Loss of Vacuolar Proton-translocating ATPase Activity in Yeast Results in Chronic Oxidative Stress * □ S Received for publication, August 30, 2006, and in revised form, December 13, 2006 Published, JBC Papers in Press, January 10, 2007, DOI 10.1074/jbc.M608293200 Elena Milgrom ‡1 , Heba Diab ‡1 , Frank Middleton § , and Patricia M. Kane ‡2 From the ‡ Departments of Biochemistry and Molecular Biology and § Neuroscience and Physiology, SUNY Upstate Medical University, Syracuse, New York 13210 Yeast mutants lacking vacuolar proton-translocating ATPase (V-ATPase) subunits (vma mutants) were sensitive to several different oxidants in a recent genomic screen (Thorpe, G. W., Fong, C. S., Alic, N., Higgins, V. J., and Dawes, I. W. (2004) Proc. Natl. Acad. Sci. U. S. A. 101, 6564 – 6569). We confirmed that mutants lacking a V 1 subunit (vma2),V o subunit, or either of the two V o a subunit isoforms are acutely sensitive to H 2 O 2 and more sensitive to menadione and diamide than wild-type cells. The vma2 mutant contains elevated levels of reactive oxygen species and high levels of oxidative protein damage even in the absence of an applied oxidant, suggesting an endogenous source of oxidative stress. vma2 mutants lacking mitochondrial DNA showed neither improved growth nor decreased sensitivity to peroxide, excluding respiration as the major source of the endogenous reactive oxygen species in the mutant. Double mutants lacking both VMA2 and components of the major cyto- solic defense systems exhibited synthetic sensitivity to H 2 O 2 . Microarray analysis comparing wild-type and vma2 mutant cells grown at pH 5, permissive conditions for the vma2 mutant, indicated high level up-regulation of several iron uptake and metabolism genes that are part of the Aft1/Aft2 regulon. TSA2, which encodes an isoform of the cytosolic thi- oredoxin peroxidase, was strongly induced, but other oxidative stress defense systems were not induced. The results indicate that V-ATPase activity helps to protect cells from endogenous oxidative stress. Both prokaryotic and eukaryotic cells have multiple mecha- nisms for preventing and counteracting the effects of oxidative stress (1). Reactive oxygen species (ROS) 3 are normal products of aerobic metabolism, but excess ROS accumulation due to perturbation of control mechanisms can result in accumulation of mutations, premature aging, and cell death (2– 4). Many of the central mechanisms for controlling redox balance in the cell are highly conserved. These mechanisms include 1) proteins involved directly in regulating redox balance or repairing effects of oxidative stress, including the well known compo- nents of the glutathione- and thioredoxin-dependent reduction systems, superoxide dismutase, catalase, and the pentose phos- phate pathway, and 2) a transcriptional response centered around recruitment of the “redox-specific” transcription fac- tors to promoters of these and other antioxidant response genes (2, 3). Only recently has the full spectrum of genes important for control of oxidative stress begun to be appreciated, through microarray analysis of responses to a variety of oxidants and screens of ordered S. cerevisiae deletion mutant arrays (5–7). Genomic screens have revealed that deletions in many genes not previously associated with redox balance result in sensitiv- ity to one or more applied oxidants (6 – 8). These screens also revealed that response to oxidative stress may be more specific than previously appreciated, since many deletion mutants show sensitivity to one type of oxidant but little sensitivity to others (7, 8). These results suggest that the cell tailors its response to different stresses rather than having a single general response pathway (9). However, certain deletion strains do show sensi- tivity to multiple different applied oxidants (7, 8). This may indicate that the deletions in these genes affect protective mechanisms that are used in multiple contexts and/or that the deletions result in a chronic state of oxidative stress that render the mutants unable to cope with further stress. Consistent with this, mutants lacking important antioxidant functions also show evidence of oxidative stress or damage even in the absence of an applied oxidative stress (10, 11). In these cases, the screens may be revealing previously unappreciated sources of oxidative stress and the cellular mechanisms that contain them. Yeast mutants lacking subunits of the vacuolar proton-trans- locating ATPase (V-ATPase) have been identified in multiple genomic screens for sensitivity to different forms of oxidative stress (6 – 8). The V-ATPase is a highly conserved proton pump responsible for acidification of organelles such as the lysosome/ vacuole, Golgi apparatus, and endosomes in all eukaryotic cells (12, 13). V-ATPases have also been recruited to a variety of more specialized functions, often involving high level expres- sion of the enzyme at the plasma membrane of polarized cells, in addition to intracellular compartments (13, 14). The yeast S. cerevisiae does not appear to express a plasma membrane form of the V-ATPase and has proven to be an excellent model for the “constitutive” functions of V-ATPases associated with acid- * This work was supported by National Institutes of Health Grant R01- GM50322 (to P. M. K.). The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. □ S The on-line version of this article (available at http://www.jbc.org) contains supplemental Fig. 1 and Tables 1 and 2. 1 These authors contributed equally to this work. 2 To whom correspondence should be addressed: Dept. of Biochemistry and Molecular Biology, SUNY Upstate Medical University, 750 E. Adams St., Syracuse, NY 13210. Tel.: 315-464-8742; Fax: 315-464-8750; E-mail: [email protected]. 3 The abbreviations used are: ROS, reactive oxygen species; V-ATPase, vacuo- lar proton-translocating ATPase; YEPD, yeast extract/peptone/dextrose medium; SC, synthetic complete medium; DHR, dihydrorhodamine 123; DNP, 2,4-dinitrophenyl; GFP, green fluorescent protein. THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 282, NO. 10, pp. 7125–7136, March 9, 2007 © 2007 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A. MARCH 9, 2007 • VOLUME 282 • NUMBER 10 JOURNAL OF BIOLOGICAL CHEMISTRY 7125

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Loss of Vacuolar Proton-translocating ATPase Activity inYeast Results in Chronic Oxidative Stress*□S
Received for publication, August 30, 2006, and in revised form, December 13, 2006 Published, JBC Papers in Press, January 10, 2007, DOI 10.1074/jbc.M608293200
Elena Milgrom‡1, Heba Diab‡1, Frank Middleton§, and Patricia M. Kane‡2
From the ‡Departments of Biochemistry and Molecular Biology and §Neuroscience and Physiology, SUNY Upstate MedicalUniversity, Syracuse, New York 13210
Yeast mutants lacking vacuolar proton-translocating ATPase(V-ATPase) subunits (vma mutants) were sensitive to severaldifferent oxidants in a recent genomic screen (Thorpe, G. W.,Fong, C. S., Alic, N., Higgins, V. J., and Dawes, I.W. (2004) Proc.Natl. Acad. Sci. U. S. A. 101, 6564–6569). We confirmed thatmutants lacking a V1 subunit (vma2�), Vo subunit, or either ofthe two Vo a subunit isoforms are acutely sensitive to H2O2 andmore sensitive to menadione and diamide than wild-type cells.The vma2� mutant contains elevated levels of reactive oxygenspecies and high levels of oxidative protein damage even in theabsence of an applied oxidant, suggesting an endogenous sourceof oxidative stress. vma2� mutants lackingmitochondrial DNAshowed neither improved growth nor decreased sensitivity toperoxide, excluding respiration as the major source of theendogenous reactive oxygen species in the mutant. Doublemutants lacking bothVMA2 and components of themajor cyto-solic defense systems exhibited synthetic sensitivity to H2O2.Microarray analysis comparing wild-type and vma2� mutantcells grown at pH 5, permissive conditions for the vma2�
mutant, indicated high level up-regulation of several ironuptake and metabolism genes that are part of the Aft1/Aft2regulon. TSA2, which encodes an isoform of the cytosolic thi-oredoxin peroxidase, was strongly induced, but other oxidativestress defense systems were not induced. The results indicatethat V-ATPase activity helps to protect cells from endogenousoxidative stress.
