MATRIX METALLOPROTEINASES 2 AND 9 IN THE COCHLEA: EXPRESSION AND ACTIVITY AFTER AMINOGLYCOSIDE EXPOSITION C. SETZ, a,b1 Y. BRAND, a,b1 V. RADOJEVIC, a,b C. HANUSEK, a,b P. J. MULLEN, a S. LEVANO, a,c A. LISTYO a,b AND D. BODMER a,b * a Department of Biomedicine, University Hospital Basel, Hebelstrasse 20, 4031 Basel, Switzerland b Clinic for Otolaryngology, Head and Neck Surgery, University Hospi- tal Basel, Petersgraben 4, 4031 Basel, Switzerland c Department of Anaesthesiology, University Hospital Basel, Spital- strasse 21, 4031 Basel, Switzerland Abstract—The matrix metalloproteinases (MMPs) are a family of proteins involved in the remodelling and homeostasis of the extracellular matrix. These proteases have been well studied in the retina and the brain, marking their importance in neuronal cell survival and death [Chintala (2006) Exp Eye Res 82:5–12; Candelario-Jalil et al. (2009) Neuroscience 158: 983–994]. The neuroepithelia of the eye and the inner ear share common characteristics. Therefore, we hypothesized that MMPs could play a similar role in the cochlea as de- scribed in the retina. We focused on the localization and function of MMP-2 and MMP-9 in the cochlea, by determining their expression and activity under normal conditions and after cochlear damage via aminoglycoside exposition. We examined their expression in 5-day-old Wistar rat cochleas by RT-PCR, real-time PCR, and Western blot. We used immu- nohistochemistry to investigate their location in the cochleas of adult C57BL/6 mice. We also determined whether or not the exposure of the organs of Corti to aminoglycosides would change MMP-2 and MMP-9 expression patterns. Western blot- ting identified MMP-2 and MMP-9 in neonatal spiral ganglion, stria vascularis, and to a lesser extent the organ of Corti. Neonatal mRNA expression of MMP-2 was approximately equivalent in all three tissues, while MMP-9 mRNA was high- est in spiral ganglion. Immunohistochemistry showed MMP-2 primarily in adult spiral ganglion neurons and inner hair cells, while MMP-9 was found mainly in spiral ganglion neurons, inner hair cells and supporting cells. Organs of Corti treated with gentamicin for 24 h showed an upregulation of MMP-2 and MMP-9 proteins, but did not show a significant upregu- lation of mRNA expression 3, 6, 12, 24, and 36 h after genta- micin exposure. Inhibition of MMP activity in organs of Corti incubated with an MMP inhibitor in organotypic cultures re- sulted in hair cell death—suggesting that a basal level of MMP activity is required for hair cell survival. © 2011 IBRO. Published by Elsevier Ltd. All rights reserved. Key words: apoptosis, extracellular matrix, hair cell, inner ear, retina. The mammalian cochlea is composed of highly specialised cells, including the outer and inner hair cells (HCs) in the organ of Corti (OC). The different cell types are embedded in a highly organized extracellular matrix (ECM). Homeo- stasis within the ECM is maintained by regulating the turnover of matrix composition. The matrix metalloprotei- nases (MMPs) are zinc-dependent proteases that play an important role in modelling the ECM. They can be divided into four main subgroups based on common structural domains: collagenases, gelatinases, stromelysins, and membrane-type MMPs (Rosenberg, 2009). The highly in- tegrated action of these enzymes remodels the matrix and performs functions in signalling, cell survival and cell death (Candelario-Jalil et al., 2009). With the exception of MMP-2, which is constitutively expressed in various tis- sues, MMPs are produced only when necessary, they can rapidly be induced in response to exogenous signals such as growth factors and cytokines (Chintala, 2006). MMPs cleave most components of the ECM including fibronectin, laminin, proteoglycans, and type IV collagen (Sternlicht and Werb, 2001; Rosenberg, 2002). In addition, MMPs can also process other pro-MMPs, cytokines, and pro-neu- rotrophins including pro-NGF and pro-BDNF (Schönbeck et al., 1998; Cauwe et al., 2007; Lee et al., 2001; Sternlicht and Werb, 2001). Regulation of MMP expression and ac- tivation is very complex and tightly controlled. MMPs are synthesized as zymogens and are secreted into the extra- cellular space in their inactive pro-form. The pro-MMPs are then activated by disruption of the zinc-thiol interaction between the catalytic site and the pro-domain. The pro- peptide of the zymogen has to be proteolytically cleaved by other MMPs or proteases for an MMP to be active (Stern- licht and Werb, 2001). The proteases plasmin, tissue plas- minogen activator (tPA), and urokinase-type plasminogen activator (uPA) are important physiological activators of 1 Both authors contributed equally to the work. *Correspondence to: D. Bodmer, Clinic for Otolaryngology, Head and Neck Surgery, University Hospital Basel, Petersgraben 4, 4031 Basel, Switzerland. Tel: 41-61-328-7603; fax: 41-61-265-4029. E-mail address: [email protected] (D. Bodmer). Abbreviations: B, brain; BCA, bicinchoninic acid; bp, base pairs; BSA, bovine serum albumin; cDNA, complementary deoxyribonucleic acid; CNFs, cochlear nerve fibers; DAPI, 4=,6-diamidino-2-phenylindole; DNase, deoxyribonuclease; ECM, extracellular matrix; EDTA, ethyl- ene diamine tetraacetic acid; FCS, fetal calf serum; GAPDH, glycer- aldehyde-3-phosphate dehydrogenase; HC, hair cell; HEPES, 4-(2- hydroxyethyl)-1-piperazineethanesulfonic acid; HL-60, human promy- elocytic leukemia HL 60 cell line; HRP, horseradish peroxidase; IHC, inner hair cell; M, marker; MMP, matrix metalloproteinase; mRNA, messenger ribonucleic acid; NCBI, National Center for Biotechnology Information; NF-B, nuclear factor “kappa-light-chain-enhancer” of ac- tivated B-cells; nt, nucleotides; NTC, no template control; OC, organ of Corti; OHC, outer hair cell; PBS, phosphate buffered saline; PBS-T, phosphate buffered saline associated with 0.1% Tween; PCR, poly- merase chain reaction; PiC, pillar cells; RT-PCR, reverse transcriptase polymerase chain reaction; SDS-PAGE, sodium dodecyl sulfate poly- acrylamide gel electrophoresis; SG, spiral ganglion; SV, stria vascu- laris; TIMPs, tissue inhibitors of metalloproteinases; T m , melting tem- perature; tPA, tissue plasminogen activator; uPA, urokinase-type plas- minogen activator. Neuroscience 181 (2011) 28 –39 0306-4522/11 $ - see front matter © 2011 IBRO. Published by Elsevier Ltd. All rights reserved. doi:10.1016/j.neuroscience.2011.02.043 28

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

2
t
s
*NSEAbCDeaheimI
Neuroscience 181 (2011) 28–39
MATRIX METALLOPROTEINASES 2 AND 9 IN THE COCHLEA:
EXPRESSION AND ACTIVITY AFTER AMINOGLYCOSIDE EXPOSITIONaareatsctbpolm
C. SETZ,a,b1 Y. BRAND,a,b1 V. RADOJEVIC,a,b
C. HANUSEK,a,b P. J. MULLEN,a S. LEVANO,a,c
A. LISTYOa,b AND D. BODMERa,b*aDepartment of Biomedicine, University Hospital Basel, Hebelstrasse0, 4031 Basel, Switzerland
bClinic for Otolaryngology, Head and Neck Surgery, University Hospi-al Basel, Petersgraben 4, 4031 Basel, Switzerland
cDepartment of Anaesthesiology, University Hospital Basel, Spital-trasse 21, 4031 Basel, Switzerland
Abstract—The matrix metalloproteinases (MMPs) are a familyof proteins involved in the remodelling and homeostasis ofthe extracellular matrix. These proteases have been wellstudied in the retina and the brain, marking their importancein neuronal cell survival and death [Chintala (2006) Exp EyeRes 82:5–12; Candelario-Jalil et al. (2009) Neuroscience 158:983–994]. The neuroepithelia of the eye and the inner earshare common characteristics. Therefore, we hypothesizedthat MMPs could play a similar role in the cochlea as de-scribed in the retina. We focused on the localization andfunction of MMP-2 and MMP-9 in the cochlea, by determiningtheir expression and activity under normal conditions andafter cochlear damage via aminoglycoside exposition. Weexamined their expression in 5-day-old Wistar rat cochleasby RT-PCR, real-time PCR, and Western blot. We used immu-nohistochemistry to investigate their location in the cochleasof adult C57BL/6 mice. We also determined whether or not theexposure of the organs of Corti to aminoglycosides wouldchange MMP-2 and MMP-9 expression patterns. Western blot-ting identified MMP-2 and MMP-9 in neonatal spiral ganglion,stria vascularis, and to a lesser extent the organ of Corti.Neonatal mRNA expression of MMP-2 was approximatelyequivalent in all three tissues, while MMP-9 mRNA was high-
1 Both authors contributed equally to the work.Correspondence to: D. Bodmer, Clinic for Otolaryngology, Head andeck Surgery, University Hospital Basel, Petersgraben 4, 4031 Basel,witzerland. Tel: �41-61-328-7603; fax: �41-61-265-4029.-mail address: [email protected] (D. Bodmer).bbreviations: B, brain; BCA, bicinchoninic acid; bp, base pairs; BSA,ovine serum albumin; cDNA, complementary deoxyribonucleic acid;NFs, cochlear nerve fibers; DAPI, 4=,6-diamidino-2-phenylindole;Nase, deoxyribonuclease; ECM, extracellular matrix; EDTA, ethyl-ne diamine tetraacetic acid; FCS, fetal calf serum; GAPDH, glycer-ldehyde-3-phosphate dehydrogenase; HC, hair cell; HEPES, 4-(2-ydroxyethyl)-1-piperazineethanesulfonic acid; HL-60, human promy-locytic leukemia HL 60 cell line; HRP, horseradish peroxidase; IHC,
nner hair cell; M, marker; MMP, matrix metalloproteinase; mRNA,essenger ribonucleic acid; NCBI, National Center for Biotechnology
nformation; NF-�B, nuclear factor “kappa-light-chain-enhancer” of ac-tivated B-cells; nt, nucleotides; NTC, no template control; OC, organ ofCorti; OHC, outer hair cell; PBS, phosphate buffered saline; PBS-T,phosphate buffered saline associated with 0.1% Tween; PCR, poly-merase chain reaction; PiC, pillar cells; RT-PCR, reverse transcriptasepolymerase chain reaction; SDS-PAGE, sodium dodecyl sulfate poly-acrylamide gel electrophoresis; SG, spiral ganglion; SV, stria vascu-laris; TIMPs, tissue inhibitors of metalloproteinases; Tm, melting tem-
aperature; tPA, tissue plasminogen activator; uPA, urokinase-type plas-minogen activator.
