Long-range chromatin interactions drive mutant Tert promoter activation Semih Can Akıncılar 1,2 , Ekta Khattar 1 , Priscilla Li Shan Boon 3 , Bilal Unal 1,2 , Melissa Jane Fullwood 3 , Vinay Tergaonkar 1,2,4* 1 Division of Cancer Genetics and Therapeutics, Laboratory of NFκB Signaling, Institute of Molecular and Cell Biology (IMCB), A*STAR (Agency for Science, Technology and Research), Singapore 138673. 2 Department of Biochemistry, Yong Loo Lin School of Medicine, National University of Singapore (NUS), Singapore 117597, Singapore 3 Cancer Science Institute, National University of Singapore, Singapore 4 Centre for Cancer Biology, University of South Australia and SA Pathology, Adelaide, Australia. Running Title: Telomerase re-activation by long-range interaction Key words: Telomerase re-activation, long-range interaction, Tert promoter mutation *Correspondence Vinay TERGAONKAR, [email protected], Institute of Molecular and Cell Biology (A*STAR), Proteos, 61, Biopolis Drive, 138673, Singapore. Ph +65-65869836; Fax +65-67791117. Email: [email protected] Disclosure of Potential Conflicts of Interest No potential conflicts of interest to be disclosed. Research. on June 20, 2018. © 2016 American Association for Cancer cancerdiscovery.aacrjournals.org Downloaded from Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Long-range chromatin interactions drive mutant Tert promoter activation
Semih Can Akıncılar1,2, Ekta Khattar1, Priscilla Li Shan Boon 3, Bilal Unal1,2, Melissa
Jane Fullwood3, Vinay Tergaonkar1,2,4*
1Division of Cancer Genetics and Therapeutics, Laboratory of NFκB Signaling, Institute of Molecular and Cell Biology (IMCB), A*STAR (Agency for Science, Technology and Research), Singapore 138673. 2Department of Biochemistry, Yong Loo Lin School of Medicine, National University of Singapore (NUS), Singapore 117597, Singapore 3 Cancer Science Institute, National University of Singapore, Singapore 4 Centre for Cancer Biology, University of South Australia and SA Pathology, Adelaide, Australia. Running Title: Telomerase re-activation by long-range interaction Key words: Telomerase re-activation, long-range interaction, Tert promoter mutation *Correspondence
Vinay TERGAONKAR, [email protected], Institute of Molecular and Cell Biology (A*STAR), Proteos, 61, Biopolis Drive, 138673, Singapore. Ph +65-65869836; Fax +65-67791117. Email: [email protected] Disclosure of Potential Conflicts of Interest No potential conflicts of interest to be disclosed.
Research. on June 20, 2018. © 2016 American Association for Cancercancerdiscovery.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177

1
Abstract
Cancer-specific Tert promoter mutations (-146C>T and -124C>T) have been linked to
reactivation of epigenetically silenced telomerase reverse transcriptase gene (Tert).
Understanding how these single nucleotide alterations drive Tert reactivation is a
fundamental unanswered question and is key for making successful therapeutics. We
show that unlike on wild-type promoter, recruitment of transcription-factor GABPA
specifically to mutant Tert promoters mediates long-range chromatin interaction,
enrichment of active histone marks and hence drives Tert transcription. CRISPR
mediated reversal of mutant Tert promoters, or deletion of its long-range interacting
chromatin, abrogates GABPA binding, long-range interactions, leading to depletion
of active histone marks, loss of Pol2 recruitment and suppression of Tert
transcription. In contrast, de-novo introduction of Tert promoter mutation enables
GABPA binding and upregulation of Tert via long-range interactions, acquisition of
active histone marks and subsequent Pol2 recruitment. This study provides a unifying
mechanistic insight into activation of mutant Tert promoters across various human
cancers.
Significance statement
This study identifies a key mechanism by which cancer specific mutant Tert
promoters cause reactivation of Telomerase reverse transcriptase (Tert). Since the
mechanism uncovered here is not utilized by promoters that drive Tert in normal
cells, this mechanism could be exploited to make inhibitors which have the potential
to block telomerase function and hence the progression of up to 90% of human
cancers.
Research. on June 20, 2018. © 2016 American Association for Cancercancerdiscovery.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177

2
Introduction
Telomerase is a reverse transcriptase that elongates telomeres and thus
maintains genomic integrity (1-3). It is minimally composed of the catalytic protein
component TERT and a template forming RNA component Terc. Although Terc is
ubiquitously expressed (4), Tert is epigenetically silenced in most adult somatic cells
limiting their replicative lifespan. Up to 90% of human cancers reactivate Tert
expression transcriptionally to reconstitute telomerase enzyme activity, which
subsequently enables replicative immortality (5).
Recently, two cancer-specific somatic mutations in the Tert promoter were
identified (6, 7). These mutations are particularly common in a subset of cancers
including melanoma (74%), glioblastoma (83%), hepatocellular carcinoma (44%) and
urothelial bladder carcinomas (53.5%) (6-10). These mutations cause a Cytosine (C)
to Thymidine (T) transition at -124bp (-124C>T) and -146bp (-146C>T) upstream of
ATG start site resulting in the creation of novel E-Twenty-Six (Ets) transcription
factor binding motifs (6). The Ets family includes 27 members amongst which,
GABPA has been shown to specifically associate with the de novo motif created by
mutations in several cancer cell types (11).
Because the mechanisms of activating transcription from wild type (WT) Tert
promoter in stem cells could be vastly different from those employed by mutant Tert
promoters, understanding how these promoters function is of critical importance.
Chromatin remodeling and epigenetic mechanisms, particularly histone modifications
and DNA methylations, have been shown to regulate Tert transcription (12). -146C>T
and -124C>T mutations are located close to the transcription start site (TSS) of the
Tert gene. It is intriguing to speculate that these promoter mutations may affect
recruitment of epigenetic regulators, which modulate the chromatin structure in order
Research. on June 20, 2018. © 2016 American Association for Cancercancerdiscovery.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177

3
to drive Tert expression. These mutations are majorly heterozygous wherein only
mutant allele specifically recruits of GABPA/B1 and RNA polymerase 2 to drive
transcription and also display active histone marks (13). But what initiates these
epigenetic changes and the nature of these epigenetic changes that eventually convert
the inactive Tert promoter to an active one are not understood at all. In this study, we
addressed these questions and observed that binding of the transcription factor
GABPA to proximal mutant Tert promoter, specifically, initiates and mediates long-
range chromatin interactions and enrichment of active histone marks on these
promoters and hence drives its transcription. The specificity of these events at mutant
Tert promoter provides insights into telomerase reactivation and suggests therapeutic
opportunities for cancer therapy for a wide range of cancers.
Results
Mutant Tert promoters display active histone marks and long-range chromatin
interactions
Cancer-specific mutations in the Tert promoter (-146C>T and -124C>T) create
consensus-binding sites for Ets transcription factors (14). Ets transcription factors are
auto-inhibited and binding of these factors alone is not sufficient to drive
transcriptional activation and it is known that they need to heterodimerize with other
factors for activating transcription (15-17). GABPA and Ets1 have been reported to
specifically interact with the proximal mutant Tert promoters (11, 18) but the
mechanism(s) by which they activate the mutant promoters is not completely
understood. It is known that levels of these factors do not dramatically differ between
normal and cancer cells. Furthermore, there are several Ets/GABPA binding sites on
the wild type promoter which remains dormant in most cell types. Clearly, just
Research. on June 20, 2018. © 2016 American Association for Cancercancerdiscovery.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177

4
creating one more binding of Ets/GABPA on the mutant Tert promoters is not the
entire picture that could explain the mechanism of Tert reactivation by these single
nucleotide changes. Are there uncharacterized events specifically initiated by the
mutant sites and their chromatin context which drives Tert transcription from mutant
Tert promoters? To address these issues, we first asked if cancer specific point
mutations alone are associated with activating epigenetic changes on the mutant Tert
promoters. Figure 1A shows the cell lines used in the study along with Tert promoter
mutation status. Figure 1B shows the schematic outline of Tert promoter. In this study
we divide the promoter in 3 distinct regions; proximal promoter (up to -1 kb), distal
promoter (up to -5kb) and long distance elements (beyond -5kb), upstream of
transcription start site (TSS). The most common Tert promoter mutations are shown
at the -146C>T and -124C>T positions with respect to the ATG translation start site.
We examined the active histone marks H3K4Me3 and H3K9Ac in BLM, A375, T98G
(bearing -146C>T mutation in Tert promoter) and U251 (bearing -124C>T mutation
in Tert promoter) cell lines up to -5kb upstream of Tert gene. Interestingly, we
observed enrichment of both H3K4Me3 and H3K9Ac in the proximal Tert promoter
region of these mutant cell lines (Fig. 1C. Sup. Fig. 1A). We next performed similar
experiments in primary melanocytes with WT Tert promoter (Fig. 1D) and
additionally, we investigated multiple cancer cells including PC3 (prostate cancer),
Fadu (head and neck cancer) and HCT116 colon cancer cell line (Fig. 1D and Sup.
Fig. 1B) wherein Tert is driven by a WT promoter. We observed a trend of lower
enrichment of the active histone marks in active WT Tert promoter from primary
melanocytes and other cancer cells (Fig. 1D). It must be noted that average Tert
expression levels from a panel of cells with mutant and active WT Tert promoter are
not correlative with the mutational status of the promoter as reported previously (Sup.
Research. on June 20, 2018. © 2016 American Association for Cancercancerdiscovery.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177

