Mass Spectrometry Characterization of the Glycation Sites of Bovine Insulin by Tandem Mass Spectrometry Sofia Guedes, Rui Vitorino, M. Rosário M. Domingues, Francisco Amado, and Pedro Domingues Department of Chemistry, University of Aveiro, Aveiro, Portugal Bovine insulin was glycated under hyperglycemic reducing conditions and in nonreducing conditions. Purification through HPLC allowed isolating glycated forms of insulin and a novel triglycated form (6224.5 Da) was purified. Endoproteinase Glu-C digestion combined with mass spectrometry (MALDI-TOF/TOF) allowed determining the exact location of the glyca- tion sites in each of the isolated glycated insulins. For the first time, a triglycated form of insulin was isolated and characterized accordingly to its glycation sites. These glucose binding sites were identified as the N-terminals of both chains (Gly1 and Phe1) and residue Lys29 of B-chain. Moreover, in diglycated insulin we found the coexistence of one specie glycated at the N-terminals of both chains (Gly1 and Phe1) and another specie containing the two glucitol adducts in B-chain (Phe1 and Lys29). Also, in monoglycated insulin generated in reducing and nonreducing conditions, one specie glycated at Phe1 and another specie glycated at Lys29, both B-chain residues coexist. (J Am Soc Mass Spectrom 2009, 20, 1319 –1326) © 2009 American Society for Mass Spectrometry O ver the last years, particular attention has been given to the role of nonenzymatic glycation in diabetic complications development, including neuropathy, nephropathy, retinopathy, and cardiovascu- lar disease [1]. Under conditions of hyperglycemia, glyca- tion compromises several functional proteins, such as hemoglobin [2, 3], glucose-6-phosphate dehydrogenase [4, 5], aldehyde reductase [6], glutathione reductase [7, 8], glutathione peroxidase [8, 9], and Cu-Zn superoxide dismutase [10, 11]. In this context, insulin glycation is of particular interest. The structurally modified peptide exhibits decreased ability to regulate plasma glucose homeostasis and is associated with reduced hepatic clearance, decreased adipose tissue lipogenesis, and a decreased glucose uptake and oxidation in isolated diaphragm and abdominal muscle in vitro [12–15]. Even though the short half-life of insulin (5–10 min), experiments performed to date indicate that a substan- tial proportion of insulin (and proinsulin) is glycated in the pancreatic cells during the stages of insulin synthe- sis and storage [16]. Glycated insulin has been mea- sured in the pancreas of various animal models of type 2 diabetes [17], and in both isolated islets and clonal -cells exposed to elevated glucose concentrations in tissue culture [18, 19]. The significant compromised biological activity of glycated insulin raises the possi- bility of its contribution to insulin resistance and glu- cose intolerance of type 2 diabetes. Earlier studies indicated that during glycation pro- cess, each insulin molecule can bind to three to eight molecules of glucose [14, 20]. More recently, indepen- dent reports generalized the view that there are three possible free amino groups in the insulin molecule, human or bovine, available for in vitro glucose binding [15, 21–23]. Nowadays, through ESI-MS/MS and MALDI-MS approaches, it is established the N-terminal Phe1 residue of the B-chain is the only glycation site of human [23] and bovine [21] insulin. Moreover, O’Harte and coworkers determined, in a novel diglycated insu- lin (under hyperglycemic reducing conditions), the N- terminals Gly1 of A-chain and Phe1 of B-chain as the sites of glycation for human insulin [15], after Glu-C enzymatic digestion and identification of digested frag- ments by plasma desorption mass spectrometry (PDMS). Since human and bovine insulin only differ in three residues (A chain: residues 8 and 10; B chain: residue 30), it is expected that glycation of bovine insulin should occur in similar positions. The present study structurally characterizes three forms of glycated bovine insulin when glycated under reducing conditions. Simultaneously, the characteriza- tion of glycated insulin produced under pseudophysi- ological conditions was also done. Our approach in- cluded HPLC purification of mono-, di-, and triglycated insulin forms, followed by enzymatic digestion and mass spectrometry (MALDI-TOF/TOF) for the unam- biguous assignment of the glycation sites. Address reprint requests to Professor Pedro Domingues, Department of Chemistry, University of Aveiro, 3810-193 Aveiro, Portugal. E-mail: [email protected] Published online March 13, 2009 © 2009 American Society for Mass Spectrometry. Published by Elsevier Inc. Received December 2, 2008 1044-0305/09/$32.00 Revised February 2, 2009 doi:10.1016/j.jasms.2009.03.004 Accepted March 5, 2009

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Mass Spectrometry Characterization of theGlycation Sites of Bovine Insulin by TandemMass Spectrometry
Sofia Guedes, Rui Vitorino, M. Rosário M. Domingues,Francisco Amado, and Pedro DominguesDepartment of Chemistry, University of Aveiro, Aveiro, Portugal
Bovine insulin was glycated under hyperglycemic reducing conditions and in nonreducingconditions. Purification through HPLC allowed isolating glycated forms of insulin and a noveltriglycated form (6224.5 Da) was purified. Endoproteinase Glu-C digestion combined withmass spectrometry (MALDI-TOF/TOF) allowed determining the exact location of the glyca-tion sites in each of the isolated glycated insulins. For the first time, a triglycated form ofinsulin was isolated and characterized accordingly to its glycation sites. These glucose bindingsites were identified as the N-terminals of both chains (Gly1 and Phe1) and residue Lys29 ofB-chain. Moreover, in diglycated insulin we found the coexistence of one specie glycated at theN-terminals of both chains (Gly1 and Phe1) and another specie containing the two glucitoladducts in B-chain (Phe1 and Lys29). Also, in monoglycated insulin generated in reducing andnonreducing conditions, one specie glycated at Phe1 and another specie glycated at Lys29,both B-chain residues coexist. (J Am Soc Mass Spectrom 2009, 20, 1319–1326) © 2009American Society for Mass Spectrometry
