DEVELOPMENTAL BIOLOGY 97, 274-290 (1983) Mammary Ductal Elongation: Differentiation of Myoepithelium and Basal Lamina during Branching Morphogenesis JAMES M. WILLIAMS AND CHARLES W. DANIEL Department of Biology, Thimann Laboratories, University of California, Santa Cruz, California 95064 Received July 28, 1982; accepted in revised form January 10, 1983 Elongation of mammary ducts in the immature mouse takes place as a result of rapid growth in end buds. These structures proliferate at the apex of elongating ducts and are responsible for penetration of the surrounding adipose stroma; by turning and branching, end buds give rise to the characteristic open pattern of the mammary ductal tree. We have used a variety of techniques to determine the cellular and structural basis for certain of these end bud activities, and now report the following. (1) The end bud tip is covered with a monolayer of epithelium, the “cap cells,” which are characterized by a relative lack of intercellular junctions and other specialized features. (2) The cap cell layer extends along the end bud flank and neck regions where it is continuous with the myoepithelium which surrounds the subtending mature duct. A linear sequence of differentiative changes occur in the cap cells in this region as they progressively alter in shape and accumulate the cytological features of mature myoepithelium. Cap cells may therefore be defined as a stem cell population providing new myoepithelial cells for ductal morphogenesis and elongation. (3) Differentiation of cap cells into myoepithelium is associated with conspicuous changes in the basal lamina. At the tip, cap cells form a 104-nm lamina similar to that described in expanding mammary alveoli and in embryonic tissues. Along the end bud flanks the basal lamina is raised from the cell surface and extensively folded, resulting in a greatly thickened lamina, measuring as much as 1.4 pm. At the surface of the subtending ducts the lamina becomes structurally simplified and resembles that at the tip, but has a significantly greater thickness, averaging 130 nm. (4) The codif- ferentiation of myoepithelium and its basement membrane is associated with changes in the surrounding stroma. Undifferentiated mesenchymal-like cells attach to the surface of the basal lamina in the midportion of the end buds and become increasingly numerous in the neck region, forming a monolayer over the myoepithelial basal lamina. These stromal cells progressively differentiated into fibroeytes which participate in collagen fibrillogenesis and give rise to the fibrous components of the stroma surrounding the mature duct. INTRODUCTION An interesting and comparatively little studied phase of mammary development occurs in the immature mouse, when mammary tissue grows rapidly to produce a tree-like pattern of ducts upon which, during preg- nancy, secretory alveoli may form. The most rapid pe- riod of ductal expansion occurs in the mouse between approximately 4 and 7 weeks of age, and is the result of intense mitotic activity in mammary end buds (Bres- ciani, 1965; Russo and Russo, 19781980). These bulbous epithelial structures appear to be specialized at their advancing edge to permit rapid penetration of the sur- rounding fatty stroma (Silberstein and Daniel, 1982b), and the posterior regions of end buds provide a supply of differentiating ductal and myoepithelial cells for elongation of subtending ducts. In addition to linear growth, mammary end buds branch dichotomously at fairly regular intervals and thereby generate the basic patterning of the gland. By turning to avoid competing tissue, they provide for regular spacing between ductal elements and, when insufficient gland-free stroma is available for continued growth, end buds disappear and their subtending ducts become mitotically inactive (Faulkin and DeOme, 1960). The basement membrane of mature mammary ducts is composed of a fibrous collagenous tunic which blends with the surrounding fatty stroma and a thinner, highly organized basal lamina which envelops the ducts and conforms closely to surface contours of the myoepithe- ha1 cells. The end bud basal lamina has recently been shown to be composed in part of glycosaminoglycans (Silberstein and Daniel, 1982b). At the tip of the ad- vancing bud hyaluronate is synthesized and appears to be associated with the process of tissue penetration. In the flanks of the end bud, where cell differentiation and ductal morphogenesis are taking place, hyaluronate synthesis gives way to the production of sulfate-rich glycosaminoglycans (S-GAGS), which are associated with collagen fibrillogenesis, cytodifferentiation, and tissue stabilization (Trelstad et aZ., 1974). The basal lamina of mammary alveoli in midpregnant mice has recently been shown to contain GAG in an organized ultrastructural pattern, and resembles the basal laminae of certain embryonic epithelia under- going branching morphogenesis (Gordon and Bernfield, 1980). Little attention has been given to the structural organization of the lamina in mammary end buds, al- though by analogy with embryonic systems it may play an important role in ductal morphogenesis. Indeed, if 274 0012-1606/83 $3.00 Copyriyht 0 1983 by Academic Press, Inc. All rights of reproduction in any form reserved.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

DEVELOPMENTAL BIOLOGY 97, 274-290 (1983)
Mammary Ductal Elongation: Differentiation of Myoepithelium and Basal Lamina during Branching Morphogenesis
JAMES M. WILLIAMS AND CHARLES W. DANIEL
Department of Biology, Thimann Laboratories, University of California, Santa Cruz, California 95064
Received July 28, 1982; accepted in revised form January 10, 1983
Elongation of mammary ducts in the immature mouse takes place as a result of rapid growth in end buds. These structures proliferate at the apex of elongating ducts and are responsible for penetration of the surrounding adipose stroma; by turning and branching, end buds give rise to the characteristic open pattern of the mammary ductal tree. We have used a variety of techniques to determine the cellular and structural basis for certain of these end bud activities, and now report the following. (1) The end bud tip is covered with a monolayer of epithelium, the “cap cells,” which are characterized by a relative lack of intercellular junctions and other specialized features. (2) The cap cell layer extends along the end bud flank and neck regions where it is continuous with the myoepithelium which surrounds the subtending mature duct. A linear sequence of differentiative changes occur in the cap cells in this region as they progressively alter in shape and accumulate the cytological features of mature myoepithelium. Cap cells may therefore be defined as a stem cell population providing new myoepithelial cells for ductal morphogenesis and elongation. (3) Differentiation of cap cells into myoepithelium is associated with conspicuous changes in the basal lamina. At the tip, cap cells form a 104-nm lamina similar to that described in expanding mammary alveoli and in embryonic tissues. Along the end bud flanks the basal lamina is raised from the cell surface and extensively folded, resulting in a greatly thickened lamina, measuring as much as 1.4 pm. At the surface of the subtending ducts the lamina becomes structurally simplified and resembles that at the tip, but has a significantly greater thickness, averaging 130 nm. (4) The codif- ferentiation of myoepithelium and its basement membrane is associated with changes in the surrounding stroma. Undifferentiated mesenchymal-like cells attach to the surface of the basal lamina in the midportion of the end buds and become increasingly numerous in the neck region, forming a monolayer over the myoepithelial basal lamina. These stromal cells progressively differentiated into fibroeytes which participate in collagen fibrillogenesis and give rise to the fibrous components of the stroma surrounding the mature duct.
INTRODUCTION
An interesting and comparatively little studied phase of mammary development occurs in the immature mouse, when mammary tissue grows rapidly to produce a tree-like pattern of ducts upon which, during preg- nancy, secretory alveoli may form. The most rapid pe- riod of ductal expansion occurs in the mouse between approximately 4 and 7 weeks of age, and is the result of intense mitotic activity in mammary end buds (Bres- ciani, 1965; Russo and Russo, 19781980). These bulbous epithelial structures appear to be specialized at their advancing edge to permit rapid penetration of the sur- rounding fatty stroma (Silberstein and Daniel, 1982b), and the posterior regions of end buds provide a supply of differentiating ductal and myoepithelial cells for elongation of subtending ducts. In addition to linear growth, mammary end buds branch dichotomously at fairly regular intervals and thereby generate the basic patterning of the gland. By turning to avoid competing tissue, they provide for regular spacing between ductal elements and, when insufficient gland-free stroma is available for continued growth, end buds disappear and their subtending ducts become mitotically inactive (Faulkin and DeOme, 1960).
The basement membrane of mature mammary ducts is composed of a fibrous collagenous tunic which blends with the surrounding fatty stroma and a thinner, highly organized basal lamina which envelops the ducts and conforms closely to surface contours of the myoepithe- ha1 cells. The end bud basal lamina has recently been shown to be composed in part of glycosaminoglycans (Silberstein and Daniel, 1982b). At the tip of the ad- vancing bud hyaluronate is synthesized and appears to be associated with the process of tissue penetration. In the flanks of the end bud, where cell differentiation and ductal morphogenesis are taking place, hyaluronate synthesis gives way to the production of sulfate-rich glycosaminoglycans (S-GAGS), which are associated with collagen fibrillogenesis, cytodifferentiation, and tissue stabilization (Trelstad et aZ., 1974).
The basal lamina of mammary alveoli in midpregnant mice has recently been shown to contain GAG in an organized ultrastructural pattern, and resembles the basal laminae of certain embryonic epithelia under- going branching morphogenesis (Gordon and Bernfield, 1980). Little attention has been given to the structural organization of the lamina in mammary end buds, al- though by analogy with embryonic systems it may play an important role in ductal morphogenesis. Indeed, if
274 0012-1606/83 $3.00 Copyriyht 0 1983 by Academic Press, Inc. All rights of reproduction in any form reserved.
