European Journal of Neuroscience, Vol. 8, pp. 1735-1 745, I996 @ European Neuroscience Association Long-term Survival of Retina Optic Nerve Section in Adult Ganglion Cells Following bcl-2 Transgenic Mice Maria Cristina Cenni', Lidia Bonfanti'~~, Jean-Claude Martinou3,Gian Michele Ratto', Enrica Strettoil and Lam berto Maffei v2 'Istituto di Neurofisiologia del CNR, 56127 Pisa Via San Zen0 51, Italy, 2Scuola Normale Superiore, Pisa, Italy, 3Glaxo Institute for Molecular Biology, Geneva, Switzerland 4Present address: Consorzio Mario Negri Sud, Chieti, Italy Keywords: axotomy, neuronal degeneration, retrograde degeneration, retina Abstract The bcl-2 gene codes for a protein that acts as a powerful inhibitor of active cell death. Since the transection of the optic nerve in adult mammalians starts a massive process of degeneration in retinal ganglion cells, we investigatedwhether the overexpression of bcl-2 in adult transgenic mice can protect the axotomited ganglion cells. We performed intracranial optic nerve transection on both wild type and transgenic adult mice, and we tested cell survival 2 or 3.5 months after axotomy. The percentage of surviving ganglion cells after optic nerve section was computed by combining the counts of the optic nerve fibres in intact nerves with the cell density measures of the ganglion cell layer of axotomized retinae. From these data we found that in transgenic mice -65% of ganglion cells survived 3.5 months after axotomy. In contrast, 2 months after surgery, 40% of ganglion cells were left in wild type retinae. We have also examined the morphology and fine structure of the proximal stump of the sectioned optic nerves by light and electron microscopy. In the transgenic mice a very large number of axons survived after surgery and they still exhibited fairly normal morphology and ultrastructure. On the other hand the wild type transected nerves had only a few visible axons that displayed clear signs of degeneration. We conclude that the overexpression of Bcl-2 protein in central neurons is a very effective strategy to ensure long-term survival in axotomized cells. Introduction The functions of the mammalian central nervous system (CNS) are irreversibly compromised by a traumatic injury as a consequence of massive neuronal cell death and of the limited regenerative capacity of the CNS. Any program aimed at improving CNS regeneration must, as a first necessary condition, ensure neuronal survival. Optic nerve section is a commonly used model for analysing anatomical and functional responses to axotomy in the CNS. The immediate consequence of this lesion is the loss of visual function from the affected eye. In the following few months, retinal ganglion cells (RGCs), whose axons compose the transected nerve, undergo a progressive process of degeneration. The death of the vast majority of RGCs obviously precludes any significant regeneration of the optic nerve. A variety of approaches have been employed to improve RGC survival after optic nerve transection (Carmignoto et al., 1989; Thanos et al., 1989; Maffei et al., 1990), but even in the most successful cases only a small number of cells were rescued (up to 22% of the total RGCs surviving after 2 months; Maffei et al., 1994). Devising better techniques to promote neuronal survival requires an improved knowledge of the death mechanisms induced by axotomy: a crucial finding was the discovery that the axon transection starts a process of apoptosis in the injured neuron (Oppenheim et al., 1990; Berkelaar et al., 1994; Garcia-Valenzuela et al., 1994; Rabacchi et al., 1994). Apoptosis is promoted or inhibited by the action of a number of genes and it is therefore conceivable that cell survival could be assisted by interfering with these regulating agents. bcl-2 is one of these controlling genes, and it codes for a protein that, through a mechanism still partially obscure, protects cells from apoptosis (Garcia et al., 1992; Allsopp et al., 1993; Mah et al., 1993). However, adult central neurons cannot exploit this protective effect since a high level of bcl-2 expression is found only during early development. To study the potentiality of the bcl-2 gene in preventing cell death after axotomy in the CNS it is necessary to find a way of activating it in the experimental model: in this study we employed a line of transgenic mice constitutively overexpressing the human bcl-2 gene in most CNS neurons, including RGCs. We analysed retinal whole-mount preparations to ascertain the degree of neuronal survival in the ganglion cell layer. In parallel, we carried out a morphological analysis of the proximal stump of the optic nerve to evaluate the integrity of the enduring ganglion cell Correspondence to: Lamberto Maffei, as above Received 27 December 1995, revised 8 March 1996. accepted 20 March 1996

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

European Journal of Neuroscience, Vol. 8, pp. 1735-1 745, I996 @ European Neuroscience Association
Long-term Survival of Retina Optic Nerve Section in Adult
Ganglion Cells Following bcl-2 Transgenic Mice
Maria Cristina Cenni', Lidia Bonfanti'~~, Jean-Claude Martinou3, Gian Michele Ratto', Enrica Strettoil and Lam berto Maff ei v 2
'Istituto di Neurofisiologia del CNR, 56127 Pisa Via San Zen0 51, Italy, 2Scuola Normale Superiore, Pisa, Italy, 3Glaxo Institute for Molecular Biology, Geneva, Switzerland 4Present address: Consorzio Mario Negri Sud, Chieti, Italy
Keywords: axotomy, neuronal degeneration, retrograde degeneration, retina
Abstract The bcl-2 gene codes for a protein that acts as a powerful inhibitor of active cell death. Since the transection of the optic nerve in adult mammalians starts a massive process of degeneration in retinal ganglion cells, we investigated whether the overexpression of bcl-2 in adult transgenic mice can protect the axotomited ganglion cells. We performed intracranial optic nerve transection on both wild type and transgenic adult mice, and we tested cell survival 2 or 3.5 months after axotomy. The percentage of surviving ganglion cells after optic nerve section was computed by combining the counts of the optic nerve fibres in intact nerves with the cell density measures of the ganglion cell layer of axotomized retinae. From these data we found that in transgenic mice -65% of ganglion cells survived 3.5 months after axotomy. In contrast, 2 months after surgery, 40% of ganglion cells were left in wild type retinae. We have also examined the morphology and fine structure of the proximal stump of the sectioned optic nerves by light and electron microscopy. In the transgenic mice a very large number of axons survived after surgery and they still exhibited fairly normal morphology and ultrastructure. On the other hand the wild type transected nerves had only a few visible axons that displayed clear signs of degeneration. We conclude that the overexpression of Bcl-2 protein in central neurons is a very effective strategy to ensure long-term survival in axotomized cells.
Introduction
The functions of the mammalian central nervous system (CNS) are irreversibly compromised by a traumatic injury as a consequence of massive neuronal cell death and of the limited regenerative capacity of the CNS. Any program aimed at improving CNS regeneration must, as a first necessary condition, ensure neuronal survival.
Optic nerve section is a commonly used model for analysing anatomical and functional responses to axotomy in the CNS. The immediate consequence of this lesion is the loss of visual function from the affected eye. In the following few months, retinal ganglion cells (RGCs), whose axons compose the transected nerve, undergo a progressive process of degeneration. The death of the vast majority of RGCs obviously precludes any significant regeneration of the optic nerve.