Over the last years, particular attention has beengiven to the role of nonenzymatic glycation indiabetic complications development, including
neuropathy, nephropathy, retinopathy, and cardiovascu-lar disease [1]. Under conditions of hyperglycemia, glyca-tion compromises several functional proteins, such ashemoglobin [2, 3], glucose-6-phosphate dehydrogenase[4, 5], aldehyde reductase [6], glutathione reductase [7,8], glutathione peroxidase [8, 9], and Cu-Zn superoxidedismutase [10, 11]. In this context, insulin glycation is ofparticular interest. The structurally modified peptideexhibits decreased ability to regulate plasma glucosehomeostasis and is associated with reduced hepaticclearance, decreased adipose tissue lipogenesis, and adecreased glucose uptake and oxidation in isolateddiaphragm and abdominal muscle in vitro [12–15].Even though the short half-life of insulin (5–10 min),experiments performed to date indicate that a substan-tial proportion of insulin (and proinsulin) is glycated inthe pancreatic cells during the stages of insulin synthe-sis and storage [16]. Glycated insulin has been mea-sured in the pancreas of various animal models of type2 diabetes [17], and in both isolated islets and clonal�-cells exposed to elevated glucose concentrations intissue culture [18, 19]. The significant compromisedbiological activity of glycated insulin raises the possi-
Address reprint requests to Professor Pedro Domingues, Department of
Chemistry, University of Aveiro, 3810-193 Aveiro, Portugal. E-mail:[email protected]© 2009 American Society for Mass Spectrometry. Published by Elsevie1044-0305/09/$32.00doi:10.1016/j.jasms.2009.03.004
bility of its contribution to insulin resistance and glu-cose intolerance of type 2 diabetes.
Earlier studies indicated that during glycation pro-cess, each insulin molecule can bind to �three to eightmolecules of glucose [14, 20]. More recently, indepen-dent reports generalized the view that there are threepossible free amino groups in the insulin molecule,human or bovine, available for in vitro glucose binding[15, 21–23]. Nowadays, through ESI-MS/MS andMALDI-MS approaches, it is established the N-terminalPhe1 residue of the B-chain is the only glycation site ofhuman [23] and bovine [21] insulin. Moreover, O’Harteand coworkers determined, in a novel diglycated insu-lin (under hyperglycemic reducing conditions), the N-terminals Gly1 of A-chain and Phe1 of B-chain as thesites of glycation for human insulin [15], after Glu-Cenzymatic digestion and identification of digested frag-ments by plasma desorption mass spectrometry (PDMS).Since human and bovine insulin only differ in threeresidues (A chain: residues 8 and 10; B chain: residue30), it is expected that glycation of bovine insulinshould occur in similar positions.
The present study structurally characterizes threeforms of glycated bovine insulin when glycated underreducing conditions. Simultaneously, the characteriza-tion of glycated insulin produced under pseudophysi-ological conditions was also done. Our approach in-cluded HPLC purification of mono-, di-, and triglycatedinsulin forms, followed by enzymatic digestion andmass spectrometry (MALDI-TOF/TOF) for the unam-
biguous assignment of the glycation sites.Published online March 13, 2009r Inc. Received December 2, 2008
Revised February 2, 2009Accepted March 5, 2009

1320 GUEDES ET AL. J Am Soc Mass Spectrom 2009, 20, 1319–1326
Materials and Methods
Chemicals and Reagents
Bovine insulin (I5500) and TFA were acquired fromSigma (St. Louis, MO). The MALDI matrices �-cyano-4-hydroxy cinnamic acid (CHCA) and sinapinic acidwere supplied by Merck (Darmstadt, Germany). Theorganic solvents such as toluene and acetonitrile (ACN)were from Riedel-deHaën (Buchs, Switzerland) andLabscan (Dublin, Ireland), respectively.
All chemicals were of analytical grade and milli-Qwater (Millipore, Bedford, MA) was used throughoutall experiments.
Insulin In Vitro Glycation
In vitro glycation was carried out by incubating insulinwith D-glucose in a protocol adapted from O’Harte etal. where reducing [23] and nonreducing conditionswere used. In nonreducing conditions, insulin (200 �gin HCl 2 mM) was incubated with D-glucose 220 mM inphosphate buffer 10 mM (pH 7.4) at 37 °C for 30 d(toluene 5 mM was added as a bacteriostatic). In reduc-ing conditions and according to the authors, insulin(100 �g in HCl 2 mM) was incubated for 24 h at 37 °Cwith D-glucose 220 mM (prepared in phosphate buffer10 mM, pH 7.4) together with a 1000-fold molar excessof the reducing agent NaBH3CN (stock in ice coldNaOH 10 mM) in phosphate buffer 10 mM (pH 7.4). Inboth cases, after incubation, reaction was stopped byaddition of acetic acid 0.5 M, and the reaction mixturewas dialyzed overnight against water (cut-off 1000 Da).Glycated and native forms were separated on a C-8analytical column (Acclaim, 4.6 � 250 mm, 5 �m particlesize; Dionex, Sunnyvale, CA) at a flow rate of 0.8 mL/min.The program was as follows: linear gradient of 0% to 35%ACN over 10 min, followed by 35% to 56% ACN over 20min, and 56% to 70% ACN over 5 min. Peaks werehand-collected, pooled from several runs, and concen-trated on a Speed-vac. Re-injection of each glycatedform using the same separation program allowed toimprove the purification of each glycated insulin form.This purification was monitored through MALDI-MS.
