MULTIPLEX REAL-TIME PCR FOR THE DETECTION OF PATHOGENIC INTESTINAL PARASITES AND COMPARISON WITH PARASITOLOGICAL TECHNIQUES MADIHAH BINTI BASUNI UNIVERSITI SAINS MALAYSIA 2012 brought to you by CORE View metadata, citation and similar papers at core.ac.uk provided by Repository@USM

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

MULTIPLEX REAL-TIME PCR FOR THE
DETECTION OF PATHOGENIC INTESTINAL
PARASITES AND COMPARISON WITH
PARASITOLOGICAL TECHNIQUES
MADIHAH BINTI BASUNI
UNIVERSITI SAINS MALAYSIA
2012
brought to you by COREView metadata, citation and similar papers at core.ac.uk
provided by Repository@USM

MULTIPLEX REAL-TIME PCR FOR THE DETECTION OF PATHOGENIC
INTESTINAL PARASITES AND COMPARISON WITH PARASITOLOGICAL
TECHNIQUES
by
MADIHAH BINTI BASUNI
Thesis submitted in fulfillment of the
requirements for the degree of
Master of Science
January 2012

ii
ACKNOWLEDGEMENTS
All praises and gratitude is due to Allah - alhamdulillah, may salawat and salam be granted
to His beloved Prophet Muhammad s.a.w., to his family and companions r.a.
My sincere thanks and appreciation is extended to my supervisor, Professor Rahmah
Noordin who is generous for sharing her wide knowledge, guidance and encouragement
throughout the duration of this work. Her comments, help, advices, suggestions and criticism
have been a great value for me. My warmest thanks also goes to my co-supervisor Dr
Zeehaida Mohamed, who has contributed significantly in my project.
My sincere appreciation is extended to all lecturers, laboratory and administration staffs of
INFORMM who have helped me from time to time during the study. Besides, I would also
like to acknowledge Dr Jamail Muhi, the Deputy Director of the State Health of Sarawak,
and his staffs; En Jamadi, Pn Lila, Pn Norkiah and Pn Rosa from Kuching Health
Department, Hospital Serian and Hospital Lundu. A lot of appreciation also goes to the
contributions from Pn Maimunah Ahmad, En Hafiz Mohamad, Cik Jannah Iman and
laboratory staffs at HUSM. My sincere thanks to Dr Hasnah from IPPT, Dr Lin Naing, Dr
Low and Dr Noorani Ahmad from School of Mathematical Sciences and Dr Sasidharan from
INFORMM for statistical consultation.
Last but not least I would like to extend my warm and sincere thanks to all my friends and
colleagues whose gave me great support, help, advices and companionship; especially Kak
Munah, Kak Anizah, Aishah Azman, Kalpu, Thanes, Izan, Kak Emi, Wani, Shue, Kak
Nurul, Tihah, Kak Syida, Zul, Farhanah, Nik Syaza, Dyana, Kak Linda, Shaza, Chun Wei,
Teng Kew, Akbar, Geita, Sin Yee, Kak Hawani, Ann, Teh, Efa, Syahira, Husna, Ju, Zubed,
Bahiyah and all friends in INFORMM and any other places during my postgraduate study,
whom I fail to mention their names due to space constraint.

iii
On a personal note, I am deeply grateful to extend my warm gratitude to my ummi Chanisah
Hj Ahmad and my abi Basuni Hj Abdul Ghani for their prayers, encouragement and love.
Special thanks to all my family members especially my beloved brothers, sisters, nephews
and nieces whom have inspired, encouraged, supported and helped me indirectly in
completing this study. Ultimately, I am deeply thankful to Allah and all of those who
supported me in any aspect during this study.
The financial supports from EU grant (COINFECT), no. INCO-C7-2006-031714 and USM
Fellowship are gratefully acknowledged.

iv
TABLE OF CONTENTS
Page
Acknowledgements ii
Table of Contents iv
List of Tables ix
List of Figures xi
List of Abbreviations xiv
Abstrak xvi
Abstract xx
CHAPTER ONE
INTRODUCTION Page
1.1 Overview of parasitic infection 1
1.2 Intestinal parasitic infection 2
1.2.1 Distribution and prevalence of intestinal parasitic infection 2
1.3 Intestinal helminth
1.3.1 Overview 3
1.3.2 General life cycle of soil-transmitted intestinal helminth 4
1.3.3 Ascaris lumbricoides 5
1.3.4 Hookworm 9
1.3.5 Strongyloides stercoralis 13
1.4 Intestinal protozoa
1.4.1 Overview 18
1.4.2 General life cycle of intestinal protozoa 19
1.4.3 Entamoeba histolytica 19
1.4.4 Giardia lamblia 26
1.4.5 Cryptosporidium parvum 31
1.5 Multiple intestinal parasitic infections cases in individuals 35
1.6 Screening methods
1.6.1 Routine laboratory practice for intestinal parasites 35
1.6.2 Other diagnostic approaches and current problems 37
1.7 Nucleic acid based diagnostic methods
1.7.1 Overview 39
1.7.2 Polymerase chain reaction (PCR) 40

v
1.7.3 Real-time PCR 41
1.7.3.1 Methods of monitoring in real-time PCR amplification 43
1.7.3.2 Advantages of real-time PCR 45
1.7.3.3 Studies on intestinal helminths and intestinal protozoa in
Malaysia
45
1.8 Statement of the problem & rationale of the study 49
1.9 Objectives of the study 51
1.10 Flow chart for research approach 52
CHAPTER TWO
MATERIALS AND METHODS
2.1 Materials
2.1.1 Stool sample 53
2.1.2 Oligonucleotide primers and probes
2.1.2.1 Primers 54
2.1.2.2 Probes 54
2.1.3 Bacterial strain 58
2.1.4 Chemicals and reagents 58
2.1.5 Sterilization 58
2.1.6 Kits 59
2.1.7 Cloning vector 59
2.1.8 Laboratory equipments 59
2.1.9 Preparation of media
2.1.9.1 Luria-Bertani (LB) broth 59
2.1.9.2 SOC medium 60
2.1.9.3 LB agar 60
2.1.9.4 LB agar with ampicillin 60
2.1.10 Preparation of buffers and reagents
2.1.10.1 Phosphate buffered saline (PBS), pH 7.2 61
2.1.10.2 TE buffer, pH 7.0 61
2.1.10.3 Ampicillin (stock solution 100 mg/ml) 61
2.1.10.4 Ethanol (70%) 62
2.1.11 Preparation of reagent for preservation of sample
2.1.11.1 Sodium acetate-acetic acid-formalin (SAF) preservative 62
2.1.12 Preparation of reagents for parasitological techniques

vi
2.1.12.1 Normal saline [sodium chloride, 0.85% (w/v)] 62
2.1.12.2 Iodine solution 62
2.1.12.3 Acid ethanol (90%) 63
2.1.12.4 Zinc sulphate (ZnSO4) solution 63
2.1.12.5 HCl-ethanol (1%) 63
2.1.12.6 Malachite green (3%) 63
2.1.12.7 Glycerol-malachite green solution 63
2.1.13 Preparation of reagents for agarose gel electrophoresis
2.1.13.1 Tris-Borate-EDTA (TBE), buffer (10X) 64
2.1.13.2 Ethidium bromide (EtBr), solution 10 mg/ml (w/v) 64
2.1.14 Preparation of reagents for DNA extraction
2.1.14.1 Polyvinylpolypyrolidone (PVPP) 2% 64
2.1.15 Preparation of reagents for cloning
2.1.15.1 5-bromo-4-chloro-3-indolyl-ß-D-galactopyranoside
solution (X-gal), 40 mg/ml
65
2.1.15.2 Calcium chloride (CaCl2), 100 mM 65
2.1.15.3 Magnesium chloride (MgCl2), 100 mM 65
2.1.15.4 Glycerol (100%) 65
2.2 Methods
2.2.1 Parasitological detection 66
2.2.1.1 Direct smear 66
2.2.1.2 Zinc sulphate concentration/floatation method 67
2.2.1.3 Trichrome staining 68
2.2.1.4 Modified Ziehl-Neelsen technique (acid-fast stain) 68
2.2.1.5 Kato-Katz technique 69
2.2.2 DNA extraction 74
2.2.3 Phocine herpesvirus 1 (PhHV-1) optimization 75
2.2.4 Polymerase chain reaction (PCR) 76
2.2.4.1 Preparation of PCR master mix 76
2.2.4.2 Conventional PCR 76
2.2.4.2.1 Optimization of primer concentration for
conventional PCR
77
2.2.4.2.2 Selection of PCR thermal profile 77
2.2.4.3 Real-time PCR assay 78
2.2.4.3.1 Optimization of primer concentrations for
real-time PCR
79

