ELSEVIER Molecular Brain Research 29 (1995) 71-80 MOLECULAR BRAIN RESEARCH Localization of the plasma in rat cerebellum, Research report membrane Ca2’-ATPase isoform PMCA3 choroid plexus and hippocampus Thomas J. Eakin a, Marta C. Antonelli e, Emilio L. Malchiodi f, Denis G. Baskin a,b,c, William L. Stahla>b,d,* a Veterans Affairs Medical Center, Seattle, WA 98108, USA ’Department of Medicine, Uniuersity of Washington School of Medicine, Seattle, WA 98195, USA ’Department of Biological Structure, University of Washington School of Medicine, Seattle, WA 98195, USA d Department of Physiology and Biophysics, University of Washington School of Medicine, Seattle, WA 98195, USA e Institute de Quimica y Fisicoquimica Bioldgicas CUBA-CONICET), Uniuersidad de Buenos Aires, Buenos Aires, Argentina f Ca’tedra de Immunologia, Fact&ad de Farmacia y Bioquimica, Uniuersidad de Buenos Aires, Buenos Aires, Argentina Accepted 4 October 1994 Abstract mRNA encoding rat plasma membrane Ca 2+-ATPase isoform PMCA3 was localized in the granule cell layer of the cerebellum and in choroid plexus by in situ hybridization with an 35S-labelled oligodeoxynucleotide probe. In order to examine whether this isoform is expressed as a protein in brain, polyclonal antibodies were raised against a peptide corresponding to a C-terminal 18 amino acid sequence of PMCA3 which had been conjugated to bovine serum albumin, Using immunoblot analysis with affinity-purified antibodies, PMCA3 protein was found in rat brain microsomes and cultured neurons. The translated protein had an observed molecular mass of approximately 135 kDa, as predicted from molecular cloning studies. The pattern of localization of PMCA3 in brain using anti-peptide antibodies was consistent with findings from in situ hybridization. PMCA3-like immunoreactive sites were found in the granule cell and molecular layers of rat cerebellum and in choroid plexus, and the pattern of staining suggests that immunoreactive sites are associated with granule cell processes. This conclusion was supported by the finding that growth-associated protein-43, a protein known to be present in axons and nerve terminals, had a pattern of distribution similar to PMCA3 in the molecular layer of cerebellum. Very low levels of PMCA3-like immunoreactivity were associated with Purkinje cell soma or processes, consistent with the low levels of PMCA3 mRNA found in these neurons. PMCA3-like immunoreactivity was lower in hippocampus than in cerebellum; hippocampal CA1 region immunoreactivity was primarily associated with dendritic fields rather than with pyramidal cell bodies. The results demonstrate that a PMCA3-like protein is expressed in neurons of rat brain and is localized primarily in cell processes. Keywords: Anti-peptide antibody; Calcium homeostasis; Calmodulin; GAP-43; Granule cell; Immunocytochemistry; In situ hybridization; mRNA, Oligonucleotide probe; Purkinje cell 1. Introduction Regulation of cytosolic Ca2+ is important in main- taining normal cellular function in the nervous system [28], and involves interaction of Ca2+-ATPase of plasma membrane (PMCA) [2,8,9,21,31,41,44,451, Ca2+- ATPase associated with sarcoplasmic (endoplasmic) reticulum [28], plasma membrane Na+-Ca2+ ex- * Corresponding author. VA Medical Center (1511, 1660 S. Columbian Way, Seattle, WA 98108, USA. Fax: (1) (206) 764-2164; E-mail:[email protected]. 0169-328X/95/$09.50 0 1995 Elsevier Science B.V. All rights reserved SSDI 0169-328X(94)00231-2 changer [28,41], and probably several calcium binding proteins and mitochondrial Ca2+ transporters [28]. The presence of these regulatory molecules may not be a fixed characteristic of cells in a general sense. Rather, each class of cells, e.g. different types of neurons, may utilize a unique combination of regulatory molecules in order to maintain homeostatic control of different lev- els of cytosolic Ca2+. In addition, different combina- tions of transport molecules and their isoforms may exist in individual parts of cells, such as in the cell body, dendrites or nerve terminals, imparting unique transport signatures to specific cell types. Control of homeostasis at the plasma membrane of