Enzymatic Digestion (Endoproteinase Glu-C)
For in-solution digestion of glycated and native insulin,samples were initially reconstituted in HCl 2 mM.Quantification was performed using the Quant-iT Pro-tein Assay kit (Invitrogen, Paisley, UK) combined withthe Qubit fluorometer. An aliquot was taken and dilutedfor enzymatic digestion with phosphate buffer 10 mM pH7.8; pH is determinant since the specificity of endoprotein-ase Glu-C for glutamic acid depends on this parameter.Endoproteinase Glu-C (Calbiochem, Merck, Darmstadt,Germany) solution was added in a final ratio of substrate:enzyme of 20:1 (wt/wt). Duplicate samples were incu-
bated overnight at 25 °C. After digestion, samples weredried under vacuum in a Speed-vac and, unless taken foranalysis, stored at �80 °C.
Reduction of Disulphide Bridges of Insulin
The disulphide bridges of monoglycated, diglycated,triglycated, and native insulin were reduced using DTT10 mM in phosphate buffer pH 8 for 1 h at 55 °C. Analiquot of each sample (10 �L) was taken and acidifiedwith TFA 1% (1 �L). These aliquots were cleaned upusing ZipTip C18 (Millipore, Bedford, MA) before massspectrometry analysis.
Mass Spectrometry
Glu-C digests of glycated and native insulin were recon-stituted in TFA 0.1%. An aliquot of each sample wasmixed (1:1) with �-CHCA matrix solution (5 mg/mL inACN 50%/TFA 0.1%) and applied onto 384-well MALDIplates. Instrument was calibrated by adding insulin (12.5ng; 0.5 �L) to each calibration spot. Peptide mass spectrawere obtained with a MALDI-TOF/TOF mass spectrom-eter (4800 Proteomics Analyzer; Applied Biosystems, Fos-ter City, CA) in the positive ion reflector mode. Spectrawere obtained in the mass range between 800 and 7000Da. Manual interpretation of tandem mass spectra wasperformed through the Data Explorer software TM ver.4.4 (Applied Biosystems).
Results and Discussion
Glycation of insulin was monitored by MALDI-MSthrough the increase in the molecular weight of insulinas a result of glucose adducts formation with thehormone. Figure 1 shows the typical mass spectra ofnative (a) and glycated insulin (b and c). In Figure 1b,we can observe a typical spectrum of glycated insulinwhere glycation of the hormone was performed byincubation with 220 mM glucose for 30 d at 37 °C in theabsence of reducing conditions. The major peak at m/z5730.66 corresponds to native bovine insulin (theoreti-cal m/z 5730.6) whereas the second peak at m/z 5892.72with a mass shift of 162 Da corresponds to a monogly-cated form of insulin. A third minor peak at m/z 6054.71is also observable and should correspond to a digly-cated form with a mass shift of 324 Da. Figure 1c, whereinsulin glycation was performed by incubation with 220mM glucose for 30 h at 37 °C in reducing conditions(NaBH3CN), shows a different mass profile, especiallyin terms of relative abundance of each specie. There arethree additional peaks in the spectrum besides the peakcorresponding to native insulin. The major peak, at m/zof 5894.72, is the protonated monoglycated form with amass shift of 164 Da (glucitol adduct). The two otherpeaks at m/z 6058.89 and 6222.82 correspond to theprotonated diglycated and triglycated forms, respec-tively. With the mono-, di-, and triglycated forms,several additional peaks are observed. These peaks are
BH3-adducts (�m � 14 Da), resulting from the in vitro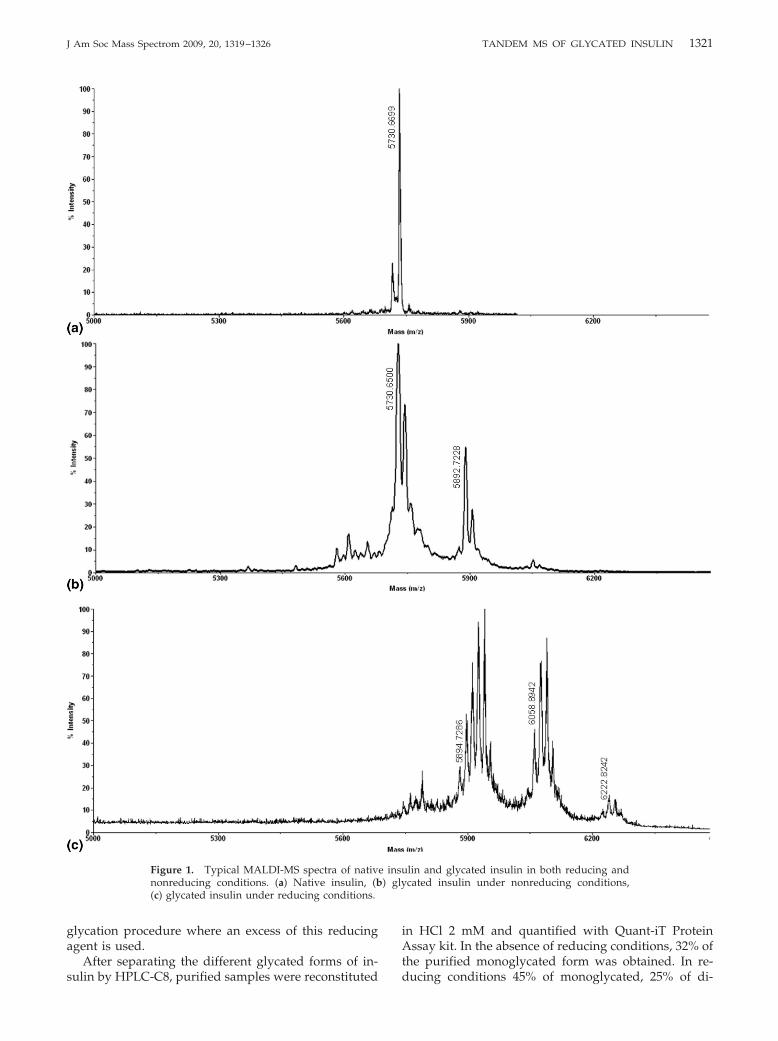
1321J Am Soc Mass Spectrom 2009, 20, 1319–1326 TANDEM MS OF GLYCATED INSULIN
glycation procedure where an excess of this reducingagent is used.
