 2013. Intra-annual
variation in river-reservoir interface fish assemblages: Implications for fish
conservation and management in regulated waters. River Research and
Applications. Available online DOI: 10.1002/rra.2667
Bunn, S. E., and A. H. Arthington. 2002. Basic principles and ecological consequences of
altered flow regimes for aquatic biodiversity. Environmental Management
30:492-507.

Texas Tech University, Matthew Acre, August 2015
39
Byström, P., and E. Garcia-Berthou. 1999. Dependent growth and size specific
competitive interaction in young fish. Oikos 86(2):217-232.
Carvalho E. D., V. F. Batista da Silva, C. Y. Fujihara, R. Henry, F. Foresti. 1998.
Diversity of fish species in River Paranapanema – Jurumirim Reservoir transition
region (São Paulo, Brazil). Italian Journal of Zoology 65:325-330. DOI:
10.1080/11250009809386841.
Dutterer, A. C., C. Mesing, R. Cailteux, M. S. Allen, W. E. Pine, and P. A. Strickland.
2012. Fish recruitment is influenced by river flows and floodplain inundation at
Apalachicola River, Florida. River Research and Applications. DOI: 10.10002/rra
Dynesius, M., and C. Nilsso. 1994. Fragmentation and flow regulation of river systems in
the northern third of the world. Science 266: 753-762.
Etnier, D. A., W. C. Starnes. 1993. The fishes of Tennessee. University of Tennessee
Press, Knoxville, Tennessee. Available: http://newfoundpress.utk.edu/pubs/fishes/
(June 2015).
Floyd, K. B., W. H. Courtenay, and R. D. Hoyt. 1984. A new larval fish trap: the
quarterfoil trap. The Progressive Fish Culturist 46:216-219.
Galat, D. L., and I. Zweimüller. 2001. Conserving large-river fishes: is the highway
analogy an appropriate paradigm? Journal of the North American Benthological
Society 20:266-279.
Gibbons, J. R. H., and J. H. Gee. 1972. Ecological segregation between Longnose and
Blacknose Dace (Genus Rhinichthys) in the Mink River, Manitoba. Journal of the
Fisheries Research Board of Canada 29(9):1245-1252.
Gido, K. B., D. L. Propst, J. D. Olden, and K. R. Bestgen. 2013. Multidecadal responses
of native and introduced fishes to natural and altered flow regimes in the
American Southwest. Canadian Journal of Fisheries and Aquatic Sciences 70:
554-564.
Guy, C. S., and M. L. Brown. 2007. Science and statistics in fisheries research. Pages 1-
31 in C. S. Guy and M. L. Brown, editors. Analysis and interpretations of
freshwater fisheries data. American Fisheries Society, Bethesda, Maryland.
Hendrickson, D. A. and A. E. Cohen. 2012. Fishes of Texas Project and Online Database
(www.fishesoftexas.org). Published by Texas Natural History Collection, a

Texas Tech University, Matthew Acre, August 2015
40
division of Texas Natural Science Center, University of Texas at Austin.
Accessed April 2015.
Junk, W. J., P. B. Bayley, and R. E. Sparks. 1989. The flood pulse concept in river-
floodplain systems. Canadian special publication of Fisheries and Aquatic
Sciences 106:110-127
Inebnit, T. E., III. 2009. Aspects of the reproductive and juvenile ecology of Alligator
Gar in the Fourche LaFave River, Arkansas. Master’s thesis. University of Central
Arkansas, Conway.
Kaemingk, M. A., B. D. S. Graeb, C. W. Hoagstrom, and D. W. Willis. 2007. Patterns of
fish diversity in a mainstem Missouri River reservoir and associated delta in
South Dakota and Nebraska, USA. River Research and Applications 23:786-791.
DOI:10.1002/rra.1002.
Kay, L. K., R. Wallus, and B. L. Yeager. 1994. Reproductive biology and early life
history of fishes in the Ohio River drainage. Volume 2: Catastomidae. Tennessee
Valley Authority, Chattanooga, Tennessee, USA.
Kluender, E. R. 2011. Seasonal habitat use of a leviathan, Alligator Gar, at multiple
spatial scales in a river-floodplain ecosystem. Master’s thesis. University of
Central Arkansas, Conway.
Leis, J. 2007. Behaviour as input for modelling dispersal of fish larvae: behaviour,
biogeography, hydrodynamics, ontogeny, physiology and phylogeny meet
hydrography. Marine Ecology Progress Series 347:185–193.
Legendre, P., and L. Legendre. 1998. Numerical Ecology, 2nd edition. Elsevier,
Amsterdam.
Lytle, D., and N. Poff. 2004. Adaptation to natural flow regimes. Trends in Ecology &
Evolution 19(2):94100.
Miller, R. R., and J. D. Williams, J. E. 1989. Williams. Extinctions of North American
Fishes during the past century. Fisheries 14:6, 22-38.
Oliveira, E. F., E. Goulart, and C.V. Minte-Vera. 2004. Fish diversity along spatial
gradients in the Itaipu Reservoir, Paraná, Brazil. Brazilian Journal of Biology
64:447-458. DOI: 10.1590/S1519-69842004000300008.

