DAFTAR ISI KATA PENGANTAR.........................................1 DAFTAR ISI.............................................2 BAB I Pendahuluan 1.1 Latar Belakang................................3 1.2 Rumusan Masalah...............................4 1.3 Tujuan .......................................4 BAB II Pembahasan 2.1 Penjelasan Respirasi Tumbuhan.................5 2.2 Reaksi Glikolisis.............................8 2.3 Lintasan Pentosa Fosfat Oksidatif (Ppo).......14 2.4 Siklus Asam Sitrat............................16 2.5 Respirasi Tumbuhan dan Jaringan...............31 2.6 Metabolisme Lemak.............................35 BAB III Penutup Simpulan..........................................37 Daftar Pustaka.........................................38 1

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

DAFTAR ISI
KATA PENGANTAR.........................................1
DAFTAR ISI.............................................2
BAB I Pendahuluan
1.1 Latar Belakang................................3
1.2 Rumusan Masalah...............................4
1.3 Tujuan .......................................4
BAB II Pembahasan
2.1 Penjelasan Respirasi Tumbuhan.................5
2.2 Reaksi Glikolisis.............................8
2.3 Lintasan Pentosa Fosfat Oksidatif (Ppo).......14
2.4 Siklus Asam Sitrat............................16
2.5 Respirasi Tumbuhan dan Jaringan...............31
2.6 Metabolisme Lemak.............................35
BAB III Penutup
Simpulan..........................................37
Daftar Pustaka.........................................38
1

BAB I
PENDAHULUAN
1.1 Latar Belakang
Dalam pengertian sehari-hari, bernafas sekedar
diartikan sebagai proses pertukaran gas di paru-paru.
Tetapi secara biologis, pengertian respirasi tidaklah
demikian. Pernafasan lebih menunjuk kepada proses
pembongkaran atau pembakaran zat sumber energi di dalam
sel-sel tubuh untuk memperoleh energi atau tenaga. Zat
makanan sumber tenaga yang paling utama adalah
karbohidrat.
Dari berbagai cabang ilmu tumbuhan yang telah
berdiri sendiri salah satunya adalah fisiologi
tumbuhan. Fisiologi tumbuhan yang mempelajari peri
kehidupan tumbuhan sudah demikian pesat berkembangnya
juga didukung oleh beberapa ilmu seperti anatomi
tumbuhan, morfologi tumbuhan, dan sistematika tumbhan.
Fisiologi tumbuhan itu sendiri merupakan ilmu yang
mempelajari atau mencari keterangan-keterangan mengenai
kehidupan tumbuhan. Untuk mempertahankan kehidupannya,
tumbuhan perlu mempunyai suatu penyediaan energi yang
berkesinambungan. Energi-energi tersebut diperoleh dari
mengambil energi kimia yang terbentuk dalam molekul
organik yang disintesis oleh fotosintesis. Suatu proses
pelepasan energi yang menyeddiakan energi bagi
keperluan sel itu diseebut dengan respirasi. Respirasi
2

sel tumbuhan berupa oksidasi molekul organik oleh
oksigen dari udara membentuk karbon dioksida dan air.
Tumbuhan juga menyerap O2 untuk pernafasannya,
umumnya diserap melalui daun (stomata). Pada keadaan
aerob, tumbuhan melakukan respirasi aerob. Bila dalam
keadaan anaerob atau kurang oksigen, jaringan melakukan
respirasi secara anaerob. Misal pada akar yang
tergenang air. Pada respirasi aerob, terjadi pembakaran
(oksidasi) zat gula (glukosa) secara sempurna, sehingga
menghasilkan energi jauh lebih besar (36 ATP) daripada
respirasi anaerob (2 ATP saja). Demikian pula respirasi
yang terjadi pada jazad renik (mikroorganisma).
Sebagian mikroorgaanisma melakukan respirasi aerobik
(dengan zat asam), anerobik (tanpa zat asam) atau cara
keduanya (aerobik fakultatif)
1.2 Rumusan Masalah
a. Apa Penjelasan Respirasi Tumbuhan?
b. Bagaimana Reaksi Glikolisis?
c. Bagaimana Lintasan Pentosa Fosfat Oksidatif (Ppo)?
d. Bagaimana Siklus Asam Sitrat?
e. Bagaimana Respirasi pada Tumbuhan?
f. Apa Penjelasan Metabolisme Lemak?
1.3 Tujuan
a. Untuk Mengetahui Penjelasan Respirasi Tumbuhan
b. Untuk Mengetahui Reaksi Glikolisis
3

c. Untuk Mengetahui Lintasan Pentosa Fosfat Oksidatif
(Ppo)
d. Untuk Mengetahui Siklus Asam Sitrat
e. Untuk Mengetahui Respirasai Pada Tumbuhan
f. Untuk Mengetahui Penjelasan Metabolisme Lemak
4

BAB II
PEMBAHASAN
2.1 Penjelasan Respirasi Tumbuhan
Respirasi adalah proses penguraian bahan makanan
yang menghasilkan energi. Fotosintesis menyediakan
molekul organik yang dibutuhkan oleh tumbuhan dan
makhluk hidup lainnya. Respirasi dan metabolisme karbon
yang terkait di dalamnya melepas energi yang tersimpan
didalam senyawa karbon dengan cara yang terkontrol
untuk digunakan sel. Pada waktu yang bersamaan,
respirasi menghasilkan banyak senyawa karbon yang
dibutuhkan sebagi prekursor untuk biosintesis senyawa
organik lainnya. Respirasi aerob merupakan proses umum
yang tejadi dalam hampir semua organisme eukariot, dan
secara umum proses respirsi diddalam tumbuhan mirip
dengan apa yang dijumpai di dalam hewan dan eukariot
tingkat rendah, tetapi beberapa aspek khusus dari
respirasi tumbuhan membedakannnya dari respirasi hewan.
Respirasi aerob adalah proses biologi yang
memobilisasidan mengoksidasi molekul organik secara
terkontrol. Selama respirasi, energi bebas dilepas dan
disimpan sementaradalam bentuk ATP yang siap digunakan
untuk aktifitas sel dan perkembangan tumbuhan.
Glukosa adalah substrat respirasi yang umum
dikenal, tetapi dalm sel tumbuhan, substrat respirasi
berasal dari sukrosa, heksosa fosafat dan triosa fosfat
5

yang berasal dari fotosintesis dan perombakan pati,
fruktosa yang mengandung polimer, gula-gula lainnya,
lemak utamanya triasilgliserol, asam–asam organik dan
kadang-kadang protein (Gambar 4.1). dari segi reaksi
kimianya dapat dituliskan sebagi berikut:
C12H12O11+¿13H2O→ 12CO2+¿48H++¿48e-
12O2+¿48H++¿48e-→24H2O
Sehingga persamaan reaksi bersih dari reaksi kimia
diatas adalah sbagai berikut:
C12H12O11 +¿12O2 →12CO2+¿11H2O
Reaksi diatas adalah kebalikan dari reaksi
fotosintesis yang merupakan reaksi-reaksi redoks dimana
sukrosa dioksidasisecara sempurna menjadi CO2,
sementara
6

Gambar 4.1. respirasi. Substrat respirasi berasal
dari proses selular lainnya yang kemudian masuk dalam
lintasan respirasi. lintasn glikolisis dan pentosa
fosfat dalam sitosol dan plastida akan mengubah gula
menjadi asma organik, mel;alui reaksi heksosaa fosfat
dan triosa fosfat akan menghasilkan NADH atau NADPH dan
ATP. Asam organik dioksidasi dalam siklus asam sitrat
mitokondria. Pada glukoneogenesiskarbon, pada pemecahan
lipid di metabolism pada siklus asam sitrat yang
kemudian digunakan untuk mensintesis gula dalam
sitosol.
Oksigen yang berperan sebagi penerima (akseptor),
direduksi menjadi air. Energi bebas yang dilepasdari
7

reaksi tersebut sebesar 5760kJ (1280 kkal) permol (342)
sukrosa yang dioksida. Melepas energi bebas secara
terkontrol, bersama-sama dengan sintesis ATP,adalah
fungsi utama dari metabolisme respirasi
Untuk mencegah kerusakan struktur sel, pelepasn
energi bebas dari molekul sukrosa dilakukan oleh sel
secar bertahap. reaksi-reaksi bertahap tersebut dapat
dikelompokkan ke empat [roses utama yaitu: glikolisis,
siklus asam sitrat, reaksi-reaksi dari lintasn pentosa
fosfat dan fosforilasioksidatif. Substrat respuirasi
masuk proses respirasi pada titik-titik yang berbeda
dari lintasn- lintasan tersebut seperti pada Gambar
4.1.
1. Glikolisis
Merupakan suatu seri reaksi-reaksi yang melibatkan
suatu grup enzim yang terdapat di sitosol dan
plastida. Sebuah molekul gula, misalanya sukrosa,
dioksida secara parsial melalui pembentukan
heksosa fosfat (gula fosfat berkarbon enam) dan
triosa fosfat (gula fosfat berkarbon tiga) untuk
menghasilkan asam organik, seperti piruvat. Proses
menghasilkan sejumlah kecil energi dalam bentuk
ATP dan molekul pereduksi, NADH (Nikotinamid
adenin dinukleotida tereduksi).
2. Lintasan pentosa fosfat
8