After separating the different glycated forms of in-
Figure 1. Typical MALDI-MS spectra of nativnonreducing conditions. (a) Native insulin, (b(c) glycated insulin under reducing conditions.
sulin by HPLC-C8, purified samples were reconstituted
in HCl 2 mM and quantified with Quant-iT ProteinAssay kit. In the absence of reducing conditions, 32% ofthe purified monoglycated form was obtained. In re-
lin and glycated insulin in both reducing andcated insulin under nonreducing conditions,
e insu) gly
ducing conditions 45% of monoglycated, 25% of di-
1322 GUEDES ET AL. J Am Soc Mass Spectrom 2009, 20, 1319–1326
glycated, and 6% of triglycated forms of insulin wereobtained.
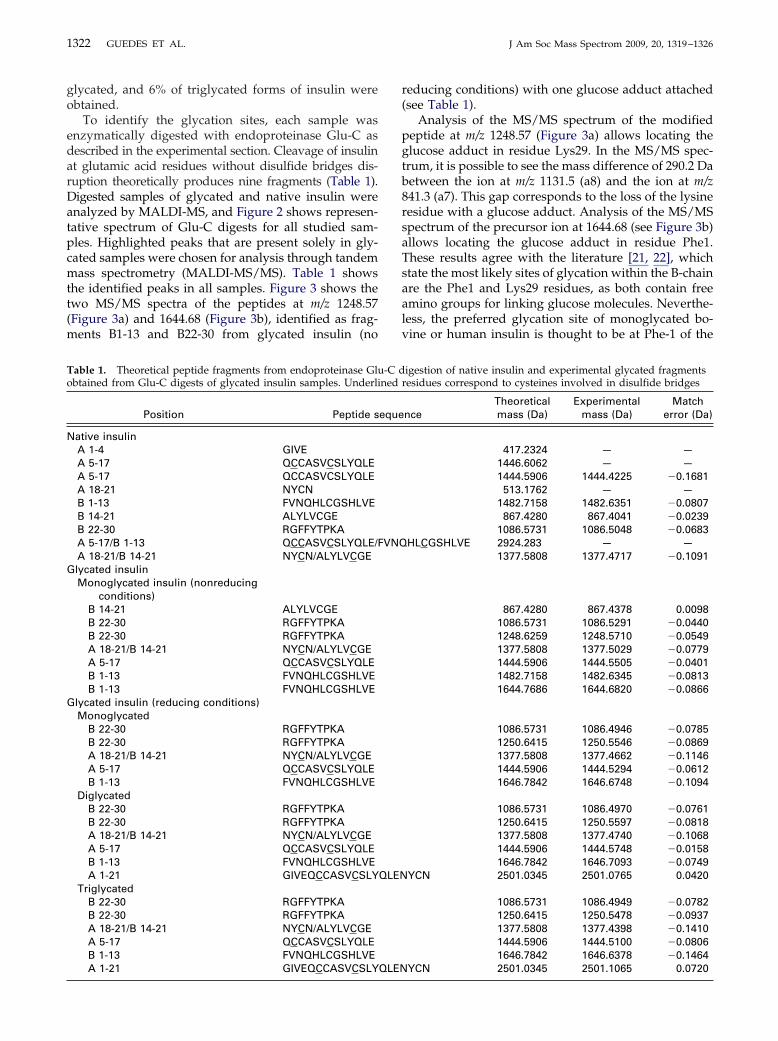
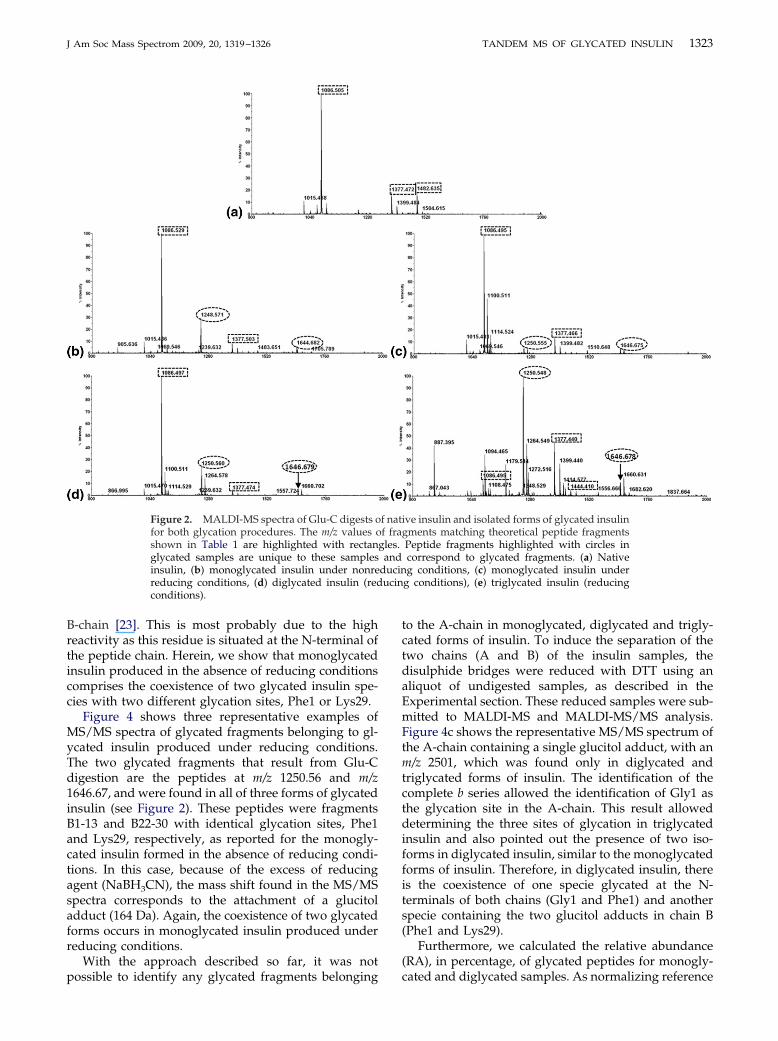
To identify the glycation sites, each sample wasenzymatically digested with endoproteinase Glu-C asdescribed in the experimental section. Cleavage of insulinat glutamic acid residues without disulfide bridges dis-ruption theoretically produces nine fragments (Table 1).Digested samples of glycated and native insulin wereanalyzed by MALDI-MS, and Figure 2 shows represen-tative spectrum of Glu-C digests for all studied sam-ples. Highlighted peaks that are present solely in gly-cated samples were chosen for analysis through