Mandatory Processing of Irrelevant Fearful Face Features in Visual Search Daniela B. Fenker 1 , Dorothee Heipertz 1 , Carsten N. Boehler 2 , Mircea A. Schoenfeld 1,2 , Tömme Noesselt 1 , Hans-Jochen Heinze 1,2 , Emrah Duezel 1,3 , and Jens-Max Hopf 1,2 Abstract ■ Faces expressing fear may attract attention in an automatic bottom–up fashion. Here we address this issue with magneto- encephalographic (MEG) recordings in subjects performing a demanding visual search combined with the presentation of irrelevant neutral or fearful faces. The impact of the facesʼ emo- tional expression on attentional selection was assessed by ana- lyzing the N2pc component—a modulation of the event-related magnetic field response known to reflect attentional focusing in visual search. We observed that lateralized fearful faces elicited an N2pc approximately between 240 and 400 msec in ventral extrastriate cortex that was independent of the N2pc reflecting target selection in visual search. Despite their clear influence on neural processing, fearful faces did not significantly influence behavioral performance. To clarify this discrepancy, we further performed an MEG experiment in which the demands of the search task were reduced. Under those conditions, lateralized fearful faces elicited an N2pc response that was again indepen- dent of the N2pc response to the search target. Behavioral per- formance was, however, influenced in a significant manner, suggesting that for behavioral effects to appear, sufficient atten- tional resources need to be left unoccupied by the search task—a notion put forward by the perceptual load theory. Our observa- tions are taken to indicate that irrelevant fearful faces influence attentional processing in extrastriate visual cortex in an automatic fashion and independent of other task-relevant attentional opera- tions. However, this may not necessarily be echoed at the be- havioral level as long as task-relevant selection operations exhaust attentional resources. ■ INTRODUCTION Given the general ecological importance of facial gestures for human social interactions, it is not surprising that faces expressing emotional states are suspected to cap- ture attention in an automatic way (Dolan & Vuilleumier, 2003; Ohman, Lundqvist, & Esteves, 2001; Vuilleumier & Schwartz, 2001; White, 1995). Although face features per se may not necessarily give rise to attentional capture (Nothdurft, 1993), the emotional valence of faces is sug- gested to be mandatorily registered outside the focus of attention (Lavie, Ro, & Russell, 2003; Vuilleumier, 2002; Morris, Ohman, & Dolan, 1999). In particular, negative emotional valence was observed to speed face identifica- tion in visual search significantly (Eastwood, Smilek, & Merikle, 2001, 2003; Fox et al., 2000; Hansen & Hansen, 1988), which was taken to indicate that negative valence may act like a pop-out feature in guiding attentional focus- ing automatically (e.g., anger superiority effect). However, the results of Hansen and Hansen (1988) obtained with natural face images (Ekman & Friesen, 1978) were chal- lenged by attesting low-level stimulus confounds (Purcell, Stewart, & Skov, 1996) or configurational shortcomings (Cave & Batty, 2006; Horstmann & Bauland, 2006). Purcell et al. (1996), for example, demonstrated that conspicuous black spots were more frequent in threatening than in happy faces, which presumably gave rise to the capture effect. Horstmann and Bauland (2006) suggested that the “anger superiority effect” demonstrated with angry faces embedded in friendly face distractors may rather reflect the fact that observers were more efficient in rejecting happy distractors than in detecting angry target faces. To avoid low-level stimulus confounds, a number of stud- ies used schematic face drawings (Eastwood et al., 2001; Ohman et al., 2001; Fox et al., 2000; White, 1995) to show that negative emotional valence undergoes automatic pri- ority decoding in visual search. These studies, however, provided rather heterogeneous and partially conflicting results and were overall not entirely conclusive. Many of the observations may simply be attributable to lower level configurational confounds and not to the emotional valence of the face drawings (Cave & Batty, 2006; Horstmann & Bauland, 2006). Regarding the automatic capture hypothesis, there are further issues to be considered. In simple search tasks, attentional capture by salient singletons has been shown 1 Otto-von-Guericke University, Magdeburg, Germany, 2 Leibniz Institute for Neurobiology, Magdeburg, Germany, 3 University College London, London, UK © 2010 Massachusetts Institute of Technology Journal of Cognitive Neuroscience 22:12, pp. 2926–2938

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Mandatory Processing of Irrelevant Fearful Face Featuresin Visual Search
Daniela B. Fenker1, Dorothee Heipertz1, Carsten N. Boehler2,Mircea A. Schoenfeld1,2, Tömme Noesselt1, Hans-Jochen Heinze1,2,
Emrah Duezel1,3, and Jens-Max Hopf1,2
Abstract
■ Faces expressing fear may attract attention in an automaticbottom–up fashion. Here we address this issue with magneto-encephalographic (MEG) recordings in subjects performinga demanding visual search combined with the presentation ofirrelevant neutral or fearful faces. The impact of the facesʼ emo-tional expression on attentional selection was assessed by ana-lyzing the N2pc component—a modulation of the event-relatedmagnetic field response known to reflect attentional focusing invisual search. We observed that lateralized fearful faces elicitedan N2pc approximately between 240 and 400 msec in ventralextrastriate cortex that was independent of the N2pc reflectingtarget selection in visual search. Despite their clear influence onneural processing, fearful faces did not significantly influencebehavioral performance. To clarify this discrepancy, we further
performed an MEG experiment in which the demands of thesearch task were reduced. Under those conditions, lateralizedfearful faces elicited an N2pc response that was again indepen-dent of the N2pc response to the search target. Behavioral per-formance was, however, influenced in a significant manner,suggesting that for behavioral effects to appear, sufficient atten-tional resources need to be left unoccupied by the search task—anotion put forward by the perceptual load theory. Our observa-tions are taken to indicate that irrelevant fearful faces influenceattentional processing in extrastriate visual cortex in an automaticfashion and independent of other task-relevant attentional opera-tions. However, this may not necessarily be echoed at the be-havioral level as long as task-relevant selection operations exhaustattentional resources. ■
INTRODUCTION
Given the general ecological importance of facial gesturesfor human social interactions, it is not surprising thatfaces expressing emotional states are suspected to cap-ture attention in an automatic way (Dolan & Vuilleumier,2003; Ohman, Lundqvist, & Esteves, 2001; Vuilleumier &Schwartz, 2001; White, 1995). Although face features perse may not necessarily give rise to attentional capture(Nothdurft, 1993), the emotional valence of faces is sug-gested to be mandatorily registered outside the focus ofattention (Lavie, Ro, & Russell, 2003; Vuilleumier, 2002;Morris, Ohman, & Dolan, 1999). In particular, negativeemotional valence was observed to speed face identifica-tion in visual search significantly (Eastwood, Smilek, &Merikle, 2001, 2003; Fox et al., 2000; Hansen & Hansen,1988), which was taken to indicate that negative valencemay act like a pop-out feature in guiding attentional focus-ing automatically (e.g., anger superiority effect). However,the results of Hansen and Hansen (1988) obtained withnatural face images (Ekman & Friesen, 1978) were chal-
lenged by attesting low-level stimulus confounds (Purcell,Stewart, & Skov, 1996) or configurational shortcomings(Cave & Batty, 2006; Horstmann & Bauland, 2006). Purcellet al. (1996), for example, demonstrated that conspicuousblack spots were more frequent in threatening than inhappy faces, which presumably gave rise to the captureeffect. Horstmann and Bauland (2006) suggested that the“anger superiority effect” demonstrated with angry facesembedded in friendly face distractors may rather reflectthe fact that observers were more efficient in rejectinghappy distractors than in detecting angry target faces.To avoid low-level stimulus confounds, a number of stud-
ies used schematic face drawings (Eastwood et al., 2001;Ohman et al., 2001; Fox et al., 2000; White, 1995) to showthat negative emotional valence undergoes automatic pri-ority decoding in visual search. These studies, however,provided rather heterogeneous and partially conflictingresults and were overall not entirely conclusive. Many ofthe observations may simply be attributable to lower levelconfigurational confounds and not to the emotional valenceof the face drawings (Cave & Batty, 2006; Horstmann &Bauland, 2006).Regarding the automatic capture hypothesis, there are
further issues to be considered. In simple search tasks,attentional capture by salient singletons has been shown
1Otto-von-Guericke University, Magdeburg, Germany, 2LeibnizInstitute for Neurobiology, Magdeburg, Germany, 3UniversityCollege London, London, UK
© 2010 Massachusetts Institute of Technology Journal of Cognitive Neuroscience 22:12, pp. 2926–2938

to be contingent on top–down expectations about thetarget (Yantis & Egeth, 1999; Folk, Remington, & Wright,1994; Yantis, 1993; Folk, Remington, & Johnston, 1992).In other words, attentional capture has been observedwhen the capturing distractorʼs feature was at least partiallytask relevant, but not when the searched-for feature set wascompletely nonoverlapping with the distractor. Hence, toverify true automatic capture, it is mandatory to demon-strate that capture appears for completely task-irrelevantfeatures (Yantis, 1996). As face information has typicallybeen task relevant in most of the abovementioned searchstudies, it remains unclear whether true automatic capturewas involved.Furthermore, for clarifying this issue, it is important to
