Maize toxin degrades peritrophic matrix proteins and stimulates compensatory transcriptome responses in fall armyworm midgut Howard W. Fescemyer a, * , Germán V. Sandoya b, 1 , Torrence A. Gill c , Seval Ozkan d , James H. Marden a , Dawn S. Luthe b a Department of Biology, The Pennsylvania State University, 208 Mueller Laboratory, University Park, PA 16802, USA b Department of Plant Science, The Pennsylvania State University, University Park, PA 16802, USA c United States Department of Agriculture USDA-ARS, Purdue University, West Lafayette, IN, USA d Department of Plant and Soil Sciences, The AgBase Databases, Institute for Genomics, Biocomputing and Biotechnology (IGBB), HPC2, Box 9555, Mississippi State University, MS 39762, USA article info Article history: Received 25 October 2012 Received in revised form 5 December 2012 Accepted 10 December 2012 Keywords: Spodoptera frugiperda Planteherbivore interaction Host plant resistance Zea mays Cysteine protease Mucin Compensatory response Gene expression abstract Understanding the molecular mechanisms underlying insect compensatory responses to plant defenses could lead to improved plant resistance to herbivores. The Mp708 inbred line of maize produces the maize insect resistant 1-cysteine protease (Mir1-CP) toxin. Reduced feeding and growth of fall army- worm larvae fed on Mp708 was previously linked to impairment of nutrient utilization and degradation of the midgut (MG) peritrophic matrix (PM) by Mir1-CP. Here we examine the biochemical and tran- scriptional responses of fall armyworm larvae to Mir1-CP. Insect Intestinal Mucin (IIM) was severely depleted from pure PMs treated in vitro with recombinant Mir1-CP. Larvae fed on Mp708 midwhorls excrete frass largely depleted of IIM. Cracks, fissures and increased porosity previously observed in the PM of larvae fed on Mp708 midwhorls could ensue when Mir1-CP degrades the IIM that cross-links chitin fibrils in the PM. Both targeted and global transcriptome analyses were performed to determine how complete dissolution of the structure and function of the PM is prevented, enabling larvae to continue growing in the presence of Mir1-CP. The MGs from fall armyworm fed on Mp708 upregulate expression of genes encoding proteins involved in PM production as an apparent compensation to replace the disrupted PM structure and restore appropriate counter-current MG gradients. Also, several families of digestive enzymes (endopeptidases, aminopeptidases, lipases, amylase) were more highly expressed in MGs from larvae fed on Mp708 than MGs from larvae fed on diets lacking Mir1-CP (artificial diet, midwhorls from Tx601 or B73 maize). Impaired growth of larvae fed on Mp708 probably results from metabolic costs associated with higher production of PM constituents and digestive enzymes in a compensatory attempt to maintain MG function. Ó 2013 Elsevier Ltd. All rights reserved. 1. Introduction Fall armyworm, Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) is a devastating pest throughout much of the western hemisphere. This insect thrives on a wide range of plants including the important crops, maize (Zea mays L.) and rice (Oryza sativa L.). Unfortunately, methods for controlling this herbivore are limited. Although chemical insecticides can be an effective control, they are Abbreviations: ARH, arylphorin; BLAST, basic local alignment search tool; BLASTn, Nucleotideenucleotide BLAST; BLASTp, Proteineprotein BLAST; BLASTx, Nucleotide 6- frame translation-protein BLAST; tBLASTx, Nucleotide 6-frame translation-nucleotide 6-frame translation BLAST; CAD, cadherin; CP, carboxypeptidases; CDA, chitin deacytlase; CTR, chymotrypsin; CHTG, Chymotrypsinogen; CYP, cytochrome P450; C T , cycle threshold; cDNA, copy deoxyribonucleic acid; DAVID, Database for Annotation, Visualization and Integrated Discovery; FBgn, FlyBase gene number; IIM, insect intestinal mucin; JA, jasmonic acid; 2-D LC-MS/MS, two dimensional liquid chromatography tandem mass spectrometry; LYSO, lysozyme; MA plots, log 2 -ratio of two expression levels/intensities versus the mean log 2 -expression of the same two expression levels/ intensities; Mir1-CP, maize insect resistant 1 cysteine protease; MG, midgut; NAC, no amplification control; NTC, no template control; PM, peritrophic matrix; PRTS, 30 k protease; Q-values, adjusted probability-values found using an optimized false discovery rate approach; qRT-PCR, quantitative reverse-transcription polymerase chain re- action; QTLs, quantitative trait loci; RNA-seq, high throughput sequencing of RNA after conversion to cDNA; TMM, trimmed mean of M values; TYRP, trypsin. * Corresponding author. Tel.: þ1 814 863 9269; fax: þ1 814 865 9131. E-mail address: [email protected] (H.W. Fescemyer). 1 Current address: The Genome Center, University of California, Davis,1636 East Alisal Street, USDA-ARS, Salinas, CA 93906, USA. Contents lists available at SciVerse ScienceDirect Insect Biochemistry and Molecular Biology journal homepage: www.elsevier.com/locate/ibmb 0965-1748/$ e see front matter Ó 2013 Elsevier Ltd. All rights reserved. http://dx.doi.org/10.1016/j.ibmb.2012.12.008 Insect Biochemistry and Molecular Biology 43 (2013) 280e291

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

at SciVerse ScienceDirect
Insect Biochemistry and Molecular Biology 43 (2013) 280e291
Contents lists available
Insect Biochemistry and Molecular Biology
journal homepage: www.elsevier .com/locate/ ibmb
Maize toxin degrades peritrophic matrix proteins and stimulatescompensatory transcriptome responses in fall armyworm midgut
Howard W. Fescemyer a,*, Germán V. Sandoya b,1, Torrence A. Gill c, Seval Ozkan d, James H. Marden a,Dawn S. Luthe b
aDepartment of Biology, The Pennsylvania State University, 208 Mueller Laboratory, University Park, PA 16802, USAbDepartment of Plant Science, The Pennsylvania State University, University Park, PA 16802, USAcUnited States Department of Agriculture USDA-ARS, Purdue University, West Lafayette, IN, USAdDepartment of Plant and Soil Sciences, The AgBase Databases, Institute for Genomics, Biocomputing and Biotechnology (IGBB), HPC2, Box 9555, Mississippi State University,MS 39762, USA
a r t i c l e i n f o
Article history:Received 25 October 2012Received in revised form5 December 2012Accepted 10 December 2012
Keywords:Spodoptera frugiperdaPlanteherbivore interactionHost plant resistanceZea maysCysteine proteaseMucinCompensatory responseGene expression
Abbreviations: ARH, arylphorin; BLAST, basic localframe translation-protein BLAST; tBLASTx, Nucleotideacytlase; CTR, chymotrypsin; CHTG, ChymotrypsinoVisualization and Integrated Discovery; FBgn, FlyBasetandem mass spectrometry; LYSO, lysozyme; MA plointensities; Mir1-CP, maize insect resistant 1 cysteineprotease; Q-values, adjusted probability-values foundaction; QTLs, quantitative trait loci; RNA-seq, high th* Corresponding author. Tel.: þ1 814 863 9269; fax
E-mail address: [email protected] (H.W. Fescemyer).1 Current address: The Genome Center, University
0965-1748/$ e see front matter � 2013 Elsevier Ltd.http://dx.doi.org/10.1016/j.ibmb.2012.12.008
a b s t r a c t
Understanding the molecular mechanisms underlying insect compensatory responses to plant defensescould lead to improved plant resistance to herbivores. The Mp708 inbred line of maize produces themaize insect resistant 1-cysteine protease (Mir1-CP) toxin. Reduced feeding and growth of fall army-worm larvae fed on Mp708 was previously linked to impairment of nutrient utilization and degradationof the midgut (MG) peritrophic matrix (PM) by Mir1-CP. Here we examine the biochemical and tran-scriptional responses of fall armyworm larvae to Mir1-CP. Insect Intestinal Mucin (IIM) was severelydepleted from pure PMs treated in vitro with recombinant Mir1-CP. Larvae fed on Mp708 midwhorlsexcrete frass largely depleted of IIM. Cracks, fissures and increased porosity previously observed in thePM of larvae fed on Mp708 midwhorls could ensue when Mir1-CP degrades the IIM that cross-linkschitin fibrils in the PM. Both targeted and global transcriptome analyses were performed to determinehow complete dissolution of the structure and function of the PM is prevented, enabling larvae tocontinue growing in the presence of Mir1-CP. The MGs from fall armyworm fed on Mp708 upregulateexpression of genes encoding proteins involved in PM production as an apparent compensation toreplace the disrupted PM structure and restore appropriate counter-current MG gradients. Also, severalfamilies of digestive enzymes (endopeptidases, aminopeptidases, lipases, amylase) were more highlyexpressed in MGs from larvae fed on Mp708 than MGs from larvae fed on diets lacking Mir1-CP (artificialdiet, midwhorls from Tx601 or B73 maize). Impaired growth of larvae fed on Mp708 probably resultsfrom metabolic costs associated with higher production of PM constituents and digestive enzymes ina compensatory attempt to maintain MG function.
� 2013 Elsevier Ltd. All rights reserved.
1. Introduction
Fall armyworm, Spodoptera frugiperda (J.E. Smith) (Lepidoptera:Noctuidae) is a devastating pest throughout much of the western
alignment search tool; BLASTn, Nude 6-frame translation-nucleotidegen; CYP, cytochrome P450; CT, cygene number; IIM, insect intestinats, log2-ratio of two expression levprotease; MG, midgut; NAC, no ausing an optimized false discover
roughput sequencing of RNA after: þ1 814 865 9131.
of California, Davis, 1636 East Alisa
All rights reserved.
hemisphere. This insect thrives on a wide range of plants includingthe important crops, maize (Zea mays L.) and rice (Oryza sativa L.).Unfortunately, methods for controlling this herbivore are limited.Although chemical insecticides can be an effective control, they are
cleotideenucleotide BLAST; BLASTp, Proteineprotein BLAST; BLASTx, Nucleotide 6-6-frame translation BLAST; CAD, cadherin; CP, carboxypeptidases; CDA, chitincle threshold; cDNA, copy deoxyribonucleic acid; DAVID, Database for Annotation,l mucin; JA, jasmonic acid; 2-D LC-MS/MS, two dimensional liquid chromatographyels/intensities versus the mean log2-expression of the same two expression levels/mplification control; NTC, no template control; PM, peritrophic matrix; PRTS, 30 ky rate approach; qRT-PCR, quantitative reverse-transcription polymerase chain re-conversion to cDNA; TMM, trimmed mean of M values; TYRP, trypsin.
l Street, USDA-ARS, Salinas, CA 93906, USA.

H.W. Fescemyer et al. / Insect Biochemistry and Molecular Biology 43 (2013) 280e291 281
expensive and not environmentally friendly, and the fall army-worm has developed resistance to many of these (Yu, 1992; Yuet al., 2003; Virla et al., 2008). Transgenic maize cultivars that ex-press a Bt-toxin are less effective against fall armyworm than otherlepidopteran herbivores (Luttrell et al., 1999; Buntin et al., 2000)and field resistance to this technology was recently observed inPuerto Rico (Storer et al., 2010). Furthermore, there are concernsabout public acceptance and the high cost of transgenic crops thatoften precludes their use by farmers in developing countries. Toextend the repertoire of potential control strategies for this herbi-vore, we are studying the mechanism of native host plant resist-ance in the maize inbred line Mp708 developed by traditional plantbreeding (Williams et al., 1990). This resistance thwarts feeding byfall armyworm and numerous other lepidopteran pests (Davis et al.,1988). When fall armyworm larvae feed on whorl tissue fromMp708, their growth is retarded by approximately 50% compared tosusceptible maize inbreds, and the time required to reach pupationincreased by several days (Chang et al., 2000). Analysis of nutri-tional indices from larvae fed on Mp708 indicated that theirnutrient utilization was severely impaired (Chang et al., 2000).
Research to understand the Mp708 mechanism slowing larvalgrowth revealed that resistance is a multigene trait regulated byseveral quantitative trait loci (QTLs) (Brooks et al., 2005, 2007).Among these traits, Mp708 has at least two unique characteristics:1] the plants are “genetically primed” to withstand larval feedingdue to constitutively elevated levels of jasmonic acid (JA) andherbivore defense gene transcripts (Shivaji et al., 2010); and 2]there is an exceedingly rapid accumulation of the maize insectresistant 1-cysteine protease (Mir1-CP) in the whorl within 1 h oflarval attack (Pechan et al., 2000). This protease is a highly potentinsecticidal protein that attacks the fall armyworm peritrophicmatrix (PM) (Pechan et al., 2002; Mohan et al., 2006), a semi-permeable protective structure that lines the larval midgut (MG)and surrounds the food bolus preventing food particles fromdirectly contacting epithelial cells and MG-microvilli (Wang andGranados, 2001). The proteolytic activity of Mir1-CP per-meabilizes the PM (Mohan et al., 2006), which ultimately impairsthe insect’s ability to utilize nutrients from its diet (Chang et al.,2000). Although it is known that Mir1-CP attacks PM proteins,we do not know if all PM proteins are equally vulnerable to pro-teolysis by Mir1-CP, or if specific PM proteins are targeted. Forexample, it has been shown that the PM structural protein, InsectIntestinal Mucin (IIM), is degraded by the metalloprotease,enhancin, produced by the Trichoplusia ni granulovirus during itsinfection of T. ni (Hübner) larvae (Lepidoptera: Noctuidae) (Wangand Granados, 1997a).
