Localized 4 Integrin Phosphorylation Directs Shear Stress–Induced Endothelial Cell Alignment Lawrence E. Goldfinger,* Eleni Tzima,* Rebecca Stockton, William B. Kiosses, Kayoko Kinbara, Eugene Tkachenko, Edgar Gutierrez, Alex Groisman, Phu Nguyen, Shu Chien, Mark H. Ginsberg Abstract—Vascular endothelial cells respond to laminar shear stress by aligning in the direction of flow, a process which may contribute to atheroprotection. Here we report that localized 4 integrin phosphorylation is a mechanism for establishing the directionality of shear stress–induced alignment in microvascular endothelial cells. Within 5 minutes of exposure to a physiological level of shear stress, endothelial 4 integrins became phosphorylated on Ser 988 . In wounded monolayers, phosphorylation was enhanced at the downstream edges of cells relative to the source of flow. The shear-induced 4 integrin phosphorylation was blocked by inhibitors of cAMP-dependent protein kinase A (PKA), an enzyme involved in the alignment of endothelial cells under prolonged shear. Moreover, shear-induced localized activation of the small GTPase Rac1, which specifies the directionality of endothelial alignment, was similarly blocked by PKA inhibitors. Furthermore, endothelial cells bearing a nonphosphorylatable 4(S 988 A) mutation failed to align in response to shear stress, thus establishing 4 as a relevant PKA substrate. We thereby show that shear-induced PKA-dependent 4 integrin phosphorylation at the downstream edge of endothelial cells promotes localized Rac1 activation, which in turn directs cytoskeletal alignment in response to shear stress. (Circ Res. 2008;103:177-185.) Key Words: integrin PKA endothelial Rac GTPase alignment E xposure of endothelial cells to laminar shear stress elicits directional cellular behaviors. The cells assume a polar- ized elongated shape parallel to the direction of flow. 1 When a wound perpendicular to the direction of flow is introduced into an endothelial cell monolayer, the cells at the upstream margin of the wound migrate approximately 1.5 times faster into the wound than those on the downstream margin. 2,3 The aligned morphology associated with laminar flow correlates with responses associated with protection from atherosclero- sis. 4 Thus, the capacity of endothelial cells to respond to laminar flow, to align along the flow, and to migrate plays a role in vascular physiology. In vitro studies have analyzed biochemical signaling events associated with flow-induced endothelial cell alignment. In response to the onset of flow a mechanosensory complex comprising VE-Cadherin, VEGFR2, and PECAM-1 leads to PI3-kinase activation, resulting in activation of integrin ad- hesion receptors. The activated integrins then form new attachments to the subendothelial extracellular matrix. 5 These adhesive events result in precise temporal modulation of Rho GTPase activity, which leads to disassembly and reassembly of actin fibers. In particular, localized activation of Rac1 GTPase at the downstream cell edges is required for the alignment of the actin fibers parallel to the direction of flow. 6 The mechanisms that control this localized Rac1 activation are obscure. The new integrin-mediated adhesions formed in response to shear stress contribute to Rac1 GTPase activation. 7 In migrating cells, 4 integrins induce highly localized Rac1 activation. 8 –11 4 integrins bind to paxillin at the trailing edge of migrating cells leading to suppression of adhesion- mediated Rac1 activation. Phosphorylation of the cytoplas- mic tail of the 4 integrin subunit by protein kinase A (PKA) at Ser 988 is localized to the leading edge of migrating cells where it blocks paxillin binding thus permitting efficient and highly localized Rac1 activation. 7,12,13 Because 4 integrins are expressed in endothelial cells, 14 we suspected that 4 integrin phosphorylation and its effects on localization of Rac1 activity may contribute to endothelial cell responses to shear stress. 15–17 Here we show that PKA-mediated 4 integrin phosphorylation is induced by shear stress at the downstream edge of endothelial cells. This spatially restricted 4 phosphorylation is required for localized activation of Rac1 and for endothelial cell alignment in response to shear. Thus, localized 4 integrin phosphorylation informs the endothelial cell about the direction of blood flow, thereby Original received December 11, 2007; resubmission received March 26, 2008; revised resubmission received May 8, 2008; accepted June 16, 2008. From the Divisions of Rheumatology and Hematology-Oncology, Department of Medicine (L.E.G., R.S., K.K., E.T., M.H.G.), the Department of Physics (E.G., A.G.), and the Department of Bioengineering and The Whitaker Institute of Biomedical Engineering (P.N., S.C.), University of California, San Diego; the Department of Cell and Molecular Physiology (E.T.), Carolina Cardiovascular Biology Center, University of North Carolina, Chapel Hill; and the Core Microscopy Facility (W.B.K.), Scripps Research Institute, La Jolla, Calif. *These authors contributed equally to this study. Correspondence to Lawrence E. Goldfinger, Sol Sherry Thrombosis Research Center and Department of Anatomy & Cell Biology, Temple University School of Medicine, 3400 N. Broad Street, OMS 415, Philadelphia, PA 19140. E-mail [email protected] © 2008 American Heart Association, Inc. Circulation Research is available at http://circres.ahajournals.org DOI: 10.1161/CIRCRESAHA.108.176354 177 by guest on July 5, 2015 http://circres.ahajournals.org/ Downloaded from by guest on July 5, 2015 http://circres.ahajournals.org/ Downloaded from by guest on July 5, 2015 http://circres.ahajournals.org/ Downloaded from by guest on July 5, 2015 http://circres.ahajournals.org/ Downloaded from by guest on July 5, 2015 http://circres.ahajournals.org/ Downloaded from by guest on July 5, 2015 http://circres.ahajournals.org/ Downloaded from by guest on July 5, 2015 http://circres.ahajournals.org/ Downloaded from by guest on July 5, 2015 http://circres.ahajournals.org/ Downloaded from by guest on July 5, 2015 http://circres.ahajournals.org/ Downloaded from by guest on July 5, 2015 http://circres.ahajournals.org/ Downloaded from by guest on July 5, 2015 http://circres.ahajournals.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Localized �4 Integrin Phosphorylation Directs ShearStress–Induced Endothelial Cell Alignment
Lawrence E. Goldfinger,* Eleni Tzima,* Rebecca Stockton, William B. Kiosses, Kayoko Kinbara,Eugene Tkachenko, Edgar Gutierrez, Alex Groisman, Phu Nguyen, Shu Chien, Mark H. Ginsberg
Abstract—Vascular endothelial cells respond to laminar shear stress by aligning in the direction of flow, a process whichmay contribute to atheroprotection. Here we report that localized �4 integrin phosphorylation is a mechanism forestablishing the directionality of shear stress–induced alignment in microvascular endothelial cells. Within 5 minutes ofexposure to a physiological level of shear stress, endothelial �4 integrins became phosphorylated on Ser988. In woundedmonolayers, phosphorylation was enhanced at the downstream edges of cells relative to the source of flow. Theshear-induced �4 integrin phosphorylation was blocked by inhibitors of cAMP-dependent protein kinase A (PKA), anenzyme involved in the alignment of endothelial cells under prolonged shear. Moreover, shear-induced localizedactivation of the small GTPase Rac1, which specifies the directionality of endothelial alignment, was similarly blockedby PKA inhibitors. Furthermore, endothelial cells bearing a nonphosphorylatable �4(S988A) mutation failed to align inresponse to shear stress, thus establishing �4 as a relevant PKA substrate. We thereby show that shear-inducedPKA-dependent �4 integrin phosphorylation at the downstream edge of endothelial cells promotes localized Rac1activation, which in turn directs cytoskeletal alignment in response to shear stress. (Circ Res. 2008;103:177-185.)
Key Words: integrin � PKA � endothelial � Rac GTPase � alignment
Exposure of endothelial cells to laminar shear stress elicitsdirectional cellular behaviors. The cells assume a polar-
ized elongated shape parallel to the direction of flow.1 Whena wound perpendicular to the direction of flow is introducedinto an endothelial cell monolayer, the cells at the upstreammargin of the wound migrate approximately 1.5 times fasterinto the wound than those on the downstream margin.2,3 Thealigned morphology associated with laminar flow correlateswith responses associated with protection from atherosclero-sis.4 Thus, the capacity of endothelial cells to respond tolaminar flow, to align along the flow, and to migrate plays arole in vascular physiology.
In vitro studies have analyzed biochemical signaling eventsassociated with flow-induced endothelial cell alignment. Inresponse to the onset of flow a mechanosensory complexcomprising VE-Cadherin, VEGFR2, and PECAM-1 leads toPI3-kinase activation, resulting in activation of integrin ad-hesion receptors. The activated integrins then form newattachments to the subendothelial extracellular matrix.5 Theseadhesive events result in precise temporal modulation of RhoGTPase activity, which leads to disassembly and reassemblyof actin fibers. In particular, localized activation of Rac1GTPase at the downstream cell edges is required for the
alignment of the actin fibers parallel to the direction of flow.6
The mechanisms that control this localized Rac1 activationare obscure.
The new integrin-mediated adhesions formed in responseto shear stress contribute to Rac1 GTPase activation.7 Inmigrating cells, �4 integrins induce highly localized Rac1activation.8–11 �4 integrins bind to paxillin at the trailing edgeof migrating cells leading to suppression of adhesion-mediated Rac1 activation. Phosphorylation of the cytoplas-mic tail of the �4 integrin subunit by protein kinase A (PKA)at Ser988 is localized to the leading edge of migrating cellswhere it blocks paxillin binding thus permitting efficient andhighly localized Rac1 activation.7,12,13 Because �4 integrinsare expressed in endothelial cells,14 we suspected that �4integrin phosphorylation and its effects on localization ofRac1 activity may contribute to endothelial cell responses toshear stress.15–17 Here we show that PKA-mediated �4integrin phosphorylation is induced by shear stress at thedownstream edge of endothelial cells. This spatially restricted�4 phosphorylation is required for localized activation ofRac1 and for endothelial cell alignment in response to shear.Thus, localized �4 integrin phosphorylation informs theendothelial cell about the direction of blood flow, thereby
Original received December 11, 2007; resubmission received March 26, 2008; revised resubmission received May 8, 2008; accepted June 16, 2008.From the Divisions of Rheumatology and Hematology-Oncology, Department of Medicine (L.E.G., R.S., K.K., E.T., M.H.G.), the Department of
Physics (E.G., A.G.), and the Department of Bioengineering and The Whitaker Institute of Biomedical Engineering (P.N., S.C.), University of California,San Diego; the Department of Cell and Molecular Physiology (E.T.), Carolina Cardiovascular Biology Center, University of North Carolina, Chapel Hill;and the Core Microscopy Facility (W.B.K.), Scripps Research Institute, La Jolla, Calif.
*These authors contributed equally to this study.Correspondence to Lawrence E. Goldfinger, Sol Sherry Thrombosis Research Center and Department of Anatomy & Cell Biology, Temple University
School of Medicine, 3400 N. Broad Street, OMS 415, Philadelphia, PA 19140. E-mail [email protected]© 2008 American Heart Association, Inc.
Circulation Research is available at http://circres.ahajournals.org DOI: 10.1161/CIRCRESAHA.108.176354
177 by guest on July 5, 2015http://circres.ahajournals.org/Downloaded from by guest on July 5, 2015http://circres.ahajournals.org/Downloaded from by guest on July 5, 2015http://circres.ahajournals.org/Downloaded from by guest on July 5, 2015http://circres.ahajournals.org/Downloaded from by guest on July 5, 2015http://circres.ahajournals.org/Downloaded from by guest on July 5, 2015http://circres.ahajournals.org/Downloaded from by guest on July 5, 2015http://circres.ahajournals.org/Downloaded from by guest on July 5, 2015http://circres.ahajournals.org/Downloaded from by guest on July 5, 2015http://circres.ahajournals.org/Downloaded from by guest on July 5, 2015http://circres.ahajournals.org/Downloaded from by guest on July 5, 2015http://circres.ahajournals.org/Downloaded from