Both prokaryotic and eukaryotic cells have multiple mecha-nisms for preventing and counteracting the effects of oxidativestress (1). Reactive oxygen species (ROS)3 are normal productsof aerobic metabolism, but excess ROS accumulation due toperturbation of controlmechanisms can result in accumulation
of mutations, premature aging, and cell death (2–4). Many ofthe centralmechanisms for controlling redox balance in the cellare highly conserved. These mechanisms include 1) proteinsinvolved directly in regulating redox balance or repairingeffects of oxidative stress, including the well known compo-nents of the glutathione- and thioredoxin-dependent reductionsystems, superoxide dismutase, catalase, and the pentose phos-phate pathway, and 2) a transcriptional response centeredaround recruitment of the “redox-specific” transcription fac-tors to promoters of these andother antioxidant response genes(2, 3).Only recently has the full spectrum of genes important for
control of oxidative stress begun to be appreciated, throughmicroarray analysis of responses to a variety of oxidants andscreens of ordered S. cerevisiae deletion mutant arrays (5–7).Genomic screens have revealed that deletions in many genesnot previously associated with redox balance result in sensitiv-ity to one or more applied oxidants (6–8). These screens alsorevealed that response to oxidative stress may be more specificthan previously appreciated, sincemany deletionmutants showsensitivity to one type of oxidant but little sensitivity to others(7, 8). These results suggest that the cell tailors its response todifferent stresses rather than having a single general responsepathway (9). However, certain deletion strains do show sensi-tivity to multiple different applied oxidants (7, 8). This mayindicate that the deletions in these genes affect protectivemechanisms that are used in multiple contexts and/or that thedeletions result in a chronic state of oxidative stress that renderthemutants unable to cope with further stress. Consistent withthis, mutants lacking important antioxidant functions alsoshowevidence of oxidative stress or damage even in the absenceof an applied oxidative stress (10, 11). In these cases, the screensmay be revealing previously unappreciated sources of oxidativestress and the cellular mechanisms that contain them.Yeastmutants lacking subunits of the vacuolar proton-trans-
locating ATPase (V-ATPase) have been identified in multiplegenomic screens for sensitivity to different forms of oxidativestress (6–8). TheV-ATPase is a highly conserved proton pumpresponsible for acidification of organelles such as the lysosome/vacuole, Golgi apparatus, and endosomes in all eukaryotic cells(12, 13). V-ATPases have also been recruited to a variety ofmore specialized functions, often involving high level expres-sion of the enzyme at the plasma membrane of polarized cells,in addition to intracellular compartments (13, 14). The yeast S.cerevisiae does not appear to express a plasmamembrane formof the V-ATPase and has proven to be an excellent model forthe “constitutive” functions of V-ATPases associatedwith acid-
* This work was supported by National Institutes of Health Grant R01-GM50322 (to P. M. K.). The costs of publication of this article were defrayedin part by the payment of page charges. This article must therefore behereby marked “advertisement” in accordance with 18 U.S.C. Section 1734solely to indicate this fact.
□S The on-line version of this article (available at http://www.jbc.org)contains supplemental Fig. 1 and Tables 1 and 2.
1 These authors contributed equally to this work.2 To whom correspondence should be addressed: Dept. of Biochemistry
and Molecular Biology, SUNY Upstate Medical University, 750 E. AdamsSt., Syracuse, NY 13210. Tel.: 315-464-8742; Fax: 315-464-8750; E-mail:[email protected].
3 The abbreviations used are: ROS, reactive oxygen species; V-ATPase, vacuo-lar proton-translocating ATPase; YEPD, yeast extract/peptone/dextrosemedium; SC, synthetic complete medium; DHR, dihydrorhodamine 123;DNP, 2,4-dinitrophenyl; GFP, green fluorescent protein.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 282, NO. 10, pp. 7125–7136, March 9, 2007© 2007 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A.
MARCH 9, 2007 • VOLUME 282 • NUMBER 10 JOURNAL OF BIOLOGICAL CHEMISTRY 7125

ification of intracellular compartments (12). All eukaryoticV-ATPases are multisubunit enzymes comprised of a complexof peripheral membrane proteins, V1, attached to a complex ofintegral membrane proteins, Vo. Deletion of any V-ATPasesubunit gene, exceptVPH1 and STV1, which encode organelle-specific isoforms of one of the Vo subunits, results in a commonVma� phenotype, characterized by a pH-dependent condi-tional lethality and sensitivity to elevated extracellular calciumconcentrations (12). In addition, vmamutants show a variety ofother defects that are not completely understood, includingpoor growth on nonfermentable carbon sources, hypersensitiv-ity to multiple drugs, and increased sensitivity to transitionmetals, such as iron, copper, and zinc (15–17).In this work, we confirm that vmamutants are highly sensi-
tive to oxidative stress, particularly H2O2, and further explorethe basis of this sensitivity.We find that the vmamutants showevidence of chronic oxidative stress, including elevated ROSlevels and protein modifications characteristic of accumulatedoxidative damage, even in the absence of any added oxidant, butthe electron transport chain is not themajor source of the stressin these cells. vmamutations result in a synthetic sensitivity toperoxide when combined with mutations in many of the com-ponents of the cytoplasmic machinery for redox control,including the Cu,Zn-superoxide dismutase, glutathione reduc-tase, glutathione synthetase, thioredoxins, thioredoxin peroxi-dases, and components of the oxidative arm of the pentosephosphate pathway, suggesting that vacuolar acidification isnot simply involved in supporting one of these mechanismscentral to redox balance. Microarray analysis under conditionspermissive for growth of the vma2mutant (pH5) indicates highlevel up-regulation of the cytosolic thioredoxin peroxidaseTsa2p and induction of the Aft1p/Aft2p regulon in the vma2mutant. Based on these results, we suggest several possibleexplanations for the sensitivity of vmamutants to applied oxi-dative stresses, such as hydrogen peroxide.
EXPERIMENTAL PROCEDURES
Materials and Strains—Hydrogen peroxide was obtainedfrom Acros. Menadione and diamide were obtained fromSigma. Dihydrorhodamine 123 was obtained from Sigma, andthe OxyBlot kit was purchased from Chemicon International.Yeast extract/peptone/dextrose (YEPD) and synthetic com-plete (SC) were prepared as described (18).All strains were obtained from theOpenBiosystems or Euro-
fan haploid deletion collections and were based on BY4741(MATa his3�1 leu2�0met15�0 ura3�0). All strains in the col-lection have a complete replacement of the open reading framewith the kanMX marker in the BY4741 background (19). TheBY4742 vma2�::Natr strain was constructed as described (20).Double mutant spores were isolated by mating the indicatedstrain from the deletion collection with the BY4742 vma2�strain, selecting diploids on YEPD containing 200 �g/ml kana-mycin and 200 �g/ml clonNAT (Werner Biolabs) and thensporulating the resulting diploids and dissecting tetrads onYEPD plates buffered to pH 5. Haploid spores resistant to bothkanamycin and clonNAT were identified.The wild-type BY4741 and the BY4741 vma2� strain were
converted to rho0 strains by extended incubationwith 25�g/ml
ethidium bromide as described (21). Loss of mitochondrialDNA was confirmed by 4�,6-diamidino-2-phenylindolestaining.Halo assays for sensitivity to various oxidants were per-
formed as described (22), except that the indicated concen-trations of oxidant were added to the filter paper disk afterthe disk was placed on an overlay of cells grown to log phasein liquid culture, mixed with an equal volume of 1% agar, andthen layered onto a YEPD plate buffered to either pH 7 or pH5. The cells were then allowed to grow at 30 °C for 2–4 days. Foreach assay, wild-type and mutant strains were spread at thesame density and grown for identical times. Relative halo sizeswere determined bymeasuring the diameter of the area aroundthe disk thatwas clear of cells and dividing thismeasurement bythe diameter of the halo for wild-type cells treated in parallelwith the same peroxide concentration.For dilution assays to test peroxide sensitivity, hydrogen per-
oxide was added to SC to give the final concentration indicated.Plates were used within 2 days of the assay. Liquid cultures ofstrains to be tested were grown to log phase, adjusted to iden-tical density, and then serially diluted (10-fold dilutions) beforespotting on the plates.Genetic and Biochemical Tests for Oxidative Stress Sensitivity
and Oxidative Damage—Dihydrorhodamine 123 (DHR) stain-ing was performed as described by Madeo et al. (23). Briefly,DHR was dissolved in ethanol to make a 2.5-mg/ml stock. Liq-uid cultures of BY4741 and BY4741 vma2� strains and the cor-responding rho0 cells were grown overnight in SC, and DHRwas added to a final concentration of 5 �g/ml. Cells wereviewed using a Zeiss Axioskop 2 fluorescencemicroscope and arhodamine filter set after 4.5 h of staining. Images were cap-tured using a Hamamatsu CCD camera. For quantitation of thepercentage of cells stained with the dye, 280–350 cells fromeach strain were scored in two independent experiments. Sam-ples were compared using the two-sample t test assuming equalvariances.Carbonylated proteins were detected by reaction with 2,4-
dinitrophenyl (DNP) hydrazine and detection of DNP-modi-fied protein by Western blotting with rabbit anti-DNP poly-clonal antibody according to protocols provided with theOxyBlot kit. Yeast cellswere prepared as described (24)with thefollowing modifications. 5 A600 units of each strain were iso-lated during log phase growth, converted to spheroplasts, andlysed by the addition of 0.5 ml of cold 0.6 M sorbitol, 1 mMEDTA, 50 mM dithiothreitol, pH 7.0, and vortex mixing for 2min. Unlysed cells were pelleted, and the supernatant was usedfor derivatization. A mock derivatization, in which all reagentsexcept DNP-hydrazine were added to the supernatant, was car-ried out in parallel for each strain. After neutralization of thereaction, equal amounts of protein for each sample were sepa-rated by SDS-PAGE and transferred to nitrocellulose forimmunoblotting.The GFP-YAP1 plasmid was a generous gift from Scott
Moye-Rowley (25). The plasmid was transformed into wild-type and vma2� strains, and transformants were selected andmaintained on fully supplemented minimal medium lackinguracil. Localization of Yap1p was visualized after overnightgrowth of the cells in SC-uracil.
