Lowering Oxygen Tension Enhances the Differentiation of Mouse Embryonic Stem Cells into Neuronal Cells Paul Mondragon-Teran, Gary J. Lye, and Farlan S. Veraitch The Advanced Centre for Biochemical Engineering, Dept. of Biochemical Engineering, University College London, Torrington Place, London WC1E 7JE U.K. DOI 10.1002/btpr.248 Published online May 19, 2009 in Wiley InterScience (www.interscience.wiley.com). Embryonic stem cells (ESC) are capable of proliferating indefinitely in vitro whilst retain- ing their ability to differentiate into cells of every adult lineage. Efficient, high yield proc- esses, which direct differentiation of ESC to specific lineages, will underpin the development of cost-effective drug screening and cell therapy products. The aim of this study was to investigate whether laboratory oxygen tension currently used for the neuronal differentiation of ESC was suboptimal resulting in inefficient process yields. An adherent monolayer proto- col for the neuronal differentiation of mouse ESC (mESC) was performed in oxygen con- trolled chambers using a chemically defined media over an 8 day period of culture. When exposed to oxygen tensions more appropriate to in vivo neuronal development (2% O 2 ), there was a 34-fold increase in the yield of viable cells from the differentiation process. Low oxygen tension inhibited cell death during an early phase (48 to 96 h) and toward the end (120 to 192 h) of the process. The percentage of cells expressing neuronal markers was determined by flow cytometry, revealing a small rise in the bIII tubulin and a threefold increase in the MAP2 populations at 2% O 2 . The total increase in the yield of viable cells expressing neuronal markers was shown to be 55-fold for bIII tubulin and 114-fold for MAP2. In conclusion, this study revealed that low oxygen tension can be used to enhance the yield of neuronal cells derived from ESCs and has implications for the development of efficient, cost-effective production processes. V V C 2009 American Institute of Chemical Engi- neers Biotechnol. Prog., 25: 1480–1488, 2009 Keywords: embryonic stem cells, neuronal differentiation, stem cell bioprocessing, bIII tubulin, MAP2, oxygen tension Introduction ESC derived from the inner cell mass of the blastocyst have the potential to differentiate into any adult cell type even after extended in vitro culture. 1,2 These cells have the scope to be a source of specialized cells for pharmaceutical screening and cell-based transplantation therapies. 3–5 The successful commercialization of ESC-derived products will be highly dependent on the development of cost-effective production processes. 6 Currently, the most inefficient phase of these bioprocesses is the differentiation step, resulting in low cell numbers and purity of the target phenotype. Much interest has been focused on ESC as a source of neurons for the treatment of neurodegenerative disorders such as Parkinson’s and Huntington’s disease. 7–9 Currently, ESC neuronal differentiation protocols are undertaken at laboratory oxygen tension (21% O 2 ). However, in vivo oxygen tensions during mammalian development tend to be much lower. Oxygen tension in the uterus of several mam- mals has been investigated with values ranging from 8.7% O 2 in rabbits and hamsters to 1.5% in monkeys. 10 From week 7 to 11 postgestation, the oxygen tension in the human placental environment is 2.4% O 2 . After 11 weeks, maternal blood flows to the embryo resulting in a rise in placental oxygen tension to 8.1% O 2 . 11–14 In addition, the concentra- tion of oxygen in the adult mammal brain is low and not uniform ranging from 0.1% O 2 in the pons area to 5.3% O 2 in gray cortex. 15–17 Recent studies have focused on the effect of oxygen ten- sion on the in vitro expansion and differentiation of foetal and postnatal neural stem cells. These findings revealed that the proliferation and differentiation of human central nervous neural stem cells was enhanced at reduced (5%) oxygen ten- sions. 18 Murine neurospheres cultured at 3% O 2 exhibited a decrease in cell death, an increase in proliferative potential, extended maintenance of multipotent potential, and higher dopaminergic commitment when compared with 21% O 2 controls. 19 In addition, low oxygen tensions (3 to 5% O 2 ) increased the yield of functional dopaminergic and serotoner- gic neurons from murine neural stem cells isolated from the developing neural crest. 16,20 In this study, we investigated whether physiological oxy- gen tensions would enhance the yield and purity of neuronal cells during ESC differentiation. We utilized a chemically defined media, monolayer protocol for the neuronal differen- tiation of mESC in conjunction with a hypoxic chamber to control the oxygen tension on the growth surface. We Correspondence concerning this article should be addressed to F. S. Veraitch at [email protected]. 1480 V V C 2009 American Institute of Chemical Engineers

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Lowering Oxygen Tension Enhances the Differentiation of Mouse Embryonic
Stem Cells into Neuronal Cells
Paul Mondragon-Teran, Gary J. Lye, and Farlan S. VeraitchThe Advanced Centre for Biochemical Engineering, Dept. of Biochemical Engineering, University College London,Torrington Place, London WC1E 7JE U.K.
DOI 10.1002/btpr.248Published online May 19, 2009 in Wiley InterScience (www.interscience.wiley.com).