5
Fig. 1C) (13, 19). These results suggest that comparing Tert levels and epigenetic
marks across cancer cell types is not ideal and the mechanistic insights regarding the
functioning of the mutant and wild type Tert promoters could only be inferred using a
set of isogenic lines wherein cause and effect of making these alterations can be
evaluated.
A recent study by Barutcu et al. suggested that GABP was specifically
enriched in Topologically Associated Domains (TADs) in breast cancer cells,
suggesting that it may also mediate interactions between domains at long distances
(20). To investigate if GABPA bound to mutant Tert promoter could engage in long-
range chromatin interactions, we performed Circular Chromosome Conformation
Capture (4C) assay (21). Figure 1E depicts the intra-chromosomal interactions of the
Tert promoter (chromosome 5) in BLM and A375 cell lines as brought forth by the
4C assay. Y-axis indicates the read counts and x axis indicates distance upstream and
downstream of the Tert promoter which are aligned with respect to the dashed red
line. The top panel (Fig. 1E) indicates the Refseq genes on chromosome 5. Significant
interactions are represented in different shades of red and blue dots for A375 and
BLM cells respectively. Darker dots indicate more significant interactions, indicated
as q values in the color-coded legends. We observed multiple significant interactions
occurring along chromosome 5 upstream and downstream of -146C>T Tert promoter.
Taken together, these findings suggest that mutant Tert promoter displays active
histone marks and long-range chromatin interaction.
Reversing the -146C>T mutant Tert promoter site to WT specifically reverses the
proximal histone marks and alters long-range chromatin interactions
Research. on June 20, 2018. © 2016 American Association for Cancercancerdiscovery.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177

6
To address if the histone marks and chromatin interactions observed on the
mutant promoters are specifically driven by the point mutation in proximal mutant
promoters, we reversed the mutated nucleotide to the WT residue by genome editing
of promoter using CRISPR/Cas9 system (Fig. 2A). Mutant Tert promoter (-146C>T –
labeled as red color) was targeted by single guide-RNA adjacent to the mutant region.
Wild type nucleotide was introduced using a repair template containing homology
regions as shown in Fig. 2A. We generated isogenic lines with mutant Tert promoter
(-146C>T) hereafter referred as BLM6 and with WT Tert promoter (-146C) hereafter
referred as BLM14. Genetic status was confirmed by Sanger sequencing as shown in
Fig.2B (top left panel). As compared to the BLM6, Tert expression was dramatically
reduced in the BLM14 cells (Sup. Fig.2A). Reversal of mutant to WT Tert promoter
also leads to reduced telomerase activity (Sup. Fig. 2B). Early passage cells did not
show significant differences in telomere length however upon long-term culture,
significant telomere attrition was observed in engineered cells (Sup. Fig. 2C-D),
suggesting the functionality of this mutation reversal. We also analyzed BLM14 cells
for Alternative Lengthening Telomeres (ALT) phenotype. There was no co-
localization between TRF2 and PML in BLM6 or BLM14 cells (Sup. Fig. 2E). U2OS
was used as the positive control where we observed very strong co-localization as
reported previously for ALT cell lines (22). We performed RNA-sequencing from
BLM6 and BLM14 cells in order to eliminate possible off-target effects of
CRISPR/Cas9 method. No significant differences were detected in genes that were
predicted to have off-target effects for guide RNA used in CRISPR/Cas9 editing by
RNA sequencing (Sup. Table1). Next, we investigated the epigenetic status of the
proximal Tert promoter in these cell lines. Interestingly, we observed that the
Research. on June 20, 2018. © 2016 American Association for Cancercancerdiscovery.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177

7
proximal active histone marks were significantly reduced upon single nucleotide
reversal (Fig. 2B).
Next, we mapped the long-range intra-chromosomal interactions in these cell
lines using the 4C assay. Fig. 2C-E shows differential intra-chromosomal interactions
of the Tert promoter between BLM6 and BLM14 cells. Red dots indicate significant
and black dots indicate non-significant interactions between mutant and WT Tert
promoter. The intra-chromosomal interactions were dramatically reduced in BLM14
(Fig. 2D) as compared to BLM6 cells (Fig. 2C). Fold interaction differences are
indicated as log2 scale that is shown in lower panel together with Refseq genes that
are located on chromosome 5 (Fig. 2E). As 4C is a semi-quantitative method, to
accurately quantify the differences of these chromatin interactions, we selected 12 of
the interactions that were obtained from 4C analysis and performed 3C-qPCR assays.
Figure 2F shows the strongest five regions interacting with the mutant Tert promoter.
The x-axis indicates the distance of the regions from -146C>T site of the Tert
promoter, which is represented as ‘0’ along with its coordinates below, and y-axis
indicates the binding strength in log10 scale. The strongest interaction was observed
between the Tert promoter and a region 300kb upstream of promoter (chr5:1,556,087-
1,558,758), hereafter referred to as T-INT1 (Fig. 2F). 3C-qPCR assay verified that the
mutant -146C>T mutant Tert promoter in BLM6 cells showed significantly higher
interaction with the T-INT1 intergenic region when compared with the -146C WT
Tert promoter in BLM14 cells (Fig. 2G). Supplementary Table 2 shows the list of
regions with genomic coordinates that were found to differentially interact with the
mutant Tert promoter in BLM6 cells. Since a GABPA binding site is the only
difference between WT and mutant Tert promoter, we speculated that histone marks
Research. on June 20, 2018. © 2016 American Association for Cancercancerdiscovery.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177

8
and long-range interactions uniquely apparent on the mutant promoters might be
initiated/mediated by GABPA binding to this site in the proximal promoter.
Silencing GABPA expression dampens the active chromatin marks as well as
long-range interactions in mutated Tert promoter
As the mutant Tert promoter creates a binding site for the Ets family member
GABPA transcription factor, we analyzed the GABPA recruitment in BLM6 and
BLM14 cells. As shown in figure 3A, GABPA was specifically enriched at the
proximal mutant Tert promoter in BLM6 cells. A similar enrichment pattern was
obtained for RNA polymerase 2 (Pol 2) (Fig. 3B). We next explored the possibility
that GABPA recruitment specifically drives the epigenetic marks in the proximal Tert
promoter region. We depleted GABPA by siRNA in BLM6 cells (Figure 3C). ChIP
performed in GABPA depleted cells which showed reduced recruitment of GABPA to
the mutant Tert promoter along with reduced Pol 2 recruitment (Fig. 3D). This was
associated with reduced Tert expression in GABPA depleted cells (Fig. 3E). Most
importantly, GABPA depletion was associated with reduced enrichment of active
histone marks in the proximal Tert promoter (Fig. 3F).
Mutant Tert promoter showed long-range interaction as observed using 4C
assays (Fig. 2C-E) and GABP has been shown to have the potential for mediating
long-range interactions (20). To investigate if GABPA is a key mediator, we
performed 3C-qPCR assay with GABPA depleted BLM6 cells. We observed
significant reduction of intra-chromosomal interaction between the Tert promoter and
the T-INT1 region upon GABPA depletion (Fig. 3G). To understand the molecular
basis of how GABPA could mediate this long-range interaction, we analyzed the T-
INT1 region for the presence of potential GABPA motifs using the transcription
Research. on June 20, 2018. © 2016 American Association for Cancercancerdiscovery.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177

9
factor motif finder database Jaspar (23). We found multiple GABPA motifs in this
region (Sup. Fig.3A). It is well known that GABPA factors work as tetramers (15-17).
As this region is about 2.5kb long and GABPA sites are spread along the whole
sequence we designed ChIP qPCR primers specific to the 5’ region, central region
and 3’ region. GABPA was specifically enriched at the 5’ region (data not shown).
We examined the GABPA binding to this region in BLM6 and BLM14 cells and
found that it was significantly increased in BLM6 cells (Sup. Fig.3B). Overall, these
results suggest that GAPBA dimers/tetramers bound at long distance, T-INT1 region
(particularly GABPA sites located in 130 to 290bp region according to
chr5:1,556,087) and -146C>T mutant site in Tert promoter provide the stable binding
required for GABPA mediated transcription and this might be the molecular basis of
the specific interaction of the mutant Tert promoter with the 300kb region upstream of
its TSS. Indeed the presence of several GABPA sites in 5’ of the T-INT1 region may
explain why this region might specifically interact with the mutant Tert promoter.
Since the WT Tert promoter does not possess the GABPA motif at -146 position, such
long-range interaction do not take place and the chromatin interaction around the Tert
promoter does not get stabilized for further modification by epigenetic factors. To our
knowledge, this is the first report showing GABPA can indeed function as a mediator
of long distance interactions.
Active histone marks are known to recruit chromatin remodelers such as
BRD4 (24), so we analyzed whether BRD4 is associated with active Tert promoter in
BLM6 cells. We observed strong enrichment of BRD4 at the Tert promoter in BLM6
cells as compared to BLM14 cells (Fig. 3H). Similar patterns of BRD4 recruitment
was observed upon reducing GABPA expression suggesting the essential role for
Research. on June 20, 2018. © 2016 American Association for Cancercancerdiscovery.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177

10
GABPA in initiating the establishment of the chromatin marks on this promoter (Fig.
3I).
The BET domain family inhibitor JQ1 has been shown to regulate GABPA
expression (25). Amongst BET family members, BRD4 has been shown to be the
most potent target of JQ1 inhibitor. Thus, to investigate whether BRD4 affects
GABPA expression, we knocked down BRD4 in BLM6 and BLM14 cells. Figure 3J
shows the knockdown efficiency in BLM6 and 14 cells. We observed significant
reduction in GABPA expression suggesting that BRD4 regulates GABPA expression
independent of Tert promoter status (Fig. 3K). Additionally, we assessed direct BRD4
recruitment to GABPA promoter and observed that it was highly enriched, suggesting
that BRD4 indeed directly regulates GABPA expression (Fig. 3L). However, reducing
BRD4 reduced Tert expression in BLM6 cells (Fig. 3M). This was corroborated with
reduction in telomerase activity upon knocking down GABPA or BRD4 in BLM6
cells (Sup. Fig. 4A). Since, there is no GABPA recruitment on Tert proximal
promoter of BLM14, GABPA reduction due to BRD4 knockdown had no effect in
BLM14 cells. Moreover, epigenetic status of Tert promoter affects the proliferation
rate in BLM6 but not the BLM14 cells with WT Tert promoter (Sup. Fig. 4B-C).
Furthermore, knocking down BRD4 led to significant reduction in active histone
marks in BLM6 cells (Fig. 3N). We also investigated the long-range interaction
changes in Tert promoter upon BRD4 knockdown. We observed significant reduction
in association of proximal Tert promoter with the T-INT1 region as measured by 3C-
qPCR assay, when BRD4 levels were reduced (Fig. 3O). As a control, we also
analyzed the effect of BRD4 knockdown on Tert expression in Fadu cancer cells
bearing active WT Tert promoter. Tert expression remained unaffected upon BRD4
knockdown in Fadu cells (Sup. Fig. 4D-E). There was no effect of knocking down
Research. on June 20, 2018. © 2016 American Association for Cancercancerdiscovery.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177