acting as a “weather vane” of shear stress–induced endothe-lial cell alignment.
Materials and MethodsComplementary DNAs and Other ReagentsGlutathione-S-transferase fusion of the CS-1 fragment of humanfibronectin was generated as described.18 Collagen, �5, and �2integrin antibodies (BD Biosciences), HP2/1 �4 integrin antibodies(Immunotech), and rat �-mouse CD31 antibodies (Invitrogen) werepurchased. PS�4 monoclonal antibodies to Ser988-phospho-�4 inte-grin were as described.19
Generation of �4 Integrin-Null and S988AKnock-In MiceC57/Bl6 mice harboring an �4 locus flanked by loxP sites were asdescribed.20 Generation of �4(S988A) mice is described in the onlinesupplement (available online at http://circres.ahajournals.org).
Primary Endothelial Cell Isolation andCell CultureJurkat T leukemia cells and human microvascular endothelial cellswere maintained as described.13,21 Primary pulmonary microvascularendothelial cells were isolated from �4(S988A) and �4 (fl/fl) mice asdescribed21 and in the online supplement.
Shear Stress Assays, Immunocytochemistry, andFluorescence Resonance Energy TransferDescribed in the online supplement.
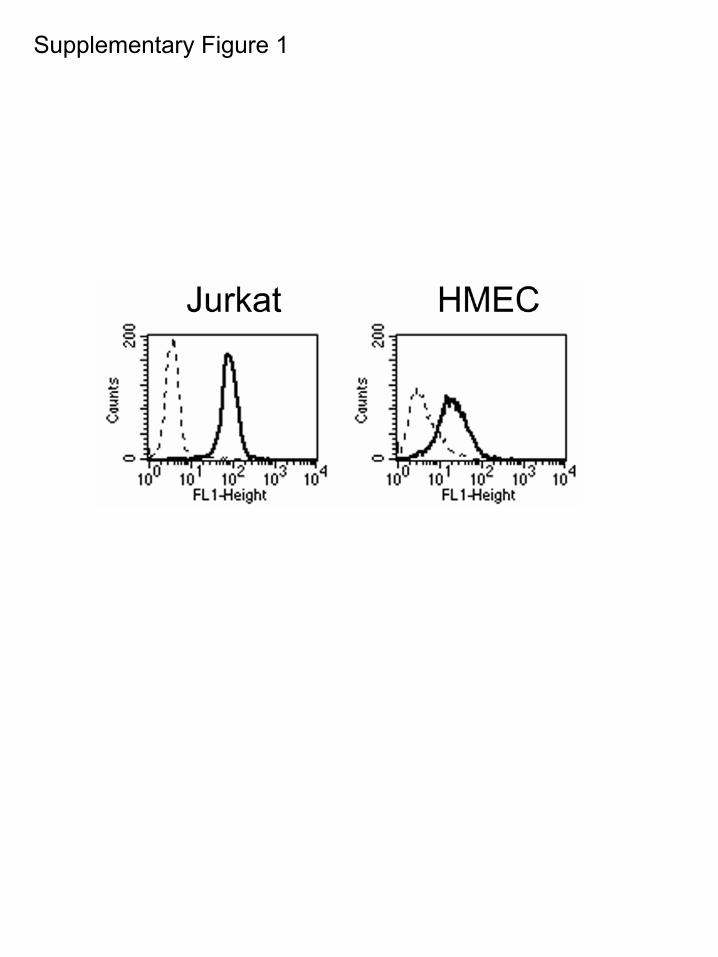
ResultsShear Stress Induces Phosphorylation of �4Integrins at the Downstream Edge ofEndothelial CellsTo assess a potential role for �4 integrin phosphorylation inendothelial cell alignment in response to shear stress, we firstverified the expression of �4 integrins in immortalizedhuman microvascular endothelial cells (HMECs) by fluores-cence-activated cell scanning analysis (FACScan). HMECsexpressed moderate levels of �4 integrin (�20% as comparedto Jurkat T cells; supplemental Figure I). To investigate therole for �4 integrin phosphorylation in endothelial cellresponses to shear stress, we plated HMECs onto coverslipscoated with the �4-binding CS-1 fragment of fibronectin andsubjected the monolayers to a laminar shear stress at 12dynes/cm2 for time intervals ranging from 5 to 30 minutes.After the shear exposure, cells were fixed immediately andstained with PS�4, a monoclonal antibody specific for �4integrin phosphorylated at Ser988.12 A dramatic increase inphosphorylation of �4 in endothelial cells was observed afterall times of exposure to shear, starting from the shortest testedtime of 5 minutes (Figure 1). We observed that �4 phosphor-ylation in response to shear was localized to cell boundaries,in particular at cell edges orthogonal to the direction of flow(Figure 1A.) Labeling with secondary antibody conjugatesalone showed undetectable staining (our unpublished results).Antibodies to �4 integrin uniformly labeled the whole cellperiphery (Figure 1C), indicating that the increased concen-tration of phospho-�4 at the cell edges was attributable toincreased �4 phosphorylation rather than a higher localconcentration of �4. Furthermore, we detected a 1.4-foldincrease in total cellular �4 phosphorylation after 5 minutes
shear stress by Western blotting of �4 immunoprecipitates(Figure 1B), confirming that shear stress upregulates �4phosphorylation.
The foregoing experiments showed that shear induced �4phosphorylation at the cell edges perpendicular to the flowdirection, but failed to establish whether it was occurringpreferentially on the proximal or distal side. To determineprecisely the relationship of shear-induced �4 phosphoryla-tion to flow direction, we generated scratch wounds orthog-onal to the flow direction in confluent monolayers ofHMECs, immediately subjected the wounded monolayers toshear stress, and stained for phospho-�4. Cells at the woundmargins on both sides of the scratch wound were scored forphospho-�4 staining. In the absence of shear, �4 phosphor-ylation was observed in �30% of cells at the wound margin.Application of shear induced increased �4 phosphorylation atthe downstream cell edges at the wound margin proximal tothe flow source but not on the upstream side of cells distal tothe flow source. The �4 phosphorylation was observed in65% (�2.9, P�0.22) of cells at the proximal wound marginbeginning 5 minutes after application of shear and persistingfor at least 30 minutes (Figure 1D). In contrast, �4 phosphor-ylation at the upstream edges of cells at the distal margin ofthe wound showed a trend toward inhibited phosphorylation(13% positive �0.86, P�0.08, Figure 1D). Similarly,phospho-�4 was rarely observed at cell edges lateral withrespect to flow direction (16% positive, unpublished results),indicating that flow increases �4 phosphorylation at thedownstream edges in endothelial cells.
Protein Kinase A Activity Is Required forShear-Induced �4 PhosphorylationPKA phosphorylates �4 integrins at Ser988 and inhibition ofPKA activity blocks �4 phosphorylation in fibroblasts and Tcells.12,19 To assess the contribution of PKA to shear-induced�4 phosphorylation in endothelial cells, we pretreated cellsplated on CS-1 with the PKA inhibitor, H-89, for 15 minutes,scratch-wounded and applied flow in medium containing theinhibitor. Cultures were fixed and stained for phospho-�4.Five-minute exposure to shear induced �4 integrin phosphor-ylation at the downstream cell edges, and addition of H-89 tothe flow medium abrogated the �4 phosphorylation response(Figure 2). At the upstream and lateral cell edges facing theinterior of a wound, where �4 phosphorylation was low,treatment with H-89 had no apparent effect on the levels ofphospho-�4 staining. As additional confirmation of this effectof PKA inhibition, we repeated these experiments using asecond pharmacological PKA inhibitor, KT-5720 at1 �mol/L; it also blocked shear-induced �4 phosphorylation(result not shown). Thus, shear-induced �4 phosphorylationrequires PKA activity.
Localized Rac1 Activation After Shear StressRequires PKA ActivityPKA-dependent �4 integrin phosphorylation helps to localizeRac1 activation to the leading edge of migrating cells.7,12
Furthermore, Rac1 becomes activated at the downstreamedge in a subconfluent endothelial monolayer within fiveminutes of exposure to laminar shear stress.6 This transient
178 Circulation Research July 18, 2008
by guest on July 5, 2015http://circres.ahajournals.org/Downloaded from