vii
2.2.4.3.2 Optimization of multiplex real-time PCR
assay
80
2.2.4.3.3 Determination of detection limit 83
2.2.5 Agarose gel electrophoresis 83
2.2.6 Cloning
2.2.6.1 Preparation of competent cells 84
2.2.6.2 Cloning PCR product into TOPO® cloning vector 85
2.2.6.3 Transformation of vector plasmid into bacterial host 85
2.2.6.4 Screening of positive clones 86
2.2.6.5 Plasmid extraction 87
2.2.6.6 Determination of purity and concentration of DNA 88
2.2.6.7 DNA sequencing 89
2.2.7 Statistical analysis 89
CHAPTER THREE
RESULTS
3.1 Cloning of specific target regions of A. lumbricoides, Ancylostoma, N.
americanus, S. stercoralis, E. histolytica, G. lamblia and C. parvum
3.1.1 Optimization of primer concentrations for conventional PCR and
selection of PCR thermal profile
90
3.1.2 Screening of positive clones 96
3.1.3 Plasmid extraction 101
3.1.4 DNA sequencing 104
3.2 Parasitological techniques detection 107
3.3 Optimization of real-time PCR assay
3.3.1 Phocine herpesvirus 1 (PhHV-1) optimization 112
3.3.2 Optimization of primer concentrations for real-time PCR assay 115
3.3.3 Optimization of multiplex real-time PCR assay 117
3.3.4 Determination of detection limit 120
3.4 Evaluation of multiplex real-time PCR assay 123
3.4.1 Analysis of real-time PCR assay and the quantitation of DNA
(Median Ct-value of positive samples and the approximate DNA
copy number)
135
3.4.2 Distribution of parasite species 138

viii
3.4.3 Statistical comparison of the techniques, sensitivity and specificity
tests
141
CHAPTER FOUR
DISCUSSION
4.1 Intestinal parasitic infections 143
4.2 Parasitological techniques
4.2.1 Microscopic examination 144
4.2.2 Concentration techniques 146
4.2.3 Other diagnostic methods 148
4.3 DNA-based diagnostic method
4.3.1 PCR assay 150
4.3.2 PCR limitations 153
4.3.3 PCR assay optimization 154
4.3.4 Real-time PCR 159
4.3.5 Multiplex real-time PCR 163
4.4 Prevalence of intestinal parasitic infections 163
4.5 Research related to the present study
4.5.1 Intestinal parasitic studies 164
4.5.2 Intestinal helminths studies 165
4.5.3 Intestinal protozoa studies 166
4.6 Conclusion and further studies 170
REFERENCES 173
LIST OF PUBLICATIONS 189
APPENDICES
A - List of chemicals and reagents
B - List of equipments
C - Table of Corbett Rotorgene

ix
LIST OF TABLES
Table Page
2.1.a Sequences of primers for intestinal helminths.
55
2.1.b Sequences of primers for intestinal protozoa.
56
2.1.c Sequences of primers for internal control.
56
2.2 Sequences of probes.
57
2.3.a DNA template added in multiplexing and background test of
intestinal helminths with and without the presence of internal
control.
81
2.3.b DNA template added in multiplexing and background test of
intestinal protozoa with and without the presence of internal
control.
82
3.1 BLAST NCBI results of the DNA sequences from
recombinant plasmids.
106
3.2.a Detection of intestinal helminths by parasitological techniques.
108
3.2.b Detection of intestinal protozoa by parasitological techniques.
109
3.3.a Detection of intestinal helminths by specific parasitological
techniques.
110
3.3.b Detection of intestinal protozoa by specific parasitological
techniques.
111
3.4 Mean and standard deviation (SD) of the Ct-values obtained
for optimization of primer concentrations in real-time PCR
assay.
116
3.5.a Ct-values of multiplex real-time PCR assay for the
multiplexing and background test of intestinal helminths.
118
3.5.b Ct-values of multiplex real-time PCR assay for the
multiplexing and background test of intestinal protozoa.
119
3.6.a Mean Ct-values and standard deviation for the determination
of detection limit for intestinal helminths.
121
3.6.b Mean Ct-values and standard deviation for the determination
of detection limit for intestinal protozoa.
122
3.7.a Detection of intestinal helminths by multiplex real-time PCR.
124

x
3.7.b Detection of intestinal protozoa by multiplex real-time PCR.
125
3.8 Median Ct-value of positive samples by real-time PCR
(positive or negative by microscopy), and approximate DNA
copies.
137
3.9 Distribution of intestinal parasites according to the species
(N=302).
140
3.10 Statistical analysis on the comparison of detection by
microscopy and real-time multiplex PCR for detection of (i)
intestinal helminths and (ii) intestinal protozoa.
142

xi
LIST OF FIGURES
Figure Page
1.1.a A. lumbricoides unfertilized egg.
7
1.1.b A. lumbricoides fertilized egg.
7
1.1.c A. lumbricoides life cycle.
8
1.2.a Hookworm larva.
11
1.2.b Hookworm egg.
11
1.2.c Hookworm life cycle.
12
1.3.a S. stercoralis rhabditiform larva (L1) in iodine stained
smear.
16
1.3.b S. stercoralis short buccal cavity in unstained wet mount.
16
1.3.c S. stercoralis filariform larva (L3) characteristic; notched
tail.
16
1.3.d S. stercoralis egg.
16
1.3.e S. stercoralis life cycle.
17
1.4.a Distribution of E. histolytica in human body.
23
1.4.b E. histolytica trophozoites in trichrome stained smear.
24
1.4.c E. histolytica cyst with four nuclei.
24
1.4.d E. histolytica life cycle.
25
1.5.a G. lamblia iodine stained cyst.
29
1.5.b G. lamblia trophozoites.
29
1.5.c G. lamblia life cycle.
30
1.6.a C. parvum oocyst in modified acid-fast stained smear.
33
1.6.b C. parvum oocyst in wet mount.
33
1.6.c C. parvum life cycle.
34
1.7 Amplification plot of real time PCR.
42
2.1.a - 2.1.g Standard procedure of Kato-Katz technique. 70

xii
3.1.a - 3.1.b Optimization of primer concentrations and selection of PCR
thermal profiles for Ancylostoma.
92
3.2.a - 3.2.b Optimization of primer concentrations and selection of PCR
thermal profiles for A. lumbrocoides.
92
3.3.a - 3.3.b Optimization of primer concentrations and selection of PCR
thermal profiles for N. americanus.
93
3.4.a - 3.4.b Optimization of primer concentrations and selection of PCR
thermal profiles for S. stercoralis.
93
3.5.a - 3.5.b Optimization of primer concentrations and selection of PCR
thermal profiles for E. histolytica.
94
3.6.a - 3.6.b Optimization of primer concentrations and selection of PCR
thermal profiles for G. lamblia.
94
3.7.a - 3.7.b Optimization of primer concentrations and selection of PCR
thermal profiles for C. parvum.
95
3.8 Screening of positive clones that has inserted target gene for
Ancylostoma.
97
3.9 Screening of positive clones that has inserted target gene for
A. lumbricoides.
97
3.10 Screening of positive clones that has inserted target gene for
N. americanus.
98
3.11 Screening of positive clones that has inserted target gene for
S. stercoralis.
98
3.12 Screening of positive clones that has inserted target gene for
G. lamblia.
99
3.13 Screening of positive clones that has inserted target gene for
C. parvum.
99
3.14 Screening of positive clones that has inserted target gene for
E. histolytica.
100
3.15 Agarose gel electrophoresis analysis of recombinant
plasmids for Ancylostoma.
102
3.16 Agarose gel electrophoresis analysis of recombinant
plasmids for N. americanus.
102
3.17 Agarose gel electrophoresis analysis of recombinant
plasmids for A. lumbrocoides.
102