Yeast V-ATPase Protects against Oxidative Stress
7126 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 10 • MARCH 9, 2007

Microarray and Northern Analyses—Total yeast RNA wasisolated from wild-type and vma2� mutant yeast cells grownto early log phase in YEPD, pH 5, as described (26). The totalRNA obtained was then further purified using a QiagenRNeasy RNA purification kit, and then the quality and quan-tity of the RNAwas analyzed using an Agilent RNA nanochipon an Agilent 2100 Bioanalyzer. Duplicate RNA samples(from two independent isolations from the vma2� strain and asingle isolation from the wild-type strain) were subjected tomicroarray analysis using Affymetrix Yeast Gen2 chips andstandard eukaryotic one-cycle amplification and labeling pro-cedures. Arrays were scanned on a GeneChip 3000 scanner(Affymetrix) and analyzed byGeneChipOperating System soft-ware (Affymetrix) to generate present and absent calls andassess sample quality. Subsequently, the .cel format image fileswere imported into GeneSpring GX (Agilent) and normalizedusing the robust multiarray average method (27). For thisreport, we determined the fraction of genes that were increasedor decreased in the mutant versus wild type strains at the two-tailed 99th percentile confidence level (defined as �2.58 S.D.values from the mean expression ratio comparing the twostrains). In the present study, this translated to cut-off values of2.1-fold up or down in assessing changes in expression in themutant strains (see supplemental Fig. 1).For Northern analysis, two independent RNA samples were
prepared from both the wild type and vma2� mutant asdescribed above. RNA was separated on a 2.2 M formaldehydedenaturing gel, transferred by capillary action toDuralonmem-brane, and fixed to the blot by UV cross-linking (260 nm for 1min), as described by Sambrook andRussell (28). Probes againstthe indicated genes were prepared by PCR amplification of250–400-bp regions of the open reading frames of the genes,followed by labeling with [�-32P]dCTP using a StratagenePrime-It random primer labeling kit. Blots were hybridizedwith the probes as described (28) and thenwashed and analyzedon an Amersham Biosciences Typhoon PhosphorImager.
RESULTS
Yeast vma Mutants Are Acutely Sensitive to Oxidative Stress—In order to confirm that the vma mutants were sensitive toexternal oxidative stress, we challenged two of the vma deletionstrains, vma2� and vma3�, with several concentrations ofH2O2. These two mutations affect the two different sectors ofthe V-ATPase; vma2� lacks the B subunit of the V1 sector butcontains assembled Vo sectors at the vacuole, and vma3� lacksthe c subunit of theVo sector and contains assembledV1 sectorsin the cytosol (29). Fig. 1A shows a halo assay for wild type,vma2�, and vma3� in which varied H2O2 concentrations wereapplied to filter disks on top of a fresh overlay of cells, and thesize of the area of inhibited growth around the disk provides ameasure of sensitivity (a larger halo indicates sensitivity to alower concentration of H2O2 as it diffuses from the filter). It isclear from Fig. 1A that both the vma2� and vma3� mutantsexhibit much larger halos than the wild-type control strain atevery peroxide concentration tested, suggesting that themutant strains are very sensitive to peroxide stress. In Fig. 1B,the sensitivity of the vma2� strain to two other oxidants, dia-mide and menadione, was compared with the wild-type strain.
The vma2� mutant is only slightly more sensitive to diamidethan the wild-type strain at pH 5, but there is an increase in itsrelative sensitivity at pH 7. In contrast, H2O2 (at the concentra-tions used in Fig. 1A) almost completely inhibits growth ofvma2� and vma3� cells at neutral pH (data not shown). Men-adione sensitivity could only be measured at elevated pH andwas clearlymore pronounced in the vma2�mutant than inwildtype. The sensitivity of the vma3�mutant to these oxidantswassimilar to or slightly greater than that of the vma2� mutant(data not shown). These data support previous indications thatvmamutants are sensitive to various forms of applied oxidativestress but also suggest that themutants are exquisitely sensitiveto H2O2.
Both the vma2� and vma3�mutations disrupt subunits thatare required for all V-ATPase activity in the cell (12). One sub-unit of the V-ATPase, the Vo a subunit, is present as twoorganelle-specific isoforms, Vph1p and Stv1p, which havesteady state localizations at the vacuole and Golgi apparatus/endosomes, respectively, in wild-type cells (30, 31). Fig. 1Cshows that both vph1� and stv1� mutants are muchmore sen-sitive to peroxide at pH 5 than wild-type cells, although neitheris as sensitive as the vma2� or vma3� mutants. These resultssuggest that both vacuolar Vph1p-containing V-ATPases andthe Golgi/endosome Stv1p containing V-ATPases contributeto resistance to oxidative stress.As a means of quantitating the relative strength of the sensi-
tivity of the vma mutants to peroxide stress, we compared therelative halo size of the vma2� challenged with 2 and 5% per-oxide to those of deletions of the two superoxide dismutases(Sod1p and Sod2p) in yeast, which are firmly established asimportant defenses against oxidative stress (32). As shown inFig. 1D, vma2� mutants have significantly larger halos thanwild-type cells challenged with 2 and 5% peroxide, consistentwith the data shown in Fig. 1A. Furthermore, the halo sizes forthe vma2 mutant are comparable with those for either super-oxide dismutase mutant, suggesting that loss of V-ATPaseactivity compromises the resistance of cells to peroxide to acomparable degree.vmaMutants Are under Chronic Oxidative Stress—Cells that
are highly sensitive to an applied oxidative stress often containelevated levels of reactive oxygen species (ROS), which can bepresent in a number of different chemical forms (33), even inthe absence of an applied stress. An increase in steady-stateROS levels can be detected with redox-sensitive dyes. In orderto compare ROS levels in the wild-type and vma mutants, weincubated the cells with DHR. DHR enters the cell but is non-fluorescent unless it is oxidized by intracellular oxidants toform rhodamine 123. Wild-type and vma2� mutants weregrown overnight in fully supplemented minimal medium, andthenDHRwas added for an additional 4.5-h incubation. Fig. 2Ashows DHR staining of one field of wild-type and of vma2�mutants. On average, only 6.75 � 1.85% of wild-type cellsshowed any staining with DHR, but 17.2 � 0.2% of vma2� cellswere stained (see also Fig. 3B). These results indicate that loss ofV-ATPase activity does result in a higher percentage of cellswith ROS, consistent with hypersensitivity to oxidative stress inthese cells.
Yeast V-ATPase Protects against Oxidative Stress
MARCH 9, 2007 • VOLUME 282 • NUMBER 10 JOURNAL OF BIOLOGICAL CHEMISTRY 7127

Cells that are under chronic oxidative stress accumulate pro-tein modifications of several different types (34, 35). One ofthese modifications is protein carbonylation, and levels of car-
bonylated protein have frequentlybeen used as a measure of chronicoxidative stress (24, 35). Wedetected protein carbonylationusing the OxyBlot system, in whichcarbonylated proteins are furtherderivatized with DNP, which canthen be detected by Western blot-ting. As shown in Fig. 2B, lysatesfrom wild-type cells contain verylittle DNP-reactive carbonylatedprotein, but the vma2� mutantcontains high levels of protein car-bonylation, even though no extra-cellular oxidant was added in theseexperiments.What Is the Source of ROS Accu-
mulation in the vmaMutants?—Re-active oxygen species can arise froma number of different sources andprocesses, but mitochondria, andspecifically the respiratory chain,are frequently one of the majorsources (36). In yeast, at least onesubunit of each complex of the elec-tron transport chain is encoded bymitochondrial DNA, and the cellscan survive by fermentative growthin the absence of mitochondrialDNA and respiratory function.Therefore, respiration can be totallyabolished by inducing the loss ofmitochondrial DNA. We selectedwild-type and vma2� mutants thathad lost mitochondrial DNA (rho0cells) and then determined whetherthe rho0 versions of the two strainsweremore or less sensitive to oxida-tive stress. As shown in Fig. 3A, lossof mitochondrial DNA did notaffect the growth of either strain atpH 5. More importantly, the rho0cells showed no improvement intheir sensitivity to oxidative stress,as would be expected if the respi-ratory chain were themajor sourceof the accumulating ROS (37). Infact, the vma2� rho0 strain wasslightly more sensitive to hydro-gen peroxide. We also comparedDHR staining between wild-typeand vma2� cells in the presenceand absence of mitochondrialDNA, and the results are shown inFig. 3B. Although the percentage
of cells showing DHR staining appeared to be somewhatlower in both the wild-type rho0 and vma2�rho0 cells, thedifference from the corresponding rho� cells was not statis-
FIGURE 1. Sensitivity of V-ATPase deletion mutants to various forms of oxidative stress. A, halo assayscomparing sensitivity of wild-type BY4741 cells (wt), BY4741 vma2�, and BY4741 vma3� cells to H2O2. Cellsgrown to logarithmic phase in YEPD buffered to pH 5 were applied to a YEPD, pH 5, plate as an overlay. Filterpaper disks were placed on top of the overlay, and 1, 2, 5, and 10% H2O2 was applied to the four disks shown.The plates were grown for 2 days at 30 °C. Clear or lightly cloudy areas surrounding each disk represent inhibitedcell growth due to H2O2 diffusion from the disk, and the size of these “halos” is dependent on the sensitivity ofthe strain to H2O2 (22). There was no growth inhibition around a control disk to which no H2O2 was applied (notshown). B, halo assays comparing the sensitivity of wild-type and vma2� cells to diamide and menadione. Theassay was prepared as in A, except that 250 mM (left) and 500 mM (right) diamide or 20 mM (left) and 40 mM (right)menadione were applied to the disks. For the middle set of panels, the plate supporting the overlay was YEPD,pH 7, rather than YEPD, pH 5. C, halo assays comparing the sensitivity of BY4741 vph1� and BY4741 stv1� toH2O2. Disks on the right were spotted with 10% H2O2, and those on the left were spotted with 5% H2O2. Theassays were done as described in A. D, comparison on relative halo sizes for different mutants in response to 5and 2% H2O2. Halo assays were performed on the indicated strains as described for A, and the size of the haloaround the 5% H2O2 (light gray) and 2% H2O2 (dark gray) filters were measured for the mutant and wild-typestrains. Relative halo size was determined by dividing the size of the mutant halo by the size of the wild-typehalo determined in parallel. Bars represent the mean � S.E. for eight independent experiments for vma2� andtwo independent experiments for sod1� and sod2�.