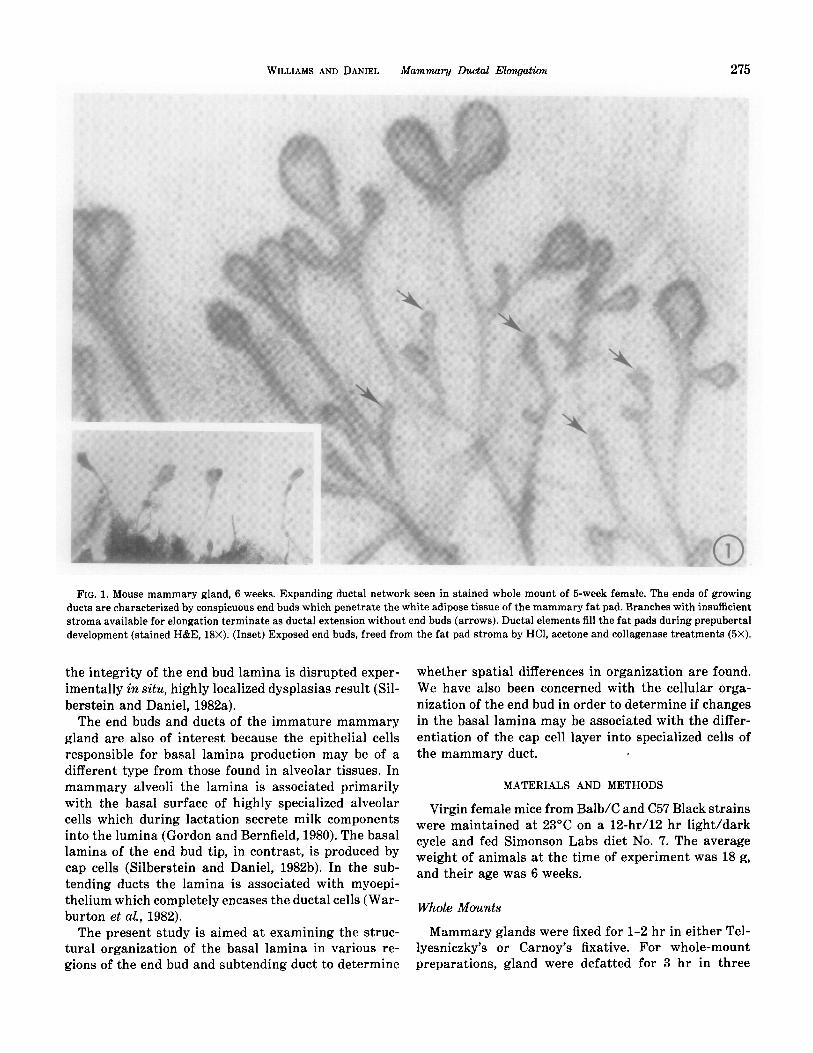
WILLIAMS AND DANIEL Mammary Ductal Elongation 275
FIG. 1. Mouse mammary gland, 6 weeks. Expanding ductal network seen in stained whole mount of 5-week female. The ends of growing ducts are characterized by conspicuous end buds which penetrate the white adipose tissue of the mammary fat pad. Branches with insufficient stroma available for elongation terminate as ductal extension without end buds (arrows). Ductal elements fill the fat pads during prepubertal development (stained H&E, 18X). (Inset) Exposed end buds, freed from the fat pad stroma by HCl, acetone and collagenase treatments (5X).
the integrity of the end bud lamina is disrupted exper- whether spatial differences in organization are found. imentally in situ, highly localized dysplasias result (Sil- We have also been concerned with the cellular orga- berstein and Daniel, 1982a). nization of the end bud in order to determine if changes
The end buds and ducts of the immature mammary in the basal lamina may be associated with the differ- gland are also of interest because the epithelial cells entiation of the cap cell layer into specialized cells of responsible for basal lamina production may be of a the mammary duct. different type from those found in alveolar tissues. In mammary alveoli the lamina is associated primarily MATERIALS AND METHODS
with the basal surface of highly specialized alveolar cells which during lactation secrete milk components into the lumina (Gordon and Bernfield, 1980). The basal lamina of the end bud tip, in contrast, is produced by cap cells (Silberstein and Daniel, 1982b). In the sub- tending ducts the lamina is associated with myoepi- thelium which completely encases the ductal cells (War- burton et cd, 1982).
The present study is aimed at examining the struc- tural organization of the basal lamina in various re- gions of the end bud and subtending duct to determine
Virgin female mice from Balb/C and C57 Black strains were maintained at 23°C on a 12-hr/12 hr light/dark cycle and fed Simonson Labs diet No. 7. The average weight of animals at the time of experiment was 18 g, and their age was 6 weeks.
Whole Mounts
Mammary glands were fixed for l-2 hr in either Tel- lyesniczky’s or Carnoy’s fixative. For whole-mount preparations, gland were defatted for 3 hr in three
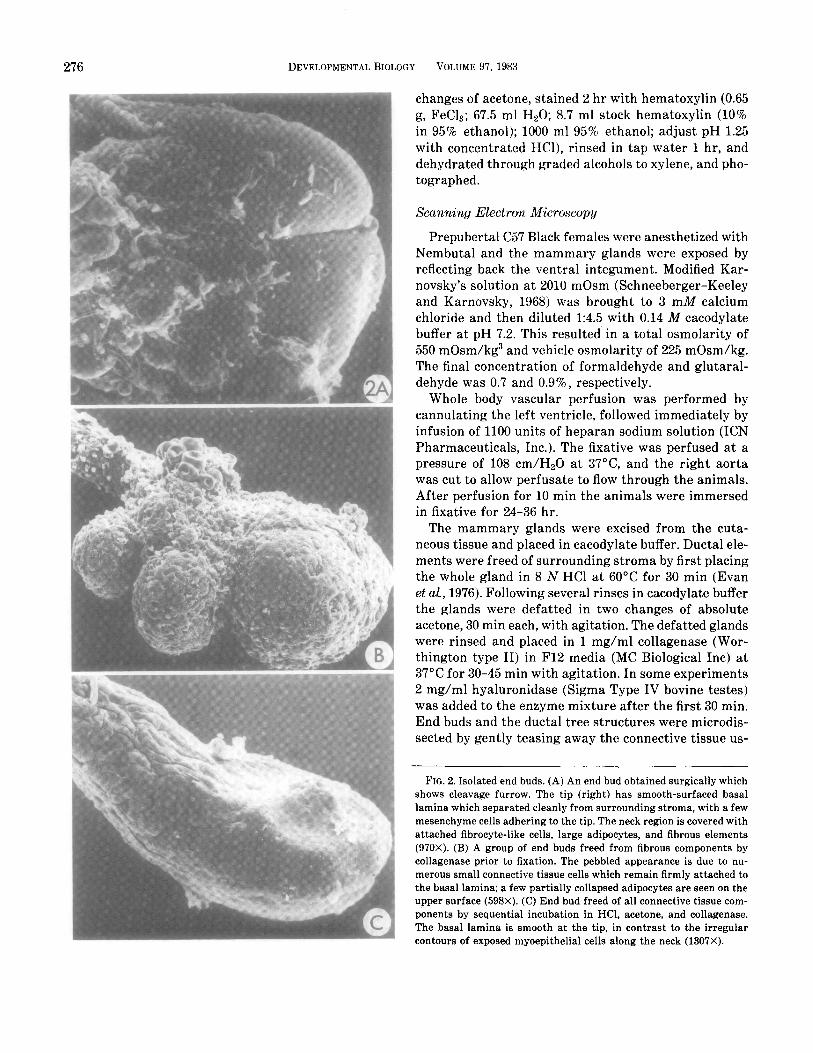
276 DEVELOPMENTAL BIOLOGY VOLUME 97, 1983
changes of acetone, stained 2 hr with hematoxylin (0.65 g, FeCl,; 67.5 ml H,O; 8.7 ml stock hematoxylin (10% in 95% ethanol); 1000 ml 95% ethanol; adjust pH 1.25 with concentrated HCl), rinsed in tap water 1 hr, and dehydrated through graded alcohols to xylene, and pho- tographed.
Scanning Electron Microscopy
Prepubertal C57 Black females were anesthetized with Nembutal and the mammary glands were exposed by reflecting back the ventral integument. Modified Kar- novsky’s solution at 2010 mOsm (Schneeberger-Keeley and Karnovsky, 1968) was brought to 3 mM calcium chloride and then diluted 1:4.5 with 0.14 M cacodylate buffer at pH 7.2. This resulted in a total osmolarity of 550 m0sm/kg3 and vehicle osmolarity of 225 mOsm/kg. The final concentration of formaldehyde and glutaral- dehyde was 0.7 and 0.9%, respectively.
Whole body vascular perfusion was performed by cannulating the left ventricle, followed immediately by infusion of 1100 units of heparan sodium solution (ICN Pharmaceuticals, Inc.). The fixative was perfused at a pressure of 108 cm/Hz0 at 37”C, and the right aorta was cut to allow perfusate to flow through the animals. After perfusion for 10 min the animals were immersed in fixative for 24-36 hr.
The mammary glands were excised from the cuta- neous tissue and placed in cacodylate buffer. Ductal ele- ments were freed of surrounding stroma by first placing the whole gland in 8 N HCl at 60°C for 30 min (Evan et al, 1976). Following several rinses in cacodylate buffer the glands were defatted in two changes of absolute acetone, 30 min each, with agitation. The defatted glands were rinsed and placed in 1 mg/ml collagenase (Wor- thington type II) in F12 media (MC Biological Inc) at 37°C for 30-45 min with agitation. In some experiments 2 mg/ml hyaluronidase (Sigma Type IV bovine testes) was added to the enzyme mixture after the first 30 min. End buds and the ductal tree structures were microdis- sected by gently teasing away the connective tissue us-
FIG. 2. Isolated end buds. (A) An end bud obtained surgically which shows cleavage furrow. The tip (right) has smooth-surfaced basal lamina which separated cleanly from surrounding stroma, with a few mesenchyme cells adhering to the tip. The neck region is covered with attached fibrocyte-like cells, large adipocytes, and fibrous elements (970X). (B) A group of end buds freed from fibrous components by collagenase prior to fixation. The pebbled appearance is due to nu- merous small connective tissue cells which remain firmly attached to the basal lamina; a few partially collapsed adipocytes are seen on the upper surface (598X). (C) End bud freed of all connective tissue com- ponents by sequential incubation in HCl, acetone, and collagenase. The basal lamina is smooth at the tip, in contrast to the irregular contours of exposed myoepithelial cells along the neck (1307X).