define the notion of automaticity thoroughly. Automaticityturns out to be a broad concept that has been definedfrom many different perspectives. A recent comprehensiveanalysis (Moors & de Houwer, 2006) reviewed definitionson the basis of notions as diverse as controllability, inten-tionality, goal dependence, consciousness, autonomy, effi-ciency, speed of processing, and more. Hence, it is criticalto clarify what particular notion of automaticity we followhere. In terms of the classification of Moors and de Houwer(2006), the present study refers to automaticity in thesense of demonstrating the presence of neural signaturesof processing face expression despite the fact that thereis neither the goal nor the intention to do so. Automaticityin the present study does not refer to whether the pro-cessing of fear features is efficient or fast or becomes con-scious in any form.Recently, a few ERP studies used task-irrelevant face
stimuli to address the automaticity issue in the sense justsketched (Eimer & Kiss, 2007; Pourtois, Grandjean, Sander,& Vuilleumier, 2004). In these studies, evidence couldbe obtained compatible with the possibility that negativeemotional valence, indeed, captures attention automatically.Finally, recent evidence from fMRI and ERP indicates thatemotional face recognition competes with other attentionaloperations for processing resources (Silvert et al., 2007;Eimer, Holmes, & McGlone, 2003; Holmes, Vuilleumier, &Eimer, 2003; Pessoa, McKenna, Gutierrez, & Ungerleider,2002). According to these observations, emotional face ex-pressionmay not capture attention automatically (Anderson,Christoff, Panitz, De Rosa, & Gabrieli, 2003) when resourcesare exhausted by some other demanding attentional opera-tion (Lavie, 2001; Lavie & Tsal, 1994).Thus, the issue whether the emotional valence of faces
grants attentional priority in a purely automatic fashion(Yantis & Johnson, 1990) is far from being settled. Herewe address this issue with high-temporal resolution MEGrecordings in human observers. In two experiments, wecombined conventional search tasks requiring subjectsto find and discriminate a colored oriented bar amongdistractor bars with a simultaneous presentation of irrele-vant face photographs expressing neutral or negativeemotional valence. Each search frame contained two pho-tographs of the same person, one presented in the left
and one in the right visual field (VF), with the emotionalexpression systematically varied between VFs. That is,a face with a negative expression could appear in theleft VF whereas a neutral expression could appear in rightVF, and vice versa. In addition, faces in both VFs coulddisplay a negative or a neutral expression. To avoid low-level stimulus confounds (Hansen & Hansen, 1988), wematched the face photographs in luminance and contrastand band-pass filtered it with a spatial Fourier filter (seeMethods), which effectively eliminated conspicuous black“mouth spots” that may potentially capture attention in-dependent of emotional valence (Purcell et al., 1996). Asa critical experimental manipulation, the location of nega-tive valence was varied independently of the location of thesearch target, which permitted to analyze the time courseof activity modulations because of the lateralized valencewith respect to the time course of activity reflecting atten-tional focusing onto the search target.
Neural activity reflecting attentional focusing was as-sessed by analyzing the magnetic analog of the so-calledN2pc component of the ERP response. The N2pc—a rel-ative negativity over the occipital scalp contralateral tothe target item between 180 and 350 msec after searchframe onset—has been first described in the ERP re-sponse by Luck and Hillyard (1994a, 1994b) and was sub-sequently characterized in the neuromagnetic response(Hopf, Boelmans, Schoenfeld, Luck, & Heinze, 2004; Hopf,Boelmans, Schoenfeld, Heinze, & Luck, 2002; Hopf et al.,2000). The N2pc reflects the operation of focusing atten-tion onto the target in visual search and was particularlyshown to permit tracking of multiple and rapid shifts ofattention in real time (Woodman & Luck, 1999, 2003b).
METHODS
Participants
Twelve healthy students (9 women, age = 18–27 years)of the Otto-von-Guericke University, Magdeburg, tookpart in Experiment 1. Eight of the students participatingin Experiment 1 and four additional students took part inExperiment 2. All subjects were right-handed, had nor-mal color vision, and had normal or corrected-to-normalvisual acuity. Subjects gave written informed consent andwere paid for participation. The study was approved bythe Ethics Committee of the Otto-von-Guericke University,Magdeburg.
Stimuli
Experiment 1
The stimulus setup is illustrated in Figure 1A–D. Eachsearch frame contained two orthogonally oriented double-colored bars in each VF superimposed onto two face photo-graphs of the same person. Color was randomly assignedto the VFs, with the bars in one VF both being either
Fenker et al. 2927

red-green or blue-yellow. The same color combinationnever appeared in both VF. The bars in each VF were pre-sentedwithin a 1°×1.5° (visual angle) region 8.5° away fromthe vertical and 3.5° below the horizontal meridian. At thestart of each trial block, one color combination was desig-nated as being the target (e.g., red/green), and subjectssearched for the vertical bar drawn in that combination.The adjacent horizontal bar was irrelevant and served as asearch distractor. The relative distribution of color withinthe bars was random, and subjects had to indicate with analternative button press whether one color of the targetcolor combination (e.g., red) appeared in the upper or lowerhalf of the vertical bar. Target color was fixed withinbut varied across different trial blocks, resulting in twopossible target definitions (vertical/green-red and vertical/blue-yellow). The presentation of a target color combinationin one VF was always accompanied by a different colorcombination in the opposite VF. This permitted to directthe focus of attention to the left or right VF by presentingidentical search frames but simply instructing subjects toattend alternative color combinations. The important ad-vantage of this approach is that low-level stimulus con-founds due to color lateralization are eliminated whencomparing the event-related magnetic field (ERMF) re-sponse elicited by left and right VF targets, thereby leavingthe purely attention-related response.
In addition to the search items, two face photographswere presented, one in the left and one in the right VF.Face photographs were taken from a subset of a hun-dred fearful and neutral faces of the Ekman and Friesen(1978) and Gur et al. (2002) collection. They subtended8.6°/10.9° (width/height) of visual angle and were pre-
sented in a region centered 7.6° away from the verticaland 3.5° below the horizontal meridian. Although thephotographs always showed the same personʼs face inboth VFs, the emotional expression was varied system-atically between VFs relative to the VF where the targetappeared. That is, in 25% of the trials, the face wouldhave a negative expression in the target VF (right VF inFigure 1B) but a neutral one in the opposite nontargetVF. In another 25% of the trials, a neutral expression inthe target VF would be combined with a negative expres-sion in opposite VF (Figure 1C). In yet another 25%, anegative emotional expression would appear in boththe target and the nontarget VF (Figure 1D), and in theremaining 25% of the trials, a neutral expression wouldappear bilaterally (Figure 1A). This permitted to assessthe effect of a lateralized emotional face expression ontothe ERMF response relative to the ERMF response re-flecting attentional focusing onto the search target (target-related N2pc). Although the bar stimuli were superimposedonto the task-irrelevant faces, they always appeared inthe buccal region without covering critical features of thefaces. To eliminate low-level stimulus confounds, we con-verted all face photographs to grayscale, matched them insize, luminance, and contrast and low-pass filtered themwith a two-dimensional fast Fourier filter (second-orderButterworth window, six cycles per image). This transfor-mation particularly eliminates the conspicuousness ofmouth spots in fearful faces, which was a critical confoundin other studies (as outlined by Purcell et al., 1996). Notethat because the fearful expression of faces is mainly con-tained in the low spatial frequency content of face photo-graphs (Vuilleumier, Armony, Driver, & Dolan, 2003),
Figure 1. (A–D) Examplesof search displays used inExperiment 1. In each VF,two orthogonally orientedbars were superimposed ona face photograph, withthe coloration of the bars(red-green, blue-yellow)differing between VFs. Onecolor combination definedthe target in a given trial block(e.g., red-green). Subjectssearched for and discriminatedthe vertical bar drawn inthat color combination. Theemotional expression of theface photographs (neutral,fearful) varied systematicallybetween VFs, such that theexpression of both facescould be (A) neutral or(D) fearful or the face in oneVF could be (B and C) fearfulwith the face in the oppositeVF being neutral. (E) Examplesearch display used in theExperiment 2.
2928 Journal of Cognitive Neuroscience Volume 22, Number 12

low-pass filtering of faces preserved this information. Allstimuli were presented on a gray background.
