Mass spectrometric characterization and physiological actions of GAHKNYLRFamide, a novel FMRFamide-like peptide from crabs of the genus Cancer Nelson D. Cruz-Bermu ´ dez,* Qiang Fu, Kimberly K. Kutz-Naber, Andrew E. Christie,à Lingjun Li and Eve Marder* *Volen Center and Department of Biology, Brandeis University, Waltham, Massachusetts, USA School of Pharmacy and Department of Chemistry, University of Wisconsin, Madison, Wisconsin, USA àDepartment of Biology and Friday Harbor Laboratories, University of Washington, Seattle, Washington, USA Abstract The stomatogastric ganglion (STG) and the cardiac ganglion (CG) of decapod crustaceans are modulated by neuroactive substances released locally and by circulating hormones re- leased from neuroendocrine structures including the pericar- dial organs (POs). Using nanoscale liquid chromatography electrospray ionization quadrupole-time-of-flight tandem mass spectrometry and direct tissue matrix-assisted laser desorp- tion/ionization Fourier transform mass spectrometry we have identified and sequenced a novel neuropeptide, GAHK- NYLRFamide (previously misassigned as KHKNYLRFamide in a study that did not employ peptide derivatization), from the POs and/or the stomatogastric nervous system (STNS) of the crabs, Cancer borealis, Cancer productus and Cancer mag- ister. In C. borealis, exogenous application of GAHKNYLRF- amide increased the burst frequency and number of spikes per burst of the isolated CG and re-initiated bursting activity in non-bursting ganglia, effects also elicited by the FMRFamide- like peptides (FLPs) SDRNFLRFamide and TNRNFLRFa- mide. In the intact STNS (which contains the STG), exogen- ous application of GAHKNYLRFamide increased the frequency of the pyloric rhythm and activated the gastric mill rhythm, effects also similar to those elicited by SDRNFLRF- amide and TNRNFLRFamide. FLP-like immunoreactivity in the POs and the STNS was abolished by pre-adsorption with the synthetic GAHKNYLRFamide. Different members of the FLP family exhibited differential degradation in the presence of extracellular peptidases. Taken collectively, the amino acid sequence of GAHKNYLRFamide, the blocking of FLP-like immunostaining, and its physiological effects on the CG and STNS suggest that this peptide is a novel member of the FLP superfamily. Keywords: cardiac ganglion, crustaceans, electrospray ion- ization quadrupole-time-of-flight mass spectrometry, neuro- peptides, pericardial organs, stomatogastric ganglion. J. Neurochem. (2006) 97, 784–799. Peptides modulate the output of neural circuits in all animals (Nusbaum et al. 2001; Marder and Thirumalai 2002). To understand fully the regulation of any neural circuit it is crucial to identify all the peptides that are used as modulators and to characterize their actions on the motor patterns produced by the circuit. The stomatogastric ganglion (STG), contained within the stomatogastric ner- vous system (STNS), and the cardiac ganglion (CG) of Received September 16, 2005; revised manuscript received December 20, 2005; accepted December 22, 2005. Address correspondence and reprint requests to Dr Lingjun Li, School of Pharmacy, University of Wisconsin, 777 Highland Avenue, Madison, WI 53705-2222, USA. E-mail: [email protected] Abbreviations used: A-l.n, anterolateral nerve; a, amide; CG, cardiac ganglion; coc, circumoesophageal connective; CoG, commissural gan- glion; DG, dorsal gastric; dgn, dorsal gastric nerve; DHB, 2,5-dihyd- roxybenzoic acid; dpon, dorsal posterior oesophageal nerve; dvn, dorsal ventricular nerve; ESI, electrospray ionization; FLP, FMRFamide-like peptide; GM, gastric mill; IC, inferior cardiac; ion, inferior oesophageal nerve; ivn, inferior ventricular nerve; LC, liquid chromatography; LG, lateral gastric; lgn, lateral gastric nerve; LP, lateral pyloric; lvn, lateral ventricular nerve; MALDI, FTMS, matrix-assisted laser desorption/ ionization Fourier transform mass spectrometry; MN, motor neuron; MS/ MS, tandem mass spectometry; mvn, medial ventricular nerve; OG, oesophageal ganglion; on, oesophageal nerve; PD, pyloric dilator; pdn, pyloric dilator nerve; P-l.n., posterolateral nerve; PO, pericardial organ; PY, pyloric; pyn, pyloric nerve; QTOF, quadrupole time of flight; SG, sinus gland; son, superior oesophageal nerve; STG, stomatogastric ganglion; stn, stomatogastric nerve; STNS, stomatogastric nervous sys- tem; TTX, tetrodotoxin; VD, ventricular dilator. Journal of Neurochemistry , 2006, 97, 784–799 doi:10.1111/j.1471-4159.2006.03747.x 784 Journal Compilation Ó 2006 International Society for Neurochemistry, J. Neurochem. (2006) 97, 784–799 Ó 2006 The Authors

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Mass spectrometric characterization and physiological actions ofGAHKNYLRFamide, a novel FMRFamide-like peptide from crabsof the genus Cancer
Nelson D. Cruz-Bermudez,* Qiang Fu,� Kimberly K. Kutz-Naber,� Andrew E. Christie,�Lingjun Li� and Eve Marder*
*Volen Center and Department of Biology, Brandeis University, Waltham, Massachusetts, USA
�School of Pharmacy and Department of Chemistry, University of Wisconsin, Madison, Wisconsin, USA
�Department of Biology and Friday Harbor Laboratories, University of Washington, Seattle, Washington, USA
Abstract
The stomatogastric ganglion (STG) and the cardiac ganglion
(CG) of decapod crustaceans are modulated by neuroactive
substances released locally and by circulating hormones re-
leased from neuroendocrine structures including the pericar-
dial organs (POs). Using nanoscale liquid chromatography
electrospray ionization quadrupole-time-of-flight tandem mass
spectrometry and direct tissue matrix-assisted laser desorp-
tion/ionization Fourier transform mass spectrometry we have
identified and sequenced a novel neuropeptide, GAHK-
NYLRFamide (previously misassigned as KHKNYLRFamide
in a study that did not employ peptide derivatization), from the
POs and/or the stomatogastric nervous system (STNS) of the
crabs, Cancer borealis, Cancer productus and Cancer mag-
ister. In C. borealis, exogenous application of GAHKNYLRF-
amide increased the burst frequency and number of spikes
per burst of the isolated CG and re-initiated bursting activity in
non-bursting ganglia, effects also elicited by the FMRFamide-
like peptides (FLPs) SDRNFLRFamide and TNRNFLRFa-
mide. In the intact STNS (which contains the STG), exogen-
ous application of GAHKNYLRFamide increased the
frequency of the pyloric rhythm and activated the gastric mill
rhythm, effects also similar to those elicited by SDRNFLRF-
amide and TNRNFLRFamide. FLP-like immunoreactivity in
the POs and the STNS was abolished by pre-adsorption with
the synthetic GAHKNYLRFamide. Different members of the
FLP family exhibited differential degradation in the presence
of extracellular peptidases. Taken collectively, the amino acid
sequence of GAHKNYLRFamide, the blocking of FLP-like
immunostaining, and its physiological effects on the CG and
STNS suggest that this peptide is a novel member of the FLP
superfamily.
Keywords: cardiac ganglion, crustaceans, electrospray ion-
ization quadrupole-time-of-flight mass spectrometry, neuro-
peptides, pericardial organs, stomatogastric ganglion.
J. Neurochem. (2006) 97, 784–799.
Peptides modulate the output of neural circuits in allanimals (Nusbaum et al. 2001; Marder and Thirumalai2002). To understand fully the regulation of any neuralcircuit it is crucial to identify all the peptides that are usedas modulators and to characterize their actions on themotor patterns produced by the circuit. The stomatogastricganglion (STG), contained within the stomatogastric ner-vous system (STNS), and the cardiac ganglion (CG) of
Received September 16, 2005; revised manuscript received December20, 2005; accepted December 22, 2005.Address correspondence and reprint requests to Dr Lingjun Li, School
of Pharmacy, University of Wisconsin, 777 Highland Avenue, Madison,WI 53705-2222, USA. E-mail: [email protected]
Abbreviations used: A-l.n, anterolateral nerve; a, amide; CG, cardiacganglion; coc, circumoesophageal connective; CoG, commissural gan-glion; DG, dorsal gastric; dgn, dorsal gastric nerve; DHB, 2,5-dihyd-roxybenzoic acid; dpon, dorsal posterior oesophageal nerve; dvn, dorsalventricular nerve; ESI, electrospray ionization; FLP, FMRFamide-likepeptide; GM, gastric mill; IC, inferior cardiac; ion, inferior oesophagealnerve; ivn, inferior ventricular nerve; LC, liquid chromatography; LG,lateral gastric; lgn, lateral gastric nerve; LP, lateral pyloric; lvn, lateralventricular nerve; MALDI, FTMS, matrix-assisted laser desorption/ionization Fourier transform mass spectrometry; MN, motor neuron; MS/MS, tandem mass spectometry; mvn, medial ventricular nerve; OG,oesophageal ganglion; on, oesophageal nerve; PD, pyloric dilator; pdn,pyloric dilator nerve; P-l.n., posterolateral nerve; PO, pericardial organ;PY, pyloric; pyn, pyloric nerve; QTOF, quadrupole time of flight; SG,sinus gland; son, superior oesophageal nerve; STG, stomatogastricganglion; stn, stomatogastric nerve; STNS, stomatogastric nervous sys-tem; TTX, tetrodotoxin; VD, ventricular dilator.
Journal of Neurochemistry, 2006, 97, 784–799 doi:10.1111/j.1471-4159.2006.03747.x
784 Journal Compilation � 2006 International Society for Neurochemistry, J. Neurochem. (2006) 97, 784–799� 2006 The Authors

decapod crustaceans have been used extensively as modelsystems for understanding the basic cellular mechanismsand synaptic physiology that underlie the generation andmodulation of neural circuit output. Work from manylaboratories has shown that both the STG and CG neuralcircuits are modulated by peptides released locally frommodulatory inputs, as well as by peptide hormonesreleased from endocrine sites and delivered to bothsystems via the hemolymph (Cooke and Hartline 1975;Christie et al. 1995; Skiebe 2001; Cooke 2002; Li et al.2002, 2003; Pulver and Marder 2002).
The FMRFamide family of peptides are well knownmodulators of neural circuit activity. Originally identifiedin the bivalve mollusk Macrocallista nimbosa (Price andGreenberg 1977), members of this peptide family havesubsequently been identified in a wide variety of organ-isms, including crustaceans. In decapod species, nopeptides possessing an FMRFamide C-terminus have beenidentified; however, a number of N-terminally extendedFLRFamides have been characterized (Trimmer et al.1987; Krajniak 1991; Mercier et al. 1993; Weimann et al.1993; Sithigorngul et al. 2002; Huybrechts et al. 2003; Liet al. 2003; Fu and Li 2005; Fu et al. 2005). Based ontheir amino acid sequences and the fact that theirphysiological actions in crustaceans are similar to FMRF-amide, these extended FLRFamides are considered to bemembers of the FMRFamide-like peptide (FLP) super-family.
Recently, several peptides containing an YLRFamide C-terminus were identified in decapod crustaceans (Sithigorn-gul et al. 2002; Fu and Li 2005; Fu et al. 2005). Thesepeptides, AYSNLNYLRFamide (Sithigorngul et al. 2002)and GYSKNYLRFamide (Fu and Li 2005; Fu et al. 2005),like the N-terminally extended FLRFamides, have beenplaced into the FLP superfamily based on amino acidsequence similarities. Here, using a combination of massspectrometric techniques, we have identified a novel YLRF-amide isoform, GAHKNYLRFamide, which is present in atleast three species of Cancer crabs: Cancer borealis, Cancerproductus and Cancer magister. Using synthetic peptide, weshow that GAHKNYLRFamide is a potent modulator of theC. borealis stomatogastric and cardiac neural circuits.Moreover, we show that the physiological effects elicitedby GAHKNYLRFamide on both circuits are similar to thoseelicited by SDRNFLRFamide and TNRNFLRFamide, twonative C. borealis FLPs (Weimann et al. 1993). In addition,the FMRFamide-like immunoreactivity observed in thepericardial organs (POs) and the STNS was abolished bypre-incubation of the antibody with synthetic peptideGAHKNYLRFamide in all three species of Cancer crabs.Collectively, our findings support the hypothesis that this andall other YLRFamides are indeed members of the FLPsuperfamily.