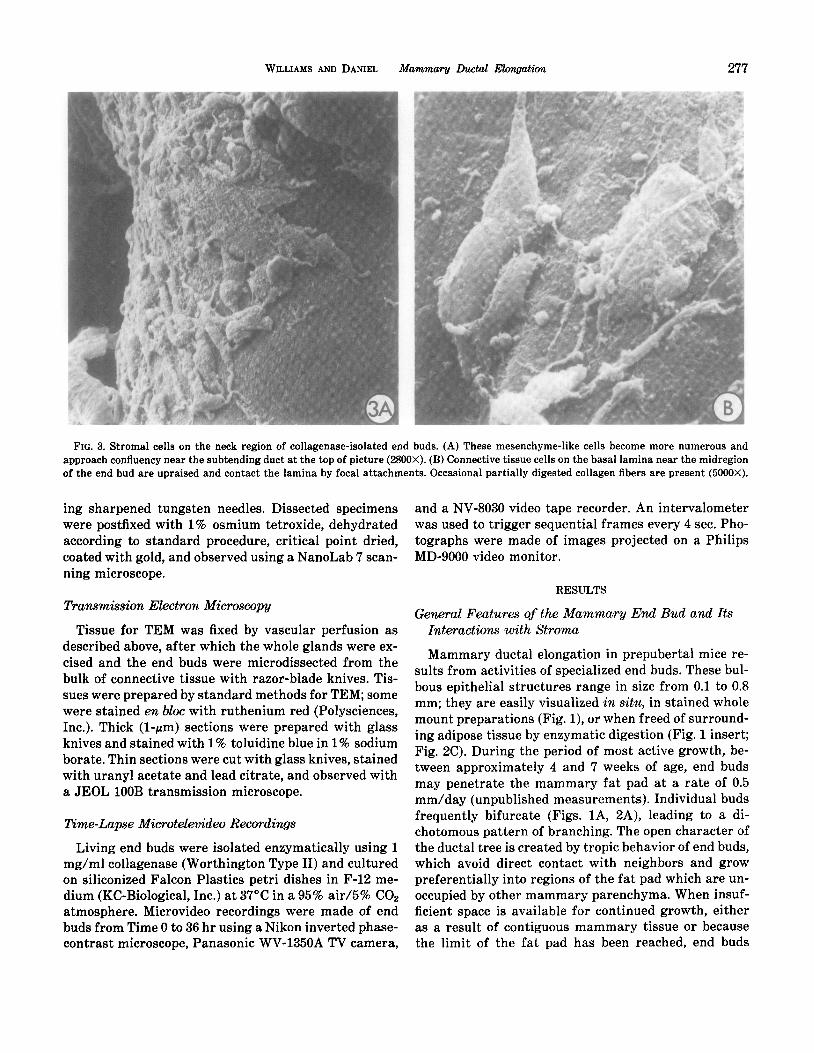
WILLIAMS AND DANIEL Mammary Ductal Elongation 277
FIG. 3. Stromal cells on the neck region of collagenase-isolated end buds. (A) These mesencbyme-like cells become more numerous and approach confluency near the subtending duct at the top of picture (2800X). (B) Connective tissue cells on the basal lamina near the midregion of the end bud are upraised and contact the lamina by focal attachments. Occasional partially digested collagen fibers are present (5000X).
ing sharpened tungsten needles. Dissected specimens were postfixed with 1% osmium tetroxide, dehydrated according to standard procedure, critical point dried, coated with gold, and observed using a NanoLab ‘7 scan- ning microscope.
Transmission Electron Microscopy
Tissue for TEM was fixed by vascular perfusion as described above, after which the whole glands were ex- cised and the end buds were microdissected from the bulk of connective tissue with razor-blade knives. Tis- sues were prepared by standard methods for TEM; some were stained en bloc with ruthenium red (Polysciences, Inc.). Thick (l-pm) sections were prepared with glass knives and stained with 1% toluidine blue in 1% sodium borate. Thin sections were cut with glass knives, stained with uranyl acetate and lead citrate, and observed with a JEOL 1OOB transmission microscope.
Time-Lapse Microtelevideo Recordings
Living end buds were isolated enzymatically using 1 mg/ml collagenase (Worthington Type II) and cultured on siliconized Falcon Plastics petri dishes in F-12 me- dium (KC-Biological, Inc.) at 3’7°C in a 95% sir/5% COP atmosphere. Microvideo recordings were made of end buds from Time 0 to 36 hr using a Nikon inverted phase- contrast microscope, Panasonic WV-1350A TV camera,
and a NV-8030 video tape recorder. An intervalometer was used to trigger sequential frames every 4 sec. Pho- tographs were made of images projected on a Philips MD-9000 video monitor.
RESULTS
General Features of the Mammary End Bud and Its Interactions with Stroma
Mammary ductal elongation in prepubertal mice re- sults from activities of specialized end buds. These bul- bous epithelial structures range in size from 0.1 to 0.8 mm; they are easily visualized in situ, in stained whole mount preparations (Fig. l), or when freed of surround- ing adipose tissue by enzymatic digestion (Fig. 1 insert; Fig. 2C). During the period of most active growth, be- tween approximately 4 and 7 weeks of age, end buds may penetrate the mammary fat pad at a rate of 0.5 mm/day (unpublished measurements). Individual buds frequently bifurcate (Figs. lA, 2A), leading to a di- chotomous pattern of branching. The open character of the ductal tree is created by tropic behavior of end buds, which avoid direct contact with neighbors and grow preferentially into regions of the fat pad which are un- occupied by other mammary parenchyma. When insuf- ficient space is available for continued growth, either as a result of contiguous mammary tissue or because the limit of the fat pad has been reached, end buds
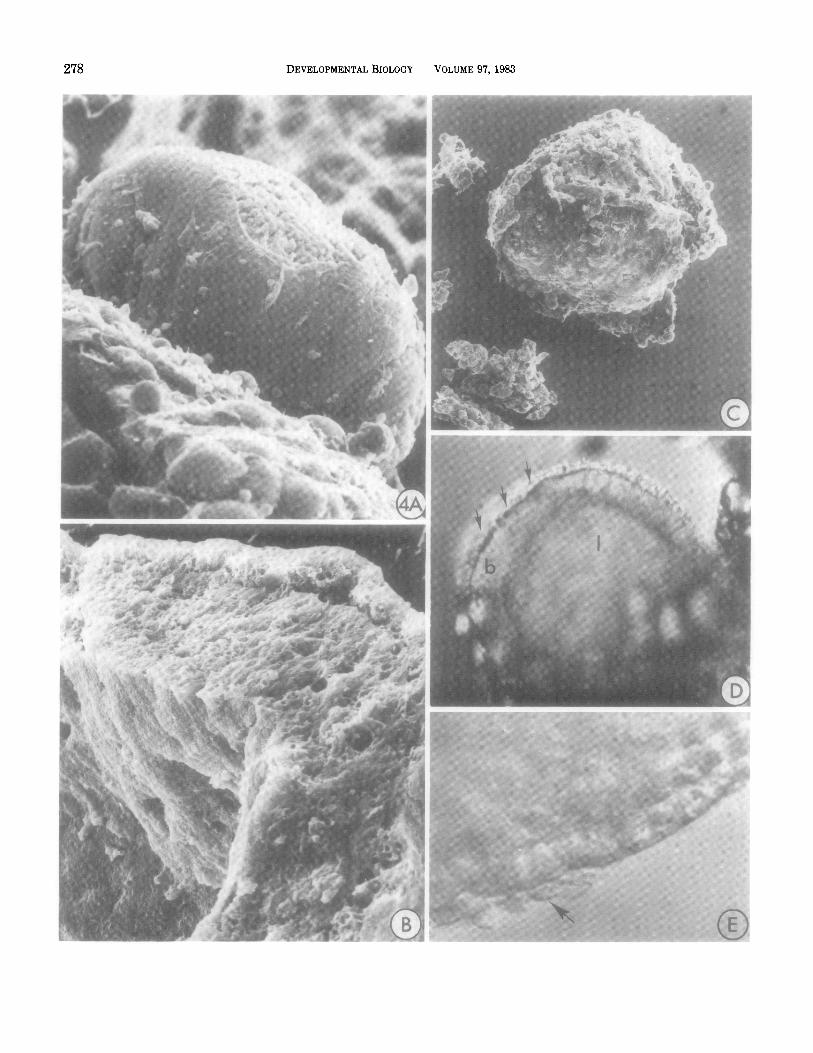
278 DEVELOPMENTALBIOLOGY VOLUME 97.1983

WILLIAMS AND DANIEL Mammary hctal Elongation 279
diminish in size and number, and the ductal tips become inactive (Fig. 1).
Characteristics of the epithelial-stromal boundaries were investigated by microdissection and treatment with various enzymes. When living end buds were teased away from surrounding tissue with fine needles and forceps the anterior half of the end bud was found to separate easily from the stroma, while the neck region and the subtending duct could be removed only by cutting or tearing the connective tissue. When these preparations were fixed by immersion, scanning electron microscopy revealed that the anterior portion of the surgically re- moved bud was covered with smooth-surfaced basal lamina and was free of adhesions (Fig. 2A). Occasional stromal cells remained attached to the lamina in this tip region; these became more numerous in the middle and neck portions where they were increasingly asso- ciated with attached fibers and adipocytes (Fig. 3). When other samples were treated with collagenase prior to fixation most of the fibers and adipocyte were removed, but numerous small stromal cells remained attached to the basal lamina (Fig. 2B). In order to avoid tissue changes taking place before fixation, other tissues were fixed in situ by whole body vascular perfusion followed by acid, acetone, and collagenase treatment, a procedure which removed both cellular and fibrous connective tis- sue elements and revealed the basal lamina of the entire end bud and associated duct (Fig. 2C). This method pre- served indentations in the end bud tip region caused by pressure of adipocytes against the rapidly advancing bud, suggesting the forces that are involved in this pro- cess of tissue penetration. These indentations were not observed in end buds fixed after enzymatic digestion (Figs. 2A, B), suggesting that smooth contours were restored during the period of incubation.
Surface details of stromal cells adhering to the basal lamina were visualized on end buds which had been surgically removed from the fat pad and fixed by im- mersion. Thinly scattered cells were attached to the tip region of actively growing end buds (Fig. 2A), but be- came more numerous in the mid portion and neck re- gion, where they formed a monolayer and often dis- played parallel orientation, resembling cells cultured to confluence on an artificial substrate (Fig. 3A). Individ- ual cells were attached to the basal lamina only in lo-
calized regions of the membrane, and most of the cell body was arched off the lamina; the cell bodies were not conspicuously flattened (Fig. 3B). The free surface of these adhering cells was characterized by small scat- tered blebs and protrusions, while the lateral surfaces displayed a moderate number of cell processes which attached to the substrate or made contact with neigh- boring cells. Transmission microscopy revealed that at- tached stromal cells in the anterior region of the end bud were structurally undifferentiated cells of mesen- chymal type (Fig. 5A), while those in the neck and duc- tal regions usually displayed cytoplasmic specializa- tions characteristic of protein synthetic fibrocysts (Fig. 11E).