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ELSEVIER Molecular Brain Research 29 (1995) 71-80
MOLECULAR BRAIN
RESEARCH
Localization of the plasma in rat cerebellum,
Research report
membrane Ca2’-ATPase isoform PMCA3 choroid plexus and hippocampus
Thomas J. Eakin a, Marta C. Antonelli e, Emilio L. Malchiodi f, Denis G. Baskin a,b,c, William L. Stahla>b,d, *
a Veterans Affairs Medical Center, Seattle, WA 98108, USA ’ Department of Medicine, Uniuersity of Washington School of Medicine, Seattle, WA 98195, USA
’ Department of Biological Structure, University of Washington School of Medicine, Seattle, WA 98195, USA d Department of Physiology and Biophysics, University of Washington School of Medicine, Seattle, WA 98195, USA
e Institute de Quimica y Fisicoquimica Bioldgicas CUBA-CONICET), Uniuersidad de Buenos Aires, Buenos Aires, Argentina f Ca’tedra de Immunologia, Fact&ad de Farmacia y Bioquimica, Uniuersidad de Buenos Aires, Buenos Aires, Argentina
Accepted 4 October 1994
Abstract
mRNA encoding rat plasma membrane Ca 2+-ATPase isoform PMCA3 was localized in the granule cell layer of the cerebellum and in choroid plexus by in situ hybridization with an 35S-labelled oligodeoxynucleotide probe. In order to examine whether this isoform is expressed as a protein in brain, polyclonal antibodies were raised against a peptide corresponding to a C-terminal 18 amino acid sequence of PMCA3 which had been conjugated to bovine serum albumin, Using immunoblot analysis with affinity-purified antibodies, PMCA3 protein was found in rat brain microsomes and cultured neurons. The translated protein had an observed molecular mass of approximately 135 kDa, as predicted from molecular cloning studies. The pattern of localization of PMCA3 in brain using anti-peptide antibodies was consistent with findings from in situ hybridization. PMCA3-like immunoreactive sites were found in the granule cell and molecular layers of rat cerebellum and in choroid plexus, and the pattern of staining suggests that immunoreactive sites are associated with granule cell processes. This conclusion was supported by the finding that growth-associated protein-43, a protein known to be present in axons and nerve terminals, had a pattern of distribution similar to PMCA3 in the molecular layer of cerebellum. Very low levels of PMCA3-like immunoreactivity were associated with Purkinje cell soma or processes, consistent with the low levels of PMCA3 mRNA found in these neurons. PMCA3-like immunoreactivity was lower in hippocampus than in cerebellum; hippocampal CA1 region immunoreactivity was primarily associated with dendritic fields rather than with pyramidal cell bodies. The results demonstrate that a PMCA3-like protein is expressed in neurons of rat brain and is localized primarily in cell processes.
Keywords: Anti-peptide antibody; Calcium homeostasis; Calmodulin; GAP-43; Granule cell; Immunocytochemistry; In situ hybridization; mRNA, Oligonucleotide probe; Purkinje cell
1. Introduction
Regulation of cytosolic Ca2+ is important in main- taining normal cellular function in the nervous system [28], and involves interaction of Ca2+-ATPase of plasma membrane (PMCA) [2,8,9,21,31,41,44,451, Ca2+- ATPase associated with sarcoplasmic (endoplasmic) reticulum [28], plasma membrane Na+-Ca2+ ex-
* Corresponding author. VA Medical Center (1511, 1660 S. Columbian Way, Seattle, WA 98108, USA. Fax: (1) (206) 764-2164; E-mail:[email protected].
0169-328X/95/$09.50 0 1995 Elsevier Science B.V. All rights reserved SSDI 0169-328X(94)00231-2
changer [28,41], and probably several calcium binding proteins and mitochondrial Ca2+ transporters [28]. The presence of these regulatory molecules may not be a fixed characteristic of cells in a general sense. Rather, each class of cells, e.g. different types of neurons, may utilize a unique combination of regulatory molecules in order to maintain homeostatic control of different lev- els of cytosolic Ca2+. In addition, different combina- tions of transport molecules and their isoforms may exist in individual parts of cells, such as in the cell body, dendrites or nerve terminals, imparting unique transport signatures to specific cell types.