Experiment 2
The general stimulus setup is illustrated in Figure 1E. Thesetup was identical to Experiment 1, except for changingthe coloration of the search items. Search frames nowcontained only uniformly colored bars, with one redand one green bar presented in opposite VFs togetherwith two orthogonally oriented blue distractor bars(one in each VF). The number and the size of the coloredbars were identical to Experiment 1. The orientation ofthe bars changed randomly from trial to trial. The sub-jectsʼ task was to discriminate the orientation of the bardrawn in the target color, which was designated at thestart of each trial block. The spatial distribution of fearfuland neutral faces was identical to Experiment 1.
Procedure
Experiment 1
At the beginning of each trial block, an instruction frameinformed participants about the target color. Each trialblock required subjects to fixate the center of the screen(white fixation dot) and to perform a sequence of 64 searchframes, with each frame presented for 400 msec in a se-quence with randomized ISIs between 1300 and 1500msec.Participants indicated via button press with the right handwhether the target color appeared in the upper (indexfinger) or lower part (middle finger) of the vertical bars.Subjects performed on eight trial blocks, such that twopossible combinations of the target color (red and blue),were searched for four times. This resulted in a total num-ber of 128 trials per condition (2 VFs × 4 distributions ofemotional valence).
Experiment 2
The experimental procedure was identical to Experiment 1,except the discrimination task. Subjects were required toreport the orientation (horizontal/vertical) of the bar drawnin the target color again with a two-alternative button pressof the right index/middle finger. Experiment 1 requiredfocusing onto subparts of the target bar and determin-ing the color distribution, the experimental task now re-quired a simple orientation discrimination of the wholetarget bar that made the discrimination task perceptuallyeasier. In terms of perceptual load theory (PLT; Lavie, Hirst,de Fockert, & Viding, 2004), perceptual load was reducedby making the perceptual task less demanding but with-out changing the number of distractors. Importantly, thischange of the discrimination task did not change top–down executive demands. Target color was randomlyassigned from trial to trial. Trial structure was identical toExperiment 1. As in Experiment 1, subjects performed
eight trial blocks amounting to a total of 128 trials percondition.
Data Recording and Analysis
The MEG and the EOG were registered simultaneouslyusing a 148-channel Bti Magnes 2500 whole-head magneto-meter (Biomagnetic Technologies Inc., San Diego, CA) anda Synamps amplifier (NeuroScan Inc., Herndon, VA). Thesignals were digitized at a rate of 254 Hz and band-passfiltered from 0.1 to 50 Hz. The horizontal and the verticalEOG were recorded bipolarly, using two electrodes be-hind the lateral orbital angles for the horizontal EOGwhereas the vertical EOG was recorded from an electrodebelow the right orbital ridge and one above the right eye.Impedances were kept less than 5 kΩ, and an electrodeplaced at FPZ served as ground. MEG signals were sub-mitted to on-line and off-line noise reduction (Robinson,1989). Epochs containing artifacts were rejected when ex-ceeding a peak-to-peak threshold of 3 pT for the MEG and100 μV for the EOG signal.
For further data analysis, ERMF waveforms were aver-aged for each experimental condition and target VF, weretime locked to search frame onset, and averaged relative toan immediately preceding baseline interval of 100 msec.For statistical validation, we quantified data as the meanmagnetic response in selected time windows and thensubjected to repeated measures ANOVAs; nonsphericitywas corrected with the Greenhouse–Geisser epsilon whennecessary. To determine the onset latency of ERMF wave-form differences, we computed t tests on the basis of meanamplitude measures in a short time window (30 msec) thatwas moved sample by sample between 150 and 450 msecafter stimulus onset. The first significant t test ( p< .05) in asequence of three or more significant consecutive t testswas taken to mark the onset of the waveform difference(Guthrie & Buchwald, 1991).
Current Source Localization
Current source localization was constrained by realisticanatomical models of volume conductor and source com-partment. To this end, anatomical and functional datahad to be in precise register, which was achieved in thefollowing way: Anatomical landmarks (left and right pre-auricular points, nasion) were digitized using a Polhemus3Space Fastrak system (Polhemus Inc., Colchester, VT)and then referenced to the sensor coordinate system.In addition, five spatially distributed coils attached tothe head were digitized, which generated magnetic fieldmarkers permitting to coregister the functional data. Cur-rent source localization was based on minimum normleast squares (MNLS) estimates as implemented in themultimodal neuroimaging software Curry 5.08 (NeuroscanInc.). MNLS represents a distributed source model (Fuchs,Wagner, Kohler, & Wischmann, 1999; Hamalainen &
Fenker et al. 2929
Ilmoniemi, 1994) that yields the distribution of currentsover the cortical surface. Realistic anatomical models ofthe source compartment and the volume conductor wereobtained by three-dimensional segmentations of the head,the cerebrospinal fluid space, and the cortical surface ofthe brain from theMontreal Neurological Institute (averageof 152 T1-weighted volumes from the ICBM project, seewww.bic.mni.mcgill.ca/cgi/icbm_view/) using the boundaryelement method (Hämäläinen & Sarvas, 1989).
RESULTS
Experiment 1
Behavioral Data
The effect of the spatial distribution of fearful face expres-sion on response time and response accuracy was analyzedwith two-way repeated measures ANOVAs with the factorsCondition (bilateral neutral, fearful face at target side, fear-ful face at nontarget side, and bilateral fearful) and TargetVF (left and right). This analysis yielded no significant maineffect of condition, neither regarding the observersʼ aver-age response time, F(3, 33) = 1.39, p > .25, nor regardingtheir response accuracy, F(3, 33) = 1.08, p > .35. Subjectsshowed almost identical response times and accuracy mea-sures in the left VF (RT[msec]/correct[%]: bilateral neu-tral = 647 msec/93%; bilateral fearful = 643 msec/96%;fearful face at target side = 645 msec/94%; fearful face atnontarget side = 646 msec/95%). Subjects were some-what faster in the right VF when fearful faces were absent
(697 msec) or in the nontarget VF (698 msec) as comparedwith when fearful faces were presented at the target side(707 msec) or bilaterally (708 msec). However, the Con-dition × Target VF interaction did not reach significance,F(3, 33) = 2.47, p = .10. Accuracy was almost identical inthe right VF (bilateral neutral = 88%, bilateral fearful =87%, fearful face at target side = 87%, and fearful face atnontarget side=88%), and theCondition×Target VF inter-action was not significant, F(3, 33) = 2.47, p = .10. Therewas, however, a significant main effect of target VF on re-sponse time, F(1, 11) = 16.79, p < .01, and accuracy, F(1,11) = 19.65, p< .01, reflecting the fact that responses weregenerally faster and more accurate for targets presented inthe left VF.