Texas Tech University, Matthew Acre, August 2015
41
Palmieri, A., F. Shah, and A. Dinar. 2001. Economics of reservoir sedimentation and
sustainable management of dams. Journal of environmental management
61(2):149–63.
Perkin, J., and T. Bonner. 2011. Long‐term changes in flow regime and fish assemblage
composition in the Guadalupe and San Marcos rivers of Texas. River Research
and Applications 27(5):566–579.
Perkin, J., and K. Gido. 2011. Stream Fragmentation Thresholds for a Reproductive
Guild of Great Plains Fishes. Fisheries 36(8):371383.
Platania, S. P., and C. S. Altenbach. 1998. Strategies and Egg Types of Seven Rio Grande
Basin Cyprinids. Copeia 3:559-569.
Poff, N. L., J. D. Allan, M. B. Bain, J. R. Karr, K. L. Prestegaard, B. D. Ritcher, R. E.
Sparks, and J. C. Stromberg. 1997. The natural flow regime. BioScience 47:769-
784.
Poff, N. L., and J. K. H. Zimmerman. 2010. Ecological responses to altered flow regimes:
a literature review to inform the science and management of environmental flows.
Freshwater Biology 55:194-205.
Pringle, C., M. Freeman, and B. Freeman. 2000. Regional effects of hydrologic
alterations on riverine macrobiota in the new world: Tropical—temperate
comparisons. BioScience 50(9):807–823.
Naiman RJ, Magnuson JJ, McKnight DM, Stanford JA. 1995. The freshwater imperative:
a research agenda. Washington (DC): Island Press.
[NRC] National Research Council. 1992. Restoration of aquatic systems: science,
technology, and public policy. Washington (DC): National Academy Press.
Rasband, W. S., ImageJ, U. S. National Institutes of Health, Bethesda, Matyland, USA,
http://imagej.nih.gov/ij/, 1997-2014.
Sammons, S. M., and M. J. Maceina. 2009. Effects of river flows on growth of Redbreast
Sunfish Lepomis auritus (Centrachidae) in Georgia rivers. Journal of Fish Biology
74:1580-1593.
Santos, A. B. I., B. F. Terra, and F. G. Araújo. 2010. Influence of the river flow on the
structure of fish assemblage along the longitudinal gradient from river to
reservoir. Zoologia 27: 732-740. DOI: 10.1590/S1984-4670210000500010.

Texas Tech University, Matthew Acre, August 2015
42
Schlosser, I. J., L. A. Toth. 1984. Niche relationships and population ecology of Rainbow
(Etheostoma caeruleum) and Fantail (E. flabellare) darters in a temporally
variable environment. Oikos 42(2):229-238.
Simon, T. P., and R. Wallus. 1989. Contributions to the early life histories of gar
(Actinopterygii: Lepisosteidae) in the Ohio and Tennessee River basins with
emphasis on larval development. Transactions Kentucky Academy of Science
50(1-2):59-74.
Simon, P., R. T. Wallus, and B. L. Yeager. 2004. Reproductive biology and early life
history of fishes in the Ohio River drainage. Volume 3: Ictaluridae. Tennessee
Valley Authority, Chattanooga, Tennessee, USA.
Simon, P., R. T. Wallus, and B. L. Yeager. 2006. Reproductive biology and early life
history of fishes in the Ohio River drainage. Volume 4: Percidae. Tennessee
Valley Authority, Chattanooga, Tennessee, USA.
Snyder, D. E., and S. C. Seal. 2008 onwards. Computer-interactive key to families of
larval fishes in freshwaters of the United States and Canada (Trinity River-Lake
Livingston larval fish collections). Larval Fish Laboratory, Colorado State
University, Fort Collins, Colorado. Available:
http://welcome.warnercnr.colostate.edu/lfl-files-to-download.html; select
distribution file fam-na.zip. (March 2013).
Tallman, R. F., and J. H. Gee. 1982. Intraspecific resource partioning in a headwaters
stream fish, the pearl dace Semotilus margarita (Cyprinidae). Environmental
Biology of Fishes 7(3):243-249.
Terra, B. F., A. B. I. Santos, and F. G. Araújo. 2010. Fish assemblage in a dammed
tropical river: an analysis along the longitudinal and temporal gradients from river
to reservoir. Neotropical Ichthyology 8:599-606.
Wallus, R. T., P. Simon, and B. L. Yeager. 1990. Reproductive biology and early life
history of fishes in the Ohio River drainage. Volume 1: Acipenseridae through
Esocidae. Tennessee Valley Authority, Chattanooga, Tennessee, USA.
Wallus, R. T., P. Simon, and B. L. Yeager. 2006. Reproductive biology and early life
history of fishes in the Ohio River drainage. Volume 5: Aphredoderidae through
Cottidae, Moronidae, and Sciaenidae. Tennessee Valley Authority, Chattanooga,
Tennessee, USA.