With exception to the order Hemiptera, all other representativesof the class Insecta contain a PM during all or most of their lifestages. The PM facilitates the digestive processes in the MG,maintains its structural integrity and protects the insect frominfection by microorganisms and parasites (Hegedus et al., 2009). Itis mainly composed of chitin, glycosaminoglycans and proteins(Wang and Granados, 2001; Hegedus et al., 2009; Toprak et al.,2010a). The proteins can be divided into nonstructural and struc-tural proteins collectively called peritrophins. Nonstructural pro-teins in the PM include chitin-modifying enzymes (e.g., chitindeacetylase, chitinase) and several families of digestive enzymes(e.g., lipases, serine proteases, exopeptidases, endopeptidases)(Toprak et al., 2010a). The composition of the PM, however, de-pends on the insect species. For example, the major proteins foundin the PM of Helicoverpa armigera (Hübner) (Lepidoptera: Noctui-dae) were chitin deactylase-like and mucin-like (Campbell et al.,2008). In the fall armyworm, anterior MG columnar cells secretea peritrophin (Bolognesi et al., 2001). Peritrophins are referred to asIIMs when they contain both chitin binding and mucin domains
(Hegedus et al., 2009; Toprak et al., 2010a) with disagreement inthe number of mucin domains required to constitute an IIM.
Besides its role in digestion, the PM plays an important role indetoxification of plant toxins consumed by the insect (Hegeduset al., 2009; Hakim et al., 2010; Toprak et al., 2010a). Thus, dis-rupting the PM, or making it more permeable, induces behavioraland physiological changes in caterpillars, such as cessation offeeding, reduced growth and increased susceptibility to toxins andpathogens (Wang and Granados, 2001; Guo et al., 2007; Hegeduset al., 2009). Although knowledge of the defensive mechanismsdeployed by maize in response to feeding by fall armyworm andother caterpillars is increasing (Lopez et al., 2007; Mohan et al.,2008; Shivaji et al., 2010; Gill et al., 2011; Smith et al., 2012), littleis known about responses to maize toxins by fall armyworm larvae.In particular, we sought to illuminate changes in gene expressionthat occur in the larval MG tissue during the larval counter defenseresponse. A better understanding of PM constituents that protectthe MG from damage will provide insights into potential new tar-gets for pest control because plant defenses often target PMstructural components such as chitin and proteins. Hence, weperformed biochemical experiments to determine the PM proteintargets of Mir1-CP and conducted both targeted and global tran-scriptome analysis of MGs from fall armyworm larvae that fed oneither resistant or susceptible maize genotypes.
2. Materials and methods
2.1. Plant and insect material
Seed for the maize (Z. mays L.) inbreds, Mp708, Tx601 and B73,were supplied by the U.S. Department of Agriculture (USDA),Agriculture Research Service (ARS), Corn Host Plant ResistanceResearch Unit at Mississippi State University (Mississippi State,MS). In the experiments reported, Mp708 developed by Williamsand colleagues (Scott et al., 1982; Williams and Davis, 1982,1984;Williams et al., 1990; Williams and Davis, 2000,2002) was used asthe insect resistant inbred line. This inbred was selected froma cross between Tx601 and the resistant inbred Mp704 (Williamset al., 1990). The B73 and Tx601 inbred lines are susceptible toaboveground damage by several lepidopteran pests while theMp708 inbred is resistant to these insects and accumulatesMir1-CPin response to caterpillar feeding (Williams et al., 1990; Pechanet al., 2000; Lopez et al., 2007). The whorls of Tx601 and B73 donot accumulate Mir1-CP (Pechan et al., 2002) and both are highlysusceptible to fall armyworm herbivory. Seeds (one to two per pot)were planted in Hagerstown loam in 2.8 l or 5 l pots. Plants weregrown under conditions simulating Pennsylvania summer condi-tions in a greenhouse. Plants were harvested at approximately theV8 stage (Ritchie et al., 1992).
The USDA-ARS Laboratory at Mississippi State University alsosupplied fall armyworm eggs that hatched into larvae used in ex-periments measuring IIM degradation, MG transcripts by quanti-tative reverse-transcription polymerase chain reaction (qRT-PCR)and RNA-seq. Larvae used in the PM and qRT-PCR experimentswere reared on a casein-based artificial diet until the second instarwhen they were placed in 30 ml clear plastic diet cups lined on thebottom with 1% agar and containing excised yellow-green mid-whorl foliage. Old foliage and frass were removed every other dayand replaced with fresh foliage. Larvae were reared in an envi-ronmental chamber at 27 �C, 14:10 (light:dark) photoperiod, and70% relative humidity. In the qRT-PCR experiment, MGs were dis-sected from larvae in the penultimate instar.
Beginning as neonates, larvae used in the RNA-seq experimentwere reared on yellow-green midwhorl foliage as described above.Midguts were dissected from larvae 2 d after molting to the last

Fig. 1. In vitro and in vivo assays of insect intestinal mucin (IIM) degradation. Theperitrophic membrane (PM) gel (Coomassie Blue stained, 12% SDS-PAGE, 10 mg totalprotein per lane) shows PMs (lacking endogenous proteases) treated in vitro (PBS, pH7.4, 28 �C, 3 h) without (� Mir) or with (þMir, 35 mg) the protease Mir1-CP. Arrowsshow the most intensely stained bands. The frass gel (silver stained, 10% SDS-PAGE,5 mg of total protein per lane) shows protein extracted from frass of last instar fallarmyworm larvae that were fed on artificial diet or whorl tissue from susceptibleTx601 (lack Mir1-CP) or resistant Mp708 (have Mir1-CP) maize inbreds from thesecond instar. The frass western blot shows cross reaction with a polyclonal IIMantibody (Wang and Granados, 2003; 1:1000 dilution) following 10% SDS-PAGE sep-aration of proteins (5 mg of total protein per lane) extracted from frass excreted by lastinstar fall armyworm larvae fed whorl tissue from Tx601 or Mp708 maize inbreds.
H.W. Fescemyer et al. / Insect Biochemistry and Molecular Biology 43 (2013) 280e291282
instar with masses between 300 and 400 mg. A piece of theabdominal body cavity was used to determine that all larvae usedfor RNA-seq had the RFLP marker associated with the corn strain(Levy et al., 2002; Nagoshi et al., 2006).
Eggs of the European corn borer [Ostrinia nubilalis (Hübner)(Lepidoptera: Crambidae)] were purchased from Benzon Research(Carlisle, PA). Larvae used in experiments requiring MG transcriptsfor qRT-PCR were reared as described for fall armyworm, exceptlarvae were not placed on yellow-green midwhorl foliage until thefourth instar and MGs were dissected from last instars. Thisexperiment with the European corn borer determined the com-parative response of another maize pest to Mir1-CP.
Midguts used for RNA-seq and qRT-PCR experiments were dis-sected from CO2 or cold anesthetized larvae, respectively, whosedorsal side was then cut longitudinally from the first thoracicsegment to the last abdominal segment, pinned open, and sub-merged in Bombyx saline or 1� phosphate buffered saline (PBS),respectively. After removingMalpighian tubules, foregut anterior tothe stomodial valve, hindgut and food bolus, the MG was trans-ferred from the body cavity and rinsed well with cold saline. Mid-guts were preserved in RNAlater� (Applied Biosystems/Ambion,Austin, TX) according to the manufacturer’s instructions or flashfrozen in liquid nitrogen and stored at �80 �C, respectively.
2.2. In vitro assay of IIM degradation
The PMs surrounding the food bolus were removed fromMGs ofmid-last instar fall armyworm larvae. Foodwas gently washed fromeach PM with PBS and deionized water. They were frozen in liquidnitrogen immediately after isolation, and stored at �80 �C. Beforetreating PMs with purified (Mohan et al., 2006) recombinant Mir1-CP, their endogenous proteases were inactivate by boiling in 50 ml2% sodium dodecyl sulfate (SDS) for 5 min followed by extensivewashing with distilled water. Water was removed by centrifugationand careful blotting with filter paper before 50 PMs were incubatedwith 35 mg of Mir1-CP in 1 ml PBS buffer (pH 7.4) at 28 �C for 3 h. Aparallel treatment of PMs without Mir1-CP was included as a con-trol. After Mir1-CP treatment, PMs were washed extensively withdeionized water. Following water removal, they were homogenizedin sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) sample buffer (Laemmli, 1970) containing 1% CHAPSO andincubated in this buffer for 1 h at room temperature. Extractedprotein was denatured by heating at 95 �C for 10 min and insolublematerial removed by centrifugation at 12,000 � g. The resultingsupernatant was stored at �20 �C until analyzed. Protein concen-tration was determined using the RC DC� Protein Assay Kit (Bio-Rad Laboratories, Hercules, CA). Composition of protein in extractsfrom both control and Mir1-CP treated samples were analyzed bySDS-PAGE (Laemmli, 1970) with proteins visualized by stainingwith Coomassie-Brilliant Blue.
An abundant protein of about 400 kDa (Fig. 1), which wasseverely depleted in Mir1-CP treated PMs, was subjected to furtheranalysis. This protein band in the SDS-PAGE gel was excised, sub-jected to in-gel trypsin digestion and resultant peptides analyzedby two dimensional liquid chromatography tandem mass spec-trometry (2-D LC-MS/MS) as described previously (Pechanova et al.,2010). A BLASTp search via the National Center for BiotechnologyInformation (NCBI) against the non-redundant protein sequencesfrom Insecta in GenBank was used to align peptides deduced fromspectra to a protein from which they likely derived.
2.3. In vivo assay of IIM degradation
Degradation of IIM inside of MGs was examined with larvae fedon Tx601 or Mp708 yellow-green whorls beginning in the second
instar (50 larvae per inbred line). After 7 d on foliage, larvae weretransferred to individual rearing cups containing artificial diet andthe first ten frass pellets were collected into 100 ml SDS-PAGEsample buffer containing 1% CHAPSO. Frass-protein was extrac-ted, assayed for protein concentration and 5 mg of protein persample was separated with SDS-PAGE as described above.
Protein in one gel was detected by silver staining (ProteoSilverPlus� Silver Stain Kit, SigmaeAldrich, Saint Louis, MO) anda western blot was prepared from a duplicate gel. Hybridization ofa 1:1000 dilution of polyclonal IIM antibody (Wang and Granados,2003) to the blot was detected via chemiluminescent (Super-Signal� West Pico Chemiluminescent Substrate, Thermo Scientific/Pierce Biotechnology, Rockford, IL) exposure of photographic filmfor 30e60 s.
2.4. RNA extraction and cDNA synthesis
The RNA-seq and qRT-PCR experiments used somewhat differ-ent protocols for RNA extraction from MG tissue. In the RNA-seqexperiment, nearly all RNAlater� was removed and, after additionof several 2.3 mm diameter stainless steel beads and freezing inliquid nitrogen, tissue was disrupted and homogenized in TRIzol�
reagent (Invitrogen, Carlsbad, CA, USA) on a mixer mill (20 Hz, 45 swithout followed by 3 min with TRIzol� using adapter blocksprechilled at �20 �C). Isolation of RNA from the homogenate fol-lowed the TRIzol� manufacturer’s protocol, except for additionalacid phenol:chloroform:isoamyl alcohol (25:24:1, v:v:v) andchloroform phase separation steps prior to RNA precipitation toreduce DNA contamination. Agilent Bioanalysis (RNA Nano Chip,Agilent Technologies, Santa Clara, CA) indicated that the isolated

H.W. Fescemyer et al. / Insect Biochemistry and Molecular Biology 43 (2013) 280e291 283
total RNA had no detectable DNA contamination and RNA integritynumbers ranged from 8.3 to 9.7.
In the qRT-PCR experiment, TRIzol� (1 ml) was added to frozentissue that was immediately disrupted and homogenized on ice bytwo 30 s bouts at 10 W (rms) of continuous sonication (Microson�XL-2000 Ultrasonic Cell Disruptor, Misonix� by Qsonica, LLC,Newtown, CT). Isolation of RNA from the homogenate followed theTRIzol� manufacturer’s protocol. Total RNA was quantified byabsorbance at 260 nm and an aliquot was treated with RQ1 RNase-free DNAse (Promega, Madison, WI) followed by heat inactivationof the DNase. First strand cDNA synthesis was done using the High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems), 1 mgof DNAse-treated RNA and the kit’s random primers supplementedwith Oligo(dT)20 at 2.5 mM.