0306-4522/11 $ - see front matter © 2011 IBRO. Published by Elsevier Ltd. All righdoi:10.1016/j.neuroscience.2011.02.043
28
est in spiral ganglion. Immunohistochemistry showed MMP-2primarily in adult spiral ganglion neurons and inner hair cells,while MMP-9 was found mainly in spiral ganglion neurons,inner hair cells and supporting cells. Organs of Corti treatedwith gentamicin for 24 h showed an upregulation of MMP-2and MMP-9 proteins, but did not show a significant upregu-lation of mRNA expression 3, 6, 12, 24, and 36 h after genta-micin exposure. Inhibition of MMP activity in organs of Cortiincubated with an MMP inhibitor in organotypic cultures re-sulted in hair cell death—suggesting that a basal level ofMMP activity is required for hair cell survival. © 2011 IBRO.Published by Elsevier Ltd. All rights reserved.
Key words: apoptosis, extracellular matrix, hair cell, innerear, retina.
The mammalian cochlea is composed of highly specialisedcells, including the outer and inner hair cells (HCs) in theorgan of Corti (OC). The different cell types are embeddedin a highly organized extracellular matrix (ECM). Homeo-stasis within the ECM is maintained by regulating theturnover of matrix composition. The matrix metalloprotei-nases (MMPs) are zinc-dependent proteases that play animportant role in modelling the ECM. They can be dividedinto four main subgroups based on common structuraldomains: collagenases, gelatinases, stromelysins, andmembrane-type MMPs (Rosenberg, 2009). The highly in-tegrated action of these enzymes remodels the matrix andperforms functions in signalling, cell survival and cell death(Candelario-Jalil et al., 2009). With the exception ofMMP-2, which is constitutively expressed in various tis-sues, MMPs are produced only when necessary, they canrapidly be induced in response to exogenous signals suchas growth factors and cytokines (Chintala, 2006). MMPscleave most components of the ECM including fibronectin,laminin, proteoglycans, and type IV collagen (Sternlichtnd Werb, 2001; Rosenberg, 2002). In addition, MMPs canlso process other pro-MMPs, cytokines, and pro-neu-otrophins including pro-NGF and pro-BDNF (Schönbeckt al., 1998; Cauwe et al., 2007; Lee et al., 2001; Sternlichtnd Werb, 2001). Regulation of MMP expression and ac-ivation is very complex and tightly controlled. MMPs areynthesized as zymogens and are secreted into the extra-ellular space in their inactive pro-form. The pro-MMPs arehen activated by disruption of the zinc-thiol interactionetween the catalytic site and the pro-domain. The pro-eptide of the zymogen has to be proteolytically cleaved byther MMPs or proteases for an MMP to be active (Stern-
icht and Werb, 2001). The proteases plasmin, tissue plas-inogen activator (tPA), and urokinase-type plasminogen
ctivator (uPA) are important physiological activators ofts reserved.

rkngahcpMseechhpe
eadrtv
pdc
fcitcA
mM(t
t(c
C. Setz et al. / Neuroscience 181 (2011) 28–39 29
the MMPs (Gasche et al., 2006). There is also protease-independent activation of MMPs possible either by oxida-tion or S-nitrosylation, which can unmask the catalyticdomain and activate MMPs without pro-domain cleavage(Gu et al., 2002; Meli et al., 2003; Pei et al., 2006). MMPactivity is controlled at multiple levels such as transcrip-tional regulation (Crawford and Matrisian, 1996), proen-zyme activation (Kleiner and Stetler-Stevenson, 1993) andby specific tissue inhibitors (TIMPs) (Murphy and Do-cherty, 1992).
Studies of MMPs in the neurosensory epithelium of theeye and brain revealed the important function of theseproteases in retinal and neuronal cell survival and death(Chintala, 2006; Candelario-Jalil et al., 2009). Studies incerebral hypoxia/ischemia in rodents and non-human pri-mates showed an elevation in MMP-2 expression after twohours of reperfusion (Chang et al., 2003; Yang et al.,2007). A marked increase in MMP-9 between 24 and 48 hafter reperfusion provides a molecular basis for both thetransient and long-term alterations that occur in the blood–brain barrier due to MMPs in reperfusion injury (Yang et al.,2007). In human ischemic strokes, active MMP-2 is in-creased first on days 2–5 compared with active MMP-9,which is elevated up to months after the ischemic episode(Clark et al., 1997). MMP-2 expression has been detectedin astrocytes and axons of the optic nerve head and inretinal ganglion cells of the human retina (Agapova et al.,2001). Furthermore, studies showed that ischemic-in-duced retinal ganglion cell loss in mice correlates with anup-regulation of MMP-9. In addition, MMP-9 knockoutmice, and mice treated with MMP synthetic inhibitor, areresistant to ischemic-induced retinal ganglion cell loss,indicating that MMP-9 plays a causative role (Chintala etal., 2002; Zhang and Chintala, 2004).
In contrast to the studies of MMPs in the brain and theetina, the role of MMPs in the inner ear is largely un-nown. Since both the retina and the inner ear arise fromeuroepithelium, both harbour sensitive sensory cells to-ether with supporting cells and both display a complexnd highly organised micro-architecture of the ECM. Weypothesized that MMPs may play an important role inochlear homeostasis. However, there are only a few re-orts of MMPs in the cochlea. A mouse model, where thepv17 gene, encoding a peroxismal protein, is inactivated,
hows a strong increase in MMP-2 expression in the innerar (Reuter et al., 1998). These mice developed a degen-ration of various inner ear structures, including a loss ofochlear neurons and degeneration of the OC along withearing loss. Previous studies also showed that mice withyperhomocysteinemia display an increase in cochlear ex-ression of MMP-2 and MMP-9 (Kundu et al., 2009). How-ver, hearing in these mice has not yet been investigated.
In this study, our experimental goal was to analyze thexpression of the two related MMPs, MMP-2 (gelatinase A)nd MMP-9 (gelatinase B), in the cochlea. These MMPsegrade gelatine, type IV collagen, collagen XVIII, fib-onectin, laminin, and different proteoglycans present inhe basement membranes (Matrisian, 1992; Stetler-Ste-
enson et al., 1993). In addition, we investigated the ex-ression of MMP-2 and MMP-9 after aminoglycoside-in-uced HC damage to further elucidate their roles in theochlea.
EXPERIMENTAL PROCEDURES
The following methods were used to answer these experimentalquestions:
● Determining expression of MMP-2 and MMP-9 mRNA in thecochlea by RT-PCR.
