MORPHOLOGY,SYSTEMATICS,EVOLUTION The Taxonomic Status of Genetically Divergent Populations of Lutzomyia longipalpis (Diptera: Psychodidae) Based on the Distribution of Mitochondrial and Isozyme Variation JAZZMIN ARRIVILLAGA, 1 JOHN- PAUL MUTEBI, 2 HERMES PIN ˜ ANGO, 3 DOUGLAS NORRIS, 2 BRUCE ALEXANDER, 4 M. DORA FELICIANGELI, 1 AND GREGORY C. LANZARO 5 J. Med. Entomol. 40(5): 615Ð627 (2003) ABSTRACT The sand ßy, Lutzomyia longipalpis (Lutz & Neiva) reputedly is a complex of cryptic species; however, there is currently no consensus as to the number of species in the complex or their geographic distributions. We conducted phylogenetic analyses of 31 populations from throughout the species range, using seven isozyme loci and genes in the mitochondrial genome. Analyses of these two independent sets of markers were largely concordant and revealed four distinct clades that support the existence of four species. The four clades have distinct geographic ranges: (1) Brazil (Species A Lu. longipalpis sensu stricto), (2) Laran (Species B Lu. pseudolongipalpis), (3) cis-Andean (Species C), and (4) trans-Andean (Species D). The cis-Andean clade may be subdivided further into two groups, one in Colombia and one in northwestern Venezuela, but their taxonomic status remains unresolved. Knowledge that Lu. longipalpis is a complex of species may ultimately shed light on anomalies in the epidemiology of visceral leishmaniasis in the New World. KEY WORDS Lutzomyia longipalpis, species complex, isozymes, mitochondrial DNA, phylogeny THE SAND FLY Lutzomyia longipalpis (Lutz and Neiva 1912) sensu lato is the primary vector of Leishmania chagasi the etiological agent of zoonotic visceral leish- maniasis (ZVL) in South and Central America. It has a wide geographical range that extends from the Yu- catan Peninsula in Mexico to northern Argentina and Paraguay (Young and Duncan 1994), including all the countries of Central America except Belize and most of those of tropical South America east of the Andes. There are no published records of Lu. longipalpis from Guyana, Surinam, or French Guiana. In addition, there are no records of this species from the PaciÞc Coast of Colombia, Ecuador, Peru, or Chile. Although Lu. lon- gipalpis has one of the widest geographical ranges of any species in the genus, its closest relatives (members of the subgenus Lutzomyia, series longipalpis) are al- most entirely restricted to Brazil, where they are as- sociated with calcareous rocks and caves (Williams 1999). This species was Þrst described as Phlebotomus lon- gipalpis by Lutz and Neiva (1912) from specimens collected from Sa ˜ o Paulo and near Cordisburgo in the state of Minas Gerais, Brazil. The original description is incomplete, no holotype was designated, and the original series of specimens have been lost. The spe- cies was redescribed by Franc ¸ a in 1920, using speci- mens from Mangaratiba-Quixada ´ (Ceara ´ ), Sa ˜ o Paulo, and Minas Gerais in Brazil as well as “Rives du Parana ´” Paraguay. Molecular and biochemical studies of Lu. longipal- pis have resulted in the emergence of three schools of thought regarding its taxonomic status: (1) that it is a complex of morphologically similar, allopatric species (Lanzaro et al. 1993, Lanzaro and Warburg 1995, Mutebi et al., 1998, 1999, Arrivillaga et al. 2000), (2) that it is a complex of species, some of which may occur sympatrically, isolation being maintained by the production of chemically distinct terpenoid phero- mones (Ward et al. 1983, 1988, Hamilton et al. 1996, Lampo et al. 1999), and (3) that it is a single hetero- geneous species with a wide geographical distribution (Bonnefoy et al. 1986, Mukhopadhay et al. 1998, Mun- stermann et al. 1998). Based on isoenzyme comparisons and cross-mating experiments using laboratory-reared sand ßies from Costa Rica, Colombia and Brazil, Lanzaro et al. (1993) suggested that Lu. longipalpis was a complex of at least three allopatric species. Subsequent isozyme studies 1 Universidad de Carabobo, BIOMED, Centro Nacional de Refer- encia de Flebotomos, La Morita, Maracay, Venezuela, Apdo. 4873 (e-mail: [email protected]). 2 University of Texas Medical Branch, Department of Pathology, Galveston, Texas 77555Ð 0609. 3 Universidad Central de Venezuela, Instituto de Zoologia Tropical, Laboratory Biologia y Ecologia de Poblaciones, Caracas, Venezuela. 4 Laborato ´ rio de Entomologia Me ´ dica, Centro de Pesquisas Rene ´ Rachou, Fundac ¸ a ˜ o Oswaldo Cruz, Av. Augusto de Lima 1715, 30190- 002 Belo Horizonte-MG, Brazil. 5 University of California, Department Entomology, Briggs Hall, Davis, California, USA, 95616 Ð 8579. 0022-2585/03/0615Ð0627$04.00/0 2003 Entomological Society of America

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

MORPHOLOGY, SYSTEMATICS, EVOLUTION
The Taxonomic Status of Genetically Divergent Populations ofLutzomyia longipalpis (Diptera: Psychodidae) Based on the
Distribution of Mitochondrial and Isozyme Variation
JAZZMIN ARRIVILLAGA,1 JOHN- PAUL MUTEBI,2 HERMES PINANGO,3 DOUGLAS NORRIS,2
BRUCE ALEXANDER,4 M. DORA FELICIANGELI,1 AND GREGORY C. LANZARO5
J. Med. Entomol. 40(5): 615Ð627 (2003)
ABSTRACT The sand ßy, Lutzomyia longipalpis (Lutz & Neiva) reputedly is a complex of crypticspecies; however, there is currently no consensus as to the number of species in the complex or theirgeographic distributions.We conducted phylogenetic analyses of 31 populations from throughout thespecies range, using seven isozyme loci and genes in themitochondrial genome. Analyses of these twoindependent sets of markers were largely concordant and revealed four distinct clades that supportthe existence of four species. The four clades have distinct geographic ranges: (1) Brazil (Species A�Lu. longipalpis sensu stricto), (2) Laran (Species B � Lu. pseudolongipalpis), (3) cis-Andean (SpeciesC), and (4) trans-Andean (Species D). The cis-Andean clade may be subdivided further into twogroups, one in Colombia and one in northwestern Venezuela, but their taxonomic status remainsunresolved. Knowledge that Lu. longipalpis is a complex of species may ultimately shed light onanomalies in the epidemiology of visceral leishmaniasis in the New World.
KEY WORDS Lutzomyia longipalpis, species complex, isozymes, mitochondrial DNA, phylogeny
THE SAND FLY Lutzomyia longipalpis (Lutz and Neiva1912) sensu lato is the primary vector of Leishmaniachagasi the etiological agent of zoonotic visceral leish-maniasis (ZVL) in South and Central America. It hasa wide geographical range that extends from the Yu-catan Peninsula in Mexico to northern Argentina andParaguay (Young andDuncan 1994), including all thecountries of Central America except Belize and mostof those of tropical South America east of the Andes.There are no published records ofLu. longipalpis fromGuyana, Surinam,orFrenchGuiana. Inaddition, thereare no records of this species from the PaciÞc Coast ofColombia, Ecuador, Peru, or Chile. Although Lu. lon-gipalpis has one of the widest geographical ranges ofany species in thegenus, its closest relatives (membersof the subgenus Lutzomyia, series longipalpis) are al-most entirely restricted to Brazil, where they are as-sociated with calcareous rocks and caves (Williams1999).