Yeast V-ATPase Protects against Oxidative Stress
7128 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 10 • MARCH 9, 2007

tically significant. In contrast, there was a significant (p �0.05) difference between the wild-type and vma2� strain,both for the rho� and the rho0 strains. These results argue
against the respiratory chain as the major source of endoge-nous oxidative stress in the vma mutants.It is still possible that mitochondria contribute to oxidative
stress in cells grown in nonfermentable carbon sources, whererespiration is required and ROS production from the respira-tory chain may be unavoidable. Like many strains that are sen-sitive to oxidative stress (32), vmamutants are unable to grow attypical concentrations of nonfermentable carbon sources,although it is possible to isolate metabolically normal mito-chondria from the mutants (38). Supek et al. (39) reported thatvmamutants were able to grow at lower concentrations of non-
FIGURE 2. V-ATPase deletion mutants show evidence of chronic oxidativestress. A, wild-type and BY4741 vma2� were stained with dihydrorhodamine123 as described under “Experimental Procedures.” After 4.5-h staining, fieldsof cells were visualized using a �40 objective, under Nomarski optics (DIC; leftfield of each set) and rhodamine fluorescence optics (right field of each set).B, wild-type and BY4741 vma2� cells in logarithmic phase growth were con-verted to spheroplasts and lysed as described under “Experimental Proce-dures.” Lysate proteins were reacted with dinitrophenyl hydrazine (�) usingthe OxyBlot kit or subjected to a mock derivatization (�). After derivatization,equal amounts of protein from each sample were loaded in each lane of anSDS-polyacrylamide gel, separated, and transferred to nitrocellulose. DNPlabeling of carbonylated proteins was detected by immunoblotting with arabbit anti-DNP polyclonal antibody followed by incubation with horseradishperoxidase-conjugated second antibody and color development using anenhanced chemiluminescence kit (Amersham Biosciences).
FIGURE 3. The electron transport chain may not be the major source ofROS in vma mutants grown in glucose. A, wild-type and BY4741 vma2�cells were depleted of mitochondrial DNA as described under “ExperimentalProcedures” to form rho0 cells, and the growth of the rho� and rho0 cells on SCand SC containing the indicated concentration of H2O2 is compared. For eachstrain, 10-fold serial dilutions from liquid cultures of cells at identical densitywere spotted on the indicated plates. Cells were grown for 2 days at 30 °C.B, the percentage of both rho� and rho0 cells stained with DHR was measuredin two independent experiments, and the mean � range of the two experi-ments is shown. C, the growth of wild type and vma2� mutants on YEPD, pH5 (optimal growth conditions), YEP containing typical concentrations of non-fermentable carbon sources (3% glycerol plus 2% ethanol or 3% lactate), andYEP with lowered concentrations of nonfermentable carbon sources (3%glycerol only or 0.3% lactate) is compared as described in A.
Yeast V-ATPase Protects against Oxidative Stress
MARCH 9, 2007 • VOLUME 282 • NUMBER 10 JOURNAL OF BIOLOGICAL CHEMISTRY 7129

fermentable carbon sources, also suggesting that defective res-piration is not the cause of the petite phenotype in the vmamutants. We assessed the growth of the vma2� mutant at var-ied concentrations of two different nonfermentable carbonsources, glycerol/ethanol and lactate. As shown in Fig. 3C, thevma2� mutant cannot grow on 3% glycerol plus 2% ethanol oron 3% lactate, although the wild-type strain grows vigorouslyunder both conditions. However, the vma2� mutant was ableto grow in the presence of 3% glycerol without added ethanol or0.3% lactate.There are numerous defense systems against oxidative
stress in yeast, including the cytosolic Cu/Zn-superoxidedismutase Sod1p, thioredoxin Trx2p and thioredoxin-dependent peroxidase Tsa1p, reduced glutathione (synthe-sized and regenerated by glutathione synthetase Gsh1p andglutathione reductase Glr1p, respectively), and the oxidativearm of the pentose phosphate pathway (33). Complementingthese cytosolic defense systems are some similar proteinslocalized to the mitochondria, including a mitochondrialthioredoxin, Trx3p, and the manganese-dependent superox-ide dismutase Sod2p. Mutations in each of these defensesystems result in increased sensitivity to challenge withextracellular oxidant (1, 33). We reasoned that doublemutants lacking both the V-ATPase and one of these defensesystems might provide clues to the source of the endogenousoxidative stress in the vma mutants. We tested this hypoth-esis by crossing the vma2� mutant to haploid deletions inseveral major cytosolic and mitochondrial antioxidantdefense systems, selecting diploids, and then sporulating thediploids and dissecting tetrads to obtain double mutantspores. Spores were allowed to germinate on YEPD mediumbuffered to pH 5, conditions that are optimal for growth ofvma deletion mutants and that appear to minimize sensitiv-ity to exogenous oxidative stress. Viable double mutantspores were obtained for crosses of vma2� with trx2�,tsa1�, gsh1�, glr1�, and zwf1�, and in Fig. 4A, the growth ofseveral of the double mutant spores on SC and SC mediumcontaining 0.5 mM H2O2 concentration is compared. In gen-eral, the double mutant spores are clearly more sensitive tohydrogen peroxide than the single mutants. Although dou-ble mutant spores were obtained by dissection on YEPD pH5, the gsh1�vma2� double mutant was not only unable togrow in the presence of low peroxide concentrations (datanot shown) but also was unable to grow at all on supple-mented minimal medium. This indicates a synthetic effect ofthe gsh1� and vma2� mutations that is manifest even in theabsence of additional oxidative stress. All of the other muta-tions tested showed increased sensitivity to oxidative stresswhen combined with the vma2� mutation, but the increasein sensitivity appears to be more pronounced for the cytoso-lic defense system mutants shown in Fig. 4A, trx2� andglr1�, than for the mitochondrial defense system mutants,trx3� and sod2�.
In similar dissections of a diploid heterozygous for sod1� andvma2�, we were not able to obtain double mutant spores, asshown in Fig. 4B. This suggests that the twomutations are syn-thetically lethal, even on the YEPD, pH 5, plates used for dissec-tion. However, both sod1� and vma2� have defects in spore
germination, and this could prevent growth of otherwise viablespores. In order to bypass the germination defect, we trans-formed the vma2� mutant with a plasmid-borne copy of wild-
FIGURE 4. vma2� mutants show synthetic sensitivity to oxidative stresswith mutants in major cytosolic antioxidant pathways. BY4742 vma2�mutant was crossed to the indicated haploid deletion mutants lacking indi-vidual cytosolic (trx2�, glr1�, gsh1�, and sod1�) or mitochondrial (trx3� andsod2�) protection mechanisms, the resulting diploids were sporulated, andtetrads were dissected. A, single and double spores from a single tetrad arecompared with the growth of the vma2� parent strain and BY4741 (wt) asdescribed in Fig. 3A. B, comparison of synthetic interaction with sod1� andsod2�. Left, the vma2� and sod1� heterozygous diploid (�pVMA2) wassporulated as described above, and tetrads were dissected on YEPD, pH 5.A–D, spores from single tetrads; the boxed positions represent missing sporespredicted from the genotypes of the other three spores to be vma2� sod1�double mutants. The heterozygous diploid was also transformed with thewild-type VMA2 gene on a plasmid before sporulation and dissection(�pVMA2). Two four-spore tetrads are shown, with the boxed spores corre-sponding to vma2� sod1� double mutants. Both spores retained the plasmidthrough dissection and germination (data not shown). Right, a vma2� andsod2� heterozygous diploid was sporulated and dissected as describedabove. In contrast to the sod1� diploid, this strain did yield four-spore tetradseven without the VMA2 plasmid present. Double mutant spores are boxed.
Yeast V-ATPase Protects against Oxidative Stress
7130 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 10 • MARCH 9, 2007

type VMA2, and then crossed, selected for diploids, and dis-sected tetrads. As shown in Fig. 4B, under these conditions,four-spore tetrads could be obtained. The VMA2-containingplasmid contained a URA3 marker, so we tested for the pres-ence of the plasmid in each spore by determining growth onmedium lacking uracil and also transferred the spores tomedium containing 5-fluoroorotic acid, a negative selection forthe URA3 marker. Two viable spores (boxed in Fig. 4B) con-tained both the vma2� and sod1� mutations and also showedsome growth on 5-fluoroorotic acid, indicating that they wereable to lose the VMA2 plasmid. These spores showed very slowgrowth and poor viability and could not be tested further foroxidant sensitivity. Therefore, sod1� vma2� double mutantspores can be obtained but have a very severe synthetic growthdefect, even in the absence of an imposed oxidative stress. Incontrast, sod2�vma2� spores, shown for comparison in Fig. 4B,were readily obtained from tetrad dissection and grew well onYEPD, pH 5, suggesting that there is a much less severe syn-thetic defect between the mitochondrial superoxide dismutasemutant and the vma2� mutant.