2.5. qRT-PCR of targeted MG transcripts
Based on previous reports (Pechan et al., 2000; Mohan et al.,2006,2008), genes involved in PM synthesis and digestion wereprimarily targeted for measurement of expression by qRT-PCR(Table 1). Nucleotide sequences for target genes in fall armyworm(Table 1; Supplemental Tables 1e3) were obtained from a closelyrelated species, H. armigera, and used in a tBLASTx search againstexpressed sequence tags (ESTs) in SPODOBASE (Landais et al., 2003;Negre et al., 2006; http://bioweb.ensam.inra.fr/spodobase) to findthe corresponding sequence in fall armyworm based on thosehaving the highest bit score. A similar approach was used for Eu-ropean corn borer except that target genes from H. armigera wereused in a tBLASTx search against only ESTs from European cornborer in GenBank. These fall armyworm- and European corn borer-sequences were then used in designing primers (SupplementalTable 4) for qRT-PCR with Primer Express (Applied Biosystems) orPrimerQuestSM (Integrated DNA Technologies, Coralville, IA).Functional annotation and classification of coding sequences usedin designing primers for qRT-PCR (Table 1; Supplemental Tables 1e3) was performed via BLASTx against the Gene Ontology (GO)database (The Gene Ontology Consortium, 2000; release 2012-07-14) using AmiGo (Carbon et al., 2009; version 1.8). A putative namefor each candidate gene was based on this annotation.
All primers (Supplemental Table 4) used in qRT-PCR had a Tm ofapproximately 60 �C, formed minimal secondary structures, andproduced amplicons between 50 and 150 bp. Cloning, Sangersequencing and BLAST annotation were used to confirm all ampli-cons. Simplex qRT-PCR assays were performed on an Applied Bio-systems 7500 Fast Real-Time PCR Instrument operated in fast modeand using amplicon detectionwith SYBR�Green. Reactions for eachcDNA sample had a final volume of 10 ml using 2 ml of cDNA diluted1:10, 500 nM forward and reverse primers, and 1� Fast SYBR�
Green Master Mix (Applied Biosystems). Each target gene PCR wasaccompanied in the same plate by an endogenous, 18S referencePCR. All sample and control reactions (i.e., NAC and NTC) were runin triplicate under the following manufacturer’s recommendedconditions; 95 �C for 20 s, and 40 cycles of 95 �C for 3 s and 60 �C for30 s. At the end of each run, a dissociation curve was generated foreach reaction using the instrument’s default thermocycle program.Single peaks and smoothly shaped graphs of the negative first de-rivative of the melting-curve were always obtained. Validationexperiments were performed for the endogenous reference geneand each target gene using serial dilutions (ranging from undilutedto a 1:10,000 dilution) of a sample formed by pooling an aliquot ofcDNA derived from each of the MGs from larvae that fed on Mp708foliage. Slopes of the curves obtained from these validation ex-periments were very similar to �3.3 and had R2 values �0.97.Relative expression of target genes was calculated using the com-parative CT method where amount of target, normalized to the 18S
rRNA endogenous reference and relative to a calibrator (larvae fedartificial diet) is given as the fold expression difference, or 2�DDCT
(Livak and Schmittgen, 2001).
2.6. RNA-seq
Equal amounts (3 mg) of total RNA from an individual MG wererandomly pooled into three replicates per treatment (i.e., Mp708 orTx601) such that each treatment replicate derived from 12 to 13MGs. Each pool of total RNA was separately enriched for poly(Aþ)RNA using the MicroPoly(A)Purist� Kit (Applied Biosystems/Ambion) according to the manufacturer’s instructions. Enrichmentinvolved two rounds of oligo(dT) selectionwith a fresh spin columnused in each round. The resulting poly(Aþ) showed little degrada-tion of the mRNAwith a broad peak ranging from 200 nt to 5000 ntcentered at about 1880 nt (Agilent Bioanalyzer RNA Pico Chip,Agilent Technologies). Estimated rRNA contamination ranged from3.2% to 6.1%.
The Penn State Genomics Core Facility (University Park, PA)prepared transcriptome libraries and performed sequencing. Li-braries were separately prepared from each of the six poly(Aþ) RNAsamples using the strand-specific SOLiD� Total RNA-Seq Kit(Applied Biosystems) according to the manufacturer’s protocol.Briefly, samples were fragmented with RNAse III and purified.Fragmented RNA (100 ng) was ligated overnight to adaptors andsubsequently reverse transcribed. Purified cDNA was size selectedand then amplified using barcoded primers. The resulting materialwas purified and assessed for proper amplification using an AgilentBioanalyzer DNA 1000 chip (Agilent Technologies).
Barcoded libraries were combined in equimolar quantities andprepared for sequencing according to the “Applied BiosystemsSOLiD 3 Plus System Templated Bead Preparation Guide.” Barcodesequencing and fragment library sequencing (50 bp) was per-formed on a full plate run of the SOLiD 3 Plus System according tothe “Applied Biosystems SOLiD 3 Plus System Instrument OperationGuide.”
The 138911 Sanger ESTs in SPODOBASE (Landais et al., 2003;Negre et al., 2006) were assembled into a reference transcriptomeusing SeqMan Pro version 8.0.2 (DNASTAR Lasergene�, Madison,WI) and default parameters as recommended by the manufacturerfor Sanger sequences. Preassembly involved a medium end trim,a vector scan and contaminant sequence screen that used a mini-mum of 5 mismatches; vector and contaminant sequence catalogswere those provided by the software. Assembly parameters werematch size of 25, minimum match of 85%, match spacing of 150,minimum sequence length of 100, gap penalty of zero, gap length of0.7 and maximum mismatch end bases of 15. Consensus callingused primary trace evidence of 50%, a majority of 75%, and qualityweights of nine before and 400 after. Calling conflicts and singlenucleotide polymorphisms (SNPs) used a minimum coverage offour, minimum inconsistency of 25% and two bases and no pairingwas specified. The assembled reference contained 7869 contigs(7,049,947 nt). Hereafter, contigs are called genes but more cor-rectly should be considered gene models.
SOLiD sequence reads were filtered and mapped (both sum-marized in Supplemental Table 5) using NextGENe� version 2.0(SoftGenetics, State College, PA). The 384 million reads(64 � 4.6 million reads per library) were filtered to accept readswhose median score threshold was �12 (ca. 94% accuracy), con-tained�25 bases and contained one ormore baseswith a score�14(ca. 96% accuracy). There were 164 million reads (27 � 1.9 millionreads per library) passing this filtering. The reads from each libraryrepresenting a replicate within a maize inbred treatment weremapped separately to the reference transcriptome using theBowtie-like algorithm in NextGENe� with the requirement that
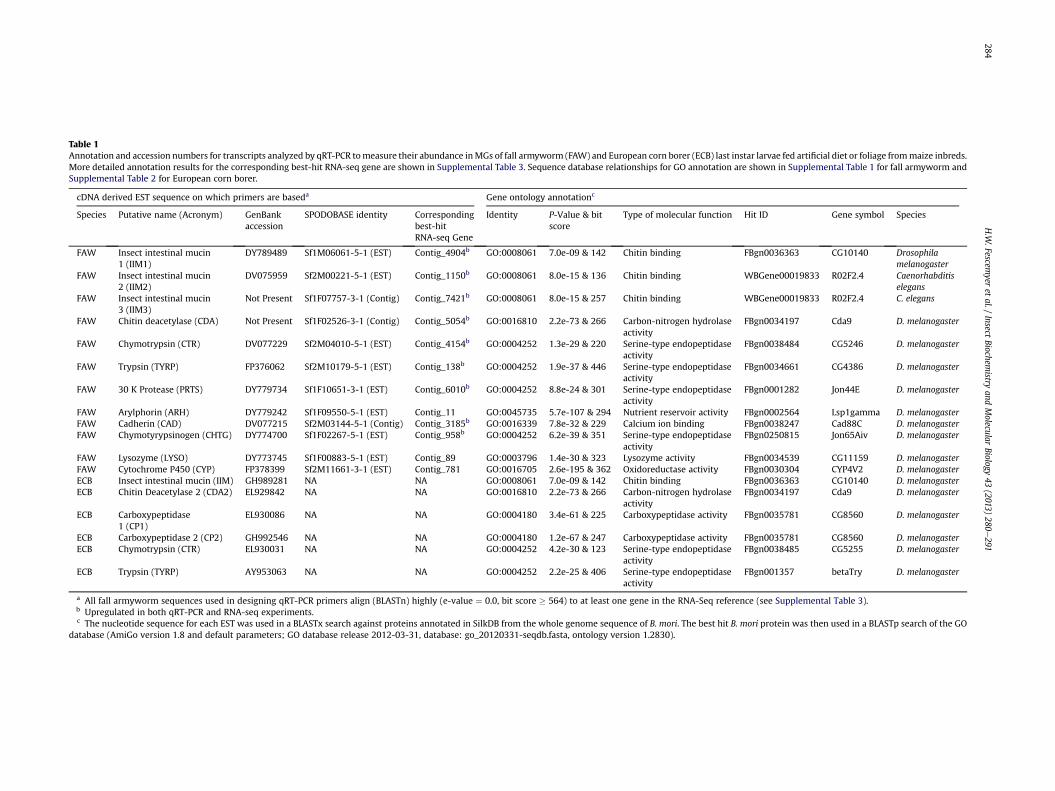
Table 1Annotation and accession numbers for transcripts analyzed by qRT-PCR tomeasure their abundance inMGs of fall armyworm (FAW) and European corn borer (ECB) last instar larvae fed artificial diet or foliage frommaize inbreds.More detailed annotation results for the corresponding best-hit RNA-seq gene are shown in Supplemental Table 3. Sequence database relationships for GO annotation are shown in Supplemental Table 1 for fall armyworm andSupplemental Table 2 for European corn borer.
cDNA derived EST sequence on which primers are baseda Gene ontology annotationc
Species Putative name (Acronym) GenBankaccession
SPODOBASE identity Correspondingbest-hitRNA-seq Gene
Identity P-Value & bitscore
Type of molecular function Hit ID Gene symbol Species
FAW Insect intestinal mucin1 (IIM1)
DY789489 Sf1M06061-5-1 (EST) Contig_4904b GO:0008061 7.0e-09 & 142 Chitin binding FBgn0036363 CG10140 Drosophilamelanogaster
FAW Insect intestinal mucin2 (IIM2)
DV075959 Sf2M00221-5-1 (EST) Contig_1150b GO:0008061 8.0e-15 & 136 Chitin binding WBGene00019833 R02F2.4 Caenorhabditiselegans
FAW Insect intestinal mucin3 (IIM3)
Not Present Sf1F07757-3-1 (Contig) Contig_7421b GO:0008061 8.0e-15 & 257 Chitin binding WBGene00019833 R02F2.4 C. elegans
FAW Chitin deacetylase (CDA) Not Present Sf1F02526-3-1 (Contig) Contig_5054b GO:0016810 2.2e-73 & 266 Carbon-nitrogen hydrolaseactivity
FBgn0034197 Cda9 D. melanogaster
FAW Chymotrypsin (CTR) DV077229 Sf2M04010-5-1 (EST) Contig_4154b GO:0004252 1.3e-29 & 220 Serine-type endopeptidaseactivity
FBgn0038484 CG5246 D. melanogaster
FAW Trypsin (TYRP) FP376062 Sf2M10179-5-1 (EST) Contig_138b GO:0004252 1.9e-37 & 446 Serine-type endopeptidaseactivity
FBgn0034661 CG4386 D. melanogaster
FAW 30 K Protease (PRTS) DY779734 Sf1F10651-3-1 (EST) Contig_6010b GO:0004252 8.8e-24 & 301 Serine-type endopeptidaseactivity
FBgn0001282 Jon44E D. melanogaster
FAW Arylphorin (ARH) DY779242 Sf1F09550-5-1 (EST) Contig_11 GO:0045735 5.7e-107 & 294 Nutrient reservoir activity FBgn0002564 Lsp1gamma D. melanogasterFAW Cadherin (CAD) DV077215 Sf2M03144-5-1 (Contig) Contig_3185b GO:0016339 7.8e-32 & 229 Calcium ion binding FBgn0038247 Cad88C D. melanogasterFAW Chymotyrypsinogen (CHTG) DY774700 Sf1F02267-5-1 (EST) Contig_958b GO:0004252 6.2e-39 & 351 Serine-type endopeptidase
activityFBgn0250815 Jon65Aiv D. melanogaster
FAW Lysozyme (LYSO) DY773745 Sf1F00883-5-1 (EST) Contig_89 GO:0003796 1.4e-30 & 323 Lysozyme activity FBgn0034539 CG11159 D. melanogasterFAW Cytochrome P450 (CYP) FP378399 Sf2M11661-3-1 (EST) Contig_781 GO:0016705 2.6e-195 & 362 Oxidoreductase activity FBgn0030304 CYP4V2 D. melanogasterECB Insect intestinal mucin (IIM) GH989281 NA NA GO:0008061 7.0e-09 & 142 Chitin binding FBgn0036363 CG10140 D. melanogasterECB Chitin Deacetylase 2 (CDA2) EL929842 NA NA GO:0016810 2.2e-73 & 266 Carbon-nitrogen hydrolase
activityFBgn0034197 Cda9 D. melanogaster
ECB Carboxypeptidase1 (CP1)
EL930086 NA NA GO:0004180 3.4e-61 & 225 Carboxypeptidase activity FBgn0035781 CG8560 D. melanogaster
ECB Carboxypeptidase 2 (CP2) GH992546 NA NA GO:0004180 1.2e-67 & 247 Carboxypeptidase activity FBgn0035781 CG8560 D. melanogasterECB Chymotrypsin (CTR) EL930031 NA NA GO:0004252 4.2e-30 & 123 Serine-type endopeptidase
activityFBgn0038485 CG5255 D. melanogaster
ECB Trypsin (TYRP) AY953063 NA NA GO:0004252 2.2e-25 & 406 Serine-type endopeptidaseactivity
FBgn001357 betaTry D. melanogaster
a All fall armyworm sequences used in designing qRT-PCR primers align (BLASTn) highly (e-value ¼ 0.0, bit score � 564) to at least one gene in the RNA-Seq reference (see Supplemental Table 3).b Upregulated in both qRT-PCR and RNA-seq experiments.c The nucleotide sequence for each EST was used in a BLASTx search against proteins annotated in SilkDB from the whole genome sequence of B. mori. The best hit B. mori protein was then used in a BLASTp search of the GO
database (AmiGo version 1.8 and default parameters; GO database release 2012-03-31, database: go_20120331-seqdb.fasta, ontology version 1.2830).
