Loss of stearoyl-CoA desaturase expression is a frequent event in prostate carcinoma Stacy Moore 1 , Beatrice Knudsen 2 , Lawrence D. True 3 , Sarah Hawley 2 , Ruth Etzioni 2 , Christian Wade 1 , David Gifford 1 , Ilsa Coleman 1 and Peter S. Nelson 1 * 1 Division of Human Biology, Fred Hutchinson Cancer Research Center, Seattle, WA, USA 2 Division of Public Health Sciences, Fred Hutchinson Cancer Research Center, Seattle, WA, USA 3 Department of Pathology, University of Washington, Seattle, WA, USA Prostate carcinogenesis is influenced by genetic alterations result- ing in a biochemical condition that favors cell proliferation and survival. Studies of prostate carcinoma using comparative genomic hybridization and cDNA microarray analysis indicate that numerous biochemical processes may be affected during cel- lular transformation and progression to an invasive phenotype. Among the consistently observed tumor-associated changes are alterations in fatty acid metabolism that influence diverse cellular activities such as signaling, energy utilization, and membrane fluidity. Increases in fatty acid synthase (FAS) levels have been shown to be one of the earliest and most frequent molecular alterations in prostate carcinogenesis. We sought to identify tu- mor-associated changes in the expression of genes with functional roles associated with lipid metabolism. Defined populations of normal and neoplastic prostate epithelium were acquired by laser capture microdissection and transcript levels were measured by cDNA microarray hybridization. We determined that stearoyl- CoA desaturase (SCD) transcripts were downregulated in cancer relative to normal epithelium. These results were confirmed by quantitative PCR. Further analysis by immunohistochemical eval- uation of radical prostatectomy samples employed a quantitative scoring system with a range of 0 –300. The median SCD expression levels were 150, 45 and 10 for normal, PIN and carcinoma sam- ples, respectively. Statistically significant differential SCD expres- sion between normal and cancerous epithelium was determined at the p 0.001 level, and between PIN and prostate carcinoma at the p 0.03 level. Of these cases, 92% overexpressed fatty acid synthase (FAS) in cancerous cells and 84.7% exhibited the signa- ture of FAS overexpression and SCD loss in prostate carcinoma as compared to normal prostate epithelium. These results indicate that loss of SCD expression is a frequent event in prostate adeno- carcinoma, and further supports a role for altered lipid metabo- lism as a factor in the process of carcinogenesis. © 2004 Wiley-Liss, Inc. Key words: prostate cancer; microarray; microdissection; lipid me- tabolism; stearoyl-CoA desaturase The development of prostate carcinoma is driven by specific genetic alterations resulting in a biochemical state that favors cell proliferation and survival. The diversity and complexity of tumor cell aberrations is exemplified by the numerous chromosomal rearrangements and gene expression changes identified using com- prehensive analytical technologies such as comparative genomic hybridization and microarray analysis. 1–3 The extent of these ge- netic alterations suggests that numerous biochemical processes may be influenced during cellular transformation and the subse- quent progression to an invasive phenotype. The identification of consistent neoplasia-associated alterations involving specific gene products or molecular pathways may provide important insights into essential events associated with carcinogenesis. Fatty acid metabolism represents a key process that influences several diverse cellular pathways and characteristics including cell signaling, energy processing and membrane fluidity. 4–6 The enzy- matic and transporter activities that modulate concentrations of lipid substrate molecules represent important regulatory elements that modulate cellular proliferation and apoptosis. Several proteins involved in the metabolism of fatty acids have been determined to be altered in neoplastic prostate cells relative to their normal counterparts. For example, increases in fatty acid synthase (FAS) levels have been shown to be one of the earliest and most frequent molecular alterations in prostate carcinogenesis. 3,7 FAS plays a pivotal role in the de novo biosynthesis of fatty acids. 8 FAS expression in primary prostate carcinomas is predictive of cancer progression, 9 and the inhibition of FAS activity induces cell death in FAS-overexpressing cancer cell lines and prolongs survival of host animals implanted with tumor xenografts. 10 –13 Increased lev- els of transcripts encoding the -methylacyl-CoA-racemase (AM- ACR) enzyme have also been shown to occur frequently in pros- tate carcinoma. 14 –17 AMACR regulates the peroxisomal -oxidation of dietary branched chain fatty acids, a biochemical process that is plausibly related to prostate carcinogenesis through the production of pro-carcinogenic oxidative intermediates. 17 In vitro studies have shown that branched fatty acids in dairy and beef products, food sources with epidemiological associations with prostate cancer risk, markedly enhance AMACR expression in prostate cancer cells. 18 Immunohistochemical studies of AMACR protein expression have demonstrated a relationship between AM- ACR protein expression and cancer grade. 19 Due to the importance of lipid metabolism in prostate carcino- genesis, we sought to identify alterations in the expression levels of additional genes with functional roles associated with lipid metabolism that had consistent alterations in mRNA levels be- tween primary prostate carcinomas and corresponding non-neo- plastic secretory cells. Defined populations of normal and neoplas- tic prostate epithelium were acquired through laser capture microdissection and transcript levels were assessed by cDNA microarray analysis after a linear amplification procedure. In ad- dition to increased expression levels of AMACR and decreased levels of GSTP1 in cancerous epithelium, we determined that transcripts encoding the stearoyl-CoA desaturase (SCD) gene were downregulated. SCD encodes a key rate-limiting enzyme involved in the synthesis of monounsaturated fatty acids. 20 Quantitative PCR measured a reduction of SCD transcripts in 10 of 12 micro- dissected carcinoma samples relative to benign epithelium. Immu- nohistochemical evaluation of 69 radical prostatectomy cases con- firmed a significant reduction or complete lack of SCD protein expression in prostate carcinoma relative to benign epithelium and an overexpression of FAS in the majority of cancers. Our results indicate that loss of SCD expression is a frequent event in prostate adenocarcinoma. Abbreviations: aRNA, amplified antisense RNA; AMACR, -methyla- cyl-CoA racemase; CaP, prostate cancer; FAS, fatty acid synthase; GSTP1, glutathione-S-transferase ; H&E, hematoxylin and eosin; LCM, laser capture microdissection; PPAR, peroxisome proliferator activated recep- tor; SCD, stearoyl-CoA desaturase. Grant sponsor: NIH; Grant number: CA97186, DK65204; Basic Sci- ences Training; Grant number: T32-DK007779. *Correspondence to: Division of Human Biology, Mailstop D4-100, Fred Hutchinson Cancer Research Center, 1100 Fairview Avenue, Seattle, WA 98109. Fax: 206-667-3377. E-mail: [email protected]. Received 23 March 2004; Accepted 4 October 2004 DOI 10.1002/ijc.20773 Published online 17 December 2004 in Wiley InterScience (www. interscience.wiley.com). Int. J. Cancer: 114, 563–571 (2005) © 2004 Wiley-Liss, Inc. Publication of the International Union Against Cancer

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Loss of stearoyl-CoA desaturase expression is a frequent event in prostate carcinomaStacy Moore1, Beatrice Knudsen2, Lawrence D. True3, Sarah Hawley2, Ruth Etzioni2, Christian Wade1, David Gifford1,Ilsa Coleman1 and Peter S. Nelson1*1Division of Human Biology, Fred Hutchinson Cancer Research Center, Seattle, WA, USA2Division of Public Health Sciences, Fred Hutchinson Cancer Research Center, Seattle, WA, USA3Department of Pathology, University of Washington, Seattle, WA, USA
Prostate carcinogenesis is influenced by genetic alterations result-ing in a biochemical condition that favors cell proliferation andsurvival. Studies of prostate carcinoma using comparativegenomic hybridization and cDNA microarray analysis indicatethat numerous biochemical processes may be affected during cel-lular transformation and progression to an invasive phenotype.Among the consistently observed tumor-associated changes arealterations in fatty acid metabolism that influence diverse cellularactivities such as signaling, energy utilization, and membranefluidity. Increases in fatty acid synthase (FAS) levels have beenshown to be one of the earliest and most frequent molecularalterations in prostate carcinogenesis. We sought to identify tu-mor-associated changes in the expression of genes with functionalroles associated with lipid metabolism. Defined populations ofnormal and neoplastic prostate epithelium were acquired by lasercapture microdissection and transcript levels were measured bycDNA microarray hybridization. We determined that stearoyl-CoA desaturase (SCD) transcripts were downregulated in cancerrelative to normal epithelium. These results were confirmed byquantitative PCR. Further analysis by immunohistochemical eval-uation of radical prostatectomy samples employed a quantitativescoring system with a range of 0–300. The median SCD expressionlevels were 150, 45 and 10 for normal, PIN and carcinoma sam-ples, respectively. Statistically significant differential SCD expres-sion between normal and cancerous epithelium was determined atthe p � 0.001 level, and between PIN and prostate carcinoma atthe p � 0.03 level. Of these cases, 92% overexpressed fatty acidsynthase (FAS) in cancerous cells and 84.7% exhibited the signa-ture of FAS overexpression and SCD loss in prostate carcinoma ascompared to normal prostate epithelium. These results indicatethat loss of SCD expression is a frequent event in prostate adeno-carcinoma, and further supports a role for altered lipid metabo-lism as a factor in the process of carcinogenesis.© 2004 Wiley-Liss, Inc.