● Determining expression levels of MMP-2 and MMP-9(mRNA and protein) in the cochlea by real-time PCR andWestern blotting.
● Determining whether exposure of OC explants to an amino-glycoside results in expression changes of MMP-2 andMMP-9 mRNA/protein in the OC by real-time PCR andWestern blotting.
● Determining the localization of MMP-2 and MMP-9 on co-chlear sections using immunohistochemistry.
● Determining the effect of MMP-inhibition in OC explants(alone and in combination with an aminoglycoside) using HCcounting.
Tissue extraction
All animal procedures were carried out according to an approvedanimal research protocol (Kantonales Veterinäramt, Basel, Swit-zerland). For every experiment, an equal gender distribution wasensured. We used animal models, which are frequently involved inexperiments concerning inner ear problems. For tissue extractionand culture, 5-day-old Wistar (WS) rat pups (Harlan, Indianopolis,USA) were used. The rats were decapitated, and cochlear micro-dissections were performed under a light microscope to isolate theOC, the spiral ganglion (SG), and the stria vascularis (SV). Braintissue was also removed from the same animals. All tissues weremaintained in chilled PBS during microdissections. Brain tissuewas used as a positive control in PCR, real-time PCR and West-ern blot, since the central nervous system has been reported toexpress MMP-2 and MMP-9 (Yong et al., 2001). In addition, wechose the human promyelocytic leukemia HL-60 cell line as apositive control for the MMP-9 Western blot.
OC culture
For experiments in which cultures of OCs were needed, OCs werefirst incubated in culture medium [Dulbecco’s Modified Eagle Me-dium supplemented with 10% FCS, 25 mM HEPES and 30 U/mlpenicillin (Invitrogen, Carlsbad, USA) at 37 °C in 5% CO2] and leftor 24 h at 37 °C in 5% CO2 for recovery. After that period, ahosen number of OCs were transferred into a new solution andncubated at different time points at 37 °C in 5% CO2. The solu-ions contained either of the following: culture medium alone,ulture medium with the presence of 0.25 mM gentamicin (Sigma-ldrich, St. Louis, USA), culture medium with the presence of 50
�M GM6001 MMP inhibitor (Merck, Darmstadt, Germany), cultureedium with the presence of 0.25 mM gentamicin and GM6001MP inhibitor, or culture medium with GM6001 negative control
Merck, Darmstadt, Germany). Each experiment was repeatedhree times.
Antibodies
The following primary antibodies were used for Western blot-ing and immunohistochemistry: anti-MMP-2 mouse monoclonalMAB3308, Chemicon, Temecula, USA), anti-MMP-9 rabbit poly-lonal (AB19016, Chemicon, Temecula, USA), and anti-�-actin
mouse monoclonal (sc-81178, Santa Cruz Biotech, Santa Cruz,
USA).
Ged
fikwea
mde1
Pgm
ukBi
T
T
C. Setz et al. / Neuroscience 181 (2011) 28–3930
RNA extraction
For PCR, 20 cochleae of 5-day-old WS rat pups were obtained,and the OC, SG and SV dissected and placed separately inRNAlater (Qiagen, Hombrechtikon, Switzerland). 20–40 mg ofbrain tissue from the same animals was also taken and served asa positive control. For real-time PCR, six OCs were incubated percondition in culture medium with or without 0.25 mM gentamicinfor 12, 24, and 36 h. In a second experiment, a 3 and 6 hincubation time was used. The organs were transferred to tubescontaining RNAlater. RNA isolation of brain tissue and inner earcomponents was performed using the RNAeasy Minikit (Qia-gen, Switzerland) including DNase treatment according to thesupplier’s instructions. An Ultra-Turrax T8 homogenizer (IKA-Werke, Staufen, Germany) was used to homogenize the tis-sues. The quantity and quality of the isolated RNA was deter-mined with NanoDrop ND 1000 (NanoDrop Technologies, DE,USA). The 260/280 nm ratio of all our samples was between 1.8and 2.1.
Primer design
Gene sequences from MMP-2 (NM_031054.2), MMP-9 (NM_031055.1), and �-actin (NM_031144.2) were accessed from
enBank. Primers for common PCR were designed using Prim-r-Blast software available at the NCBI. Our criteria for primeresign included Tm values between 58 and 60 °C, a minimum
length of 20 nt, and a product size of 100 –500 bp with anabsence of long G-C stretches. Primers were designed to crossat least one exon junction for the specific amplification of cDNAto avoid amplification of genomic DNA. The primers sequences,annealing temperatures and product sizes, are provided in Table1. For real-time PCR, primers were designed using the UniversalProbeLibrary Assay Design Center (Roche Applied Biosciences,Rotkreuz, Switzerland).
PCR
Total RNA (1 �g) was reverse transcribed to cDNA with therst-strand cDNA synthesis kit (Roche Applied Biosciences, Rot-reuz, Switzerland) according to the supplier’s instructions. PCRas performed using the PCR Master Mix (Roche Applied Biosci-nces, Rotkreuz, Switzerland) with primers specific for MMP-2nd MMP-9. �-actin primers were used as a positive control for
cDNA synthesis. The primer sets are described in Table 1.PCR reactions were run three independent times in the Ep-
pendorf Mastercycler (Eppendorf, Hamburg, Germany) under thefollowing conditions.
For MMP-2, the initial denaturation as started at 94 °C for 3min, followed by 35 cycles. Each cycle consisted of denaturationat 94 °C for 30 s, annealing at 60 °C for 30 s and extension at72 °C for 30 s, a final extension step was performed at 72 °C for5 min.
For MMP-9, touchdown PCR parameters were used. The initialdenaturation was started at 94 °C for 3 min. The first seven cyclesconsisted of denaturation at 94 °C for 30 s, annealing at 64 °C for
able 1. Primer sequences used for MMP-2, MMP-9 and �-actin
Gene Primer name Sequence 5= ¡ 3=
MMP-2 F2 ACCAGAACACCATCGAGACCR2 GAGTCCACAGCTCATCGTCA
MMP-9 F9 ACCACTAAAGGTCGCTCGGAR9 AGTACTGCTTGCCCAGGAAG
�-actin FA ACGGTCAGGTCATCACTATCGRA ATCCTGTCAGCAATGCCTGGG
30 s and extension at 72 °C for 30 s. An annealing temperature of
61 °C was used for cycles 8 to 14, and for cycles 15 to 21 theannealing temperature was decreased to 58 °C. The last 19 cycleswere performed at 55 °C. A final extension step was performed at72 °C for 5 min.
For �-actin, the initial denaturation was started at 94 °C for 3in that was followed by 30 cycles. Each cycle consisted ofenaturation at 94 °C for 30 s, annealing at 58 °C for 45 s andxtension at 72 °C for 30 s, with a final extension step at 72 °C formin.
The PCR products were stained with SybrGreen I (Molecularrobes, OR, USA), separated by electrophoresis on a 2% agaroseel and visualized under UV light. Omission of cDNA in the PCRixture served as negative control.
The PCR fragments of MMP-2 and MMP-9 were sequencedsing the PCR primers, the BigDye terminator cycle sequencingit and the ABI Prism 3100 Avant Genetic Analyzer (Appliediosystems, Foster City, USA) according to the manufacturer’s
nstructions.
Real-time PCR
Total RNA (500 ng) was reverse transcribed into cDNA with thefirst strand cDNA synthesis kit (Roche Applied Biosciences, Rot-kreuz, Switzerland) according to supplier’s instructions. The reac-tion took place in an ABI Prism 7900HT Sequence Detection
Fig. 1. Detection of MMP-2 and MMP-9 mRNA in the cochlea of5-d-old Wistar rats. Total RNA of the OC, SG and SV was reversedtranscribed into cDNA and amplified by PCR. Total brain (B) RNAserved as a positive control. Omission of RT reaction served as notemplate control (NTC). The DNA ladder (M) is indicated with its
Annealing temperature Exons Product length
60 2 375 bp3
58 6 169 bp7
58 3 208 bp4
TGGTTACGAAGCA
lengths. �-actin was used to control cDNA synthesis.
Mp
ifvBra
b
ie
C. Setz et al. / Neuroscience 181 (2011) 28–39 31
System (Applied Biosystems, Foster City, USA) using a Fast StartUniversal SYBR Green Master (Rox) (Roche Applied Biosciences,Rotkreuz, Switzerland). The primer sequences were: MMP-2 for-ward, 5=-GCACCACCGAGGATTATGAC-3=, MMP-2 reverse 5=-CACCCACAGTGGACATAGCA-3=, MMP-9 forward 5=-CCTCTG-CATGAAGACGACATAA-3=, MMP-9 reverse 5=-GGTCAGGTT-TAGAGCCACGA-3= (Microsynth, St. Gallen, Switzerland). Eachreaction contained 300 nM of primer. The cycling parameters were10 min at 95 °C, then 40 cycles of 15 s at 95 °C and 60 s at 60 °C.We calculated relative quantities of specifically amplified cDNAwith the comparative threshold cycle method. GAPDH acted asan endogenous reference. Relative expression of MMP-2 andMMP-9 mRNA in the different compartments of the cochleawere compared to brain tissue. No-template and no-reverse-transcription controls served as negative controls and excludedDNA contamination. All experiments were repeated three timesin triplicate.
