MAI MUUTTUNUT PILU MULUT MOONHITI US009943555B2 ( 12 ) United States Patent Falb et al . ( 10 ) Patent No . : US 9 , 943 , 555 B2 ( 45 ) Date of Patent : * Apr . 17 , 2018 ( 54 ) BACTERIA ENGINEERED TO REDUCE HYPERPHENYLALANINEMIA 2014 / 0079701 Al 2015 / 0238545 A1 2015 / 0359894 Al 3 / 2014 Miller et al . 8/ 2015 Borody 12 / 2015 Weinrich et al . ( 71 ) Applicant : Synlogic , Inc ., Cambridge , MA ( US ) FOREIGN PATENT DOCUMENTS ( 72 ) Inventors : Dean Falb , Sherborn , MA ( US ); Vincent M . Isabella , Cambridge , MA ( US ); Jonathan W . Kotula , Somerville , MA ( US ); Paul F . Miller , Salem , CT ( US ); Yves Millet , Newton , MA ( US ); Sarah Rowe , Somerville , MA ( US ) CN WO WO WO WO WO WO WO 1154845 A WO 2009 / 004595 A2 WO 2013 / 192543 A2 WO 2014 / 018832 A1 WO 2014 / 066945 Al WO 2014 / 138324 AL WO 2016 / 183532 AL WO 2016 / 210373 A2 7 / 1997 1 / 2009 12 / 2013 1 / 2014 5 / 2014 9 / 2014 11 / 2016 12 / 2016 ( 73 ) Assignee : Synlogic , Inc ., Cambridge , MA ( US ) OTHER PUBLICATIONS ( * ) Notice : Subject to any disclaimer , the term of this patent is extended or adjusted under 35 U .S .C . 154 (b ) by 0 days . This patent is subject to a terminal dis claimer . ( 21 ) Appl . No . : 15 / 260 , 211 ( 22 ) Filed : Sep . 8 , 2016 ( 65 ) Prior Publication Data US 2017 / 0014457 A1 Jan . 19 , 2017 Related U .S . Application Data ( 63 ) Continuation of application No . 15 / 154 , 934 , filed on May 13 , 2016 . ( 60 ) Provisional application No . 62 / 161 , 137 , filed on May 13 , 2015 , provisional application No . 62 / 256 , 052 , filed on Nov . 16 , 2015 . Coban et al Screening of phenylpyruvic acid producers and opti mization of culture conditions in bench scale bioreactors Bioprocess Biosyst Eng ( 2014 ) 37 : 2343 - 2352 .* Xingyuan et al . 2000 New Strategeutics of Gene Therapy for Hyperphenylalaninemia Rats Beijing Red - Cross Chaoyang Hospi tal , Capital University ofMedical Sciences Beijing , 100020 ; China . Translation .* Xingyuan et al . , A new strategeutics of gene therapy for hyperphenylalaninemia rats , Beijing Red Cross Chaoyang Hospital , Capital University of Medical Sciences Beijing , 100020 China Abstract .* IMarbach et al ac operon induction in Escherichia coli : Systematic comparison of IPTG and TMG induction and influence of the transacetylase LacA Journal of Biotechnology 157 ( 2012 ) 82 - 88 .* Chen et al High - level Expression of Phenylalanine Ammonia - lyase in Lactococcus lactis via Synthesized Sequence Based on Bias Codons Chinese Journal of Biotechnology , Mar . 2006 , vol . 22 , No . 2 , pp . 187 - 190 ( translation ) .* Pascalle et al Controlled Gene Expression Systems for Lactococcus lactis with the Food - Grade Inducer Nisin Applied and Environmen tal Microbiology , Oct . 1996 , p . 3662 - 3667 . * Folling et al The discovery of phenylketonuria Acta Paldlatr Suppl 407 : 4 - 10 . 1994 .* Xingyuan et al . A New Strategeutics of Gene Therapy for Hyperphenylalaninemia RatsBeijing Red - Cross Chaoyang Hospi tal , Capital University of Medical Sciences Beijing , 100020 ; China . * Marbach et al ac operon induction in Escherichia coli : Systematic comparison of IPTG and TMG induction and influence of the transacetylase LacA Journal of Biotechnology 157 ( 2012 ) 82 - 88 .* Al Hafid , N and J. Christodoulou ( Oct . 2015 ) “ Phenylketonuria : a review of current and future treatments " Transl Pediatr , 4 ( 4 ): 304 317 . Albiniak , A .M . et al . ( 2013 ) “ High - level secretion of a recombinant protein to the culture medium with a Bacillus subtilis twin - arginine translocation system in Escherichia coli ” FEBS J, 280 : 3810 - 3821 . Altenhoeffer et al . ( Apr . 9 , 2004 ) “ The probiotic Escherichia coli strain Nissle 1917 interferes with invasion of human intestinal epithelial cells by different enteroinvasive bacterial pathogens ” FEMS Immunol Med Microbiol , 40 (3 ) : 223 - 229 . ( Continued ) ( 51 ) Int . Cl . A61K 35 / 741 ( 2015 . 01 ) CO7K 14 / 245 ( 2006 . 01 ) C12R 1/ 19 ( 2006 . 01 ) A61K 35 / 74 ( 2015 . 01 ) C12N 9/ 88 ( 2006 . 01 ) C12N 9 / 06 ( 2006 . 01 ) C12N 15 / 70 ( 2006 . 01 ) A61K 35 / 00 ( 2006 . 01 ) ( 52 ) U .S . CI . CPC . .. .. . .. ... A61K 35 / 74 ( 2013 . 01 ) ; COZK 14 / 245 ( 2013 . 01 ) ; C12N 9/ 0014 ( 2013 . 01 ); C12N 9 / 0022 ( 2013 . 01 ) ; C12N 9 / 88 ( 2013 . 01 ) ; C12N 15 / 70 ( 2013 . 01 ) ; C12Y 104 / 03002 ( 2013 . 01 ); C12 Y 403 / 01024 ( 2013 . 01 ); A61K 2035 / 11 ( 2013 . 01 ) ( 58 ) Field of Classification Search CPC .. . . .. . A61K 35 / 741 ; CO7K 14 / 245 ; C12R 1/ 19 See application file for complete search history . Primary Examiner — Maria G Leavitt ( 74 ) Attorney , Agent , or Firm — Finnegan , Henderson , Farabow , Garrett & Dunner LLP ( 56 ) References Cited U .S . PATENT DOCUMENTS 5 , 589 , 168 A 5 , 989 , 463 A 6 , 203 , 797 B1 6 , 835 , 376 B1 7 , 731 , 976 B2 12 / 1996 Allen et al . 11 / 1999 Tracy et al . 3 / 2001 Perry 12 / 2004 Neeser et al . 6 / 2010 Cobb et al . ( 57 ) ABSTRACT Genetically engineered bacteria , pharmaceutical composi tions thereof , and methods of modulating and treating dis eases associated with hyperphenylalaninemia are disclosed . 14 Claims , 90 Drawing Sheets

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

MAI MUUTTUNUT PILU MULUT MOONHITI US009943555B2
( 12 ) United States Patent Falb et al .
( 10 ) Patent No . : US 9 , 943 , 555 B2 ( 45 ) Date of Patent : * Apr . 17 , 2018
( 54 ) BACTERIA ENGINEERED TO REDUCE HYPERPHENYLALANINEMIA
2014 / 0079701 Al 2015 / 0238545 A1 2015 / 0359894 Al
3 / 2014 Miller et al . 8 / 2015 Borody
12 / 2015 Weinrich et al . ( 71 ) Applicant : Synlogic , Inc . , Cambridge , MA ( US ) FOREIGN PATENT DOCUMENTS ( 72 ) Inventors : Dean Falb , Sherborn , MA ( US ) ;
Vincent M . Isabella , Cambridge , MA ( US ) ; Jonathan W . Kotula , Somerville , MA ( US ) ; Paul F . Miller , Salem , CT ( US ) ; Yves Millet , Newton , MA ( US ) ; Sarah Rowe , Somerville , MA ( US )
CN WO WO WO WO WO WO WO
1154845 A WO 2009 / 004595 A2 WO 2013 / 192543 A2 WO 2014 / 018832 A1 WO 2014 / 066945 Al WO 2014 / 138324 AL WO 2016 / 183532 AL WO 2016 / 210373 A2
7 / 1997 1 / 2009
12 / 2013 1 / 2014 5 / 2014 9 / 2014 11 / 2016 12 / 2016 ( 73 ) Assignee : Synlogic , Inc . , Cambridge , MA ( US )
OTHER PUBLICATIONS ( * ) Notice : Subject to any disclaimer , the term of this patent is extended or adjusted under 35 U . S . C . 154 ( b ) by 0 days . This patent is subject to a terminal dis claimer .
( 21 ) Appl . No . : 15 / 260 , 211
( 22 ) Filed : Sep . 8 , 2016
( 65 ) Prior Publication Data US 2017 / 0014457 A1 Jan . 19 , 2017
Related U . S . Application Data ( 63 ) Continuation of application No . 15 / 154 , 934 , filed on
May 13 , 2016 . ( 60 ) Provisional application No . 62 / 161 , 137 , filed on May
13 , 2015 , provisional application No . 62 / 256 , 052 , filed on Nov . 16 , 2015 .
Coban et al Screening of phenylpyruvic acid producers and opti mization of culture conditions in bench scale bioreactors Bioprocess Biosyst Eng ( 2014 ) 37 : 2343 - 2352 . * Xingyuan et al . 2000 New Strategeutics of Gene Therapy for Hyperphenylalaninemia Rats Beijing Red - Cross Chaoyang Hospi tal , Capital University of Medical Sciences Beijing , 100020 ; China . Translation . * Xingyuan et al . , A new strategeutics of gene therapy for hyperphenylalaninemia rats , Beijing Red Cross Chaoyang Hospital , Capital University of Medical Sciences Beijing , 100020 China Abstract . * IMarbach et al ac operon induction in Escherichia coli : Systematic comparison of IPTG and TMG induction and influence of the transacetylase LacA Journal of Biotechnology 157 ( 2012 ) 82 - 88 . * Chen et al High - level Expression of Phenylalanine Ammonia - lyase in Lactococcus lactis via Synthesized Sequence Based on Bias Codons Chinese Journal of Biotechnology , Mar . 2006 , vol . 22 , No . 2 , pp . 187 - 190 ( translation ) . * Pascalle et al Controlled Gene Expression Systems for Lactococcus lactis with the Food - Grade Inducer Nisin Applied and Environmen tal Microbiology , Oct . 1996 , p . 3662 - 3667 . * Folling et al The discovery of phenylketonuria Acta Paldlatr Suppl 407 : 4 - 10 . 1994 . * Xingyuan et al . A New Strategeutics of Gene Therapy for Hyperphenylalaninemia RatsBeijing Red - Cross Chaoyang Hospi tal , Capital University of Medical Sciences Beijing , 100020 ; China . * Marbach et al ac operon induction in Escherichia coli : Systematic comparison of IPTG and TMG induction and influence of the transacetylase LacA Journal of Biotechnology 157 ( 2012 ) 82 - 88 . * Al Hafid , N and J . Christodoulou ( Oct . 2015 ) “ Phenylketonuria : a review of current and future treatments " Transl Pediatr , 4 ( 4 ) : 304 317 . Albiniak , A . M . et al . ( 2013 ) “ High - level secretion of a recombinant protein to the culture medium with a Bacillus subtilis twin - arginine translocation system in Escherichia coli ” FEBS J , 280 : 3810 - 3821 . Altenhoeffer et al . ( Apr . 9 , 2004 ) “ The probiotic Escherichia coli strain Nissle 1917 interferes with invasion of human intestinal epithelial cells by different enteroinvasive bacterial pathogens ” FEMS Immunol Med Microbiol , 40 ( 3 ) : 223 - 229 .
( Continued )
( 51 ) Int . Cl . A61K 35 / 741 ( 2015 . 01 ) CO7K 14 / 245 ( 2006 . 01 ) C12R 1 / 19 ( 2006 . 01 ) A61K 35 / 74 ( 2015 . 01 ) C12N 9 / 88 ( 2006 . 01 ) C12N 9 / 06 ( 2006 . 01 ) C12N 15 / 70 ( 2006 . 01 ) A61K 35 / 00 ( 2006 . 01 )
( 52 ) U . S . CI . CPC . . . . . . . . . . . A61K 35 / 74 ( 2013 . 01 ) ; COZK 14 / 245
( 2013 . 01 ) ; C12N 9 / 0014 ( 2013 . 01 ) ; C12N 9 / 0022 ( 2013 . 01 ) ; C12N 9 / 88 ( 2013 . 01 ) ; C12N 15 / 70 ( 2013 . 01 ) ; C12Y 104 / 03002
( 2013 . 01 ) ; C12 Y 403 / 01024 ( 2013 . 01 ) ; A61K 2035 / 11 ( 2013 . 01 )
( 58 ) Field of Classification Search CPC . . . . . . . A61K 35 / 741 ; CO7K 14 / 245 ; C12R 1 / 19 See application file for complete search history . Primary Examiner — Maria G Leavitt
( 74 ) Attorney , Agent , or Firm — Finnegan , Henderson , Farabow , Garrett & Dunner LLP ( 56 ) References Cited
U . S . PATENT DOCUMENTS 5 , 589 , 168 A 5 , 989 , 463 A 6 , 203 , 797 B1 6 , 835 , 376 B1 7 , 731 , 976 B2
12 / 1996 Allen et al . 11 / 1999 Tracy et al . 3 / 2001 Perry
12 / 2004 Neeser et al . 6 / 2010 Cobb et al .
( 57 ) ABSTRACT Genetically engineered bacteria , pharmaceutical composi tions thereof , and methods of modulating and treating dis eases associated with hyperphenylalaninemia are disclosed .
14 Claims , 90 Drawing Sheets

US 9 , 943 , 555 B2 Page 2
( 56 ) References Cited
OTHER PUBLICATIONS Andersen P . S . et al . ( Apr . 1995 ) “ Uracil uptake in Escherichia coli K - 12 : isolation of uraA mutants and cloning of the gene ” J Bacteriol , 177 ( 8 ) : 2008 - 2013 . Arai et al . ( Aug . 28 , 1995 ) “ Expression of the nir and nor genes for denitrification of Pseudomonas aeruginosa reguires a novel CRP / FNR - related transcriptional regulator , DNR , in addition to ANR ” FEBS Lett , 371 ( 1 ) : 73 - 76 . Argos , P . ( 1989 ) “ A possible homology between immunodeficiency virus p24 core protein and picornaviral VP2 coat protein : prediction of HIV p24 antigenic sites ” EMBO J , 8 ( 3 ) : 779 - 785 . Arthur et al . ( Oct . 5 , 2012 ) “ Intestinal inflammation targets cancer inducing activity of the microbiota ” Science , 338 ( 6103 ) : 120 - 123 . NIH Public Access Author Manuscript : available in PMC May 6 , 2013 ( 11 pages ) . Baek , J . O . et al . ( Apr . 2011 ) " Expression and characterization of a second L - amino acid deaminase isolated from Proteus mirabilis in Escherichia coli ” J Basic Microbiol , 51 : 129 - 135 . Bifulco , D . et al . ( 2013 ) “ A thermostable L - aspartate oxidase : a new tool for biotechnological applications " Appl Microbiol Biotechnol , 97 : 7285 - 7295 . Boysen , A . et al . ( Apr . 2010 ) “ Translational Regulation of Gene Expression by an Anaerobically Induced Small Non - coding RNA in Escherichia coli ” J Biol Chem , 285 ( 14 ) : 10690 - 10702 . Callura et al . ( Sep . 7 , 2010 ) “ Tracking , tuning , and terminating microbial physiology using synthetic riboregulators ” Proc Natl Acad Sci USA , 107 ( 36 ) : 15898 - 15903 . Castiglione et al . ( Sep . 2009 ) “ The transcription factor DNR from Pseudomonas aeruginosa specifically reguires nitric oxide and haem for the activation of a target promoter in Escherichia coli ” Microbiology , 155 ( Pt 9 ) : 2838 - 2844 . Chang ( 2007 ) “ Use of Enzyme Artificial Cells for Genetic Enzyme Defects ” In Artificial Bells : Biotechnology , Nanomedicine , Regen erative Medicine , Blood Substitutes , Bioencapsulation , and Cell / Stem Cell Therapy . Regenerative Medicine , Artificial Cells and Nanomedicine - vol . 1 . Singapore : World Scientific Publishing , pp . 147 - 159 . Clarkson et al . ( 1971 ) “ Diaminopimelic Acid and Lysine Auxotrophs of Pseudomonas aeruginosa 8602 " J Gen Microbiol , 66 : 161 - 169 . Collinson , I . et al . ( 2015 ) “ Channel crossing : how are proteins shipped across the bacterial plasma membrane ? ” Philos Trans R SOC B , 370 : 20150025 [ online ] . Retrieve from : http : / / rstb . royalsocietypublishing . org / , on Jun . 16 , 2016 ( 13 pages ) . Costa , T . R . D . et al . ( May 2015 ) " Secretion systems in Gram negative bacteria : structural and mechanistic insights ” Nat Rev Microbiol , 13 ( 6 ) : 343 - 359 . Cuevas - Ramos et al . ( Jun . 22 , 2010 ) “ Escherichia coli induces DNA damage in vivo and triggers genomic instability in mamma lian cells " Proc Natl Acad Sci USA , 107 ( 25 ) : 11537 - 11542 . Danino , T . et al . ( May 2015 ) “ Programmable probiotics for detec tion of cancer in urine ” Sci Transl Med , 7 ( 289 ) : 289ra84 [ online ] . Retrieved from : www . sciencetranslational medicine . org , Jul . 30 , 2015 ( 11 pages ) . Deutscher ( Apr . 2008 ) “ The mechanism of carbon catabolite repres sion in bacteria ” Curr Opin Microbiol , 11 ( 2 ) : 87 - 93 . Dinleyici et al . ( Nov . 2014 ) " Saccharomyces boulardii CNCM 1 - 745 in different clinical conditions ” Expert Opin Biol Ther , 14 ( 11 ) : 1593 - 1609 . Dobbelaere , D . et al . ( 2003 ) “ Evaluation of nutritional status and pathophysiology of growth retardation in patients with phenylketonuria ” J Inherit Metab Dis , 26 ( 1 ) : 1 - 11 . Duerre , J . and S . Chakrabarty ( Feb . 1975 ) “ L - Amino Acid Oxidases of Proteus rettgeri ” J Bacteriol , 121 ( 2 ) : 656 - 663 . Durand , S . and G . Storz ( Mar . 2010 ) “ Reprogramming of Anaerobic Metabolism by the FnrS Small RNA ” Mol Microbiol , 75 ( 5 ) : 1215 1231 . NIH Public Access Author Manuscript ; available in PMC Sep . 17 , 2010 ( 28 pages ) .
Eiglmeier et al . ( Jul . 1989 ) “ Molecular genetuc analysis of FNR dependent promoters ” Mol Microbiol , 3 ( 7 ) : 869 - 878 . Estrem , S . T . et al . ( Aug . 1998 ) “ Identification of an UP element consensus sequence for bacterial promoters " Proc Natl Acad Sci USA , 95 ( 17 ) : 9761 - 9766 . Galimand et al . ( Mar . 1991 ) “ Positive FNR - like control of anaerobic arginine degradation and nitrate respiration in Pseudomonas aeruginosa ” J Bacteriol , 173 ( 5 ) : 1598 - 1606 . Gardner et al . ( 2000 ) “ Construction of a genetic toggle switch in Escherichia coli ” Nature , 403 : 339 - 342 . GenBank Database Accession No . AA86752 ( Feb . 3 , 1996 ) “ amino acid deaminase [ Proteus mirabilis HI4320 ” [ online ] . National Cen ter for Biotechnology Information ( NCBI ) , U . S . National Library of Medicine ; http : / / www . ncbi . nlm . nih . gov / protein / AAA86752 ( 1 page ) . GenBank Database Accession No . AAH26251 . 1 ( Jul . 15 , 2006 ) “ Phenylalanine hydroxylase [ Homo sapiens ] ” [ online ] . National Center for Biotechnology Information ( NCBI ) , U . S . National Library of Medicine ; http : / / www . ncbi . nlm . nih . gov / protein / AAH26251 ( 2 pages ) . GenBank Database Accession No . ABA23593 . 1 ( Jan . 28 , 2014 ) “ histidine ammonia - lyase [ Anabaena variabilis ATCC 29413 ] ” [ online ] . National Center for Biotechnology Information ( NCBI ) , U . S . National Library of Medicine ; http : / / www . ncbi . nlm . nih . gov / protein / ABA23593 ( 2 pages ) . GenBank Database Accession No . ACD36582 . 1 ( Aug . 15 , 2011 ) “ L - amino acid deaminase [ Proteus mirabilis ) ” [ online ] . National Center for Biotechnology Information ( NCBI ) , U . S . National Library of Medicine ; http : / / www . ncbi . nlm . nih . gov / protein / ACD36582 ( 1 page ) . GenBank Database Accession No . BAA90864 . 1 ( Feb . 18 , 2000 ) “ L - amino acid deaminase [ Proteus vulgaris ] ” [ online ] . National Center for Biotechnology Information ( NCBI ) , U . S . National Library of Medicine ; http : / / www . ncbi . nlm . nih . gov / protein BAA90864 ( 1 page ) . GenBank Database Accession No . CAE15566 . 1 ( Feb . 27 , 2015 ) “ Histidine ammonia - lyase ( histidase ) [ Photorhabdus luminescens subsp . laumondii TT01 ] ” [ online ] . National Center for Biotechnol ogy Information ( NCBI ) , U . S . National Library of Medicine ; http : / / www . ncbi . nlm . nih . gov / protein / CAE15566 ( 2 pages ) . GenBank Database Accession No . EDV65095 . 1 ( Jun . 20 , 2008 ) " arromatic amino acid transport protein AroP [ Escherichia coli F11 ] ” [ online ] . National Center for Biotechnology Information ( NCBI ) , U . S . National Library of Medicine ; http : / / www . ncbi . nlm . nih . gov / protein / EDV65095 ( 2 pages ) . GenBank Database Accession No . EU669819 . 1 ( Aug . 15 , 2011 ) “ Proteus mirabilis L - amino acid deaminase gene , complete cds ” [ online ] . National Center for Biotechnology Information ( NCBI ) , U . S . National Library of Medicine ; http : / / www . ncbi . nlm . nih . gov / nuccore / EU669819 ( 2 pages ) . GenBank Database Accession No . U35383 . 1 ( Feb . 3 , 1996 ) “ Proteus mirabilis amino acid deaminase ( aad ) gene , complete cds ” [ online ] . National Center for Biotechnology Information ( NCBI ) , U . S . National Library of Medicine ; http : / / www . ncbi . nlm . nih . gov / nuccore / U35383 ( 2 pages ) . Gerdes et al . ( Oct . 2006 ) “ Essential genes on metabolic maps ” Curr Opin Biotechnol , 17 ( 5 ) : 448 - 456 . Gilbert , H . J . et al . ( Jan . 1985 ) “ Molecular cloning of the phenyl alanine ammonia lyase gene from Rhodosporidium toruloides in Escherichia coli K - 12 ” Bacteriol , 161 ( 1 ) : 314 - 320 . Görke and Stülke . ( Aug . 2008 ) “ Carbon catabolite repression in bacteria : many ways to make the most out of nutrients ” Nat Rev Microbiol , 6 ( 8 ) : 613 - 624 . Hasegawa et al . ( Sep . 15 , 1998 ) “ Activation of a consensus FNR dependent promoter by DNR of Pseudomonas aeruginosa in response to nitrite ” FEMS Microbiol Lett , 166 ( 2 ) : 213 - 217 . He , G . et al . ( Apr . 13 , 1999 ) “ Noninvasive measurement of ana tomic structure and intraluminal oxygenation in the gastrointestinal tract of living mice with spatial and spectral EPR imaging ” Proc Natl Acad Sci USA , 96 ( 8 ) : 4586 - 4591 . Hoeks , M . P . et al . ( Jan . 2009 ) “ Adult issues in phenylketonuria ” Neth J Med , 67 ( 1 ) : 2 - 7 .

US 9 , 943 , 555 B2 Page 3
( 56 ) References Cited OTHER PUBLICATIONS
Hoeren et al . ( Nov . 15 , 1993 ) “ Sequence and expression of the gene encoding the respiratory nitrous - oxide reductase from Paracoccus denitrificans ” Eur J Biochem , 218 ( 1 ) : 49 - 57 . Hosseini et al . ( May 2011 ) " Propionate as a health - promoting microbial metabolite in the human gut ” Nutr Rev , 68 ( 5 ) : 245 - 258 . Hou , Y . et al . ( Oct . 2015 ) “ Production of phenylpyruvic acid from L - phenylalanine using an L - amino acid deaminase from Proteus mirabilis : comparison of enzymatic and whole - cell biotransforma tion approaches " Appl Microbiol Biotechnol , 99 ( 20 ) : 8391 - 8402 . Isabella et al . ( Jan . 20 , 2011 ) “ Deep sequencing - based analysis of the anaerobic stimulon in Neisseria gonorrhoeae ” BMC Genomics , 12 : 51 ( 24 pages ) . Ivanovaska , V . et al . ( 2014 ) “ Pediatric Drug Formulations : A Review of Challenges and Progress ” Pediatrics , 134 : 361 - 372 . Kobe , B . et al . ( Jun . 1997 ) “ Regualtion and crystallization of phosphorylated and dephosphorylated forms of truncated dimeric phenylalanine hydroxylase ” Protein Sci , 6 ( 6 ) : 1352 - 1357 . Kwok , S . C . et al . ( Jan . 29 , 1985 ) “ Nucleotide sequence of a full - length complementary DNA clone and amino acid sequence of human phenylalanine hydroxylase ” Biochemistry , 24 ( 3 ) : 556 - 561 . Lee , D . H . et al . ( 2011 ) " Cumulative Number of Cell Divisions as a Meaningful Timescale for Adaptive Laboratory Evolution of Escherichia coli ” PLOS ONE , 6 : e26172 , http : / / dx . doi . org / 10 . 1371 / journal . pone . 0026172 ( 8 pages ) . Leonard , “ Disorders of the urea cycle and related enzymes " in Inborn Metabolic Diseases , 4th ed . Heidelberg : Springer Medizin Verlag , 2006 ; pp . 263 - 272 . Longo , N . et al . ( Jul . 5 , 2014 ) “ Phase 1 Trial of Subcutaneous rAVPAL - PEG in Subjects with Phenylketonuria ” Lancet , 384 ( 9937 ) : 37 - 44 . HHS Public Access Author Manuscript ; available in PMC Jul . 5 , 2015 ( 18 pages ) . Lopez and Anderson ( Dec . 2015 ) " Synthetic Auxotrophs with Ligand - Dependent Essential Genes for a BL21 ( DE3 ) Biosafety Strain " ACS Synthetic Biology , 4 ( 12 ) : 1279 - 1286 . MacDonald , M . J . and G . B . D ' Cunha ( 2007 ) “ A modern view of phenylalanine ammonia lyase ” Biochem Cell Biol , 85 ( 3 ) : 273 - 282 . MacLeod , E . L . et al . ( Jun . 2010 ) “ Nutritional Management of Phenylketonuria " Ann Nestle Eng , 68 ( 2 ) : 58 - 69 . Meadow , P . and W . Work ( 1959 ) “ Biosynthesis of diaminopimelic acid and lysine in Escherichia coli ” Biochem J , 72 ( 3 ) : 396 - 400 . Moffitt , M . C . et al . ( Jan . 30 , 2007 ) “ Discovery of two cyanobacterial phenylalanine ammonia lyases : kinetic and structural characteriza tion ” Biochemistry , 46 ( 4 ) : 1004 - 1012 . Moore et al . ( Nov . 3 , 2006 ) “ Regulation of FNR dimerization by subunit charge repulsion ” J Biol Chem , 281 ( 44 ) : 33268 - 33275 . Nougayrede et al . ( Aug . 11 , 2006 ) “ Escherichia coli induces DNA double - strand breaks in eukaryotic cells ” Science , 313 ( 5788 ) : 848 851 . Olier et al . ( Nov . - Dec . 2012 ) “ Genotoxicity of Escherichia coli Nissle 1917 strain cannot be dissociated from its probiotic activity ” Gut Microbes , 3 ( 6 ) : 501 - 509 . Pelmont , J . et al . ( 1972 ) “ L - aminoacide oxydases des enveloppes de Proteus mirabilis : propriétés générales ( L - amino acid oxidases of Proteus mirabilis : general properties ) ” Biochimie 54 ( 10 ) : 1359 1374 ( French ; English summary on p . 1359 ) . Pi , J . et al . ( Jun . 1991 ) " Cloning and sequencing of the phep gene , which encodes the phenylalanine - specific transport system of Escherichia coli ” J Bacteriol , 173 ( 12 ) : 3622 - 3629 . Pi , J . et al . ( Nov . 1998 ) “ Functional consequences of changing proline residues in the phenylalanine - specific permease of Escherichia coli ” J Bacteriol , 180 ( 21 ) : 5515 - 5519 . Pi , J . and J . Pittard ( May 1996 ) “ Topology of the phenylalanine specific permease of Escherichia coli ” J Bacteriol ; 178 ( 9 ) : 2650 2655 . Pugsley , A . P . ( Mar . 1993 ) “ The complete general secretory pathway in gram - negative bacteria ” Microbiol Rev , 57 ( 1 ) : 50 - 108 . Purcell , O . et al . ( 2013 ) “ Towards a whole - cell modeling approach for synthetic biology ” Chaos , 23 ( 2 ) : 025112 ( 8 pages ) .
Ray et al . ( Nov . 15 , 1997 ) “ The effects of mutation of the anr gene on the aerobic respiratory chain of Pseudomonas aeruginosa ” FEMS Microbiol Lett , 156 ( 2 ) : 227 - 232 . Reeves , A . Z . et al . ( Apr . 2015 ) “ Engineering E . coli into a protein delivery system for mammalian cells ” ACS Synth Biol , Just Accepted Manuscript , DOI : 10 : 1021 / acssynbio . 5b00002 [ online ] . Retrieved from : http : / / pubs . acs . org , on Apr . 20 , 2015 ( 26 pages ) . Final publication in vol . 5 , pp . 644 - 654 . Reister et al . ( Oct . 10 , 2014 ) “ Complete genome sequence of the Gram - negative probiotic Escherichia coli strain Nissle 1917 ” J Biotechnol , 187 : 106 - 107 . Rembacken et al . ( Aug . 21 , 1999 ) “ Non - pathogenic Escherichia coli versus mesalazine for the treatment of ulcerative colitis : a randomised trial ” Lancet , 354 ( 9179 ) : 635 - 639 . REFSEQ Database Accession No . NP _ 415108 . 1 ( Dec . 16 , 2014 ) “ phenylalanine transporter [ Escherichia coli str . K - 12 substr . MG16551 ” [ online ] . National Center for Biotechnology Information ( NCBI ) , U . S . National Library of Medicine ; http : / / www . ncbi . nlm . nih . gov / protein / NP _ 415108 ( 3 pages ) . REFSEQ Database Accession No . WP _ 011146484 . 1 ( May 24 , 2013 ) “ histidine ammonia - lyase [ Photorhabdus luminescens ) ” [ online ] . National Center for Biotechnology Information ( NCBI ) , U . S . National Library of Medicine ; http : / / www . ncbi . nlm . nih . gov / protein / WP _ 011146484 ( 1 page ) . Rigel , N . W . and Braunstein ( 2008 ) “ A new twist on an old path way — accessory secretion systems ” Mol Microbiol , 69 ( 2 ) : 291 - 302 . Saier Jr . , M . H . ( 2006 ) “ Protein Secretion and Membrane Insertion Systems in Gram - Negative Bacteria ” J Membrane Biol , 214 : 75 - 90 . Saier Jr . , M . H . ( 2006 ) “ Protein Secretion Systems in Gram - Nega tive Bacteria . Gram - negative bacteria possess many protein secre tion - membrane insertion systems that apparently evolved indepen dently ” Microbe , 1 ( 9 ) : 414 - 419 . Salmon , K . et al . ( Aug . 8 , 2003 ) “ Global gene expression profiling in Escherichia coli K12 . The effects of oxygen availability and FNR ” J Biol Chem , 278 ( 32 ) : 29837 - 29855 . Sarkissian , C . N . et al . ( Mar . 1999 ) “ A different approach to treat ment of phenylketonuria : Phenylalanine degradation with recom binant phenylalanine ammonia lyase ” Proc Natl Acad Sci USA , 96 ( 5 ) : 2339 - 2344 . Sarkissian , C . N . et al . ( Jun . 2007 ) “ Quantitation of phenylalanine and its trans - cinnamic , benzoic and hippuric acid metabolites in biologival fluids in a single GC - MS analysis ” J Mass Spectrom , 42 ( 6 ) : 811 - 817 . Sarkissian , C . N . et al . ( Nov . 2011 ) “ Evaluation of orally adminis tered PEGylated phenylalanine ammonia lyase in mice for the treatment of Phenylketonuria ” Mol Genet Metab , 104 ( 3 ) : 249 - 254 . NIH Public Access Author Manuscript ; available in PMC Nov . 1 , 2012 ( 15 pages ) . Sat et al . ( Mar . 2003 ) “ The Escherichia coli mazEF suicide module mediates thymineless death ” J Bacteriol , 185 ( 6 ) : 1803 - 1807 . Sawers ( Jun . 1991 ) " Identification and molecular characterization of a transcritional regulator from Pseudomonas aeruginosa PAO1 exhibiting structural and functional similarity to the FNR protein of Escherichia coli ” Mol Microbiol , 5 ( 6 ) : 1469 - 1481 . Schultz ( Jul . 2008 ) “ Cinical use of E . coli Nissle 1917 in inflam matory bowel disease ” Inflamm Bowel Dis , 14 ( 7 ) : 1012 - 1018 . Silhavy , T . J . et al . ( 2010 ) “ The bacterial cell envelope ” Cold Spring Harb Perspect Biol , 2 , a000414 ( 17 pages ) . Sonnenborn and Schulze ( 2009 ) “ The non - pathogenic Escherichia coli strain Nissle 1917 _ features of a versatile probiotic ” Microbial Ecology in Health and Disease , 21 : 122 - 158 . Stanley , S . A . et al . ( Oct . 2003 ) “ Acute infection and macrophage subversion by Mycobacterium tuberculosis require a specialized secretion system ” PNAS , 100 ( 22 ) : 13001 - 13006 . Steele , R . D . ( Jun . 1986 ) “ Blood - brain barrier transport of the alpha - keto acid analogs of amino acids ” Fed Proc , 45 ( 7 ) : 2060 2064 . Trunk et al . ( Jun . 2010 ) “ Anaerobic adaptation in Pseudomonas aeruginosa : definition of the Anr Dnr regulons ” Environ Microbiol , 12 ( 6 ) : 1719 - 1733 .