A variety of approaches have been employed to improve RGC survival after optic nerve transection (Carmignoto et al., 1989;
Thanos et al., 1989; Maffei et al., 1990), but even in the most successful cases only a small number of cells were rescued (up to 22% of the total RGCs surviving after 2 months; Maffei et al., 1994). Devising better techniques to promote neuronal survival requires an improved knowledge of the death mechanisms induced by axotomy:
a crucial finding was the discovery that the axon transection starts a process of apoptosis in the injured neuron (Oppenheim et al., 1990; Berkelaar et al., 1994; Garcia-Valenzuela et al., 1994; Rabacchi et al., 1994). Apoptosis is promoted or inhibited by the action of a number of genes and it is therefore conceivable that cell survival could be assisted by interfering with these regulating agents. bcl-2 is one of these controlling genes, and it codes for a protein that, through a mechanism still partially obscure, protects cells from apoptosis (Garcia et al., 1992; Allsopp et al., 1993; Mah et al., 1993). However, adult central neurons cannot exploit this protective effect since a high level of bcl-2 expression is found only during early development. To study the potentiality of the bcl-2 gene in preventing cell death after axotomy in the CNS it is necessary to find a way of activating it in the experimental model: in this study we employed a line of transgenic mice constitutively overexpressing the human bcl-2 gene in most CNS neurons, including RGCs.
We analysed retinal whole-mount preparations to ascertain the degree of neuronal survival in the ganglion cell layer. In parallel, we carried out a morphological analysis of the proximal stump of the optic nerve to evaluate the integrity of the enduring ganglion cell
Correspondence to: Lamberto Maffei, as above
Received 27 December 1995, revised 8 March 1996. accepted 20 March 1996
1736 Long-term survival of axotomized RGCs in bcl-2 transgenic mice
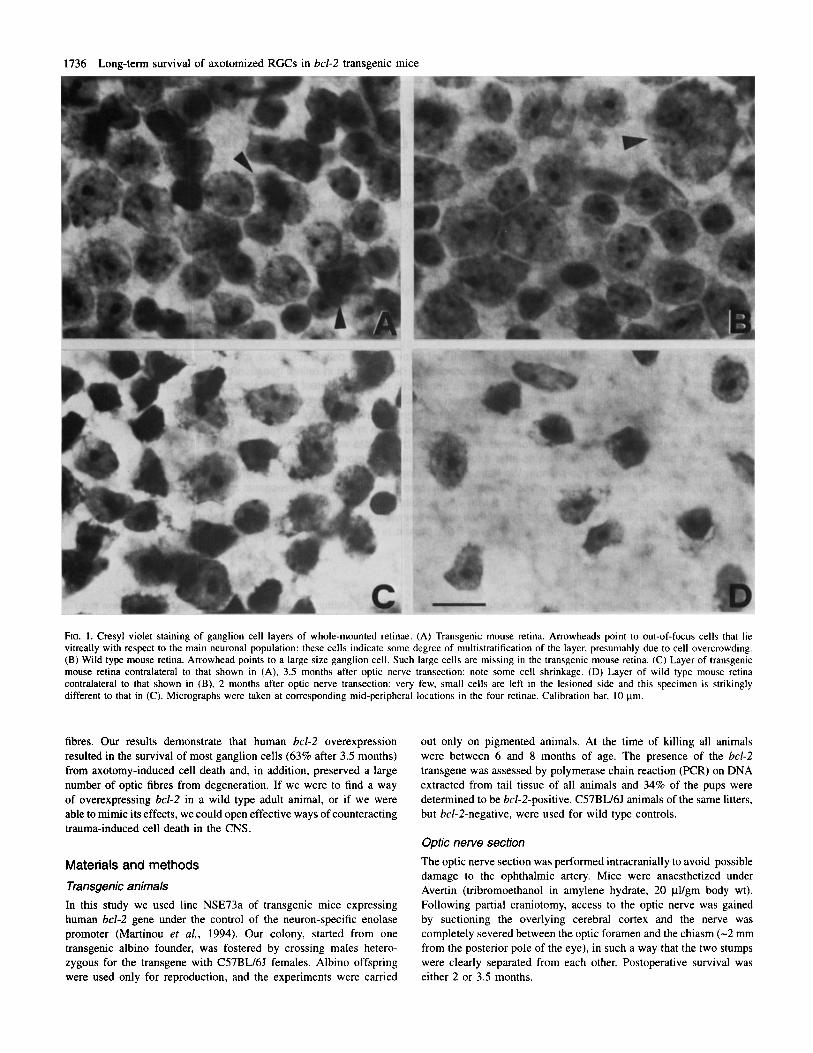
FIG. 1. Cresyl violet staining of ganglion cell layers of whole-rnounted retinae. (A) Transgenic mouse retina. Arrowheads point to out-of-focus cells that lie vitreally with respect to the main neuronal population: these cells indicate some degree of multistratification of the layer, presumably due to cell overcrowding. (B) Wild type mouse retina. Arrowhead points to a large size ganglion cell. Such large cells are missing in the transgenic mouse retina. (C) Layer of transgenic mouse retina contralateral to that shown in (A), 3.5 months after optic nerve transection: note some cell shrinkage. (D) Layer of wild type mouse retina contralateral to that shown in (B), 2 months after optic nerve transection: very few, small cells are left in the lesioned side and this specimen is strikingly different to that in (C). Micrographs were taken at corresponding mid-peripheral locations in the four retinae. Calibration bar. 10 pm.
fibres. Our results demonstrate that human bcl-2 overexpression resulted in the survival of most ganglion cells (63% after 3.5 months) from axotomy-induced cell death and, in addition, preserved a large number of optic fibres from degeneration. If we were to find a way of overexpressing bcl-2 in a wild type adult animal, or if we were able to mimic its effects, we could open effective ways of counteracting trauma-induced cell death in the CNS.
Materials and methods Transgenic animals In this study we used line NSE73a of transgenic mice expressing human bcl-2 gene under the control of the neuron-specific enolase promoter (Martinou e l nl., 1994). Our colony, started from one transgenic albino founder, was fostered by crossing males hetero- zygous for the transgene with C57BU6J females. Albino offspring were used only for reproduction, and the experiments were carried
out only on pigmented animals. At the time of killing all animals were between 6 and 8 months of age. The presence of the bcl-2 transgene was assessed by polymerase chain reaction (PCR) on DNA extracted from tail tissue of all animals and 34% of the pups were determined to be bcl-2-positive. C57BU6J animals of the same litters, but bcl-2-negative, were used for wild type controls.
