Maintenance of CD34 Expression During Proliferation of CD34 + Cord Blood Cells on Glycosaminoglycan Surfaces SUNDARARAJAN V. MADIHALLY, a ALAN W. FLAKE, b HOWARD W.T. MATTHEW a a Department of Chemical Engineering and Materials Science, Wayne State University, Detroit, Michigan, USA; b Children’s Hospital of Philadelphia, Abramson Pediatric Research Center, Philadelphia, Pennsylvania, USA Key Words. Glycosaminoglycan · Heparin · Chondroitin sulfate · CD34 + cells · Chitosan · Proliferation ABSTRACT Recent studies have indicated that glycosaminogly- can (GAG) interactions with hematopoietic progenitors play a significant role in the regulation of hematopoiesis. However, the details of these interac- tions are not clear. In this study, we examined the role of soluble and immobilized GAGs in the proliferation of CD34 + cells. Chitosan, a cationic polysaccharide, was used to immobilize GAGs in ionic complex membranes. The GAGs studied were heparin, hyaluronate, and chondroitin sulfates A, B, and C. CD34-enriched umbilical cord blood cells were seeded onto tissue culture plates coated with the GAG-chitosan complex membranes. Cultures were maintained in medium supplemented with stem cell factor and interleukin 3 for up to six weeks, during which total and CD34 + cell num- bers were determined by flow cytometry. Total cell num- ber expansion ranged from 25-fold to 40-fold after six weeks. However, only heparin and chondroitin sulfate B (CSB) surfaces retained a significant CD34 + fraction. All other surfaces exhibited declines in CD34 expression, with negligible CD34 + percentages remaining after four weeks. In contrast, heparin and CSB surfaces exhibited CD34 + fractions as high as 90% after four weeks. GAG desorption studies indicated that the observed effects were partly mediated by desorbed GAGs in a concentration dependent manner. Subsequent studies showed that sustained high (160 mg/ml) heparin levels had toxic effects, while the same concentration of CSB exhibited more rapid early proliferation of CD34 + cells. In conclusion, this culture system has demonstrated the ability to produce simultaneous proliferation and CD34 + cell enrichment of a partially purified cord blood population by controlling the nature and levels of GAG moieties to which the cells are exposed. The results indicate that specific GAGs can significantly influence the growth and differentiation characteristics of cultured CD34 + cells. Stem Cells 1999;17:295-305 STEM CELLS 1999;17:295-305 Correspondence: Dr. Howard Matthew, Wayne State University, Chemical Engineering & Materials Science Dept., 5050 Anthony Wayne Drive, Detroit, Michigan 48202, USA. Accepted for publication July 24, 1999. ©AlphaMed Press 1066-5099/99/$5.00/0 INTRODUCTION The hematopoietic microenvironment plays a major role in controlling proliferation and lineage differentiation during hematopoiesis [1-5]. Many studies have demon- strated that the activities of hematopoietic progenitors are regulated by multiple receptor-ligand interactions with soluble, cell-surface-bound, and extracellular-matrix (ECM)-bound cytokines. However, there is a paucity of information regarding the detailed mechanisms of these interactions. In fact, the evidence shows that many impor- tant hematopoietic cytokines bind with varying affinities to a variety of ECM molecules [5-9]. The specific effects of these binding interactions vary widely depending on the cytokine and ECM molecules involved. Such effects may range from activity reductions to activity enhancement. In addition, the nature of the effect (enhancement or inhibition) may also depend on the relative concentrations of participating molecules. Proteoglycans and the ECM polysaccharides termed glycosaminoglycans (GAGs) are major components of the hematopoietic matrix [10, 11]. A number of studies have indicated that glycosaminoglycans bind and present cytokines to hematopoietic cells in a more highly active form [6, 7, 9, 12-14]. Most notably, the results show that

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Maintenance of CD34 Expression During Proliferation of CD34+ Cord Blood Cells on Glycosaminoglycan Surfaces
SUNDARARAJAN V. MADIHALLY ,a ALAN W. FLAKE ,b HOWARD W.T. M ATTHEW a
aDepartment of Chemical Engineering and Materials Science, Wayne State University, Detroit, Michigan, USA;bChildren’s Hospital of Philadelphia, Abramson Pediatric Research Center, Philadelphia, Pennsylvania, USA
Key Words. Glycosaminoglycan · Heparin · Chondroitin sulfate · CD34+ cells · Chitosan · Proliferation
ABSTRACT
Recent studies have indicated that glycosaminogly-can (GAG) interactions with hematopoietic progenitorsplay a significant role in the regulation ofhematopoiesis. However, the details of these interac-tions are not clear. In this study, we examined the roleof soluble and immobilized GAGs in the proliferation ofCD34+ cells. Chitosan, a cationic polysaccharide, wasused to immobilize GAGs in ionic complex membranes.The GAGs studied were heparin, hyaluronate, andchondroitin sulfates A, B, and C. CD34-enrichedumbilical cord blood cells were seeded onto tissueculture plates coated with the GAG-chitosan complexmembranes. Cultures were maintained in mediumsupplemented with stem cell factor and interleukin 3 forup to six weeks, during which total and CD34+ cell num-bers were determined by flow cytometry. Total cell num-ber expansion ranged from 25-fold to 40-fold after sixweeks. However, only heparin and chondroitin sulfate B(CSB) surfaces retained a significant CD34+ fraction. All
other surfaces exhibited declines in CD34 expression,with negligible CD34+ percentages remaining after fourweeks. In contrast, heparin and CSB surfaces exhibitedCD34+ fractions as high as 90% after four weeks.
GAG desorption studies indicated that the observedeffects were partly mediated by desorbed GAGs in aconcentration dependent manner. Subsequent studiesshowed that sustained high (160 mg/ml) heparin levelshad toxic effects, while the same concentration of CSBexhibited more rapid early proliferation of CD34+ cells.In conclusion, this culture system has demonstrated theability to produce simultaneous proliferation andCD34+ cell enrichment of a partially purified cord bloodpopulation by controlling the nature and levels of GAGmoieties to which the cells are exposed. The resultsindicate that specific GAGs can significantly influencethe growth and differentiation characteristics ofcultured CD34+ cells. Stem Cells 1999;17:295-305
STEM CELLS 1999;17:295-305
Correspondence: Dr. Howard Matthew, Wayne State University, Chemical Engineering & Materials Science Dept.,5050 Anthony Wayne Drive, Detroit, Michigan 48202, USA. Accepted for publication July 24, 1999. ©AlphaMed Press1066-5099/99/$5.00/0
INTRODUCTION
The hematopoietic microenvironment plays a majorrole in controlling proliferation and lineage differentiationduring hematopoiesis [1-5]. Many studies have demon-strated that the activities of hematopoietic progenitors areregulated by multiple receptor-ligand interactions withsoluble, cell-surface-bound, and extracellular-matrix(ECM)-bound cytokines. However, there is a paucity ofinformation regarding the detailed mechanisms of theseinteractions. In fact, the evidence shows that many impor-tant hematopoietic cytokines bind with varying affinities toa variety of ECM molecules [5-9]. The specific effects of
these binding interactions vary widely depending on thecytokine and ECM molecules involved. Such effects mayrange from activity reductions to activity enhancement. In addition, the nature of the effect (enhancement orinhibition) may also depend on the relative concentrationsof participating molecules.