tandemmass spectrometry (MALDI-MS/MS). Table 1 showsthe identified peaks in all samples. Figure 3 shows thetwo MS/MS spectra of the peptides at m/z 1248.57(Figure 3a) and 1644.68 (Figure 3b), identified as frag-ments B1-13 and B22-30 from glycated insulin (no
Table 1. Theoretical peptide fragments from endoproteinase Gluobtained from Glu-C digests of glycated insulin samples. Underli
Position Peptide s
Native insulinA 1-4 GIVEA 5-17 QCCASVCSLYQLEA 5-17 QCCASVCSLYQLEA 18-21 NYCNB 1-13 FVNQHLCGSHLVEB 14-21 ALYLVCGEB 22-30 RGFFYTPKAA 5-17/B 1-13 QCCASVCSLYQLE/FA 18-21/B 14-21 NYCN/ALYLVCGE
Glycated insulinMonoglycated insulin (nonreducing
conditions)B 14-21 ALYLVCGEB 22-30 RGFFYTPKAB 22-30 RGFFYTPKAA 18-21/B 14-21 NYCN/ALYLVCGEA 5-17 QCCASVCSLYQLEB 1-13 FVNQHLCGSHLVEB 1-13 FVNQHLCGSHLVE
Glycated insulin (reducing conditions)Monoglycated
B 22-30 RGFFYTPKAB 22-30 RGFFYTPKAA 18-21/B 14-21 NYCN/ALYLVCGEA 5-17 QCCASVCSLYQLEB 1-13 FVNQHLCGSHLVE
DiglycatedB 22-30 RGFFYTPKAB 22-30 RGFFYTPKAA 18-21/B 14-21 NYCN/ALYLVCGEA 5-17 QCCASVCSLYQLEB 1-13 FVNQHLCGSHLVEA 1-21 GIVEQCCASVCSLY
TriglycatedB 22-30 RGFFYTPKAB 22-30 RGFFYTPKAA 18-21/B 14-21 NYCN/ALYLVCGEA 5-17 QCCASVCSLYQLEB 1-13 FVNQHLCGSHLVE
A 1-21 GIVEQCCASVCSLYQLENreducing conditions) with one glucose adduct attached(see Table 1).