Materials and methods
Animals and dissection
C. borealis were obtained from the Marine Biological Laboratories
(Woods Hole, MA, USA) and Commercial Lobster (Boston, MA,
USA). C. productus and C. magister were collected by hand, ring-
trap or dredge in the Puget Sound area and San Juan Archipelago of
Washington State (USA). Regardless of species, animals were
maintained without food in artificial seawater tanks at 10–12�C.For the isolation of tissues, animals were cold-anesthetized by
packing them in ice for 15–30 min, after which the heart, foregut
eyestalks and lateral walls of the pericardial chamber were isolated.
The cardiac nervous system was isolated from the heart as described
below, while the intact STNS or selected regions of it, namely the
STG and/or the commissural ganglia (CoGs), were isolated from the
foregut. Sinus glands (SGs) were isolated from the eyestalks. POs
were obtained from the lateral walls of the pericardial chamber.
Irrespective of tissue, all dissections were carried out in chilled
physiological saline (440 mM NaCl, 13 mM KCl, 26 mM MgCl2,
13 mM CaCl2, 11 mM Trizma base, 5 mM maleic acid, pH 7.45).
FMRFamide-like peptide immunohistochemistry
To map the distribution of FLPs in neural tissues of C. borealis, C.magister and C. productus, STNSs, POs, SGs and cardiac nervous
systems were isolated and processed for FMRFamide immuno-
histochemistry as whole mounts. The labeling methods used have
been documented extensively in previous publications (Christie et al.2004a; Fu et al. 2005; Messinger et al. 2005). The primary antibody
used was a rabbit polyclonal antibody generated against the synthetic
peptide FMRFamide conjugated to bovine thyroglobulin via 4,4-
difluoro-3,3-dinitrophenyl sulfone (Immunostar Inc., Hudson, WI,
USA) at final dilutions of 1 : 300–1 : 500 with incubation times of
24–72 h. The secondary antibodies used were either donkey anti-
rabbit IgG conjugated to Alexa Fluor 488 (Molecular Probes, Eugene,
OR, USA) or donkey anti-rabbit IgG conjugated to Alexa Fluor 594
(Molecular Probes), each used at a final concentration of 1 : 300 with
an incubation time of 12–24 h. After immunolabeling, preparations
were viewed and data collected using a Bio-Rad MRC 600 laser
scanning confocal microscope (Bio-Rad Microscience Ltd., Hemel
Hempstead, UK), a Bio-Rad Radiance 2000 laser scanning confocal
microscope or a Nikon Eclipse E600 epifluorescence microscope
(Tokyo, Japan). Descriptions of the hardware and software used for
imaging on these systems are extensively documented in previous
publications (Christie et al. 1997, 2003, 2004a; Messinger et al.2005).
Antibody adsorption studies
To determine whether GAHKNYLRFamide might be among the
peptides detected by our FMRFamide immunohistochemistry system,
antibody adsorptions using this peptide were conducted. Specifically,
the FMRFamide antibody (1 : 500 dilution) was incubated with
10)5 M GAHKNYLRFamide for 2 h at room temperature (� 24�C)before incubation with tissue for 24 h. All other processing was
identical to that used for standard whole-mount labeling.
Direct tissue sample preparation
For direct tissue mass spectrometric analysis, samples were prepared
as described by Kutz et al. (2004). Briefly, dissected tissue was
Neuromodulation by GAHKNYLRFamide 785
� 2006 The AuthorsJournal Compilation � 2006 International Society for Neurochemistry, J. Neurochem. (2006) 97, 784–799

rinsed in acidified methanol, containing 90% methanol (Fisher
Scientific, Pittsburgh, PA, USA), 9% glacial acetic acid (Fisher
Scientific) and 1% deionized water. Tissue was then desalted in
dilute matrix-assisted laser desorption/ionization (MALDI) matrix
consisting of 10 mg/mL 2,5-dihydroxybenzoic acid (DHB) (MP
Biomedicals Inc., Irvine, CA, USA) prepared in deionized water. A
spot containing 0.6 lL saturated DHB [150 mg/mL in 50 : 50 v/v
water : purge trap-grade methanol (Fisher Scientific)] was placed on
one facet of the IonSpec MALDI sample probe. Before the DHB
crystallized fully, the tissue was placed carefully on the facet and an
additional 0.6-lL drop of saturated DHB was placed on top of the
tissue to help affix the tissue to the target. The saturated DHB was
then allowed to crystallize at room temperature.
Tissue extraction
Peptide extraction from pooled tissue was performed using ice-cold
acidified methanol. Approximately 30 POs were removed from their
acidified methanol storage buffer and placed in a 0.1-mL tissue
grinder (Wheaton, Inc., Millville, NJ, USA) along with 100 lLstorage buffer. The tissue was then homogenized completely, after
which the extraction liquid was transferred to a 1.5-mL microcen-
trifuge tube (Fisher Scientific) and centrifuged at 16 100 g for
10 min in an Eppendorf 5415 D microcentrifuge (Brinkmann
Instruments Inc., Westbury, NY, USA). The supernatant was
retained and the resulting pellet re-extracted with acidified methanol
and re-spun. Supernatants were combined and concentrated to
dryness with a Savant SC 110 SpeedVac concentrator (Thermo
Electron Corporation, West Palm Beach, FL, USA). Finally, a
minimal amount (20–300 lL) of resuspension solution (deionized
H2O with 0.1% trifluoroacetic acid or 0.1% formic acid) was added
to the extract. This resuspended extract was then vortexed and
briefly centrifuged with supernatant to be used for HPLC separation
followed by MALDI–Fourier transform mass spectrometry (FTMS)
screening or liquid chromatography (LC) tandem mass spectometry
(MS/MS) analysis.
Reductive methylation of tissue extract
Reductive methylation of tissue extract was performed using a
procedure described previously (Fu and Li 2005). The PO extracts
from C. borealis or C. productus were derivatized using this
method. Ten microliters of crude extract were mixed with 15 lLacetate buffer (1 M, pH 4.8), followed by the addition and mixing of
5 lL formaldehyde (37% in H2O v/v; Sigma-Aldrich, St Louis,
MO, USA), and subsequent addition of 2 lL 5 M NaBH3CN
(Sigma-Aldrich). The labeling reaction was allowed to take place for
6 h at room temperature. Excess formaldehyde was quenched by the
addition of 4 lL ammonia (37% in H2O v/v; Sigma-Aldrich). The
resulting solution was stored at )20�C before LC MS/MS analysis.
Formaldehyde-D2 labeling was performed in a similar manner,
except that 7 lL of formaldehyde-D2 (25% in H2O v/v; Isotec,
Sigma-Aldrich) were used instead.
Acetylation of the crude extract
The acetylation of tissue extracts was performed using a procedure
reported previously (Che and Fricker 2002). Ten microliters of C.productus PO extract were mixed with 0.5 M ammonium bicarbon-
ate solution (Fisher Scientific), followed by the addition of 1 lLacetic anhydride (Fisher Scientific). The solution was left at room
temperature for 10 min, and 2 lL of ammonia solution (37% in
H2O v/v; Sigma-Aldrich) were then added to maintain the pH. This
procedure was repeated three times, after which the resulting
solution was stored at )20�C before LC MS/MS analysis.
Peptide standards
The peptide standards SDRNFLRFamide and TNRNFLRFamide
were purchased from American Peptide Company (Sunnyvale, CA,
USA) and GAHKNYLRFamide was synthesized at the Biotechno-
logy Center of the University of Wisconsin (Madison, WI, USA).
Peptide degradation
Equal concentrations (10 lM) of synthetic peptide standards
SDRNFLRFamide, TNRNFLRFamide and GAHKNYLRFamide
were incubated in physiological saline in the presence of freshly
dissected POs from C. magister. The same peptide mixture was
incubated in tissue-free physiological saline as a control. In all
experiments, a 100-lL aliquot was prepared and incubated for 3 h at
both room temperature and 4�C. A 10-lL aliquot of the saline
solution was withdrawn every hour and 0.5 lL was spotted along
with the same volume of DHB MALDI matrix for mass spectro-
metric analysis.
MALDI FTMS
MALDI FTMS experiments were performed on an IonSpec Fourier
transform mass spectrometer (Lake Forest, CA, USA) equipped with
a 7.0-Tesla actively shielded superconducting magnet. The FTMS
instrument contained an external ion source utilizing a quadrupole
ion guide to transfer the ions to the ion cyclotron resonance (ICR)
cell, which was differentially pumped. The sample probe was a
10-faceted stainless steel target. A 337-nm nitrogen laser (Laser
Science, Inc., Franklin, MA, USA) was used for ionization/
desorption. All mass spectra were collected in positive ion mode
using the in-cell accumulation method. The latter method was
written using IonSpec version 7.0 software, as described previously
(Kutz et al. 2004). The pulse sequence involved the use of seven
laser ionization/desorption events, each being optimized for ions at
m/z 1000 to be transported down the quadrupole. This method was
used to increase the ion concentration before detection.
Capillary LC electrospray ionization (ESI) quadrupole-time-of-
flight (QTOF) MS/MS
Nanoscale LC QTOF MS/MS was performed using a Waters
capillary LC system coupled to a QTOF Micro mass spectrometer
(Waters Corp., Milford, MA, USA). Chromatographic separations
were performed on a reverse-phase capillary column (Atlantis�
dC18; 75 lm internal diameter · 100 mm long, 3 lm particle size;
Waters Corp.). The mobile phases used were: A, deionized H2O
with 5% acetonitrile and 0.1% formic acid; B, acetonitrile with 5%
deionized H2O and 0.1% formic acid; and C, deionized H2O with
0.1% formic acid. An aliquot of 1.4 lL of tissue extract was injected
and loaded onto the trap column (PepMapTM C18; 300lm column
internal diameter · 1 mm, 5 lm particle size; LC Packings,
Sunnyvale, CA, USA) using mobile phase C at a flow rate of
30 lL/min for 3 min. Following this, the stream select module was
switched to a position at which the trap column became in line with
the analytical capillary column, and a linear gradient of mobile
phases A and B was initiated. A splitter was added between the
786 N. D. Cruz-Bermudez et al.
Journal Compilation � 2006 International Society for Neurochemistry, J. Neurochem. (2006) 97, 784–799� 2006 The Authors

mobile phase mixer and the stream select module to reduce the flow
rate from 18 lL/min to 200 nL/min.
The nanoflow ESI source conditions were set as follows:
capillary voltage 3800 V, sample cone voltage 40 V, extraction
cone voltage 1 V, source temperature 120�C, cone gas (N2) 13 L/h.
For the reference spray, the same settings were used except that the
sample cone voltage was set at 10 V and reference scans were
performed every 10 s. A data-dependent acquisition was employed
for the MS survey scan and the selection of precursor ions and
subsequent MS/MS of the selected parent ions. The MS scan range
was from m/z 100–2000 and the MS/MS scan was from m/z 50–
2000. The MS/MS de novo sequencing was performed with a
combination of manual sequencing and automatic sequencing by
PepSeq software (Waters Corp.).
Isolation of CG
To isolate the CG for physiological experimentation, the anterior
portion of the carapace, the stomach and the hepatopancreas were
removed. Cuts were made laterally along the dorsal carapace on
either side of the heart chamber and a small piece of the carapace
with heart attached was separated from the animal. Muscles around
the heart were excised, and the hypodermis with the attached heart
was removed from the carapace by blunt dissection and the heart
subsequently pinned ventral side up in a Sylgard-coated (Dow
Corning, Midland, MI, USA) dish. A V-shaped cut was made from
the sternal artery on the ventral wall of the heart towards each of the
ventral ostia to expose the Y-shaped CG, which sits directly on the
dorsal inner wall of the heart muscle. The ganglion was freed from
the muscle by transecting each of the nerve roots at the most distal
point at which the root penetrated the muscle. In some preparations,
a small non-contracting piece of muscle was left attached to the
posterior end of the main trunk. The isolated ganglion was then
pinned down dorsal side up in a Sylgard-coated 25-mL dish
(60 · 15 mm) containing chilled (10–12�C) saline.