Lintasn ini juga beraa dalam sitosol dan plastida.
Atom karbon dari glukosa 6 fosfat awalnya
dioksidasi menjadi ribulosa 5 fosfat, suatu
molekul organik yang memiliki 5 atom karbon. Atom
karbon kemudian hilang dalam bentuk CO2 dan dua
molekul pereduksi, dalam bentuk NADPH, dihasilkan.
3. Siklus asam sitrat
Daalm siklus ini perivat dioksidasi secara
sempurna menjadi CO2 dan dihasilkan banyak molekul
pereduksi (16 NADH +¿4 FADH2 untuk tiap molekul
sukrosa yang dioksidasi). Reaksi-reaksi dalam
siklus asam sitratmelibatkan enzim-enzim yang
terdapat didalam matrik mitokondria, kecuali
enzimsuksinat dehidrogenase yang terdapat pada
membran dalam mitokondria.
4. Fosforilasi oksidatif
Pada tahap ini elektron di pindahkan melalui suatu
rantai transport elektron yang terdiri dari
sekumpulan protein transport elektron yang
terdapat pada membran dalam mitokondria. Sistem
transport ini memindahkan elektron dari NADH (dan
molekul pereduksi sejenis) yang dihasilkan dari
glikolisis, lintasan pentosa fosfat, dan siklus
asam sitrat ke molekul oksigen. Pada saat
pemindahan elektron terjadi pembebasan energi
yang kemudian disimpan dalam bentuk ATP yang
dibentuk dari ADP dan Pi dengan katalisator ATP
9

sintase. Reaksi-reaksi redoks bersama-sama dengan
sintesis ATP disebut fosforilasi oksidatif.
Nikotinamid adenin dinukleotida (NAD+/NADH) adalah
suatu kofaktor organik (koenzim)yang berasosiasi dengan
banyak enzim yang mengkatasis reaksi-reaksi redoks di
dalam sel. NAD+ adalah bentuk kofaktor yang teroksidasi
dan Dapat menenerima dua elektron untuk membentuk NADH
seperti reaksi dibawah ini:
NAD++¿2e− +¿H+→ NADHReaksi diatas dapat berlangsung bolak-balik. Potensial
reduksi standar dari reaksi redoks diatas sekitar -320
mV, yang berarti NADH merupakan reduktan (pendonor
elektron) yang cukup kuat. Jadi NADH merupakan molekul
yang baik untuk menyimpan energi bebas yang dibawah
oleh elektron yang dilepas selama proses glikolisis dan
siklus asam sitrat. Oksidasi NADH oleh oksigen melalaui
rantai transport elektron melepas energi bebas (220 kJ
per mol, atau 52 kkal permol)dan dapt digunakan untuk
mensistesis ATP. Serkarang kita bisa formulasikan
reaksi respirasi sel dari satu molekul sukrosa dengan
melibattkan komponen enrgi sebagai berikut:
C12H12O11 +¿12O2 →12CO2+¿11 H2O
60 ADP +¿60 Pi →60ATP+¿60 H2O
2.2 Glikolisis
10

Pada tahap awal glikolisis (dari kata latin,
gykos= gula; lysiss= pemecaahan), karbohidrat diubah
menjadi heksosa fosfat, yang kemudian dipecah menjadi
dua molekul triosa fosfat. Selanjutnya kedua molekul
triosa fosfat tersebut dioksidasi menjadi dua molekul
piruvat. Disamping menyiapkan substrat untuk oksidasi
dalam siklus asam sitrat, glikolisis juga menghasilkan
sejumlah kecil ATP dan NADH
Ketika oksigen molekular tidak tersedia , seperti
akar tumbuhan yang terendam, glikolisisdapat menjadi
sumber energi utama bagi sel. Dalam kondisi seperti
ini, fermentasi yang berlangsung di dalam sitosol,
mendaur ulang NADH yang dihasilkan dari glikolisis
dengan cara mereduksi piruvat.
Glikolisis terjadi pada semua organisme (prokariot
dan eukariot). Secara prinsip reaksi-reaksi dalam
glikolisis dan fermentasi dalam tumbuhan hampir sama
dengan yang terjadi dalam sel hewan ( Gambar 4.2).
tetapi glikolisis pada tumbuhan memiliki mekanisme
pengaturan yang khas, berlangsung secara pararel antara
glikolisis sitosol dan plastida, serta jalur-jalur
alternatif dari reaksi-reaksi alternatif dari reaksi-
reaksi glikolisis di dalam sitosol.
11

Gambar 4.2. lintasan glikolisis
Pada hewan substrat utam respirasi adalh glukosa
dan produk akhirnya adalah piruvat. Pada tumbuhan,
sukrosa merupaka bentuk gula utama yang
ditranslokasikan dan oleh karenanya merupakan jenis
gula utam ayang diimpor oleh jaringan/ organ non
fotositesis, maka sukrosa yang di yakini merupakan
substrat utama respirasi tumbuan dan hasil akhirnya
tidak hanya piruvat , tetapi juga dapaat berupa asm
organik lainnya yaitu malat.
12

Pada tahap awal gl ikolisis, sukrosa dipecah
menjadi dua monosakarida yaitu glukosa dan fruktosa,
yang dapt segera masuk kelintasan glikolisis. Terdapat
dua lintasan yang memecah sukrosa didlam tumbuhan.
Pertama, pada sebagian besar jaringan tumbuhan, enzim
sukrosa sintase yang ada didalam sitosol, digunakan
unuk memecah sukrosa dengan cara menggabungkan sukrosa
dengan UDP menjadi fruktosa dan UDP gukosa dan
pirofosfat (PPi) menjadi UTP dan glukosa 6 fosfat
( Gambar 4.2). kedua pada beebrap jaringan tumbuhan ,
enxzim invertase yang terdapat pad dinding sel, vakuola
atau sistosol menghidrolisis sukrosa menjadi dua
heksosa, yaitu fruktosa dan glukosa, yang kemudian di
fosforilasi dalam suatu reaksi yang menggunakan ATP.
Plastida, seperti kloroplas dan amiloplas, dapat
juga menyediakan substarat untuk glikolisis. Pati
disintesis dan dipecahkan hanya didalam plastid.
Plastid mengubah pati menjadi triosa fosfat menggunakan
enzim –enzim glikolisis yang mengubah heksosa fosfat
menjadi triosa fosafat. Senyawa karbon yang diperoleh
dari pemecahan pati masuk kedalam lintasn glikolisis
didalam sitososl. Senyawa karbon tersebut terutama
daalm bentuk heksosa fosfat yang ditranslokasikan dari
amiloplas atau trosa fosafat yang ditranslokasikan dari
kloroplas. Hasil fotosintesis dapat juga langsung masuk
lintasn glikolisis dalm bentuk triosa fosfat.
13

Pada tahap awal glikolisis, tiap unit heksosa
difosforilasi dua kali dipecah menjadi dua molekul
triosa fosfat. Bergantung apakah pemecahan tersebut
menggunakan enzim sukrosa sintase atau invertase,
serangkaian reaksi pemecaahn tersebut menggunakan dua
sampai empat molekul ATP untuk tiap unit sukrosa.
Reaksi-reaksi tersebt juga melibatkan dua dari tiga
reaksi tidak dapat balik pentin dari lintasan
glikolisis yang dikatalis oleh enzim heksokinase dan
fosfofruktokinase (Gmabar 4.2). Reaksi
fosfofruktokinase adalah stu dari titik kendali
glikolisis baik pada tumbuhan maupun hewan.
Ketika molekul gliseraldehid 3 fosfat terbentuk,
lintasn glikolisis memulai memanen energi pada tahap
permanen energi dari glikolisis. Enzim gliseraldehid 3
fosfat dehidrogenase mengkatalis oksidasi aldehid untuk
memebntuk asasm karboksilat dan mereduksi NAD+menjadi
NADH. Reaksi tersebut melepas energi bebas dalam jumlah
cukup untukmelakukan fosforilasi gliseraldehid 3
fosfat mnggunakan fosfat inorganik dan membentuka 1,3
bisfosfogliserat. Molekul ini merupakan suatu pemberi
gugus fosfat yang cukup kuat karena memiliki standar
energi bebas untuk hidrolisis yang cukup tinggi(-49,3
kJ per mol atau -11,8 kkal per mol).
Pada tahap berikutnya gugus fosfat dari ataom
karbon ke 1 dari 1,3 bisfosfogliserat dipindahkan ke
14