xiii
3.18 Agarose gel electrophoresis analysis of recombinant
plasmids for S. stercoralis.
102
3.19 Agarose gel electrophoresis analysis of recombinant
plasmids for E. histolytica.
103
3.20 Agarose gel electrophoresis analysis of recombinant
plasmids for G. lamblia.
103
3.21 Agarose gel electrophoresis analysis of recombinant
plasmids for C. parvum.
103
3.22.a Positive amplification curves and the Ct-values of real-time
PCR for determination of optimized PhHV-1 dilution prior
to DNA extraction.
113
3.22.b Analysis of PCR product for determination of optimized
PhHV-1 dilution prior to DNA extraction based on agarose
gel electrophoresis.
114
3.23.a Quantitation data for Cycling B red channel (for detection of
PhHV-1).
126
3.23.b Quantitation data for Cycling B orange channel (for
detection of A. lumbricoides).
127
3.23.c Quantitation data for Cycling B green channel (for detection
of N. americanus).
128
3.23.d Quantitation data for Cycling B yellow channel (for
detection of Ancylostoma).
129
3.23.e Quantitation data for Cycling B crimson channel (for
detection of S. stercoralis).
130
3.24.a Quantitation data for Cycling B red channel (for detection of
PhHV-1).
131
3.24.b Quantitation data for Cycling B yellow channel (for
detection of E. histolytica).
132
3.24.c Quantitation data for Cycling B green channel (for detection
of G. lamblia).
133
3.24.d Quantitation data for Cycling B orange channel (for
detection of C. parvum).
134

xiv
LIST OF ABBREVIATIONS
Description Abbreviations
1 5-bromo-4-chloro-3-indolyl-ß-D-galactopyranoside X-gal
2 Acquired Immune Deficiency Syndrome AIDS
3 Base pair bp
4 Basic Local Alignment Search Tool BLAST
5 Black hole quencher BHQ
6 Bovine serum albumin BSA
7 Calcium chloride CaCl2
8 Calcium chloride dehydrate CaCl2.2H2O
9 Celcius C
10 Centers for Disease Control and Prevention CDC
11 Centimeter cm
12 Complementary DNA cDNA
13 Cycle threshold Ct
14 Deoxyribonucleic acid DNA
15 Direct immunofluorescence assay DFA
16 Disodium hydrogen phosphate Na2HPO4
17 Double-stranded DNA dsDNA
18 Eggs per gram EPG
19 Enzyme-linked immunosorbent assay ELISA
20 Ethidium bromide EtBr
21 Ethylenediaminetetraacetic acid EDTA
22 Fluorescence resonance energy transfer FRET
23 Full examination microscopic examination FEME
24 High-performance liquid chromatography HPLC
25 Hydrochloric acid HCl
26 Immunofluorescence assay IFA
27 Indirect hemagglutination assay IHA
28 Internal transcribed spacer ITS
29 Iodine I2
30 Larvae stage 1 L1
31 Larvae stage 2 L2
32 Larvae stage 3 L3
33 Larvae stage 4 L4
34 Loop-mediated isothermal DNA amplification LAMP
35 Magnesium chloride MgCl2
36 Microliter µl
37 Micrometer µm
38 Micromolar µM
39 Mililiter ml/mL
40 Milligram mg
41 Millimeter mm
42 Milimolar mM
43 Minor groove binder MGB

xv
44 Multiplex tandem PCR MT-PCR
45 Nanometer nm
46 Nanomolar nM
47 National Center for Biotechnology Information NCBI
48 Non fluorescent quencher NFQ
49 Non template control NTC
50 Number No.
51 Optical density OD
52 Phosphate buffered saline PBS
53 Plaque forming unit PFU
54 Polyvinylpyrrolidone PVPP
55 Polymerase chain reaction PCR
56 Potassium dihydrogen phosphate KH2PO4
57 Potassium iodide KI
58 Quantitative polymerase chain reaction qPCR
59 Random amplification of polymorphic DNA RAPD
60 Restriction fragment length polymorphism RFLP
61 Revolutions per minute rpm
62 Ribonucleic acid RNA
63 Ribosomal DNA rDNA
64 Single-stranded DNA ssDNA
65 Small subunit ribosomal RNA SSUrRNA
66 Sodium acetate NaOAc
67 Sodium chloride NaCl
68 Soil-transmitted helminth STH
69 Specific gravity SG
70 Synergy Brands, Inc. SYBR
71 Tetramethylrhodamine TAMRA
72 Tris-borate-EDTA TBE
73 Ultra violet UV
74 Unit U
75 Weight per volume w/v
76 World Health Organization WHO
77 Zinc sulphate ZnSO4
78 Zinc sulphate heptahydrate ZnSO4.7H2O

xvi
PENGESANAN PARASIT PATOGENIK USUS MELALUI KAEDAH ‘REAL
-TIME MULTIPLEKS PCR’ DAN PERBANDINGAN DENGAN TEKNIK-
TEKNIK PARASITOLOGI
ABSTRAK
Jangkitan parasit dalam usus oleh helmin (cacing) dan protozoa adalah antara
jangkitan yang paling biasa berlaku dan kekal sebagai masalah besar di kalangan
masyarakat awam terutamanya di negara membangun. Jangkitan parasit usus yang
berkaitan dengan helmin biasanya menyebabkan kesakitan di bahagian abdomen dan
disertai oleh anereksia, loya dan cirit-birit, manakala protozoa usus lazimnya
menyebabkan cirit-birit.
Kaedah pengesanan rutin yang dijalankan untuk mengesan jangkitan parasit usus
biasanya bergantung kepada pengesanan dengan mikroskop. Bagaimanapun, kaedah
ini tidak sensitif dan memerlukan kepakaran mikroskopi untuk mengelakkan
‘misdiagnosis’ mahupun ‘overdiagnosis’. Kekurangan ini telah mencetuskan
pembangunan kaedah yang lebih sensitif menggunakan DNA. PCR adalah satu
kaedah yang terbukti sensitif dan spesifik untuk mengesan kehadiran organism yang
patogenik di dalam usus. Memandangkan PCR secara konvensional mengambil masa
panjang serta mudah terdedah kepada pencemaran, maka pembangunan kaedah
pengesanan ‘real-time PCR’ yang pantas dan mampu memberikan data secara
langsung dan kuantitatif ke atas produk PCR adalah bermanfaat.
Dalam kajian ini, satu kaedah ‘real-time multipleks PCR’ untuk mengesan spesies
helmin usus yang berkaitan rapat iaitu Ascaris lumbricoides, Strongyloides

