Macronuclear Development in Conjugants of Tetrahymena thermophila, which Were Artificially Separated at Meiotic Prophase MAURYLA KIERSNOWSKA;' ANDRZEJ KACZANOWSKIa and JOANNA MORGA' "Department of' Cvtophyriology, University of Wursuw, 00-927 Warsaw,, Polund ABSTRACT. Conjugant pairs of Tetrahyrnenu rhermophilu were mechanically separated by vigorous pipetting at the early stages of meiotic prophase. The complete sequence of conjugational nuclear events including the appearance of pronuclei, development of the new macronuclei (postzygotic development), and resorption of the old macronuclei was observed in the separated cells, without pro- nuclear exchange. The pronuclei in the separated cells were recognised by the presence of components of the extranuclear cytoskeleton, which were labelled with anti-tubulin and anti-fenestrin antibodies in the same way as in undisturbed conjugants. The apical region of the separated conjugants (the post-junction area), corresponding to the junction area of conjugants was labelled with anti-fenestrin antibody and maintained the properties required for the nuclear development. The results of the genetic study were consistent with a hypothesis that cytogamy (pronuclear fusion) was induced in the separated conjugants. Therefore, the lasting cell contact is not necessary for the successful completion of conjugational nuclear events. Key Words. Cell junction, conjugation, cytogamy ONJUGAL pairing in Tetrahymena thermophila induces a C sequence of nuclear events: meiosis, prezygotic mitosis, pronuclear exchange and fusion, two postzygotic divisions, macronuclear development, and resorption of the old macro- nucleus (Cole and Soelter 1997; Cole et al. 1997; Kaczanowski and Kaczanowska 1996; Kaczanowski and Kiersnowska 1996; Kaczanowski et al. 1991; Martindele, Allis, and Bruns 1982; Nanney 1953; Sugai and Hiwatashi 1974; reviewed in Bruns 1986; Orias 1986). Development of macronuclear anlagen de- pends upon anterior positioning of two products of the post- zygotic divisions while two other products positioned posteri- orly are inhibited in this differentiation and remain as micro- nuclei (Kaczanowski and Kiersnowska 1996; Kaczanowski et al. 1991; Martindale, Allis, and Bruns 1982; Nanney 1953). Under standard conditions, conjugating cells remain paired in T. thermophila for about 10-11 h (Gaertig and Kaczanowski 1987; Kiersnowska, Kaczanowski, and De Haller 1993; Martin- dele, Allis, and Bruns 1982). By then, the separating conjugant cells contain macronuclear anlagen with 4C DNA content (Doerder and De Bault 1975; Kaczanowski et al. 1991) and are already advanced in resorption of the old macronucleus (stage 11 of macronuclear development (Cole and Soelter 1997; Kac- zanowski et al. 1991; Martindele, Allis, and Bruns 1982; Nan- ney 1953). The conjugation cell junction, which is a physical requirement for the reciprocal exchange of pronuclei, may also be a source of the subsequent developmental signals required for postzygotic development. In previous studies, cytogamy in T. thermophila was induced by treatment of the conjugants with the osmotic shock (Cole and Bruns 1992; Orias, Hamilton, and Flacks 1979) and this procedure was used to successfully screen for mutants (Cole and Soelter 1997; Cole et al. 1997; Melia, Cole, and Turkewitz 1998). This shock, applied to conjugants that were at the 3rd prezygotic division, or at the stage of pro- nuclei, prevented the pronuclear transfer, but allowed macro- nuclear development. These experiments leave open the possi- bility that, even in the absence of the pronuclear exchange, postzygotic development requires maintaining of cell contact at least until the 3rd prezygotic division (executed about 4 h after pair formation). In this study, conjugants of T. thermophila were mechanically separated as early as during meiotic pro- phase and then observed for further development to determine whether cytogamy may be induced in this way and whether Corresponding Author: M. Kiersnowska-Telephone number: 48-22- 620-0381 ext. 410; FAX number: 48-22-826-8624; Ernail: procyt@ warman.com.pl I Current address: Department of Immunology, Institute of Oncology, Warsaw, Poland. lasting cell contact is a prerequisite for progress through pre- and postzygotic conjugation nuclear events in Tetrahymena. MATERIALS AND METHODS Stocks. Two heterokaryon strains of different mating types and carrying the drug markers for cycloheximide resistance (chx-I) and for 6-methylpurine resistance (mpr-I) in their mi- cronuclei were used (Table l). They were kindly supplied by Dr J. Gaertig (University of Georgia, Athens, GA). Cells with micronuclear defects: strain A*V and the amicronucleate strain BI3840 were used as cytologically marked strains for studies on karyotypes. They were kindly obtained from Dr. J. Frankel (University of Iowa) and from Dr. A. Kaney (Bryn Mawr Col- lege, PA). Growth and induction of mating. The growth medium used throughout this study was PPY medium [1% proteose peptone (Difco) supplemented with 0.15% yeast extract (Difco)]. Stock cultures were maintained at room temperature, and experiments were performed at 30 "C. For starvation and conjugation, 10 mM Tris-HC1 medium (pH 7.4) was used. To induce mating, cells were starved for 20 h in the starvation medium and cells of different mating types were mixed in approximately equal numbers according to Bruns and Brussard (1974). Mechanical separation of conjugating pairs. To induce separation of the pairs, 5 ml of cell samples containing early conjugants (1.5-3.5 h. after mixing the cells) were transfered to a 12-ml vol. centrifuge tube and then vigorously pipetted with a 5-ml automatic pipette. In some experiments cells were also centrifuged for 2 min at 300 g. The efficiency of the cell separation was monitored under a dissecting microscope. The samples in which no more than 5% pairs remained were trans- ferred to small Erlenmeyer flasks and put on the shaker (EL- PAN, Poland) at 30 "C, in order to prevent further cell pairing. The shaking frequency was 220 g. Cytological methods. For routine staining, cell samples were air-dried on coverslips, fixed with Carnoy's fixative, and stained with Giemsa according to Gaertig, Kiersnowska, and Iftode (1988). For recording karyotypes of exconjugant strains, they were crossed with amicronucleate strain BI3840 (Kaney and Speare 1983) or with A*V strain which has extremely small aneuploid micronuclei (Doerder and Shabatura 1980). Four to five-hour-old conjugants were fixed with Shaudinn's fixative and stained with Giemsa according to Martindale, Allis, and Bruns (1982). Cytological stages of conjugation were distin- guished according to Martindale, Allis, and Bruns (1982), in- cluding stages I-VI of the meiotic prophase. Immunolabelling. Cortical structures. For visualisation of the cortical structures of the conjugants and the cytoskeletal structures involved in nuclear events, 3 antibodies were used: 139

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Macronuclear Development in Conjugants of Tetrahymena thermophila, which Were Artificially Separated at Meiotic Prophase
MAURYLA KIERSNOWSKA;' ANDRZEJ KACZANOWSKIa and JOANNA MORGA' "Department of' Cvtophyriology, University of Wursuw, 00-927 Warsaw,, Polund
ABSTRACT. Conjugant pairs of Tetrahyrnenu rhermophilu were mechanically separated by vigorous pipetting at the early stages of meiotic prophase. The complete sequence of conjugational nuclear events including the appearance of pronuclei, development of the new macronuclei (postzygotic development), and resorption of the old macronuclei was observed in the separated cells, without pro- nuclear exchange. The pronuclei in the separated cells were recognised by the presence of components of the extranuclear cytoskeleton, which were labelled with anti-tubulin and anti-fenestrin antibodies in the same way as in undisturbed conjugants. The apical region of the separated conjugants (the post-junction area), corresponding to the junction area of conjugants was labelled with anti-fenestrin antibody and maintained the properties required for the nuclear development. The results of the genetic study were consistent with a hypothesis that cytogamy (pronuclear fusion) was induced in the separated conjugants. Therefore, the lasting cell contact is not necessary for the successful completion of conjugational nuclear events.