molekul ADP untuk membentuk ATP dan molekul 3
fosfogliserat. Untuk tiap sukrosa yang masuk glikolisis
akan dihasilkan empat ATP dari reaksi ini. Sintesis ATP
dengan car tersebut disebut fosforilasi tingkat
substrat, yang melibatkan pemindahan langsung sebuah
gugus fosfat dari molekul substrat ke ADP untuk
membentuk ATP. Sintesis ATP melalui proses ini berbeda
mekanismenya dengan sintesis ATP melalui fosforilasi
oksidatif di mktokondria aatu sintesi ATP pada proses
fotosintesi.
Pada dua reaksi berikutnya, fosfat dari molekul 3
fosfogliserat dipindahkan keatom karbon ke dua dan
kemudian sebuah molekul air dilepas menghasilka senyawa
yang disebut fosfoenol piruvat (PEP). Gugus fosfat dari
PEP memiliki standar energi bebas yang tinggi (-61,9 kJ
per mol atau -14,8 kkal permol), sehingga PEP merupakan
suatu donor fosfat yang baik untuk membentuk ATP.
Dengan menggunakan PEP sebagai substrat, enzim piruvat
kinase memindahkan gugus fosfat dari PEP ke ADP untuk
memebentuk ATP dan piruvat. Tahap terakhir ini, yang
merupakan tahap tidak dapat balik ketiga yang penting
dalam glikolisis, menghasilkan empat molekul ATP untuk
tiap molekul sukrosa yang masuk kedalam lintasan
glikolisis.
Di dalam tumbuhan, selain PEP diubah ke bentuk
piruvat , PEP juga dapat mengalami karboksilasi untuk
15

membentuk asam organik oksaloasetat(OAA) dengan bantuan
enzim malat dehidrogenase dan menggunakan NADH. Malat
yang dihasilkan dapat disimpan didalam vakuola atau
ditransport ke mitokondria dan masuk kesiklus asam
sitrat.
2.2.1 Fermentasi berlangsung pada kondisi anaerob
Pada kondisi tidak ad oksigen sepertipad tumbuhan
yang tergenang, siklus asma sitrat dan fosforilai
oksidatif tidak dapat berlangsung. Glikolisis tidak
dapat terus berjalan karen asuplai NAD+terbatas, dan
apabial semua NAD+ telah direduksi menjadi NADH, mak
salah satu tahap glikolisis yang dikatalis oleh enzim
gliseraldehid 3 fosfat dehidrogenase tidak dapat
berlangsung. Unuk mengatasi masaalh ini, tumbuhan dan
organisme lain dapat melakukan metabolisme piruvat
dengan jalanlain, yang disebut fermentasi atau
metabolisme fermentatif.
Terdapat dua jenis metabolisme fermentatif, yaitu
fermentasi alkohol dan fermentasi asam laktat. Pada
fermentasi alkohol(umum terjadi didalam tumbuhan),
enzim piruvat dehidrogenae dan alkohol dehidrogenase
mengubah piruvat menjadi alkohol dan CO2 serta
mengoksidasi NADH. Pada fermentasi asam laktat (umum
terjadi pada sel otot hewan, tetapi juga dijumpai di
tumbuhan),enzim laktat dehidrogenase menggunakan NADH
untuk mereduksi piruvat menjadi laktat. Jadi kedua
16

fermentasi tersebut adalh proses meproduksi kembali
NAD+.
Pada jagung, respon awal terhadap kondisi oksigen
terbatas adalah terjadinya fermentasi asam laktat,
tetapi kemudian dilanjutkan dengan fermentasi alkohol.
Hal ini disebabkan alkohol merupakan senyawa yang
kurang toksik dan dapt berdifusi ke luar sel, sementara
asam laktat dapat terakumulasi dan mendorong terjadinya
kondisi asam di dalam sitosol.
Fermentasi tidak melepaskan semua energi yang
tersedia di dalam molekul gila, oleh karenanya
efisiensi fermentasi lebih rendah dibanding efisensi
respirasi aerob. Efisiensi didefinisikan sebagi jumlah
energi yang tersimpan dalam bentuk ATP relatif terhadap
potensi energi yang tersedia di dalam sebuah molekul
sukrosa. Perubahan energi bebas standar (∆G°) u ntuk
oksidsi sempurna dari sukrosa adlah -5760 kJ mol-1(1380
kkal mol-1). Nilai ∆G° untuk sintesis ATP adalh 32kJ
mol-1 (7,7 kkal mol-1). Akan tetapi dibawah kondisi yang
umumnya terrjadi didalam sel hewan atau tumbuhan,
sintesis ATP memerlukan masukan energi bebas kira-kira
sebesar 50 kJ mol-1(12 kkal mol-1). Misalkan ATP bersih
untuk tiap molekul sukrosa yang difermentasikan adalah
empat molekul ATP, maka efisiensi fermentasi hanya
sebesar 4%. Sebagian energi yang tersedia di dalam
molekul sukrosa masih tetapi dalam molekul laktat atau
17

etanol. Selama respirasi aerob, pirufat yang di
produksi dari glikolisis di transport kedalam
mitokondria dan menghasilkan lebih banyak energi dalam
bentuk ATP.
18

2.3 Lintasan Pentosa Fosfat Oksidatif (Ppo)
Glikolisis bukan satu-satunya lintasan oksidasi
gula di dalam sel tumbuhan. Lintasan PPO dapat
merupakan lintasna lain dari oksidasi gula (Gambar
4.3). lintasan ini berlangsung didalaam sitosol dan
plastida. Umumnya lintasan PPO di plastida lebih
dominan dibanding yang terjadi didalam sitosol.
Dari Gambar 4.3 terlihat bahwa dua reaksi pertam
dari lintasan PPO merupakan tahap oksidasi yang
mengubah glukosa 6 fosfat menjadi ribulosa 5 fosfat
menjadi intermediat glikolisis seperti gliseraldehid 3
fosfat dan fruktossa 6 fosaft. Karena glukosa 6 fosfat
dapat dibentuk kembali dari gliseraldehid 3 fosfat dan
fruktosa 6 fosfat oleh enzim –enzim glikolisis, maka
untuk enam kali siklus, reaksi lintasn Ppo dapat
dituliskan sebagai berikut:
6 glukosa 6 P + 12NADP+ + 7H2O →
5 glukosa 5 P + 6CO2 + Pi +
12NADPH + 12H+
Lintasan PPO memiliki beberapa peranan didalam
metabolisme tumbuhan, antara lain:
1. NADPH yang berhasil dapat digunakan dalam reaksi-
reaksi reduksi yang berjkaitan dengan reaksi-
reaksi biosintesis yang terjadi di dalam sitosol.
Pada amiloplas atau kloroplas yang berfungsi
19
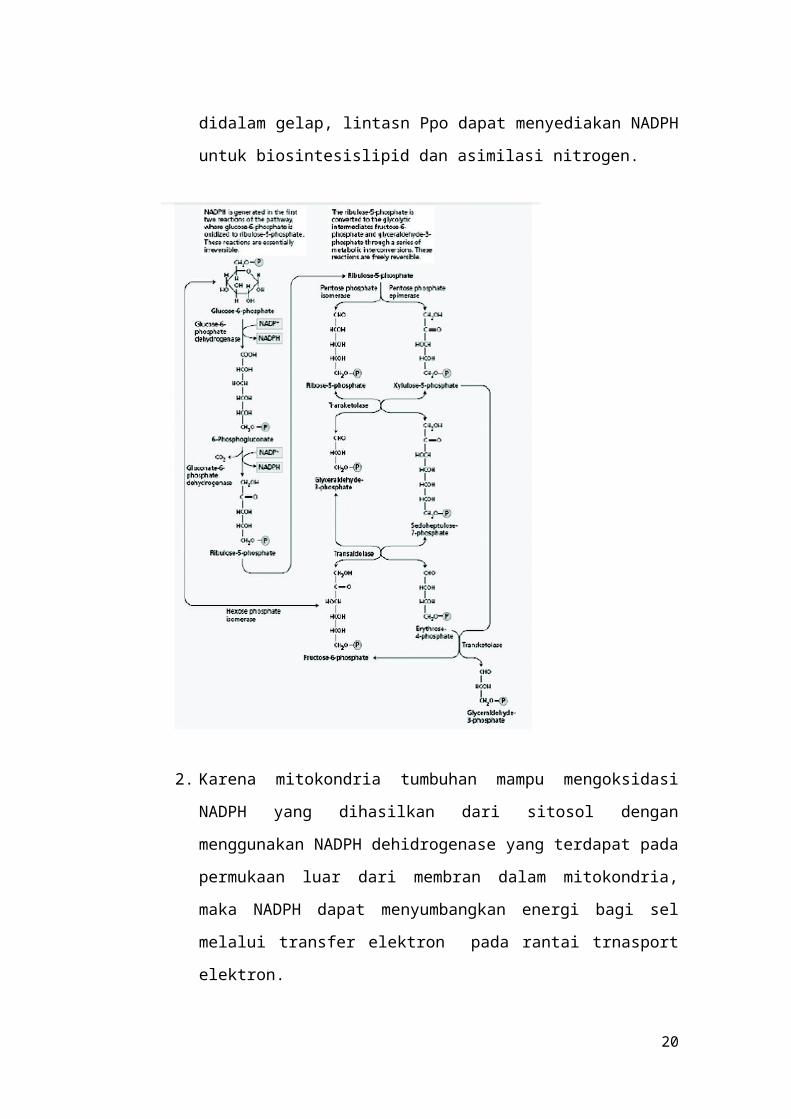
didalam gelap, lintasn Ppo dapat menyediakan NADPH
untuk biosintesislipid dan asimilasi nitrogen.
2. Karena mitokondria tumbuhan mampu mengoksidasi
NADPH yang dihasilkan dari sitosol dengan
menggunakan NADPH dehidrogenase yang terdapat pada
permukaan luar dari membran dalam mitokondria,
maka NADPH dapat menyumbangkan energi bagi sel
melalui transfer elektron pada rantai trnasport
elektron.
20