H.W
.Fescemyer
etal./
InsectBiochem
istryand
Molecular
Biology43
(2013)280
e291
284
H.W. Fescemyer et al. / Insect Biochemistry and Molecular Biology 43 (2013) 280e291 285
85% of 12 or more nucleotides comprising a read must match thereference. A read was allowed to map only once (i.e., no ambiguousmapping). Of reads passing through filtering, 109 million(18 � 1.2 million reads per library) mapped to the reference withcoverage of 163 � 8 reads/gene/library, read length of 34 � 0.2 nt,and 3.6 � 0.06 million of the 7,049,947 nt in the reference covered.The number of mapped reads per gene in each library, representinga replicate within a maize inbred treatment, were summed byNextGENe� as read counts per gene. These data representingtranscript abundance were subsequently used in data analyses. Theraw sequence data (NCBI short read archive [SRA] accessionSRP017381), processed data (filtered and mapped reads), finalprocessed data (filtered and normalized read counts along withreference contig sequences and annotation), and metadata fromthis study were deposited to the NCBI Gene Expression Omnibus(GEO) under accession number GSE38744.
2.7. Data analyses
Values for fold difference from qRT-PCR data were natural log-arithm transformed to increase homogeneity of variance. A stan-dard least squares general linearmodel (GLM) considering the fixedfactor food (N ¼ 4; larvae fed artificial diet or foliage from maizeinbreds Mp708, Tx601, B73) with three replicates per food treat-ment was calculated for each genewhose transcript abundancewasmeasured in the qRT-PCR experiment. Tukey’s test was used todetermine significant differences (a ¼ 0.05) between the means ofmain effects. Probability (P) values in the results section are for theindependent effects type III sums of squares F tests from theseGLMs performedwith JMP�, version 5 (SAS Institute Inc., Cary, NC).
For the RNA-seq analysis, we used the R software package(v2.15.0) to confirm the overall data quality (frequency distributionhistogram plots, hexagonal binning plots with R hexbin packagev1.26.0, MA plots with R edgeR package v2.6.7 (Robinson et al.,2010), complete linkage cluster dendrogram plots, frequency his-tograms of Fisher’s exact test P-values with R SAGEnhaft packagev1.22.0), filter low expressing genes, and normalize the read counts.Low expressing genes were filtered by summing raw read countsper gene across replicate libraries within a maize inbred treatment;
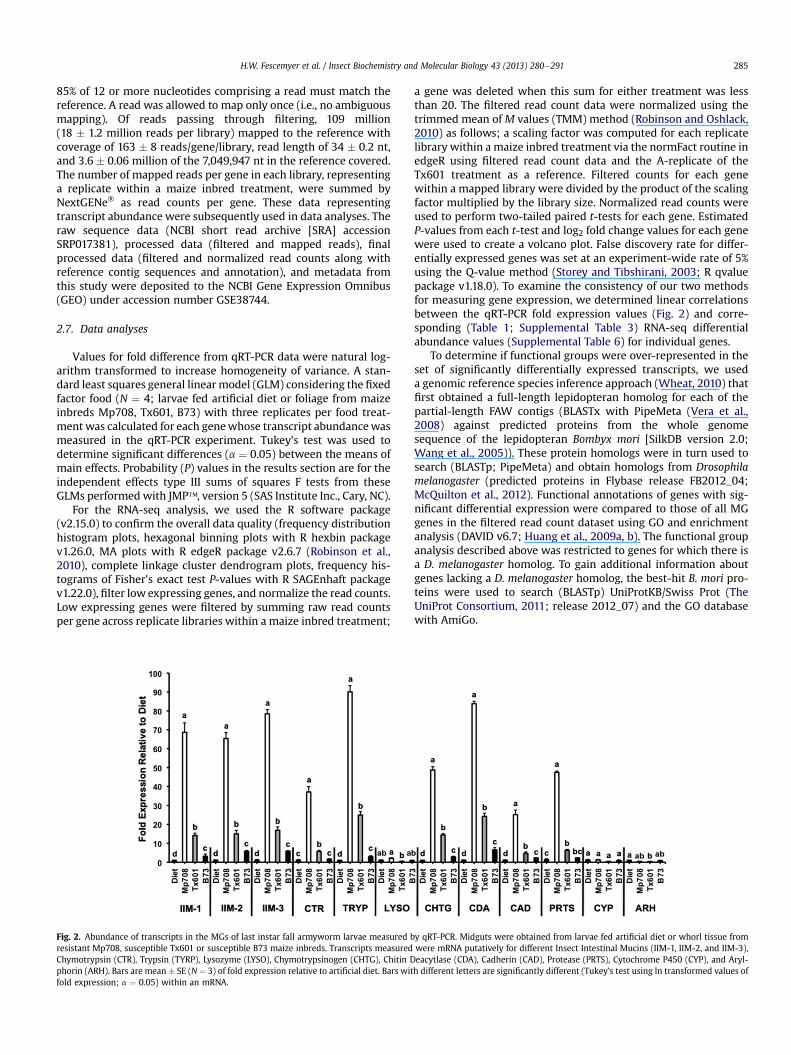
Fig. 2. Abundance of transcripts in the MGs of last instar fall armyworm larvae measured bresistant Mp708, susceptible Tx601 or susceptible B73 maize inbreds. Transcripts measuredChymotrypsin (CTR), Trypsin (TYRP), Lysozyme (LYSO), Chymotrypsinogen (CHTG), Chitin Dphorin (ARH). Bars are mean � SE (N ¼ 3) of fold expression relative to artificial diet. Bars witfold expression; a ¼ 0.05) within an mRNA.
a gene was deleted when this sum for either treatment was lessthan 20. The filtered read count data were normalized using thetrimmed mean of M values (TMM) method (Robinson and Oshlack,2010) as follows; a scaling factor was computed for each replicatelibrary within a maize inbred treatment via the normFact routine inedgeR using filtered read count data and the A-replicate of theTx601 treatment as a reference. Filtered counts for each genewithin a mapped library were divided by the product of the scalingfactor multiplied by the library size. Normalized read counts wereused to perform two-tailed paired t-tests for each gene. EstimatedP-values from each t-test and log2 fold change values for each genewere used to create a volcano plot. False discovery rate for differ-entially expressed genes was set at an experiment-wide rate of 5%using the Q-value method (Storey and Tibshirani, 2003; R qvaluepackage v1.18.0). To examine the consistency of our two methodsfor measuring gene expression, we determined linear correlationsbetween the qRT-PCR fold expression values (Fig. 2) and corre-sponding (Table 1; Supplemental Table 3) RNA-seq differentialabundance values (Supplemental Table 6) for individual genes.
To determine if functional groups were over-represented in theset of significantly differentially expressed transcripts, we useda genomic reference species inference approach (Wheat, 2010) thatfirst obtained a full-length lepidopteran homolog for each of thepartial-length FAW contigs (BLASTx with PipeMeta (Vera et al.,2008) against predicted proteins from the whole genomesequence of the lepidopteran Bombyx mori [SilkDB version 2.0;Wang et al., 2005)). These protein homologs were in turn used tosearch (BLASTp; PipeMeta) and obtain homologs from Drosophilamelanogaster (predicted proteins in Flybase release FB2012_04;McQuilton et al., 2012). Functional annotations of genes with sig-nificant differential expression were compared to those of all MGgenes in the filtered read count dataset using GO and enrichmentanalysis (DAVID v6.7; Huang et al., 2009a, b). The functional groupanalysis described above was restricted to genes for which there isa D. melanogaster homolog. To gain additional information aboutgenes lacking a D. melanogaster homolog, the best-hit B. mori pro-teins were used to search (BLASTp) UniProtKB/Swiss Prot (TheUniProt Consortium, 2011; release 2012_07) and the GO databasewith AmiGo.
y qRT-PCR. Midguts were obtained from larvae fed artificial diet or whorl tissue fromwere mRNA putatively for different Insect Intestinal Mucins (IIM-1, IIM-2, and IIM-3),eacytlase (CDA), Cadherin (CAD), Protease (PRTS), Cytochrome P450 (CYP), and Aryl-h different letters are significantly different (Tukey’s test using ln transformed values of
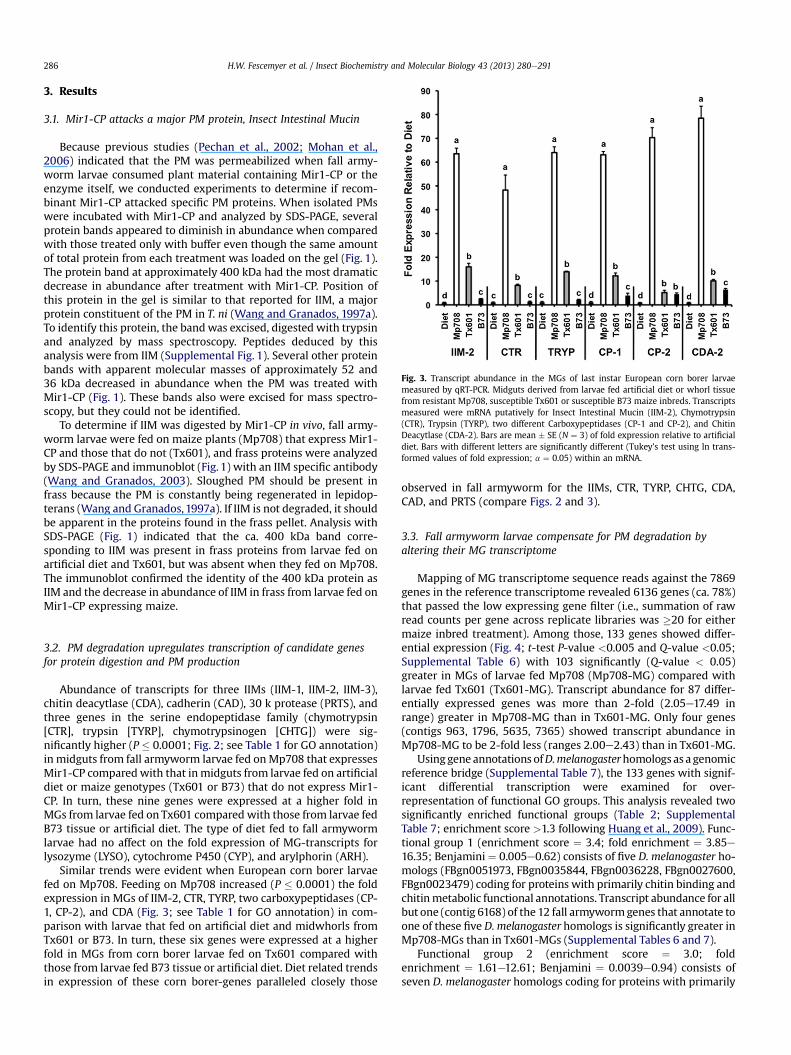
Fig. 3. Transcript abundance in the MGs of last instar European corn borer larvaemeasured by qRT-PCR. Midguts derived from larvae fed artificial diet or whorl tissuefrom resistant Mp708, susceptible Tx601 or susceptible B73 maize inbreds. Transcriptsmeasured were mRNA putatively for Insect Intestinal Mucin (IIM-2), Chymotrypsin(CTR), Trypsin (TYRP), two different Carboxypeptidases (CP-1 and CP-2), and ChitinDeacytlase (CDA-2). Bars are mean � SE (N ¼ 3) of fold expression relative to artificialdiet. Bars with different letters are significantly different (Tukey’s test using ln trans-formed values of fold expression; a ¼ 0.05) within an mRNA.