Key Words. Cell junction, conjugation, cytogamy
ONJUGAL pairing in Tetrahymena thermophila induces a C sequence of nuclear events: meiosis, prezygotic mitosis, pronuclear exchange and fusion, two postzygotic divisions, macronuclear development, and resorption of the old macro- nucleus (Cole and Soelter 1997; Cole et al. 1997; Kaczanowski and Kaczanowska 1996; Kaczanowski and Kiersnowska 1996; Kaczanowski et al. 1991; Martindele, Allis, and Bruns 1982; Nanney 1953; Sugai and Hiwatashi 1974; reviewed in Bruns 1986; Orias 1986). Development of macronuclear anlagen de- pends upon anterior positioning of two products of the post- zygotic divisions while two other products positioned posteri- orly are inhibited in this differentiation and remain as micro- nuclei (Kaczanowski and Kiersnowska 1996; Kaczanowski et al. 1991; Martindale, Allis, and Bruns 1982; Nanney 1953). Under standard conditions, conjugating cells remain paired in T. thermophila for about 10-11 h (Gaertig and Kaczanowski 1987; Kiersnowska, Kaczanowski, and De Haller 1993; Martin- dele, Allis, and Bruns 1982). By then, the separating conjugant cells contain macronuclear anlagen with 4C DNA content (Doerder and De Bault 1975; Kaczanowski et al. 1991) and are already advanced in resorption of the old macronucleus (stage 11 of macronuclear development (Cole and Soelter 1997; Kac- zanowski et al. 1991; Martindele, Allis, and Bruns 1982; Nan- ney 1953). The conjugation cell junction, which is a physical requirement for the reciprocal exchange of pronuclei, may also be a source of the subsequent developmental signals required for postzygotic development. In previous studies, cytogamy in T. thermophila was induced by treatment of the conjugants with the osmotic shock (Cole and Bruns 1992; Orias, Hamilton, and Flacks 1979) and this procedure was used to successfully screen for mutants (Cole and Soelter 1997; Cole et al. 1997; Melia, Cole, and Turkewitz 1998). This shock, applied to conjugants that were at the 3rd prezygotic division, or at the stage of pro- nuclei, prevented the pronuclear transfer, but allowed macro- nuclear development. These experiments leave open the possi- bility that, even in the absence of the pronuclear exchange, postzygotic development requires maintaining of cell contact at least until the 3rd prezygotic division (executed about 4 h after pair formation). In this study, conjugants of T. thermophila were mechanically separated as early as during meiotic pro- phase and then observed for further development to determine whether cytogamy may be induced in this way and whether
Corresponding Author: M. Kiersnowska-Telephone number: 48-22- 620-0381 ext. 410; FAX number: 48-22-826-8624; Ernail: procyt@ warman.com.pl
I Current address: Department of Immunology, Institute of Oncology, Warsaw, Poland.
lasting cell contact is a prerequisite for progress through pre- and postzygotic conjugation nuclear events in Tetrahymena.
MATERIALS AND METHODS Stocks. Two heterokaryon strains of different mating types
and carrying the drug markers for cycloheximide resistance (chx-I) and for 6-methylpurine resistance (mpr-I) in their mi- cronuclei were used (Table l). They were kindly supplied by Dr J. Gaertig (University of Georgia, Athens, GA). Cells with micronuclear defects: strain A*V and the amicronucleate strain BI3840 were used as cytologically marked strains for studies on karyotypes. They were kindly obtained from Dr. J. Frankel (University of Iowa) and from Dr. A. Kaney (Bryn Mawr Col- lege, PA).
Growth and induction of mating. The growth medium used throughout this study was PPY medium [1% proteose peptone (Difco) supplemented with 0.15% yeast extract (Difco)]. Stock cultures were maintained at room temperature, and experiments were performed at 30 "C. For starvation and conjugation, 10 mM Tris-HC1 medium (pH 7.4) was used. To induce mating, cells were starved for 20 h in the starvation medium and cells of different mating types were mixed in approximately equal numbers according to Bruns and Brussard (1974).
Mechanical separation of conjugating pairs. To induce separation of the pairs, 5 ml of cell samples containing early conjugants (1.5-3.5 h. after mixing the cells) were transfered to a 12-ml vol. centrifuge tube and then vigorously pipetted with a 5-ml automatic pipette. In some experiments cells were also centrifuged for 2 min at 300 g. The efficiency of the cell separation was monitored under a dissecting microscope. The samples in which no more than 5% pairs remained were trans- ferred to small Erlenmeyer flasks and put on the shaker (EL- PAN, Poland) at 30 "C, in order to prevent further cell pairing. The shaking frequency was 220 g.
Cytological methods. For routine staining, cell samples were air-dried on coverslips, fixed with Carnoy's fixative, and stained with Giemsa according to Gaertig, Kiersnowska, and Iftode (1988). For recording karyotypes of exconjugant strains, they were crossed with amicronucleate strain BI3840 (Kaney and Speare 1983) or with A*V strain which has extremely small aneuploid micronuclei (Doerder and Shabatura 1980). Four to five-hour-old conjugants were fixed with Shaudinn's fixative and stained with Giemsa according to Martindale, Allis, and Bruns (1982). Cytological stages of conjugation were distin- guished according to Martindale, Allis, and Bruns (1982), in- cluding stages I-VI of the meiotic prophase.
Immunolabelling. Cortical structures. For visualisation of the cortical structures of the conjugants and the cytoskeletal structures involved in nuclear events, 3 antibodies were used:
139

I40 J. EUKARYOT. MICROBIOL., VOL. 47, NO. 2, MARCH-APRIL 2000
Table 1. Strains of Tetrahymena thermophilu used in this study."
Strain Micronuclear genotype
~
Macronuclea?' genome & phenotype Mating type Reference
( C H X I , cy-sensitive, VI) Bruns 1986 CU427 c.hx- Ilchx- I CU428 m p r l - l l m p r l - I ( M P R I , mp-sensitive, VII) Bruns 1986 813840 amicronuclear strain (IV) Kaney and Spedre 1983 A*V micronuclear defective strain (V) Doerder and Shabatura 1980
" Designations of heterokaryon strains were used according to the proposal of The Seventh International Meeting on Ciliate Molecular Biology Nomenclature (1998).
Macronuclear gene alleles, drug sensitivity, and mating types (Roman numerals) are given in brackets.
I ) a commercial (Sigma) monoclonal anti-tyrosine tubulin (clone TUB-1A2); 2) a rabbit anti-dog brain tubulin, kindly supplied by Dr. M. De Brabander and Dr. R. Nuydens from Janssen Research Foundation, Belgium and 3 ) a monoclonal antibody (mAb 3A7) against the ciliate protein p-64, called fe- nestrin (Nelsen et a]. 1994), kindly supplied by Dr. J. Frankel from the University of Iowa. The cells were pelleted by cen- trifugation (300-400 g), permeabilized and fixed; the details of this procedure depended upon primary antibodies as listed be- low.
Labelling with anti-tubulin antibodies. Cells were permea- bilized with 0 . 2 4 3 % Triton X-100 in PHEM buffer (60 mM PIPES, 25 mM HEPES, l0mM EGTA, 2 mM MgCl,), pH 6.9 (Shliva and Van Blerkom 1981) for 2-4 min at room temper- ature and then fixed for 30-45 min with 2% paraformaldehyde in a PHEM buffer (for labelling with a monoclonal anti-tubulin antibody) or with 1 % glutaraldehyde (for labelling with a poly- clonal anti-tubulin antibody). After fixation the cells were washed in three 10-min rinses with buffer A (phosphate-buff- ered saline: 130 mM NaCl, 2 mM KC1, 8 mM Na,HPO,, 2mM KH,PO, with 1% bovine serum albumin, 10 mM EGTA, 2 mM MgCl, and 0.1% Tween). To remove the autofluorescence of the glutaraldehyde, cells were incubated for 15 min in NaBH, (2 mg/ml in buffer A), then washed in three 5-min rinses. Pel- lets were suspended in the primary antibodies and incubated
overnight at 4 "C. Anti-tyrosine tubulin and anti-dog brain tu- bulin antibodies were diluted 1:150 and 1:30, respectively, in buffer A without MgCI, and EGTA.