Taken together, these results indicate that most of themajor cytosolic mechanisms for handling oxidative stress areoperating in the vma2� mutant. In fact, they are critical forits optimal growth, because double mutants exhibit in-creased sensitivity to low concentrations of H2O2 and poorergrowth even in the absence of added oxidant in some cases.Therefore, it is unlikely that the sensitivity of the V-ATPasemutants to oxidative stress is coming from their inability tosupport one of these mechanisms, but instead, loss ofV-ATPase activity must help to create an additional sourceof increased oxidative stress. Loss of mitochondrial protec-tion systems aggravates the sensitivity of the vmamutants tooxidative stress but less severely than loss of the parallelcytoplasmic systems.Microarray Analysis Indicates Dramatically Perturbed
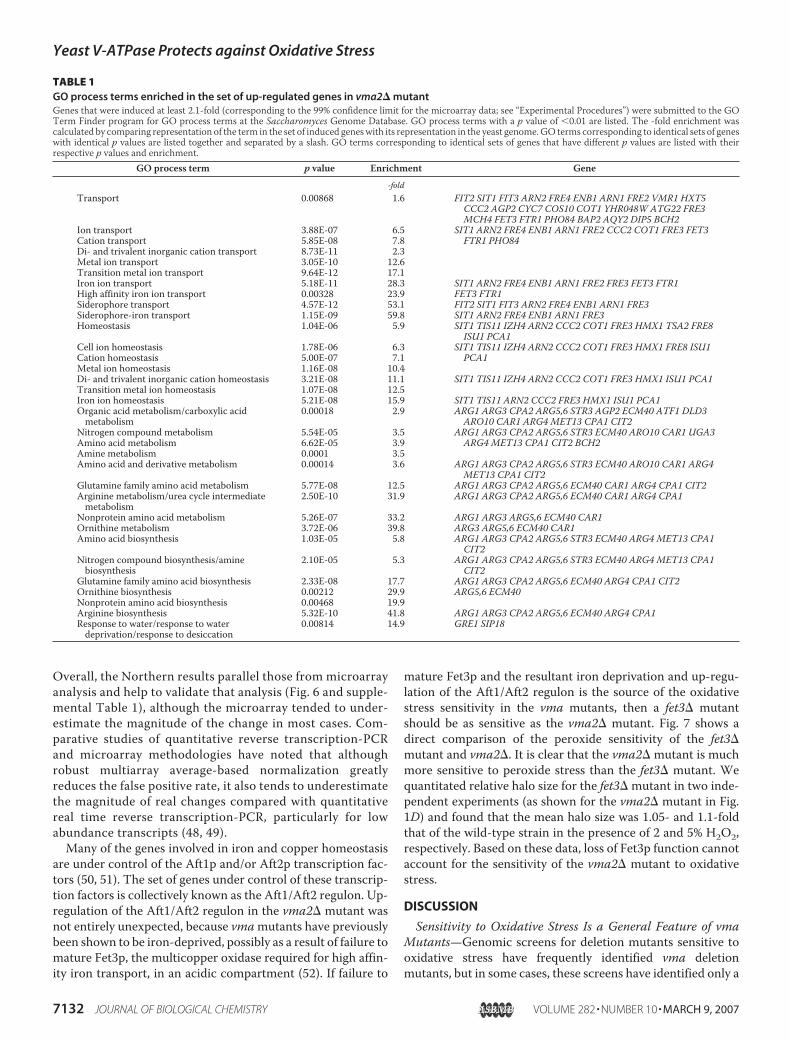
Metal Ion Homeostasis and Up-regulation of TSA2 in avma2� Mutant—The overall transcriptional profile of a cellcan provide critical insights into its physiological state. In aneffort to better understand potential sources of oxidativestress in the vma mutants, we grew both wild-type cells anda congenic vma2� mutant to log phase in YEPD, pH 5, pre-pared RNA, and then performedmicroarray analysis to com-pare the transcriptional profiles of the two strains. Genesthat were up-regulated or down-regulated 2.1-fold or more(calculated to be at the 99th percentile confidence level asdescribed under “Experimental Procedures”) in the vma2�strain are listed in supplemental Table 1. These data wereanalyzed using the Saccharomyces Genome Database GeneOntology (GO) Term Finder program to extract GO termsthat are enriched in the up-regulated or down-regulatedgene sets relative to the entire set of yeast genes on themicroarray. The results from the set of up-regulated geneswere particularly notable in the context of the oxidativestress sensitivity of the vma2 mutant. Table 1 lists the GOprocess terms that were calculated to have a p value (proba-bility of random association) of �0.01 for this set of genes aswell as the enrichment of each term in the data set relative toits representation in the genome. It is clear from this analysis
that there is a highly significant enrichment for genes in twogeneral categories: 1) metal ion (particularly iron) transportand homeostasis and 2) arginine/glutamine/ornithine bio-synthesis and metabolism.To begin to assess the extent to which the vma2� mutant is
responding transcriptionally to oxidative stress, we comparedthe set of genes up-regulated or down-regulated by at least4-fold in the vma2� strain to the published sets of geneschanged by at least 4-fold in response to a grx5� mutation or20-min treatment with 0.3 mM H2O2 (5, 40). The grx5 nullmutant has been used as a model of yeast cells under chronic,modest oxidative stress (40). Grx5p is a mitochondrial glutare-doxin, and in its absence, cells accumulate carbonylated pro-teins even in the absence of applied oxidative stress (41), symp-tomatic of chronic oxidative stress (see Fig. 2). Transcriptionalresponse to an acute challenge toH2O2wasmeasured byGaschet al. (5) in the context of determining common features inresponse to environmental stress. Only 5323 genes were avail-able in all three data sets for comparison, but of these, 264 genesshowed a 4-fold change in at least one of the three data sets, andthree-way correlations of these genes are shown in Fig. 5. Therewas very little correlation between the acutely stressed H2O2-treated cells and the chronically stressed grx5 mutant (R2 �0.0038). The vma2�mutant data correlated better with each ofthese data sets, showing anR2 � 0.060with the grx5 data and anR2� 0.092with theH2O2 treatment data. Taken together, thesedata indicate that the transcriptional response to loss of Vma2phas elements in common with responses seen in these modelsfor chronic and acute oxidative stress but also has clear differ-ences. Tabulated data used to generate Fig. 5 are shown in sup-plemental Table 2. As part of this analysis, we were also able toidentify which genes were changed by at least 4-fold in morethan one data set. Only eight genes were changed by at least4-fold in more than one data set, and all were increased. Fiveof these, FIT3, FIT2, ARN1, ARN2, and YBR047w, have pre-viously been implicated in iron and/or copper homeostasis(42, 43). Each of these genes was up-regulated by at least2.9-fold in all three data sets, suggesting that they are strongcommon components.Because of the potential for links between perturbed metal
ion homeostasis and oxidative stress, we probedNorthern blotsof two independent RNA preparations from the wild-type andvma2� genes for several of the genes that are implicated iniron/copper homeostasis and that showed altered expression inthe microarray. The results are shown in Fig. 6. The Northernblots show a clear up-regulation of mRNAs for FIT2, whichencodes one of a family of cell wall mannoproteins that assist iniron uptake; SIT1 and ENB1, which both encode iron-sid-erophore transporters; FRE2, an iron reductase at the plasmamembrane;HMX1, an ER-localized heme degradation enzymerequired for recovery of iron fromheme; andFET3, required forhigh affinity iron transporter at the plasmamembrane (44–46).In contrast, the major high affinity transporter of copper at theplasmamembrane, encoded byCTR1 (47), is down-regulated inthe vma2� cells. Although most enzymes previously identifiedas up-regulated in response to an applied oxidative stress arenot significantly up-regulated, there is a large up-regulation ofTSA2, which encodes an isoform of thioredoxin peroxidase.