US 9 , 943 , 555 B2 Page 4
( 56 ) References Cited
OTHER PUBLICATIONS
Ukena et al . ( Dec . 12 , 2007 ) “ Probiotic Escherichia coli Nissle 1917 inhibits leaky gut by enhancing mucosal integrity ” PLoS One , 2 ( 12 ) : e1308 . [ online ] DOI : 10 . 1371 / journal . pone . 0001308 ( 11 pages ) . Unden et al . ( Jul . 4 , 1997 ) “ Alternative respiratory pathways of Escherichia coli : energetics and transcritional regulation in response to electron acceptors ” Biochim Biophys Acta , 1320 ( 3 ) : 217 - 234 . UniProtKB / Swiss - Prot Database Accession No . Q3M5Z3 . 1 ( Nov . 11 , 2015 ) “ RecName : Full - Phenylalanine ammonia - lyase ” National Center for Biotechnology Information ( NCBI ) , U . S . National Library of Medicine , http : / / www . ncbi . nlm . nih . gov / pro tein / Q3M5Z3 ( 7 pages ) . Vockley , J . et al . ( Feb . 2014 ) “ Phenylalanine hydroxylase defi ciency : diagnosis and management guideline ” Genet Med , 16 ( 2 ) : 188 - 200 . Wanner , L . A . et al . ( Jan . 1995 ) “ The phenylalanine ammonia - lyase gene family in Arabidopsis thaliana ” Plant Mol Biol , 27 ( 2 ) : 327 338 . Williams , J . S . et al . ( Aug . 2005 ) “ The gene stlA encodes a phenyl alanine ammonia - lyase that is involved in the production of a stilbene antibiotic in Photorhabdus luminescens TT01 ” Microbiol ogy , 151 ( Pt 8 ) : 2543 - 2550 . Winteler et al . ( Mar . 1996 ) “ The homologous regulators ANR of Pseudomonas aeruginosa and FNR of Escherichia coli have over lapped but distinct specificities for anaerobically inducible promot ers ” Microbiology , 142 ( Pt 3 ) : 685 - 693 . Wright et al . ( Mar . 20 , 2015 ) “ GeneGuard : A modular plasmid system designed for biosafety ” ACS Synth Biol , 4 ( 3 ) : 307 - 316 . Wu et al . ( Oct . 7 , 2015 ) “ Direct regulation of the natural compe tence egulator gene tfoX by cyclic AMP ( CAMP ) and CAMP receptor protein in Vibrios ” Sci Rep , 5 : 14921 ( 15 pages ) . Xiang , L and B . S . Moore ( Jun . 2005 ) “ Biochemical characterization of a prokaryotic phenylalanine ammonia lyase ” J Bacteriol , 187 ( 12 ) : 4286 - 4289 . Zhang and Lin ( 2009 ) “ DEG 5 . 0 , a database of essential genes in both prokaryotes and eukaryotes ” Nucl Acids Res , 37 ( suppl . 1 ) : D455 - D458 . U . S . Appl . No . 62 / 183 , 935 , filed Jun . 24 , 2015 , by Kotula et al . U . S . Appl . No . 62 / 184 , 811 , filed Jun . 25 , 2015 , by Falb et al . U . S . Appl . No . 62 / 263 , 329 , filed Dec . 4 , 2015 , by Kotula et al .
Becker , S . et al . ( Aug . 1996 ) “ O , as the Regulatory Signal for FNR - Dependent Gene Regulation in Escherichia coli ” J Bacteriol , 178 ( 15 ) : 4515 - 4521 . Blau , N . and N . Longo ( 2015 ) “ Alternative therapies to address the unmet medical needs of patients with phenylketonuria ” Expert Opin Pharmacother , 16 ( 6 ) : 791 - 800 . Braat , H . et al . ( 2006 ) “ A Phase I Trial With Transgenic Bacteria Expressing Interleukin - 10 in Crohn ' s Disease ” Clin Gastroenterol Hepatol , 4 : 754 - 759 Christodoulou , J . et al . ( Nov . 2012 ) “ Enzyme substitution therapy for phenylketonuria delivered orally using a genetically modified probiotic : Proof of principle ” 62nd Annual Meeting of the American Society of Human Genetics , Nov . 6 - 10 , 2012 , San Francisco , CA ; Program No . 166 , Nov . 8 , 2012 . Huibregtse , I . L . et al . ( 2012 ) “ Genetically Modified Lactococcus lactis for Delivery of Human Interleukin - 10 to Dendritic Cells " Gastroenterol I Res Pract , vol . 2012 , Article ID 639291 ( 7 pages ) . International Patent Application No . PCT / US2016 / 032562 , filed May 13 , 2016 , by Synlogic , Inc . : International Search Repeat arid Written Opinion : dated Aug . 22 , 2016 . International Patent Application No . PCT / US2016 / 062369 , filed Nov . 16 , 2016 , by Synlogic , Inc . : International Search Report and Written Opinion ; dated Mar . 10 , 2017 . Kang , T . S . et al . ( 2010 ) “ Converting an injectable protein thera peutic into an oral form : Phenylalanine ammonia lyase for phenylketonuria ” Mol Genet Metabol , 99 : 4 - 9 . Liu , J . et al . ( 2002 ) “ Study on a Novel Strategy to Treatment of Phenylketonuria ” Art Cells , Blood Subs , and Immob Biotech , 30 ( 4 ) : 243 - 257 . Mengesha , A . et al . ( 2006 ) “ Development of a flexibie and potent hypoxiainducible promoter for tumor - targeted gene expression in attenuated Salmonella ” Cancer Biology & Therapy , 5 ( 9 ) 1120 - 1128 . Sleator , R . D . and C . Hill ( 2009 ) “ Rational Design of Improved Pharmabiotics ” J Biomed Biotechnol , vol . 2009 , Article ID 275287 ( 7 pages ) . Steidler , L . at al . ( Jul . 1 , 2003 ) “ Biological containment of geneti cally modified Lactococcus lactis for intestinal delivery of human interleukin 10 " Nat Biotechnol , 21 ( 7 ) : 785 - 789 . Strauch . K . L . et al . ( Feb . 1985 ) “ Oxygen Regulation in Salmonella typhimurium ” J Baceriol , 161 ( 2 ) : 673 - 680 . Unden , G . et al . ( 2002 ) “ Control of FNR Function of Escherichia coli by O2 and Reducing Conditions ” J Mol Microbiol Biotechnol , 4 ( 3 ) 263 - 268 .
* cited by examiner
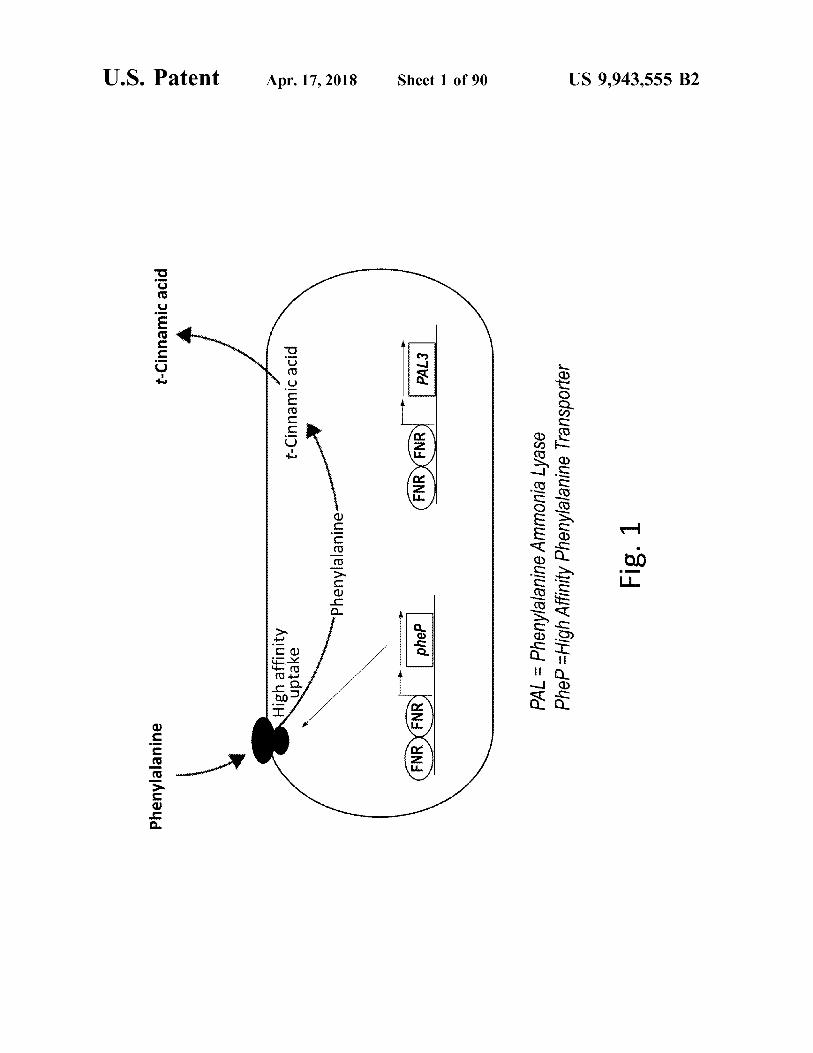
Phenylalanine
t - Cinnamic acid
atent
High affinity uptake
t - Cinnamic acid
Apr . 17 , 2018
www . phenylalanine
FNR FNR ) | Pher
( FNR XFNR ) | PAL3
Sheet 1 of 90
PAL = Phenylalanine Ammonia Lyase PheP = High Affinity Phenylalanine Transporter Fig . 1
US 9 , 943 , 555 B2
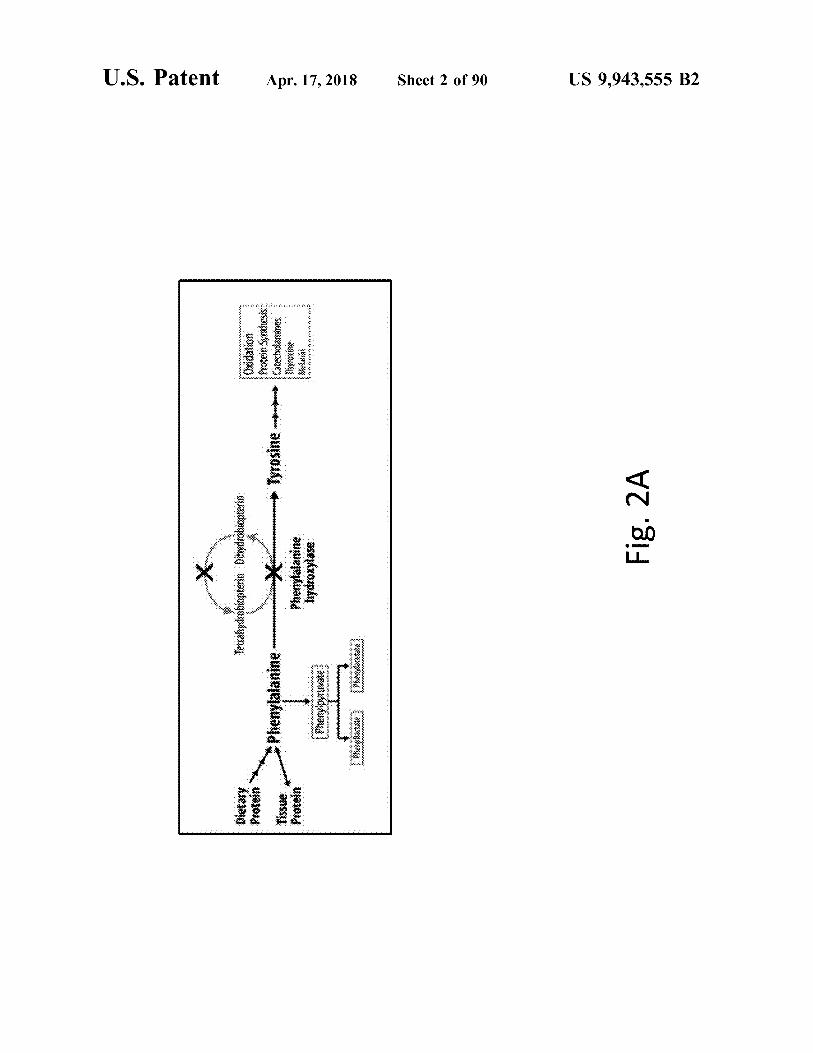
US 9 , 943 , 555 B2
V2 : 8 ! 4
Sheet 2 of 90
w
www .
Apr . 17 , 2018
BERELLO LUPUS
* * * * * * * * * * * * * * * * * *
RAK
BejkxOSPX sujuejekian . X
ujajoxd * mil
au soit
Jujuejejávald
SURP * * ) Elepo
dajold Kilimo
antspokeo UBICCITUSPÁLENI
atent
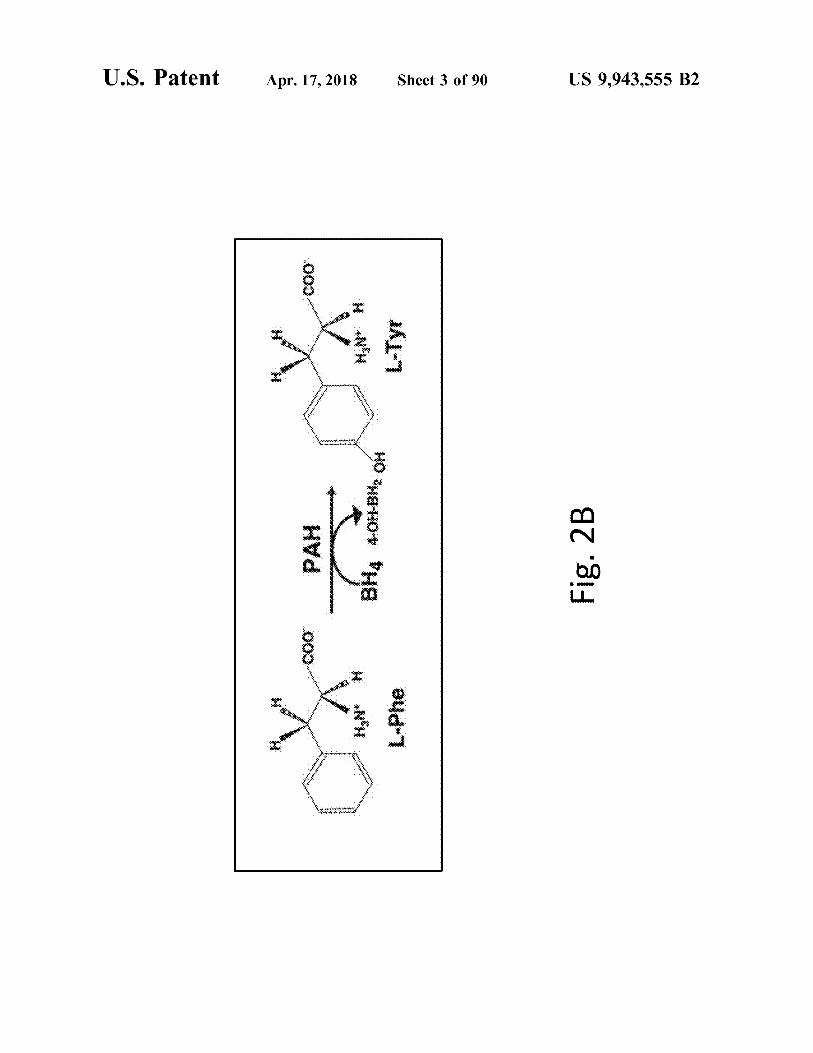
atent
Plantation in
coo
PAH
Apr . 17 , 2018
potrete perare
preferens
?????????????????? ; a
W
MN * H
ontre
HxN * H
L - Phe
BHA 4 - 01 - BH , OH
. . . .
L - Tyr
Sheet 3 of 90
Fig . 2B
US 9 , 943 , 555 B2
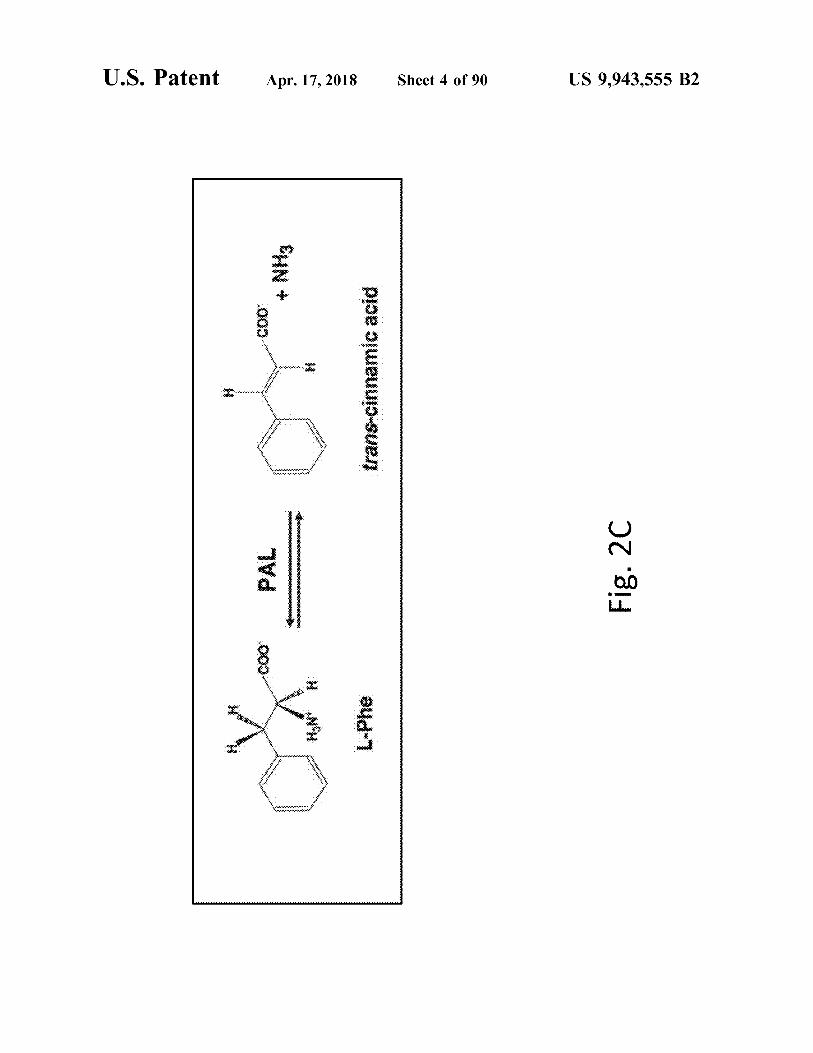
US 9 , 943 , 555 B2
O7813
Sheet 4 of 90
pioe oueuujo - sues
a4d - 1
Apr . 17 , 2018
N NH
- - -
www V
* p * * * * * * * * *
* *
* * * *
opi .
* *
* * * * *
atent
atent
???????????? : ???????
Apr . 17 , 2018
?? ????? ?????
?? ?? ???????
???? ?????????
?
??? NE Phenylalanine
Phenylpyruvate
Sheet 5 of 90
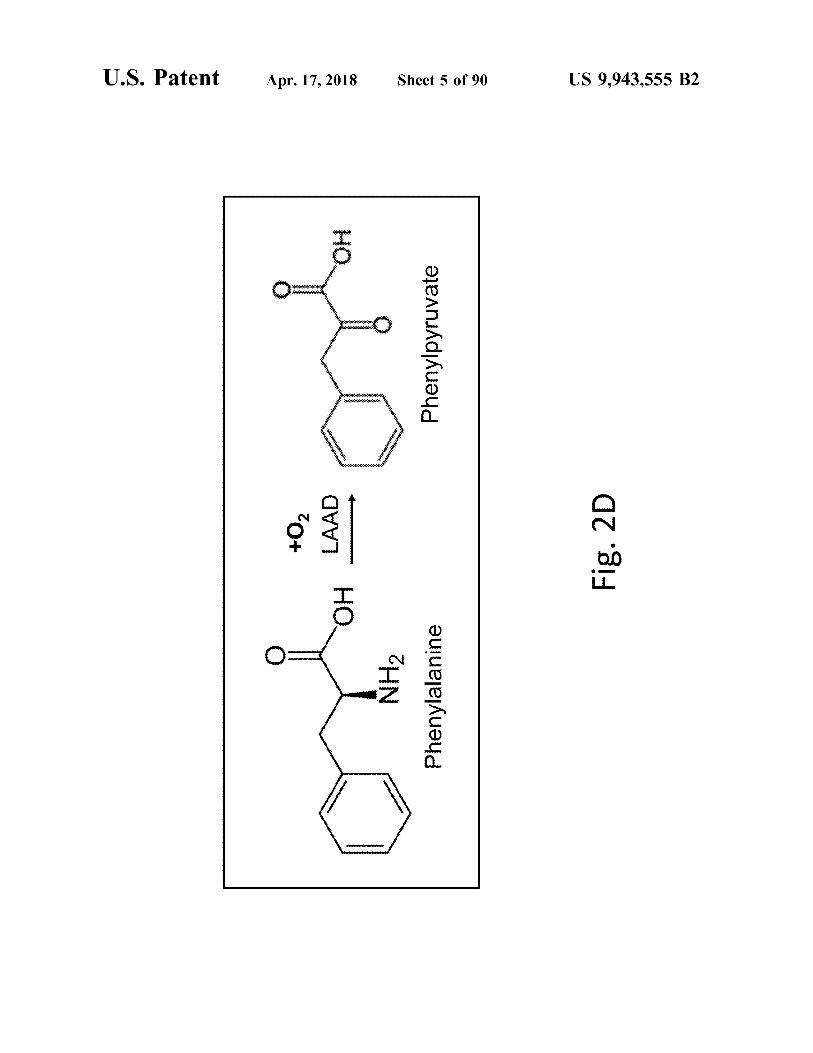
Fig . 2D
US 9 , 943 , 555 B2
US 9 , 943 , 555 B2
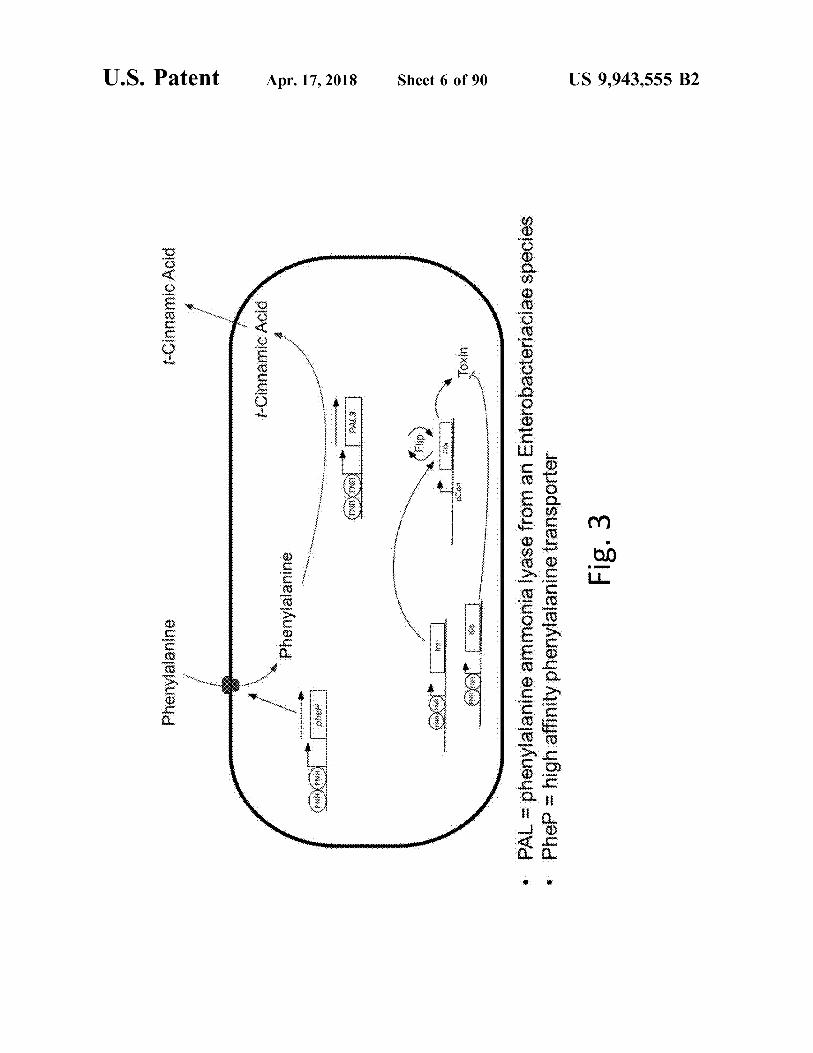
Fig . 3
• Phep = high affinity phenylalanine transporter
• PAL = phenylalanine ammonia lyase from an Enterobacteriaciae species
wwwwww .
minnes
ruimtes . * * * * *
* * - - - *
www .
Toxin
123 - - -
- 1
O200 D
0 . 0000C
* * * *
*
* * * * * * * * *
Sheet 6 of 90
. ???????????????????????????????
rispondentes de
* *
*
BB 3
Daisissa wiririiniinin * *
* * *
vari
* * *
Apr . 17 , 2018
* * * varivoorras
190 . 00 star
-
Phenylalanine .
- Cinnamic Acid
atent
8 - Cinnamic Acid
???????? { A & ????
Plasma Membrane
Outer Membrane
atent Apr . 17 , 2018
arabinose
LAADI
arac arac Para
| ParaBAD
Sheet 7 of 90
Phenylalanine —
Phenylpyruvate -
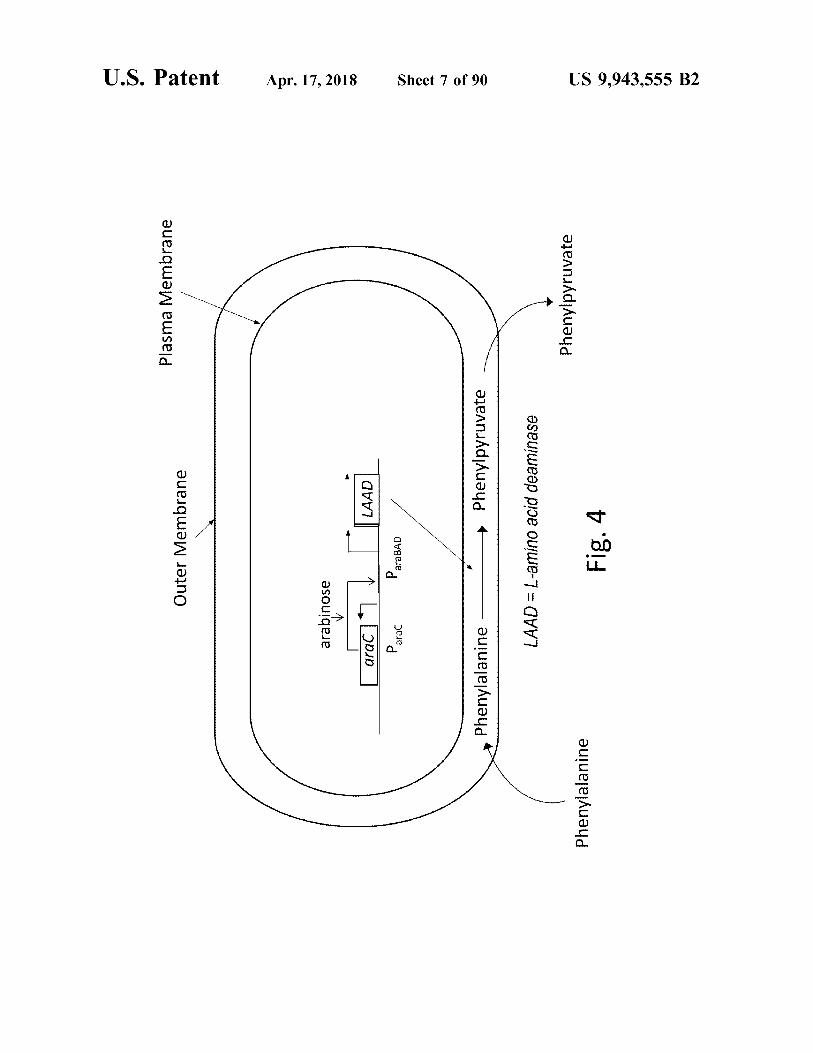
LAAD = L - amino acid deaminase
Phenylpyruvate
Phenylalanine
Fig . 4
US 9 , 943 , 555 B2
Phenylalanine
t - cinnamic acid
Plasma Membrane Outer Membrane
atent
High affinity uptake
t - cinnamic acid
Apr . 17 , 2018
Phenylalanine FNR XFNR
phep
FNRY FNR FNR
PAL3 PAL37
arabinose arabinose | aractv | LAAD Parac Parabad arac
LAAD
Sheet 8 of 90
Phenylalanine
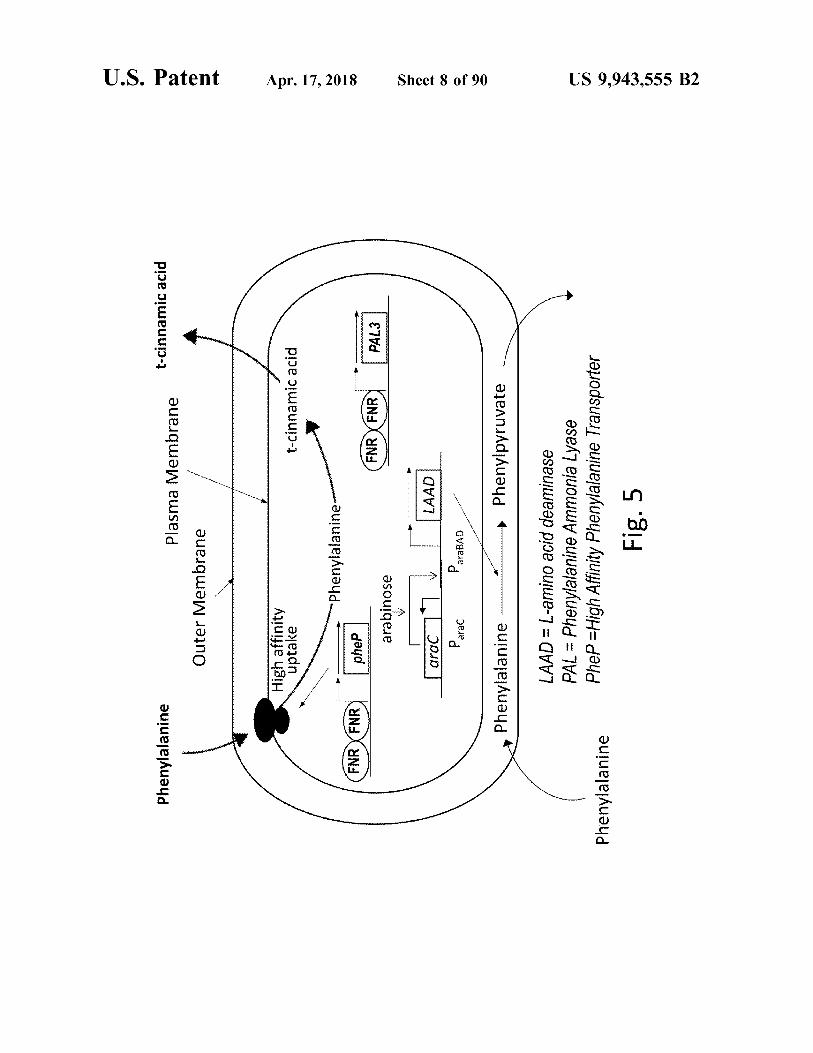
Phenylpyruvate LAAD = L - amino acid deaminase PAL = Phenylalanine Ammonia Lyase Phep = High Affinity Phenylalanine Transporter
Fig . 5
Phenylalanine
US 9 , 943 , 555 B2
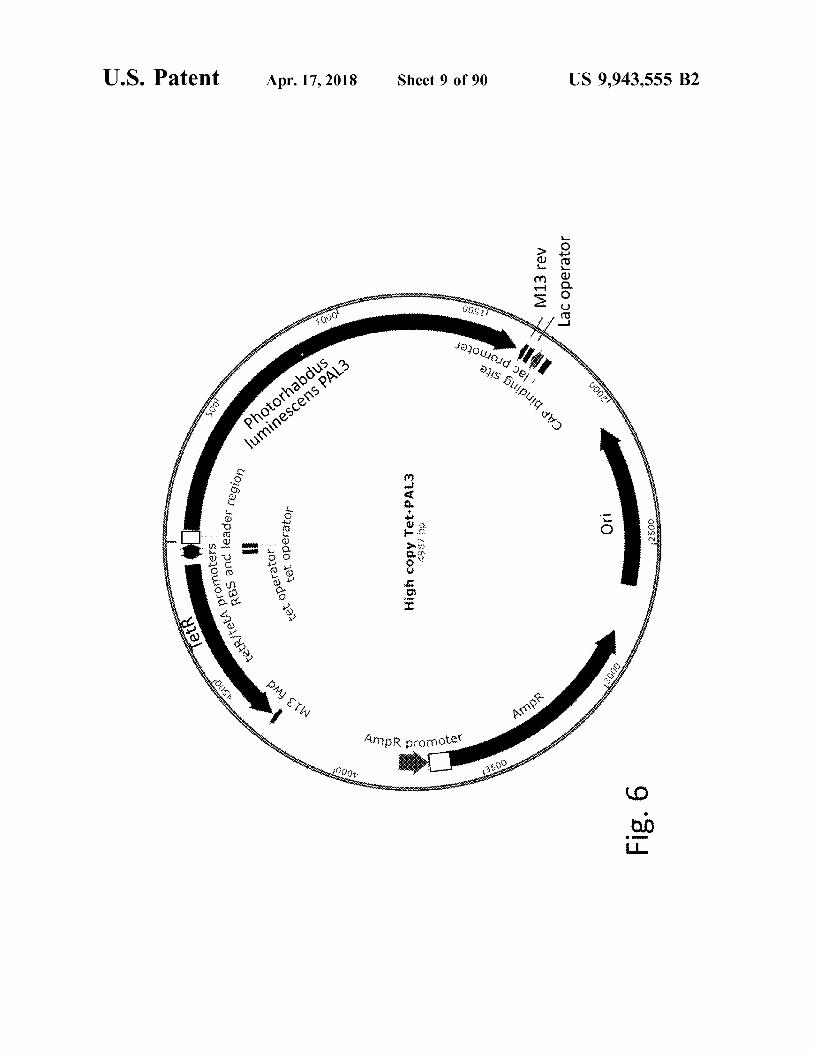
Tetr
atent
500
tetr / tetan Sjejowalay
500
es and leader regis tet operator tet operator
M13 fwd
luminescens PAL3 Photorhabdus
Apr . 17 , 2018
1000 ?
4000
AmpR promoter
High copy Tet - PAL3 09 ( 865
Sheet 9 of 90
within of 3500
lac promoter
Ampo
CAP binding Silk 011
XM13 rev Lac operator
000 ? 1
Ori
13000
Fig . 6
US 9 , 943 , 555 B2
2500
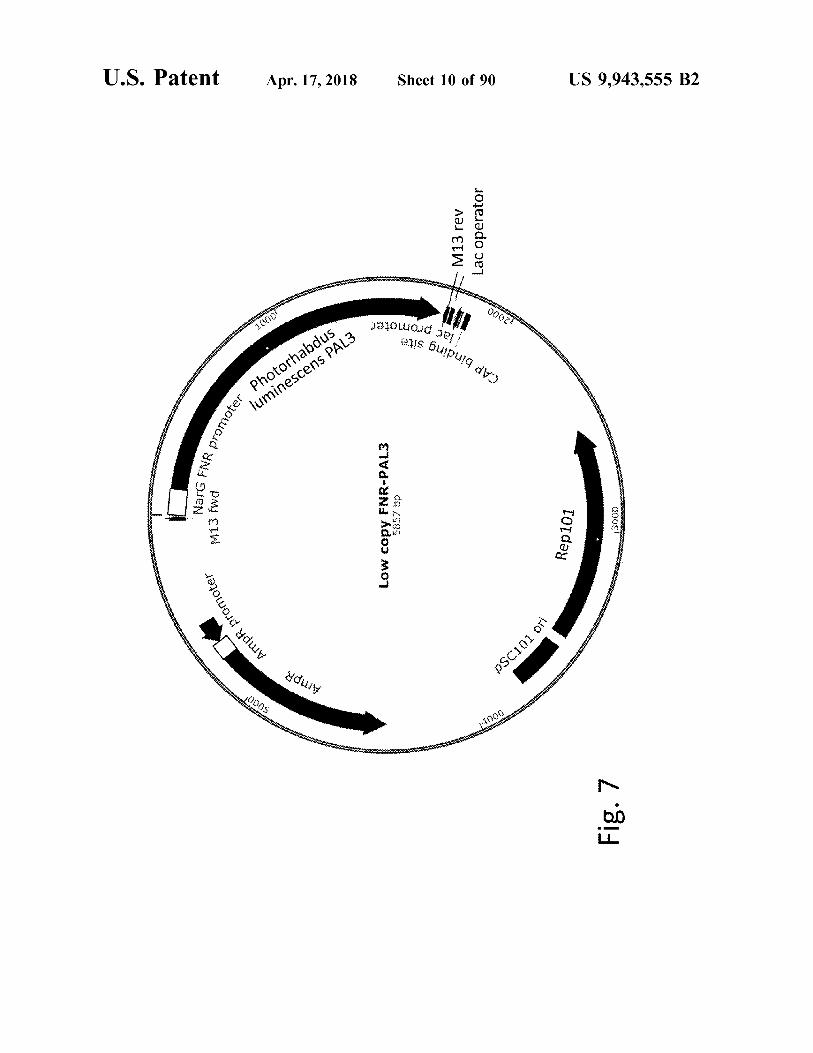
atent Apr . 17 , 2018 Apr 17 , 2018 Sheet 10 of 90 US 9 , 943 , 555 B2
M13 rev Lac operator 0007 1000 meter lac pro
po binding site CAP binal ter Photorhabdus luminescens PAL3 we
NarG FNR prome M13 fwd Low copy FNR - PALS dQ2585 Rep101 00091
promoter Ampk promo DSC101 Ori 2
ydurv e tradem 50007 1000
Fig . 7
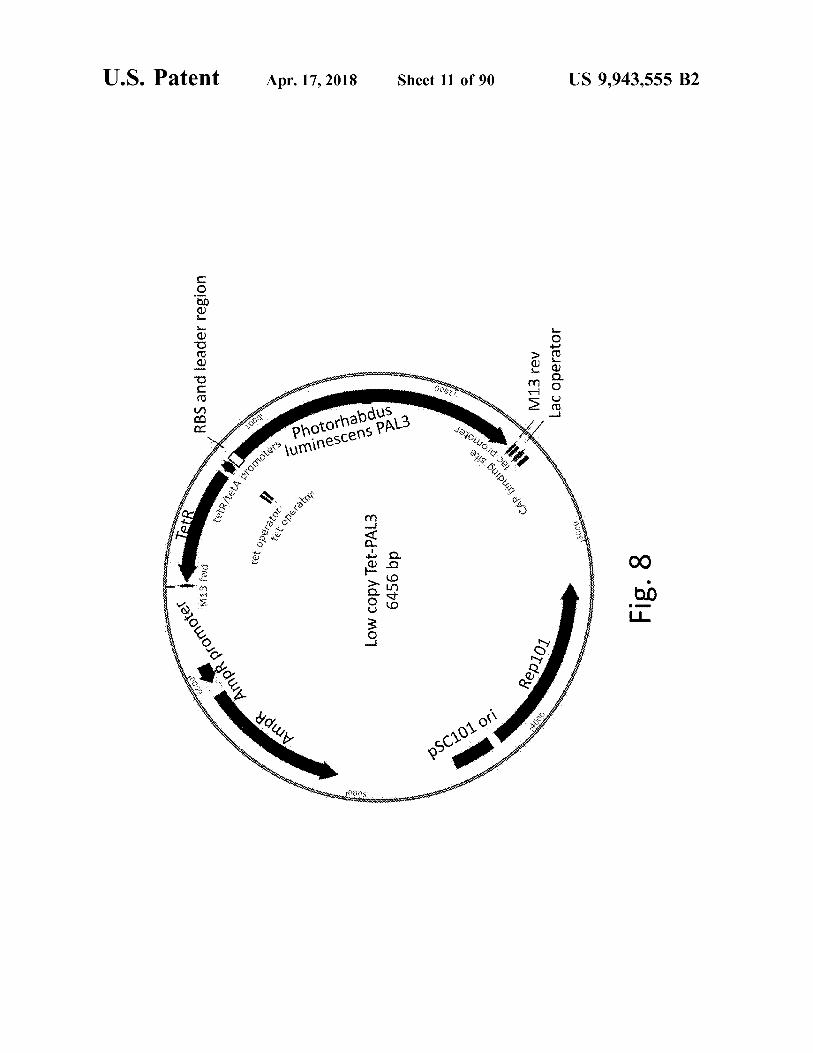
1000e AmpR promoter
tR
atent
RBS and leader region
M13 fwa
. . .
tet
1000
AmpR
Petriteta promoter far tet operator
luminescens PAL3 Photorhabdus
Apr . 17 , 2018
soos
Low copy Tet - PAL3 6456 bp
2000 .
PSC101 ori
Sheet 11 of 90
CAP Dinding sites 0 promoter 1011
060 * . .
Rep101
M13 rev Lac operator
Fig . 8
US 9 , 943 , 555 B2
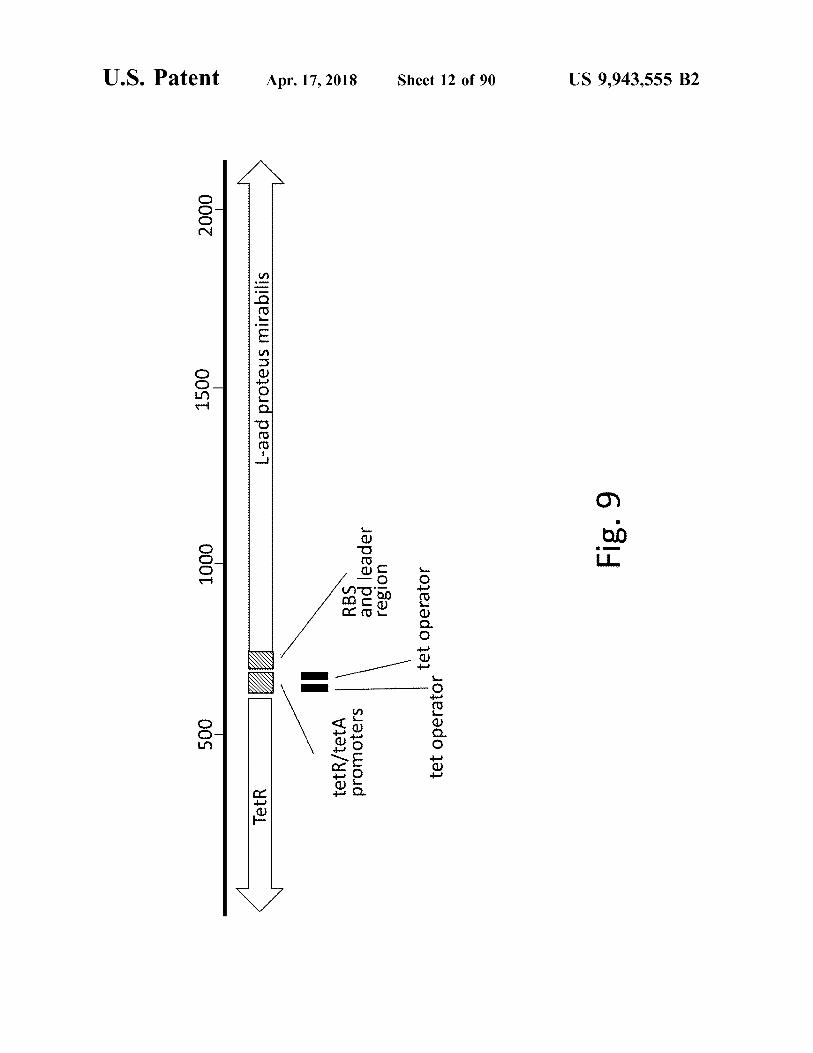
500 500
1000 1000
1500 1500
atent
2000
TetR TetR
L - aad proteus mirabilis
Apr . 17 , 2018
tetR / teta promoters tetrihtoters
RBS |
and leader region
tet operator
tet operator
Sheet 12 of 90
Fig . 9
US 9 , 943 , 555 B2
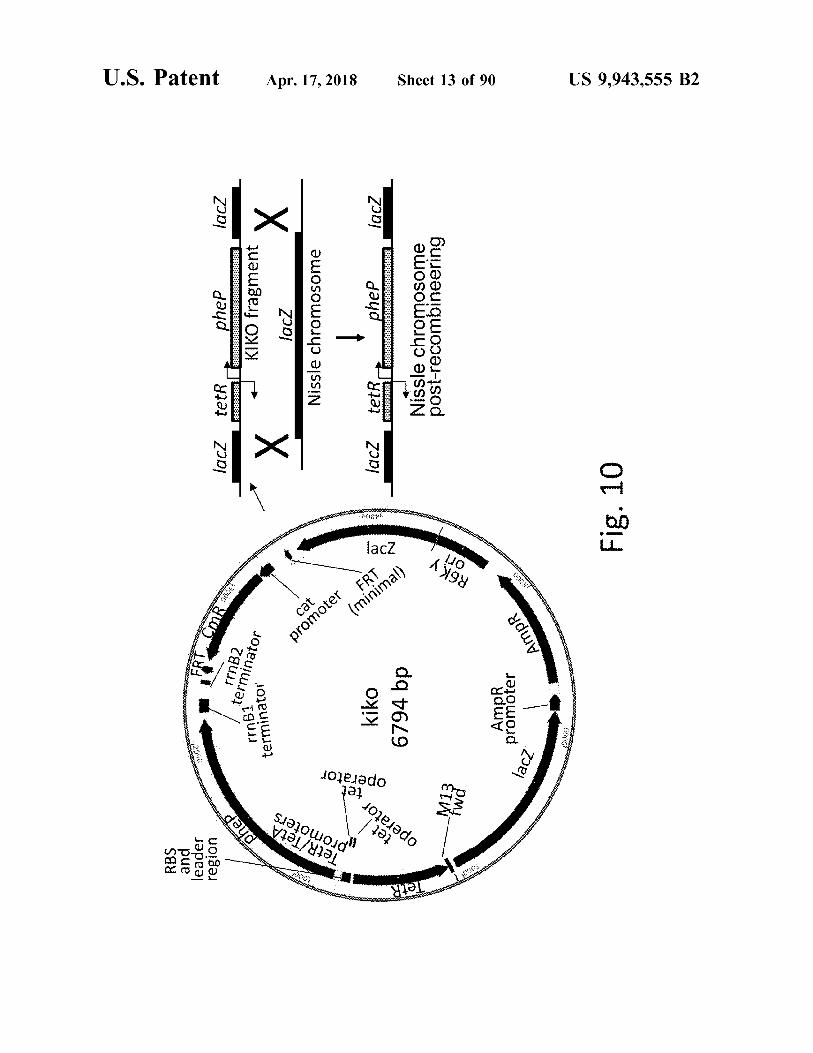
RBS and leader region
2007 ?
atent
* *
FRT CMR
phep
lacz
DONDE * *
le
rrnB1 terminator rrnB2 terminator
“
X
lacz lacz tetRo phep + KIKO fragment lacz Nissle chromosome Ix !
cat
TetR / TETA promoters
Apr . 17 , 2018
operator
promoter FRT ( minimal )
lacz
tetR
tet operator ret
lacz
lacz
kiko 6794 bp
111
TetR
Nissle chromosome post - recombineering
M13 fwd
ROKY
Sheet 13 of 90
AmpR promoter AmpR
0009
lacz 60
Fig . 10
US 9 , 943 , 555 B2
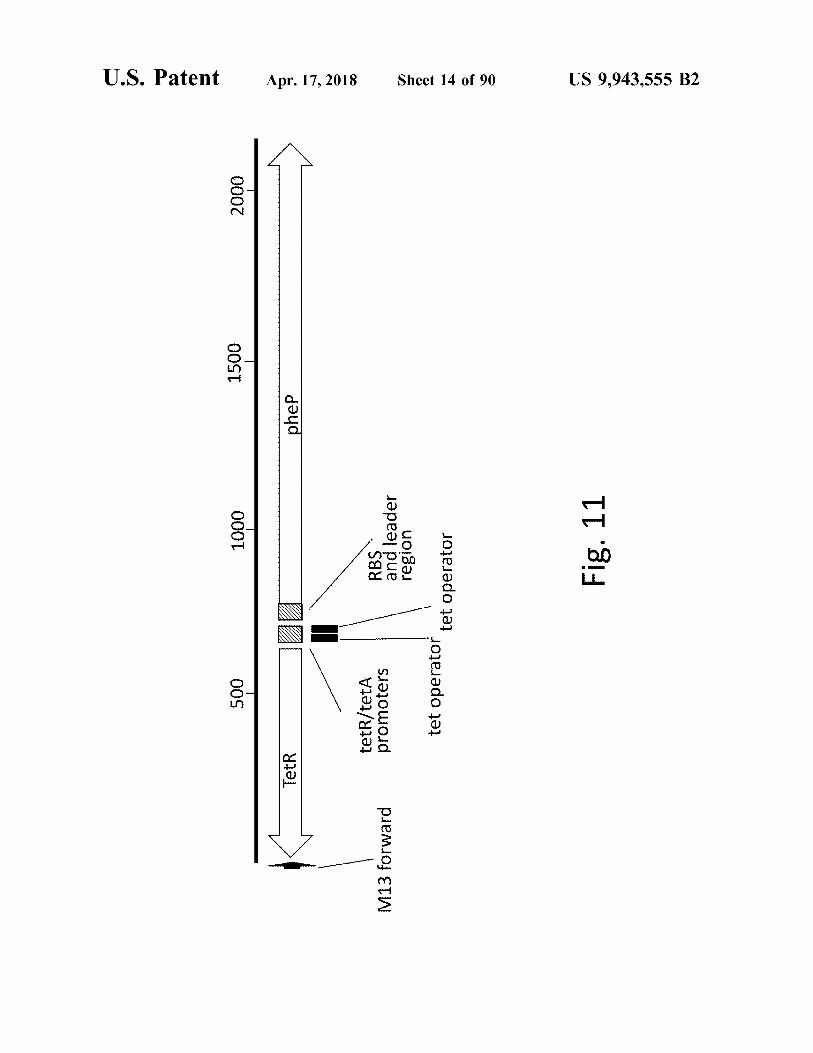
atent
500 500
1000 1000
1500 1500
2000 2000
TetR
phep
Apr . 17 , 2018
M13 forward
tetR / teta promoters RBS and leader region
tet operator te
' tet operator
Sheet 14 of 90
Fig . 11
US 9 , 943 , 555 B2
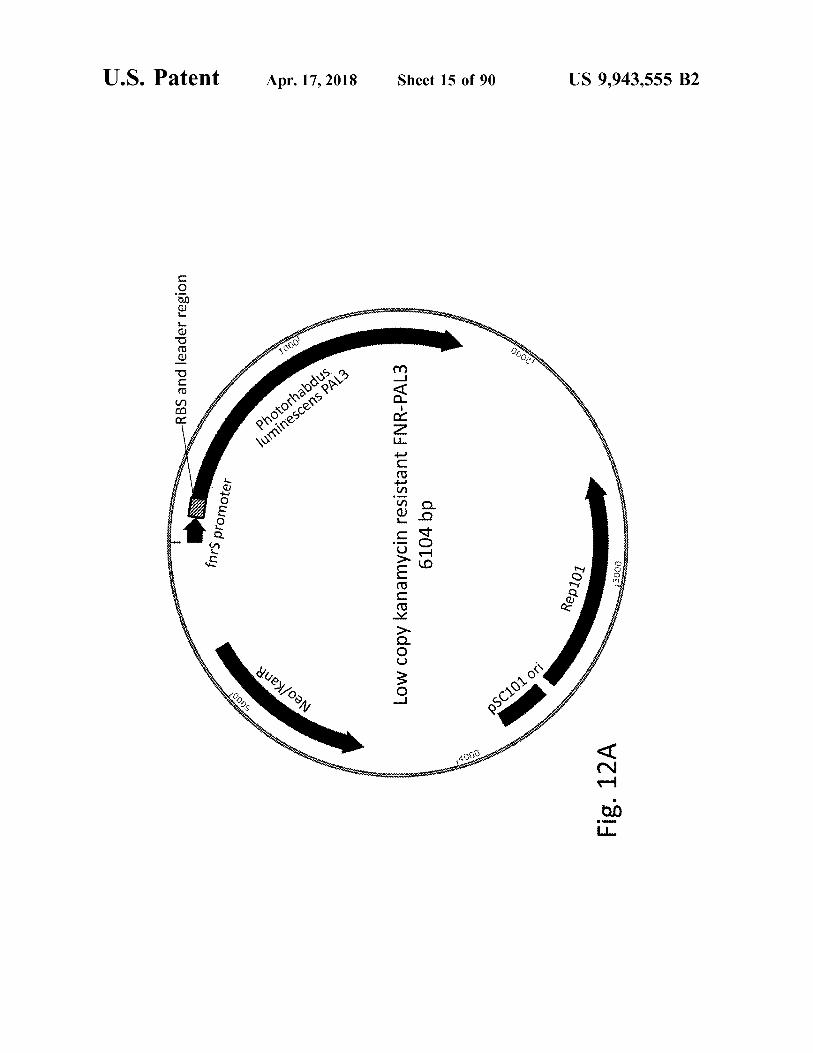
atent Apr . 17 , 2018 Sheet 15 of 90 US 9 , 943 , 555 B2
RBS and leader region 1000 ) 0002
Photorhabdus luminescens PAL3 S
fnrs promoter Low copy kanamycin resistant FNR - PAL3 6104 bp 3000 Rep101
Neo / KanR 5000 PSC101 ori
met het 4000
Fig . 12A
500
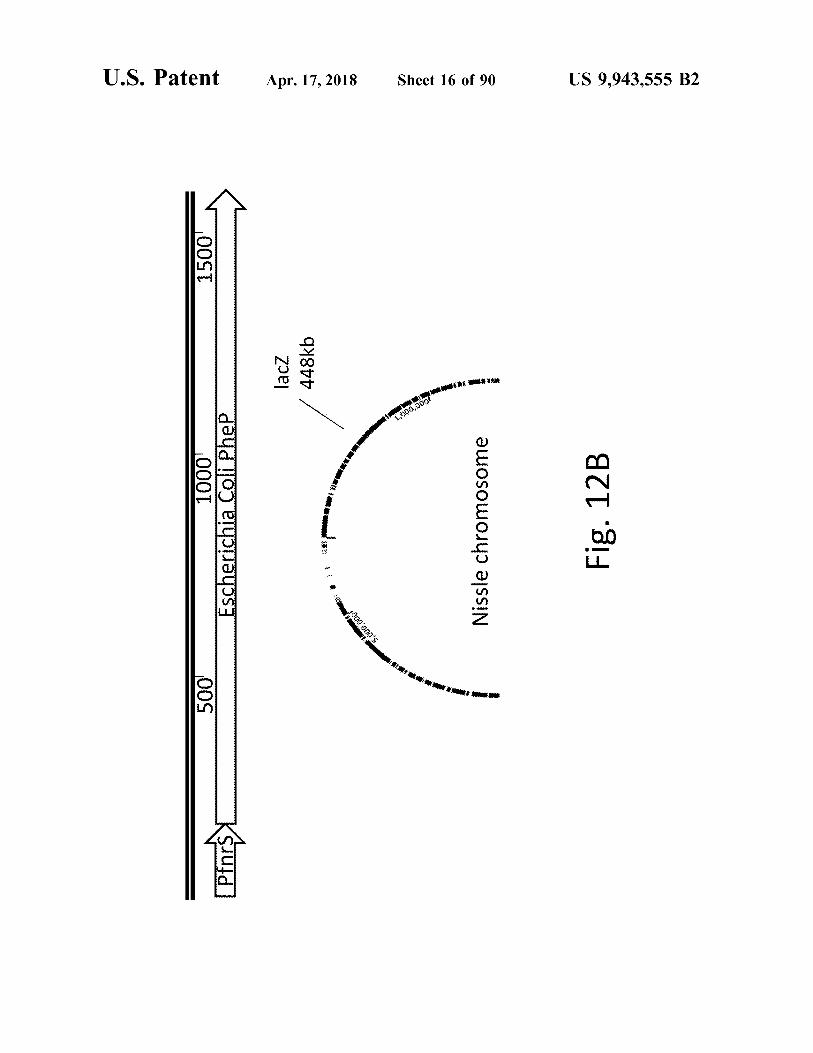
1500
atent
Pins Pin by
1000 Escherichia coli Phep Escherichia coli Phep
?
– lacz 448kb
Apr . 17 , 2018 Sheet 16 of 90
Nissle chromosome
sim
u
Fig . 12B
US 9 , 943 , 555 B2
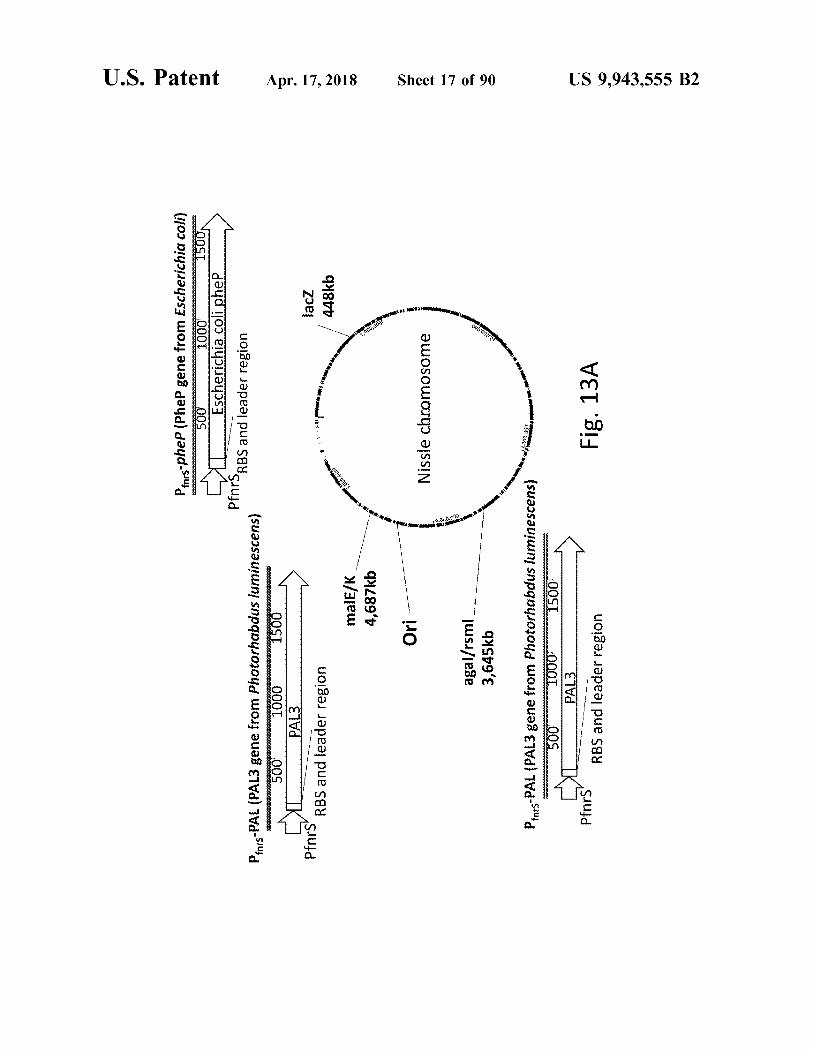
atent
Pas - pheP ( Phel gene from Escherichia coli )
500 500
1000 1000 1500K
Escherichia coli phep
PfnrSRBS and leader region
Pfars - PAL ( PAL3 gene from Photorhabdus luminescens ) 500 1000 1500
L PAL3
PAL3
Pfnrs !
lacz
RBS and leader region
448kb
malE / K 4 , 687kb
Apr . 17 , 2018
. : : .
02
Ori - -
Nissle chromosome
Sheet 17 of 90
agal / rsmi 3 , 645kb
http : .
500
mana
1000 Pfars - PAL ( PAL3 gene from Photorhabdus luminescens ) * * *
1500
TT PAL3 RBS and leader region
Fig . 13A
PAL3
Pfnrs
US 9 , 943 , 555 B2
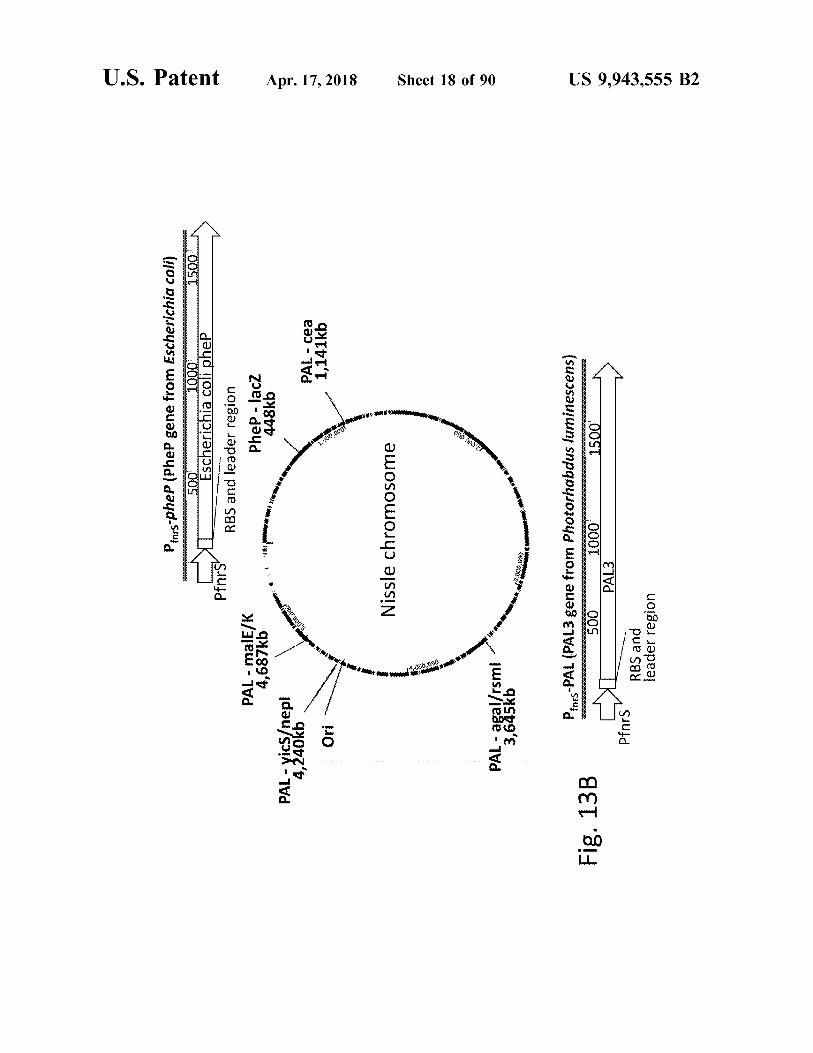
Pos - pheP ( Phee gene from Escherichia coli )
500
1000
15001
Escherichia coli pheP RBS and leader region
Phep - lacz 448kb
atent
Pfnrs PAL - malE / K 4 , 687kb
PAL - yics / nepl 4 , 240kb
Oria
, 000 ,
5
PAL - cea 1 , 141kb
Apr . 17 , 2018
1
Nissle chromosome
Sheet 18 of 90
PAL - agal / rsmi 3 , 645kb
Fig . 13B
1000
1500
Penrs - PAL ( PAL3 gene from Photorhabdus luminescens ) 500
NT
PAL3 PAL3
Pfnrs
RBS and leader region
US 9 , 943 , 555 B2
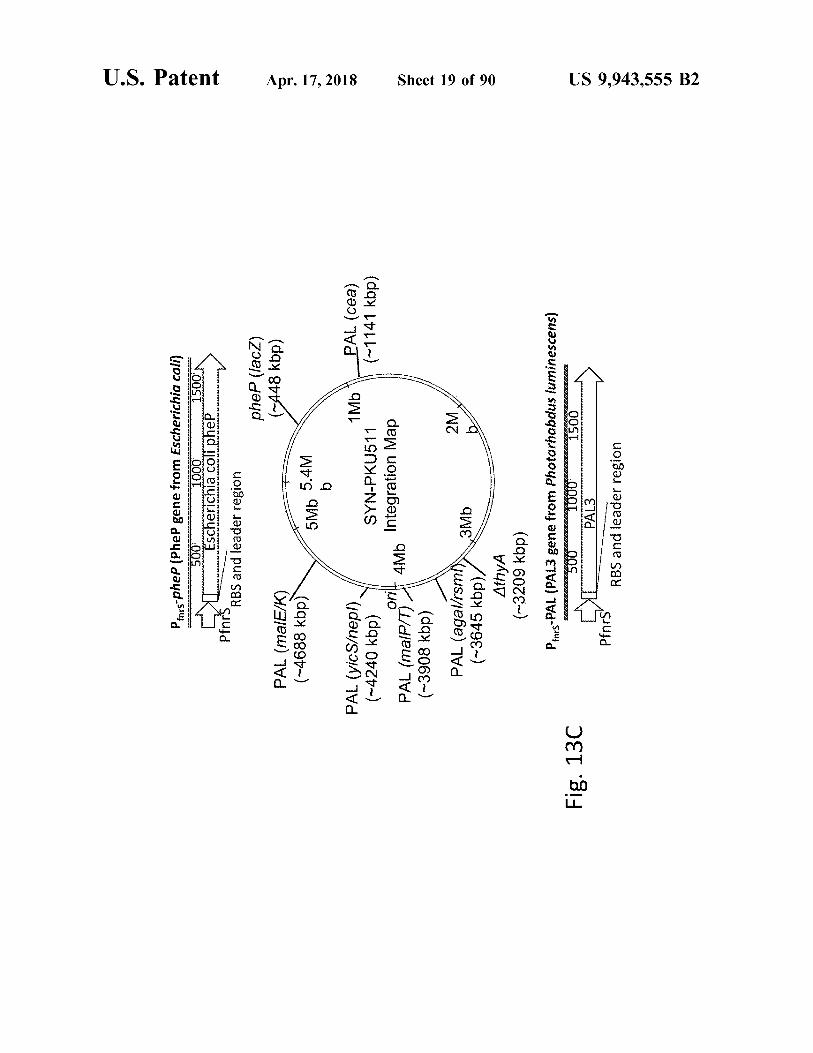
U . S . Patent atent
Ptnrs
Pas - phep ( Phee gene from Escherichia coli ) 500m 1000
1500N
DO Escherichia coli phep RBS and leader region
pheP ( lacZ )
PAL ( malE / K )
( - 448 kbp )
( - 4688 kbp )
5Mb 5 . 4M
Apr . 17 , 2018
PAL ( yicS / n
1 Mb PAL ( cea )
( - 4240 kbp ) y SYN - PKU511 ( ~ 1141 kbp ) orilt 4Mb Integration Map
PAL ( malP / T ) 4Mb ( ~ 3908 kbp )
PAL ( agal / rsm 3Mb ( ~ 3645 kbp ) /
AthyA ( - 3209 kbp )
Pfines - PAL ( PAL3 gene from Photorhabdus luminescens ) 500 1000 1500
TTPAL3 Pfnrs ! RBS and leader region
Sheet 19 of 90
Fig . 130
PAL3
US 9 , 943 , 555 B2
atent
500
1000 000
1500 1500
2000
500
???
???
Apr . 17 , 2018
arac arac
]
[
Laad prote L - aad proteus mirabilis
RBS and leader region
Sheet 20 of 90
Fig . 14
US 9 , 943 , 555 B2
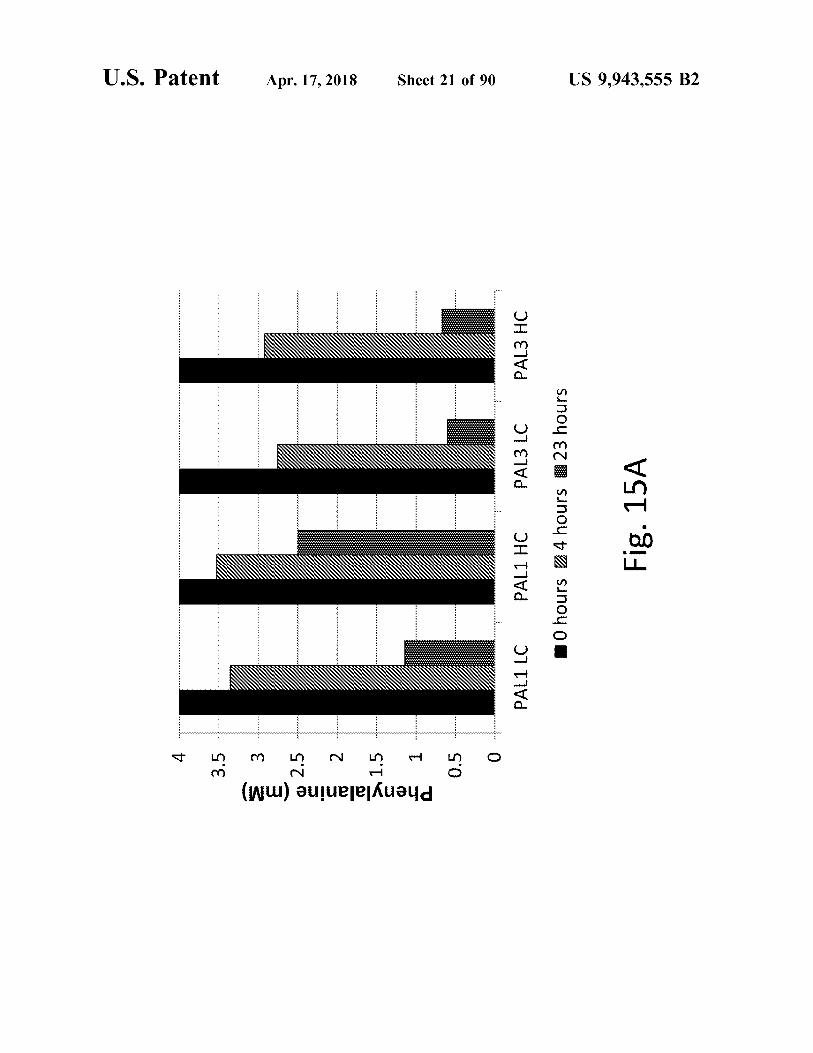
o & Phenylalanine ( mM )
B L M ? ? ?
PAL1 LC ??????????????????????????????????????????????????????????????????????? ????????????????????????????????????????? ????????????????????????????????????
????????????????????????????????????????? ?
PAL1 HC ??????????????????????????? ???????????????? ????????????
??? ???????????????? ????????????? ??????????????????
Fig . 15A j0 hours Z24 hours 23 hours
? ? ? ???????????????????????????? ? ???? PAL3 LC ??????????????????????????????????
??????????? ??????????????????????????? ?????????????????????????????????
PAL3 HC ?????????????????????
z8 SSSEt66 Sn 06 J0 Iz 100?S 8I07 CLI adv 1???
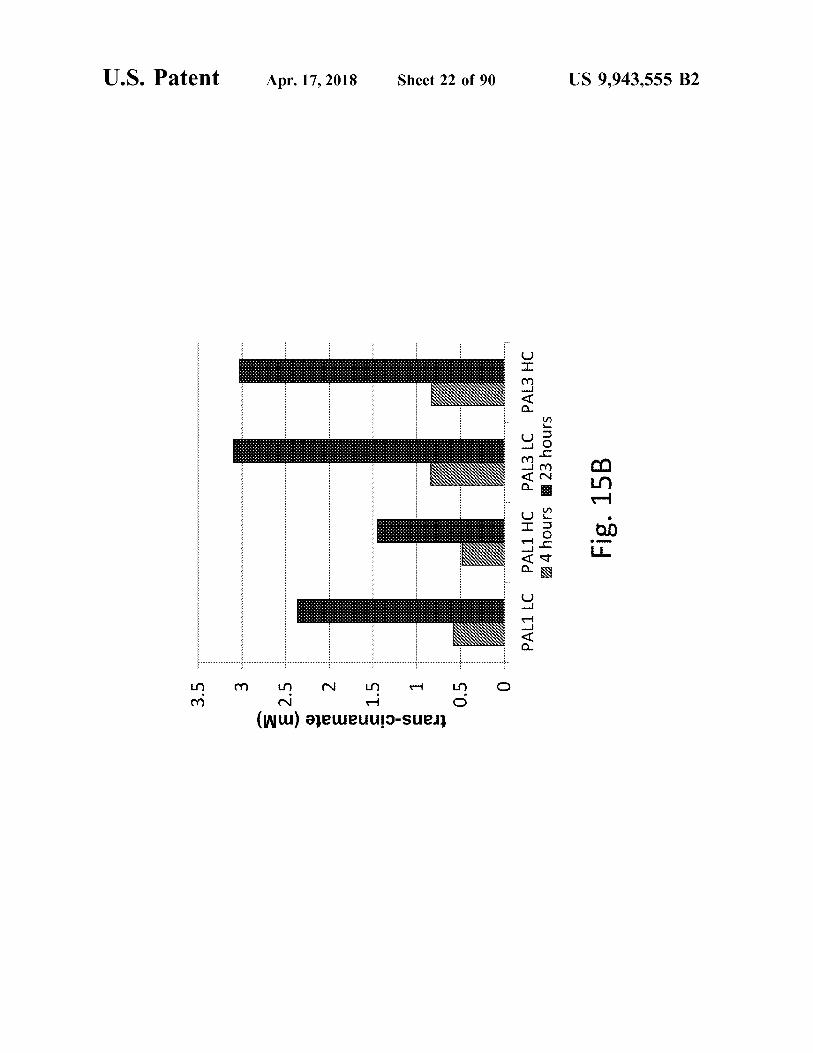
?
atent
? ?
Apr . 17 , 2018 wawcow -
trans - cinnamate ( mm ) ?
- - - - - - - - - - - - - - - - -
? ??
ZZLI2222222222
Sheet 22 of 90
? ?
PAL1 L
PAL1 HC PAL3 LC PAL3 HC
Z 4 hours 23 hours Fig . 15B
US 9 , 943 , 555 B2
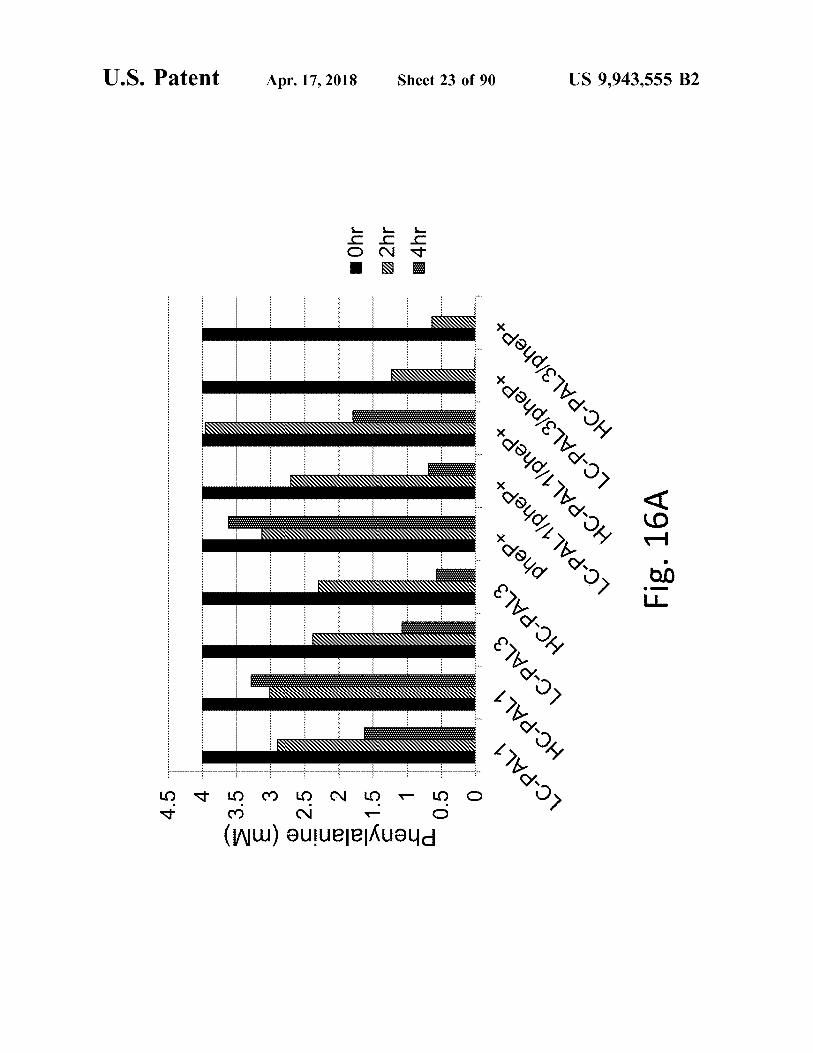
Phenylalanine ( mm ) å vänä ? en 2222222222211111111111111LLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLL ????????????????????????????
???????????????????????? ZZZZZZZZZZZZZZ22222222222
LC - PAL1 . HC - PAL1 LC - PAL3 HC - PAL3
pheP + HC - PAL1 / pheP + LC - PAL1 / pheP + .
| HC - PAL3 / pheP + | LC - PAL3 / pheP +
? 22222222222222222222LILLLLLLLLLLL L LLL ? ?????????????????????
22222222222222 L LLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLL
Fig . 16A 222 LLLLLLLLLLLLLLLLLLLL L L
LLLLLLLLLLLLLL22222222222222222222222ZZZZZZ ?????????????????????????????
212 LLLLLLLL LLLLLLLLLLL LLLLLLLLLL
? ???????????????????????????
12112212112112112112112212112112112112112112112112212112112112112212 LLLLLLLLLLLLLLLLLLLL LLLLLLLLL2222222222222222222222222222222222LLLLL LLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLL
??? ?????????????????????????????????????????
ZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZ 1 LLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLL ZUZAZIZZIUZULU
2222222ZZZZZ LLLLLLLLLLLLLLLLLLLL 12 ??
14hr 22hr Ohr
za SSS?€ + 6 * 6 SN 06 JO ET J??US 810Z ' LI ady jual
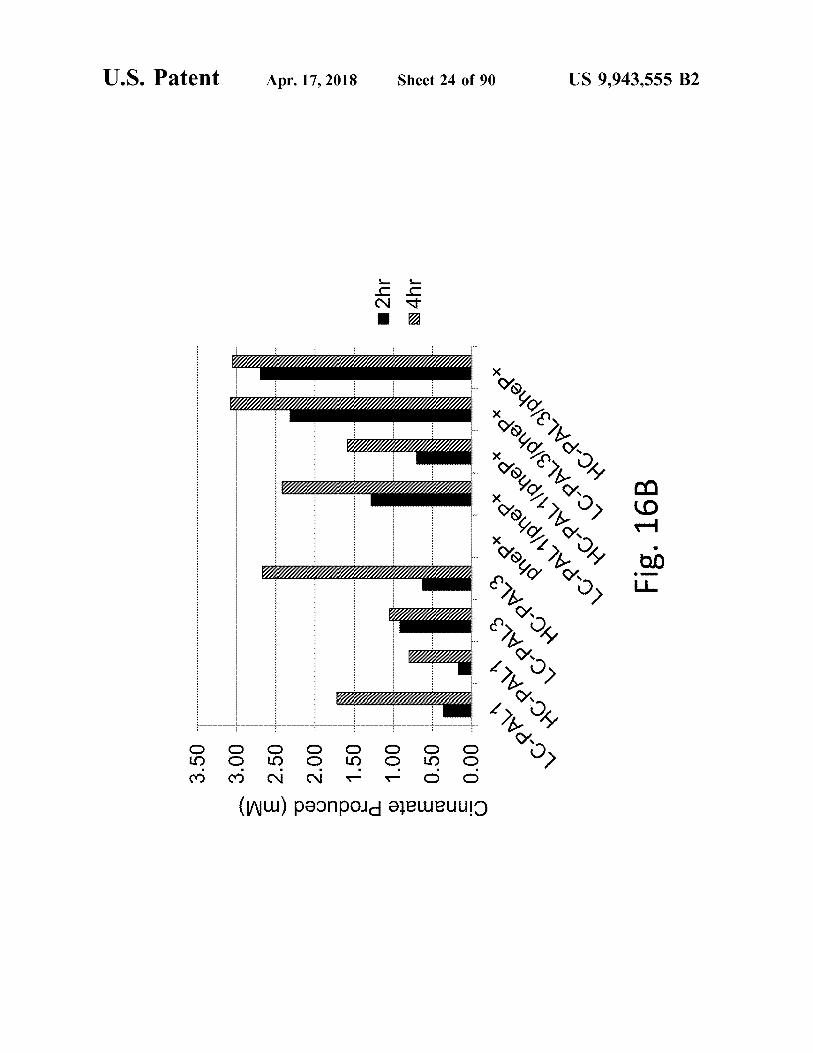
atent Apr . 17 , 2018
Cinnamate Produced ( MM ) a gaña o
- - -
- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - - - - - - - - - - - - -
2hr N 4hr
LC - PAL1 . HC - PAL1 .
HC - PAL1 / pheP + "
LC - PAL3 HC - PAL3
pheP + LC - PAL1 / pheP +
| LC - PAL3 / pheP + HC - PAL3 / pheP +
Sheet 24 of 90
?
Fig . 16B
US 9 , 943 , 555 B2
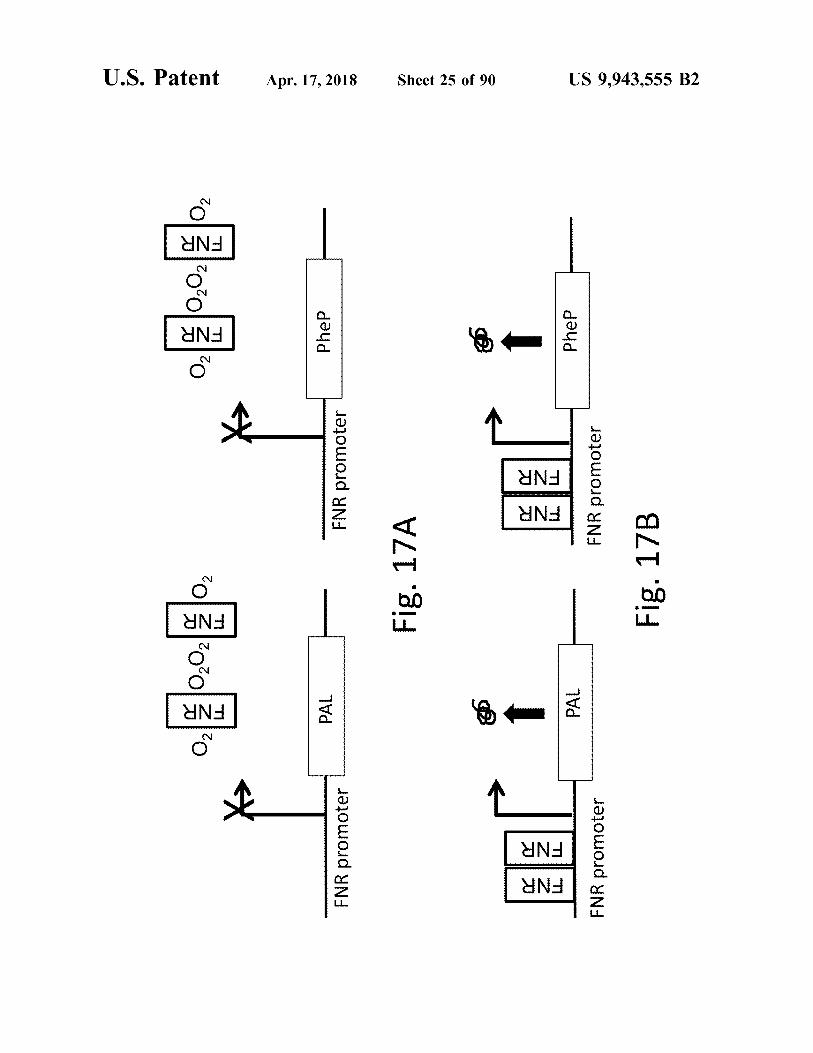
ENR
FNR
FNR
FNR
atent 1
* ° ( E ) 5•2• Im
x 8 . 01 I have
Apr . 17 , 2018
PAL PAL
Phen
FNR prom FNR promoter
FNR promoter
Fig . 17A Fig . 17A
Sheet 25 of 90 Sheet 25 at oo
FNR
FNR
ENR
ENR
PAL
PheP PheP
-
FNR promoter
FNR promoter Fig . 17B
US 9 , 943 , 555 B2
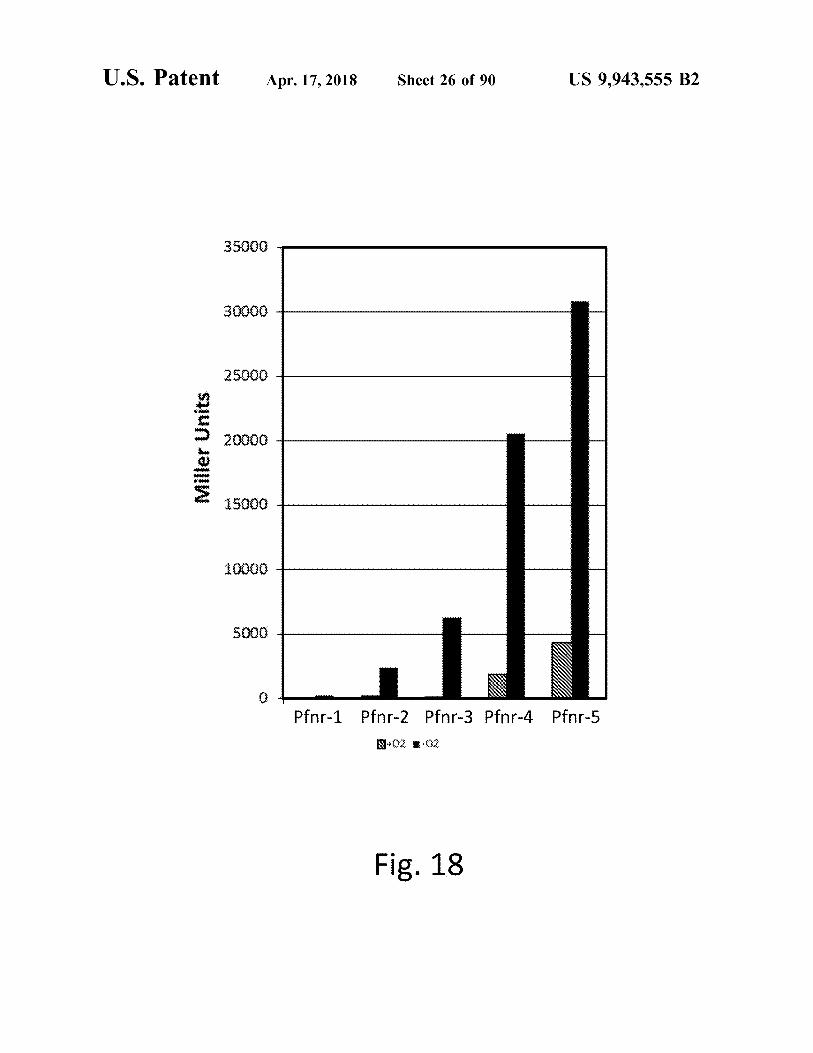
atent Apr . 17 , 2018 Sheet 26 of 90 US 9 , 943 , 555 B2
35000
30000
25000
20000 Miller Units 11111111111111111111111111 15000
10000 1111111111111 111111111111111
Pfnr - 1 Pfnr - 2 Pfnr - 3 Pfnr - 4 Pfnr - 5 N 0 * - 02
Fig . 18
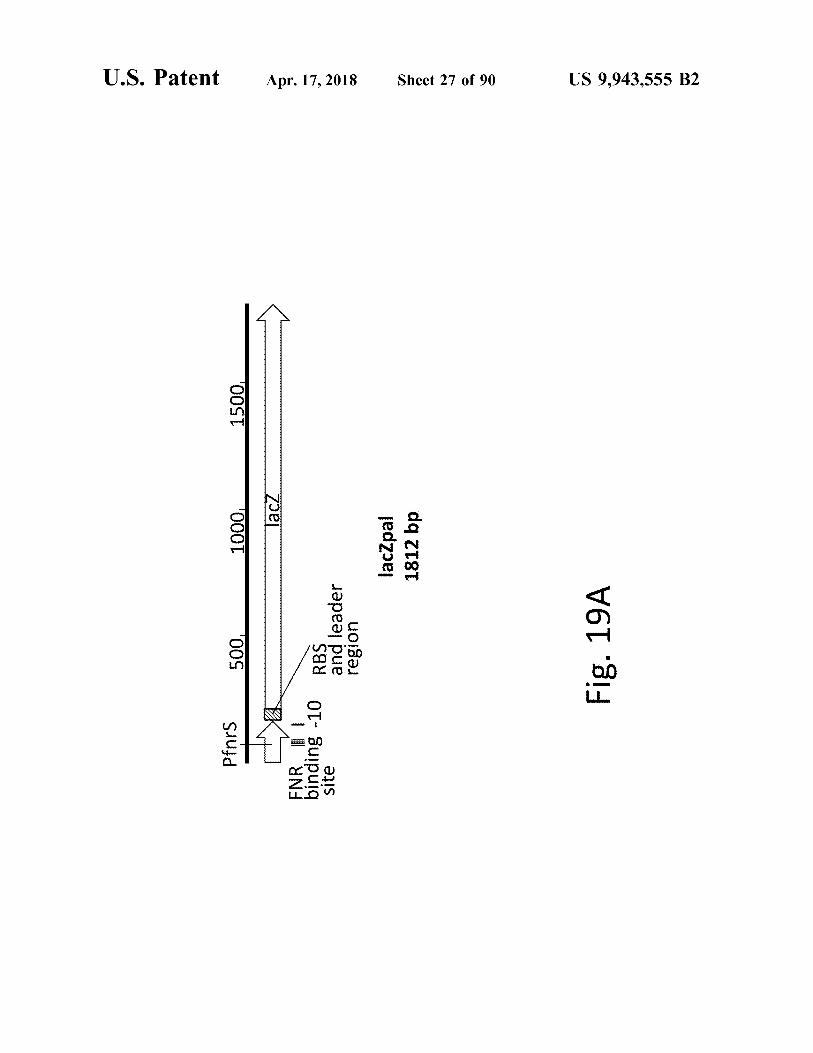
atent
Suufd
500
1500 .
1000 lacz
Ha ENR 1 binding - 10 site
Apr . 17 , 2018
RBS and leader region
lacZpal 1812 bp
Sheet 27 of 90
Fig . 19A
US 9 , 943 , 555 B2
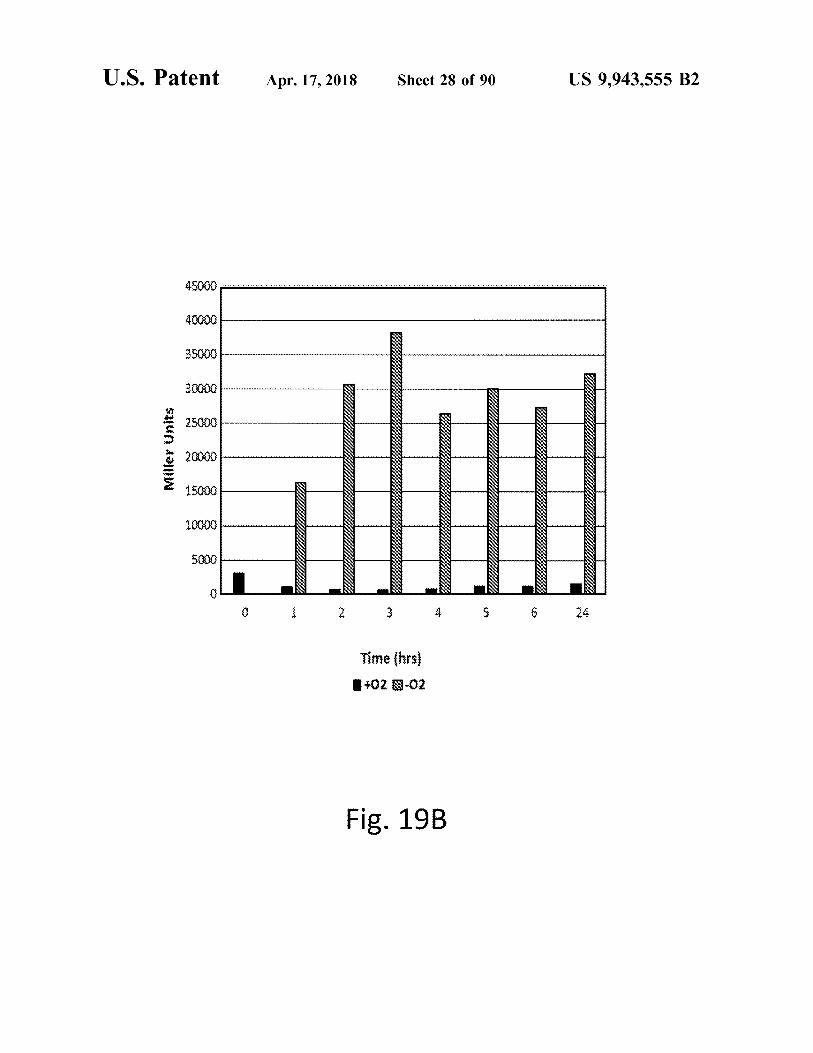
atent Apr . 17 , 2018 Sheet 28 of 90 US 9 , 943 , 555 B2
45000 .
40000 IZ
35000
30000 ZY
TI
27
12212 Miller Units Jl IYYYYLILLLLLLLLLLLLL ULUUUUUUU
4 227
22
21 112
1 . 2222
10000 212
1 . 2222 222
5000 12 . 1
12 12227 2212
0 1 2 3 4 5 6 24
Time ( hrs ) 1 + 02 9 - 02
Fig . 19B
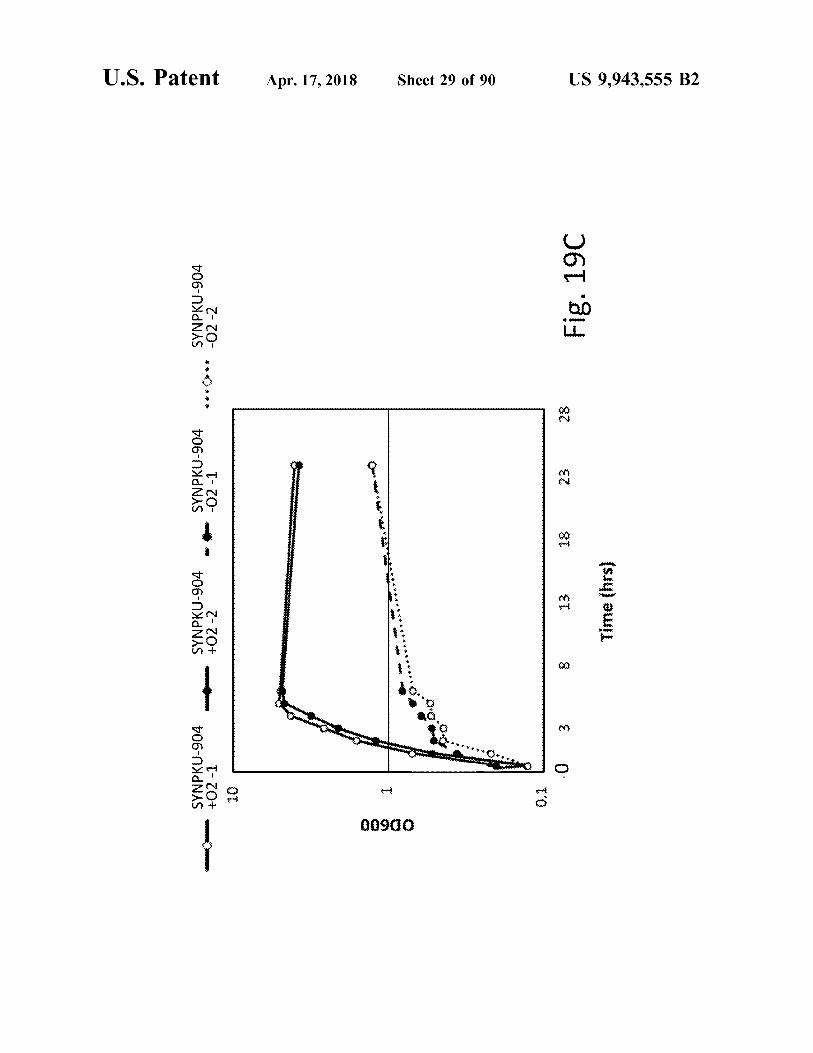
U . S . Patent
SYNPKU - 904 + O2 - 1
SYNPKU - 904 + O2 - 2
SYNPKU - 904 - 02 - 1
* * * * * *
SYNPKU - 904 02 - 2
10
Apr . 17 , 2018
00900
Sheet 29 of 90
0 . 1
0
3
8
13
18
23
28
Fig . 19C
Time ( hrs )
US 9 , 943 , 555 B2
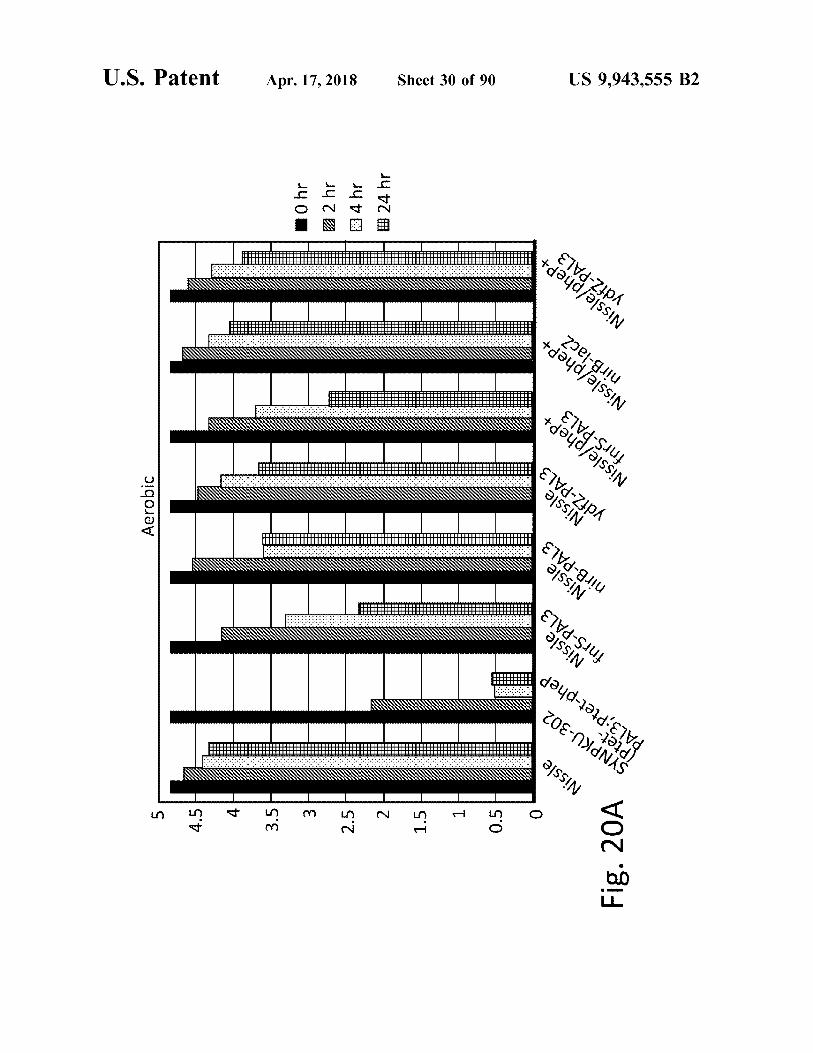
Fig 20 o © - 8 A 6 _ n
?????????????????????? ? ? ? ? ? ? ? ? ? ?
: Nissle | | | 1 |
| | |
| |
1 |
1 |
1 1 1 1 1 1 1 1 1 1 I I 1 1 1 1 1 1 1 1 1 1 1 | | | | | | 1
| 1 |
1 |
1 |
1 |
IIIIIIIII | 1 1 1 1 1 1 1 1 1 1 | | | | | | | | | | | | | | | | | | | | | | | | II I II I II 1 1 1 1 1 1 1 1 1
( Ptet SNNPKU - 302 ? ????????
? ??????? ????? ?????????? ?????????????????????? VAL3 Ptet - phep
|
???????????? ?????
Nissle fnrS - PAL3 1 1 1 1 1 1 1 1 1 1 1 1 | 1 1
| 1
1 1 1
1 1
1 1
1 1
1 1
1 1
1 1
1 1
1 1
1 1
1 1 1 1 1 1
????????????????????????????? ?????
Nissle ??B - PAL3 ?
E ? ?????????????????????? ????????????????? ???????????????
" " " "
Aerobic ??????????????????? ????????? ????????
????
Nissle ydfZ - PAL3 # 1 1 1 1 1 1 1 1 1 1
??? ???????????????????????????????? : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : ?????? ? ??? ???????? ? ?
??????? Msgen? * ????????????????
| | | | | | | 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 | | | | | | | | | |
IIIIIIIIIIII
??? ?
???????? ? ???? ? ? ? ? ? ? ????????????????????
??????????????????????????????????????? ????????????????????? ????????????????
| NisslepheP + TirB - lac? . LLL ILL FILL ILL LLLLL LL ALL LL LLLLL L LLLL LLL LL ? .
|
" . . . . . . ' 1 . . ' . . . . . . . . . .
??? ? ? ? ?
NisslepheP + ydt - PAL3 | 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 | 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
?? 24 hr E3 4 hr A2 hr 10 hr
US 9 , 943 , 555 B2 Sheet 30 of 90 Apr . 17 , 2018 atent
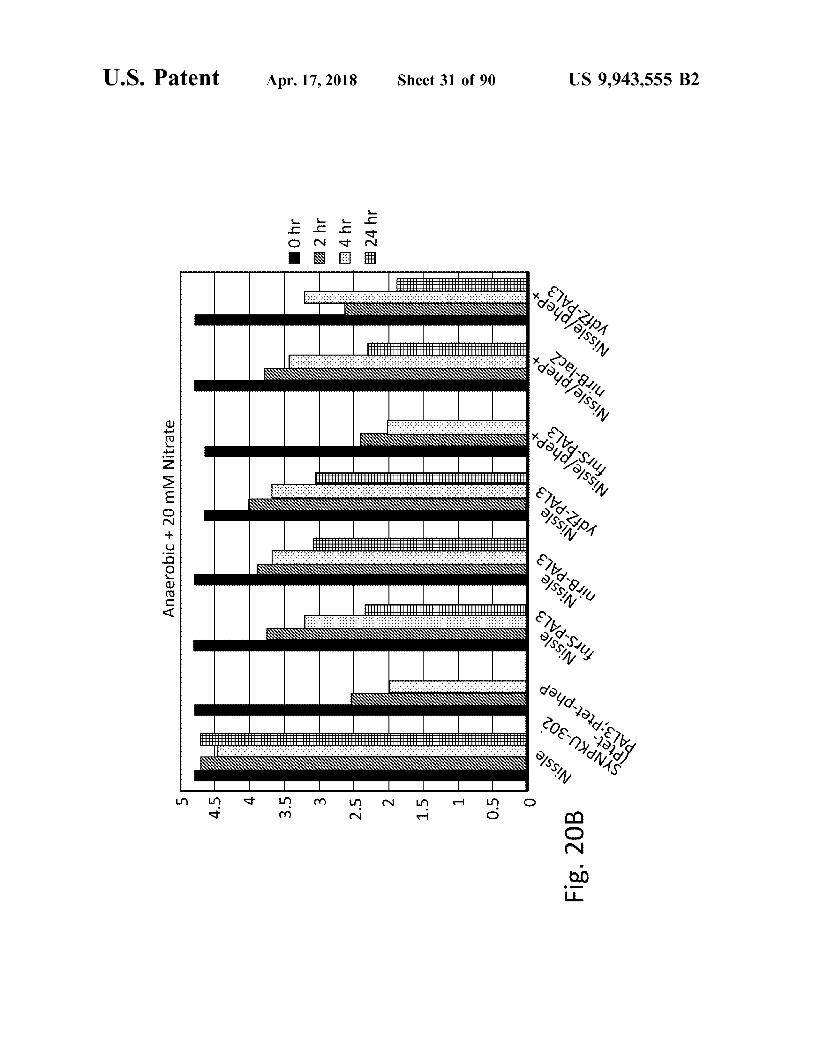
Fig . 20B
w o ú?nä w ? ?? ? ? II III
1722ZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZ ??
222222222222222222222222222222222222222222222222222222222222 2222222222 2222222 2117
UUUUUUUUUU UUUUUUUUUUUUUUUUU 1UUIIIIIIIIIIIIIIIIII DUUUUUUUU
Nissle SYNPKU - 302 S3 , Ptet - phep
Nissle 222222222222222222222222222222222222222222222222222222222222222222222222222222222222222222222222222222222222222222222 fnrS - PAL3
U UUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUU 111111111111111111111111111111111111111111111
UUUUUUUUUUUUUUUUUUUUUU
2222222222
Nissle nir B - PAL3
IIIIIIIIIIIIIIIII UUUUUUU
Anaerobic + 20 mM Nitrate z zzZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZ LL LLLLLLLLLLLLLLLLLLLLLLLLL LLLLLLLLLLLLLLLLLLLLLLLLL 2222 22LLLLLLLLLLLLLLLLLL LLLLLLLLLLLLLLLLL LLLLLLLLLLLLLLL 2222222222222222222222222222222222222222
zzzzzZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZ222222 LLLLLLLLLLLLLLLLLL222222 22222222222222222222222222ZZZZ
Nissle ydfZ - PAL3 IIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIII UUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUU
11111111111 ??
Nissle / phe fnrS - PAL3 2222222222222
22222222222222 2 . 2222222
??????????????????????????????????????????????????????
212 LLLLLLLLLLLLL LLLLLLLLLLL LLLLLL22112112222222222222222222222222 222222222 2 22222
NisslepheP + nirB - lacz UUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUU
1UUUUUUUUUUUUUUUUUUUUUUUUUUUUUUUU
222222222222222222222222222 22222 2 22LLLLLLL2Z222222222222ZZZZZZZZZZZ NisslepheP + ydfZ - PAL3 IIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIII IIIIIIIIIIIIIII JUU IIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIII
24 hr 3 4hr 2hr 10hr
US 9 , 943 , 555 B2 Sheet 31 of 90 Apr . 17 , 2018 atent
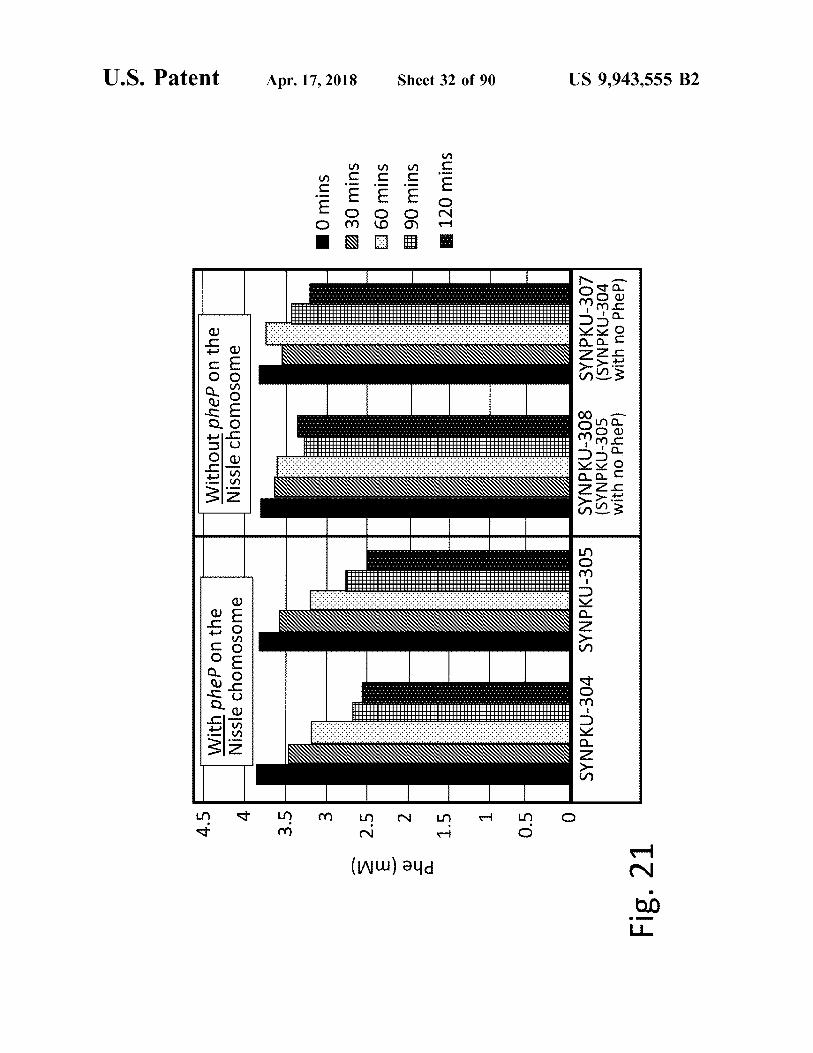
atent Apr . 17 , 2018 Sheet 32 of 90 US 9 , 943 , 555 B2
10 mins ZZ30 mins [ 60 mins E # 90 mins ® 120 mins [
' ' ' ' ' ' ' ' ' ' ' ' ' ' ' '
' ' ' '
' ' ' '
• •
' ' ' ' ' '
' ' ' ' ' '
• - -
' '
' '
-
' '
' '
-
' ' ' '
' ' ' '
' '
' '
' ' ' '
' ' ' '
• '
" -
' '
' •
' '
' •
' '
' •
' ' ' ' '
' ' • • •
' ' '
' ' •
' '
' •
' '
' •
' '
' -
' '
' -
' ' ' '
' ' -
' ' ' '
' ' - -
' '
' -
' '
' -
' '
' -
' '
' -
' ' ' '
' ' - -
' '
' -
' '
' -
' '
' -
' ' '
-
' '
' -
•
' ' ' ' ' '
' ' ' - - -
" " "
' '
' -
"
' '
' -
' '
' -
"
' '
' -
' '
' -
' ' ' '
' ' - -
' ' ' '
' ' - -
" "
'
' '
' -
'
' '
' -
"
' '
' -
'
' '
' -
' ' ' ' '
' ' - - -
' '
" -
' '
' -
' ' ' ' ' ' • - - - ,
" " | | | | | | | | | |
1 |
| 1
| 1
| 1
| 1
| 1
| 1
| 1
| 1
| 1
| 1
| |
| |
| | 1
| | I
1 | I '
In '
1 | 1
| | I I | | I I 1 1 1 1 1 I 1 1 1 1 1 1 1 1 1 1 1 1 1
• ' . SYNPKU - 307 ( SYNPK? - 304
with no PheP ) ??????????????????????????? ???????????????????
???????????????????????????????????????????? ????????????????????????????????????????????????????????????? ????????????????????????????????????????? ???????????????????????????????????????????????????
Without phep on the Nissle chomosome ' ' '
- '
- '
- -
' -
' -
-
' -
' -
-
' -
' -
-
' - '
- -
'
' - -
' - -
-
' ' -
' ' -
- -
' - '
- -
' - -
' - -
-
' - '
- -
' -
-
'
' - -
' - -
-
' -
' -
-
'
' ' - - -
' ' - - -
- -
' ' - -
' ' - -
- -
'
' - '
- -
' - -
' - -
-
' - '
- -
' - -
' - -
-
' - '
- -
' ' - -
' ' - -
- - [
' -
' -
- ]
' -
' -
-
' -
' -
- [
' -
' -
-
' -
' - -
- I
' -
' -
-
' - '
- -
' -
' -
-
' - '
- -
' ' •
' ' - -
- -
' •
' -
-
' •
' -
-
' ' • •
' ' - -
- -
' •
' -
-
' • •
' - -
-
' •
' -
-
'
' •
' -
-
' •
' -
-
' •
' -
-
' ' ' '
- - -
' ' '
- -
' '
' -
' ' ' ' ' '
' ' ' - - -
- - -
' ' '
-
' '
' -
-
' ' '
' - -
- -
' ' '
- -
' ' '
' - -
- - -
- - - - - - - - - - - - - -
1 1 1
1 1
1 1 | 1 1 1 1
| 1 1
1 1 1
1 1
| 1 1 1 1 1
??????????????? ?????????????????????????????????????????????? ???????????????????????????????????????????????? :
( SYNPKU - 305 with no Phep )
SYNPK0 - 305 | SYNPKU - 308 - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
' -
' -
' -
' -
' -
' -
' -
' -
' -
' -
' -
' -
' -
' -
' -
' -
' -
' -
' -
' -
' -
' ' ' ' ' ' ' ' ' '
' ' ' ' ' " " " " " " " " " " " " " " " " " " " " " " " " " " "
?
1 | | | | | I I 1 1 1 1
I I I 1 1 1 [ [ [ [ [ [ LL
| 1
1 1 1 1 1 1 1 [ [ [ [ [ [ [
L IL
| II | | | | II | | | I 1 1 | | | IIIIII | | | | | | | | hhl With phep on the Nissle chomosome ' ' ' ' ' ' ' ' '
- - - ' ' '
- '
• •
' -
' •
•
' ' - -
' ' • •
• •
' -
' •
•
' - -
' • •
•
'
' -
' •
•
' -
' •
•
' - '
• •
' -
' •
•
'
' -
' •
•
' ' - - ' '
• • • •
'
' -
' •
•
' -
' •
•
' ' - -
' ' • •
• •
' -
' •
' -
' •
•
' -
' •
•
' - '
, •
' -
' ,
•
' -
' ,
•
' -
' ,
•
' -
' , ,
•
' -
' ,
•
'
' -
' ,
•
' -
' ,
•
'
' -
' ,
•
' '
' ,
•
' ' '
' ' ,
'
' '
' ,
•
' ' '
, •
'
' ' ' '
' ' , ,
• •
' '
' ,
•
'
' ' '
' , ,
•
' • • • , , ,
• • • •
|
| | | | | | | | | | | | 1 1 1 1 1 1 1 1 1 1
" " " SYNPKU - 304 ?
????????????????????????????? ????????????????????????????????????????????????????????????????? ? ? ? ? ? ? ? ???????????????????????????????????
???????????????????????????????????????????????????????? ???????????????????
? ? ? ????????????
6 A ? ? ? ? Phe ( mM )
Fig . 21
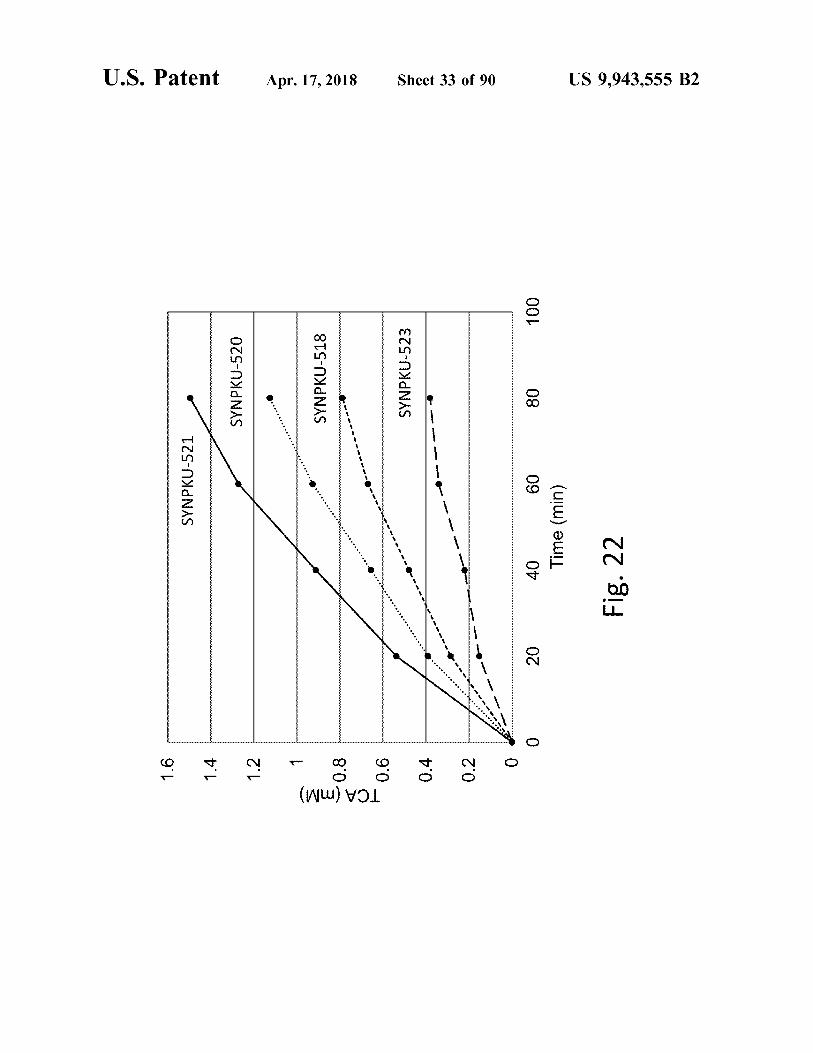
ã w
w
w wwwwwwwwwww
wwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwww w
wwwwwwwwwwwwwwwwwwwww
IZS - NXONAS
atent
SYNPKU - 520
ã
SYNPKU - 518
- TCA ( MM )
Apr . 17 , 2018
SYNPKU - 523
Å
Sama mee en
home annen for
sammen new to
Sheet 33 of 90
i o 0
20
40 60 Time ( min )
80
100
Fig . 22
US 9 , 943 , 555 B2
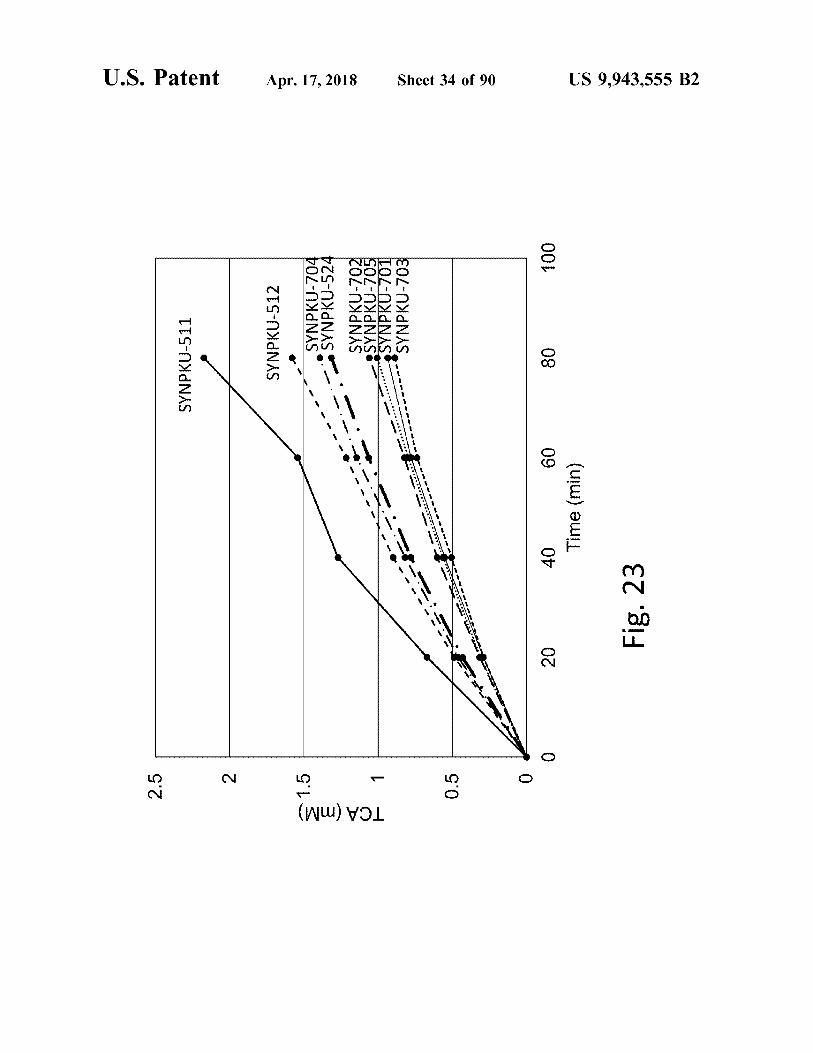
ö
5
SYNPKU - 511
atent
N
SYNPKU - 512 SYA SYNPKU - 704 SYNPKU - 524
ö TCA ( mM )
-
Apr . 17 , 2018
: :
SYNPKU - 702 SYNPKU - 705 SYNPKU - 701 SYNPKU - 703
- - - - - - - - SYNPKU - 703
Z
Sheet 34 of 90
ó o
20
40 60 Time ( min )
80
100
Fig . 23
US 9 , 943 , 555 B2
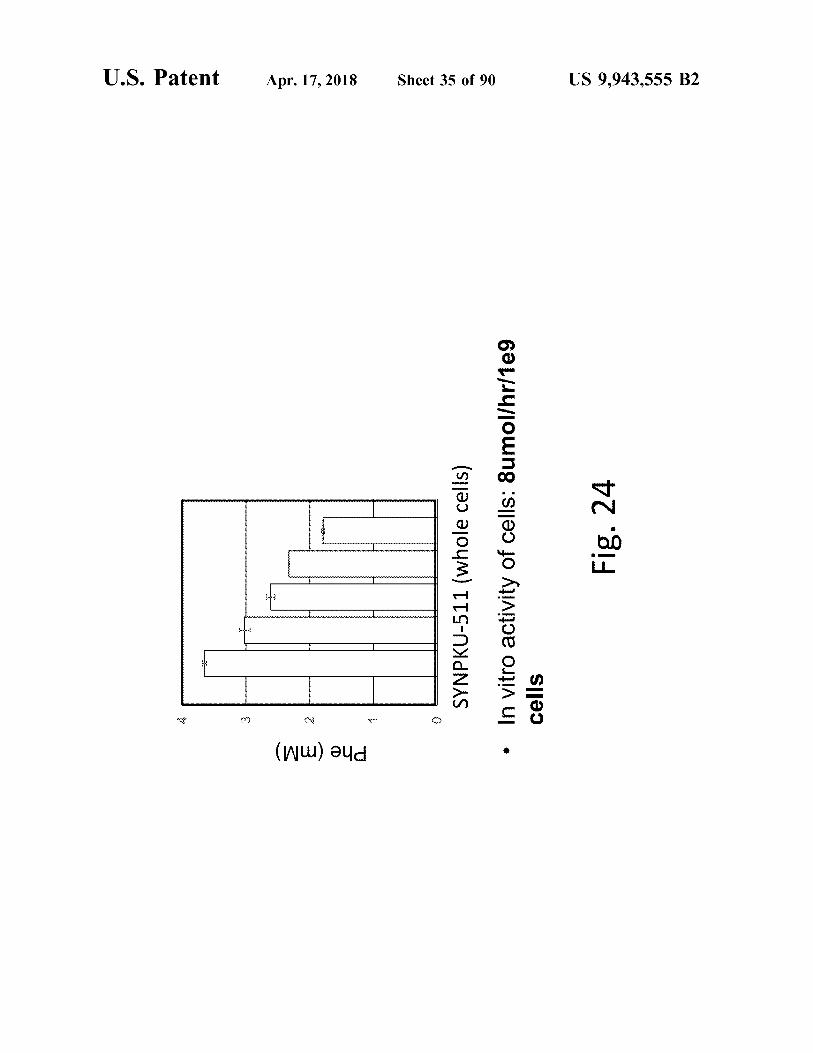
atent
Phe ( mm )
Apr . 17 , 2018
SYNPKU - 511 ( whole cells ) • In vitro activity of cells : Sumol / hr / 1e9
cells
Sheet 35 of 90
Fig . 24
US 9 , 943 , 555 B2
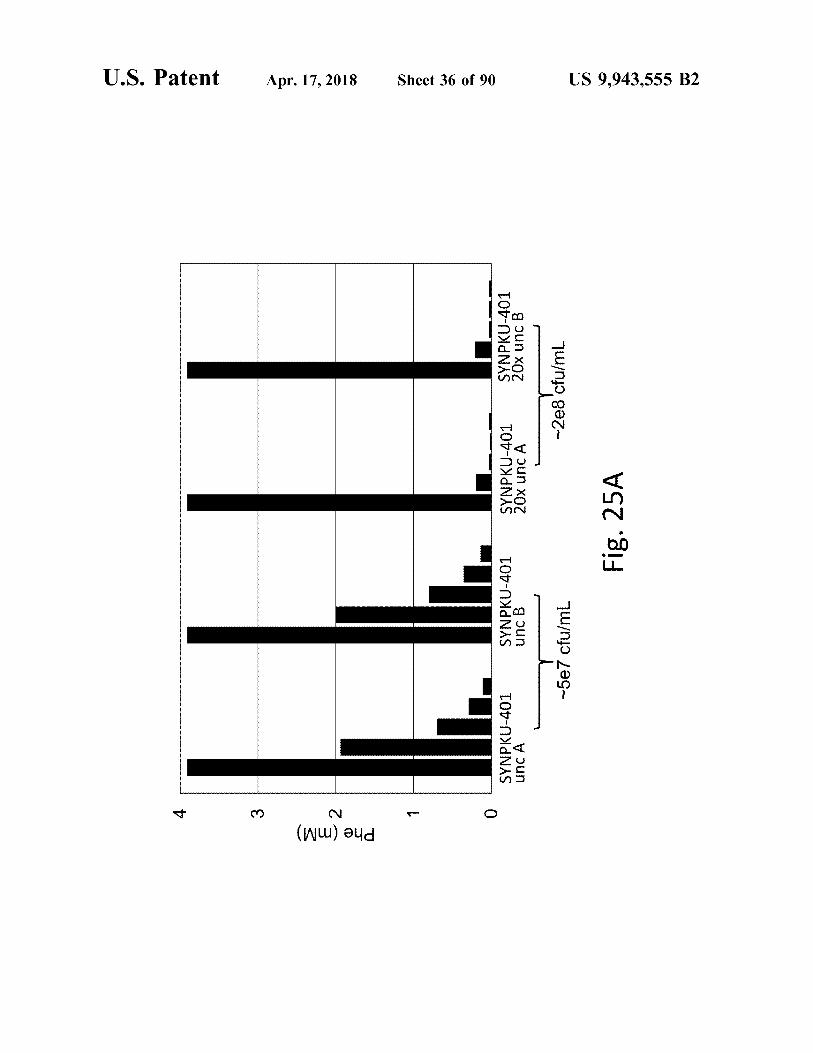
atent
w Phe ( mm )
1111 Apr . 17 , 2018 Sheet 36 of 90
SYNPKU - 401 unc A
SYNPKU - 401 unc B
SYNPKU - 401 20x uncA
SYNPKU - 401 20x uncB ~ 2e8cfu / mL
- 5e7 cfu / mL
Fig . 25A
US 9 , 943 , 555 B2
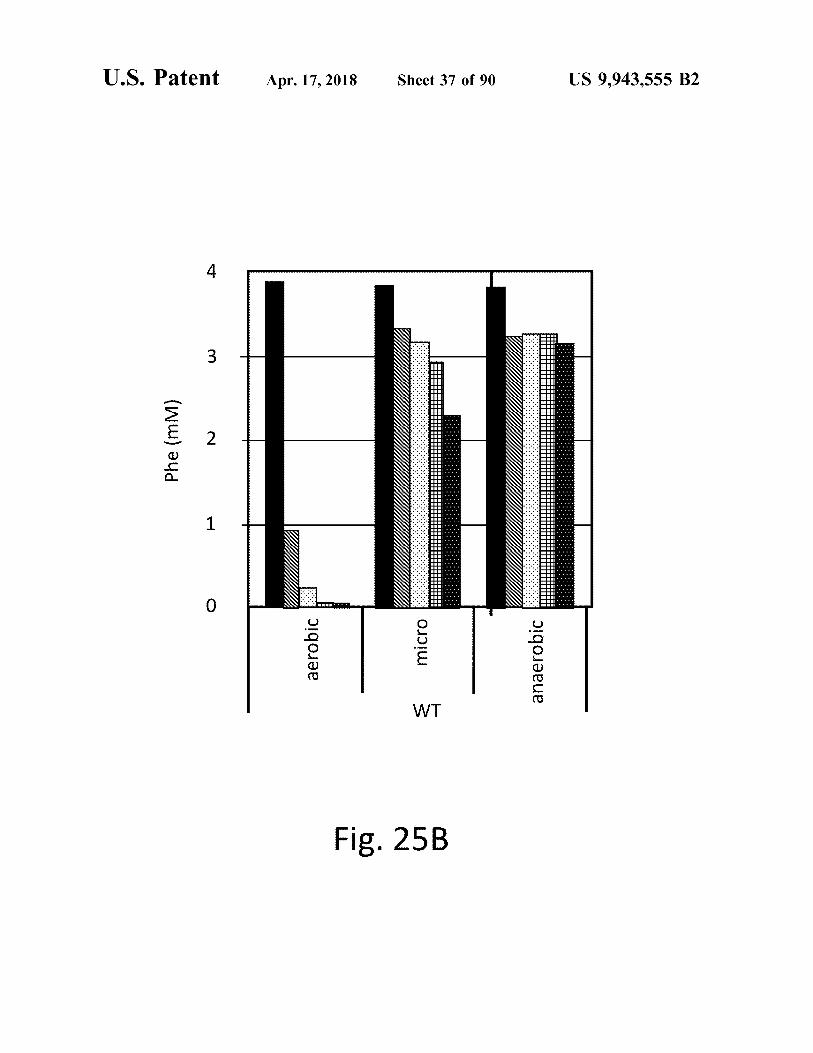
atent Apr . 17 , 2018 Sheet 37 of 90 US 9 , 943 , 555 B2
• |
???? ? ??? _ en | ?
' ' ' '
?
• ' '
' • '
' '
' '
' ' ' ' |
|
|
|
|
I II ' ' ' _ N
| '
? | | I ' 1 1 1 " " " "
Phe ( mM ) [ [ ] l
' "
'
' "
'
' "
'
' "
' "
1 ' [ [ [ [ [ [ m
' ' ' " ' ' ' ' ••• "
' ' '
' ' ' " '
? ' ' ' '
I II ' ' ' '
" • "
I | | III " " " "
[ I " " " "
| | | • ' ' '
1 1
' " " " 1
?? ?? ?
| ? 1 1
|
1
?? ?? I II
[ [ ] ?? ' ' ' '
_ o ???
??
? ?? ' ' " ' ' l
aerobic micro anaerobic ?
Fig . 258
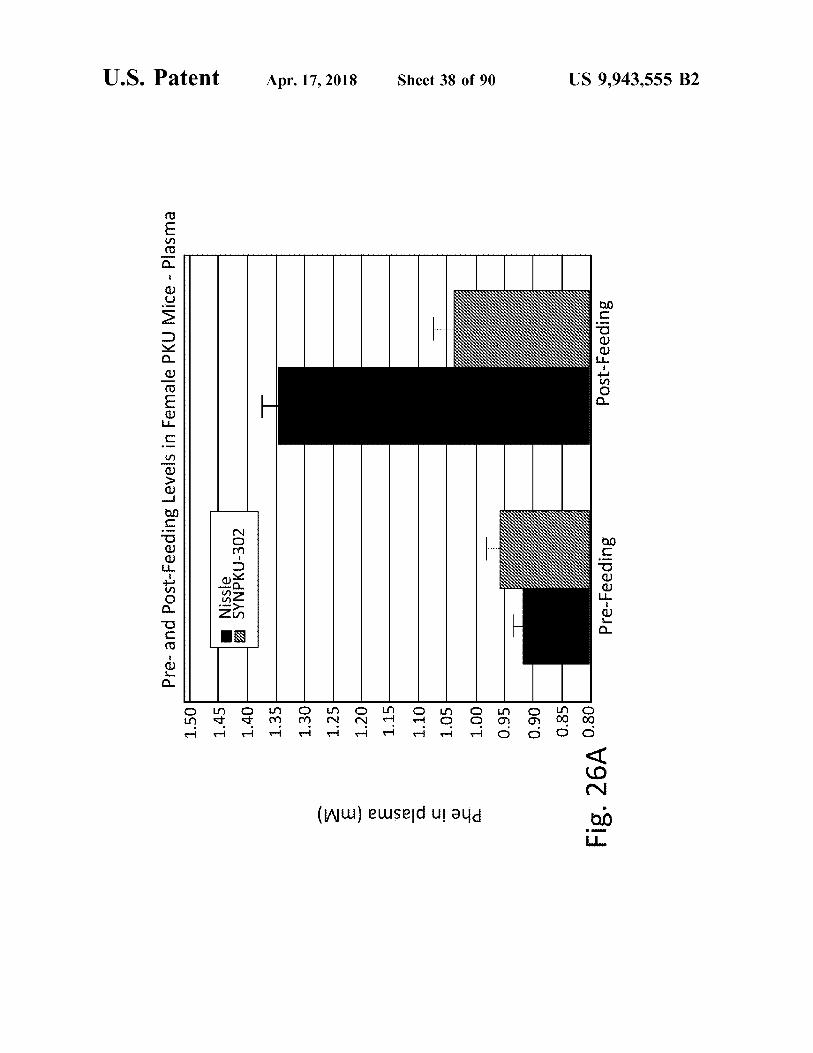
Pre - and Post - Feeding Levels in Female PKU Mice - Plasma
1 . 50
jual
1 . 45
Nissle Z SYNPKU - 302
1 . 40 1 . 35 1 . 30
810Z ' LI ady
1 . 25 1 . 20 1 . 15
Phe in plasma ( mm )
1 . 10 1 . 05
06 JO 8€ v??YS
1 . 00 0 . 95
22 .
12
2222
2222 . zzzzzz
222 22222222
122222222222222222222222LLLLLLLLLLLL izLLLLLLLLLLLLLLLLLLL LLLLLLLLLLLLLLLLLLLLLLLL LLLLLLLL
0 . 90 0 . 85
Fig . 26A 0 . 80
Pre - Feeding
Post - Feeding
za SSS?€ + 6 * 6 SN
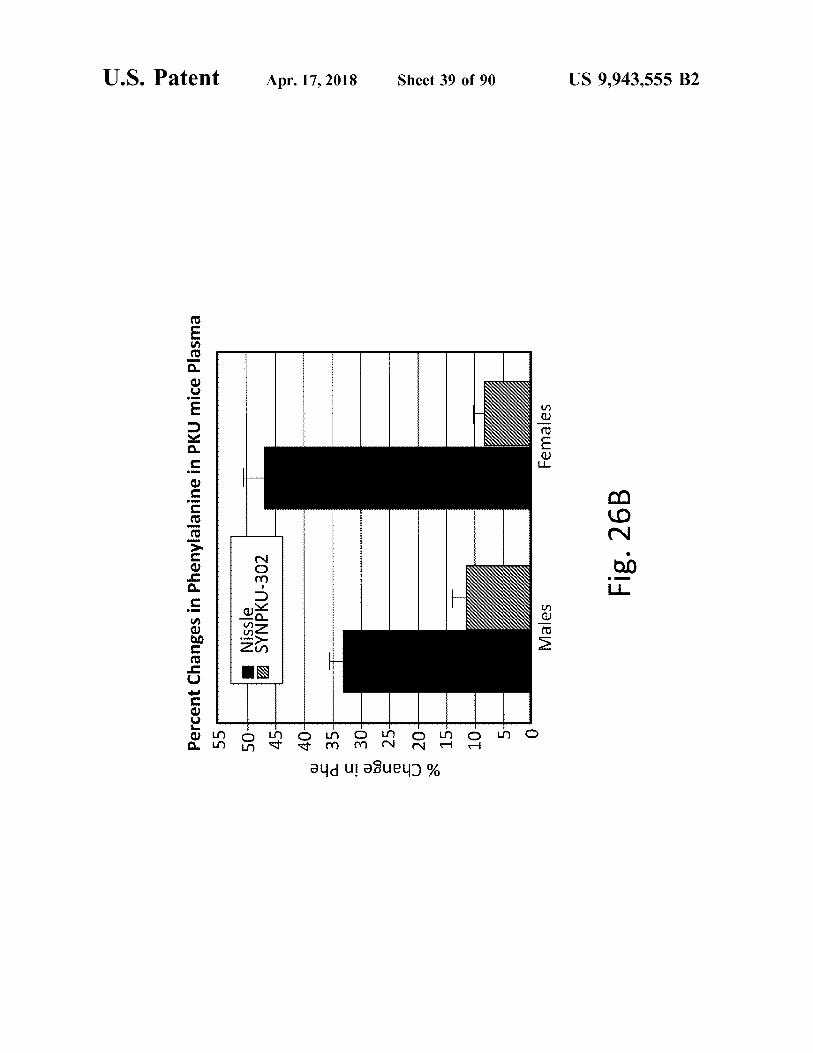
% Change in Phe 6 H 8 9 8 7 6 7 8 9 8 o C
Males IIIIIIIIIIIII
SYNPKU - 302 Nissle .
?
22222222222222222222222222222222
?????
Fig . 26B Percent Changes in Phenylalanine in PKU mice Plasma Females
22LLLLLLLLLLLLLLL ? ????? 222222 LLLLLLLLL
za SSS?€ + 6 * 6 SN 06 JO 6E J??US 810Z ' LI ady jual
atent Apr . 17 , 2018 Sheet 40 of 90 US 9 , 943 , 555 B2
wwwwwwwww w wwwwwwww pomimmmmmmmmmmmmmmmmmmmmmmmmmmmmmmmmmmmming
* * * wwwwwwwwwwwwww Website
22222222222 11122
177777777777777 42222
LLLLLLLL21 12 ( ww ) ( audy ( WW ) ( oudy
? 1111111111 LIITILIT 1122
27
HO SYN - PKU901 SYN - PKU303 H2O SYN - PKU901 SYN - PKU303
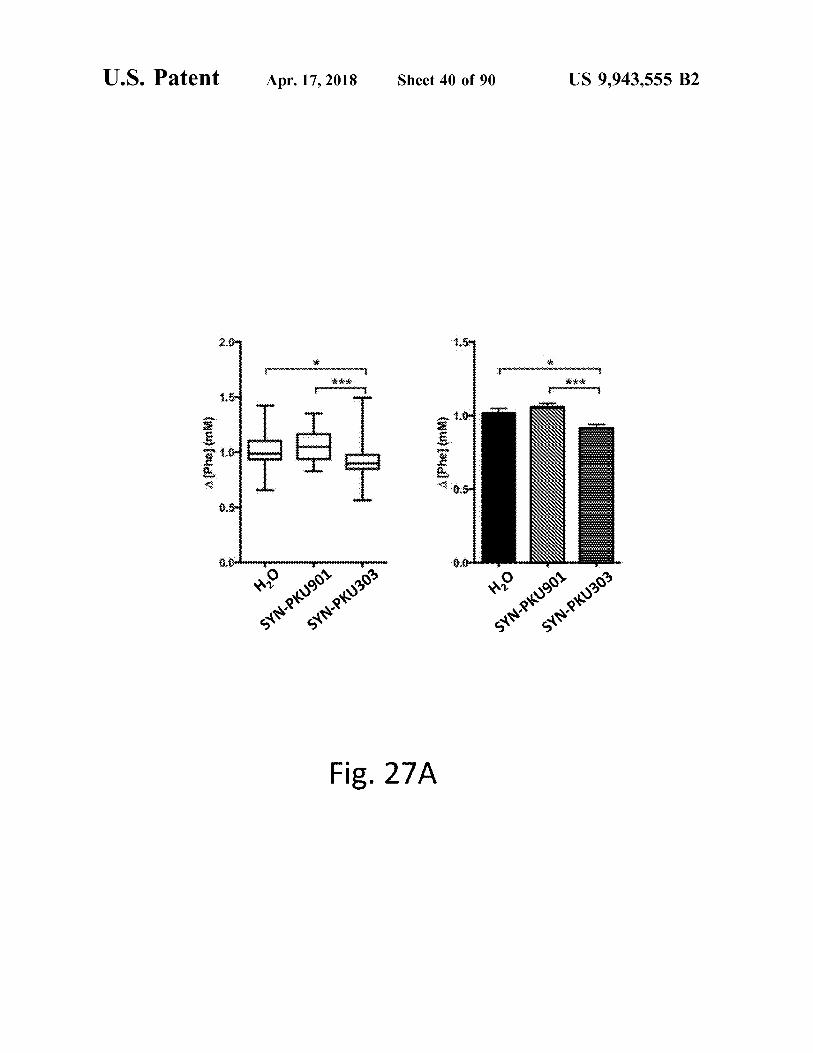
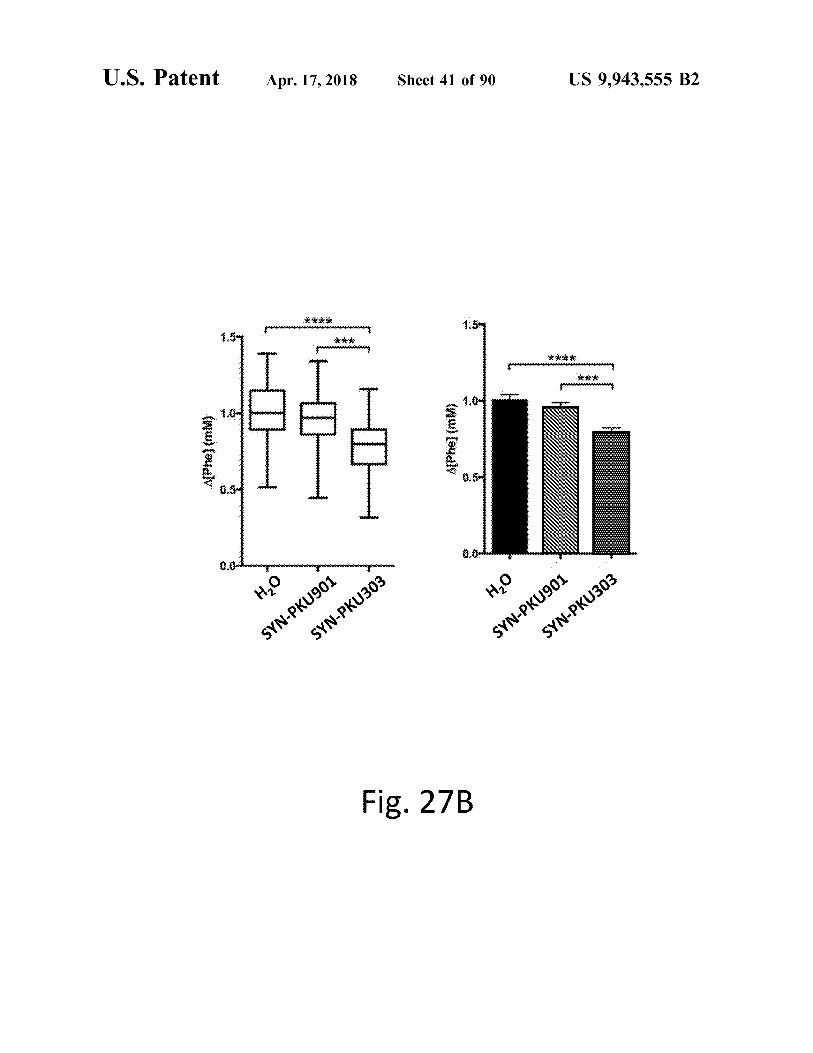
Fig . 27A
atent Apr . 17 , 2018 Sheet 41 of 90 US 9 , 943 , 555 B2
* * wwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwwww
an h
ht
ZZZZZ TILLILITLLLLLLL
?????
wu ) ( oudly ( Nw ) ( audiv 2222222222 1111111111 1 + 1 + 1 + 1 + 1 + 1 + 1 + 1 + 1 + 1 + 1 + 1
' o mmmm
H2O SYN - PKU901 SYN - PKU303 ?? 106nyd - NAS ???? » d - NAS
Fig . 27B
U . S . Paten atent Apr . 17 , 2018 Sheet 42 of 90 US 9 , 943 , 555 B2
. . . .
?????? ?????? 21ss A [ Phe ] ( mm )
ITT 1
TTT IIIIII
ITIT II I TIITTIIN IIIIIIII s
LLLLLLLLLLL IT IIIII
sssssssssss I IT
1 INDI TIT
I 1 TIT
?????? 11 . 0 . 0 - H2O SYN - PKU901 SYN - PKU303 SYN - PKU304
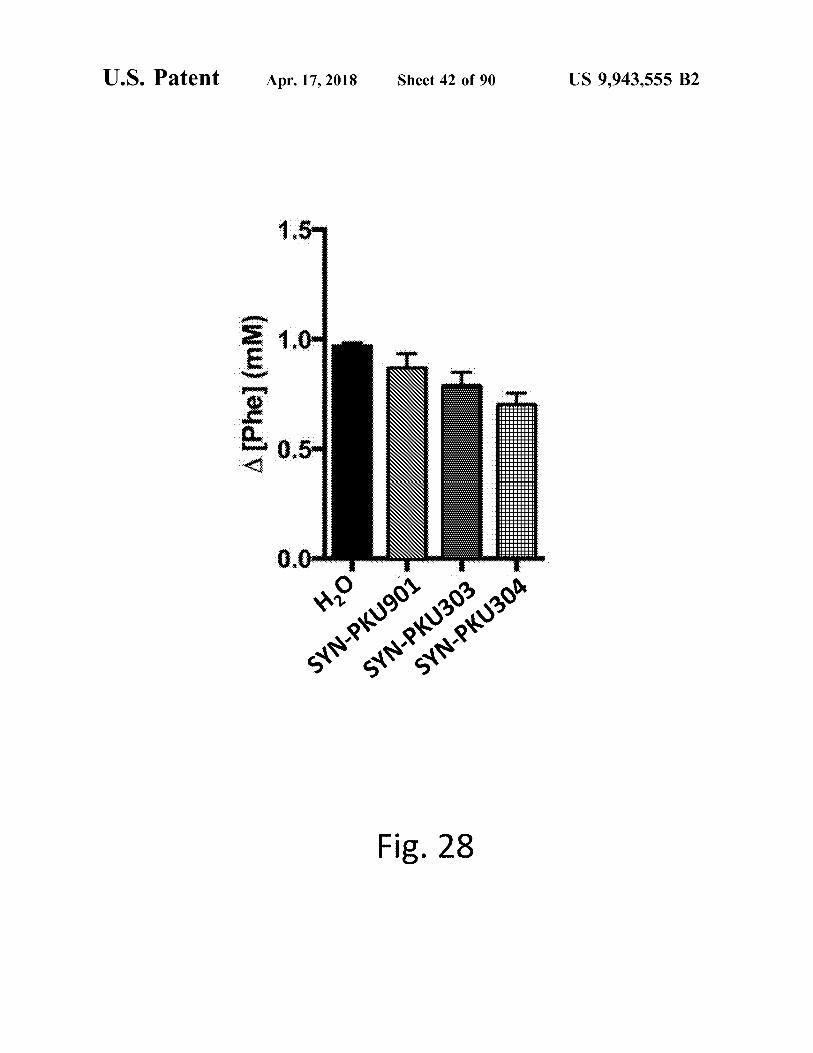
Fig . 28
2hrs
4hrs
* * *
*
* * * *
atent
* * * *
r
* * * * * *
??? * *
AIPhel { mM )
AIPhe ( mM )
ITH | LL IF
Apr . 17 , 2018
0 . 5
0 . 5
…
H . 0 SYN - PKU901 ? SYN - PKU303 ? | SYN - PKU304?
| H2O SYN - PKU901 ) | SYN - PKU303 ? | SYN - PKU304 \
Sheet 43 of 90
0 . 0
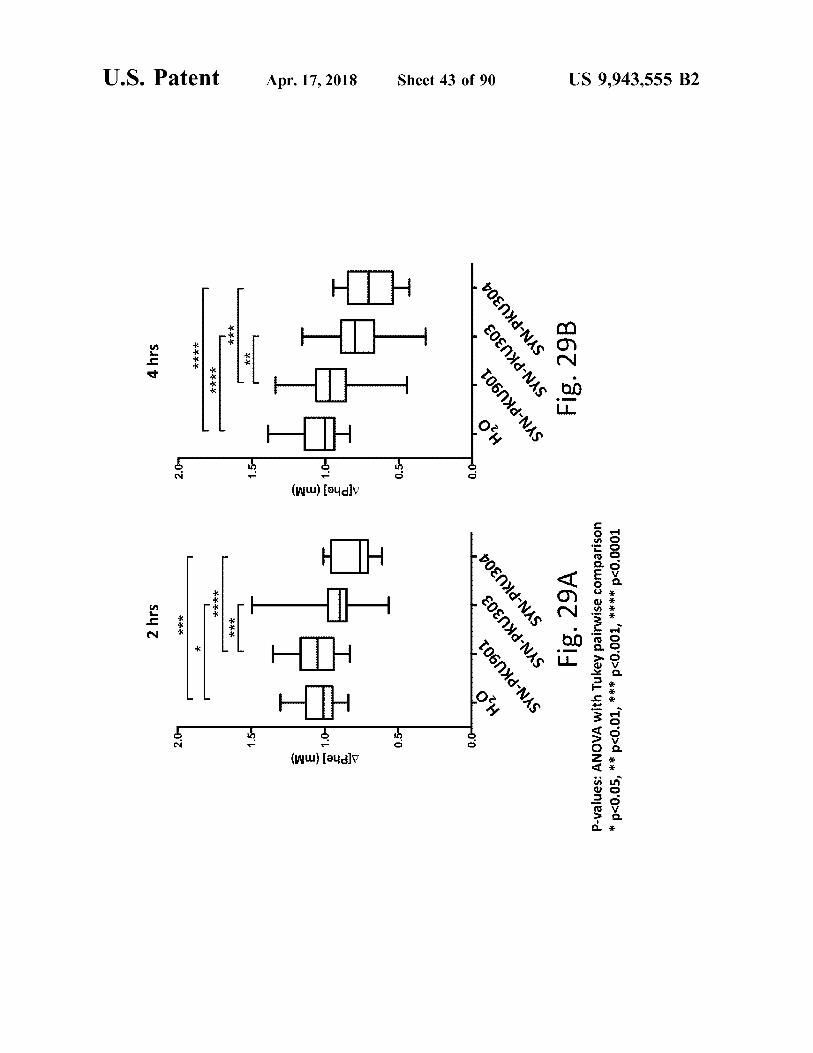
Fig . 29B
Fig . 29A
P - values : ANOVA with Tukey pairwise comparison * p < 0 . 05 , * * p < 0 . 01 . * * * p < 0 . 001 . * * * * p < 0 . 0001
US 9 , 943 , 555 B2
2 hrs
4 hrs
2 . 0
* * *
atent
*
* * * *
IL
I
* * * *
* * * *
1
* * *
* * *
* *
1 . 5
1 : : .
A [ Phe ] ( MM )
: : : : ?
A [ Phe ] ( mm )
:
11 . : ) > > > : .
Apr . 17 , 2018
: 1
:
0 . 5
-
,
H20 SYN - PKU901 SYN - PKU303 SYN - PKU304
H20 SYN - PKU901 SYN - PKU303 SYN - PKU304
Sheet 44 of 90
0 . 000
0 . 02
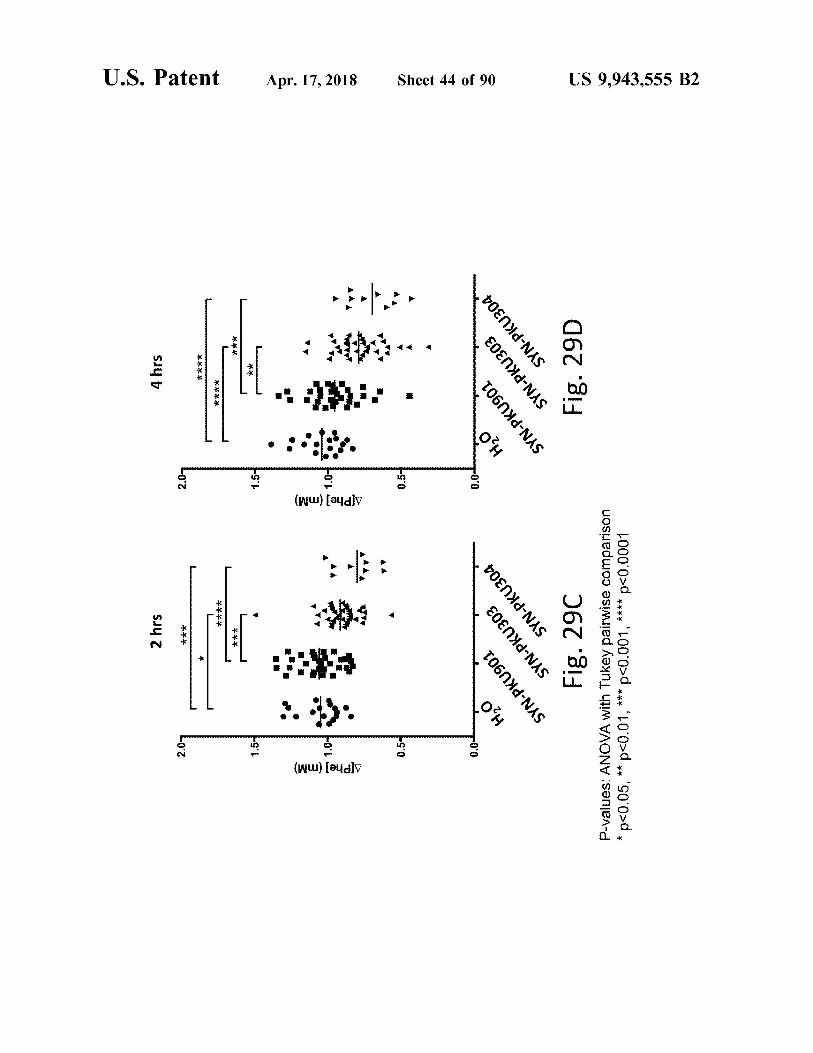
M Fig . 29C
Fig . 29D
P - values : ANOVA with Tukey pairwise comparison * p < 0 . 05 , * * p < 0 . 01 , * * * p < 0 . 001 , * * * * p < 0 . 0001
US 9 , 943 , 555 B2
atent
1 . 5mm
11111111111111111
A [ Phe ] ( MM )
1 . 0
Huiviviviviviviviviviviviviviviviviviviviviu
??????? ?????????????????? ?????? ???????????? ? ? ?
HOH HH HT
A [ Phe ) ( mm )
ZZZZZZZZZZZZZZZZZZZZZZZZZZZZ
Apr . 17 , 2018
HHHMMMMMMMMMMMMMMMMMMMMMMMMMM
IIIIIIIIIIIIIIIIIIIIIIIIIIII MUUUUUUUUUUUUUUUUUU
HIIH
0 . 5
Z
SYN - PKU901 2x109 CFUS SYN - PKU304 5x108 CFUS * SYN - PKU304 1x109 CFUS SYN - PKU304 2x10°CFUS
0 . 0m SYN - PKU901 2x109 CFUS SYN - PKU304 5x108 CFUS SYN - PKU304 1x10 CFUS SYN - PKU304 2x109 CFUS
H2O
o°H
Sheet 45 of 90
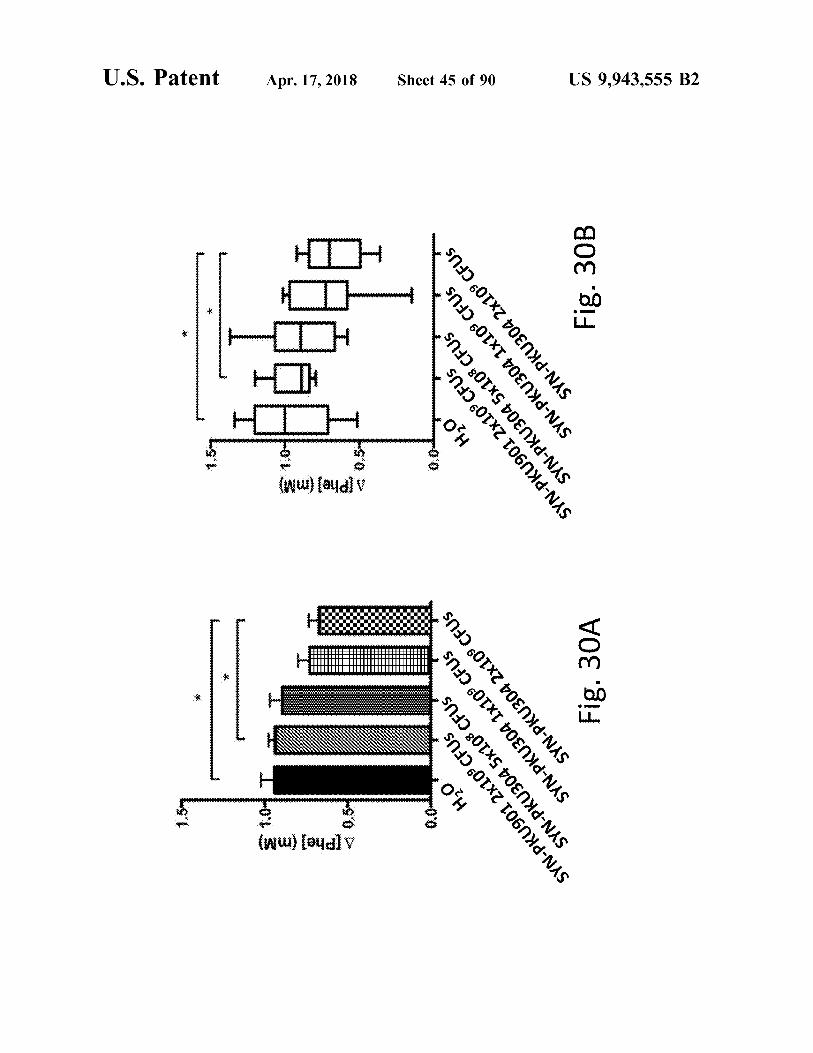
Fig . 30A
Fig . 30B
US 9 , 943 , 555 B2
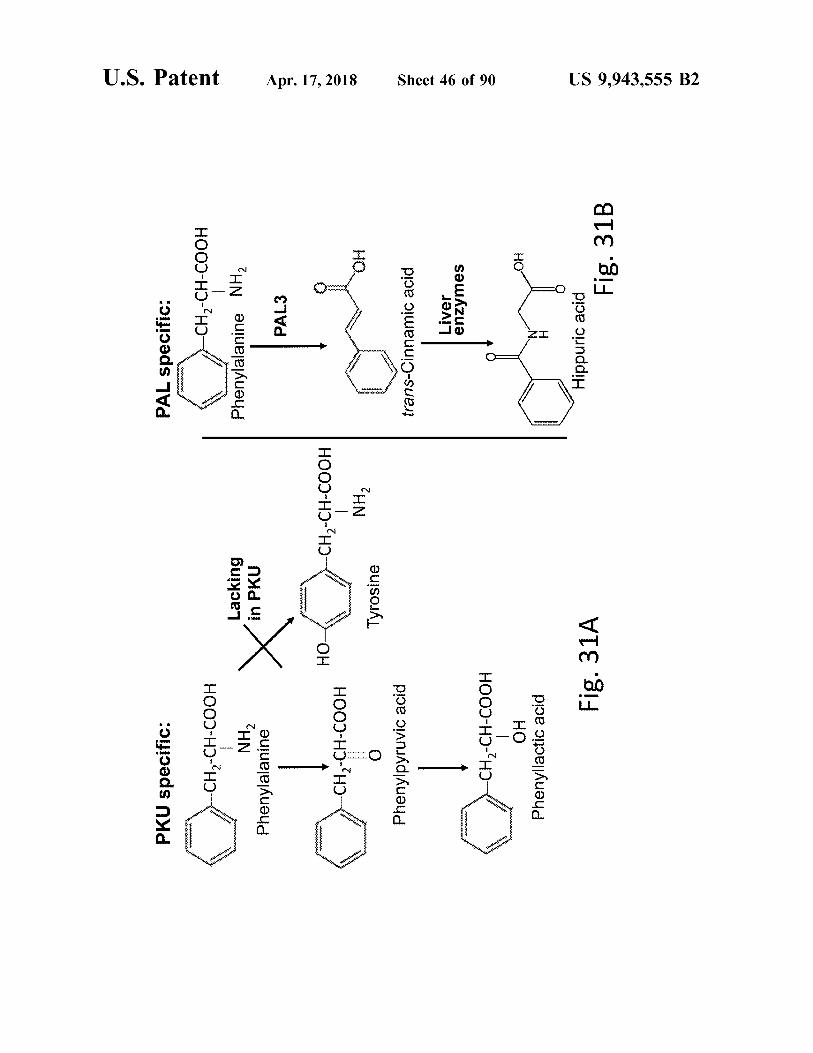
PKU specific :
PAL specific : - CH2 - CH - COOH
atent
pad
- CH2 - CH - COOH NH2 Phenylalanine
Lacking in PKU
Phenylalanine PAL3
Apr . 17 , 2018
HOM
- CH2 - CH - COOH
CH2 - CH - COOH
Tyrosine Tyrosine NHL
Phenylpyruvic acid
trans - Cinnamic acid Liver enzymes
Sheet 46 of 90
_ CH2 - CH - COOH ?? Phenyllactic acid Fig . 31A
Hippuric acid
Fig . 31B
US 9 , 943 , 555 B2
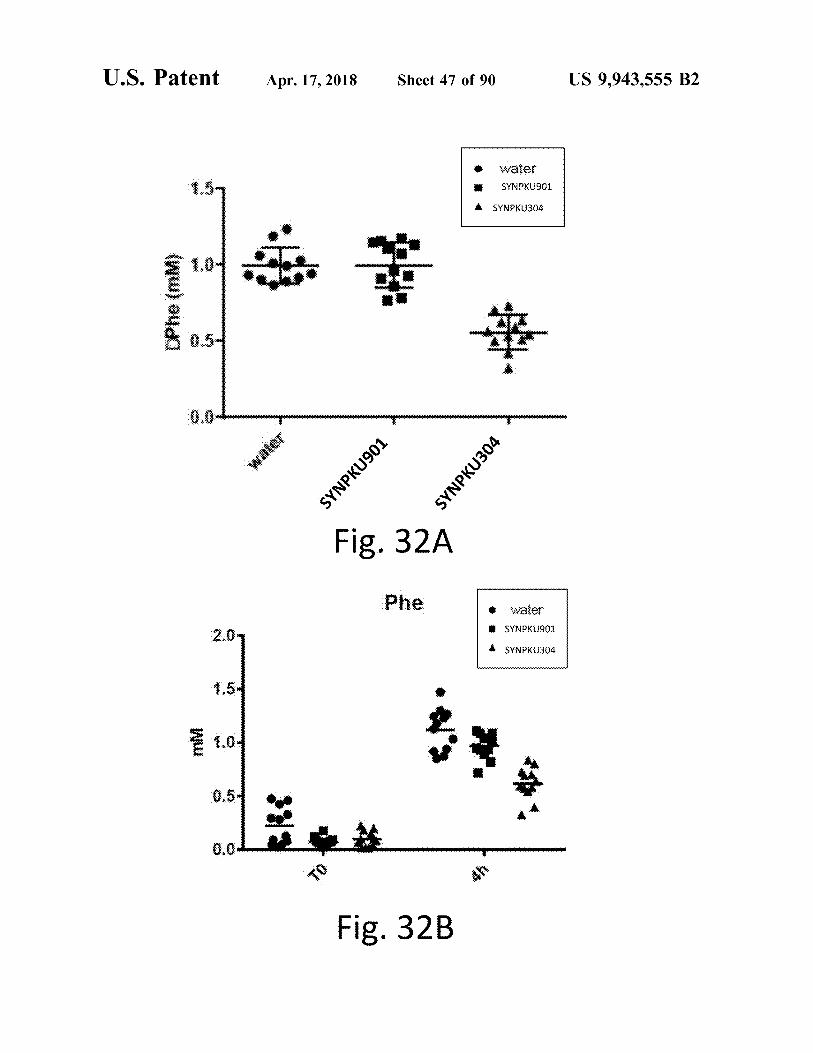
U . S . Patent atent Apr . 17 , 2018 Sheet 47 of 90 US 9 , 943 , 555 B2
Water SYNPKU901
the SYNPKU304 There DPhe ( MM )
water SYNPKU901 SYNPKU304 Fig . 32A
Phe ! !
# water # SYNPKU901
SYNPKU304
WW
AK : #
Fig . 32B
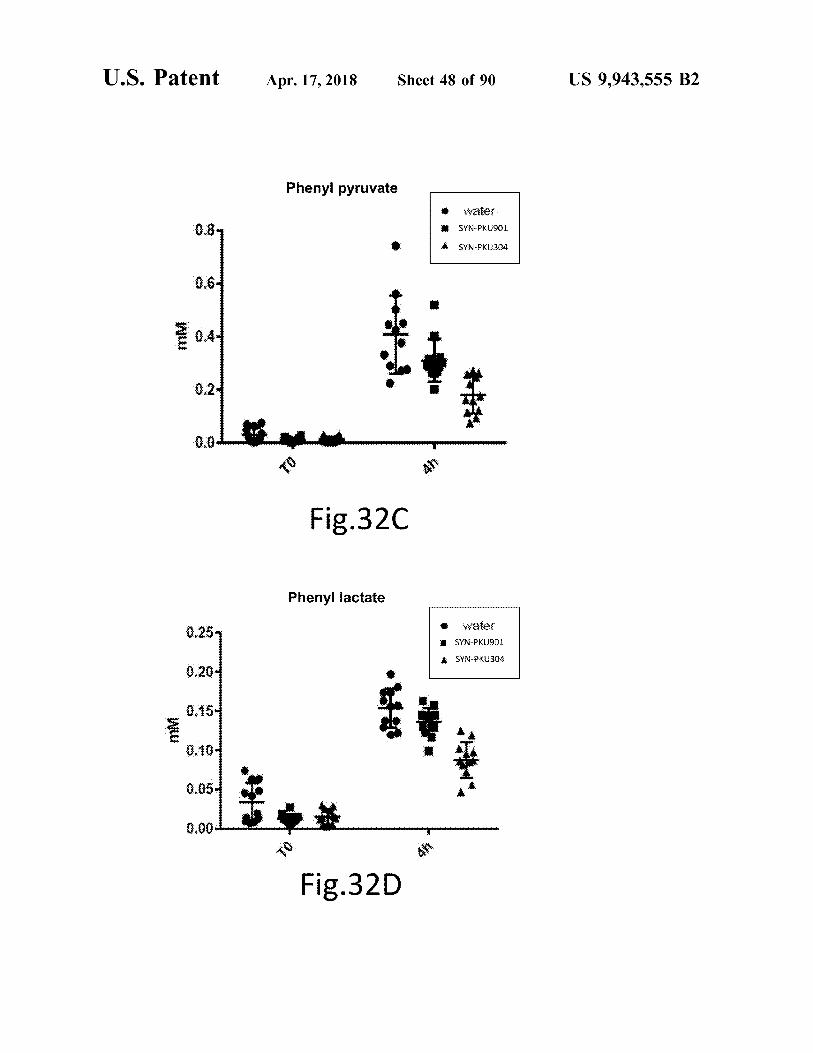
atent Apr . 17 , 2018 Sheet 48 of 90 US 9 , 943 , 555 B2
Phenyi pyruvate Water SYN - PXU901 SYN - PKU304
WW
Fig . 320
Phenyl lactate
SYN - PKU901 SYN - PKU304 0 . 20
WU 0 . 05
0 . 00
Fig . 32D
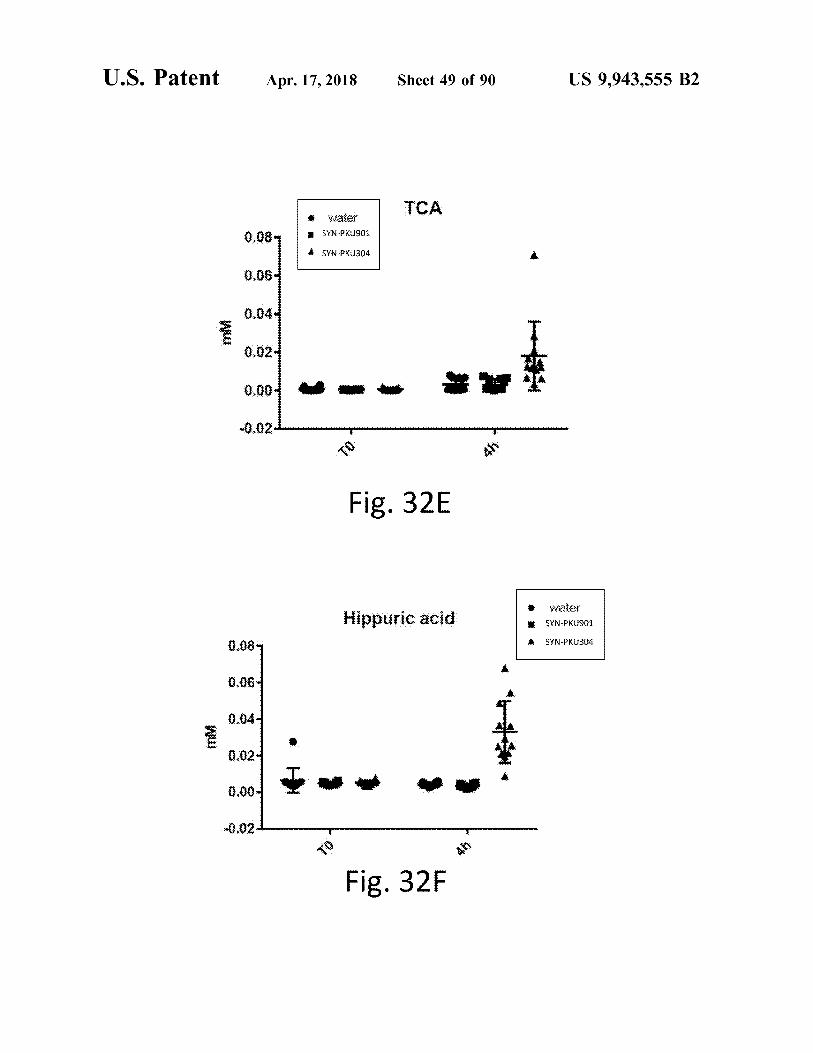
atent Apr . 17 , 2018 Sheet 49 of 90 US 9 , 943 , 555 B2
TCA Water * SYN - PKU901 0 . 08
SYN - PKU304
0 . 06
0 . 04 . WW
0 . 024 - - - vw - 0 . 02
Fig . 32E
Hippurie neid Hippuric acid water SÝN - PKU901 a 0 . 08 # SYN - PKU304
0 . 06 *
Wu _ _ _
* * * *
LLLLLLLLS
Fig . 32F
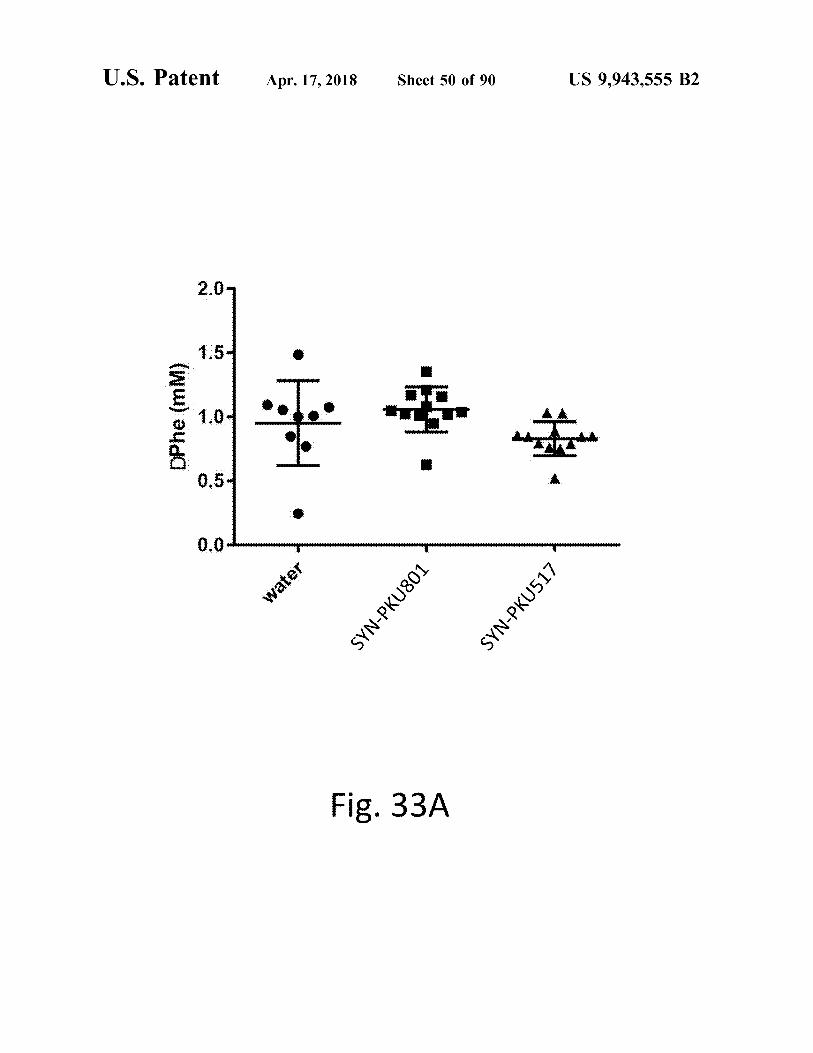
atent Apr . 17 , 2018 Sheet 50 of 90 US 9 , 943 , 555 B2
den
DPhe ( mm ) ? ? * * * i
o
water SYN - PKU801 SYN - PKU517
Fig . 33A
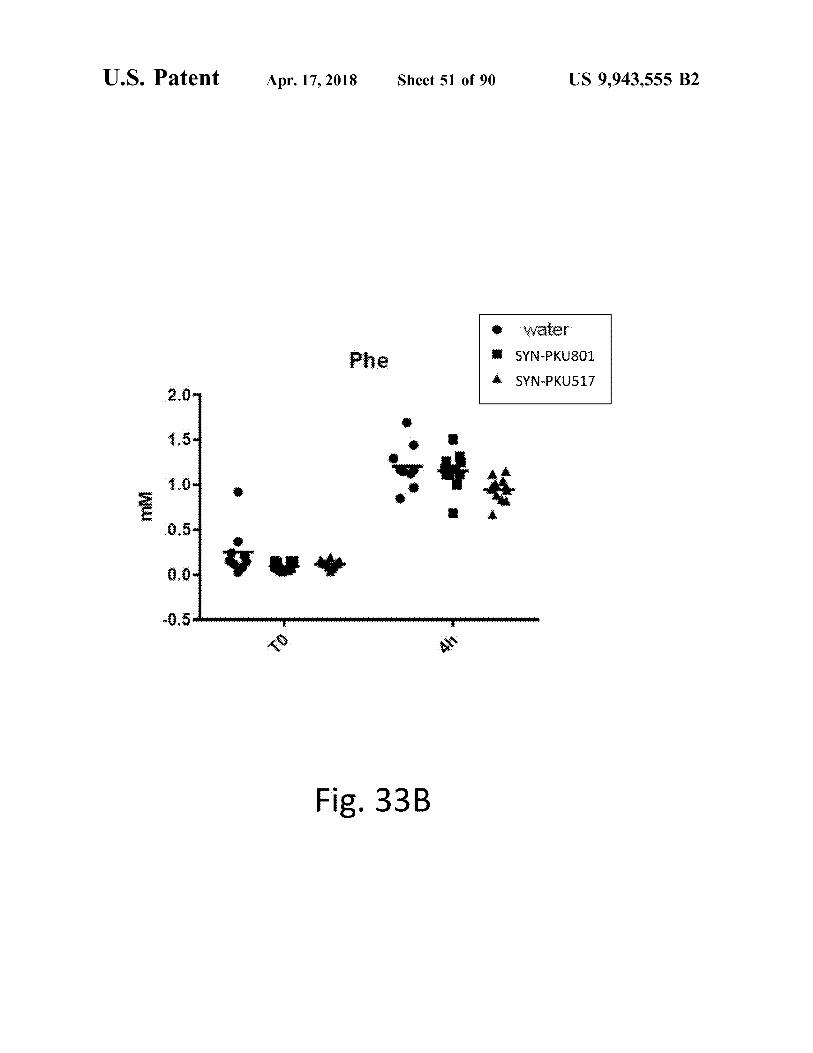
atent Apr . 17 , 2018 Sheet 51 of 90 US 9 , 943 , 555 B2
Water
Phe SYN - PKU801
SYN - PKU517 1
r . é
Ww
CA
Fig . 33B
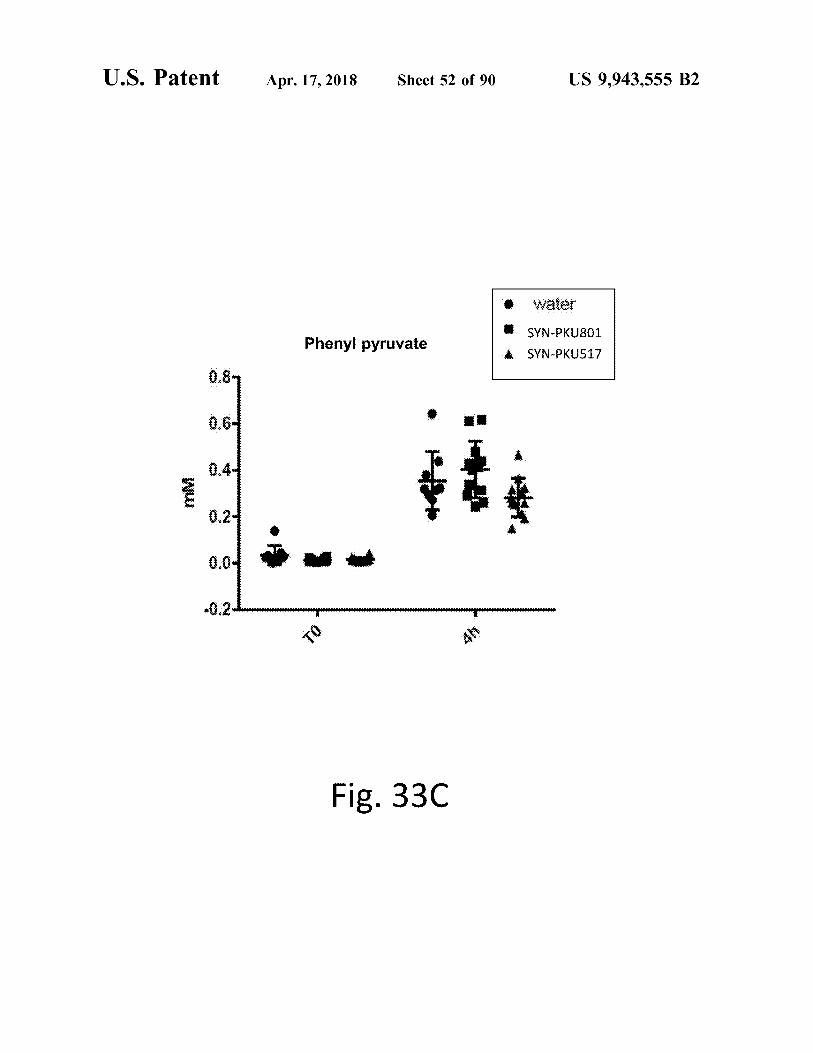
atent Apr . 17 , 2018 Sheet 52 of 90 US 9 , 943 , 555 B2
Water SYN - PKU801 Phenyl pyruvate SYN - PKQ517
??
r ? ????? ?
????? -
Fig . 33C
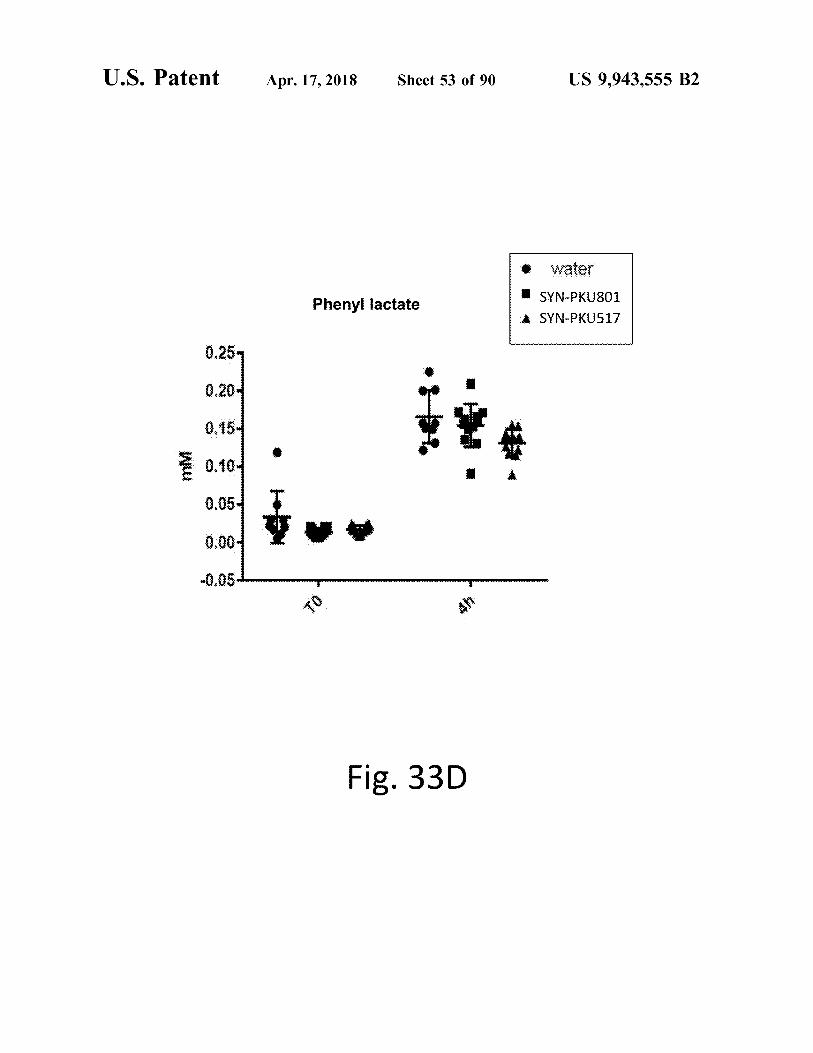
atent Apr . 17 , 2018 Sheet 53 of 90 US 9 , 943 , 555 B2
Water
Phenyl lactate SYN - PKU801 SYN - PKU517
0 . 257 .
0 . 20 4
0 . 15 WW 0 . 10
0 . 05
0 . 00
- 0 . 05
Fig . 33D
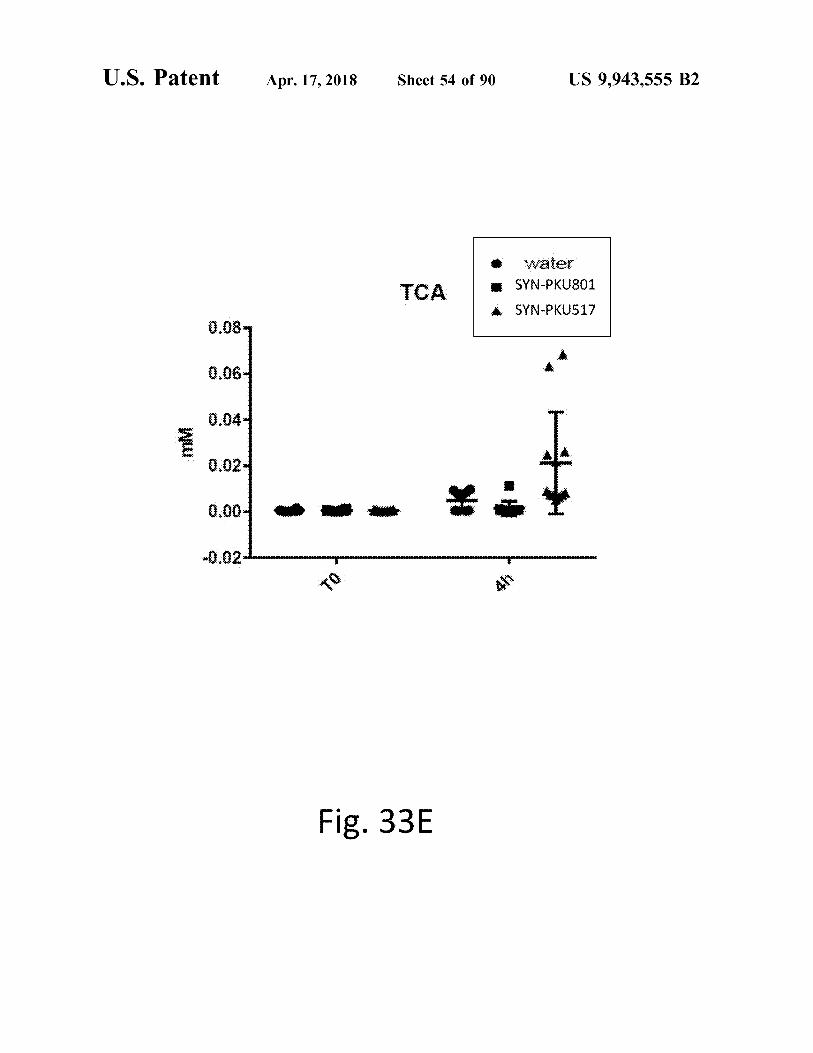
atent Apr . 17 , 2018 Sheet 54 of 90 US 9 , 943 , 555 B2
TCA Water SYN - PKU801 SYN - PKU517
0 . 087 wwwww
0 . 06
Ww 0 . 02
. . . mer * 0 . 004 vivivivivivi 0 . 02
Fig . 33E
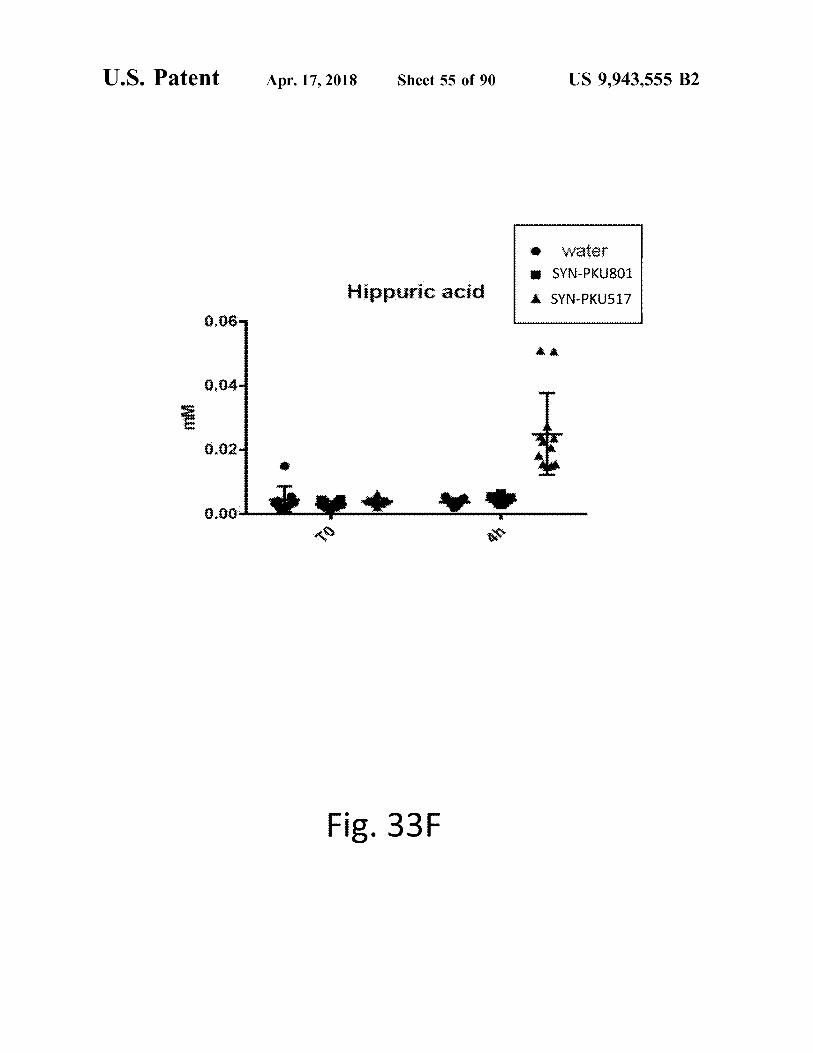
U . S . Patent atent Apr . 17 , 2018 Sheet 55 of 90 US 9 , 943 , 555 B2
Water SYN - PKU801
A SYN - PKU517 Hippuric acid
wu
0 . 00 L
Fig . 33F
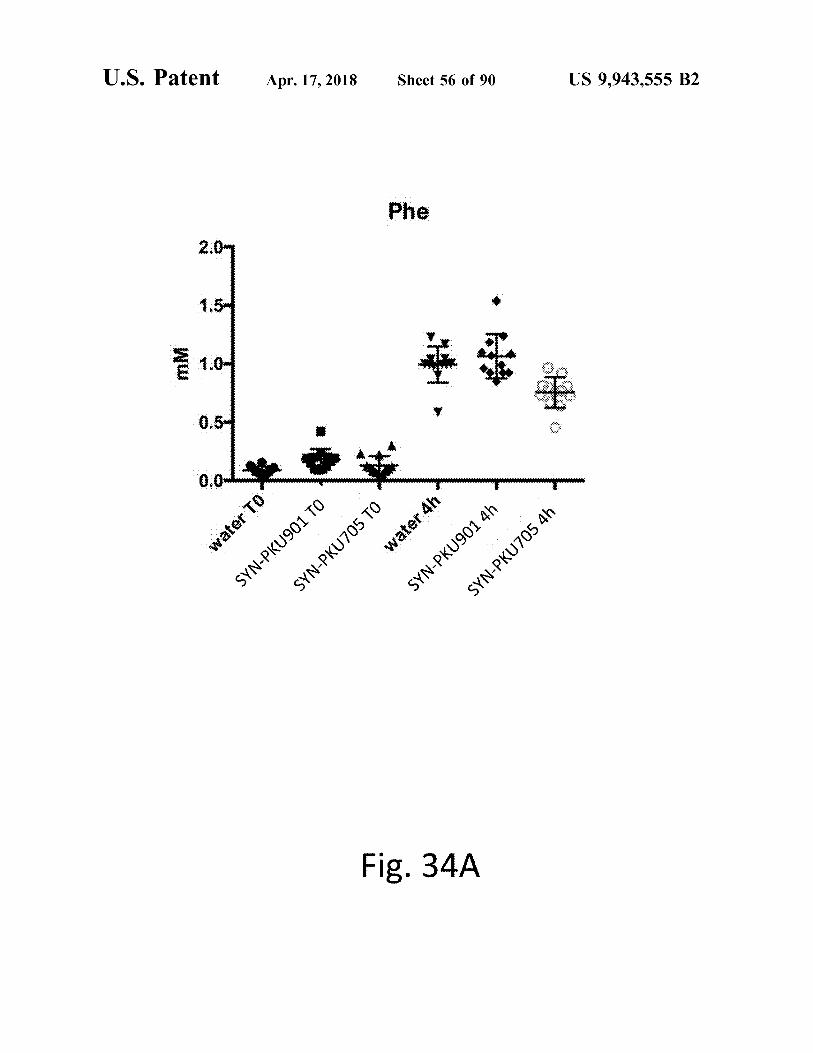
atent Apr . 17 , 2018 Sheet 56 of 90 US 9 , 943 , 555 B2
Phe
? ?i #
mM Se of
F
* water TO water SYN - PKU705 TO water 4h SYN - PKU901 4h SYN - PKU705 4h SYN - PKU901 TO
Fig . 34A
atent Apr . 17 , 2018 Sheet 57 of 90 US 9 , 943 , 555 B2
A [ Phe ]
# mM
water SYN - PKU901 SYN - PKU705
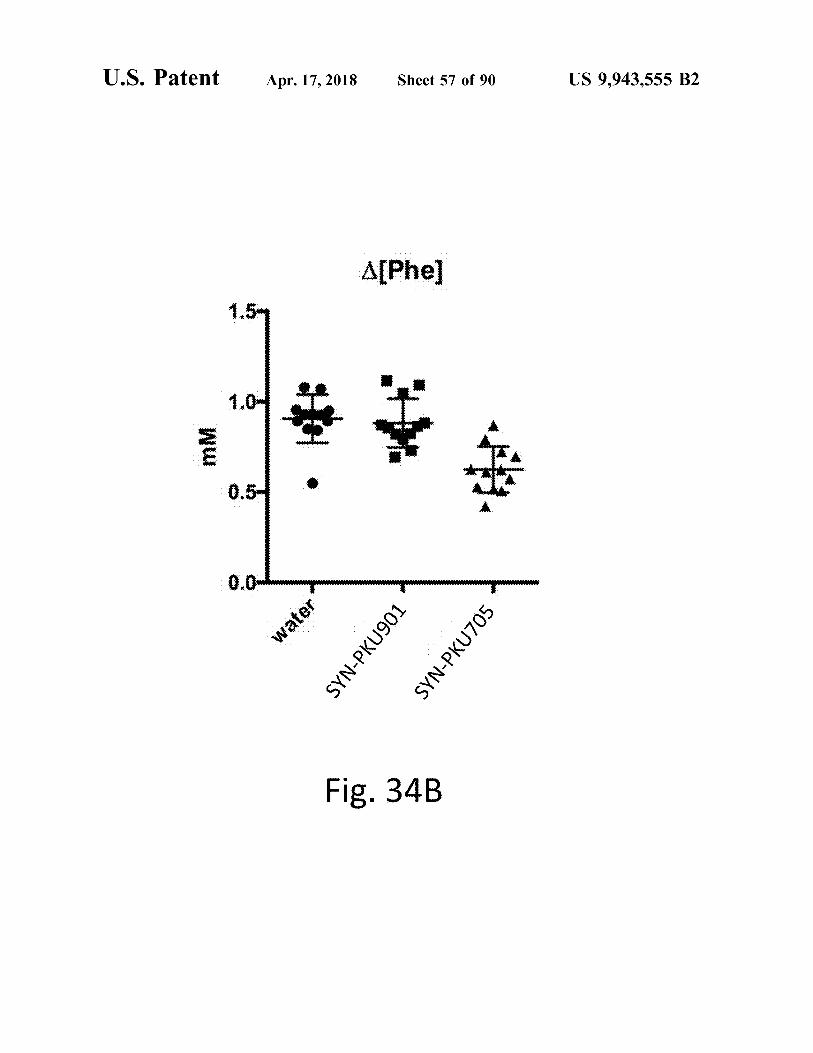
Fig . 34B
atent Apr . 17 , 2018 Sheet 58 of 90 US 9 , 943 , 555 B2
. . . . . . . . . . . . . .
0 . 015
0 . 0104 mm
0 . 005 < * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * . . . . . . . . .
0 . 000
water TO SYN - PKU705 TO SYN - PKU901 4h SYN - PKU705 4h SYN - PKU901 TO
Fig . 340
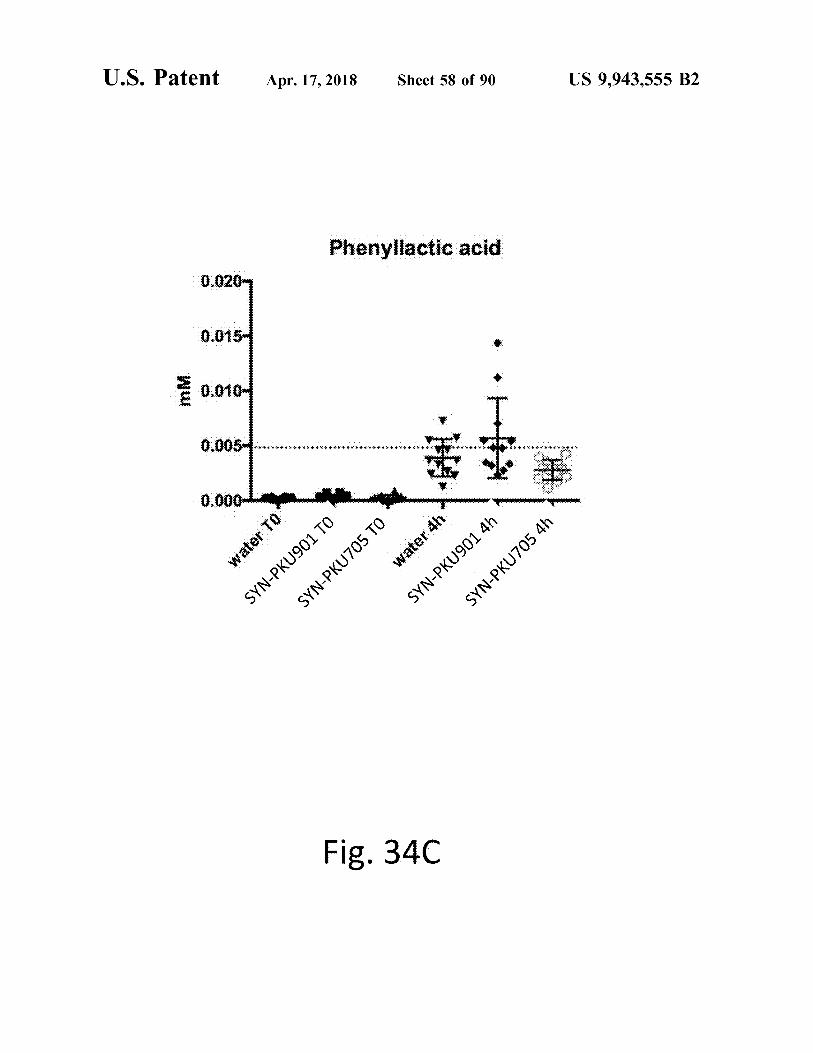
atent Apr . 17 , 2018 Sheet 59 of 90 US 9 , 943 , 555 B2
Phenylpyruvic acid
WW
0 . 0 .
Water TO SYN - PKU901 TO SYN - PKU705 TO
Water 4h SYN - PKU705 4h SYN - PKU901 4h
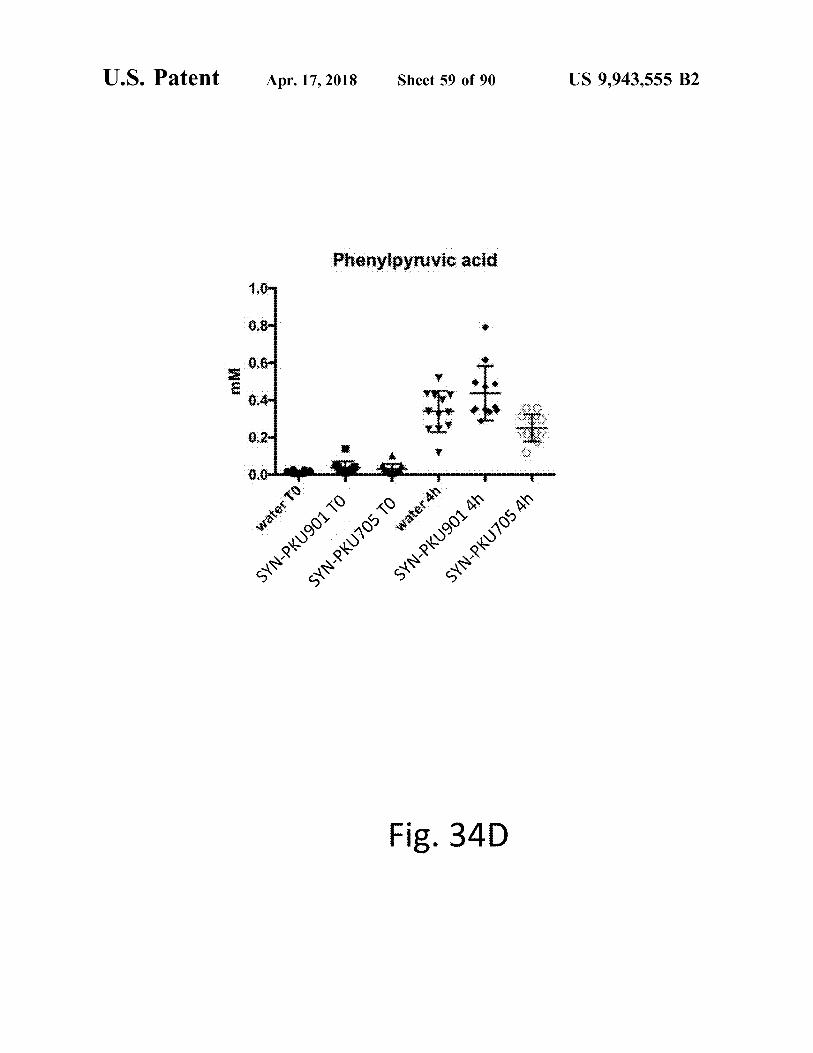
Fig . 34D
atent Apr . 17 , 2018 Sheet 60 of 90 US 9 , 943 , 555 B2
TCA 0 . 015
0 . 010
0 . 005
0 . 000 1111 . . .
Water TO SYN - PKU901 TO . . . .
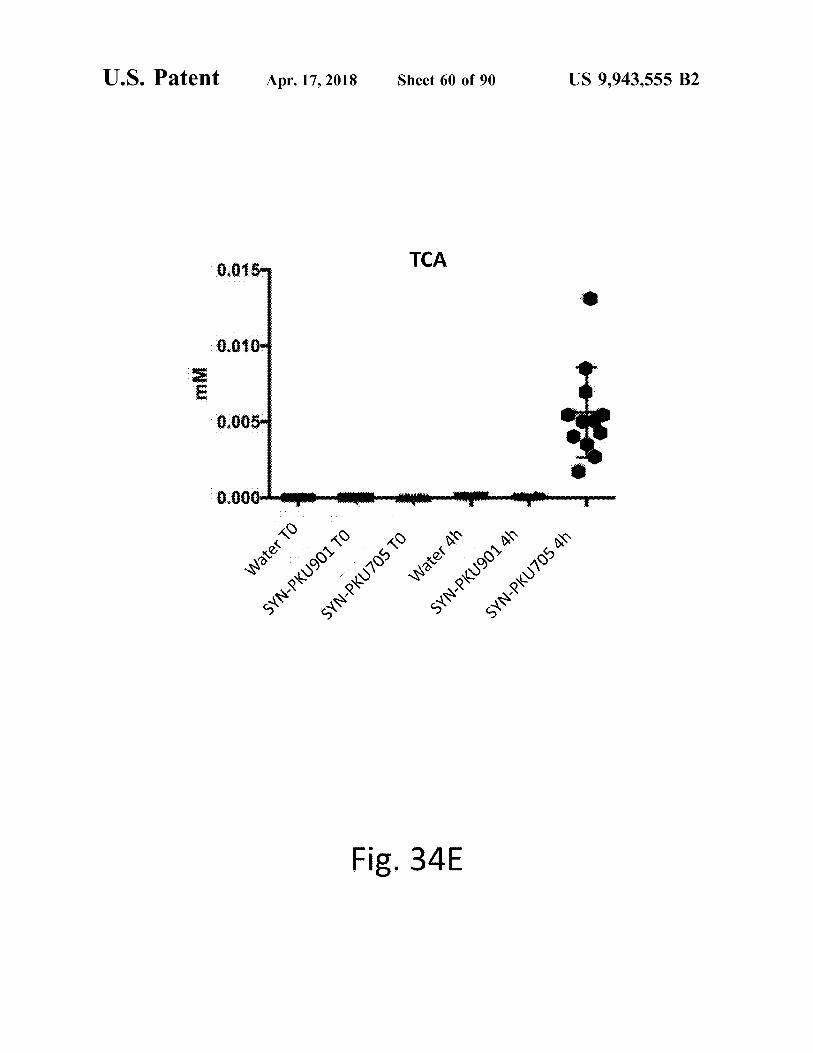
was SYN - PKU705 TO Water 4h SYN - PKU901 4h SYN - PKU705 4h
Fig . 34E
atent Apr . 17 , 2018 Sheet 61 of 90 US 9 , 943 , 555 B2
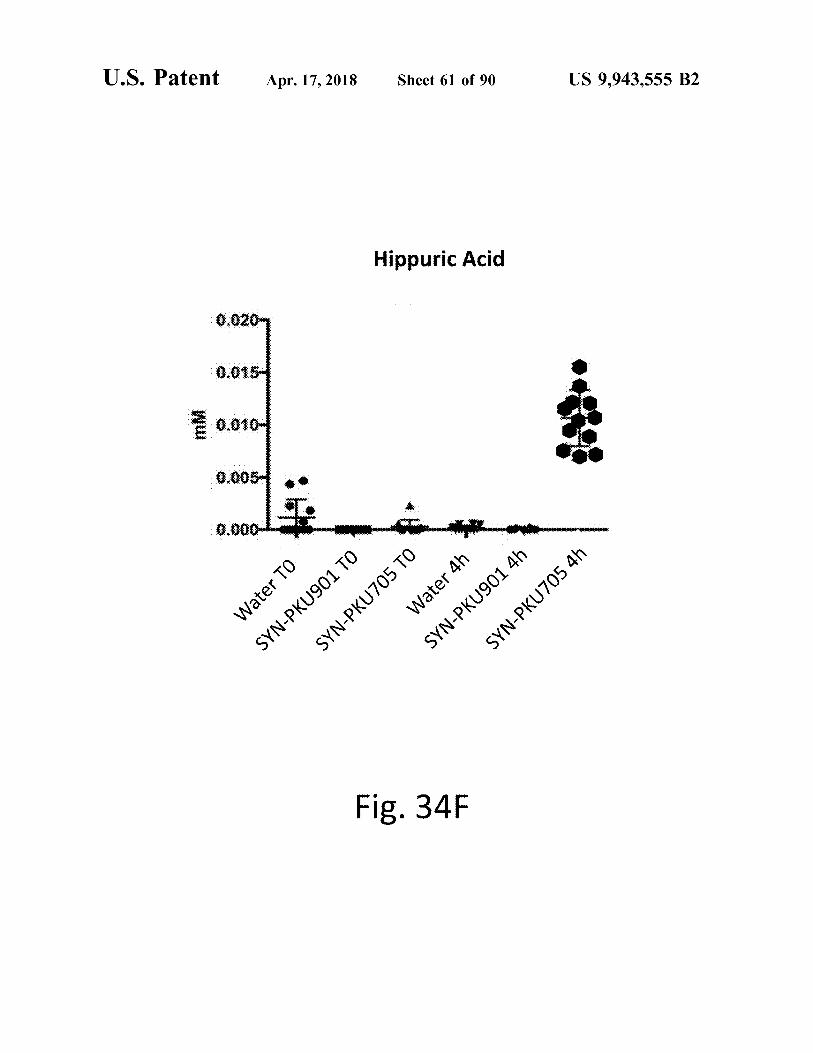
Hippuric Acid
0 . 020 ,
0 . 015
0 . 010
0 . 005
0 . 000
Water TO 40 106nyd - NAS SYN - PKU705 4h Water 4h . SYN - PKU901 TO SYN - PKU 705 TO
Fig . 34F
atent
Statisttintson
styttiin Hotlin
About
retur Het interim that
Whething
NH2 phenylalanine
Apr . 17 , 2018
wwwwwww
probetrotter
Within
??? VENH2 www
Sheet 62 of 90
wwww NH2
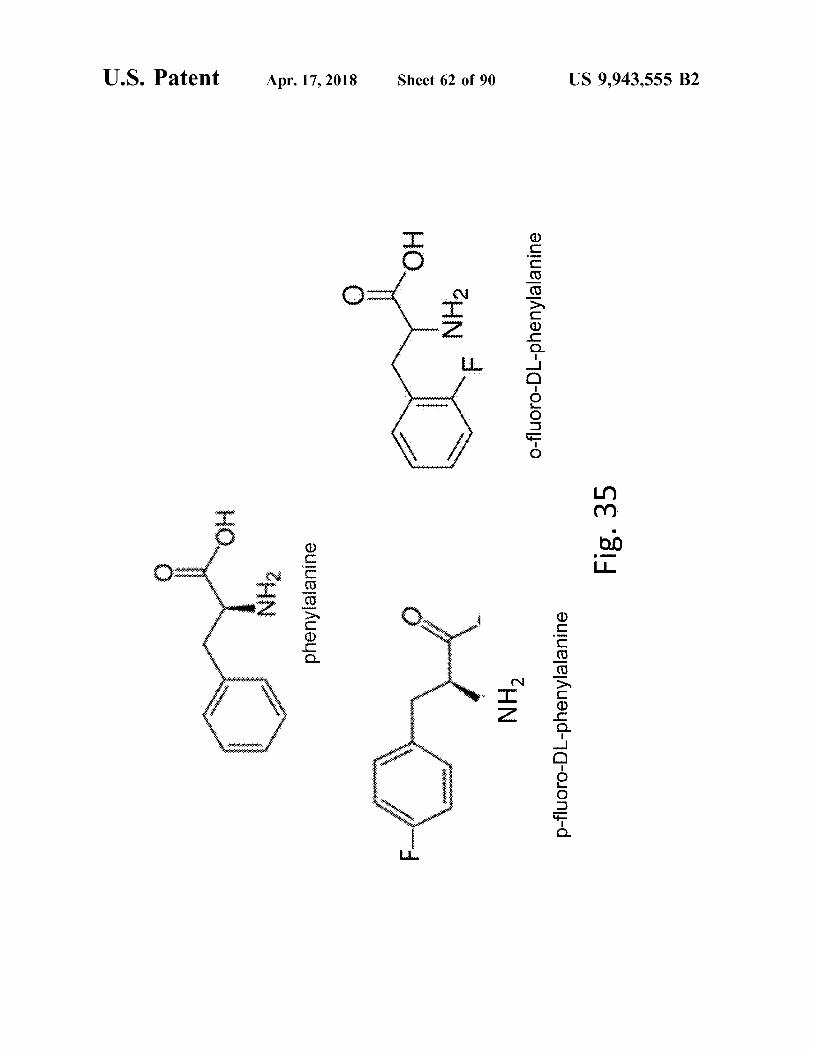
o - fluoro - DL - phenylalanine
p - fluoro - DL - phenylalanine
Fig . 35
US 9 , 943 , 555 B2
araC / BAD
insB / 1
male / K ( ~ 5327k !
( - 4688 kbp )
~ 71 . 4kbp )
atent
lacz ( ~ 448 kbp )
metJ / BL ( ~ 4551 kbp )
argE / C ( ~ 4590kb :
5 . 4 Mb
V5 Mb
dapa 748 kb
5 Mb
rhtC / rhtB 104409 kbp )
hemc / cya ( ~ 4387 kbp )
Apr . 17 , 2018
??? .
1 M6L
( 1141 kbp )
E . coli 1917 Nissle chromosome
- ( - 132kbp )
4 Mb
BPKL A raspekêp ) porno romanesti
Micrones ) proglalkbp )
par monde 3 Mb Fig . 36 ( ~ 330 % Abp )
.
Sheet 63 of 90
mety / argG ( ~ 3671 kbp )
2 Mb
Fig . 36 sediment
US 9 , 943 , 555 B2
atent Apr . 17 , 2018
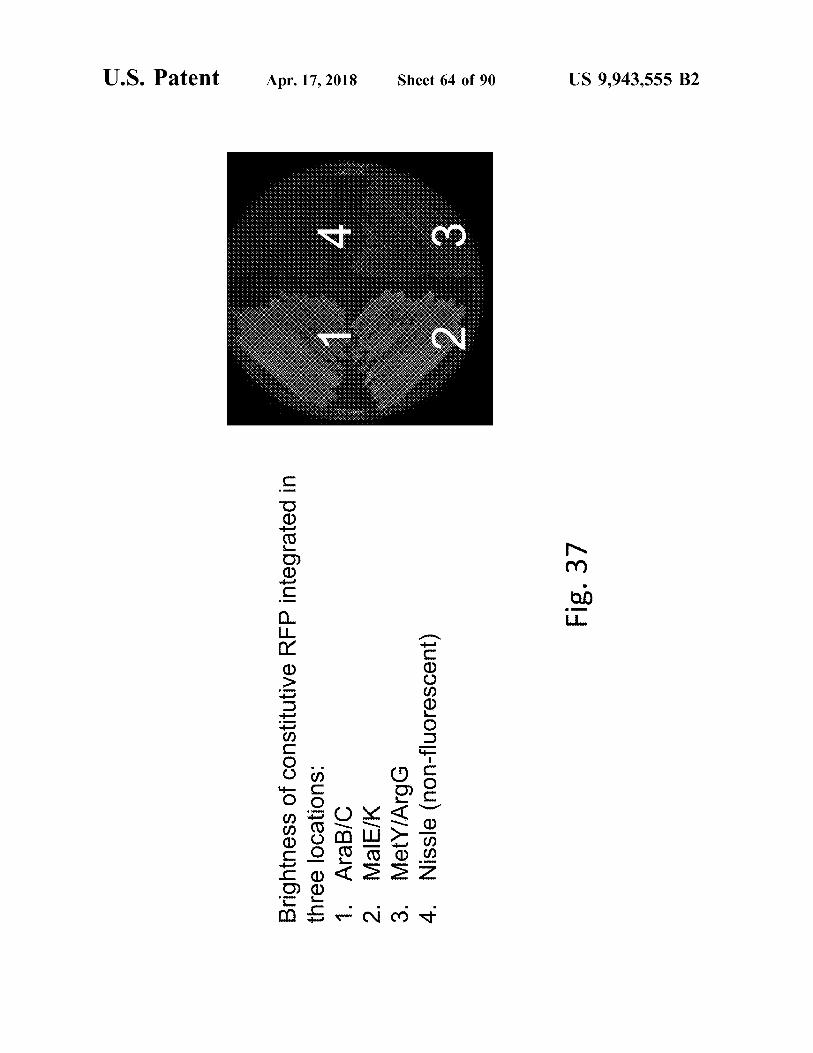
Brightness of constitutive RFP integrated in
three locations : 1 . AraB / C 2 . MalE / K 3 . MetY / ArgG
4 . Nissle ( non - fluorescent )
Sheet 64 of 90
Fig . 37
US 9 , 943 , 555 B2
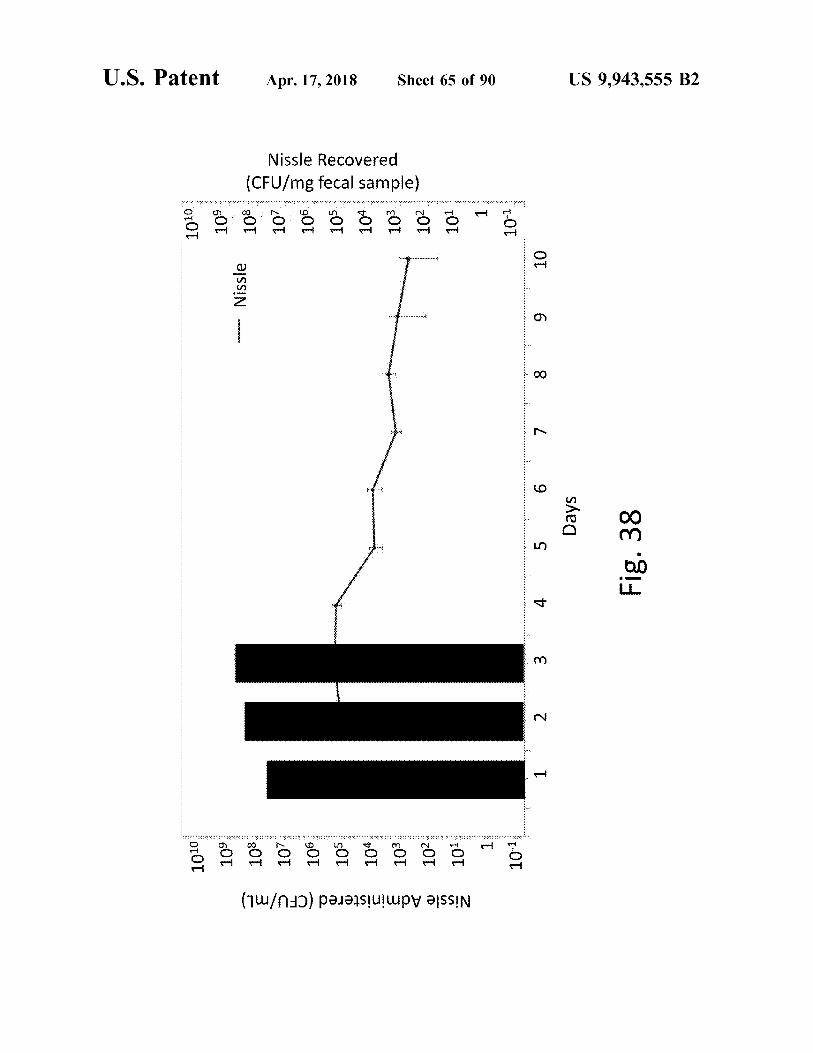
US 9 , 943 , 555 B2
Fig . 38 Days
- 6 8
9 ster
:
…
… .
.
. . . … . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . - - - - - - - - - - - - - - - - - - . . . . . . . . . . . . . . . . . . . . . . . . . . .
iiiiiii
10 - 11
: :
| : :
?
Sheet 65 of 90
: : : : : : :
nonersion
inseconnet
: : : :
www
.
Pierrrrrrrr
:
Nissle Recovered ( CFU / mg fecal sample )
Apr . 17 , 2018
Nissle Administered ( CFU / mL )
: : : : :
. . .
: : : : :
Nissle
: : : : :
wiwiwii
: :
atent
?
or : :
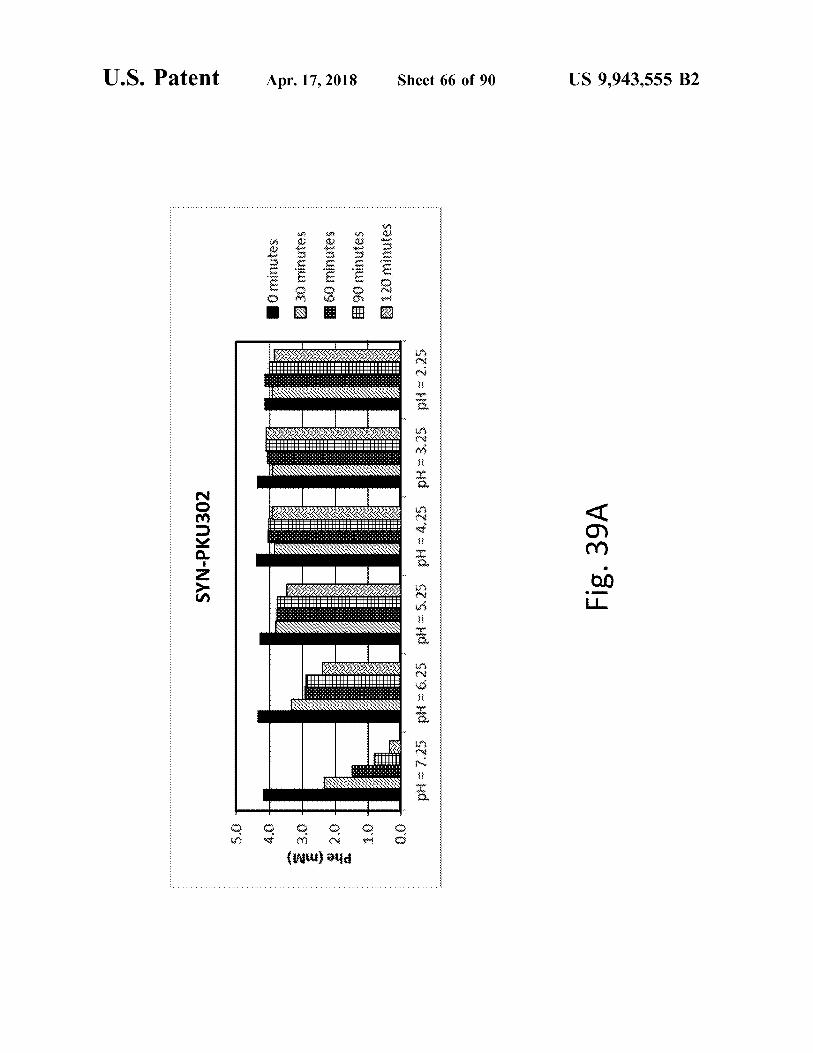
US 9 , 943 , 555 B2
Fig . 39A
Sheet 66 of 90
SC 2 Hd 92€ Hd 925 - Hd ST Sud S29 Hd SZZ = Hd
SainUWOZI
ETU sainuu 062 Sanuju 099
Apr . 17 , 2018
( WW ) aud
SajnuW0€ 2 sajnuw01
atent
SYN - PKU302
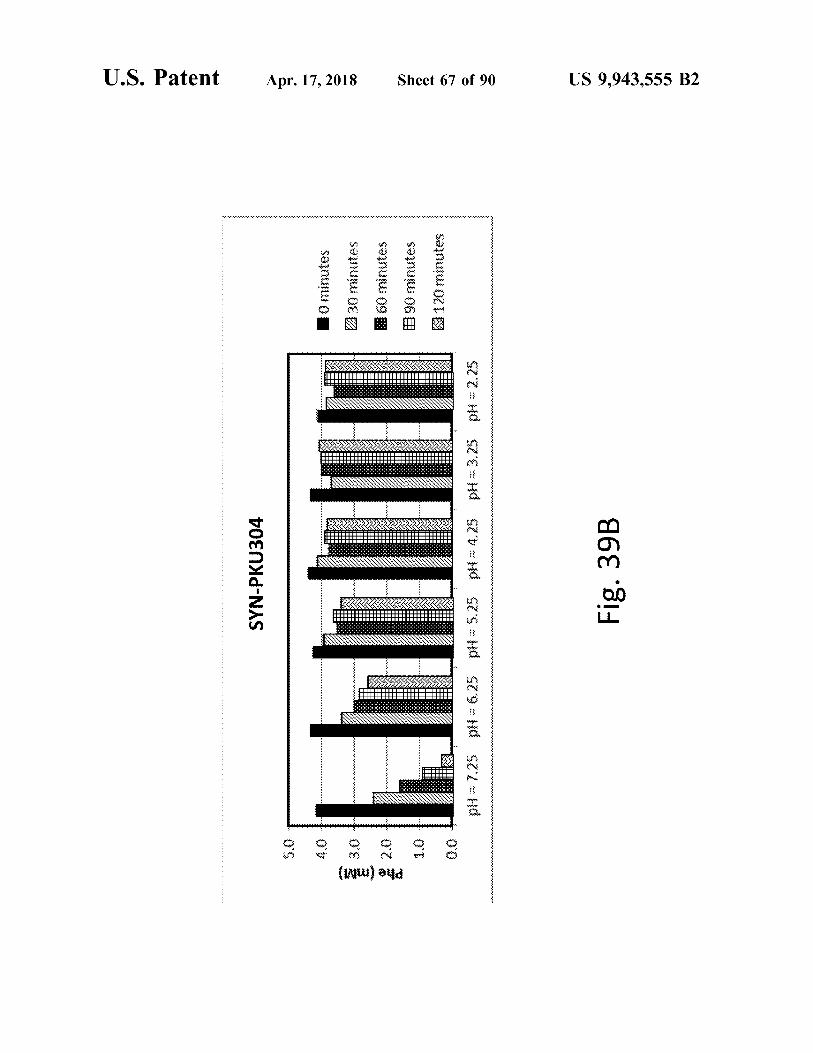
atent
SYN - PKU304
Apr . 17 , 2018
10 minutes
T ILIT
SumUW OE U
T
( WW ) aud
??????????
MMMMMMMMMMM
17111XXXXXXX
IITTITTIIIIIIIII
1111IIIIIIIIIIIII11111
60 minutes 90 minutes 0 120 minutes
Sheet 67 of 90
p
= 7 , 2
tk - 6 . 3
t? -
25
ph? : 4 . 25
ph? : 3 . 2
t? - 3 . 23
Fig . 39B
US 9 , 943 , 555 B2
Synthetic Biotics : Single Product ; Multiple MoAs
atent
MOA 1
Apr . 17 , 2018
MOA |
MOA 2 MOA 2
E . coli 1917 Nissle chromosome
Sheet 68 of 90
MOA 5
MOA 3
MOA 4 Fig . 40
US 9 , 943 , 555 B2
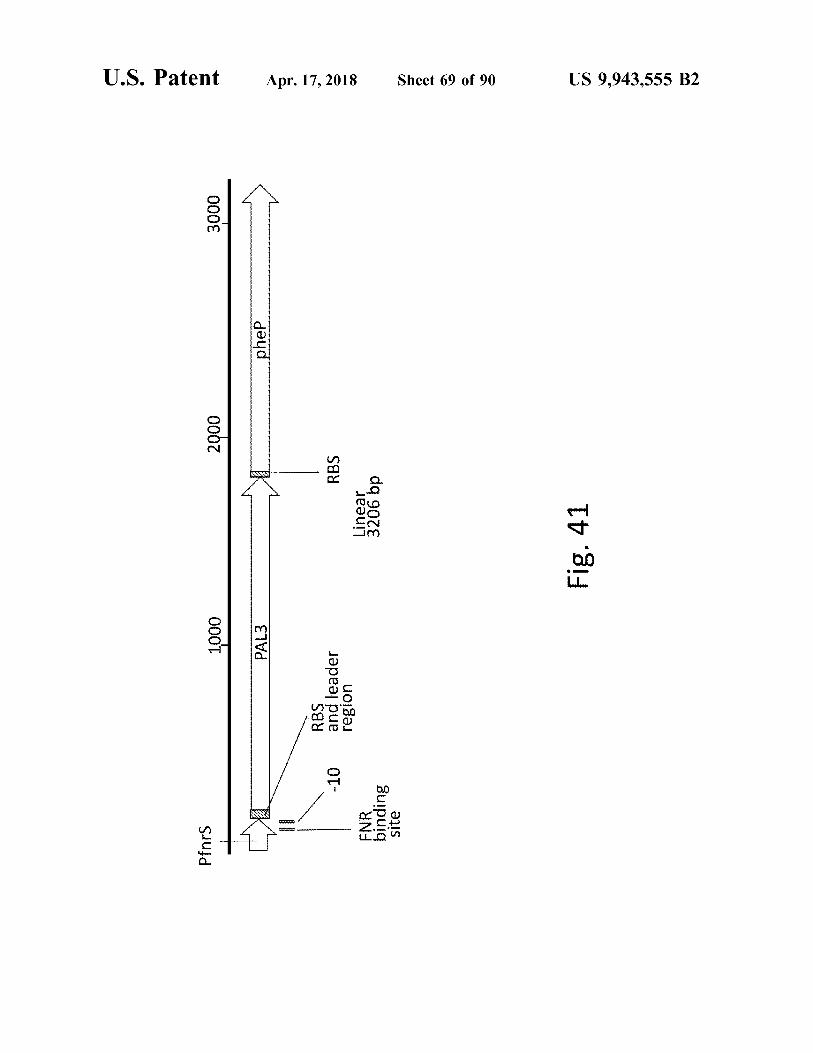
atent
Pfnrs
1000
2000
3000
-
PAL3 PAL3
-
I
_
phep phep
RBS and leader region
Apr . 17 , 2018
RBS Linear 3206 bp
FNR binding site
Sheet 69 of 90
Fig . 41
US 9 , 943 , 555 B2
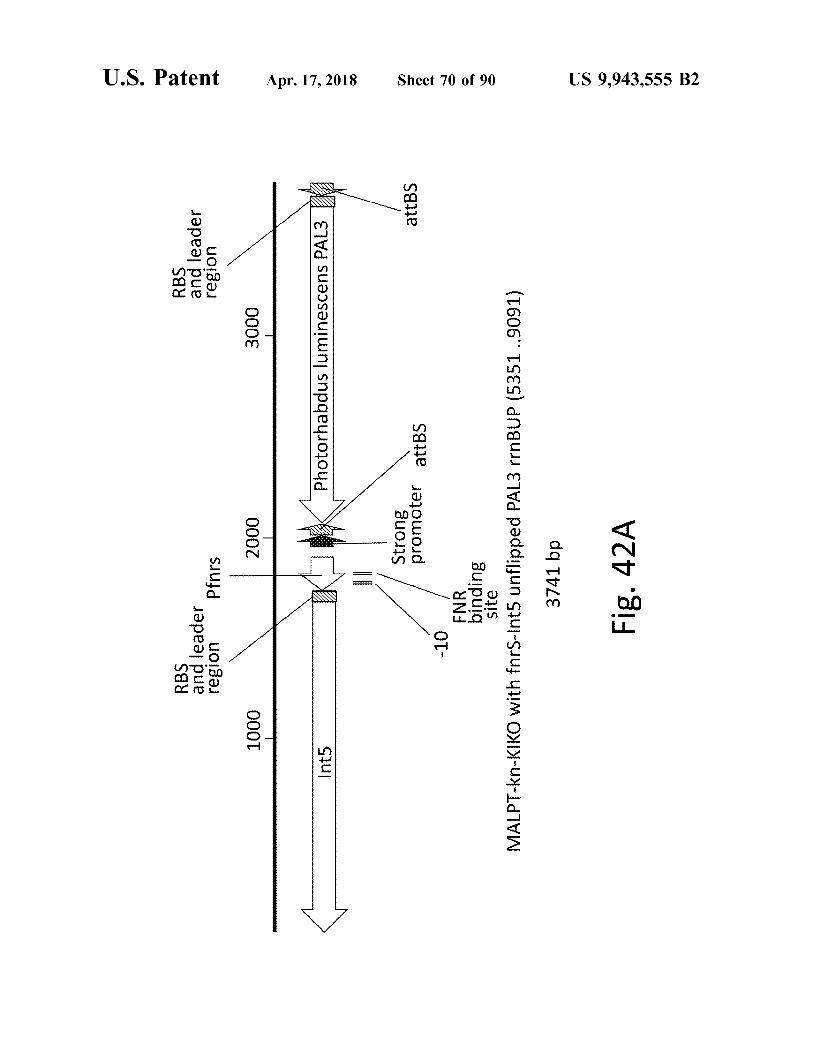
RBS and leader region Pfnrs
2000 2000
RBS and leader region
atent
1000 1000
3000
Apr . 17 , 2018
Int5
Photorhabdus luminescens PAL3
Strong promoter attBS
attBS
- 10
Sheet 70 of 90
FNR binding site MALPT - kn - KIKO with fnrS - Int5 unflipped PAL3 rrnBUP ( 5351 . . 9091 ) 3741 bp Fig . 42A
US 9 , 943 , 555 B2
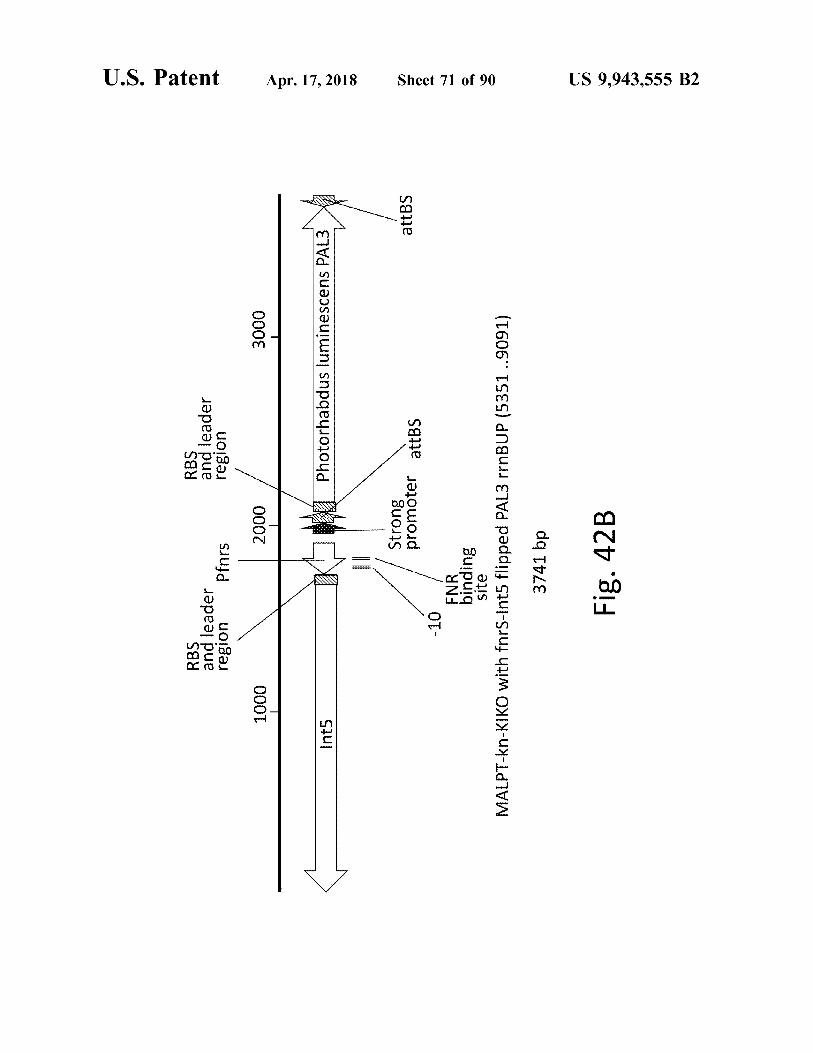
RBS . and leader region Pfnrs
RBS and leader region
atent
1000
2000
3000
Apr . 17 , 2018
Int5 Ints
Photorhabdus luminescens PAL3 X attBS
Strong promoter attbs
- 10
FNR binding site MALPT - kn - KIKO with fors - Int5 flipped PAL3 rrnBUP ( 5351 . . 9091 ) 3741 bp
Sheet 71 of 90
Fig . 42B
US 9 , 943 , 555 B2
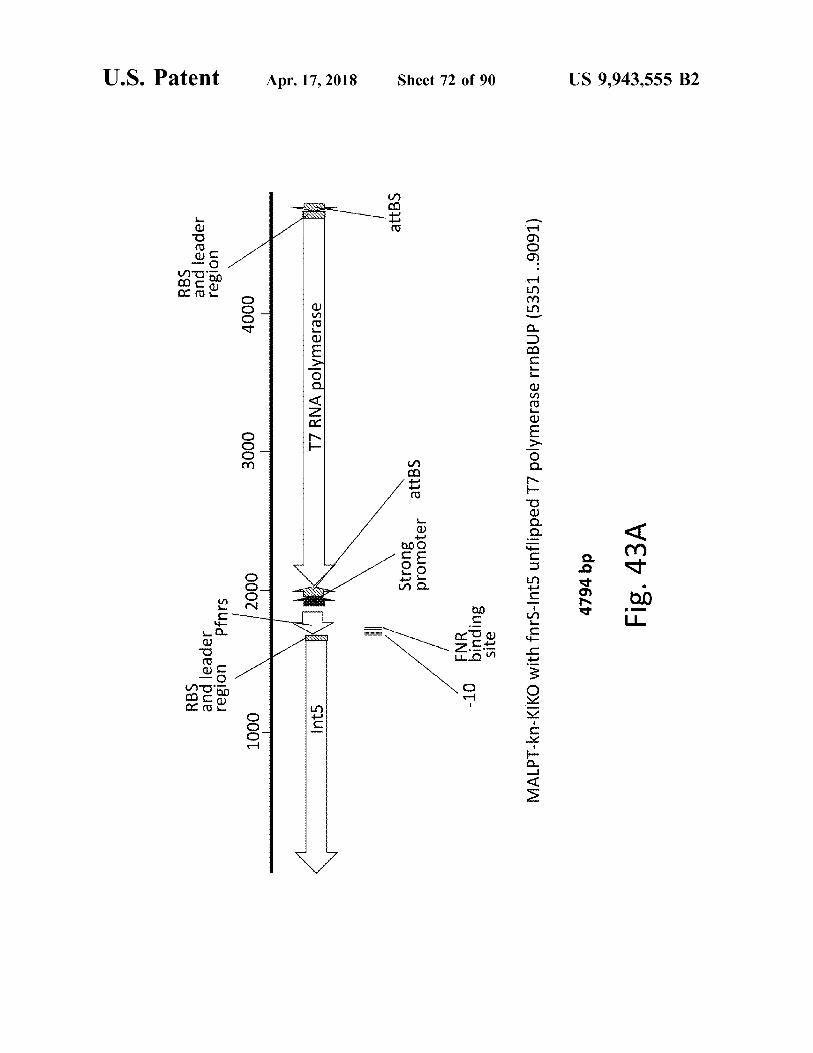
RBS and leader region Pfnrs
e RBS and leader region madera
atent
1000 1000
2000 2000
3000 3000
4000
Int5
T7 RNA polymerase 7 RNA polymerase
Apr . 17 , 2018
attBS
promoter attBS Stronger attBS
Sheet 72 of 90
FNR - 10
binding site
MALPT - kn - KIKO with fors - Int5 unflipped T7 polymerase rrnBUP ( 5351 9091 ) 4794 bp Fig . 43A
US 9 , 943 , 555 B2
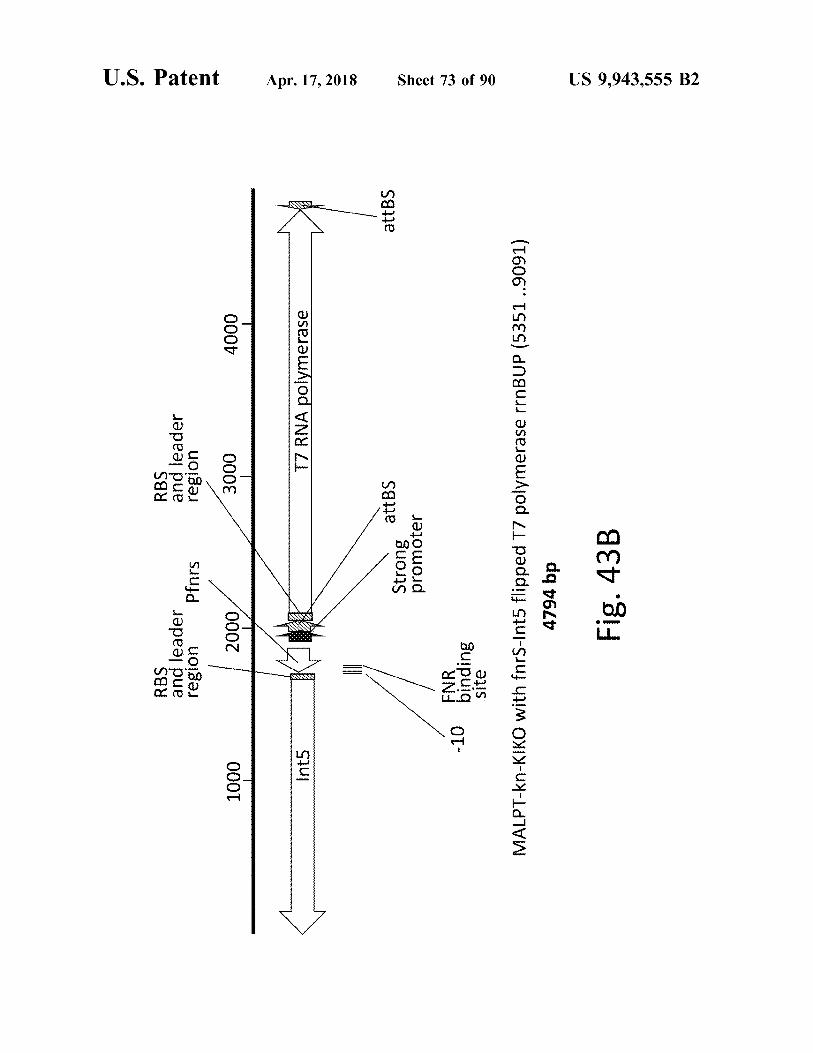
RBS and leader region Pfnrs
RBS and leader region 3000 3000
atent
1000 1000
2000 2000
4000
Int5
T7 RNA polymerase
Apr . 17 , 2018
attes
attBS
Strong promoter
- 10
FNR
Sheet 73 of 90
- 10
binding site
MALPT - kn - KIKO with fors - Int5 flipped T7 polymerase rrnBUP ( 5351 . . 9091 ) 4794 bp Fig . 43B
US 9 , 943 , 555 B2
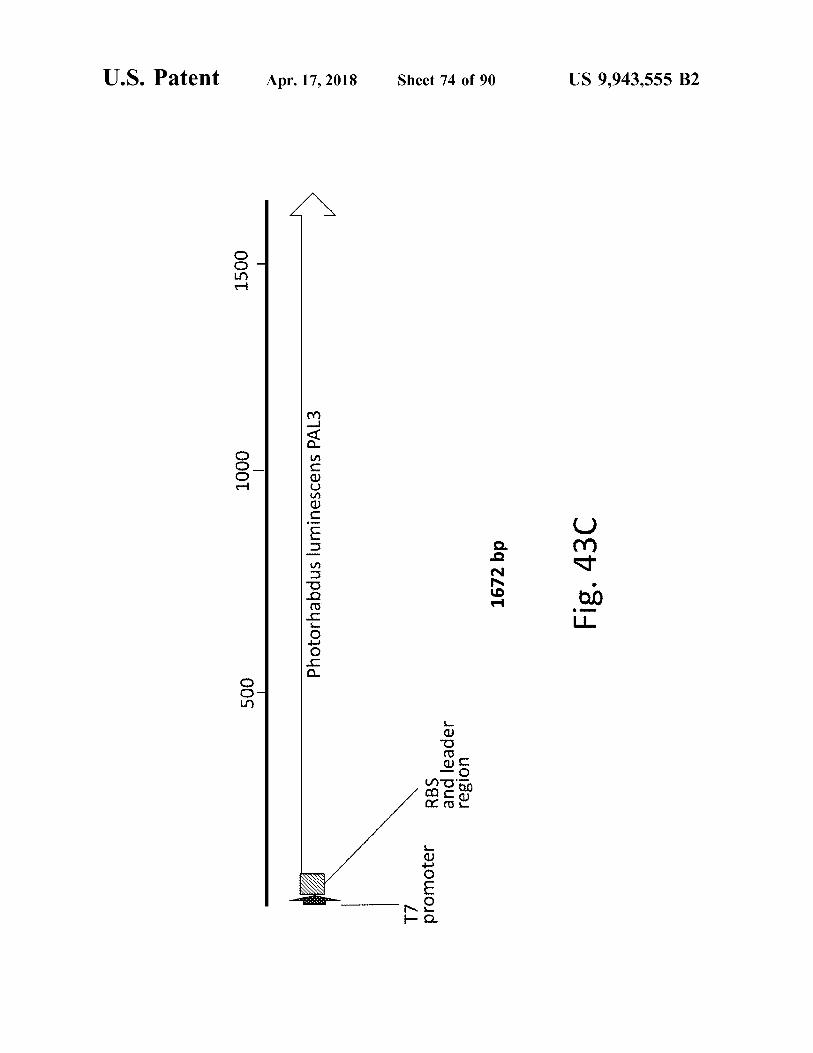
atent
500 500
1000 1000
1500 1500
Photorhabdus luminescens PAL3
Apr . 17 , 2018
T7 promoter RBS and leader region
Sheet 74 of 90
1672 bp Fig . 430
US 9 , 943 , 555 B2
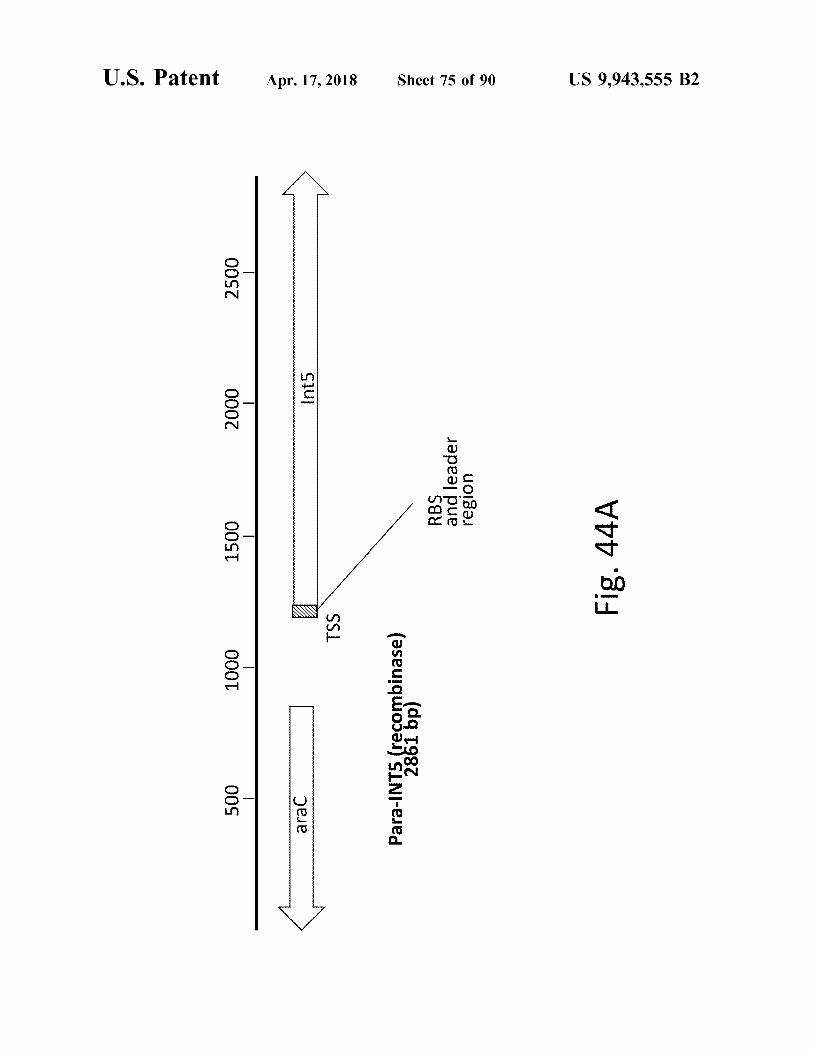
atent
500
1000
1500
2000
2500
ara arac
o
Int5 Ints
Apr . 17 , 2018
TSS
Para - INT5 ( recombinase ) 2861 bp )
RBS and leader region
Sheet 75 of 90
Fig . 44A
US 9 , 943 , 555 B2
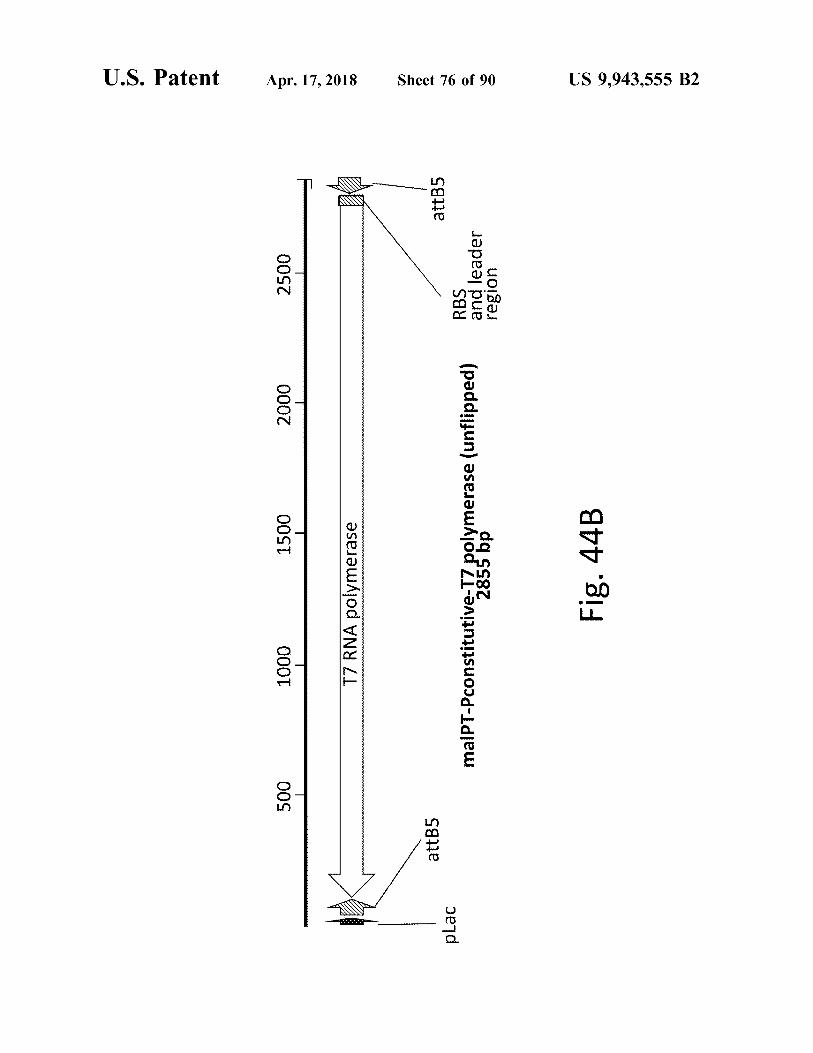
atent
500
1000
1500
2000
2500
Apr . 17 , 2018
T7 RNA polymerase
attB5 attB5 plac
attB5
RBS and leader region
Sheet 76 of 90
malPT - Pconstitutive - T7 polymerase ( unflipped ) 285 Fig . 44B
US 9 , 943 , 555 B2
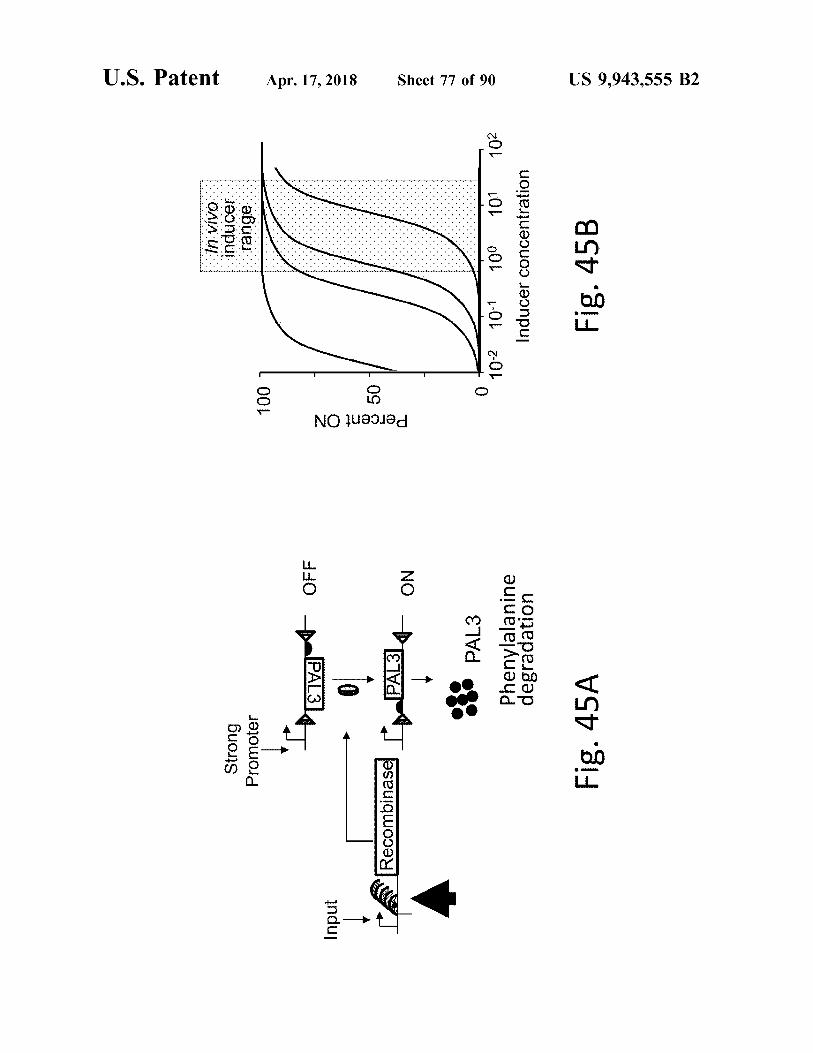
atent
In vivo inducer
Strong Promoter
TTTTTTTTT
range . .
-
-
1001
4
OFF
PAL3
OFF
Apr . 17 , 2018
Input
Percent ON
Reco
nbinase Iba PAL3 / 4
ON
TTTTTTTTTTTTTTT
·
- -
·
Sheet 77 of 90
· · · ·
* * * PAL3
O
+
102
Phenylalanine degradation
10 - 2 10 - 1 100 101 Inducer concentration
Fig . 45A
Fig . 45B
US 9 , 943 , 555 B2
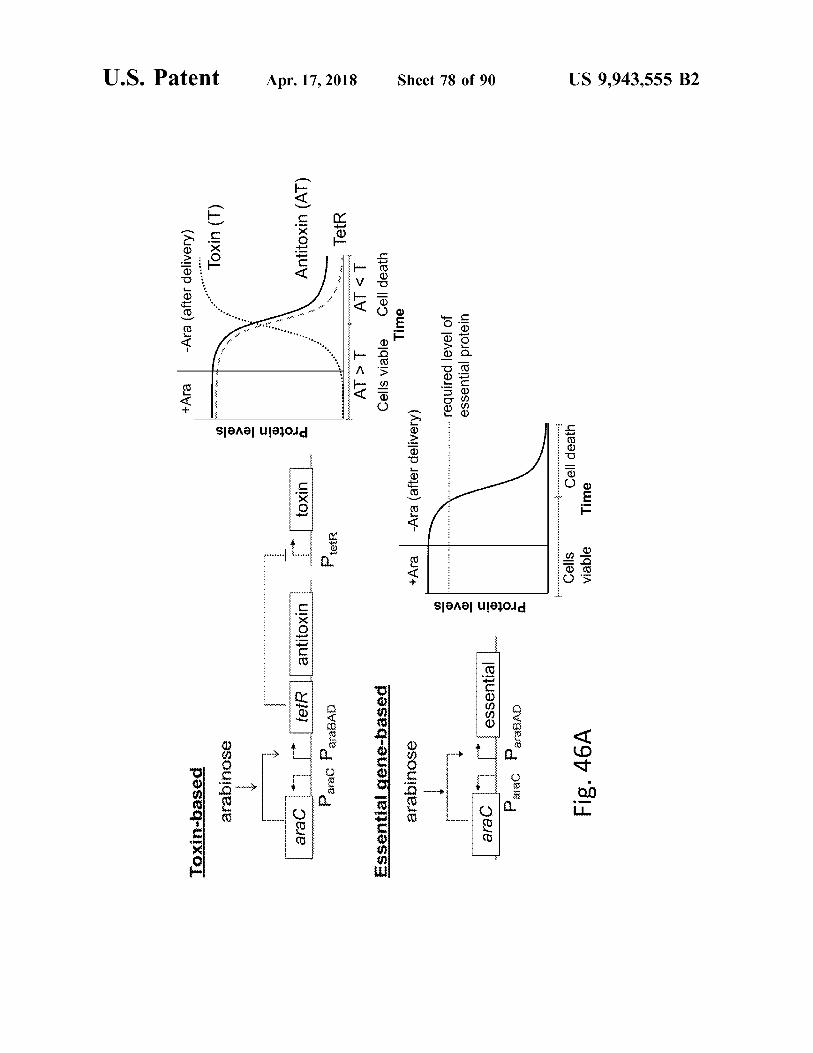
+ Ara
Toxin - based arabinose
Ara ( after delivery )
Toxin ( T )
atent
N
N
002
w
*
* 774 time *
Protein levels
tetR
antitoxin
toxin
* * 1
arac
arac 11 tetrantitoxin | Parac ParaBAD
Antitoxin ( AT )
* * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * *
Apr . 17 , 2018
ter
Preto
* * * *
TetR
wwwwwwwwwwwwwwwwwwwwwwwwwwwww
Essential gene - based arabinose
ATT ATT Cells viable cell death Time
Ara ( after delivery )
+ Ara
Y
.
Sheet 78 of 90
. . .
.
. .
.
. . . .
.
. . . .
.
. . . required level of essential protein
.
| arac 1 essential Parac ParalAD
Protein levels
wwwwwwwwwwwwwwwwwwwww
Fig . 46A
Cells viable
Cell death Time
US 9 , 943 , 555 B2
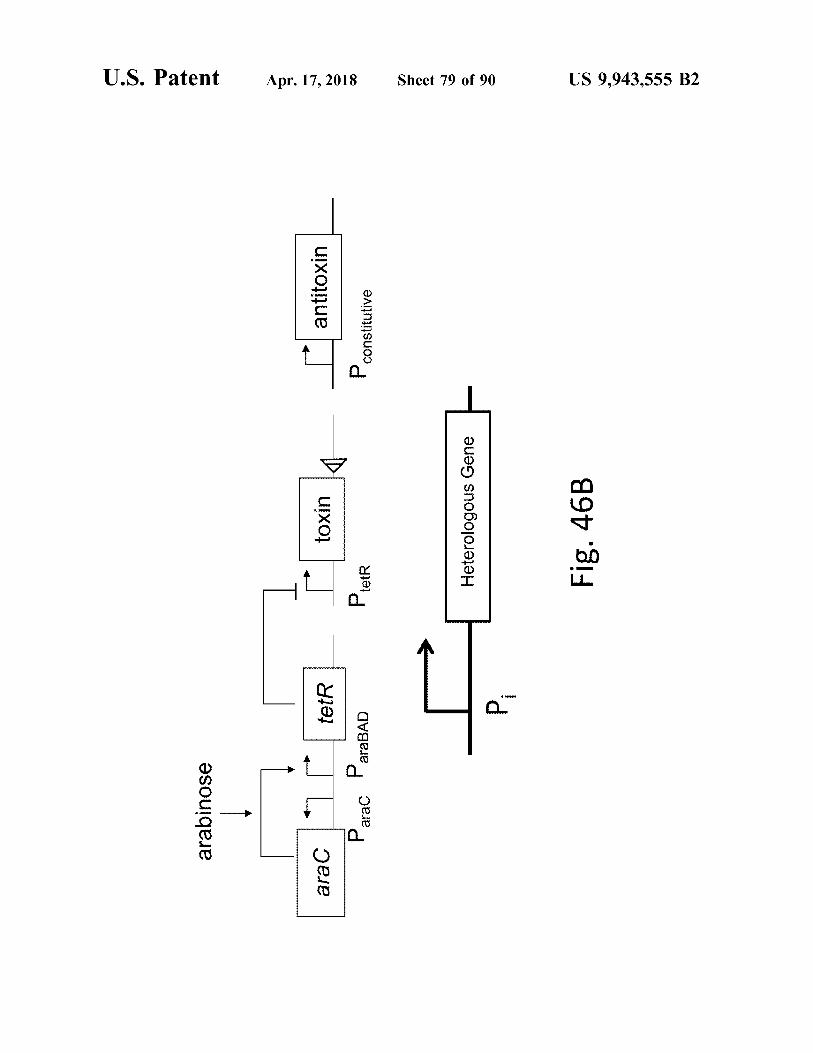
atent Apr . 17 , 2018 Sheet 79 of 90 Sheet 79 of 90 US 9 , 943 , 555 B2
antitoxin filmen )
constitutive
toxin Heterologous Gene Fig . 46B
tetR ranija
se sono le
arabinose tetR
Parac ParaBAD arac
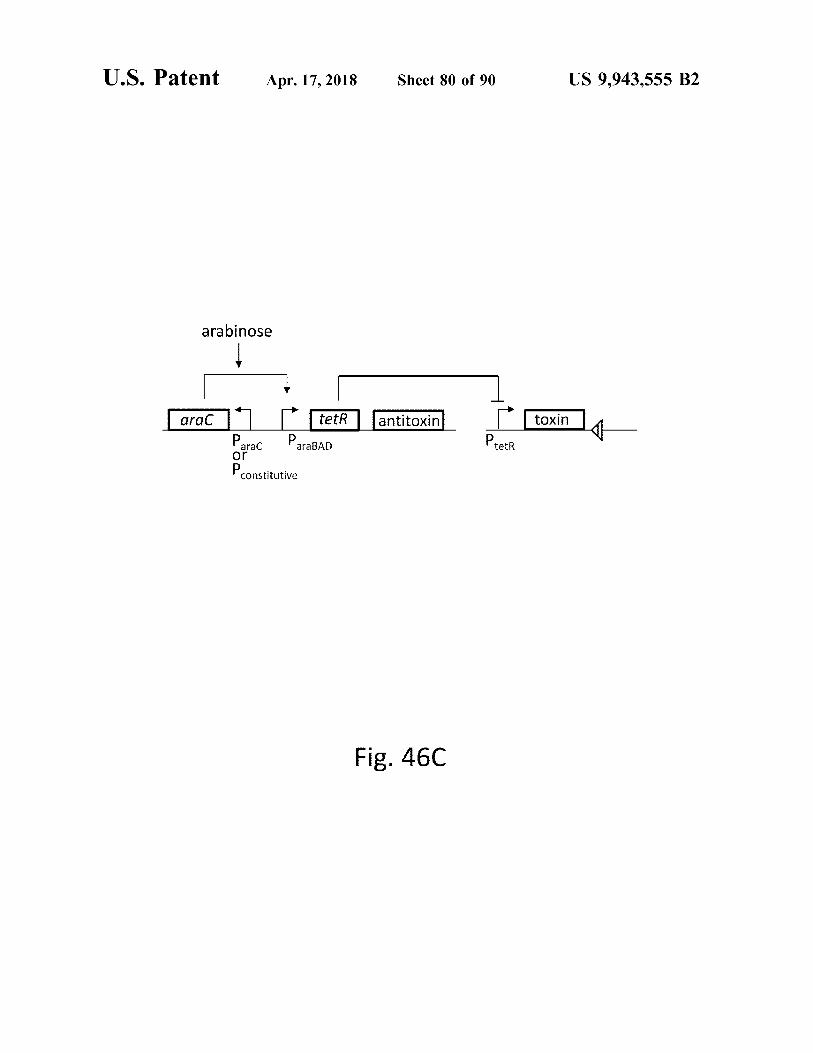
atent Apr . 17 , 2018 Sheet 80 of 90 US 9 , 943 , 555 B2
arabinose
| arac 17 | tetr ] [ antitoxin ] I toxin tetR Para? ParaBAD
or
constitutive
Fig . 46C
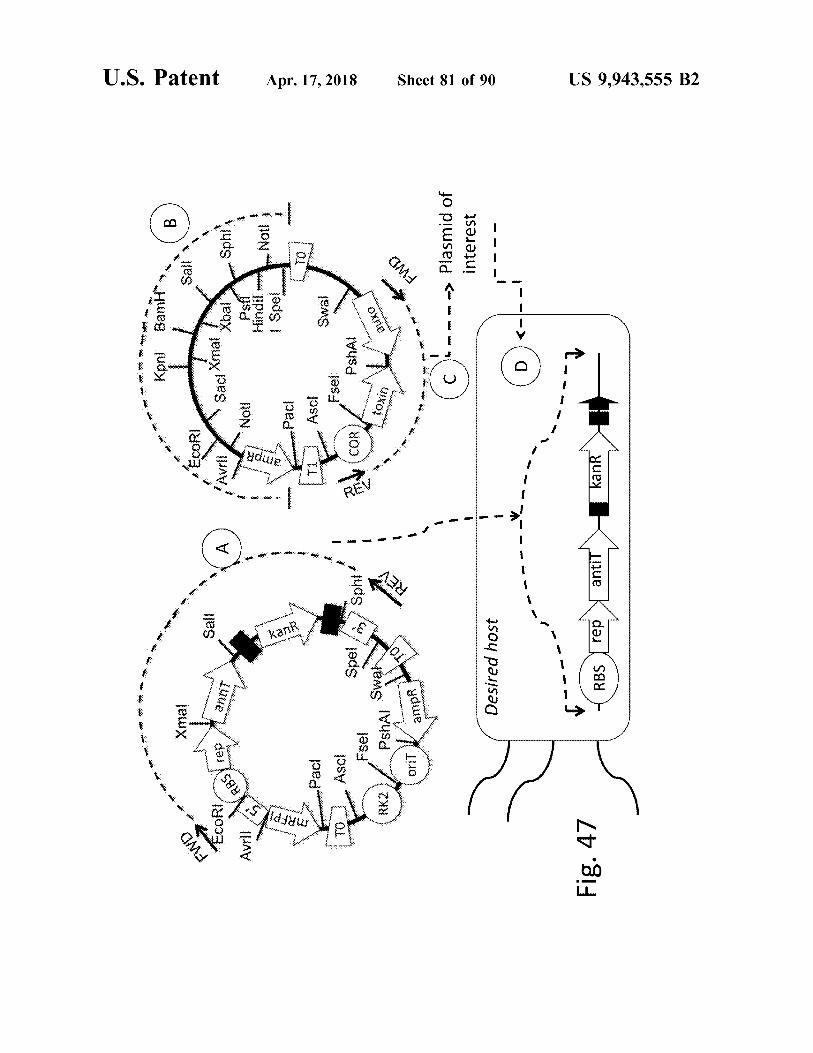
Kpni Bamit
were in the morning
FWD
Xmal
thin der
atent
Sall Sail
ECORI Avril
antt
Avril et
Sphi '
mentioned Not
www
????? Wewe ORI
Xbal Sac Xmal bar
Psti Hindi ( Spel
paci ASCI
Swal
Fisel PsHAI
Notti
dow
kand
TO
i
Paci
Apr . 17 , 2018
minder
TO ) Asct
Fsel pel RK2 ) Psha , Swah
Sphi !
!
* * * * *
toxin
representera
? ??? ???? ????
FWD no es
anipa
con we w
- - - > Plasmid of interest
Sheet 81 of 90
Desired host
were content
Fig . 47 Fig . 47
RBS
rep
antit
RBS repanti
kanR kank
US 9 , 943 , 555 B2
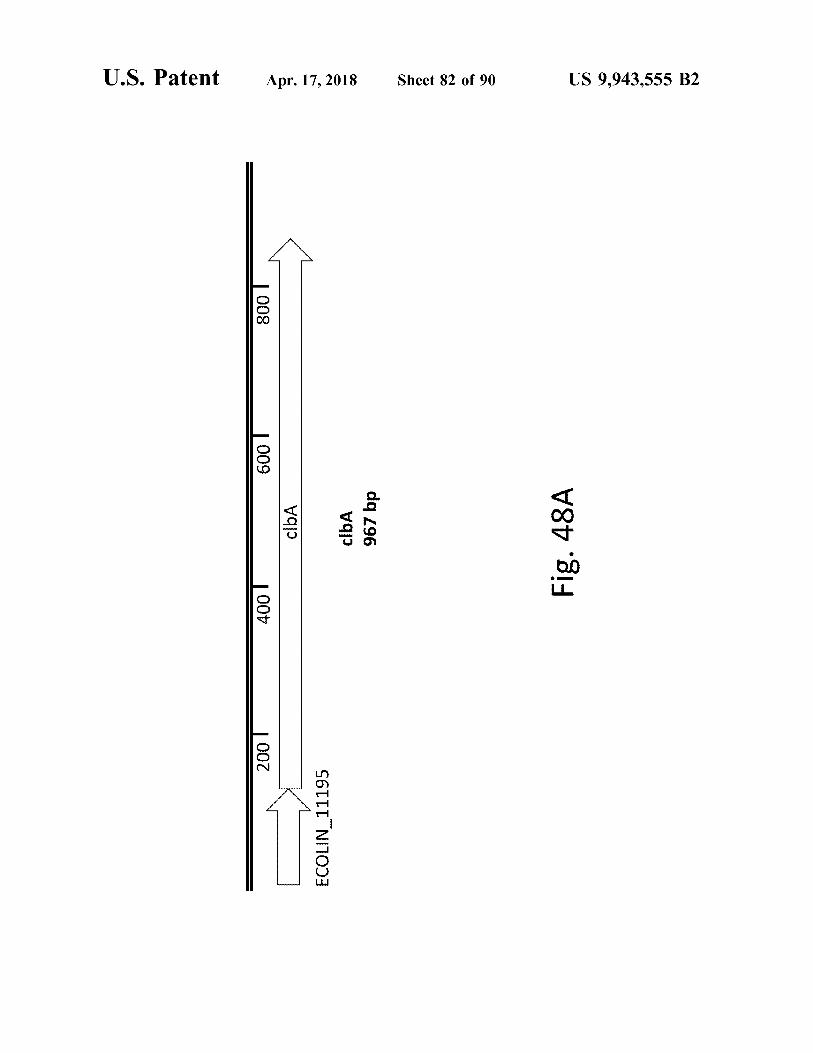
U . S . Patent Apr . 17 , 2018 Sheet 82 of 90 US 9 , 943 , 555 B2
800
600T 6001 clbA clbA 967 bp
Fig . 48A 2001 4001
2001 2001 ECOLIN _ 11195
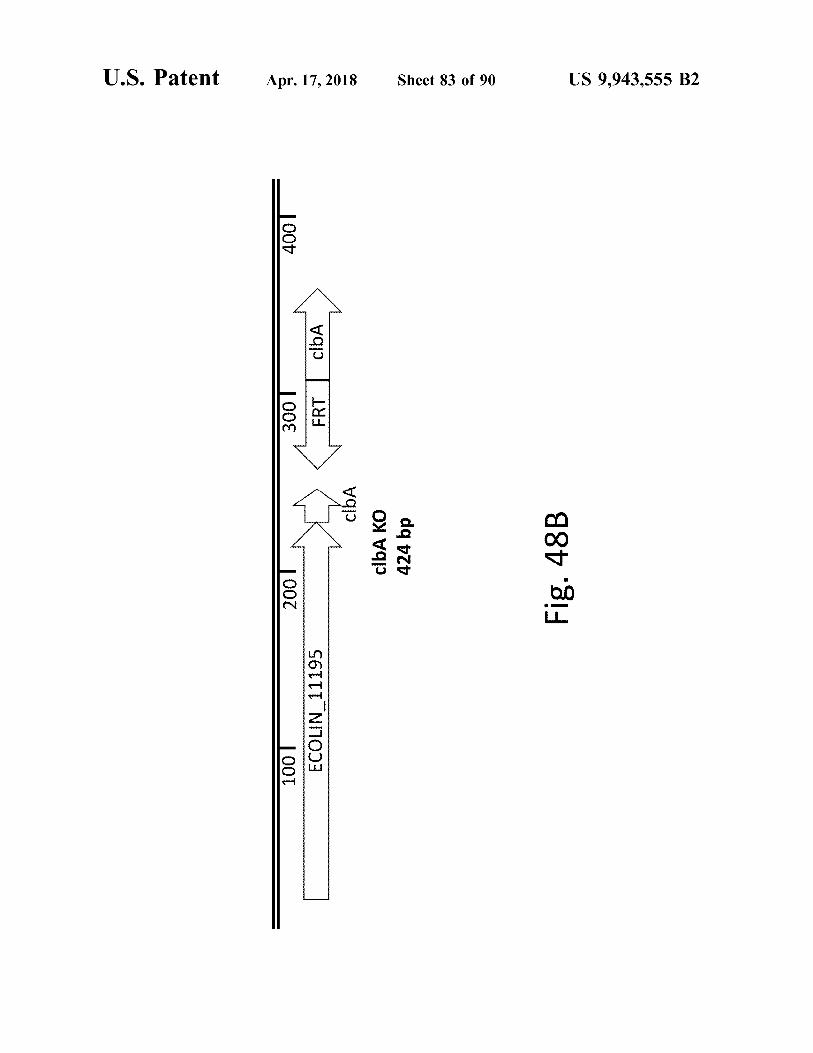
atent
200
3001
400
100 100 ECOLIN 11195
95
200
FRT FRT
clbA clba
Apr . 17 , 2018
clba clbA KO 424 bp
Sheet 83 of 90
Fig . 48B
US 9 , 943 , 555 B2