This species was Þrst described as Phlebotomus lon-gipalpis by Lutz and Neiva (1912) from specimenscollected from Sao Paulo and near Cordisburgo in thestate of Minas Gerais, Brazil. The original descriptionis incomplete, no holotype was designated, and theoriginal series of specimens have been lost. The spe-cies was redescribed by Franca in 1920, using speci-mens from Mangaratiba-Quixada (Ceara), Sao Paulo,andMinasGerais in Brazil as well as “Rives du Parana”Paraguay.Molecular and biochemical studies of Lu. longipal-
pis have resulted in the emergence of three schools ofthought regarding its taxonomic status: (1) that it is acomplex of morphologically similar, allopatric species(Lanzaro et al. 1993, Lanzaro and Warburg 1995,Mutebi et al., 1998, 1999, Arrivillaga et al. 2000), (2)that it is a complex of species, some of which mayoccur sympatrically, isolation beingmaintained by theproduction of chemically distinct terpenoid phero-mones (Ward et al. 1983, 1988, Hamilton et al. 1996,Lampo et al. 1999), and (3) that it is a single hetero-geneous species with a wide geographical distribution(Bonnefoy et al. 1986,Mukhopadhay et al. 1998,Mun-stermann et al. 1998).Based on isoenzyme comparisons and cross-mating
experiments using laboratory-reared sand ßies fromCosta Rica, Colombia andBrazil, Lanzaro et al. (1993)suggested thatLu. longipalpiswas a complex of at leastthree allopatric species. Subsequent isozyme studies
1 Universidad de Carabobo, BIOMED, Centro Nacional de Refer-encia de Flebotomos, La Morita, Maracay, Venezuela, Apdo. 4873(e-mail: [email protected]).
2 University of Texas Medical Branch, Department of Pathology,Galveston, Texas 77555Ð0609.
3 UniversidadCentral deVenezuela, Instituto deZoologiaTropical,Laboratory Biologia y Ecologia de Poblaciones, Caracas, Venezuela.
4 Laboratorio de Entomologia Medica, Centro de Pesquisas ReneRachou, Fundacao Oswaldo Cruz, Av. Augusto de Lima 1715, 30190-002 Belo Horizonte-MG, Brazil.
5 University of California, Department Entomology, Briggs Hall,Davis, California, USA, 95616Ð8579.
0022-2585/03/0615Ð0627$04.00/0 � 2003 Entomological Society of America

Tab
le1
.C
olle
ctio
nsi
tes,
sam
ple
size
sfo
ris
ozym
ean
dm
itoc
hond
rial
anal
yses
and
hapl
otyp
efr
eque
ncie
sfo
r3
1po
pula
tion
sof
Lu.
long
ipal
pis
Population
Location
Isozymen
(S.E.)
COIregionn(h
aplotypefreq.)
12Sregionn(h
aplotypefreq.)
16Sregionn
(hap
lotypefreq.)
Colombia
Bucaraman
ga
07�08�
N,73
�09�
W58.4(10.4)
n�
38(O
�35,P
�3)
n�
14(A
�14)
n�
14(A
�14)
PaloGordo
07�08�
N,73
�09�
W40.3(7.6)
Neiva
02�56�
N,75
�18�
W11.3(1.4)
n�
12(Q
�12)
n�
10(A
�10)
n�
12(A
�14)
Durania
08�08�
N,73
�09�
W27.9(2.9)
--
-CostaRica
Brasilito
10�28�
N,85
�18�
W71.3(11.2)
--
-Liberia
10�28�
N,85
�18�
W71.4(11.1)
n�
39(R
�39)
n�
13(F
�13)
n�
12(A
�12)
Honduras
Ran
choGrande
14�02�
N,87
�41�
W41.3(10.4)
--
-Orocu
ina
13�29�
N,87
�06�
W47.7(7.7)
--
-TigerIsland
13�17�
N,87
�37�
W38.7(7.3)
--
-San
FranciscodelCoray
13�40�
N,87
�30�
W36.3(7.3)
--
-Pavan
a13
�25�
N,87
�20�
W18.1(2.1)
n�
25(S
�25)
n�
16(G
�16)
n�
12(A
�12)
San
Juan
Bau
tista
13�34�
N,87
�16�
W18.9(2.3)
--
-LosGuatales
13�21�
N,87
�37�
W28.1(4.6)
--
-Nicaragua
Poch
omil
11�45�
N,86
�30�
W18.1(2.1)
--
-Las
Huertas
13�02�
N,86
�34�
W49.0(8.8)
--
-Venezu
ela
Curarigua
09�59�
N,69
�55�
W15.1(1.8)
n�
39(A
�36,B
�2,
C�
1)n
�16
(A�
2,B
�1,
C�
1,D
�12)
n�
12(A
�10,B
�2)
Cojedes
09�40�
N,68
�40�
W15.0(1.6)
--
-Trujillo
09�25�
N,70
�30�
W15.4(1.7)
n�
32(E
�30,F
�2)
n�
14(G
�14)
n�
12(A
�12)
ElPaso
09�59�
N,69
�55�
W16.4(1.6)
n�
29(A
�26,B
�1,
D�
2)n
�20
(D�
20)
-Brazil
Pakaraima
05�30�
N,60
�40�
W-
n�
28(L
�25,M
�3)
n�
12(F
�12)
n�
17(A
�17)
San
tarem
02�26�
S,54
�41�
W61.6(3.8)
n�
13(H
�12,I
�1)
--
Salvaterra
00�46�
S,48
�31�
W54.3(5.9)
n�
34(G
�34)
--
Baturite
04�20�
S,38
�53�
W52.3(2.8)
n�
32(K
�32)
n�
14(E
�14)
n�
17(A
�17)
Jaco
bina
11�11�
S,40
�31�
W65.3(2.8)
n�
16(J
�16)
n�
16(K
�16)
n�
16(A
�16)
Lap
inhacaves
19�03�
S,43
�57�
W63.7(3.1)
n�
35(N
�35)
n�
9(A
�9)
n�
16(A
�16)
Fortaleza
03�43�
S,38
�30�
W59.9(4.9)
--
-Sobral
03�42�
S,40
�21�
W60.3(5.0)
--
-Cam
ara
00�46�
S,48
�31�
W62.9(4.7)
--
-Bacab
al00
�46�
S,48
�31�
W59.1(3.2)
--
-Itap
ipoca
03�30�
S,39
�35�
W60.6(4.4)
--
-MontesClaros
16�43�
S,43
�52�
W62.1(5.2)
--
-
616 JOURNAL OF MEDICAL ENTOMOLOGY Vol. 40, no. 5

compared wild-caught specimens from different lo-calities within Colombia (Lanzaro et al. 1997), Brazil(Mutebi et al. 1999), and Central America (Mutebi etal. 1998), the results of which provided additionalsupport for the hypothesis that one species occurs ineach of the three regions. Arrivillaga et al. (1999a,b)and Arrivillaga and Feliciangeli (2000) presented ev-idence for two cryptic species within the Lu. longi-palpis complex in Venezuela, based on two diagnosticisozyme loci and morphometric analysis of larvalmouthpart characters. Lampo et al. (1999) reportedthe occurrence of two species occurring sympatricallyat Curarigua (El Paso), Venezuela, based on the sametwo isozyme loci. One of these species showed simi-larities to both Brazilian and Colombian populations,whereas the other appeared to be indigenous to Ven-ezuela.Although the patchy distribution of American vis-
ceral leishmaniasis and different manifestations of Le.chagasi infection may be explained by reasons otherthan the involvement of several species within a Lu.longipalpis complex, the public health importance ofthe disease is such that the true identity of the vectorin each focus needs to be determined. Visceral leish-maniasis has been reported from all countries whereLu. longipalpis is present, with the exception of Pan-ama (WHO 1990). An atypical, nodular form of cu-taneous leishmaniasis caused by Le. chagasi has beenreported in some areas of Central America (Zeledonet al., 1989; Ponce et al., 1991, Belli et al., 1999). Failureof the parasite to visceralize in many patients in Cen-
tral Americamay be linked to lower concentrations ofthe vasodilator maxadilan in the saliva of Lu. longipal-pis from this region (Warburg et al., 1994, Yin et al.,2000).Simultaneous analyses of genetic markers in the
nuclear and mitochondrial genomes can provide use-ful insights into the evolutionary history and taxo-nomic status of populations. In the current study weinvestigated geographical variation in the cytochromec oxidase I gene (COI), as well as the 12S and 16Sribosomal genes all within themitochondrial genome.In addition, we studied several enzyme encoding nu-cleargeneswith thegoalof clarifying thephylogeneticand taxonomic relationships among populations ofLu.longipalpis.