3. Lintasan PPO menghasilkan ribulosa 5 fosfat yang
merupakan prekursordari biosintesi ribosa dan
deoksiribosa yang diperluka untuk sintesi RNA dan
DNA.
4. Intermediet lainnya, eritrosa 4 fosfat dapat
bergabung dengan PEP untuk menghasilkan senyawa
fenolik tumbuhan, termasuk juga asam amino
aromatik dan prekursor –prekursor untuk lignin,
flavonoid, dan fitoaleskin.
5. Selama thap awal daun-daun berbuah menjadi hijau
diman pada jaringan daun tersebut fotosintesis
belum sepenuhnya berlangsung, lintasan PPO
diyakini sebagi penghasil intermedietsiklus
kalvin.
Lintasan PPO dikendalikan oleh reaksi awal dari
lintasan yang dikatalis oleh enzim glukosa 6 fosfat
dehidrogenase, diman aktifitasnya di hambat oleh rasio
yang tinggi dari NADPH terhadap NADP+
2.4 Siklus Asam Sitrat
Selama abad ke-19, ahli–ahli biologimeneliti
pengaruh keberadaan oksigen terhadap metabolisme
respirasi. Pada kondisi tidak ada oksigen,
selmeproduksi etanol atau asam laktat, sementara
apabila oksigen, sel mengkonsumsi oksigen,dan
memproduksi CO2 dan H2O. Pada tahun 1973, seorang ahli
21

kimia inggris kelahiran jerman, Hans A. Krebs,
melaporkan penelitiannya tentang siklus asam sitrat
(juga disebut siklus asam trikarboksilat atau siklus
krebs). Siklus asam sitrat menerangkan bagaiman piruvat
diubah menjadi molekul CO2 dan H2O dalam suatu
rangkaian reaksi kimia yang bersiklus. Siklus asam
sitrat berlangsung didalam matriks mitokondria.
Siklus asam sitrat disebut juga siklus asam
trikarboksilat untuk menunjukkan pentingnya dua molekul
trikarboksilat, citrat, dan isotrat sebagai intermediet
22

Gambar 4.4. reaksi siklus asam sitrat pada tanaman.
Piruvat dioksidasi menjadi 3 molekul CO2. Elektron
–elektron yang dilepaskan selama oksidasi tersebut
digunakan untuk mereduksi 4 molekul NAD+ menjadi
NADH dan 1 molekul FAD menjadi FADH2.
Pertama dari siklus ini. Untuk berlangsungnya siklus
asam sitrat , piruvat yang dihasilkan dari glikolisis
harus di transport kedalam matriks mitokondria melalui
23

protein transport khusus yang terdapat pada membaran
dalam mitokondria.
Setelah piruvat ada di dalam matriks, piruvat
kemudian di dekarboksilasi dalmsuari retu reaksi
oksidasi oleh enzim piruvat dehidrogenase. Hasil dar
reaksi ini adalah NADH, CO2dan asam asetatdalam bentuk
asetil KoA. Piruvat dehidrogenase merupakan kompleks
enzim yang mengkatalis keseluruhan tiga proses ini,
yaitu dekarboksilasi, oksidasi dan konjugasi asam
asetat dengan KoA (gambar 4.4).
Pada reaksiberikutnya enzim sitrat sintase
menggabungkan grup asetil dari asetil KoA dengan OAA,
suatu asam dikarboksilat beratom karbon empat, untuk
menghasilkan sitrat yang kemudian diisomerasi oleh
enzim aconitase untukeghasilakn isositrat. Dua tahap
reaksi berikutnya adalah reaksi dekarboksilai oksidatif
berurutan yang masing-masing menghasilkan suatu molekul
NADH, melepas satu molekul CO2 dan membentuk suksinil
KoA. Sampai tahap ini sudah tiga molekul CO2 dilepas
untuk tiap piruvat ynag masuk ke mitokondria, atau
molekul CO2 dari tiap molekul sukrosa yang di oksidasi
Tahap berikutnya adalah oksidasi suksimil KoA
menjadi OAA, seingga memungkinkan siklus asam sitrat
terus berputar. Mula-mual energi bebas yang tersedia
dari ikatan trioster pada suksinil KoA disimpan didalam
ATP dari ADP dan Pi melalui prose fosforilasi tingkat
substrat yang di katalis oleh enzim suksinil KoA
24

sintase. Suksinat yang dhasilkan dioksidasi menjadi
fumarat oleh enzim suksinat dehidrogenase, yang berada
pada membran dalam mitokondria dan merupakan enzim
yang juga berperan dalam mitokondria dan merupakn
enzim yang juga berperan did alam rantai transport
elektron.
Elektron dan proton dilepas dari suksinat dari
suatu reaksi redoks yang melibatkan FAD (flavin adenin
dinukloetida). FAD secara kovalen terikat pada sisi
aktif dari suksinat dehidrogenase menjadi OAA dan
menghasilkan satu molekul NADH. Sebagi kesimpulan,
setiap piruvat yang masuk mitokondria akan menghasilkan
tiga molekul CO2, empat molekul NADH, satu molekul
FADH2, dan satu molekul ATP.
2.4.1 Siklus Asam Sitrat Pada Tumbuhan Memiliki
Keunikan
Siklus asam soitrat tumbuhan (Gambar 4.4) tidak
identik dengna siklus asam sitrat yang terjadi pada
mitokondria hewan. Sebagi contoh pada tahapan yang
dikatalis oleh enzim suksinil Koa sintetase, di
tumbuhan menghasilkan ATP sedangkan pada hewan
menghasilkan GTP.
Perbedaan keduanya adalah aktifitas enzim NAD+
malat yang tidak dijumpai pada organisme lain. Enzim
ini mengkatalis reaksi dekarboksilasi oksidatif dari
malat menjadi piruvat:
25

Malat + NAD+ → piruvat + CO2+ NADH
Keberadaan enzim NAD+ malat memungkinkan mitokondria
tumbuhan mampu melakukan lintasan alternatif dari
metabolisme PEP yang dihasilkan meleui glikolisis.
Seperti diketahui, malat dapat disintesis dari PEP di
sitosol dengan bantuan enzim PEP karboksilase dan malt
dehidrogenase. Malat kemudian ditransport kedalam
matriks mitokondria , dimana enzim NAD+ malat dapat
mengoksidasinya menjadi piruvat. Reaksi ini
memungkinkan oksidasi sempurna dari intermediat-
intermediat siklus asam sitrat, seperti malat atau
sistrat.
Alternatif lain darijalur metabolisme PEP adalah
digunakannya malat sebagi pengganti intermediet siklus
asam sitrat. Malat yang masuk ke mitokondria hasil
PEPkarboksilase dapat mengisi kekuranagan malat akibat
keluarnya intermediat respirasi dari siklus. Misal 2
oksoglutarat dapat keluar siklus dan digunakan dalam
asimilasi nitrogen. Hal ini daapt menyebabknan
kekurangan malat untuk kelangsungan siklus. Adanya
malat yang masuk ke matriks dari sitososl hasil
metabolisme PEP dapat memnuhi kekurangan malat
tersebut, sehingga siklus tetap berjalan.
2.4.2 Rantai Transport Elektron Dan Sintesis Atp
ATP adalah pembawa energi yang digunakn oleh sel
untuk menggerakkaan proses- prose kehidupan, dan enrgi
26

yangdisimpan dalam bentuk NADH dan FADH2 harus diubah
ke bentuk ATP. Proses bergantung oksigen ini, disebut
fosforilasi oksidatif, terjadi pada membran dalam
mitokondria.
Setiap molekul sukrosa yang dioksidasi melalui
glikolisis dan siklus asam sitrat menghasilkan 4
molekul NADH di dalam sitosol dan 16 molekul NADH dan 4
molekul FADH2 di dalam matriks mitokondria. Molekul-
molekul pembawa elektron ini harus
Gambar 4.5. organisasi rantai transport Elektron dan
sintesis ATP pada membran dalam mitokondria
tanaman . ranati transpotrt elektron pada
mitokondria tanaman mempunyai 5 enzim tambahan.
Inhibitor spesifik, rotenon untuk kompleks I,
27