Key words: prostate cancer; microarray; microdissection; lipid me-tabolism; stearoyl-CoA desaturase
The development of prostate carcinoma is driven by specificgenetic alterations resulting in a biochemical state that favors cellproliferation and survival. The diversity and complexity of tumorcell aberrations is exemplified by the numerous chromosomalrearrangements and gene expression changes identified using com-prehensive analytical technologies such as comparative genomichybridization and microarray analysis.1–3 The extent of these ge-netic alterations suggests that numerous biochemical processesmay be influenced during cellular transformation and the subse-quent progression to an invasive phenotype. The identification ofconsistent neoplasia-associated alterations involving specific geneproducts or molecular pathways may provide important insightsinto essential events associated with carcinogenesis.
Fatty acid metabolism represents a key process that influencesseveral diverse cellular pathways and characteristics including cellsignaling, energy processing and membrane fluidity.4–6 The enzy-matic and transporter activities that modulate concentrations oflipid substrate molecules represent important regulatory elementsthat modulate cellular proliferation and apoptosis. Several proteinsinvolved in the metabolism of fatty acids have been determined tobe altered in neoplastic prostate cells relative to their normalcounterparts. For example, increases in fatty acid synthase (FAS)levels have been shown to be one of the earliest and most frequentmolecular alterations in prostate carcinogenesis.3,7 FAS plays a
pivotal role in the de novo biosynthesis of fatty acids.8 FASexpression in primary prostate carcinomas is predictive of cancerprogression,9 and the inhibition of FAS activity induces cell deathin FAS-overexpressing cancer cell lines and prolongs survival ofhost animals implanted with tumor xenografts.10–13 Increased lev-els of transcripts encoding the �-methylacyl-CoA-racemase (AM-ACR) enzyme have also been shown to occur frequently in pros-tate carcinoma.14–17 AMACR regulates the peroxisomal�-oxidation of dietary branched chain fatty acids, a biochemicalprocess that is plausibly related to prostate carcinogenesis throughthe production of pro-carcinogenic oxidative intermediates.17 Invitro studies have shown that branched fatty acids in dairy and beefproducts, food sources with epidemiological associations withprostate cancer risk, markedly enhance AMACR expression inprostate cancer cells.18 Immunohistochemical studies of AMACRprotein expression have demonstrated a relationship between AM-ACR protein expression and cancer grade.19
Due to the importance of lipid metabolism in prostate carcino-genesis, we sought to identify alterations in the expression levelsof additional genes with functional roles associated with lipidmetabolism that had consistent alterations in mRNA levels be-tween primary prostate carcinomas and corresponding non-neo-plastic secretory cells. Defined populations of normal and neoplas-tic prostate epithelium were acquired through laser capturemicrodissection and transcript levels were assessed by cDNAmicroarray analysis after a linear amplification procedure. In ad-dition to increased expression levels of AMACR and decreasedlevels of GSTP1 in cancerous epithelium, we determined thattranscripts encoding the stearoyl-CoA desaturase (SCD) gene weredownregulated. SCD encodes a key rate-limiting enzyme involvedin the synthesis of monounsaturated fatty acids.20 QuantitativePCR measured a reduction of SCD transcripts in 10 of 12 micro-dissected carcinoma samples relative to benign epithelium. Immu-nohistochemical evaluation of 69 radical prostatectomy cases con-firmed a significant reduction or complete lack of SCD proteinexpression in prostate carcinoma relative to benign epithelium andan overexpression of FAS in the majority of cancers. Our resultsindicate that loss of SCD expression is a frequent event in prostateadenocarcinoma.
Abbreviations: aRNA, amplified antisense RNA; AMACR, �-methyla-cyl-CoA racemase; CaP, prostate cancer; FAS, fatty acid synthase; GSTP1,glutathione-S-transferase �; H&E, hematoxylin and eosin; LCM, lasercapture microdissection; PPAR, peroxisome proliferator activated recep-tor; SCD, stearoyl-CoA desaturase.
Grant sponsor: NIH; Grant number: CA97186, DK65204; Basic Sci-ences Training; Grant number: T32-DK007779.
*Correspondence to: Division of Human Biology, Mailstop D4-100,Fred Hutchinson Cancer Research Center, 1100 Fairview Avenue, Seattle,WA 98109. Fax: 206-667-3377. E-mail: [email protected].
Received 23 March 2004; Accepted 4 October 2004DOI 10.1002/ijc.20773Published online 17 December 2004 in Wiley InterScience (www.
interscience.wiley.com).
Int. J. Cancer: 114, 563–571 (2005)© 2004 Wiley-Liss, Inc.
Publication of the International Union Against Cancer

Material and methodsProstate tissue acquisition, microdissection, RNA extraction andaRNA amplification
Prostate tissue was obtained from patients undergoing radicalprostatectomy in accordance with an IRB-approved collectionprotocol. Tissue was snap-frozen in a liquid nitrogen/isopentanebath and cryopreserved in embedding medium at �80°C. Patho-logical grading was assigned using H&E-stained sections. Sevenmicron thick sections were cut from frozen tissue blocks, appliedto uncharged microscope slides, fixed in 95% ethanol/5% dieth-ylpyrocarbonate (DEPC)-treated water, and stored desiccated at�80°C. Before microdissection, sections were rehydrated inRNase-free water and stained with HistoGene Stain Solution (Ar-turus, Mountain View, CA) for 30 sec. After rinsing in RNase-freewater, sections were dehydrated using 4 steps of 10 sec each in100% ethanol, 100% ethanol, xylene and xylene, followed byair-drying. Microdissection was carried out using a PixCell II lasercapture microdissection (LCM) system, following the manufactur-er’s protocols (Arcturus). Approximately 2,000 neoplastic andnormal prostate epithelial cells were separately collected fromeach patient sample and placed into 30 �l of sample lysis buffer(Arcturus).