Proteoglycans and the ECM polysaccharides termedglycosaminoglycans (GAGs) are major components of thehematopoietic matrix [10, 11]. A number of studies haveindicated that glycosaminoglycans bind and presentcytokines to hematopoietic cells in a more highly activeform [6, 7, 9, 12-14]. Most notably, the results show that

Madihally, Flake, Matthew 296
heparan sulfate is required for maintenance of long-termculture-initiating cells (LTC-ICs) [9]. The mechanism ofaction seems to be via selective enhancement of certainactivities of early-acting cytokines. Additionally, it hasbeen shown that heparin (a more highly sulfated version ofheparan sulfate) can directly activate some growth factorreceptors. However, the detailed mechanisms of theseeffects are poorly understood. In particular, the degree ofspecificity of the GAG-cytokine interactions and questionsof opposing effects on different progenitors and lineagesremain largely unresolved. In light of this uncertainty,culturing hematopoietic cells on ECM polysaccharidesurfaces should provide more insight into the roles of thesemolecules in hematopoiesis.
Within the ECM, GAGs are immobilized by combina-tions of covalent, ionic, and hydrogen bonding interactions.In this interfacial environment, the three-dimensionalnature of GAG-growth factor interactions may be some-what different from the corresponding interactions insolution. However, most in vitro culture systems eitherincorporate GAGs with other complicating factors (i.e., stroma-based systems), or supply GAGs in purelysoluble form (cytokine-based, stroma-free systems). In thisstudy, the effects of specific GAGs on the expansion ofCD34+ cord blood cells were studied in stroma-free culturesusing a model GAG surface. GAGs were immobilized ontoculture surfaces by forming insoluble ionic complexes withthe amino-polysaccharide chitosan [15-17]. Humanumbilical cord blood CD34+ cells were seeded onto thecomplex surfaces, and their proliferation and retention ofCD34 expression were followed.
MATERIALS AND METHODS
Chitosan (66% deacetylated), glacial acetic acid,hyaluronic acid (HA) from human umbilical cord, heparinfrom porcine intestinal mucosa, chondroitin sulfate A(CSA) from bovine articular cartilage, chondroitin sulfate B(CSB) from bovine intestinal mucosa, and chondroitin sul-fate C (CSC) from shark cartilage, were purchased fromSigma Chemical Co. (St. Louis, MO). Type I collagen wasextracted from rat tail tendon by the method of ElsdaleandBardas modified by Dunn[18]. Alcian blue 8GX and tolu-idine blue were obtained from Polysciences, Inc.(Warrington, PA). All culture medium components werepurchased from Life Technologies (Rockville, MD).
Preparation of Immobilized GAG Culture MembranesPolysaccharide complex membranes were deposited
onto tissue culture polystyrene dishes using the ionicinteraction between a GAG solution and a pre-depositedchitosan-acetic acid membrane. To prepare sterile chitosan
solution (2 wt% in 0.2 M acetic acid), 2 g of chitosan wasautoclaved in 100 ml of water and then dissolved by adding1.2 g of sterile filtered glacial acetic acid and stirring for 2 h.Sterile GAG solutions (1 wt%) were prepared by dissolvingthe GAG in saline (0.9% NaCl) buffered with 50 mMHEPES (pH 7.4) and then autoclaving. The GAGs evaluatedin this study were heparin, HA, CSA (chondroitin-4-sulfate),CSB (dermatan sulfate), and CSC (chondroitin-6-sulfate).
Cell cultures were conducted in 24-well culture plates,with all culture surfaces represented on each plate by threereplicate wells. Control culture surfaces were of threetypes: untreated culture wells, wells coated with type Icollagen, and chitosan-coated wells. To prepare thecollagen-coated wells, 0.25 ml of a 1 mg/ml solution of col-lagen in 1 mM HCl was evaporated to dryness in each welland then equilibrated with HEPES buffered saline. Purechitosan culture surfaces were prepared by covering thebottom of each well with 50 µl of sterile chitosan solution.The solution was then evaporated to dryness at room tem-perature in a biological laminar flow hood. This processproduced a coating of dry chitosan acetate. The coatedwells were then washed with 0.5 ml of 0.1 M NaOH toremove the acetate. This was followed by equilibration with1.5 ml of 50 mM HEPES buffered saline (pH 7.4). GAG-chitosan complex surfaces were prepared by firstcoating wells with 50 µl of sterile chitosan solution andagain evaporating to dryness. The coated wells werecovered with 0.5 ml of sterile GAG solution and allowed to equilibrate for ~6 h. This procedure resulted in thedeposition of a GAG-chitosan complex membrane on thedish. The formed complex membranes were then washed four times with 1.5 ml of HEPES buffered saline. In theRESULTS section, membranes formed by this procedure aredesignated “regular” membranes.
Preparation of GAG-Depleted Complex MembranesHeparin-chitosan and CSB-chitosan complex mem-
branes were prepared in 24-well plates as described above.The membranes were then washed six times with phosphatebuffered saline (PBS) (pH 7.4). For each wash, 0.5 ml ofPBS was added to each well and the plate was incubated at37°C for 4 h, after which the solution was aspirated. Cellswere seeded onto the membranes after the sixth wash.
Procurement and Culture of CD34+ CellsHuman umbilical cord blood was collected post-partum
in accordance with a protocol approved by the Wayne StateUniversity Institutional Review Board. CD34+ cells wereisolated and partially purified using an immunomagneticseparation kit (mini MACS, Miltenyi Biotec; Auburn, CA).In brief, anticoagulated cord blood (heparin, 1 mg/ml) was

297 Glycosaminoglycan Effects on CD34+ Cell Growth
mixed with an equal volume of Iscove’s modifiedDulbecco’s medium (IMDM) and layered onto Histopaquedensity medium (D1.077, Sigma Chemical Co.). After cen-trifugation at 1,500 rpm for 35 min, mononuclear cells wereretrieved from the interphase band and washed three timeswith tissue culture medium. Immunomagnetic separationwas then conducted following the manufacturer’s instruc-tions. An aliquot of the CD34-enriched cell suspension wastaken for analysis prior to cell seeding. From a typical run,~1 million mononuclear cells were recovered from 20 ml ofblood with CD34+ percentages ranging from 30% to 60%.
The isolated cells were seeded into 24-well tissueculture plates containing the control and GAG complexsurfaces. Seeding density was 15,000-30,000 cells per wellwith 0.5 ml of culture medium per well. The culturemedium consisted of IMDM supplemented with 10% fetal calf serum, 1% bovine serum albumin (BSA), 5 × 10-5 Mb-mercaptoethanol, 100 U/ml penicillin, 100 µg/mlstreptomycin, 2.5 µg/ml amphotericin B, 5 ng/ml stem cellfactor (SCF), and 10 ng/ml interleukin 3 (IL-3). Half-vol-ume medium changes were done at three-day intervalswithout cell removal. The total culture duration was sixweeks.