Embryonic stem cells (ESC) are capable of proliferating indefinitely in vitro whilst retain-ing their ability to differentiate into cells of every adult lineage. Efficient, high yield proc-esses, which direct differentiation of ESC to specific lineages, will underpin the developmentof cost-effective drug screening and cell therapy products. The aim of this study was toinvestigate whether laboratory oxygen tension currently used for the neuronal differentiationof ESC was suboptimal resulting in inefficient process yields. An adherent monolayer proto-col for the neuronal differentiation of mouse ESC (mESC) was performed in oxygen con-trolled chambers using a chemically defined media over an 8 day period of culture. Whenexposed to oxygen tensions more appropriate to in vivo neuronal development (2% O2),there was a 34-fold increase in the yield of viable cells from the differentiation process. Lowoxygen tension inhibited cell death during an early phase (48 to 96 h) and toward the end(120 to 192 h) of the process. The percentage of cells expressing neuronal markers wasdetermined by flow cytometry, revealing a small rise in the bIII tubulin and a threefoldincrease in the MAP2 populations at 2% O2. The total increase in the yield of viable cellsexpressing neuronal markers was shown to be 55-fold for bIII tubulin and 114-fold forMAP2. In conclusion, this study revealed that low oxygen tension can be used to enhancethe yield of neuronal cells derived from ESCs and has implications for the development ofefficient, cost-effective production processes. VVC 2009 American Institute of Chemical Engi-neers Biotechnol. Prog., 25: 1480–1488, 2009Keywords: embryonic stem cells, neuronal differentiation, stem cell bioprocessing, bIIItubulin, MAP2, oxygen tension
Introduction
ESC derived from the inner cell mass of the blastocysthave the potential to differentiate into any adult cell typeeven after extended in vitro culture.1,2 These cells have thescope to be a source of specialized cells for pharmaceuticalscreening and cell-based transplantation therapies.3–5 Thesuccessful commercialization of ESC-derived products willbe highly dependent on the development of cost-effectiveproduction processes.6 Currently, the most inefficient phaseof these bioprocesses is the differentiation step, resulting inlow cell numbers and purity of the target phenotype.
Much interest has been focused on ESC as a source ofneurons for the treatment of neurodegenerative disorderssuch as Parkinson’s and Huntington’s disease.7–9 Currently,ESC neuronal differentiation protocols are undertaken atlaboratory oxygen tension (�21% O2). However, in vivooxygen tensions during mammalian development tend to bemuch lower. Oxygen tension in the uterus of several mam-mals has been investigated with values ranging from 8.7%O2 in rabbits and hamsters to 1.5% in monkeys.10 Fromweek 7 to 11 postgestation, the oxygen tension in the human
placental environment is 2.4% O2. After 11 weeks, maternalblood flows to the embryo resulting in a rise in placentaloxygen tension to 8.1% O2.
11–14 In addition, the concentra-tion of oxygen in the adult mammal brain is low and notuniform ranging from 0.1% O2 in the pons area to 5.3% O2
in gray cortex.15–17
Recent studies have focused on the effect of oxygen ten-sion on the in vitro expansion and differentiation of foetaland postnatal neural stem cells. These findings revealed thatthe proliferation and differentiation of human central nervousneural stem cells was enhanced at reduced (5%) oxygen ten-sions.18 Murine neurospheres cultured at 3% O2 exhibited adecrease in cell death, an increase in proliferative potential,extended maintenance of multipotent potential, and higherdopaminergic commitment when compared with 21% O2
controls.19 In addition, low oxygen tensions (3 to 5% O2)increased the yield of functional dopaminergic and serotoner-gic neurons from murine neural stem cells isolated from thedeveloping neural crest.16,20
In this study, we investigated whether physiological oxy-gen tensions would enhance the yield and purity of neuronalcells during ESC differentiation. We utilized a chemicallydefined media, monolayer protocol for the neuronal differen-tiation of mESC in conjunction with a hypoxic chamber tocontrol the oxygen tension on the growth surface. We
Correspondence concerning this article should be addressed to F. S.Veraitch at [email protected].
1480 VVC 2009 American Institute of Chemical Engineers

decided to use 2% O2, which lies close to the placental oxy-gen tension postgestation and mid-way within the rangesreported for the adult mammalian brain.11–17 After 8 days ofdifferentiation, we found that 2% O2 enhanced the yield ofcells expressing neuron specific markers (bIII-tubulin andMAP2). These results highlight that mimicking in vivo oxy-gen tensions during early embryo development can be usedto achieve significant enhancements in the yield of neuronalcells from ESC differentiation protocols.
Materials and Methods
Undifferentiated cell culture
E14Tg2a mESC, kindly donated by Stem Cell Sciences(Cambridge, UK), were cultured in an Heraeus Hera-Cell150 incubator (Jencons-PLS, West Sussex, UK) at 37�C and5% CO2. During routine maintenance, the cells were grownon T25 Iwaki tissue culture-treated flasks (SLS, Nottingham,UK) coated with 0.1% gelatin (Sigma, Poole, UK) in thepresence of 5 mL culture media. The culture media used forthe maintenance of undifferentiated cell lines consisted ofthe following: 380 mL of Glasgow minimal essential media(GMEM) supplemented with 400 lL of 0.1 M 2-b mercapto-ethanol (both Sigma), 40 mL foetal bovine serum, 4 mLnonessential amino acids, 4 mL of 200 mM L-glutamine, and4 mL of 100 mM sodium pyruvate (all Invitrogen, Paisley,UK). The culture media was supplemented with 0.5 mL of1 � 106 units mL�1 leukaemia inhibitory factor (LIF, Milli-pore, Hertfordshire, UK) after filter sterilization. After 2days, the cells were washed with Dulbecco’s phosphate buf-fered saline without Ca2þ/Mg2þ (DPBS, Sigma), detachedwith 0.025% trypsin, 0.04% ethylenediaminetetraacetic acid(EDTA, both Invitrogen), and 0.9% chick serum (Sigma)dissolved in DPBS, quenched with fresh growth medium,centrifuged for 3 min at 300g, resuspended in growth media,and then reinoculated into a new T25 flask precoated withgelatin.