antimycin untuk kompleks III, sianida untuk
komplek IV, dan asam silisil hidroksamat (SHAM)
untuk oksidase alternatif
Dioksidasi kembali, karena kalau tidak maaka
keseluruhna proses respirasi akan terhenti. Rantai
transport elektron mengkatalis aliran elektron dari
NADH(dan FADH2) ke oksigen, suatu penerima elektron
terakhir dari proses respirasi. Untuk oksidasi satu
molekul NADH, reaksinya dapat dituliskan sebagai
berikut:
NADH + H+ +1/2O2 → NAD+ + H2O
Dari potensial reduksi pasangan NADH-NAD+ (-320
mV) dan pasangan H2O-1/2 O2 (+810 mV), dapat dihitung
bahwa energi bebas standar yang dilepas selam
keseluruhan reaksi adala kira-kira 220 kJ mol-1 (52
kkal mol-1) untuk tiap dua elektron. Sedangkan energi
bebas yang dilepas dalam oksidasi suksinat sebesar 152
kJ mol-1 (36 kkal mol-1)untuk tiap dua elektron.
Peranan dari rantai transport elektron adalah untuk
mengoksidasi NADH (dan FADH2) dan menggunakan sebagian
energi bebas yang di lepaskan untuk menimbulakn gardien
elektrokimia proton antara matriks dan ruang antar
membran.
Rantai transport elektron tumbuhan memiliki
kompleks pembawa elektron yang sama dengan apa yang
28

dijumpai pada organisme lain. Protein-protein transport
elektron tersebut dapat dikelompokkan kedalam 4 komplek
yang semuanya terdapat pada membran dalam mitokondria
(Gambar 4.5). keempat kompleks protein tersebut adalah:
Komplek I (NADH dehidrogenase). Elektron dari NADH di
oksidsi oleh komplek I yang memiliki pembawa elektron
yang disebut FMN (Flavin monoukleotida) dan beberapa
pusat besi sulfur. Kompleks I kemudian memindahkan
elektron ke ubiquinon. Empat proton dipompakan dari
matriks ke ruang antar membran untuk tiap pasang
elektron yang melewati kompleks ini.
Ubiquinon adalah pembawa proton dan elektron
terlarut lemak dan terletak di daalam membran dalam
mitokondrian. Ubiquinon dapat berdifusi didalam bagian
hidrifobik dari membran lapis ganda.
Komplek II (Suksinat dehidrogenase). Oksigen suksinat
dalam siklus asam sitrat yang menghsilkan FADH2,
mentransfer elektron ke ubiquinon. Kompleks ini tidak
memomopa proton.
Komplek III (komplek sitokrom bc1). Kompeks ini
mengoksidasi ubiquinon dan memindahkan elektron melalui
dua molekul sitokrom tibe b (b565 dan b560) dan sitokrom c1
kesitokrom c. Empat proton dipompakan oleh kompleks ini
untuk tiap pasang elektron yang ditransfer.
29

Komplek IV (sitokrom c oksidase). Komplek ini memiliki
dua pusat Cu (CuA dan CuB) dan sitokrom a dan a3.
Kompleks IV adalah komplek yang terakhir dan
menggunakan empat elektron untuk mereduksi oksigen
membentuk dua molekul air. Dua proton dipompakan untuk
tiap pasang elektron.
Disamping kesamaan kompleks protein transfer antara
rantai respirasi hewan dan tumbuahn, tumbuhan memiliki
keunikan pada beberpa enzim transport elektron yang
tidak dijumpai pada hewan, yaitu:
1. Dua NAD(P)H dehidrogenase. Keduanya bergantung
Ca2+. Kompleks ini berada pada permukaan luar dari
membran dalam mitokondria menghadap keruang antar
membran dan dapat mengoksidasi NADH dari sitosol.
Elektron dari NA(P)H dehidrogenase ini masuk
kerantai transport elektron langsung pada pool
ubiquinon.
2. Mitokondria tumbuhan memiliki dua lintasan
oksidasi NADH dari matriks. Aliran elektron
melalui komplek I sensitif terhadap senyawa
inhibitor seperti rotenon dan piericidin.
Mitokondria tumbuhan juga memiliki enzim
dehidrogenase resitsen rotenon. Peranan dari
lintasan ini sebagai bypass manakala kompleks I
dalam keadaan overload, seprti pada kondisi
fotorespirasi.
30

3. Adanya NAD(P)H dehidrogenase yang terdapat pada
permukaan membran dalam yang menghadap ke matriks.
4. Sebagian besar tumbuhan memiliki lintasan
respirasi alternatif untuk mereduksi oksigen.
Lintasa ini disebut lintasan oksidase alternatif
yang tidak sensitif terhadap penghambatan sianida,
azida dan karbon monooksida.
Pada fosforilasi oksidatif, transfer elektron ke
oksigen melalui kompleks I sampai IV bersamaan dengan
sintesis ATP dari ADP dan Pi melalui ATP sintse. Jumlah
ATP yang disentesis bergantung pada asal donor
elektron. Pada percobaan yang dilakukan dengan
menggunakan mitokondria yang diisolasi, elektron yang
berasal dari NADH matriks (NADH dari siklus asam
sitrat) memberikan rasio ADP:O (jumlah ATP yang
disintesis untuk tiap pasang elektron yang ditransfer
ke oksigen) adalah 2,4-2,7. NADH yang berasal dari
suksinat dan dari luar mitokondria masing masing
memiliki nialai ADP:O antara 1,6-1.8, sementara
askorbat memberikan ADP:O 0,8-0,9. Hasil percobaan ini
juga sesuia dengan perhitungan biaya pembuatan ATP
berdasrkan jumlah H+ yang dipompkan meleawati kompleks
I, III, IV yang menghasilkan kesimpulan bahwa setiap
satu molekul ATP yang disentesis memerlukan 4H+.
Mekanisme sintesis ATP didalam mitokondria
didasarkan pada hipotesis kemiosmosis yang pertama kali
31

diusulkan oleh Peter Mitchell pada tahun 1961. Menurut
teori tersebut, orientasi pembawa elektron didalam
membaran dalam mitokondria memungkinkan transfer proton
(H+) melintasi membran dalam mitokondria selama aliran
elektron. Sejumlah studi membuktikan bahwa transport
elektron mitokondria berkaitan erta dengan transfer
proton dari matriks mitokondria keruang antar membran.
Bagaimana transport elektron dapat berpasangan denagn
translokasi proton sampai saat ini masih belum jelas
mekanismenya.
Karena membran dalam mitokondria impermeabel
terhadap proton, maka suatu gradient elektronika proton
dapat terbentuk. Energi bebas yang dibutuhkan untuk
membentuk gradien elektrokimia proton berasal dari
energi bebas yang dilepas selama transport elektron.
Energi bebas yang terkandung dari gradien elektrokimia
ini dapat digunakan untuk melakukan kerja kimia
(sintesis ATP). Sinetsis ATP ini juga dapat berlangsung
karena adanya komplek enzim F0F1 –ATP sintase yang
terdapat pada membran dalam mitokondria.
F0F1–ATP sintase adalah kompleks enzim yang terdiri
dari dua komponen utama, yaitu F1 dan F0. F1 merupakan
komplek protein periferal membran yang tersusun
sedikitnya oleh lima sub unit protein yang berbeda dan
mengandung sisi katalitik untuk mengubah ADP dan Pi
menjadi ATP. Komplek ini menenpel pada integral pada
permukaan matriks dari membran dalam mitokondria.
32

Lewatnya proton melalui saluran tersebut bersama-
sama dengan pengaktifan situs katalik dari F1 dari
memungkinkan sintesis ATP yang secara simultan
menggunakan gradien elektrokimia proton. Untuk tiap
molekul ATP yang disentesis, 3 proton dilewatkan
melalui F0 dari ruang antar membran menuju matrik
mitokondria. Mekanisme kemiosmotik dari sintesis ATP
memiliki dua implikasi. Pertama, tempat pembentukan ATP
pada membran dalam mitokondria adalah ATP sintase,
bukan kompleks I, III atau IV. Kedua, teori kemiosmotik
menerangkan mekanisme kebocoran proton yang merangsang
laju transpor elektron tetapi menurunkan sintesis ATP.
2.4.3 Transport pada Membran Dalam Menukar Substrat dan
Produk
Gradien elektrokimia proton juga berperan dalam
pergerakan asam organik hasil siklus asam sitrat,
substrat dan produk dari sintesis ATP dari dan kedalam
mitokondria. Meskipun ATP disentesis didalam matriks
mitokondria, sebagian besar ATP digunakan diluar
mitokondria, sehingga mekanisme yang efisien diperlukan
untuk memasukkan ADP ke mitoondria dan mengeluarkan ATP
dari mitokondria.
Transport adenilat melibatkan protein membran
dalam mitokondria yang lan, yaitu ADP/ATP transporter.
Transporter ini mengkatalis pertukaran ADP dan ATP
melintasi membran dalam mitokondria (gambar 4.6).
33

