LYMPHOKINES IN RHEUMATOID SYNOVITIS P. Stastny, M. Rosenthal, M, Andreis, D. Cooke, and M. Ziff Department of Internal Medicine The University of Texas Health Science Center at Dallas Dallas, Texas 75235 The rheumatoid synovial membrane exhibits many indications of the pres- ence of a local immune response. It is heavily populated by lymphocytes, plasma cells, and macrophagesl and has been shown to actively synthesize immunoglobulins.2 The cells that congregate in perivascular collections and frequently develop structures that resemble lymphoid follicles include, in certain regions, numerous plasma cells and, in others, lymphocytes undergoing blastic transformation in association with collections of macrophages, which suggest the morphologic appearance of a reaction of cellular hypersensitivity in the rheumatoid process.:' Also, lymphocytes with surface markers characteristic of T cells4 and with cytotoxic activity for tissue culture monolayer cells,j, in addition to lymphocytes that bear surface immunoglobulin, have been identi- fied in the rheumatoid joint. Thus, it appears that both the antibody-forming system and the machinery of cellular hypersensitivity are activated in the rheumatoid synovium. The studies to be described were conducted to further investigate the participation of cell-mediated immunity in chronic synovitis, both in rabbits with experimental antigen-induced arthritis and in humans with spontaneous rheumatoid disease. The processes of cellular immunity are known to participate in a variety of immune phenomena. They are the basis for the development of chronic in- flammatory (delayed hypersensitivity) reactions in the skin 5 and in other sites, such as the eye* or the bladder.$' The participation of cellular immunity in host resistance to certain infections,l" allograft rejection,ll and tumor im- munity l2 is well established. Perhaps the most striking role of the lymphocytes of the thymus-derived system is their participation in the regulation and modula- tion of the immune response. T cells may enhance the potential for antibody production by their helper effects l3 and also reduce the magnitude of the immune response when they act as suppressor cells.14 Both the differentiation of lymphocytes along different pathways and the expression of effector functions of cell-mediated immunity appear to be mediated by substances produced by sensitized lymphocytes that have been called l y m p h ~ k i n e s . ~ ~ These products include factors that lead to the accumulation and stimulation of macrophages and factors that act on lymphocytes to cause them to proliferate and differen- tiate along various effector pathways. Lymphokine-like materials, which include macrophage migration inhibitory factor, lymphocyte blastogenic factor,lG and immunoglobulin synthesis-enhancing factors,'? have been found in synovial fluids and in supernatants of explants of inflamed synovial tissues in human patients with arthritis. In experimental animals, a synovitis was produced by injection of lymphokine-rich lymphocyte supernatants into normal joints.'\ In the present studies, lymphokine-like mate- rials were found in the joints of most patients with rheumatoid arthritis, in some patients with other forms of chronic inflammatory arthritis, but only rarely in joints from patients with degenerative joint disease. 117

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

LYMPHOKINES IN RHEUMATOID SYNOVITIS
P. Stastny, M. Rosenthal, M, Andreis, D. Cooke, and M. Ziff
Department of Internal Medicine The University of Texas Health Science Center at Dallas
Dallas, Texas 75235
The rheumatoid synovial membrane exhibits many indications of the pres- ence of a local immune response. It is heavily populated by lymphocytes, plasma cells, and macrophagesl and has been shown to actively synthesize immunoglobulins.2 The cells that congregate in perivascular collections and frequently develop structures that resemble lymphoid follicles include, in certain regions, numerous plasma cells and, in others, lymphocytes undergoing blastic transformation in association with collections of macrophages, which suggest the morphologic appearance of a reaction of cellular hypersensitivity in the rheumatoid process.:' Also, lymphocytes with surface markers characteristic of T cells4 and with cytotoxic activity for tissue culture monolayer cells,j, in addition to lymphocytes that bear surface immunoglobulin, have been identi- fied in the rheumatoid joint. Thus, it appears that both the antibody-forming system and the machinery of cellular hypersensitivity are activated in the rheumatoid synovium. The studies to be described were conducted to further investigate the participation of cell-mediated immunity in chronic synovitis, both in rabbits with experimental antigen-induced arthritis and in humans with spontaneous rheumatoid disease.
The processes of cellular immunity are known to participate in a variety of immune phenomena. They are the basis for the development of chronic in- flammatory (delayed hypersensitivity) reactions in the skin 5 and in other sites, such as the eye* or the bladder.$' The participation of cellular immunity in host resistance to certain infections,l" allograft rejection,ll and tumor im- munity l2 is well established. Perhaps the most striking role of the lymphocytes of the thymus-derived system is their participation in the regulation and modula- tion of the immune response. T cells may enhance the potential for antibody production by their helper effects l3 and also reduce the magnitude of the immune response when they act as suppressor cells.14 Both the differentiation of lymphocytes along different pathways and the expression of effector functions of cell-mediated immunity appear to be mediated by substances produced by sensitized lymphocytes that have been called l y m p h ~ k i n e s . ~ ~ These products include factors that lead to the accumulation and stimulation of macrophages and factors that act on lymphocytes to cause them to proliferate and differen- tiate along various effector pathways.
Lymphokine-like materials, which include macrophage migration inhibitory factor, lymphocyte blastogenic factor,lG and immunoglobulin synthesis-enhancing factors,'? have been found in synovial fluids and in supernatants of explants of inflamed synovial tissues in human patients with arthritis. In experimental animals, a synovitis was produced by injection of lymphokine-rich lymphocyte supernatants into normal joints.'\ In the present studies, lymphokine-like mate- rials were found in the joints of most patients with rheumatoid arthritis, in some patients with other forms of chronic inflammatory arthritis, but only rarely in joints from patients with degenerative joint disease.
117