Culture with Soluble GAGs The effects of soluble heparin and CSB were evaluated in
noncoated tissue culture wells using GAG-supplementedmedium. The test GAGs were individually dissolved in cul-ture medium at concentrations of 7 µg/ml or 160 µg/ml. Cellswere seeded into noncoated wells using the GAG-supple-mented medium, and cultures were conducted as describedabove. For these studies, the control condition consisted ofcells in noncoated wells with the normal culture medium.
Analysis of Cell PhenotypeCulture plates were sacrificed after two, four, and six
weeks of culture for cell counting and phenotype analysis.Nonadherent and weakly adherent cells were collected fromindividual wells by pipette rinsing with PBS. Cells were pel-leted once and resuspended in 0.6 ml of PBS containing 1%bovine albumin. Staining was conducted at 4°C using a fluo-rescently labeled anti-CD34 antibody (HPCA-2-PE, BectonDickinson; San Jose, CA). After 30 min of incubation, cellswere washed and resuspended in PBS/BSA. Cells werecounted and analyzed for CD34 expression using a flowcytometer (Becton Dickinson).
GAG Quantification AssaysGAGs in solutions and in membrane washes were quan-
tified by an alcian blue-based colorimetric method [19]. A 30µl volume of the sample or a GAG standard solution was
transferred into a 96-well microassay plate and mixed with240 µl of alcian blue dye solution (1.4 mg/ml in 0.05 Msodium acetate). After 10 min of equilibration, the absorbanceof the solution at 480 nm was measured. The GAG concen-tration was determined from a standard curve prepared usingsolutions of the test GAG (0 to 30 µg/ml) in PBS.
The GAG content of complex membranes was deter-mined using a colorimetric method based on binding of tolu-idine blue [20, 21]. To assay bound GAG in complexmembranes on a 24-well plate, the overlying solution was firstaspirated and the membrane rinsed once with PBS. A 600 µlvolume of toluidine blue solution (0.075 mg/ml in water) wasthen applied for 10 min. The decrease in absorbance of thedye solution at 630 nm was measured and correlated withGAG mass from a standard curve. To prepare the standardcurve, 2 to 10 µg of the test GAG in 150 µl of water wasmixed with 150 µl of 0.15 mg/ml toluidine blue solution.Next, 300 µl of cyclohexane was added, and the mixture wasvortexed. The mixture was allowed to separate, and theorganic layer with associated toluidine blue-GAG complexwas removed by aspiration. The decrease in the absorbance ofthe aqueous layer at 630 nm was then measured.
Heparin Desorption Kinetics To evaluate the kinetics of heparin desorption from com-
plex membranes, 500 µl of PBS were applied to each of aseries of heparin-chitosan membranes in 24-well plates. Theplates were incubated at 37°C, and solutions were collectedat time intervals up to 96 h. The heparin content of solutionsand membranes was then measured as described above.
Statistical AnalysisCell expansion on GAG-chitosan and other test
surfaces was compared to expansion on the control plasticsurface by applying Student’s t-test to corresponding timepoints. Significance was tested at a probability level of 5%.
RESULTS
Culture Membrane Formation and Morphology In solution, chitosan is a positively charged polymer
and is capable of forming insoluble ionic complexes with negatively charged polymers such as the glyco-saminoglycans. Similarly, dehydrated chitosan salts can becomplexed in situ by rehydrating with an aqueous solutionof a negatively charged polymer. We have previously usedthis ionic interaction to microencapsulate primary hepato-cytes [15, 22]. Similar chitosan-GAG complexes have alsobeen applied as aids to wound healing and tissue regenera-tion [23-25]. Membranes produced by the GAG-chitosancomplex formation procedure appeared granular and textured

Madihally, Flake, Matthew 298
under the phase contrast light microscope. Numerous ridgesand convolutions could be clearly seen. In contrast,untreated chitosan surfaces were smooth and transparent.Complex membranes exhibited uniform staining with tolu-idine blue, indicating uniform GAG distribution. After sixweeks of culture, toluidine blue staining was still signifi-cant, indicating that measurable quantities of GAGremained throughout the culture period.
Cell Morphology and Adhesion While the significance of cell shape in these studies is
uncertain, it is generally believed that the most primitivehematopoietic progenitors are poorly adherent and thusmay be more likely to exhibit a spherical morphology. In
light of this, we briefly report our morphological observa-tions. In general, the vast majority of proliferating cellswere nonadherent. In addition to individual cells, some for-mation of both detached and adherent cell aggregates wasobserved on the CSC-chitosan surfaces. The number andsize of these aggregate colonies increased during the cultureperiod (Fig. 1). Cells of a variety of shapes and sizes werealso observed on the CSC-chitosan surfaces in the latterhalf of the culture period. In the case of noncoated poly-styrene surfaces, multiple cell shapes and sizes were alsoobserved, but few aggregates were seen up to week 6. Onthe heparin-chitosan surfaces, no colony formation oraggregation was seen, and all cells maintained their originalspherical shape. On the chitosan and HA-chitosan surfaces,
Figure 1. Cord blood CD34+ cells on GAG-chitosan membranes. Phase contrast micrographs. A: CSB, week 4. B: heparin, week 4. C: attachedcolonies on CSC, week 4. D: CSB-depleted membranes, week 6. Note the adherent cell population.
299 Glycosaminoglycan Effects on CD34+ Cell Growth
some aggregate formation was observed at the end of twoweeks, but aggregate frequency subsequently declined. Inthe case of CSB, a small number of aggregates wasobserved in the latter half of the culture.
With the exception of the aggregate colonies describedabove, cell attachment to the GAG-chitosan membraneswas negligible for most of the culture period. On the morehighly textured heparin-chitosan and CSB-chitosan mem-branes, many cells appeared to be wedged in membraneniches and valleys, but these could be easily detached withgentle washing. Growth of a fully spread, strongly adherentcell population was observed in the noncoated andcollagen-coated wells during weeks 3 to 6 of culture.
To examine the growth contributions of the adherentcells, two- and four-week-old plates, from which thenonadherent and weakly adherent cells had been removedby washing, were supplied with fresh medium for an addi-tional two weeks. No significant cell growth was observed,indicating that the critical proliferative cell population waseither nonadherent or only weakly adherent.