Control of homeostasis at the plasma membrane of

72 T.J. Eakin et al. /Molecular Bruin Research 29 (I 995) 71-80
neurons may primarily involve PMCA and the Naf- Ca2+ exchanger. Relative contributions to calcium homeostasis by these transporters has been studied in acutely dissociated cells and primary cultures of neu- rons [2,3,19,26,43]. Benham et al. [2] showed that PMCA is a more likely candidate as the major regula- tor of calcium homeostasis in rat sensory neurons un- der normal physiological conditions, and that the role of the Na+-Ca2+ exchanger is less certain. However, the relative contributions of PMCA and Na+-Ca2+ exchange to calcium homeostasis has been difficult to assess in the absence of specific inhibitors for these transporters.
Understanding the role of different transporters is also complicated by the existence of multiple isoforms of the transport molecules and their alternatively spliced variants. In the case of PMCA in rat brain, mRNAs encoding four PMCA isoforms, ranging in size from approximately 4 to 7 kb have been identified [6,7,20,34] and recent reports using polymerase chain reaction amplification indicate that at least 10 alterna- tively spliced mRNA variants exist [6,23,40]. Based on Northern blot and polymerase chain reaction analyses, PMCAl and PMCA4 mRNAs appear to be present in all tissues [20,23]. PMCA2 mRNA is found mainly in brain and heart [20], and PMCA3 mRNA is expressed predominantly in brain and skeletal muscle [20]. In the case of the PMCA3 isoform, six alternatively spliced mRNA variants, designated a-f, have been described [6,23], but only PMCA3a and PMCA3b were found at significant levels in rat brain. In situ hybridization (ISH) studies [36,38,39] in rat central nervous system (CNS) indicate that PMCA isoforms are associated primarily with neurons. PMCAl mRNA is broadly distributed, but the highest levels are in CA1 pyramidal neurons of hippocampus. PMCA2 mRNA is most abundant in Purkinje cells of the cerebellum, and PMCA3 mRNA levels are high in choroid plexus, cerebellum and hippocampus. PMCA4 mRNA is ex- pressed at high levels in piriform cortex, amygdaloid nucleus and lamina 2 and 6 of cerebral cortex [39].
The presence of PMCA-like protein has recently been demonstrated in brain tissue [141. On western blots of CNS membrane proteins, monoclonal antibod- ies against purified erythrocyte PMCA bound to bands with molecular masses of approximately 135,000 to 140,000, which is in the range expected for PMCA isoforms [9]. Immunocytochemical studies showed that a PMCA-immunoreactive protein was present in rat cerebellum [14] and choroid plexus [4] and established that an epitope common to several PMCA isoforms is present in mammalian brain [4,14]. However, the pres- ence of translated protein of specific isoforms has not been determined. In order to address this problem we have produced a polyclonal antibody which is se- quence-specific for PMCA3. This isoform which is ex-
pressed at high levels in brain and muscle has been localized in cerebellum, hippocampus and choroid plexus using immunocytochemistry.
2. Materials and methods
2.1. Materials
[12’I]Calmodulin was obtained from New England Nuclear (Boston, MA). Oligonucleotide probes to spe- cific PMCA mRNA isoforms were described and char- acterized in a previous study [38]. Probe PMCA3-D2, antisense to nucleotides 1823 to 1855 (a translated portion of PMCA3 mRNA), was labeled with [a- thio3”S]-dATP (New England Nuclear). Immunocyto- chemistry kits were from Vector Labs (Burlingame, CA). Immunological chemicals were from Pierce (Rockford, IL) unless otherwise specified. Protein A- Sepharose CL-4B and EAH Sepharose 4B were from Pharmacia (Piscataway, NJ). HN33p hippocampal neu- ron cultures were obtained from Dr. Bruce Wainer [25]. NG108-15 neuron-glial hybrid cells were a gift of Dr. Ken Mackie, University of Washington. Cell cul- ture reagents were from Gibco/BRL (Gaithersburg, MD) unless otherwise specified. Enhanced Chemilumi- nescence reagents were from Amersham (Arlington Heights, IL). Anti-Na,K-ATPase a3 fusion protein an- tibody was from Upstate Biotechnology, Inc. (Lake Placid, NY). GAP-43 antibody was from Boehringer Mannheim (Indianapolis, IN).