Optic nerve section The optic nerve section was performed intracranially to avoid possible damage to the ophthalmic artery. Mice were anaesthetized under Avertin (tribromoethanol in amylene hydrate, 20 FVgm body wt). Following partial craniotomy, access to the optic nerve was gained by suctioning the overlying cerebral cortex and the nerve was completely severed between the optic foramen and the chiasm (-2 mm from the posterior pole of the eye), in such a way that the two stumps were clearly separated from each other. Postoperative survival was either 2 or 3.5 months.

Long-term survival of axotomized RGCs in bcl-2 transgenic mice 1737
FIG. 2. Soma size distribution of neurons in the RGC layer of transgenic and wild type mouse retinae. Distribution histograms for a wild type (W, average size 10.8 pm, n = 639) and a transgenic mouse (0, average size 9.30 pm, n = 996) retina. The distribution in the wild type retina is markedly bimodal and it can be very well fitted by the sum of two gaussian distributions (continuous line), with averages of 8.7 and 13.4 pm and standard deviations of 3.5 and 6.4 pm. In accordance with the findings of Drager and Olsen (1981) all the displaced amacrine cells are contained in the first mode, while the second mode is entirely composed of ganglion cells. The distribution in the transgenic retina is essentially unimodal.
Nissl staining Animals were deeply anaesthetized with an intra-peritoneal injection of Avertin and immediately perfused transcardially with normal saline, followed by 4% paraformaldehyde/O.l M cacodylate buffer, pH 7.2. After the execution of a small cut in the sclera to record the retina orientation, the eye bulb was enucleated, opened at the ora serrata and the whole retina was dissected. Afterwards the tissue was mounted, ganglion cells side up, on gelatinized slides, fixed with 2.5% glutaraldehyde, then with formalin-ethanol solution (l:9). Finally the whole-mounted retina was stained with cresyl violet (0. I%), dehydrated, mounted and examined. In total, we analysed the retinae from 14 animals, and the detailed composition of each experimental group is given in the Results. Since the retina was anchored to the gelatinized slide during the entire histological preparation, we expected minimal tissue shrinkage. Nevertheless we evaluated shrinkage in a limited number of cases (n = 4) by comparing the retina surface before and after the histological procedure. In all cases linear shrinkage was found to be <5% and it was henceforth ignored. Besides, since shrinkage affects in a complementary way retinal surface and cell density, the estimate of total cell numbers is not influenced by this artefact.
Density measures Cell density in the ganglion cell layer was measured by a computer- aided procedure. In brief the outline of the retina was acquired in a vectorial form by a computer-controlled microscope (Ratto and Usai, 1991) and the cell density was measured in several regularly distributed fields (size 96 X 66 pm; 2 M 6 fields for each retina, average 26). The computerized system generated a reference system centred in the optic disk of each retina and kept track of the x-y co-ordinates of the centre of the analysed fields and of the associated cell densities. Counting was performed by a human operator on the image of the field displayed on a TV screen. Cells intersecting the field edges were counted only along two congruent sides of the area. In about one-third of cases the counts for three contiguous fields were averaged together to return the cell number for that location. For each retina
we counted on average 2% of the total cells (percentage range: 0.8- 7.3%) and altogether we counted -63 OOO cells in 25 retinae. The variance of the associated Poisson distribution for each retina ranged from 1.2 to -3.5% (average 2.1%) of the total cell number. This was well below the variability found between different retinae belonging to the same experimental group.
The small fraction of non-neuronal cells in the ganglion cell layer has been excluded by using morphological criteria already described (Drager and Olsen, 1981; Perry et al., 1983). Analysis was continued by exporting the retina profile, the counts and the co-ordinates of each measured field to a general-purpose data analysis program (Origin, MicroCal Software Inc., Northampton, MA) that was used to generate the isodensity maps.
The total cell number in the ganglion cell layer of each retina was computed either by multiplying the retinal surface by the average cell density, or by measuring the area of each zone of the isodensity map and by multiplying it for the relative density. The two methods returned very similar values (within 5% one from the other) and they were used interchangeably without further distinction. Areas were measured with an image analyser (MCID-M4, Imaging Research Inc., St Catharines, Ontario).
Electron microscopy of the optic nerve Classically the number of RGCs has been determined by employing a retrograde tracer, either injected in the RGC targets or placed in the form of a pellet on the optic nerve stump. Unfortunately neither HRF’ nor DiI (4Di-Asp) uniformly label the entire ganglion cell population (Carmignoto et al., 1989; Mey and Thanos, 1993). Therefore these methods may introduce a variable and unpredictable bias in the ganglion cell count. To overcome these problems we decided to estimate the ganglion cell number by counting the axons in the optic nerves.
Mice were deeply anaesthetized with an intra-peritoneal injection of Avertin, then they were perfused transcardially with saline solution followed by fixative solution containing 4% paraformaldehyde, 0.1 M sodium cacodylate buffer, pH 7.2, and 0.05 M calcium chloride.

1738
TABLE 1. Data from semithin and ultrathin sections of optic nerves of transgenic and wild type mice
Long-term survival of axotomized RGCs in bcl-2 transgenic mice
~~
Transgenic Wild type
Nerve cross-sectional area (mm2) 0.128 2 0.0013 0.064 5 0.0012 Axon number (10’) 112.2 ? 17.4 45.4 ? 4.0 No. of nerves 4 3
degree of homogeneity when comparing distributions from different retinae. Images from -30 fields (size 120 X 80 pm) were acquired and analysed for soma size with the image analyser. The soma size was computed as the longest cord that was traced within the perimeter of the cell body. On average we measured -800 cells for each of the 10 analysed retinae (range 430-1450 cells).
Nerve areas were measured from semithin sections adjacent to the ultrathin sections used for fibre counting. All morphometrical parameters were measured with the image analyser. Average values are shown together with standard deviations.
The optic nerves were carefully dissected and post-fixed in 4% glutaraldehyde in the same buffer. A few unoperated mice employed only for counting the optic nerve fibres were perfused with a mixture of 2% paraformaldehyde, 2.5% glutaraldehyde in cacodylate buffer, and post-fixed in the same fixative. After 12-1 8 h of fixation at 4°C. the optic nerves were immersed in cacodylate buffer and trimmed perpendicularly to their long axes at 1.5 mrn intervals from the head of the optic nerve to the optic chiasm. Each block of tissue was processed separately. The specimens were post-fixed in 2% cold osmium tetroxide, stained en bloc with 1% uranyl acetate in maleate buffer, dehydrated in an ethanol series and flat embedded in Epon- Araldite. The optic nerves of three wild type, unoperated mice, were used for counting the optic nerve fibres. Fibre counts in transgenic mice were carried out on four nerves: one originated from a single, unoperated animal, while the others were the right optic nerves of three mice operated on the left side. No signs of degeneration were detected in both the right retinae and optic nerves of these animals.