Neuronal differentiation
Neuronal differentiation was based on the methods devel-oped by Ying et al.21 After 2 days of cell culture in T25flasks, undifferentiated mESCs were washed with 5 mLDPBS without Ca2þ/Mg2þ and trypsinized for 3 min at37�C. Trypsin was quenched with GMEM complete culturemedia without LIF and the cell suspension was centrifuged,resuspended in neuronal differentiation culture media(NDIFF-RHBA, Stem Cells Sciences, Edinburgh, UK), andthen plated at 5 � 103 cells cm�2 in Iwaki tissue culture-treated 6-well plates (SLS, Nottingham, UK), which hadbeen precoated with gelatin. The total media volume perwell was 2 mL, which was renewed every 2 days over an 8day period. Neuronal differentiation was performed at twodifferent oxygen tensions: normal culture conditions 20% O2
supplemented with 5% CO2 in an Heraeus Hera-Cell 150 in-cubator and within the sealed hypoxia chamber (see belowfor description), which was gassed from a premixed cylinderof 2% O2, 5% CO2, and 93% N2 (BOC, London, UK). A 5cm Petri dish containing 3 mL of sterile water was placed inthe hypoxia chamber with the culture plates to providehumidification. Once the chamber had been sealed, it wasplaced in the incubator to maintain the temperature at 37�C.Medium exchanges were carried out in a laminar flow hoodat laboratory oxygen conditions. Viable cell concentration
and viability throughout the differentiation process weremeasured using Guava’s ViaCount assay (Guava Technolo-gies, CA), which uses two proprietary dyes to discriminatebetween viable and nonviable cells as per manufacturer’sinstructions. Samples were analyzed on a Guava EasyCyte96-well flow cytometer using ExpressPlus software to calcu-late the viable cell concentration and viability (both GuavaTechnologies).
Fabrication and operation of the hypoxic chamber
To achieve low oxygen tension culture conditions, ahypoxic chamber was fabricated. A cylindrical chamber witha 7.5 cm radius at its base and 14.5 cm high was assembledusing acrylic as the material of construction. The top of thechamber was removable and contained two ports: one forgas inlet and another for gas out. This design ensured thatthe top could be removed before placing up to 3 � 6 wellplates and a Petri dish in the chamber. The top was thentightly sealed using an o-ring sealing mechanism and eightclamping screws before directly purging the chamber withgas from a premixed cylinder.
Oxygen measurements
The oxygen tension inside the hypoxia chamber was meas-ured using the Presens Oxy-4 system in conjunction with Sp-pst3-NAU-D5-NOP oxygen sensor spots (both Presens,Regensburg, Germany). Presens’ detection principle is basedon the quenching of luminescence caused by collisionbetween molecular oxygen and oxygen sensitive luminescentdye molecules. The luminescent dye molecules are immobi-lized in the form of a spot and coated with an optical insula-tion to avoid interference from intrinsic fluorescence. Onespot was attached to the wall of the hypoxic chamber tomonitor the gas phase. A second spot was used to monitorthe oxygen tension in the liquid phase. This was placed atthe base of a 6-well plate before being submerged with 2mL of differentiation media. The Oxy-4 system and sensorspots were calibrated before each experiment in accordancewith the manufacturer’s instructions, which can be found atwww.presens.de.
Immunocytochemistry
For immunocytochemistry (ICC) analysis, NDIFF-RHBAmedia was removed and cells were washed with DPBSbefore being fixed with 4% para-formaldehyde (Sigma) for20 min at room temperature. The cells were then washedtwice with DPBS and incubated for 30 min with blockingsolution consisting of DPBS, 0.25% Triton �100, and 2%goat serum (both Sigma). Cells were incubated at room tem-perature for 60 min with the primary antibodies anti-MAP2(1:500), anti-bIII tubulin (1:400, both Sigma), and anti-Nes-tin (1:300, Millipore) diluted in blocking solution. Isotypecontrols, used at the same dilution as the primary antibodies,were mouse IgG2b for bIII tubulin and mouse IgG1 (bothSigma) for Nestin and MAP2. After incubation with the pri-mary antibodies, the cells were washed twice with DPBS(Sigma) and incubated for 60 min at room temperature withan anti-mouse IgG (Fc Specific) FITC conjugate (Sigma)diluted 1:400 in blocking solution. All cells were counter-stained with DAPI (1 lg mL�1 for 5 min) prior to imaging.Samples were analyzed after processing using an invertedfluorescent Nikon Eclipse TE2000-U microscope. The
Biotechnol. Prog., 2009, Vol. 25, No. 5 1481
number and maximum diameter of neural rosettes from 30random fields of view expressing bIII tubulin was assessedin triplicate wells using image processing analysis in Java(Image J, NIH).
Flow cytometry
On day 8, cells were stained for nestin, bIII tubulin, andMAP2 using the ICC method described above, without theinclusion of DAPI, followed by trypsination (5 min at 37�C)to obtain a single cell suspension. Cells were gently resus-pended using a Pasteur glass pipette before analysis usingflow cytometry. Cells incubated with the isotype controlswere used to gate the negative population allowing the anal-ysis of the percentage of the population expressing each ofthe markers.
Statistical analysis
Unless otherwise stated, all cell culture experiments wereconducted in triplicate. Significant differences between datapoints were calculated using a two-tailed, paired, studentt-test.