pergerakan ATP4- bermuatan negatif keluar dan ADP3- ke
dalam yang menyisakan satu muatan negatif diruang antar
membran, digerakkan oleh gradien potensial elektrik
yang disebabkan oleh pemompaan proton.
Pengambilan fosfat iorganik (Pi) melibatkan
protein transporter fosfat dengan menggunakan komponen
gradien proton (∆pH) untuk menggerakkan pertukaran Pi-
yang masuk kedalam matriks dengan OH- yang keluar dari
matriks. Selama terjadi ∆pH antara ruang anatar membran
dan matriks mitokondria, kandungan Pi didalam matriks
akan tetap tinggi. Mekanisme yang sama juga berlaku
untuk pengambilan piruvat oleh matriks mitokondria yang
menukarnya dengan OH-. Total biaya untuk mengambil satu
gugus fosfat (I OH- keluar, yang sama dengan I H+
masuk) dan pertukaran ADP dan ATP (satu muatan negatif
keluar, yang sama dengan satu muatan positif masuk)
adalah 1 H+. Proton ini seharusnya juga dimasukkan
kedalam perhitungan biaya untuk mensintesis ATP. Jadi
biaya total ATP adalah 3 H+ digunakan oleh ATP sintase
ditambah 1 H+ untuk pertukaran melintasi membran atau
total biaya 4 H+.
34
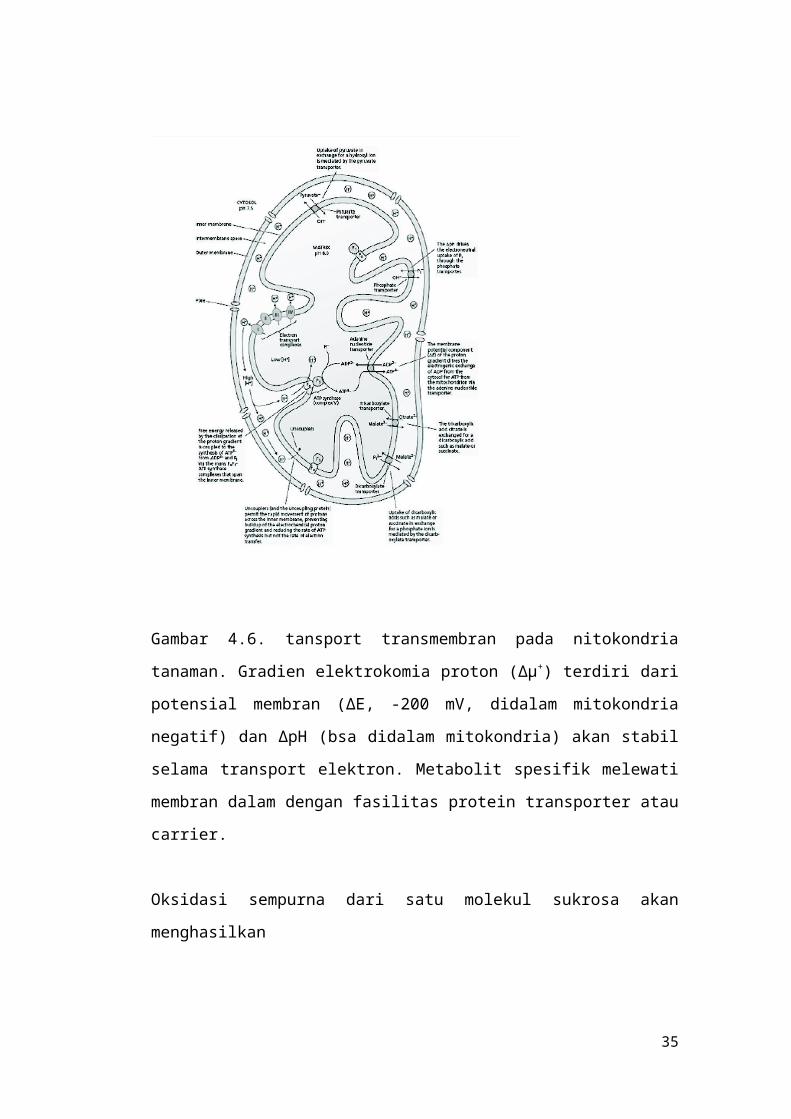
Gambar 4.6. tansport transmembran pada nitokondria
tanaman. Gradien elektrokomia proton (∆µ+) terdiri dari
potensial membran (∆E, -200 mV, didalam mitokondria
negatif) dan ∆pH (bsa didalam mitokondria) akan stabil
selama transport elektron. Metabolit spesifik melewati
membran dalam dengan fasilitas protein transporter atau
carrier.
Oksidasi sempurna dari satu molekul sukrosa akan
menghasilkan
35

8 molekul ATP melalui fosforilaso subsrat (4
dalam glikolisis dan 4 pada siklus asam sitrat)
4 molekul NADH dari sitosol (hasil gilkolisis)
16 molekul NADH dan 4 molekul FADH2 didalam
matrik mitokondria
Berdasarkan teori rasio ADP:O, maka kira-kira 52
molekul ATP akan dihasikan melalui fosforilasi
oksidatif dari satu molekul sukrosa, sehingga secara
keseluruhan kira-kira 60 ATP dihasilkan dari satu
molekul sukrosa yang dioksidasi secara aerob.
Dengan menggunakan 50 kJ mol-1 (12 kkal mol-1)
energi bebas untuk sintesis ATP secara in vivo, maka
sebanyak 3010 kJ mol-1 (720 kkal mol-1) energi bekas
untuk tiap mol sukrosa yang dioksidasi secara aerob.
jumlah tersebut sekitar 52% dari total energi yang
dikandung oleh satu mol sukrosa apabila dioksida secara
sempurna, sisanya dilepas dalam bentuk panas.
Bandingkan dengan jumlah energi bebas yang dapat
disimpan bentuk ATP dari sukrosa yang difermentasi
2.4.4 Efisiensi Konservasi Energi pada Tumbuahan Dapat
Berubah-ubah
Efisiensi konservasi energi didialam tumbuhan bisa
berubah-ubah. Bahkan tumbuhan tumbuhan memiliki protein
pada membran dalam mitokondria yang dapat menurunkan
jumlah ATP yang dihasilkan dari proses respirasi aerob
36

tersebut. Hal ini memungkinkan karena tumbuhan harus
memiliki fleksibilitas adaptasi terhadap faktor
pembatas lingkungan tumbuhnya, oleh karenanya bagi
tumbuhan kemampuan beradabtasi dengan korbanan
menurunkan jumlah ATP yang dihasilkan lebih penting
daripada efisiensi konservasi energi. Ada beberapa cara
tumbuhan menurunkan produksi ATP melalui mekanisme
nonfosforilasi, antara lain:
Oksidase alternatve. jika sianida (1mM) ditambahkan
pada jaringan hewan yang aktif berespirasi, sitokrom
oksidase c akan terhambat kerjanya dan laju respirasi
menurun dengan cepat sampai dibawah 1% dari laju
semula. Tetapi sebagian besar jaringan tumbuhan
menunjukkan tingkat respirasi yang resisten sianida
sebesar 10 sampai 25%, bahkan beberapa jaringan dapat
mencapai 100%, dari laju respirasi jaringan yang tidak
terhambat. Enzim yang bertanggung jawab terhadap
pengambilan oksigen pada kondisi ini telah
didefinisikasi sebagai enzim oksidase aternative, suatu
oksidase yang resisten sianida yang terdapat pada
membran dalam mitokondria.
enzim ini bekerja dimulai pada level pool elektron
ubiquinon. oksidase alternative menerima elektron
langsung dan kemudian mentransfer empat elektron ke
molekul oksigen untuk membentuk air. Ketika elektron
dilewatkan ke oksidase alternative dari pool ubiquinon,
dua situs pompa proton (kompleks III dan IV) di lewati.
37

karena tidak ada situs konservasi energi antara
oksidase alternative dan oksigen ,maka energi bebas
yang biasanya disimpan didalam ATP akan di lepas
sebagai panas ketika elektron-elektron dilewatkan
melalui jalur oksidase alternative. pertanyaannya
kemudaian bagaimana mungkin suatu proses yang
kelihatannya membuang-buang energi tetapi diperlukan
dalam metabolisme tumbuhan?
contoh kegunaan oksidase alternative yang paling
dikenal dapat dijumapai pada aktivitas oksiadse
alternatif selama perkembangan bunga Lily (Sauromatum
guttatum). Hanya beberapa saat sebelum penyerbukan ,
jaringan bunga (disebut appendix) yang menopang bunga
jantan dan betina, menunjukkan peningkatan laju
respirasi yang sangat cepat memlaui jalur oksidase
alternative. Akibatnya suhu pada bagian atas appendix
meningkat sebesar 25ºC diatas suhu normal selama
periode 7 jam. selama produksi panas ini, senyawa-
senyawaamine, indol, dan terpen diuapkan dari bunga
Lily tersebut dan dapat berfungsi sebagai penarik
serangga polinator. Pada sebagian besar tumbuhan, laju
respirasi dan laju oksidase alternative masih terlalu
rendah untuk menghasilkan panas yang begitu tinggi.
lalu apakah fungsi lain dari oksidase alternative?
lintasan alternative dapat berfungsi sebagai
lintasan “limpahan energi” hasil oksidasi substrat yang
terakumulasi setelah digunakan untuk tumbuh,
38