MEG Data
Effect of target lateralization. To assess neural activityreflecting attentional focusing onto the target, we analyzedthe N2pc—a component that is derived as the differ-ence between ERMF responses elicited by left and rightVF targets (target-related N2pc). We first focused on searchframes with the same emotional face expression in bothVFs (bilateral neutral and bilateral fearful) because thisprovides a clean measure of the target-related N2pc, whichis unbiased by potential-lateralized responses because ofpresentation of a fearful face in only one VF. Figure 2Ashows waveforms elicited by search targets in the left (solidlines) and right (dashed lines) VFs together with a fielddistribution map of the left-minus-right ERMF difference
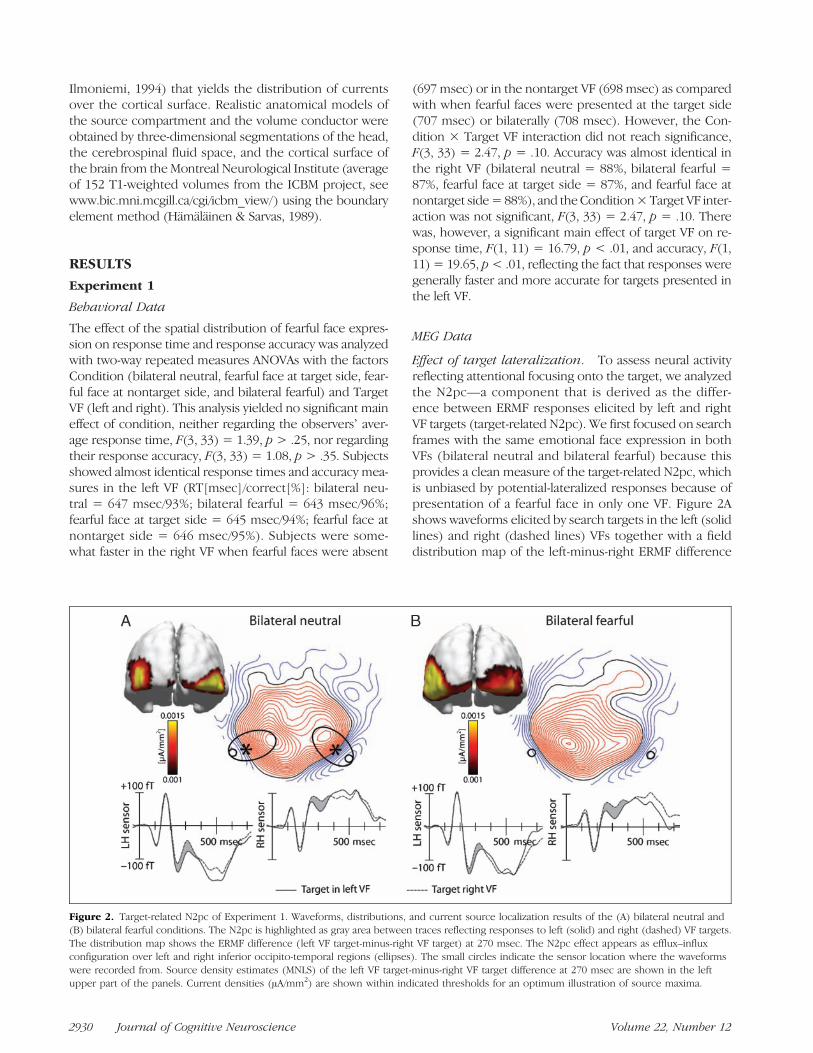
Figure 2. Target-related N2pc of Experiment 1. Waveforms, distributions, and current source localization results of the (A) bilateral neutral and(B) bilateral fearful conditions. The N2pc is highlighted as gray area between traces reflecting responses to left (solid) and right (dashed) VF targets.The distribution map shows the ERMF difference (left VF target-minus-right VF target) at 270 msec. The N2pc effect appears as efflux–influxconfiguration over left and right inferior occipito-temporal regions (ellipses). The small circles indicate the sensor location where the waveformswere recorded from. Source density estimates (MNLS) of the left VF target-minus-right VF target difference at 270 msec are shown in the leftupper part of the panels. Current densities (μA/mm2) are shown within indicated thresholds for an optimum illustration of source maxima.
2930 Journal of Cognitive Neuroscience Volume 22, Number 12

(at 270 msec) of the bilateral neutral condition. Waveformsare shown from selected sensors over the left and rightoccipito-temporal cortex (circles), which reveal a clearN2pc effect. The N2pc is visible as the waveform differ-ence between traces (gray area) approximately from 220to 380 msec after search frame onset. It should be notedthat in contrast to the N2pc of the ERP, the magnetic ana-log of the N2pc does not appear as activity modulation con-tralateral to the target item. Instead, it appears as similarpolarity modulation at occipito-temporal sensors in theleft and right hemispheres when responses elicited by leftand right VF targets are compared. Specifically, a left VF tar-get elicits a negativity (downward deflection in Figure 2A)at both left and right occipito-temporal sensors relative toa right VF target. For simplicity, we will keep the term N2pcto refer to the magnetic analog of this component. In thefield distribution map, the N2pc appears as efflux–influxconfiguration over left and right inferior occipito-temporalregions (encircled by ellipses), with red lines reflectingthe efflux part and blue lines the influx part. As detailedpreviously (Hopf, Vogel, Woodman, Heinze, & Luck, 2002;Hopf et al., 2000), the current origin underlying such fieldconfiguration is predicted to arise from underneath thetransition zone between the efflux and the influx com-ponents (asterisks). Current source density estimates ofthe left-minus-right ERMF difference confirm this predictionin showing current maxima in the left and right inferioroccipito-temporal cortex (upper left insert). Figure 2B illus-trates the results for the bilateral fearful condition. As inthe bilateral neutral condition, a clear N2pc effect is visible.Importantly, the waveforms as well as the field distribu-tion of the left-minus-right ERMF difference show an N2pceffect that is very similar to the neutral condition. Sourcedensity estimates of the left-minus-right ERMF difference re-veal current sourcemaxima in left and right ventral occipito-temporal cortex.For statistical validation, we computed a repeated
measures ANOVA with the factors Target VF (left/right),Sensor Hemisphere (left/right), and Condition (bilateralneutral/bilateral fearful) on the basis of mean amplitudesbetween 250 and 350 msec. This revealed a significanteffect of Target VF, F(1, 11) = 21.7, p < .001, which vali-dates the presence of the N2pc. There was no main effectof Condition, F(1, 11) = 0.18, p > .65, and no TargetVF × Condition interaction, F(1, 11) = 0.16, p > .7, indi-cating that the N2pc amplitude did not significantly dif-fer between conditions. Waveforms generally differedbetween left and right hemisphere sensors, which werereflected by a significant effect of Sensor Hemisphere,F(1, 11) = 10.7, p < .01.In sum, the field distribution and the source localization
results indicate that visual search in the control and thebilateral fearful condition produced a typical N2pc effectapproximately between 220 and 380 msec that is perfectlyin line with our previous characterization of the N2pcin the magnetic response (Hopf, Vogel, et al., 2002; Hopfet al., 2000). Notably, there was no difference when com-
paring the N2pc responses elicited by the control and thebilateral fearful condition.
The effect of lateralized fearful faces. To analyze theeffect of irrelevant fearful faces on attentional focusing,we considered search frames with a fearful face expres-sion only in the left or right VF. Specifically, the magneticresponse elicited by a fearful face in the left VF (Figure 1C)was compared with the response elicited by a fearful facein the right VF (Figure 1B), when both times the targetwas presented in the right VF. Analogously, the responseto a fearful face in the left and right VFs was comparedwhen the search target appeared in the left VF. Ideally,those comparisons equal attention effects related to focus-ing onto the search target but leave effects related to thelateralized fearful faces. Figure 3 summarizes the resultsseparately for targets in the left (Figure 3A) and right (Fig-ure 3B) VFs. For both target VFs, we obtained a clear mod-ulation of the ERMF response that starts around 240 msec.Although smaller in amplitude, this modulation is similarin time course, polarity, and field distribution (left-minus-right difference) to the N2pc seen when comparing leftversus right VF targets (cf. Figure 2). Furthermore, sourcedensity estimates of the left-minus-right ERMF differencereveal current maxima in ventral occipito-temporal cortexregions typical for the N2pc. Taken together, those resultsindicate that fearful faces, in fact, elicit an N2pc effect ontheir own, which is independent of the neural processesmediating the focusing of attention onto the search target.
To validate the N2pc to the lateralized fearful facesstatistically, we computed an overall repeated measuresANOVA with the factors Face Lateralization (fearful faceleft and fearful face right), Sensor Hemisphere (left andright), and Target VF (left and right) on the mean ERMF re-sponse between 250 and 350 msec. This analysis revealeda significant main effect of Face Lateralization, F(1, 11) =16.9, p < .005, indicating the presence of an N2pc due tothe fearful faces. There was also a significant effect of Tar-get VF, F(1, 11) = 6.2, p < .05, reflecting the presence ofan N2pc to the lateralized targets. There was, however,no interaction between Face Lateralization × Target VF,F(1, 11) = 1.9, p > .2, which indicates that the size of theN2pc effect to fearful faces was independent of the localiza-tion of the search target.