Western blotting
A total of 10 animals were decapitated and 20 cochleae werecarefully microdissected in chilled PBS. For each cochlea, theOC was separated from the SG and the SV. For Westernblotting, in which OC incubation was needed, 10 OCs wereincubated in culture medium alone and 10 OCs were incubatedin the presence of 0.25 mM gentamicin for 24 h. Inner earexplants were then homogenized in CelLytic buffer containing
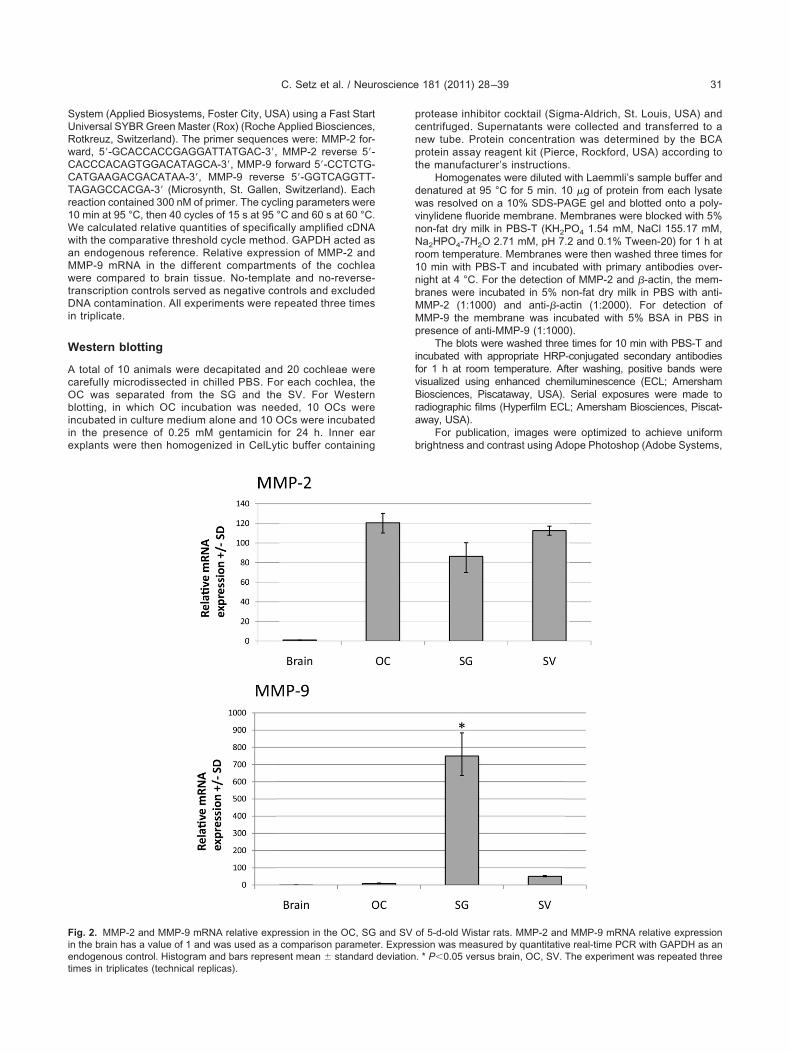
Fig. 2. MMP-2 and MMP-9 mRNA relative expression in the OC, SGn the brain has a value of 1 and was used as a comparison parametendogenous control. Histogram and bars represent mean � standard
times in triplicates (technical replicas).
protease inhibitor cocktail (Sigma-Aldrich, St. Louis, USA) andcentrifuged. Supernatants were collected and transferred to anew tube. Protein concentration was determined by the BCAprotein assay reagent kit (Pierce, Rockford, USA) according tothe manufacturer’s instructions.
Homogenates were diluted with Laemmli’s sample buffer anddenatured at 95 °C for 5 min. 10 �g of protein from each lysatewas resolved on a 10% SDS-PAGE gel and blotted onto a poly-vinylidene fluoride membrane. Membranes were blocked with 5%non-fat dry milk in PBS-T (KH2PO4 1.54 mM, NaCl 155.17 mM,Na2HPO4-7H2O 2.71 mM, pH 7.2 and 0.1% Tween-20) for 1 h atroom temperature. Membranes were then washed three times for10 min with PBS-T and incubated with primary antibodies over-night at 4 °C. For the detection of MMP-2 and �-actin, the mem-branes were incubated in 5% non-fat dry milk in PBS with anti-MMP-2 (1:1000) and anti-�-actin (1:2000). For detection of
MP-9 the membrane was incubated with 5% BSA in PBS inresence of anti-MMP-9 (1:1000).
The blots were washed three times for 10 min with PBS-T andncubated with appropriate HRP-conjugated secondary antibodiesor 1 h at room temperature. After washing, positive bands wereisualized using enhanced chemiluminescence (ECL; Amershamiosciences, Piscataway, USA). Serial exposures were made to
adiographic films (Hyperfilm ECL; Amersham Biosciences, Piscat-way, USA).
For publication, images were optimized to achieve uniformrightness and contrast using Adope Photoshop (Adobe Systems,
of 5-d-old Wistar rats. MMP-2 and MMP-9 mRNA relative expressionsion was measured by quantitative real-time PCR with GAPDH as an. * P�0.05 versus brain, OC, SV. The experiment was repeated three
and SVr. Expresdeviation

fi
C. Setz et al. / Neuroscience 181 (2011) 28–3932
San Jose, CA, USA). Each blot is representative of three inde-pendent experiments.
Immunohistochemistry
Animal and tissue preparation. The experiments were per-formed on cochleae of wild-type 12-month-old adult C57BL/6mice. Mice were euthanized with an overdose of sodium pento-barbital (100 mg/kg, Streuli AG, Uznach, Switzerland) and tran-scardially perfused with 50 ml of phosphate-buffered 4% parafor-maldehyde (pH 7.4 at 40 °C). The inner ear was carefully re-moved. Decalcification was carried out in a light-protected flask for10 days in a solution of 120 mM EDTA (Merck, NJ, USA) indistilled water (pH 6.8). After decalcification, cochleae were pre-pared for paraffin embedding. Briefly, cochleae were dehydratedin graded ethanol solutions (at 70, 80, 95, and 3�100% each for1 h, 3� xylol at 1 h and 2� paraplast at �60 °C for 1 h and, finally,paraplast at �60 °C for 10 h), and embedded in paraffin at 56 °C.For histological evaluation, sagittal paraffin sections of 10 �mthicknesses were cut on a Leitz microtome and mounted onSuperfrost plus slides (Menzel, Braunschweig, Germany). Sec-tions were deparaffinized, rehydrated, and were proceeded toantigen unmasking by boiling in 10 mM sodium citrate buffer pH6.0 and then maintained at the sub-boiling temperature for 10 min.The slides were then cooled and washed in PBS-T for 5 min.
82 kD
64 kD
B OC
MMP-2: proac�ve 7
49 kD
37 kDB OC
β-ac�n: 41 kD
M
M
115 kD
82 kDB
MMP-9: proac�ve 92 k
HL-60
B
β-ac�n: 41 kD49 kD
37 kDHL-60
M
M
Fig. 3. Western blot analysis of MMP-2 and MMP-9 expression in the Ocorresponding to the inactive proform (72 and 92 kDa respectively) anbe visualized. Marker (M) proteins are indicated with their size. �-actinsecond �-actin band can be visualized in the human promyelocytic leu
gure legend, the reader is referred to the Web version of this article.Tissue staining. Microtome sections were incubated for 1 hat room temperature in blocking solution containing PBS-T and3% goat serum followed by the first antibody overnight at 4 °C.The following antibodies were used: anti-MMP-2 mouse monoclo-nal (MAB3308, Chemicon, Temecula, USA), anti-MMP-9 rabbitpolyclonal (AB19016, Chemicon, Temecula, USA). After threewashes in PBS-T, the sections were incubated for 1 h at roomtemperature with the appropriate secondary antibodies (1:250,Alexa conjugated, Molecular Probes, Eugene, USA) diluted inPBS-T with 1% normal goat serum for 2 h at room temperature.After washing in PBS, the sections were counterstained with DAPIand mounted on glass slides with Mowiol.