penyimpanan dan pembukaan ATP. Jadi, sepertinya
lintasan alternative dapat berlangsung kalau lintasan
rantai transport elektron utama telah jenuh. Tetapi
sekarang telah diketahui dengan jelas bahwa lintasan
oksidase alternative dapat berlangsung sebelum lintasan
utama jenuh. jadi lintasan oksidase alternatif dapat
berlangsung sebelum lintasan utama jenuh. jadi lintasan
oksidase alternatif adalah perannya dalam respon
tumbuhan terhadap stres yang dapat menghambat respirasi
mitokondria, seperti defisiensi unsur fosfor,
kekeringan, suhu dingin, cekaman osmotik dan stres
lainnya. Dengan cara membuka keran elektron, lintasan
oksidase alternatif dapat mencegah over reduksi dari
pool ubiquinon, yang kalau tidak dikontrol akan dapat
menghasilkan oksigen reaktif seperti anion superoxida
dan radikal hidroksil yang dpat merusak respirasi.
Protein “uncouller”. Pada membran dalam mitokondria
hewan dijumpai protein membran yang berperan sebagai
protein “ouncopler” yang kerjanya mirip pada
mitokondria hewan. kerja protein ini dapat diinduksi
oleh stres dan mungkin berperan seperti halnya oksidase
alternatif, yaitu untuk mencegah terjadinya over
reduksi dari rantai transport elektron. Belum diketahui
dengan jelas, mengapa tumbuhan memiliki kedua mekanisme
pencegahan over reduksi tersebut.
39

NADH dehidrogenase resisten rotenon ( Internal NDin
(NADH). ND in (NADH) adalah salah satu dari beberapa
NADH dehidroenase yang dijumpai pada mitokondria
tumbuhan. Enzim ini berfungsi sebagai bypass elektron
apabila kompleks I kelebihan elektron. tidak seperti
pada kompleks I, pada saat elektron melewati NDin (NADH)
ini tidak dipompakan proton. Komplek I memiliki
afinitas yang lebih tinggi terhadap NADH dibanding NDin
(NADH). pada saat suplai NADH rendah didalam matriks,
biasanya ADP dalam keadaan tersedia, Kompleks I akan
mendominasi pengaliran elektron dari NADH ke rantai
transport elektron. Akan tetapi ketika ADP menjadi
lebih aktif. Peran pentingnya enzim ini secara
fisiologis masih belum diketahui dengan jelas.
2.4.5 Respirasi Mitokondria Dikendalikan oleh
Metabolit-metabolit Kunci
ADP dan Pi merupakan subsrat dari sintesis ATP
yang menjadi pengatur kunci dari laju glikolisis
didalam sitosol, siklus asam sitrat dan fosforilasi
oksidatif di dalam mitokondria. Beberapa titik-titik
pengendali dari ketiga tahap respirasi dapat dilihat
pada Gambar 4.7.
40

Gambar 4.7. pengaturan aktivitas piruvat dehidrogenase
(PDH) dengan fosforilasi reversibel dan
metabolit lainnya.
salah satu situs pengendali siklus asam sitrat
adalah kompleks enzim piruvat dehidrogenase, yang
secara dapat balik dapat difosforilasi baik oleh suatu
41

kiase dan fosfatase. Piruvat dapat menghambat kerja
dari kinase, sehingga memungkinkan enzim piruvat
dehidrogenase dapat bekerja ketika piruvat tersedia.
Beberapa enzim dalam siklus asam sitrat termasuk
termasuk di dalamnya piruvat dehidrogenase dan 2 -
oxogludehidrogenase secara langsung dihambat oleh
suplai NADH.
Oksidasi siklus asam sitrat dan rantai respirasi
secara dinamis dikendalikan oleh tingkat adenin
nukleotida di dalam sel. Pada saat kebutuhan akan ATP
di dalam sitosol menurun relatif terhadap laju sintesis
ATP di dalam mitokondria, ADP yang tersedia
akanberkurang, dan rantai transport elektron akan
berjalan lambat, yang berakibatakan meningkatnya NADH
di dalam matriks. Tingkat NADH yang tinggi ini akan
menyebabkan aktifitas beberapa enzim dehidrogenase pada
siklus asam sitrat yang menjadi terhambat.
Pembentukan intermediat siklus asam sitrat dan
turunannya, seperti sitrat dan glutamat, menghambat
kerja piruvat kinase di sitosol, sehingga meningkatkan
konsentrasi PEP di dalam sitosol,yang berakibat
menurunnya laju konversi fruktosa- 6-fosfat menjadi
fruktosa-1,6-bisfosfat, sehingga menghambat glikolisis.
Sebagai kesimpulan, laju respirasi tumbuhan
dikendalikan dari “bawah ke atas” oleh suplai ADP di
dalam mitokondria. ADP dapat mengatur laju transfer
elektron dan sintesis ATP, yang selanjutnya
42

mengaturaktifitas siklus asam sitrat, yang akhirnya
mengatur laju dari glukolisis.
2.4.6 Hubungan Respirasi dengan Lintasan Metabolisme
Lain di dalam Tumbuhan
Glikolisis, lintasan pentosa fosfat, dan siklus
asam sitrat berkaitan erat
dengan beberapa lintasan metabolik lain di dalam
tumbuhan
(Gambar 4.8). Lintasan-lintasan di dalam respirasi
merupakan pusat penghasil berbagai jenis metabolit
tumbuhan, termasuk asam amino, lipid dan senyawa
terkait, isoprenoid, porfirin dan lain-lain seperti
pada Gambar 4.8. Detail proses metabolisme dari
biosintesis senyawa-senyawa tersebut tidak akan dibahas
pada kuliah Fisiologi tumbuhan dasar ini.
43

Gambar 4.8. Glikolisis , lintasan pentosa fosfat dan
siklus asam sitrat berkaitan dengan prekursor
beberapa lintasan biosintesis pada tanaman
tingkat tinggi. Lintasan –lintasan menggambarkan
bahwa biosintesis pada tanaman membutuhkan
karbon dan tidak semua karbon yang masuk dalam
glikolisis dioksidasi menjadi CO2.
44

2.5 Respirasi Pada Tumbuhan
Sejauh ini kita telah membicarakan respirasi yang
berlangsung pada tingkat organel dan sel. Lalu
bagaimana pengetahuan ini dapat dikaitkan denganfungsi
tumbuhan secar keseluruhan diaalam? Pada bagian ini
kita akan membahas respirasi tumbuhan dalam berbagai
kondisis di alam.
2.5.1 Tumbuhan Merespirasikan Hampir setengah dari
hasil fotosintesis
Banyak faktor dapat mempengaruhi laju respirasi
dan kebiasaan hiduptumbuhan atau organ tumbuhan,
seperti spesies dan kebiasaan hidup dari tumbuhan,
jenis dan umur organ tumbuhan, faktor lingkungan
sepertikonsentrasi oksigen, suhu, hara, dan suplai air.
Laju respirasi tumbuhan secara keseluruhan umunya
lebih rendah dibanding laju respirasi jaringan hewan.
Perbedaan ini dikarenakan volume sel tumbuhan terisi
oleh vakuola yang besar dan dinding sel yang kedunya
tidak memiliki mitokondria . Akan tetapi, laju
respirasi pada beberapa jaringan tumbuhan sama
tingginya dengan laju respirasi jaringan hewan. Pada
kenyataannya , mitokondria tumbuhan berespirasi lebih
cepat dari mitokondria hewan.
Meskipun secara umum tumbuhan laju respirasi yang
rendah , kontribusi respirasi terhadap keseluruhan
45

penggunaan karbon didalam tumbuhan sangatlah penting,
karena semua jaringan tumbuhan berespirasi selam 24 jam
per hari. Bahkan respirasi jaringan fotosintetik dapat
menggunakan sejumlah besar hasil fotosintetis. Hasil
penelitian menunjukkan bahwa 30-60% hasil fotosintetis
tumbuhan herba hilang melalui respirasi. Pada pohon
muda, kira-kira sepertiga dari fotosintat hilang
direspirasikan dan menjadi dua kali lipat pada pohon
yang lebih tua, sebagai akibat menurunnya rasio
jaringan fotosintetik dan nonfotosintetik. Di daerah
tropik,70-80% dari hasil fotosintesis harian hilang
melalui respirasi karena tingginya laju respirasi
gelap akiabt suhu malam yang tinggi.
2.5.2 Respirasi Selama Fotosintetis
Mitokondria terlibat dalam metabolisme dari organ
fotosintetik, baik melalui respirasi
cahaya( fotrespirasi) maupun respirasi gelap.
Fotorespirasi menghasilkan mengoksidasi hasil
fotosintesis menjadi glisin yang kemudian dioksidasi
menjadi serin didalam mitokondria. Pada saat yang
bersamaan, mitokondria juga melakukan respirasi siklus
asam sitrat(respirasi gelap).
Pada organ fotosintetik,seperti daun, aktifitas
pirufat dehidrogenase dalm kondisi cahaya menurun
sebesar 25%, yang mengakibatkan laju respirasi secar
keseluruhan menurun. Respirasi mitokondria pada organ
46