H.W. Fescemyer et al. / Insect Biochemistry and Molecular Biology 43 (2013) 280e291286
3. Results
3.1. Mir1-CP attacks a major PM protein, Insect Intestinal Mucin
Because previous studies (Pechan et al., 2002; Mohan et al.,2006) indicated that the PM was permeabilized when fall army-worm larvae consumed plant material containing Mir1-CP or theenzyme itself, we conducted experiments to determine if recom-binant Mir1-CP attacked specific PM proteins. When isolated PMswere incubated with Mir1-CP and analyzed by SDS-PAGE, severalprotein bands appeared to diminish in abundance when comparedwith those treated only with buffer even though the same amountof total protein from each treatment was loaded on the gel (Fig. 1).The protein band at approximately 400 kDa had the most dramaticdecrease in abundance after treatment with Mir1-CP. Position ofthis protein in the gel is similar to that reported for IIM, a majorprotein constituent of the PM in T. ni (Wang and Granados, 1997a).To identify this protein, the band was excised, digested with trypsinand analyzed by mass spectroscopy. Peptides deduced by thisanalysis were from IIM (Supplemental Fig. 1). Several other proteinbands with apparent molecular masses of approximately 52 and36 kDa decreased in abundance when the PM was treated withMir1-CP (Fig. 1). These bands also were excised for mass spectro-scopy, but they could not be identified.
To determine if IIM was digested by Mir1-CP in vivo, fall army-worm larvae were fed on maize plants (Mp708) that express Mir1-CP and those that do not (Tx601), and frass proteins were analyzedby SDS-PAGE and immunoblot (Fig. 1) with an IIM specific antibody(Wang and Granados, 2003). Sloughed PM should be present infrass because the PM is constantly being regenerated in lepidop-terans (Wang and Granados, 1997a). If IIM is not degraded, it shouldbe apparent in the proteins found in the frass pellet. Analysis withSDS-PAGE (Fig. 1) indicated that the ca. 400 kDa band corre-sponding to IIM was present in frass proteins from larvae fed onartificial diet and Tx601, but was absent when they fed on Mp708.The immunoblot confirmed the identity of the 400 kDa protein asIIM and the decrease in abundance of IIM in frass from larvae fed onMir1-CP expressing maize.
3.2. PM degradation upregulates transcription of candidate genesfor protein digestion and PM production
Abundance of transcripts for three IIMs (IIM-1, IIM-2, IIM-3),chitin deacytlase (CDA), cadherin (CAD), 30 k protease (PRTS), andthree genes in the serine endopeptidase family (chymotrypsin[CTR], trypsin [TYRP], chymotrypsinogen [CHTG]) were sig-nificantly higher (P � 0.0001; Fig. 2; see Table 1 for GO annotation)in midguts from fall armyworm larvae fed onMp708 that expressesMir1-CP comparedwith that inmidguts from larvae fed on artificialdiet or maize genotypes (Tx601 or B73) that do not express Mir1-CP. In turn, these nine genes were expressed at a higher fold inMGs from larvae fed on Tx601 compared with those from larvae fedB73 tissue or artificial diet. The type of diet fed to fall armywormlarvae had no affect on the fold expression of MG-transcripts forlysozyme (LYSO), cytochrome P450 (CYP), and arylphorin (ARH).
Similar trends were evident when European corn borer larvaefed on Mp708. Feeding on Mp708 increased (P � 0.0001) the foldexpression in MGs of IIM-2, CTR, TYRP, two carboxypeptidases (CP-1, CP-2), and CDA (Fig. 3; see Table 1 for GO annotation) in com-parison with larvae that fed on artificial diet and midwhorls fromTx601 or B73. In turn, these six genes were expressed at a higherfold in MGs from corn borer larvae fed on Tx601 compared withthose from larvae fed B73 tissue or artificial diet. Diet related trendsin expression of these corn borer-genes paralleled closely those
observed in fall armyworm for the IIMs, CTR, TYRP, CHTG, CDA,CAD, and PRTS (compare Figs. 2 and 3).
3.3. Fall armyworm larvae compensate for PM degradation byaltering their MG transcriptome
Mapping of MG transcriptome sequence reads against the 7869genes in the reference transcriptome revealed 6136 genes (ca. 78%)that passed the low expressing gene filter (i.e., summation of rawread counts per gene across replicate libraries was �20 for eithermaize inbred treatment). Among those, 133 genes showed differ-ential expression (Fig. 4; t-test P-value <0.005 and Q-value <0.05;Supplemental Table 6) with 103 significantly (Q-value < 0.05)greater in MGs of larvae fed Mp708 (Mp708-MG) compared withlarvae fed Tx601 (Tx601-MG). Transcript abundance for 87 differ-entially expressed genes was more than 2-fold (2.05e17.49 inrange) greater in Mp708-MG than in Tx601-MG. Only four genes(contigs 963, 1796, 5635, 7365) showed transcript abundance inMp708-MG to be 2-fold less (ranges 2.00e2.43) than in Tx601-MG.
Usinggene annotations ofD.melanogasterhomologs as a genomicreference bridge (Supplemental Table 7), the 133 genes with signif-icant differential transcription were examined for over-representation of functional GO groups. This analysis revealed twosignificantly enriched functional groups (Table 2; SupplementalTable 7; enrichment score >1.3 following Huang et al., 2009). Func-tional group 1 (enrichment score ¼ 3.4; fold enrichment ¼ 3.85e16.35; Benjamini ¼ 0.005e0.62) consists of five D. melanogaster ho-mologs (FBgn0051973, FBgn0035844, FBgn0036228, FBgn0027600,FBgn0023479) coding for proteins with primarily chitin binding andchitinmetabolic functional annotations. Transcript abundance for allbut one (contig 6168) of the 12 fall armywormgenes that annotate toone of these five D. melanogaster homologs is significantly greater inMp708-MGs than in Tx601-MGs (Supplemental Tables 6 and 7).
Functional group 2 (enrichment score ¼ 3.0; foldenrichment ¼ 1.61e12.61; Benjamini ¼ 0.0039e0.94) consists ofseven D. melanogaster homologs coding for proteins with primarily
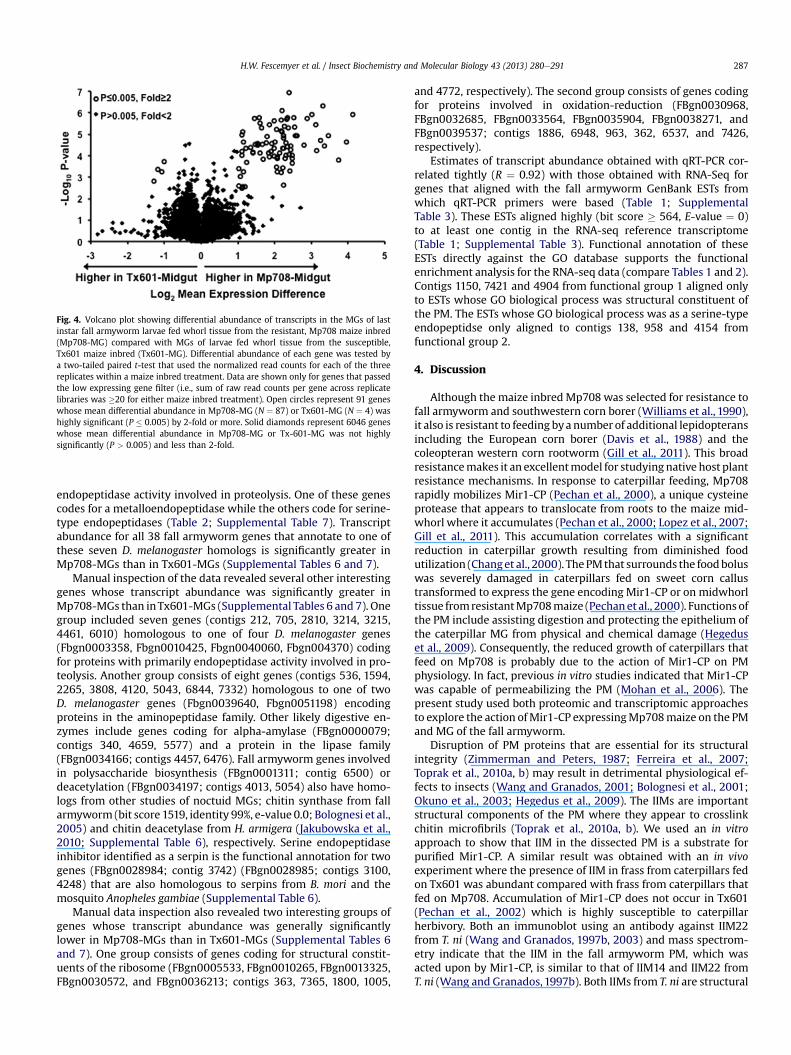
Fig. 4. Volcano plot showing differential abundance of transcripts in the MGs of lastinstar fall armyworm larvae fed whorl tissue from the resistant, Mp708 maize inbred(Mp708-MG) compared with MGs of larvae fed whorl tissue from the susceptible,Tx601 maize inbred (Tx601-MG). Differential abundance of each gene was tested bya two-tailed paired t-test that used the normalized read counts for each of the threereplicates within a maize inbred treatment. Data are shown only for genes that passedthe low expressing gene filter (i.e., sum of raw read counts per gene across replicatelibraries was �20 for either maize inbred treatment). Open circles represent 91 geneswhose mean differential abundance in Mp708-MG (N ¼ 87) or Tx601-MG (N ¼ 4) washighly significant (P � 0.005) by 2-fold or more. Solid diamonds represent 6046 geneswhose mean differential abundance in Mp708-MG or Tx-601-MG was not highlysignificantly (P > 0.005) and less than 2-fold.
H.W. Fescemyer et al. / Insect Biochemistry and Molecular Biology 43 (2013) 280e291 287
endopeptidase activity involved in proteolysis. One of these genescodes for a metalloendopeptidase while the others code for serine-type endopeptidases (Table 2; Supplemental Table 7). Transcriptabundance for all 38 fall armyworm genes that annotate to one ofthese seven D. melanogaster homologs is significantly greater inMp708-MGs than in Tx601-MGs (Supplemental Tables 6 and 7).
Manual inspection of the data revealed several other interestinggenes whose transcript abundance was significantly greater inMp708-MGs than inTx601-MGs (Supplemental Tables 6 and 7). Onegroup included seven genes (contigs 212, 705, 2810, 3214, 3215,4461, 6010) homologous to one of four D. melanogaster genes(Fbgn0003358, Fbgn0010425, Fbgn0040060, Fbgn004370) codingfor proteins with primarily endopeptidase activity involved in pro-teolysis. Another group consists of eight genes (contigs 536, 1594,2265, 3808, 4120, 5043, 6844, 7332) homologous to one of twoD. melanogaster genes (Fbgn0039640, Fbgn0051198) encodingproteins in the aminopeptidase family. Other likely digestive en-zymes include genes coding for alpha-amylase (FBgn0000079;contigs 340, 4659, 5577) and a protein in the lipase family(FBgn0034166; contigs 4457, 6476). Fall armyworm genes involvedin polysaccharide biosynthesis (FBgn0001311; contig 6500) ordeacetylation (FBgn0034197; contigs 4013, 5054) also have homo-logs from other studies of noctuid MGs; chitin synthase from fallarmyworm (bit score 1519, identity 99%, e-value 0.0; Bolognesi et al.,2005) and chitin deacetylase from H. armigera (Jakubowska et al.,2010; Supplemental Table 6), respectively. Serine endopeptidaseinhibitor identified as a serpin is the functional annotation for twogenes (FBgn0028984; contig 3742) (FBgn0028985; contigs 3100,4248) that are also homologous to serpins from B. mori and themosquito Anopheles gambiae (Supplemental Table 6).
Manual data inspection also revealed two interesting groups ofgenes whose transcript abundance was generally significantlylower in Mp708-MGs than in Tx601-MGs (Supplemental Tables 6and 7). One group consists of genes coding for structural constit-uents of the ribosome (FBgn0005533, FBgn0010265, FBgn0013325,FBgn0030572, and FBgn0036213; contigs 363, 7365, 1800, 1005,
and 4772, respectively). The second group consists of genes codingfor proteins involved in oxidation-reduction (FBgn0030968,FBgn0032685, FBgn0033564, FBgn0035904, FBgn0038271, andFBgn0039537; contigs 1886, 6948, 963, 362, 6537, and 7426,respectively).
Estimates of transcript abundance obtained with qRT-PCR cor-related tightly (R ¼ 0.92) with those obtained with RNA-Seq forgenes that aligned with the fall armyworm GenBank ESTs fromwhich qRT-PCR primers were based (Table 1; SupplementalTable 3). These ESTs aligned highly (bit score � 564, E-value ¼ 0)to at least one contig in the RNA-seq reference transcriptome(Table 1; Supplemental Table 3). Functional annotation of theseESTs directly against the GO database supports the functionalenrichment analysis for the RNA-seq data (compare Tables 1 and 2).Contigs 1150, 7421 and 4904 from functional group 1 aligned onlyto ESTs whose GO biological process was structural constituent ofthe PM. The ESTs whose GO biological process was as a serine-typeendopeptidse only aligned to contigs 138, 958 and 4154 fromfunctional group 2.