Total RNA was extracted and DNase-treated from each LCMsample using a spin column isolation method, according to themanufacturer’s protocols (Nanoprep kit, Stratagene, La Jolla, CA).RNA was eluted in 20 �l of RNase-free water and vacuum-evaporated to a volume of 10 �l. T7-based RNA amplification wascarried out using a MessageAmp aRNA amplification kit (Ambion,Austin, TX) according to the manufacturer’s suggested protocol.The entire first round of amplified RNA (aRNA) was used as atemplate for a second round of amplification. Two rounds ofamplification yielded 20–100 �g of aRNA.
Gene expression analysis by cDNA microarray hybridizationA non-redundant set of approximately 6,200 cDNA clones was
identified from the Prostate Expression DataBase (PEDB), a publicsequence repository of Expressed Sequence Tag (EST) data de-rived from human prostate cDNA libraries.21 Microarrays wereconstructed as we have described previously; each cDNA is rep-resented twice per array.22,23 cDNA probes were synthesized from2 �g of aRNA amplified from normal and neoplastic epitheliumusing Cy3-dCTP or Cy5-dCTP (Amersham Bioscience, Piscat-away, NJ) fluorescent dyes. Probes were competitively hybridizedto microarrays under a coverslip for 16 hr at 63°C. Fluorescentarray images were collected for Cy3 and Cy5 using a GenePix4000A fluorescent scanner (Axon Instruments, Foster City, CA),and image intensity data were extracted and analyzed using GE-NEPIX PRO 3.0 microarray analysis software. Each experimentwas repeated with a switch in fluorescent labels to account for dyeeffects.
Data analysis was carried out as we have described previously.23
Briefly, data were filtered to remove values from poorly hybridizedcDNAs either flagged by the GenePix software, or exhibitingintensity levels �2 SD above the background local to each spot.Intensity ratios for each cDNA hybridized with probes derivedfrom the neoplastic and normal samples were calculated as log2(neoplastic/normal). Data from the 4 replicate cDNAs for eachexperiment were combined and the average log2 ratios were usedfor comparative analyses. cDNAs with average log2 ratios of �1or ��1 (2-fold change) were classified as differentially-expressedbetween normal and neoplastic epithelium.
Quantitative PCRcDNA was generated from 1–5 �g aRNA, or 30 �g total control
RNA using 2 �g random hexamers for priming reverse transcrip-tion by SuperScript II (400 U per reaction; Invitrogen, Carlsbad,CA). Primers and salts were removed using a Microcon 30 filter(Millipore, Bedford, MA). Quantitative PCR (QPCR) reactionswere done in triplicate per experiment, using approximately 5 ng
of cDNA, 0.1–0.3 �M of each primer, and SYBR Green PCRmaster mix (Applied Biosystems, Foster City, CA) in a 50 �Lreaction volume. cDNA generation and PCR amplifications wererepeated three times. Reactions were carried out and analyzedusing an Applied Biosystems 7700 sequence detector. Sampleswere normalized to the cycle threshold value obtained during theexponential amplification of GAPDH. Expression levels of AM-ACR, GSTP1 and SCD were calculated. Values are reported as theratio of gene expression in neoplastic to normal epithelium. Errorbars represent the error of the mean from the 3 experiments. Primersets were tested using serial 10-fold dilutions of template. For the10-fold dilutions, the difference in threshold cycle number wasapproximately 3.2, indicating high PCR efficiency. Control reac-tions with RNA or water as template did not produce significantamplification products. Amplification of a single PCR-product perreaction was monitored by generation of a single dissociationcurve. The sequences of primers used in our study were: GAPDHforward, 5�-CCTCAACGACCACTTTGTCA-3�; GAPDH reverse,5�-TTACTCCTTGGAGGCCATGT-3�; AMACR forward, 5�-AGT-AACTCGGGGCCTGTTTC-3�; AMACR reverse, 5�-CTGGATGT-TGCTGTGTGTTG-3�; GSTP1 forward, 5�-AGGCAAGACCTTCA-TTGTGG-3�; GSTP1 reverse, 5�-TCATGGATCAGCAGCAAGTC-3�; SCD forward, 5�-TTGGAGAAGCGGTGGATAAC-3�; SCDreverse, 5�-AAAAATCCCACCCAATCACA-3�.
Northern analysisA human multiple tissue Northern blot was obtained from
CLONTECH and hybridized with SCD cDNA probe labeled with[�-32P]dCTP by random priming using the Rediprime II randomprimer labeling system (Amersham) according to the manufactur-er’s protocol. Filters were imaged and quantitated using a phos-phor-capture screen and ImageQuant software (Molecular Dynam-ics).
Tissue microarray immunohistochemistryA tissue microarray (TMA) was constructed with 0.6 mm
diameter triplicate cores of tumor and normal tissue from 69prostatectomy cases containing Gleason sum score of 6, 7 or 8cancer using an ATA-27 tissue arrayer (Beecher Instruments,Sun Prairie, WI). The tissue was obtained under an IRB ap-proved protocol after removal of patient identifiers. For immu-nohistochemical analysis, the TMA slide was deparaffinizedand rehydrated. For SCD antigen retrieval, the tissue was firsttreated with 0.0005% proteinase K (Boehringer Mannheim,Indianapolis, IN) in 50 mM Tris buffer pH 8.0 and then placedin a steamer for 20 minutes. FAS antigen was retrieved in themicrowave for 15 min in citrate buffer pH 6.0. The SCDpolyclonal rabbit antibody (Alpha Diagnostic International Inc.,San Antonio, TX ) was diluted 1:50 in PBS/1% BSA and theFAS monoclonal antibody (clone 23, BD Bioscience, Pharmin-gen, San Diego, CA) was diluted 1:2,000. Both antibodies wereincubated with slides at room temperature for 60 min. Antibodystaining was developed with the VECTOR ABC kit.
Stained slides were imaged using the BLISS system (BachusLaboratories, Lombard, IL). Cores were visualized with a Webbased image display at 20 magnification. Tumor and normaltissue was assessed on each core using an H&E-stained sequentialarray slide. The tumor was graded and the normal tissue wasevaluated for the degree of atrophy. During initial phases of theanalysis we noted maximal SCD expression in fully differentiatedsecretory epithelium. Atrophic epithelium demonstrated decreasedor absent SCD immunoreactivity presumably due to the absence ofdifferentiated secretory epithelial cells. Cores containing partiallyor fully atrophic epithelium were therefore excluded from the finalanalysis. SCD expression was scored as the percentage of epithe-lial cells with granular cytoplasmic SCD expression. Basal epithe-lial cells were considered internal negative controls.
564 MOORE ET AL.

Interpretation and statistical analysis of immunohistochemistrystaining
A scoring system incorporating staining intensity parametersand percentage of cell reactivity was used to facilitate the com-parison of SCD and FAS expression in normal epithelium, PIN andinvasive carcinoma. Each tissue core was used as an independentsample. Cores comprised of both normal epithelium and PIN orcancer were given a separate score for each histology. Four cate-gories of staining were defined: no stain, faint, moderate andintense. The percent of cells falling into each of these categorieswas estimated by the study pathologist and used to compute asingle score as follows: Score 0 (percent with no stain) � 1 (percent with faint stain) � 2 (percent with moderate stain) �3 (percent with intense stain). The minimal score was 0 and themaximal score was 300. When 2 tissue types (e.g., PIN, cancer)were present in the same TMA score, a score was calculated foreach tissue type.