Labelling with anti-fenestrin antibody. Pelleted cells, chilled on ice, were treated for 15 min with cold 0.3% Triton X-100 ,
in 50% ETOH, washed in three 10-min rinses at room temper- ' ature in buffer B (phosphate-buffered saline: 130 mM NaCI, 2 mM KCI, 8 mM Na,HPO,, 2mM KH,PO, with 1 % bovine se- rum albumin, and 0.1% Tween) and then incubated overnight (4 "C) with anti-fenestrin diluted 1:40 in buffer B.
After incubation with anti-tubulin or anti-fenestrin antibod- ies, cells were washed 4X in buffer A or buffer B, respectively, and resuspended for 1 h in FITC-labelled secondary antibodies (1: 100 anti-rabbit Ig-G in buffer A or anti-mouse Ig-G in buffer B). The cells were washed in three 10-min rinses of buffers as above and, in two 5-min rinses of PBS (phosphate-buffered saline: 130 mM NaCl, 2 mM KC1, 8 mM Na,HPO,, 2mM KH,PO,) and then stained with 3 pg/ml DAPl or 1 pg/ml pro- pidium iodide (PI) and washed with PBS. The antiquenching agent DABCO (1,4-diazabicyclo [2,2,2] octane, SIGMA; Bat- taglia et al. 1994) was added before microscopic observations. The cells were analyzed and photographed with a Jenamed fluo- rescent microscope or with a Zeiss LSM 150 confocal micro- scope.
The procedure of preparing cells for immunolabeling does not disrupt joined pairs: only one single cell derived from a disrupted pair was found among 524 conjugant pairs in two control cell samples fixed and immunostained 6 h after conju- gation (i.e. 1/1048 conjugant cells). The frequency of cells in pairs in these control cell samples after cell preparation was as high as before pelleting the cells (about 90%).
Genetic methods. Synclonal or polyclonal cultures were grown in drops of PPY medium arranged on plastic Petri dishes. Cells were tested for drug resistance by replica plating to the medium containing the drug: cycloheximide (25 pM) or meth- ylpurine (15 pM) as recommended by Orias and Bruns (1976).
RESULTS Separation of control (undisturbed) conjugants. During
the normal course of conjugation most cells pair between 1-4 h after mixing and separate between 11-14 h. The cells separate at stage I1 of macronuclear development with two enlarged and faintly stained macronuclear anlagen and two micronuclei.
Fig. 1. Early stages of pair formation of Tetrahymrna rhermophila. A. The cells touched at the transformed tips (hatched area) and slide tip to tip (arrows). These connnections were unstable; some of the cells changed their presumptive mates. B. The angle the paired cells was relatively wide (>90") and the paired cells twisted against each other (dashed arrows). The transparent regions of the tranformed tips were
less and the area of the cell junction was increased. The transformed cell tips were just visible. D. The angle between the cells was decreased to about 45". The transformed tips became invisible. Most of these pairs were lying at the bottom of the dish making very limited movements.
These cells contain a remnant of the old macronucleus, which is shifted posteriorly and condensed, or the old macronucleus is competely resorbed,
The cells were crossed and the cell samples were stained with Giemsa at 12, and 13 h after mixing. In thelO-h sample,
cells were found among the 780 conjugants (less than 1%); 98% of the Paired cells were at stage I1 Of macronuclear ment. After 11, 12, and 13 h of conjugation respectively, 7% (1 17/1631), 17%(100/576), and 28% (143/ 516) of the conju-
clearly seen, C, The angle between the cells was decreased to 90" or the frequency Of pairing was 84% but Only sing1e exconjugant
KIERSNOWSKA ET AL.-SEPARATED CONJUGANTS 141
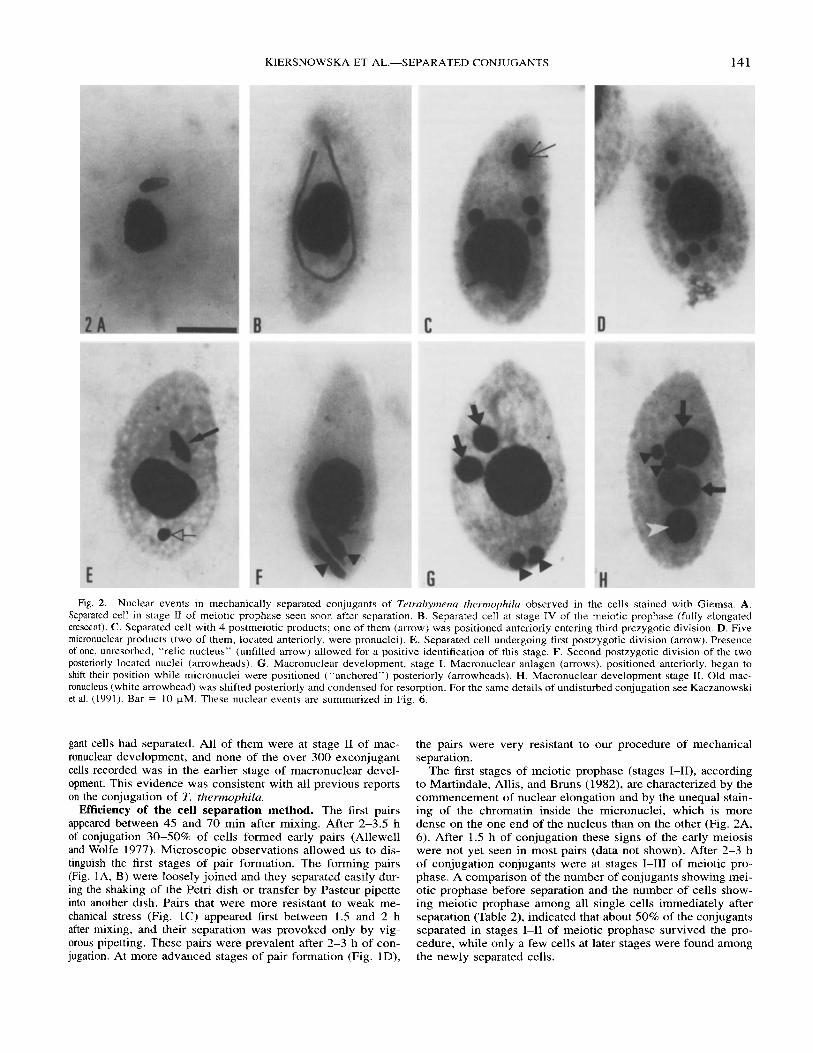
Fig. 2. Nuclear events in mechanically separated conjugants of Terrahymena thermophila observed in the cells stained with Giemsa. A. Separated cell in stage I1 of meiotic prophase seen soon after separation. B. Separated cell at stage IV of the meiotic prophase (fully elongated crescent). C. Separated cell with 4 postmeiotic products; one of them (arrow) was positioned anteriorly entering third prezygotic division. D. Five micronuclear products (two of them, located anteriorly, were pronuclei). E. Separated cell undergoing first postzygotic division (arrow). Presence of one, unresorbed, “relic nucleus” (unfilled arrow) allowed for a positive identification of this stage. F. Second postzygotic division of the two posteriorly located nuclei (arrowheads). G. Macronuclear development, stage I. Macronuclear anlagen (arrows), positioned anteriorly, began to shift their position while micronuclei were positioned (“anchored”) posteriorly (arrowheads). H. Macronuclear development stage 11. Old mac- ronucleus (white arrowhead) was shifted posteriorly and condensed for resorption. For the same details of undisturbed conjugation see Kaczanowski et al. (1991). Bar = 10 pM. These nuclear events are summarized in Fig. 6.
gant cells had separated. All of them were at stage I1 of mac- ronuclear development, and none of the over 300 exconjugant cells recorded was in the earlier stage of macronuclear devel- opment. This evidence was consistent with all previous reports on the conjugation of T. thermophila.