Slices were visualized on an Olympus AX-70 microscopeequipped with a spot digital camera. Recorded images were ad-justed for brightness and contrast with Image-Pro plus and Pho-toshop image processing software. Inner ears from three animalswere analyzed.
Hair cell counting
OCs from 5-day-old Wistar rats were fixed in 4% paraformalde-hyde containing PBS and permeabilized with 5% Triton X-100 inPBS containing 10% FCS. OCs were incubated with a 1:100dilution of Texas Red X-phalloidin (Molecular Probes, Eugene,USA) for 45 min at room temperature. OCs were cultivated underfive different conditions for 24 h: culture medium alone; culture
SV
ac�ve 64 kD
SV
SG SV
�ve 86 kD
SG SV
MMP-2 proac�ve
MMP-2 ac�ve
MMP-9 proac�ve
MMP-9 ac�ve
V and brain (B) of 5-d-old Wistar rats. For MMP-2 and MMP-9, a bandcorresponding to the activated form (64 and 86 kDa respectively) cann at 41 kDa to illustrate equal protein loading in the different lanes. A-60 cell line lysates. For interpretation of the references to color in this
SG
2 kD /
SG
OC
D / ac
OC
C, SG, Sd a band
is showkemia HL
c(t
(SrboDcm
tfifo
Mct
RscwMottnc
BM
TtifMrpsSafi
Aoes(
Gd
TtrmHtlrw0Ct
C. Setz et al. / Neuroscience 181 (2011) 28–39 33
medium containing 0.25 mM gentamicin; culture medium contain-ing 50 �M GM6001 MMP Inhibitor; culture medium containingboth 0.25 mM gentamicin and 50 �M GM6001 MMP Inhibitorsolution. The specificity of GM6001 was tested using N-t-butoxy-carbonyl-L-leucyl-L-tryptophan methylamide, a closely relatedompound without MMP inhibitor activity, as a negative controlWebber et al., 2002). Six OCs were taken for each condition andhe experiment was repeated three times.
We visualized the OCs using a fluorescence microscopeOlympus IX71) and photographed them with an AxioCam (Zeiss,an Diego, USA). Surviving HCs were counted in a section cor-
esponding to 20 inner HCs at three different sites located on theasal and middle turns of each OC. The HC-counting resultsbtained from each group were analyzed by one-way ANOVA.ifferences between groups were considered statistically signifi-ant when the P-value was less than 0.05. Data are presented asean � standard deviation (SD).
RESULTS
MMP-2 and MMP-9 genes are expressed in thecochlea
MMP-2 and MMP-9 mRNA were detected in the OC, theSG, and in the SV of 5-day-old Wistar rat pups usingspecific primer sets (Table 1). One single band of thecorrect size for every tissue was observed (Fig. 1). Theamplification of �-actin confirmed a successful cDNA syn-hesis. The specificity of the designed primers was con-rmed using cDNA from rat brain tissue. The correct PCRragments of MMP-2 and MMP-9 were confirmed by dide-xy sequencing.
MP-2 mRNA is homogenously expressed in theochlea, while MMP-9 gene expression is higher inhe SG than in both the OC and the SV
eal-time PCR was used to determine the gene expres-ion of MMP-2 and MMP-9 in the three different cochlearomponents. Brain cDNA from 5-day-old Wistar rat pupsas used as a comparative mRNA expression parameter.MP-2 mRNA was found to be expressed equally through-ut the cochlea, and was over 80-fold higher than in brainissue. MMP-9 mRNA expression was particularly high inhe SG (Fig. 2). For every PCR amplification, we took 500g of cDNA from brain and 500 ng cDNA from each co-hlear component.
oth proactive and active forms of MMP-2 andMP-9 are expressed in the cochlea
o investigate expression of MMP-2 and MMP-9 protein inhe cochlea, we performed an immunoblot analysis withsolated total protein extracts from the OC, SG, and SVrom 5-day-old Wistar rat pups. Specific detection ofMP-2 and MMP-9 was observed in protein extracts from
at brain. No protein detection was observed by omittingrimary antibody, suggesting its specificity (data nothown). MMP-2 and MMP-9 were detected in the OC, theG, and the SV. In both cases, we used specific primaryntibodies against their inactive proform and their activeorm. For MMP-2, we detected a 72-kDa band correspond-
ng to the inactive proform in the three parts of the cochlea. tsecond band at 64 kDa corresponding to the active formf the protein was detected only in brain and SG homog-nates. For MMP-9, two bands were identified that repre-ented the inactive proform (96 kDa) and the active form82 kDa) in the cochlea (Fig. 3).
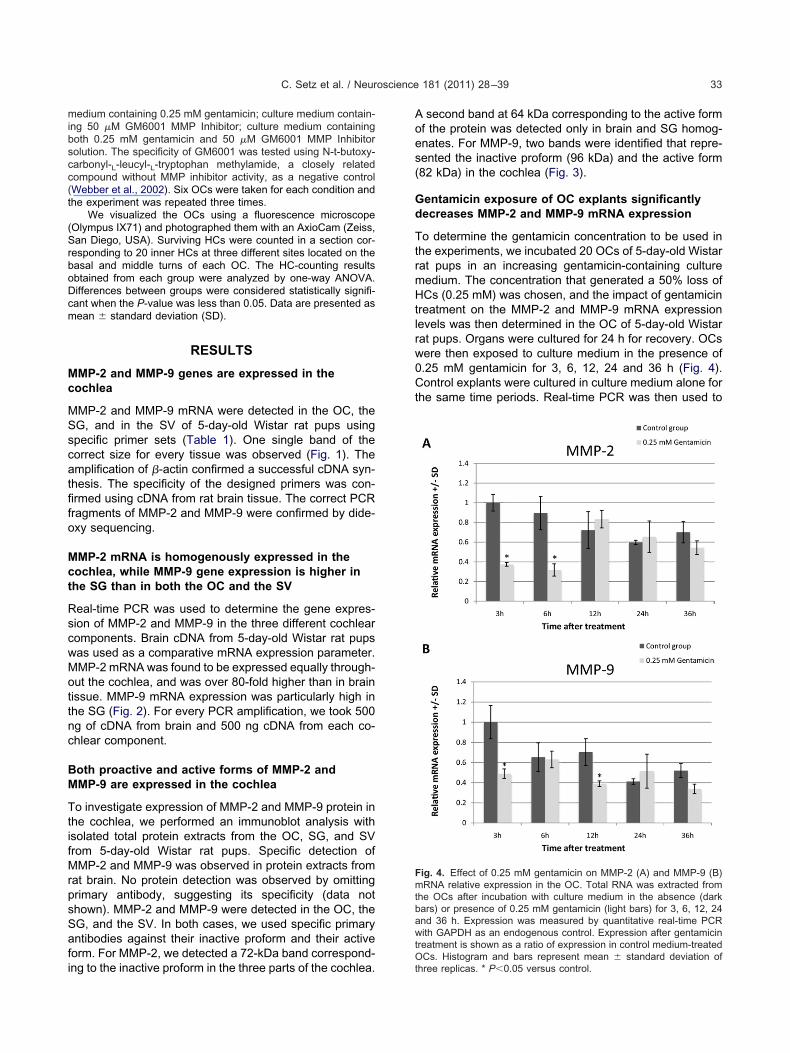
entamicin exposure of OC explants significantlyecreases MMP-2 and MMP-9 mRNA expression
o determine the gentamicin concentration to be used inhe experiments, we incubated 20 OCs of 5-day-old Wistarat pups in an increasing gentamicin-containing cultureedium. The concentration that generated a 50% loss ofCs (0.25 mM) was chosen, and the impact of gentamicin
reatment on the MMP-2 and MMP-9 mRNA expressionevels was then determined in the OC of 5-day-old Wistarat pups. Organs were cultured for 24 h for recovery. OCsere then exposed to culture medium in the presence of.25 mM gentamicin for 3, 6, 12, 24 and 36 h (Fig. 4).ontrol explants were cultured in culture medium alone for
he same time periods. Real-time PCR was then used to
Fig. 4. Effect of 0.25 mM gentamicin on MMP-2 (A) and MMP-9 (B)mRNA relative expression in the OC. Total RNA was extracted fromthe OCs after incubation with culture medium in the absence (darkbars) or presence of 0.25 mM gentamicin (light bars) for 3, 6, 12, 24and 36 h. Expression was measured by quantitative real-time PCRwith GAPDH as an endogenous control. Expression after gentamicintreatment is shown as a ratio of expression in control medium-treatedOCs. Histogram and bars represent mean � standard deviation of
hree replicas. * P�0.05 versus control.
cnnMttawM
t 41 kDat n of this
C. Setz et al. / Neuroscience 181 (2011) 28–3934
determine quantitative mRNA levels of MMP-2 and MMP-9in all the samples (GAPDH served as control). No signifi-cant difference in the MMP-2 and MMP-9 mRNA expres-sion was found in OCs treated with gentamicin comparedwith those organs incubated with culture medium aloneafter 24 and 36 h gentamicin treatment. A decrease inMMP-2 mRNA expression was observed after 3 and 6 hgentamicin treatment (Fig. 4A) and a decrease in MMP-9mRNA expression was observed after 3 and 12 h genta-micin treatment (Fig. 4B).