The box plots summarize the distribution of Score for eachgroup of cores, Normal, PIN, Cancer. The “box” shows the medianScore as a dashed line and the 25th percentile and 75th percentileof the Score distribution as the lower and upper parts of the box.The median is the Score at the 50% percentile: half of all cores geta Score higher than the median, and half get a Score lower. The25th (75th) percentile is the point at which 25% (75%) of the coresscore lower (and 75% or 25% score higher). Thus, the area in the“box” represents the middle 50% of the cores. The “whiskers”shown above and below the boxes represent the largest and small-est observed Scores that are �1.5 box lengths from the end of thebox. Circles (outliers) represent cores with Scores �1.5 boxlengths from the end of the box.
We used generalized estimating equations (GEE) to conduct aregression analysis of the association between score (dependentvariable) and tissue type (normal epithelium, PIN, cancer). Themodels fit were of the form: Scope Intercept � Slope*(tissue
FIGURE 1 – Analysis of SCD expression in microdissected normal and neoplastic prostate epithelium. (a) cDNA microarray expression valuesfor glutathione-S-transferase � (GSTP1), �-methylacyl-CoA racemase (AMACR) and stearoyl-CoA-desaturase (SCD) in 12 matched micro-dissected pairs of normal and neoplastic prostate epithelium. The scale represents fold-change in cancer epithelium relative to normal epithelium.(b) Quantitative PCR expression values for GSTP1 (hatched bar), AMACR (open bar), and SCD (solid bar) in the same 12 matched pairs ofnormal and neoplastic prostate epithelium used in the microarray studies.
565SCD LOSS IN PROSTATE CARCINOMA
type).24,25 This technique accounts for the correlation that arisesamong observations from the same patient.
ResultscDNA array expression profiling of microdissected prostateepithelium identifies decreased SCD expression in prostatecarcinoma
The cellular architecture and composition of the human prostategland is complex, comprising several different cell populationspresent in varying numbers in any particular region. The bulktissue preparations commonly used in studying epithelial tumors,such as the prostate, often contain large numbers of ‘contaminat-ing’ cells, including fibromuscular stromal components, leuko-cytes and endothelial cells. To facilitate the comparative analysisof gene expression between normal and neoplastic prostate epithe-lium, we used laser capture microdissection methods to acquirepure populations of epithelial cells. Approximately 2,000 epithelialcells with normal or neoplastic morphology were captured fromeach of 12 radical prostatectomy specimens. After total RNAisolation, a linear RNA amplification procedure was employed togenerate sufficient RNA for labeling and cDNA microarray hy-bridization.26 Several genes previously identified to be differen-tially expressed between normal prostate epithelium and prostatecarcinoma were found to be altered in this analysis. These includedincreased expression of AMACR in carcinoma, and decreasedexpression of GSTP1 in cancer relative to benign cells (Fig. 1a).Due to the recognized importance of enzymes regulating lipidmetabolism, we searched the expression profiles for genes withfunctional roles known to be involved in lipid biosynthesis orutilization and identified the decreased expression of transcriptsencoding stearoyl-coenzyme A desaturase (SCD) in 8 of 12 car-cinoma samples with 5 of 12 exhibiting more than a 50% reductionin expression (Fig. 1a).
QPCR analysis of genes differentially expressed in neoplasticprostate epithelium
To confirm the differential expression of SCD, GSTP1 andAMACR in normal and neoplastic prostate epithelium, we carriedout quantitative PCR analysis on total RNA samples obtained bymicrodissection. The QPCR analysis confirmed the microarrayresults, though in general the differential expression measurementswere greater as measured by QPCR relative to microarray values(Fig. 1b). GSTP1 expression was reduced in cancer epitheliumrelative to normal epithelium in all 12 samples analyzed. AMACRexpression was increased in cancer epithelium relative to normalepithelium in all cases. Transcripts encoding SCD were downregu-lated in 10 of 12 carcinoma samples relative to normal epitheliumconfirming the microarray results. SCD transcript levels weregenerally reduced 3–4-fold in neoplastic epithelium with a differ-ential expression range between 2-fold and 30-fold (Fig. 1b).
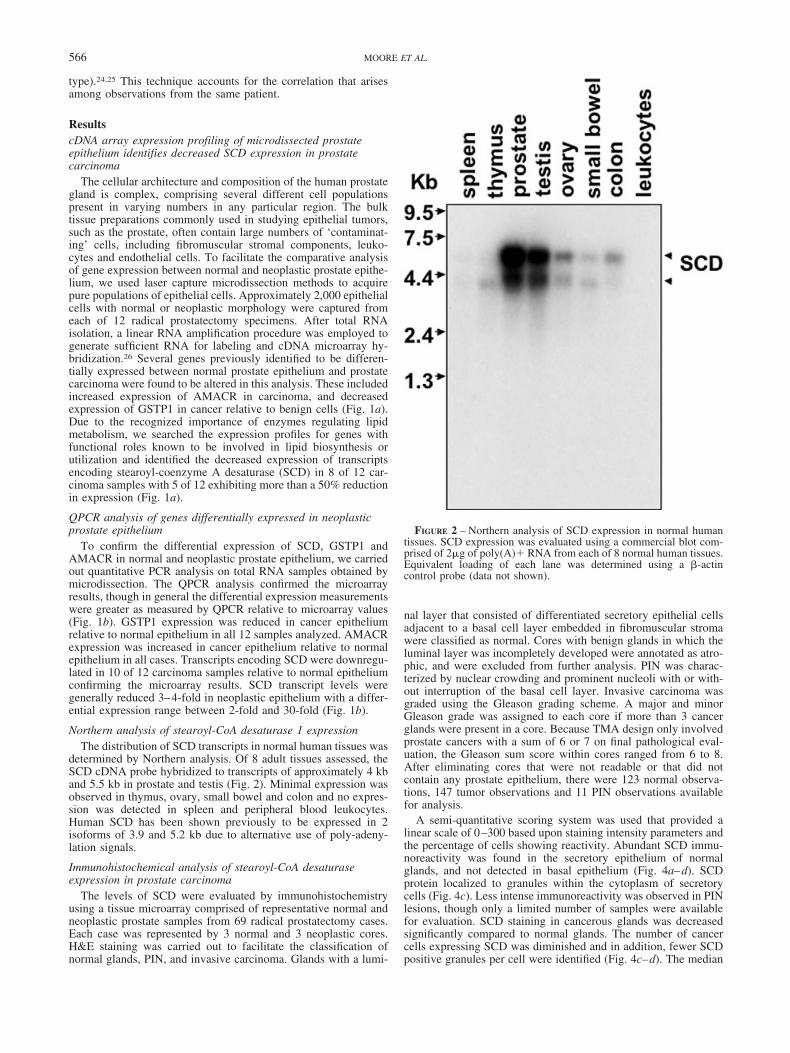
Northern analysis of stearoyl-CoA desaturase 1 expressionThe distribution of SCD transcripts in normal human tissues was
determined by Northern analysis. Of 8 adult tissues assessed, theSCD cDNA probe hybridized to transcripts of approximately 4 kband 5.5 kb in prostate and testis (Fig. 2). Minimal expression wasobserved in thymus, ovary, small bowel and colon and no expres-sion was detected in spleen and peripheral blood leukocytes.Human SCD has been shown previously to be expressed in 2isoforms of 3.9 and 5.2 kb due to alternative use of poly-adeny-lation signals.