Materials and Methods
Sand Fly Collections. Specimens of Lu. longipalpiswere collected from 31 different localities in six coun-tries (Table 1; Fig. 1). Flies were aspirated directlyfrom domestic animals (horses, cows, pigs, and chick-ens) or from surfaces in and around their enclosures.In someplaces these collectionswere augmentedwithcollections fromCDC light traps. Three other sand ßyspecies were analyzed as outgroups: Phlebotomus pa-patasi (Scopoli) from a colony originating from Israel,Lutzomyia evansi (Nunez Tovar) collected in CostaRica and Lutzomyia gomezi (Nitzulescu) collected inVenezuela.
Fig. 1. Collection sites for the 31 populations Lu. longipalpis included in this study.
September 2003 ARRIVILLAGA ET AL.: MOLECULAR-BIOCHEMICAL ANALYSIS OF THE Lu. longipalpis 617

mtDNA Variation
Polymerase Chain Reaction (PCR) Amplification.GenomicDNAwas extracted andpuriÞed from frozensingle sand ßies using previously described methods(Bender et al. 1983). Three target DNA fragmentswere ampliÞed: COI, 12S and 16S mtDNAs. AmpliÞ-cation reactions were carried out in 25 �l containing0.5 U of Taq polymerase (Biolase), one X enzymebuffer with MgCl2 (2.5 �l/reaction), 100 nmol dNTP,and 3�lDNAas template. AmpliÞcation and sequenc-ing of puriÞed products were achieved using the fol-lowing primers: COI region (Kambhapati and Smith1995) TGATCAAATTTATAAT (�) and GGTA-AAATT AAAATATAAACTTC (�); 12S region (Si-monet al. 1994)TACTATGTTACGACTTAT(�) andAAACTAGGATTAGATACCC (�); 16S region (Si-mon et al. 1994) TTACGCTGTTA TCCTAA (�) andCACCTGTTTAACAAAAACAT (�).
SSCP and Sequencing Analyses. Polymorphismwasdetectedby single strandconformationpolymorphism(SSCP) analysis for each specimen (12Ð39 individu-als)perpopulation for 13populations (Table1;Fig. 1).Gels for SSCPwere composedof 0.6XTBE(1XTBE�53 mM Trizma base, 53 mM boric acid, 1.5 mM EDTAph 8.0), 7.9% acrylamide, and 0.21% N,N�-methylle-nebisacrylamide. Following PCR ampliÞcation 7 �l ofproduct was removed to a 500 �l tube containing 9 �lof denaturing loading mix (20 mM NaOH, 90% form-amide, 0.05% bromophenol blue, and 0.05% xylenecyanol). The tube was tapped to mix the contents,spun brießy in a microcentrifuge, heated to 95�C for3 min in a thermal cycler and plunged into ice for atleast 10 min. From this cooled mixture, 16 �l wasloaded into the SSCP gel. Electrophoresis was carriedout on vertical slab gels (16 � 18 cm) that were runin circulating electrode buffer (0.5X TBE) refriger-ated at 4�C.Currentwasmaintained at 20mAuntil thexylene cyanol dye had migrated to the bottom of thegel. Gels were then silver stained, dried, scored, andphotographed. Polymorphism was detected by SSCPin the following way: Preliminary studies veriÞed thereproducibility of SSCP migration patterns. This in-formation was used to develop an optimal protocol.SSCP was Þrst used to identify unique haplotypeswithin single populations. These were then used tocreate haplotype proÞles for each populationscreened. Population proÞles were compared by re-running putative haplotypes, from all populations, ona single gel. Haplotypes were veriÞed by direct cyclesequencing on anABI-377 automated sequencer (Per-kin-Elmer).
Sequence Analyses. Sequences for all COI, 12S and16S mitochondrial haplotypes initially were alignedusing Clustal V (Higgins et al. 1992) and manuallyadjusted for obvious misalignments. Gaps weretreatedasmissingdata.All phylogenetic analyseswerecomputed using PAUP 4.0b2 (Swofford 1999). Neigh-bor-joining (NJ) analysis (Saito and Nei 1997) wascomputed under the assumptions of the Kimura-2-parameter model (K2P). Maximum parsimony (MP)analysis was computed with equally weighted and
reweighted characters (Farris 1983, Goloboff 1993)based on the consistency index (Farris 1969) by heu-ristic search using the tree bisection-reconnection(TBR) branch-swapping algorithm with 100 randomstepwise additions. This adjusts the weight of charac-ters (nucleotides) based on their Þt to the most par-simonious tree after deleting uninformative charac-ters (Carpenter 1996). ConÞdence values wereestimated by bootstrapping (1,000 replications forCOI and 100 for 12S and 16S). The nucleotide com-position, pairwise base differences, and genetic dis-tance (K2P) were determined with PAUP 4.Ob2.
Isozyme Variation
Isozyme Analysis. Sand ßies from 30 populations(Table 1; Fig. 1) were assayed for genetic variation atseven isozyme loci using standard starch gel electro-phoresis. Individual sand ßies were homogenized in 8�l ofdistilledwater.Thehomogenatewas fractionatedbyelectrophoresis on 12.5%(wt:vol) horizontal starchgels. The seven enzymes assayed were: hexokinase(E. C. 2.7.1.1), isocitrate dehydrogenase- one (E. C.1.1.1.42), malic acid dehydrogenase-1 (E.C. 1.1.1.37),malic acid dehydrogenase- two (E. C. 1.1.1.37), malicenzyme-1 (E. C. 1.1.140), phosphoglucoisomerase(E. C. 5.3.1.9), and alpha-trehalase (E. C. 3. 2.1.28).Running buffers and histochemical staining methodswere as described by Lanzaro et al. (1993). Differ-ences inmigrationdistances of bands at the same locuswere designated as different alleles, and multiplebands at the same locus in an individual were assumedto be heterozygous. Loci were designated with posi-tive or negative codes depending on whether theymigrated to the anode or cathode. For each locus, themost frequent electromorph was given an rf value of1.00. Sand ßies from three laboratory stocks [i.e., Mel-gar (Colombia), Liberia (Costa Rica) and LapinhaCaves (Brazil)] used by Lanzaro et al. (1993) wereincluded on each gel to aid in identiÞcation of alleles.