Efficiency of the cell separation method. The first pairs appeared between 45 and 70 min after mixing. After 2-3.5 h of conjugation 30-50% of cells formed early pairs (Allewell and Wolfe 1977). Microscopic observations allowed us to dis- tinguish the first stages of pair formation. The forming pairs (Fig. IA, B) were loosely joined and they separated easily dur- ing the shaking of the Petri dish or transfer by Pasteur pipette into another dish. Pairs that were more resistant to weak me- chanical stress (Fig. 1C) appeared first between 1.5 and 2 h after mixing, and their separation was provoked only by vig- orous pipetting. These pairs were prevalent after 2-3 h of con- jugation. At more advanced stages of pair formation (Fig. lD),
the pairs were very resistant to our procedure of mechanical separation.
The first stages of meiotic prophase (stages 1-11), according to Martindale, Allis, and Bruns (1982), are characterized by the commencement of nuclear elongation and by the unequal stain- ing of the chromatin inside the micronuclei, which is more dense on the one end of the nucleus than on the other (Fig. 2A, 6). After 1.5 h of conjugation these signs of the early meiosis were not yet seen in most pairs (data not shown). After 2-3 h of conjugation conjugants were at stages 1-111 of meiotic pro- phase. A comparison of the number of conjugants showing mei- otic prophase before separation and the number of cells show- ing meiotic prophase among all single cells immediately after separation (Table 2) , indicated that about 50% of the conjugants separated in stages 1-11 of meiotic prophase survived the pro- cedure, while only a few cells at later stages were found among the newly separated cells.
142 J . EUKARYOT. MICROBIOL., VOL. 47, NO. 2, MARCH-APRIL 2000
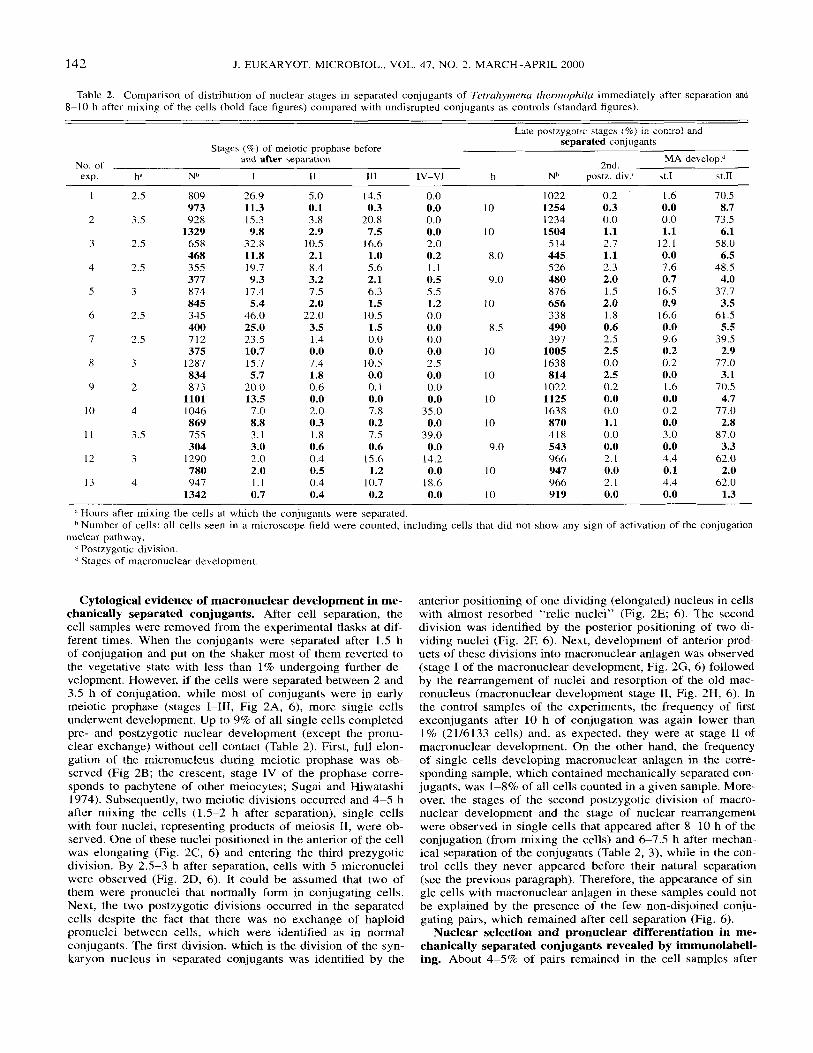
Table 2. Comparison of distribution of nuclear stages in separated conjugants of Tetrahymma thermophilu immediately after separation and 8-10 h after mixing of the cells (bold face figures) compared with undisrupted conjugants as controls (standard figures).
Late postzygotic stages (%) i n control and separated conjugants Stages (9%) of meiotic prophase before
and after separation MA develop? No. of 2nd. exp. h ‘ Nh I I1 111 IV-VI h Nb postz. div.‘ st.1 st.11
1 2.5
2 3.5
3 2.5
4 2.5
5 3
6 2.5
7 2.5
8 3
9 2
10 4
I I 3.5
12 3
13 4
809 973 928
1329 658 468 355 377 874 845 345 400 712 375
1287 834 873
1101 1046 869 755 304
1290 780 947
1342
26.9 11.3 15.3 9.8
32.8 11.8 19.7 9.3
17.4 5.4
46.0 25.0 2.3.5 10.7 15.7 5.7
20.0 13.5 7.0 8.8 3. I 3.0 2.0 2.0 1.1 0.7
5.0 0.1 3.8 2.9
10.5 2.1 8.4 3.2 7.5 2.0
22.0 3.5 I .4 0.0 7.4 1.8 0.6 0.0 2.0 0.3 I .8 0.6 0.4 0.5 0.4 0.4
14.5 0.3
20.8 7.5
16.6 1.0 5.6 2.1 6.3 1.5
10.5 1.5 0.0 0.0
10.5 0.0 0.1 0.0 7.8 0.2 7.5 0.6
15.6 1.2
10.7 0.2
0.0 0.0 0.0 0.0 2.0 0.2 1.1 0.5 5.5 1.2 0.0 0.0 0.0 0.0 2.5 0.0 0.0 0.0
35.0 0.0
39.0 0.0
14.2 0.0
18.6 0.0
10
10
8.0
9.0
1 0
8.5
10
10
10
10
9.0
10
10
1022 0.2 1254 0.3 1234 0.0 1504 1.1
5 14 2.7 445 1.1 526 2.3 480 2.0 876 I .5 656 2.0 338 1.8 490 0.6 397 2.5
1005 2.5 1638 0.0 814 2.5
1022 0.2 1125 0.0 1638 0.0 870 1.1 418 0.0 543 0.0 966 2.1 947 0.0 966 2.1 919 0.0
I .6 70.5 0.0 8.7 0.0 73.5 1.1 6.1
12.1 58.0 0.0 6.5 7.6 48.5 0.7 4.0
16.5 37.7 0.9 3.5
16.6 61.5 0.0 5.5 9.6 39.5 0.2 2.9 0.2 77.0 0.0 3.1 1.6 70.5 0.0 4.7 0.2 77.0 0.0 2.8 3.0 87.0 0.0 3.3 4.4 62.0 0.1 2.0 4.4 62.0 0.0 1.3
,’ Hours after mixing the cells at which the conjugants were separated. Number of cells: all cells seen in a microscope field were counted, including cells that did not show any sign of activation of the conjugation
Postzygotic division. Stages of macronuclear development.
nuclear pathway.