Analysis of the MS/MS spectrum of the modifiedpeptide at m/z 1248.57 (Figure 3a) allows locating theglucose adduct in residue Lys29. In the MS/MS spec-trum, it is possible to see the mass difference of 290.2 Dabetween the ion at m/z 1131.5 (a8) and the ion at m/z841.3 (a7). This gap corresponds to the loss of the lysineresidue with a glucose adduct. Analysis of the MS/MSspectrum of the precursor ion at 1644.68 (see Figure 3b)allows locating the glucose adduct in residue Phe1.These results agree with the literature [21, 22], whichstate the most likely sites of glycation within the B-chainare the Phe1 and Lys29 residues, as both contain freeamino groups for linking glucose molecules. Neverthe-less, the preferred glycation site of monoglycated bo-vine or human insulin is thought to be at Phe-1 of the
igestion of native insulin and experimental glycated fragmentsresidues correspond to cysteines involved in disulfide bridges
nceTheoreticalmass (Da)
Experimentalmass (Da)
Matcherror (Da)
417.2324 — —1446.6062 — —1444.5906 1444.4225 �0.1681513.1762 — —
1482.7158 1482.6351 �0.0807867.4280 867.4041 �0.0239
1086.5731 1086.5048 �0.0683HLCGSHLVE 2924.283 — —
1377.5808 1377.4717 �0.1091
867.4280 867.4378 0.00981086.5731 1086.5291 �0.04401248.6259 1248.5710 �0.05491377.5808 1377.5029 �0.07791444.5906 1444.5505 �0.04011482.7158 1482.6345 �0.08131644.7686 1644.6820 �0.0866
1086.5731 1086.4946 �0.07851250.6415 1250.5546 �0.08691377.5808 1377.4662 �0.11461444.5906 1444.5294 �0.06121646.7842 1646.6748 �0.1094
1086.5731 1086.4970 �0.07611250.6415 1250.5597 �0.08181377.5808 1377.4740 �0.10681444.5906 1444.5748 �0.01581646.7842 1646.7093 �0.0749
YCN 2501.0345 2501.0765 0.0420
1086.5731 1086.4949 �0.07821250.6415 1250.5478 �0.09371377.5808 1377.4398 �0.14101444.5906 1444.5100 �0.08061646.7842 1646.6378 �0.1464
-C dned
eque
VNQ
QLEN
YCN 2501.0345 2501.1065 0.0720
1323J Am Soc Mass Spectrom 2009, 20, 1319–1326 TANDEM MS OF GLYCATED INSULIN
B-chain [23]. This is most probably due to the highreactivity as this residue is situated at the N-terminal ofthe peptide chain. Herein, we show that monoglycatedinsulin produced in the absence of reducing conditionscomprises the coexistence of two glycated insulin spe-cies with two different glycation sites, Phe1 or Lys29.
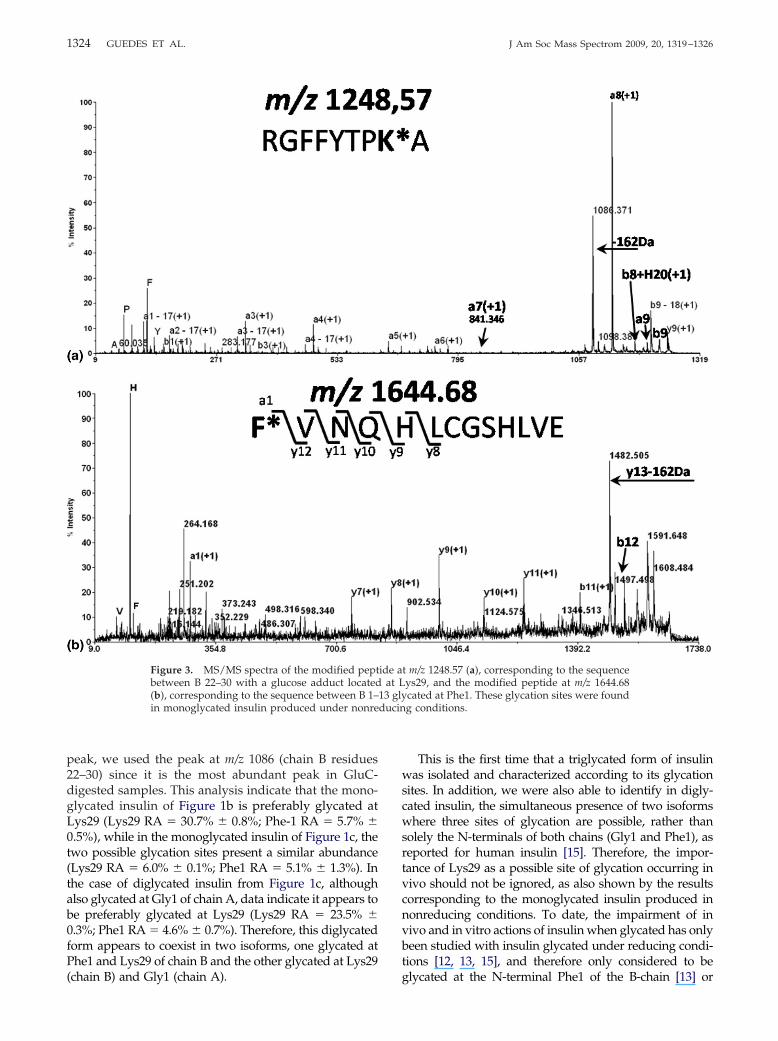
Figure 4 shows three representative examples ofMS/MS spectra of glycated fragments belonging to gl-ycated insulin produced under reducing conditions.The two glycated fragments that result from Glu-Cdigestion are the peptides at m/z 1250.56 and m/z1646.67, and were found in all of three forms of glycatedinsulin (see Figure 2). These peptides were fragmentsB1-13 and B22-30 with identical glycation sites, Phe1and Lys29, respectively, as reported for the monogly-cated insulin formed in the absence of reducing condi-tions. In this case, because of the excess of reducingagent (NaBH3CN), the mass shift found in the MS/MSspectra corresponds to the attachment of a glucitoladduct (164 Da). Again, the coexistence of two glycatedforms occurs in monoglycated insulin produced underreducing conditions.