Given that irrelevant fearful faces elicited an N2pc re-sponse that is independent of the N2pc response to thesearch targets, it would be informative to determine therelative time course of both N2pc responses. To this end,we computed sample-wise sliding t tests (see Methods)comparing ERMF waveforms elicited by left versus rightVF targets and by left versus right VF fearful faces be-tween 150 and 450 msec. Figure 3C summarizes respectiveresults. Time ranges of significant waveform differences( p < .05) reflecting the N2pc are highlighted by horizon-tal bars. The analysis was based on waveforms collapsedacross left and right hemisphere sensors. Apparently, theN2pc to fearful faces appears in the same time range as
Fenker et al. 2931
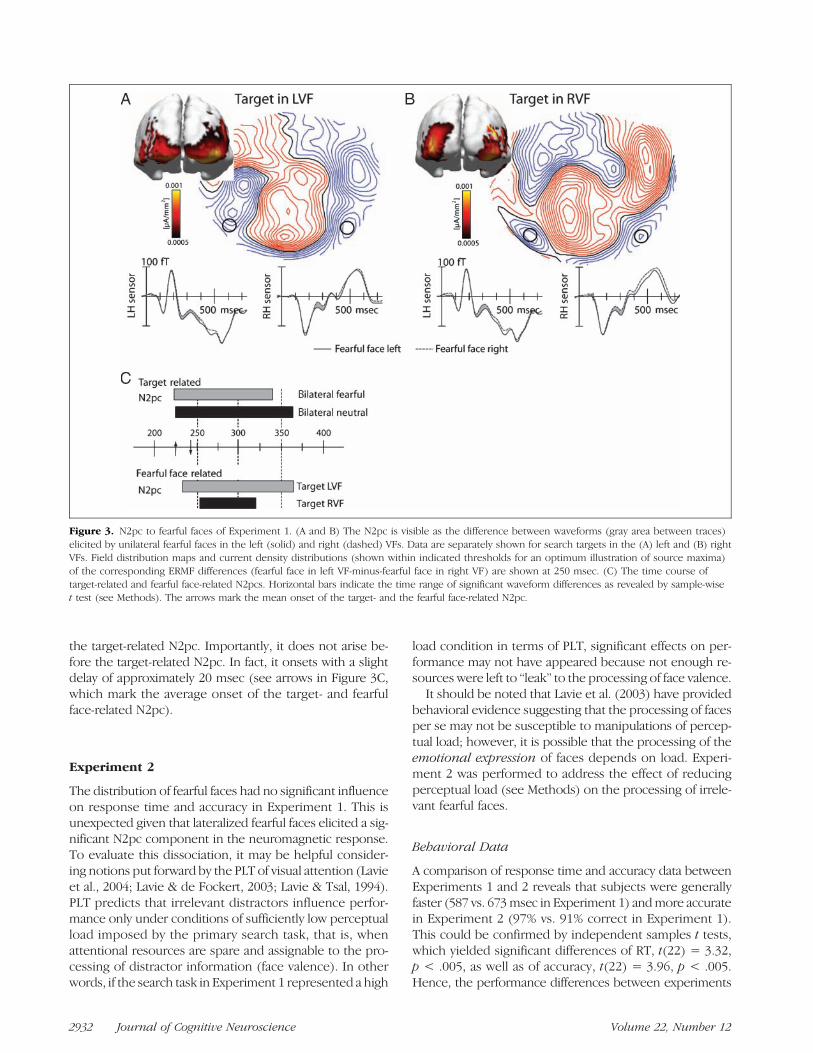
the target-related N2pc. Importantly, it does not arise be-fore the target-related N2pc. In fact, it onsets with a slightdelay of approximately 20 msec (see arrows in Figure 3C,which mark the average onset of the target- and fearfulface-related N2pc).
Experiment 2
The distribution of fearful faces had no significant influenceon response time and accuracy in Experiment 1. This isunexpected given that lateralized fearful faces elicited a sig-nificant N2pc component in the neuromagnetic response.To evaluate this dissociation, it may be helpful consider-ing notions put forward by the PLT of visual attention (Lavieet al., 2004; Lavie & de Fockert, 2003; Lavie & Tsal, 1994).PLT predicts that irrelevant distractors influence perfor-mance only under conditions of sufficiently low perceptualload imposed by the primary search task, that is, whenattentional resources are spare and assignable to the pro-cessing of distractor information (face valence). In otherwords, if the search task in Experiment 1 represented a high
load condition in terms of PLT, significant effects on per-formance may not have appeared because not enough re-sources were left to “leak” to the processing of face valence.It should be noted that Lavie et al. (2003) have provided
behavioral evidence suggesting that the processing of facesper se may not be susceptible to manipulations of percep-tual load; however, it is possible that the processing of theemotional expression of faces depends on load. Experi-ment 2 was performed to address the effect of reducingperceptual load (see Methods) on the processing of irrele-vant fearful faces.
Behavioral Data
A comparison of response time and accuracy data betweenExperiments 1 and 2 reveals that subjects were generallyfaster (587 vs. 673msec in Experiment 1) andmore accuratein Experiment 2 (97% vs. 91% correct in Experiment 1).This could be confirmed by independent samples t tests,which yielded significant differences of RT, t(22) = 3.32,p < .005, as well as of accuracy, t(22) = 3.96, p < .005.Hence, the performance differences between experiments
Figure 3. N2pc to fearful faces of Experiment 1. (A and B) The N2pc is visible as the difference between waveforms (gray area between traces)elicited by unilateral fearful faces in the left (solid) and right (dashed) VFs. Data are separately shown for search targets in the (A) left and (B) rightVFs. Field distribution maps and current density distributions (shown within indicated thresholds for an optimum illustration of source maxima)of the corresponding ERMF differences (fearful face in left VF-minus-fearful face in right VF) are shown at 250 msec. (C) The time course oftarget-related and fearful face-related N2pcs. Horizontal bars indicate the time range of significant waveform differences as revealed by sample-wiset test (see Methods). The arrows mark the mean onset of the target- and the fearful face-related N2pc.
2932 Journal of Cognitive Neuroscience Volume 22, Number 12

validate our experimental manipulation in aiming at lower-ing the perceptual demands of the discrimination task. Im-portantly, a repeated measures ANOVA with the factorsCondition (bilateral neutral, fearful face at target side, fearfulface at nontarget side, and bilateral fearful) and Target VF(left and right) revealed that the distribution of fearful facessignificantly influenced response time, F(3, 33) = 5.62, p<.05, in Experiment 2. The Condition × Target VF interac-tion was not significant, F(3, 33) = 0.84, p > .35. A sub-sequent post hoc analysis revealed that the effect onresponse time was mainly due to the fact that subjects weresignificantly slower when fearful faces appeared in the targetVF (594 msec) as opposed to the nontarget VF (580 msec)(t = 3.23, p < .01). Response times for the bilateral fearfuland bilateral neutral conditions were both 587 msec. Re-sponse time was generally faster in the right VF than inthe left VF, which is reflected by a significant main effectof Target VF, F(1, 11)=9.14, p< .01. In contrast to responsetime, accuracy was completely uninfluenced by the distri-bution of fearful faces (bilateral neutral = 97.0%, bilateralfearful = 97.2%, fearful face target side = 97.3%, fearful facenontarget side = 96.9%). Neither the factor Condition, F(3,9) = 0.37, p > .65, nor the Condition × Target VF inter-action, F(3, 33) = 2.43, p > .15, was significant. There wasalso no main effect of VF, F(1, 11) = 0.08, p > .75. What isnotable is that although subjects were generally faster inresponding to left VF targets in Experiment 1, they werenow somewhat faster in the right VF. This difference isunexpected but may reflect qualitative differences in per-ceptual task. Experiment 2 required a simple orientation dis-crimination, whereas Experiment 1 required to discriminatethe relative position of color within a bar. The latter task isequivalent to discriminating spatial phase, and spatial phasediscrimination has been shown to be better in the left thanin the right VF (Fiorentini & Berardi, 1984). It is possible thatthe change of VF asymmetries between experiments relatesto this difference of perceptual tasks.Taken together, Experiment 2 revealed thatwhen theper-
ceptual demands of target discrimination are reduced incomparison to Experiment 1, significant performance ef-fects on response time appear. This is consistent with thepossibility that the task requirements of Experiment 2 leavemore attentional resources unoccupied that could spill tothe processing of irrelevant fearful faces.
MEG Data
Figure 4 summarizes the MEG results. Figure 4A showsthe target-related N2pc effect, that is, waveforms elicitedby targets in the left and right VFs of the bilateral neutral(left) and bilateral fearful (right) conditions. The N2pc ef-fect is visible as waveform difference (gray area betweentraces) approximately between 240 and 370 msec. A com-parison with corresponding waveforms of Experiment 1(Figure 2A and B) reveals that the effects are rather similarboth in terms of the size of the N2pc as well as its timecourse. Moreover, current source density estimates of the
N2pc revealed similar current maxima in the left and rightinferior occipito-temporal cortex. As in Experiment 1, sta-tistical validation of the N2pc effect was performed onthe basis of mean amplitude measures between 250 and350 msec. A repeated measures ANOVA with the factorsTarget VF (left/right), Sensor Hemisphere (left/right), andCondition (bilateral neutral/bilateral fearful) revealed a sig-nificant effect of Target VF, F(1, 11) = 11.2, p< .01, whichvalidates the presence of the N2pc. There was no maineffect of Condition, F(1, 11) = 0.21, p> .65, and no TargetVF × Condition interaction, F(1, 11) = 1.77, p > .20,indicating that the N2pc amplitude did not significantlydiffer between conditions. Waveforms generally differedbetween left and right hemisphere sensors, which werereflected by a significant effect of Sensor Hemisphere,F(1, 11) = 5.75, p < .05.