118 Annals New York Academy of Sciences
MATERIALS AND METHODS
Antigeri-lfidrtced Arthritis ir i Rabbits
New Zealand white rabbits of either sex that weighed between 2.5 and 3.0 kg were used in these experiments.'!' They were immunized with 5 mg of antigen (egg albumin or bovine serum albumin) in complete Freund's adjuvant and were subsequently challenged intraarticularly with 2.5 mg of the respective antigen in 0.5 ml of sterile pyrogen-free saline. Synovial tissues were obtained for study 2, 4, and 6 weeks after challenge (FIGURE 1) .
Preparation of Lyr,iphokirta-Ricli Lymphocyte Supernatririts
Rabbits were immunized as described by Andreis et d." with 3.2 mg ot keyhole limpet hemocyanin ( K L H ) in complete Freund's adjuvant. Lymph node cells, obtained 10-40 days after immunization, were cultured at a con- centration of 20 X 10'' cells/ml in tissue culture medium 199 (TC 199) that contained 5% heat-inactivated rabbit serum and 2 mg/ml KLH. They were incubated for 48 hr at 37" C in 5% CO,, 95% air. At the end of culture, supernatants were harvested, and K L H was removed by solid immunoabsorption with rabbit anti-KLH antibody coupled to Agar0seo.l' Control supernatants were produced by culturing lymph node cells without antigen, adding antigen at the end of culture after removal of the lymphocytes, and then processing to remove the K L H by immunoabsorption (FIGURE 2 ) .
Synovitd Fluids trrrd Tissues f r o t ~ Pntients
Synovial fluids were obtained from 37 patients with classic or definite rheumatoid arthritis ( R A ) , 18 patients with osteoarthritis ( O A ) , and 15 patients with other arthritides, which included gout (4) , Reiter's syndrome (4), gonococcal arthritis (3 ) , villonodular synovitis (2) , psoriatic arthritis ( l ) , and ankylosing spondylitis ( 1 ) . The fluids were centrifuged to remove cells and debris (FIGURE 3 ) and dialyzed against phosphate-buffered saline. The hy- aluronate was then removed either by acidification to pH 5 and removal of the
FIGURE 1. Production of MIF by rabbit synovial tissue. Specimens of synovium were obtained 2, 4, and 6 weeks after intraarticular injection of antigen into animals previously im- munized with the same antigen in complete Freund's adjuvant.

Stastny er al.: Lymphokines & Rheumatoid Synovitis 119
FIGURE 2 . Production of active and control supernatants for the study of lympho- kines.
resultant mucin precipitate or by treatment with hyaluronidase (0.2 mg/ml for 2 hr at 37" C). They were then centrifuged at 15,000 g to remove debris, and the clear supernatants were filtered through a Sephadexa or agar gel column, as will be described, to test the fractions for lyniphokine activity.
Tissues were obtained during surgical synovectomy from 14 patients with classic or definite RA. The specimens were cut into 2-3-mm pieces, and samples of about 400 mg wet weight were cultured in 2 ml of TC 199 with Earle's salts, buffered with 20 mM Hepes buffer and 20 mM sodium bicarbonate. which also contained penicillin, streptomycin, mycostatin, and 15% fetal calf serum. Supernatants were harvested after 48 hr of culture at 37" C in a humidi- fied atmosphere of 5 % CO,, 9 5 % air.
Chrornatogrciphic Procedures
Five-milliliter volumes of synovial fluid were placed on 2.5 X 92.5-cm columns of Sephadex G-200, Sephadex G-100, or Agarose ASM. Elution was performed with 0.1 M tris, 0.2 M sodium chloride buffer. Fractions of 2 ml were collected after reading the optical density profile at 280 nm. The fractions that comprised each of the protein peaks and buffer controls were pooled and dialyzed against distilled water in the cold for 72 hr and lyophilized. Markers of known molecular weight were utilized to estimate the size of materials eluted from the columns. They were IgM ( M W 900,000), IgG ( M W 150,000), serum albumin (MW 68,000), ovalbumin ( M W 45,000), and chymotrypsinogen ( M W 23,000). Prior to testing for lymphokine activity, the lyophilized speci- mens were reconstituted in tissue culture medium with 15% fetal calf serum to the original volume of joint fluid placed on the cotumn.

120 Annals New York Academy of Sciences
FIGURE 3 . Methods used for the study of lymphokines in human syno- vial fluids.
Assay for Macrophage Migration Inhibition
Synovial fluid fractions or culture supernatants and their respective medium controls were made up in TC 199 and 15% fetal calf serum and were sterilized by passage through a 0.22 pm Millipores filter. Macrophages were obtained from the peritoneum of Hartley strain guinea pigs 72 hr after injection of light mineral oil. The exudate cells were washed once and suspended to a concen- tration of 10% by volume in TC 199 and 15% fetal calf serum. Glass capil- laries were filled, heat sealed at one end, and centrifuged at 200 g for 3 min. Each capillary was then cut at the edge of the packed cells and placed in Sykes-Moore culture chambers (Bellco Glass Inc.) where they were held in place by a small amount of sterile silicone. The chambers were filled with samples and control fluids and incubated at 37" C for 48 hr. Migration areas were determined at 24 and 48 hr by tracing their outline on a microscope projection screen (Hudson Photographic Industries) and measured by pla- nimetry. The effect of test samples on macrophage migration was determined by comparison with simultaneously prepared medium controls and was calcu- iated as follows:
mean migration of test sample mean migration of control
% macrophage migration = x 100.
Assay f o r Lymphocyte Blastogenic Factor
Normal Balb/C mouse spleens were used to prepare suspensions of indi- cator lymphocytes. These were washed once and suspended in medium RPMI 1640 (Gibco) without serum, which contained penicillin and streptomycin, at a concentration adjusted to give 10 x loc. nucleated spleen cells in 1.5 ml. To 1.5 ml of cell suspension was then added 0.5 ml of synovial fluid diluted 1:50 in RPMI 1640 medium, chromatographic fractions, synovial membrane cdture