Wild - type clbA ( SEQ ID NO : 64 )
atent
tac
7
aata
( ?
0 t
(
a
t
t
a
a
c
actt
0 caaatatcacataatcttaacatatcaataaacacagtaaagtttcatgtgaaaaacatcaaaca taaa
ataca ??
taaa
aaacc tgaggattgatatattaattggacatactagtttttttcatcaaaccagtagagataacttcott cactatctcaatgaggaagaaataaaacgctatgatcagtttcattttgtgagtgataaagaact ctatattttaagccgtatcctgctcaaaacagcactaaaaagatatcaacctgatgtctcattac aatcatggcaatttagtacgtgcaaatatggcaaaccatttatagtttttcctcagttggcaaaa aagattttttttaacctttcccatactatagatacagtagccgttgctattagttctcactgega gottggtgtcgatattgaacaaataagagatttagacaactcttatctgaatatcagtcagcatt tttttactocacaggaagctactaacatagtttcacttcctcottatgaaggtcaattacttttt tggaaaatgtggacgctcaaagaagcttacatcaaatatcgaggtaaaggcctatctttaggact ggattotattgaatttcatttaacaaataaaaaactaacttcaaaatatagagattcacctottt atttctctcaatggaaaatatgtaactcatttctcgcattagcctctccactcatcacccctaaa ataactattaagctatttcctatgcagtcccaactttatcaccacgactatcagctaattcatto gtcaaatgggcagaattgaatcgccacggataatctagacacttctgagccgtcgataatattga ttttcatattccgtcggtggtgtaagtatcccgcataatcgtgccattcacatttag Apr . 17 , 2018
clbA knockout ( SEQ ID NO : 65 )
Sheet 84 of 90
aaaaaac a
O
at
rt
* Cat t
c
a
a t
a
tt a
a t
ot C
t
c
t
cta
G
T
G
??? ggatggggggaaacatggataagttcaaagaaaaaaacccgttatctctgcgtgaaagacaagta ???? ttgcgcatgctggcacaaggtgatgagtactctcaaatatcacataatcttaacatatcaataaa cacagtaaagtttcatgtgaaaaacatcaaacataaaatacaagctcggaatacgaatcacgcta tacacattgctaacaggaatgagattatctaaatgaggattgaTGTGTAGGCTGGAGCTGCTTCG AAGTTCCTATACTTTCTAGAGAATAGGAACTTCGGAATAGGAACTTCGGAATAGGAACTAAGGAG GATATTCATATGtcgtcaaatgggcagaattgaatcgccacggataatctagacacttctgagco gtcgataatattgattttcatattccgtcggtgg G
T
A
Fig . 49
US 9 , 943 , 555 B2
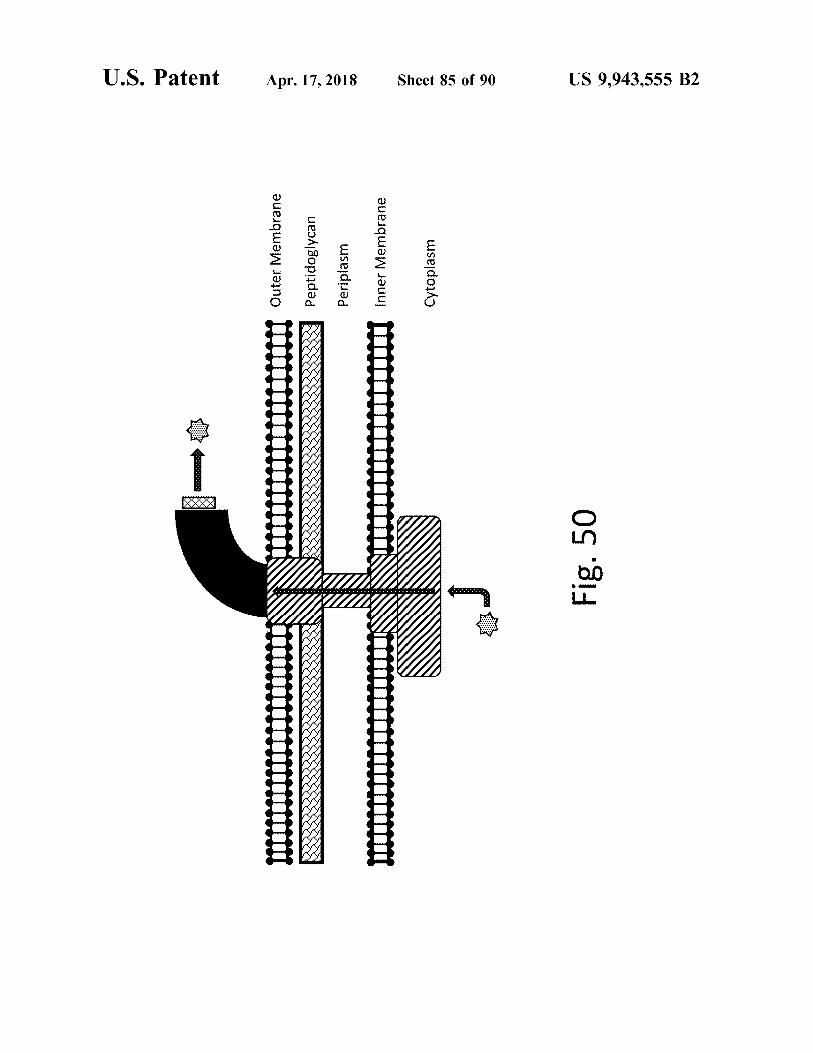
atent Apr . 17 , 2018 Sheet 85 of 90 US 9 , 943 , 555 B2
i Outer Membrane Outer Membrane Peptidoglycan Periplasm Inner Membrane Cytoplasm
1
Fig . 50
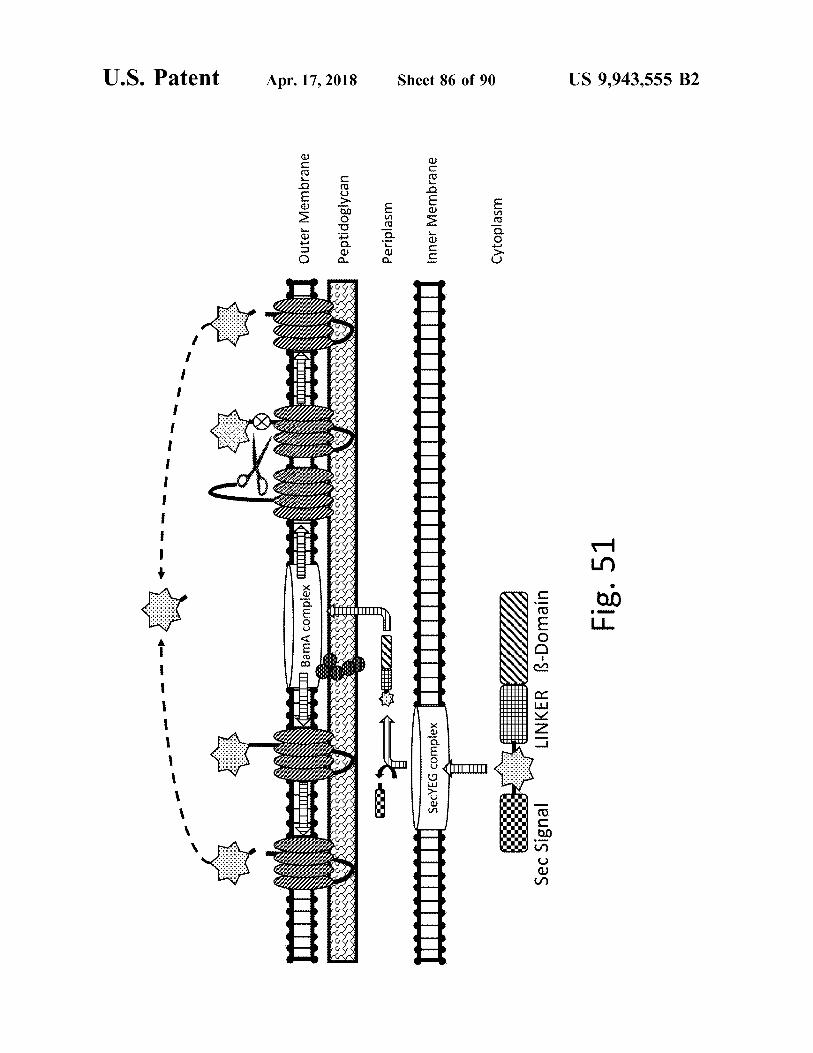
- - - - - - -
atent
??? BamA complex ????
Outer Membrane
Apr . 17 , 2018
|
Peptidoglycan
??III
Periplasm
H
I
I
SecYEG complex
Mit
SecYEG complex
Inner Membrane Inner Membrane
Sheet 86 of 90
Cytoplasm
Sec Signal
LINKER B - Domain
Fig . 51
US 9 , 943 , 555 B2
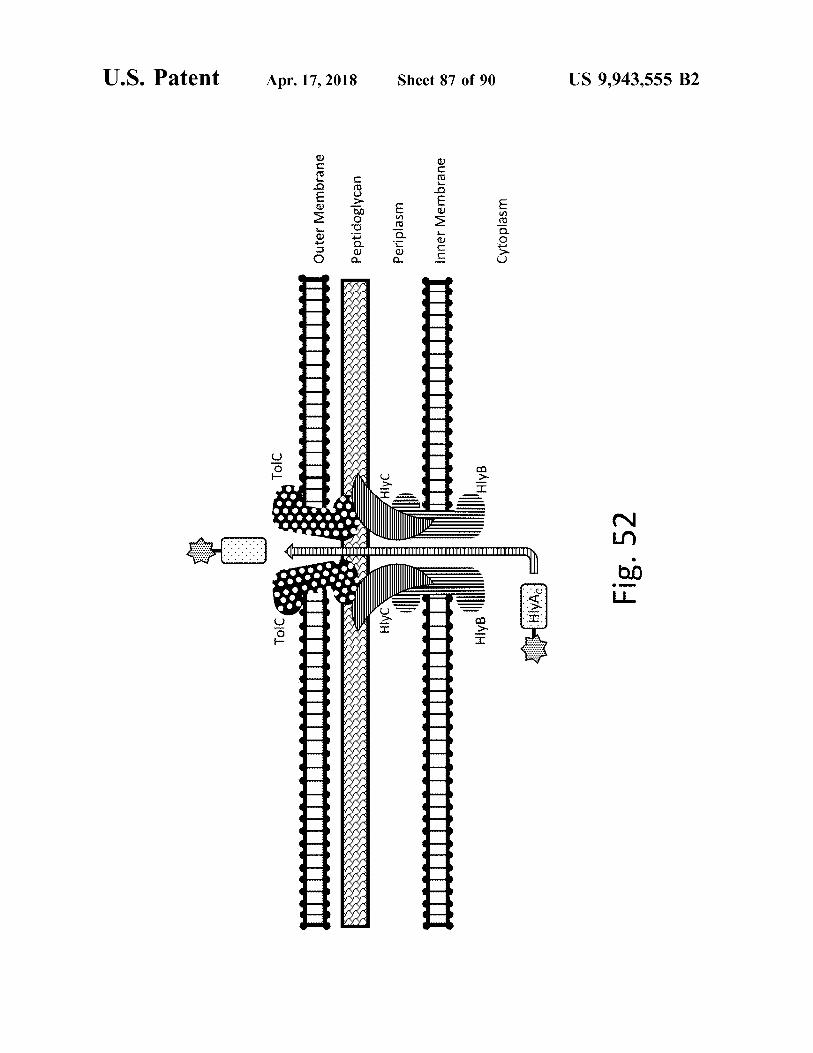
atent
TOIC
Tolco W
Outer Membrane 1111 Outer Membrane
Apr . 17 , 2018
ny
Peptidoglycan
Hlyc
Hlyc
Periplasm
null11111111111111111 1
Inner Membrane Inner Membrane
Sheet 87 of 90
Hl??
' HlyB
Cytoplasm
HIYA : 5 Fig . 52
US 9 , 943 , 555 B2
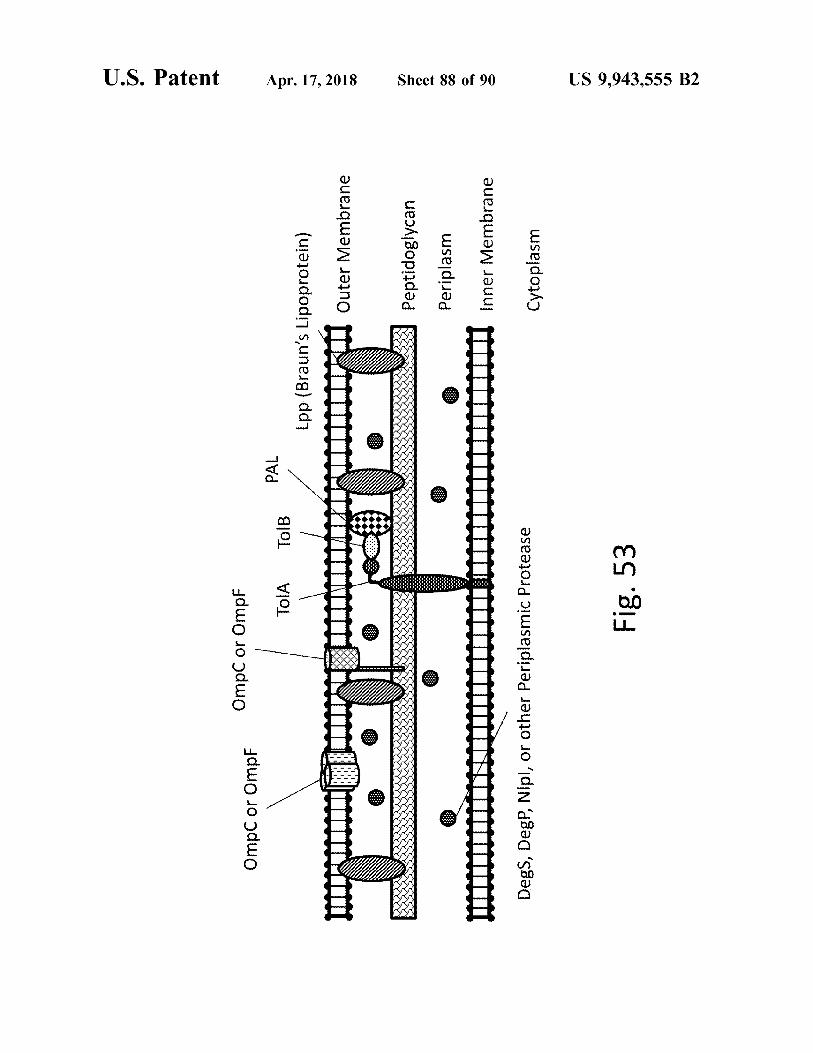
atent
Omocor Omot
Ompc or Ompt
PAL
TOIA
TOIB
.
Lpp ( Braun ' s Lipoprotein )
Outer Membrane
Apr . 17 , 2018
NUORIL I
Outer Membrane
P
o
1
S
VVVVV
peptidoglycan Peptidoglycan Periplasm Periplasm
0 0
mutatni mutiti W
Sheet 88 of 90
IU Inner Membrane
Degs , DegP , Nlpl , or other Periplasmic Protease
Cytoplasm
Fig . 53
US 9 , 943 , 555 B2
atent Apr . 17 , 2018 Sheet 89 of 90 US 9 , 943 , 555 B2
DEFINE
RE?? DESIGN INTEGRATE ???? sic BUILD
E
Fig . 54
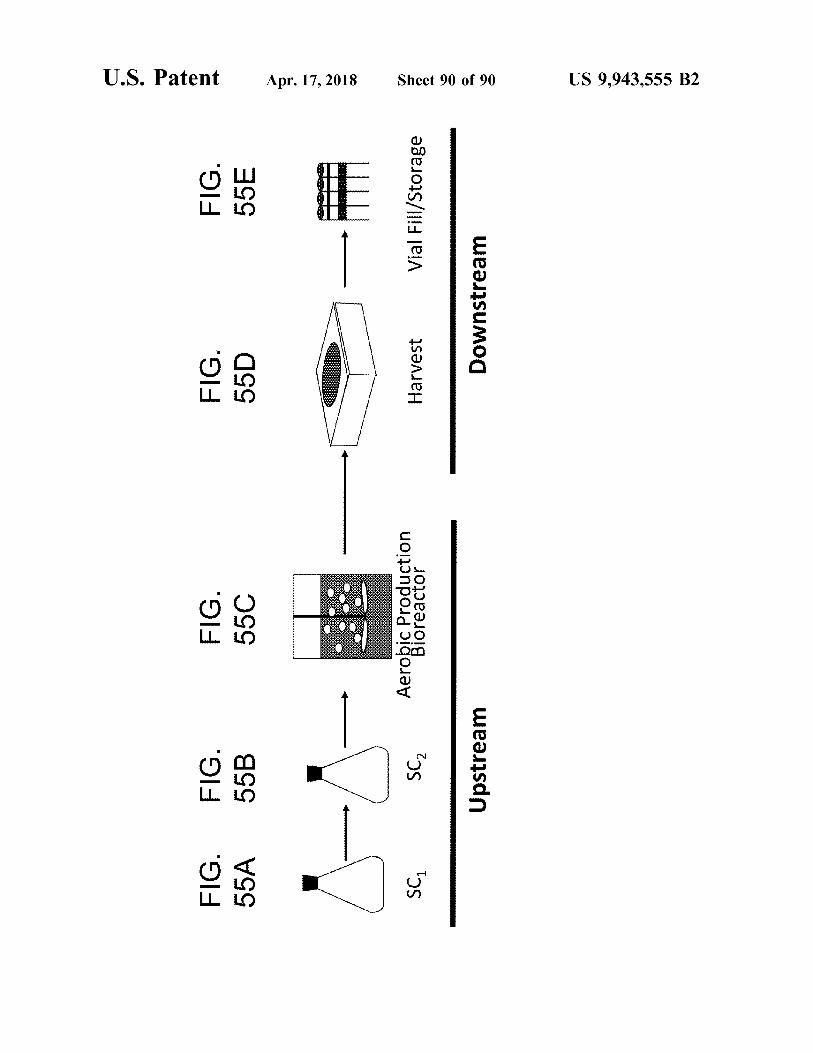
U . S . Patent
FIG . 55A
FIG . 55B
FIG . 55C
FIG . 55D
FIG . 55E
1 - 1 -
-
- F
1
Apr . 17 , 2018
SC ,
SC ,
Aerobic Peraduction
Harvest
Vial Fill / Storage
Sheet 90 of 90
Upstream
Downstream
US 9 , 943 , 555 B2