4. Discussion
Although the maize inbredMp708 was selected for resistance tofall armyworm and southwestern corn borer (Williams et al., 1990),it also is resistant to feeding bya number of additional lepidopteransincluding the European corn borer (Davis et al., 1988) and thecoleopteran western corn rootworm (Gill et al., 2011). This broadresistancemakes it an excellentmodel for studyingnative host plantresistance mechanisms. In response to caterpillar feeding, Mp708rapidly mobilizes Mir1-CP (Pechan et al., 2000), a unique cysteineprotease that appears to translocate from roots to the maize mid-whorl where it accumulates (Pechan et al., 2000; Lopez et al., 2007;Gill et al., 2011). This accumulation correlates with a significantreduction in caterpillar growth resulting from diminished foodutilization (Changet al., 2000). ThePMthat surrounds the foodboluswas severely damaged in caterpillars fed on sweet corn callustransformed to express the gene encoding Mir1-CP or on midwhorltissue fromresistantMp708maize (Pechanet al., 2000). Functions ofthe PM include assisting digestion and protecting the epithelium ofthe caterpillar MG from physical and chemical damage (Hegeduset al., 2009). Consequently, the reduced growth of caterpillars thatfeed on Mp708 is probably due to the action of Mir1-CP on PMphysiology. In fact, previous in vitro studies indicated that Mir1-CPwas capable of permeabilizing the PM (Mohan et al., 2006). Thepresent study used both proteomic and transcriptomic approachesto explore the action ofMir1-CP expressingMp708maize on the PMand MG of the fall armyworm.
Disruption of PM proteins that are essential for its structuralintegrity (Zimmerman and Peters, 1987; Ferreira et al., 2007;Toprak et al., 2010a, b) may result in detrimental physiological ef-fects to insects (Wang and Granados, 2001; Bolognesi et al., 2001;Okuno et al., 2003; Hegedus et al., 2009). The IIMs are importantstructural components of the PM where they appear to crosslinkchitin microfibrils (Toprak et al., 2010a, b). We used an in vitroapproach to show that IIM in the dissected PM is a substrate forpurified Mir1-CP. A similar result was obtained with an in vivoexperiment where the presence of IIM in frass from caterpillars fedon Tx601 was abundant compared with frass from caterpillars thatfed on Mp708. Accumulation of Mir1-CP does not occur in Tx601(Pechan et al., 2002) which is highly susceptible to caterpillarherbivory. Both an immunoblot using an antibody against IIM22from T. ni (Wang and Granados, 1997b, 2003) and mass spectrom-etry indicate that the IIM in the fall armyworm PM, which wasacted upon by Mir1-CP, is similar to that of IIM14 and IIM22 fromT. ni (Wang and Granados, 1997b). Both IIMs from T. ni are structural
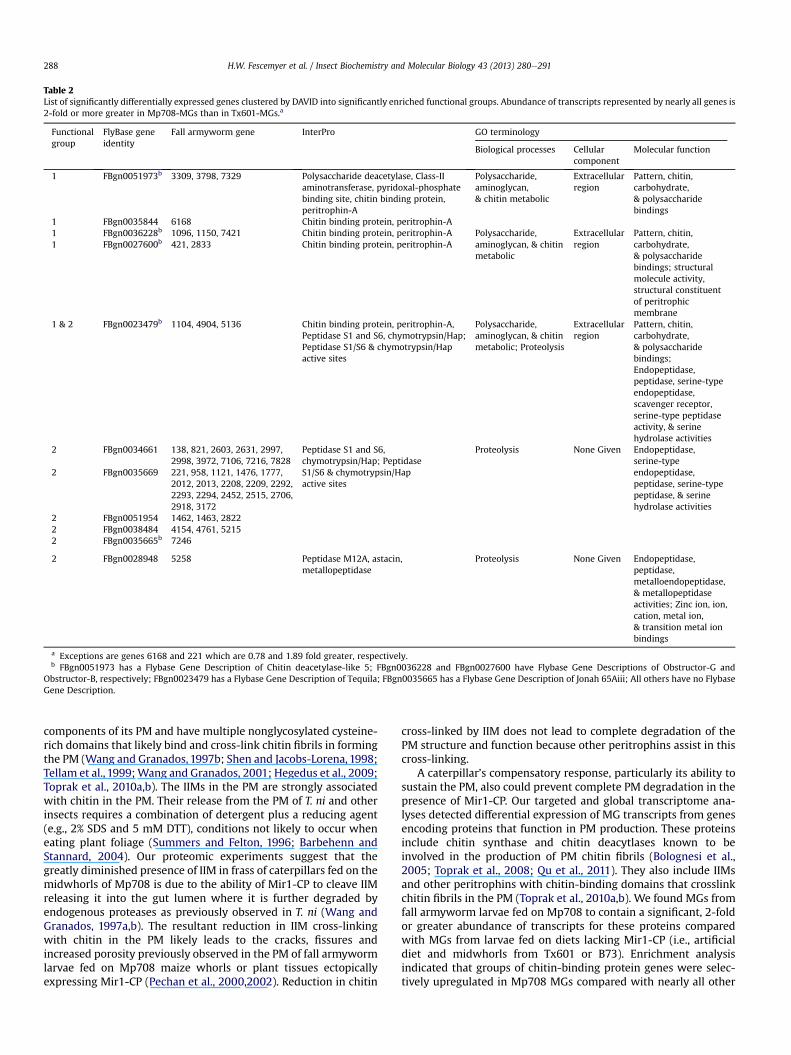
Table 2List of significantly differentially expressed genes clustered by DAVID into significantly enriched functional groups. Abundance of transcripts represented by nearly all genes is2-fold or more greater in Mp708-MGs than in Tx601-MGs.a
Functionalgroup
FlyBase geneidentity
Fall armyworm gene InterPro GO terminology
Biological processes Cellularcomponent
Molecular function
1 FBgn0051973b 3309, 3798, 7329 Polysaccharide deacetylase, Class-IIaminotransferase, pyridoxal-phosphatebinding site, chitin binding protein,peritrophin-A
Polysaccharide,aminoglycan,& chitin metabolic
Extracellularregion
Pattern, chitin,carbohydrate,& polysaccharidebindings
1 FBgn0035844 6168 Chitin binding protein, peritrophin-A1 FBgn0036228b 1096, 1150, 7421 Chitin binding protein, peritrophin-A Polysaccharide,
aminoglycan, & chitinmetabolic
Extracellularregion
Pattern, chitin,carbohydrate,& polysaccharidebindings; structuralmolecule activity,structural constituentof peritrophicmembrane
1 FBgn0027600b 421, 2833 Chitin binding protein, peritrophin-A
1 & 2 FBgn0023479b 1104, 4904, 5136 Chitin binding protein, peritrophin-A,Peptidase S1 and S6, chymotrypsin/Hap;Peptidase S1/S6 & chymotrypsin/Hapactive sites
Polysaccharide,aminoglycan, & chitinmetabolic; Proteolysis
Extracellularregion
Pattern, chitin,carbohydrate,& polysaccharidebindings;Endopeptidase,peptidase, serine-typeendopeptidase,scavenger receptor,serine-type peptidaseactivity, & serinehydrolase activities
2 FBgn0034661 138, 821, 2603, 2631, 2997,2998, 3972, 7106, 7216, 7828
Peptidase S1 and S6,chymotrypsin/Hap; PeptidaseS1/S6 & chymotrypsin/Hapactive sites
Proteolysis None Given Endopeptidase,serine-typeendopeptidase,peptidase, serine-typepeptidase, & serinehydrolase activities
2 FBgn0035669 221, 958, 1121, 1476, 1777,2012, 2013, 2208, 2209, 2292,2293, 2294, 2452, 2515, 2706,2918, 3172
2 FBgn0051954 1462, 1463, 28222 FBgn0038484 4154, 4761, 52152 FBgn0035665b 7246
2 FBgn0028948 5258 Peptidase M12A, astacin,metallopeptidase
Proteolysis None Given Endopeptidase,peptidase,metalloendopeptidase,& metallopeptidaseactivities; Zinc ion, ion,cation, metal ion,& transition metal ionbindings
a Exceptions are genes 6168 and 221 which are 0.78 and 1.89 fold greater, respectively.b FBgn0051973 has a Flybase Gene Description of Chitin deacetylase-like 5; FBgn0036228 and FBgn0027600 have Flybase Gene Descriptions of Obstructor-G and
Obstructor-B, respectively; FBgn0023479 has a Flybase Gene Description of Tequila; FBgn0035665 has a Flybase Gene Description of Jonah 65Aiii; All others have no FlybaseGene Description.
H.W. Fescemyer et al. / Insect Biochemistry and Molecular Biology 43 (2013) 280e291288
components of its PM and have multiple nonglycosylated cysteine-rich domains that likely bind and cross-link chitin fibrils in formingthe PM (Wang and Granados, 1997b; Shen and Jacobs-Lorena, 1998;Tellam et al., 1999;Wang and Granados, 2001; Hegedus et al., 2009;Toprak et al., 2010a,b). The IIMs in the PM are strongly associatedwith chitin in the PM. Their release from the PM of T. ni and otherinsects requires a combination of detergent plus a reducing agent(e.g., 2% SDS and 5 mM DTT), conditions not likely to occur wheneating plant foliage (Summers and Felton, 1996; Barbehenn andStannard, 2004). Our proteomic experiments suggest that thegreatly diminished presence of IIM in frass of caterpillars fed on themidwhorls of Mp708 is due to the ability of Mir1-CP to cleave IIMreleasing it into the gut lumen where it is further degraded byendogenous proteases as previously observed in T. ni (Wang andGranados, 1997a,b). The resultant reduction in IIM cross-linkingwith chitin in the PM likely leads to the cracks, fissures andincreased porosity previously observed in the PM of fall armywormlarvae fed on Mp708 maize whorls or plant tissues ectopicallyexpressing Mir1-CP (Pechan et al., 2000,2002). Reduction in chitin
cross-linked by IIM does not lead to complete degradation of thePM structure and function because other peritrophins assist in thiscross-linking.
A caterpillar’s compensatory response, particularly its ability tosustain the PM, also could prevent complete PM degradation in thepresence of Mir1-CP. Our targeted and global transcriptome ana-lyses detected differential expression of MG transcripts from genesencoding proteins that function in PM production. These proteinsinclude chitin synthase and chitin deacytlases known to beinvolved in the production of PM chitin fibrils (Bolognesi et al.,2005; Toprak et al., 2008; Qu et al., 2011). They also include IIMsand other peritrophins with chitin-binding domains that crosslinkchitin fibrils in the PM (Toprak et al., 2010a,b). We found MGs fromfall armyworm larvae fed on Mp708 to contain a significant, 2-foldor greater abundance of transcripts for these proteins comparedwith MGs from larvae fed on diets lacking Mir1-CP (i.e., artificialdiet and midwhorls from Tx601 or B73). Enrichment analysisindicated that groups of chitin-binding protein genes were selec-tively upregulated in Mp708 MGs compared with nearly all other

H.W. Fescemyer et al. / Insect Biochemistry and Molecular Biology 43 (2013) 280e291 289
MG genes in our reference. Compartmentalization of digestiveevents by preserving the counter-current MG gradients betweenendoperitrophic and ectoperitrophic spaces is an important func-tion of the PM (Bolognesi et al., 2001,2008). This compartmentali-zation is probably disrupted by the increased porosity of the PM asa result of Mir1-CP’s action on IIM. Upregulating expression in MGcells of genes encoding proteins involved in PM production sug-gests that insects fed on maize containing Mir1-CP compensate byproducing new PM in an effort to cover the disrupted PM structure.
Like most lepidopterans, the fall armyworm has a Type 1 PMwhose components are secreted around the food bolus by thecollumner cells in the anterior two thirds of the MG epithelium(Harper and Granados, 1999; Bolognesi et al., 2001). These cells alsosecrete digestive enzymes compartmentalized between endoperi-trophic and ectoperitrophic spaces (Ortego et al., 1996) with some(e.g., amylase, carboxypeptidase, aminopeptidase, lipase, chymo-trypsin and trypsin) also immobilized on the PM (Ferreira et al.,1994; Bolognesi et al., 2001; Jordo et al., 1999; Toprak et al., 2008,2010b). We foundMGs from fall armyworm larvae fed on Mp708 tocontain a significant, 2-fold or greater abundance of transcripts forseveral digestive enzymes compared with MGs from larvae fed ondiets lacking Mir1-CP. These enzymes include alpha-amylase,a metalloendopeptidase (i.e., astacin), and those in the amino-peptidase, serine-type endopeptidase (i.e., chymotrypsin, typsin,etc.) and lipase families. Like genes encoding chitin-binding pro-teins, enrichment clustering of genes encoding digestive enzymesindicates that they are being selectively upregulated comparedwith nearly all other MG genes in our reference. Differentialexpressionwas not detected for genes encoding proteins associatedwith microapocrine secretion (e.g., gelsolin, annexin) (Ferreiraet al., 2007; Bolognesi et al., 2001), perhaps because they werenot present in the EST reference library against which sequencereads were mapped. Fall armyworm larvae that were fed on dietcontaining chitin-binding calcafluor lack a PM and, similar to larvaefed on diets containing Mir1-CP, have impaired growth due toincreased metabolic cost to convert food into body mass (Bolognesiet al., 2008). They also lack the counter-current MG gradients thatenable enzyme recycling (Bolognesi et al., 2001,2008) probablyresulting in increased digestive enzyme excretion. A similar situa-tion appears to occur in larvae whose PM is disrupted by Mir1-CP.Our findings suggest that impaired growth of these larvae probablyresults from metabolic costs associated with upregulation of pro-teins involved in PM production and digestion in addition to theattempt to maintain MG gradient homeostasis.