Cap Cells
Directly beneath the basal lamina a loosely organized epithelial layer was identified (Fig. 4A), which usually was slightly separated from the end bud mass (Fig. 4B). In living preparations viewed with interference con- trast optics these outermost cells were readily distin- guished from the relatively compact mass of cells deeper in the end bud body, and were seen to form a continuous cortex one to two cell layers thick covering the end bud tip and flanks (Figs. 4D, E). We have termed this cor- tical layer “cap cells.”
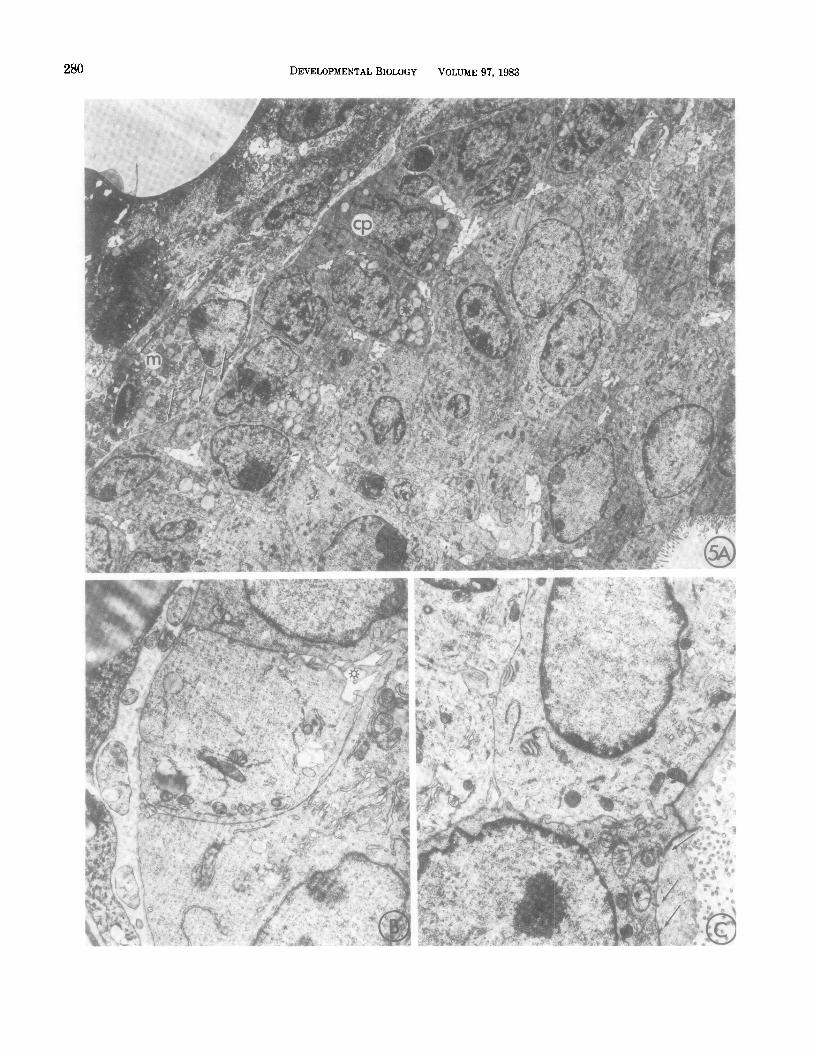
A number of features distinguish cap cells from pre- viously reported mammary cell types. Cap cells appear to lack specialized cell junctions or adhesions, and in- tercellular spaces were frequently observed on their lat- eral borders into which cell processes extended (Figs. 5A, B). A lack of adhesivity was particularly apparent on the apical surfaces, which were frequently separated from underlying cells of the end bud body by fluid-filled spaces (Figs. 4B, 5A, B). When unfixed end buds were treated with testicular hyaluronidase (2 mg/ml for 30 min) and observed microscopically, cap cells could be seen to detach, singly or in groups, from the underlying epithelium and to float away as the basal lamina was digested (Fig. 4C).
Cap cell nuclei appear round or oval in thin section and display prominent nucleoli, with sparse hetero- chromatin in the vicinity of the nuclear envelope. The cytoplasm is rich in free ribosomes, with small amounts
FIG. 4. End buds seen in scanning electron microscope preparations or with interference contrast optics. (A) Side view of an end bud surgically removed from the fat pad stroma; only a few connective tissue cells remain attached to the exposed basal lamina anteriorly, but posteriorly the neck and duct region could not be mechanically separated from the stroma. At the tip the basal lamina has been torn, exposing underlying cap cells (1800X). (B) On the fractured surface of an end bud the distinct cap cell layer is separated from the inner epithelium and lumen by a series of irregular spaces (3340X). (C) Collagenase- and hyaluronidase-treated end bud with connective tissue and portions of the basal lamina removed. Cap cells are easily dislodged from the inner epithelium when the basal lamina is digested (210X). (D) A living end bud viewed with interference contrast shows a clearly defined cap cell layer (arrows), body (b), and lumen (1) of the end bud (140X). (E) At higher magnification, a few mesenchyme cells are seen to remain attached after dissection in culture medium (arrowhead, 280X).
280 DEVELOPMENTALBIOLOGY VOLUME 97,1983
WILLIAMS AND DANIEL Mammary Ductal Elongation 281
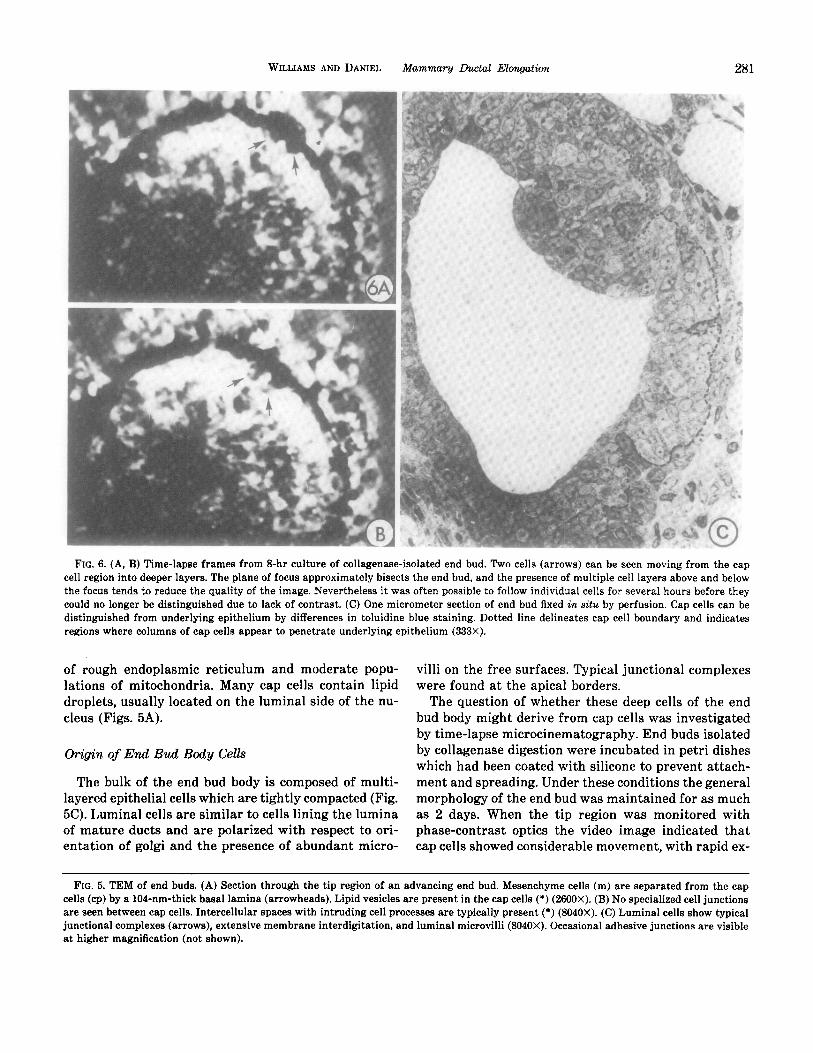
FIG. 6. (A, B) Time-lapse frames from 8-hr culture of collagenase-isolated end bud. Two cells (arrows) can be seen moving from the cap cell region into deeper layers. The plane of focus approximately bisects the end bud, and the presence of multiple cell layers above and below the focus tends to reduce the quality of the image. Nevertheless it was often possible to follow individual cells for several hours before they could no longer be distinguished due to lack of contrast. (C) One micrometer section of end bud fixed in situ by perfusion. Cap cells can be distinguished from underlying epithelium by differences in toluidine blue staining. Dotted line delineates cap cell boundary and indicates regions where columns of cap cells appear to penetrate underlying epithelium (333X).
of rough endoplasmic reticulum and moderate popu- lations of mitochondria. Many cap cells contain lipid droplets, usually located on the luminal side of the nu- cleus (Figs. 5A).
Origin of End Bud Body Cells
The bulk of the end bud body is composed of multi- layered epithelial cells which are tightly compacted (Fig. 5C). Luminal cells are similar to cells lining the lumina of mature ducts and are polarized with respect to ori- entation of golgi and the presence of abundant micro-
villi on the free surfaces. Typical junctional complexes were found at the apical borders.
The question of whether these deep cells of the end bud body might derive from cap cells was investigated by time-lapse microcinematography. End buds isolated by collagenase digestion were incubated in petri dishes which had been coated with silicone to prevent attach- ment and spreading. Under these conditions the general morphology of the end bud was maintained for as much as 2 days. When the tip region was monitored with phase-contrast optics the video image indicated that cap cells showed considerable movement, with rapid ex-
FIG. 5. TEM of end buds. (A) Section through the tip region of an advancing end bud. Mesenchyme cells (m) are separated from the cap cells (cp) by a 104-nm-thick basal lamina (arrowheads). Lipid vesicles are present in the cap cells (*) (2600X). (B) No specialized cell junctions are seen between cap cells. Intercellular spaces with intruding cell processes are typically present (*) (8040X). (C) Luminal cells show typical junctional complexes (arrows), extensive membrane interdigitation, and luminal microvilli (8040X). Occasional adhesive junctions are visible at higher magnification (not shown).