Population Genetic Analysis. Individual ßy geno-types were organized into text input Þles and popu-lations were analyzed for genetic variability, compli-ance to Hardy-Weinberg expectations, and geneticrelatedness using the computer program, BIOSYS-1.(Swofford and Selander 1989). Allele frequencies andNeiÕs genetic distance were calculated. Phenetic anal-ysis was completed using a cluster analysis employingtheunweightedpair-groupmethod(unweightedpair-group method with arithmetic average) of NeiÕs ge-netic distance (1978).
Isozyme Phylogenetic Analyses. Raw input Þles forthe BIOSYS-1 program were converted to PHYLIP3.5c version format (Phylogeny Inference Package;Felsenstein, 1995) and the continuous character max-imum likelihood (CONTML)programwasperformedfor gene frequencies (all alleles at each locus). Lu.evansiwas used as an outgroup, global rearrangementswereallowed, and the inputorderof species (OTUÕs�populations in this case) were randomized. Geneticdistances were computed using the GENDIST pro-gram (PHYLIP 3.5c) assuming that each locus con-
618 JOURNAL OF MEDICAL ENTOMOLOGY Vol. 40, no. 5
tained all alleles, and Nei distances were calculatedfrom the original data set (allele frequency). A searchfor the best hypothetical phylogenetic tree was per-formed using the FITCH program (PHYLIP 3.5c) us-ing the distances obtained from GENDIST as input;negative branches were not allowed, and Lu. evansiwas Þxed as reference group (outgroup). Global re-arrangements were allowed and the input order ofOTUÕswere randomizedwith Þve jumbles during ran-domization. Further analysis was done with theKITSCHprogram(PHYLIP 3.5c)whichuses the sameadditive model employed in FITCH but assumes anevolutionary clock.
Results
mtDNA Variation
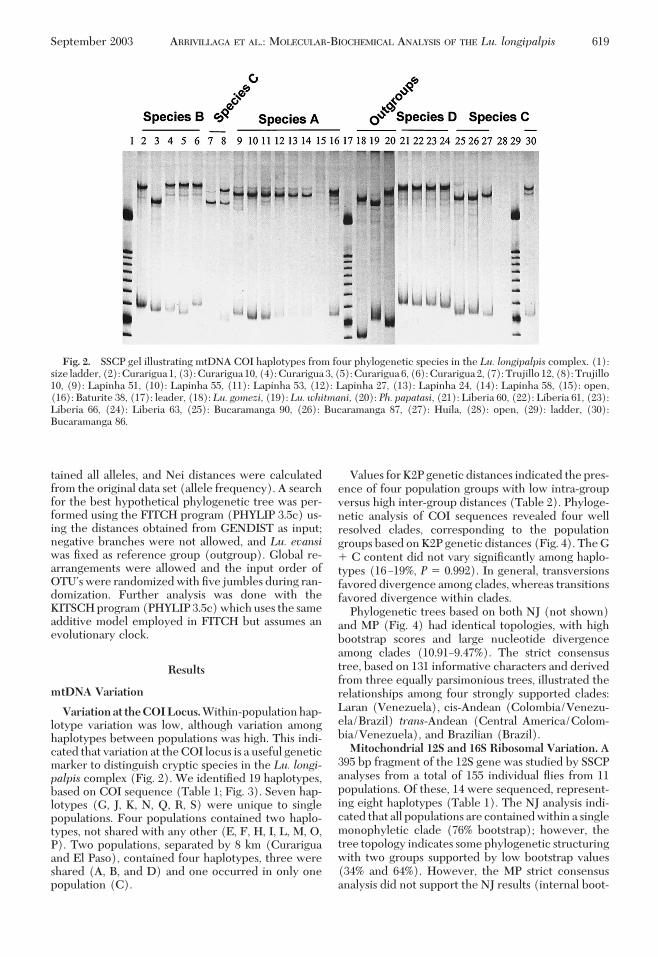
Variationat theCOILocus.Within-populationhap-lotype variation was low, although variation amonghaplotypes between populations was high. This indi-cated that variation at theCOI locus is a useful geneticmarker to distinguish cryptic species in the Lu. longi-palpis complex (Fig. 2). We identiÞed 19 haplotypes,based on COI sequence (Table 1; Fig. 3). Seven hap-lotypes (G, J, K, N, Q, R, S) were unique to singlepopulations. Four populations contained two haplo-types, not shared with any other (E, F, H, I, L, M, O,P). Two populations, separated by 8 km (Curariguaand El Paso), contained four haplotypes, three wereshared (A, B, and D) and one occurred in only onepopulation (C).
Values forK2P genetic distances indicated the pres-ence of four population groups with low intra-groupversus high inter-group distances (Table 2). Phyloge-netic analysis of COI sequences revealed four wellresolved clades, corresponding to the populationgroups based onK2P genetic distances (Fig. 4). TheG� C content did not vary signiÞcantly among haplo-types (16Ð19%, P � 0.992). In general, transversionsfavored divergence among clades, whereas transitionsfavored divergence within clades.Phylogenetic trees based on both NJ (not shown)
and MP (Fig. 4) had identical topologies, with highbootstrap scores and large nucleotide divergenceamong clades (10.91Ð9.47%). The strict consensustree, based on 131 informative characters and derivedfrom three equally parsimonious trees, illustrated therelationships among four strongly supported clades:Laran (Venezuela), cis-Andean (Colombia/Venezu-ela/Brazil) trans-Andean (Central America/Colom-bia/Venezuela), and Brazilian (Brazil).
Mitochondrial 12S and 16S Ribosomal Variation. A395 bp fragment of the 12S gene was studied by SSCPanalyses from a total of 155 individual ßies from 11populations. Of these, 14 were sequenced, represent-ing eight haplotypes (Table 1). The NJ analysis indi-cated that all populations are containedwithin a singlemonophyletic clade (76% bootstrap); however, thetree topology indicates somephylogenetic structuringwith two groups supported by low bootstrap values(34% and 64%). However, the MP strict consensusanalysis did not support the NJ results (internal boot-
Fig. 2. SSCP gel illustrating mtDNA COI haplotypes from four phylogenetic species in the Lu. longipalpis complex. (1):size ladder, (2):Curarigua1, (3):Curarigua10, (4):Curarigua3, (5):Curarigua6, (6):Curarigua2, (7):Trujillo12, (8):Trujillo10, (9): Lapinha 51, (10): Lapinha 55, (11): Lapinha 53, (12): Lapinha 27, (13): Lapinha 24, (14): Lapinha 58, (15): open,(16): Baturite 38, (17): leader, (18): Lu. gomezi, (19): Lu. whitmani, (20): Ph. papatasi, (21): Liberia 60, (22): Liberia 61, (23):Liberia 66, (24): Liberia 63, (25): Bucaramanga 90, (26): Bucaramanga 87, (27): Huila, (28): open, (29): ladder, (30):Bucaramanga 86.