2.2. In situ hybridization
ISH procedures were previously described [15,16, 35,37,38]. In brief, slides with one rat brain cryosection (12 pm thick) were warmed to room temperature. Sections were fixed (4% paraformaldehyde), dehy- drated (70 to 100% ethanol), delipidated (chloroform), and air-dried. The ‘5S-labeled probe (PMCA3-D2) was added to hybridization buffer (43% deionized for- mamide, 10% dextran sulfate, 300 mM NaCl, 10 mM Tris, 1 mM EDTA, 1 x Denhardt’s solution (0.02% each of bovine serum albumin, Ficoll, and polyvinylpy- rollidone), 0.5 mg yeast tRNA/ml, and 10 mM dithio- threitol) and then applied to the slide. Sections were covered with Parafilm (American National Can, Greenwich, CT) and placed in moist chambers at 37°C. Following an overnight incubation, the Parafilm was removed and sections were washed twice in 1 x stand- ard saline citrate (1 X SSC = 150 mM NaCl, 15 mM sodium citrate) for 30 min at 55°C and once for 1 h at room temperature. Slides were dipped in NTB 2 emul- sion (Kodak, Rochester, NY) diluted 1:l with 600 mM ammonium acetate and exposed for 28 days. The emul- sion was developed in Kodak D-19 developer, diluted 1:l with distilled water, for 4 min at 16°C. Sections

T.J. Eakin et al. /Molecular Brain Research 29 (1995) 71-80 73
were counterstained with Cresyl violet acetate and coverslips were applied. Grain counts were done with an MCID imaging system (Imaging Research, St. Catherines, ONT) as previously described [38]. In brief cells were located under bright-field optics and grain counts were collected and analyzed from the digitized image under dark-field optics using version Ml soft- ware. Generally, 2-4 tissue sections (30 cells of a given type per tissue section) from a single exposure time were analyzed.
2.3. Preparation of antigens
A peptide for residues 1142-1159 of PMCA3 (Ser- Ala-Ala-Lys-Pro-Thr-Ser-Ala-Ala-Gly-Asn-Pro-Ser- Gly-Glu-Ser-Ile-Pro-(Cys)), which is specific for a PMCA3 epitope and not present in other PMCA iso- forms [23], was synthesized commercially (the C-termi- nal Cys was added for conjugation to carrier) and coupled to bovine serum albumin (BSA) using the Pierce Imject Activated Immunogen Conjugation Kit. Approximately 2 mg of peptide was dissolved in 200 to 500 ~1 conjugation buffer (83 mM sodium phosphate buffer, 0.1 M EDTA, 0.9 M NaCl, 0.02% NaN,, pH 7.2). Separately, 2 mg maleimide activated carrier pro- tein, BSA or keyhole limpet hemocyanin (KLH), was dissolved in 200 ~1 distilled H,O, allowed to react with the peptide for 2 hr at room temperature and purified by gel filtration. Fractions of peptide-carrier conju- gates were pooled and stored at - 80°C. The antiserum used in the present study was named anti-
PMCA3,,,,-,,,,.
2.4. Antisera
Female New Zealand white rabbits (2.5 kg) were injected with peptide-BSA conjugate, initially in com- plete Freund’s adjuvant (0.5 mg peptide/ml) at multi- ple sites (2 ml total injection) and were boosted on days 7, 14 and 28 with conjugate in incomplete Freund’s adjuvant. Testing on day 35 by ELISA showed opti- mum response.