282 DEVELOPMENTALBIOLOGY VOLUME 97,1983

WILLIAMS AND DANIEL Mammary Lhctd Elongation 283
bd .
FIG. 8. A composite drawing made from light and electron microscope analysis. At the top of the drawing stromal components are represented. Adipocytes (a) abut against cap cells at the tip (left). Fibrous components and fibrocytes (f) comprise the connective tissue tunic around the neck region. The basal lamina (bl) is represented as cut away to expose the underlying cap cells (cp). Cap cells are cuboidal but become progressively flattened toward the mid region of the end bud, differentiate into and are continuous with myoepithelial cells (mc) in the neck region. The basal lamina overlying myoepithelial cells in the mid region (bl’) is 14 times thicker than that at the tip (see Figs. llc and 12B). Mitosis is seen in the cap and body cells (bd). Cells which border the fluid filled lumen have a prominent microvilli.
tension and retraction of cell processes. Analysis of stop- motion frames revealed that certain cap cells appeared to migrate from their peripheral location into the deeper regions of the end bud, where they either moved out of the plane of focus or could no longer be followed due to lack of contrast with surrounding cells (Figs. 6A, B). Penetration of cap cells into deeper regions of the end bud was also suggested in l-pm-thick sections of plastic- embedded material, in which toluidine blue staining provided contrast between cap and body cells (Fig. SC).
Cap Cells ~krentiate into Mgoepithelium
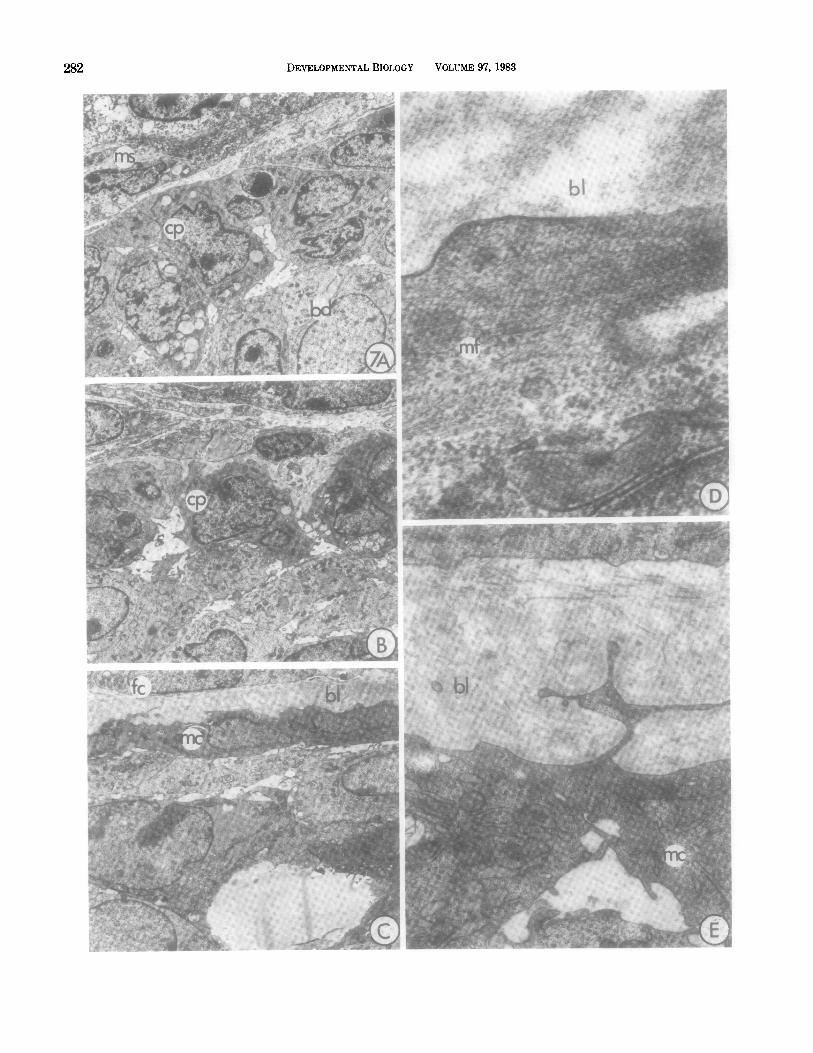
The cap cell layer (Fig. 7A) is clearly continuous with the ductal myoepithelial cells, which form the outer- most epithelial layer of mature ducts. In the end bud tip the cap layer consists of low columnar cells which become progressively cuboidal (Fig. 7B) and finally spindle shaped (Fig. 7C) as the contour of the end bud is followed to its lateral sides. These progressive changes can also be traced in l-pm sections of plastic-embedded
material (Fig. 6C), and are summarized diagramatically in Fig. 8. In the lateral regions of the end bud the cap cell cytoplasm becomes increasingly electron dense with abundant free ribosomes and moderate rough endo- plasmic reticulum. As differentiation proceeds, the en- doplasmic reticulum become dilated (Fig. 7E). In the constricted neck region these cells become distinctly myoepithelial, with peripheral cytoplasmic caveoli, ser- rated basal borders, and irregular nuclei containing conspicuous peripheral heterochromatin (Figs. 7C, E). Bundles of cytoplasmic filaments, the presumptive myo- filaments, begin to appear in the neck region after the cells have completed their shape changes (Fig. 7D).
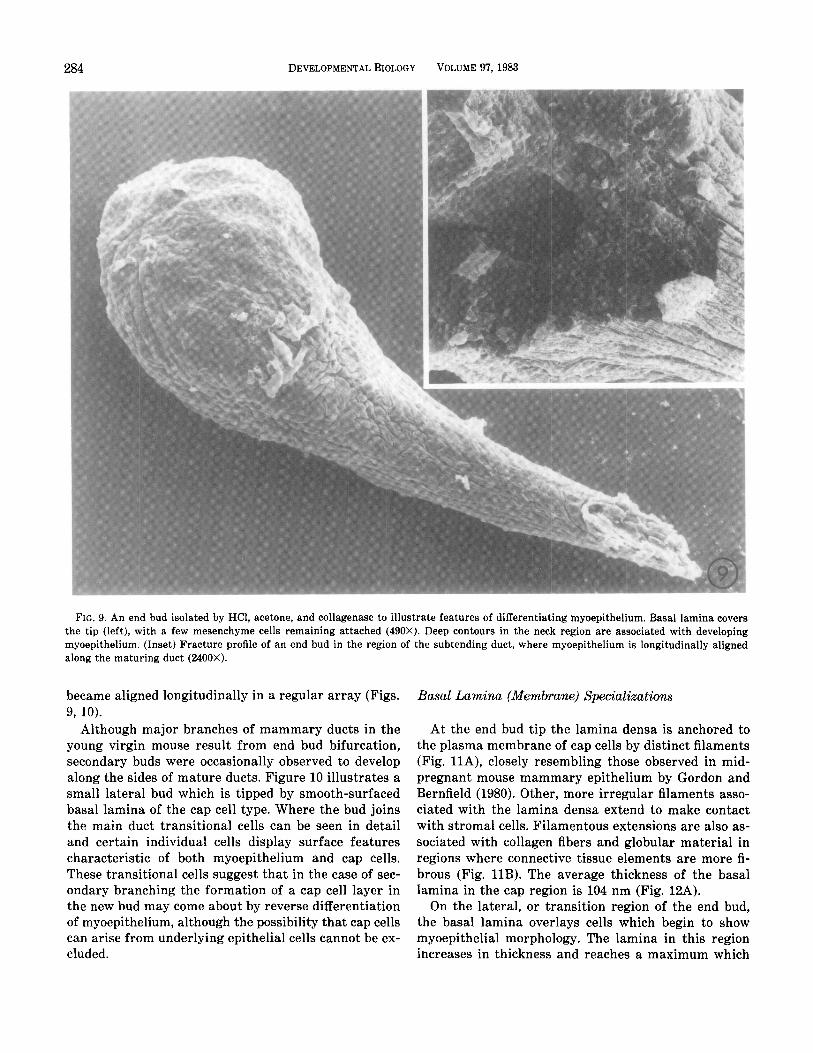
The progressive differentiation of ductal myoepi- thelium was also examined by scanning electron mi- croscopy of specimens which had been freed of connec- tive tissue (Fig. 9). Basal lamina in the tip region had a pebbled appearance, reflecting the shape of underlying cap cells. In the neck area the transitional cells were convoluted, with deep folds and irregular margins. In the region of differentiated duct the myoepithelial cells
FIG. 7. Progressive changes in cap cells (cp) which have an undifferentiated character in the tip region (A), but in the end bud flanks (B) begin to develop irregular surface features and electron-dense cytoplasm. Toward the neck, in the region of the subtending duct (C), they have developed many of the characteristics of mature myoepithelial cells (mc). At higher magnification other features of transition stages are visible. (D, E) including an increased density of cytoplasmic organelles, the gradual accumulation of cytoplasmic myofilaments (mf), dilated endoplasmic reticulum, electron-dense cytoplasm, and extensively folded basal lamina. ms, mesenchyme cell; bl, basal lamina; fc, fibrocyte; bd, body cell (A-C, 2500X; D, 76,500X; E, 18,760X’).
284 DEVELOPMENTAL BIOLOGY VOLUME 97,1983
FIG. 9. An end bud isolated by HCl, acetone, and collagenase to illustrate features of differentiating myoepithelium. Basal lamina covers the tip (left), with a few mesenchyme cells remaining attached (490X). Deep contours in the neck region are associated with developing myoepithelium. (Inset) Fracture profile of an end bud in the region of the subtending duct, where myoepithelium is longitudinally aligned along the maturing duct (2400X).
became aligned longitudinally in a regular array (Figs. 9, 10).