localized Rac1 activation is required for subsequent stressfiber alignment. Using a FRET-based assay, we found thatshear stress induced a polarized increase of activated GTP-bound Rac1 at the downstream edge of HMECs as previouslyreported.6 This polarized Rac activation occurred concur-rently with shear-mediated �4 integrin phosphorylation (Fig-ures 3 and 1D). Treatment with the PKA inhibitor (H-89) thatblocked shear-induced �4 phosphorylation inhibited the lo-calized Rac1 activation, indicating that PKA activity isrequired for the shear induction of polarized Rac1 activationat the downstream edge of endothelial cells (Figure 3).
Shear-Induced Cell Alignment of the ActinCytoskeleton Requires PKA ActivityEndothelial cells respond to prolonged exposure to shearstress by remodeling their actin cytoskeleton along a domi-
nant longitudinal cell axis, a phenomenon referred to asendothelial cell alignment.22 This response requires dynamicspatio-temporal regulation of Rac1 activation, as expressionof either constitutively active or dominant negative Rac1blocks the alignment.6 Because PKA activity was required forpolarized Rac1 activation, we assessed whether PKA activityis necessary for alignment. HMECs were subjected to shear,and actin filaments were labeled with rhodamine-phalloidin.Cells subjected to prolonged (20 hour) laminar shear stress at12 dynes/cm2 developed an elongated bipolar shape withactin stress fibers aligned in the direction of flow. However,alignment and elongation did not occur if H-89 was added tothe flow medium (Figure 4A). To quantify this morphologicalobservation, we assessed stress fiber alignment by measuringthe angles of actin filament bundles relative to the direction offlow.
Figure 1. �4 integrin is phosphorylated at downstream cell edges in response toshear stress. A and C, HMECs were seeded onto CS-1-coated glass slides, confluent(A) or scratch wounded (C), and fixed without any exposure to flow (T�0) or immedi-ately after 5-minute exposure to flow of culture media at 12 dynes/cm2 (T�5). Sam-ples were labeled with PS�4 antibodies (A, upper panels in C) or total �4 integrin(lower panel in C). The black arrow indicates laminar flow direction. White arrowheadsindicate regions of concentrated �4 integrin phosphorylation or total �4 integrin. Bar, 10 �m. B, �4 immunoprecipitates from HMECssubjected to shear for 0 or 5 minutes, blotted with total �4 or PS�4 antibodies. Densitometric ratios of band intensities (PS�4/total,shear:static) are indicated. D, HMECs plated on CS-1–coated slides were scratch wounded (top-to-bottom), subjected to flow (left-to-right), then stained with PS�4 antibodies. Cells at the wound margin were scored by blinded observers for cell periphery phospho-�4staining. The black arrow shows laminar flow direction. White arrows indicate wound margins. Arrowheads indicate regions of concen-trated �4 integrin phosphorylation.
Goldfinger et al �4 Integrin Phosphorylation in Endothelium 179
by guest on July 5, 2015http://circres.ahajournals.org/Downloaded from