11
BRD4 or GABPA on alternate splicing of Tert mRNA (Sup. Fig. 4F). This suggests
that depletion of GABPA or BRD4 affects Tert transcription from mutant Tert
promoter. These results could be used to guide us towards therapeutic modalities.
Reversing -146C>T Tert promoter mutation to wild type in A375 melanoma and
T98G glioblastoma cells reduces the active chromatin marks and affects long-
range chromatin interactions
To further validate the novel molecular mechanism mediated by long-range
interactions observed in our assays, and to find the general significance of these
findings, we performed CRISPR mediated reversal of -146C>T mutation in A375
melanoma cells to generate mutant WT Tert promoter containing cells (A375 -146C).
Reversal of Tert promoter mutation in A375 cells reduced the Tert expression,
telomerase activity and significantly diminished the active histone marks (Fig. 4A-C).
ChIP-qPCR analysis showed that Pol 2 binding was significantly reduced in A375 -
146C cells (Fig. 4D). Furthermore, GABPA and BRD4 were significantly enriched in
A375 -146C>T cells as compared to A375 -146C cells (Fig. 4E-F). We performed
3C-qPCR assay in these cells and the A375 -146C cells showed significant reduction
in interaction with the T-INT1 region (Fig. 4G). We also generated WT Tert promoter
containing glioblastoma cell line T98G (T98G -146C) using CRISPR/Cas9 method
(18). T98G cells naturally contain a -146C>T mutation in Tert promoter. We found
that T98G -146C cells showed reduced active histone marks, Pol2, GABPA and
BRD4 in the proximal Tert promoter region (Fig. 4H-J). The 3C-qPCR assay revealed
that the WT Tert promoter in T98G -146C cells showed significant reduction in
interaction with the T-INT1 region as compared with mutant Tert promoter (Fig. 4K).
We conclude that the mechanism by which -146C>T mutation operates is most likely
conserved across cancer types.
Research. on June 20, 2018. © 2016 American Association for Cancercancerdiscovery.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177

12
Introduction of -146C>T mutant site in the wild type Tert promoter generates
long-range chromatin interactions and increases proximal histone marks
To investigate if the cancer-associated single point mutations are sufficient for
causing the changes described above, we next introduced the -146C>T mutation in
HCT116 colon cancer cells with WT Tert promoter to create isogenic lines (HCT116
-146C and HCT116 -146C>T) (Fig. 5A). Cell lines were verified by sequencing and
HCT116 -146C>T cell line was homozygous (data not shown). Introduction of Tert
promoter mutation in HCT116 cells increased the Tert expression, TRAP activity and
significantly enriched the active histone marks (Fig. 5B-D). ChIP analysis showed
that Pol 2 binding was significantly increased at the mutant Tert promoter (Fig. 5E).
Furthermore, GABPA and BRD4 were significantly enriched in mutant Tert promoter
as compared to WT Tert promoter (Fig. 5F-G). GABPA was also strongly associated
with T-INT1 region in HCT116 -146C>T cells (Sup. Fig. 5). We further investigated
whether HCT116 -146C and HCT116 -146C>T cells show differential long-range
interaction. 3C-qPCR assay revealed that mutant Tert promoter in HCT116 -146C>T
cells showed significant interaction with the T-INT1 region (Fig. 5H) which was also
the strongest interaction observed in 3C analysis of BLM cells (Fig. 2G).
As summarized in Fig. 5I, reversing -146C>T mutation to -146C wild type in
BLM, A375 and T98G cells causes reduction in GABPA recruitment and loss of
long-range interaction. This is concomitant with reduction in active histone marks,
Pol 2 recruitment, Tert expression and telomerase activity. In contrast, introduction of
-146C>T mutation in HCT116 cells causes increase in GABPA recruitment and
formation of long-range interaction. This is concomitant with increase in active
Research. on June 20, 2018. © 2016 American Association for Cancercancerdiscovery.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177

13
histone marks, Pol 2 recruitment, Tert expression and telomerase activity as compared
to wild type isogenic counterpart. We conclude that -146C>T Tert promoter mutation
could be sufficient to initiate long-range interaction of this promoter and it could also
be sufficient for activating this promoter via epigenetic modifications initiated and
promoted by stabilizing long-range interaction with T-INT1 region of Tert promoter
(Fig. 1B).
Deletion of T-INT1 region abrogates transcription from both -124C>T and -146
C>T mutant Tert promoters
To verify whether mutant Tert promoters are indeed activated via long-range
interaction with T-INT1 region, we created cells wherein we deleted the T-INT1
region while keeping the proximal mutant Tert promoter intact. We also investigated
the generality of our observations by also including a cell line with -124C>T mutation
in these assays. Figure 6A shows the location of mutation site (-146C>T or -124C>T)
on Tert promoter and the location of the T-INT1 region. Putative GABPA motifs in
T-INT1 region are shown as red dots. CRISPR mediated removal of the interacting
region from three cell lines (BLM and A375 cell lines which have -146C>T mutation
in Tert promoter and U251 cell line which has -124C>T mutation in Tert promoter)
led to the generation of respective isogenic lines that do have T-INT1 region (T-INT1
WT) or cells where the T-INT1 region is deleted (T-INT1 KO) (Figure 6A). Genetic
status was confirmed by genotyping PCR as shown in Sup. Fig. 6A. In BLM cells,
upon removal of T-INT1 region, Tert expression and telomerase activity reduced
dramatically (Fig. 6B-C) without any change in GABPA expression (Sup. Fig. 6B).
We observed a significant reduction in active histone mark H3K4Me3 in proximal
Tert promoter (Fig. 6D). Also Pol 2, GABPA and BRD4 recruitment to Tert promoter
Research. on June 20, 2018. © 2016 American Association for Cancercancerdiscovery.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177

14
was also reduced significantly (Fig. 6E-F and Sup. Fig. 6C). Removal of T-INT1
interaction region in A375 was also associated with reduced Tert expression and
telomerase activity (Fig. 6G-H). Active histone mark H3K4Me3 was reduced
significantly in the proximal Tert promoter of A375 (Fig. 6I). Pol 2 and GABPA
recruitment to Tert promoter was also reduced significantly (Fig. 6J-K).
Interestingly, we observed dramatic reduction in Tert expression and
telomerase activity in U251 T-INT1 KO cell line which harbors -124C>T mutation
(Fig. 6L-M). Similar to melanoma cells, deletion of T-INT1 region decreased the
enrichment of H3K4Me3, Pol2 and GABPA in the proximal Tert promoter of U251
cells (Fig. 6N-P)
These results demonstrate that GABPA recruitment to the mutant Tert
promoter enables long-range interaction of proximal Tert promoter with T-INT1
region and facilitates generation of associated epigenetic changes that drive its
transcription. These results also suggest that this is a general mechanism used by both
-124C>T and the -146C>T mutant promoters. Our results are presented in the form of
a model in figure 7.
Discussion
Cancer specific Tert promoter mutations offer an invaluable starting point to
understand the mechanistic basis of activation of the dormant Tert promoter in
cancers. Our study reveals a novel mechanism utilized by GABPA to activate Tert
transcription. Using 3C assay we have shown that GABPA mediates long-range
chromatin interaction to form a stable complex that drives Tert transcription. Reversal
of mutation in Tert promoter or removal of mutant Tert promoter interacting region
(T-INT1) using genome editing results in loss of GABPA binding, depletion of
Research. on June 20, 2018. © 2016 American Association for Cancercancerdiscovery.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177

15
epigenetic marks and reduction of Tert transcription. Interestingly, introduction of -
146C>T Tert promoter mutation in HCT116 cells could specifically enrich histone
marks and enable long-range interaction. Our findings suggest that GABPA binding
to mutant Tert promoter is an initiating and sufficient event to drive the epigenetic
status of Tert promoter.
Introduction of -146C>T and -124C>T Tert promoter mutations in human
embryonic stem cells and their differentiation them into fibroblasts and neural
progenitor cells revealed that these mutations were capable of overcoming epigenetic
silencing (26). Moreover the Tert expression in these cells was comparable to that
found in cancer cells and could significantly delay telomere shortening induced
senescence. Furthermore, the high frequency of mutations in Tert promoter majorly at
two nucleotide positions (-146C>T, -124C>T) strongly implicates them as driver
events, appearing upon tumor initiation or possibly later in tumor development (27,
28).
Human Tert promoter harbors G-rich region of 12 sequential G-tracts,
encompassing three Sp1 binding sites, and has the ability to form multiple G-
quadruplexes. Masking of the Sp1 binding sites by G quadruplex structure is
predicted to produce significant inhibition of Tert promoter activity (29). The G-
quadruplex structure may also have an important role in regulation of mutant Tert
promoter and it is suggested that Tert promoter mutations may have an impact on this
predicted secondary structure and that the complex relationship between secondary
structure and GABP recruitment could also play a role in regulating Tert expression
(11). The complex regulatory crosstalk between long-range interaction and G
quadruplex structure in regulating mutant versus WT Tert promoter needs further
investigation.
Research. on June 20, 2018. © 2016 American Association for Cancercancerdiscovery.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177