Yeast V-ATPase Protects against Oxidative Stress
MARCH 9, 2007 • VOLUME 282 • NUMBER 10 JOURNAL OF BIOLOGICAL CHEMISTRY 7131
Overall, the Northern results parallel those from microarrayanalysis and help to validate that analysis (Fig. 6 and supple-mental Table 1), although the microarray tended to under-estimate the magnitude of the change in most cases. Com-parative studies of quantitative reverse transcription-PCRand microarray methodologies have noted that althoughrobust multiarray average-based normalization greatlyreduces the false positive rate, it also tends to underestimatethe magnitude of real changes compared with quantitativereal time reverse transcription-PCR, particularly for lowabundance transcripts (48, 49).Many of the genes involved in iron and copper homeostasis
are under control of the Aft1p and/or Aft2p transcription fac-tors (50, 51). The set of genes under control of these transcrip-tion factors is collectively known as the Aft1/Aft2 regulon. Up-regulation of the Aft1/Aft2 regulon in the vma2� mutant wasnot entirely unexpected, because vmamutants have previouslybeen shown to be iron-deprived, possibly as a result of failure tomature Fet3p, the multicopper oxidase required for high affin-ity iron transport, in an acidic compartment (52). If failure to
mature Fet3p and the resultant iron deprivation and up-regu-lation of the Aft1/Aft2 regulon is the source of the oxidativestress sensitivity in the vma mutants, then a fet3� mutantshould be as sensitive as the vma2� mutant. Fig. 7 shows adirect comparison of the peroxide sensitivity of the fet3�mutant and vma2�. It is clear that the vma2� mutant is muchmore sensitive to peroxide stress than the fet3� mutant. Wequantitated relative halo size for the fet3� mutant in two inde-pendent experiments (as shown for the vma2� mutant in Fig.1D) and found that the mean halo size was 1.05- and 1.1-foldthat of the wild-type strain in the presence of 2 and 5% H2O2,respectively. Based on these data, loss of Fet3p function cannotaccount for the sensitivity of the vma2� mutant to oxidativestress.
DISCUSSION
Sensitivity to Oxidative Stress Is a General Feature of vmaMutants—Genomic screens for deletion mutants sensitive tooxidative stress have frequently identified vma deletionmutants, but in some cases, these screens have identified only a
TABLE 1GO process terms enriched in the set of up-regulated genes in vma2� mutantGenes that were induced at least 2.1-fold (corresponding to the 99% confidence limit for the microarray data; see “Experimental Procedures”) were submitted to the GOTerm Finder program for GO process terms at the Saccharomyces Genome Database. GO process terms with a p value of �0.01 are listed. The -fold enrichment wascalculated by comparing representation of the term in the set of induced geneswith its representation in the yeast genome.GO terms corresponding to identical sets of geneswith identical p values are listed together and separated by a slash. GO terms corresponding to identical sets of genes that have different p values are listed with theirrespective p values and enrichment.
GO process term p value Enrichment Gene-fold
Transport 0.00868 1.6 FIT2 SIT1 FIT3 ARN2 FRE4 ENB1 ARN1 FRE2 VMR1 HXT5CCC2 AGP2 CYC7 COS10 COT1 YHR048W ATG22 FRE3MCH4 FET3 FTR1 PHO84 BAP2 AQY2 DIP5 BCH2
Ion transport 3.88E-07 6.5 SIT1 ARN2 FRE4 ENB1 ARN1 FRE2 CCC2 COT1 FRE3 FET3FTR1 PHO84Cation transport 5.85E-08 7.8
Di- and trivalent inorganic cation transport 8.73E-11 2.3Metal ion transport 3.05E-10 12.6Transition metal ion transport 9.64E-12 17.1Iron ion transport 5.18E-11 28.3 SIT1 ARN2 FRE4 ENB1 ARN1 FRE2 FRE3 FET3 FTR1High affinity iron ion transport 0.00328 23.9 FET3 FTR1Siderophore transport 4.57E-12 53.1 FIT2 SIT1 FIT3 ARN2 FRE4 ENB1 ARN1 FRE3Siderophore-iron transport 1.15E-09 59.8 SIT1 ARN2 FRE4 ENB1 ARN1 FRE3Homeostasis 1.04E-06 5.9 SIT1 TIS11 IZH4 ARN2 CCC2 COT1 FRE3 HMX1 TSA2 FRE8
ISU1 PCA1Cell ion homeostasis 1.78E-06 6.3 SIT1 TIS11 IZH4 ARN2 CCC2 COT1 FRE3 HMX1 FRE8 ISU1
PCA1Cation homeostasis 5.00E-07 7.1Metal ion homeostasis 1.16E-08 10.4Di- and trivalent inorganic cation homeostasis 3.21E-08 11.1 SIT1 TIS11 IZH4 ARN2 CCC2 COT1 FRE3 HMX1 ISU1 PCA1Transition metal ion homeostasis 1.07E-08 12.5Iron ion homeostasis 5.21E-08 15.9 SIT1 TIS11 ARN2 CCC2 FRE3 HMX1 ISU1 PCA1Organic acid metabolism/carboxylic acidmetabolism
0.00018 2.9 ARG1 ARG3 CPA2 ARG5,6 STR3 AGP2 ECM40 ATF1 DLD3ARO10 CAR1 ARG4 MET13 CPA1 CIT2
Nitrogen compound metabolism 5.54E-05 3.5 ARG1 ARG3 CPA2 ARG5,6 STR3 ECM40 ARO10 CAR1 UGA3ARG4 MET13 CPA1 CIT2 BCH2Amino acid metabolism 6.62E-05 3.9
Amine metabolism 0.0001 3.5Amino acid and derivative metabolism 0.00014 3.6 ARG1 ARG3 CPA2 ARG5,6 STR3 ECM40 ARO10 CAR1 ARG4
MET13 CPA1 CIT2Glutamine family amino acid metabolism 5.77E-08 12.5 ARG1 ARG3 CPA2 ARG5,6 ECM40 CAR1 ARG4 CPA1 CIT2Arginine metabolism/urea cycle intermediatemetabolism
2.50E-10 31.9 ARG1 ARG3 CPA2 ARG5,6 ECM40 CAR1 ARG4 CPA1
Nonprotein amino acid metabolism 5.26E-07 33.2 ARG1 ARG3 ARG5,6 ECM40 CAR1Ornithine metabolism 3.72E-06 39.8 ARG3 ARG5,6 ECM40 CAR1Amino acid biosynthesis 1.03E-05 5.8 ARG1 ARG3 CPA2 ARG5,6 STR3 ECM40 ARG4 MET13 CPA1
CIT2Nitrogen compound biosynthesis/aminebiosynthesis
2.10E-05 5.3 ARG1 ARG3 CPA2 ARG5,6 STR3 ECM40 ARG4 MET13 CPA1CIT2
Glutamine family amino acid biosynthesis 2.33E-08 17.7 ARG1 ARG3 CPA2 ARG5,6 ECM40 ARG4 CPA1 CIT2Ornithine biosynthesis 0.00212 29.9 ARG5,6 ECM40Nonprotein amino acid biosynthesis 0.00468 19.9Arginine biosynthesis 5.32E-10 41.8 ARG1 ARG3 CPA2 ARG5,6 ECM40 ARG4 CPA1Response to water/response to waterdeprivation/response to desiccation
0.00814 14.9 GRE1 SIP18
Yeast V-ATPase Protects against Oxidative Stress
7132 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 10 • MARCH 9, 2007

subset of the vmamutants (7), raising the possibility that resist-ance to oxidative stress is a “moonlighting” function of certainV-ATPase subunits rather than a general role of the V-ATPase.
We deliberately chose to assess sensitivity to oxidative stress instrains containing deletions in a V1 sector subunit (vma2�) anda Vo sector subunit (vma3�) that are both found in all yeastV-ATPases as well as the twoVo a subunit isoforms (vph1� andstv1�), each found in a subset of V-ATPases. We found that allwere sensitive, suggesting that sensitivity is a general feature ofloss of V-ATPase activity. The vma2� and vma3� mutantsweremore sensitive than the vph1� and stv1�mutants, but thisis expected, because the latter twomutants only lack a subset ofV-ATPase complexes. The vma3�mutantmay be slightlymoresensitive to peroxide (Fig. 1A) and other oxidants (data notshown) than the vma2� mutant. This difference is harder toexplain, because both mutations abolish all V-ATPase activity.However, Vma3p has also been suggested to play a role invacuole-vacuole fusion independent from V1 subunits (53),and it is possible that an additional function could accountfor the small increase in sensitivity to oxidative stress. Takentogether with the previous data, these results suggest thatloss of V-ATPase function is responsible for the sensitivity tooxidative stress and that loss of function in both the vacuolarand Golgi/endosome compartments may contribute to thissensitivity.Increased susceptibility to oxidative stress may also account
for other characteristics of vma mutants. The inability of vmamutants to grow on nonfermentable carbon sources has longbeen rather puzzling, particularly sinceOhya et al. (38) demon-strated that themutant cells have normal rates of respiration aswell as normal levels of several respiratory chain enzymes inisolated mitochondria. These experiments argued against theinability to synthesize respiratory enzymes (e.g. as a result ofdefects in metal ion trafficking or distribution (54)) as thesource of the growth defect in the vma strains. However, manymutants sensitive to oxidative stress fail to grow on nonfer-mentable carbon sources, presumably because they cannot tol-erate the levels of superoxide and other ROS produced as sideproducts of respiration (7). We have also found that vmamutants are very slow to return to log phase growth after theyhave grown to high density.4 This is also characteristic of cer-tain mutants sensitive to oxidative stress, because they poorlytolerate the diauxic switch that accompanies consumption ofavailable glucose and the metabolic transition from fermenta-tion to consumption of ethanol (55–58). The ability of the vmamutants to grow on low, but not high, concentrations of non-fermentable carbon sources (Fig. 4A) (39) may also be consist-ent with their sensitivity to oxidative stress. Supek et al. (39),who first observed this phenomenon, hypothesized that vacu-olar function was required for storage (or possibly metabolism)of excess respiratory metabolites. These “excess metabolites”could be ROS produced as a side product of rapid respiration.However, the growth of the vma mutants on low concentra-tions of nonfermentable carbon sources also seems to be some-what paradoxical, because there is general agreement that athigh levels of respiratory activity lower levels of ROS are pro-duced (36, 59). Further work will be necessary to fully under-stand the connections between mitochondrial function andoxidative stress in the vmamutants.