The response to lateralized fearful faces is shown in Fig-ure 4B. As in Experiment 1, the lateralized presentation offearful faces produces an N2pc response that is indepen-dent of the location of the search target. Again, the effectis generally smaller than the N2pc to the search targets, butit appears consistently and in roughly the same time range.Current source density estimates reveal current maximain ventral occipito-temporal cortex, typical for the N2pcand corresponding with the maxima observed in Experi-ment 1. For statistical validation, we computed an overallrepeated measures ANOVA with the factors Face Lateraliza-tion (fearful face left and fearful face right), Sensor Hemi-sphere (left and right), and Target VF (left and right) on themean ERMF response between 250 and 350 msec. Thisanalysis revealed a significant main effect of Face Laterali-zation, F(1, 11) = 7.86, p < .02, confirming the presenceof an N2pc due to the fearful faces. The factor Target VFwas also significant, F(1, 11) = 6.47, p < .05, reflectingthe presence of an N2pc to the lateralized targets. As inExperiment 1, no interaction between face lateralizationand target VF was observed, F(1, 11) = 0.06, p > .8, indi-cating that the N2pc to fearful faces was independent of thelocation of the search target.
We finally determined the time course of the N2pc re-sponses to the target and to the lateralized fearful faceson the basis of sample-wise sliding t tests (see Methods)analogous to Experiment 1 (Figure 1D). Time ranges ofsignificant waveform differences reflecting the N2pc arehighlighted by horizontal bars in Figure 4C. The N2pc tofearful faces appears roughly in the same time range as thetarget-related N2pc, and as in Experiment 1, the onset ofthe N2pc to fearful faces is delayed by 25 msec, that is, byan amount that is comparable to Experiment 1 (20 msec).Also, the onset of the target-related N2pc of the bilateralfearful condition arises with a small delay (18 msec) relativeto the bilateral neutral condition—a delay not observedin Experiment 1 (Figure 3C). To summarize, irrelevant fear-ful faces elicited an N2pc response that is completely in-dependent of the target-related N2pc response. Despitethe reduced perceptual demands on target discriminationin Experiment 2, size, time course, and current origin of
Fenker et al. 2933

the N2pc response to the fearful faces are very similar inboth experiments.
DISCUSSION
The presented data from both experiments clearly demon-strate that a lateralized presentation of an irrelevant fearful
face elicits a small but significant N2pc effect in ERMF re-sponse approximately between 240 and 400 msec thatshows a typical current origin in ventral extrastriate cortex(Hopf et al., 2000). The results of Experiment 2 indicatethat this effect is not altered when changing the perceptualdemands of the task-relevant search. The N2pc has beenshown to represent a reliable signature of rapid shifts of
Figure 4. Target-related N2pc of the (A) bilateral neutral and bilateral fearful conditions together with the N2pc elicited by (B) lateralizedfearful faces of Experiment 2. The N2pc is highlighted as gray area between traces. Source density estimates (MNLS) were computed from leftVF target-minus-right VF target difference at 260 msec and from the difference fearful face in left VF-minus-fearful face in right VF at 280 msec.(C) The time course of target-related and fearful face-related N2pcs of Experiment 2. Horizontal bars indicate the time range of significantwaveform differences as revealed by sample-wise t test. Arrows mark the mean onset of the target- and the fearful face-related N2pc.
2934 Journal of Cognitive Neuroscience Volume 22, Number 12

the attentional focus in visual search (Woodman & Luck,1999, 2003b), even under conditions where target infor-mation does not reach the stage of conscious representa-tion (Woodman & Luck, 2003a). Given that face valence inthe present experiments was completely task irrelevantand nonpredictive with respect to the location of thesearch target, the observation of an N2pc appears to at-test true automatic capture (Yantis, 1996), at least with re-spect to neuromagnetic correlates of attentional focusing.In other words, faces with negative emotional valence—even when being completely task irrelevant—do influenceattentional processing in extrastriate visual areas to a cer-tain degree in a mandatory manner. As detailed in theintroduction, previous demonstrations of automatic atten-tional capture by fearful faces were challenged (Purcellet al., 1996) on the basis of the possibility that low-levelfeatural differences between fearful and neutral faces wereaccounting for the behavioral effects. The present experi-ments extend those studies in showing that fearful facescapture attention when respective low-level confoundsare eliminated. Experiment 2 reveals that when the per-ceptual demands of the search task are low, the presenceof an irrelevant fearful face in the target VF reduces searchspeed, consistent with the notion that fearful faces im-pose a distracting influence on focusing onto the searchtarget.The present findings are generally in line with recent
observations by Eimer and Kiss (2007) who reportedthat irrelevant fearful faces presented in the visual periph-ery elicited an N2pc in the ERP response while subjectsperformed a task requiring the detection of occasionalluminance changes at fixation. A similar conclusion wasdrawn by Pourtois et al. (2004). They observed that whenpresenting an irrelevant fearful face versus a neutralface at the location of a subsequent target item, the ERPresponse to the target was enhanced around 130 msec(P1 attention effect), indicating that fearful faces ledto automatic location precuing. It should be noted thatalthough the present data demonstrate automatic atten-tional capture by fearful in comparison to neutral faces,the capturing effect may not be specific to the fear fea-ture of faces. It is likely that other emotional expressionslike sadness or happiness produce similar capture. Infact, a recent ERP study by Brosch, Sander, Pourtois, andScherer (2008) demonstrates that irrelevant face stimulieliciting positive emotions (nurturance-inducing babyfaces) produce attentional capture as revealed by a P1 at-tention effect analogous to the one seen in Pourtois et al.(2004).The present MEG studies combine the presentation of
emotional face features with the simultaneous require-ment to perform a demanding visual search task. Thisadds to these previous studies the possibility to evaluatethe time course of operations underlying the automaticcapture by fearful face features relative to the time courseof neural processes subserving attentional focusing invisual search. The present data show that the N2pc to
irrelevant fearful faces arises with an onset not earlierthan the N2pc reflecting target selection. In fact, the timecourse is delayed relative to task-relevant processes of at-tentional focusing onto the bars by a comparable amount(20 and 25 msec) in both experiments, suggesting thatperceptual load does not influence the relative time courseof neural processing devoted to the target versus thefearful faces. Hence, fearful face expression modulatesneural processing in extrastriate cortex automatically, butnot with temporal priority. In Experiment 2, a small de-lay of the target-related N2pc is seen for the bilateral fear-ful relative to the bilateral neutral condition (Figure 4C).Response times to the bilateral neutral and fearful condi-tions, however, are not different, indicating that this delaymay not relate to the response time slowing to unilat-eral fearful faces presented in the target versus the non-target VF.
Evidence for mandatory and automatic processing offear features in extrastriate cortex has also been providedwith fMRI recordings. Vuilleumier, Armony, Driver, andDolan (2001), for example, observed that fearful facesenhanced the BOLD response not only in the amygdalabut also in the fusiform cortex even when faces appearedat irrelevant spatial locations. On the basis of convergingevidence from fMRI lesion studies (Vuilleumier, Richardson,Armony, Driver, & Dolan, 2004), the authors speculate thatfeedback projections from the amygdala to extrastriatevisual cortex entail modulatory influences on visual pro-cessing that are fast and independent of modulations aris-ing from typical attentional control structures in parietaland frontal cortex (Vuilleumier & Driver, 2007). Analysesof fMRI-based functional connectivity have further cor-roborated that notion (Noesselt, Driver, Heinze, & Dolan,2005). Given the independence of the N2pc responsesto fearful faces and to the search target, the present dataare clearly in line with the possibility that the N2pc to fear-ful faces reflects top–down modulatory influences from theamygdala. However, the onset latency of approximately240 msec suggests a considerable delay relative to theinitial feed-forward sweep of processing in visual cortex(Foxe & Simpson, 2002). The N2pc to fearful faces is de-layed even with respect to the typical face-related responsein the electro-magnetic signal (N170 and N200) (Allison,Puce, Spencer, & McCarthy, 1999; Bentin, McCarthy, Perez,Puce, & Allison, 1996) and is therefore less consistent withface valence information being transferred by a fast routethat bypasses the canonical visual pathway (Pessoa et al.,2002). A delayed onset of this order, however, appears tobe in line with the time course of modulatory effects ob-served with ERPs (Krolak-Salmon, Fischer, Vighetto, &Mauguiere, 2001) or recently with intracranial recordings(Krolak-Salmon, Henaff, Vighetto, Bertrand, & Mauguiere,2004). Krolak-Salmon et al. (2004), for example, report thatfaces expressing emotion (fearful or happy) produced amarked enhancement of neural activity in the amygdalaand fusiform cortex but approximately 180 msec onwardafter face presentation.