Stastny et a l . : Lymphokines & Rheumatoid Synovitis 121
supernatants, or similarly diluted medium controls (FIGURE 4). Cultures were performed in triplicate, each in a total volume of 2 ml, and incubated in 15 ml disposable plastic tubes (Falcon Plastics) for 48 hr at 37" C in 5 % CO,, 95% air. After 48 hr, 1.0 pCi of tritiated thymidine, specific activity 2 Ci/mM was added to each tube and allowed to incubate further for 24 hr. At the end of culture, the cells were collected on glass fiber filters ( Reeve-Angel, no. 934AH) and washed with 20 ml saline, followed by 10 ml of 5% trichloroacetic acid (TCA) and 10 ml of methanol. The filters were dried in an oven and counted with Aquasol scintillation fluid (New England Nuclear).
Assay f o r B-Cell-Stimulating Factor
The effect of synovial fluids, synovial tissue supernatants, or their fractions on immunoglobulin synthesis by human peripheral blood lymphocytes was determined by measuring the incorporation of "C-labeled amino acids into newly synthesized immunoglobulins I 7 n by a method developed by Jasin et aL2"I *l Peripheral blood lymphocytes from either normal donors or from patients with classic or definite RA were used as indicator cells.17, lia They were cultured with synovial fluid or with chromatographic fractions that were added to produce final concentrations of 1 :200 and 1 :40, respectively. After incubation, aliquots of supernatants were used to determine the incorporated radioactivity into total protein, measured as TCA-precipitable material and into immunoglobulin as material precipitated in slight antibody excess, with poly- valent goat antihuman y-globulin. To control for nonspecific precipitation, ovalbumin-antiovalbumin was precipitated in another aliquot of each super- natant, and the resultant counts were subtracted from the values obtained in the y-globulin precipitates.
RESULTS
Production of Macrophage Migration Inhibitory Factor in Rabbits with Experimental Arthritis
Macrophage migration inhibitory activity was observed in the supernatants of synovial membrane cultures of 19 of 25 joints from rabbits with antigen- induced experimental arthritis (TABLE 1 ) . Supernatants of cultures of synovial
J o i n t F lu id o r
Synov ia l Culture
'H-Thymidine Normal Mouse Sp leen C e l l s 10' C e l l s
Medium Control
\ FIGURE 4. Assay for lymphocyte blastogenic factor.