The average angles from the flow direction of actinfilaments in control and H-89–treated cells under staticconditions were 42�1.6 (17.15°�S.D.) and 41�1.1°(11.58°�S.D.), respectively, both very close to the value of45° expected for randomly oriented fibers. Thus, actin fibersin static cells were not aligned. Actin fibers in control cellssubjected to shear aligned to an average angle of 14.3�0.69°(9.23°�S.D.) from the flow direction, indicating a major biasin the alignment of actin filaments toward the flow direction.Furthermore, the reduction in standard deviation(17.1539.23) indicates a marked reduction in the variabilityof the orientation of the actin fibers, providing an independentmeasure of the alignment response. In contrast, actin fila-ments in sheared H-89–treated cells aligned to a much lesserextent with an average radial displacement of 38�1°(10.61°�S.D.) (Figure 4B and supplemental Table I). Similarto H-89, addition of KT-5720 also dramatically reduced thealignment (supplemental Table I). Thus, PKA activity isrequired for actin stress fiber reorientation and morphologicalalignment of endothelial cells in response to shear stress.
�4 Integrin Is Required forShear-Induced AlignmentThe experiments reported above showed that PKA activationwas required for localized �4 integrin phosphorylation, forlocalized GTP loading of Rac1, and for cytoskeletal align-ment in response to shear stress. These results suggested that�4 integrin phosphorylation may provide cues for endothelialcell reorientation and stress fiber alignment. Integrin ligationby the subendothelial matrix is necessary for shear-inducedRac1 activation,6 and thus presumably for alignment. There-fore we hypothesized that blocking �4 integrin binding toligands in the extracellular matrix would inhibit stress fiberalignment induced by shear. To test this hypothesis, we
subjected confluent HMEC monolayers cultured on slidescoated with fibronectin (a ligand for �4 and �5 integrins23) toshear stress for 20 hours in medium containing either controlIgG or function-blocking antibodies to �4, �5, or �2 integrinsubunits. In all cases the cells remained attached to thesubstratum in monolayers. After shear exposure, cellularalignment was observed in the presence of control IgG orantibodies to �2 or �5 integrin subunits, with the mean stressfiber angles relative to the flow direction of 20.8�0.42°(2.95°�S.D.), 23�1.6° (11.46°�S.D.), and 26�1.6°(13.84°�S.D.), respectively (Figure 5). However, blockingantibodies to �4 integrins inhibited endothelial elongationand stress fiber reorientation in the flow direction, with themean angle�SEM of 48�2.4° (16.68°�S.D.; Figure 5). Inseparate control experiments, these antibodies blocked theability of endothelial cells to adhere to the relevant substratesCS-1 (anti-�4), the cell-binding domain of fibronectin (anti-�5), or collagen (anti-�2; supplemental Figure II). These dataindicate that �4 integrin interaction with the subendothelialmatrix is required for shear-induced cytoskeletal alignment inmicrovascular endothelial cells.
To verify separately that �4 integrins are required forshear-induced alignment, we generated endothelial cells de-ficient in �4 integrin expression and compared the alignmentresponses of these cells with those of �4-expressing endothe-lial cells. To do this, we isolated pulmonary endothelial cells(MLECs) from the lungs of wild-type mice and mice with aconditional �4-null allele in which exon 28 (which includesthe �4 polyadenylation signal) is flanked by loxP recombi-nation sites.20 CD31-positive cell cultures were then infectedwith adenovirus encoding Cre recombinase, leading to loss ofsurface �4 in the mutant cells as determined by FACS(supplemental Figure III); however, these cells adhered andspread on fibronectin-coated surfaces (Figure 5B), presum-
Figure 2. Protein kinase A activity is required for shear-induced �4 phosphorylation. Monolayers of HMECs seeded on CS-1 werescratch wounded and subjected to flow in medium �100 �mol/L H-89 for 5 minutes, then fixed and stained with PS�4 antibodies(upper panels) or �4 antibodies (lower panel). The black arrow shows laminar flow direction. Arrowheads indicate �4 phosphorylationat the cell periphery. Bar, 10 �m.
180 Circulation Research July 18, 2008
by guest on July 5, 2015http://circres.ahajournals.org/Downloaded from