For each nerve used for fibre counts, the block containing the initial segment was oriented in the cutting head of a Leica Ultratome V ultramicrotome and sectioned with a diamond knife, with the cutting face perpendicular to the long axis of the nerve, as defined by the nerve surface. For each sample a single semithin ( I pm) section and one adjacent ultrathin section (silver-gold, -90 nm thick) were obtained. The semithin sections were stained with Azur II/ toluidine blue, mounted in DePeX, coverslipped and examined with a Zeiss Axiophot light microscope interfaced with the image analyser for measure of the optic nerve area. The ultrathin sections were stretched with xylene vapours to minimize compression, collected on uncoated, 200 mesh copper grids, stained with uranyl acetate and lead citrate for 4 min and photographed with a Jeol 1200 EX11 electron microscope. Photographs were taken uniformly all over the surface of each section, using the copper grid as a sampling matrix: 4-5 pictures were shot at regular intervals from each window of the grid. A total number of 35-50 pictures were taken for each nerve at a magnification of 6000. A carbon replica grating was photographed at the same enlargement, and used to print the micrographs at a final magnification of -13 000. On each print a rectangular area of -230 pm2 was traced and all the fibres within that area and those touching either the top or the left border were counted. On average, 6% of the area of each transgenic optic nerve and 13% of the area of each wild type nerve were photographed and counted. The total number of fibres in each nerve was obtained by multiplying the average density for the area of the nerve section.
Measure of soma size From inspection of the isodensity maps of each analysed retina we choose a mid-peripheral area where the cell density was close to the average density for the entire retina. We did so to maintain a certain
Results We crossed a line of the transgenic mouse line NSE73a with C57BL/ 65 mice. DNA was extracted from 89 offspring and analysed by PCR. Thirty mice tested positive for the human bcl-2 gene. In these transgenic mice the human bcl-2 is constitutively expressed in the majority of central neurons including RGCs (Martinou et al. 1994). We tested and quantified the protective effects of the overexpression of human Bcl-2 protein on axotomized ganglion cells by analysing the retinal response to optic nerve transection on transgenic and wild type adult mice. We performed intracranial optic nerve transection on 6 transgenic animals: four of them were analysed after 3.5 months and two after 2 months. As controls, four wild type mice were analysed 2 months after axotomy. All the retinae of these 10 animals, together with a number of additional transgenic (n = 3) and wild type (n = 2) mouse retinae, were dissected and processed for cresyl violet staining.
Figure 1 shows the RGC layer of the normal and operated sides of transgenic and wild type retinae. As expected from the natural cell death reduction occumng in this transgenic breed (Martinou et al., 1994) the RGC layer is markedly denser, up to the point that occasionally cell bodies appeared to be multistratified, while space between cells is restricted. A possible consequence of the cell crowding is a conspicuous lack of large neurons in the transgenic retina in comparison to the wild type retina, as shown by size distributions in Figure 2. The difference between Figure 1C and Figure 1D demonstrates the dramatic effect of bcl-2 overexpression on cell survival: whereas the transgenic mouse retina (Fig. 1C) shows a modest decrease in cell density 3.5 months after axotomy, the wild type retina (Fig. ID) has very few residual cells 2 months after axotomy.
The ganglion cell layer contains two different groups of neurons: RGCs and displaced amacrine cells. Since Nissl staining equally marks both populations we could not estimate the ganglion cell number by this preparation alone and we decided to count the optic nerve axons. Unfortunately this method works only in the unlesioned animals since the wild type proximal nerve stump is essentially devoid of fibres. Therefore we estimated the RGCs survival as follows: we obtained the number of RGCs in the unlesioned retinae by counting the fibres in the optic nerve, then we subtracted this from the entire ganglion cell layer population obtained from Nissl staining, to return the displaced amacrine cell number. Finally, since amacrine cells are not affected by the optic nerve transection (Perry et al., 1983), we computed the number of RGCs that survived axotomy by subtracting the number of displaced amacrine cells we found in the control retinae from the Nissl counts of axotomized retinae. All animals were of similar age (6-8 months). Given the large inter-strain variability of RGCs number (Williams et al., 1990; Rice et al., 1995) we did not employ the data already present in the literature and instead used age-matched, bcl-2-negative littermates of the transgenic mice as wild type controls.
Fibre counts were made on 30-50 electron micrographs for each nerve printed at a final magnification of -13 000. In both transgenic and wild type mice the large majority of the optic nerve fibres were

Long-term survival of axotomized RGCs in bcl-2 transgenic mice 1739
FIG. 3. Isodensity maps for two pairs of transgenic (top) and wild type (bottom) retinae. The maps on the left show the RGC layer cell density in the control retinae, while the maps on the right show the cell distributions, 3.5 and 2 months after transection respectively. In both map pairs the shift from one grey level to the adjacent indicates a 5% change in cell density with respect to the peak density of the corresponding unlesioned retina. Therefore the darker appearance of the map on the top right compared with the map on the bottom right map demonstrates the larger percentage of surviving cells in the transgenic retina. The inverted centre-periphery gradient, evident in the top-right map, has been observed in all of the axotomized transgenic retinae. The total number of neurons in the RGC layer of these four retinae are 182 OOO and 138 OOO for the control and lesioned transgenic retinae, and 136 OOO and 76 400 for the control and transected wild type retinae respectively. The correspondence between grey levels and cell densities is shown below each retinal map. The first number next to a grey level indicates the cell density upper limit (cells/mm2). The second number expresses the cell density relative to the peak density of the unlesioned retina. The white area at the centre of each map is the optic disk. N, T, D and V indicate the nasal, temporal, dorsal and ventral retina respectively. The isodensity maps were computed as explained in Materials and methods. The Figure was composed with Adobe Photoshop, where the retinae were rotated as appropriate and text and calibrations were added. Calibration bar, 3 mm.
myelinated. This characteristic provided an unequivocal criterion for their identification. The axoplasm was characterized by a pale matrix and it contained uniformly dispersed microtubules, bundles of intermediate filaments, dark mitochondria and irregular membranous profiles. A certain number of fibres, apparently unmyelinated, dis- played a subaxolemmal layer of electron dense material: this is a characteristic feature of myelinated fibres at the node of Ranvier (Peters et al., 1976). Other fibres, more numerous in the transgenic
optic nerves, appeared surrounded by a glial process but lacked a myelin sheath. All processes, with pale axoplasm and displaying both microtubules and filaments, were included in the counts. Astrocytic processes were easily identified by the presence of abundant filaments; they exhibited a dark matrix and normally lacked microtubules. Finally, a few dark profiles containing microtubules but lacking filaments, were classified as glial processes, possibly oligodendrocytic, and were not counted.