Models supporting the various functions of the PM that enhancedigestive efficiency suggest that these functions could be extendedto any insect feeding on any diet (Bolognesi et al., 2008). In theEuropean corn borer, our candidate gene analysis found abundanceof transcripts for an IIM, a chitin deacytlase, and the digestive en-zymes in the serine endopeptidase (e.g., chymotrypsin, tyrpsin) andcarboxypeptidase families to be significantly greater in MGs fromlarvae fed on Mp708 compared with MGs from larvae fed on dietslacking Mir1-CP. In previous research, Mp708 showed resistance toEuropean corn borer (Davis et al., 1989) and other insect pests ofmaize (Davis et al., 1988; Gill et al., 2011). These results and thelikelihood of evolutionarily conserved PM functions (Bolognesiet al., 2008) suggest that upregulation of PM production anddigestive enzymes is a universal component of a caterpillar’scompensatory response to the presence of Mir1-CP or other cys-teine proteases (Li et al., 2009) in its diet. Transcriptional regulationof MG genes may be a common means by which insects adapt toplant defenses that affect MG physiology.
Expressing protease inhibitors is another means by which in-sects’ counter/compensate for the presence of plant defensiveproteases. Lepidopterans that feed on laticiferous plants protect
their PM from degradation by expressing cysteine protease-inhibitors (Li et al., 2009). These inhibitors were not detected inthe present study possibly because theywere not present in the ESTreference library against which sequence reads were mapped.Although fall armyworm has a wide host range, it is rarely found onlaticiferous plants (Luginbill, 1928) and may not express cysteineprotease-inhibitors. However, the abundance of transcripts asso-ciated with genes annotating as serine protease inhibitors or ser-pins was significantly greater in MGs from larvae fed on Mp708compared with those from larvae fed on Tx601. Insect serpins areinvolved in immune responses in plasma and in tissues (Tong andKanost, 2005; An and Kanost, 2010). Preliminary scanning elec-tron micrographs indicated possible damage to the MG epitheliumof fall armyworm fed on Mp708. Perhaps it is possible that thepredicted serpin has an immunological role in protecting the dis-rupted MG epithelium from microbial infection. Alternatively, theserpins may protect the MG cell surface from luminal serine pro-teinases. A proteinaceous mucus layer absent in insects confers thisprotection to mammalian gut cells. Further evidence for disruptionof the MG epithelium is that transcript abundance of a geneencoding a cadherin (Contig 3185; Fig. 2, Table 1, and SupplementalTable 7) is significantly higher in larvae fed on Mp708 comparedwith those from larvae fed on Tx601. Cadherins play importantroles in cell adhesion by ensuring that cells within tissues arebound together. Moreover, Mir1-CP was found to synergize activityof a Bacillus thuringiensis toxin (Mohan et al., 2008) whose mode ofaction includes direct attack of the MG by forming ion poresthrough binding to cadherins (Gómez et al., 2002; Vachon et al.,2012).
Most of the differentially expressed genes (N ¼ 133 for Q-value � 0.05) have homologs present in the PM or MG tissue ofother lepidopterans including H. armigera (Campbell et al., 2008;Pauchet et al., 2008,2010), Mamestra configurata Walker (Noctui-dae) (Toprak et al., 2008,2010b), Epiphyas postvittana (Walker)(Tortricidae) (Simpson et al., 2007), and B. mori (Kajiwara et al.,2005). This homology along with our finding that transcriptabundance of similar MG genes also increased in European cornborer fed on Mp708 suggests that transcriptional responses of fallarmyworm to the IIM-degrading Mir1-CP toxin are common acrosslepidopteran pests in general.
Degradation of IIM in the PM was shown to be an importanttarget substrate for Mir1-CP expressed in Mp708. Other PM pro-teins may also be degraded by the presence of Mir1-CP. Althoughyet to be identified, these additional protein targets of Mir1-CP arealso probably peritrophin structural components because theysurvived the method (boiling in 2% SDS) we used to purify the PM.It appears thatMir1-CPmay specifically disrupt the PM by targetingproteins that crosslink chitin fibrils. Both targeted and globaltranscriptome analysis revealed that disruption of the PM by foliagecontaining Mir1-CP is associated primarily with upregulation ofgenes in the MG that encode proteins involved in PM productionand food digestion. Ramping up production of these proteinsprobably enables some larvae to sustain nutrient acquisition andgrowth when feeding on resistant Mp708 maize. Compensatorytranscriptional regulation of the gene products most affected byplant defenses is probably a common mechanism by which insectstolerate and perhaps evolve host plant specificity.
Acknowledgments
The authors thank Paul W. Williams and SusanWolf at the USDAARS in Mississippi State, MS for supplying fall armyworm eggs andinbred maize seed; Ping Wang for the IIM antibody; Department ofEntomology at Penn State, Gary Felton and Michelle Peiffer forinvaluable help and facilities for rearing and dissecting insects;

H.W. Fescemyer et al. / Insect Biochemistry and Molecular Biology 43 (2013) 280e291290
Courtney Brenner, Daniel Matasic andMonicaWatson for assistancewith RNA isolation; and Craig Praul and Candace Price for technicalwork to prepare transcriptome libraries and SOLiD sequencing atthe Penn State Genomics Core Facility. GVS thanks FundaciónAlfonsoMartín Escudero for the fellowship as a Postdoctoral Scholarat Penn State. This research was supported by The Huck Institutes ofthe Life Sciences at Penn State, US NSF IOS-0641219 to DSL, andUSDA AFRI (2010-65106-20656) to DSL, HWF and JHM. Additionalsupport for HWF and JHM is from US NSF IOS-0950416.
Appendix A. Supplementary material
Supplementary material associated with this article can befound, in the online version, at http://dx.doi.org/10.1016/j.ibmb.2012.12.008.
References
An, C., Kanost, M.R., 2010. Manduca sexta serpin-5 regulates prophenoloxidaseactivation and the toll signaling pathway by inhibiting hemolymph proteinaseHP6. Insect Biochem. Mol. Biol. 40, 683e689.
Barbehenn, R.V., Stannard, J., 2004. Antioxidant defense of the midgut epitheliumby the peritrophic envelope in caterpillars. J. Insect Physiol. 50, 783e790.
Bolognesi, R., Ribeiro, A.F., Terra, W.R., Ferreira, C., 2001. The peritrophic membraneof Spodoptera frugiperda: secretion of peritrophins and role in immobilizationand recycling digestive enzymes. Arch. Insect Biochem. Physiol. 47, 62e75.
Bolognesi, R., Arakane, Y., Muthukrishnan, S., Kramer, K.J., Terra, W.R., Ferreira, C.,2005. Sequences of cDNAs and expression of genes encoding chitin synthaseand chitinase in the midgut of Spodoptera frugiperda. Insect Biochem. Mol. Biol.35, 1249e1259.
Bolognesi, R., Terra, W.R., Ferreira, C., 2008. Peritrophic membrane role inenhancing digestive efficiency. Theoretical and experimental models. J. InsectPhysiol. 54, 1413e1422.
Brooks, T.D., Willcox, M.C., Williams, W.P., Buckley, P.M., 2005. Quantitative trait lociconferring resistance to fall armyworm and southwestern corn borer leaffeeding damage. Crop Sci. 45, 2430e2434.
Brooks, T.D., Bushman, B.S., Williams, W.P., McMullen, M.D., Buckley, P.M., 2007.Genetic basis of resistance to fall armyworm (Lepidoptera: Noctuidae) andsouthwestern corn borer (Lepidoptera: Crambidae) leaf-feeding damage inmaize. J. Econ. Entomol. 100, 1470e1475.
Buntin, G.D., Lee, R.D., Wilson, D.M., McPherson, R.M., 2000. Evaluation of Yieldgardtransgenic resistance for control of fall armyworm and corn earworm (Lep-idoptera: Noctuidae) on corn. Fla. Entomol. 84, 37e42.
Campbell, P.M., Cao, A.T., Hines, E.R., East, P.D., Gordon, K.H.J., 2008. Proteomicanalysis of the peritrophic matrix from the gut of the caterpillar, Helicoverpaarmigera. Insect Biochem. Mol. Biol. 38, 950e958.
Carbon, S., Ireland, A., Mungall, C.J., Shu, S., Marshall, B., Lewis, S., the AmiGO Hub,the Web Presence Working Group, 2009. AmiGO: online access to ontology andannotation data. Bioinformatics 25, 288e289.
Chang, Y.M., Luthe, D.S., Davis, F.M., Williams, W.P., 2000. Influence of whorl regionfrom resistant and susceptible corn genotypes on fall armyworm (Lepidoptera:Noctuidae) growth and development. J. Econ. Entomol. 93, 477e483.
Davis, F.M., Williams, W.P., Mihm, J.A., Barry, B.E., Overman, L.J., Wiseman, B.R.,Riley, T.J., 1988. Resistance to multiple lepidopterous species in tropical derivedcorn germplasm. Miss. Agri. For. Exp. Stn. Tech. Bull. 157, 1e6.
Davis, F.M., Ng, S.S., Williams, W.P., 1989. Mechanisms of resistance in corn to leaffeeding by southwestern corn borer and European corn borer (Lepidoptera:Pyralidae). J. Econ. Entomol. 82, 919e922.
Ferreira, C., Capella, A.N., Sitnik, R., Terra,W.R.,1994.Digestive enzymes inmidgut cells,endo- and ectoperitrophic contents, and peritrophic membranes of Spodopterafrugiperda (Lepidoptera) larvae. Arch. Insect Biochem. Physiol. 26, 299e313.
Ferreira, A.H.P., Cristofolettia, P.T., Lorenzinic, D.M., Guerra, L.O., Paiva, P.B.,Briones, M.R.S., Terra, W.R., Ferreira, C., 2007. Identification of midgut micro-villar proteins from Tenebrio molitor and Spodoptera frugiperda by cDNA libraryscreenings with antibodies. J. Insect Physiol. 53, 1112e1124.
Gill, T.A., Sandoya, G., Williams, P., Luthe, D.S., 2011. Belowground resistance towestern corn rootworm in lepidopteran-resistant maize genotypes. J. Econ.Entomol. 104, 299e307.
Gómez, I., Sánchez, J., Miranda, R., Bravo, A., Soberón, M., 2002. Cadherin-like re-ceptor binding facilitates proteolytic cleavage of helix a-1 in domain I andoligomer pre-pore formation of Bacillus thuringiensis Cry1Ab toxin. FEBS Lett.513, 242e246.
Guo, H.F., Fang, J.C., Wang, J.P., Zhong, W.F., Liu, B.S., 2007. Interaction of Xestia c-nigrum granulovirus with peritrophic matrix and Spodoptera litura nucleopo-lyhedrovirus in Spodoptera litura. J. Econ. Entomol. 100, 20e25.
Harper, M.S., Granados, R.R., 1999. Peritrophic membrane structure and formationof larval Trichoplusia ni with an investigation on the secretion patterns of a PMmucin. Tissue Cell 31, 202e211.
Hakim, R.S., Baldwin, K., Smagghe, G., 2010. Regulation of midgut growth, devel-opment, and metamorphosis. Annu. Rev. Entomol. 55, 593e608.
Hegedus, D., Erlandson, M., Gillott, C., Toprak, U., 2009. New insights into peri-trophic matrix synthesis, architecture, and function. Annu. Rev. Entomol. 54,285e302.
Huang, D.W., Sherman, B.T., Lempicki, R.A., 2009a. Systematic and integrativeanalysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 4,44e57.
Huang, D.W., Sherman, B.T., Lempicki, R.A., 2009b. Bioinformatics enrichment tools:paths toward the comprehensive functional analysis of large gene lists. NucleicAcids Res. 37, 1e13.
Jakubowska, A.K., Caccia, S., Gordon, K.H., FerrÈ, J., Herrero, S., 2010. Down-regulation of a chitin deacetylase-like protein in response to baculovirusinfection and its application for improving baculovirus infectivity. J. Virol. 84,2547e2555.
Jordo, B.P., Capella, A.N., Terra, W.R., Ribeiro, A.F., Ferreira, C., 1999. Nature of theanchors of membrane-bound aminopeptidase, amylase, and trypsin andsecretory mechanisms in Spodoptera frugiperda (Lepidoptera) midgut cells.J. Insect Physiol. 45, 29e37.
Kajiwara, H., Ito, Y., Imamaki, A., Nakamura, M., Mita, K., Ishizaka, M., 2005. Proteinprofile of silkwormmidgut of fifth-instar day-3 larvae. J. Electrophor. 49, 61e69.
Laemmli, U.K., 1970. Cleavage of Structural Proteins during the Assembly of theHead of Bacteriophage T4. Nature 227, 680e685.
Landais, I., Ogliastro, M., Mita, K., Nohata, J., Lopez-Ferber, M., Duonor-Cérutti, M.,Shimada, T., Fournier, P., Devauchelle, G., 2003. Annotation pattern of ESTs fromSpodoptera frugiperda Sf9 cells and analysis of the ribosomal protein genesreveal insect-specific features and unexpectedly low codon usage bias. Bioin-formatics 19, 2343e2350.