Cytological evidence of macronuclear development in me- chanically separated conjugants. After cell separation, the cell samples were removed from the experimental flasks at dif- ferent times. When the conjugants were separated after 1.5 h of conjugation and put on the shaker most of them reverted to the vegetative state with less than 1% undergoing further de- velopment. However, if the cells were separated between 2 and 3.5 h of conjugation, while most of conjugants were in early meiotic prophase (stages 1-111, Fig 2A, 6), more single cells underwent development. Up to 9% of all single cells completed pre- and postzygotic nuclear development (except the pronu- clear exchange) without cell contact (Table 2). First, full elon- gation of the micronucleus during meiotic prophase was ob- served (Fig 2B; the crescent, stage IV of the prophase corre- sponds to pachytene of other meiocytes; Sugai and Hiwatashi 1974). Subsequently, two meiotic divisions occurred and 4-5 h after mixing the cells (1.5-2 h after separation), single cells with four nuclei, representing products of meiosis 11, were ob- served. One of these nuclei positioned in the anterior of the cell was elongating (Fig. 2C, 6) and entering the third prezygotic division. By 2.5-3 h after separation, cells with 5 micronuclei were observed (Fig. 2D, 6). It could be assumed that two of them were pronuclei that normally form in conjugating cells. Next, the two postzygotic divisions occurred in the separated cells despite the fact that there was no exchange of haploid pronuclei between cells, which were identified as in normal conjugants. The first division, which is the division of the syn- karyon nucleus in separated conjugants was identified by the
anterior positioning of one dividing (elongated) nucleus in cells with almost resorbed “relic nuclei” (Fig. 2E; 6). The second division was identified by the posterior positioning of two di- viding nuclei (Fig. 2E 6). Next, development of anterior prod- ucts of these divisions into macronuclear anlagen was observed (stage I of the macronuclear development, Fig. 2G, 6) followed by the rearrangement of nuclei and resorption of the old mac- ronucleus (macronuclear development stage 11, Fig. 2H, 6). In the control samples of the experiments, the frequency of first exconjugants after 10 h of conjugation was again lower than 1% (2116133 cells) and, as expected, they were at stage I1 of macronuclear development. On the other hand, the frequency of single cells developing macronuclear anlagen in the corre- sponding sample, which contained mechanically separated con- jugants, was 1-8% of all cells counted in a given sample. More- over, the stages of the second postzygotic division of macro- nuclear development and the stage of nuclear rearrangement were observed in single cells that appeared after 8-10 h of the conjugation (from mixing the cells) and 6-7.5 h after mechan- ical separation of the conjugants (Table 2, 3), while in the con- trol cells they never appeared before their natural separation (see the previous paragraph). Therefore, the appearance of sin- gle cells with macronuclear anlagen in these samples could not be explained by the presence of the few non-disjoined conju- gating pairs, which remained after cell separation (Fig. 6).
Nuclear selection and pronuclear differentiation in me- chanically separated conjugants revealed by immunolabell- ing. About 4-5% of pairs remained in the cell samples after

KIERSNOWSKA ET AL.-SEPARATED CONJUGANTS 143
Table 3. Distribution of the postzygotic stages in control (standard figures) and separated (bold face figures) conjugants of Tetrahymenu hermophila 6.5-10 h after mixing the cells.
Poatzygotic stages (%) in control and separated conjugants
No. of After 1st 2nd Ma development exp.& h N division division st.1 St .11
5 7 674 5.6 7.5 16.0 7.4 407 4.9 0.5 1.0 0.0
10 876 4.2 I .5 16.5 37.7 656 1.8 2.0 0.9 3.5
6 6.5 254 18.9 6.0 17.5 0.0 590 2.0 0.5 4.2 2.2
8.5 338 3.0 1.8 16.6 61 .5 490 1.2 0.6 0.0 5.5
a Data from Experiments 5 and 6 in Table 2.
cell separation and the probability of disruption of these pairs during fixation and preparation for immunolabelling was very low (1/1000, see previous section). Thus, the probability of finding single conjugant cells disrupted during preparation was very low.
In the separated conjugants, as in normal conjugants (Gaertig and Fleury 1992; Takagi, Numata, and Watanabe 1991), a poly- clonal anti-dog brain tubulin antibody labelled the perinuclear microtubules around the nucleus recruited to the postmeiotic division and the products of this division (pronuclei). Micro- tubular strands connected the anteriorly localized products of the second (data not shown) and third prezygotic divisions with
the apical cell region above the oral apparatus (Fig. 3A, B). This apical region, called here the post-junction area, corre- sponds to the junction area of the pairing cells (Fig. 3, 4A, 5A, C).
When the posterior product of the postmeiotic mitosis reached the anterior of the cell, the anterior product was oval- shaped (Fig. 3B, 5D), surrounded by a thick sheet of microtu- bules, and was closer to the post-junction area. During normal, undisturbed conjugation the same change in the shape (from spherical to oval) of the anterior pronuclei occurs just before the passage of these pronuclei to the partner cell (Nelsen et al. 1994).
The two overlapping nuclei, enclosed by the microtubules extending to the post-junction area, were observed in separated conjugants collected 6 h after mixing (Fig. 3C) and it seemed that these microtubules enclosed the fusing pronuclei rather than the dividing synkaryon. The restitution of the intracyto- plasmic microtubular network characteristic for vegetative cells appeared at stage I1 of macronuclear development (data not shown) as in normal conjugants (Gaertig and Fleury 1992). Ac- cording to Nelsen et al.(1994), the conjugation cell junction and the material surrounding pronuclei are specifically labelled with the mAb 3A7 antibody directed against a 64-kd protein, fenes- trin. In this study, the mAb 3A7 antibody labelled the pronuclei and post-junction area in mechanically separated cells (Fig. 4A). The pronuclei were connected to each other and to the post-junction area anteriorly to the oral apparatus with a fine anti-fenestrin-stained material.
Evidence of oral reorganization and resumption of feed- ing in the separated conjugants. The regression of the old
Fig. 3. Optical section of the separated conjugants of Tetrahymenu thermophika stained with a polyclonal antiserum raised against dog-brain tubulin (visualized with FITC-conjugated secondary antiserum) and costained with propidium iodide. The conjugants were separated 2.5 h after mixing of cells. The cells were collected 2.5 h later and immunostained. A and B. Two products of the third prezygotic division of the selected, postmeiotic nucleus were surrounded by microtubules (arrowheads). A represents 4 (from 20, 2;: 0.45 kM) superimposed, optical sections. B represents 2 (from 22, z: 0.35 pM) superimposed optical sections. C . The two overlapping nuclei (arrows) were enclosed by the microtubules (arrowheads). Note the connection of the microtubules with the post- junction area (thick arrow). C represents one (from 24) optical section (z: 0.3 (LM). Bar = 5pM.
144 J. EUKARYOT. MICROBIOL., VOL. 47, NO. 2, MARCH-APRIL 2000
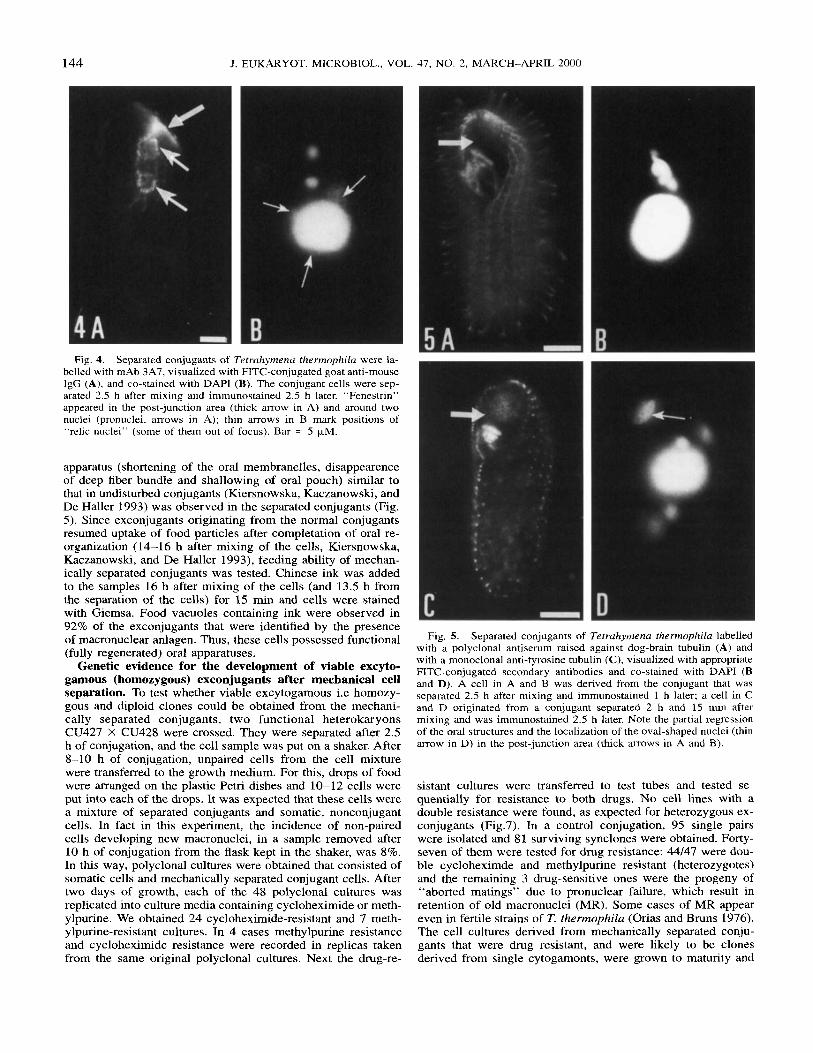
Fig. 4. Separated conjugants of Tetruhymenu thermophilu were la- belled with mAb 3A7, visualized with FITC-conjugated goat anti-mouse IgG (A), and co-stained with DAPI (B). The conjugant cells were sep- arated 2.5 h after mixing and immunostained 2.5 h later. “Fenestrin” appeared in the post-junction area (thick arrow in A) and around two nuclei (pronuclei, arrows in A); thin arrows in B mark positions of “relic nuclei” (some of them out of focus). Bar = 5 kM.