Isolation of STNS
To isolate the STNS for physiological experimentation, the foregut
was removed and the complete STNS isolated by manual dissection.
The isolated STNS was then pinned out in a Sylgard-coated petri
dish containing chilled saline. After pinning, the STG was
desheathed to facilitate both access of the neuropil to exogenously
applied substances and to aid in the penetration of the intrinsic STG
somata for intracellular recording. In experiments in which the STG
was isolated from anterior ganglia inputs, a well was built around
the stomatogastric nerve (stn) and the saline in the well was replacedwith an isotonic sucrose solution (750 mM) containing 10)6 M
tetrodotoxin (TTX).
Data acquisition and analysis
Petroleum jelly wells were built around the nerves to monitor the
electrical activity using stainless steel extracellular pin electrodes.
During recordings, preparations were continuously superfused with
chilled physiological saline (12�C) by means of a Peltier cooling
system, and were monitored using a thermoelectric probe in the
bath. For bath applications, peptides were diluted in saline
immediately before each application and were superfused by means
of a switching port on the inflow of a superfusion system. In some
experiments, peptides were pressure applied from a puffer micro-
pipette using a Picospritzer II (General Valve Corp., Fairfield, NJ,
USA). Intracellular recordings from the CG and STG neurons were
done using 20–40-MW glass microelectrodes filled with 0.6 M
K2SO4 and 20 mM KCl, and an Axoclamp 2A (Axon Instruments,
Foster City, CA, USA). Signals were amplified and filtered using an
A-M Systems 1700 Differential AC amplifier (Carlsborg, WA,
USA). Data were recorded to a computer hard drive using a Digidata
1322A data acquisition board and pClamp 8 software (Axon
Instruments). Data files were converted and analyzed in Spike 2
(version 4; Cambridge Electronic Design, Cambridge, UK). Statis-
tical tests were performed and graphs were prepared in SigmaPlot
(version 8; SPSS Inc., Chicago, IL, USA) and StatView (version 5;
SAS Institute, Cary, NC, USA), and figures were prepared in Canvas
(version 9; ACD Systems of America, Inc., Miami, FL, USA). Data
are presented as plots of the cycle frequency (the inverse of the cycle
period). Paired t-tests for statistical significance were performed on
the cycle period. Time stretches of several minutes in which the
range of values did not change visibly were assumed to represent the
steady state and used to determine the means. All histograms
represent the mean ± SEM unless stated otherwise.
Results
De novo sequencing of a new FLP by QTOF MS/MS
A novel FLP with a molecular mass of 1103.6 Da was firstde novo sequenced from the C. borealis PO extract bynanoscale LC ESI QTOF MS/MS. The MS/MS fragmenta-tion spectrum of this peptide is shown in Fig. 1(a) and aputative sequence of [GA/Q/K]H[K/Q]NYLRFamide wasderived. The low mass fragment ions, such as a1 and b1 ions,are missing in the spectrum. The a1 ion (G) is 30.03437 Daand thus cannot be detected during MS/MS scan (m/z 50–2000). The b1 ion is usually missing in the low-energycollision-induced dissociation MS/MS spectra (Papayanno-poulos 1995). The y8 ion is also weak. GA (or AG,128.058578 Da) has a mass very close to single amino acidresidue Q (128.05858 Da). The differentiation of such asmall mass difference is impossible with the mass measure-ment accuracy of any currently available mass spectrometer.Furthermore, it was previously reported that glycine in thesequence generally produces less abundant fragment ionsupon cleavage and might be difficult to detect (Papayanno-poulos 1995). Thus, based solely on the MS/MS spectrum ofthe native peptide, the first two amino acid residues cannotbe unequivocally determined. Another ambiguity is thedifferentiation of isobaric amino acid residues lysine(128.09496 Da) and glutamine (128.05858 Da), which havevery close masses. In fact, this novel peptide was previouslymisassigned as KHKNYLRFamide as a result of theseambiguities and incomplete fragmentation at the first twoamino acid residues (Kutz et al. 2004).
These sequence ambiguities were resolved by the use ofisotopic formaldehyde labeling. The reductive methylation
Neuromodulation by GAHKNYLRFamide 787
� 2006 The AuthorsJournal Compilation � 2006 International Society for Neurochemistry, J. Neurochem. (2006) 97, 784–799

labels the amino group at the N-terminus and the side chain oflysinewith twomethyl groups (or (CHD2)2- for formaldehyde-D2 labeling). After modification, the masses of the a and b ionseries increase by 28 Da (or 32 Da for formaldehyde-D2
labeling). It also produces an intense a1 ion and a strong y(n-1)ion. As shown in Figs 1(b) and 1(c), the a1 and y8 ions becomestronger. The 28-Da (32-Da for formaldehyde-D2 labeling)mass increment also enabled the detection of the a1 ion (58 and62 Da respectively) within theMS/MS scan range, yielding anunequivocal identification of glycine at the N-terminus.
The presence of lysine at the fourth position was alsodetermined by formaldehyde labeling. The side chain of thelysine residue was labeled and an additional 28-Da mass
increase was seen for the b4 ion. The masses of the y6 toy8 ions also increased by an additional 28 Da as a resultof this labeling. Thus, formaldehyde labeling suggests thatthe amino acid sequence of the peptide at 1103.6 Da isGAHKNYLRFamide rather than the previously assignedKHKNYLRFamide (Kutz et al. 2004).
Acetic anhydride labeling was also used to confirm thesequence assignment. This labeling adds one acetyl group(+ 42 Da) to the N-terminal amino group or the lysine sidechain. It can also form an ester with the hydroxyl benzene ofthe tyrosine side chain. Therefore, three acetyl modificationswere observed (+ 126 Da). As shown in Fig. 1(d), the MS/MS fragmentation spectrum confirmed our sequence assign-ment. This experiment highlights the unique advantage of theQTOF instrument in that low mass a1 and b1 ions can betransmitted and detected, allowing the unequivocal deter-mination of the N-terminal sequence. The nearly identicalfragmentation patterns between putative GAHKNYLRF-amide from C. borealis tissue extract and the syntheticpeptide further confirm the sequence derivation of the newpeptide (Fig. 2).
This peptide was also identified in the PO extract fromC. productus by LC ESI MS/MS (data not shown).
Direct tissue analysis by MALDI FTMS
To examine the distribution of this novel RFamide peptide inthe STNS and major neuroendocrine organs in C. borealis,we performed direct tissue analysis using MALDI FTMS.GAHKNYLRFamide was detected in both the CoG and theSTG (data not shown). In contrast to its presence in the POs,GAHKNYLRFamide was not present in the other majorneurosecretory site, the sinus gland (data not shown).
GAHKNYLRFamide is highly conserved among the threeCancer species. Figure 3 shows a comparison of the MALDI
Fig. 1 MS/MS fragmentation spectra of a novel FLP from C. borealis
pericardial organ extract. (a) Native peptide, GAHKNYLRFamide; (b)
(CH3)2GAHK(CH3)2NYLRFamide, with formaldehyde-H2 labeling; (c)
(CHD2)2GAHK(CHD2)2NYLRFamide, with formaldehyde-D2 labeling;
(d) G*AHK*NY*LRFamide with acetic anhydride labeling. Note that
acetylation (represented by *) at the N-terminus, lysine side chain and
tyrosine residue yielded a total mass shift of 126 Da. Modification sites
are indicated by asterisks in the sequences.
Fig. 2 MS/MS fragmentation spectra of GAHKNYLRFamide
(552.792+) eluted from LC MS/MS analysis of C. borealis PO tissue
extract (a) and synthetic peptide standard (b). The fragmentation
patterns of the peptide isolated from the C. borealis PO extract and the
synthetic peptide standard match well, confirming the peptide se-
quence assignment. Major sequence-specific fragment ions (immo-
nium ions H, R, b- and y-type ions) are identical in both spectra.
788 N. D. Cruz-Bermudez et al.
Journal Compilation � 2006 International Society for Neurochemistry, J. Neurochem. (2006) 97, 784–799� 2006 The Authors

FTMS peptide profiles of the POs from C. borealis, C.productus and C. magister. As seen, similar peptide profileswere observed with GAHKNYLRFamide (1104.6+) found inthe POs from all three species. The ubiquitous presence ofthis peptide in the POs of multiple species and its tissuedistribution within the STNS suggests a possible regulatoryrole in the neural networks in the STNS and in the CG.
FLP labeling is present in the PO and STNS, but not in
the SG or cardiac nervous system, of Cancer species
Previously, immunohistochemical methods were used todemonstrate the presence of FLPs in the POs of C. borealisand C. productus, and the absence of members of this familyin SGs of these species (Christie et al. 1995; Fu et al. 2005).The distribution of FLPs in the STNS of C. borealis wasmapped using the same methodology (Marder et al. 1987;Christie et al. 2004b). Here, we used FMRFamide-likeimmunostaining to assay the distribution of FLPs in thePOs, STNS, SGs and/or cardiac nervous systems of C.borealis, C. magister and C. productus.
Using whole mounts, we found that FLP-like immunore-activity is also present in the POs of C. magister (n ¼ 6). Asin previously investigated species such as C. borealis and C.productus (Christie et al. 1995; Fu et al. 2005), this labelingconsisted of immunopositive axons in the core of the nerveroots that form the PO, as well as fine fibers and superficiallylocated release terminals that branched from these axons(data not shown).
In the STNSs of both C. magister (n ¼ 4) and C.productus (n ¼ 12), extensive FLP-like labeling was seen.The immunopositive structures in both species were verysimilar to one another, as well as to those previouslyreported for C. borealis (Marder et al. 1987; Christie et al.2004b). Specifically, these profiles included stained somatain the CoGs (� 30 somata per ganglion in each species),oesophageal ganglion (OG) (four somata in C. borealisand C. magister and six somata in C. productus) andinferior ventricular nerve (ivn)/brain (two somata in eachspecies), immunopositive axons in the circumoesophagealconnective (cocs) (numerous axons projecting both anteriorand posterior to the CoG in each coc in each species),inferior oesophageal nerve (ions) (3–4 axons per ion ineach species), ivn (3–4 axons in each species), superioroesophageal nerve (sons) (8–10 axons per son in eachspecies), dorsal posterior oesophageal nerve (dpons) (alarge fascicle of axons per dpon in each species),oesophageal nerve (on) (3–4 axons in each species), stn(� 10 axons in each species) and dorsal ventricular nerve(dvn) (two axons in each species), and labeled neuropil inthe CoGs, STG and at the son/stn junction and in the sons(commonly several patches in each son near the dponjunction). As reported previously (Christie et al. 2004a), inC. productus, but not the other two species, four axonswere also labeled in each anterior cardiac nerve, arborizingthere to produce the neuroendocrine anterior cardiacplexus. Examples of FLP-like immunolabeling in theSTNS of C. productus are shown in Fig. 4.
As in C. borealis and C. productus (Christie et al. 1995;Fu et al. 2005), no FLP-like labeling was seen in the SGs ofC. magister (n ¼ 6; data not shown). Likewise no FLP-likeimmunoreactivity was seen in the cardiac nervous systems ofC. borealis, C. magister or C. productus (n ¼ 2, n ¼ 3 andn ¼ 5 respectively; data not shown).