Immunohistochemical analysis of stearoyl-CoA desaturaseexpression in prostate carcinoma
The levels of SCD were evaluated by immunohistochemistryusing a tissue microarray comprised of representative normal andneoplastic prostate samples from 69 radical prostatectomy cases.Each case was represented by 3 normal and 3 neoplastic cores.H&E staining was carried out to facilitate the classification ofnormal glands, PIN, and invasive carcinoma. Glands with a lumi-
nal layer that consisted of differentiated secretory epithelial cellsadjacent to a basal cell layer embedded in fibromuscular stromawere classified as normal. Cores with benign glands in which theluminal layer was incompletely developed were annotated as atro-phic, and were excluded from further analysis. PIN was charac-terized by nuclear crowding and prominent nucleoli with or with-out interruption of the basal cell layer. Invasive carcinoma wasgraded using the Gleason grading scheme. A major and minorGleason grade was assigned to each core if more than 3 cancerglands were present in a core. Because TMA design only involvedprostate cancers with a sum of 6 or 7 on final pathological eval-uation, the Gleason sum score within cores ranged from 6 to 8.After eliminating cores that were not readable or that did notcontain any prostate epithelium, there were 123 normal observa-tions, 147 tumor observations and 11 PIN observations availablefor analysis.
A semi-quantitative scoring system was used that provided alinear scale of 0–300 based upon staining intensity parameters andthe percentage of cells showing reactivity. Abundant SCD immu-noreactivity was found in the secretory epithelium of normalglands, and not detected in basal epithelium (Fig. 4a–d). SCDprotein localized to granules within the cytoplasm of secretorycells (Fig. 4c). Less intense immunoreactivity was observed in PINlesions, though only a limited number of samples were availablefor evaluation. SCD staining in cancerous glands was decreasedsignificantly compared to normal glands. The number of cancercells expressing SCD was diminished and in addition, fewer SCDpositive granules per cell were identified (Fig. 4c–d). The median
FIGURE 2 – Northern analysis of SCD expression in normal humantissues. SCD expression was evaluated using a commercial blot com-prised of 2�g of poly(A)� RNA from each of 8 normal human tissues.Equivalent loading of each lane was determined using a �-actincontrol probe (data not shown).
566 MOORE ET AL.

SCD expression measured 150, 45 and 10 for normal prostate, PINand cancer, respectively (Fig. 4e). In this limited sample set, therewas no correlation between SCD expression and the Gleason sumscore in cancer-containing cores. Statistically significant differen-tial SCD expression between normal prostate and prostate carci-noma was determined at the p 0.001 level, and between PIN andprostate carcinoma at the p 0.03 level. The comparison betweennormal prostate and PIN and between PIN and prostate carcinomawas statistically significant at the p 0.05 level.
We observed weak stromal cell reactivity with the SCD-1 anti-body in most tissue samples. It is likely that this reactivity repre-sents non-specific binding of the SCD-1 antibody. A control,leaving out the primary antibody was negative. We cannot com-pletely exclude the possibility that a small a mount of SCD-1expression occurs in stromal cells. If this is the case, there is nodifference between normal and cancer stroma. It is therefore un-likely that SCD-1 in the stroma is related to the presence or thebiologic behavior of prostate cancer.
To assess the association between SCD and FAS expression, aparallel TMA slide was stained with the FAS antibody. Data forFAS and SCD expression were available for 85 cancer-containingcores. In 78 of 85 cores (92%), over expression of FAS (totalscore �100) was observed. Of the 78 FAS overexpressing cores,72 (92%) demonstrated decreased SCD expression (total score �100) occurred in cancerous compared to normal glands (Fig. 4f).A 95% confidence interval for the percent of cores staining posi-tive for FAS and negative for SCD extends from 0.83–0.97. Theseresults demonstrated that the vast majority of the tumor samplesstudied exhibited FAS overexpression and loss of SCD.
Discussion
Stearoyl-CoA desaturase (SCD; EC 1.14.99.5) is an iron-con-taining enzyme involved in mediating the rate-limiting step ofunsaturated fatty acid biosynthesis. SCD catalyzes the �-9-cisdesaturation of palmitic acid (C16:0) and stearic acid (C18:0) tothe monounsaturated fatty acids palmitoleic acid (C16:1) and oleicacid (C18:1) respectively (Fig. 3).27,28 These monounsaturatedfatty acids are principle components of membrane phospholipids,triglycerides and cholesterol esters.29 The ratio of saturated andunsaturated fatty acids has been implicated in the regulation of cellgrowth and differentiation through effects on the physical proper-ties of cell-membrane fluidity and signal transduction.4,30 Smallchanges in this ratio, and in the amount of available unsaturatedfatty acids can affect the ability of the cell to respond to externalstimuli. Alterations in cellular fatty acid composition have beenassociated with a range of pathologies including neurological andvascular diseases, diabetes, obesity, hypertension and cancer.29
In humans, a single functional gene encoding SCD has beenidentified that maps to chromosome 10 and produces 2 transcriptsof 3.9 and 5.2kb through the use of alternative polyadenylationsignals.31 The mouse expresses 4 SCD isoforms designated Scd1,Scd2, Scd3 and Scd4.32 Murine Scd1 and Scd2 share 85% and82% nucleotide sequence identity with human SCD, respectively.In vivo and in vitro studies have demonstrated that SCD geneexpression is regulated normally by dietary constituents, hormonalfactors and developmental processes. SCD expression is nega-tively regulated by polyunsaturated fatty acids, cholesterol andarachidonic acid as measured by decreased SCD promoter activity,mRNA and protein levels.33 The expression of SCD is positivelyregulated by a lipogenic diet through induction of sterol responseelement binding protein (SREBP), a major coordinate regulator ofmultiple enzymes involved in fatty acid metabolism.33 TGF-�,retinoids and peroxisome proliferator activated receptor (PPAR)agonists such as Clofibrate and Gemfibrozil have also been shownto induce SCD expression.34–36 Consistent with these observa-tions, a sterol response element (SRE) and peroxisome proliferatorresponse element (PPRE) have been identified in the SCD pro-moter region.36 In vitro studies of SCD overexpression resulted an
increase in the plasma membrane content of monounsaturated fattyacids relative to saturated fatty acids in addition to alterations inmembrane organization with decreases in membrane-ordered re-gions.37
Recent studies describing the molecular and physiological ef-fects of SCD deficiency in animal models are enlightening. Ntambiet al.38 demonstrated that mice with a targeted disruption of Scd1exhibited reduced body adiposity, increased insulin sensitivity, andresistance to diet-induced weight gain. The protection from obesityinvolved increased energy expenditure and increased oxygen con-sumption. The expression of several genes involved in the oxida-tion of lipids was upregulated, whereas lipid synthesis genes weredownregulated. These observations suggest that a consequence ofScd1 deficiency is an activation of lipid oxidation in addition toreduced triglyceride synthesis and storage. Studies of murine mod-els with leptin deficiency demonstrated that leptin specificallyrepresses Scd1 transcript levels and enzymatic activity resulting inlean and hypermetabolic animals.39 In the context of our results, anincreased incidence of prostate carcinoma or other malignancies inmice with germline alterations in Scd expression was not de-scribed. Mice express 4 isoforms of Scd, however, and redundanteffects may mask some pathological features of Scd1 loss in theprostate.