2.5. Enzyme-linked immunosorbent assay of antisera
A standardized procedure using PMCA3 peptide bound to titer plate wells was followed. Each well of a 96-well plate was treated with 50 ~1 peptide (10 pg/ml) diluted in 0.05 M phosphate-buffered saline (PBS); in some cases, peptide-KLH conjugates were used. Plates were incubated overnight at 4°C and were blocked with 50 ~1 1% non-fat dry milk (Carnation, Los Angeles, CA) in PBS for 2 h at 37°C. Wells were washed 3 times with 150 ~1 0.05% Tween-20 in PBS (Tween-PBS). Then 50 ~1 antibody diluted (1:lOO to 1:200,000) in Tween-PBS was added and incubated for 2 hr at room temperature. Wells were washed 3 times with 150 ~1
Tween-PBS and 50 ~1 goat anti-rabbit IgG conjugated to alkaline phosphatase (Sigma, St. Louis, MO) diluted 1:lOOO with 1% BSA in PBS was added and incubated for 2 h at room temperature. Wells were washed 6 times with 150 ~1 Tween-PBS per well. For the alka- line phosphatase reaction a solution of 50 ~1 p- nitrophenylphosphate (1 mg/ml) in 10% diethanol- amine, 0.5 mM MgCl,, 0.02% NaN,, pH 9.8, was added and incubated for 30 min. Absorbency was de- termined at 410 nm and optical density readings were plotted versus dilutions of primary antibody to deter- mine the titer for the antiserum, i.e. the dilution at which 50% maximum optical density was observed.
2.6. IgG and antibody purification
IgG was isolated from antisera with significant ELISA titers using Protein A-Sepharose CL-4B. Typi- cally 3 ml of serum was continuously circulated for several hours at 4°C through a column made from 1.5 g freeze-dried Protein A-Sepharose CL-4B (swollen in buffer). The IgG fraction was eluted with 0.1 M glycine, pH 3.0 and neutralized immediately to pH 7.0 with 1 M Tris base. The IgG fraction was dialyzed versus 10 mM Tris-HCl, pH 7.4 overnight at 4°C.
EAH Sepharose 4B (pre-swollen) was washed with 0.5 M NaCl, then distilled water, and finally resus- pended in 5 ml distilled water and adjusted to pH 4.5 with HCl. Peptide (18 mg) was dissolved in 1 ml water and adjusted to pH 4.5. Separately 70 mg N-ethyl-N’- (3-dimethylaminopropyl) carbodiimide (Sigma) was dis- solved in water and adjusted to pH 4.5. Peptide, car- bodiimide and EAH-Sepharose 4B were combined and mixed on an orbital shaker for 24 h at room tempera- ture. The gel was washed with 200 ml 0.5 M NaCl and 200 ml distilled water on a filter funnel and poured to form an approximately 10 cm column bed using a 0.7 cm X 15 cm BioRad EconoColumn (BioRad Laborato- ries, Hercules, CA) which was washed with 50 mM Tris, 150 mM NaCl, 0.05% NaN,, pH 7.8. The IgG fraction was continuously circulated through the pep- tide-Sepharose column for several hours at 4°C and was washed with Tris-NaCl-azide buffer to remove unbound IgG. Affinity-purified antibodies were ob- tained by eluting with 0.1 M glycine as above for the IgG purification. The recovered antibodies were dia- lyzed overnight versus 10 mM Tris-HCl, pH 7.4 at 4°C and were concentrated using a Speed-Vat (Savant In- struments, Farmingdale, NY) and resuspended in the same starting serum volume with PBS. There was no remaining activity against BSA in the purified antibod- ies, as assessed by ELISA.
2.7. Cell culture and lysis of cells
HN33p hippocampal neurons [25] were grown in Dulbecco’s Modified Eagle’s Medium (DMEM) con-

74 T.J. Eakin et al. /Molecular Brain Research 29 (1995) 71-80
taining 10% fetal calf serum (Hyclone Laboratories, Logan, UT), 1% penicillin/streptomycin (Pen/Strep) and 1% r_-glutamine. NG108-15 cells were grown in DMEM, 10% fetal calf serum, HAT medium (Sigma), 1% L-glutamine, 1% Pen/Strep and Na pyruvate (5.5 mg/500 ml) in 5 to 10% CO,. Cells were detached by washing several times with Hank’s Balanced Salt Solu- tion and were collected by centrifugation. They were resuspended in a lysis buffer made by mixing 450 ~1 of 1% Nonidet P-40, 0.4% sodium dodecyl sulfate (SDS), 66 mM EDTA, 10 mM Tris-HCl, pH 7.4 and 50 ~1 protease inhibitor cocktail (before dilution: 25 mM EDTA, 100 mM N-ethylmaleimide, 2 mM phenyl- methyl sulfonyl fluoride and 20 pg pepstatin/5 ml). These were centrifuged in a microfuge at 4°C for 10 min at 15,000 rpm and supernatant was used for elec- trophoresis. Protein concentrations were determined using the Pierce BCA microtiter plate assay.