Adsorption of FMRFamide-like immunostaining by
GAHKNYLRFamide
As GAHKNYLRFamide possesses a similar C-terminus tothe FLPs, we hypothesized that some of the FLP-likeimmunoreactivity seen in the POs and STNS might reflectthe presence of this peptide in these tissues. To test thishypothesis, we conducted antibody adsorptions using syn-thetic GAHKNYLRFamide as a blocking agent. In bothtissues and in all species, adsorption of the FMRFamideantibody with 10)5 M GAHKYLRFamide before use in
Fig. 3 MALDI FTMS direct tissue peptide profiling of (a) C. borealis
(b) C. productus and (c) C. magister POs. Signals correspond to the
protonated molecular ions, [M + H]+, where M is the molecular weight
of each peptide. Similar yet distinct peptide profiles are observed for
each species, with GAHKNYLRFamide (1104.6+) peptide found in all
three species (highlighted with a star). The 1146.6+ peak in panels (a)
to (c) is GYSKNYLRFamide.
Neuromodulation by GAHKNYLRFamide 789
� 2006 The AuthorsJournal Compilation � 2006 International Society for Neurochemistry, J. Neurochem. (2006) 97, 784–799
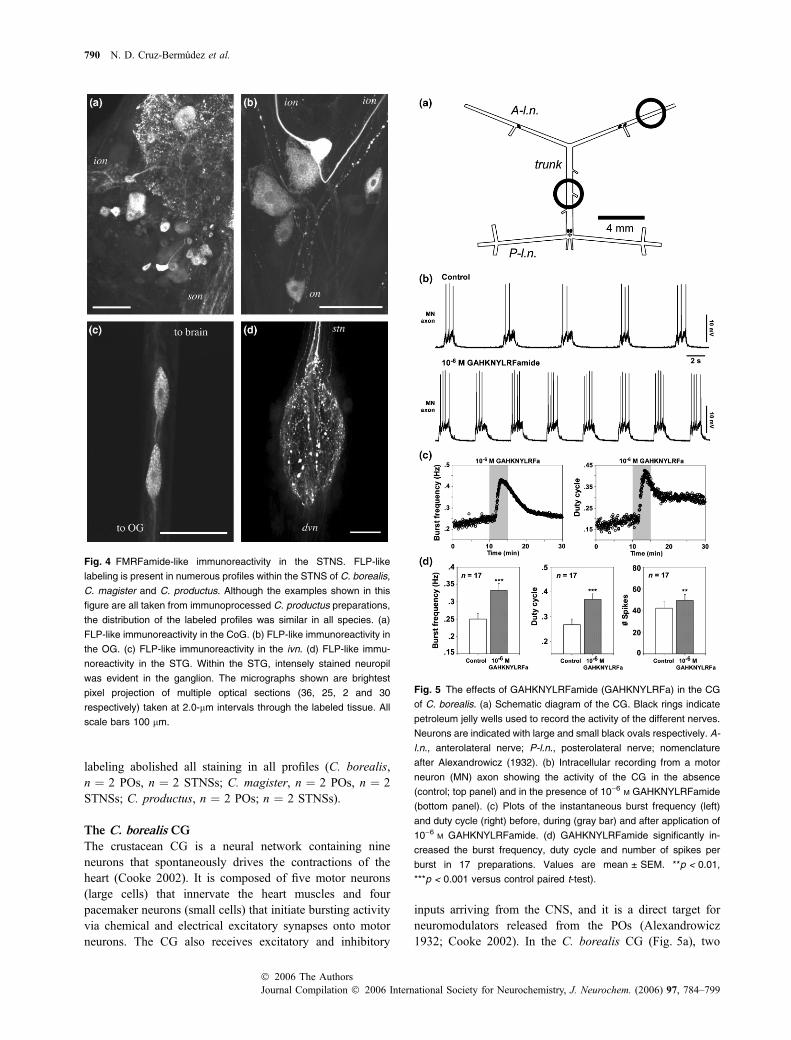
labeling abolished all staining in all profiles (C. borealis,n ¼ 2 POs, n ¼ 2 STNSs; C. magister, n ¼ 2 POs, n ¼ 2STNSs; C. productus, n ¼ 2 POs; n ¼ 2 STNSs).
The C. borealis CG
The crustacean CG is a neural network containing nineneurons that spontaneously drives the contractions of theheart (Cooke 2002). It is composed of five motor neurons(large cells) that innervate the heart muscles and fourpacemaker neurons (small cells) that initiate bursting activityvia chemical and electrical excitatory synapses onto motorneurons. The CG also receives excitatory and inhibitory
inputs arriving from the CNS, and it is a direct target forneuromodulators released from the POs (Alexandrowicz1932; Cooke 2002). In the C. borealis CG (Fig. 5a), two
Fig. 4 FMRFamide-like immunoreactivity in the STNS. FLP-like
labeling is present in numerous profiles within the STNS of C. borealis,
C. magister and C. productus. Although the examples shown in this
figure are all taken from immunoprocessed C. productus preparations,
the distribution of the labeled profiles was similar in all species. (a)
FLP-like immunoreactivity in the CoG. (b) FLP-like immunoreactivity in
the OG. (c) FLP-like immunoreactivity in the ivn. (d) FLP-like immu-
noreactivity in the STG. Within the STG, intensely stained neuropil
was evident in the ganglion. The micrographs shown are brightest
pixel projection of multiple optical sections (36, 25, 2 and 30
respectively) taken at 2.0-lm intervals through the labeled tissue. All
scale bars 100 lm.
Fig. 5 The effects of GAHKNYLRFamide (GAHKNYLRFa) in the CG
of C. borealis. (a) Schematic diagram of the CG. Black rings indicate
petroleum jelly wells used to record the activity of the different nerves.
Neurons are indicated with large and small black ovals respectively. A-
l.n., anterolateral nerve; P-l.n., posterolateral nerve; nomenclature
after Alexandrowicz (1932). (b) Intracellular recording from a motor
neuron (MN) axon showing the activity of the CG in the absence
(control; top panel) and in the presence of 10)6 M GAHKNYLRFamide
(bottom panel). (c) Plots of the instantaneous burst frequency (left)
and duty cycle (right) before, during (gray bar) and after application of
10)6 M GAHKNYLRFamide. (d) GAHKNYLRFamide significantly in-
creased the burst frequency, duty cycle and number of spikes per
burst in 17 preparations. Values are mean ± SEM. **p < 0.01,
***p < 0.001 versus control paired t-test).
790 N. D. Cruz-Bermudez et al.
Journal Compilation � 2006 International Society for Neurochemistry, J. Neurochem. (2006) 97, 784–799� 2006 The Authors
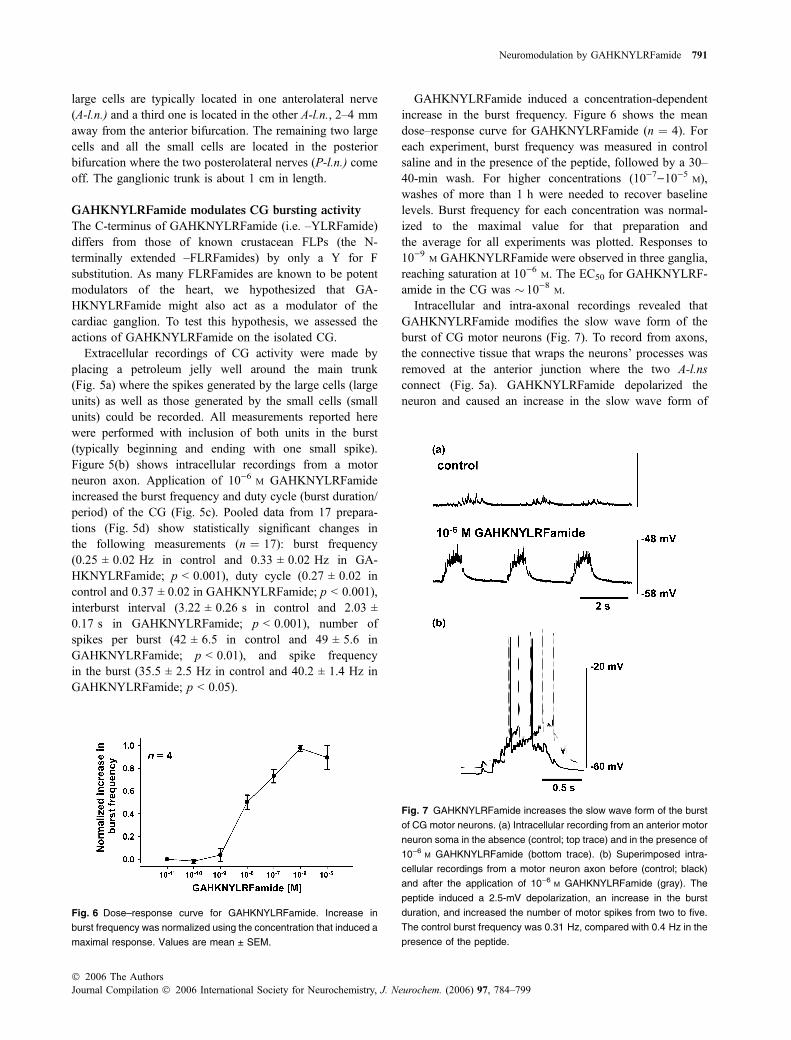
large cells are typically located in one anterolateral nerve(A-l.n.) and a third one is located in the other A-l.n., 2–4 mmaway from the anterior bifurcation. The remaining two largecells and all the small cells are located in the posteriorbifurcation where the two posterolateral nerves (P-l.n.) comeoff. The ganglionic trunk is about 1 cm in length.
GAHKNYLRFamide modulates CG bursting activity
The C-terminus of GAHKNYLRFamide (i.e. –YLRFamide)differs from those of known crustacean FLPs (the N-terminally extended –FLRFamides) by only a Y for Fsubstitution. As many FLRFamides are known to be potentmodulators of the heart, we hypothesized that GA-HKNYLRFamide might also act as a modulator of thecardiac ganglion. To test this hypothesis, we assessed theactions of GAHKNYLRFamide on the isolated CG.
Extracellular recordings of CG activity were made byplacing a petroleum jelly well around the main trunk(Fig. 5a) where the spikes generated by the large cells (largeunits) as well as those generated by the small cells (smallunits) could be recorded. All measurements reported herewere performed with inclusion of both units in the burst(typically beginning and ending with one small spike).Figure 5(b) shows intracellular recordings from a motorneuron axon. Application of 10)6 M GAHKNYLRFamideincreased the burst frequency and duty cycle (burst duration/period) of the CG (Fig. 5c). Pooled data from 17 prepara-tions (Fig. 5d) show statistically significant changes inthe following measurements (n ¼ 17): burst frequency(0.25 ± 0.02 Hz in control and 0.33 ± 0.02 Hz in GA-HKNYLRFamide; p < 0.001), duty cycle (0.27 ± 0.02 incontrol and 0.37 ± 0.02 in GAHKNYLRFamide; p < 0.001),interburst interval (3.22 ± 0.26 s in control and 2.03 ±0.17 s in GAHKNYLRFamide; p < 0.001), number ofspikes per burst (42 ± 6.5 in control and 49 ± 5.6 inGAHKNYLRFamide; p < 0.01), and spike frequencyin the burst (35.5 ± 2.5 Hz in control and 40.2 ± 1.4 Hz inGAHKNYLRFamide; p < 0.05).
GAHKNYLRFamide induced a concentration-dependentincrease in the burst frequency. Figure 6 shows the meandose–response curve for GAHKNYLRFamide (n ¼ 4). Foreach experiment, burst frequency was measured in controlsaline and in the presence of the peptide, followed by a 30–40-min wash. For higher concentrations (10)7)10)5 M),washes of more than 1 h were needed to recover baselinelevels. Burst frequency for each concentration was normal-ized to the maximal value for that preparation andthe average for all experiments was plotted. Responses to10)9 M GAHKNYLRFamide were observed in three ganglia,reaching saturation at 10)6 M. The EC50 for GAHKNYLRF-amide in the CG was � 10)8 M.
Intracellular and intra-axonal recordings revealed thatGAHKNYLRFamide modifies the slow wave form of theburst of CG motor neurons (Fig. 7). To record from axons,the connective tissue that wraps the neurons’ processes wasremoved at the anterior junction where the two A-l.nsconnect (Fig. 5a). GAHKNYLRFamide depolarized theneuron and caused an increase in the slow wave form of
Fig. 6 Dose–response curve for GAHKNYLRFamide. Increase in
burst frequency was normalized using the concentration that induced a
maximal response. Values are mean ± SEM.