US 9 , 943 , 555 B2 BACTERIA ENGINEERED TO REDUCE gastritis and severe allergic reactions ( e . g . , wheezing , light
HYPERPHENYLALANINEMIA headedness , nausea , flushing of the skin ) . The enzyme phenylalanine ammonia lyase ( PAL ) is
The present application is a continuation of U . S . appli capable of metabolizing phenylalanine to non - toxic levels of cation Ser . No . 15 / 154 , 934 , filed May 13 , 2016 , which 5 ammonia and transcinnamic acid . Unlike PAH , PAL does claims the benefit of priority to U . S . Provisional Patent not require THB cofactor activity in order to metabolize Application No . 62 / 161 , 137 , filed May 13 , 2015 , and U . S . phenylalanine . Studies of oral enzyme therapy using PAL Provisional Patent Application No . 62 / 256 , 052 , filed Nov . have been conducted , but “ human and even the animal 16 , 2015 , the contents of which are hereby incorporated by studies were not continued because PAL was not available in reference herein in their entirety . 10 sufficient amounts at reasonable cost ” ( Sarkissian et al . ,
This disclosure relates to compositions and therapeutic 1999 ) . A pegylated form of recombinant PAL ( PEG - PAL ) is methods for reducing hyperphenylalaninemia . In certain also in development as an injectable form of treatment . aspects , the disclosure relates to genetically engineered However , most subjects dosed with PEG - PAL have suffered bacteria that are capable of reducing hyperphenylalaninemia from injection site reactions and / or developed antibodies to in a mammal . In certain aspects , the compositions and 15 this therapeutic enzyme ( Longo et al . , 2014 ) . Thus , there is methods disclosed herein may be used for treating diseases significant unmet need for effective , reliable , and / or long associated with hyperphenylalaninemia , e . g . , phenylketonu term treatment for diseases associated with hyperphenylala ria . ninemia , including PKU .
Phenylalanine is an essential amino acid primarily found L - amino acid deaminase ( LAAD ) catalyzes oxidative in dietary protein . Typically , a small amount is utilized for 20 deamination of phenylalanine to generate phenylpyruvate , protein synthesis , and the remainder is hydroxylated to and trace amounts of ammonia and hydrogen peroxide . tyrosine in an enzymatic pathway that requires phenylala - Phenylpyruvic acid ( PPA ) is widely used in the pharmaceu nine hydroxylase ( PAH ) and the cofactor tetrahydrobiop tical , food , and chemical industries , and PPA is the starting terin . Hyperphenylalaninemia is a group of diseases associ - material for the synthesis of D - phenylalanine , a raw inter ated with excess levels of phenylalanine , which can be toxic 25 mediate in the production of many chiral drugs and food and cause brain damage . Primary hyperphenylalaninemia is additives . LAAD has therefore been studied in the context of caused by deficiencies in PAH activity that result from industrial PPA production ( Hou et al . 2015 , Appl Microbiol mutations in the PAH gene and / or a block in cofactor Biotechnol . 2015 October ; 99 ( 20 ) : 8391 - 402 ; “ Production of metabolism . phenylpyruvic acid from L - phenylalanine using an L - amino
Phenylketonuria ( PKU ) is a severe form of hyperphenyl - 30 acid deaminase from Proteus mirabilis : comparison of enzy alaninemia caused by mutations in the PAH gene . PKU is an matic and whole - cell biotransformation approaches ” ) . Phe autosomal recessive genetic disease that ranks as the most nylpyruvate is unable to cross the blood brain barrier ( Steele , common inborn error of metabolism worldwide ( 1 in 3 , 000 Fed Proc . 1986 June ; 45 ( 7 ) : 2060 - 4 ; “ Blood - brain barrier births ) , and affects approximately 13 , 000 patients in the transport of the alpha - keto acid analogs of amino acids , ” United States . More than 400 different PAH gene mutations 35 indicating that this conversion is useful in controlling the have been identified ( Hoeks et al . , 2009 ) . Current PKU neurological phenotypes of PKU . therapies require substantially modified diets consisting of In some embodiments , the disclosure provides genetically protein restriction . Treatment from birth generally reduces engineered bacteria that encode and express a phenylalanine brain damage and mental retardation ( Hoeks et al . , 2009 ; metabolizing enzyme ( PME ) . In some embodiments , the Sarkissian et al . , 1999 ) . However , the protein - restricted diet 40 disclosure provides genetically engineered bacteria that must be carefully monitored , and essential amino acids as encode and express phenylalanine ammonia lyase and / or well as vitamins must be supplemented in the diet . Further phenylalanine hydroxylase and / or L - aminoacid deaminase more , access to low protein foods is a challenge as they are and are capable of reducing hyperphenylalaninemia . more costly than their higher protein , nonmodified counter - In certain embodiments , the genetically engineered bac parts ( Vockley et al . , 2014 ) . 45 teria are non - pathogenic and may be introduced into the gut
In children with PKU , growth retardation is common on in order to reduce toxic levels of phenylalanine . In certain a low - phenylalanine diet ( Dobbelaere et al . , 2003 ) . In adult - embodiments , the phenylalanine ammonia lyase and / or phe hood , new problems such as osteoporosis , maternal PKU , nylalanine hydroxylase and / or L - aminoacid deaminase is and vitamin deficiencies may occur ( Hoeks et al . , 2009 ) . stably produced by the genetically engineered bacteria , Excess levels of phenylalanine in the blood , which can 50 and / or the genetically engineered bacteria are stably main freely penetrate the blood - brain barrier , can also lead to tained in vivo and / or in vitro . In certain embodiments , the neurological impairment , behavioral problems ( e . g . , irrita genetically engineered bacteria further comprise a phenyl bility , fatigue ) , and / or physical symptoms ( e . g . , convulsions , alanine transporter gene to increase their uptake of phenyl skin rashes , musty body odor ) . International guidelines alanine . The invention also provides pharmaceutical com recommend lifelong dietary phenylalanine restriction , which 55 positions comprising the genetically engineered bacteria , is widely regarded as difficult and unrealistic ( Sarkissian et and methods of modulating and treating disorders associated al . , 1999 ) , and “ continued efforts are needed to overcome the with hyperphenylalaninemia . biggest challenge to living with PKU — lifelong adherence to the low - phe diet ” ( Macleod et al . , 2010 ) . BRIEF DESCRIPTION OF THE FIGURES
In a subset of patients with residual PAH activity , oral 60 administration of the cofactor tetrahydrobiopterin ( also FIG . 1 depicts a synthetic biotic for treating phenylke referred to as THB , BH4 , Kuvan , or sapropterin ) may be tonuria ( PKU ) and disorders characterized by hyperphenyl used together with dietary restriction to lower blood phe alaninemia . nylalanine levels . However , cofactor therapy is costly and FIG . 2 A depicts a schematic of phenylalanine hydroxy only suitable for mild forms of phenylketonuria . The annual 65 lase action in phenylketonuria ( PKU ) . FIG . 2B depicts a cost of Kuvan , for example , may be as much as $ 57 , 000 per schematic of phenylalanine hydroxylase ( PAH ) action . FIG . patient . Additionally , the side effects of Kuvan can include 2C depicts a schematic of phenylalanine ammonia lyase