How might a reduction of SCD expression contribute to thedevelopment of human prostate carcinoma? At this time, sev-eral possible mechanisms are intriguing, but speculative. Onecellular attribute implicated in neoplastic growth involves theregulation of cellular signaling pathways through targeted par-
FIGURE 3 – Biochemical pathway of fatty acid metabolism involvingstearoyl-CoA-desaturase. Stearoyl-CoA-desaturase (SCD) catalyzesthe desaturation of the unsaturated fatty acids palmitate (C16:0) andstearate (C18:0) to the 9 monounsaturated fatty acids palmitoleate(C16:1) and oleate (C18:1), respectively.59 In prostate carcinoma,increased expression of FAS (vertical dashed arrows) and decreasedexpression of SCD (vertical dashed arrow) would produce increasedlevels of palmitate. FAS, fatty acid synthase; SCD, stearoyl-CoA-desaturase; GPAT, glycerol-3-phosphate acyltransferase.
567SCD LOSS IN PROSTATE CARCINOMA

FIGURE 4.
568 MOORE ET AL.

titioning of signaling molecules to regions of the plasma mem-brane. Several signal transducers responding to receptor-generated signals are localized to specialized plasma membranedomains comprised of cholesterol and glycosphingolipids.40
These domains are detergent insoluble and have been termedrafts.41 The spatial organization and concentration of selectedproteins that partition to rafts may serve to increase the effi-ciency and specificity of signal transduction by facilitatingprotein interactions and preventing inappropriate cross-talk be-tween pathways.40 Importantly, a significant characteristic dic-tating the partitioning of signaling proteins to lipid raft mi-crodomains involves the addition of palmitate to cysteineresidues, a reaction termed palmitoylation.42,43 Numerous sig-naling proteins have been shown to undergo regulation bypalmitoylation including Wnt, RAS and regulators of G proteinsignaling (RGS proteins).44 – 47 Loss of SCD expression wouldbe expected to increase cellular palmitate levels. Further en-hancement of the palmitate pool in neoplastic prostate epithe-lium would also result from overexpression of the fatty acidsynthase enzyme (Fig. 3), a finding observed in a majority ofprimary prostate carcinomas.7,48
It has been shown that a cell survival signal mediated byepidermal growth factor receptor (EGFR) activation of thePI3K/Akt pathway is operative in prostate cancer cells, and isdependent upon localization to lipid rafts.49 Pharmacologicaldisruption of lipid rafts inhibited signaling via EGFR and led toapoptosis. Overexpression of SCD also leads to a reduction ofdetergent insoluble lipid rafts,37 and thus could result in loss ofcell survival signals. Conversely, a loss of SCD expressionmight be expected to provide an increase in lipid rafts with theability to activate anti-apoptotic pathways. Additional evidencesupporting a role for the influence of cellular lipid compositionon signaling pathways is provided by studies of tumor cellsensitivity to cytotoxicity mediated by tumor necrosis factor(TNF). Murine tumor cell lines engineered to express the yeast�-9-desaturase gene, Ole 1, contained increased plasma mem-brane levels of unsaturated oleic acid and decreased levels ofsaturated stearic acid.30 These altered fatty acid ratios wereassociated with increased membrane fluidity and a markedlyenhanced sensitivity to killing by TNF. One mechanism pro-
posed for TNF cytotoxicity involves lipid peroxidation result-ing from increased production of oxygen free radicals in themitochondria of TNF-sensitive cells.50 Membranes containingextra double bonds in the form of unsaturated fatty acids maybecome more accessible to the action of free radicals. It followsthat tumor cells expressing SCD at a low level, with saturatedrather than unsaturated fatty acids comprising membrane struc-ture, may have a survival advantage toward host anti-tumormechanisms including TNF-mediated cell killing.
Several biochemical regulators of tumor cell growth havealso been shown to modulate SCD expression. TGF-� andretinoids increase cellular levels of SCD and also inhibit tumorgrowth through effects on cell cycle, cell differentiation andapoptosis.51,52 Our results open the possibility that the loss ofSCD expression in tumor cells may serve to modulate thesegrowth inhibitory effects. Ligands of the PPAR family of nu-clear receptors also upregulate SCD expression, and showgrowth inhibitory activities toward a wide variety of tumortypes in vitro and in vivo.53 Ligands of PPAR in particularhave been the focus of study in prostate carcinoma and dem-onstrate substantial growth inhibitory effects toward prostatecancer cell lines.54 Phase II clinical trials of troglitazone, aPPAR ligand used for treating Type 2 diabetes, exhibitedmeasurable biological activities in patients with advanced pros-tate carcinoma.54 It is plausible that a component of PPAR affects may be mediated through SCD activity, and downregu-lation of SCD expression in neoplastic cells could abrogategrowth suppression. The significance of PPARs in prostatecarcinogenesis remains controversial,55 however, and thismechanism may represent only one of several pathways influ-encing SCD activity. Although signaling regulators of tumorcell growth have been shown to modulate SCD expression,alterations in SCD levels influence signaling pathways impor-tant for cell growth and metabolism. SCD deficiency enhancessignaling through the insulin receptor (IR) pathway as demon-strated by an increase in basal phosphorylation of the IR andinsulin receptor substrates-1 and -2 (IRS-1 and IRS-2), in-creased association of IRS-1 and IRS-2 with PI3-kinase, andincreased phosphorylation of Akt.56 Activation of the PI3-kinase/Akt pathway has been shown to be an important pathwayregulating proliferation, apoptosis and growth in many cancersincluding prostate carcinoma.57
Our observation that reduced SCD expression is associated withprostate carcinoma adds another member to a growing list ofcancer-associated alterations in components of lipid metabolismthat includes FAS,48 AMACR15 and sterol regulatory elementbinding protein (SREBP).58 The relationships between these pro-teins in the context of prostate carcinogenesis remains to bedetermined. Our findings offer many testable hypothesis regardingthe role of known SCD substrates and products in regulatingcellular growth. It is also possible that the desaturase activity ofSCD may operate on additional substrates not yet known to beassociated with carcinogenesis.
Acknowledgements
We wish to thank K. Adolphson for assistance with immuno-histochemistry. This work was supported by NIH grants CA97186and DK65204 to P.S.N. S.M. was supported by a Basic SciencesTraining Grant in Urology (T32-DK007779).
References
1. Visakorpi T, Kallioniemi AH, Syvanen AC, Hyytinen ER, Karhu R,Tammela T, Isola JJ, Kallioniemi OP. Genetic changes in primary andrecurrent prostate cancer by comparative genomic hybridization. Can-cer Res 1995;55:342–7.
2. Nelson PS, Clegg N, Arnold H, Ferguson C, Bonham M, White J,Hood L, Lin B. The program of androgen-responsive genes in
neoplastic prostate epithelium. Proc Natl Acad Sci USA 2002;99:11890 –5.
3. Welsh JB, Sapinoso LM, Su AI, Kern SG, Wang-Rodriguez J,Moskaluk CA, Frierson HF Jr., Hampton GM. Analysis of geneexpression identifies candidate markers and pharmacological targetsin prostate cancer. Cancer Res 2001;61:5974–8.