ably via other fibronectin-binding integrins. Wild-type and�4-null MLECs were plated onto fibronectin-coated slidesand subjected to laminar shear stress for 20 hours. Wild-typeMLECs either infected with Cre or with empty adenovirusaligned in response to shear stress (Figure 5B). In contrast,the �4-null (�4 fl/fl CRE) endothelial cells maintained apolygonal morphology and did not align in the flow direction.These cells also displayed few discernable stress fibers,suggesting a disruption in the ability of actin to reorganize in�4-null cells in response to shear. Thus, �4 integrin contrib-utes to shear stress-induced morphological alignment inprimary pulmonary endothelial cells.
�4 Integrin Phosphorylation Is Necessaryfor AlignmentAs shown above, PKA activity is required for �4 phosphor-ylation in response to shear in wounded monolayers and forcell alignment in prolonged shear. Therefore, we considered
that �4 phosphorylation may be an early event in establishingthe directionality of shear stress-induced cytoskeletal align-ment. To test this possibility, we isolated endothelial cellsfrom the lungs of mice expressing �4 integrin that harbors aSer9883Ala mutation (S988A), which disrupts the PKA phos-phorylation site in �4.
The antiphospho-�4 PS�4 antibody did not react in West-ern blots of lysates from endothelial cells derived from thesemice, nor did these cells display any edge PS�4 staining insheared, wounded cultures (our unpublished results andsupplemental Figure IV). Whereas wild-type endothelial cellsexposed to laminar shear for 20 hours aligned, �4(S988A)endothelial cells did not (Figure 6A). As shown in Figure 6B,the average angle from the flow direction of actin filaments inwild-type and mutant cells under static conditions was37.6�0.85° (10.7°�S.D.) and 44�1.° (12.2°�S.D.), respec-tively (Figure 6B). Filaments in wild-type cells subjected toshear stress were orientated at an average angle of16.9�0.25° (3.2°�S.D.) from the flow direction, which wasclose to the value obtained earlier (14.3�0.69 (9.23°�S.D.),indicating that most actin filaments aligned parallel to theflow direction. In contrast, filaments in �4(S988A) endothelialcells subjected to shear stress maintained an average angle of47�0.92° (14.3°�S.D.; Figure 6B), showing that they didnot align after shear. Thus, �4 integrin phosphorylation isrequired for stress fiber alignment induced by shear stress.
As a further test of the role of �4 integrin phosphorylationin the endothelial cell responses to shear stress, we assessedthe effect of the �4(S988A) mutation on shear-induced Rac1activation in endothelial cells. HMECs were cotransfectedwith a Rac FRET reporter plasmid and plasmids encodingwild-type �4 or �4(S988A). These cells were plated on �4ligands, scratch wounded, and immediately subjected to 12dynes/cm2 shear stress for 5 minutes, fixed, and analyzed forRac1 activation. In cells expressing recombinant wild-type�4 integrins, Rac1 activation was observed at the downstreamedge of 78�8.2% of cells at the wound margins, consistentwith earlier observations (Figures 3 and 7A). In contrast,Rac1 activation was observed at the downstream edge in only5�5% of the cells expressing recombinant �4(S988A) (Figure7B). Thus, ectopic expression of �4(S988A) in endothelialcells exerts a dominant inhibitory effect on the localization ofshear-induced Rac1 activation. These results demonstrate that�4 integrin phosphorylation is required for shear-inducedpolarized Rac1 activation and consequently for endothelialcell alignment.
DiscussionEndothelial cell alignment in the direction of blood flow hasbeen known for many years; recent work indicated thatlocalized activation of Rac1 GTPase at the downstream sideof the endothelial cell is a critical event in flow-inducedalignment.6 Here we report that localized �4 integrin phos-phorylation leads to this localized Rac1 activation and sub-sequent stress fiber alignment and endothelial cell elongationparallel to the flow direction in response to shear stress. �4integrins were phosphorylated within 5 minutes of shearstress exposure and phosphorylation occurred predominantlyat the downstream edges of the cells. Inhibition of PKA
Figure 3. Rac1 activation after shear stress requires PKA activ-ity. HMECs were transfected with Rac1(wt)-GFP and seeded onCS-1-coated slides, shear-loaded with Alexa-PBD and scratchwounded, then subjected to flow for 5 minutes �H-89. Rac1activation was assessed by FRET. Arrowheads indicate the cellperiphery. The black arrow indicates laminar flow direction forall samples. Bar, 10 �m.
Goldfinger et al �4 Integrin Phosphorylation in Endothelium 181
by guest on July 5, 2015http://circres.ahajournals.org/Downloaded from

blocked �4 phosphorylation and prevented both localizedRac1 activation and stress fiber alignment in the flowdirection. Furthermore, �4 integrins are required for endothe-lial cell alignment because deletion of �4 or addition ofantibodies against �4 inhibited stress fiber alignment. Mostimportantly, PKA phosphorylation of �4 is involved inalignment because endothelial cells bearing �4(S988A), amutation which disrupts the PKA phosphorylation site, fail toalign in the flow direction. Together these results show thatshear-induced PKA-dependent �4 phosphorylation is local-ized to the downstream edge of endothelial cells. The local-ized �4 phosphorylation leads to localized Rac1 activation atthe downstream edge, enabling endothelial cell reorientationin the flow direction. Previous studies showed how a mech-anosensory complex led to integrin activation that resulted inRac1 activation.5,6 The present studies elucidate the pathwaywhereby �4 integrin phosphorylation informs the endothelialcell about the flow direction by localizing this Rac1 activa-
tion to the downstream edge, thereby acting as a “weathervane” of shear-induced endothelial cell reorientation.
The tangential drag forces imposed by laminar shear stressinduce �4 integrin phosphorylation at Ser988 in endothelialcells. Using a phospho-specific anti-�4 antibody, we ob-served phosphorylation as early as 5 minutes after applicationof 12 dynes/cm2 shear stress, which is within the ranges ofshear stresses in medium-sized arteries.24 Previous studieshave suggested that other key signaling events occur withinsimilar time frames, including c-Src activation (1 minute,peak at 10 minutes),25 VEGFR2 phosphorylation (1 minute,peak at 5 minutes),26,27 and Ras28 and Rac1 activation (5minutes).6 Furthermore, �4 is phosphorylated at Ser988, aknown PKA phosphorylation site,19 and blocking PKA abol-ishes phosphorylation. The dependence of �4 phosphoryla-tion on the application of shear and on PKA activity suggeststhat shear stress may activate PKA; however, we cannotexclude the possibility that there is tonic PKA activity and
Figure 4. Shear-induced stress fiber alignment requires PKA activity. A, HMECs were plated on CS-1 and subjected to flow for 20hours in the continuous presence or absence of 100 �mol/L H-89, then fixed and labeled with rhodamine-phalloidin. Bar, 10 �m. B,Average angles of stress fibers (mapped onto a 0 to 90° range) from a line parallel to the flow direction and the S.D. of the angles areshown as the angular and radial positions of the corresponding bullets.
Figure 5. �4 integrin is required for shear-induced alignment. A, HMECs seeded on fibronectin were subjected for 20 hours to flowwith growth medium containing control IgG or blocking antibodies to human �2, �4, or �5 integrins, then fixed and labeled withrhodamine-phalloidin. Bars, 10 �m. Average angles of stress fibers from a line parallel to the flow direction and the S.D. of the anglesare shown as the angular and radial positions of the corresponding bullets. B, MLECs isolated from wild-type mice treated with vector(wt) or wild-type and �4 fl/fl mice treated with Cre recombinase (wt CRE and �4 fl/fl CRE, respectively) were plated on fibronectin andsubjected to flow for 20 hours, then labeled with rhodamine-phalloidin. Bar, 10 �m.
182 Circulation Research July 18, 2008
by guest on July 5, 2015http://circres.ahajournals.org/Downloaded from

that shear stress acts to suppress phosphatase activity. Nev-ertheless, we favor the former possibility because PKA isknown to be activated in endothelial cells by shear stress,which also induces phosphorylation of several PKA sub-strates such as VASP29 and endothelial nitric oxide syn-thase.30 Shear stress exerts force on endothelial cell attach-ments to the substrate, attachments mediated by integrins.Mechanical strain on integrins can result in enhancement ofintracellular cAMP concentration leading to PKA activa-tion.31 The role of integrin attachments in initiating �4phosphorylation warrants future study. In sum, we concludethat fluid shear stress results in PKA-dependent �4 integrinphosphorylation in microvascular endothelial cells.
Localized phosphorylation of �4 integrins induced byshear stress is an important cue for alignment directionality.
After 5 minutes in shear, phosphorylation was observed onlyat the downstream cell edge. Furthermore, stress fiber align-ment required both the presence of �4 integrin and itsphosphorylation by PKA. �4 integrins strongly promote cellmigration. The �4 cytoplasmic tail is sufficient for a promi-gratory response,8 and we have shown that phosphorylation atSer988 at the cell’s leading edge is the key determinant of thisfunction; Ser988 is also the only identified �4 phosphorylationsite in vivo.12,13 Shear stress accelerates endothelial woundclosure.32 When a wound is orthogonal to the flow, cells onthe wound margin proximal to the flow source (in which �4is phosphorylated) migrate into the wound space faster thancells on the distal margin (in which �4 phosphorylation doesnot occur).2,3 Those latter cells have to move against the flow.Indeed, reendothelialization occurs fastest parallel to the flow
Figure 7. Nonphosphorylatable �4 integrin inhibits shear-induced Rac1 activation.HMECs cotransfected with Rac1(wt)-GFP and �4(wt) or �4(S988A) were shear-loadedwith recombinant PAK-1-binding domain of p21 coupled to Alexa dye (Alexa-PBD),plated on CS-1-coated slides, scratch wounded, and subjected to flow for 5 min-utes. Fixed cells were assayed for Rac FRET. A, Representative images of cells atthe wound edge after flow exposure. Bar, 25 �m. B, Quantification of polarizedRac1 activation by shear. Images of cells at the wound edges were divided intoquadrants and FRET signal at the cell periphery was assessed by visual inspectionfor at least 15 cells per sample. Shown are percentages of cells in which FRET wasobserved in the quadrant facing the wound (“Toward” in A) in cells overexpressing�4(wt) or �4(S988A) as indicated, �SEM; **P�0.0002; *P�0.00001.
Figure 6. �4 integrin phosphorylation is necessary for alignment. A, MLECs isolated from wild-type and �4(S988A) mice plated onfibronectin were subjected to flow for 20 hours, then labeled with rhodamine-phalloidin. Bar, 10 �m. B, Average angles of stress fibersfrom a line parallel to the flow direction and the standard deviations of the angles are shown as the angular and radial positions of thecorresponding bullets.
Goldfinger et al �4 Integrin Phosphorylation in Endothelium 183
by guest on July 5, 2015http://circres.ahajournals.org/Downloaded from