US 9 , 943 , 555 B2
( PAL ) action . FIG . 2D depicts a schematic of L - amino acid the strain is SYN - PKU511 . FIG . 13C depicts a schematic deaminase ( LAAD ; e . g . , from Proteus mirabilis ) action . diagram of one preferred embodiment of the disclosure ,
FIG . 3 depicts a synthetic biotic for treating phenylke - wherein the E . coli Nissle chromosome is engineered to tonuria ( PKU ) and disorders characterized by hyperphenyl contain five copies of PAL under the control of an oxygen alaninemia . 5 level - dependent promoter ( e . g . , PfnrS - PAL3 ) inserted at
FIG . 4 depicts a synthetic biotic for treating phenylke different integration sites on the chromosome ( malE / K , tonuria ( PKU ) and disorders characterized by hyperphenyl - vicS / nepl . malP / T . agal / rsml . and cea ) , and one copy of a alaninemia . phenylalanine transporter gene under the control of an FIG . 5 depicts a synthetic biotic for treating phenylke oxygen level - dependent promoter ( e . g . , PfnrS - pheP ) tonuria ( PKU ) and disorders characterized by hyperphenyl - 10 inserted at a different integration site on the chromosome alaninemia . ( lacZ ) . The genome is further engineered to include a thyA FIG . 6 depicts the gene organization of an exemplary auxotrophy , in which the thyA gene is deleted and / or construct comprising a gene encoding PAL3 and a Tet promoter sequence on a high - copy plasmid e . g . , as com replaced with an unrelated gene , as well as a kanamycin prised in SYN - PKU202 , SYN - PKU303 . resistance gene .
FIG . 7 depicts the gene organization of an exemplary FIG . 14 depicts the gene organization of a non - limiting construct comprising a gene encoding PAL3 and an FNR exemplary construct comprising a gene encoding araC and promoter sequence on a low - copy plasmid , e . g . , as com a gene encoding LAAD from Proteus mirabilis and an prised in SYN - PKU304 , SYN - PKU307 , SYN - PKU305 , arabinose inducible promoter ( ParaBAD ) sequence for chro SYN - PKU306 . 20 mosomal insertion into the endogenous arabinose operon for
FIG . 8 depicts the gene organization of an exemplary chromosomal integration , e . g . , as comprised in SYN construct comprising a gene encoding PAL3 and a Tet PKU705 . promoter sequence on a low - copy plasmid , e . g . , SYN . FIG . 15A depicts phenylalanine concentrations in samples PKU302 , SYN - PKU201 . comprising bacteria expressing PAL1 or on low - copy ( LC ;
FIG . 9 depicts the gene organization of an exemplary 25 SYN - PKU101 ) or high - copy ( HC ; SYN - PKU102 ) plasmids construct , e . g . , comprised in SYN - PKU401 , comprising a or PAL3 on low - copy ( LC ; SYN - PKU201 ) or high - copy cloned LAAD gene under the control of a Tet promoter ( HC ; SYN - PKU202 ) plasmids , induced with anhydrous sequence and a Tet repressor gene . tetracycline ( ATC ) , and then grown in culture medium
FIG . 10 depicts a schematic representation of the con - supplemented with 4 mM ( 660 , 000 ng / mL ) of phenylala struction of a phep knock - in strain , wherein recombineering 30 nine . Samples were removed at 0 hrs , 4 hrs , and 23 hrs . is used to insert a second copy of phep into the Nissle lacZ Phenylalanine concentrations were determined by mass gene . spectrometry . FIG . 15B depicts cinnamate levels in samples
FIG . 11 depicts the gene organization of an exemplary at 4 hrs and 23 hrs post - induction . In PAL3 - expressing construct comprising a gene encoding PheP , a gene encoding strains , the PAL3 gene is derived from Photorhabdus lumi TetR , and a tet promoter sequence for chromosomal inser - 35 nescens , an enterobacterium in the same taxonomic subdi tion e . g . , as for example comprised in SYN - PKU203 , SYN - vision as Escherichia coli . PKU401 , SYN - PKU402 , SYN - PKU302 , and SYN . FIG . 16A depicts phenylalanine concentrations in samples PKU303 . comprising bacteria expressing PAL1 or PAL3 on low - copy
FIG . 12A depicts the gene organization of an exemplary ( LC ) or high - copy ( HC ) plasmids , or further comprising a construct , comprising a cloned PAL3 gene under the control 40 copy of phep driven by the Tet promoter integrated into the of an FNR promoter sequence , on a low - copy , kanamycin - chromosome . Bacteria were induced with ATC , and then resistant plasmid ( PSC101 origin of replication . Under grown in culture medium supplemented with 4 mM ( 660 , anaerobic conditions , PAL3 degrades phenylalanine to non 000 ng / mL ) of phenylalanine to an OD600 of 2 . 0 . Samples toxic trans - cinnamate . FIG . 12B depicts an additional copy were removed at 0 hrs , 2 hrs , and 4 hrs post - induction and of the endogenous E . coli high affinity phenylalanine trans - 45 phenylalanine concentrations were determined by mass porter , phep , driven by the PfnrS promoter and inserted into spectrometry . Notably , the additional copy of pheP permitted the lacZ locus on the Nissle chromosome . the degradation of phenylalanine ( 4 mM ) in 4 hrs . FIG . 16B FIGS . 13A , 13B , and 13C depict schematic diagrams of depicts cinnamate levels in samples at 2 hrs and 4 hrs
non - limiting embodiments of the disclosure . FIG . 13A post - induction . In some embodiments , cinnamate may be depicts phenylalanine degradation components integrated 50 used as an alternative biomarker for strain activity . PheP into the E . coli Nissle chromosome . In some embodiments , overexpression improves phenylalanine metabolism in engi engineered plasmid - free bacterial strains are used to prevent neered bacteria . Strains analyzed in this data set are SYN plasmid conjugation in vivo . In some embodiments , mul - PKU101 , SYN - PKU102 , SYN - PKU202 , SYN - PKU201 , tiple insertions of the PAL gene result in increased copy SYN - PKU401 , SYN - PKU402 , SYN - PKU203 , SYN number and / or increased phenylalanine degradation activity . 55 PKU302 , SYN - PKU303 . In some embodiments , a copy of the endogenous E . coli high FIGS . 17A and 17B depict the state of one non - limiting affinity phenylalanine transporter , phep , is driven by the embodiment of the PAL construct under non - inducing ( FIG . PfnrS promoter and is inserted into the lacZ locus . FIG . 13B 17A ) and inducing ( FIG . 17B ) conditions . FIG . 17A depicts depicts a schematic diagram of one non - limiting embodi - relatively low PAL and PheP production under aerobic ment of the disclosure , wherein the E . coli Nissle chromo - 60 conditions due to oxygen ( 0 ) preventing FNR from some is engineered to contain four copies of PfnrS - PAL dimerizing and activating PAL and / or phep gene expression . inserted at four different insertion sites across the genome FIG . 17B depicts up - regulated PAL and PheP production ( malE / K , yicS / nepl , agal / rsml , and cea ) , and one copy of a under anaerobic conditions due to FNR dimerizing and phenylalanine transporter gene inserted at a different inser - inducing FNR promoter - mediated expression of PAL and tion site ( lacZ ) . In this embodiment , the PAL gene is PAL3 65 pheP ( squiggle above “ PAL ” and “ pheP ” ) . Arrows adjacent derived from P . luminescens , and the phenylalanine trans - to a single rectangle , or a cluster of rectangles , depict the porter gene is phep derived from E . coli . In one embodiment , promoter responsible for driving transcription ( in the direc