16
Furthermore, it would also be interesting to understand what co-factor(s)
cooperate with GABPA or other Ets family members on the -146C>T and the -
124C>T mutant sites. Clearly having the know-how and the ability to target distinct
mechanisms which operate in cancer cells which are driven by mutant Tert promoter
would be a step forward in designing therapeutics against telomerase in cancer.
Targeting mutant Tert promoter specific pathways that do not impinge on the WT
Tert promoter will help in designing drugs that are less toxic and can be taken over
longer term at a higher dose. With the realization that telomerase inhibition could
yield a magic bullet to treat most human cancers of any origin and carrying any
genetic alteration. Many pharma companies developed and tested various versions of
telomerase inhibitors for over the last decade. Many refinements were made and the
most successful inhibitor by Genron was believed to reach the clinic. But this
inhibitor which inhibits enzymatic function of telomerase, failed clinical trials early
last year. Based on the understanding of how mutant Tert promoters operate, a new
set of inhibitors that limit Tert transcription only in cancer cells with -124C>T and -
146C>T mutations could be designed rapidly. Since these new transcriptional
inhibitors will work to limit levels of TERT and hence telomerase (indirectly limiting
the activity) only in cells with mutation, namely the cancer cells, they may not cause
toxicity to stem cells or other normal somatic cells where continued telomerase
activity is necessary for physiology.
Research. on June 20, 2018. © 2016 American Association for Cancercancerdiscovery.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177

17
Material and methods Cell lines and reagents: A375 and HCT116 cell lines were originally purchased from
ATCC and were a gift from Dr. Shang Li (DUKE-NUS Medical School, Singapore).
Fadu cells were obtained from ATCC. BLM and primary melanocytes were a gift
from Dr. Birgitte Lane (Institute of Medical Biology, Singapore), T98G-WT
(CRISPR engineered wild type Tert promoter) was produced previously (18). PC3
was a gift from Dr. Ernesto Guccione (Institute of Molecular and Cell Biology,
Singapore). All cells were maintained in Dulbecco’s Modified Eagle’s Medium
(DMEM) except PC3 that was maintained in RPMI and supplemented with 10% FBS
(Hyclone), penicillin and streptomycin (Gibco) using standard tissue culture
techniques. Cell lines were not authenticated by us.
Genomic DNA isolation and Sanger Sequencing: Genomic DNAs of cell lines were
isolated with tail lysis buffer supplemented with Proteinase K. DNA was precipitated
with isopropanol and Tert promoter was sequenced as described previously (18) .
Reversal of the Tert promoter mutation by CRISPR/Cas9 editing: -146C>T Tert
promoter mutation in BLM, A375 cell lines were converted back to wild type
promoter sequence (-146C) and -146C was mutated to -146C>T in HCT116 cells as
described previously (18). Briefly, guide RNA specific to Tert promoter was cloned
into pX458 (GFP) plasmid and was co-transfected with repair template (130bp)
containing wild type (-146C) or mutant (-146C>T) Tert promoter sequence. GFP
positive cells were seeded into 96 well plates (1 cell/well) by FACS after 48h and
each clone was screened by PCR and Sanger sequencing.
Tert interaction region removal by CRISPR/Cas9 editing: pX458-GFP plasmid
was modified by removing Cas9 and GFP with EcorI-AgeI restriction enzymes and
inserted Ds-Red monomer in order to prevent off-target effect of excess Cas9 enzyme
Research. on June 20, 2018. © 2016 American Association for Cancercancerdiscovery.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177

18
in the cells (30) and also to be able to observe transfection efficiency of the both
plasmids. Guide-RNA-1 CACCGCCACAAGGAATGCCGTACAT and Guide-
RNA-2 CACCGCCGGCGTGGAGCAATTCCAC targeting Tert interaction region
(Chr5:1,556,087-1,558,758) were cloned into pX458 (GFP) and pX458 (Ds-Red)
plasmids respectively. Cells were transfected in 6-well plate by X-tremeGENE 9
transfection reagent (Roche). GFP and Ds-Red positive cells were sorted into 96 well
plates (1 cell/well) after 48h by FACS and each clone was genotyped by PCR with
primers surrounding T-INT1 region Forward: GTCTGCATGCAATGCTGTC
reverse: GGGGACATTTATGTCTTCTGC. T-INT1 WT PCR product was ~4530bp
and T-INT1 KO PCR product was 550bp.
RNA-Sequencing: RNA-sequencing library was prepared from two replicates of
BLM6 and BLM14 cell line by Illumina Truseq Total RNA sequencing kit according
to manufacturer’s instructions.
RNA-Sequencing Analysis: Raw reads were aligned to the human genome hg19 by
Tophat2. Differential gene expression was performed by Cufflinks and Cuffdiff with
default parameters.
RNA interference and gene expression: For siRNA treatment, cells were transfected
when they reached ~ 60% confluency with si-Control (Dharmacon; D001810-10), si-
Gabpa (Dharmacon; L011662) or si-BRD4 (Dharmacon; L-004937) using X-
tremeGENE siRNA transfection reagent (Roche) according to manufacturer’s
recommendations. Medium was replaced with fresh medium after 8h of incubation
and cells were harvested 24h-48h post-transfection. For gene expression analysis,
RNAs and cDNAs were prepared as described previously (31).
Chromatin Immunoprecipitation (ChIP) assays: ChIP was performed as described
previously (32). Anti-H3K4Me3 (Millipore; 04-745), anti-H3K9Ac (Millipore; 07-
Research. on June 20, 2018. © 2016 American Association for Cancercancerdiscovery.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177

19
352), anti-H3K14Ac (Millipore; 07-353), anti-Pol2 (Santa-Cruz; sc-899), anti-
GABPA (Santa-Cruz; sc-22810), anti-BRD4 (Bethyl; A301-985A) and IgG (Santa-
Cruz) antibodies were used for the immunoprecipitation. After elution, chip eluate
was used for ChIP-qPCR with primers (33) targeting TSS, 0.8kb, 1kb, 2.5kb and 5kb
upstream regions of Tert gene and/or Tert interaction region. Primers were indicated
in Sup. Table 3.
Circular Chromosome Capture Assay (4C): 4C was performed as described
previously (34). Briefly, 107 cells were fixed with 1% formaldehyde for 10 min and
nuclei pellets were isolated after cell lysis with cold lysis buffer (50mM Tris-Hcl pH
7.5, 150mM NaCl, 5mM EDTA, 0.5% NP-40, 1% Triton X-100) supplemented with
protease inhibitors. First step digestion was performed over-night at 37°C with 200
unit HindIII enzyme. Digestion efficiency was measured by RT-qPCR with HindIII
site specific primers. After phenol-chloroform extraction, DNA was ligated over-night
at 16°C by T4 DNA ligase (Fermentas). Following de-crosslinking, DNA was
processed for second digestion with 50 units of DpnII enzyme for over-night at 37°C.
After final ligation 4C template DNAs were quantified by Qubit dsDNA High
sensitivity kit (Thermo Fischer) and preceded for library preparation for sequencing
using Tert promoter specific primers (Sup. Table 3) with Illumina Nextera adapters.
4C sequencing analysis: The quality of the fastq files were evaluated using FastQC
v0.11.3 (35). Illumina Nextera adaptor sequences were removed using scythe v0.991
(36). Tagdust2.3 was used to extract reads that were sandwiched between the bait
region (TERT-promoter + HindIII) and a DpnII cut site (37). The extracted reads
were aligned to the hg19 genome using bowtie2 (38) with very sensitive parameters in
unpaired mode. The aligned reads were filtered for mapping qualities of > 30. r3CSeq
(39) was used in batch mode to call interactions for A375, BLM cells. Plots were
Research. on June 20, 2018. © 2016 American Association for Cancercancerdiscovery.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177

20
generated by FourCSeq of the differential intra-chromosomal interactions of the Tert
promoter between replicates of BLM6 CRISPR mutant and BLM14 CRISPR wild
type (40). The green line shows the distance dependent fit and the blue dashed line
indicates the fit of z-score > 2. Interactions detected by z-score > 2 (p-adjusted value
< 0.05) for at least one replicate are shown as red dots. Fragments not called as
interactions and that do not show significant change between conditions are shown as
black dots. The calculated log2 fold change is shown above the Hg19 Refseq genes in
the regions 1 megabase from the Tert promoter.
Chromosome Conformation Capture Assay (3C): 3C was performed as described
previously (41). Cells were processed in the same way as described in 4C. Following
HindIII digestion and ligation, 3C template DNAs were quantified by Qubit dsDNA
High sensitivity kit (Thermo Fischer). In order to optimize real time PCRs and to
estimate minimal amount of 3C template for quantification in a linear range, we
prepared control template including equal amounts of target regions that we obtained
in 4C analysis. These regions were amplified by region specific primers and purified
from gel and mixed in equimolar amounts. After digestion of these fragments with
HindIII, fragments were ligated by T4 DNA ligase. 3C DNA templates were used for
normalization of 3C-qPCRs. 3C-qPCR reactions were performed by Sybr-Greener kit
(Invitrogen) and samples were normalized by the levels of Tert promoter region 3C-
qPCR primers were indicated in Sup. Table3.
Real Time telomeric repeat amplification protocol (TRAP): Real time TRAP
assay was performed as described previously (42) .
Colony formation assay: Colony formation assay was performed by crystal violet
BLM6 and BLM14 cells were transfected with siControl, siGABPA and siBRD4.
After 24h 500 cells were seeded into 6 well plate. Culture media was refreshed every
Research. on June 20, 2018. © 2016 American Association for Cancercancerdiscovery.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177