xvii
stercoralis, Necator americanus dan Ancylostoma duodenale dan satu kaedah ‘real-
time multipleks PCR’ untuk mengesan protozoa usus yang juga berkaitan rapat iaitu
Entamoeba histolytica, Cryptosporidium parvum dan Giardia lamblia telah
dioptimumkan dan dinilai. Keputusan yang diperolehi telah dibandingkan dengan
keputusan yang didapati menggunakan teknik parasitologi.
Sebanyak 302 sampel tinja telah dipungut daripada pesakit yang mengalami masalah
gastrousus daripada Hospital Serian, Hospital Lundu dan Hospital Universiti Sains
Malaysia. Teknik parasitologi yang dilakukan ke atas sampel tersebut adalah kaedah
smir langsung, pengapungan / pemendapan dengan larutan zink sulfat, pewarnaan
trikrom untuk E. histolytica dan G. lamblia, pewarnaan ‘acid fast’ untuk C. parvum
dan kaedah Kato-Katz untuk mengesan ova helmin.
Primer yang digunakan dalam kajian ini telah diuji menggunakan lima tahap
kepekatan primer dan dua profil suhu PCR. Produk PCR diklon ke dalam vektor
pengklonan TOPO® dan plasmid DNA yang diperolehi disimpan sebagai kawalan
positif untuk pembinaan lengkung piawai (‘standard curve’). Daripada lengkung
piawai yang diperolehi, had pengesanan bagi setiap organisma adalah seperti berikut:
satu salinan DNA untuk G. lamblia, 10 salinan DNA untuk Ancylostoma, A.
lumbrocoides dan S. stercoralis, dan 102 salinan DNA untuk N. americanus, E.
histolytica dan C. parvum. Untuk kaedah ‘real-time multipleks PCR’,
pengoptimuman had kepekatan primer juga dilakukan untuk mengelakkan
perencatan kaedah PCR yang mungkin berlaku disebabkan oleh templat DNA yang
mempunyai ketumpatan tinggi. Phocine herpesvirus 1(PhHV-1) dengan kepekatan
yang optimum iaitu 10-2
PFU/ml telah dimasukkan bersama dalam setiap

xviii
pengekstrakan templat, ini bertujuan untuk mengesan sebarang keputusan negatif
palsu yang mungkin disebabkan oleh kehadiran bahan perencat dalam sampel tinja
ataupun disebabkan kegagalan proses PCR.
Setiap tindak balas yang dijalankan diuji dengan satu atau beberapa templat DNA
dalam satu tiub, samaada dengan kehadiran DNA daripada PhHV-1 ataupun tanpa
kehadiran DNA tersebut. Dua pengesanan menggunakan kaedah ‘real-time
multipleks PCR’ dilakukan ke atas semua DNA, iaitu satu pengesanan untuk helmin
dan pengesanan untuk protozoa. Keputusan yang diperolehi dibandingkan dengan
keputusan teknik parasitologi berdasarkan ujian sensitiviti, spesifisiti dan Chi-square.
Berdasarkan keputusan daripada 302 sampel yang diuji, untuk kaedah pengesanan
cacing, 13 (4.3%) sampel positif dikesan melalui kaedah mikroskopi manakala
94 (31.1%) sampel positif dikesan menggunakan kaedah ‘real-time PCR’. Untuk
pengesanan protozoa pula, lima (1.7%) sampel positif dikesan melalui kaedah
mikroskopi manakala 23 (7.6%) sampel positif dikesan menggunakan kaedah ‘real-
time PCR’ (p <0.05). Jangkitan oleh dua atau lebih parasit usus (samaada helmin
atau protozoa) dicatatkan dalam 29 daripada 110 sampel yang positif (26%)
menggunakan kaedah ‘real-time PCR’ manakala kaedah mikroskopi tidak dapat
mengesan kehadiran jangkitan berganda.
Kedua-dua kaedah ‘real-time multipleks PCR’ yang telah dioptimumkan dalam
kajian ini telah berjaya mengesan semua organisma yang disasarkan. Kaedah-kaedah
ini juga telah dinilai ke atas sampel pesakit dan berjaya mengesan empat jenis cacing
usus dan tiga jenis protozoa usus yang penting. Untuk pengesanan cacing, kaedah

xix
PCR berjaya mengesan 7.2 kali lebih banyak sampel positif manakala untuk
pengesanan protozoa, kaedah PCR mengesan 4.6 kali lebih banyak sampel positif
berbanding teknik parasitologi.
Kesimpulannya, kaedah ‘real-time PCR’ dalam kajian ini menyediakan satu kaedah
alternatif untuk diagnosis parasit usus. Kaedah ini boleh diaplikasikan untuk
diagnosis rutin pesakit, pemantauan rawatan dan kajian epidemiologi berkaitan
jangkitan parasit usus.

xx
MULTIPLEX REAL-TIME PCR FOR THE DETECTION OF PATHOGENIC
INTESTINAL PARASITES AND COMPARISON WITH
PARASITOLOGICAL TECHNIQUES
ABSTRACT
Intestinal parasitic infections by helminths and protozoa are among the most
prevalent infections and remain a major public health burden in underdeveloped
countries. Most intestinal helminth infections cause abdominal pain accompanied by
aneroxia, nausea and diarrhea while intestinal protozoa cause diarrheal diseases.
Routine diagnostic methods for intestinal parasitic infections which rely heavily on
microscopic detection are insensitive, and require well-trained microscopists to avoid
misdiagnosis or overdiagnosis of the infection. These limitations have led to the
development of highly sensitive DNA-based assays. PCR has been proven to be
sensitive and specific for detection of enteric pathogens. Since conventional PCR is
time consuming and prone to cross-contamination, it is desirable to develop a real-
time PCR assay which is rapid and can provide quantitative and real-time
information on the amplified products.
In this study, a real-time multiplex PCR assay for the detection of closely related
intestinal helminths namely Ascaris lumbricoides, Strongyloides stercoralis, Necator
americanus and Ancylostoma duodenale and an assay for the detection of three
closely related intestinal protozoa which comprised Entamoeba histolytica,
Cryptosporidium parvum and Giardia lamblia were optimized and evaluated.

xxi
A total of 302 stool samples were collected from patients with gastrointestinal
problems from Hospital Serian, Hospital Lundu and Hospital Universiti Sains
Malaysia. Parasitological techniques were performed by direct wet smear, zinc
sulphate floatation/sedimentation, trichrome staining for E. histolytica and G.
lamblia, acid-fast staining for C. parvum and Kato Katz technique for helminth ova.
The primers were tested at five concentrations and two PCR thermal profiles. The
PCR products were cloned into TOPO® cloning vector and the DNA plasmids were
used as positive control for standard curve construction. The detection limit for each
organism was as follows: one DNA copy for G. lamblia; 10 copies for each
Ancylostoma, A. lumbrocoides and S. stercoralis; and 102 copies for each N.
americanus, E. histolytica and C. parvum. For real-time multiplex PCR assay
optimizations, primer limitation steps were performed in order to avoid any
inhibition due to high abundant template. Phocine herpesvirus 1 (PhHV-1) with an
optimum dilution of 10-2
PFU/ml was included at the template preparation stage in
order to detect false negative results due to the presence of any inhibitor compounds
or PCR failure. Each assay was tested with one or multiple DNA template with or
without the addition of PhHV-1 DNA in each reaction. Two real-time multiplex PCR
assays i.e. for detection of intestinal helminths and intestinal protozoa were
performed on all DNA samples. The real-time PCR results were compared with those
obtained by parasitological techniques based on sensitivity, specificity and Chi-
square tests.
For the detection of intestinal helminths, out of 302 samples, microscopic
examination detected 13 (4.3%) positive samples while real-time PCR assay detected

xxii
94 (31.1%). For the detection of intestinal protozoa, microscopic examination
detected five (1.7%) positive cases and real-time PCR detected 23 (7.6%) with
p<0.05. Multiple infections by two and more organisms (either helminths or
protozoa) were recorded in 29 out of 110 positive cases (26%) by real-time multiplex
PCR, while no cases of multiple infections were reported by microscopic
examination.
The real-time multiplex PCR assays optimized in this study successfully detected all
the target organisms. They were also successfully evaluated on patients’ samples for
the detection of four important intestinal helminths and three common intestinal
protozoa. The PCR assay detected 7.2 times more positive samples for intestinal
helminths and 4.6 times for intestinal protozoa as compared to parasitological
techniques.
In conclusion, the real-time PCR assays described in this study provide an alternative
laboratory diagnostic method for intestinal parasitic infections and would be useful
for treatment monitoring and epidemiological studies.