apparatus (shortening of the oral membranelles, disappearence of deep fiber bundle and shallowing of oral pouch) similar to that in undisturbed conjugants (Kiersnowska, Kaczanowski, and De Haller 1993) was observed in the separated conjugants (Fig. 5). Since exconjugants originating from the normal conjugants resumed uptake of food particles after completation of oral re- organization (14-16 h after mixing of the cells, Kiersnowska, Kaczanowski, and De Haller 1993), feeding ability of mechan- ically separated conjugants was tested. Chinese ink was added to the samples 16 h after mixing of the cells (and 13.5 h from the separation of the cells) for 15 min and cells were stained with Giemsa. Food vacuoles containing ink were observed in 92% of the exconjugants that were identified by the presence of macronuclear anlagen. Thus, these cells possessed functional (fully regenerated) oral apparatuses.
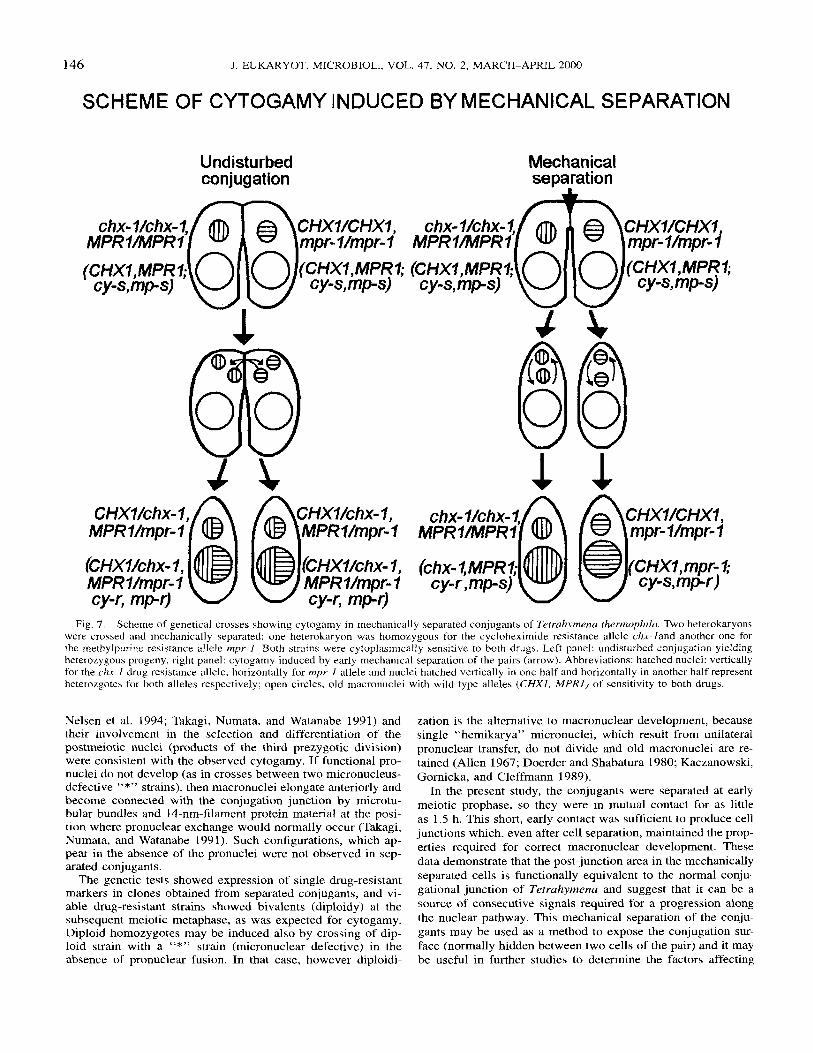
Genetic evidence for the development of viable excyto- gamous (homozygous) exconjugants after mechanical cell separation. To test whether viable excytogamous i.e homozy- gous and diploid clones could be obtained from the mechani- cally separated conjugants, two functional heterokaryons CU427 X CU428 were crossed. They were separated after 2.5 h of conjugation, and the cell sample was put on a shaker. After 8-10 h of conjugation, unpaired cells from the cell mixture were transferred to the growth medium. For this, drops of food were arranged on the plastic Petri dishes and 10-12 cells were put into each of the drops. It was expected that these cells were a mixture of separated conjugants and somatic, nonconjugant cells. In fact in this experiment, the incidence of non-paired cells developing new macronuclei, in a sample removed after 10 h of conjugation from the flask kept in the shaker, was 8%. In this way, polyclonal cultures were obtained that consisted of somatic cells and mechanically separated conjugant cells. After two days of growth, each of the 48 polyclonal cultures was replicated into culture media containing cycloheximide or meth- ylpurine. We obtained 24 cycloheximide-resistant and 7 meth- ylpurine-resistant cultures. In 4 cases methylpurine resistance and cycloheximide resistance were recorded in replicas taken from the same original polyclonal cultures. Next the drug-re-
Fig. 5. Separated conjugants of Tetruhymenu rhermophila labelled with a polyclonal antiserum raised against dog-brain tubulin (A) and with a monoclonal anti-tyrosine tubulin (C), visualized with appropriate FITC-conjugated secondary antibodies and co-stained with DAPI (B and D). A cell in A and B was derived from the conjugant that was separated 2.5 h after mixing and immunostained 1 h later; a cell in C and D originated from a conjugant separated 2 h and 15 min after mixing and was immunostained 2.5 h later. Note the partial regression of the oral structures and the localization of the oval-shaped nuclei (thin arrow in D) in the post-junction area (thick arrows in A and 8 ) .