21
two days and after 7 days cells were fixed with 75% ethanol for 30 min and stained
with 0.2% crystal violet dye.
Immunofluorescence: ALT-associated PML bodies were visualized via TRF2 and
PML immunofluorescence staining. Cells were grown on chamber slides (Millipore
EZ slides) and fixed with 4% paraformaldehyde for 10 minutes at room temperature.
After washing with PBS, cells were permeabilized with 0.2% Triton X-100 solution
in PBS for 5 minutes at room temperature and were blocked with PBS supplemented
with 0.2% Triton X-100 and 1% BSA (blocking buffer) for 30 minutes at room
temperature. Following incubation, cells were incubated with primary antibodies
TRF2 (Millipore-05-521) and PML (Santa-Cruz-sc-5621), 1/200 and 1/150 dilutions
respectively, in blocking buffer over night at 4°C. Cells were washed 4 times with
blocking buffer for 8 minutes and incubated with secondary antibodies AF488 and
AF555 (1/2000 dilution) (Invitrogen A11001 and A21428) for 1h at room
temperature. After 4 times washing, image acquisition was performed with Zeiss
LSM800, Plan-Apochromat 63x/1.40 aperture of the objective lenses.
Statistical Analysis: Student’s t-test (two-tailed) was performed to determine the
significance of difference for ChIP-qPCR qPCR and RT-TRAP experiments. Results
of each ChIP, gene expression, 4C, 3C assays were obtained from at least two or three
independent experiments as indicated in the figure legends.
Accession Numbers: All data has been uploaded to GEO (GSE77265).
Acknowledgments
S.C.A and B.U. are supported by the SINGA scholarship. We thank the Agency for Science Technology and Research, Singapore (A*Star) for funding and support to the V.T. laboratory. We thank Phua Qian Hua for her help during the genotyping process and Dr. Shang Li (DUKE-NUS Medical School, Singapore) for the southern blot telomere length experiment.
Research. on June 20, 2018. © 2016 American Association for Cancercancerdiscovery.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177

22
Grant Support This work supported by the core budget from Institute of Molecular and Cell Biology (IMCB), Singapore. P.B. and M.J.F are supported by the National Research Foundation (NRF) Singapore through an NRF Fellowship awarded to M.J.F (NRF-NRFF2012-054), and NTU school of biological sciences start-up funds awarded to M.J.F., as well as by funding given to the Cancer Science Institute, NUS, by the NRF and the Ministry of Education, Singapore under the Research Center of Excellence funding, and the RNA Biology Center at the Cancer Science Institute of Singapore, NUS, as part of funding under the Singapore Ministry of Education’s Tier 3 grants. References 1. Moyzis RK, Buckingham JM, Cram LS, Dani M, Deaven LL, Jones MD, et al. A highly conserved repetitive DNA sequence, (TTAGGG)n, present at the telomeres of human chromosomes. Proceedings of the National Academy of Sciences of the United States of America. 1988;85:6622-6. 2. Blackburn EH. The end of the (DNA) line. Nature structural biology. 2000;7:847-50. 3. O'Sullivan RJ, Karlseder J. Telomeres: protecting chromosomes against genome instability. Nat Rev Mol Cell Biol. 2010;11:171-81. 4. Yi X, Shay JW, Wright WE. Quantitation of telomerase components and hTERT mRNA splicing patterns in immortal human cells. Nucleic Acids Res. 2001;29:4818-25. 5. Shay JW, Wright WE. Senescence and immortalization: role of telomeres and telomerase. Carcinogenesis. 2005;26:867-74. 6. Horn S, Figl A, Rachakonda PS, Fischer C, Sucker A, Gast A, et al. TERT promoter mutations in familial and sporadic melanoma. Science. 2013;339:959-61. 7. Huang FW, Hodis E, Xu MJ, Kryukov GV, Chin L, Garraway LA. Highly recurrent TERT promoter mutations in human melanoma. Science. 2013;339:957-9. 8. Killela PJ, Reitman ZJ, Jiao Y, Bettegowda C, Agrawal N, Diaz LA, Jr., et al. TERT promoter mutations occur frequently in gliomas and a subset of tumors derived from cells with low rates of self-renewal. Proc Natl Acad Sci U S A. 2013;110:6021-6. 9. Heidenreich B, Nagore E, Rachakonda PS, Garcia-Casado Z, Requena C, Traves V, et al. Telomerase reverse transcriptase promoter mutations in primary cutaneous melanoma. Nat Commun. 2014;5:3401. 10. Akincilar SC, Unal B, Tergaonkar V. Reactivation of telomerase in cancer. Cell Mol Life Sci. 2016;73:1659-70. 11. Bell RJ, Rube HT, Kreig A, Mancini A, Fouse SD, Nagarajan RP, et al. Cancer. The transcription factor GABP selectively binds and activates the mutant TERT promoter in cancer. Science. 2015;348:1036-9. 12. Sui X, Kong N, Wang Z, Pan H. Epigenetic regulation of the human telomerase reverse transciptase gene: A potential therapeutic target for the treatment of leukemia (Review). Oncol Lett. 2013;6:317-22.
Research. on June 20, 2018. © 2016 American Association for Cancercancerdiscovery.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177

23
13. Stern JL, Theodorescu D, Vogelstein B, Papadopoulos N, Cech TR. Mutation of the TERT promoter, switch to active chromatin, and monoallelic TERT expression in multiple cancers. Genes Dev. 2015;29:2219-24. 14. Hollenhorst PC, McIntosh LP, Graves BJ. Genomic and biochemical insights into the specificity of ETS transcription factors. Annu Rev Biochem. 2011;80:437-71. 15. Skalicky JJ, Donaldson LW, Petersen JM, Graves BJ, McIntosh LP. Structural coupling of the inhibitory regions flanking the ETS domain of murine Ets-1. Protein Sci. 1996;5:296-309. 16. Cowley DO, Graves BJ. Phosphorylation represses Ets-1 DNA binding by reinforcing autoinhibition. Genes Dev. 2000;14:366-76. 17. Chinenov Y, Henzl M, Martin ME. The alpha and beta subunits of the GA-binding protein form a stable heterodimer in solution. Revised model of heterotetrameric complex assembly. J Biol Chem. 2000;275:7749-56. 18. Li Y, Zhou QL, Sun W, Chandrasekharan P, Cheng HS, Ying Z, et al. Non-canonical NF-kappaB signalling and ETS1/2 cooperatively drive C250T mutant TERT promoter activation. Nat Cell Biol. 2015;17:1327-38. 19. Nault JC, Mallet M, Pilati C, Calderaro J, Bioulac-Sage P, Laurent C, et al. High frequency of telomerase reverse-transcriptase promoter somatic mutations in hepatocellular carcinoma and preneoplastic lesions. Nat Commun. 2013;4:2218. 20. Barutcu AR, Lajoie BR, McCord RP, Tye CE, Hong D, Messier TL, et al. Chromatin interaction analysis reveals changes in small chromosome and telomere clustering between epithelial and breast cancer cells. Genome Biol. 2015;16:214. 21. Zhao Z, Tavoosidana G, Sjolinder M, Gondor A, Mariano P, Wang S, et al. Circular chromosome conformation capture (4C) uncovers extensive networks of epigenetically regulated intra- and interchromosomal interactions. Nat Genet. 2006;38:1341-7. 22. Bryan TM, Englezou A, Gupta J, Bacchetti S, Reddel RR. Telomere elongation in immortal human cells without detectable telomerase activity. EMBO J. 1995;14:4240-8. 23. Mathelier A, Fornes O, Arenillas DJ, Chen CY, Denay G, Lee J, et al. JASPAR 2016: a major expansion and update of the open-access database of transcription factor binding profiles. Nucleic acids research. 2016;44:D110-5. 24. Dey A, Chitsaz F, Abbasi A, Misteli T, Ozato K. The double bromodomain protein Brd4 binds to acetylated chromatin during interphase and mitosis. Proc Natl Acad Sci U S A. 2003;100:8758-63. 25. Bhadury J, Nilsson LM, Muralidharan SV, Green LC, Li Z, Gesner EM, et al. BET and HDAC inhibitors induce similar genes and biological effects and synergize to kill in Myc-induced murine lymphoma. Proceedings of the National Academy of Sciences of the United States of America. 2014;111:E2721-30. 26. Chiba K, Johnson JZ, Vogan JM, Wagner T, Boyle JM, Hockemeyer D. Cancer-associated TERT promoter mutations abrogate telomerase silencing. Elife. 2015;4. 27. Vinagre J, Almeida A, Populo H, Batista R, Lyra J, Pinto V, et al. Frequency of TERT promoter mutations in human cancers. Nat Commun. 2013;4:2185.
Research. on June 20, 2018. © 2016 American Association for Cancercancerdiscovery.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177

24
28. Bell RJ, Rube HT, Xavier-Magalhaes A, Costa BM, Mancini A, Song JS, et al. Understanding TERT Promoter Mutations: A Common Path to Immortality. Mol Cancer Res. 2016;14:315-23. 29. Palumbo SL, Ebbinghaus SW, Hurley LH. Formation of a unique end-to-end stacked pair of G-quadruplexes in the hTERT core promoter with implications for inhibition of telomerase by G-quadruplex-interactive ligands. J Am Chem Soc. 2009;131:10878-91. 30. Ran FA, Hsu PD, Wright J, Agarwala V, Scott DA, Zhang F. Genome engineering using the CRISPR-Cas9 system. Nat Protoc. 2013;8:2281-308. 31. Koh CM, Khattar E, Leow SC, Liu CY, Muller J, Ang WX, et al. Telomerase regulates MYC-driven oncogenesis independent of its reverse transcriptase activity. J Clin Invest. 2015;125:2109-22. 32. Ghosh A, Saginc G, Leow SC, Khattar E, Shin EM, Yan TD, et al. Telomerase directly regulates NF-kappaB-dependent transcription. Nat Cell Biol. 2012;14:1270-81. 33. Zhao Y, Cheng D, Wang S, Zhu J. Dual roles of c-Myc in the regulation of hTERT gene. Nucleic acids research. 2014;42:10385-98. 34. Splinter E, de Wit E, van de Werken HJ, Klous P, de Laat W. Determining long-range chromatin interactions for selected genomic sites using 4C-seq technology: from fixation to computation. Methods. 2012;58:221-30. 35. Andrews S. FastQC: a quality control tool for high throughput sequence data. 2010; Available from: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ 36. Buffalo V. Scythe. 2012; Available from: https://github.com/vsbuffalo/scythe 37. Lassmann T. TagDust2: a generic method to extract reads from sequencing data. BMC Bioinformatics. 2015;16:24. 38. Langmead B, Salzberg SL. Fast gapped-read alignment with Bowtie 2. Nat Methods. 2012;9:357-9. 39. Thongjuea S, Stadhouders R, Grosveld FG, Soler E, Lenhard B. r3Cseq: an R/Bioconductor package for the discovery of long-range genomic interactions from chromosome conformation capture and next-generation sequencing data. Nucleic Acids Res. 2013;41:e132. 40. Klein FA, Pakozdi T, Anders S, Ghavi-Helm Y, Furlong EE, Huber W. FourCSeq: analysis of 4C sequencing data. Bioinformatics. 2015;31:3085-91. 41. Hagege H, Klous P, Braem C, Splinter E, Dekker J, Cathala G, et al. Quantitative analysis of chromosome conformation capture assays (3C-qPCR). Nat Protoc. 2007;2:1722-33. 42. Akincilar SC, Low KC, Liu CY, Yan TD, Oji A, Ikawa M, et al. Quantitative assessment of telomerase components in cancer cell lines. FEBS Lett. 2015;589:974-84.
Research. on June 20, 2018. © 2016 American Association for Cancercancerdiscovery.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177