Although major branches of mammary duets in the young virgin mouse result from end bud bifurcation, secondary buds were occasionally observed to develop along the sides of mature ducts. Figure 10 illustrates a small lateral bud which is tipped by smooth-surfaced basal lamina of the cap cell type. Where the bud joins the main duct transitional cells can be seen in detail and certain individual cells display surface features characteristic of both myoepithelium and cap cells. These transitional cells suggest that in the case of sec- ondary branching the formation of a cap cell layer in the new bud may come about by reverse differentiation of myoepithelium, although the possibility that cap cells can arise from underlying epithelial cells cannot be ex- cluded.
Basal Lamina (Membrane) Specializations
At the end bud tip the lamina densa is anchored to the plasma membrane of cap cells by distinct filaments (Fig. llA), closely resembling those observed in mid- pregnant mouse mammary epithelium by Gordon and Bernfield (1980). Other, more irregular filaments asso- ciated with the lamina densa extend to make contact with stromal cells. Filamentous extensions are also as- sociated with collagen fibers and globular material in regions where connective tissue elements are more fi- brous (Fig. 11B). The average thickness of the basal lamina in the cap region is 104 nm (Fig. 12A).
On the lateral, or transition region of the end bud, the basal lamina overlays cells which begin to show myoepithelial morphology. The lamina in this region increases in thickness and reaches a maximum which
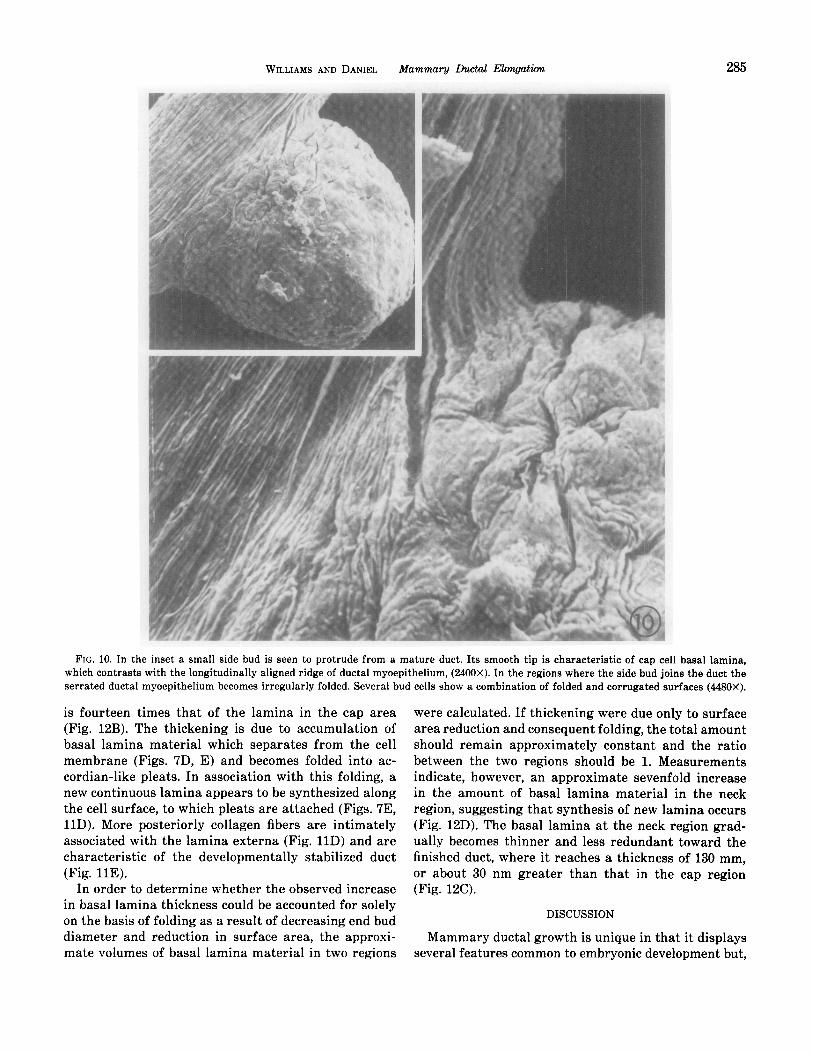
WILLIAMS AND DANIEL Mammary Ductal Elongation 285
FIG. 10. In the inset a small side bud is seen to protrude from a mature duct. Its smooth tip is characteristic of cap cell basal lam ina, which contrasts with the longitudinally aligned ridge of ductal myoepithelium, (2400X). In the regions where the side bud joins the duct the serrat ed ductal myoepithelium becomes irregularly folded. Several bud cells show a combination of folded and corrugated surfaces (448~ OX).
is fourteen times that of the lamina in the cap area (Fig. 12B). The thickening is due to accumulation of basal lamina material which separates from the cell membrane (Figs. 7D, E) and becomes folded into ac- cordian-like pleats. In association with this folding, a new continuous lamina appears to be synthesized along the cell surface, to which pleats are attached (Figs. 7E, 11D). More posteriorly collagen fibers are intimately associated with the lamina externa (Fig. 11D) and are characteristic of the developmentally stabilized duct (Fig. 11E).
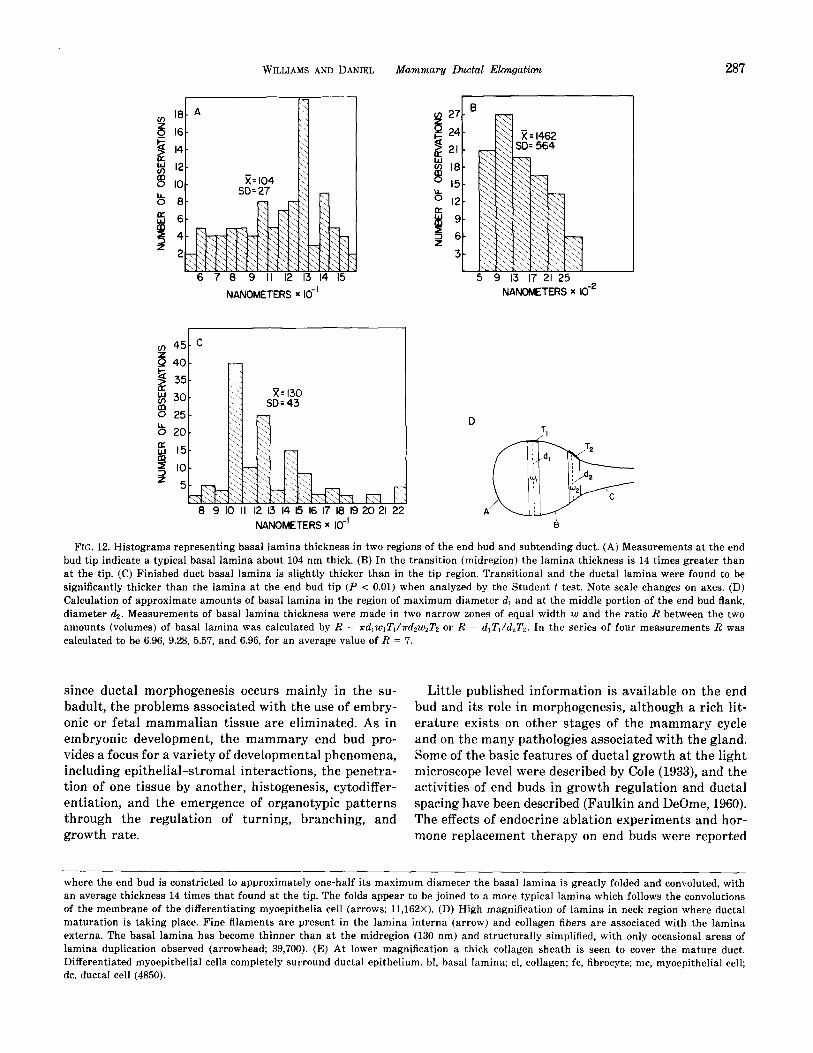
In order to determine whether the observed increase in basal lamina thickness could be accounted for solely on the basis of folding as a result of decreasing end bud diameter and reduction in surface area, the approxi- mate volumes of basal lamina material in two regions
were calculated. If thickening were due only to surface area reduction and consequent folding, the total amount should remain approximately constant and the ratio between the two regions should be 1. Measurements indicate, however, an approximate sevenfold increase in the amount of basal lamina material in the neck region, suggesting that synthesis of new lamina occurs (Fig. 12D). The basal lamina at the neck region grad- ually becomes thinner and less redundant toward the finished duct, where it reaches a thickness of 130 mm, or about 30 nm greater than that in the cap region (Fig. 12C).
DISCUSSION
Mammary ductal growth is unique in that it displays several features common to embryonic development but,
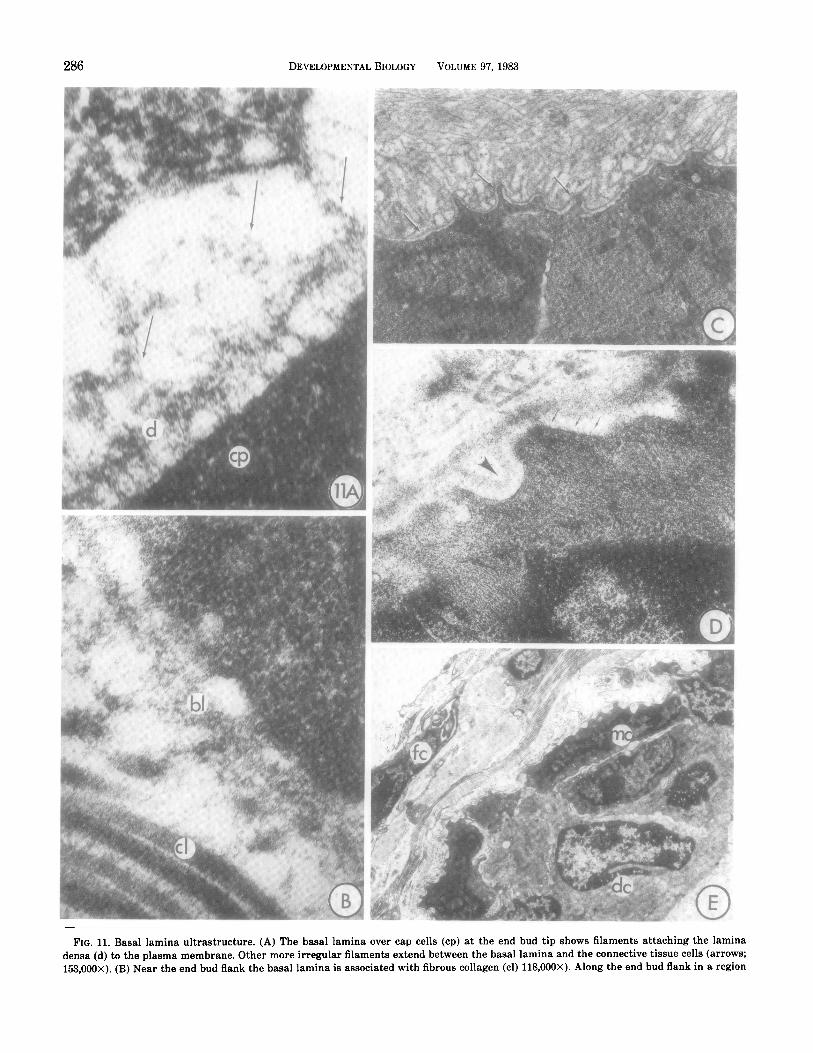
286 DEVELOPMENTAL BIOLOGY VOLUME 9'7,1983
FIG. 11. Basal lamina ultrastructure. (A) The basal lamina over cap cells (cp) at the end bud tip shows filaments attaching the lamina densa (d) to the plasma membrane. Other more irregular filaments extend between the basal lamina and the connective tissue cells (arrows; 153,000X). (B) Near the end bud flank the basal lamina is associated with fibrous collagen (cl) 118,000X). Along the end bud flank in a region
WILLIAMS AND DANIEL
6 7 8 9 II 12 13 14 I5
NANOMETERS x IO-’
Mammary Ductal Elongation 287
m 27- 6 t 24-
2 21-
: 18.