Fig. 7 GAHKNYLRFamide increases the slow wave form of the burst
of CG motor neurons. (a) Intracellular recording from an anterior motor
neuron soma in the absence (control; top trace) and in the presence of
10)6 M GAHKNYLRFamide (bottom trace). (b) Superimposed intra-
cellular recordings from a motor neuron axon before (control; black)
and after the application of 10)6 M GAHKNYLRFamide (gray). The
peptide induced a 2.5-mV depolarization, an increase in the burst
duration, and increased the number of motor spikes from two to five.
The control burst frequency was 0.31 Hz, compared with 0.4 Hz in the
presence of the peptide.
Neuromodulation by GAHKNYLRFamide 791
� 2006 The AuthorsJournal Compilation � 2006 International Society for Neurochemistry, J. Neurochem. (2006) 97, 784–799
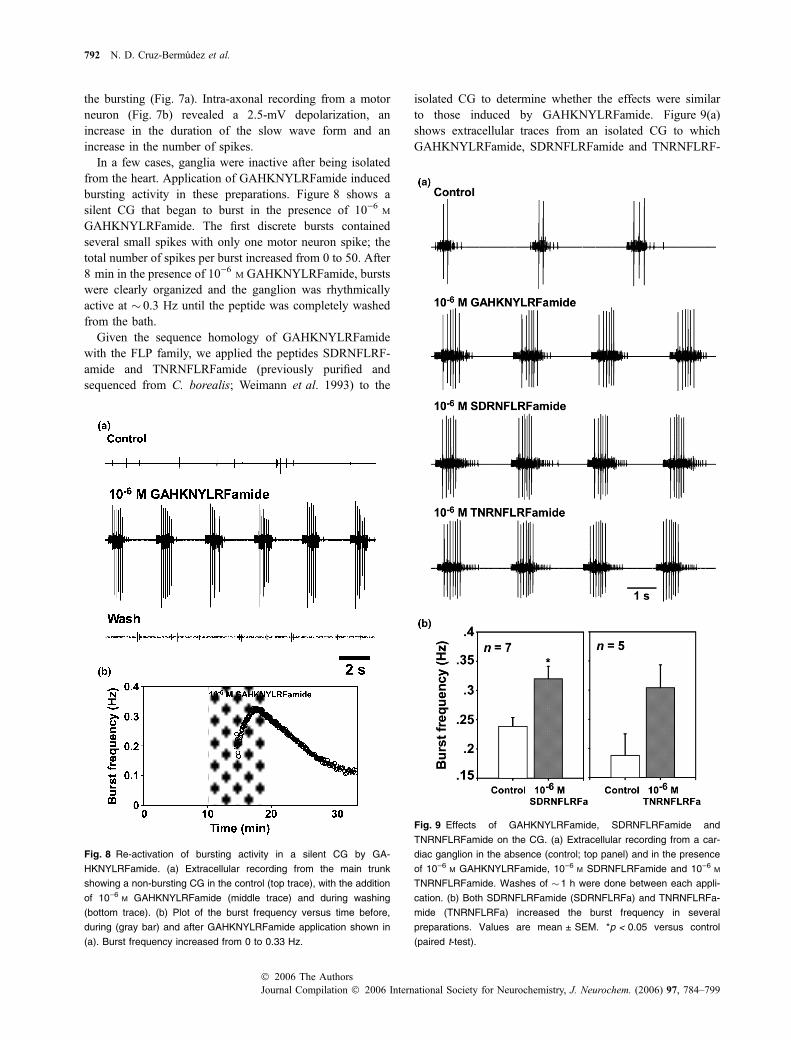
the bursting (Fig. 7a). Intra-axonal recording from a motorneuron (Fig. 7b) revealed a 2.5-mV depolarization, anincrease in the duration of the slow wave form and anincrease in the number of spikes.
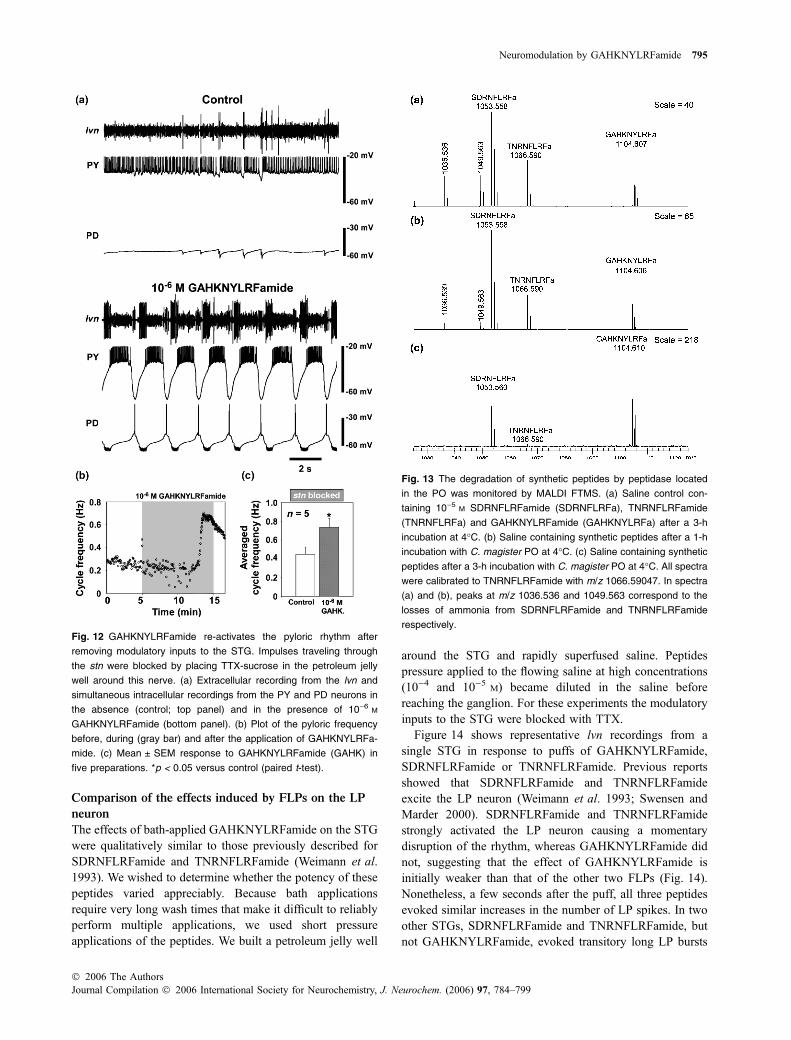
In a few cases, ganglia were inactive after being isolatedfrom the heart. Application of GAHKNYLRFamide inducedbursting activity in these preparations. Figure 8 shows asilent CG that began to burst in the presence of 10)6 M
GAHKNYLRFamide. The first discrete bursts containedseveral small spikes with only one motor neuron spike; thetotal number of spikes per burst increased from 0 to 50. After8 min in the presence of 10)6 M GAHKNYLRFamide, burstswere clearly organized and the ganglion was rhythmicallyactive at � 0.3 Hz until the peptide was completely washedfrom the bath.
Given the sequence homology of GAHKNYLRFamidewith the FLP family, we applied the peptides SDRNFLRF-amide and TNRNFLRFamide (previously purified andsequenced from C. borealis; Weimann et al. 1993) to the
isolated CG to determine whether the effects were similarto those induced by GAHKNYLRFamide. Figure 9(a)shows extracellular traces from an isolated CG to whichGAHKNYLRFamide, SDRNFLRFamide and TNRNFLRF-
Fig. 8 Re-activation of bursting activity in a silent CG by GA-
HKNYLRFamide. (a) Extracellular recording from the main trunk
showing a non-bursting CG in the control (top trace), with the addition
of 10)6 M GAHKNYLRFamide (middle trace) and during washing
(bottom trace). (b) Plot of the burst frequency versus time before,
during (gray bar) and after GAHKNYLRFamide application shown in
(a). Burst frequency increased from 0 to 0.33 Hz.
Fig. 9 Effects of GAHKNYLRFamide, SDRNFLRFamide and
TNRNFLRFamide on the CG. (a) Extracellular recording from a car-
diac ganglion in the absence (control; top panel) and in the presence
of 10)6 M GAHKNYLRFamide, 10)6 M SDRNFLRFamide and 10)6 M
TNRNFLRFamide. Washes of �1 h were done between each appli-
cation. (b) Both SDRNFLRFamide (SDRNFLRFa) and TNRNFLRFa-
mide (TNRNFLRFa) increased the burst frequency in several
preparations. Values are mean ± SEM. *p < 0.05 versus control
(paired t-test).
792 N. D. Cruz-Bermudez et al.
Journal Compilation � 2006 International Society for Neurochemistry, J. Neurochem. (2006) 97, 784–799� 2006 The Authors
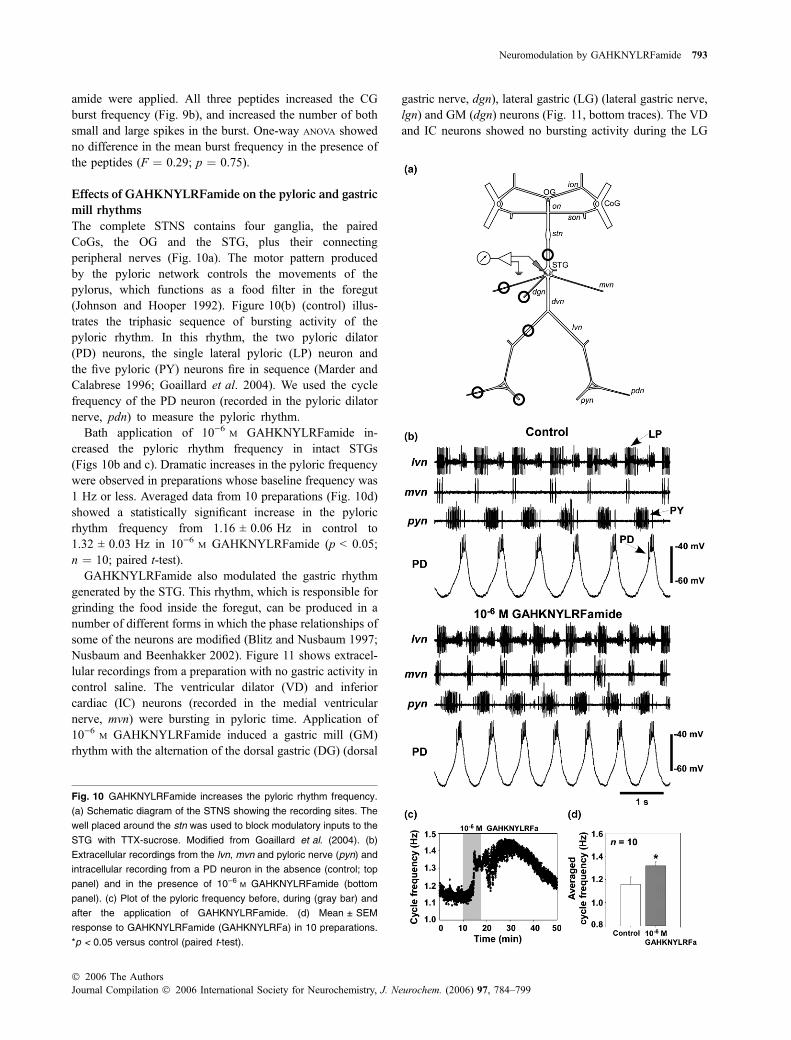
amide were applied. All three peptides increased the CGburst frequency (Fig. 9b), and increased the number of bothsmall and large spikes in the burst. One-way ANOVA showedno difference in the mean burst frequency in the presence ofthe peptides (F ¼ 0.29; p ¼ 0.75).