sistant cultures were transferred to test tubes and tested se- quentially for resistance to both drugs. No cell lines with a double resistance were found, as expected for heterozygous ex- conjugants (Fig.7). In a control conjugation, 95 single pairs were isolated and 81 surviving synclones were obtained. Forty- seven of them were tested for drug resistance: 44/47 were dou- ble cycloheximde and methylpurine resistant (heterozygotes) and the remaining 3 drug-sensitive ones were the progeny of “aborted matings” due to pronuclear failure, which result in retention of old macronuclei (MR). Some cases of MR appear even in fertile strains of T. thermophila (Orias and Bruns 1976). The cell cultures derived from mechanically separated conju- gants that were drug resistant, and were likely to be clones derived from single cytogamonts, were grown to maturity and
KIERSNOWSKA ET AL -SEPARATED CONJUGANTS
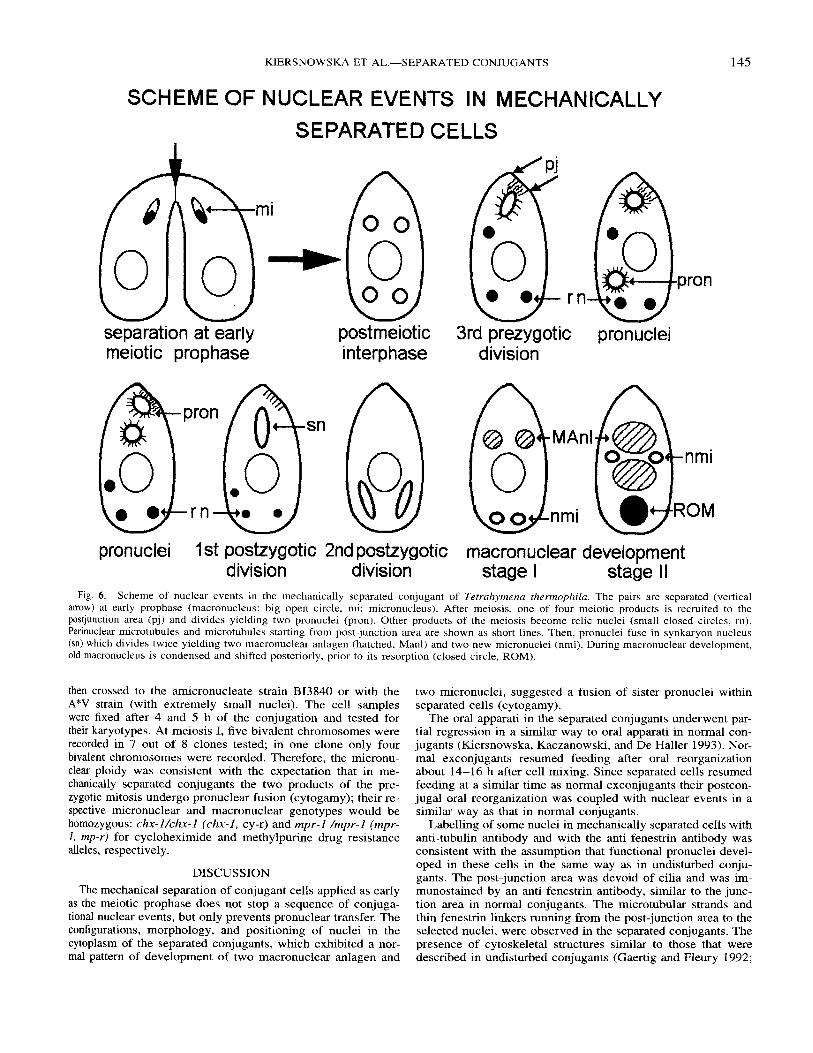
SCHEME OF NUCLEAR EVENTS IN MECHANICALLY SEPARATED CELLS
pron
separation at early meiotic prophase
pron
- rn -
.sn
post meiot ic i n te rphase
3rd prezygotic pronuclei division
145
pronuclei 1 st postzygotic 2nd postzygotic macronuclear development division division stage I stage II
Fig. 6. Scheme of nuclear events in the mechanically separated conjugant of Tetrahymenu thermophila. The pairs are separated (vertical arrow) at early prophase (macronucleus: big open circle, mi: micronucleus). After meiosis, one of four meiotic products is recruited to the postjunction area (pj) and divides yielding two pronuclei (pron). Other products of the meiosis become relic nuclei (small closed circles, rn). Perinuclear microtubules and microtubules starting from post-junction area are shown as short lines. Then, pronuclei fuse in synkaryon nucleus (sn) which divides twice yielding two macronuclear anlagen (hatched, Manl) and two new micronuclei (nmi). During macronuclear development, old macronucleus is condensed and shifted posteriorly, prior to its resorption (closed circle, ROM).
then crossed to the amicronucleate strain B13840 or with the A*V strain (with extremely small nuclei). The cell samples were fixed after 4 and 5 h of the conjugation and tested for their karyotypes. At meiosis I, five bivalent chromosomes were recorded in 7 out of 8 clones tested; in one clone only four bivalent chromosomes were recorded. Therefore, the micronu- clear ploidy was consistent with the expectation that in me- chanically separated conjugants the two products of the pre- zygotic mitosis undergo pronuclear fusion (cytogamy); their re- spective micronuclear and macronuclear genotypes would be homozygous: chx-Uchx-1 (chx-I, cy-r) and mpr-1 /mpr-I (mpr- 1, mp-r) for cycloheximide and methylpurine drug resistance alleles, respectively.
DISCUSSION The mechanical separation of conjugant cells applied as early
as the meiotic prophase does not stop a sequence of conjuga- tional nuclear events, but only prevents pronuclear transfer. The configurations, morphology, and positioning of nuclei in the cytoplasm of the separated conjugants, which exhibited a nor- mal pattern of development of two macronuclear anlagen and
two micronuclei, suggested a fusion of sister pronuclei within separated cells (cytogamy).
The oral apparati in the separated conjugants underwent par- tial regression in a similar way to oral apparati in normal con- jugants (Kiersnowska, Kaczanowski, and De Haller 1993). Nor- mal exconjugants resumed feeding after oral reorganization about 14-16 h after cell mixing. Since separated cells resumed feeding at a similar time as normal exconjugants their postcon- jugal oral reorganization was coupled with nuclear events in a similar way as that in normal conjugants.
Labelling of some nuclei in mechanically separated cells with anti-tubulin antibody and with the anti-fenestrin antibody was consistent with the assumption that functional pronuclei devel- oped in these cells in the same way as in undisturbed conju- gants. The post-junction area was devoid of cilia and was im- munostained by an anti-fenestrin antibody, similar to the junc- tion area in normal conjugants. The microtubular strands and thin fenestrin linkers running from the post-junction area to the selected nuclei, were observed in the separated conjugants. The presence of cytoskeletal structures similar to those that were described in undisturbed conjugants (Gaertig and Fleury 1992;
146 J. EUKARYOT. MICROBIOL., VOL. 47, NO. 2, MARCH-APRIL 2000
SCHEME OF CYTOGAMY INDUCED BY MECHANICAL SEPARATION
CHXVchx- l , , q f)WXWhx- I , MPR7/mpr-7 @ @ MPRl/mpr-l
Mechanical separation
1 1 A h
Fig. 7. Scheme of genetical crosses showing cytogamy in mechanically separated conjugants of Tetrcrhymena thermophila. Two heterokdryons were crossed and mechanically separated: one heterokaryon was homozygous for the cycloheximide resistance allele chx- land another one for the methylpurine resistance allele mpr-I. Both strains were cytoplasrnically sensitive to both drugs. Left panel: undisturbed conjugation yielding heteroLygous progeny, right panel: cytogamy induced by early mechanical separation of the pairs (arrow). Abbreviations: hatched nuclei: vertically for the chx-1 drug resistance allele, horizontally for mpr-l allele and nuclei hatched vertically in one half and horizontally in another half represent heterozgotes for both alleles respectively; open circles, old macronuclei with wild type alleles ( C H X I , M P R l ) of sensitivity to both drugs.
Nelsen et al. 1994; Takagi, Numata, and Watanabe 1991) and their involvement in the selection and differentiation of the postmeiotic nuclei (products of the third prezygotic division) were consistent with the observed cytogamy. If functional pro- nuclei do not develop (as in crosses between two micronucleus- defective “*” strains), then macronuclei elongate anteriorly and become connected with the conjugation junction by microtu- bular bundles and 14-nm-filament protein material at the posi- tion where pronuclear exchange would normally occur (Takagi, Numata, and Watanabe 1991). Such configurations, which ap- pear in the absence of the pronuclei were not observed in sep- arated conjugants.
The genetic tests showed expression of single drug-resistant markers in clones obtained from separated conjugants, and vi- able drug-resistant strains showed bivalents (diploidy) at the subsequent meiotic metaphase, as was expected for cytogamy. Diploid homozygotes may be induced also by crossing of dip- loid strain with a “*” strain (micronuclear defective) in the absence of pronuclear fusion. In that case, however diploidi-
zation is the alternative to macronuclear development, because single “hemikarya” micronuclei, which result from unilateral pronuclear transfer, do not divide and old macronuclei are re- tained (Allen 1967; Doerder and Shabatura 1980; Kaczanowski, Gornicka, and Cleffmann 1989).