Fenker et al. 2935

Dissociation between Neuromagnetic andBehavioral Data
Despite the presence of N2pc effects to irrelevant fearfulfaces in both experiments, behavior was not significantlyinfluenced by the distribution of fearful faces in Experi-ment 1. In contrast, reducing the perceptual demands oftarget discrimination in Experiment 2 produced a signifi-cant effect on response time—an observation that fits withnotions of the PLT of visual attention (Lavie et al., 2004;Lavie & de Fockert, 2003; Lavie & Tsal, 1994). PLT suggeststhat under high perceptual load, distractors may not haveany effect on performance because perceptual capacity isexhausted, but when load is sufficiently reduced, idle re-sources may spill over to process distractor information.We may, therefore, conclude that irrelevant fearful facesmodulate neural operations underlying attentional focus-ing and target discrimination in extrastriate cortex man-datorily, but this modulation may not relate to effects onthe behavioral level unless the perceptual demands of theattention task are sufficiently low.
It should be noted that this conclusion partially conflictswith fMRI studies that have investigated this issue (Bishop,Jenkins, & Lawrence, 2007; Pessoa, Padmala, & Morland,2005; Pessoa et al., 2002). Those studies observed thatBOLD effects to irrelevant emotional faces in the amygdalaand extrastriate cortex are eliminated when attentional re-sources are exhausted by a demanding discrimination task.Although the present data suggest such load dependencyat the behavioral level in line with Pessoa et al. (2005),we find neural correlates in the neuromagnetic responseunder high load conditions of Experiment 1. Of course, itis possible that the search task in Experiment 1 did not ex-haust attentional resources as much as the orientation dis-crimination task in the Pessoa et al. (2002) study. Also,there are critical differences in experimental design andmethodology. In the studies of Bishop et al. (2007) andPessoa et al., (2002, 2005), faces were presented at fixation,whereas in the present study faces appeared at peripheralVF locations. That is, the way faces influenced attentionalselection presumably differed between studies. Further-more, studies performed with fMRI may not be as sensitiveas MEG recordings in assessing brief and transient activitydifferences. Further research is clearly needed to clarifythe discrepancies.
Nevertheless, the apparent dissociation between neuro-physiological measures of attention and performance hasa notable implication. Under high perceptual load perfor-mance, effects are negligible while neural correlates re-flecting attentional selection in extrastriate visual areas areclearly observed. Reducing perceptual load leads to signifi-cant behavioral effects but does not change the neural cor-relates of attention in extrastriate cortex. This suggests thatthe ultimate locus where resources trade for controlling be-havior (in the sense of PLT)may not be the locus of sensory/attentional selection reflected by the N2pc. This is notableas the N2pc is known to reflect the attenuation of distractor
interference in visual search (Hopf, Boelmans, et al., 2002;Luck, Girelli, McDermott, & Ford, 1997), and PLT in factsuggests that load effects pertain to the reduction of dis-tractor interference during stimulus encoding. Althoughthis appears to be puzzling, it is important to acknowledgethat the perceptual encoding of stimuli is likely mediatedby multiple selection processes and the N2pc does not re-flect all of them (Hopf et al., 2000). It is possible that behav-ioral effects under low load result from a spill of resourcesfrom perceptual processes that attentional selection re-flected by the N2pc does not draw on.In sum, despite variable effects on the behavioral level,
the presented MEG data indicate that completely irrelevantfearful faces influence attentional processing in extrastriatecortex in an automatic way. This automatic influence turnsout to be unaltered by a change of the perceptual demandsof the task-relevant search. Automaticity, however, doesnot imply temporal priority. Fearful face associated pro-cessing in extrastriate cortex is found to arise with a slightdelay relative to task-relevant attentional processing. Ourobservations add to a growing body of neurophysiologicalresearch suggesting privileged processing of emotionalstimuli (Brosch et al., 2008; Eimer & Kiss, 2007; Anderson,2005; Noesselt et al., 2005; Vuilleumier, 2002).
Conclusion
Although it is widely suggested that fearful faces captureattention in an automatic fashion, this notion has beenquestioned after a critical evaluation of the available behav-ioral evidence (Cave & Batty, 2006; Horstmann & Bauland,2006; Purcell et al., 1996). Here, we add to this debate byproviding neurophysiological evidence that irrelevant fearfulfaces, indeed, “capture” sensory processing subserving atten-tional selection in extrastriate visual cortex in an automaticway, even when effects on behavior are nonsignificant.
Acknowledgments
This research was made possible with the support provided bythe DFG-Grant, KFO 163, TP4.
Reprint requests should be sent to Jens-Max Hopf, Departmentof Neurology, Otto-von-Guericke University, Leipziger Strasse44, 39120 Magdeburg, Germany, or via e-mail: [email protected].
REFERENCES
Allison, T., Puce, A., Spencer, D. D., & McCarthy, G. (1999).Electrophysiological studies of human face perception:I. Potentials generated in occipitotemporal cortex by faceand non-face stimuli. Cerebral Cortex, 9, 415–430.
Anderson, A. K. (2005). Affective influences on the attentionaldynamics supporting awareness. Journal of ExperimentalPsychology: General, 134, 258–281.
Anderson, A. K., Christoff, K., Panitz, D., De Rosa, E., &Gabrieli, J. D. (2003). Neural correlates of the automaticprocessing of threat facial signals. Journal of Neuroscience,23, 5627–5633.
2936 Journal of Cognitive Neuroscience Volume 22, Number 12

Bentin, S., McCarthy, G., Perez, E., Puce, A., & Allison, T.(1996). Electrophysiological studies of face perception inhumans. Journal of Cognitive Neuroscience, 8, 551–565.
Bishop, S. J., Jenkins, R., & Lawrence, A. D. (2007). Neuralprocessing of fearful faces: Effects on anxiety are gradedby perceptual capacity limitations. Cerebral Cortex, 17,1595–1603.
Brosch, T., Sander, D., Pourtois, G., & Scherer, K. R. (2008).Beyond fear: Rapid spatial orienting toward positiveemotional stimuli. Psychological Science, 19, 362–370.
Cave, K. R., & Batty, M. J. (2006). From searching for featuresto searching for threat: Drawing the boundary betweenpreattentive and attentive vision. Visual Cognition, 14,629–646.
Dolan, R. J., & Vuilleumier, P. (2003). Amygdala automaticityin emotional processing. Annals of the New York Academyof Sciences, 985, 348–355.
Eastwood, J. D., Smilek, D., & Merikle, P. M. (2001). Differentialattentional guidance by unattended faces expressingpositive and negative emotion. Perception &Psychophysics, 63, 1004–1013.
Eastwood, J. D., Smilek, D., & Merikle, P. M. (2003).Negative facial expression captures attention and disruptsperformance. Perception & Psychophysics, 65, 352–358.
Eimer, M., Holmes, A., & McGlone, F. P. (2003). The role ofspatial attention in the processing of facial expression: AnERP study of rapid brain responses to six basic emotions.Cognitive, Affective & Behavioral Neuroscience, 3, 97–110.
Eimer, M., & Kiss, M. (2007). Attentional capture bytask-irrelevant fearful faces is revealed by the N2pccomponent. Biological Psychology, 74, 108–112.
Ekman, P., & Friesen, W. V. (1978). Pictures of facial affect.Palo Alto, CA: Consulting Psychologists Press.
Fiorentini, A., & Berardi, N. (1984). Right-hemispheresuperiority in the discrimination of spatial phase.Perception, 13, 695–708.
Folk, C. L., Remington, R. W., & Johnston, J. C. (1992).Involuntary covert orienting is contingent on attentionalcontrol settings. Journal of Experimental Psychology:Human Perception and Performance, 18, 1030–1044.
Folk, C. L., Remington, R. W., & Wright, J. H. (1994). Thestructure of attentional control: Contingent attentionalcapture by apparent motion, abrupt onset, and color.Journal of Experimental Psychology: Human Perceptionand Performance, 20, 317–329.
Fox, E., Lester, V., Russo, R., Bowles, R. J., Pichler, A., &Dutton, K. (2000). Facial expressions of emotion: Are angryfaces detected more efficiently? Cognition & Emotion, 14,61–92.
Foxe, J. J., & Simpson, G. V. (2002). Flow of activation fromV1 to frontal cortex in humans. A framework for defining“early” visual processing. Experimental Brain Research,142, 139–150.
Fuchs, M., Wagner, M., Kohler, T., & Wischmann, H. A. (1999).Linear and nonlinear current density reconstructions.Journal of Clinical Neurophysiology, 16, 267–295.
Gur, R. C., Sara, R., Hagendoorn, M., Marom, O., Hughett, P.,Macy, L., et al. (2002). A method for obtaining 3-dimensionalfacial expressions and its standardization for use inneurocognitive studies. Journal of Neuroscience Methods,115, 137–143.
Guthrie, D., & Buchwald, J. S. (1991). Significance testing ofdifference potentials. Psychophysiology, 28, 240–244.
Hamalainen, M. S., & Ilmoniemi, R. J. (1994). Interpretingmagnetic fields of the brain: Minimum norm estimates.Medical & Biological Engineering & Computing, 32, 35–42.