1
CHAPTER ONE
INTRODUCTION
1.1 Overview of parasitic infection
Parasitic infections are often caused by two most common type of organisms which
are protozoa and helminths. Protozoa is single-celled organism meanwhile helminth
is multicellular organism. Parasitic infection occurs when organisms live on or
within a living host (usually mammals e.g. human). In general, parasite harms its
host or survives by utilizing the host nutrients (http://www.cdc.gov/parasites).
Parasitic infections not only cause morbidity but also mortality in humans (Haque,
2007). Children and the elderly are highly susceptible to parasitic infections
compared to other age groups due to their weakened immune systems and less
capability to respond effectively to infections (Nagamani et al., 2007).
Immunocompromised or immunosuppressed people in cases of injury, surgery or
chronic illness are also more vulnerable to acquire serious infections (Graczyk and
Fried, 2007). In addition, children who are more exposed to soil when playing, have
higher chances to be infected with soil-transmitted helminths. Parasitic infections are
also responsible for many ill health conditions. The symptoms usually depend on the
type of the parasite and the affected organ of the host. Moreover, prolonged or
undetected infection may cause systemic problems which can affect the whole
system of the human body. Most of the parasites undergo a complicated life cycle
which includes the sexual and asexual types of reproduction within mammalian or
invertebrate hosts.

2
1.2 Intestinal parasitic infections
Parasitic infections caused by intestinal helminths and protozoan parasites are among
the most prevalent infections among humans in underdeveloped and developing
countries (Haque, 2007). It is also among the leading causes of death and disease in
tropical and subtropical regions of the world. Intestinal parasites refer to organisms
that live in the intestine and consume the nutrition of the host. Common
complications due to intestinal parasitic infections are abdominal discomfort,
dysentery or mechanical irritation of intestinal mucosa and other general symptoms
such as bloating, diarrhea and fever. Furthermore, when the burden of the infection is
prominent, intestinal parasites may also cause serious health conditions and problems
such as malnutrition, mental retardation and death especially in children (Amuta et
al., 2009).
1.2.1 Distribution and prevalence of intestinal parasitic infections
Intestinal parasites are distributed worldwide and many countries have high
prevalence rates of the infection. It is estimated that there are more than three billion
people infected with intestinal parasites throughout the world (Balcioglu et al., 2007;
Kurt et al., 2007). Several factors affect the distribution and the prevalence of
intestinal parasites. Personal hygiene, dietary habits, education level of the
community, socio-economic status and climates are among the most common factors
that influence the prevalence of intestinal parasitic infections (Balcioglu et al., 2007;
Mahsol et al., 2008). Furthermore, overcrowded areas with inadequate hygiene and
sanitation may increase transmission of parasitic infections. Therefore, the
prevalence of intestinal parasitic infection is usually high in urban slum communities

3
where the socio-economic status is low, environmental sanitation and living
condition is poor and water supply is unsafe with unhygienic personal habits (Noor
Azian et al., 2007).
1.3 Intestinal helminths
1.3.1 Overview
Intestinal helminths are macroparasites or multicellular organisms. Intestinal
helminths include nematodes (roundworms), cestodes (tapeworms) and trematodes
(flatworms) which live in the human gut. The common nematodes are the large
roundworm Ascaris lumbricoides, the whipworm Trichuris trichiura, two species of
blood-feeding hookworm Necator americanus and Ancylostoma duodenale, and the
threadworm Strongyloides stercoralis (Chigozie et al., 2007; Haque, 2007). They are
known as geohelminths or soil-transmitted helminths (STHs).
STHs infect more than a billion people and among the most common cause of
chronic infection to human (de Gruijter et al., 2005; Wiria et al., 2010). Recent
estimates suggest that A. lumbricoides infects 1221 million people, T. trichiura
infects 795 million, and hookworms infect 740 million people (de Silva et al., 2003).
Furthermore, infections with S. stercoralis, which affect 30 to 100 million people
worldwide, are probably even more underestimated (Wiria et al., 2010).
Commonly, infection with STH does not typically result in clinical disease.
However, the effect of infections depends on several factors namely the helminth

4
species, the intensity of infection and the host immunological status. Majority of the
infected individuals exhibit asymptomatic infections. This is because the pathology
of STH is strongly related to the intensity of infection which is the number of worms,
and usually, only a few worms are found in human body especially in early infection.
The most common symptoms of intestinal helminth infection include diarrhea, foul
breath, headache, nausea, abdominal pain and itching. Constipation and bloating can
also arise, due to the intestinal organs obstruction (Baba et al., 2009). Other
symptoms associated with intestinal helminth infections are anaemia and asthma (da
Silva et al., 2008; Dori et al., 2011).
1.3.2 General life cycle of soil-transmitted intestinal helminths
Intestinal helminth infection can be transmitted by fecal-oral route in the course of
contact with parasite eggs or larvae in contaminated soil. Usually, infection occurs
through accidental ingestion of A. lumbricoides or T. trichiura eggs or penetration of
the skin by hookworm or S. stercoralis larvae. Intestinal helminths do not divide in
the host (Haque, 2007) but they reproduce sexually. Male and female adult worms
mate in the intestinal of the host and produce eggs. Adult worms inhabit in specific
part of the host intestine; A. lumbricoides, hookworm and S. stercoralis live in the
small intestine while T. trichiura inhabits the colon. Adult worms can produce large
number of eggs and are able to survive for several years in the host, depending on the
species. The female worms then discharge their eggs or larvae in human feces (Garg
et al., 2005).

5
Depending on the environmental conditions, the eggs (of A. lumbricoides and T.
trichiura) can remain viable in the soil for several months while larvae (of
hookworms and S. stercoralis) can remain viable for several weeks. Higher humidity
of external environment is associated with faster development of the eggs to hatch
into larvae. Some of the helminth larvae (namely A. duodenale and S. stercoralis)
can undergo hypobiosis in the human body for several months. Hypobiosis is an
arrested development at a specific point in the nematode life cycle. In this stage, the
larvae do not grow and stop moving. They can survive for weeks or months before
resuming development and may be resistant to some antihelminthics.
1.3.3 Ascaris lumbricoides
Ascaris lumbricoides is one of the most common and most widespread human
infections, although Ascaris suum (the pig nematode) has also produced human
infection. It is estimated that A. lumbricoides causes 1.2 million cases of acute illness
and 10000 deaths annually. A. lumbricoides infection is rarely found in developed
countries. However the infection rate is increased with travelling and migration in
developing countries. The infection by A. lumbricoides is known as ascariasis. It
occurs by ingestion of the infective Ascaris eggs which hatch into larvae in the small
intestine. The larvae migrate to the caecum and proximal colon where they penetrate
the mucosa. The larvae then move via the portal blood to reach the liver. After
migration in the liver, the larvae advance to the lungs, penetrate the alveolar space
and move to the pharynx where they are swallowed and return to the small intestine.
Upon reaching the small intestine, they develop into adult worms, mate and produce
eggs (Dold and Holland, 2011). The morphology of A. lumbricoides eggs in

6
diagnosis stage are shown in Figures 1.1.a - 1.1.b while the complete life cycle of
Ascaris is further explained in Figure 1.1.c.
Infection of A. lumbricoides may be asymptomatic. Though, it may become life
threatening as seen with hepatobiliary and intestinal obstructions due to heavy worm
burdens (de Silva et al., 1997; Crompton et al., 1999). Hepatobiliary and intestinal
obstruction are the main causes of morbidity and mortality due to ascariasis.
Otherwise the infection may contribute to impaired nutrition, development, and
educational progress especially in children (Hlaing, 1993).
In the stage when the larvae migrate to the pulmonary system, eosinophilic
pneumonitis or Loeffler syndrome may develop. It happens when the eosinophils
accumulate in the lung due to tissue destruction and releasing of the larval antigens.
During this period, the patient may also develop asthma with hypersecretion of
mucus, bronchiolar inflammation or has sputum containing eosinophils. In allergy-
prone people, urticaria or itchiness of the skin may also occur. Infrequently, the
larvae emerge through fistulae or fallopian tubes, urinary bladder, lungs or heart and
pancreatic or bile duct (Noor Azian et al., 2007; Varkey et al., 2007). Women are
believed to be more affected since the progesterone hormones play a role in inducing
Oddi’s sphincter relaxation which allows the nematode to access the biliary duct
(Galzerano et al., 2010). However, the most intense infections usually occur in
children aged five to fifteen years due to higher exposure to contaminated
environment and low immune system of the children. Treatment with albendazole
was reported to give 93% reduction in egg count and a better cure rate (85% to
100%) than ivermectin treatment (78.4%) (Belizario et al., 2003).