122 Annals New York Academy of Sciences
membranes from normal joints and saline-injected joints displayed n o migration inhibitory activity. Tissue from only one of 13 joints injected with urate crystals yielded active material. The macrophage migration inhibitory activity in synovial supernatants from antigen-induced arthritis was nondialyzable and was eluted from Sephadex G-200 with an albumin marker.
Synovitis Produced by Injection of Lymphokine-Rich Supernatants of Antigen-Stimulated Lymphocytes
In these experiments,lX injections of active and control supernatants were given three times at 48-hr intervals, and the suprapatellar pouch was washed with heparinized saline to recover leukocytes 2 or 5 days after the last injection. Injection of lymphokine-rich supernatants into normal rabbit knee joints pro- duced marked exudation of leukocytes into the joint cavity ( FIGURE 5). Upon
TABLE 1
PRODUCTION OF MACROPHACE MIGRATION INHIBITORY FACTOR BY SYNOVIAL TISSUE FROM RABBITS WITH EXPERIMENTAL ARTHRITIS
Type of Synovitis
No. of Experiments
% Migration Inhibition
(mean & SE )
Noninjected Saline Urate ::: Antigen 'i
10 6
13 25
1.3k0.7 1.320.9 3 .8e3.2
3 1.7? 4.3
'k Urate controls received 5.0 mg of sterile monosodium urate crystals 12-30 hr
i Tissues were obtained 2-6 weeks after intraarticular antigen challenge in rabbits before removal of joint tissues.
immunized with the same antigen in complete Freund's adjuvant in the footpads.
examination of stained smears, it was found that the exudate consisted almost entirely of mononuclear cells.
The synovial membrane of joints injected three times at 48-hr intervals with lymphokine-rich lymphocyte supernatant exhibited hyperplasia of the lining layer and infiltration of the sublining layer with large mononuclear cells, mainly macrophages and fibroblasts. These changes are illustrated in FIGURE 6. The appearance of infiltration macrophages and fibroblasts as seen in the electron microscope is shown in FIGURE 7.
Macrophage Migration Inhibitory Factors in Human Synovial Fluids and Synovial Membrane Cultures
When rheumatoid synovial fluids were fractionated on Sephadex G-200, macrophage migration inhibitory activity eluted with the albumin marker. Such activity was observed in 18 of 22 patients with RA. It was observed in four
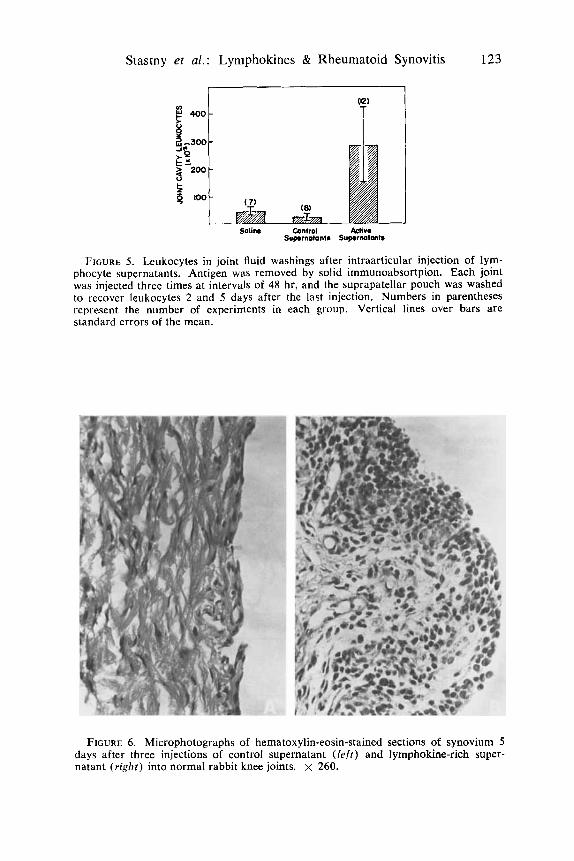
Stastny et al.: Lymphokines & Rheumatoid Synovitis 123
cn) 400 T
Salin Control M V . Suprnotantr Sup.mnatonIm
FIGURE 5. Leukocytes in joint fluid washings after intraarticular injection of lym- phocyte supernatants. Antigen was removed by solid immunoabsortpion. Each joint was injected three times at intervals of 48 hr, and the suprapatellar pouch was washed to recover leukocytes 2 and 5 days after the last injection. Numbers in parentheses represent the number of experiments in each group. Vertical lines over bars are standard errors of the mean.
FIGURE 6 . Microphotographs of hematoxylin-eosin-stained sections of synovium 5 days after three injections of control supernatant ( l e f t ) and lymphokine-rich super- natant ( r i g h t ) into normal rabbit knee joints. x 260.
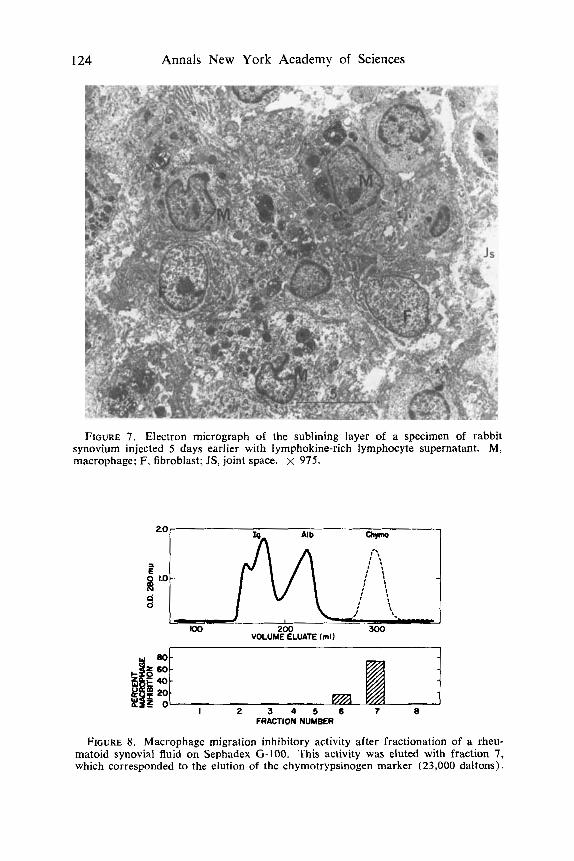
124 Annals New York Academy of Sciences
FIGURE 7. Electron micrograph of the sublining layer of a specimen of rabbit synovium injected 5 days earlier with lymphokine-rich lymphocyte supernatant. M, macrophage; F, fibroblast; JS, joint space. X 975.
100
I 2 3 4 5 6 7 e FRACTION NUMBER
FIGURE 8. Macrophage migration inhibitory activity after fractionation of a rheu- matoid synovial fluid on Sephadex G-100. This activity was eluted with fraction 7, which corresponded to the elution of the chyrnotrypsinogen marker (23,000 daltons).