MMP-2 and MMP-9 protein levels are increased in theOC after gentamicin exposure
The effect of gentamicin on the protein levels of MMP-2and MMP-9 in the OC of 5-day-old Wistar rat pups wasexamined. After 24 h treatment of 10 OCs with 0.25 mMgentamicin, an increase in the levels of MMP-2 and MMP-9was observed in gentamicin treated OCs compared to thecontrol. Both the proactive and the active forms of MMP-2and MMP-9 were found to increase (Fig. 5).
MMP-2 and MMP-9 are localized in OCs and inthe SG
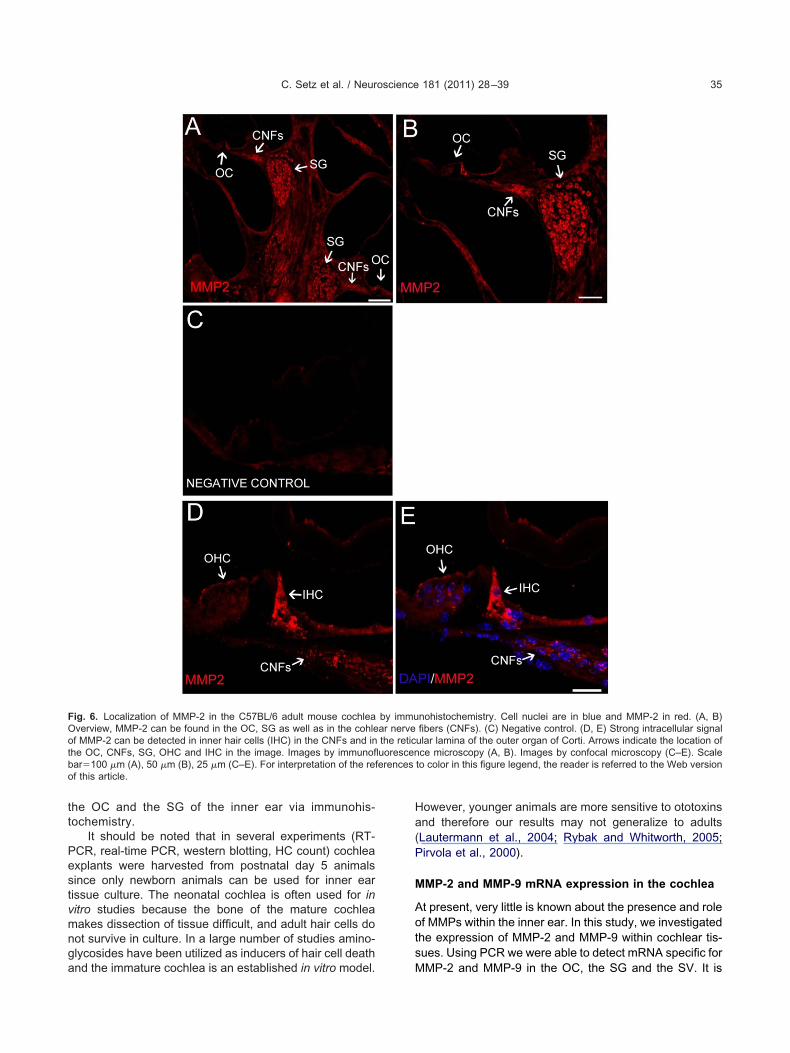
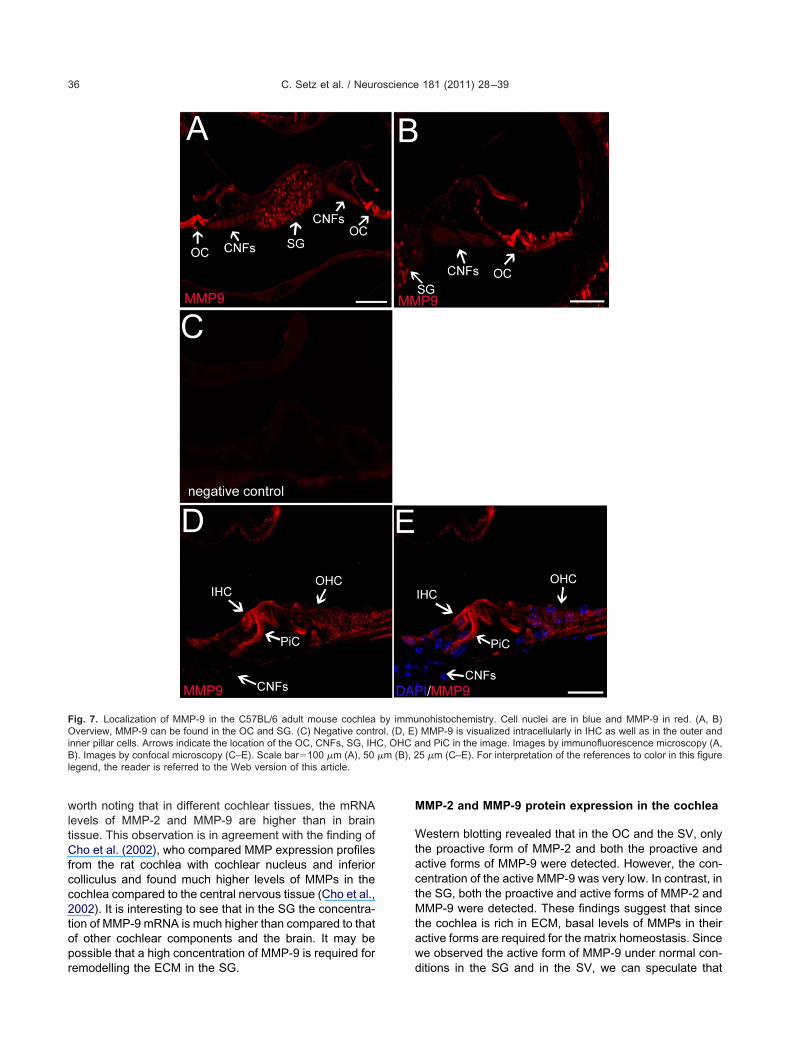
We used immunohistochemical staining to show thatMMP-2 (Fig. 6) and MMP-9 (Fig. 7) were present in thecochlea of C57BL/6 mice. MMP-2 was found to be stronglyexpressed in the inner HCs, spiral ganglion, cochlearnerve fibers and the reticular lamina of the outer organ ofCorti (Fig. 6). MMP-9 immunoreactivity was observed in-tracellularly in the inner HCs, supporting cells, as well asthe SG. In contrast to MMP-2, no MMP-9 immunoreactivity
Fig. 5. Western blot analysis of MMP-2 and MMP-9 expression in thepresence of 0.25 mM gentamicin. �-actin is shown in the lower panel ao color in this figure legend, the reader is referred to the Web versio
was observed in the cochlear nerve fibers (Fig. 7).
Inhibition of MMP-activity results in HC death
OCs treated for 24 h with MMP inhibitor resulted in HCloss. N-[(2R)-2-(hydroxamidocarbonylmethyl)-4-methyl-pentanoyl]-L-tryptophan methylamide, is a potent inhibitorof collagenases which blocks specifically the activity ofMMP-1, 2, 3, 8 and 9. In contrast, organs treated for 24 h withN-t-butoxycarbonyl-L-leucyl-L-tryptophan methylamide, alosely related compound to the MMP inhibitor used, showedo HC loss (Figs. 8 and 9). No difference was found in theumber of HCs lost when OCs were incubated only withMP inhibitor compared with organs treated with gen-
amicin alone (Figs. 8 and 9). In addition, organs main-ained in culture medium containing an MMP inhibitornd gentamicin experienced no additional HC loss,hen compared to those explants treated either withMP inhibitor or gentamicin alone (Figs. 8 and 9).
DISCUSSION
In this study, we demonstrate that MMP-2 and MMP-9are expressed within the cochlea. Real-time PCR re-vealed different expression levels of these two MMPs inthe OC, the SG and the SV. Western blotting demon-strated that both active and inactive forms of MMP-2 andMMP-9 are present within the cochlea. After exposure togentamicin, the concentration of both forms increased inthe OC. We demonstrated that inhibition of MMP activityresults in HC loss within the OC, and exposure of OCexplants to both MMP inhibitor and gentamicin does notresult in increased HC loss when compared to exposureof either MMP inhibitor or gentamicin alone. Finally, both
r 24 h exposure to culture medium alone or culture medium with theto illustrate equal protein loading. For interpretation of the references
article.