2.8. Electrophoresis and Western blots
Samples of rat brain microsomal membranes [331 or lysed cell supernatant were solubilized in reducing buffer (final concentrations: 0.125 M Tris, pH 6.8, 4% sodium dodecyl sulfate, 20% glycerol, 10% P-mercap- toethanol and 0.04% Bromophenol blue) for 1 h at 37°C. Electrophoresis was carried out on discontinuous gels (4% stacking gel, 7.5% separating gel) with 25 pg protein per lane at 150 V (constant) for approximately 1 h. Proteins were transferred to nitrocellulose in 25 mM Tris, 192 mM glycine, 20% (v/v> methanol, pH 8.3 at 100 V (constant) for 1 h. For immunodetection of proteins, Western blot lanes were incubated with Blotto (10 mM Tris, 150 mM NaCl, 5% non-fat dry milk (Carnation), 0.05% Tween-20, pH 7.5) at 4°C overnight. Nitrocellulose strips were then incubated with diluted primary antisera in 10 mM Tris, 150 mM NaCl, 0.1% Tween-20, 1% BSA for 1 h at room temperature and washed once for 5 min in 10 mM Tris, 150 mM NaCl, pH 7.5 (TBN), 3 times for 5 min per wash in TBN containing 0.1% Tween-20 (TBNT) and once for 5 min in TBN. Horseradish peroxidase (HRP)-conjugated protein G (BioRad, diluted 1:4000 in Blotto) was added and incubated for 1 h. Nitrocellulose strips were washed 4 times for 5 min per wash in TBNT and once for 5 min in TBN. Immunoreactive sites were detected by incubating with Amersham Enhanced Chemilumines- cence detection reagent (ECL) for approximately 2 min. Strips were apposed to Kodak XOMat X-ray film for 1 to 10 min exposure.
2.9. /‘251]calmodulin detection of PMCA on blots
Nitrocellulose blot lanes were incubated in 5% BSA, 10 mM Tris, 150 mM KCl, 1 mM CaCl,, 0.2% Tween- 20, 0.02% NaN,, pH 7.5, overnight at room tempera- ture. The blots were incubated in the above buffer
containing 1 X lo6 dpm [‘251]calmodulin per 2 ml overnight at room temperature. Blots were washed 4 times at 30 min per wash in the same buffer without BSA at room temperature and exposed to Kodak XO- Mat film for 4 to 12 h.
2.10. Immunocytochemistry
Rats were anesthetized with an intraperitoneal in- jection of 0.8 ml pentobarbital (65 mg/ml) and per- fused through the heart with a mixture of 4% para- formaldehyde in 0.1 M PBS, pH 7.4 for 15 to 20 min at room temperature. After initial fixation brain and liver tissue were removed and divided into 3 to 4 parts and placed in cold fixative for 2 hr on ice. Tissue was washed several times with fresh PBS and then overnight at 4°C. Subsequently tissue parts were either frozen in Freon or dehydrated in ethanol and embedded in paraffin. For immunocytochemistry, 20 pm cryosec- tions or 6 pm paraffin sections were used.
Generally, the recommendations of Hofmann et al. [221 for light microscope immunocytochemistry were followed. Cryostat tissue sections or paraffin tissue sections were rinsed 3 times at room temperature in wash buffer (WB) consisting of 0.3 M Tris, 0.15 M NaCI, 1% BSA, 0.05% Triton X-100, pH 7.5 and then treated for 30 min with blocking buffer (BB) consisting of WB, but with 0.5% Triton X-100 in place of 0.05% detergent. After rinsing 3 times with WB, tissue sec- tions were incubated with primary antiserum (typically 1:lOO to 1:5,000 dilutions in WB) or with pre-absorbed antiserum overnight at 4°C. Pre-absorbed antiserum was prepared by the addition of 1 mg PMCA3,,42_,,5v peptide to 1 ml of a 1:500 dilution of primary anti- serum and mixing at 4°C overnight. Sections were rinsed 3 times with WB, and incubated with biotiny- lated anti-rabbit antiserum (1:200) for 60 min at room temperature. After washing 3 times with WB at room temperature, slides were incubated for 60 min with HRP-avidin complex (1:50 dilution in WB). These were rinsed at least 3 times in WB at room tempera- ture and treated with diaminobenzidine (25 pg/lOO ml 0.05 M Tris-buffered saline, pH 7.5 containing 83 ~1 fresh 30% hydrogen peroxide), for 2 to 3 min at room temperature. They were washed once with Tris buffer (0.05 M, pH 7.5) and then once with distilled water at room temperature. Sections were sometimes lightly counter-stained with Gill’s hematoxylin. Slides were mounted with Permount (Fisher, Pittsburgh, PA).