Results
Oxygen tension control
To test the ability of the hypoxic chamber to maintain adesired oxygen tension, the chamber was first purged withpremixed gas containing 2, 3, 10, 11, 18, 20, and 21% oxy-gen (data not shown). All the premixes contained 5% CO2
with nitrogen making up the remainder of the composition.The chamber was gassed at a flow rate of 7.5 L min�1 for 5min before sealing the chamber and monitoring the oxygentension on the chamber wall and at the growth surface of6-well plates submerged in 2 mL of growth medium. Aftergassing, the expected oxygen tensions were reached instanta-neously on the wall of the chamber. Equilibrium betweenthe gas phase and liquid phase took a number of hoursdepending on the concentration of oxygen in the premix. Wealso monitored the oxygen tension at the growth surface of
6-well plate cultures in normal incubators. Here, weobserved a drop from 21% to 20% upon transfer from thelaminar flow hood to the incubator. This was probably dueto the fact that air from the laboratory is mixed with CO2
(5%) in the incubator resulting in a small drop in oxygentension. These results revealed that the oxygen tension inCO2 supplemented incubators under normal conditions is�20% (19.8 � 0.2% O2) not 21% O2. Further testing of thehypoxic chamber established that the rate of media evapora-tion was comparable in the hypoxia chamber when comparedwith media in humidified incubators operating at 37�C and5% CO2 (data not shown). Figure 1 shows the oxygen ten-sion profile at the growth surface of a 6-well plate during an8-day neuronal differentiation in the hypoxic chamber main-tained at 2% O2 (2.2 � 0.2% O2). Upon inoculation, it took�5 h for the oxygen tension on the growth surface to equili-brate to 2% O2. The oxygen tension was maintained constantat 2% throughout the next 2 days indicating that differentiat-ing mESC were not consuming enough oxygen to cause asignificant drop in oxygen tension at the growth surface. Af-ter 2 day, the plates were removed from the hypoxic cham-bers to replace the medium, causing an increase in theoxygen tension to 21%. Once the medium had beenexchanged, the plates were placed back in the hypoxic cham-bers and took �5 h to equilibrate to 2% O2 at the growthsurface. These results reveal that hypoxic chambers onlymaintain the oxygen tension during the incubation phase andthat medium exchange in a normal laminar flow hood causesa rapid rise in oxygen tension at the growth surface. Theinfluence of these transient shifts in oxygen tension remainsto be determined.
Neuronal differentiation at 2% O2 increases the yieldof viable cells
Neuronal differentiation of mouse embryonic stem cellsunder normal culture conditions (20% O2) and physiologicalconditions (2% O2) was performed using the E14Tg2a cellline. Neuronal development in monolayer culture was fol-lowed by light microscopy every 48 h during the 192 hprotocol (Figure 2). These representative images show the
Figure 1. Oxygen control during neuronal differentiation of mESC at 2% O2.
The trace shows the oxygen tension at the growth substrate which was measured continuously using a precalibrated Presens spot. Every 48 h, theculture medium was renewed manually in a normal laminar flow hood.
1482 Biotechnol. Prog., 2009, Vol. 25, No. 5

gradual expansion of attached cellular aggregates under bothconditions. At 2% O2, there was an observable rise in thenumber of aggregates forming and maturing as the differen-tiation process proceeded. The viable cell concentration andviability of attached cells was assessed during the differen-tiation protocol every 24 h (Figure 3). Under both oxygentensions, the cells exhibited biphasic growth curves. Thiswas caused by a drop in viable cell concentration between48 and 96 h. Increased viable cell numbers were observedthroughout the differentiation protocol at 2% O2 when com-pared with the 20% O2 control. After 192 h of differentia-tion, there were 8.2 � 102 � 3.7 � 102 cells cm�2 at 20%O2 and 2.8 � 104 � 5.6 � 103 cells cm�2 at 2% O2. Theseresults indicate that lowering the oxygen tension to 2% gaverise to a significant (P \ 0.01) 34-fold increase in cell pro-duction. Under both conditions, the viability of the attachedcells remained above 80% throughout the differentiation.However, the differentiation protocol was associated withthe release of significant numbers of cells into suspension.This was probably caused by the stringent, serum-free,microenvironment, which results in the preferential selection
Figure 2. Representative phase contrast images taken every 2 days during the 8 day neuronal differentiation protocol at both 2% O2
and 20% O2.
Figure 3. Effect of oxygen tension on the viable cell concentra-tion (closed symbols) and viability (open symbols) ofthe attached population during neuronal differentia-tion of mESC.
Cultures were monitored every 24 h at both 2% O2 (h) and20% O2 (~). Error bars represent one standard deviation aboutthe mean of three independent data points (n ¼ 3).
Biotechnol. Prog., 2009, Vol. 25, No. 5 1483
of neuronal lineages.21 The continual release of dead cellsmeans that under control conditions (20% oxygen), therewas no net gain in the number of cells (Figure 2). The via-bility of released cells was similar for both oxygen condi-tions with a maximum value of 25% at 24 h decreasing to�3% after 96 h (Figure 4a). These results indicate that aproportion of viable cells were released or unable to attachto the growth surface during the early stages of the differen-tiation. From 72 h onward, the majority of the cells releasedwere dead. Lowering the oxygen tension did not affect theviability of the released cells, demonstrating that theincreased cell numbers observed in Figure 3 were not due toan inhibitory action on the release of viable cells. Figure 4bshows the viability for the total system (attached þ releasedcells). Although the viability under both conditions was com-parable after 24 h, exposure to 2% O2 inhibited cell deathduring the two distinct phases. After 96 h, the viability ofcells exposed to 20% O2 had dropped to 13.4 � 2.5%. Atthe same time, point a significantly higher percentage ofcells (36.0 � 1.7%) exposed to 2% O2 were viable (P \0.01). During the last 24 h of the process, there was a dropin viability under 20% O2 condition from 53.8 � 1.2% to27.7 � 1.0%. Over the same time period, the viability ofcells cultured at 2% O2 rose from 68.8 � 1.5% to 73.3 �2.7%.These results show that low oxygen tensions inhibitcell death during two distinct phases of this neuronal differ-entiation process.