25
Figure Legends Figure 1. Mutant Tert promoter displays active histone marks and distinct long-range interactions: (A) Cell lines that were used in the study with their origin and
Tert promoter status. WT refers to wild type Tert promoter; WT-ALT refers to wild
type Tert promoter with alternate lengthening of telomeres. (B) Schematic view of
Tert promoter. TSS represents the transcription start site. Tert promoter mutations
were indicated as -146C>T and -124C>T. Tert promoter is shown in three parts
including proximal promoter (up to -1kb), distal promoter (-1kb to -5kb) and long
distance elements. (C) ChIP was performed in A375 and BLM melanoma cells
against histone marks (H3K4Me3 and H3K9Ac). (D) ChIP was performed in primary
melanocytes, Fadu and PC3 cell lines against histone marks (H3K4Me3 and
H3K9Ac). Graph shows qPCR analysis with % Input obtained across various regions
of Tert promoter with the distances indicated in boxes below X-axis. Error bars
indicate the mean ± SD of the two independent experiments. (E) 4C sequencing for
long-range interactions of Tert promoter. Plot generated by r3CSeq of chromatin
interactions of the Tert promoter for A375 and BLM cell lines. The top panel shows
the Refseq genes in chromosome 5. The line plots show detected interactions 500 kb
upstream and downstream of the Tert promoter. X axis indicates the distance from
Tert promoter which was shown as ‘0’. Y axis was the read counts for each
interaction. Different shades of red and blue color dots indicate positive interactions
for A375 and BLM cells (average of two replicates) respectively. Darker color
indicates more significant interactions according to q values indicated in the legend. P
values were calculated by two tailed Student’s t test method.
Figure 2. Reversing the Tert promoter mutation to wild-type reverses the active chromatin marks and alters long-range chromatin interactions. (A) CRISPR/Cas9 mediated reversal strategy of mutated -146C>T residue (shown as red
color) to wild type -146C residue (shown as orange color) by repair template
harboring -146C residue. We obtained BLM cells which have undergone CRISPR
process and are mutant for -146C residue and are labeled as BLM6. BLM cells which
have undergone CRISPR process and mutated back to wild-type for -146T residue to -
Research. on June 20, 2018. © 2016 American Association for Cancercancerdiscovery.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177

26
146C residue are labeled as BLM14. (B) DNA chromatograms spanning the Tert
promoter region in BLM6 and BLM14 are shown. ChIP was performed in BLM6 and
BLM14 cells against histone marks (H3K4Me3 and H3K9Ac). Graph shows qPCR
analysis with % Input obtained across various regions of Tert promoter with distances
indicated in boxes below x-axis. Error bars indicate the mean ± SD of the two
independent experiments. P values were calculated by two tailed Student’s t test
method. (C-E) Plot generated by FourCSeq of the differential intra-chromosomal
interactions of the Tert promoter between replicates of BLM6 and BLM14. The green
line shows the distance dependent fit and the blue dashed line indicates the fit of z-
score > 2. Interactions detected by z-score > 2 (p-adjusted value < 0.05) for at least
one replicate are shown as red dots. Fragments not called as interactions and that do
not show significant change between conditions are shown as black dots. (E) The
calculated log2 fold change is shown above the hg 19 Refseq genes in the regions 1
megabase from the Tert promoter. (F) Quantifications of 5 interactions, obtained from
(C-E), were measured by 3C-qPCR in BLM6 cells. (G) Quantification of Tert
promoter interaction with 300kb upstream (chr5:1,556,087-1,558,758) DNA region
(T-INT1) measured by 3C-qPCR in BLM6 and BLM14 cells. Error bars indicate the
mean ± SD of the 3 independent experiments. P values were calculated by two tailed
Student’s t test method.
Figure 3. Silencing GABPA expression dampens the active chromatin marks as well as long-range interactions in mutated Tert promoter. (A-B) ChIP was
performed in BLM6 and BLM14 cells against GABPA and Pol 2 followed by qPCR
with primers specific for Tert promoter region proximal to TSS. Graph shows qPCR
results with fold recruitment over IgG or % input method as indicated in y-axis. (C) GABPA expression analysis in BLM6 cells transfected siControl (siCont) and
siGABPA. (D) ChIP was performed in BLM6 cells transfected with siCont and
siGABPA against IgG, GABPA and Pol 2 followed by qPCR with primers specific
for Tert promoter region proximal to TSS. (E) Tert expression analysis in BLM6 cells
transfected with siCont and siGABPA. (F) ChIP was performed in BLM6 cells
transfected siCont and siGABPA against histone marks (H3K4Me3 and H3K9Ac)
followed by qPCR with primers specific for Tert promoter region indicated in boxes
below x-axis. (G) Quantification of Tert promoter interaction with 300kb upstream
(chr5:1,556,087-1,558,758) DNA region measured by 3C-qPCR in BLM6 cells upon
Research. on June 20, 2018. © 2016 American Association for Cancercancerdiscovery.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177

27
GABPA knockdown. (H) ChIP was performed in BLM6 and BLM14 cells against
BRD4 followed by qPCR with primers specific for Tert promoter region proximal to
TSS. (I) ChIP was performed in BLM6 cells transfected with siControl (siCont) and
siGABPA against BRD4 followed by qPCR with primers specific for Tert promoter
region proximal to TSS. (J) BRD4 expression analysis in BLM6 and BLM14 cells
transfected siCont and siBRD4. (K) GABPA expression analysis in BLM6 and 14
cells transfected with siCont and siBRD4. (L) ChIP was performed against IgG and
BRD4 in BLM6 cells followed by qPCR with primers specific for GABPA promoter.
(M) Tert expression analysis in BLM6 and BLM14 cells transfected with siCont and
siBRD4. (N) ChIP was performed against histone marks (H3K4Me3 and H3K9Ac)
followed by qPCR with primers specific for proximal Tert promoter region in BLM6
cells transfected with siCont and siBRD4. (O) Quantification of Tert promoter
interaction with 300kb upstream T-INT1 region measured by 3C-qPCR in BLM6 and
BLM14 cells with BRD4 knockdown. Error bars indicate the mean ± SD of the two
independent experiments. P values were calculated by two tailed Student’s t test
method.
Figure 4: Reversing the Tert promoter mutation to wild type in A375 melanoma and T98G glioblastoma cell lines reduces the active chromatin marks and affects long-range chromatin interactions. We obtained A375 cells which have undergone
CRISPR process and are mutant for -146C residue and are labeled as A375 -146C>T.
A375 cells which have undergone CRISPR process and mutated back to wild type for
-146T residue to -146C residue are labeled as A375 -146C . (A) Graph shows qPCR
analysis of Tert expression normalized to actin levels. (B) Graph shows telomerase
activity (TRAP) in A375 -146C>T and A375 -146C cells. (C-F) ChIP was performed
in A375 -146C>T and A375 -146C cells against histone marks (H3K4Me3 and
H3K9Ac), Pol2, GABPA and BRD4. Graph shows qPCR analysis with primers
specific to Tert promoter region proximal to TSS. Results were calculated with %
input or fold recruitment over IgG as indicated in y-axis. (G) 3C-qPCR assay was
performed in A375 -146C>T and A375 -146C cells. Quantification of Tert promoter
interaction with T-INT1 region is shown. (H-J) ChIP was performed in T98G -
146C>T and T98G -146C cells against histone marks (H3K4Me3 and H3K9Ac), Pol2
and GABPA. Graph shows qPCR analysis with % input obtained with primers
specific to Tert promoter region proximal to TSS. (K) 3C-qPCR assay was performed
Research. on June 20, 2018. © 2016 American Association for Cancercancerdiscovery.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177

28
in T98G -146C>T and T98G -146C cells. Quantification of Tert promoter interaction
with chr5:1,556,087-1,558,758 region (T-INT1) is shown. n=3 for all the
experiments, error bars indicate mean±SD of 3 independent experiments. P values
were calculated by student’s t test method.
Figure 5: Introducing -146C>T Tert promoter mutation in HCT116 cells increases the active chromatin marks and enables long-range chromatin interaction. (A) CRISPR/Cas9 mediated conversion of -146C residue (orange color)
to mutant -146C>T residue (red color) by repair template harboring -146T nucleotide
is shown. We obtained HCT116 -146C cells which have undergone CRISPR process
and are wild type for -146C residue and are labeled as HCT116 -146C. HCT116 cells
which have undergone CRISPR process and are mutated for -146C residue to -146T
residue and are labeled as HCT116 -146C>T. (B) Graph shows qPCR analysis of
Tert expression normalized to actin levels. (C) Graph shows telomerase activity
(TRAP) in HCT116 -146C and HCT116 -146C>T cells. (D-G) ChIP was performed
in HCT116 -146C and HCT116 -146C>T cells against histone mark H3K4Me3,
H3K9Ac, Pol 2, GABPA and BRD4. Graph shows qPCR analysis with % input
obtained with primers specific to Tert promoter region proximal to TSS. (H) 3C-
qPCR assay was performed in HCT116 -146C and HCT116 -146C>T cells.
Quantification of Tert promoter interaction with T-INT1 region is shown. n=3 for all
the experiments, error bars indicate mean±SD of 3 independent experiments. P values
were calculated by student’s t test method. (I) Figure summarizes the results of
GABPA enrichment, formation of long-range interaction, enrichment of active
histone marks and Pol2, Tert expression and telomerase activity that were obtained
from isogenic cell lines generated by CRISPR/Cas9 editing. “↑” indicates increase
and “↓” indicates decrease.
Figure 6: Removal of T-INT1 region in melanoma and glioblastoma cell lines reverses the active chromatin marks and decreases Tert expression and telomerase activity. (A) Removal of T-INT1 region strategy in -146C>T mutant cell
line is shown by CRISPR/Cas9 editing (left). Removal of T-INT1 region strategy in -
124C>T mutant cell line is shown by CRISPR/Cas9 editing (right). Red dots indicate
putative GABPA motifs in T-INT1 region. We obtained T-INT1 WT cells which have
undergone CRISPR process and are wild type for Chr5:1,556,087-1,558,758 region
Research. on June 20, 2018. © 2016 American Association for Cancercancerdiscovery.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177