direction following endothelial wounds in vivo, indicating aflow-induced enhancement of cell migration.33,34 Shear stresscan promote migration of endothelial cells from the upstreamedge of wounds; our studies now suggest that localized �4phosphorylation can contribute to this enhanced directionalmigration.
The restriction of �4 phosphorylation to the downstreamside of endothelial cells under shear serves to localize othersignaling responses required for proper cytoskeletal align-ment. Blocking �4 phosphorylation disrupts localized Rac1activation to the downstream edge, which is essential forendothelial cytoskeletal alignment.6 One clue to the mecha-nism for this effect on Rac1 comes from our previous studiesin migrating cells. In particular, �4 cytoplasmic tail phos-phorylation at Ser988 by PKA enables that integrin to activateRac1 because it prevents the binding of a protein complexthat blocks Rac1 activation.7,12 Other signaling events areinduced or enhanced by shear and play a role in cellmigration. For example, PI3-kinase activity is increased byshear.35 PI3-K is also typically localized to the leading edgeof migrating cells36 and can promote Rac1 activation, but it isnot required for the alignment response.37 Both cell migrationand morphological alignment under shear are driven bycytoskeletal rearrangements22,38 in response to Rho GTPases.�4 phosphorylation by shear stress coordinates localized RhoGTPase signaling to favor cytoskeletal alignment along theflow and migration in the flow direction.
�4 integrins and PKA play important roles in endothelialfunctions such as neovascularization. Endothelial cells ex-press �4 integrins in vitro and in vivo, and fibronectin, aligand for �4 integrins, is highly expressed in the vascula-ture.15,39,40 �4 integrins mediate endothelial cell adhesion,spreading, proliferation, and migration in vitro.39,41 In vivo,�4 expression is required for hematopoietic and endothelialprogenitor cell homing and for efficient angiogenesis indeveloping embryos and in tumors. �4 antagonists also blockangiogenesis in a chick chorioallantoic membrane modelconfirming this function of �4 integrins, although the require-ment for �4 in angiogenesis may result from combinedcontributions in vascular endothelial and smooth musclecells, as well as from paracrine effects in macrophages.41–43
Similarly, PKA regulates vascular endothelial cell adhesion,migration and survival,44,45 and angiogenesis.46,47 New bloodvessel formation is stimulated by shear stress and requiresflow conditions which promote cytoskeletal parallel align-ment, indicating the importance of endothelial alignment inangiogenesis.24,48 Therefore shear-induced PKA-dependent�4 phosphorylation may be an important regulatory step inendothelial functions during vascular development andremodeling.
Sources of FundingThis work was supported by American Heart Association SDG0435295 (to L.E.G.) and 0635228N (to E.T.), National ScienceFoundation NIRT 0608863 (to A.G.), and NIH grants HL088632 (toE.T.), GM 68524 (to E.G.), HL085159 (to S.C.), and AR27214,HL078784, and HL31950 (to M.H.G.). E.T. is an Ellison MedicalFoundation New Scholar in Aging.
DisclosuresNone.
References1. Noria S, Cowan DB, Gotlieb AI, Langille BL. Transient and steady-state
effects of shear stress on endothelial cell adherens junctions. CircRes. 1999;85:504–514.
2. Gojova A, Barakat AI. Vascular endothelial wound closure under shearstress: role of membrane fluidity and flow-sensitive ion channels. J ApplPhysiol. 2005;98:2355–2362.
3. Hsu PP, Li S, Li YS, Usami S, Ratcliffe A, Wang X, Chien S. Effects offlow patterns on endothelial cell migration into a zone of mechanicaldenudation. Biochem Biophys Res Commun. 2001;285:751–759.
4. Cunningham KS, Gotlieb AI. The role of shear stress in the pathogenesisof atherosclerosis. Lab Invest. 2005;85:9–23.
5. Tzima E, del Pozo MA, Shattil SJ, Chien S, Schwartz MA. Activation ofintegrins in endothelial cells by fluid shear stress mediates Rho-dependentcytoskeletal alignment. EMBO J. 2001;20:4639–4647.
6. Tzima E, del Pozo MA, Kiosses WB, Mohamed SA, Li S, Chien S,Schwartz MA. Activation of Rac1 by shear stress in endothelial cellsmediates both cytoskeletal reorganization and effects on gene expression.EMBO J. 2002;21:6791–6800.
7. Nishiya N, Kiosses WB, Han J, Ginsberg MH. An alpha4 integrin-paxillin-Arf-GAP complex restricts Rac activation to the leading edge ofmigrating cells. Nat Cell Biol. 2005;7:343–352.
8. Kassner PD, Kawaguchi S, Hemler ME. Minimum � chain cytoplasmictail sequence needed to support integrin-mediated adhesion. J Biol Chem.1994;269:19859–19867.
9. Yang JT, Rayburn H, Hynes RO. Cell adhesion events by alpha4 integrinsare essential in placental and cardiac development. Development. 1995;121:549–560.
10. Sengbusch JK, He W, Pinco KA, Yang JT. Dual functions of �4�1integrin in epicardial development: initial migration and long-termattachment. J Cell Biol. 2002;157:873–882.
11. Arroyo AG, Yang JT, Rayburn H, Hynes RO. Alpha4 integrins regulatethe proliferation/differentiation balance of multilineage hematopoieticprogenitors in vivo. Immunity. 1999;11:555–566.
12. Goldfinger LE, Han J, Kiosses WB, Howe AK, Ginsberg MH. Spatialrestriction of {alpha}4 integrin phosphorylation regulates lamellipodialstability and {alpha}4{beta}1-dependent cell migration. J Cell Biol.2003;162:731–741.
13. Han J, Rose DM, Woodside DG, Goldfinger LE, Ginsberg MH. Integrinalpha4 beta1-dependent cell migration requires both phosphorylation andde-phosphorylation of the alpha4 cytoplasmic domain. J Biol Chem.2003;287:34845–34853.
14. Priestley GV, Ulyanova T, Papayannopoulou T. Sustained alterations inbiodistribution of stem/progenitor cells in Tie2Cre� alpha4(f/f) mice arehematopoietic cell autonomous. Blood. 2007;109:109–111.
15. Sheppard AM, Onken MD, Rosen GD, Noakes PG, Dean DC. Expandingroles for alpha 4 integrin and its ligands in development. Cell AdhesCommun. 1994;2:27–43.
16. Kil SH, Krull CE, Cann G, Clegg D, Bronner-Fraser M. The alpha4subunit of integrin is important for neural crest cell migration. Dev Biol.1998;202:29–42.
17. Pinco KA, Liu S, Yang JT. alpha4 integrin is expressed in a subset ofcranial neural crest cells and in epicardial progenitor cells during earlymouse development. Mech Dev. 2001;100:99–103.
18. Jongewaard IN, Tsai PM, Smith JW. The type III connecting segment offibronectin contains an aspartic acid residue that regulates the rate ofbinding to integrin alpha 4 beta 1. Cell Adhes Commun. 1996;3:487–495.
19. Han J, Liu S, Rose DM, Schlaepfer DD, McDonald H, Ginsberg MH.Phosphorylation of the integrin �4 cytoplasmic domain regulates paxillinbinding. J Biol Chem. 2001;276:40903–40909.
20. Feral CC, Rose DM, Han J, Fox N, Silverman GJ, Kaushansky K,Ginsberg MH. Blocking the alpha 4 integrin-paxillin interaction selec-tively impairs mononuclear leukocyte recruitment to an inflammatorysite. J Clin Invest. 2006;116:715–723.
21. Reutershan J, Stockton R, Zarbock A, Sullivan GW, Chang D, Scott D,Schwartz MA, Ley K. Blocking p21-activated kinase reduceslipopolysaccharide-induced acute lung injury by preventing poly-morphonuclear leukocyte infiltration. Am J Respir Crit Care Med. 2007;175:1027–1035.
184 Circulation Research July 18, 2008
by guest on July 5, 2015http://circres.ahajournals.org/Downloaded from