fotosintetik berperan dalam :1. Mensuplai ATP, dan 2.
Mensuplai kerangka karbon untuk biosintetis, seperti 2-
oxoglutarat untuk assimilasi nitogen.
2.5.3 Laju Respirasi Jaringan dan Organ Tumbuhan
Laju respirasi jaringan bergantung pada aktifitas
metabolik dari jaringan tersebut. Secara umum semakin
tinggi aktifitas metabolisme dari suatu jaringan,
semakin tinggi laju respirasi dari jaringan tersebut.
Tunas yang sedang berkembang menunjukkan laju respirasi
jaringan vegetatif akan menurundari titik pertumbuhan
ke daerah yang lebih terdiferensiasi. Jaringan tumbuhan
yang telah dewasa , biasanya laju respirasinya tetap
atau menurun dengan lambat dengan bertambahnya umur dan
tingkat penuaan. Kekecualian terjadi pada kondisi
klimakterik, diman apeningkatan laju respirasi terjadi
pada proses pemasakn buah dan penuaan daun yan gugr/
lepas dan bungah potong. Hal ini dikarenakan adanya
produksi etilen endogenous yang dapatt meningkatkan
aktifiats lintasan alternatif resisten sianida pada
membran dalam mitokondria
2.5.4 Fungsi Mitokondria Sangat Penting Selama
Perkembangan Polen
Jumlah mitokondria dan ekspresi protein respirasi
di dalam anter yang sedang berkembang sangat tinggi.
Hal ini dikarenakan proses perkembangan polen adalah
proses yang memerlukan banyak energi. Karakteter
47

fisiologis yang berkaitan langsung dengan mitikondria
tumbuhan adalah karakter sterilitas jantan sitoplasma
(cytoplasmic male steriliti atau cms). Tanaman yang memiliki
karakter cms tidaka daapt menghasilkan polen yang
viabel, oleh karenanya tumbuhan seperti itu disebut
tanaman jantan steril. Istilah sitoplasma menunjukkan
pada pola pewarisan sifat ini tidak mengikuti pola
pewarisan mendel, tetapisifat ini diwariskan secar
maternal dari genom mitokondria. Tanaman cms sangat
penting dalam pemulaian tanaman karena galur jantan
steril yang stabil dapat digunakan untuk menghasilkan
benih hibrida.
Karakter cms pada tanaman disebabkan oleh adnya
mutasi pada DNA mitokondria yang menyandikan subunit
kompleks protein yang terlibat dalm fosforilasi
oksidatif. Pada jagung, tanaman cms-T memiliki perubahn
susunan DNA sedemikian rupa sehingga dihasilkan protein
URF13. Protein ini ketika berinteraksi dengan toksin
Hm-T, yang di hasilkan oleh cendawan Bipolaris maydis yang
menyerang jagung, dapat menyebabakan kebocoran membran
dalam mitokondria.
2.5.5 Faktor lingkungan yang berpengaruh terhadap laju
respirasi
1. Oksigen
Oksigen dapat mempengaruhi respirasi tumbuhan
karena peranannya sebagi substrat dari keseluruhan
48

proses respirasi. Pada suhu 25°C konsentrasi
keseimbangan oksigen didalam larutan jenuh udara
(21%O2)sekitar 250 uM. Nilai Km dari oksigen dalam
reaksi yang dikatalisis oleh sitopkrom c oksidase
dibawah 1 uM, sehingga seharusnya tidak ada
ketergantungan laju respirasi terhadap konsentrasi
oksigen di atmosfer. Tetapai kenyataanya, laju
respirasi jaringan dan potongan jaringan menurun
pada konsentrasi oksigen atmosfer berturut-turut
5% dan 2-3%. Hal ini menunjukkan bahwa difusi
oksigen melalui fase cair di dalm jaringan menjadi
terbatas. Oleh karena itu keberadaan ruang udara
antar sel sangatlah penting bagi ketersediaan
oksigen untuk mitokondria. Apabila tidak ada
difusi gas maka respirasi sel tumbuhan akan
terhambat.
2. Air
Keterbatasan difusi oksigen akan lebih terhambat
pada organ tumbuhan yang tumbuh pada medium cair.
Pada budidaya hidronik, larutan hara harus terus
mendapat aerasi untuk menjaga tingkat oksigen yang
tinggi di sekitar akar. Masalah suplai oksigen
juga dapat dialami oleh tumbuhan yang tumbuh pada
tanah yang sangat basah atau tergenang. Suplai
oksigen yang berkurang karena keterbatasan difusi
melalui medium cair( jenuh air, rendah oksigen)
dapat menghambat respirasi tumbuhan.
49

3. Suhu
Laju respirasi umumnya meningkat dengan
meningkatnya suhu. Pada suhu antara 0- 30°C
peningkatan laju respirasi untuk tiap 10°C
peningkatan dia tas suhu ambien sebesar 2°C. Di
atas suhu 30°C peningkatan laju respirasi menjadi
lebih lambat sampai mencapai konsta pada suhu 40-
50°C, akan menurun pada suhu yang lebih tinggi.
Suhu malam yang tinggi juga dapat meningkat laju
respirasi dari tumbuhan-tumbuhan tropik.
4. Konsentrasi CO2
Konsentrasi CO2 3-5% dapat menghambat laju
respirasi. Konsentrasi tersebut sebenarnya jauh
lebih dari kandungan normal CO2 atmosfer yang
hanya 0,036% (360 ppm). Percobaan dengan
menumbuhkan tumbuhan pada atmosfer dengan 700 ppm
CO2 menunjukan laju respirasi yang 15- 20% lebih
rendah dari respirasi tumbuhan pada kondisi
atmosfer normal.
2.6 Metabolisme Lemak
Hewan menggunakan lemak untuk penyimpanan energi,
tanaman menggunakan mereka terutama untuk penyimpanan
karbon. Lemak dan minyak yang penting bentuk
penyimpanan berkurang karbon di banyak biji,
termasukmereka spesies pertanian penting seperti
kedelai, bunga matahari, kacang tanah, dan kapas.
Minyak sering melayani utama fungsi penyimpanan pada
50

tanaman yang menghasilkan nondomesticated biji kecil.
Beberapa buah-buahan, seperti zaitun dan alpukat, juga
lemak toko dan minyak.Pada bagian akhir ini bab ini
kami akan menjelaskan biosintesis dari dua jenis
gliserolipid: the trigliserida (yang lemak dan minyak
yang tersimpan dalam biji) dan gliserolipid kutub (yang
membentuk bilayers lipid dari membran sel) (Gambar
11.14). Kita akan melihat bahwa biosintesis
triasilgliserol dan gliserolipid kutub membutuhkan
kerjasama dari dua organel: plastida dan retikulum
endoplasma. Tanaman juga dapat menggunakan lemak dan
minyak untuk produksi energi. Kita dengan demikian akan
memeriksa proses yang kompleks dimana berkecambah biji
memperoleh energi metabolisme dari oksidasi lemak dan
minyak.
Lemak dan Minyak menyimpan sejumlah besar energi
Lemak dan minyak milik lipid kelas umum,
struktural beragam kelompok senyawa hidrofobik yang
larut dalam pelarut organik dan sangat tidak larut
dalam air. Lipid merupakan bentuk yang lebih berkurang
karbon dari karbohidrat, sehingga oksidasi lengkap 1 g
lemak atau minyak (yang berisi sekitar 40 kJ, atau 9,3
kkal, energi) dapat menghasilkanjauh lebih ATP dari
oksidasi 1 g pati(sekitar 15,9 kJ, atau 3,8 kkal).
51

Sebaliknya, biosintesislemak, minyak, dan molekul
terkait, seperti fosfolipidmembran, membutuhkan
investasi besar Sejalan energi metabolik.
Lipid lainnya adalah penting bagi struktur dan
fungsi tanaman tetapi tidak digunakan untuk penyimpanan
energi. ini termasuk lilin, yang membentuk kutikula
pelindung yang mengurangi kehilangan air dari jaringan
tanaman terkena, dan terpenoid (juga dikenal sebagai
isoprenoidnya), yang meliputi karotenoid terlibat dalam
fotosintesis dan sterol hadir dalam banyak membran
tanaman.
52
Related Documents