Stastny et at.: Lymphokines & Rheumatoid Synovitis 125
TABLE 2
FROM SYNOVIAL FLUIDS MACROPHAGE MIGRATION INHIBITORY ACTIVITY OF SEPHADEX (3-200 FRACTION 111
% Macrophage No. with No. Migration Migration
Diagnosis Studied (meankSE) < 80% of Control
Rheumatoid arthritis 22 47+4 18
Osteoarthritis 15 85&6 3
Various inflammatory
p < 0.001
NS
arthritides 11 83'7 4
of 1 1 patients with various inflammatory arthritides and in three of 15 patients with OA (TABLE 2 ) .
When synovial fluids from patients with RA were fractionated on Sephadex G-100, the macrophage migration inhibitory activity eluted in fraction 7, which corresponds to the elution of the chymotrypsinogen marker (FIGURE 8 ) . Thus, the migration inhibitory factor in rheumatoid synovial fluid was eluted similarly to MIF produced from human peripheral blood lymphocytes stimulated with streptokinase-streptodornase (SKSD) , as reported by Rocklin et aLZ2
To determine whether immune complexes, known to exist in some rheuma- toid synovial fluids, could inhibit macrophage migration, in separate experiments rheumatoid synovial fluids were fractionated on Agarose ASM. Each of five such fluids displayed migration inhibitory activity in the region of elution of serum albumin. In addition, in two of these experiments, migration inhibitory activity was present in peak 2 , which corresponds to the position of elution of IgM (FIGURE 9) . Thus, it is possible that inhibition of migration of macro- phages in some rheumatoid joint fluids might in part be due to the presence of immune complexes.
Synovial tissues obtained at surgical synovectomy were cultured for 48 hr,
FIGURE 9. Macrophage migration inhibitory activity after fractionation of a rheumatoid synovial fluid on agarose A5m. This activity was eluted with fraction 7, which corresponded to
140 220 300 the elution of serum albumin, and w, ii 100 1.0L2!J VOLUME ELUATE (ml) with fraction 2, which contains mate-
d
rials of the size of IgM. &f g ggm 40 as 20 I
I 2 3 4 5 6 7 FRACTION NUMBER

126 Annals New York Academy of Sciences
TABLE 3 BLASTOGENIC FACTOR I N SYNOVIAL FLUIDS
Synovial [''HITdR Incorporation Fluid WBC (cpm/lO' mouse Stimulation
Patient Diagnosis ( x lO-'/rnm'') spleen cells) lndex
Medium
E.S. F.T. B.M. T.V. L.B. L.J. N.M.
control - RA + I< A RA RA OA t OA OA
- 11.9 36.0 20.6 10.8
1 . 1 0.3 0.3
584 4,743
18,58 1 10,656 12,500
3 64 2,0 1 n 1,098
- 8.0
31.8 18.2 21.4 0.6 3.4 I .9
:* Stimulation index=cpm with synovial fluid/cpm control. + RA, rheumatoid arthritis; OA, osteoarthritis.
and supernatants were dialyzed and tested for migration inhibitory activity. Nine of 11 specimens were found to have macrophage migration inhibitory activity. In four of them, the activity was associated with the third protein peak eluted from Sephadex G-200.
Blastogenic Factor in Synovial Fluids cind Synovial Culture Supernatants
Normal mouse spleen cells were cultured with synovial effusions from 19 patients with RA. Blastogenic activity, measured by an increase in tritiated thymidine incorporation, was observed in 18 of 19 rheumatoid effusions and in two of 10 nonrheumatoid fluids. Typical results of such experiments are summarized in TABLE 3 . After fractionation on Sephadex G-200, the lympho- cyte blastogenic activity was associated with the third protein peak. Similarly, culture supernatants of three rheumatoid synovial tissue specimens were found to have lymphocyte blastogenic activity. This was also eluted with the third protein peak from Sephadex G-200 (FIGURE 10).
FIGURE 10. Blastogenic activity after fraction of rheumatoid synovial fluid on Sephadex G-200. Blastogenic activity eluted with the fraction that corresponded to the elution of the serum albumin marker.

Stastny et al. : Lymphokines & Rheumatoid Synovitis 127
B-Cell-Stimulating Factor in Synovial Fluids and Synovial Cultcrre Supernatants
Human peripheral blood lymphocytes were used as targets for the assay of B-cell-stimulating activity, which was determined by measuring incorporation of 14C-labeled amino acids into newly synthesized immunoglobulins.17" En- hancement of immunoglobulin synthesis to more than twofold the corresponding control value was observed in seven of 10 different rheumatoid synovial fluids tested. Two of four synovial fluids from patients with inflammatory arthritis other than RA exhibited a similar enhancement of immunoglobulin synthesis. None of seven fluids from patients with OA showed this effect. In other experi- ments, two culture supernatants of rheumatoid synovial tissues were tested for their effect on immunoglobulin synthesis. They were also found to enhance "C-labeled amino acid incorporation into Ig, 2.5- and 2.6-fold over the control values.
DISCUSSION
Studies were undertaken to determine whether lymphokines or mediators of cellular immunity play a role in chronic synovitis. In onc group of experi- ments, a macrophage migration inhibitory factor was found in cultures of synovial tissues from rabbits with antigen-induced experimental synovitis. The inflammatory reaction in the synovial membrane in this experimental model is known to depend in part on the cellular immune zi In this model, sensitized lymphocytes are stimulated by antigen injected into joints. Because lymphocytes from such rabbits can regularly be shown to produce migration inhibitory factor and other lymphokine products when stimulated by antigen in vitro,*' it is not surprising that such agents are also produced in vivo. In fact, lymphokine production in vivo has been demonstrated by others, in the lymph nodes of sensitized sheep injected with PPD 4.i and in extracts of delayed hypersensitivity skin reaction sites.2G
In other experiments, the possible role of lymphokines in the development of chronic synovitis was investigated. Normal rabbit joints were injected with lymphokine-rich supernatants of cultured sensitized lymph node cells stimulated by antigen. The fluids obtained were rendered antigen-free by solid immunoabsorption and then introduced into normal joints three times at 48-hr intervals. When the joints were examined several days later, an exudate of mononuclear cells was found in the joint cavity, and the tissue displayed hyperplasia of the lining layer and infiltration with large mononuclear cells in the sublining layer, which consisted mainly of macrophages and fibroblasts. Accumulation of histiocytes or macrophages is the most characteristic feature of the delayed hypersensitivity response.?' The results of these experiments suggest that in active synovitis, such changes could well be mediated by the local production and release of lymphokine materials. Bennett and Bloom 2s
have previously found that injection of lymphokine-rich supernatants into the skin of guinea pigs produced an inflammatory reaction characterized as an abbreviated delayed hypersensitivity response.2g Lymphocytes and plasma cells, which accumulate in large numbers in active antigen-induced synovitis, were quite rare in the lymphokine-induced lesions, presumably because of the absence of antigenic stimulation. It appears that collection of lymphocytes in the