1740 Long-term survival of axotomized RGCs in bcl-2 transgenic mice
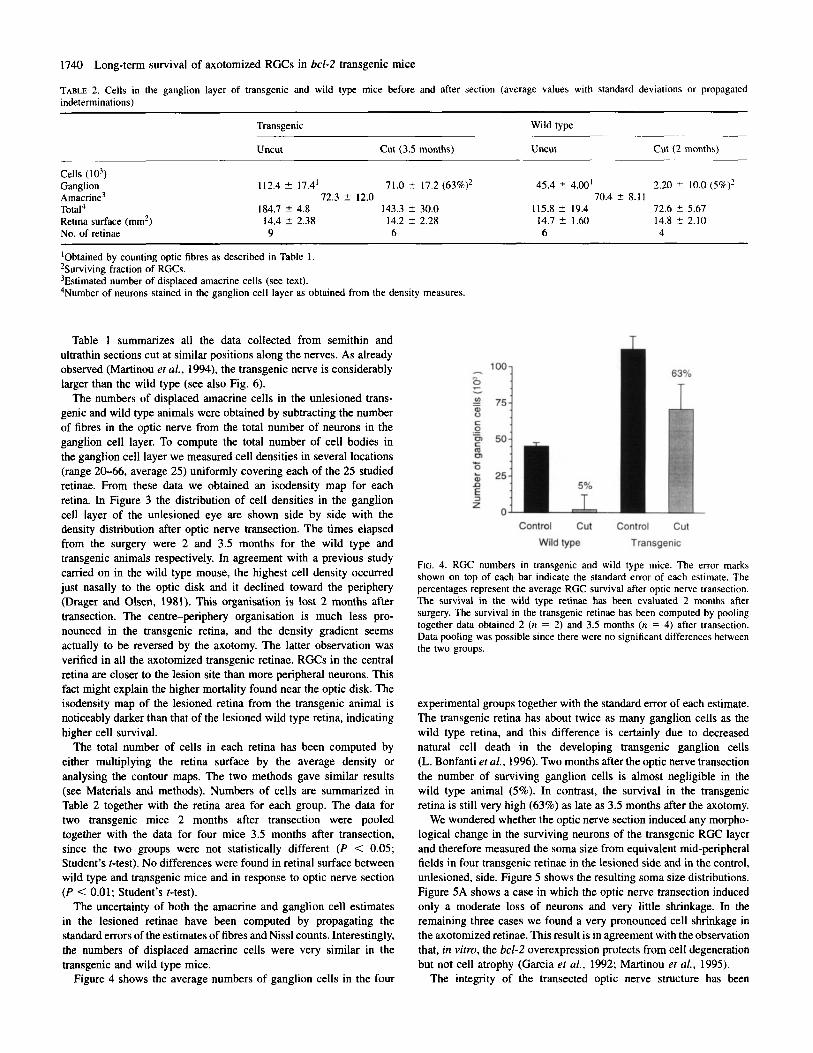
TABLE 2. Cells in the ganglion layer of transgenic and wild type mice before and after section (average values with standard deviations or propagated indeterminations)
Transgenic Wild type
Uncut Cut (3.5 months) Uncut Cut (2 months)
Cells (lo3) Ganglion Amacnne3
2.20 ? 10.0 (59)? 112.4 +- 17.4’ 71.0 t 17.2 (63%)2 45.4 5 4.00‘ 72.3 2 12.0 70.4 f 8.1 1
72.6 t 5.67 Total4 184.7 +- 4.8 143.3 2 30.0 115.8 2 19.4 Retina surface (mm’) 14.4 5 2.38 14.2 t 2.28 14.7 2 1.60 14.8 -C 2.10 No. of retinae 9 6 6 4
‘Obtained by counting optic fibres as described in Table I . 2Surviving fraction of RGCs. 3Estimated number of displaced amacrine cells (see text). 4Number of neurons stained in the ganglion cell layer as obtained from the density measures.
Table 1 summarizes all the data collected from semithin and ultrathin sections cut at similar positions along the nerves. As already observed (Martinou er al., 1994), the transgenic nerve is considerably larger than the wild type (see also Fig. 6).
The numbers of displaced amacrine cells in the unlesioned trans- genic and wild type animals were obtained by subtracting the number of fibres in the optic nerve from the total number of neurons in the ganglion cell layer. To compute the total number of cell bodies in the ganglion cell layer we measured cell densities in several locations (range 20-66, average 25) uniformly covering each of the 25 studied retinae. From these data we obtained an isodensity map for each retina. In Figure 3 the distribution of cell densities in the ganglion cell layer of the unlesioned eye are shown side by side with the density distribution after optic nerve transection. The times elapsed from the surgery were 2 and 3.5 months for the wild type and transgenic animals respectively. In agreement with a previous study carried on in the wild type mouse, the highest cell density occurred just nasally to the optic disk and it declined toward the periphery (Drager and Olsen, 1981). This organisation is lost 2 months after transection. The centre-periphery organisation is much less pro- nounced in the transgenic retina, and the density gradient seems actually to be reversed by the axotomy. The latter observation was verified in all the axotomized transgenic retinae. RGCs in the central retina are closer to the lesion site than more peripheral neurons. This fact might explain the higher mortality found near the optic disk. The isodensity map of the lesioned retina from the transgenic animal is noticeably darker than that of the lesioned wild type retina, indicating higher cell survival.
The total number of cells in each retina has been computed by either multiplying the retina surface by the average density or analysing the contour maps. The two methods gave similar results (see Materials and methods). Numbers of cells are summarized in Table 2 together with the retina area for each group. The data for two transgenic mice 2 months after transection were pooled together with the data for four mice 3.5 months after transection, since the two groups were not statistically different (P < 0.05; Student’s t-test). No differences were found in retinal surface between wild type and transgenic mice and in response to optic nerve section (P < 0.01; Student’s t-test).
The uncertainty of both the amacrine and ganglion cell estimates in the lesioned retinae have been computed by propagating the standard errors of the estimates of fibres and Nissl counts. Interestingly, the numbers of displaced amacrine cells were very similar in the transgenic and wild type mice.
Figure 4 shows the average numbers of ganglion cells in the four
FIG. 4. RGC numbers in transgenic and wild type mice. The error marks shown on top of each bar indicate the standard error of each estimate. The percentages represent the average RGC survival after optic nerve transection. The survival in the wild type retinae has been evaluated 2 months after surgery. The survival in the transgenic retinae has been computed by pooling together data obtained 2 (n = 2) and 3.5 months (n = 4) after transection. Dats pooling was possible since there were no significant differences between the two groups.
experimental groups together with the standard error of each estimate. The transgenic retina has about twice as many ganglion cells as the wild type retina, and this difference is certainly due to decreased natural cell death in the developing transgenic ganglion cells (L. Bonfanti eral., 1996). Two months after the optic nerve transection the number of surviving ganglion cells is almost negligible in the wild type animal (5%) . In contrast, the survival in the transgenic retina is still very high (63%) as late as 3.5 months after the axotomy.