September 2003 ARRIVILLAGA ET AL.: MOLECULAR-BIOCHEMICAL ANALYSIS OF THE Lu. longipalpis 619

Fig. 3. Unique mtDNA haplotypes found for the CO I gene fragment from 13 populations of Lu. longipalpis. Eachhaplotype was assigned a letter designation (AÐS). Haplotype frequencies for each population are presented in Table 1.Sequence for a�495bp fragmentof theCOIgene ispresented. (- indicates gaps indels).Abbreviationsdesignatinghaplotypesrefer to site of origin as follows: jacobina � Jacobina, Brazil; Cu � Curarigua, Venezuela; La � Lapinha Caves, Brazil; Tru �Trujillo, Venezuela; Pa � Pavana, Honduras; Li � Liberia, Costa Rica; Bu � Bucaramanga, Colombia; baturite � Baturite,Brazil; ne�Huila, Brazil; ro�Pacaraima, Brazil; santa� Santarem,Brazil; salva� Salvaterra, Brazil. Numbers following somenames indicate that more than one haplotype was recovered from that site.
620 JOURNAL OF MEDICAL ENTOMOLOGY Vol. 40, no. 5

strapping �50%), indicating no divergence amongpopulations based on 107 informative characters(trees not presented).SSCP analysis of a 360 bp fragment of the 16S
mtDNA locus was conducted on 140 ßies from 10populations. Sequence obtained for 11 individuals re-vealed only two haplotypes (Table 1). MP and NJanalyses, employing 25 informative characters, pro-ducedphylogenetic treeswithno signiÞcantbootstrap
values, indicating no divergence at this locus amongthe populations studied (trees not presented).
COI versus 12S/16S Based Phylogenies. Phyloge-netic analysis of mtDNA ribosomal sequences re-vealed a high degree of homogeneity among popula-tions of Lu. longipalpis collected over an extensivegeographical area. In contrast, phylogenetic analysisof COI sequence, conducted on the same set of pop-ulations, revealed signiÞcant levels of divergence.This
Table 2. Kimura-2- parameter (K2P) genetic distances among four clades based on cythocrome c oxidase I from 13 populations ofLu. longipalpis. The number of populations for each clade are designated N. Standard error in parenthesis
Laran cis-Andes trans-Andes Brazil
Laran (N � 2) 0.013 (0.006) 0.125 (0.02) 0.129 (0.006) 0.123 (0.003)cis-Andes (N � 4) Ñ 0.055 (0.022) 0.130 (0.016) 0.101 (0.012)trans-Andes (N � 4) Ñ Ñ 0.077 (0.029) 0.101 (0.01)Brazil (N � 5) Ñ Ñ Ñ 0.0324 (0.01)
Fig. 4. Strict Consensus tree obtained by MP analysis based on 131 mtDNA COI informative characters. Numbers onbranches are bootstrap values based on 1,000 replicates. Labels at the terminus of each branch provide the name of thepopulation. The speciÞc haplotype is indicated in parentheses by an alphabetical designation corresponding to those in Fig.4 and in Table 1. Lu. evansi represents the in group and Ph. papatasi the outgroup.
September 2003 ARRIVILLAGA ET AL.: MOLECULAR-BIOCHEMICAL ANALYSIS OF THE Lu. longipalpis 621

differencemay be the consequence of a slower rate ofevolution of the ribosomal region.
Isozyme Variation
Genetic distance coefÞcients, D (Nei 1978), basedon analysis of seven enzyme encoding loci revealedhigh levels of genetic differentiation among popula-tion groups originating from Þve regions; Colombia,Central America, Brazil and two areas in Venezuela(Table 3). Cluster analysis was conducted on a matrixof D values for all pairwise comparisons among 30populations employing the unweighted pair group al-gorithms using arithmetic averages (unweighted pair-groupmethodwith arithmetic average) of Sneath andSokal (1973). In the resulting dendrogram (Fig. 5)populations in each of the Þve geographic regionsclustered together. Levels of genetic distance amongthe Þve groups (D � 0.12Ð0.49) were high and con-
sistent with values previously reported for sibling spe-cies in the genus Lutzomyia (Lanzaro and Warburg1995). Within regions, values for D were consistentlybelow 0.02, typical for levels of divergence betweenlocal populations within a single insect species. Thetotal FST for all populations for all loci was high 0.62,indicating genetic sub-structuring amongpopulations.Effective migration rates (Nem) among all 30 Þeldcollected populations were extremely low, Nem �0.15, indicating restricted gene ßow among popula-tions. Several enzyme loci diagnostic for the Þve pop-ulation groups identiÞed in this study includedMdh-2,Gpi, andHk(Table 4).Gels illustratingMdh-2 andGpizymograms are presented in Fig. 6.
Isozyme Phylogeny
CONTML Analysis. A single rooted tree was ob-tained (Fig. 7A) from 5,547 trees analyzed based on
Table 3. Genetic distance (Nei 1978) based on seven isozyme loci, within and between species in the Lu. longipalpis species complex.Range in parentheses
SpeciesNo. of
populationsSpecies B (Laran)
Species A(Brazilian)
Species C1
(cis-Andean:Colombian)
Species C2
(cis-Andean:Venezuelan)
Species D(trans-Andean)
Species B 2 0.004 (0.004Ð0.004)Species A 11 .325 (0.276Ð0.391) 0.003 (0.000Ð0.015)Species C1 11 0.487 (0.438Ð0.539) 0.150 (0.133Ð0.166) 0.000 (0.000Ð0.001)Species C2 2 0.415 (0.345Ð0.493) 0.147 (0.116Ð0.175) 0.141 (0.107Ð0.174) 0.015 (0.015Ð0.015)Species D 4 0.466 (0.370Ð0.547) 0.145 (0.113Ð0.168) 0.133 (0.084Ð0.165) 0.121 (0.073Ð0.172) 0.017 (0.003Ð0.030)
Fig. 5. A unweighted pair-group method with arithmetic average dendrogram constructed from values for pairwisegenetic distance (Nei 1978) for 30 Þeld populations of Lu. longipalpis in Colombia, Venezuela, Central America and Brazil.Labels on branches indicate the geographic region of origin for each population group.
622 JOURNAL OF MEDICAL ENTOMOLOGY Vol. 40, no. 5

Table 4. Allele frequencies for 3 isozyme loci that discriminated 5 clades from field-collected Lu. longipalpis in South and CentralAmerica. Frequencies of the most common alleles are highlighted in bold print. Clades in this table are based on cluster analysis of Nei’sgenetic distance D (Fig. 5)
Locus
Clade
cis-Andean(Colombia)
trans-Andean Brazilian cis-Andean (Venezuela) Laran
Mdh-2(N) (226) (714) (791) (40) (40)A 0.998 0.996 0.994 0.900 0.087B 0.002 0.004 0.004 0.063 0.063C 0 0 0.001 0 0D 0 0 0 0.038 0.850
Gpi(N) (162) (523) (641) (40) (39)A 0.892 0.004 0.004 0.063 0.026B 0.022 0.973 0.011 0.025 0.013C 0.040 0.005 0.013 0.863 0.154D 0.022 0 0.002 0.013 0.026E 0.003 0.001 0 0 0.026F 0.003 0.007 0.007 0 0G 0.012 0 0 0.038 0H 0 0 0.001 0 0I 0 0.011 0.962 0 0.756J 0.006 0 0 0 0
Hk(N) (116) (336) (742) (34) (39)A 0.897 0.991 0.993 0.956 0.051B 0.078 0.007 0.004 0.015 0C 0.026 0.001 0.003 0.029 0.949
Fig. 6. (A) A starch gel stained for malic acid dehydrogenase-2 (Mdh-2, E.C. 1.1.1.37). Samples include specimens fromfour populations in Venezuela. Samples from Trujillo and Cojedes represent species C and carry the A allele at this locus.Samples from El Paso and Curarigua represent species B and carry the B allele. CO and BZ are colony controls fromMelgar,Colombia andLapinhaCaves, Brazil respectively. (B)A starch gel stained for the enzymephosphoglucoisomerase (Gpi, E. C.5.3.1.9). The gel contains 30 Lu. longipalpis individuals from eight populations. Bucaramanga and Palo Gordo are in Colombiaand represent speciesC1; Liberia andBrasilito are inCostaRica and represent speciesD; Salvaterra andFortaleza are inBraziland represent species A; and Curarigua is in Venezuela and represents species B. There are six alleles illustrated on this gel.The A allele is the most common allele in species C; the B allele is the most common in species A and B; and the I allele ismost common in species D. The F, S1 and S2 alleles are less common.