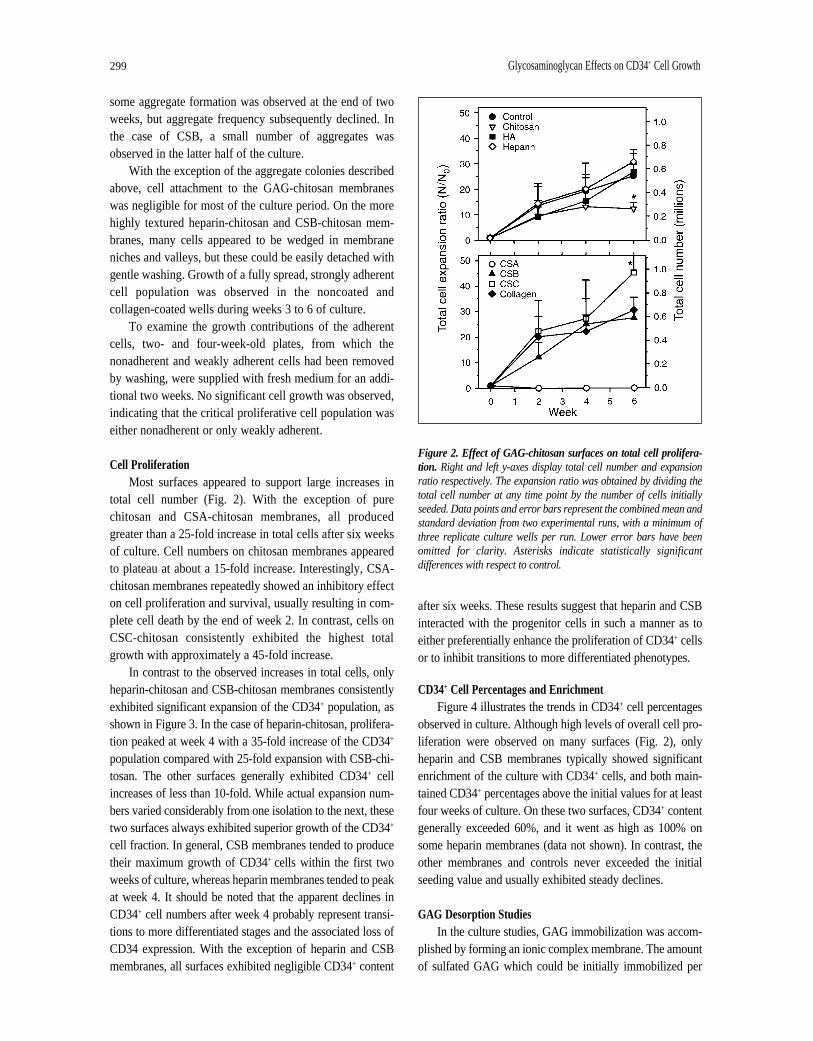
Cell Proliferation Most surfaces appeared to support large increases in
total cell number (Fig. 2). With the exception of purechitosan and CSA-chitosan membranes, all producedgreater than a 25-fold increase in total cells after six weeksof culture. Cell numbers on chitosan membranes appearedto plateau at about a 15-fold increase. Interestingly, CSA-chitosan membranes repeatedly showed an inhibitory effecton cell proliferation and survival, usually resulting in com-plete cell death by the end of week 2. In contrast, cells onCSC-chitosan consistently exhibited the highest totalgrowth with approximately a 45-fold increase.
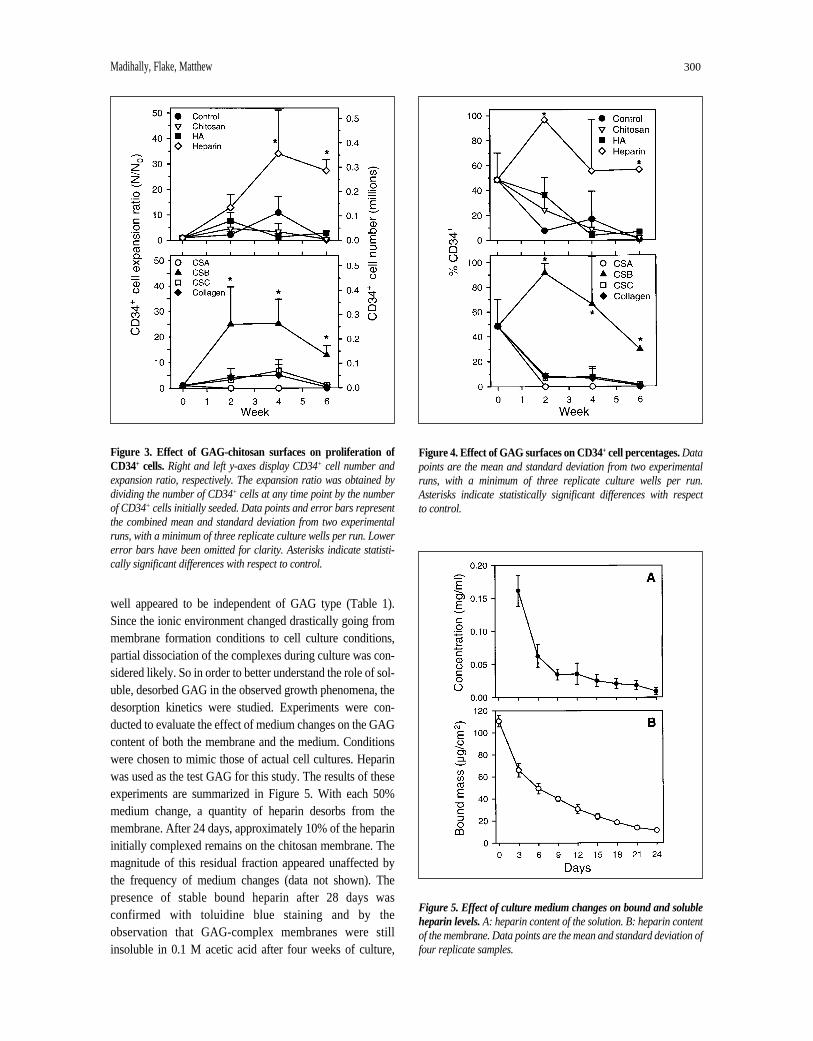
In contrast to the observed increases in total cells, onlyheparin-chitosan and CSB-chitosan membranes consistentlyexhibited significant expansion of the CD34+ population, asshown in Figure 3. In the case of heparin-chitosan, prolifera-tion peaked at week 4 with a 35-fold increase of the CD34+
population compared with 25-fold expansion with CSB-chi-tosan. The other surfaces generally exhibited CD34+ cellincreases of less than 10-fold. While actual expansion num-bers varied considerably from one isolation to the next, thesetwo surfaces always exhibited superior growth of the CD34+
cell fraction. In general, CSB membranes tended to producetheir maximum growth of CD34+ cells within the first twoweeks of culture, whereas heparin membranes tended to peakat week 4. It should be noted that the apparent declines inCD34+ cell numbers after week 4 probably represent transi-tions to more differentiated stages and the associated loss ofCD34 expression. With the exception of heparin and CSBmembranes, all surfaces exhibited negligible CD34+ content
after six weeks. These results suggest that heparin and CSBinteracted with the progenitor cells in such a manner as toeither preferentially enhance the proliferation of CD34+ cellsor to inhibit transitions to more differentiated phenotypes.
CD34+ Cell Percentages and Enrichment Figure 4 illustrates the trends in CD34+ cell percentages
observed in culture. Although high levels of overall cell pro-liferation were observed on many surfaces (Fig. 2), onlyheparin and CSB membranes typically showed significantenrichment of the culture with CD34+ cells, and both main-tained CD34+ percentages above the initial values for at leastfour weeks of culture. On these two surfaces, CD34+ contentgenerally exceeded 60%, and it went as high as 100% onsome heparin membranes (data not shown). In contrast, theother membranes and controls never exceeded the initialseeding value and usually exhibited steady declines.