29
(T-INT1) are labeled as T-INT1 WT. T-INT1 KO cells which have undergone
CRISPR process and interaction region Chr5:1,556,087-1,558,758 was removed are
labeled as T-INT1 KO. (B) Graph shows qPCR analysis of Tert expression
normalized to actin levels in BLM cells. (C) Graph shows telomerase activity (TRAP)
in BLM T-INT WT and BLM T-INT1 KO cells. (D-F) ChIP was performed in BLM
T-INT1 WT and BLM T-INT1 KO cells against H3K4Me3, Pol2 and GABPA
followed by qPCR with primers specific to Tert promoter region proximal to TSS.
Graph shows qPCR results with % input method. (G) Graph shows qPCR analysis of
Tert expression normalized to actin levels in A375 T-INT1 WT and A375 T-INT1
KO cells. (H) Graph shows telomerase activity (TRAP) in A375 T-INT1 WT and
A375 T-INT1 KO cells. (I-J) ChIP was performed in A375 T-INT1 WT and A375 T-
INT1 KO cells against H3K4Me3 and Pol2 followed by qPCR with primers specific
to Tert promoter region proximal to TSS. Graph shows qPCR results with % input
method. (K) ChIP was performed in A375 T-INT1 WT and A375 T-INT1 KO cells
against GABPA followed by qPCR with primers specific to Tert promoter region
proximal to TSS. Results were analyzed with fold recruitment over IgG with primers
specific to Tert promoter region proximal to TSS. (L) Graph shows qPCR analysis of
Tert expression normalized to actin levels in U251 T-INT1 WT and U251 T-INT1
KO cells. (M) Graph shows telomerase activity (TRAP) in U251 T-INT1 WT and
U251 T-INT1 KO cells. (N-O) ChIP was performed in U251 T-INT1 WT and U251
T-INT KO cells against H3K4Me3 and Pol2 followed by qPCR with primers specific
to Tert promoter region proximal to TSS. Results were analyzed with % input
method. (P) ChIP was performed in U251 T-INT1 WT and U251 T-INT1 KO cells
against GABPA followed by qPCR with primers specific to Tert promoter region
proximal to TSS and results were analyzed with fold recruitment over IgG. N=3 for
all the experiments, error bars indicate mean±SD of 3 independent experiments. P
values were calculated by student’s t-test method.
Figure 7: Molecular mechanism of Tert reactivation by cancer specific Tert promoter mutations. Cells with -146C>T and -124C>T mutations exhibit GABPA
motif that leads to enrichment of active histone marks in the Tert promoter and
formation of a novel intra-chromosomal long-range interaction, between Tert
promoter and chr5:1,556,087-1,558,758 (T-INT1) region, through GABPA
heterodimerization. These epigenetic features drive transcriptional activation of Tert
Research. on June 20, 2018. © 2016 American Association for Cancercancerdiscovery.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177

30
gene by recruitment of Pol2 to the promoter. CRISPR/Cas9 editing of mutant Tert
promoter to wild type, abrogates long range interaction, decreases active histone mark
occupancy in the proximal Tert promoter which causes dramatic reduction of Tert
gene expression. Telomerase active cells with wild type Tert promoter employs
moderate levels of active histone marks which is sufficient to drive transcription of
Tert. Introduction of promoter mutation to these cells increases enrichment of active
histone marks on the proximal Tert promoter and mediates long-range interaction that
enhances expression of Tert gene.
Research. on June 20, 2018. © 2016 American Association for Cancercancerdiscovery.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177

A
C -146C>T Tert promoter
E
Fig1
D -146C Tert promoter
Distal Tert promoter
ATG TSS
-146C>T -78 +1 -2500 -5000 -124C>T
Long Distance Elements
-1000 -800
Proximal Tert promoter
Cell Line Cancer Type Promoter Status
BLM Melanoma -146C>T
A375 Melanoma -146C>T
T98G Glioblastoma -146C>T
U251 Glioblastoma -124C>T
HCT116 Colon WT
Fadu Head and Neck WT
PC3 Prostate WT
HeLa Cervix WT
P.Melanocytes WT
U2OS Osteosarcoma WT-ALT
B
Research. on June 20, 2018. © 2016 American Association for Cancercancerdiscovery.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177

Fig2 A
ATG TSS -146C>T
-78 +1 -2500 -5000 -124
Long Distance Elements
ATG TSS -146C -78 +1 -2500 -5000 -124
Long Distance Elements
Repair Template
-1000 -800
-1000 -800
CRISPR/Cas9
Editing
BLM6 (-146C>T Tert Promoter)
BLM14 (-146C Tert Promoter)
G
C
D
E
F
T-I
NT
1
B
Research. on June 20, 2018. © 2016 American Association for Cancercancerdiscovery.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177
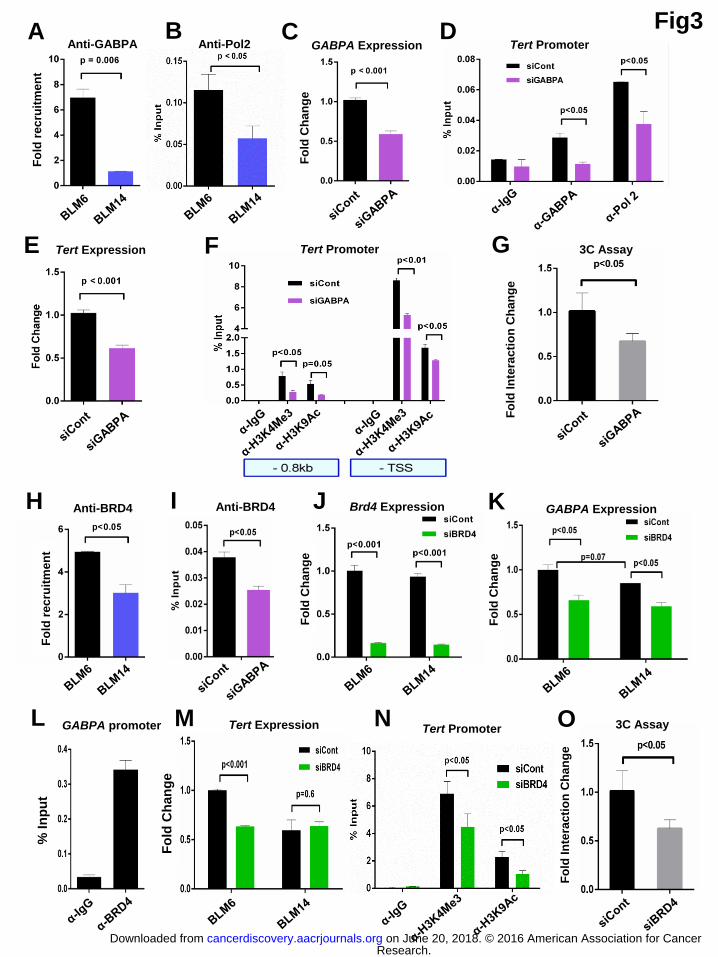
Fig3
Tert Expression E
A Anti-GABPA
Fo
ld r
ec
ruit
men
t C
GABPA Expression
Fo
ld C
ha
ng
e
F Tert Promoter
D Tert Promoter
G
Fo
ld In
tera
cti
on
Ch
an
ge
3C Assay
Anti-BRD4
Fo
ld r
ecru
itm
en
t
H Anti-BRD4 I Brd4 Expression
Fo
ld C
han
ge
Fo
ld In
tera
cti
on
Ch
an
ge
3C Assay O Tert Expression M
Fo
ld C
ha
ng
e
L GABPA promoter
% I
np
ut
GABPA Expression K F
old
Ch
an
ge
J
Anti-Pol2
Tert Promoter
B
N
Research. on June 20, 2018. © 2016 American Association for Cancercancerdiscovery.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177