3. Results
3.1. Localization of PMCAS mRNA by in situ hybridiza- tion
Hybridization of sSS-labelled oligodeoxynucleotide antisense probe for PMCA3 mRNA was prominent in
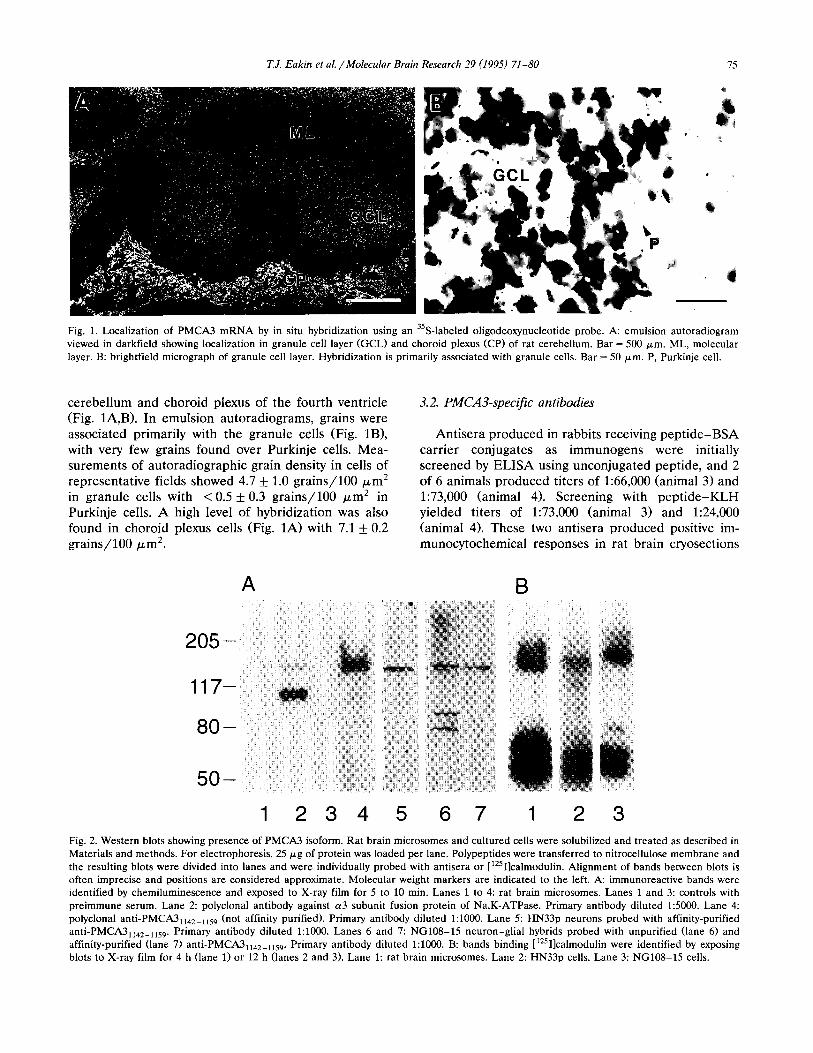
T.J. Eakin et al. /Molecular Brain Research 29 (1995) 71-80 75
Fig. 1. Localization of PMCA3 mRNA by in situ hybridization using an 3SS-labeled oligodeoxynucleotide probe. A: emulsion autoradiogram viewed in darkfield showing localization in granule cell layer (GCLI and choroid plexus (CP) of rat cerebellum. Bar = 500 Frn. ML, molecular layer. B: brightfield micrograph of granule cell layer. Hybridization is primarily associated with granule cells. Bar = 50 pm. P, Purkinje cell.
cerebellum and choroid plexus of the fourth ventricle (Fig. lA,B). In emulsion autoradiograms, grains were associated primarily with the granule cells (Fig. lB), with very few grains found over Purkinje cells. Mea- surements of autoradiographic grain density in cells of representative fields showed 4.7 + 1.0 grains/100 pm* in granule cells with < 0.5 k 0.3 grains/100 pm* in Purkinje cells. A high level of hybridization was also found in choroid plexus cells (Fig. 1A) with 7.1 * 0.2 grains/100 pm*.