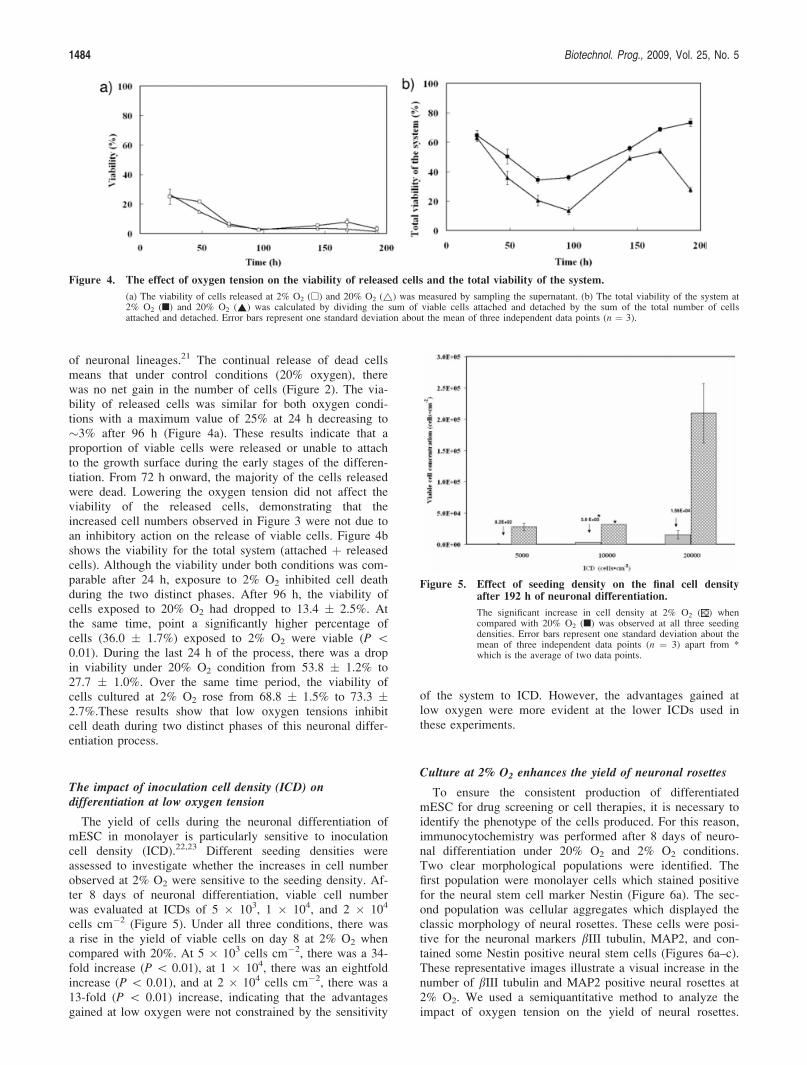
The impact of inoculation cell density (ICD) ondifferentiation at low oxygen tension
The yield of cells during the neuronal differentiation ofmESC in monolayer is particularly sensitive to inoculationcell density (ICD).22,23 Different seeding densities wereassessed to investigate whether the increases in cell numberobserved at 2% O2 were sensitive to the seeding density. Af-ter 8 days of neuronal differentiation, viable cell numberwas evaluated at ICDs of 5 � 103, 1 � 104, and 2 � 104
cells cm�2 (Figure 5). Under all three conditions, there wasa rise in the yield of viable cells on day 8 at 2% O2 whencompared with 20%. At 5 � 103 cells cm�2, there was a 34-fold increase (P \ 0.01), at 1 � 104, there was an eightfoldincrease (P \ 0.01), and at 2 � 104 cells cm�2, there was a13-fold (P \ 0.01) increase, indicating that the advantagesgained at low oxygen were not constrained by the sensitivity
of the system to ICD. However, the advantages gained atlow oxygen were more evident at the lower ICDs used inthese experiments.
Culture at 2% O2 enhances the yield of neuronal rosettes
To ensure the consistent production of differentiatedmESC for drug screening or cell therapies, it is necessary toidentify the phenotype of the cells produced. For this reason,immunocytochemistry was performed after 8 days of neuro-nal differentiation under 20% O2 and 2% O2 conditions.Two clear morphological populations were identified. Thefirst population were monolayer cells which stained positivefor the neural stem cell marker Nestin (Figure 6a). The sec-ond population was cellular aggregates which displayed theclassic morphology of neural rosettes. These cells were posi-tive for the neuronal markers bIII tubulin, MAP2, and con-tained some Nestin positive neural stem cells (Figures 6a–c).These representative images illustrate a visual increase in thenumber of bIII tubulin and MAP2 positive neural rosettes at2% O2. We used a semiquantitative method to analyze theimpact of oxygen tension on the yield of neural rosettes.
Figure 4. The effect of oxygen tension on the viability of released cells and the total viability of the system.
(a) The viability of cells released at 2% O2 (h) and 20% O2 (~) was measured by sampling the supernatant. (b) The total viability of the system at2% O2 (n) and 20% O2 (~) was calculated by dividing the sum of viable cells attached and detached by the sum of the total number of cellsattached and detached. Error bars represent one standard deviation about the mean of three independent data points (n ¼ 3).
Figure 5. Effect of seeding density on the final cell densityafter 192 h of neuronal differentiation.
The significant increase in cell density at 2% O2 ( ) whencompared with 20% O2 (n) was observed at all three seedingdensities. Error bars represent one standard deviation about themean of three independent data points (n ¼ 3) apart from *which is the average of two data points.