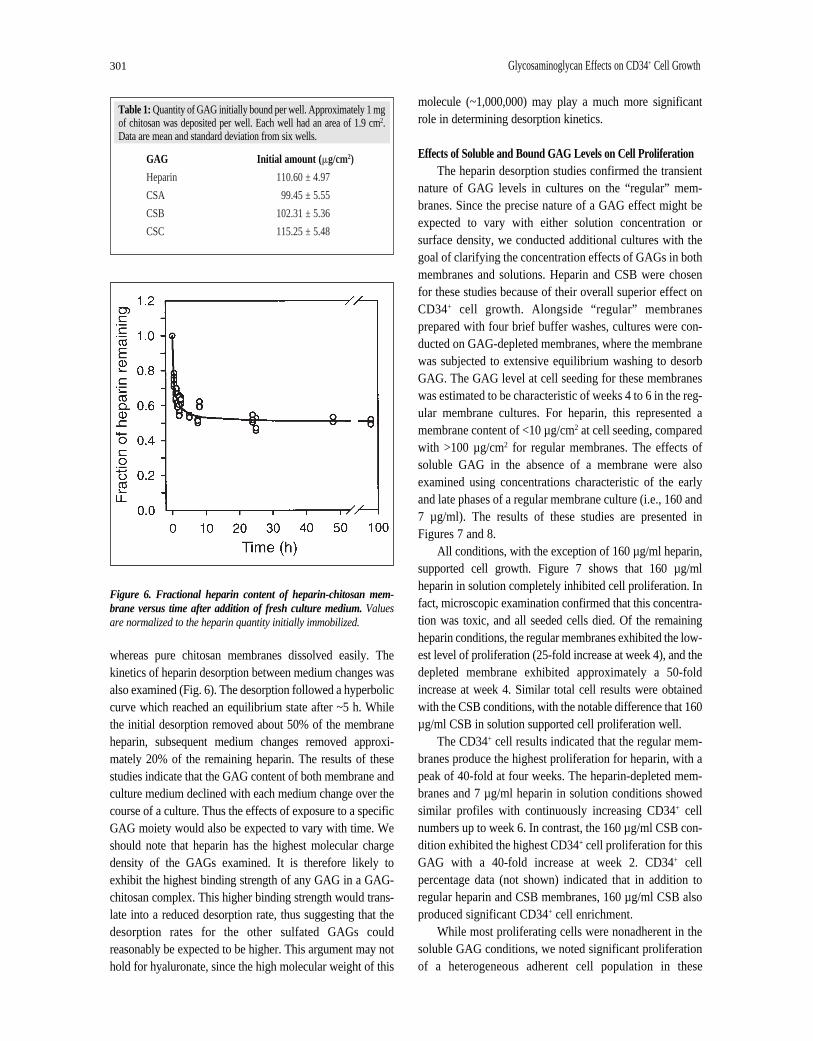
GAG Desorption StudiesIn the culture studies, GAG immobilization was accom-
plished by forming an ionic complex membrane. The amountof sulfated GAG which could be initially immobilized per
Figure 2. Effect of GAG-chitosan surfaces on total cell prolifera-tion. Right and left y-axes display total cell number and expansionratio respectively. The expansion ratio was obtained by dividing thetotal cell number at any time point by the number of cells initiallyseeded. Data points and error bars represent the combined mean andstandard deviation from two experimental runs, with a minimum ofthree replicate culture wells per run. Lower error bars have beenomitted for clarity. Asterisks indicate statistically significantdifferences with respect to control.
Madihally, Flake, Matthew 300
well appeared to be independent of GAG type (Table 1).Since the ionic environment changed drastically going frommembrane formation conditions to cell culture conditions,partial dissociation of the complexes during culture was con-sidered likely. So in order to better understand the role of sol-uble, desorbed GAG in the observed growth phenomena, thedesorption kinetics were studied. Experiments were con-ducted to evaluate the effect of medium changes on the GAGcontent of both the membrane and the medium. Conditionswere chosen to mimic those of actual cell cultures. Heparinwas used as the test GAG for this study. The results of theseexperiments are summarized in Figure 5. With each 50%medium change, a quantity of heparin desorbs from themembrane. After 24 days, approximately 10% of the heparininitially complexed remains on the chitosan membrane. Themagnitude of this residual fraction appeared unaffected bythe frequency of medium changes (data not shown). Thepresence of stable bound heparin after 28 days wasconfirmed with toluidine blue staining and by theobservation that GAG-complex membranes were stillinsoluble in 0.1 M acetic acid after four weeks of culture,
Figure 3. Effect of GAG-chitosan surfaces on proliferation ofCD34+ cells.Right and left y-axes display CD34+ cell number andexpansion ratio, respectively. The expansion ratio was obtained bydividing the number of CD34+ cells at any time point by the numberof CD34+ cells initially seeded. Data points and error bars representthe combined mean and standard deviation from two experimentalruns, with a minimum of three replicate culture wells per run. Lowererror bars have been omitted for clarity. Asterisks indicate statisti-cally significant differences with respect to control.
Figure 4. Effect of GAG surfaces on CD34+ cell percentages.Datapoints are the mean and standard deviation from two experimentalruns, with a minimum of three replicate culture wells per run.Asterisks indicate statistically significant differences with respect to control.
Figure 5. Effect of culture medium changes on bound and solubleheparin levels.A: heparin content of the solution. B: heparin contentof the membrane. Data points are the mean and standard deviation offour replicate samples.
301 Glycosaminoglycan Effects on CD34+ Cell Growth
whereas pure chitosan membranes dissolved easily. Thekinetics of heparin desorption between medium changes wasalso examined (Fig. 6). The desorption followed a hyperboliccurve which reached an equilibrium state after ~5 h. Whilethe initial desorption removed about 50% of the membraneheparin, subsequent medium changes removed approxi-mately 20% of the remaining heparin. The results of thesestudies indicate that the GAG content of both membrane andculture medium declined with each medium change over thecourse of a culture. Thus the effects of exposure to a specificGAG moiety would also be expected to vary with time. Weshould note that heparin has the highest molecular chargedensity of the GAGs examined. It is therefore likely toexhibit the highest binding strength of any GAG in a GAG-chitosan complex. This higher binding strength would trans-late into a reduced desorption rate, thus suggesting that thedesorption rates for the other sulfated GAGs couldreasonably be expected to be higher. This argument may nothold for hyaluronate, since the high molecular weight of this
molecule (~1,000,000) may play a much more significantrole in determining desorption kinetics.