Effects of GAHKNYLRFamide on the pyloric and gastric
mill rhythms
The complete STNS contains four ganglia, the pairedCoGs, the OG and the STG, plus their connectingperipheral nerves (Fig. 10a). The motor pattern producedby the pyloric network controls the movements of thepylorus, which functions as a food filter in the foregut(Johnson and Hooper 1992). Figure 10(b) (control) illus-trates the triphasic sequence of bursting activity of thepyloric rhythm. In this rhythm, the two pyloric dilator(PD) neurons, the single lateral pyloric (LP) neuron andthe five pyloric (PY) neurons fire in sequence (Marder andCalabrese 1996; Goaillard et al. 2004). We used the cyclefrequency of the PD neuron (recorded in the pyloric dilatornerve, pdn) to measure the pyloric rhythm.
Bath application of 10)6 M GAHKNYLRFamide in-creased the pyloric rhythm frequency in intact STGs(Figs 10b and c). Dramatic increases in the pyloric frequencywere observed in preparations whose baseline frequency was1 Hz or less. Averaged data from 10 preparations (Fig. 10d)showed a statistically significant increase in the pyloricrhythm frequency from 1.16 ± 0.06 Hz in control to1.32 ± 0.03 Hz in 10)6 M GAHKNYLRFamide (p < 0.05;n ¼ 10; paired t-test).
GAHKNYLRFamide also modulated the gastric rhythmgenerated by the STG. This rhythm, which is responsible forgrinding the food inside the foregut, can be produced in anumber of different forms in which the phase relationships ofsome of the neurons are modified (Blitz and Nusbaum 1997;Nusbaum and Beenhakker 2002). Figure 11 shows extracel-lular recordings from a preparation with no gastric activity incontrol saline. The ventricular dilator (VD) and inferiorcardiac (IC) neurons (recorded in the medial ventricularnerve, mvn) were bursting in pyloric time. Application of10)6 M GAHKNYLRFamide induced a gastric mill (GM)rhythm with the alternation of the dorsal gastric (DG) (dorsal
gastric nerve, dgn), lateral gastric (LG) (lateral gastric nerve,lgn) and GM (dgn) neurons (Fig. 11, bottom traces). The VDand IC neurons showed no bursting activity during the LG
Fig. 10 GAHKNYLRFamide increases the pyloric rhythm frequency.
(a) Schematic diagram of the STNS showing the recording sites. The
well placed around the stn was used to block modulatory inputs to the
STG with TTX-sucrose. Modified from Goaillard et al. (2004). (b)
Extracellular recordings from the lvn, mvn and pyloric nerve (pyn) and
intracellular recording from a PD neuron in the absence (control; top
panel) and in the presence of 10)6 M GAHKNYLRFamide (bottom
panel). (c) Plot of the pyloric frequency before, during (gray bar) and
after the application of GAHKNYLRFamide. (d) Mean ± SEM
response to GAHKNYLRFamide (GAHKNYLRFa) in 10 preparations.
*p < 0.05 versus control (paired t-test).
Neuromodulation by GAHKNYLRFamide 793
� 2006 The AuthorsJournal Compilation � 2006 International Society for Neurochemistry, J. Neurochem. (2006) 97, 784–799

and GM bursts. In most preparations, the DG neuron wasstrongly active in the presence of GAHKNYLRFamide and itstopped bursting after the peptide was removed from thebath. Similar gastric activity was induced by the peptidesSDRNFLRFamide and TNRNFLRFamide in C. borealis(Weimann et al. 1993).
Effects of bath-applied GAHKNYLRFamide after
blockade of the modulatory inputs
The pyloric and gastric rhythms depend on modulatoryinputs that project through the stn to the STG. When theactivity in this nerve is blocked using TTX-sucrose, therhythm slows down or completely terminates. Application ofdifferent amines and peptides can re-instate rhythmic pyloricactivity in silent preparations (Marder and Hooper 1985;Marder and Weimann 1992). Application of 10)6 M GA-HKNYLRFamide reinitiated the pyloric rhythm in isolatedSTGs. Figure 12 shows recordings from the lateral ventric-ular nerve (lvn) and intracellularly from the PY and PDneurons. In control conditions (Fig. 12a, top three traces) thePY neuron was depolarized and it was firing tonically. A fewLP spikes were observed in the lvn trace and the PD neuronwas silent. In the presence of GAHKNYLRFamide(Fig. 12a, lower three traces), the LP, PY and PD neuronsre-started a low-frequency triphasic pattern. GAHKNYLRF-amide also increased the pyloric rhythm frequency in slowpreparations after the stn had been blocked (Fig. 12b).Pooled data (Fig. 12c) showed that the pyloric frequencywith the stn blocked increased from 0.45 ± 0.08 Hz incontrols to 0.74 ± 0.09 Hz in 10)6 M GAHKNYLRFamide(p < 0.05; n ¼ 5; paired t-test). The gastric rhythm was alsoactivated in preparations with the stn blocked (data notshown).
Differential degradation of FLPs monitored by MALDI
FTMS
It was shown previously that extracellular peptidase activitycould inactivate neuropeptide function in the crustacean
nervous system as well as fine tune motor pattern modulation(Coleman et al. 1994; Wood and Nusbaum 2002). Giventhat similar physiological effects of GAHKNYLRFamideand the other two previously established FLPs are observedin the CG and STG, we set out to evaluate possibledifferential degradation of different FLPs in the presence ofextracellular peptidases. By monitoring the relative abun-dance of the synthetic peptides by mass spectrometry atdifferent time points, the relative rates of degradation of thepeptides can be compared. The scale of the spectrum isdetermined by the amount of ions being detected, with thelower scale corresponding to more ions during the detectionevent.
Figure 13 shows the degradation of three synthetic FLPsover time when exposed to the peptidases present in tissuedissected from a crab. The saline control (Fig. 13a)showed the relative abundances of the peptides in theabsence of peptidase degradation. SDRNFLRFamide waspresent at the highest abundance, then GAHKNYLRF-amide, closely followed by TNRNFLRFamide. Peakscorresponding to the neutral loss of ammonia were presentfor SDRNFLRFamide and TNRNFLRFamide owing tomultiple asparagine and arginine residues present in thesequences.
Peptide mixtures in saline that had been incubated with POtissue for 1 h (Fig. 13b) showed a decrease in relativeabundance of all the synthetic peptides by an increase in thescale of the spectrum to 65. Furthermore, the abundance ofTNRNFLRFamide relative to GAHKNYLRFamide wasdecreased.
A marked change in total abundance of the ions wasobserved after 3 h incubation of saline containing syntheticpeptides with PO tissue (Fig. 13c), with an overall scale of218. Furthermore, a significant change in the relative abun-dance was noted, with SDRNFLRFamide and TNRNFLRF-amide now at lower abundance than GAHKNYLRFamide,indicating that the latter peptide is more resistant to peptidasedegradation than the other two FLPs.
Fig. 11 Activation of the GM rhythm by
GAHKNYLRFamide. Extracellular record-
ings from the mvn, dgn and lgn nerves in
the absence (control; top panel) and in the
presence of 10)6 M GAHKNYLRFamide
(bottom panel). The DG neuron (largest unit
in the dgn) was consistently activated by
GAHKNYLRFamide (eight of 10 STGs). In
this preparation (ions were cut off), the
rhythm was generated by the alternation of
the DG, LG (large unit in the lgn) and GM
neurons.
794 N. D. Cruz-Bermudez et al.
Journal Compilation � 2006 International Society for Neurochemistry, J. Neurochem. (2006) 97, 784–799� 2006 The Authors
Comparison of the effects induced by FLPs on the LP
neuron
The effects of bath-applied GAHKNYLRFamide on the STGwere qualitatively similar to those previously described forSDRNFLRFamide and TNRNFLRFamide (Weimann et al.1993). We wished to determine whether the potency of thesepeptides varied appreciably. Because bath applicationsrequire very long wash times that make it difficult to reliablyperform multiple applications, we used short pressureapplications of the peptides. We built a petroleum jelly well
around the STG and rapidly superfused saline. Peptidespressure applied to the flowing saline at high concentrations(10)4 and 10)5 M) became diluted in the saline beforereaching the ganglion. For these experiments the modulatoryinputs to the STG were blocked with TTX.
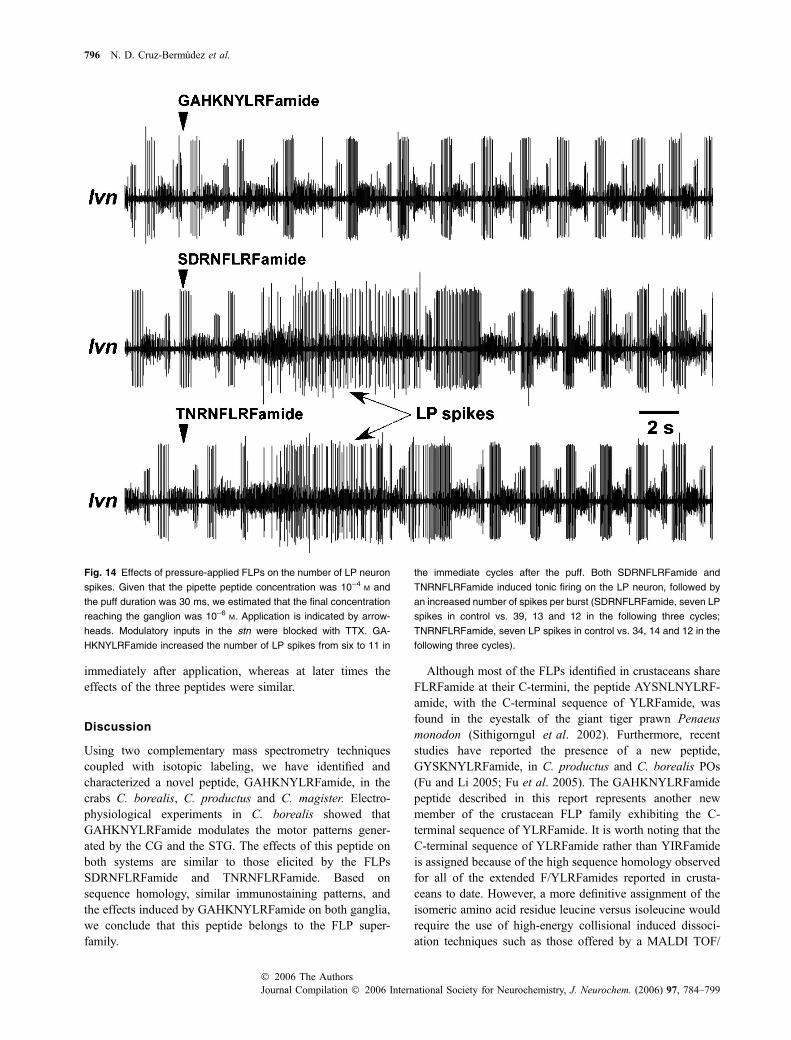
Figure 14 shows representative lvn recordings from asingle STG in response to puffs of GAHKNYLRFamide,SDRNFLRFamide or TNRNFLRFamide. Previous reportsshowed that SDRNFLRFamide and TNRNFLRFamideexcite the LP neuron (Weimann et al. 1993; Swensen andMarder 2000). SDRNFLRFamide and TNRNFLRFamidestrongly activated the LP neuron causing a momentarydisruption of the rhythm, whereas GAHKNYLRFamide didnot, suggesting that the effect of GAHKNYLRFamide isinitially weaker than that of the other two FLPs (Fig. 14).Nonetheless, a few seconds after the puff, all three peptidesevoked similar increases in the number of LP spikes. In twoother STGs, SDRNFLRFamide and TNRNFLRFamide, butnot GAHKNYLRFamide, evoked transitory long LP bursts
Fig. 12 GAHKNYLRFamide re-activates the pyloric rhythm after
removing modulatory inputs to the STG. Impulses traveling through
the stn were blocked by placing TTX-sucrose in the petroleum jelly
well around this nerve. (a) Extracellular recording from the lvn and
simultaneous intracellular recordings from the PY and PD neurons in
the absence (control; top panel) and in the presence of 10)6 M
GAHKNYLRFamide (bottom panel). (b) Plot of the pyloric frequency
before, during (gray bar) and after the application of GAHKNYLRFa-
mide. (c) Mean ± SEM response to GAHKNYLRFamide (GAHK) in
five preparations. *p < 0.05 versus control (paired t-test).