We wondered whether the optic nerve section induced any morpho- logical change in the surviving neurons of the transgenic RGC layer and therefore measured the soma size from equivalent mid-peripheral fields in four transgenic retinae in the lesioned side and in the control, unlesioned, side. Figure 5 shows the resulting soma size distributions. Figure 5A shows a case in which the optic nerve transection induced only a moderate loss of neurons and very little shrinkage. In the remaining three cases we found a very pronounced cell shrinkage in the axotomized retinae. This result is in agreement with the observation that, in vitro, the bcl-2 overexpression protects from cell degeneration but not cell atrophy (Garcia et al., 1992; Martinou et al., 1995).
The integrity of the transected optic nerve structure has been

Long-term survival of axotomized RGCs in bcl-2 transgenic mice 1741
FIG. 5 . Soma size distribution of neurons in the RGC layer of transgenic mouse retinae. Distribution histograms for four pairs of retinae from four operated transgenic mice: 0, cell size distribution in the control retina; W. cell size distribution in the transected retina (A, B and C: 3.5 months after surgery; D: 2 months after surgery).
evaluated by light and electron microscopy of the proximal stumps (Fig. 6). When we compared the structure of nerves in a transgenic mouse and in a wild type mouse, 3.5 and 2 months after transection respectively, a large number of intact myelinated fibres was still observed in the transgenic nerve (Fig. 6E), while a picture of general degeneration was apparent in the wild type mouse nerve (Fig. 6F). In the wild type the axon number was dramatically decreased, while myelinic sheaths appeared darker and altered in structure and astroglial cells appeared enlarged, with their processes filling the space among the degenerating fibres (glial scar).
The difference between transgenic and wild type nerves after transection was even more dramatic at the electron microscopy level (Fig. 7). Three and a half months after axotomy the surviving fibres in the transgenic nerve were numerous and exhibited fairly normal ultrastructure: although microtubule concentration seemed lowered, axons still had mitochondria, intermediate filaments and myelinic sheaths. The optic nerve of wild type mice showed mostly degenerating fibres and very few large axons. It has to be noted that we had to limit the analysis of the wild type mouse optic nerve stump to samples obtained 2 months after transection, since after that time the degeneration was total.
Discussion
Most central adult neurons die after being struck by an acute injury, and this degenerative process is one of the major causes of the lack of regeneration in the CNS. A representative example is the fate encountered by RGCs after axotomy: in mammals optic nerve transection starts a process of active cell death that leads to the degeneration of most RGCs within a few weeks (Berkelaar et al., 1994; Garcia-Valenzuela et al., 1994). This rapid and extensive degeneration obviously frustrates whatever intrinsic regenerative capability ganglion cells might have (Doster et aZ., 1991).
The crucial characteristic of the active cell death process is its being controlled by the concerted action of a number of genes, some promoting, some inhibiting the process itself (for reviews see Osborne and Schwartz, 1994; Davies, 1995; Korsmeyer, 1995). A number of experiments, mainly performed in vitro, have shown that one of the regulating genes, bcl-2, has a powerful inhibitory action on active cell death (Garcia et al., 1992; Hockenbery et al., 1993; Greenlund et al., 1995; Farlie et al., 1995). The present study shows that the overexpression of the human bcZ-2 gene effectively protects the adult RGC population from axotomy-induced cell death in viva

1742 Long-term survival of axotomized RGCs in bcl-2 transgenic mice
FIG. 6. Light micrographs of transgenic and wild type optic nerves. (A and B) Semithin cross sections of (A) transgenic and (B) wild type mouse optic nerve. (C and D) Higher power views of same preparations as in (A) and (B) respectively: note that the general organisation of the two nerves is similar. However the nerve of the wild type animal exhibits fibres of large size which are absent in the transgenic counterpart. (E and F) Proximal stumps of (E) the transgenic and (F) the wild type mouse optic nerve, 3.5 and 2 months after transection. Many fibres are still visible in (E); in (F) arrows point to degenerating myelinated fibres and there is a general degeneration of the nervous structure, with astrocytes (a) occupying the largest fraction of the nerve surface. Calibration bars: 100 pm (A, B). 10 pm (C-F).

Long-term survival of axotomized RGCs in bcl-2 transgenic mice 1743
FIG. 7. Electron microscopy of transected optic nerves in transgenic and wild type animals. (A and B) Low power electron micrographs of (A) transgenic mouse optic nerve 3.5 months after transection (proximal stump, near the optic nerve head) and (B) wild type mouse optic nerve, 2 months after transection. Note the large number of surviving myelinated fibres in (A). In (B), at the same location, astroglial processes (ap) occupy most of the nerve surface, arrowheads point to degenerating myelin sheaths and rare large size axons survive (*). (C) Same preparation as in (A): surviving axons of transgenic mouse optic nerve show normal myelinic sheaths, mitochondria (m) and intermediate filaments (arrows). Arrowheads point to bundles of filaments inside an astrocytic process. (D) Same preparation: astrocytic processes appear enlarged and densely packed with filaments (arrowheads). Arrows indicate a gap junction between two astrocytes. nu: astrocytic nucleus. Calibration bars: I pm (A, B), 0.5 ym (C), 0.2 ym (D).

1744 Long-term survival of axotomized RGCs in bcl-2 transgenic mice
Retinae of transgenic bcl-2 mice
In the transgenic mouse generated by Martinou and collaborators (Dubois-Dauphin et al., 1994; Martinou et al., 1994), the gene coding the human Bcl-2 protein is steadily expressed in neurons starting from embryonic day 13. Immunohistochemical studies showed that bcl-2 is present throughout most of the retina (Martinou et al., 1994). Since all retinal neurons, including RGCs, are drastically reduced by naturally occurring cell death (Sidman, 1961), it is foreseeable that the transgenic mouse retina has more neurons than the wild type. In agreement with this prediction, we found that the transgenic optic nerve has more than twice as many fibres as the wild type nerve, and this increase is reflected by the optic nerve larger size. Correspondingly we observed a considerable increment in the density of the ganglion cell layer of the transgenic retina. This higher density has been already observed by Martinou and collaborators in the central retina, but no distinction was made between ganglion and amacrine cells (Martinou et al., 1994). By putting together the optic nerve data and the analysis of the cell density we found that the absolute number of displaced amacrine cells is basically identical in the wild type and transgenic animals. This is a somewhat surprising finding, since amacrine cells are also reduced by developmental cell death. We propose two possible explanations for this observation: (i) not all amacrine cells express amounts of bcl-2 sufficient to receive complete protection from cell death [indeed, the published data leave some doubts whether all amacrine cells strongly express human bcl-2 (Martinou et al., 1994)] and (ii) since most displaced amacrine cells reach the RGC layer when the ganglion cells are already there, it is conceivable that the increased cell density of the transgenic retina might hamper the amacrine cells migration. The increase in cell density of the transgenic RGC layer results in a reduction of both neuronal size and intercellular space. Notwithstanding the greater number of cells, the transgenic retinae are not wider than the wild type retihae.
bcl-2 overexpression protects RGCs from axotomy-induced cell death
In normal animals the expression of bcl-2 in central neurons is high during embryonal development and drops after birth (Merry et al., 1994). In contrast bcl-2 remains highly expressed in peripheral neurons throughout the entire lifetime. It is tempting to hypothesize that the expression of bcl-2 in the adult peripheral nervous system (PNS) might contribute to endow the peripheral neurons with their intrinsic resistance from trauma-induced cell death. This idea prompted us to ascertain whether bcl-2 overexpression protects central neurons from axotomy-induced cell death.