7
Figure 1.1.a
Ascaris lumbricoides unfertilized egg. Elongated and covered by a
visible mammillated layer.
Figure 1.1.b
Ascaris lumbricoides fertilized egg. Rounded, has a thick shell and
smaller than unfertilized egg.
Source: http://www.phsource.us/PH/PARA/Diagnosing_Medical_Parasites.pdf

8
Figure 1.1.c
Ascaris lumbricoides life cycle 1. The life cycle begins with the
production of 40 to 60 eggs by adult females living in the small
intestine and deposited into external environment (soil) 2. Eggs
become infectious within several weeks and transmitted to the human
host by ingestion or by inhalation of contaminated soil or dust 3.
Within the host, Ascaris larvae hatch in the jejunum, penetrate the
intestinal wall, and migrate by hepatic venules to the right heart and
pulmonary circulation 4. They subsequently break through into
alveolar spaces, ascend the trachea, and are swallowed back into the
intestine 5. They undergo a final moult and develop into adult worms
(15 cm to 40 cm), which mate and generate a new generation of eggs.
Notes Under normal condition, the time from ingestion of eggs to development of
new eggs is 10 to 12 weeks. Adult worms live for approximately one year
before expelled from human body. Occasionally, male-only adult worms
infection occur with yielding no eggs in stool while female-only adult worms
infection produce infertile eggs, which never become infectious.
Source: https://online.epocrates.com/u/2924908/Ascariasis/Basics/Etiology

9
1.3.4 Hookworm
Primarily two species of hookworms infect humans; Ancylostoma duodenale and
Necator americanus. The World Health Organization has catogerized A. duodenale
and N. americanus as very destructive parasites (WHO, 2005). N. americanus tends
to be prevalent in tropical climates, while A. duodenale is commonly found in a
cooler and drier environment. However, their geographical distributions overlap
considerably, and both species are endemic to many areas (Albonico et al., 1998;
Brooker et al., 2004; Hotez et al., 2005). Currently, it is estimated that 740 million
people are infected worldwide, and more than 80 million of them are severely
affected clinically (Bethony et al., 2006).
Both species of the hookworm are transmitted through percutaneous route. The eggs
are excreted in human feces. However, as opposed to Ascaris, hookworm eggs hatch
into infective larvae in the feces or appropriate soil conditions. The infective larvae
then infect humans by active invasion of the skin. They find and recognize their hosts
by the behavioural phases of activation, directed crawling and penetration (Haas et
al., 2005). The infective, third-stage filariform larvae (L3) penetrate human skin and
migrate via the circulatory system and the lung to finally reside as the adult stage (8
mm to 20 mm in length) usually in the duodenum. The adult worms can persist in the
host within one to two years before being eliminated from the intestine (Gasser et al.,
2008).
Iron-deficiency anaemia caused by intestinal blood loss is the most outstanding
feature of hookworm infection. Blood loss occurs when the adults worms attach via
their buccal capsule to the small intestines, rupture capillaries and suck the blood

10
from the intestinal mucosa (Albonico et al., 1998). The level of blood loss is
proportionally related to the number of adult worms that inhabit in the host intestine.
The risk and severity of iron-deficiency anaemia is described by the balance between
iron intake and iron loss from intestinal bleeding. In heavy infections, hookworm can
cause physical and mental retardation and deaths in children as well as adverse
maternal-fetal outcomes in pregnant women (Hotez et al., 2005).
A. duodenale is capable of oral infection through accidental ingestion of the eggs
(Hoagland and Schad, 1978; Schad, 1991). Furthermore, A. duodenale larvae may
undergo a dormant state (arrested stage of development in the musculature and/or
intestine) after penetrating into the skin (Albonico et al., 1998). Infection by A.
duodenale may occur via transmammary routes; whereas N. americanus requires the
transpulmonary migration routes (Setasuban et al., 1980).
Currently, the control of hookworms has relied predominantly on antihelminthics
drugs such as albendazole, mebendazole, pyrantel pamoate or levamisole (Bethony et
al., 2006). The morphology and the life cycle of hookworm are summarised in
Figures 1.2.a, 1.2.b and 1.2.c.

11
Figure 1.2.a
Hookworm larva. General morphology of both hookworm larvae is
similar. Approximate length; 10 to 13µm for female and 8 to 11 µm
for male worm.
Figure 1.2.b Hookworm egg. General morphology of both hookworm eggs is
similar. Oval shaped, 56 to 75 µm by 36 to 40 µm in size, with
transparent and smooth thin shell.
Source: http://www.phsource.us/PH/PARA/Diagnosing_Medical_Parasites.pdf

12
Figure 1.2.c Hookworm life cycle 1. Hookworm eggs are passed in the stool. Under
favourable condition; warmth, humidity and shade, the eggs develop
into rhabditiform (L1) larvae 2. Rhabditiform larvae hatch within one
to two days 3. The larvae grow and develop in the feces and/or the
soil. After five to ten days (including two moults) they become
filariform (L3) larvae which are infective to the human host. These
ensheathed larvae can survive for three to four weeks under favourable
environmental conditions 4. Upon contact with the suitable host, the
larvae penetrate through the skin of the host 5. The larvae transported
via the blood stream to the heart and then to the lungs. They migrate
into the pulmonary alveoli, ascend the airways to the pharynx 6. They
are then swallowed and reach the small intestine and develop into
mature adults. The adult worms attach to the small intestinal wall and
feed on blood 7. The worms copulate, and the females produce
fertilized eggs, which are released in the feces.
Source: http://sprojects.mmi.mcgill.ca/tropmed/disease/intest-hookworm/life.htm

13
1.3.5 Strongyloides stercoralis
Strongyloides sp. can infect mammals, birds, reptiles and amphibians. To date, only
two species of Strongyloides have been known to be able to infect humans;
Strongyloides stercoralis and Strongyloides fuelleborni. However the most common
and clinically important pathogenic species in humans is S. stercoralis whereas S.
fuelleborni is found periodically in Africa and Papua New Guinea (Ashford et al.,
1992). S. stercoralis inhabits the small intestine of the host. Abdominal pain, nausea,
vomiting, wheezing, and asthma are the common symptoms of S. stercoralis
infection. During the infection, immunocompetent individuals may produce a
moderate clinical symptom of diarrhea whereas it can be fatal in
immunocompromised patients (Adenusi et al., 2003; Olsen et al., 2009; Feely et al.,
2010).
Strongyloides can undergo two types of development which are known as
heterogenic development and homogenic development. In heterogonic development,
Strongyloides eggs hatch in the feces and develop into L1 (first-stage larvae). L1
undergo L2, L3 and L4 stages and develop into rhabditiform male and female worms
which are the free-living nematodes. These rhabditiform or the free-living nematodes
will mate and the female lays eggs which hatch to release L1 which moult to an L2,
then into infective filariform L3. These infective L3 are long lived and can persist in
the environment until they come across a suitable host. This type of development is
also known as indirect or sexual development. On the other way, homogenic
development occurred when the L1 larvae is directly moult via L2 into infective L3
and persist in the environment until they encounter a suitable host. This type of
development is also known as direct or asexual development (Viney and Lok., 2007).