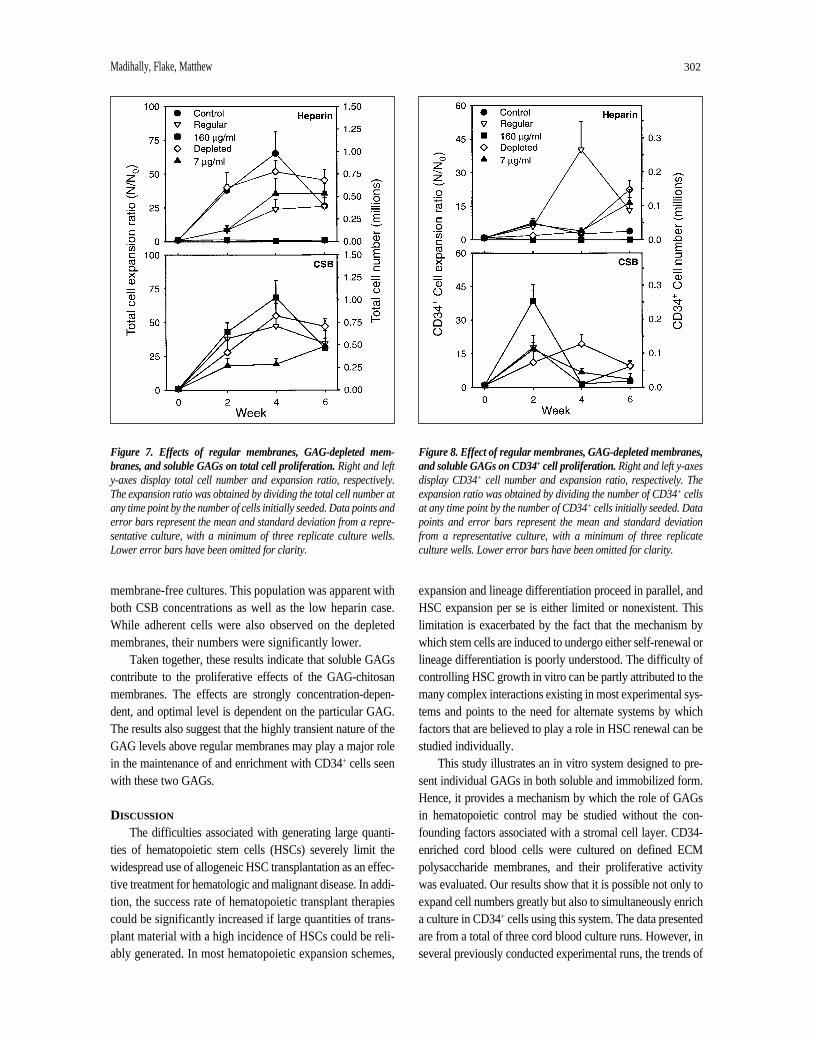
Effects of Soluble and Bound GAG Levels on Cell ProliferationThe heparin desorption studies confirmed the transient
nature of GAG levels in cultures on the “regular” mem-branes. Since the precise nature of a GAG effect might beexpected to vary with either solution concentration orsurface density, we conducted additional cultures with thegoal of clarifying the concentration effects of GAGs in bothmembranes and solutions. Heparin and CSB were chosenfor these studies because of their overall superior effect onCD34+ cell growth. Alongside “regular” membranesprepared with four brief buffer washes, cultures were con-ducted on GAG-depleted membranes, where the membranewas subjected to extensive equilibrium washing to desorbGAG. The GAG level at cell seeding for these membraneswas estimated to be characteristic of weeks 4 to 6 in the reg-ular membrane cultures. For heparin, this represented amembrane content of <10 µg/cm2 at cell seeding, comparedwith >100 µg/cm2 for regular membranes. The effects ofsoluble GAG in the absence of a membrane were alsoexamined using concentrations characteristic of the earlyand late phases of a regular membrane culture (i.e., 160 and7 µg/ml). The results of these studies are presented inFigures 7 and 8.
All conditions, with the exception of 160 µg/ml heparin,supported cell growth. Figure 7 shows that 160 µg/mlheparin in solution completely inhibited cell proliferation. Infact, microscopic examination confirmed that this concentra-tion was toxic, and all seeded cells died. Of the remainingheparin conditions, the regular membranes exhibited the low-est level of proliferation (25-fold increase at week 4), and thedepleted membrane exhibited approximately a 50-foldincrease at week 4. Similar total cell results were obtainedwith the CSB conditions, with the notable difference that 160µg/ml CSB in solution supported cell proliferation well.
The CD34+ cell results indicated that the regular mem-branes produce the highest proliferation for heparin, with apeak of 40-fold at four weeks. The heparin-depleted mem-branes and 7 µg/ml heparin in solution conditions showedsimilar profiles with continuously increasing CD34+ cellnumbers up to week 6. In contrast, the 160 µg/ml CSB con-dition exhibited the highest CD34+ cell proliferation for thisGAG with a 40-fold increase at week 2. CD34+ cellpercentage data (not shown) indicated that in addition toregular heparin and CSB membranes, 160 µg/ml CSB alsoproduced significant CD34+ cell enrichment.
While most proliferating cells were nonadherent in thesoluble GAG conditions, we noted significant proliferationof a heterogeneous adherent cell population in these
Table 1: Quantity of GAG initially bound per well. Approximately 1 mgof chitosan was deposited per well. Each well had an area of 1.9 cm2.Data are mean and standard deviation from six wells.
GAG Initial amount ( mg/cm2)
Heparin 110.60 ± 4.97
CSA 99.45 ± 5.55
CSB 102.31 ± 5.36
CSC 115.25 ± 5.48
Figure 6. Fractional heparin content of heparin-chitosan mem-brane versus time after addition of fresh culture medium.Valuesare normalized to the heparin quantity initially immobilized.
Madihally, Flake, Matthew 302
membrane-free cultures. This population was apparent withboth CSB concentrations as well as the low heparin case.While adherent cells were also observed on the depletedmembranes, their numbers were significantly lower.
Taken together, these results indicate that soluble GAGscontribute to the proliferative effects of the GAG-chitosanmembranes. The effects are strongly concentration-depen-dent, and optimal level is dependent on the particular GAG.The results also suggest that the highly transient nature of theGAG levels above regular membranes may play a major rolein the maintenance of and enrichment with CD34+ cells seenwith these two GAGs.
DISCUSSION
The difficulties associated with generating large quanti-ties of hematopoietic stem cells (HSCs) severely limit thewidespread use of allogeneic HSC transplantation as an effec-tive treatment for hematologic and malignant disease. In addi-tion, the success rate of hematopoietic transplant therapiescould be significantly increased if large quantities of trans-plant material with a high incidence of HSCs could be reli-ably generated. In most hematopoietic expansion schemes,
expansion and lineage differentiation proceed in parallel, andHSC expansion per se is either limited or nonexistent. Thislimitation is exacerbated by the fact that the mechanism bywhich stem cells are induced to undergo either self-renewal orlineage differentiation is poorly understood. The difficulty ofcontrolling HSC growth in vitro can be partly attributed to themany complex interactions existing in most experimental sys-tems and points to the need for alternate systems by whichfactors that are believed to play a role in HSC renewal can bestudied individually.