128 Annals New York Academy of Sciences
synovium, as seen in antigen-induced arthritis,”’ required injection of antigen into an immunized host.
Little is known about the role of lymphokines in viva. Mediators of cellular immunity, however, have been shown to cause striking histologic changes in lymph nodes in vivo.3z Regarding their possible role in chronic synovitis, in- ferences can be drawn from activities observed in in vitro experiments with synovial fluid and synovial tissue culture supernatants. For the most part, however, most of these inferences remain speculative. Of the biologic effects of the lymphokines listed in TABLE 4, accumulation of macrophages in the
TABLE 4 BIOLOGIC EFFECTS OF SUPERNATANTS OF STIMULATED LYMPHOCYTES
Test Conditions Target Effect Reference
in Vitro
macrophages chemotaxis migration inhibition stimulation
fibroblasts cytotoxicity increased collagen formation
osteoclasts resorption of bone lymphocytes stimulation of DNA synthesis
stimulation of protein synthesis enhancement of Ig and specific
antibody production
32 1 6 4 1 45 41 34
35 this report 17 17, this report
in Vivo
knee joints hyperplasia lining layer I 18, this report accumulation of macrophages
collection of mononuclear cells in joint fluid
synovium is likely to be due to a chemotactic Stimulation of macro- phages attracted to the synovium may be of significance in activating these cells. Activated macrophages may release enzymes and other agents that may partici- pate in the injury to connective tissues observed in severe arthritis. Stimulation of fibroblasts 33 and of osteoclasts 34 could participate in fibrosis and bone erosion. Finally, the present experiments suggest that proliferation of lympho- cytes and enhancement of Ig synthesis are caused by lymphokine-like material in joint effusions. Much more work will be needed, however, to determine which of the tissue culture phenomena described are important in vivo, how they interreact, and how they relate to distinct molecules produced by lym- phocytes.
Interpretation of the in vitro demonstration of lymphokines in biologic fluids involves difficulties. Macrophage migration inhibitory factors have recently been shown to originate not only from T lymphocytes but also from B cells,36- .*l nonlymphoid cells lines in culture,”’ and polymorphonuclear leuko- c y t e ~ . ~ ~ Antigen-antibody complexes may also inhibit macrophage migrati~n.~!’ Thus, it appears that several different stimuli acting upon the macrophage can

Stastny et al.: Lymphokines & Rheumatoid Synovitis 129
elicit the same response. Some of these materials can be easily separated from each other by simple physicochemical procedures,4o whereas others may be more difficult to distinguish. The chromatographic studies performed thus far indicate that macrophage migration inhibitory activity in rheumatoid synovial fluid may be associated in part with a fraction that contains immune complexes. The major part of the activity was, however, associated with fractions smaller than immunoglobulin. It is of interst that after fractionation on Sephadex G-1 00, synovial fluid MIF eluted, like MIF derived from antigen-stimulated human lymphocytes,zz in the region of the chymotrypsinogen marker, which has a molecular weight of 23,000 daltons.
Blastogenic factor has thus far not been reported to be produced by cells other than T lymphocytes.-" Similarly, the B-cell-stimulating factor 42, 43 seems to be a T-cell product. Antigen-antibody complexes have, however, been previ- ously shown to induce blastic transformation.' In fact, after fractionation of rheumatoid synovial fluids on Agarose A5M, some of the stimulating activity was associated with heavy fractions that contained immunoglobulin complexes.li The majority of the activity, however, was associated with material of the size of serum albumin or somewhat smaller. These factors were both devoid of specificity; they stimulated DNA synthesis in mouse lymphocytes and Ig in lymphocytes from normal human donors. It would appear likely, therefore, that both blastogenic factor and the Ig synthesis stimulatory factor eluted with it are lymphokine-like agents. Which cells in the inflamed synovial membrane are the source of each of these agents is at present not known. It seems likely that they are derived in large part from activated T cells. Perhaps more im- portant is the fact that these materials exhibit identifiable biologic activities in vitro. If similar effects are exerted in vivo, it would appear likely that these factors participate in the initiation and perpetuation of chronic synovitis.
SUMMARY
In antigen-induced experimental arthritis of rabbits, a macrophage migration inhibitory factor was released from the inflamed synovial tissues. A migration inhibitory factor, blastogenic factor, and B-cell-stimulating factor were also found in human rheumatoid synovial fluids and culture supernatants of rheuma- toid tissue explants. Joint fluids from patients with inflammatory conditions other than RA sometimes also displayed these activities. OA fluids were usually inactive. At present, little is known of the origin or role in vivo of the lym- phokine-like activities observed in the joints of rheumatoid patients.
In related experiments, injection of lymphokine-rich antigen-free lymphocyte supernatants into normal rabbit knee joints produced a synovitis characterized by lining layer hyperplasia and infiltration of the sublining layer by macrophages. The lymphocytic and plasmacytic components seen in active antigen-induced synovitis were absent. It seems likely that some of the changes observed in active chronic synovitis are mediated by soluble factors of the lymphokine variety.
REFERENCES
1. NORTON, W. L. & M. ZIFF. 1966. Electron microscopic observations on the rheumatoid synovial membrane. Arthritis Rheumat. 9: 580.