Levy, H.C., Garcia-Maruniak, A., Maruniak, J.E., 2002. Strain identification of Spo-doptera frugiperda (Lepidoptera: Noctuidae) insects and cell line: PCR-RFLP ofcytochrome oxidase subunit I gene. Fla. Entomol. 85, 186e190.
Li, C., Song, X., Li, G., Wang, P., 2009. Midgut cysteine protease-inhibiting activity inTrichoplusia ni protects the peritrophic membrane from degradation by plantcysteine proteases. Insect Biochem. Mol. Biol. 39, 726e734.
Livak, K.J., Schmittgen, T.D., 2001. Analysis of relative gene expression data usingreal-time quantitative PCR and the 2�DDCT method. Methods 25, 402e408.
Lopez, L., Camas, A., Shivaji, R., Ankala, A., Williams, P., Luthe, D., 2007. Mir1-CP,a novel defense cysteine protease accumulates in maize vascular tissues inresponse to herbivory. Planta 226, 517e527.
Luginbill, P., 1928. The Fall Armyworm. USDA Technical Bulletin 34, p. 91.Luttrell, R.G., Wan, L., Knighten, K., 1999. Variation in susceptibility of noctuid
(Lepidoptera) larvae attacking cotton and soybean to purified endotoxin pro-teins and commercial formulations of Bacillus thuringiensis. J. Econ. Entomol. 92,21e32.
Mohan, S., Ma, P.W.K., Pechan, T., Bassford, E.R., Williams, W.P., Luthe, D.S., 2006.Degradation of the S. frugiperda peritrophic matrix by an inducible maize cys-teine protease. J. Insect Physiol. 52, 21e28.
Mohan, S., Ma, P.W.K., Williams, W.P., Luthe, D.S., 2008. A naturally occurring plantcysteine protease possesses remarkable toxicity against insect pests and syn-ergizes Bacillus thuringiensis toxin. PLOS ONE 3, e1786.
McQuilton, P., Pierre St., S.E., Thurmond, J., the FlyBase Consortium, 2012. FlyBase101 e the basics of navigating FlyBase. Nucleic Acids Res. 40 (Database issue),D706eD714.
Nagoshi, R.N., Meagher, R.L., Adamczyk, J.J., Braman, S.K., Brandenburg, R.L.,Nuessly, G., 2006. New restriction fragment length polymorphisms in thecytochromeoxidase I gene facilitate host strain identification of fall armyworm(Lepidoptera: Noctuidae) populations in the southeastern United States. J. Econ.Entomol. 99, 671e677.
Negre, N., Hotelier, T., Volkoff, A.N., Gimenez, S., Cousserans, F., Mita, K., Sabau, X.,Rocher, J., Lopez-Ferber, M., d’Alencon, E., Audant, P., Sabourault, C.,Bidegainberry, V., Hilliou, F., Fournier, P., 2006. Spodobase: an EST database forthe lepidopteran crop pest Spodoptera. BMC Bioinform. 7, 1471e2210.
Okuno, S., Takatsuka, J., Nakai, M., Ototake, S., Masui, A., Kunimi, Y., 2003. Viral-enhancing activity of various stilbene-derived brighteners for a Spodoptera lit-ura (Lepidoptera: Noctuidae) nucleopolyhedrovirus. Biol. Control 26, 146e152.
Ortego, F., Novillo, C., CastaÒera, P., 1996. Characterization and distribution ofdigestive proteases of the stalk corn borer, Sesamia nonagrioides Lef. (Lep-idoptera: Noctuidae). Arch. Insect Biochem. Physiol. 32, 163e180.
Pauchet, Y., Muck, A., Svatos, A., Heckel, D.G., Preiss, S., 2008. Mapping the larvalmidgut lumen proteome of Helicoverpa armigera, a generalist herbivorous in-sect. J. Proteome Res. 7, 1629e1639.
Pauchet, Y., Wilkinson, P., Vogel, H., Nelson, D.R., Reynolds, S.E., Heckel, D.G.,Ffrench-Constant, R.H., 2010. Pyrosequencing the Manduca sexta larval midguttranscriptome: messages for digestion, detoxification and defence. Insect Mol.Biol. 19, 61e75.
Pechan, T., Ye, L., Chang, Y., Mitra, A., Lin, L., Davis, F.M., Williams, W.P., Luthe, D.S.,2000. A unique 33-kD cysteine proteinase accumulates in response to larvalfeeding in maize genotypes resistant to fall armyworm and other Lepidoptera.Plant Cell 12, 1031e1040.
Pechan, T., Cohen, A., Williams, W.P., Luthe, D.S., 2002. Insect feeding mobilizesa unique plant defense protease that disrupts the peritrophic matrix of cater-pillars. Proc. Natl. Acad. Sci. USA 99, 13319e13323.
Pechanova, O., Pechan, T., Ozkan, S., McCarthy, F.M., Williams, W.P., Luthe, D.S., 2010.Proteome profile of the developing maize (Zea mays L.) rachis. Proteomics 10,3051e3055.

H.W. Fescemyer et al. / Insect Biochemistry and Molecular Biology 43 (2013) 280e291 291
Qu, M., Liu, T., Yang, J., Yang, Q., 2011. The gene, expression pattern and subcellularlocalization of chitin synthase B from the insect Ostrinia furnacalis. Biochem.Biophys. Res. Commun. 404, 302e307.
Ritchie, S.W., Hanway, J.J., Benson, G.O., 1992. How a Corn Plant Develops. Iowa StateUniversity of Science and Technology, Ames, IA, U.S.A. Cooperative ExtensionService Special Report 48. http://www.soilcropandmore.info/crops/Corn/How-Corn-Grows/index.htm.
Robinson, M.D., Oshlack, A., 2010. A scaling normalization method for differentialexpression analysis of RNA-seq data. Genome Biol. 11, R25.
Robinson, M.D., McCarthy, D.J., Smyth, G.K., 2010. EdgeR, a bioconductor package fordifferential expression analysis of digital gene expression data. Bioinformatics26, 139e140.
Scott, G.E., Davis, F.M., Williams, W.P., 1982. Registration of Mp701 and Mp702germplasm lines of maize. Crop Sci. 22, 1270.
Shen, Z., Jacobs-Lorena, M., 1998. A type I peritrophic matrix protein from themalaria vector Anopheles gambiae binds to chitin. Cloning, expression, andcharacterization. J. Biol. Chem. 273, 17665e17670.
Shivaji, R., Camas, A., Ankala, A., Engelberth, J., Tumlinson, J.H., Williams, W.P.,Wilkinson, J.R., Luthe, D.S., 2010. Plants on constant alert: elevated levels ofjasmonic acid and jasmonate-induced transcripts in caterpillar-resistant maize.J. Chem. Ecol. 36, 179e191.
Simpson, R.M., Newcomb, R.D., Gatehouse, H.S., Crowhurst, R.N., ChagnÈ, D.,Gatehouse, L.N., Markwick, N.P., Beuning, L.L., Murray, C., Marshall, S.D.,Yauk, Y.-K., Nain, B., Wang, Y.-Y., Gleave, A.P., Christeller, J.T., 2007. Expressedsequence tags from the midgut of Epiphyas postvittana (Walker) (Lepidoptera:Tortricidae). Insect Mol. Biol. 16, 675e690.
Smith, W.E.C., Shivaji, R., Williams, W.P., Luthe, D.S., Sandoya, G.V., Smith, C.L.,Sparks, D.L., Brown, A.E., 2012. A maize line resistant to herbivory constitutivelyreleases (E)-b-caryophyllene. J. Econ. Entomol. 105, 120e128.
Storer, N.P., Babcock, J.M., Schlenz, M., Meade, T., Thompson, G.D., Bing, J.W.,Huckaba, R.M., 2010. Discovery and characterization of field resistance to Btmaize: Spodoptera frugiperda (Lepidoptera: Noctuidae) in Puerto Rico. J. Econ.Entomol. 103, 1031e1038.
Storey, J.D., Tibshirani, R., 2003. Statistical significance for genome wide studies.Proc. Natl. Acad. Sci. USA 100, 9440e9445.
Summers, C.B., Felton, G.W., 1996. Peritrophic envelope as a functional antioxidant.Arch. Insect Biochem. Physiol. 32, 131e142.
Tellam, R.L., Wijffels, G., Willadsen, P., 1999. Peritrophic matrix proteins. InsectBiochem. Mol. Biol. 29, 87e101.
The Gene Ontology Consortium, 2000. Gene ontology: tool for the unification ofbiology. Nat. Genet. 25, 25e29.
The UniProt Consortium, 2011. Ongoing and future developments at the UniversalProtein Resource. Nucleic Acids Res. 39, D214eD219.
Tong, Y., Kanost, M.R., 2005. Manduca sexta serpin-4 and serpin-5 inhibit theprophenol oxidase activation pathway: cDNA cloning, protein expression, andcharacterization. J. Biol. Chem. 280, 14923e14931.
Toprak, U., Baldwin, D., Erlandson, M., Gillott, C., Hou, X., Coutu, C., Hegedus, D.D.,2008. A chitin deacetylase and putative insect intestinal lipases are componentsof the Mamestra configurata (Lepidoptera: Noctuidae) peritrophic matrix. InsectMol. Biol. 17, 573e585.
Toprak, U., Erlandson, M., Hegedus, D., 2010a. Peritrophic matrix proteins. TrendsEntomol. 6, 23e51.
Toprak, U., Baldwin, D., Erlandson, M., Gillott, C., Hegedus, D.D., 2010b. Insect in-testinal mucins and serine proteases associated with the peritrophic matrixfrom feeding, starved and moulting Mamestra configurata larvae. Insect Mol.Biol. 19, 163e175.
Vachon, V., Laprade, R., Schwartz, J.-L., 2012. Current models of the mode of actionof Bacillus thuringiensis insecticidal crystal proteins: a critical review. J. Invertbr.Pathol. 111, 1e12.
Vera, J.C., Wheat, C.W., Fescemyer, H.W., Frilander, M.J., Crawford, D.L., Hanski, I.,Marden, J.H., 2008. Rapid transcriptome characterization for a nonmodel or-ganism using 454 pyrosequencing. Mol. Ecol. 17, 1636e1647.
Virla, E.G., Ilvarez, A.N., Loto, F., Pera, L.M., BaigorÌ, M., 2008. Fall armyworm strains(Lepidoptera: Noctuidae) in Argentina, their associate host plants and responseto different mortality factors in laboratory. Fla. Entomol. 91, 63e69.
Wang, P., Granados, R.R., 1997a. An intestinal mucin is the target substrate fora baculovirus enhancin. Proc. Natl. Acad. Sci. USA 94, 6977e6982.
Wang, P., Granados, R.R., 1997b. Molecular cloning and sequencing of a novelinvertebrate intestinal mucin cDNA. J. Biol. Chem. 272, 16663e16669.
Wang, P., Granados, R.R., 2001. Molecular structure of the peritrophic membrane(PM): Identification of potential PM target sites for insect control. Arch. InsectBiochem. Physiol. 47, 110e118.
Wang, P., Granados, R.R., 2003. Rapid and efficient isolation of highly specific an-tibodies from an antiserum against a pool of proteins. Biotech. Histochem. 78,201e205.
Wang, J., Xia, Q., He, X., Dai, M., Ruan, J., Chen, J., Yu, G., Yuan, H., Hu, Y., Li, R.,Feng, T., Ye, C., Lu, C., Wang, J., Li, S., Wong, G.K.-S., Yang, H., Wang, J., Xiang, Z.,Zhou, Z., Yu, J., 2005. SilkDB: a knowledgebase for silkworm biology and ge-nomics. Nucleic Acids Res. 33 (Database issue), D399eD402.
Wheat, C.W., 2010. Rapidly developing functional genomics in ecological modelsystems via 454 transcriptome sequencing. Genetica 138, 433e451.
Williams, P., Davis, F.M., 1982. Registration of Mp704 germplasm line of maize. CropSci. 22, 1269e1270.
Williams, P., Davis, F.M., 1984. Registration of Mp705, Mp706, and Mp707 germ-plasm lines of maize. Crop Sci. 24, 1217.
Williams, W.P., Davis, F.M., Windham, G.L., 1990. Registration of Mp708 germplasmline of maize. Crop Sci. 30, 757.
Williams, P., Davis, F.M., 2000. Registration of maize germplasm Mp713 and Mp714.Crop Sci. 40, 584.
Williams, P., Davis, F.M., 2002. Registration of maize germplasm line Mp716. CropSci. 42, 671e672.
Yu, S.J., 1992. Detection and biochemical characterization of insecticide resistance infall armyworm (Lepidoptera: Noctuidae). J. Econ. Entomol. 85, 675e682.
Yu, S.J., Nguyen, S.N., Abo-Elghar, G.E., 2003. Biochemical characterization ofinsecticide resistance in the fall armyworm, Spodoptera frugiperda (J.E. Smith).Pestic. Biochem. Physiol. 77, 1e11.
Zimmerman, D., Peters, W., 1987. Fine structure and permeability of peritrophicmembranes of Calliphora erythrocephala (Meigen) (Insecta: Diptera) afterinhibition of chitin and protein synthesis. Comp. Biochem. Physiol. 86B,353e360.
Related Documents