This study illustrates an in vitro system designed to pre-sent individual GAGs in both soluble and immobilized form.Hence, it provides a mechanism by which the role of GAGsin hematopoietic control may be studied without the con-founding factors associated with a stromal cell layer. CD34-enriched cord blood cells were cultured on defined ECMpolysaccharide membranes, and their proliferative activitywas evaluated. Our results show that it is possible not only toexpand cell numbers greatly but also to simultaneously enricha culture in CD34+ cells using this system. The data presentedare from a total of three cord blood culture runs. However, inseveral previously conducted experimental runs, the trends of
Figure 7. Effects of regular membranes, GAG-depleted mem-branes, and soluble GAGs on total cell proliferation.Right and lefty-axes display total cell number and expansion ratio, respectively.The expansion ratio was obtained by dividing the total cell number atany time point by the number of cells initially seeded. Data points anderror bars represent the mean and standard deviation from a repre-sentative culture, with a minimum of three replicate culture wells.Lower error bars have been omitted for clarity.
Figure 8. Effect of regular membranes, GAG-depleted membranes,and soluble GAGs on CD34+ cell proliferation.Right and left y-axesdisplay CD34+ cell number and expansion ratio, respectively. Theexpansion ratio was obtained by dividing the number of CD34+ cellsat any time point by the number of CD34+ cells initially seeded. Datapoints and error bars represent the mean and standard deviationfrom a representative culture, with a minimum of three replicateculture wells. Lower error bars have been omitted for clarity.

303 Glycosaminoglycan Effects on CD34+ Cell Growth
heparin and CSB superiority are consistent and reproducible.Those additional runs are not incorporated, because they didnot include all the GAGs presented here and some were ofshorter duration. Since we wished to evaluate the differencesbetween GAGs, we have presented only those runs in whichall the GAGs were studied simultaneously.
A number of interesting observations were made withregard to the relative effects of various GAGs on cellproliferation. First, the difference in response to CSA(chondroitin-4-sulfate) and CSC (chondroitin-6-sulfate)was striking given that the two GAG species differ only inthe location of the sulfate group on the disaccharide repeat-ing unit. CSC consistently exhibited the highest total cellproliferation of any GAG, although it performed poorlywith regard to maintenance of the CD34+ population. Incontrast, the apparent toxicity of CSA to these cellssuggests that the effect may have been mediated by interac-tions involving highly specific recognition of CSA. SinceCSA is not known to be inherently toxic to any cell type,the possibility of an apoptotic mechanism may bear furtherinvestigation. The superiority of CSB in CD34 maintenancealso points to the existence of highly specific interactions.CSB differs from CSA and CSC by the location of thesulfate group and the presence of L-iduronic acid as aconstituent monosaccharide [26]. Thus, it is likely thatunique binding characteristics with cytokines and/orreceptors may be key to its effect. In comparing heparin and CSB, we noted that although CSB generally reached its maximum CD34+ expansion before heparin (week 2 versus week 4), there appeared to be no correlationbetween the proliferation rate and the CD34+ percentagewith either GAG.
The observed total cell expansion was similar to thatreported for the expansion of CD34+ cells in stroma-free,cytokine-based (specifically, IL-3 and SCF) systems [27-29]. The results of De Bruyn et al.[27] indicated thatthe expansion of CD34+ cells peaked at day 14, in all com-binations of SCF, IL-3, IL-6, GM-CSF, and anti-transform-ing growth factor-b. After day 14, the population declined.In our experiments, expansion of the CD34+ population wasobserved with certain GAGs up to four weeks. Some cul-tures on heparin surfaces maintained high percentages ofCD34+ cells with expansion even up to six weeks. Theseconditions, specifically those incorporating heparin andCSB, were able to produce a selective expansion of themore primitive progenitors while apparently inhibitingextensive differentiation. In spite of the fact that CD34expression was maintained on heparin and CSB surfaces,the precise functional characteristics of the cells expandedin our cultures have not yet been determined. Gupta et al.
[9] reported that heparin did not support maintenance of the LTC-ICs. Our results show that for heparin, the CD34+
culture outcomes ranged from minimal proliferation toextensive proliferation to full toxicity as a function of theheparin levels. This concentration effect is supported byreports that variations in GAG levels may play a significantrole in the regulation of hematopoiesis [30, 31]. Notably,the activities of IL-3 and GM-CSF were shown to increaseas the concentration of heparan sulfate increased [30].Activity reached a maximum with increasing GAG concen-tration followed by a decline in activity with furtherincrease in the GAG level.
Our heparin desorption studies indicated that while thecomplex membranes were synthesized in such a manner asto ensure that the cell contact surface was comprisedprimarily of the GAG component, most of the GAGleached into solution over the course of the culture. Thissuggests that the observed effects may in fact be due mainlyto soluble GAG interactions as opposed to interactions withsurface-bound molecules. In addition, the desorptionexperiments showed that solution concentrations fall by anorder of magnitude during culture. Thus, the relativeimportance of surface-bound versus soluble GAG maychange with time, leading to a greater role for thatimmobilized on the surface at later times in culture.
The mechanism by which GAGs may alter the pro-liferative disposition of an HSC-enriched population in oursystem is not yet clear. Clearly, the ability of GAG speciesto bind and enhance the activities of some hematopoieticgrowth factors is recognized and can be expected to play arole. By the same token, GAGs may shield, sequester, orotherwise reduce the availability of other growth factors.For any given growth factor, these GAG-mediated effectson HSCs can reasonably be expected to vary with the GAGconcentration. In light of these possibilities, additionalstudies are clearly needed to identify concentrationthresholds and the precise nature of the molecular interac-tions involved. Likewise, the relative roles of immobilizedand soluble GAG must be further clarified.
Our observations on the effects of soluble GAGs andGAG-depleted membranes suggest a number of potentialmechanisms by which selective proliferation of and enrich-ment with CD34+ cells may have occurred in our system.The toxicity of high heparin levels suggests that the highsolution concentration during the early stages of culture onregular membranes may have selectively killed or inhibitedthe proliferation of committed progenitors. Furthermore,the lower levels found later in culture may have providedappropriate enhancement of early progenitor growth. TheGAG-chitosan membrane may also have had an effect

Madihally, Flake, Matthew 304
separate to the growth factor activating ability of the GAGcomponent. Prior work in our laboratory has shown that GAG-chitosan membranes limit the attachment andinhibit proliferation of a number of cell types, includingfibroblasts, smooth muscle cells, and endothelial cells.Thus, it is likely that the surface would have limitedattachment and spreading of contaminating stromal com-ponents and more differentiated cells in our CD34+
preparation. Limited cell spreading is often accompaniedby reduced function. Since such cells are known tosecrete differentiation-triggering soluble factors, reducedfunction may have correlated with a reduced differentia-tion signal, thus allowing proportionately greater self-renewal of CD34+ cells. This concept is supported by theobservation that the reduction in GAG content later incultures coincided with the appearance of an adherentpopulation. In keeping with this idea, reductions inCD34+ cell content between weeks 4 and 6 may havebeen partially caused by an increasing differentiation-inducing signal from the growing adherent population.Finally, it has recently been shown that heparin is capa-ble of activating a fibroblast growth factor receptor [32].While this receptor may not be of major significance for
HSC proliferation, the fact raises the possibility that sim-ilar direct GAG effects may occur in this system at someoptimal concentration.