A B C
D E F G
H I J K
Tert Expression
Fig4 Anti-H3K4Me3 – Anti-H3K9Ac
Anti-Pol2 Anti-GABPA Anti-BRD4 3C Assay
Anti-GABPA Anti-Pol2 3C Assay
TRAP Assay
A375
-146C>T
A375
-146C
T98G
-146C>T
T98G
-146C
α-H3K4Me3 α-H3K9Ac
Anti-H3K4Me3 – Anti-H3K9Ac
α-H3K4Me3 α-H3K9Ac
A375 -146C>T
A375 -146C
A375
-146C>T A375
-146C
A375
-146C>T A375
-146C
A375
-146C>T A375
-146C
T98G
-146C>T
T98G
-146C
T98G
-146C>T
T98G
-146C
A375
-146C>T
A375
-146C
A375
-146C>T
A375
-146C
Research. on June 20, 2018. © 2016 American Association for Cancercancerdiscovery.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177
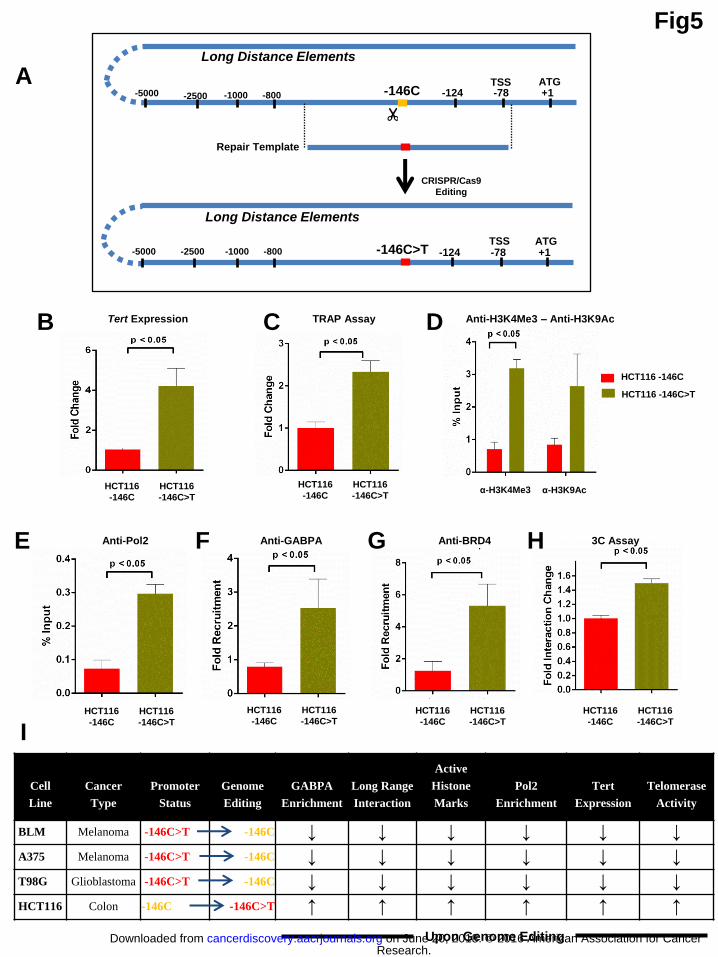
Fig5
B C D
E F G H
Tert Expression TRAP Assay Anti-H3K4Me3 – Anti-H3K9Ac
Anti-Pol2 Anti-GABPA Anti-BRD4 3C Assay
HCT116
-146C>T
HCT116
-146C α-H3K4Me3 α-H3K9Ac
HCT116 -146C>T
HCT116 -146C
A ATG TSS -146C -78 +1 -2500 -5000 -124
Long Distance Elements
ATG TSS -146C>T -78 +1 -2500 -5000 -124
Long Distance Elements
Repair Template
-1000 -800
-1000 -800
HCT116
-146C>T
HCT116
-146C
HCT116
-146C>T
HCT116
-146C
HCT116
-146C>T
HCT116
-146C HCT116
-146C>T
HCT116
-146C
HCT116
-146C>T
HCT116
-146C
CRISPR/Cas9
Editing
Cell
Line
Cancer
Type
Promoter
Status
Genome
Editing
GABPA
Enrichment
Long Range
Interaction
Active
Histone
Marks
Pol2
Enrichment
Tert
Expression
Telomerase
Activity
BLM Melanoma -146C>T -146C ↓ ↓ ↓ ↓ ↓ ↓
A375 Melanoma -146C>T -146C ↓ ↓ ↓ ↓ ↓ ↓
T98G Glioblastoma -146C>T -146C ↓ ↓ ↓ ↓ ↓ ↓
HCT116 Colon -146C -146C>T ↑ ↑ ↑ ↑ ↑ ↑
I
Upon Genome Editing Research.
on June 20, 2018. © 2016 American Association for Cancercancerdiscovery.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177

Fig6
Tert Expression Anti-H3K4Me3 Anti-Pol2 Anti-GABPA
Tert Expression TRAP Assay Anti-H3K4Me3 Anti-Pol2 Anti-GABPA
A
B C D E F
G H I J K
L
TRAP Assay
Tert Expression
BLM
T-INT1
KO
BLM
T-INT1
WT
TRAP Assay Anti-H3K4Me3 Anti-Pol2 Anti-GABPA
M N O P
ATG TSS
-146C>T -78 +1 -2500 -5000
Long Distance
Elements
ATG TSS -146C>T -78 +1 -2500 -5000 -1000 -800
-1000 -800
Chr5:
1,556,087
Chr5:
1,558,858
T-INT1 region
Multiple
GABPA
motif
CRISPR/Cas9
Interaction
Region
Removal
ATG TSS
-124C>T -78 +1 -2500 -5000
Long Distance
Elements
ATG TSS -124C>T -78 +1 -2500 -5000
Long Distance
Elements
-1000 -800
-1000 -800
Chr5:
1,556,087
Chr5:
1,558,858
T-INT1 region
Multiple
GABPA
motif
CRISPR/Cas9
Interaction
Region
Removal
Long Distance
Elements
BLM
T-INT1
KO
BLM
T-INT1
WT
BLM
T-INT1
KO
BLM
T-INT1
WT
BLM
T-INT1
KO
BLM
T-INT1
WT
BLM
T-INT1
KO
BLM
T-INT1
WT
A375
T-INT1
KO
A375
T-INT1
WT
A375
T-INT1
KO
A375
T-INT1
WT
A375
T-INT1
KO
A375
T-INT1
WT
A375
T-INT1
KO
A375
T-INT1
WT
A375
T-INT1
KO
A375
T-INT1
WT
U251
T-INT1
KO
U251
T-INT1
WT
U251
T-INT1
KO
U251
T-INT1
WT
U251
T-INT1
KO
U251
T-INT1
WT
U251
T-INT1
KO
U251
T-INT1
WT
U251
T-INT1
KO
U251
T-INT1
WT
Research. on June 20, 2018. © 2016 American Association for Cancercancerdiscovery.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177
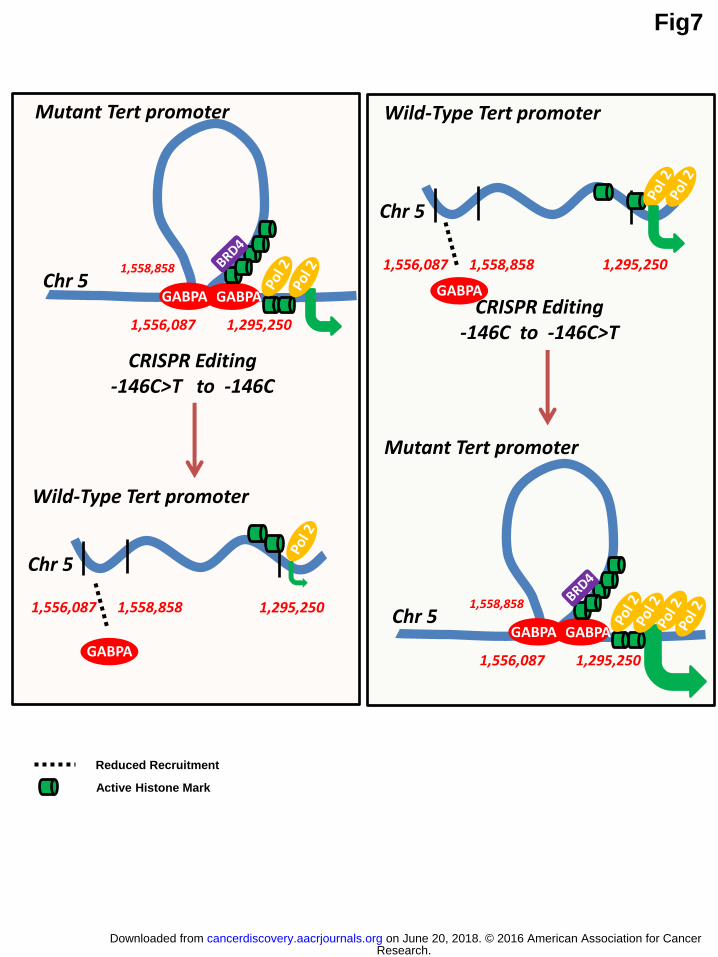
Fig7
1,556,087 1,558,858 1,295,250
Chr 5
Wild-Type Tert promoter
1,556,087 1,558,858 1,295,250
Chr 5
Wild-Type Tert promoter
GABPA
CRISPR Editing -146C>T to -146C
CRISPR Editing -146C to -146C>T
Reduced Recruitment
Active Histone Mark
GABPA GABPA
1,556,087
1,558,858
1,295,250
Chr 5
Mutant Tert promoter
GABPA GABPA
1,556,087
1,558,858
1,295,250
Chr 5
Mutant Tert promoter
GABPA
Research. on June 20, 2018. © 2016 American Association for Cancercancerdiscovery.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177

Published OnlineFirst September 20, 2016.Cancer Discov Semih Akincilar, Ekta Khattar, Priscilla Li Shan Boon, et al. activationLong-range chromatin interactions drive mutant Tert promoter
Updated version
10.1158/2159-8290.CD-16-0177doi:
Access the most recent version of this article at:
Material
Supplementary
http://cancerdiscovery.aacrjournals.org/content/suppl/2016/09/17/2159-8290.CD-16-0177.DC1
Access the most recent supplemental material at:
Manuscript
Authoredited. Author manuscripts have been peer reviewed and accepted for publication but have not yet been
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerdiscovery.aacrjournals.org/content/early/2016/09/17/2159-8290.CD-16-0177To request permission to re-use all or part of this article, use this link
Research. on June 20, 2018. © 2016 American Association for Cancercancerdiscovery.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 20, 2016; DOI: 10.1158/2159-8290.CD-16-0177
Related Documents









![Long Noncoding RNAs, Chromatin, and Developmentdownloads.hindawi.com/journals/tswj/2010/180798.pdf · active chromatin modifications and a more open chromatin conformation[26,39,40,41,42].](https://static.cupdf.com/doc/110x72/5f8885d811957319d07a36bf/long-noncoding-rnas-chromatin-and-active-chromatin-modifications-and-a-more-open.jpg)