In the present study, the conjugants were separated at early meiotic prophase, so they were in mutual contact for as little as 1.5 h. This short, early contact was sufficient to produce cell junctions which, even after cell separation, maintained the prop- erties required for correct macronuclear development. These data demonstrate that the post-junction area in the mechanically separated cells is functionally equivalent to the normal conju- gational junction of Tetrahymena and suggest that it can be a source of consecutive signals required for a progression along the nuclear pathway. This mechanical separation of the conju- gants may be used as a method to expose the conjugation sur- face (normally hidden between two cells of the pair) and it may be useful in further studies to determine the factors affecting

KIERSNOWSKA ET AL.-SEPARATED CONJUGANTS 147
this junction or its role in signaling processes of nuclear de- velopment.
ACKNOWLEDGMENTS This work was supported by a grant from KBN (The Com-
mittee of Scientific Research, Poland), No. 6P04C 071 10 to Dr. Janina Kaczanowska. Thanks are due to Drs. J. Kacza- nowska and to D. Wloga for discussions and valuable com- ments.
LITERATURE CITED Allen, S. L. 1967. Genomic exclusion. A rapid mean for inducing ho-
mozygous diploid lines in Tetruhymenu pyrformis , syngen 1. Sci- ence, 155575-577.
Allewell, N. & Wolfe, J. 1977. A kinetic analysis of a memory of a developmental interaction. Mating interaction in Tetruhpmenu ppri- formis. E.wp. Cell Res., 109: 15-24.
Battaglia, M., Pozzi, D., Grimaldi, S. & Parasassi, T. 1994. Hoechst 33258 staining for detecting mycoplasma contamination in cell cul- tures: a method for reducing fluorescence photobleaching. Biotech. Histochrm., 69: 152- 156.
Bruns. I? J. 1986. Genetic organization in Tetruhymmu. in: Cali, J. (ed., The Molecular Biology of Ciliated Protozoa. Academic Press, New York. p. 27-44.
Bruns, I? J . & Brussard, T. B. 1974. Pair formation in Tetrahymenu pyrifhrmis, an inducible developmental system. J . Exp. Zoof . , 188: 337-344.
Cole, E. & Bruna, P J. 1992. Uniparental cytogamy: a novel efficient method for bringing mutations of Tetruhymenu into homozygous ex- pression with precocious sexual maturity. Genetics, 132: 1017-103 1 .
Cole, E. & Soelter, T. A. 1997. A mutational analysis of conjugation in Tetruhymenu thermophilu. 2. Phenotypes affecting middle and late development: third prezygotic nuclear division, pronuclear exchange, pronuclear fusion and postzygotic development. Dev. B i d . , 189:233- 245.
Cole, E., Cassidy-Hanky, D., Hemish, J., Tuan, J . & Bruns, P. J . 1997. A mutational analysis of conjugation in Tetruhymenu thermophilu. 1. Phenotypes affecting early development: meiosis to nuclear selection. Dev. Riot.. 189:215-232.
Doerder E I? & De Bault, L. E. 1975. Cytofluorometric analysis of nuclear DNA during meiosis fertilization and macronuclear devel- opment in the ciliate 7etruhvmenu pyr$ormis, syngen I . J . Cell Sci., 17:471-493.
Doerder, F. I? & Shabatura, S. K. 1980. Genomic exclusion in Tefru- hymrnu thermophilu. A cytogenetic and cytofluorometric study. Drv. Genet., 1:205-218.
Gaertig, J. & Fleury, A. 1992. Spatio-temporal reorganization of intra- cytoplasmic microtubules is associated with nuclear selection and dif- ferentiation during the developmental process in the ciliate Tetruhy- metzci thrrmophilu. Protoplu.smu, 167:74-87.
Gaertig, J . & Kaczanowski, A. 1987. Correlation between shortened period of cell pairing during geneomic exclusion and the block in postransfer nuclear development in Tetruhymenu thermophilu. Dev. Growth und Diner., 29:553-562.
Gaertig, J., Kiersnowska, M. & Iftode, E 1988. Induction of cybrid strains of Tetruhymenu by electrofusion. J . Cell Sci., 89:253-261.
Kaczanowski, A. & Kaczanowska, J. 1996. Induction of blocks in nu- clear division and overcondensation of meiotic chromosomes with cycloheximide during conjugation of Tetruhymenu thermophilu. J . Eukutyot. (Euk.) Microbiul., 43:380-388.
Kaczanowski, A. & Kiersnowska, M. 1996. The differentiation of post- zygotic posterior nuclei into micronuclei in Tetruhymenu thermophilu is not affected by electrofusion with anterior pole of another conju- gant. Europ. J . Protistol., 32: 1-1 2.
Kaczanowski, A,, Gornicka, I. & Cleffmann, G. 1989. Arrest in micro- nuclear DNA replication during genomic exclusion in Tetruhymenu produces haploid strains. Genetics, 121:37-45.
Kaczanowski, A,, Ramel, M., Kaczanowska, J . & Wheatley, D. 1991. Macronuclear differentiation in conjugating pairs of Tetruhymenu treated with antitubuline drug nocodazole. Exp. Cell R e x , 195:330- 337.
Kaney, A. R. & Speare, V. J. 1983. An amicronuclear mutant of Tet- ruhymena thermophila. Exp. Cell Res., 143:461-467.
Kiersnowska, M., Kaczanowski, A. & De Haller, G. 1993. Inhibition of oral morphogenesis during conjugation of Tetruhymenu thermophilu and its resumption after cell separation. Europ. J . Protistol., 29:359- 369.
Martindale, D. W., Allis, C. D. & Bruns, P. J . 1982. Conjugation in Tetruhpmenu thermophilu: a temporal analysis of cytological stages. Exp. Cell Kes., 140:227-236.
Melia, S. M., Cole, E. S. & Turkewitz, A. I? 1998. Mutational analysis of regulated exocytosis in Tetruhymenu. J . Cell Sci., 111: 13 1-140.
Nanney, D. L. 1953. Nucelocytoplsmic interaction during conjugation in Tetruhymmu. Biol. Bull., 105: 133-148.
Nelsen, M. E., Wiliams, N. E., Yi, H., Knaak, J. & Frankel, J. 1994. “Fenestrin” and conjugation in Tetruhymenu thermophilu. J . Eukur- yot. (Euk.) Microhiol., 41:483-495.
Orias, E. 1986. Ciliate conjugation. In: Gall, J. (ed.), The Molecular Biology of Ciliated Protozoa, Academic Press, New York. p. 45-84.
Orias, E. & Bruns, I? J. 1976. Induction and isolation of mutants in Tetruh.ymenu. In: Prescott, D. M. (ed.), Methods In Cell Biology. Academic Press, New York. 13:247-283.
Orias, E., Hamilton, E. P. & Flacks, M. 1979. Osmotic shock prevents nuclear exchange and produces whole-genome hoinozygotes in con- jugating Tetruhymenu. Science, 203:660-663.
Shliva, M. & Van Blerkom, J . 198 1 . Structural interaction of cytoskel- eta1 components. J.Cell Biol., 90:222-235.
Sugai, T. & Hiwatashi, K. 1974. Cytologic and autoradiography of the micronucleus at the meiotic prophase in Tetruhvmenu pyr(fi)rmi.s. J . Protozool., 2 1542-548.
Takagi, I., Numata, 0. & Watanabe, Y. 1991. Involvement of 14-nm filament-forming protein and tubulin in gametic pronuclear behavior during conjugation in Tetruhymenu. J . Protozool., 38:34 1-35 1.
The Seventh International Meeting on Ciliate Molecular Biology No- menclature. 1998. Proposed genetic nomenclature rules for Tetruhy- menu thermophilu, Paramecium primuureliu and Purumecium tetruu- reliu. Genetics. 149:459-462.
Received 2-19-90, 4-2-99, 10-6-09; accepted 10-6-09
Related Documents