September 2003 ARRIVILLAGA ET AL.: MOLECULAR-BIOCHEMICAL ANALYSIS OF THE Lu. longipalpis 623
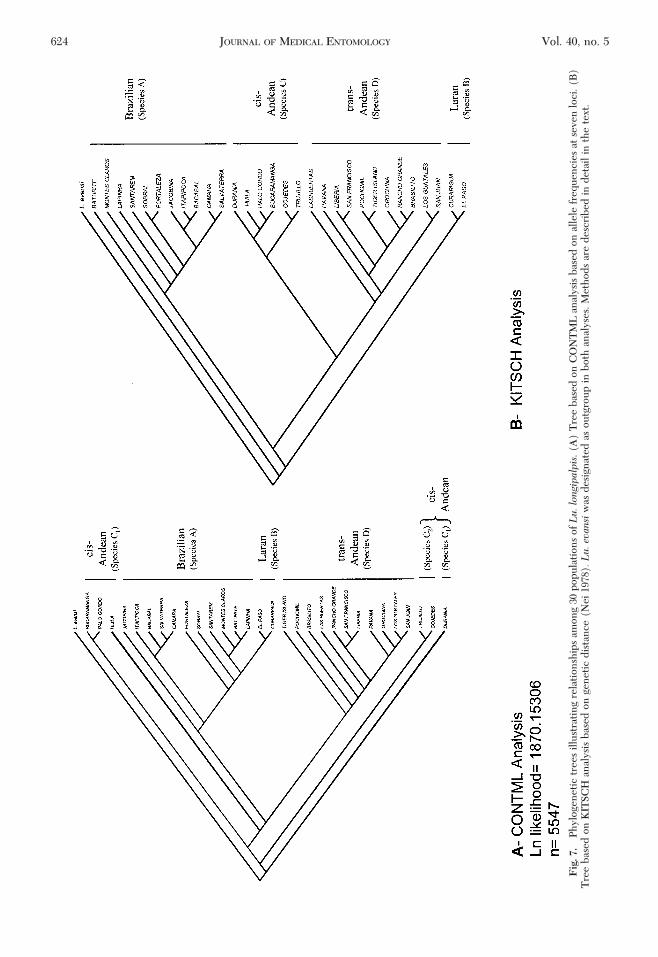
Fig
.7.
Phylogenetictreesillustratingrelationshipsam
ong30
populationsofLu.lo
ngi
palp
is.(A
)TreebasedonCONTMLan
alysisbasedonallele
frequenciesat
sevenloci.(B
)TreebasedonKITSCH
analysisbasedongeneticdistance
(Nei1978).
Lu.ev
ansiwas
designatedas
outgroupin
both
analyses.Methodsaredescribedin
detailin
thetext.
624 JOURNAL OF MEDICAL ENTOMOLOGY Vol. 40, no. 5

allozyme frequencies. The tree topology showsfour independent, monophyletic clades: cis-AndeanÐColombia represents an ancestral, Andean gene pool,closely related to three recently evolved and mono-phyletic internal groups: Brazilian, Laran (northwestVenezuelan populations) and trans-AndeanÐ CentralAmerican populations. The two monophyletic lin-eages (Brazilian and trans-Andean) contain complexinternal branching (Fig. 7A). The Brazilian popula-tions consist of two internal groups. Themost recentlyevolved in-group is represented by two Venezuelanpopulations that make up the Laran Clade. However,the internal “paraphyletic” grouping indicates a dif-ferent divergence time for these groups. The two in-ternalBraziliangroupscontainpopulations thatdonothave well-deÞned geographical distributions, indicat-ing extensive gene ßow between them. The trans-Andean Clade contains a cluster of closely relatedCentral American populations and appears to repre-sent an ancestral sister-groupof the cis-AndeanClade.This arrangement indicates extensive gene ßowamong populations in Central America.
KITSCH AnalysisThis analysis revealed four monophyletic clades
(Fig. 7B). The most ancestral sister groups are theBrazilian and Laran clades. The most recently di-verged group is represented by the trans-AndeanClade, recently diverged from the cis-Andean Clade.
Analyses of Isozyme Variation versus PhylogeneticAnalysis of Isozymes. Different topologies were ob-tained from the phylogenetic analyses performed(CONTML and KITSCH). However, similar conclu-sions regarding the relationships among populationsand their taxonomic status can be drawn from the twotrees illustrated in Fig. 7. Both analyses revealed fourclades that may be interpreted as phylogenetic spe-cies.Thephenogram,basedonunweightedpair-groupmethod with arithmetic average cluster analysis ofgenetic distance (Fig. 5) clearly resolved the samegroups, although it further split the cis-Andean Cladeinto two groups (Colombian and Venezuelan).
DiscussionmtDNA and isozyme variation were used as com-
plementary tools for the identiÞcation of species
Fig. 8. Geographical distribution of species in the Lu. longipalpis complex. (A) Distribution of species based onunweighted pair-group method with arithmetic average cluster analysis of genetic distance (Nei 1978) computed fromallozyme frequencies (seven loci) among 30 populations (Fig. 5). (B) Distribution of species based on phylogenetic analysisof isozyme data [same data as in (A), Fig. 7B]. (C) Distribution of species based on phylogenetic (maximum parsimony)of sequence for the mitochondrial cytochrome c oxidase I locus recovered from 13 populations of Lu. longipalpis (Fig. 4).