130 Annals New York Academy of Sciences
2.
3.
4.
5.
6.
7 .
8.
9.
10.
1 1 .
12.
13.
14.
15.
16.
17.
SMILEY, J . D., C. SACHS & M. ZIFF. 1968. I n v i m synthesis of immunoglobulin by rheumatoid synovial membrane. J. Clin. Invest. 47: 624.
KOBAYASHI, I. & M. ZIFF. 1973. Electron microscopic studies of lymphoid cells in the rheumatoid synovial membrane. Arthritis Rheumat. 16: 471.
WINCHESTER, R. J., J . B. WINFIELD, F. SIEGAL, P. WERNET, Z. BENTWICH & H. G. KUNKEL. 1974. Analyses of lymphocytes from patients with rheumatoid arthritis and systemic lupus erythematosus: occurrence of interfering cold- reactive antilymphocyte antibodies. J. Clin. Invest. 54: 1082.
HEDBERG, H. & B. KALLEA. 1964. Studies on mononuclear cells obtained from synovial fluid of patients with different types of arthritis. Cytotoxic effect on tissue-cultured human fibroblasts. Acta Pathol. 62: 177.
MACLENNAN, I. C. M. & G . LOEWI. 1968. The effect of peripheral lymphocytes from patients with inflammatory joint disease on human target cells in vitro. Clin. Exp. Immunol. 3: 385.
BENACERRAF, B. & I. GREEN. 1969. Cellular hypersensitivity. Annu. Rev. Med. 20: 141.
SCKLOSSMAN, S. & C . A. STETSON. 1957. Vascularization of the cornea during delayed hypersensitivity reactions. J. Immunol. 79: 208.
COE, J . E. & J. D. FELDMAN. 1966. Extracutaneous delayed hypersensitivity, particularly in the guinea pig bladder. Immunology 10: 127.
MACKANESS, G. B. 1970. The monocyte in cellular immunity. Sem. Hematol. 7: 172.
RUSSEL, P. S. & A. P. MONACO. 1965. The Biology of Tissue Transplantation. Little, Brown and Company. Boston, Mass.
BERNSTEIN, I. D., D. E. THOR, B. ABAR & H. J. REPP. 1971. Tumor immunity: tumor suppression in vivo initiated by soluble products of specifically stimu- lated lymphocytes. Science 172: 729.
MILLER, J. F. A. P., A. BASTEN, J . SPRENT & C. CHEEM. 1971. Interaction be- tween lymphocytes in immune responses. Cell Immunol. 2: 469.
KATZ, D. H. & B. BENACERRAF. 1972. The regulatory influence of activated T cells on B cell responses to antigen. Advan. Immunol. 15: 2.
DUMONDE, D. C., R. A. WOLSTENCROFT, G. S. PANAYI, M. MATTHEW, J . MORLEY & W. T. HOWSON. 1969. Lymphokines: non-antibody mediators of cellular immunity generated by lymphocyte activation. Nature (London) 224: 38.
STASTNY, P., M. ROSENTHAL, M. ANDREIS & M. ZIFF. 1974. Lymphokines in the rheumatoid joint. Arthritis Rheumat. In press.
ROSENTHAL, M., P. STASTNY & M. ZIFF. 1974. Enhancement of immunoglobulin and rheumatoid factor synthesis by fractions of rheumatoid synovial fluid and rheumatoid synovial tissue culture supernatants. J. Rheumatol. l(supp1.): 30.
17a. ROSENTHAL. M.. P. STASTNY & M. ZIFF. 1975. In DreDaration. 18.
19.
20.
21.
22.
23.
24.
25.
ANDREIS, M., P: STASTNY & M. ZIFF. 1974. Expeiimental arthritis produced by injection of mediators of delayed hypersensitivity. Arthritis Rheumat. 17: 537.
STASTNY, P., T. D. COOKE & M. ZIFF. 1973. Production of a macrophage migra- tion inhibitory factor in rabbits with experimental arthritis. Clin. Exp. Im- munol. 14: 141.
JASIN, H. E. & M. ZIFF. 1969. Immunoglobulin and specific antibody synthesis in a chronic inflammatory focus: antigen-induced synovitis. J. Immunol. 102: 355.
COOKE, T. D. & H. E. JASIN. 1972. The pathogenesis of chronic inflammation in experimental antigen-induced arthritis. Arthritis Rheumat. 15: 327.
ROCKLIN, R. D., H. G. REMOLD & J . R. DAVIS. 1972. Characterization of hu- man migration inhibitory factor (MIF) for antigen-stimulated lymphocytes. Cell. Immunol. 5: 436.
DUMONDE, D. C. & L. E. GLYNN. 1962. The production of arthritis in rabbits by an immunological reaction to fibrin. Brit. J . Exp. Pathol. 43: 373.
LOEWI, G. 1968. Experimental immune inflammation in the synovial mem- brane. I. The immunological mechanism. Immunology 15: 417.
HAY, J. D., P. J. LACHMAN & Z. TRAKA. 1973. The appearance of migration in-