1484 Biotechnol. Prog., 2009, Vol. 25, No. 5
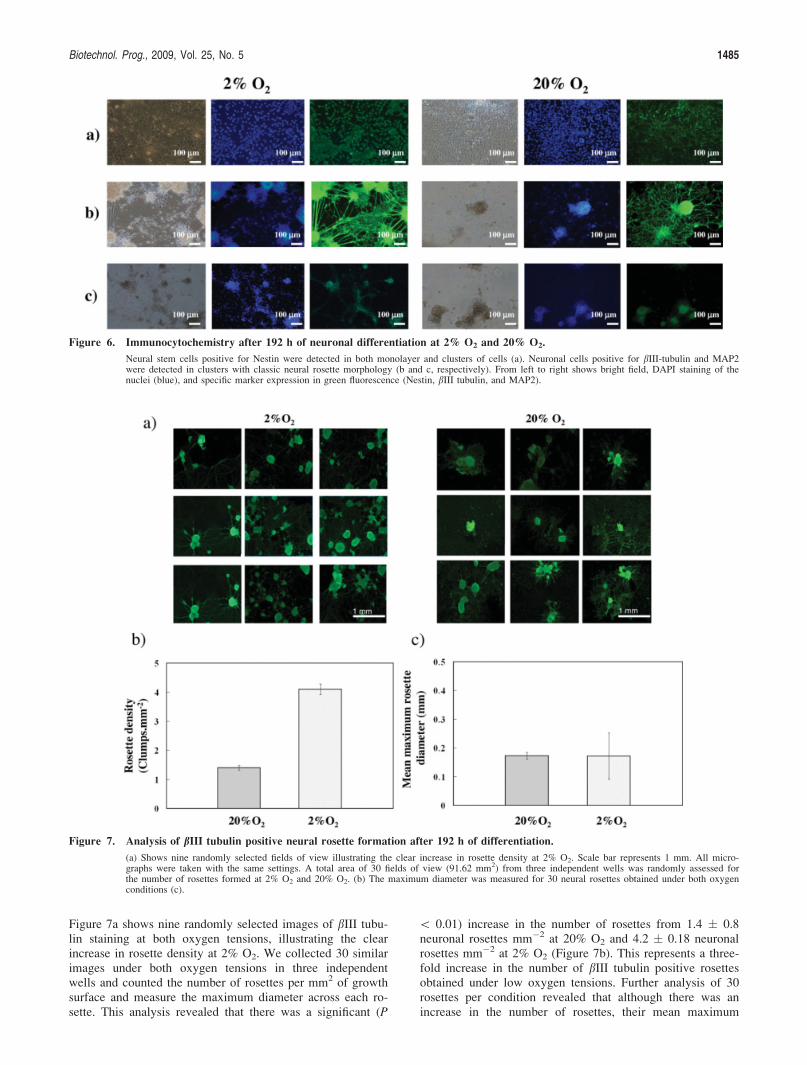
Figure 7a shows nine randomly selected images of bIII tubu-lin staining at both oxygen tensions, illustrating the clearincrease in rosette density at 2% O2. We collected 30 similarimages under both oxygen tensions in three independentwells and counted the number of rosettes per mm2 of growthsurface and measure the maximum diameter across each ro-sette. This analysis revealed that there was a significant (P
\ 0.01) increase in the number of rosettes from 1.4 � 0.8neuronal rosettes mm�2 at 20% O2 and 4.2 � 0.18 neuronalrosettes mm�2 at 2% O2 (Figure 7b). This represents a three-fold increase in the number of bIII tubulin positive rosettesobtained under low oxygen tensions. Further analysis of 30rosettes per condition revealed that although there was anincrease in the number of rosettes, their mean maximum
Figure 6. Immunocytochemistry after 192 h of neuronal differentiation at 2% O2 and 20% O2.
Neural stem cells positive for Nestin were detected in both monolayer and clusters of cells (a). Neuronal cells positive for bIII-tubulin and MAP2were detected in clusters with classic neural rosette morphology (b and c, respectively). From left to right shows bright field, DAPI staining of thenuclei (blue), and specific marker expression in green fluorescence (Nestin, bIII tubulin, and MAP2).
Figure 7. Analysis of bIII tubulin positive neural rosette formation after 192 h of differentiation.
(a) Shows nine randomly selected fields of view illustrating the clear increase in rosette density at 2% O2. Scale bar represents 1 mm. All micro-graphs were taken with the same settings. A total area of 30 fields of view (91.62 mm2) from three independent wells was randomly assessed forthe number of rosettes formed at 2% O2 and 20% O2. (b) The maximum diameter was measured for 30 neural rosettes obtained under both oxygenconditions (c).
Biotechnol. Prog., 2009, Vol. 25, No. 5 1485

diameters were comparable under both oxygen conditionswith diameter of 0.173 � 0.012 mm for 20% O2 and 0.172� 0.081 mm for 2% O2 (Figure 7c).
Enhanced yield of neuronal cells at low oxygen tensions
For a quantitative analysis of neuronal differentiation, weused flow cytometry to assess the percentage of cellsexpressing Nestin, bIII tubulin, and MAP2 after 8 days ofdifferentiation (Figure 8). There was a threefold increase inthe percentage of cells expressing MAP2 and a smaller butsignificant increase (22%, P \ 0.01) in the percentage ofcells expressing bIII tubulin when cultured at 2% O2. Lower-ing the oxygen tension caused a small drop in the percentageof cells expressing the neural stem cell marker Nestin. Toassess the impact of oxygen tension on the yield of the pro-cess, we calculated the number of cells expressing Nestin,bIII tubulin, and MAP2 on day 8 by multiplying the viablecell number with the percentage of cells expressing eachmarker. As shown in Figure 9, lowering the oxygen tensioncaused an increase in total number of neuronal cells (bIIItubulin and MAP2 positive) and neural stem cells (Nestinpositive). Monolayer neuronal differentiation at 2% O2
resulted in a 55-fold increase (P \ 0.01) in the number ofcells expressing bIII tubulin. This was higher than the three-fold increase in yield which was estimated using image anal-
ysis of rosette numbers in Figure 7. The comparable sizes ofrosettes under both conditions indicated that the density ofbIII tubulin positive cells within the rosettes may have been
Figure 8. Flow cytometry histograms showing the percentage of cells expressing the neural stem cell marker Nestin and the neuronalmarkers bIII tubulin and MAP2 at 2% O2 and 20% O2.
Errors represent one standard deviation about the mean of three independent data points (n ¼ 3) apart from bIII tubulin where n ¼ 6. AppropriateMouse IgG1 and IG2b were used as isotype controls.
Figure 9. The effect of ( ) 2% O2 and (n) 20% O2 on theyield of neural stem cells (nestin positive) and neu-rons (bIII tubulin and MAP2 positive) after 192 h ofneuronal differentiation.