September 2003 ARRIVILLAGA ET AL.: MOLECULAR-BIOCHEMICAL ANALYSIS OF THE Lu. longipalpis 625

boundaries amongmembers of theLu. longipalpis spe-cies complex. Results using the two approaches werelargely concordant. We recognized four cryptic spe-cies in what we considered the Lu. longipalpis speciescomplex: Species A, the Brazilian Clade, is repre-sented by 11 populations sampled throughout Brazil;Species B, the Laran Clade, is represented by popu-lations fromEl Paso and Curaruiga (La Rinconada) innorthwest Venezuela; Species C, the cis-AndeanClade, is representedbypopulations inColombia (Bu-caramanga, Palo Gordo, Neiva, Durania), Venezuela(Trujillo and Cojedes), and northern Brazil (Pacara-ima); and Species D, the trans-Andean Clade is rep-resented by 11 populations from various parts ofCentral America. The distribution of our species Aincludes the areas from which Lu. longipalpis wasoriginally described and should be considered to rep-resent Lutzomyia longipalpis sensu stricto.Our speciesB has been formally described as a new species, Lutz-omyia pseudolongipalpis Arrivillaga and Feliciangeli(Arrivillaga and Feliciangeli 2001). Formal descrip-tions including assignment of new scientiÞc names arecurrently underway for species C and D.There are some inconsistencies in the outcome of
our analyses. Phenetic analysis (unweighted pair-group method with arithmetic average) based on ge-netic distance (DNei) computed from allozyme fre-quencies grouped the 30 populations into Þve ratherthan four groups (Fig. 5). The cis-Andean Clade, spe-cies C was split. We designated species C1 to includespecimens collected from Bucaramanga, Palo Gordo,Durania, and Neiva in Colombia and species C2 in-cluded populations from Trujillo and Cojedes in Ven-ezula 5). This arrangement was based largely on threediscriminating markers. In the isozyme-based phylo-genetic analysis the differences because of autapo-morphic characters, such as MDH- two and HK, pro-vide no phylogenetic information among populations.The GPI character is a synapomorphy and is the mostinformative character in the MP analysis (data notshown). Consequently the phylogenetic approach re-solved four groups, rather than Þve (Fig. 7B). How-ever, examination of Fig. 7B revealed that C1 and C2
are split and their placement indicates that the twogroups were recently diverged. This arrangement isnot supported by the COI mitochondrial sequencedata. For the present, we are suggesting that fourspecies, rather than Þve, be considered until this dis-crepancy is resolved. The mitochondrial phylogenyindicates spatial overlap in the distributions of speciesC and D that is not evident in the analyses based onallozymes.
Acknowledgments
In Brazil, we thank Italo Sherlock, of FIOCRUZ, Salvador;Raimundo Nonato de Souza and Irineu Guerreiro de Souzaof FIOCRUZ, Fortaleza; Irolanda da Rocha Barata, EvaldoCarneiro das Chagas and Lourdes Maria Garcez Silveira ofInstituto Evandro Chagas, Belem; Elizabeth Rangel, ClaudioMeneses and Diamar C. Pinto of the Entomology Depart-ment FIOCRUZ, Rio de Janeiro for Þeld and technical as-
sistance. We are greatly indebted to the authorities atFIOCRUZ in Salvador, Fortaleza, and Rio de Janeiro as wellas the Instituto Evandro Chagas, Belem for providing us withtransportation. We are grateful to Francisco Freitas, Her-berto Carvalho Dantas Filho, Leonidas Monte Batista, Ma-noelCaetanoDuarte, andOsvaldoCarlos Silva da Silveira fortheir Þeld assistance inSantarem.Wealso thankPauloCalacaand Evangley Rios for Þeld assistance in Montes Claros. InVenezuela we thank Milagros Oviedo of Universidad de LosAndes for the Þeld collaboration in Trujillo localities, FreddyArias, Arturo Bravo, Maria Martinez, Florencio Mendoza,and CarlosMendez of the Centro Nacional de Referencia deTaxonomõa de Flebotomos for technical assistance, YadiraRangel for assistance with isozyme analysis and Juan CarlosNavarro for recommendations for the phylogenetic analyses.In Costa Rica we thank Marco Herrero and Julio Rojas ofUniversidad Nacional in Heredia; in Honduras we thankCarlos Ponce of the Central Laboratory, Ministry of Health,Tegucigalpa. InNicaraguawewould like to acknowledge theassistance of Alejandro Belli and Sonia Valle of the CentroNacional de Diagnostico y Referencia, Ministero de Salud,Managua. Finally, we thank Bill Sweeney and Scott MarriottofUTMB,Galveston, for technical support. This researchwassupported, in part, by a grant from the JohnD. andCatherineT. MacArthur Foundation program entitled “MolecularBiology of Parasite Vectors,” by Grant No. AI39540 from theNational Institutes of Health to G.C.L and by funds fromConselho Nacional de Desenvolvimento CientõÞco e Tecno-logico (CNPq), Fundacao Nacional de Saude (FUNASA) toBruceAlexander and a fellowship fromTDR-WHOto J. C.A.Travel to Roraima, Brazil was funded in part by FundacaoInstituto Oswaldo Cruz
References Cited
Arrivillaga, J. C., and M. D. Feliciangeli. 2000. Diferencia-cion morfologica larval entre poblaciones de Lutzomyialongipalpis (Lutz &Neiva, 1912) de Venezuela (Diptera:Psychodidae. Bol. Entomol. Venez. 15: 229Ð234.
Arrivillaga, J. C., and M. D. Feliciangeli. 2001. Lutzomyiapseudolongipalpis: the Þrst new species within the longi-palpis (Diptera: Psychodidae: Phlebotominae) complexfrom La Rinconada, Curarigua, Lara State, Venezuela.J. Med. Entomol. 38: 783Ð790.
Arrivillaga, J.C., J.C.Navarro, andM.D.Feliciangeli. 1999a.Morfologia y quetotaxia del tagma cefalico de Lutzomyia(Diptera: Psychodidae). Bol. Entomol. Venez. 14: 1Ð12.
Arrivillaga, J. C., Y. Rangel, M. D. Feliciangeli, and G. Lan-zaro. 1999b. Morphogenetic studies of Lutzomyia longi-palpis (Psychodidae: Phlebotominae). J. Am.Mosq. Con-tro. Assoc. 15: 410Ð411.
Arrivillaga, J. C., Y. Rangel, M. Oviedo, and M. D. Felician-geli. 2000a. Genetic Divergence among Venezuelanpopulations ofLutzomyia longipalpis (Dipteras: Psychod-idae: Phlebotominae). J. Med. Entomol. 37: 325Ð330.
Arrivillaga, J. C., Y. Rangel, M. Oviedo, and M. D. Felician-geli. 2000b. Correlated morphological and genetic di-versity among Lutzomyia longipalpis (Diptera: Psychod-idae) collections in Venezuela. J. Am.Mosq. Cont. Assoc.16: 171Ð174.
Belli, A., D. Garcia, X. Palacios, et al. 1999. Widespreadatypical cutaneous leishmaniasis caused by Leishmani(L.) chagasi in Nicaragua. Am. J. Trop. Med. Hyg. 61:380Ð385.
Bender, W., P. Spierer, and D. S. Hogness. 1983. Chromo-somal walking and jumping to isolate DNA from the aceand rosy loci and the bithorax complex in Drosophilamelanogaster. J. Mol. Bio. 168: 17Ð33.
626 JOURNAL OF MEDICAL ENTOMOLOGY Vol. 40, no. 5

Bonnefoy, S. M., F. Tibayrenc, J. P. Le Pont, P. Dujardin, P.Desjeux, and F. J. Ayala. 1986. An isozymic study ofLutzomyia longipalpis (Diptera: Psychodidae), the vectorof visceral leishmaniasis in the “ Yungas” (Bolivia). Cah.O.R.S.T. O. M. Ser. Entomol. Med. Parasitol. 24: 213Ð217.
Carpenter, J. M. 1996. Uninformative bootstrapping. Cla-distics 12: 177Ð181.
Farris, J. S. 1969. A successive approximations approach tocharacter weighting. Syst. Zool. 26: 269Ð276.
Farris, J. S. 1983. The logical basis of phylogenetic analysis,pp. 1Ð36. InN. I. Platnick andV.A. Funk [eds.], Advancesin Cladistics. Columbia University Press, New York.
Felsenstein, J. 1995. Phylip 3.5c version. Phylogenetic in-ference package�. Copyright 1986Ð1995 by JosephFelsenstein and the University of Washington.
Franca, C. 1920. Observations sur le genre Phlebotomus.II.ÑPhlebotomus du Nouveau Monde (Phlebotomes duBresil et du Paraguay). Bull. Soc. Portug. Sci. Nat. 8:215Ð236.