OC afte
MMP-2 and MMP-9 were found to be expressed within
Ha(P
M
Aots
ferenceso
C. Setz et al. / Neuroscience 181 (2011) 28–39 35
the OC and the SG of the inner ear via immunohis-tochemistry.
It should be noted that in several experiments (RT-PCR, real-time PCR, western blotting, HC count) cochleaexplants were harvested from postnatal day 5 animalssince only newborn animals can be used for inner eartissue culture. The neonatal cochlea is often used for invitro studies because the bone of the mature cochleamakes dissection of tissue difficult, and adult hair cells donot survive in culture. In a large number of studies amino-glycosides have been utilized as inducers of hair cell death
Fig. 6. Localization of MMP-2 in the C57BL/6 adult mouse cochleaOverview, MMP-2 can be found in the OC, SG as well as in the cohleof MMP-2 can be detected in inner hair cells (IHC) in the CNFs and inthe OC, CNFs, SG, OHC and IHC in the image. Images by immunoflbar�100 �m (A), 50 �m (B), 25 �m (C–E). For interpretation of the ref this article.
and the immature cochlea is an established in vitro model. M
owever, younger animals are more sensitive to ototoxinsnd therefore our results may not generalize to adultsLautermann et al., 2004; Rybak and Whitworth, 2005;irvola et al., 2000).
MP-2 and MMP-9 mRNA expression in the cochlea
t present, very little is known about the presence and rolef MMPs within the inner ear. In this study, we investigatedhe expression of MMP-2 and MMP-9 within cochlear tis-ues. Using PCR we were able to detect mRNA specific for
nohistochemistry. Cell nuclei are in blue and MMP-2 in red. (A, B)fibers (CNFs). (C) Negative control. (D, E) Strong intracellular signal
ular lamina of the outer organ of Corti. Arrows indicate the location ofce microscopy (A, B). Images by confocal microscopy (C–E). Scale
to color in this figure legend, the reader is referred to the Web version
by immuar nervethe reticuorescen
MP-2 and MMP-9 in the OC, the SG and the SV. It is
l
C. Setz et al. / Neuroscience 181 (2011) 28–3936
worth noting that in different cochlear tissues, the mRNAlevels of MMP-2 and MMP-9 are higher than in braintissue. This observation is in agreement with the finding ofCho et al. (2002), who compared MMP expression profilesfrom the rat cochlea with cochlear nucleus and inferiorcolliculus and found much higher levels of MMPs in thecochlea compared to the central nervous tissue (Cho et al.,2002). It is interesting to see that in the SG the concentra-tion of MMP-9 mRNA is much higher than compared to thatof other cochlear components and the brain. It may bepossible that a high concentration of MMP-9 is required for
Fig. 7. Localization of MMP-9 in the C57BL/6 adult mouse cochleaOverview, MMP-9 can be found in the OC and SG. (C) Negative contrinner pillar cells. Arrows indicate the location of the OC, CNFs, SG, IHB). Images by confocal microscopy (C–E). Scale bar�100 �m (A), 50egend, the reader is referred to the Web version of this article.
remodelling the ECM in the SG.
MMP-2 and MMP-9 protein expression in the cochlea
Western blotting revealed that in the OC and the SV, onlythe proactive form of MMP-2 and both the proactive andactive forms of MMP-9 were detected. However, the con-centration of the active MMP-9 was very low. In contrast, inthe SG, both the proactive and active forms of MMP-2 andMMP-9 were detected. These findings suggest that sincethe cochlea is rich in ECM, basal levels of MMPs in theiractive forms are required for the matrix homeostasis. Sincewe observed the active form of MMP-9 under normal con-
nohistochemistry. Cell nuclei are in blue and MMP-9 in red. (A, B)MMP-9 is visualized intracellularly in IHC as well as in the outer andnd PiC in the image. Images by immunofluorescence microscopy (A,5 �m (C–E). For interpretation of the references to color in this figure
by immuol. (D, E)C, OHC a�m (B), 2
ditions in the SG and in the SV, we can speculate that

bwiWsosttab
C. Setz et al. / Neuroscience 181 (2011) 28–39 37
basal levels of active MMP-9 are required for matrix ho-meostasis in these tissues. In contrast, the active form ofMMP-2 can only be observed in the SG, while in the OCand the SV, it is only present in its proactive form. It ispossible that in the OC, most of the MMPs are in aninactive form since very little ECM is present. Therefore theneed for matrix remodelling under normal (non-pathologi-cal) conditions is low. We speculate that under pathologi-
B
A. Control
B. MMP inhibitor nega�ve control
C. Gentamicin
D. MMP inhibitor
E. Gentamicin + MMP inhibitor
Fig. 8. Effect of MMP inhibitor on gentamicin-induced hair cell (HC)damage. Photograph of phalloidin-labeled OCs. The three outer HCrows and the single inner HC row can be well recognized in controlsand in explants treated with the closely related compound to theMMP inhibitor without the MMP activity (A, B). OCs exposed to 50�M MMP-Inhibitor demonstrate to suffer from HC damage (D) aswell as 0.25 mM gentamicin-treated OC (C). Treatment with 50 �MMMP inhibitor and 0.25 mM gentamicin resulted in no additional HCloss (E). Bar, 20 �m. For interpretation of the references to color inthis figure legend, the reader is referred to the Web version of thisarticle.
cal conditions fast activation of proactive MMPs is required
to maintain matrix homeostasis to protect the OC fromdamage. It is worth noting that the protein expressionlevels of MMP-2 and MMP-9 cannot be compared with themRNA levels expressed in the brain and each compart-ment of inner ear including OC, SG, and SV. Post-tran-scriptional events such as different protein stability of theMMPs examined in different inner ear compartments maybe responsible for the observed phenomenon.
MMP inhibition results in HC death
We observed HC loss within the OC after MMP inhibition.It seems that a basal level of MMP activity is required in thenormal OC for proper function, and when MMP activity wasblocked, the resulting disturbance of the ECM homeostasisresulted in HC loss.
Inhibition of MMP activity in OC explants resulted in thesame degree of HC loss whether the explants were ex-posed to the inhibitor alone or the MMP-inhibitor was com-bined with gentamicin. This suggests that the apoptoticmechanisms triggered by the lack of MMP activity might bethe same as those triggered by aminoglycoside exposition.Another possibility is that gentamicin-induced disturbancesin MMPs proactive and active levels is compensated byMMP inhibition. Therefore, we do not observe additionalHC damage when the two compounds are administeredtogether.
The results of MMP inhibition in vitro suggest that aasal level of MMP activity is required for HC survival, buthat might be the mechanisms of HC death after MMP
nhibition? Currently we can only speculate. In a report,oo et al. (2008) showed, that inhibition of MMP activity
ignificantly suppresses the expression of inducible nitricxidase synthase through mechanisms involving suppres-ion of nuclear factor “kappa-light-chain-enhancer” of ac-ivated B-cells (NF-�B). Our group also recently showedhat inhibition of NF-�B activity result in HC loss (Nagy etl., 2005). Therefore, it might be possible that MMP inhi-ition suppresses NF-�B activity, which then results in HC
loss. Another possibility is that MMPs contribute to thestability of the reticular lamina, which protects the cells ofthe organ from potassium toxicity. Inhibition could exposethe hair cell somata to potassium.
MMP-2 and MMP-9 after aminoglycoside-inducedHC loss
It is interesting to note that OCs exposed to gentamicinresult in an increased expression level of the proactiveform of MMP-2 and MMP-9, compared to explants incu-bated in culture medium alone (Fig. 5). In contrast, adifferent observation has been made in heterozygousknockout mice for cysthatione beta-synthase (Kundu et al.,2009). These mice suffered from hyperhomocysteinemia,and the authors of this study observed higher levels of boththe proactive and active forms of MMP-2 and MMP-9 incochlear tissue compared to wild-type mice. However, theyused whole cochlear organ lysates for the immunoblots,while we separately analysed the three different cochlear
components (OC, SG and SV).
MpcsMtOecptreel(
wscat
C. Setz et al. / Neuroscience 181 (2011) 28–3938
Gene expression profiles over time for MMP-2 andMP-9 in OC explants after gentamicin exposure dis-layed no up regulation in mRNA levels. In fact, a de-rease in MMP-2 and MMP-9 mRNA levels could behown after 3 and 6 h for MMP-2 and after 3 and 12 h forMP-9. All other time points showed no difference be-
ween OC explants held in culture medium compared withC explants incubated in culture medium with the pres-nce of 0.25 mM gentamicin. From these results, we con-lude that the increased levels of MMP-2 and MMP-9rotein after gentamicin exposure are mainly due to post-ranscriptional regulation (such as protein stability) than toegulation at the transcriptional level, where we wouldxpect increased mRNA levels compared to the controlxplants. It is well-known that MMP activity and expression
evels can be regulated on the post-transcriptional levelYu et al., 1997).