These values were obtained by multiplying the total number ofcells obtained at 192 h by the percentage of Nestin, bIII tubu-lin, and MAP2 expressing cells. Error bars represent one stand-ard deviation about the mean of three independent data points(n ¼ 3).
1486 Biotechnol. Prog., 2009, Vol. 25, No. 5

much higher at low oxygen tensions. The yield analysis inFigure 8 also revealed that there was a 114-fold increase (P\ 0.01) in the number of cells expressing MAP2 at 2% O2.These results indicate that lowering the oxygen tensionenhanced the yield of cells expressing neuronal markersthrough a cumulative increase in both the yield of viablecells and in the percentage of the population expressingthese markers.
Discussion
The development of cost-effective treatments and screen-ing tools using ESC will be dependent on the developmentof efficient bioprocesses. One of the most inefficient stepscurrently is directed differentiation. Present protocols sufferfrom low yields and purity of the target phenotype. In addi-tion, the differentiation processes can be time consumingwhich may increase production costs and the risk of contam-ination. Efforts have been made to address these issues bymimicking elements of the microenvironment during embry-onic development. Operating at near-physiological oxygentensions has also been shown to modulate the differentiationof ESC into embryoid bodies, hematopoietic progenitors,cardiomyocytes, and chondrocytes.24–27 The work presentedin this study shows that the lowering oxygen tension tophysiological levels can greatly enhance the yield of neuro-nal cells from ESC differentiation processes.
Differentiation at 2% O2 had a significant impact on thenumbers of viable cells present throughout the 8 day differ-entiation process. Between 48 and 96 h, there was a drop incell number which was associated with a simultaneous risein the release of dead cells. Two percent O2 inhibited butdid not completely ablate the rise in cell death during thisphase. Toward the end of the process (168–192 h), there wasan increase in cell death resulting in a drop in cell numbersat 20% O2. Lowering the oxygen tension inhibited cell deathand coincided with an increase in the number of viable cellduring this phase. Neural stem cells (NSC) have also beenshown to be stimulated under hypoxic conditions. Morrisonet al.20 revealed that 5% O2 enhanced the ability of NSCsisolated from the sciatic nerve of E14.5 rats to survive andform multipotent colonies when plated at clonal cell den-sities. Studies using NSCs derived from E12 rat mesencepha-lon revealed that low oxygen tension inhibited cell deathduring expansion and differentiation.16 Our findings establishthat the proliferative and protective effects of low oxygentension in NSCs are also found during the directed neuronaldifferentiation of mESCs. Low oxygen tensions alsoenhanced the proliferation of human NSC,28 suggesting thatthis approach may be used to increase the yield of humanESC neuronal differentiation processes. Therefore, it will becritical to establish whether low oxygen has a beneficialeffect during the differentiation of clinically relevant humancells.
We observed a large increase in the percentage of cellsexpressing the mature neuronal marker MAP2 and a muchsmaller increase in the expression of bIII-tubulin at 2% O2.Horie et al.29 showed that low oxygen tension enhanced theproportion of bIII-tubulin cells during the differentiationof mouse neurospheres. However, astrocyte productionremained unaffected and there was a reduction in the pro-duction of gabaminergic neurons, indicating that loweringthe oxygen tension could result in the enrichment of specifictypes of neuronal cells. This may explain why we observed
significant enrichment of the MAP2 population when com-pared with bIII-tubulin at 2% O2. At both 2 and 20% O2, wedid not observe the expression of markers for astrocytes(GFAP), serotonergic neurons (serotonin), gabaminergic neu-rons (GABA), and oligodendrocytes (O4) (data not shown),although previous studies have shown that the differentiationprotocol can be extended by 6 days resulting in the expres-sion of these markers.21 Thus, it would be interesting tocontinue the differentiation protocol under low oxygen con-ditions to investigate whether specific types of specializedneurons were being enriched.
In conclusion, lowering the oxygen tension to 2% O2
increased the number and the percentage of cells expressingneuronal markers resulting in a significant enhancement inthe yield of the differentiation process. These results showthat mimicking physiological oxygen tensions during embryodevelopment will be a powerful tool for enhancing the effi-ciency, increasing the purity, and reducing the manufacturingcosts associated with ESC differentiation processes.
Acknowledgments
The authors wish to acknowledge Hazel Thomson, RuiriFriel, and Tim Allsopp (Stem Cell Sciences, Cambridge, UK)for their advice and technical support. The authors also grate-fully acknowledge the financial support from the then UKTechnology Strategy Board (TSB project No: TP/2/BP/6/I/10419). The authors like to thank the UK Joint InfrastructureFund (JIF), the Science and Research Investment Fund (SRIF),and the Gatsby Charitable Foundation for funds to establish theUCL Centre for Micro Biochemical Engineering. PMT wouldlike to thank the Mexican Science and Technology Agency(CONACyT) for the award of a doctoral fellowship.
Literature Cited
1. Evans MJ, Kaufman MH. Establishment in culture of pluripo-tential cells from mouse embryos. Nature. 1981;292:154–156.
2. Thomson JA, Itskovitz-Eldor J, Shapiro SS, Waknitz MA,Swiergiel JJ, Marshall VS, Jones JM. Embryonic stem cell linesderived from human blastocysts. Science. 1998;282:1145–1147.
3. Studer L. Stem cells with brainpower. Nat Biotechnol. 2001;19:1117–1118.
4. McNeish J. Embryonic stem cells in drug discovery. Nat RevDrug Discov. 2004;3:70–80.
5. Vats A, Tolley NS, Bishop AE, Polak JM. Embryonic stemcells and tissue engineering: delivering stem cells to the clinic.J R Soc Med. 2005;98:346–350.
6. Thomson H. Bioprocessing of stem cells for drug discovery.Trends Biotechnol. 2007;25:224–230.
7. Brustle O, McKay RD. Neuronal progenitors as tools for cellreplacement in the nervous system. Curr Opin Neurobiol.1996;6:688–695.