With the approach described so far, it was not
Figure 2. MALDI-MS spectra of Glu-C digests ofor both glycation procedures. The m/z values oshown in Table 1 are highlighted with rectanglycated samples are unique to these samplesinsulin, (b) monoglycated insulin under nonrereducing conditions, (d) diglycated insulin (redconditions).
possible to identify any glycated fragments belonging
to the A-chain in monoglycated, diglycated and trigly-cated forms of insulin. To induce the separation of thetwo chains (A and B) of the insulin samples, thedisulphide bridges were reduced with DTT using analiquot of undigested samples, as described in theExperimental section. These reduced samples were sub-mitted to MALDI-MS and MALDI-MS/MS analysis.Figure 4c shows the representative MS/MS spectrum ofthe A-chain containing a single glucitol adduct, with anm/z 2501, which was found only in diglycated andtriglycated forms of insulin. The identification of thecomplete b series allowed the identification of Gly1 asthe glycation site in the A-chain. This result alloweddetermining the three sites of glycation in triglycatedinsulin and also pointed out the presence of two iso-forms in diglycated insulin, similar to the monoglycatedforms of insulin. Therefore, in diglycated insulin, thereis the coexistence of one specie glycated at the N-terminals of both chains (Gly1 and Phe1) and anotherspecie containing the two glucitol adducts in chain B(Phe1 and Lys29).
Furthermore, we calculated the relative abundance(RA), in percentage, of glycated peptides for monogly-
ive insulin and isolated forms of glycated insulingments matching theoretical peptide fragmentsPeptide fragments highlighted with circles incorrespond to glycated fragments. (a) Nativeg conditions, (c) monoglycated insulin under
g conditions), (e) triglycated insulin (reducing
f natf fra
gles.and
ducinucin
cated and diglycated samples. As normalizing reference
ducin
1324 GUEDES ET AL. J Am Soc Mass Spectrom 2009, 20, 1319–1326
peak, we used the peak at m/z 1086 (chain B residues22–30) since it is the most abundant peak in GluC-digested samples. This analysis indicate that the mono-glycated insulin of Figure 1b is preferably glycated atLys29 (Lys29 RA � 30.7% � 0.8%; Phe-1 RA � 5.7% �0.5%), while in the monoglycated insulin of Figure 1c, thetwo possible glycation sites present a similar abundance(Lys29 RA � 6.0% � 0.1%; Phe1 RA � 5.1% � 1.3%). Inthe case of diglycated insulin from Figure 1c, althoughalso glycated at Gly1 of chain A, data indicate it appears tobe preferably glycated at Lys29 (Lys29 RA � 23.5% �0.3%; Phe1 RA � 4.6% � 0.7%). Therefore, this diglycatedform appears to coexist in two isoforms, one glycated atPhe1 and Lys29 of chain B and the other glycated at Lys29
Figure 3. MS/MS spectra of the modified peptbetween B 22–30 with a glucose adduct located(b), corresponding to the sequence between B 1–in monoglycated insulin produced under nonre
(chain B) and Gly1 (chain A).
This is the first time that a triglycated form of insulinwas isolated and characterized according to its glycationsites. In addition, we were also able to identify in digly-cated insulin, the simultaneous presence of two isoformswhere three sites of glycation are possible, rather thansolely the N-terminals of both chains (Gly1 and Phe1), asreported for human insulin [15]. Therefore, the impor-tance of Lys29 as a possible site of glycation occurring invivo should not be ignored, as also shown by the resultscorresponding to the monoglycated insulin produced innonreducing conditions. To date, the impairment of invivo and in vitro actions of insulin when glycated has onlybeen studied with insulin glycated under reducing condi-tions [12, 13, 15], and therefore only considered to be
t m/z 1248.57 (a), corresponding to the sequenceys29, and the modified peptide at m/z 1644.68cated at Phe1. These glycation sites were foundg conditions.
ide aat L
13 gly
glycated at the N-terminal Phe1 of the B-chain [13] or

1325J Am Soc Mass Spectrom 2009, 20, 1319–1326 TANDEM MS OF GLYCATED INSULIN
glycated at the N-terminals of both chains (Gly1 and Phe1)[15]. It would be of interest to develop further work withglycated insulin at Lys29, since insulin glycation in vivo isassociated with characteristic complications of type 2diabetes such as glucose intolerance and insulin resis-tance. Elucidating the importance of the structural alter-ations, which occur in consequence of glycation, in themodulation of insulin action should continue to be ofgreat interest.
AcknowledgmentsThe authors express their appreciation for the financial supportprovided by the “Fundação para a Ciência e Tecnologia” (FCT-grant SFRH/BD/19,769/2004) and PTDC/QUI/68,408/2006.
References1. Brownlee, M. Negative Consequences of Glycation. Metabolism. 2000,
49(2) (Suppl 1), 9–13.2. Bookchin, R. M.; Gallop, P. M. Structure of Hemoglobin AIc: Nature of
the N-Terminal beta Chain Blocking Group. Biochem. Biophys. Res.Commun. 1968, 32(1), 86–93.
3. Sen, S.; Kar, M.; Roy, A.; Chakraborti, A. S. Effect of NonenzymaticGlycation on Functional and Structural Properties of Hemoglobin.Biophys. Chem. 2005, 113(3), 289–298.
4. Ganea, E.; Harding, J. J. Inactivation of Glucose-6-Phosphate Dehydro-genase by Glycation. Biochem. Soc. Trans. 1994, 22(4), 445S.
5. Ganea, E.; Harding, J. J. Trehalose and 6-Aminohexanoic Acid Stabilize andRenature Glucose-6-Phosphate Dehydrogenase Inactivated by Glycationand by Guanidinium Hydrochloride. Biol. Chem. 2005, 386(3), 269–278.
6. Takahashi, M.; Lu, Y. B.; Myint, T.; Fujii, J.; Wada, Y.; Taniguchi, N. InVivo Glycation of Aldehyde Reductase, a Major 3-Deoxyglucosone
Figure 4. MS/MS spectrum of the modified pepbetween B 22–30 with a glucose adduct locatemodified peptide at m/z 1646.67, which correspo(c) shows an MS/MS spectrum representative2501.10, where the glycation site is located atinsulin produced under reducing conditions.