Dubois-Dauphine and collaborators showed that the neurons origin- ating the facial nerve survive peripheral axotomy in the neonatal transgenic mouse, while they undergo massive cell death in the neonatal wild type animal (Dubois-Dauphine et al., 1994). A similar result has been obtained by Farlie et al. (1995) who found a protective effect on motoneurons following section of the sciatic nerve in new- born transgenic mice overexpressing bcl-2. However, in the adult wild type mouse facial nerve transection does not cause generalized cell death and this might occur because, although the originating facial nucleus belongs to the CNS, the nerve glial environment is characteristic of the PNS (Schwann cells instead of oligodendrocites). For these reasons the facial nerve section is not an ideal model in which to study axotomy-induced degeneration in the adult CNS. A similar argument also holds for the section of the sciatic nerve. On the other hand, transection of the optic nerve, like transection of any other central nerve, leads to massive degeneration both in developing
and adult animals with no significant indications of regeneration, making it a more appropriate model in which to study the possible inhibitory role of bcl-2 overexpression on axotomy-induced cell death in the CNS. Our data show that bcl-2 overexpression rescues most adult RGCs from cell death, ensuring the long-term survival of both cell bodies and axons after optic nerve section.
Other studies have been directed at finding strategies to prevent ganglion cell death induced by optic nerve section. Whereas a number of techniques showed some promise in temporarily retarding cell death, very few offered any real hope of achieving significant long- term survival. Most of these studies have been based on the hypothesis that axotomy-induced cell death is caused by neurotrophic factor deprivation due to the target removal. Intra-ocular injections of trophic factors, microglia suppressing factors, target-derived fragments and Schwann cells succeeded in ensuring a limited degree of long-term survival to the axotomized ganglion cells (Carmignoto et al., 1989; Maffei et al., 1990; Mey and Thanos, 1993; Thanos et al., 1993; Mansour-Robaey et al., 1994; Huxlin et al., 1995). The best long- term results have been obtained by Schwann cell injections: Maffei and collaborators succeeded in rescuing up to 22% of the axotomized ganglion cells for 14 weeks following optic nerve section (Maffei et al., 1990). In another study injections of microglia suppressing factors rescued 34% of cells but this survival was tested only 28 days after the optic nerve was cut (Thanos et al., 1993). These survival rates are relatively low when compared with our average survival of 63% after 15 weeks from the transection. In addition, a large number of severed fibres survived in the proximal stumps of the optic nerves of transgenic animals. This indicates a better survival of the whole neuronal structure after axotomy. Such an extensive fibre protection has never been observed either in vitro or in vivo.
In other experiments the axotomized retina was connected either to the optic tectum or to pretectum by a graft of peripheral nerve (Vidal-Sanz et al., 1987; Thanos 1992). Some of the surviving ganglion cells regenerated axons that, by connecting to the target, restored a rudimentary light-driven response. This recovery was obtained despite the fact that the RGC survival was only lo%, showing that RGCs have some intrinsic regenerative capability. The high survival found in the bcl-2 transgenic mouse would substantially increase the pool of cells potentially capable of regenerating.
We observed that neurons in the RGC layer exhibited soma shrinkage, in spite of the conspicuous RGC survival in the transgenic retina. This result is analogous to the finding that, although it protects isolated neurons from cell death induced by neurotrophic factor deprivation, bcl-2 does not prevent cell atrophy (Garcia et al., 1992; Farlie et al., 1995). A different situation is encountered when the protection is given by Schwann cell injections (Maffei et al., 1990). In this experiment, even if the fraction of surviving RGCs is much smaller than reported here, the remaining neurons exhibited very little shrinkage. This observation suggests that the protective effects of bcl-2 and trophic factors proceed along at least partially separated pathways.
The shrinkage of ganglion cells may cast some shadow over the functional integrity of the surviving neurons, since cell death is preceded by atrophy of the neurites and shrinkage of the cell body (Oppenheim, 1991; Greenlund et al.. 1995). An effective response to this question can only come from physiological studies on the transgenic mouse. Indeed, the recordings of the ganglion cell compon- ent of the electroretinogram (P-ERG) are still basically unchanged 3.5 months after the optic nerve transection (Porciatti et al., 1995). In contrast the P-ERG has totally disappeared 2 months after transection in the wild type animal. Such electrophysiological record- ings indicate that the rescued axotomized RGCs are still functional

Long-term survival of axotomized RGCs in bcl-2 transgenic mice 1745
and that they preserve enough intraretinal synaptic connections to account for the observed light responsiveness.
Acknowledgements We are grateful to Mr A. Bertini, Mr P. Taccini and Mr A. Tacchi for technical help, and to Professor D. Burr for reading the manuscript. This study has been partially supported by the International Institute for Paraplegia, grant P24.
Abbreviations CNS central nervous system PCR polymerase chain reaction PNS peripheral nervous system RGC retinal ganglion cell
References Allsopp. T. E., Wyatt, S.. Patterson, H. F. and Davies, A. M. (1993) The
proto-oncogene bcl-2 can selectively rescue neurotrophic factor-dependent neurons from apoptosis. Cell, 73, 295-307.
Berkelaar. M.. Clarke. D. B., Wang, Y.-C., Bray, G. M. and Aguayo, A. J. (1994) Axotomy results in delayed death and apoptosis of retinal ganglion cells in adult rats. J. Neurosci., 14, 43684374.
Bonfanti, L., Strettoi, E., Chierzi, S., Cenni, M. C., Liu, X.-H., Martinou, J.-C., Maffei. L. and Rabacchi, S. A. (1996) Protection of retinal ganglion cells from natural and axotomy-induced cell death in neonatal transgenic mice overexpressing bcl-2. J. Neurosci.. 16, in press.
Carmignoto, G., Maffei, L., Candeo, P.. Canella, R. and Comelli, C. (1989) Effect of NGF on the survival of rat retinal ganglion cells following optic nerve section. J. Neurosci. 9, 1263-1272.
Davies, A. M. (1995) The BcI-2 family of proteins and the regulation of neuronal survival. Trends Neurosci., 18, 355-358.
Doster, S. K., Lozano. M. A., Aguayo, A. J. and Willard, M. B. (1991) Expression of the growth-associated protein Gap-43 in adult rat retinal ganglion cells following axon injury. Neuron. 6, 635-647.