Hämäläinen, M. S., & Sarvas, J. (1989). Realistic conductivitygeometry model of the human head for interpretation of
neuromagnetic data. IEEE Transactions on BiomedicalEngineering, 36, 165–171.
Hansen, C. H., & Hansen, R. D. (1988). Finding the face in thecrowd: An anger superiority effect. Journal of Personalityand Social Psychology, 54, 917–924.
Holmes, A., Vuilleumier, P., & Eimer, M. (2003). The processingof emotional facial expression is gated by spatial attention:Evidence from event-related brain potentials. BrainResearch, Cognitive Brain Research, 16, 174–184.
Hopf, J. M., Boelmans, K., Schoenfeld, A., Luck, S. J., & Heinze,H.-J. (2004). Attention to features precedes attention tolocations in visual search: Evidence from electromagneticbrain responses in humans. Journal of Neuroscience, 24,1822–1832.
Hopf, J. M., Boelmans, K., Schoenfeld, A. M., Heinze, H.-J.,& Luck, S. J. (2002). How does attention attenuatetarget-distractor interference in vision? Evidence frommagnetoencephalographic recordings. Cognitive BrainResearch, 15, 17–29.
Hopf, J. M., Luck, S. J., Girelli, M., Hagner, T., Mangun,G. R., Scheich, H., et al. (2000). Neural sources of focusedattention in visual search. Cerebral Cortex, 10, 1233–1241.
Hopf, J. M., Vogel, E., Woodman, G., Heinze, H. J., & Luck, S. J.(2002). Localizing visual discrimination processes in timeand space. Journal of Neurophysiology, 88, 2088–2095.
Horstmann, G., & Bauland, A. (2006). Search asymmetrieswith real faces: Testing the anger-superiority effect.Emotion, 6, 193–207.
Krolak-Salmon, P., Fischer, C., Vighetto, A., & Mauguiere, F.(2001). Processing of facial emotional expression:Spatio-temporal data as assessed by scalp event-relatedpotentials. European Journal of Neuroscience, 13, 987–994.
Krolak-Salmon, P., Henaff, M. A., Vighetto, A., Bertrand, O.,& Mauguiere, F. (2004). Early amygdala reaction to fearspreading in occipital, temporal, and frontal cortex: A depthelectrode ERP study in human. Neuron, 42, 665–676.
Lavie, N. (2001). Capacity limits in selective attention:Behavioral evidence and implications for neural activity.In J. Braun, C. Koch, & J. L. Davis (Eds.), Visual attentionand cortical circuits (pp. 49–68). Cambridge, MA:MIT Press.
Lavie, N., & de Fockert, J. W. (2003). Contrasting effects ofsensory limits and capacity limits in visual selectiveattention. Perception & Psychophysics, 65, 202–212.
Lavie, N., Hirst, A., de Fockert, J. W., & Viding, E. (2004).Load theory of selective attention and cognitive control.Journal of Experimental Psychology: General, 133,339–354.
Lavie, N., Ro, T., & Russell, C. (2003). The role of perceptualload in processing distractor faces. Psychological Science,14, 510–515.
Lavie, N., & Tsal, Y. (1994). Perceptual load as a majordeterminant of the locus of selection in visual attention.Perception & Psychophysics, 56, 183–197.
Luck, S. J., Girelli, M., McDermott, M. T., & Ford, M. A.(1997). Bridging the gap between monkey neurophysiologyand human perception: An ambiguity resolution theory ofvisual selective attention. Cognitive Psychology, 33, 64–87.
Luck, S. J., & Hillyard, S. A. (1994a). Electrophysiologicalcorrelates of feature analysis during visual search.Psychophysiology, 31, 291–308.
Luck, S. J., & Hillyard, S. A. (1994b). Spatial filtering duringvisual search: Evidence from human electrophysiology.Journal of Experimental Psychology: Human Perceptionand Performance, 20, 1000–1014.
Moors, A., & de Houwer, J. (2006). Automaticity: A theoreticaland conceptual analysis. Psychological Bulletin, 132,297–326.
Fenker et al. 2937

Morris, J. S., Ohman, A., & Dolan, R. J. (1999). A subcorticalpathway to the right amygdala mediating “unseen” fear.Proceedings of the National Academy of Sciences, U.S.A.,96, 1680–1685.
Noesselt, T., Driver, J., Heinze, H. J., & Dolan, R. (2005).Asymmetrical activation in the human brain duringprocessing of fearful faces. Current Biology, 15, 424–429.
Nothdurft, H.-C. (1993). Faces and facial expressions do notpop out. Perception, 22, 1287–1298.
Ohman, A., Lundqvist, D., & Esteves, F. (2001). The face inthe crowd revisited: A threat advantage with schematicstimuli. Journal of Personality and Social Psychology,80, 381–396.
Pessoa, L., McKenna, M., Gutierrez, E., & Ungerleider, L. G.(2002). Neural processing of emotional faces requiresattention. Proceedings of the National Academy ofSciences, U.S.A., 99, 11458–11463.
Pessoa, L., Padmala, S., & Morland, T. (2005). Fate ofunattended fearful faces in the amygdala is determinedby both attentional resources and cognitive modulation.Neuroimage, 28, 249–255.
Pourtois, G., Grandjean, D., Sander, D., & Vuilleumier, P.(2004). Electrophysiological correlates of rapid spatialorienting towards fearful faces. Cerebral Cortex, 14,619–633.
Purcell, D. G., Stewart, A. L., & Skov, R. B. (1996). It takes aconfounded face to pop out of a crowd. Perception, 25,1091–1108.
Robinson, S. E. (1989). Environmental noise cancellation forbiomagnetic measurements. In S. J. Williamson, M. Hoke,G. Stroink, & M. Kotani (Eds.), Advances in biomagnetism(pp. 721–724). New York: Plenum Press.
Silvert, L., Lepsien, J., Fragopanagos, N., Goolsby, B., Kiss, M.,Taylor, J. G., et al. (2007). Influence of attentional demandson the processing of emotional facial expressions in theamygdala. Neuroimage, 38, 357–366.
Vuilleumier, P. (2002). Facial expression and selective attention.Current Opinion in Psychiatry, 15, 291–300.
Vuilleumier, P., Armony, J. L., Driver, J., & Dolan, R. J. (2001).Effects of attention and emotion on face processing in thehuman brain: An event-related fMRI study. Neuron, 30,829–841.
Vuilleumier, P., Armony, J. L., Driver, J., & Dolan, R. J.(2003). Distinct spatial frequency sensitivities for processingfaces and emotional expressions. Nature Neuroscience, 6,624–631.
Vuilleumier, P., & Driver, J. (2007). Modulation of visualprocessing by attention and emotion: Windows on causalinteractions between human brain regions. PhilosophicalTransactions of the Royal Society of London, Series B,Biological Sciences, 362, 837–855.
Vuilleumier, P., Richardson, M. P., Armony, J. L., Driver, J., &Dolan, R. J. (2004). Distant influences of amygdala lesion onvisual cortical activation during emotional face processing.Nature Neuroscience, 7, 1271–1278.
Vuilleumier, P., & Schwartz, S. (2001). Emotional facialexpressions capture attention. Neurology, 56, 153–158.
White, M. (1995). Preattentive analysis of facial expressionsof emotion. Cognition & Emotion, 9, 439–460.
Woodman, G. F., & Luck, S. J. (1999). Electrophysiologicalmeasurement of rapid shifts of attention during visualsearch. Nature, 400, 867–869.
Woodman, G. F., & Luck, S. J. (2003a). Dissociations amongattention, perception, and awareness during object-substitutionmasking. Psychological Science, 14, 605–611.
Woodman, G. F., & Luck, S. J. (2003b). Serial deploymentof attention during visual search. Journal of ExperimentalPsychology: Human Perception and Performance, 29,121–138.
Yantis, S. (1993). Stimulus-driven attentional capture andattentional control settings. Journal of ExperimentalPsychology: Human Perception and Performance, 19,676–681.
Yantis, S. (1996). Attentional capture in vision. In A. Kramer,M. Coles, & G. Logan (Eds.), Converging operations in thestudy of selective visual attention (pp. 45–76). Washington,DC: American Psychological Association.
Yantis, S., & Egeth, H. E. (1999). On the distinction betweenvisual salience and stimulus-driven attentional capture.Journal of Experimental Psychology: Human Perceptionand Performance, 25, 661–676.
Yantis, S., & Johnson, D. N. (1990). Mechanisms of attentionalpriority. Journal of Experimental Psychology: HumanPerception and Performance, 16, 812–825.
2938 Journal of Cognitive Neuroscience Volume 22, Number 12
Related Documents