14
Only the female worms of Strongyloides will be infective to the host. The female can
either turn into L3 (infective) or free-living adult (non pathogenic) while the male
worm can only develop into a free-living nematode. The infective larvae (L3) can
penetrate into the skin host and migrate through the lungs until they are swallowed to
reach the small intestine or else, they migrate in the host and moult via an L4 stage to
develop adult parasitic female worms (Olsen et al., 2009).
S. stercoralis also undergoes a dormant state as the same as A. duodenale. This
dormancy of infective L3 larvae in human can cause chronic infection. At this stage,
the infective L3 larvae can also cause transmammary transmission by re-activation of
the larvae at lactation after their migration and arrestment in the mammary glands
(Miller, 1981). Autoinfection can also occur in Strongyloides life cycle.
Autoinfection is repeated generations of development in the same host individual.
During autoinfection, the fast development of female larvae into infective L3 occurs
within the gut and penetrates directly into the tissues of the primary host. Therefore,
the whole life cycle is occurred within the host. During this time, infective L3 larvae
may distribute through many organs and tissues of the host as well as the
development of new female larvae in the gut (Vadlamudi et al., 2006; Viney and
Lok, 2007).
Strongyloides infections can be detected by the presence of Strongyloides eggs or
larvae in the feces. Direct examination of thick smear on fresh fecal sample will
reveal L1 stages and, or eggs. However, eggs are rarely found as they hatch in the
intestine. High intensity of Strongyloides L1 can be detected by direct smear.
Meanwhile, for lower intensity infections, larvae can be collected from feces using

15
concentration techniques. Fecal samples can also be grown to obtain the infective
L3s. In severe and disseminated infection, sputum testing is also done to detect
Strongyloides larvae (Smith et al., 1985; Maayan et al., 1987).
The drug of choice to treat Strongyloides infection is thiabendazole. However,
ivermectin is also effective especially for disseminated infection (Datry et al., 1994;
Viney and Lok, 2007). The morphology and the life cycle of S. stercoralis are
summarised in Figures 1.3.a, 1.3.b, 1.3.c, 1.3.d and 1.3.e.

16
Figure 1.3.a Strongyloides stercoralis rhabditiform larva (L1) in iodine stained
smear. Large genital primordium (arrow) with short buccal cavity, and
sharp tail.
Figure 1.3.b Strongyloides stercoralis short buccal cavity in unstained wet mount of
stool.
Figure 1.3.c Strongyloides stercoralis filariform larva (L3) characteristic; notched
tail.
Figure 1.3.d Strongyloides egg. Oval and thin shelled, similar morphology with
hookworm egg but smaller in size (50 to 58 by 30 to 34 µm). It is
rarely found in stool as they hatch in the intestine.
Source: http://www.med.cmu.ac.th/dept/parasite/nematode/ssrlarva.htm
http://www.tropicalmed.eu/Page/WebObjects/PageTropE.woa/wa/displayPage?name=Readi
ngCultureAgarMicro

17
Figure 1.3.e Strongyloides stercoralis life cycle. The shaded box indicates the host.
The figure shows three possible development of first stage larvae (L1)
1. The autoinfective cycle, unique to S. stercoralis, in which
development from the L1 to the autoinfective L3 (aL3) occurs within
the gut of the primary host. Autoinfection can lead to explosive cycles
of development and a highly pathogenic disseminated infection. All
post-parasitic male L1 develop to free-living adult males. Post-
parasitic female L1 passed in the feces may undergo development by
either of two alternative pathways 2. The homogonic cycle involves
direct development to the infective L3 (iL3) 3. Heterogonic
development involves development to the free-living female and,
following mating, production of a generation of free-living progeny.
All progeny of the free-living adults develop to the iL3.
Source: http://www.ncbi.nlm.nih.gov/books/NBK19663/figure/A14969/?report=objectonly

18
1.4 Intestinal protozoa
1.4.1 Overview
Protozoa are unicellular microparasites. Majority of the protozoa are non-pathogenic
commensals. However, some of them can cause severe disease under certain
circumstances. For example, Giardia (waterborne protozoa) produces mild diarrhea
in immunocompetent individuals though in immunocompromised people, the
infection can result in a severe disease. Intestinal protozoa are commonly found in
tropical countries or areas with poor sanitary conditions. They are the more common
causes of gastrointestinal infections in the developed countries as compared to
helminths (Haque, 2007). Most intestinal protozoa has complex life cycle which
enables the protozoa to adapt and replicate at high rates within the host (Garg et al.,
2005; Haque, 2007).
Entamoeba histolytica, Giardia lamblia, and Cryptosporidium parvum have been
recognized as the three most important intestinal parasitic protozoa that cause
diarrhea in human and other mammals (Marshall et al., 1997; Garcia et al., 2000;
Guy et al., 2004; Verweij et al., 2004; Wang et al., 2004; Coklin et al., 2007; Haque,
2007). These protozoan parasites are acquired orally through contaminated food and
water. In severe cases, they can produce chronic diarrhea which may lead to
malabsorption, weight loss, and dehydration. The diseases they cause are known as
amebiasis, giardiasis, and cryptosporidiosis respectively.

19
1.4.2 General life cycle of intestinal protozoa
A typical protozoan life cycle consists of cysts and trophozoites. Many intestinal
protozoa exhibit a similar life cycle. After ingestion by the host, the cysts transform
into trophozoites. Trophozoites is an active phase, in which protozoa are motile,
acquires nutrient and undergoes asexual replication. If desiccation or low nutrients
occurs, the trophozoites will secrete a thick wall that functions to protect them from
dehydration and enter into a dormant period. The trophozoites develop into cysts
instead of undergoing replication and is excreted with the feces, then ingested by the
next host. The cysts will convert back into trophozoites when conditions are
favourable.
Source: http://www.tulane.edu/~wiser/protozoology/notes/intes.html
1.4.3 Entamoeba histolytica
Amebiasis is a major cause of morbidity and mortality in the developing world
(Furrows et al., 2004). Currently, it is seen more frequently in developed country
through the increase in travel (Tanyuksel et al., 2005) and resulting in 100000 deaths
per year (Verweij et al., 2004). Several members of the genus Entamoeba infect
humans. E. histolytica and E. dispar is morphologically similar and previously
considered to be the same species but, genetic and biochemical data indicate that this

20
two species are distinct. However, E. histolytica or E. dispar can be distinguished
from another species of Entamoeba by having peripheral chromatin in the nuclear
and a smaller karyosome than Entamoeba coli and Entamoeba hartmanni.
Among various Entamoeba sp., only E. histolytica is considered pathogenic. E.
histolytica can cause a severe intestinal disease characterized by amebic dysentery.
Humans are the only host of E. histolytica. E. histolytica exhibits a typical fecal-oral
life cycle which consists of infectious cysts that are passed in the feces and
trophozoites that replicate within the large intestine. Infection occurs through the
ingestion of cysts primarily by consuming contaminated food and water. Once the
cysts ingested, excystation occurs to release the trophozoites. The trophozoites then
secrete enzymes that digest the intestinal lining and leads to a perforated colon and
peritonitis; an inflammation of the lining of the abdominal wall. Adherence of the
trophozoites, cytotoxicity, and disruption of the tissues are important factors in the
pathogenesis of E. histolytica (Ravdin, 1986; Petri et al., 2002).
In severe cases, dehydration and anaemia may result from the loss of fluids and
blood especially in children. Instead of infection in the intestinal tract of the host, the
amebas can also metastasize to other organs such as liver and lungs and produce
extraintestinal amebiasis. The liver is the most commonly affected organ and this is
probably due to the direct transport of trophozoites from the large intestine to the
liver via the hepatic portal vein and causing amebic liver abscesses. Haematogenous
spread of trophozoites to other sites, such as the lungs, brain, spleen or pericardium