FIGURE 4 – Immunohistochemical analysis of SCD protein expres-sion. (a) Section of formalin fixed paraffin embedded tissue stainedwith the SCD antibody. Normal epithelial glands (NE) are embeddedin a fibromuscular stroma and express SCD in secretory epithelialcells, as indicated by a brown immunohistochemical staining reaction(100). (b) High power view of a normal gland showing SCD ex-pression in the cytoplasm and localized to cytoplasmic granules(600). (c) Decreased SCD expression in prostate cancer. Low powerview of an SCD negative prostate cancer with a GS of 6 (CE)infiltrating in between SCD positive normal glands (NE). Magnifica-tion 100. (d) Higher power view of granular SCD immunoreac-tivity predominantly in the normal secretory epithelium with loss ofexpression in adjacent cancer epithelium. Magnification 400. (e)Quantitative analysis of SCD expression in normal and neoplasticprostate tissue. Boxes show the 25–75th percentiles and median levelof expression (center dashed line). The whiskers extend 1.5 times theInter-quartile range from the end of the box. (f) FAS expression inprostate cancer cores. FAS and SCD expression were scored using thesame scoring scheme. A score of �100 indicated FAS overexpressionand a score of � 100 the loss of SCD expression.
569SCD LOSS IN PROSTATE CARCINOMA

4. Stubbs CD, Smith AD. Essential fatty acids in membrane: physicalproperties and function. Biochem Soc Trans 1990;18:779–81.
5. Faergeman NJ, Knudsen J. Role of long-chain fatty acyl-CoA esters inthe regulation of metabolism and in cell signaling. Biochem J 1997;323:1–12.
6. Tang DG, La E, Kern J, Kehrer JP. Fatty acid oxidation and signalingin apoptosis. Biol Chem 2002;383:425–42.
7. Swinnen JV, Roskams T, Joniau S, Van Poppel H, Oyen R, Baert L,Heyns W, Verhoeven G. Overexpression of fatty acid synthase is anearly and common event in the development of prostate cancer. Int JCancer 2002;98:19–22.
8. Kuhajda FP. Fatty-acid synthase and human cancer: new perspectiveson its role in tumor biology. Nutrition 2000;16:202–8.
9. Shurbaji MS, Kalbfleisch JH, Thurmond TS. Immunohistochemicaldetection of a fatty acid synthase (OA-519) as a predictor of progres-sion of prostate cancer. Hum Pathol 1996;27:917–21.
10. Pizer ES, Chrest FJ, DiGiuseppe JA, Han WF. Pharmacologicalinhibitors of mammalian fatty acid synthase suppress DNA replicationand induce apoptosis in tumor cell lines. Cancer Res 1998;58:4611–5.
11. Li JN, Gorospe M, Chrest FJ, Kumaravel TS, Evans MK, Han WF,Pizer ES. Pharmacological inhibition of fatty acid synthase activityproduces both cytostatic and cytotoxic effects modulated by p53.Cancer Res 2001;61:1493–9.
12. Pizer ES, Wood FD, Heine HS, Romantsev FE, Pasternack GR,Kuhajda FP. Inhibition of fatty acid synthesis delays disease progres-sion in a xenograft model of ovarian cancer. Cancer Res 1996;56:1189–93.
13. De Schrijver E, Brusselmans K, Heyns W, Verhoeven G, Swinnen JV.RNA interference-mediated silencing of the fatty acid synthase geneattenuates growth and induces morphological changes and apoptosisof LNCaP prostate cancer cells. Cancer Res 2003;63:3799–804.
14. Xu J, Stolk JA, Zhang X, Silva SJ, Houghton RL, Matsumura M,Vedvick TS, Leslie KB, Badaro R, Reed SG. Identification of differ-entially expressed genes in human prostate cancer using subtractionand microarray. Cancer Res 2000;60:1677–82.
15. Rubin MA, Zhou M, Dhanasekaran SM, Varambally S, Barrette TR,Sanda MG, Pienta KJ, Ghosh D, Chinnaiyan AM. �-Methylacylcoenzyme A racemase as a tissue biomarker for prostate cancer.JAMA 2002;287:1662–70.
16. Luo J, Duggan DJ, Chen Y, Sauvageot J, Ewing CM, Bittner ML,Trent JM, Isaacs WB. Human prostate cancer and benign prostatichyperplasia: molecular dissection by gene expression profiling. Can-cer Res 2001;61:4683–8.
17. Luo J, Zha S, Gage WR, Dunn TA, Hicks JL, Bennett CJ, Ewing CM,Platz EA, Ferdinandusse S, Wanders RJ, Trent JM, Isaacs WB, et al.Alpha-methylacyl-CoA racemase: a new molecular marker for pros-tate cancer. Cancer Res 2002;62:2220–6.
18. Mobley JA, Leav I, Zielie P, Wotkowitz C, Evans J, Lam YW,L’Esperance BS, Jiang Z, Ho SM. Branched fatty acids in dairy andbeef products markedly enhance alpha-methylacyl-CoA racemase ex-pression in prostate cancer cells in vitro. Cancer Epidemiol Biomar-kers Prev 2033;12:775–83.
19. Kuefer R, Varambally S, Zhou M, Lucas PC, Loeffler M, Wolter H,Mattfeldt T, Hautmann RE, Gschwend JE, Barrette TR, Dunn RL,Chinnaiyan AM, et al. �-Methylacyl-CoA racemase: expression lev-els of this novel cancer biomarker depend on tumor differentiation.Am J Pathol 2002;161:841–8.
20. Miyazaki M, Ntambi JM. Role of stearoyl-coenzyme A desaturase inlipid metabolism. Prostaglandins Leukot Essent Fatty Acids 2003;68:113–21.
21. Hawkins V, Doll D, Bumgarner R, Smith T, Abajian C, Hood L,Nelson PS. PEDB: the Prostate Expression Database. Nucleic AcidsRes 1999;27:204–8.
22. Lin B, White JT, Ferguson C, Wang S, Vessella R, Bumgarner R,True LD, Hood L, Nelson PS. Prostate short-chain dehydrogenasereductase 1 (PSDR1): a new member of the short-chain steroid dehy-drogenase/reductase family highly expressed in normal and neoplasticprostate epithelium. Cancer Res 2001;61:1611–8.
23. Nelson PS, Clegg N, Arnold H, Ferguson C, Bonham M, White J,Hood L, Lin B. The program of androgen-responsive genes in neo-plastic prostate epithelium. Proc Natl Acad Sci USA2002;99:11890–5.
24. Zeger SL, Liang KY. Longitudinal data analysis for discrete andcontinuous outcomes. Biometrics 1986;42:121–30.
25. Zeger SL, Liang KY, Albert PS. Models for longitudinal data: ageneralized estimating equation approach. Biometrics 1988;44:1049–60.
26. Van Gelder RN, von Zastrow ME, Yool A, Dement WC, Barchas JD,Eberwine JH. Amplified RNA synthesized from limited quantities ofheterogeneous cDNA. Proc Natl Acad Sci USA 1990;87:1663–7.
27. Enoch HG, Strittmatter P. Role of tyrosyl and arginyl residues in ratliver microsomal stearyl coenzyme A desaturase. Biochemistry 1978;17:4927–32.
28. Strittmatter P, Spatz L, Corcoran D, Rogers MJ, Setlow B, Redline R.Purification and properties of rat liver microsomal stearyl coenzyme Adesaturase. Proc Natl Acad Sci USA 1974;71:4565–9.