4 E. Milgrom, H. Diab, F. Middleton, and P. M. Kane, unpublished data.
FIGURE 5. Correlation of expression changes in the vma2� mutant,grx5�, and wild-type (wt) cells treated for 20 min with 0.3 mM H2O2.Genes showing at least a 4-fold change of expression between vma2� andwild-type cells (this study), grx5� (a model of wild-type chronic oxidativestress), and wild-type cells (40) or wild-type cells with and without a 20-mintreatment with 0.3 mM H2O2 (a model of acute oxidative stress (5)) were iden-tified, and the expression of these genes among the three experiments wascompared. 264 genes changed by at least 4-fold in at least one of the data setsand thus are included in the comparison. Log2 changes for each of thesegenes are plotted against the other two and used to calculate the indicated R2
value.
Yeast V-ATPase Protects against Oxidative Stress
MARCH 9, 2007 • VOLUME 282 • NUMBER 10 JOURNAL OF BIOLOGICAL CHEMISTRY 7133

Potential Sources of OxidativeStress in vma Mutants—Althoughsensitivity to oxidative stress mayaccount for the inability of the vmamutants to grow on standard con-centrations of nonfermentable car-bon sources, superoxide productionfrom the respiratory chain, gener-ally considered a major source ofcellular ROS (60), does not accountfor the peroxide sensitivity of thevma2� mutant. A number of differ-entmutants that are sensitive to oxi-dative stress show improved growthin the absence of mitochondrialDNA (37, 61). Eviction of mito-chondrial DNA from vma2�mutant to form the vma2�rho0mutant did not improve the generalgrowth properties of the mutant orits resistance to peroxide challenge(Fig. 4B). rho0 strains have previ-ously been shown to have loweredresistance to peroxide in some cases(62), but the basis of this phenome-non is not well understood. Therewas relatively little decrease in ROSstaining in the vma2�rho0 strain,again suggesting that respiration isnot the major source of ROS.
We reasoned that the vmamutants might be affecting one ofthe other major mechanisms for resistance to oxidative stress(e.g. by perturbing metal ion access for assembly of superoxidedismutase). Althoughwe cannot completely eliminate this pos-sibility, the strong synthetic phenotypes of mutants lackingboth vma2� and several other mechanisms argue that loss ofV-ATPase function creates an additional source of stress.The strong up-regulation of the Aft1/Aft2 regulon in the
vma2� mutant also highlights the potential importance of theV-ATPase inmetal ion homeostasis and suggests some possiblesources of oxidative stress sensitivity. Davis-Kaplan et al. (52)previously showed that vmamutants have low levels of cellulariron that may be attributed to the requirement for an acidicpost-Golgi compartment for insertion of the copper clusterinto apo-Fet3p. Iron deprivation and/or loss of Fet3p functionhas also been associatedwith increased sensitivity to other tran-sitionmetals because of up-regulation of less specific transport-ers (63). Because some of these metals are redox-active, theycould contribute to oxidative stress.However, we donot believethat the iron deprivation that occurs in vma mutants is solelyresponsible for the oxidative stress. First, if loss of Fet3p matu-ration were the root cause of the sensitivity of vma2� to oxida-tive stress, then the fet3� mutant should be as sensitive to oxi-dative stress as the vma2�mutant, but it is not (Fig. 7). Second,we found that although the addition of iron and copper to anumber of mutants exhibiting a Vma� phenotype improvedtheir growth, it did not improve the growth of the vmamutants(16). In contrast, the addition of excess iron reduced the toxicity
FIGURE 6. Northern blots of genes related to iron metabolism or oxidative stress. Each panel comparestwo independent preparations of wild-type and vma2� RNA, probed with a 32P-labeled fragment of the indi-cated gene and actin as a control. Identical amounts (10 –20 �g, depending on the gene) of wild-type andvma2� RNA were loaded for each blot. Gene-specific probes and Northern blots were prepared as describedunder “Experimental Procedures.” The blots shown were quantitated using an Amersham BiosciencesTyphoon PhosphorImager, and the average ratio of signals in the vma2� and wild-type sample pairs, normal-ized to the actin loading control, is shown � the range of the two sample pairs. Microarray ratios (supplementalTable 1) for these genes are included for comparison.
FIGURE 7. Loss of Fet3p function does not result in strong sensitivity tooxidative stress. Halo assays for wild-type cells and the congenic fet3� andvma2� mutants were prepared as described in the legend to Fig. 1 and under“Experimental Procedures,” with 2 and 5% H2O2 applied to the filter disk.Lawns of cells were grown for 2 days.
Yeast V-ATPase Protects against Oxidative Stress
7134 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 10 • MARCH 9, 2007

of other metals by competition under conditions where lessspecific transporters were up-regulated (63). In addition,although the Aft1/Aft2 regulon clearly has an iron-sensitiveinput, it is also induced in conditions other than direct irondeprivation, including chronic oxidative stress (40). In fact,cells become somewhatmore sensitive to oxidative stress in theabsence of Aft1p and much more sensitive in the absence ofboth Aft1p and Aft2p (50). This suggests that up-regulation ofthe Aft1/Aft2 regulon could be a response to the oxidativestress in the vma2� mutant rather than a cause.Significantly, in the vma2�mutant, we did not see a dramatic
up-regulation of the oxidative stress response genes controlledby Yap1p and Skn7p, which are strongly up-regulated inresponse to an acute oxidative stress (5, 64). Consistent withthis, we did not see a strong nuclear concentration of GFP-Yap1p (25) in the vma2mutant grown in the absence of appliedoxidant, although GFP-Yap1p was translocated to the nucleuswithin 15 min of H2O2 addition (data now shown). Microarrayanalysis of the grx5� mutant indicated little up-regulation ofYap1- or Skn7-dependent genes but did show a strong up-reg-ulation of the Aft1/Aft2 regulon, similar to that seen in thevma2�mutant (40). However, the comparisonwithmicroarraydata from cells treated for 20 min with H2O2 (Fig. 5) does sug-gest that there are at least some common features between tran-scriptional response to vma2� and an acute oxidative challengewith H2O2.In conclusion, we do not fully understand the source of the
chronic oxidative stress seen in the vma2� mutant, but we canpropose several potential sources. First, it is entirely possiblethat improper distribution or utilization of one or more metalscontributes to the oxidative stress, although a simple explana-tion based on iron deprivation does not seem sufficient. In arecent genomic dissection of the S. cerevisiae “ionome,” theV-ATPase was identified as a major player in uptake and distri-bution of multiple different metals (15). It is also possible thataltered pH homeostasis in the vma mutants leads to an envi-ronment that extends the lifetime of certain ROS or otherwisechanges their metabolism. Regardless of the mechanism, theV-ATPase appears to play a unique and essential role in resist-ance to oxidative stress that may be important in other eukary-otic cells as well.
Acknowledgments—We thank Scott Moye-Rowley (University ofIowa) for the GFP-Yap1 plasmid and Jerry Kaplan (University ofUtah) for iron measurements and helpful suggestions.
REFERENCES1. Carmel-Harel, O., and Storz, G. (2000)Annu. Rev.Microbiol. 54, 439–4612. Jamieson, D. J. (1998) Yeast 14, 1511–15273. Costa, V., andMoradas-Ferreira, P. (2001)Mol. AspectsMed. 22, 217–2464. Stadtman, E. R., and Berlett, B. S. (1998) Drug Metab. Rev. 30, 225–2435. Gasch, A. P., Spellman, P. T., Kao, C. M., Carmel-Harel, O., Eisen, M. B.,
Storz, G., Botstein, D., and Brown, P. O. (2000) Mol. Biol. Cell 11,4241–4257
6. Outten, C. E., Falk, R. L., and Culotta, V. C. (2005)Biochem. J. 388, 93–1017. Thorpe, G.W., Fong, C. S., Alic, N., Higgins, V. J., and Dawes, I.W. (2004)
Proc. Natl. Acad. Sci. U. S. A. 101, 6564–65698. Higgins, V. J., Alic, N., Thorpe, G. W., Breitenbach, M., Larsson, V., and
Dawes, I. W. (2002) Yeast 19, 203–214
9. Temple, M. D., Perrone, G. G., and Dawes, I. W. (2005) Trends Cell Biol.15, 319–326
10. Chang, E. C., Crawford, B. F., Hong, Z., Bilinski, T., and Kosman, D. J.(1991) J. Biol. Chem. 266, 4417–4424
11. Gralla, E. B., and Valentine, J. S. (1991) J. Bacteriol. 173, 5918–592012. Kane, P. M. (2006)Microbiol. Mol. Biol. Rev. 70, 177–19113. Nishi, T., and Forgac, M. (2002) Nat. Rev. Mol. Cell Biol. 3, 94–10314. Wieczorek, H., Brown, D., Grinstein, S., Ehrenfeld, J., and Harvey, W. R.