21
and cutaneous lesions can also occur, although it is extremely rare (Pelton et al.,
2006; Shenoy et al., 2010; Maldonado et al., 2011).
Only about 10% of the infected individuals will develop symptomatic invasive
amebiasis and the remaining will undergo a non-invasive infection. The non-invasive
infection is often asymptomatic but can cause diarrhea or other gastrointestinal
symptoms (Tanyuksel et al., 2005). This non-invasive infection can persist to an
invasive amebiasis when the trophozoites invade the intestinal mucosa and kill the
epithelial cells to produce ulcers and dysentery.
Previous studies show anti-ameba humoral responses in both asymptomatic and
symptomatic E. histolytica infections (Sánchez et al., 2002). Thus, this suggests that
even in asymptomatic cases, there is a limited amount of invasion of the
trophozoites. The symptom of infection depends on the organ infected. Hepatic
infections are characterized by hepatomegaly, liver tenderness, and pain in the upper
right quadrant, fever and anorexia while the symptoms of pulmonary amebiasis
include cough, chest pain, dyspnea (difficult breathing) and fever. The sputum of the
infected patient may be purulent or blood-stained and contain trophozoites (Wiser,
2011).
The best diagnosis of amebiasis requires the demonstration of E. histolytica cysts or
trophozoites in feces or tissues (Fotedar et al., 2007). Fresh stools can be
immediately examined for motile trophozoites or preserved, stained and
microscopically examined for cysts or trophozoites. Cysts will tend to predominate

22
in formed stools and trophozoites in diarrheic stools. Trophozoites with ingested
erythrocytes are commonly found in the dysenteric feces (Wiser, 2011).
The extraintestinal infection is diagnosed by sigmoidoscopy for ulcers, especially in
more severe disease (Madanagopalan et al., 1968; Harries, 1982). Besides, the
aspirates or biopsies can be examined microscopically for trophozoites that are most
likely to be found at the abscess wall. Other methods such as antigen and antibody
detection tests using enzyme-linked immunosorbent assay (ELISA), direct
fluorescent antibody (DFA) or indirect haemagglutination (IHA) are available for
detection of E. histolytica (Fotedar et al., 2007).
The choice of drug for amebiasis depends on the clinical stage of the infection and
the location of infection (lumen or tissue). The treatment is using antibiotics which
kill the organism in the body, and followed or combined with an agent (lumenal anti-
amebic drugs) which kills the parasite throughout the intestine. Several antibiotics
are recommended for all symptomatic infections including metronidazole, tinidazole,
tetracycline, and chloroquine (Kimura et al., 2007). Iodoquinol, paromomycin or
diloxanidefuroate are the luminal agents to treat asymptomatic cases (Petri and
Singh, 1999; Wiser, 2011). In endemic areas where the rates of re-infection are high
and treatments are expensive, only symptomatic cases would be treated. However,
lumenal anti-amebic drugs are given to asymptomatic cyst passers to prevent the
progression to severe disease and to control the spread of the disease (Wiser, 2011).
The distribution of E. histolytica in human body, the morphology and the life cycle
of E. histolytica are summarised in Figures 1.4.a, 1.4.b, 1.4.c and 1.4.d.

23
Figure 1.4.a Distribution of Entamoeba histolytica in human body A. E. histolytica
is found primarily in the colon where it can live as a non-pathogenic
commensal or invade the intestinal mucosa B. The ameba can
metastasize to other organs via a blood stream; primarily involving the
portal vein and liver C. The ameba can also spread via a direct
expansion causing a pulmonary infection, cutaneous lesions or
perianal ulcers.
Source: http://www.tulane.edu/~wiser/protozoology/notes/intes.html
A
A A
A
B
B
B
C
C

24
Figure 1.4.b Entamoeba histolytica trophozoites in trichrome stained smear. Two
diagnostic characteristics are observed. Two trophozoites have
ingested erythrocytes, and all three have nuclei with small, centrally
located karyosomes.
Figure 1.4.c Entamoeba histolytica cyst with four nuclei.
Source: http://umanitoba.ca/science/zoology/faculty/dick/z346/entahome.html
25
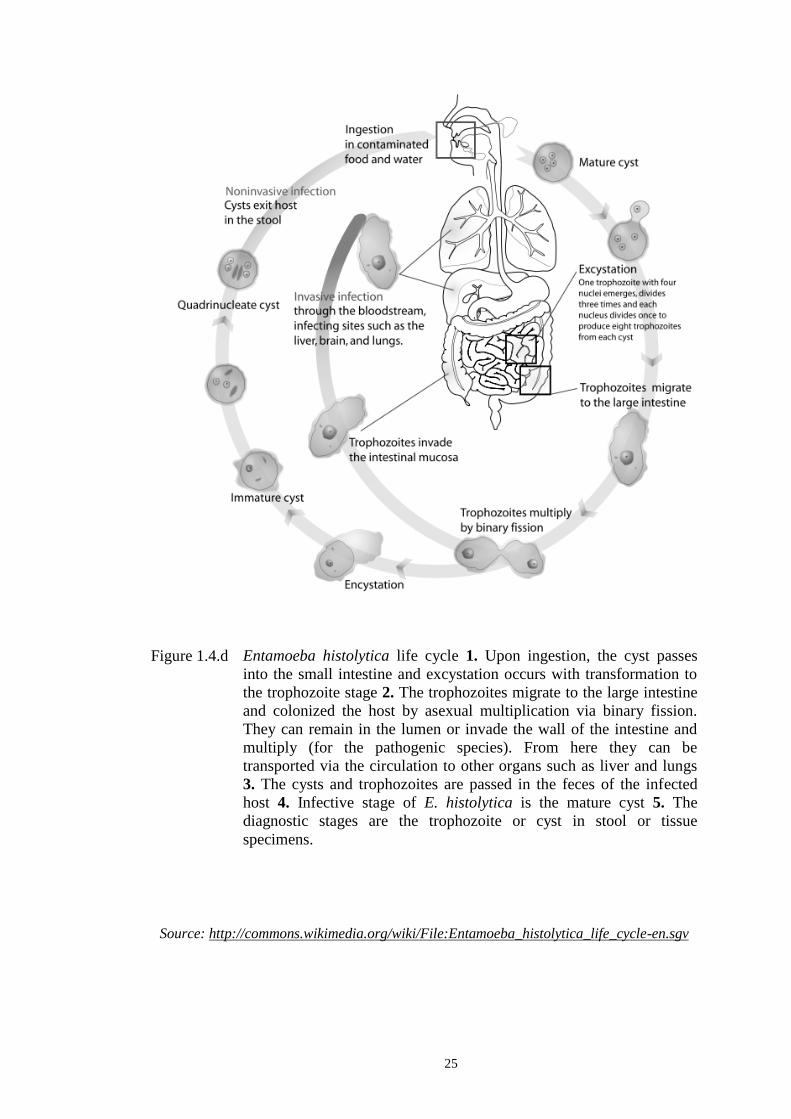
Figure 1.4.d Entamoeba histolytica life cycle 1. Upon ingestion, the cyst passes
into the small intestine and excystation occurs with transformation to
the trophozoite stage 2. The trophozoites migrate to the large intestine
and colonized the host by asexual multiplication via binary fission.
They can remain in the lumen or invade the wall of the intestine and
multiply (for the pathogenic species). From here they can be
transported via the circulation to other organs such as liver and lungs
3. The cysts and trophozoites are passed in the feces of the infected
host 4. Infective stage of E. histolytica is the mature cyst 5. The
diagnostic stages are the trophozoite or cyst in stool or tissue
specimens.
Source: http://commons.wikimedia.org/wiki/File:Entamoeba_histolytica_life_cycle-en.sgv
Related Documents