We conclude that MMP-2 and MMP-9 are expressedithin different cochlear locations and that aminoglyco-ide-exposition alters MMP-2 and MMP-9 levels in theochlea together with an altered distribution of the inactivend the proactive form of these proteins. Finally, we showhat MMP inhibition results in loss of auditory HCs.
Acknowledgments—Supported by grants from the Novartis Stif-tung für medizinisch-biologische Forschung, Basel, Switzerlandand the Schwerhörigenverein Nordwestschweiz.
REFERENCES
Agapova OA, Ricard CS, Salvador-Silva M, Hernandez MR (2001)Expression of matrix metalloproteinases and tissue inhibitors ofmetalloproteinases in human optic nerve head astrocytes. Glia33:205–216.
Candelario-Jalil E, Yang Y, Rosenberg GA (2009) Diverse roles ofmatrix metalloproteinases and tissue inhibitors of metalloprotei-nases in neuroinflammation and cerebral ischemia. Neuroscience158:983–994.
Cauwe B, Van den Steen PE, Opdenakker G (2007) The biochemical,biological, and pathological kaleidoscope of cell surface substratesprocessed by matrix metalloproteinases. Crit Rev Biochem MolBiol 42:113–185.
Chang DI, Hosomi N, Lucero J, Heo JH, Abumiya T, Mazar AP, del
Fig. 9. Quantitative analysis of surviving HCs. Histograms show no diffwith OCs treated with MMP-Inhibitor or both (P�0.05). Histogram andgroups to control/MMP inhibitor negative control.
Zoppo GJ (2003) Activation systems for latent matrix metallopro-
teinase-2 are upregulated immediately after focal cerebralischemia. J Cereb Blood Flow Metab 23:1408–1419.
Chintala SK, Zhang X, Austin JS, Fini ME (2002) Deficiency in matrixmetalloproteinase gelatinase B (MMP-9) protects against retinalganglion cell death after optic nerve ligation. J Biol Chem 277:47461–47468.
Chintala SK (2006) The emerging role of proteases in retinal ganglioncell death. Exp Eye Res 82:5–12.
Cho Y, Gong TW, Stöver T, Lomax MI, Altschuler RA (2002) Geneexpression profiles of the rat cochlea, cochlear nucleus, and infe-rior colliculus. J Assoc Res Otolaryngol 3:54–67.
Clark AW, Krekoski CA, Bou SS, Chapman KR, Edwards DR (1997)Increased gelatinase A (MMP-2) and gelatinase B (MMP-9) activ-ities in human brain after focal ischemia. Neurosci Lett 238:53–56.
Crawford HC, Matrisian LM (1996) Mechanisms controlling the tran-scription of matrix metalloproteinase genes in normal and neoplas-tic cells. Enzyme Protein 49:20–37.
Gasche Y, Soccal PM, Kanemitsu M, Copin JC (2006) Matrix metal-loproteinases and diseases of the central nervous system with aspecial emphasis on ischemic brain. Front Biosci 11:1289–1301.
Gu Z, Kaul M, Yan B, Kridel SJ, Cui J, Strongin A, Smith JW, Lidding-ton RC, Lipton SA (2002) S-nitrosylation of matrix metalloprotei-nases: signaling pathway to neuronal cell death. Science 297:1186–1190.
Kleiner DE Jr, Stetler-Stevenson WG (1993) Structural biochemistryand activation of matrix metalloproteases. Curr Opin Cell Biol 5:891–897.
Kundu S, Tyagi N, Sen U, Tyagi SC (2009) Matrix imbalance byinducing expression of metalloproteinase and oxidative stress incochlea of hyperhomocysteinemic mice. Mol Cell Biochem 332:215–224.
Lautermann J, Dehne N, Schacht J, Jahnke K (2004) Aminoglycoside-and cisplatin-ototoxicity: from basic science to clinics. Laryngorhi-nootologie 83:317–323.
Lee R, Kermani P, Teng KK, Hempstead BL (2001) Regulation of cellsurvival by secreted proneurotrophins. Science 294:1945–1948.
Matrisian LM (1992) The matrix-degrading metalloproteinases. Bioes-says 14:455–463.
Meli DN, Christen S, Leib SL (2003) Matrix metalloproteinase-9 inpneumococcal meningitis: activation via an oxidative pathway. J In-fect Dis 187:1411–1415.
Murphy G, Docherty AJ (1992) The matrix metalloproteinases andtheir inhibitors. Am J Respir Cell Mol Biol 7:120–125.
Nagy I, Monge A, Albinger-Hegyi A, Schmid S, Bodmer D (2005)NF-kappaB is required for survival of immature auditory hair cellsin vitro. J Assoc Res Otolaryngol 6:260–268.
Pei P, Horan MP, Hille R, Hemann CF, Schwendeman SP, Mallery SR
the number of surviving HCs in OCs exposed to gentamicin comparedpresent mean � standard deviation. * P�0.05 between the indicated
erence inbars re
(2006) Reduced nonprotein thiols inhibit activation and function of

C. Setz et al. / Neuroscience 181 (2011) 28–39 39
MMP-9: implications for chemoprevention. Free Radic Biol Med41:1315–1324.
Pirvola U, Xing-Qun L, Virkkala J, Saarma M, Murakata C, CamorattoAM, Camoratto AM, Walton KM, Ylikoski J (2000) Rescue ofhearing, auditory hair cells, and neurons by CEP-1347/KT7515, aninhibitor of c-Jun N-terminal kinase activation. J Neurosci 20:43–50.
Reuter A, Nestl A, Zwacka RM, Tuckermann J, Waldherr R, Wag-ner EM, Höyhtyä M, Meyer zum Gottesberge AM, Angel P,Weiher H (1998) Expression of the recessive glomerulosclerosisgene Mpv17 regulates MMP-2 expression in fibroblasts, thekidney, and the inner ear of mice. Mol Biol Cell 9:1675–1682.
Rosenberg GA (2009) Matrix metalloproteinases and their multipleroles in neurodegenerative diseases. Lancet Neurol 8:205–216.
Rosenberg GA (2002) Matrix metalloproteinases in neuroinflamma-tion. Glia 39:279–291.
Rybak LP, Whitworth CA (2005) Ototoxicity: therapeutic opportunities.Drug Discov Today 10:1313–1321.
Schönbeck U, Mach F, Libby P (1998) Generation of biologically activeIL-1 beta by matrix metalloproteinases: a novel caspase-1-inde-pendent pathway of IL-1 beta processing. J Immunol 161:3340–3346.
Sternlicht MD, Werb Z (2001) How matrix metalloproteinases regulatecell behavior. Annu Rev Cell Dev Biol 17:463–516.
Stetler-Stevenson WG, Liotta LA, Kleiner DE Jr (1993) Extracellularmatrix 6: role of matrix metalloproteinases in tumor invasion andmetastasis. FASEB J 7:1434–1441.
Webber CA, Hocking JC, Yong VW, Stange CL, McFarlane S (2002)Metalloproteases and guidance of retinal ganglion axons in thedeveloping visual system. J Neurosci 22:8091–8100.
Woo MS, Park JS, Choi IY, Kim WK, Kim HS (2008) Inhibition ofMMP-3 or -9 suppresses lipopolysaccharide-induced expression ofproinflammatory cytokines and iNOS in microglia. J Neurochem106:770–780.
Yang Y, Estrada EY, Thompson JF, Liu W, Rosemberg GA (2007)Matrix metalloproteinase-mediated disruption of tight junction pro-teins in cerebral vessels is reversed by synthetic matrix metallo-proteinase inhibitor in focal ischemia in rat. J Cereb Blood FlowMetab 27:697–709.
Yong VW, Power C, Forsyth P, Edwards DR (2001) Metalloprotei-nases in biology and pathology of the nervous system. Nat RevNeurosci 2:502–511.
Yu M, Sato H, Seiki M, Spiegel S, Thompson EW (1997) Calcium influxinhibits MT1-MMP processing and blocks MMP-2 activation. FEBSLett 412:568–572.
Zhang X, Chintala SK (2004) Influence of interleukin-1 beta inductionand mitogen-activated protein kinase phosphorylation on optic
nerve ligation-induced matrix metalloproteinase-9 activation in theretina. Exp Eye Res 78:849–860.(Accepted 17 February 2011)
Related Documents