Fig. 13 The degradation of synthetic peptides by peptidase located
in the PO was monitored by MALDI FTMS. (a) Saline control con-
taining 10)5 M SDRNFLRFamide (SDRNFLRFa), TNRNFLRFamide
(TNRNFLRFa) and GAHKNYLRFamide (GAHKNYLRFa) after a 3-h
incubation at 4�C. (b) Saline containing synthetic peptides after a 1-h
incubation with C. magister PO at 4�C. (c) Saline containing synthetic
peptides after a 3-h incubation with C. magister PO at 4�C. All spectrawere calibrated to TNRNFLRFamide with m/z 1066.59047. In spectra
(a) and (b), peaks at m/z 1036.536 and 1049.563 correspond to the
losses of ammonia from SDRNFLRFamide and TNRNFLRFamide
respectively.
Neuromodulation by GAHKNYLRFamide 795
� 2006 The AuthorsJournal Compilation � 2006 International Society for Neurochemistry, J. Neurochem. (2006) 97, 784–799
immediately after application, whereas at later times theeffects of the three peptides were similar.
Discussion
Using two complementary mass spectrometry techniquescoupled with isotopic labeling, we have identified andcharacterized a novel peptide, GAHKNYLRFamide, in thecrabs C. borealis, C. productus and C. magister. Electro-physiological experiments in C. borealis showed thatGAHKNYLRFamide modulates the motor patterns gener-ated by the CG and the STG. The effects of this peptide onboth systems are similar to those elicited by the FLPsSDRNFLRFamide and TNRNFLRFamide. Based onsequence homology, similar immunostaining patterns, andthe effects induced by GAHKNYLRFamide on both ganglia,we conclude that this peptide belongs to the FLP super-family.
Although most of the FLPs identified in crustaceans shareFLRFamide at their C-termini, the peptide AYSNLNYLRF-amide, with the C-terminal sequence of YLRFamide, wasfound in the eyestalk of the giant tiger prawn Penaeusmonodon (Sithigorngul et al. 2002). Furthermore, recentstudies have reported the presence of a new peptide,GYSKNYLRFamide, in C. productus and C. borealis POs(Fu and Li 2005; Fu et al. 2005). The GAHKNYLRFamidepeptide described in this report represents another newmember of the crustacean FLP family exhibiting the C-terminal sequence of YLRFamide. It is worth noting that theC-terminal sequence of YLRFamide rather than YIRFamideis assigned because of the high sequence homology observedfor all of the extended F/YLRFamides reported in crusta-ceans to date. However, a more definitive assignment of theisomeric amino acid residue leucine versus isoleucine wouldrequire the use of high-energy collisional induced dissoci-ation techniques such as those offered by a MALDI TOF/
Fig. 14 Effects of pressure-applied FLPs on the number of LP neuron
spikes. Given that the pipette peptide concentration was 10)4 M and
the puff duration was 30 ms, we estimated that the final concentration
reaching the ganglion was 10)6 M. Application is indicated by arrow-
heads. Modulatory inputs in the stn were blocked with TTX. GA-
HKNYLRFamide increased the number of LP spikes from six to 11 in
the immediate cycles after the puff. Both SDRNFLRFamide and
TNRNFLRFamide induced tonic firing on the LP neuron, followed by
an increased number of spikes per burst (SDRNFLRFamide, seven LP
spikes in control vs. 39, 13 and 12 in the following three cycles;
TNRNFLRFamide, seven LP spikes in control vs. 34, 14 and 12 in the
following three cycles).
796 N. D. Cruz-Bermudez et al.
Journal Compilation � 2006 International Society for Neurochemistry, J. Neurochem. (2006) 97, 784–799� 2006 The Authors

TOF instrument (Nachman et al. 2005). FLPs with YL/IRFamide at the C-terminus have been found mostly innematodes. For example, KHEYLRFamide has been repor-ted in Caenorhabditis elegans, Ascaris suum, Panagrellusredivivus and Haemonchus contortus (Geary et al. 1999). Alarge number of FLPs ending with YLRFamide, includingFKHEYLRFamide, EKHEYLRFamide, KHEEYLRFamide,KSQYIRFamide and QPKARSGYIRFamide, were foundin C. elegans (Geary et al. 1999; Li et al. 1999a, 1999b).Additionally, SDPYLRFamide was found in the pond snailLymnaea stagnalis (Linacre et al. 1990).
For a structurally diverse peptide family such as theFLP family, an interesting question to address is whetherdifferent members of the same family have synergistic orantagonistic physiological effects upon release. Despitestructural similarities between FLPs, individual chemicalisoforms can have distinct and diverse effects. In nematodes,it has been shown that peptides with similar amino acidsequences may have contrasting physiological effects(Fellowes et al. 2000; Davis and Stretton 2001; Bowmanet al. 2002; Kim and Li 2004). Differential expression ofsubsets of a peptide family may be one mechanism usedto subtly manipulate the functional properties of a cell.Therefore, it is essential to unambiguously identify all ofthe chemical isoforms of a given peptide family andcharacterize their physiological actions individually.
Although it is not surprising that the effects of GA-HKNYLRFamide on the STNS and the CG are very similarto those of SDRNFLRFamide and TNRNFLRFamide, theirdifferential degradation by extracellular peptidases mightprovide a potential mechanism for differential actions onneural circuits. Previous studies have documented functionalinactivation of neuropeptides via enzymatic degradation andfine tuning peptidergic modulation through extracellularpeptidase activity (Coleman et al. 1994; Wood and Nusbaum2002). The present peptide degradation experiments providethe first evidence that different members of the FLP familyexhibit differential sensitivity to extracellular peptidaseactivity. The slower degradation rate of GAHKNYLRFamidecompared with TNRNFLRFamide and SDRNFLRFamidemight suggest a long-lasting and hormonal effect in additionto the local modulatory effect. Taken together, these resultssuggest that the effects of GAHKNYLRFamide on the STGmight last longer because it is degraded at a slower rate bypeptidase, although it is a less potent activator of the LPneuron than the other two FLPs studied here. It is possiblethat, depending on the properties of individual neurons in thenetwork and the overall environment of the ganglion, aparticular FLP is more likely to influence the output of thenetwork in a given physiological context.
Studies in the STG have demonstrated that motor circuitscan be massively modulated by neuroactive substances(Marder and Weimann 1992). Many neuropeptides (andamines) can re-activate the pyloric rhythm in quiescent
preparations and induce plateau potentials in some neurons(Hooper and Marder 1984; Weimann et al. 1993, 1997; Liet al. 2003). However, the output of the network is mediatedby the action of each modulator on specific STG neurons(Swensen and Marder 2000, 2001). For instance,TNRNFLRFamide can modulate the LP and PY neurons, buthas no effect on the PD neurons (Swensen and Marder 2001).Are these same neurons equally sensitive to GAHKNYLRF-amide? It has been hypothesized that FLPs are likely to bind tothe same receptor, but that each one has a different affinity forthe receptor (Jorge-Rivera and Marder 1996). Thus, it wouldbe interesting to investigate whether all members of the FLPfamily modulate the same neurons in the STG and whether allthese FLPs activate the same ionic conductance, describedpreviously by Swensen and Marder (2000).
We have shown here that GAHKNYLRFamide has exci-tatory effects on the CG of the species from which it wasisolated. The increased burst frequency, number of spikes perburst and intraburst spike frequency induced by GAHK-NYLRFamide in the CG are similar to the actions induced bySDRNFLRFamide and TNRNFLRFamide in the C. borealisCG. TNRNFLRFamide and SDRNFLRFamide also increaseCG burst frequency in other species of crustacean (Saver et al.1999). These results on isolated ganglia are consistent with thefinding that most FLPs increase the rate of contraction ofisolated hearts in different species (Mercier and Russenes1992; Wilkens and McMahon 1992; McGaw et al. 1995;Worden et al. 1995), which might result from an increase insynaptic transmission from the CG motor neurons and/or theaction of these peptides on the heart muscle.
Recently, it was shown that SDRNFLRFamide directlymodulates the Homarus americanus myocardium by increas-ing intracellular Ca2+ (released internally from the sarco-plasmic reticulum) or through the sarcolemma (Wilkenset al. 2005). Thus, the emerging scenario is that FLPs andpossibly other modulators can act on both the CG and heartmuscle to increase cardiac function in crustaceans. This samesynergetic modulation is known to occur in the stomach,where substances released by modulatory inputs in the STGcan also modulate many muscles of the stomach (Jorge-Rivera and Marder 1996, 1997; Jorge-Rivera 1997; Jorge-Rivera et al. 1998). One hypothesis is that, when the motorpatterns generated by the STG are weak, hormonallydelivered substance can strengthen muscle contractions andensure movement (Jorge-Rivera and Marder 1996). Like-wise, it is possible that, in any particular endogenousphysiological state, slow active hearts increase their outputwhen neuropeptides such as GAHKNYLRFamide or otherFLPs are released by the POs.
Acknowledgements
This work was supported by the National Institute of Neurological
Disorders and Stroke grant NS 17813 (EM), an Individual National
Neuromodulation by GAHKNYLRFamide 797
� 2006 The AuthorsJournal Compilation � 2006 International Society for Neurochemistry, J. Neurochem. (2006) 97, 784–799

Research Service Award (NRSA) award NS 052923 and an
American Psychological Association Predoctoral Fellowship in
Neuroscience (NDC-B), the start-up funds from the School of
Pharmacy and Wisconsin Alumni Research Foundation at the
University of Wisconsin (LL), a research award from the American
Society for Mass Spectrometry (sponsored by Thermo Electron
Corp.) (LL), National Science Foundation CAREER Award (CHE-
0449991) (LL), National Institutes of Health Chemistry–Biology
Interface Training Grant (T32 G008505) (KKK), and a fellowship
from Merck Research Laboratories (QF). AEC acknowledges
financial support from the University of Washington and Friday
Harbor Laboratories. John Edwards, Curtis Easton, Christopher
Goiney and Eahn Holm are thanked for their help with some of the
anatomical experiments.
References
Alexandrowicz J. S. (1932) The innervation of the heart of the crustacea.I. Decapoda. Q. J. Micr. Sci. 75, 181–255.
Blitz D. M. and Nusbaum M. P. (1997) Motor pattern selection viainhibition of parallel pathways. J. Neurosci. 17, 4965–4975.
Bowman J. W., Friedman A. R., Thompson D. P., Maule A. G.,Alexander-Bowman S. J. and Geary T. G. (2002) Structure–activityrelationships of an inhibitory nematode FMRFamide-related pep-tide, SDPNFLRFamide (PF1), on Ascaris suum muscle. Int. J.Parasitol. 32, 1765–1771.
Che F. Y. and Fricker L. D. (2002) Quantitation of neuropeptides inCpe(fat)/Cpe(fat) mice using differential isotopic tags and massspectrometry. Anal. Chem. 74, 3190–3198.
Christie A. E., Skiebe P. and Marder E. (1995) Matrix of neuromodu-lators in neurosecretory structures of the crab, Cancer borealis.J. Exp. Biol. 198, 2431–2439.
Christie A. E., Baldwin D. H., Marder E. and Graubard K. (1997)Organization of the stomatogastric neuropil of the crab, Cancerborealis, as revealed by modulator immunocytochemistry. CellTissue Res. 288, 135–148.
Christie A. E., Edwards J. M., Cherny E., Clason T. A. and Graubard K.(2003) Immunocytochemical evidence for nitric oxide- and carbonmonoxide-producing neurons in the stomatogastric nervous systemof the crayfish Cherax quadricarinatus. J. Comp. Neurol. 467,293–306.
Christie A. E., Cain S. D., Edwards J. M. et al. (2004a) The anteriorcardiac plexus: an intrinsic neurosecretory site within the stoma-togastric nervous system of the crab Cancer productus. J. Exp.Biol. 207, 1163–1182.