Texas Tech University, Matthew Acre, August 2015
43
Wallus, R. T., P. Simon, and B. L. Yeager. 2008. Reproductive biology and early life
history of fishes in the Ohio River drainage. Volume 6: Elassomatidae and
Centrarchidae. Tennessee Valley Authority, Chattanooga, Tennessee, USA.
Ward, J. V., and J. A. Stanford. 1995. The serial discontinuity concept: extending the
model to floodplain rivers. Regulated Rivers: Research and Management 10:159-
168.
Ward, J. V., and J. A. Stanford. 1995. Ecological connectivity in alluvial river
ecosystems and its disruption by flow regulation. Regulated Rivers: Research and
Management 11:105-119.
Ward, J. V. 1998. Riverine landscapes: Biodiversity patterns, disturbance regimes, and
aquatic conservation. Biological Conservation 83(3):269-278.
Welcomme, R. L., K. O. Winemiller, and I. G. Cowx. 2006. Fish environmental guilds as
a tool for assessment of ecological condition of rivers. River research and
Applications 22:377-396.
Welker, M. T., C. L. Pierce, and D. H. Wahl. 1994. Growth and survival of larval fishes:
roles of competition and zooplankton abundance. Transactions of the American
Fisheries Society 123(5):703-717.
Werner, E. E., and D. J. Hall. 1977. Competition and habitat shift in two sunfishes
(Centrarchidae). Ecology 58(4):869-876.
Yang S., X. Gao, M. Li, B. Ma, and H. Liu. 2012. Interannual variations of the fish
assemblage in the transitional zone of the Three Gorges Reservoir: persistence
and stability. Environmental Biology of Fishes 93:295-304. DOI:
10.1007/s10641-011-9936-6.

Texas Tech University, Matthew Acre, August 2015
44
Appendix
Table A.1 Mean (±SE) CPUE and H’ for larval fishes captured in 2013 and 2014 from the Trinity River, Texas by
site. The riverine sites from the middle Trinity River represented by the river backwaters (RIVBW) and the river
main channel (RIVMC), and the river-reservoir interface (RRI) sites are represented by the RRI backwaters
(RRIBW), and the RRI main channel (RRIMC).
Texas Tech University, Matthew Acre, August 2015
45
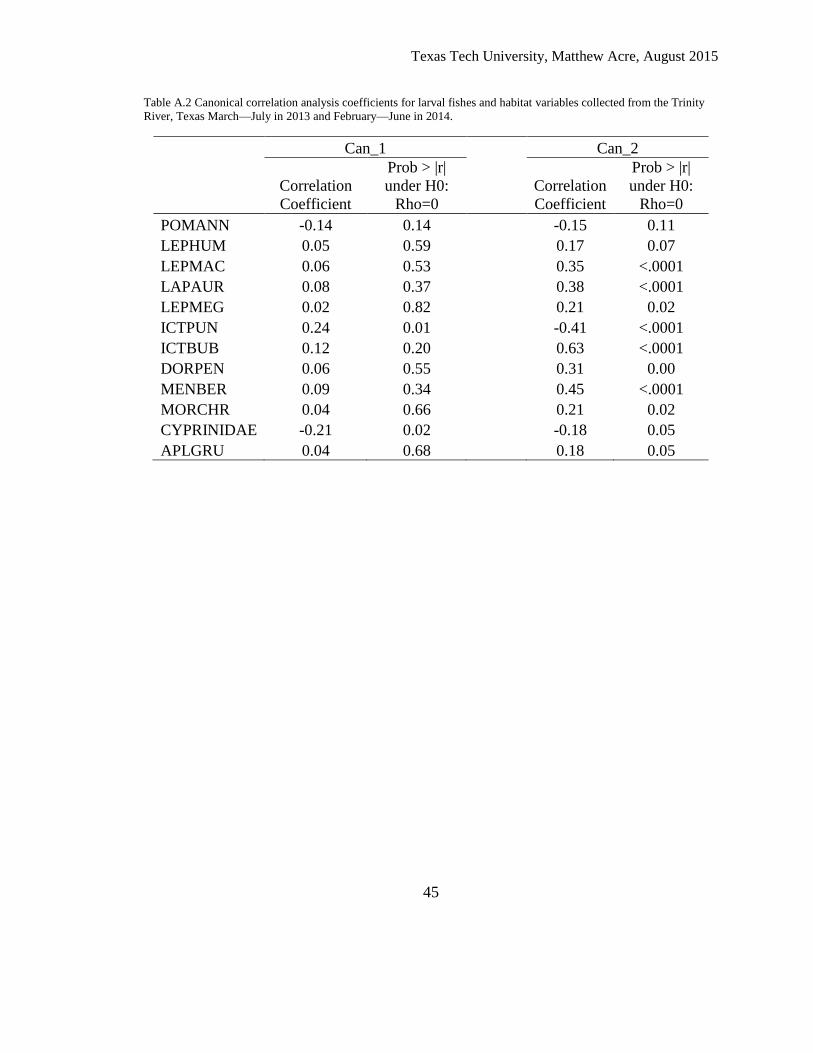
Table A.2 Canonical correlation analysis coefficients for larval fishes and habitat variables collected from the Trinity
River, Texas March—July in 2013 and February—June in 2014.
Can_1 Can_2
Correlation
Coefficient
Prob > |r|
under H0:
Rho=0
Correlation
Coefficient
Prob > |r|
under H0:
Rho=0
POMANN -0.14 0.14 -0.15 0.11
LEPHUM 0.05 0.59 0.17 0.07
LEPMAC 0.06 0.53 0.35 <.0001
LAPAUR 0.08 0.37 0.38 <.0001
LEPMEG 0.02 0.82 0.21 0.02
ICTPUN 0.24 0.01 -0.41 <.0001
ICTBUB 0.12 0.20 0.63 <.0001
DORPEN 0.06 0.55 0.31 0.00
MENBER 0.09 0.34 0.45 <.0001
MORCHR 0.04 0.66 0.21 0.02
CYPRINIDAE -0.21 0.02 -0.18 0.05
APLGRU 0.04 0.68 0.18 0.05