US 9 , 943 , 555 B2
tion of the arrow ) of such gene ( s ) . Arrows above each phenylalanine was quantified by mass spectrometry . SYN rectangle depict the expression product of each gene . PKU511 comprises 5 integrated copies of an anaerobically
FIG . 18 depicts B - galactosidase levels in samples com ( FNR ) controlled gene encoding phenylalanine ammonia prising bacteria harboring a low - copy plasmid expressing lyase ( PAL ) at 5 chromosomal locations and an anaerobi lacZ from an FNR - responsive promoter selected from the 5 cally controlled gene encoding a high affinity Phe trans exemplary FNR promoters shown Table 3 ( Pfnr1 - 5 ) . Dif - porter ( pheP ) integrated in the lacZ locus . ferent FNR - responsive promoters were used to create a FIGS . 25A and 25B depict phenylalanine concentrations library of anaerobic - inducible reporters with a variety of in cultures of a synthetic probiotic strain , SYN - PKU401 , in cultures o expression levels and dynamic ranges . These promoters which comprises a high copy pUC57 - plasmid with LAAD included strong ribosome binding sites . Bacterial cultures 10 driven by a Tet inducible promoter , cells were grown in were grown in either aerobic ( + 0 , ) or anaerobic conditions flasks shaking at 37 C , and induced with TCA at early log ( 02 ) . Samples were removed at 4 hrs and the promoter phase for a duration of 2 hours . Cells were spun down and activity based on B - galactosidase levels was analyzed by performing standard B - galactosidase colorimetric assays . re - suspended in assay buffer containing phenylalanine . Cells
FIG . 19 A depicts a schematic representation of the lacZ 15 were measured at various cell concentrations and at varying gene under the control of an exemplary FNR promoter oxygen levels . Cells were either incubated aerobically ( 1 ml ) ( P ) . LacZ encodes the B - galactosidase enzyme and is a in a 14 ml culture tube , shaking at 250 rpm . For microaero common reporter gene in bacteria . FIG . 19B depicts FNR bic conditions , cells ( 1 ml ) were incubated in a 1 . 7 ml promoter activity as a function of ß - galactosidase activity in conical tube without shaking . Cells were incubated anaero SYN - PKU904 . SYN - PKU904 , an engineered bacterial 20 bically in a Coy anaerobic chamber supplying 90 % N2 , 5 % strain harboring a low - copy fnrS - lacZ fusion gene , was CO2 , and 5 % H2 . Aliquots were removed from cell assays grown in the presence or absence of oxygen . Values for every 30 min for 2 hrs for phenylalanine quantification by standard B - galactosidase colorimetric assays are expressed mass spectrometry . FIG . 25A depicts phenylalanine concen in Miller units ( Miller , 1972 ) . These data suggest that the trations under aerobic conditions using two cell densities . A fnrS promoter begins to drive high - level gene expression 25 and B are duplicates under the same experimental condi within 1 hr . under anaerobic conditions . FIG . 19C depicts tions . The activity in aerobic conditions is ~ 50 umol / hr . / 1e9 the growth of bacterial cell cultures expressing lacZ over cells . FIG . 25B depicts phenylalanine concentrations of time , both in the presence and absence of oxygen . aerobically , microaerobically , or anaerobically grown cells .
FIGS . 20A and 20B depict phenylalanine levels produced FIG . 26A shows phenylalanine concentrations before and under aerobic ( FIG . 20A ) or anaerobic conditions ( FIG . 30 after feeding in an in vivo mouse model of PKU . At the 20B ) in samples of wild - type Nissle , samples of bacteria beginning of the study , homozygous BTBR - Pahenu2 mice comprising a low - copy plasmid expressing PAL3 from the were given water supplemented with 100 micrograms / mL Tet promoter or exemplary FNR promoters , or further com - ATC and 5 % sucrose . Mice were fasted by removing chow prising a copy of phep driven by the Tet promoter and overnight ( 10 hrs ) , and blood samples were collected by integrated into the chromosome . Samples were incubated in 35 mandibular bleeding the next morning in order to determine culture medium supplemented with ATC and 4 mM ( 660 , baseline phenylalanine levels . Mice were given chow again , 000 ng / mL ) of phenylalanine . Samples were removed at 0 gavaged with 100 microliters ( 5x10° CFU ) of bacteria hrs , 2 hrs , 4 hrs , and 24 hrs . Phenylalanine concentration was ( SYN - PKU302 or control Nissle ) after 1 hr . , and allowed to determined by mass spectrometry . These data suggest that feed for another 2 hrs . Serum phenylalanine concentrations the FNR - responsive fnrS promoter is as effective at activat - 40 were determined 2 hrs post - gavage . FIG . 26B shows the ing PAL3 expression as a tetracycline - inducible promoter percent ( % ) change in blood phenylalanine concentrations under anaerobic conditions . before and after feeding as a male or female group average
FIG . 21 depicts phenylalanine concentrations in cultures ( p < 0 . 01 ) . of synthetic probiotic strains , with and without an additional FIGS . 27A and 27B depict blood phenylalanine concen copy of pheP inserted on the chromosome . After 1 . 5 hrs of 45 trations relative to baseline following subcutaneous phenyl growth , cultures were placed in Coy anaerobic chamber alanine challenge in an in vivo mouse model of PKU . Mice supplying 90 % N , 5 % CO . , and 5 % H , . After 4 hrs of were orally gavaged with 200 uL of H , O ( n = 30 ) , SYN induction , bacteria were resuspended in assay buffer con - PKU901 ( n = 33 ) , or SYN - PKU303 ( n = 34 ) at 30 and 90 taining 4 mM phenylalanine . Aliquots were removed from minutes post - phenylalanine injection ( 0 . 1 mg / gram of aver cell assays every 30 min for 3 hrs for phenylalanine quan - 50 age group body weight ) . FIGS . 27A and 27B show blood tification by mass spectrometry . Phenylalanine degradation phenylalanine concentrations at 2 hrs and 4 hrs post - phe rates in strains comprising an additional copy of phep nylalanine injection , respectively . These data indicate that ( SYN - PKU304 and SYN - PKU305 ; left ) were higher than oral administration of the engineered probiotic strain SYN strains lacking an additional copy of pheP ( SYN - PKU308 PKU303 significantly reduces blood phenylalanine levels in and SYN - PKU307 ; right ) . 55 mice , compared to mice administered mock treatment ( H2O )
FIG . 22 depicts trans - cinnamate concentrations ( PAL or the parental strain ( SYN - PKU901 ) ( * , p < 0 . 05 ; * * * , activity ) for strains comprising single PAL3 insertions at p < 0 . 001 ; * * * * , p < 0 . 00001 ) . SYN - PKU303 is capable of various locations on the chromosome . intercepting enterorecirculating phenylalanine .
FIG . 23 depicts trans - cinnamate concentrations ( PAL FIG . 28 depicts blood phenylalanine concentrations rela activity ) for strains comprising multiple PAL3 insertions at 60 tive to baseline following subcutaneous phenylalanine chal various locations on the chromosome . lenge in an in vivo mouse model of PKU . Mice were orally
FIG . 24 depicts phenylalanine concentrations in cultures gavaged with 200 uL of H2O ( n = 30 ) , SYN - PKU901 ( n = 33 ) , of synthetic probiotic strain SYN - PKU511 over time . After SYN - PKU303 ( n = 34 ) , or SYN - PKU304 ( n = 34 ) at 30 and 90 2 . 5 hrs of growth , cultures were placed in Coy anaerobic minutes post - phenylalanine injection ( 0 . 1 mg / gram of aver chamber supplying 90 % N2 , 5 % CO2 , and 5 % H2 . After 3 . 5 65 age group body weight ) . Blood phenylalanine concentra hrs of induction in phenylalanine containing medium , whole tions post phenylalanine injection indicate that SYN cell extracts were prepared every 30 min for 3 hrs and PKU304 ( low copy plasmid containing forS - PAL ) is at least