Stastny et al. : Lyniphokines & Rheumatoid Synovitis 131
hibition factor and a mitogen in lymph draining tuberculin reactians. Eur. J. Immunol. 3: 127.
COHEN, S., P. A. WARD, T. YOSHIDA & C. L. BUREK. 1973. Biologic activity of extracts of delayed hypersensitivity skin reaction sites. Cell. Immunol. 9: 363.
LUBAROFF, D. M. & B. H. WAKSMAN. 1968. Bone marrow as a source of cells in reactions of cellular hypersensitivity. 1. Passive transfer of tuberculin sen- sitivity in syngeneic systems. J . Exp. Med. 128: 1425.
BENNETT, B. & B. R. BLOOM. 1968. Reactions in wiwo and in tzifro produced by a soluble substance associated with delayed-type hypersensitivity. Proc. Nat. Acad. Sci. USA 59: 756.
PICK, E., J. KREJCI, K. CECH & I. L. TURK. 1969. Interaction between ‘sensitized lymphocytes’ and antigen in virro. I . The release of a skin reactive factor. Immunology 17: 741.
GLYNN, L. E. 1968. The chronicity of inflammation and its significance in rheu- matoid arthritis. Ann. Rheumat. Diseases 27: 105.
WARD, P. A,, H. G. REMOLD & J . R. DAVID. 1969. Leukotactic factor produced by sensitized lymphocytes. Science 162: 1079.
NATHAN, C . F., M. L. KARNOVSKY & J. R. DAVID. 1971. Alterations of macro- phage function by mediators from lymphocytes. J. Exp. Med. 133: 1356.
JOHNSON, R. L. & M. ZIFF. 1974. Lymphokine stimulation of collagen syn- thesis. J. Rheumatol. l(supp1.): 29(abs.).
LUBEN, R. A., G. R. MUNDY, C . L. TRUMMEL & L. G. RAISZ. 1974. Partial purification of osteoclast-activating factor from phytohemagglutinin-stimulated human leukocytes. J. Clin. Invest. 53: 1473.
KELLY, R. H., T. A. WOLSTENCROFT, D. C. DUMONDE & B. M. BALFOUR. 1972. Role of lymphocyte activation products (LAP) in cell mediated immunity. TI. Effect of lymphocyte activation products on lymph node architecture and evidence for peripheral release of LAP following antigenic stimulation. Clin. Exp. Tmmunol. 10: 49.
YOSHIDA, T., H. SONOZAKI & S. COHEN. 1973. The production of migration in- hibition factor by and T cells of the guinea pig. J. Exp. Med. 138: 784.
PAPACEORGIOU, W., L. HENLEY & P. R. GLADE. 1972. Production and charac- terization of migration inhibitory factor(s) ( M l F ) of established lymphoid and non-lymphoid cell lines. J. Tmmunol. 108: 494.
STASTNY, P. & M. ZIFF. 1970. Inhibitor of macrophage migration produced by polymorphonuclear leukocytes. I . Reticuloendothel. SOC. 7: 140.
SPITLER, L., H . HUBER & H. H. FUDENBERG. 1969. Inhibition of capillary migra- tion migration by antigen-antibody complexes. J. Immunol. 102: 404.
STASTNY, P. & M. ZIFF. 1971. Macrophage migration inhibition and cytotoxi- city in acute and chronic inflammation. In Trnmunopathology of Inflammation. B. K. Ferscher & J. C . Houck, Eds. : 66. Excerpta Medica. Amsterdam, The Netherlands.
ROCKLIN, R. D., R. P. MACDERMOTT, L. CHEN, S. F. SCHLOSSMAN & J. R. DAVID. 1974. Studies on mediator production bv highly purified human T and B lym- phocytes. J. Exp. Med. 140: 1303.
ROSENTHAL, M., P. STASTNY & M. ZIFF. 1973. Stimulation of r-globulin syn- thesis and specific antibody production by a factor released by activated lym- phocytes. J . Immunol. 111: 11 19.
GEHA, R. S., E. SCHNEEBERCER, F. S. ROSEN & E. MERLER. 1973. Interaction of human thymus derived and non-thymus derived lymphocytes in vitro. Induc- tion of proliferation and antibody synthesis in B lymphocytes by a soluble factor released from antigen stimulated T lymphoctyes. J . Exp. Med. 138: 1230.
BLOCH-SHTACHER, N., K. HIRSCHHORN & J . W. UHR. 1968. The response of lymphocytes from non-immunized humans to antigen-tntibody complexes. Clin. Exp. Tmmunol. 3: 889.
STASTNY, P. 1974. Unpublished observations.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
3 6.
37.
38.
39.
40.
41.
42.
43.
44.
45.
Related Documents


![Journal of Foot and Ankle Research - Systematic review on ......Background Foot problems are frequently identified in patients with rheumatoid arthritis (RA) [1–5]. Synovitis of](https://static.cupdf.com/doc/110x72/60cf72f68917a17d390ff64c/journal-of-foot-and-ankle-research-systematic-review-on-background-foot.jpg)

![Rheumatoid arthritis in adults: management...rheumatoid arthritis (RA) who are found to have synovitis on clinical examination.[2009] 1.1.3 Consider measuring anti-CCP antibodies in](https://static.cupdf.com/doc/110x72/60d0e92fa524842b50414dfa/rheumatoid-arthritis-in-adults-management-rheumatoid-arthritis-ra-who-are.jpg)