Drager, U. C. and Olsen, J. F. (1981) Ganglion cell distribution in the retina of the mouse. Invest. Ophthalmol. Vis. Sci., 20, 285-295.
Dubois-Dauphin, M., Frankowski, H., Tsujimoto, Y., Huarte, J. and Martinou, J.-C. (1994) Neonatal motoneurons overexpressing the bcl-2 protooncogene in transgenic mice are protected from axotomy-induced cell death. Proc. Narl Acad. Sci. USA, 94, 3309-3313.
Farlie. A., Dringen, R., Rees, S. M., Kannourakis, G. and Bernard, 0. (1995) Transgene expression can protect neurons against development and induced cell death. Proc. Natl Acad. Sci. USA, 92, 43974401.
Garcia, I., Martinou, I., Tsujimoto. Y. and Martinou, J.-C. (1992) Prevention of programmed cell death of sympathetic neurons by bcl-2 proto-oncogene. Science, 258, 302-304.
Garcia-Valenzuela, E., Gorczyca, W., Darzynkiewicz, Z. and Sharma, S. C. (1994) Apoptosis in adult retinal ganglion cells after axotomy. J. Neurobiol., 25, 43 1438.
Greenlund, L. 1.. Deckewerth, T. L. and Johnson, E. M. (1995) Superoxide dismutase delays neuronal apoptosis: a role for reactive oxygen species in programmmed neuronal death. Neuron, 14, 303-3 15.
Hockenbery, D. M., Oltai. Z., Yin X.-M., Milliman, C. and Korsmeyer, S. J. (1993) Bcl-2 functions in an antioxidant pathway to prevent apoptosis. Cell, 75, 241-25 1.
Huxlin, K. R., Dreher. B., Schulz, M., Jervie Sefton, A. and Bennett, M. R. (1995) Effect of collicular proteoglycan on the survival of adult rat retinal ganglion cells following axotomy. Eur: J. Neurosci., 7, 96-107.
Korsmeyer, S. J. (1995) Regulators of cell death. Trends Genet., 11, 101-105. Maffei, L., Carmignoto, G., Perry, V. H., Candeo, P. and Ferrari, G. (1990)
Schwann cells promote the survival of rat retinal ganglion cells after optic nerve section. Proc. Narl Acad. Sci. USA, 87, 185551859,
Mah, S. P., Zhong, L. T., Liu, Y., Roghani, A., Edwards, R. H. and Bredesen, D. E. (1993) The protooncogene bcl-2 inhibits apoptosis in PC12 cells. J . Neurochem., 60, 1183-1 186.
Mansour-Robaey, S., Clarke, D. B., Wang, Y.-C., Bray, G. M. and Aguayo, A. J. (1994) Effects of ocular injury and administration of brain-derived neurotrophic factor on survival and regrowth of axotomized retinal ganglion cells. Proc. Narl Acad. Sci. USA, 91, 1632-1636.
Martinou, I., Fernandez, P.-A., Missotten, M., White, E.. Allet, B., Sadoul, R. and Martinou, J.-C. (1995) Viral proteins EIB19K and p35 protect sympathetic neurons from cell death induced by NGF deprivation. J. Cell
Martinou, J.-C., Dubois-Dauphin, M., Staple, J., Rodriguez, I., Frankowski, H., Missoten, M., Albertini, P., Talabot, D., Catsicas, S., Pietra, C. and Huarte, J. ( I 994) Overexpression of Bcl-2 in transgenic mice protects neurons from naturally occurring cell death and experimental ischemia. Neuron, 13. 1017-1030.
Mey, J. and Thanos, S. (1993) lntravitreal injections of neurotrophic factors support the survival of axotomized retinal ganglion cells in adult rats in vivo. Brain Res., 602. 304-317.
Merry, D. E., Veis, J. D., Hickey, W. F. and Korsmeyer. S. (1994) Bcl-2 protein expression is widespread in the developing nervous system and retained in the adult PNS. Developmenr, 120, 302-31 I .
Oppenheim, R. W. (1991) Cell death during development of the nervous system. Annu. Rev. Neurosci., 14, 453-501.
Oppenheim, R. W., Prevette, D., Tytell, M. and Homma, S. (1990) Naturally occurring and induced neuronal death in the chick embryo in vivo requires protein and RNA synthesis: evidence for the role of cell death genes. Dev. Biol., 138, 104-113.
Osbome, B. S. and Schwartz, L. M. (1994) Essential genes that regulate apoptosis. Trends Cell Biol., 4, 394-398.
Perry, V. H., Henderson, Z. and Linden, R. (1983) Postnatal changes in retinal ganglion cells and optic axon populations in the pigmented rat. J. Comp. Neurol., 219, 3546-3568.
Peters, A., Palay, L. and Webster, H. de F. (1976) The fine structure of the nervous system. In The Neurons and the Supporring Cells. Saunders, Philadelphia.
Porciatti. V., Pizzorusso. T., Martinou, J.-C. and Maffei. L. (1995) Visual responses of axotomized retinal ganglion cells are preserved in transgenic mice overexpressing bcl-2. Soc. Neurosci. Absrr., 21(2), 1555.
Rabacchi, S. A., Bonfanti, L., Liu, X.-H. and Maffei, L. (1994) Apoptotic cell death induced by optic nerve lesion in the neonatal rat. J. Neurosci., 14, 5292-5301.
Ratto, G. M. and Usai, C. (1991) Computer aided tracing and encoding of axonal arborizations. J. Neurosci. Methods, 36, 3 3 4 3 .
Rice, D. S.. Williams, R. W. and Goldowitz, D. (1995) Genetic control of retinal projections in inbred strains of albino mice. J. Comp. Neurol., 354, 459-469.
Sidman. R. (1961) Histogenesis of mouse retina studied with thymidine-H3. In Smelser, G. K. (ed.), The Structure of the Eye. Academic Press, New York, pp. 487-506.
Thanos, S., Bahr, M.. Barde, Y.-A. and Vanselow, J. (1989) Survival and axonal elongation of adult rat retinal ganglion cells. In vitro effects of lesioned sciatic nerve and brain-derived neurotrophic factor. Eur: J. Neurosci., 1, 19-26.
Thanos, S., Mey, J. and Wild, M. (1993) Treatment of the adult retina with microglia-suppressing factors retards axotomy-induced neuronal degradation and enhances axonal regeneration in vivo and in vim. J. Neurosci., 13, 455466.
Vidal-Sanz, M., Bray, G. M., Villegas-Peres, M. P., Thanos, S. and Aguayo, A. J. (1987) Axonal regeneration and synapse formation in the superior colliculus by retinal ganglion cells in the adult rat. J. Neurosci., 7, 2894-2909.
Williams, M. A., Piiion, L. G. P., Linden, R. and Pinto, L. H. (1990) The pearl mutation accelerates the schedule of natural cell death in the early postnatal retina. Exp. Brain Res., 82, 393400.
Biol., 128, 201-208.
Related Documents