22. McCue S, Noria S, Langille BL. Shear-induced reorganization of endo-thelial cell cytoskeleton and adhesion complexes. Trends CardiovascMed. 2004;14:143–151.
23. Hynes RO. Integrins: bidirectional, allosteric signaling machines. Cell.2002;110:673–687.
24. Resnick N, Yahav H, Shay-Salit A, Shushy M, Schubert S, ZilbermanLC, Wofovitz E. Fluid shear stress and the vascular endothelium: forbetter and for worse. Prog Biophys Mol Biol. 2003;81:177–199.
25. Jalali S, Li YS, Sotoudeh M, Yuan S, Li S, Chien S, Shyy JY. Shear stressactivates p60src-Ras-MAPK signaling pathways in vascular endothelialcells. Arterioscler Thromb Vasc Biol. 1998;18:227–234.
26. Chen KD, Li YS, Kim M, Li S, Yuan S, Chien S, Shyy JY. Mechano-transduction in response to shear stress. Roles of receptor tyrosinekinases, integrins, and Shc. J Biol Chem. 1999;274:18393–18400.
27. Jin ZG, Ueba H, Tanimoto T, Lungu AO, Frame MD, Berk BC. Ligand-independent activation of vascular endothelial growth factor receptor 2 byfluid shear stress regulates activation of endothelial nitric oxide synthase.Circ Res. 2003;93:354–363.
28. Li YS, Shyy JY, Li S, Lee J, Su B, Karin M, Chien S. The Ras-JNKpathway is involved in shear-induced gene expression. Mol Cell Biol.1996;16:5947–5954.
29. Wei L, Muller S, Ouyang J, Stoltz JF, Wang X. Changes of vasodilator-stimulated phosphoprotein (VASP) and its phosphorylation in endothelialcells exposed to laminar flow. Clin Hemorheol Microcirc. 2003;28:113–120.
30. Boo YC, Hwang J, Sykes M, Michell BJ, Kemp BE, Lum H, Jo H. Shearstress stimulates phosphorylation of eNOS at Ser(635) by a protein kinaseA-dependent mechanism. Am J Physiol Heart Circ Physiol. 2002;283:H1819–H1828.
31. Meyer CJ, Alenghat FJ, Rim P, Fong JHJ, Fabry B, Ingber DE.Mechanical control of cyclic AMP signalling and gene transcriptionthrough integrins. Nature Cell Biol. 2000;2:666–668.
32. Albuquerque ML, Waters CM, Savla U, Schnaper HW, Flozak AS. Shearstress enhances human endothelial cell wound closure in vitro. Am JPhysiol Heart Circ Physiol. 2000;279:H293–H302.
33. Schwartz SM, Gajdusek CM, Reidy MA, Selden SC III, HaudenschildCC. Maintenance of integrity in aortic endothelium. Fed Proc. 1980;39:2618–2625.
34. Haudenschild CC, Schwartz SM. Endothelial regeneration. II. Restitutionof endothelial continuity. Lab Invest. 1979;41:407–418.
35. Decave E, Rieu D, Dalous J, Fache S, Brechet Y, Fourcade B, Satre M,Bruckert F. Shear flow-induced motility of Dictyostelium discoideumcells on solid substrate. J Cell Sci. 2003;116:4331–4343.
36. Funamoto S, Meili R, Lee S, Parry L, Firtel RA. Spatial and temporalregulation of 3-phosphoinositides by PI 3-kinase and PTEN mediateschemotaxis. Cell. 2002;109:611–623.
37. Wojciak-Stothard B, Ridley AJ. Shear stress-induced endothelial cellpolarization is mediated by Rho and Rac but not Cdc42 or PI 3-kinases.J Cell Biol. 2003;161:429–439.
38. Ridley AJ, Schwartz MA, Burridge K, Firtel RA, Ginsberg MH, BorisyG, Parsons JT, Horwitz AR. Cell migration: integrating signals from frontto back. Science. 2003;302:1704–1709.
39. Massia SP, Hubbell JA. Vascular endothelial cell adhesion and spreadingpromoted by the peptide REDV of the IIICS region of plasma fibronectinis mediated by integrin alpha 4 beta 1. J Biol Chem. 1992;267:14019–14026.
40. Brezinschek RI, Brezinschek HP, Lazarovits AI, Lipsky PE,Oppenheimer-Marks N. Expression of the beta 7 integrin by humanendothelial cells. Am J Pathol. 1996;149:1651–1660.
41. Calzada MJ, Zhou L, Sipes JM, Zhang J, Krutzsch HC, Iruela-Arispe ML,Annis DS, Mosher DF, Roberts DD. Alpha4beta1 integrin mediatesselective endothelial cell responses to thrombospondins 1 and 2 in vitroand modulates angiogenesis in vivo. Circ Res. 2004;94:462–470.
42. Grazioli A, Alves CS, Konstantopoulos K, Yang JT. Defective bloodvessel development and pericyte/pvSMC distribution in alpha 4 integrin-deficient mouse embryos. Dev Biol. 2006;293:165–177.
43. Jin H, Su J, Garmy-Susini B, Kleeman J, Varner J. Integrin alpha4beta1promotes monocyte trafficking and angiogenesis in tumors. Cancer Res.2006;66:2146–2152.
44. Netherton SJ, Sutton JA, Wilson LS, Carter RL, Maurice DH. Bothprotein kinase A and exchange protein activated by cAMP coordinateadhesion of human vascular endothelial cells. Circ Res. 2007;101:768–776.
45. Bodnar RJ, Yates CC, Wells A. IP-10 blocks vascular endothelial growthfactor-induced endothelial cell motility and tube formation via inhibitionof calpain. Circ Res. 2006;98:617–625.
46. Kim S, Bakre M, Yin H, Varner JA. Inhibition of endothelial cell survivaland angiogenesis by protein kinase A. J Clin Invest. 2002;110:933–941.
47. Bakre MM, Zhu Y, Yin H, Burton DW, Terkeltaub R, Deftos LJ, VarnerJA. Parathyroid hormone-related peptide is a naturally occurring, proteinkinase A-dependent angiogenesis inhibitor. Nat Med. 2002;8:995–1003.
48. Vyalov S, Langille BL, Gotlieb AI. Decreased blood flow rate disruptsendothelial repair in vivo. Am J Pathol. 1996;149:2107–2118.
Goldfinger et al �4 Integrin Phosphorylation in Endothelium 185
by guest on July 5, 2015http://circres.ahajournals.org/Downloaded from