8. McDonald WJ, Liu X, Qu Y, Liu S, Mickey S, Turetsky D,Gottlieb DI, Choi DW. Transplanted embryonic stem cells sur-vive, differentiate and promote recovery in injured rat spinalcord. Nat Med. 1999;5:1410–1412.
9. McLeod M, Hong M, Sen A, Sadi D, Ulalia R, Behie LA,Mendez I. Transplantation of bioreactor-produced neural stemcells into the rodent brain. Cell Transplant. 2006;15:689–697.
10. Fischer B, Bavister BD. Oxygen tension in the oviduct anduterus of rhesus monkeys, hamsters and rabbits. J Reprod Fertil.1993;99:673–679.
11. Rodesch F, Simon P, Donner C, Jauniaux E. Oxygen measure-ments in endometrial and trophoblastic tissues during earlypregnancy. Obstet Gynecol. 1992;80:283–285.
12. Genbacev O, Zhou Y, Ludlow JW, Fisher SJ. Regulation ofhuman placental development by oxygen tension. Science.1997;277:1669–1672.
Biotechnol. Prog., 2009, Vol. 25, No. 5 1487

13. Jauniaux E, Watson AL, Hempstock J, Bao YP, Skepper JN,Burton GJ. Onset of maternal arterial blood flow and placentaloxidative stress a possible factor in human early pregnancy fail-ure. Am J Pathol. 2000;157:2111–2122.
14. Burton GJ, Jaunaiux E. Maternal vascularization of the humanplacenta: does the embryo develop in a hypoxic environment?Gynecol Obstet Fertil. 2001;29:503–508.
15. Andreasen A, Danscher G, Juhl S, Stoltenberg M, RevsbechNP, Jensen H, Jensen KB. Distinct differences in partial oxygenpressure at micrometer ranges in the rat hippocampal region.J Neurosci Methods. 1997;72:15–21.
16. Studer L, Csete M, Lee SH, Kabbani N, Walikonis J, Wold B,Mckay R. Enhanced proliferation, survival, and dopaminergicdifferentiation of CNS precursors in lowered oxygen. J Neuro-sci. 2000;20:7377–7383.
17. Erecinska M, Silver AI. Tissue oxygen and brain sensitivity tohypoxia. Respir Physiol. 2001;128:263–276.
18. Pistollato F, Chen H, Schwartz P, Basso G, Panchision D. Oxy-gen tension controls the expansion of human CNS precursorsand the generation of astrocytes and oligondendrocytes. MolCell Neurosci. 2007;35:424–435.
19. Milosevic J, Schwartz S, Krohn K, Poppe M, Storch A,Schwartz J. Low atmospheric oxygen avoids maturation, senes-cence and cell death of murine mesencephalic neural precursors.J Neurochem. 2005;92:718–729.
20. Morrison SJ, Csete M, Groves AK, Melega W, Wold B, Ander-son DJ. Culture in reduced levels of oxygen promotes clono-genic sympathoadrenal differentiation by isolated neural creststem cells. J Neurosci. 2000;20:7370–7376.
21. Ying QL, Stavridis M, Griffiths D, Li M, Smith A. Conversionof embryonic stem cells into neuroectodermal precursors inadherent monoculture. Nat Biotechnol. 2003;21:183–186.
22. Diogo MM, Henrique D, Cabral JM. Optimization and integra-tion of expansion and neural commitment of mouse embryonicstem cells. Biotechnol Appl Biochem. 2008;49:105–112.
23. Veraitch FS, Scott R, Wong JW, Lye GJ, Mason C. The impactof manual processing in the expansion and directed differentia-tion of embryonic stem cells. Biotechnol Bioeng. 2008;99:1216–1229.
24. Adelman DM, Maltepe E, Simon MC. Multilineage embryonichematopoiesis requires hypoxic ARNT activity. Genes Dev.1999;13:2478–2483.
25. Bauwens C, Yin T, Dang S, Peerani R, Zandstra PW. Develop-ment of a perfusion fed bioreactor for embryonic stem cell-derived cardiomyocyte generation: oxygen-mediated enhance-ment of cardiomyocyte output. Biotechnol Bioeng. 2005;90:452–461.
26. Chen L, Fink T, Ebbesen P, Zachar V. Optimized chondrogene-sis of ATCD5 cells through sequential regulation of oxygenconditions. Tissue Eng. 2006;12:559–567.
27. Cameron CM, Harding F, Hu WS, Kaufman DS. Activation ofhypoxic response in human embryonic stem cell–derived embry-oid bodies. Exp Biol Med. 2008;233:1044–1057.
28. Storch A, Paul G, Csete M, Boehm OB, Carvey PM, Kupsch A,Schwarz J. Long-term proliferation and dopaminergic differen-tiation of human mesencephalic neural precursor cells. Exp Neu-rol. 2001;170:317–325.
29. Horie N, So K, Moriya T, Kitagawa N, Tsutsumi K, Nagata I,Shinohara K. Effect of oxygen concentration on the proliferationand differentiation of mouse neural stem cells in vitro. Cell MolNeurobiol. 2008;28:833–845.
Manuscript received Oct. 28, 2008, and revision receivedMar. 10, 2009.
1488 Biotechnol. Prog., 2009, Vol. 25, No. 5
Related Documents











![STEM CELLS EMBRYONIC STEM CELLS/INDUCED PLURIPOTENT STEM CELLS Stem Cells.pdf · germ cell production [2]. Human embryonic stem cells (hESCs) offer the means to further understand](https://static.cupdf.com/doc/110x72/6014b11f8ab8967916363675/stem-cells-embryonic-stem-cellsinduced-pluripotent-stem-cells-stem-cellspdf.jpg)