Goloboff, P. A. 1993. Estimating character weights duringtree search. Cladistics 9: 83Ð91.
Hamilton, J.G.C., R. D. Ward, M. J. Dougherty, et al. 1996.Comparison of the sex-pheromone components ofLutzomyia longipalpis from areas of visceral and atypicalcutaneous leishmaniasis in Honduras and Costa Rica.Ann. Trop. Med. Parasitol. 90: 533Ð541.
Higgins, D. G., A. J. Bleasby, and R. Fuchs. 1992. Clustal Vimproved software for multiple sequence alignment.Comp. Appl. Biol. 8: 189Ð191.
Kambhampati, S., and P. T. Smith. 1995. PCR primers forthe ampliÞcation of four insect mitochondrial gene frag-ments. Insect. Mol. Biol. 4: 223Ð236.
Lampo, M., D. Torgerson, L. M. Marquez, M. Rinaldi, C. Z.Garcia, and A. Arab. 1999. Occurence of sibling speciesof Lutzomyia longipalpis (Diptera: Psychodidae) in Ven-ezuela: Þrst evidence from reproductively oslated sym-patric populations. Am. J. Trop.Med.Hyg. 61: 1004Ð1009.
Lanzaro, G. C., K. Ostrovska, M. Herrero, P. G. Lawyer, andA. Warburg. 1993. Lutzomyia longipalpis is a speciescomplex: Genetic divergence and interspeciÞc hybridsterility among three populations. Am. J. Trop.Med.Hyg.48: 839Ð847.
Lanzaro,G.C., andA.Warburg. 1995. Genetic variability inphlebotomine sandßies: possible implications for leish-maniasis epidemiology. Parasitol. Today 11: 151Ð154.
Lanzaro,G.C., B. Alexander, J. P.Mutebi, J.Montoya-LermaJ. Montoya-Lerma, and Warburg, A. 1997. Genetic vari-ation amongnatural and laboratory colonypopulations ofLutzomyia longipalpis from Colombia. Mem. OswaldoCruz. 93: 65Ð70.
Mukhopadhyay, J. K., K. Ghosh, A. Azevedo, E. Rangel, andL. E. Munstermann. 1998. Genetic polymorphism ofmorphological andbiochemical characters inNatal Brazilpopulations of Lutzomyia longipalpis (Diptera: Psychod-idae). J. Am. Mosq. Cont. Assoc. 14: 277Ð282.
Munstermann, L.E.M., A. C. Morrison, C. Ferro, R. Pardo,and M. Torres. 1998. Genetic structure of local popula-tions of Lutzomyia longipalpis (Diptera: Psychodidae) inCentral Colombia. J. Med. Entomol. 35: 82Ð89.
Mutebi, J. P., E. Rowton, M. Herrero, et al. 1998. Geneticvariability among populations of the sandßy Lutzomyia(Lutzomyia) longipalpis (Diptera: Psychodidae) fromCentral America. J. Med. Entomol. 35(2): 169Ð174.
Mutebi, J. P., B. Alexander, I. Sherlock, et al. 1999. Breedingstructure of the sand ßy Lutzomyia longipalpis (Lutz &Neiva) in Brazil. Am. J. Trop. Med. Hyg. 61: 149Ð157.
Nei, M. 1978. Estimation of average heterozygosity and ge-netic distance from a small number of individuals. Ge-netics 89: 583Ð590.
Ponce, C., E. Ponce, A. Morrison, et al. 1991. Leishmaniadonovani chagasi: new clinical variant of cutaneous leish-maniasis in Honduras. Lancet 33: 67Ð70.
Saito, N., and M. Nei. 1997. The neighbor-joining method anew method for reconstructing phylogenetic trees. Mol.Bio. Evol. 4: 406Ð425.
Simon, C., F. Frati, A. Beckenbach, B. Crespi, H. Liu, and P.Flook. 1994. Evolution weighting and phylogenetic util-ity of mitochondrial gene sequences and compilation ofconserved polymerase chain reaction primers. Ann. En-tomol. Soc. Am. 87: 651Ð700.
Sneath, P.H.A., and R. R. Sokal. 1973. Numerical taxonomy.W.H.Freeman, SanFrancisco. Steiner,W.W.M., andD. J.Joslyn. 1979. Electrophoretic techniques for the geneticstudy of mosquitoes. Mosq. News 39: 35Ð54.
Swofford, D. L. 1999. Phylogenetic Analysis Using Parsimony(PAUP), version 4.0b. 2. Sinauer, Inc. Suderland, MA.
Swofford,D.L., andR.B. Selander. 1989. BIOSYS-1.A com-puter program for the analysis of allelic variation in pop-ulationgenetics andbiochemical systematics. IllinoisNat-ural History Survey, Urbana, IL.
Warburg, A., E. Saravia, G. C. Lanzaro, R. G. Titus, and F.Neva. 1994. Saliva of Lutzomyia longipalpis sibling spe-cies differs in its composition and capacity to enhanceleishmaniasis. Phil. Trans. R. Soc. Lond. B. 345: 223Ð270.
Ward, R. D., A. L. Ribeiro, P. D. Ready, and A. Murtagh.1983. Reproductive isolation between different forms ofLutzomyia longipalpis (Lutz & Neiva) (Diptera: Psy-chodidae), the vector of Leishmania donovani chagasiCunha & Chagas and its signiÞcance to kala-azar distri-bution in South America. Mem. Inst. Oswaldo Cruz. 78:269Ð280.
Ward, R. D., A. Phillips, B. Burnet, and C. Marcondes. 1988.TheLutzomyia longipalpis complex reproduction anddis-tribution, pp. 257Ð269. In Biosystematics of Haematopha-gous Insects. Service. M. W., 37th edition. ClarendonPress, Oxford, England.
World Health Organization. 1990. Leishmaniasis Control.Technical report series No. 793. WHO. Geneva.
Williams, P. 1999. Patterns in the geographical distributionof members of the genus Lutzomyia Franca (Diptera:Psychodidae-Phlebotominae),pp. 455Ð502. In J.F.Burger[ed.], Contributions to the Knowledge of Diptera. ACollection of Articles on Diptera Commemorating theLife and Work of Graham B. Fairchild. Mem. Entomol.,International 14, Associated Publishers, Gainesville, FL.
Yin, H., Norris, D. E., and Lanzaro, G. C. 2000. Siblingspecies in the Lutzomyia longipalpis complex differ inlevels of mRNA expression for the salivary peptide, max-adilan. Insect Mol. Biol. 9: 309Ð314.
Young, D. M., and M. A. Duncan. 1994. Guide to the iden-tiÞcation and geographic distribution of Lutzomyia sand-ßies in Mexico, The West Indies, Central and SouthAmerica (Diptera: Psychodidae) Mem Am. Entomol.Inst. No. 54. Associated Publishers, Gainesville. FL.
Zeledon, R., H. Hidalgo, A. Viquez, and A. Urbina. 1989.Atypical cutaneous leishmaniasis in a semiarid region ofnorth-west Costa Rica. Trans. R. Soc. Trop.Med. Hyg. 83:789.
Received for publication 5 September 2001; accepted 5 De-cember 2002.
September 2003 ARRIVILLAGA ET AL.: MOLECULAR-BIOCHEMICAL ANALYSIS OF THE Lu. longipalpis 627
Related Documents