g l5-
b l2-
8 g- I 6- Z
3 t
NANCMETERS x d2
NANOMETERS x IO-’
FIG. 12. Histograms representing basal lamina thickness in two regions of the end bud and subtending duct. (A) Measurements at the end bud tip indicate a typical basal lamina about 104 nm thick. (B) In the transition (midregion) the lamina thickness is 14 times greater than at the tip. (C) Finished duct basal lamina is slightly thicker than in the tip region. Transitional and the ductal lamina were found to be significantly thicker than the lamina at the end bud tip (P < 0.01) when analyzed by the Student t test. Note scale changes on axes. (D) Calculation of approximate amounts of basal lamina in the region of maximum diameter dI and at the middle portion of the end bud flank, diameter 4. Measurements of basal lamina thickness were made in two narrow zones of equal width w and the ratio R between the two amounts (volumes) of basal lamina was calculated by R = K~~w~Z’~/K&UJ’~ or R = d,Z’,/&T,. In the series of four measurements R was calculated to be 6.96, 9.28, 5.57, and 6.96, for an average value of R = 7.
since ductal morphogenesis occurs mainly in the su- badult, the problems associated with the use of embry- onic or fetal mammalian tissue are eliminated. As in embryonic development, the mammary end bud pro- vides a focus for a variety of developmental phenomena, including epithelial-stromal interactions, the penetra- tion of one tissue by another, histogenesis, cytodiffer- entiation, and the emergence of organotypic patterns through the regulation of turning, branching, and growth rate.
Little published information is available on the end bud and its role in morphogenesis, although a rich lit- erature exists on other stages of the mammary cycle and on the many pathologies associated with the gland. Some of the basic features of ductal growth at the light microscope level were described by Cole (1933), and the activities of end buds in growth regulation and ductal spacing have been described (Faulkin and DeOme, 1960). The effects of endocrine ablation experiments and hor- mone replacement therapy on end buds were reported
where the end bud is constricted to approximately one-half its maximum diameter the basal lamina is greatly folded and convoluted, with an average thickness 14 times that found at the tip. The folds appear to be joined to a more typical lamina which follows the convolutions of the membrane of the differentiating myoepithelia cell (arrows; 11,162X). (D) High magnification of lamina in neck region where ductal maturation is taking place. Fine filaments are present in the lamina interna (arrow) and collagen fibers are associated with the lamina externa. The basal lamina has become thinner than at the midregion (130 nm) and structurally simplified, with only occasional areas of lamina duplication observed (arrowhead; 39,700). (E) At lower magnification a thick collagen sheath is seen to cover the mature duct. Differentiated myoepithelial cells completely surround ductal epithelium. bl, basal lamina; cl, collagen; fc, fibrocyte; me, myoepithelial cell; de, ductal cell (4850).

288 DEVELOPMENTALBIOLOGY VOLUME 97,1983
(Nandi, 19.58), DNA synthesis and cell cycle character- istics have been studied (Bresciani, 1965; Russo and Russo, 1978,198O; Russo et al, 1979). More recently the role of the extracellular matrix and the basal lamina in the mammary gland have received attention (Gordon and Bernfield, 1980; Silberstein and Daniel, 1982). The present paper is the first detailed report of cellular and structural features of the mammary end bud and their relation to morphogenetic activity.
Cap Cells
We have identified a layer of cuboidal epithelial cells at the end bud tip which are considered to be a stem cell population, on the basis of the following evidence: (1) Cap cells are undifferentiated by morphological cri- teria, and in contrast to previously described mammary cells, are loosely arranged, without specialized intra- cellular junctions for adhesion or communication. (2) They are continuously proliferating (Daniel, 1975). (3) The cap cell layer is continuous with the myoepithelium surrounding the subtending duct, and both are distinct from the compact epithelium of the end bud medulla or from luminal cells of the duct. (4) Cap cells appear to migrate to deeper regions of the end bud, but no evidence is available as to whether they subsequently differentiate and contribute to the population of mam- mary ductal-type cells.
Morphological evidence for the cap cell origin of duc- tal myoepithelium is supported by the observation that intermediate forms can be distinguished. Cuboidal or low columnar cap cells gradually become altered into cells with highly irregular surface features in the flank region, and in the neck assume a slender, spindle shape aligned longitudinally along the maturing duct. These transitional cells displayed a gradual and progressive increase in cytological markers of myoepithelial cell dif- ferentiation including desmosomes, membrane caveoli, ridges, and finally myofilaments. If myoepithelial cells were to differentiate in the medullary regions of the end bud and migrate subsequently to the surface it is most unlikely that such a linear accumulation of dif- ferentiated characteristics along the cortex would be observed. These results cannot exclude the possibility of some exchange of myoepithelial cells between cor- tical and deep layers as suggested by Radnor (1972), and indeed electron-dense cells resembling myoepi- thelium are occasionally found throughout the duct. There seems little doubt, however, that cap cells are progenitors to myoepithelium.
Although we cannot be certain that cap cells con- tribute to medullary cells of the end bud and ultimately, to luminal cells of the duct, such a lineage is suggested by both microscopy and time-lapse observations which
indicate that after cell division some daughter cells from the cap layer may migrate into the end bud medulla. These observations cannot be carried out in vivo and it is possible that they were in some way related to the end bud’s response to conditions of culture (Richards et aZ., 1982). Nor was it possible to follow such processes as differentiation of microvilli or formation of junc- tional complexes in cells which migrated. Nonetheless there is precedence for postulating a pluripotent mam- mary stem cell. Bennett et al. (1978) have described three different types of cells in clones of Rama 25 cells, a line originally derived from a DMBA-induced rat mammary tumor. These cuboidal cells have been shown to undergo differentiation in monlayer culture into elongated and droplet cells with myoepithelial and secretory charac- teristics, respectively, and in collagen gel cultures Rama 25 cells reconstitute a system of branching tubules (Bennett, 1980). Only speculative notions could be ad- vanced concerning the nature of these cuboidal precur- sor cells, but Bennett et al. (1978) and Rudland et al. (1980) postulated the existence of pluripotent progen- itor cells in both embryonic and end bud tissues of nor- mal gland.
In addition to providing a source of ductal cells it is likely that the cap layer plays an important role in growth regulation and pattern formation. The mam- mary end bud is highly responsive to its local tissue environment, and since the cap cells are separated from stroma only by the basement membrane, end bud re- sponses to chemical or physical cues probably involve modulation of cap cell activities which ultimately affect the direction, pattern, and rate of growth. It is signif- icant that a discrete cap cell layer cannot be distin- guished (unpublished observations) in ductal tips in which growth is terminated either by lack of available stroma or by aging in response to serial transplantation (Daniel et aL, 1968).
Basal Lam&a Specialixaticms
In considering differences between the basal lamina of end buds and the alveoli it should be taken into ac- count that the lamina of alveolar tissue may be the product of mammary secretory cells themselves, since the alveolar surface is only partially covered by myoepi- thelium (Murakami et aZ., 1981). In contrast, ductal cells in the major branches of mammary duets are not in contact with the basal lamina, which is synthesized ei- ther by cap cells at the end bud tip or posteriorly, by the confluent population of myoepithelium which sur- rounds the mature ducts (Warburton et aL, 1982). There is evidence in the rat that both cell types are capable of synthesizing basement membrane components (Kid- well et al, 1980; Liotta et aL, 1979).

WILLIAMS AND DANIEL Mammwy Lhctal Elongation 289
We have shown that the basal lamina displays strik- ing heterogeneity in structure and composition. At the advancing tip the cap cells produce a basal lamina 104 nm in thickness, which consists of periodically arranged stalks in the lamina interna and anionic globular ma- terial in the lamina externa. These structural features are similar to those described in alveoli of pregnant mammary gland (Gordon and Bernfield, 1980) and in embryonic epithelium undergoing branching and fold- ing (Cohn et aL, 1977). Autoradiographic studies of the cap cell basal lamina indicate that hyaluronic acid is synthesized while relatively little sulfated GAG is formed (Silberstein and Daniel, 1982b). We suggest that the chemical and physical properties of this cap cell lamina are adapted both for rapid turnover associated with growth and for penetration of stroma, where the hyaluronate component of the advancing edge may function to prevent adhesions and reduce frictional re- sistance. This interpretation is supported by the specific observation that during microdissection the end bud tip separates easily from surrounding stroma, and by the more general finding that in embryos hyaluronate is frequently associated with cell mobility (Toole et aZ., 1977).