(1999) BioEssays 21, 637–64815. Eide, D. J., Clark, S., Nair, T. M., Gehl, M., Gribskov, M., Guerinot, M. L.,
and Harper, J. F. (2005) Genome Biol. 6, R7716. Sambade, M., Alba, M., Smardon, A. M., West, R. W., and Kane, P. M.
(2005) Genetics 170, 1539–155117. Parsons, A. B., Brost, R. L., Ding, H., Li, Z., Zhang, C., Sheikh, B., Brown,
G. W., Kane, P. M., Hughes, T. R., and Boone, C. (2004) Nat. Biotechnol.22, 62–69
18. Sherman, F. (1991)Methods Enzymol. 194, 3–2119. Winzeler, E. A., Shoemaker, D. D., Astromoff, A., Liang, H., Anderson, K.,
Andre, B., Bangham, R., Benito, R., Boeke, J. D., Bussey, H., Chu, A. M.,Connelly, C., Davis, K., Dietrich, F., Dow, S.W., El Bakkoury,M., Foury, F.,Friend, S. H., Gentalen, E., Giaever, G., Hegemann, J. H., Jones, T., Laub,M., Liao, H., Liebundguth, N., et al. (1999) Science 285, 901–906
20. Tong, A. H., Evangelista, M., Parsons, A. B., Xu, H., Bader, G. D., Page, N.,Robinson, M., Raghibizadeh, S., Hogue, C. W., Bussey, H., Andrews, B.,Tyers, M., and Boone, C. (2001) Science 294, 2364–2368
21. Fox, T. D., Folley, L. S., Mulero, J. J., McMullin, T. W., Thorsness, P. E.,Hedin, L. O., andCostanzo,M. C. (1991)Methods Enzymol. 194, 149–165
22. Haarer, B. K., and Amberg, D. C. (2004)Mol. Biol. Cell 15, 4522–453123. Madeo, F., Frohlich, E., Ligr, M., Grey, M., Sigrist, S. J., Wolf, D. H., and
Frohlich, K. U. (1999) J. Cell Biol. 145, 757–76724. Dirmeier, R., O’Brien, K. M., Engle, M., Dodd, A., Spears, E., and Poyton,
R. O. (2002) J. Biol. Chem. 277, 34773–3478425. Coleman, S. T., Epping, E. A., Steggerda, S. M., and Moye-Rowley, W. S.
(1999)Mol. Cell Biol. 19, 8302–831326. Schmitt, M. E., Brown, T. A., and Trumpower, B. L. (1990) Nucleic Acids
Res. 18, 3091–309227. Irizarry, R. A., Hobbs, B., Collin, F., Beazer-Barclay, Y. D., Antonellis, K. J.,
Scherf, U., and Speed, T. P. (2003) Biostatistics 4, 249–26428. Sambrook, J., and Russell, D. W. (2001)Molecular Cloning: A Laboratory
Manual, 3rd Ed., pp. 7.21–7.45, Cold Spring Harbor Laboratory Press,Cold Spring Harbor, NY
29. Doherty, R. D., and Kane, P. M. (1993) J. Biol. Chem. 268, 16845–1685130. Kawasaki-Nishi, S., Bowers, K., Nishi, T., Forgac, M., and Stevens, T. H.
(2001) J. Biol. Chem. 276, 47411–4742031. Manolson, M. F., Wu, B., Proteau, D., Taillon, B. E., Roberts, B. T., Hoyt,
M. A., and Jones, E. W. (1994) J. Biol. Chem. 269, 14064–1407432. Liu, X. F., Elashvili, I., Gralla, E. B., Valentine, J. S., Lapinskas, P., and
Culotta, V. C. (1992) J. Biol. Chem. 267, 18298–1830233. Moradas-Ferreira, P., Costa, V., Piper, P., and Mager, W. (1996)Mol. Mi-
crobiol. 19, 651–65834. Cabiscol, E., Piulats, E., Echave, P., Herrero, E., and Ros, J. (2000) J. Biol.
Chem. 275, 27393–2739835. Berlett, B. S., and Stadtman, E. R. (1997) J. Biol. Chem. 272, 20313–2031636. Barros,M.H., Bandy, B., Tahara, E. B., andKowaltowski, A. J. (2004) J. Biol.
Chem. 279, 49883–4988837. Guidot, D.M.,McCord, J.M.,Wright, R.M., andRepine, J. E. (1993) J. Biol.
Chem. 268, 26699–2670338. Ohya, Y., Umemoto, N., Tanida, I., Ohta, A., Iida, H., and Anraku, Y.
(1991) J. Biol. Chem. 266, 13971–1397739. Supek, F., Supekova, L., and Nelson, N. (1994) J. Biol. Chem. 269,
26479–2648540. Belli, G., Molina, M. M., Garcia-Martinez, J., Perez-Ortin, J. E., and Her-
rero, E. (2004) J. Biol. Chem. 279, 12386–1239541. Rodriguez-Manzaneque, M. T., Ros, J., Cabiscol, E., Sorribas, A., and Her-
rero, E. (1999)Mol. Cell Biol. 19, 8180–819042. Philpott, C. C., Protchenko, O., Kim, Y. W., Boretsky, Y., and Shakoury-
Elizeh, M. (2002) Biochem. Soc. Trans. 30, 698–70243. van Bakel, H., Strengman, E., Wijmenga, C., and Holstege, F. C. (2005)
Yeast V-ATPase Protects against Oxidative Stress
MARCH 9, 2007 • VOLUME 282 • NUMBER 10 JOURNAL OF BIOLOGICAL CHEMISTRY 7135

Physiol. Genomics 22, 356–36744. Kosman, D. J. (2003)Mol. Microbiol. 47, 1185–119745. Protchenko, O., Ferea, T., Rashford, J., Tiedeman, J., Brown, P. O., Bot-
stein, D., and Philpott, C. C. (2001) J. Biol. Chem. 276, 49244–4925046. Protchenko, O., and Philpott, C. C. (2003) J. Biol. Chem. 278,
36582–3658747. Dancis, A., Haile, D., Yuan, D. S., and Klausner, R. D. (1994) J. Biol. Chem.
269, 25660–2566748. Mimmack, M. L., Brooking, J., and Bahn, S. (2004) Biol. Psychiatry 55,
337–34549. Yuen, T., Wurmbach, E., Pfeffer, R. L., Ebersole, B. J., and Sealfon, S. C.
(2002) Nucleic Acids Res. 30, e4850. Blaiseau, P. L., Lesuisse, E., and Camadro, J. M. (2001) J. Biol. Chem. 276,
34221–3422651. Rutherford, J. C., Jaron, S., and Winge, D. R. (2003) J. Biol. Chem. 278,
27636–2764352. Davis-Kaplan, S. R.,Ward, D.M., Shiflett, S. L., andKaplan, J. (2004) J. Biol.
Chem. 279, 4322–432953. Peters, C., Bayer,M. J., Buhler, S., Andersen, J. S.,Mann,M., andMayer, A.
(2001) Nature 409, 581–588
54. Kaplan, J.,McVeyWard,D., Crisp, R. J., and Philpott, C. C. (2006)Biochim.Biophys. Acta 1763, 646–651
55. Drakulic, T., Temple, M. D., Guido, R., Jarolim, S., Breitenbach, M.,Attfield, P. V., and Dawes, I. W. (2005) FEMS Yeast Res. 5, 1215–1228
56. Bonawitz, N. D., Rodeheffer, M. S., and Shadel, G. S. (2006)Mol. Cell Biol.26, 4818–4829
57. Longo, V. D., Gralla, E. B., and Valentine, J. S. (1996) J. Biol. Chem. 271,12275–12280
58. Longo, V. D., Liou, L. L., Valentine, J. S., and Gralla, E. B. (1999) Arch.Biochem. Biophys. 365, 131–142
59. Lin, S. J., Kaeberlein, M., Andalis, A. A., Sturtz, L. A., Defossez, P. A.,Culotta, V. C., Fink, G. R., and Guarente, L. (2002) Nature 418, 344–348
60. Chance, B., Sies, H., and Boveris, A. (1979) Physiol. Rev. 59, 527–60561. Haynes, C. M., Titus, E. A., and Cooper, A. A. (2004) Mol. Cell 15,
767–77662. Grant, C. M., MacIver, F. H., and Dawes, I. W. (1997) FEBS Lett. 410,
219–22263. Li, L., and Kaplan, J. (1998) J. Biol. Chem. 273, 22181–2218764. Lee, J., Godon, C., Lagniel, G., Spector, D., Garin, J., Labarre, J., and Tole-
dano, M. B. (1999) J. Biol. Chem. 274, 16040–16046
Yeast V-ATPase Protects against Oxidative Stress
7136 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 10 • MARCH 9, 2007
Related Documents








![AtHMA3, a P1B-ATPase Allowing Cd/Zn/Co/Pb … › content › plantphysiol › 149 › 2 › 894.full.pdfAtHMA3, a P 1B-ATPase Allowing Cd/Zn/Co/Pb Vacuolar Storage in Arabidopsis1[W]](https://static.cupdf.com/doc/110x72/5f0eb3fb7e708231d44085a9/athma3-a-p1b-atpase-allowing-cdzncopb-a-content-a-plantphysiol-a-149-a.jpg)