29. Ntambi JM. Regulation of stearoyl-CoA desaturase by polyunsatu-rated fatty acids and cholesterol. J Lipid Res 1999;40:1549–58.
30. Gyorfy Z, Benko S, Kusz E, Maresca B, Vigh L, Duda E. Highlyincreased TNF sensitivity of tumor cells expressing the yeast delta9-desaturase gene. Biochem Biophys Res Commun 1997;241:465–70.
31. Zhang L, Ge L, Parimoo S, Stenn K, Prouty SM. Human stearoyl-CoAdesaturase: alternative transcripts generated from a single gene byusage of tandem polyadenylation sites. Biochem J 1999;340:255–64.
32. Miyazaki M, Jacobson MJ, Man WC, Cohen P, Asilmaz E, FriedmanJM, Ntambi JM. Identification and characterization of murine SCD4,a novel heart-specific stearoyl-CoA desaturase isoform regulated byleptin and dietary factors. J Biol Chem 2003;278:33904–11.
33. Bene H, Lasky D, Ntambi JM. Cloning and characterization of thehuman stearoyl-CoA desaturase gene promoter: transcriptional acti-vation by sterol regulatory element binding protein and repression bypolyunsaturated fatty acids and cholesterol. Biochem Biophys ResCommun 2001;284:1194–8.
34. Miller CW, Waters KM, Ntambi JM. Regulation of hepatic stearoyl-CoA desaturase gene 1 by vitamin A. Biochem Biophys Res Commun1997;231:206–10.
35. Samuel W, Nagineni CN, Kutty RK, Parks WT, Gordon JS, ProutySM, Hooks JJ, Wiggert B. Transforming growth factor-beta regulatesstearoyl coenzyme A desaturase expression through a Smad signalingpathway. J Biol Chem 2002;277:59–66.
36. Miller CW, Ntambi JM. Peroxisome proliferators induce mouse liverstearoyl-CoA desaturase 1 gene expression. Proc Natl Acad Sci USA1996;93:9443–8.
37. Sun Y, Hao M, Luo Y, Liang CP, Silver DL, Cheng C, Maxfield FR,Tall AR. Stearoyl-CoA desaturase inhibits ATP-binding cassettetransporter A1-mediated cholesterol efflux and modulates membranedomain structure. J Biol Chem 2003;278:5813–20.
38. Ntambi JM, Miyazaki M, Stoehr JP, Lan H, Kendziorski CM, YandellBS, Song Y, Cohen P, Friedman JM, Attie AD. Loss of stearoyl-CoAdesaturase-1 function protects mice against adiposity. Proc Natl AcadSci USA 2002;99:11482–6.
39. Cohen P, Miyazaki M, Socci ND, Hagge-Greenberg A, Liedtke W,Soukas AA, Sharma R, Hudgins LC, Ntambi JM, Friedman JM. Rolefor stearoyl-CoA desaturase-1 in leptin-mediated weight loss. Science2002;297:240–3.
40. Moffett S, Brown DA, Linder ME. Lipid-dependent targeting of Gproteins into rafts. J Biol Chem 2000;275:2191–8.
41. Simons K, Ikonen E. Functional rafts in cell membranes. Nature1997;387:569–72.
42. Linder ME, Deschenes RJ. New insights into the mechanisms ofprotein palmitoylation. Biochemistry 2003;42:4311–20.
43. Melkonian KA, Ostermeyer AG, Chen JZ, Roth MG, Brown DA.Role of lipid modifications in targeting proteins to detergent-resistantmembrane rafts. Many raft proteins are acylated, while few areprenylated. J Biol Chem 1999;274:3910–7.
44. Willert K, Brown JD, Danenberg E, Duncan AW, Weissman IL, ReyaT, Yates JR 3rd, Nusse R. Wnt proteins are lipid-modified and can actas stem cell growth factors. Nature 2003;423:448–52.
45. Baker TL, Zheng H, Walker J, Coloff JL, Buss JE. Distinct rates ofpalmitate turnover on membrane-bound cellular and oncogenic H-ras.J Biol Chem 2003;278:19292–300.
46. Hiol A, Davey PC, Osterhout JL, Waheed AA, Fischer ER, Chen CK,Milligan G, Druey KM, Jones TL. Palmitoylation regulates regulatorsof G-protein signaling (RGS) 16 function. I. Mutation of amino-terminal cysteine residues on RGS16 prevents its targeting to lipidrafts and palmitoylation of an internal cysteine residue. J Biol Chem2003;278:19301–8.
47. Casey PJ. Protein lipidation in cell signaling. Science 1995;268:221–5.
48. Rossi S, Graner E, Febbo P, Weinstein L, Bhattacharya N, Onody T,Bubley G, Balk S, Loda M. Fatty acid synthase expression definesdistinct molecular signatures in prostate cancer. Mol Cancer Res2003;1:707–15.
49. Zhuang L, Lin J, Lu ML, Solomon KR, Freeman MR. Cholesterol-rich lipid rafts mediate Akt-regulated survival in prostate cancer cells.Cancer Res 2002;62:2227–31.
50. Grooten J, Goossens V, Vanhaesebroeck B, Fiers W. Cell membranepermeabilization and cellular collapse, followed by loss of dehydro-genase activity: early events in tumour necrosis factor-induced cyto-toxicity. Cytokine, 1993;5:546–55.
51. Siegel PM, Massague J. Cytostatic and apoptotic actions of TGF-� inhomeostasis and cancer. Nat Rev Cancer 2003;3:807–20.
52. Altucci L, Gronemeyer H. The promise of retinoids to fight againstcancer. Nat Rev Cancer 2001;1:181–93.
53. Koeffler HP. Peroxisome proliferator-activated receptor gamma andcancers. Clin Cancer Res 2003;9:1–9.
570 MOORE ET AL.

54. Mueller E, Smith M, Sarraf P, Kroll T, Aiyer A, Kaufman DS, Oh W,Demetri G, Figg WD, Zhou XP, Eng C, Spiegelman BM, et al. Effects ofligand activation of peroxisome proliferator-activated receptor gamma inhuman prostate cancer. Proc Natl Acad Sci USA 2000;97:10990–5.
55. Saez E, Olson P, Evans RM. Genetic deficiency in Pparg does not alterdevelopment of experimental prostate cancer. Nat Med 2003;9:1265–6.
56. Rahman SM, Dobrzyn A, Dobrzyn P, Lee SH, Miyazaki M, NtambiJM. Stearoyl-CoA desaturase 1 deficiency elevates insulin-signalingcomponents and downregulates protein-tyrosine phosphatase 1B inmuscle. Proc Natl Acad Sci USA 2003;100:11110–5.
57. Vivanco I, Sawyers CL. The phosphatidylinositol 3-Kinase AKTpathway in human cancer. Nat Rev Cancer 2002;2:489–501.
58. Ettinger SL, Sobel R, Whitmore TG, Akbari M, Bradley DR,Gleave ME, Nelson CC. Dysregulation of sterol response element-binding proteins and downstream effectors in prostate cancer dur-ing progression to androgen independence. Cancer Res 2004;64:2212–21.
59. Kim YC, Ntambi JM. Regulation of stearoyl-CoA desaturase genes:role in cellular metabolism and pre-adipocyte differentiation. Bio-chem Biophys Res Commun 1999;266:1–4.
571SCD LOSS IN PROSTATE CARCINOMA
Related Documents