US 9 , 943 , 555 B2
as effective as SYN - PKU303 ( high copy plasmid containing centrations of phenylalanine ( FIG . 33A ) , and absolute val Tet - PAL ) in reducing circulating Phe levels in the entero ues of phenylalanine and PKU specific and PAL specific recirculation model . metabolites ( FIGS . 33B , 33C , 33D , 33E , and 33F ) following
FIGS . 29A and 29B depict blood phenylalanine concen subcutaneous phenylalanine challenge in an in vivo mouse trations relative to baseline following subcutaneous phenyl - 5 model of PKU . Mice were orally gavaged with a total of alanine challenge in an in vivo mouse model of PKU . Mice 8004 of H2O ( n = 9 ] ) , SYN - PKU801 ( n = 12 ) , or 800 uL of were orally gavaged with H2O , SYN - PKU901 , SYN SYN - PKU517 ( n = 12 ) ( 3 . 6e10 cfu / mouse ) at 30 and 90 PKU303 , or SYN - PKU304 at 30 and 90 minutes post minutes post - phenylalanine injection . FIG . 33A depicts phenylalanine injection ( 0 . 1 mg / gram of average group body blood phenylalanine concentrations relative to baseline ; weight ) . FIGS . 29A and 29B show blood phenylalanine 10 total metabolic activity for SYN - PKU517 was calculated as concentrations at 2 hrs and 4 hrs post - phenylalanine injec 39 . 6 umol / hr . and the total reduction in Aphe was 17 % tion , respectively . These data indicate that oral administra tion of engineered probiotic strains SYN - PKU303 and SYN relative to SYN - PKU801 ( P < 0 . 05 ) . FIG . 33B depicts the PKU304 significantly reduces blood phenylalanine levels in blood phenylalanine concentration at 0 and 4 hours post mice compared to mice administered mock treatment ( H . O ) 15 phenylalanine injection . FIG . 33C depicts the blood phe or the parental strain ( SYN - PKU901 ) ( * p < 0 . 05 ; * * nylpyruvate concentration at 0 and 4 hours post phenylala p < 0 . 01 ; * * * , p < 0 . 001 ; * * * * , p < 0 . 0001 ) . FIGS . 29C and 29D nine injection . FIG . 33D depicts the blood phenyllactate depict scatter plots of the data shown in FIGS . 29A and 29B . concentration at 0 and 4 hours post phenylalanine injection .
FIGS . 30A and 30B depict blood phenylalanine concen FIG . 33E depicts the blood t - cinnamic acid concentration at trations relative to baseline following subcutaneous phenyl - 20 0 and 4 hours post phenylalanine injection . FIG . 33F depicts alanine challenge in an in vivo mouse model of PKU . Mice the blood hippuric acid concentration at 0 and 4 hours post were orally gavaged with 200 uL of H2O ( n = 12 ) , 200 uL of phenylalanine injection . SYN - PKU901 ( n = 12 ) , or 100 , 200 , or 400 uL of SYN - FIGS . 34A , 34B , 34C , 34D , 34E , and 34F depict blood PKU304 ( n = 12 in each dose group ) at 30 and 90 minutes phenylalanine concentrations relative to baseline and con post - phenylalanine injection ( 0 . 1 mg / gram of average group 25 centrations of phenylalanine ( FIG . 34A ) , and absolute val body weight ) . FIGS . 30A and 30B show a dose - dependent ues of phenylalanine and PKU specific and PAL specific decrease in blood phenylalanine levels in SYN - PKU304 - metabolites ( FIGS . 34B , 340 , 34D , 34E , and 34F ) following treated mice compared to mice administered mock treatment subcutaneous phenylalanine challenge in an in vivo mouse ( HO ) or the parental strain ( SYN - PKU901 ) ( * 30 % model of PKU . Mice were orally gavaged with a total of 800 decrease ; p < 0 . 05 ) . This experiment represents one of eight 30 uL of H2O ( n = 12 ) , SYN - PKU901 ( n = 12 ) , or 800 uL of studies of this same design , and each one shows that SYN - PKU705 ( n = 12 ) ( 3 . 6e10 cfu / mouse ) at 30 and 90 SYN - PKU304 is capable of intercepting enterorecirculating minutes post - phenylalanine injection . FIG . 34A depicts phenylalanine . blood phenylalanine concentrations relative to baseline ;
FIGS . 31A and 31B depicts a schematic of PKU specific total metabolic activity for SYN - PKU705 was calculated as and PAL specific phenylalanine metabolites . FIG . 31A 35 133 . 2 umol / hr . and the total reduction in Aphe was 30 % depicts a schematic of the conversion of phenylalanine to relative to SYN - PKU901 ( P < 0 . 05 ) . FIG . 34B depicts the phenylpyruvic acid and phenyllactic acid in the absence of blood phenylalanine concentration at 0 and 4 hours post functional PAH . FIG . 31B depicts a schematic of the con - phenylalanine injection . FIG . 34C depicts the blood phe version of phenylalanine to trans - cinnamic acid by PAL3 , nylpyruvate concentration at 0 and 4 hours post phenylala which is further metabolized to hippuric acid by liver 40 nine injection . FIG . 34D depicts the blood phenyllactate enzymes . These metabolites can be detected by mass spec - concentration at 0 and 4 hours post phenylalanine injection . trometry as described in Examples 24 - 26 or by other means . FIG . 34E depicts the blood t - cinnamic acid concentration at
FIGS . 32A , 32B , 32C , 32D , 32E , and 32F depict blood and 4 hours post phenylalanine injection . FIG . 34F depicts phenylalanine concentrations relative to baseline and con - the blood hippuric acid concentration at 0 and 4 hours post centrations of phenylalanine ( FIG . 32A ) , and absolute val - 45 phenylalanine injection . ues of phenylalanine and PKU specific and PAL specific FIG . 35 depicts phenylalanine and 2 toxic analogs , metabolites ( FIGS . 32B , 32C , 32D , 32E , and 32F ) following p - fluoro - DL - phenylalanine , and o - fluoro - DL - phenylalanine , subcutaneous phenylalanine challenge in an in vivo mouse which are useful for an untargeted approach to select PAL model of PKU . Mice were orally gavaged with a total of enzymes with increased activity . P - fluoro - DL - phenylala 8004 of H2O ( n = 12 ) , SYN - PKU901 ( n = 12 ) , or 8004 of 50 nine , and o - fluoro - DL - phenylalanine are incorporated into SYN - PKU304 ( n = 12 ) ( 2 . 9e10 cfu / mouse ) at 30 and 90 cellular protein in the place of phenylalanine , resulting in minutes post - phenylalanine injection . FIG . 32A depicts cell death . Since these compounds are readily taken up by blood phenylalanine concentrations relative to baseline ; Phep , and can act as a substrate for PAL as shown below , total metabolic activity for SYN - PKU304 was calculated as they can be employed in genetic selection and screening for 81 . 2 umol / hr . and the total reduction in Aphe was 45 % 55 the identification of strains with improved Phe consumption relative to SYN - PKU901 ( P < 0 . 05 ) . FIG . 32B depicts the activity . Mutations allowing more efficient PAL metabolism blood phenylalanine concentration at 0 and 4 hours post may prevent the incorporation of the phenylalanine analog phenylalanine injection . FIG . 32C depicts the blood phe into cellular protein , therefore allowing growth under higher nylpyruvate concentration at 0 and 4 hours post phenylala concentrations of the analog . nine injection . FIG . 32D depicts the blood phenyllactate 60 FIG . 36 depicts a map of exemplary integration sites concentration at 0 and 4 hours post phenylalanine injection within the E . coli 1917 Nissle chromosome . These sites FIG . 32E depicts the blood t - cinnamic acid concentration at indicate regions where circuit components may be inserted O and 4 hours post phenylalanine injection . FIG . 32F depicts into the chromosome without interfering with essential gene the blood hippuric acid concentration at 0 and 4 hours post expression . Backslashes ( l ) are used to show that the inser phenylalanine injection . 65 tion will occur between divergently or convergently
FIGS . 33A , 33B , 33C , 33D , 33E , and 33F depict blood expressed genes . Insertions within biosynthetic genes , such phenylalanine concentrations relative to baseline and con - as thyA , can be useful for creating nutrient auxotrophies . In