Along the constricting end bud flanks the basal lam- ina is extraordinarily hypertrophied. Several lines of evidence indicate that this extracellular material is of basal lamina type and of epithelial origin: (1) The excess material is not amorphous and usually takes the form of folded sheets. These sheets are of approximately the same thickness as the lamina densa of the basal lamina attached to the epithelial cells, and in many cases are continuous with it (Figs. 7D, E, llC, D). (2) The hy- pertrophied material has similar ruthenium red stain- ing characteristics as the cell-associated basal lamina, and stains similarly with alcian blue (Silberstein and Daniel, 1982b). (3) Thickening and folding of other basal laminae have been reported in association with diabetes (Vracko, 1978), gynecomastia (Hassan and Olaizola, 1979), malignant mammary tumors (Pitelka et aL, 1980), tumors of Drosophila imaginal discs (Williams, 1980), and benign lesions (Jao et aL, 1978). In normal tissues extensive lamina folding has been reported in the re- gressing mammary gland after weaning (Pitelka and Hamamoto, 1977), and less dramatic hypertrophy of the basal lamina is found in the dental basement membrane of embryonic mice (Meyer et aL, 1981) in the basement membrane of the human apical ectodermal ridge (Kel- ley and Fallon, 1976), and in development of the mouse salivary gland (Bernfield and Wessels, 1970). To our knowledge, however, there are not previous reports of basal laminae of these unusual dimensions (almost 1.5 pm) or configurations associated with normal devel- opmental processes.
Although some of the basal lamina folding appears due to a reduction of surface area, our calculations (Fig. 12D) indicate that synthesis is also required. This is consistent with the high level of 35S04 incorporation into the lamina in this region (Silberstein and Daniel, 1982b). It is reasonable to suppose that the presence of a greatly thickened, anionic lamina may influence the microenvironment of underlying cells. Altered concen- trations of cations such as Ca2+ could affect the poly- merization of cytoskeletal elements in the differentiat- ing myoepithelium, as example. While answers to these problems await further investigation, the mammary end bud presents interesting opportunities for study of the functional and morphogenetic role of the basal lamina.
We appreciate the excellent technical assistance given by Terry Brown and Phyllis Strickland toward this project. We also thank Kay Gillis for illustrating the end bud. Many thanks to Jan Nowell for her instruction and critiques of electron microscopy. Thanks to Mar- shall Silvan for his suggestions on quantitative aspects of this project. This work was supported by PHS Grant AG 01050 from the National Institute on Aging.
REFERENCES
BENNETT, D. C. (1980). Morphogenesis of branching tubules in culture of cloned mammary epithelial cells. Nature (Lcmdon) 285, 657-659.
BENNETT, D. C., PEACHEY, L. A., DURBIN, H., and RUDLAND, P. S. (1978). A possible mammary stem cell line. CeU 15, 283-298.
BERNFIELD, M. R., and WESSELS, N. K. (1970). Intra- and extracellular control of epithelial morphogenesis. Den Biol (Suppl.) 4, 195-249.
BRESCIANI, F. (1965). Effect of ovarian hormones on duration of DNA synthesis in cells of the C3H mouse mammary gland. Exp. Cell Res. 48, 13-82.
COHN, R. M., BANERJEE, S. D., and BERNFIELD, M. R. (1977). Basal lamina of embryonic salivary epithelia. Nature of glycosaminogly- cans and organization of extracellular materials. J. Cell. Biol 73, 464-478.
COLE, H. 0. (1933). The mammary gland of the mouse during the estrous cycle, pregnancy and lactation. Proc. Roy. Sot. B 114, 136- 161.
DANIEL, C. W. (1975). Regulation of cell division in aging mouse mam- mary epithelium. Advan Exp. Med Biol. 52, 1-19.
DANIEL, D. W., DEOME, K. B., YOUNG, J. T., BLAIR, P. B., and FAIJLKIN, L. J. (1968). The in vivo life span of normal and preneoplastic mouse mammary glands: A serial transplantation study. Proc. Nat. Acad. Sci. USA 61,53-60.
EVAN, A. P., DAIL, W. G., DAMMROSE, D., and PALMER, C. (1976). Scan- ning electron microscopy of cell surfaces following removal of ex- tracellular material. Anat Res. 185, 433-446.
FAULKINS, L. J., and DEOME, K. B. (1960). Regulation of growth and spacing of gland elements in the mammary fat pad of the C3H mouse. J. Nat. Cancer Inst. 24, 953-963.
GORDON, J. R.. and BERNFIELD, M. R. (1980). The basal lamina of the postnatal mammary epithelium contains glycosaminoglycan in a precise ultrastructural organization. Dev. Biol 74, 118-135.
HASSAN, M. O., and OLAIZOLA, M. Y. (1979). Ultrastructural obser- vations on gynecomastia. Arch. Patkol. Lab. Med. 103. 624-630.
JAO, W., VASQUEZ, T., KEH, P. C., and GOULD, V. E. (1978). Myoepi- thelial differentiation and basal lamina deposition in tibroadenoma and adenosis of the breast. J. Pathol. 126, 107-112.

290 DEVELOPMENTAL BIOLOGY VOLUME 97,1983
KELLY, R. O., and FALLON, J. F. (1976). Ultrastructural analysis of the apical ectodermal ridge during vertebrate limb morphogenesis. Dev. Biol. 51,241-256.
KIDWELL, W. R., WICHA, M. S., SALOMON, D., and LIO~A, L. A. (1980). Hormonal controls of collagen substratum formation by cultured mammary cells: implications for growth and differentiation. In “Control Mechanisms in Animal Cells” (L. Jimenez de Asva, et a& eds.), pp. 333-340. Raven Press, New York.
LIO~A, L. A., WICHA, M. S., FIODART, J. M., RENNARD, S. I., GARBISA, S., and KIDWELL, W. R. (1979). Hormonal requirements for base- ment membrane collagen deposition by cultured rat mammary ep- ithelium. Lob. Invest. 41, 511-518.
MEYER, J. M., STAUBLI, A., and RUCH, J. V. (1981). Ruthenium red staining and tannic acid fixation of dental basement membrane. Cell Tissue Res. 220.589-59’7.
MURAKAMI, M., SUGITO, A., ABE, J., HAMASAKI, M., and SHIMOKA, T. (1981). SEM observations of some exocrine glands, with special ref- erence to configuration of the associated myoepithelial cells. Biomed. Res. 2(Suppl.), 96-102.
NANDI, S. (1958). Endocrine control of mammary gland development and function in the C3H/He Crgl mouse. J. Nat. Cancer Inst. 21, 1039-1055.
PITELKA, D. R., and HAMAMOTO, S. T. (1977). Form and function in mammary epithelium: the interpretation of ultrastructure. J. Dairy Sci. 60, 643-654.
PITELKA, D. R., HAMAMOTO, S. J., and TAGGART, B. N. (1980). Basal lamina and tissue recognition in malignant mammary tumors. Can- cer Res. 40, 1600-1611.
RADNOR, C. P. (1972). Myoepithelial cell differentiation in rat mam- mary glands. .Z. Anut. 111, 381-398.
RICHARDS, J., GUZMAN, R., KONRAD, M., YANG, J., and NANDI, S. (1982). Growth of mouse mammary gland end buds cultured in a collagen matrix. Exp. Cell Res. 141, 433-443.
RUDLAND, P. S., ORMEROD, E. J., and PATERSON, F. G. (1980). Stem cells in rat mammary development and cancer: a review. J. Roy. Sot. Med 73, 437-442.
Russo, I. H., and Russo, J. (1978). Developmental stage of the rat
mammary gland as determinant of its susceptibility to 7,12-di- methylben$a]anthracene. J. Nat. Cancer Inst. 61,1439-1449.
Russo, J., and Russo, I. H. (1980). Influence of differentiation and cell kinetics on the susceptibility of the rat mammary gland to carci- nogenesis. Cancer Res. 40, 2677-2687.
Russo, J., WILGUS, G., and Russo, I. H. (1979). Susceptibility of the mammary gland to carcinogenesis. I. Differentiation of the mam- mary gland as a determinant of tumor incidence and type of lesion. Amer. J. PathoL 96, 721-735.
SCHNEEBERGER-KEELEY, E. E., and KARNOVSKY, M. J. (1968). The ul- trastructural basis of alveolar capillary membrane permeability to peroxidase used as a tracer. J. CeU Biol. 37, 781-793.
SILBERSTEIN, G. B., and DANIEL, C. W. (1982a). Elvax 40P Implants: Sustained, local release of bioactive materials influencing mam- mary ductal development. Dev. Biol. 93,272-278.
SILBERSTEIN, G. B., and DANIEL, C. W. (1982b). Glycosaminoglycans in the basal lamina and extracellular matrix of the developing mouse mammary duct. Dev. Biol 90, 215-222.
TOOLE, B. P., OKAYAMA, M., ORKIN, R. W., YASHIMURA, M., MUTO, M., and NAJI, A. (1977). Developmental rates of hyaluronate and chon- droitin sulfate proteoglycans. In “Cell and Tissue Interaction” (Lash, J. W. and Burger, M. M., eds.), pp. 139-154. Raven Press, New York.
TRELSTAD, R. L., HAYASHI, K., and TOOLE, B. P. (1974). Epithelial collagen and glycosaminoglycans in the embryonic cornea: Molec- ular order and morphogenesis in the basement membrane. J. Cell Biol. 62, 815-830.
VRACKO, R. (1978). Effects of aging and diabetes on basal lamina thickness of six cell types. In “Biology and Chemistry of Basement Membranes” (N. A. Kefalides, ed.), pp. 483-493. Academic Press, New York.
WARBURTON, M. J., MITCHELL, D., ORMEROD, E. J., and RUDLAND, P. (1982). Distribution of myoepithelial cells and basement membrane proteins in the resting, pregnant, lactating, and involuting rat mammary gland. J. Hid&em. Cytochem, 30,667-676.
WILLIAMS, J. M. (1980). “A Study of Tumorigenesis in Imaginal Disc of Drosophila melanogaster Bearing the Temperature Sensitive Mu- tation shibire’“‘. M.A. Thesis, University of California, Santa Cruz.
Related Documents