Texas Tech University, Matthew Acre, August 2015
46
Table A.3 Taxonomic richness and mean species richness ± SD of push net (PN) and light trap (LT) samples taken
from riverine backwaters (RIVBW), riverine main channel (RIVMC), river-reservoir interface backwaters (RRIBW),
and river-reservoir interface main channel (RRIMC) from the Trinity River and Lake Livingston, Texas March—July
in 2013 and February—June in 2014.
Mesohabitat Gear
Species
Count
Mean ± SD
RIVBW PN 22 0.77 ± 1.04
LT 7 0.16 ± 0.48
RIVMC PN 8 0.48 ± 0.80
LT 2 0.10 ± 0.31
RRIBW PN 23 1.70 ± 1.38
LT 14 0.63 ±1.10
RRIMC PN 16 1.30 ±1.23
LT 10 0.78 ± 0.99

Texas Tech University, Matthew Acre, August 2015
47
Table A.4 Fish collected in the Trinity River, Texas March—July in 2013 and February—June in 2014 by site and year. An ‘x’ signifies a species
was captured at that site and a blank space indicates that the species was not captured.
2013 2014 2013 2014 2013 2014 2013 2014 2013 2014 2013 2014 2013 2014 2013 2014 2013 2014 2013 2014 2013 2014 2013 2014
Spotted Gar x
Threadfin Shad x x x x x x x x x x x x x x x x x x x x x x
Cyprinidae x x x x x x x x x x x x x x x x x x x x x
Smallmouth Buffallo x x x x x x x x x
Spotted Sucker* x x x x
Blue Catfish x x x x x x
Channel Catfish x x x x x x x x x
Tadpole Madtom x
Flathead Catfish x
Inland Silverside x x x x x x x x x x x x x x x x x x x x x x x
Sheepshead Minnow x x
White Bass x x x x x x x x x x x x x x x x x
Redbreast Sunfish x x x x x x
Green Sunfish x x x x x x
Warmouth x x x x x
Orangespotted Sunfish x x x x x x x x x x x x x x x x
Bluegill x x x x x x x x x x x x x x x x
Longear Sunfish x x x x x x x x x x x
Sunfush species x x x x x
Largemouth Bass x x x x
White Crappie x x x x x x x x x
Black Crappie x
Bluntnose Darter x x x x x x
Harlequin Darter x
Bigscale Logperch x x x x x x
Percina spp . x x x
Freshwater Drum x x x x x x x x x
RRIBW3 RRIBW4 RRIBW5 RRIBW6 RRIBW7 RRIMCRIVBW9 RIVBW12 RIVBW13 RIVBW14 RIVMC RRIBW2
RRIRiver
Related Documents