It should be noted that since expression of the CD33and CD38 antigens was not evaluated in our expanded cellpopulation, the precise nature of the CD34+ cells at fourweeks is not known. It is possible that the expanded popu-lation may represent mostly early progenitors already com-mitted to a differentiation track, as opposed to true,uncommitted “stem cells.” In conclusion, this culture sys-tem has demonstrated the ability to produce simultaneousproliferation and CD34+ cell enrichment of a partially puri-fied cord blood population by controlling the nature andlevels of GAG moieties to which the cells are exposed.
ACKNOWLEDGMENTS
The authors wish to acknowledge the advice and edito-rial assistance of Dr. James Eliasonof the KarmanosCancer Institute. We also wish to acknowledge MaureenHoosang, R.N.and staff of the Hutzel Hospital, Labor andDelivery Center for assistance in the collection of cordblood samples. Support for this research was provided bythe National Institutes of Health grant #RO3-DK52278.
REFERENCES
1 Dexter TM. Growth and differentiation in the haemopoieticsystem. Biochem Soc Trans 1991;19:303-306.
2 Ploemacher RE, Van’t Hull E, Van Soest PL. Studies of hemo-poietic microenvironments: effects of acid mucopolysaccha-rides and dextran sulfate on erythroid and granuloiddifferentiation in vitro. Exp Hematol 1978;6:311-320.
3 Gallagher JT, Spooncer E, Dexter TM. Role of the cellularmatrix in haemopoiesis. I. Synthesis of glycosaminogly-cans by mouse bone marrow cell cultures. J Cell Sci1983;63:155-171.
4 Testa NG, Dexter TM. The biology of long-term bone marrowcultures and its application to bone marrow transplantation.Curr Opin Oncol 1991;3:272-278.
5 Yoder MC, Williams DA. Matrix molecule interactions withhematopoietic stem cells. Exp Hematol 1995;23:961-967.
6 Bruno E, Luikart SD, Long MW et al. Marrow-derived heparansulfate proteoglycan mediates the adhesion of hematopoieticprogenitor cells to cytokines. Exp Hematol 1995;23:1212-1217.
7 Gordon MY, Riley GP, Watt SM et al. Compartmentalizationof a haematopoietic growth factor (GM-CSF) by glyco-saminoglycans in the bone marrow microenvironment.Nature 1987;326:403-405.
8 Graham GJ, Wright EG, Hewick R et al. Identification andcharacterization of an inhibitor of haemopoietic stem cellproliferation. Nature 1990;344:442-444.
9 Gupta P, McCarthy JB, Verfaillie CM. Stromal fibroblastheparan sulfate is required for cytokine-mediated ex vivomaintenance of human long-term culture-initiating cells.Blood 1996;87:3229-3236.
10 Jaques LB. Glycosaminoglycans as polyelectrolytes: rejuve-nation of original concepts. Semin Thromb Hemost1991;17(suppl 1):1-4.
11 Kjellen L, Lindahl U. Proteoglycans: structure and interac-tions. Ann Rev Biochem 1991;60:443-475.
12 Roberts R, Gallagher J, Spooncer E et al. Heparan sulphatebound growth factors: a mechanism for stromal cell mediatedhaemopoiesis. Nature 1988;332:376-378.
13 Siczkowski M, Clarke D, Gordon MY. Binding of primitivehematopoietic progenitor cells to marrow stromal cellsinvolves heparan sulfate. Blood 1992;80:912-919.
14 Spooncer E, Gallagher JT, Krizsa F et al. Regulation of haemopoiesis in long-term bone marrow cultures. IV.Glycosaminoglycan synthesis and the stimulation ofhaemopoiesis by beta-D-xylosides. J Cell Biol 1983;96:510-514.
15 Matthew HWT, Salley SO, Peterson WD et al. Complex coac-ervate microcapsules for mammalian cell culture and artificialorgan development. Biotechnol Prog 1993;9:510-519.
16 Chandy T, Sharma CP. Chitosan as a biomaterial. BiomaterArtif Cells Artif Organs 1990;18:1-24.

305 Glycosaminoglycan Effects on CD34+ Cell Growth
17 Hayashi T. Biodegradable polymers for biomedical uses.Prog Poly Sci 1994;19:663-702.
18 Dunn JCY, Tompkins RG, Yarmush ML. Long-term invitro function of adult hepatocytes in a collagen sandwichconfiguration. Biotechnol Prog 1991;7:237-245.
19 Gold EW. A simple spectrophotometric method for estimat-ing glycosaminoglycan concentrations. Anal Biochem1979;99:183-188.
20 Smith PK, Mallia AK, Hermanson GT. Colorimetric methodfor the assay of heparin content in immobilized heparinpreparations. Anal Biochem 1980;109:466-473.
21 Kiraly K, Lapvetelainen T, Arokoski J et al. Application of selected cationic dyes for the semiquantitative estimation ofglycosaminoglycans in histological sections of articular cartilageby microspectrophotometry. Histochem J 1996;28:577-590.
22 Matthew HW, Basu S, Peterson WD et al. Performance of plasma-perfused, microencapsulated hepatocytes:prospects for extracorporeal liver support. J Pediatr Surg1993;28:1423-1427; discussion 1427-1428.
23 Denuziere A, Ferrier D, Damour O et al. Chitosan-chondroitin sulfate and chitosan-hyaluronate polyelectrolytecomplexes: biological properties. Biomaterials 1998;19:1275-1285.
24 Kratz G, Back M, Arnander C et al. Immobilised heparinaccelerates the healing of human wounds in vivo. Scand JPlast Reconstr Surg Hand Surg 1998;32:381-385.
25 Kratz G, Arnander C, Swedenborg J et al. Heparin-chitosancomplexes stimulate wound healing in human skin. Scand J Plast Reconstr Surg Hand Surg 1997;31:119-123.
26 Lindahl U, Hook M. Glycosaminoglycans and their bindingto biological macromolecules. Annu Rev Biochem1978;47:385-417.
27 De Bruyn C, Delforge A, Bron D et al. Ex vivo expansion of CD34+ CD38- cord blood cells. J Hematother 1997;6:93-102.
28 Moore MA, Hoskins I. Ex vivo expansion of cord blood-derived stem cells and progenitors, Blood Cells1994;20:468-479.
29 Traycoff CM, Abboud MR, Laver J et al. Ex vivo expansionof CD34+ cells from purified adult human bone marrow andumbilical cord blood hematopoietic progenitor cells. Prog Clin Biol Res 1994;389:385-391.
30 Alvarez-Silva M, Borojevic R. GM-CSF and IL-3 activitiesin schistosomal liver granulomas are controlled by stroma-associated heparan sulfate proteoglycans. J Leukoc Biol1996;59:435-441.
31 Seiffert D. The glycosaminoglycan binding site governsligand binding to the somatomedin B domain of vitronectin. J Biol Chem 1997;272:9971-9978.
32 Gao G, Goldfarb M. Heparin can activate a receptor tyrosinekinase. EMBO J 1995;14:2183-2190.
Related Documents