Supplementary information
Supplementary Materials & Methods
Generation of α4 integrin S988A knock-in mice: A genomic fragment
encompassing exons 25-28 of mouse α4 integrin was cloned into Bluescript
plasmid. A single Ser-to-Ala point mutation was created at position 988, and a
neo cassette flanked by loxP sequences was introduced between exon 27 and
exon 28 for drug selection. The targeting construct was electroporated into
embryonic stem cells and drug-resistant clones were screened by Southern
blotting for recombination between the targeting construct and the α4 genomic
locus. Positive clones were injected into blastocysts, which were then implanted
into pseudo-pregnant female mice. Chimeric mice (agouti) were crossed with
mice of the same genetic background as the blastocysts. Progeny from these
crosses were screened by genomic PCR for heterozygous S988A mutation.
Heterozygous S988A progeny were crossed to obtain homozygous littermates.
Generation of α4 fl/fl (null) pulmonary endothelial cells: α4 fl/fl cells were treated
with soluble Cre recombinase to remove exon 28 and the polyadenylation tail
from the α4 gene locus. Elimination of α4 protein expression was confirmed by
FACScan using R1/2 antibodies to mouse α4 integrins (Abcam, Cambridge, MA).
Shear stress assays: Cells were plated on glass slides and subjected to laminar
shear in a parallel plate flow chamber as described previously1, or they were
plated on coverslips which were fitted into microfluidic devices, as described
elsewhere (Tkachenko et al., manuscript in preparation). Flow rates were

adjusted to maintain laminar shear stress of 12 dynes/cm2. Actin staining and
analysis are as described2.
Immunocytochemistry: Sterile glass coverslips or slides were coated with ECM
proteins (CS-1, fibronectin or collagen) at 5 μg/ml overnight at 4°C, then blocked
for one hour with 1 mg/ml BSA. Cells were suspended with trypsin/EDTA and
plated onto coated cover slips and treated as described previously3.
Fluorescence Resonance Energy Transfer: FRET assays and calculations to
account for bleedthrough and background were performed as described
previously2. The corrected 8-bit FRET images typically had a fluorescence
intensity range of 0–100 and were displayed using pseudocolor, where blue was
closest to 0 and red closest to 100.

Supplementary Figures and Tables
Online Table 1. Average stress fiber angles relative to the direction of flow.
Cells were subjected to shear stress under the conditions indicated. Stress fibers
were labeled with rhodamine-phalloidin and angles from lines parallel to the flow
direction were measured.
Supplementary Figure 1: Jurkat cells and human microvascular endothelial
(HME) cells were surface-labeled with mouse IgG (dotted lines) or HP2/1 mouse
anti-human α4 integrin and subjected to FACS analysis.
Supplementary Figure 2: HMECs were pre-incubated for 15 min with blocking
antibodies against the indicated integrins and seeded onto surfaces coated with
the cell-binding fragment of fibronectin (911), the α4-binding fragment of
fibronectin (CS-1), collagen, or poly-L-Lysine for 60 min. Cells were fixed and
labeled with crystal violet to indicate relative numbers of adherent cells.
Supplementary Figure 3: Cells in Figure 5B were suspended with trypsin and
labeled with R1/2 antibodies to mouse α4 integrin and analyzed by FACS.
Supplementary Figure 4: Primary endothelial cell cultures derived from mice
expressing wild type α4 or α4 harboring a Ser Ala mutation at position 988
were seeded on CS-1, scratch wounded and subjected to shear stress for 5 min.
Sheared cells were fixed and stained with PSα4. White arrows indicate enhanced
plasma membrane α4 phosphorylation.

References
(1) McCue S, Noria S, Langille BL. Shear-induced reorganization of endothelial cell cytoskeleton and adhesion complexes. Trends Cardiovasc Med. 2004;14:143-51.
(2) Tzima E, del Pozo MA, Kiosses WB, Mohamed SA, Li S, Chien S,
Schwartz MA. Activation of Rac1 by shear stress in endothelial cells mediates both cytoskeletal reorganization and effects on gene expression. EMBO J. 2002;21:6791-800.
(3) Goldfinger LE, Han J, Kiosses WB, Howe AK, Ginsberg MH. Spatial restriction of {alpha}4 integrin phosphorylation regulates lamellipodial stability and {alpha}4{beta}1-dependent cell migration. J Cell Biol. 2003;162:731-41.

Online Table 1
Average stress fiber angles from parallel to thedirection of flow.
28.0341.6 ± 3.96No
22.5946.8 ± 2.62YesKT-5720, 1 μM
11.5841.37 ± 1.10No
10.6137.63 ± 1.01YesH-89, 30 μM
17.1541.48 ± 1.64No
9.2314.31 ± 0.69YesVector control
St. Dev. (°)Average angle from orthogonal (°)Flow (12 dynes/cm2)Treatment
Supplementary Figure 1

0
0.5
1
1.5
2
2.5
α5 α4 α2 α5 α4 α2 α5 α4 α2 α5 α4 α2
9-11 CS-1 Collagen poly-L-LysBlocking Ab:
OD
590
Supplementary Figure 2

wt wt CRE α4 fl/fl CRE
Supplementary Figure 3

Supplementary Figure 4
wt S988A
Flow

GinsbergEugene Tkachenko, Edgar Gutierrez, Alex Groisman, Phu Nguyen, Shu Chien and Mark H.
Lawrence E. Goldfinger, Eleni Tzima, Rebecca Stockton, William B. Kiosses, Kayoko Kinbara,Alignment
Induced Endothelial Cell−4 Integrin Phosphorylation Directs Shear StressαLocalized
Print ISSN: 0009-7330. Online ISSN: 1524-4571 Copyright © 2008 American Heart Association, Inc. All rights reserved.is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation Research
doi: 10.1161/CIRCRESAHA.108.1763542008;103:177-185; originally published online June 26, 2008;Circ Res.
http://circres.ahajournals.org/content/103/2/177World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circres.ahajournals.org/content/suppl/2008/06/26/CIRCRESAHA.108.176354.DC1.htmlData Supplement (unedited) at:
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer about this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further informationEditorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center, not theCirculation Researchin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on July 5, 2015http://circres.ahajournals.org/Downloaded from
Related Documents