US 9 , 943 , 555 B2 10
some embodiments , an individual circuit component is FIG . 45A depicts a schematic of a recombinase - based inserted into more than one of the indicated sites . switch to activate PAL3 expression using different inducible
FIG . 37 depicts three bacterial strains which constitu - promoters and ribosome binding sites . Recombinase expres tively express red fluorescent protein ( RFP ) . In strains 1 - 3 , sion causes recombinatorial flipping of the PAL3 gene to the the rfp gene has been inserted into different sites within the 5 ON orientation , leading to the production of PAL3 and to the bacterial chromosome , and results in varying degrees of degradation of phenylalanine . In some embodiments , brightness under fluorescent light . Unmodified E . coli Nissle recombinase - based switches are tuned to respond to specific ( strain 4 ) is non - fluorescent . levels of an inducer . FIG . 45B depicts the relationship
FIG . 38 depicts a graph of Nissle residence in vivo . between the concentration of an inducer and the percentage Streptomycin - resistant Nissle was administered to mice via 10 of PAL3 - containing constructs in the ON orientation . The oral gavage without antibiotic pre - treatment . Fecal pellets shaded area shows the predicted efficacy range of the from 6 total mice were monitored post - administration to inducer in vivo . determine the amount of administered Nissle still residing FIG . 46A depicts another non - limiting embodiment of the within the mouse gastrointestinal tract . The bars represent disclosure , wherein the expression of a heterologous gene is the number of bacteria administered to the mice . The line 15 activated by an exogenous environmental signal . In the represents the number of Nissle recovered from the fecal absence of arabinose , the AraC transcription factor adopts a samples each day for 10 consecutive days . conformation that represses transcription . In the presence of
FIGS . 39A and 39B depict phenylalanine concentrations arabinose , the AraC transcription factor undergoes a con in SYN - PKU302 cultures over time . After 1 . 5 hrs of growth , formational change that allows it to bind to and activate the ATC was added to cultures of SYN - PKU302 , and SYN - 20 ParaBAD promoter ( ParaBAD ) , which induces expression of PKU304 cultures were placed in Coy anaerobic chamber the Tet repressor ( TetR ) and an anti - toxin . The anti - toxin supplying 90 % N2 , 5 % CO2 , and 5 % H2 . After 4 hrs of builds up in the recombinant bacterial cell , while TetR induction , bacteria were resuspended in assay buffer con - prevents expression of a toxin ( which is under the control of taining 4 mM phenylalanine and at different pH ( pH range a promoter having a TetR binding site ) . However , when 7 . 25 - 2 . 25 ) . Aliquots were removed from cell assays every 25 arabinose is not present , both the anti - toxin and TetR are not 30 min for 2 hrs for phenylalanine quantification by mass expressed . Since TetR is not present to repress expression of spectrometry . Phenylalanine degradation rates decreased as the toxin , the toxin is expressed and kills the cell . FIG . 46A pH of the assay buffer decreased in both strains , SYN - also depicts another non - limiting embodiment of the disclo PKU302 ( FIG . 39A ) and SYN - PKU304 ( FIG . 39B ) . sure , wherein the expression of an essential gene not found
FIG . 40 depicts an exemplary schematic of the E . coli 30 in the recombinant bacteria is activated by an exogenous 1917 Nissle chromosome comprising multiple mechanisms environmental signal . In the absence of arabinose , the AraC of action ( MoAs ) . transcription factor adopts a conformation that represses
FIG . 41 depicts the gene organization of an exemplary transcription of the essential gene under the control of the construct in which the PAL3 and phep genes are co - tran - araBAD promoter and the bacterial cell cannot survive . In scribed under the control of an exemplary FNR promoter 35 the presence of arabinose , the AraC transcription factor ( Pfurs ) . undergoes a conformational change that allows it to bind to FIGS . 42A and 42B depict the gene organization of an and activate the araBAD promoter , which induces expres
exemplary construct in which the Int5 recombinase gene is s ion of the essential gene and maintains viability of the operably linked to an exemplary FNR promoter ( Ptnrs ) , and bacterial cell . the PAL3 gene is operably linked to a strong constitutive 40 FIG . 46B depicts a non - limiting embodiment of the promoter . FIG . 42A depicts a schematic diagram of the disclosure , where an anti - toxin is expressed from a consti PAL3 gene , flanked by Int5 sites , in the OFF orientation ( 3 ' tutive promoter , and expression of a heterologous gene is to 5 ' ) . When Int5 gene expression is activated under anaero - activated by an exogenous environmental signal . In the bic conditions , recombinatorial flipping of PAL3 to the ON absence of arabinose , the AraC transcription factor adopts a orientation ( 5 ' to 3 ' ; FIG . 42B ) leads to the production of 45 conformation that represses transcription . In the presence of PAL3 and to phenylalanine metabolism . Any strong consti - arabinose , the AraC transcription factor undergoes a con tutive promoter sequence may be used . formational change that allows it to bind to and activate the
FIGS . 43A , 43B , and 43C depict the gene organization of araBAD promoter , which induces expression of TetR , thus an exemplary construct in which the Int5 recombinase gene preventing expression of a toxin . However , when arabinose is operably linked to an FNR promoter ( Plus ) , and the gene 50 is not present , TetR is not expressed , and the toxin is encoding T7 RNA polymerase is flanked by recombinase expressed , eventually overcoming the anti - toxin and killing sites and operably linked to a strong constitutive promoter . the cell . The constitutive promoter regulating expression of FIG . 43A depicts a schematic diagram of the T7 RNA the anti - toxin should be a weaker promoter than the pro polymerase gene , flanked by Int5 sites , in the OFF orienta moter driving expression of the toxin . The araC gene is tion . When Int5 gene expression is activated under anaerobic 55 under the control of a constitutive promoter in this circuit . conditions , the T7 RNA polymerase gene is flipped to the FIG . 46C depicts another non - limiting embodiment of the ON orientation ( FIG . 43B ) . In engineered bacterial strains disclosure , wherein the expression of a heterologous gene is comprising a copy of PAL3 under the control of a 17 - driven activated by an exogenous environmental signal . In the promoter ( P . 77 ; FIG . 43C ) , T7 RNA polymerase expression absence of arabinose , the AraC transcription factor adopts a leads to the production of PAL3 and to phenylalanine 60 conformation that represses transcription . In the presence of metabolism . arabinose , the AraC transcription factor undergoes a con
FIGS . 44A and 44B depict the gene organization of an formational change that allows it to bind to and activate the exemplary construct in which the Int5 recombinase gene is araBAD promoter , which induces expression of the Tet operably linked to an ParaBAD promoter ( P AD ) , and the repressor ( TetR ) and an anti - toxin . The anti - toxin builds up gene encoding T7 RNA polymerase is flanked by recombi - 65 in the recombinant bacterial cell , while TetR prevents nase sites and operably linked to a strong constitutive expression of a toxin ( which is under the control of a promoter . promoter having a TetR binding site ) . However , when

US 9 , 943 , 555 B2 12
arabinose is not present , both the anti - toxin and TetR are not starter culture 1 ( SC1 ) : loop full - glycerol stock , duration expressed . Since TetR is not present to repress expression of overnight , temperature 37° C . , shaking at 250 rpm . FIG . 55B the toxin , the toxin is expressed and kills the cell . The araC depicts the parameters for starter culture 2 ( SC2 ) : 1 / 100 gene is either under the control of a constitutive promoter or dilution from SC1 , duration 1 . 5 hours , temperature 37° C . , an inducible promoter ( e . g . , AraC promoter ) in this circuit . 5 shaking at 250 rpm . FIG . 55C depicts the parameters for the
FIG . 47 depicts the use of GeneGuards as an engineered production bioreactor : inoculum — SC2 , temperature 37° C . , safety component . All engineered DNA is present on a pH set point 7 . 00 , pH dead band 0 . 05 , dissolved oxygen set plasmid which can be conditionally destroyed . See , e . g . , point 50 % , dissolved oxygen cascade agitation / gas FLO , Wright et al . , 2015 . agitation limits 300 - 1200 rpm , gas FLO limits 0 . 5 - 20 stan
FIG . 48A depicts a schematic diagram of a wild - type clbA 10 dard liters per minute , duration 24 hours . FIG . 55D depicts construct . FIG . 48B depicts a schematic diagram of a clbA the parameters for harvest : centrifugation at speed 4000 rpm knockout construct . and duration 30 minutes , wash 1x10 % glycerol / PBS , cen
FIG . 49 depicts exemplary sequences of a wild - type clb trifugation , re - suspension 10 % glycerol / PBS . FIG . 55E construct and a clbA knockout construct . depicts the parameters for vial fill / storage : 1 - 2 mL aliquots ,
FIG . 50 depicts a schematic of a secretion system based 15 - 80° C . on the flagellar type III secretion in which a modified flagellum is used to secrete a therapeutic peptide of interest DESCRIPTION OF EMBODIMENTS by recombinantly fusing the peptide to an N - terminal fla gellar secretion signal of a native flagellar component so that The present disclosure includes genetically engineered the intracellularly expressed chimeric peptide can be mobi - 20 bacteria , pharmaceutical compositions thereof , and methods lized across the inner and outer membranes into the sur - of modulating and treating disorders associated with hyper rounding host environment . phenylalaninemia . In some embodiments , the genetically
FIG . 51 depicts a schematic of a type V secretion system engineered bacteria comprise a gene encoding non - native for the extracellular production of recombinant proteins in phenylalanine ammonia lyase ( PAL ) and are capable of which a therapeutic peptide ( star ) can be fused to an N - ter - 25 processing and reducing phenylalanine in a mammal . Thus , minal secretion signal , a linker and the beta - domain of an the genetically engineered bacteria and pharmaceutical com auto - secreter . In this system , the N - terminal signal sequence positions comprising those bacteria may be used to metabo directs the protein to the SecA - YEG machinery , which lize phenylalanine in the body into non - toxic molecules in moves the protein across the inner membrane into the order to treat and / or prevent conditions associated with periplasm , followed by subsequent cleavage of the signal 30 hyperphenylalaninemia , including PKU . In certain aspects , sequence . The beta - domain is recruited to the Bam complex the compositions comprising the genetically engineered where the beta - domain is folded and inserted into the outer bacteria may be used in the methods of the disclosure to treat membrane as a beta - barrel structure . The therapeutic peptide and / or prevent disorders associated with hyperphenylala is then threaded through the hollow pore of the beta - barrel ninemia . structure ahead of the linker sequence . The therapeutic 35 In order that the disclosure may be more readily under peptide is freed from the linker system by an autocatalytic stood , certain terms are first defined . These definitions cleavage or by targeting of a membrane - associated peptidase should be read in light of the remainder of the disclosure and ( scissors ) to a complementary protease cut site in the linker . as understood by a person of ordinary skill in the art . Unless
FIG . 52 depicts a schematic of a type I secretion system , defined otherwise , all technical and scientific terms used which translocates a passenger peptide directly from the 40 herein have the same meaning as commonly understood by cytoplasm to the extracellular space using HlyB ( an ATP - a person of ordinary skill in the art . Additional definitions binding cassette secreter ; HlyD ( a membrane fusion pro - are set forth throughout the detailed description . tein ) ; and Tolc ( an outer membrane protein ) which form a “ Hyperphenylalaninemia , " “ hyperphenylalaninemic , " channel through both the inner and outer membranes . The and “ excess phenylalanine ” are used interchangeably herein secretion signal - containing C - terminal portion of HlyA is 45 to refer to increased or abnormally high concentrations of fused to the C - terminal portion of a therapeutic peptide ( star ) phenylalanine in the body . In some embodiments , a diag to mediate secretion of this peptide . nostic signal of hyperphenylalaninemia is a blood phenyl
FIG . 53 depicts a schematic of the outer and inner alanine level of at least 2 mg / dL , at least 4 mg / dL , at least membranes of a gram - negative bacterium , and several dele - 6 mg / dL , at least 8 mg / dL , at least 10 mg / dL , at least 12 tion targets for generating a leaky or destabilized outer 50 mg / dL , at least 14 mg / dL , at least 16 mg / dL , at least 18 membrane , thereby facilitating the translocation of a thera - mg / dL , at least 20 mg / dL , or at least 25 mg / dL . As used peutic polypeptides to the extracellular space , e . g . , thera - herein , diseases associated with hyperphenylalaninemia peutic polypeptides of eukaryotic origin containing disul - include , but are not limited to , phenylketonuria , classical or phide bonds . Deactivating mutations of one or more genes typical phenylketonuria , atypical phenylketonuria , perma encoding a protein that tethers the outer membrane to the 55 nent mild hyperphenylalaninemia , nonphenylketonuric peptidoglycan skeleton , e . g . , lpp , ompC , ompA , ompF , tol? , hyperphenylalaninemia , phenylalanine hydroxylase defi tolB , pal , and / or one or more genes encoding a periplasmic ciency , cofactor deficiency , dihydropteridine reductase defi protease , e . g . , degs , degP , nlpl , generates a leaky phenotype . ciency , tetrahydropterin synthase deficiency , and Segawa ' s Combinations of mutations may synergistically enhance the disease . Affected individuals can suffer progressive and leaky phenotype . 60 irreversible neurological deficits , mental retardation ,
FIG . 54 depicts a schematic of non - limiting processes for encephalopathy , epilepsy , eczema , reduced growth , micro designing and producing the genetically engineered bacteria cephaly , tremor , limb spasticity , and / or hypopigmentation of the present disclosure . ( Leonard 2006 ) . Hyperphenylalaninemia can also be sec
FIGS . 55A , B , C , D , and E depict a schematic of non - ondary to other conditions , e . g . , liver diseases . limiting manufacturing processes for upstream and down - 65 “ Phenylalanine ammonia lyase ” and “ PAL ” are used to stream production of the genetically engineered bacteria of refer to a phenylalanine metabolizing enzyme ( PME ) that the present disclosure . FIG . 55A depicts the parameters for converts or processes phenylalanine to trans - cinnamic acid








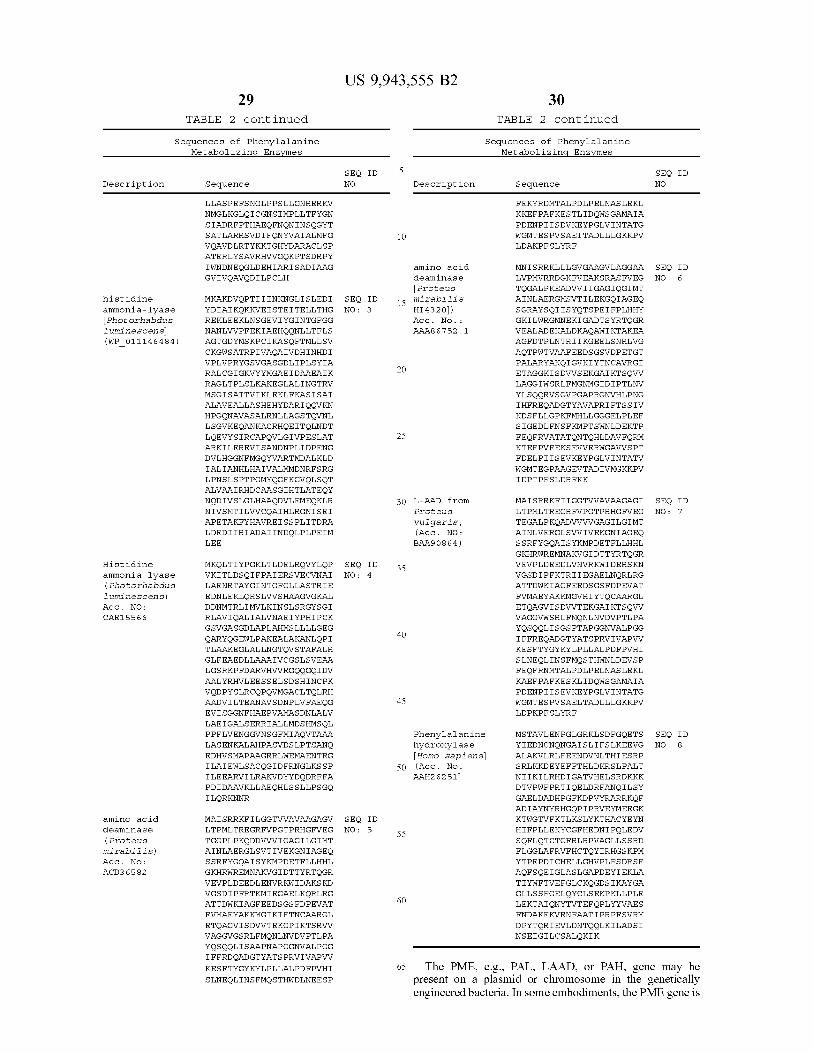


























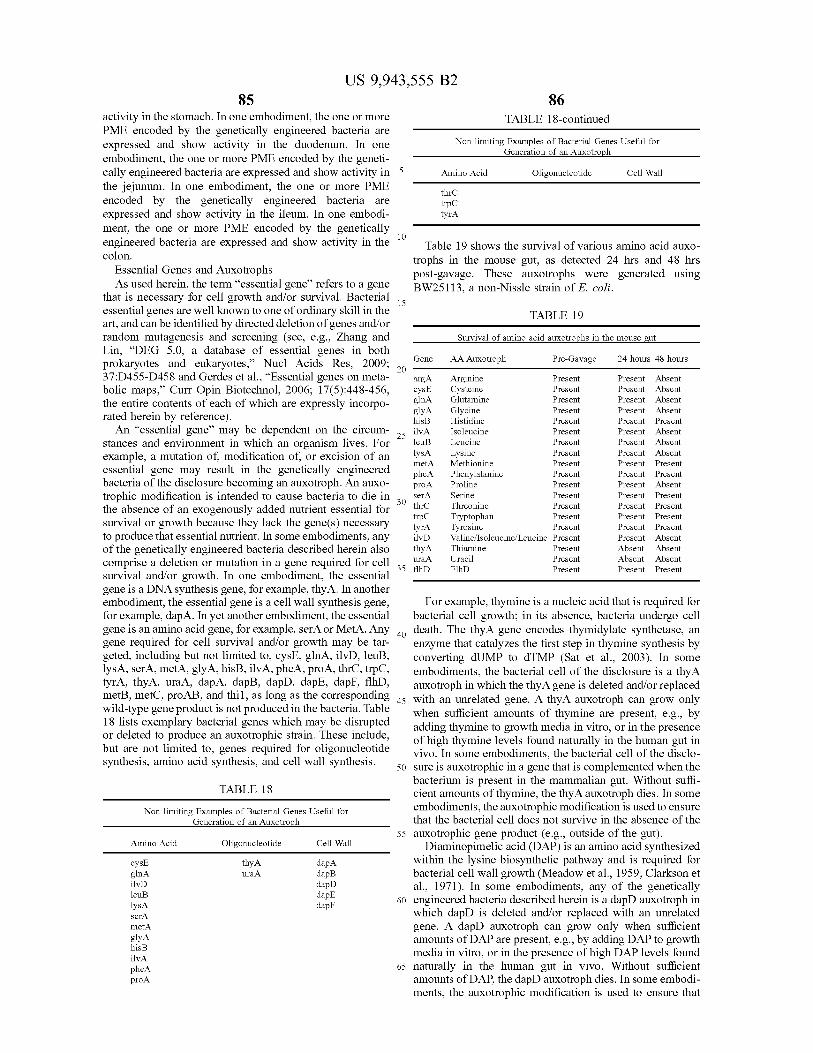









































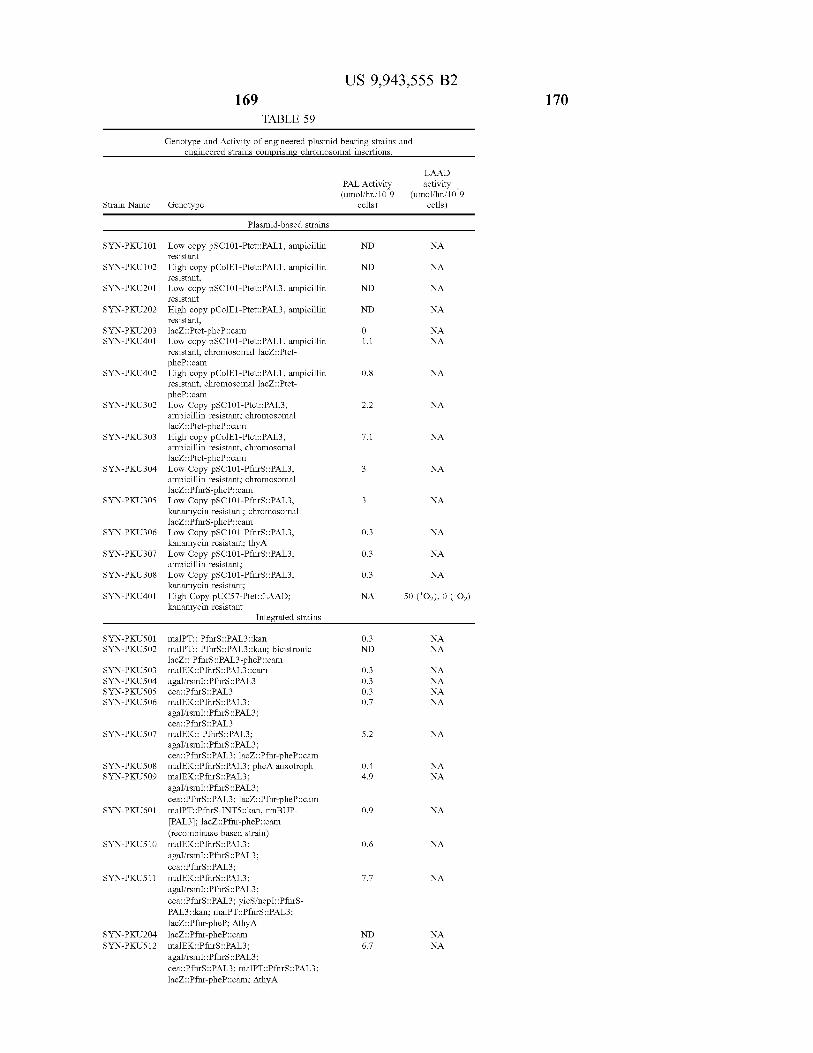
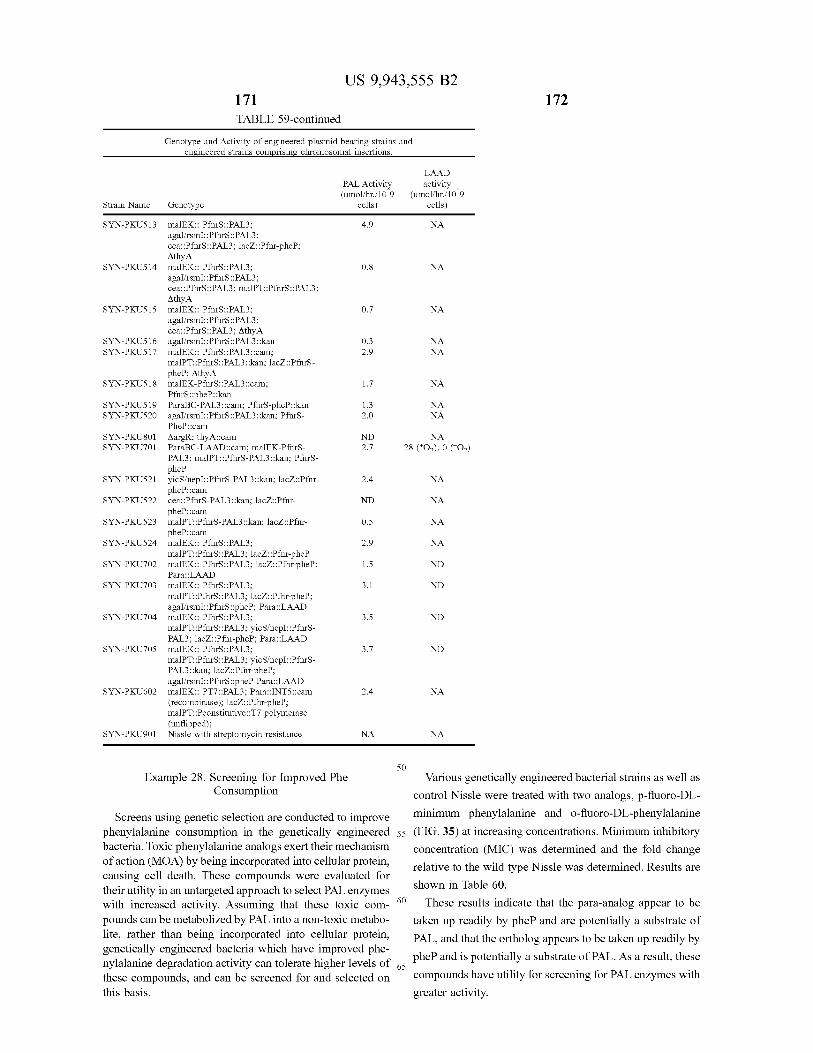




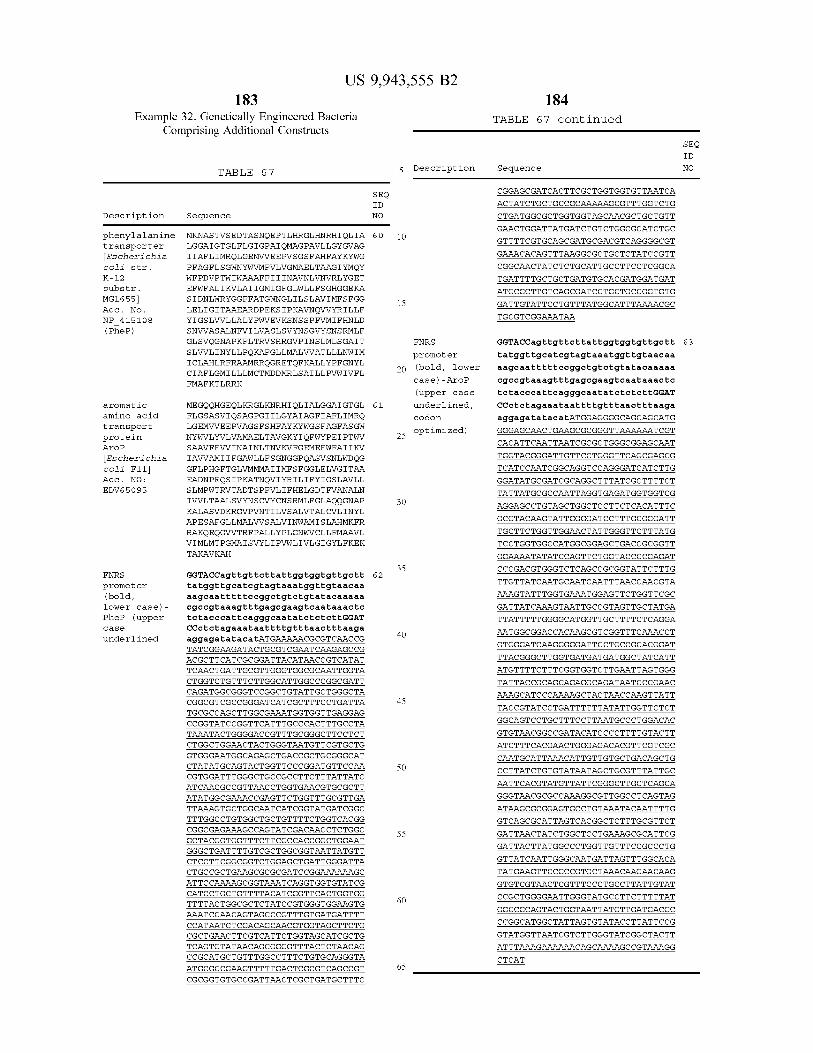








































































Related Documents