Reducing Enzyme: Identification of Glycation Sites. Biochemistry. 1995,34(4), 1433–1148.
7. Blakytny, R.; Harding, J. J. Glycation (Nonenzymic Glycosylation) Inacti-vates Glutathione Reductase. Biochem J. 1992, 288(Pt 1), 303–307.
8. Niwa, T.; Tsukushi, S. 3-Deoxyglucosone and AGEs in Uremic Compli-cations: Inactivation of Glutathione Peroxidase by 3-Deoxyglucosone.Kidney Int. Suppl. 2001, 78, S37–41.
9. Baldwin, J. S.; Lee, L.; Leung, T. K.; Muruganandam, A.; Mutus, B.Identification of the Site of Nonenzymatic Glycation of GlutathionePeroxidase: Rationalization of the Glycation-Related Catalytic Alter-ations on the Basis of Three-Dimensional Protein Structure. Biochim.Biophys. Acta. 1995, 1247(1), 60–64.
10. Arai, K.; Iizuka, S.; Tada, Y.; Oikawa, K.; Taniguchi, N. Increase in theGlucosylated Form of Erythrocyte Cu-Zn-Superoxide Dismutase inDiabetes and Close Association of the Nonenzymatic Glucosylationwith the Enzyme activity. Biochim. Biophys. Acta. 1987, 924(2), 292–296.
11. Oda, A.; Bannai, C.; Yamaoka, T.; Katori, T.; Matsushima, T.; Yamashita, K.Inactivation of Cu, Zn-Superoxide Dismutase by In Vitro Glycosylationand in Erythrocytes of Diabetic Patients. Horm. Metab. Res. 1994, 26(1), 1–4.
12. Abdel-Wahab, Y. H.; O’Harte, F. P.; Boyd, A. C.; Barnett, C. R.; Flatt,P. R. Glycation of Insulin Results in Reduced Biological Activity in Mice.Acta Diabetol. 1997, 34(4), 265–270.
13. Boyd, A. C.; Abdel-Wahab, Y. H.; McKillop, A. M.; McNulty, H.;Barnett, C. R.; O’Harte, F. P.; Flatt, P. R. Impaired Ability of GlycatedInsulin to Regulate Plasma Glucose and Stimulate Glucose Transportand Metabolism in Mouse Abdominal Muscle. Biochim. Biophys. Acta.2000, 1523(1), 128–134.
14. Dolhofer, R.; Wieland, O. H. Preparation and Biological Properties ofGlycosylated Insulin. FEBS Lett. 1979, 100(1), 133–136.
15. O’Harte, F. P.; Boyd, A. C.; McKillop, A. M.; Abdel-Wahab, Y. H.;McNulty, H.; Barnett, C. R.; Conlon, J. M.; Hojrup, P.; Flatt, P. R.Structure, Antihyperglycemic Activity and Cellular Actions of a NovelDiglycated Human Insulin. Peptides. 2000, 21(10), 1519–1526.
16. Schalkwijk, C. G.; Brouwers, O.; Stehouwer, C. D. Modulation of InsulinAction by Advanced Glycation End Products: A New Player in theField. Horm. Metab. Res. 2008, 40(9), 614–619.
17. Abdel-Wahab, Y. H.; O’Harte, F. P.; Ratcliff, H.; McClenaghan, N. H.;Barnett, C. R.; Flatt, P. R. Glycation of Insulin in the Islets of Langerhansof Normal and Diabetic Animals. Diabetes. 1996, 45(11), 1489–1496.
18. Abdel-Wahab, Y. H.; O’Harte, F. P.; Barnett, C. R.; Flatt, P. R. Glycationof Insulin in a Cultured Insulin-Secreting Cell Line. Biochem. Soc. Trans.1997, 25(1), 128S.
19. Abdel-Wahab, Y. H.; O’Harte, F. P.; Barnett, C. R.; Flatt, P. R. Charac-
at m/z 1250.56 (a), corresponding to the sequenceLys29. (b) Shows the MS/MS spectrum of the
the sequence between B 1–13 glycated at Phe1.e A-chain with a single glucitol adduct at m/z
These glycation sites were found in glycated
tided atnds toof thGly1.
terization of Insulin Glycation in Insulin-Secreting Cells Maintained inTissue Culture. J. Endocrinol. 1997, 152(1), 59–67.
1326 GUEDES ET AL. J Am Soc Mass Spectrom 2009, 20, 1319–1326
20. Anzenbacher, P.; Kalous, V. Binding of D-Glucose to Insulin. Biochim.Biophys. Acta. 1975, 386(2), 603–607.
21. Farah, M. A.; Bose, S.; Lee, J. H.; Jung, H. C.; Kim, Y. Analysis of
Glycated Insulin by MALDI-TOF Mass Spectrometry. Biochim. Biophys.Acta. 2005, 1725(3), 269–282.22. McKillop, A. M.; Meade, A.; Flatt, P. R.; O’Harte, F. P. Evaluation of theSite(s) of Glycation in Human Proinsulin by Ion-Trap LCQ ElectrosprayIonization Mass Spectrometry. Regul. Pept. 2003, 113(1/3), 1–8.
23. O’Harte, F. P.; Hojrup, P.; Barnett, C. R.; Flatt, P. R. Identification of theSite of Glycation of Human Insulin. Peptides. 1996, 17(8), 1323–1330.
Related Documents