Christie A. E., Stein W., Quinlan J. E., Beenhakker M. P., Marder E. andNusbaum M. P. (2004b) Actions of a histaminergic/peptidergicprojection neuron on rhythmic motor patterns in the stomatogastricnervous system of the crab Cancer borealis. J. Comp. Neurol. 469,153–169.
Coleman M. J., Konstant P. H., Rothman B. S. and Nusbaum M. P.(1994) Neuropeptide degradation produces functional inactivationin the crustacean nervous system. J. Neurosci. 14, 6205–6216.
Cooke I. M. (2002) Reliable, responsive pacemaking and pattern gen-eration with minimal cell numbers: the crustacean cardiac gan-glion. Biol. Bull. 202, 108–136.
Cooke I. M. and Hartline D. K. (1975) Neurohormonal alterationof integrative properties of the cardiac ganglion of the lobsterHomarus americanus. J. Exp. Biol. 63, 33–52.
Davis R. E. and Stretton A. O. (2001) Structure–activity relationships of18 endogenous neuropeptides on the motor nervous system of thenematode Ascaris suum. Peptides 22, 7–23.
Fellowes R. A., Maule A. G., Marks N. J., Geary T. G., Thompson D. P.and Halton D. W. (2000) Nematode neuropeptide modulation ofthe vagina vera of Ascaris suum: in vitro effects of PF1, PF2, PF4,AF3 and AF4. Parasitology 120, 79–89.
Fu Q. and Li L. (2005) De novo sequencing of neuropeptides usingreductive isotopic methylation and investigation of ESI QTOF MS/MS fragmentation pattern of neuropeptides with N-terminaldimethylation. Anal. Chem. 77, 7783–7795.
Fu Q., Kutz K. K., Schmidt J. J., Hsu Y.-W. A., Messinger D. I., CainS. D., de la Iglesia H. O., Christie A. E. and Li L. (2005) Hormonecomplement of the Cancer productus sinus gland and pericardialorgan: an anatomical and mass spectrometric investigation.J. Comp. Neurol. 493, 606–625.
Geary T. G., Marks N. J., Maule A. G., Bowman J. W., Alexander-Bowman S. J., Day T. A., Larsen M. J., Kubiak T. M., Davis J. P.and Thompson D. P. (1999) Pharmacology of FMRFamide-relatedpeptides in helminths. Ann. N. Y. Acad. Sci. 897, 212–227.
Goaillard J. M., SchulZ. D. J., Kilman V. L. and Marder E. (2004)Octopamine modulates the axons of modulatory projection neu-rons. J. Neurosci. 24, 7063–7073.
Hooper S. L. and Marder E. (1984) Modulation of a central patterngenerator by two neuropeptides, proctolin and FMRFamide. BrainRes. 305, 186–191.
Huybrechts J., Nusbaum M. P., Bosch L. V., Baggerman G., De Loof A.and Schoofs L. (2003) Neuropeptidomic analysis of the brain andthoracic ganglion from the Jonah crab, Cancer borealis. Biochem.Biophys. Res. Commun 308, 535–544.
Johnson B. and Hooper S. (1992) Overview of the stomatogastric ner-vous system, in Dynamic Biological Networks (Harris-Warrick R.,Marder E., Selverston A. and Moulins M., eds), pp. 1–30. MITPress, Cambridge.
Jorge-Rivera J. C. (1997) Modulation of stomatogastric musculature inthe crab Cancer borealis. PhD Thesis. Neuroscience GraduateProgram, 114.
Jorge-Rivera J. C. and Marder E. (1996) TNRNFLRFamide andSDRNFLRFamide modulate muscles of the stomatogastric systemof the crab Cancer borealis. J. Comp. Physiol. A 179, 741–751.
Jorge-Rivera J. C. and Marder E. (1997) Allatostatin decreases stoma-togastric neuromuscular transmission in the crab, Cancer borealis.J. Exp. Biol. 200, 2937–2946.
Jorge-Rivera J. C., Sen K., Birmingham J. T., Abbott L. F. and Marder E.(1998) Temporal dynamics of convergent modulation at a crusta-cean neuromuscular junction. J. Neurophysiol. 80, 2559–2570.
Kim K. and Li C. (2004) Expression and regulation of an FMRFamide-related neuropeptide gene family in Caenorhabditis elegans.J. Comp. Neurol. 475, 540–550.
Krajniak K. G. (1991) The identification and structure–activity relationsof a cardioactive FMRFamide-related peptide from the blue crabCallinectes sapidus. Peptides 12, 1295–1302.
Kutz K. K., Schmidt J. J. and Li L. (2004) In situ tissue analysis ofneuropeptides by MALDI FTMS in-cell accumulation. Anal.Chem. 76, 5630–5640.
Li C., Kim K. and Nelson L. S. (1999a) FMRFamide-related neuro-peptide gene family in Caenorhabditis elegans. Brain Res. 848,26–34.
Li C., Nelson L. S., Kim K., Nathoo A. and Hart A. C. (1999b) Neu-ropeptide gene families in the nematode Caenorhabditis elegans.Ann. N. Y. Acad. Sci. 897, 239–252.
Li L., Pulver S. R., Kelley W. P., Thirumalai V., Sweedler J. V. andMarder E. (2002) Orcokinin peptides in developing and adultcrustacean stomatogastric nervous systems and pericardial organs.J. Comp. Neurol. 444, 227–244.
Li L., Kelley W. P., Billimoria C. P., Christie A. E., Pulver S. R.,Sweedler J. V. and Marder E. (2003) Mass spectrometric investi-
798 N. D. Cruz-Bermudez et al.
Journal Compilation � 2006 International Society for Neurochemistry, J. Neurochem. (2006) 97, 784–799� 2006 The Authors

gation of the neuropeptide complement and release in the peri-cardial organs of the crab, Cancer borealis. J. Neurochem. 87,642–656.
Linacre A., Kellett E., Saunders S., Bright K., Benjamin P. R. and BurkeJ. F. (1990) Cardioactive neuropeptide Phe-Met-Arg-Phe-NH2
(FMRFamide) and novel related peptides are encoded in multiplecopies by a single gene in the snail Lymnaea stagnalis. J. Neurosci.10, 412–419.
Marder E. and Calabrese R. L. (1996) Principles of rhythmic motorpattern generation. Physiol. Rev. 76, 687–717.
Marder E. and Hooper S. L. (1985) Neurotransmitter modulation of thestomatogastric ganglion of decapod crustaceans, in Model NeuralNetworks and Behavior (Selverston A. I., ed.), pp. 319–337.Plenum Press, New York.
Marder E. and Thirumalai V. (2002) Cellular, synaptic and networkeffects of neuromodulation. Neural Netw. 15, 479–493.
Marder E. and Weimann J. M. (1992) Modulatory control of multipletask processing in the stomatogastric nervous system, in Neuro-biology of Motor Progamme Selection (Kien J., McCrohan C. andWinlow B., eds), pp. 3–19. Pergamon Press, New York.
Marder E., Calabrese R. L., Nusbaum M. P. and Trimmer B. (1987)Distribution and partial characterization of FMRFamide-like pep-tides in the stomatogastric nervous systems of the rock crab,Cancer borealis, and the spiny lobster, Panulirus interruptus.J. Comp. Neurol. 259, 150–163.
McGaw I. J., Wilkens J. L., McMahon B. R. and Airriess C. N. (1995)Crustacean cardioexcitatory peptides may inhibit the heart in vivo.J. Exp. Biol. 198, 2547–2550.
Mercier A. J. and Russenes R. T. (1992) Modulation of crayfish hearts byFMRFamide-related peptides. Biol. Bull. 182, 333–340.
Mercier A. J., Orchard I., TeBrugge V. and Skerrett M. (1993) Isolationof two FMRFamide-related peptides from crayfish pericardialorgans. Peptides 14, 137–143.
Messinger D. I., Kutz K. K., Le T., Verley D. R., Hsu Y. W., Ngo C. T.,Cain S. D., Birmingham J. T., Li L. and Christie A. E. (2005)Identification and characterization of a tachykinin-containingneuroendocrine organ in the commissural ganglion of the crabCancer productus. J. Exp. Biol. 208, 3303–3319.
Nachman R. J., Russell W. K., Coast G. M., Russell D. H. and Predel R.(2005) Mass spectrometric assignment of Leu/Ile in neuropeptidesfrom single neurohemal organ preparations of insects. Peptides 26,2151–2156.
Nusbaum M. P. and Beenhakker M. P. (2002) A small-systems approachto motor pattern generation. Nature 417, 343–350.
Nusbaum M. P., Blitz D. M., Swensen A. M., Wood D. and Marder E.(2001) The roles of co-transmission in neural network modulation.Trends Neurosci. 24, 146–154.
Papayannopoulos I. A. (1995) The interpretation of collision-induceddissociation tandem mass spectra of peptides.Mass. Spectrom. Rev.14, 49–73.
Price D. A. and Greenberg M. J. (1977) Structure of a molluscan car-dioexcitatory neuropeptide. Science 197, 670–671.
Pulver S. R. and Marder E. (2002) Neuromodulatory complement of thepericardial organs in the embryonic lobster, Homarus americanus.J. Comp. Neurol. 451, 79–90.
Saver M. A., Wilkens J. L. and Syed N. I. (1999) In situ and in vitroidentification and characterization of cardiac ganglion neurons inthe crab, Carcinus maenas. J. Neurophysiol. 81, 2964–2976.
Sithigorngul P., Pupuem J., Krungkasem C., Longyant S., Chaivisut-hangkura P., Sithigorngul W. and Petsom A. (2002) Seven novelFMRFamide-like neuropeptide sequences from the eyestalk of thegiant tiger prawn Penaeus monodon. Comp. Biochem. Physiol. BBiochem. Mol. Biol. 131, 325–337.
Skiebe P. (2001) Neuropeptides are ubiquitous chemical mediators:using the stomatogastric nervous system as a model system. J. Exp.Biol. 204, 2035–2048.
Swensen A. M. and Marder E. (2000) Multiple peptides converge toactivate the same voltage-dependent current in a central pattern-generating circuit. J. Neurosci. 20, 6752–6759.
SwensenA.M. andMarder E. (2001)Modulators with convergent cellularactions elicit distinct circuit outputs. J. Neurosci. 21, 4050–4058.
Trimmer B. A., Kobierski L. A. and Kravitz E. A. (1987) Purification andcharacterization of FMRFamide-like immunoreactivity substancesfrom the lobster nervous system: isolation and sequence analysis oftwo closely related peptides. J. Comp. Neurol. 266, 16–26.
Weimann J. M., Marder E., Evans B. and Calabrese R. L. (1993) Theeffects of SDRNFLRFamide and TNRNFLRFamide on the motorpatterns of the stomatogastric ganglion of the crab Cancer borealis.J. Exp. Biol. 181, 1–26.
Weimann J. M., Skiebe P., Heinzel H.-G., Soto C., Kopell N., Jorge-Rivera J. C. and Marder E. (1997) Modulation of oscillator inter-actions in the crab stomatogastric ganglion by crustacean cardio-active peptide. J. Neurosci. 17, 1748–1760.
Wilkens J. L. and McMahon B. R. (1992) Intrinsic properties andextrinsic neurohormonal control of crab cardiac hemodynamics.Experimentia 48, 827–833.
Wilkens J. L., Shinozaki T., Yazawa T. and ter Keurs H. E. (2005) Sitesand modes of action of proctolin and the FLP F2 on lobster cardiacmuscle. J. Exp. Biol. 208, 737–747.
Wood D. E. and Nusbaum M. P. (2002) Extracellular peptidase activitytunes motor pattern modulation. J. Neurosci. 22, 4185–4195.
Worden M. K., Kravitz E. A. and Goy M. F. (1995) Peptide F1, an N-terminally extended analog of FMRFamide, enhances contractileactivity inmultiple target tissues in lobster. J. Exp. Biol. 198, 97–108.
Neuromodulation by GAHKNYLRFamide 799
� 2006 The AuthorsJournal Compilation � 2006 International Society for Neurochemistry, J. Neurochem. (2006) 97, 784–799
Related Documents