3.2. PMCAJ-specific antibodies
Antisera produced in rabbits receiving peptide-BSA carrier conjugates as immunogens were initially screened by ELISA using unconjugated peptide, and 2 of 6 animals produced titers of 1:66,000 (animal 3) and 1:73,000 (animal 4). Screening with peptide-KLH yielded titers of 1:73,000 (animal 3) -and 1:24,000 (animal 4). These two antisera produced positive im- munocytochemical responses in rat brain cryosections
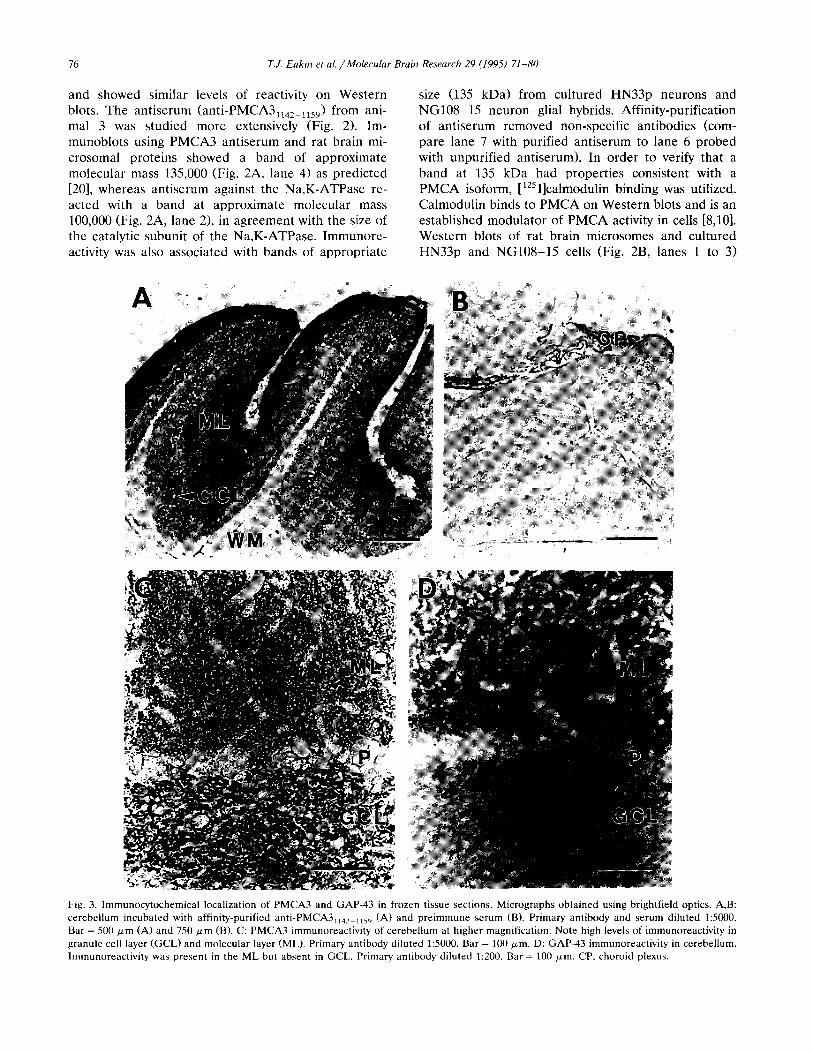
1234 567 123 Fig. 2. Western blots showing presence of PMCA3 isoform. Rat brain microsomes and cultured cells were solubilized and treated as described in Materials and methods. For electrophoresis, 25 pg of protein was loaded per lane. Polypeptides were transferred to nitrocellulose membrane and the resulting blots were divided into lanes and were individually probed with antisera or [ ‘2511calmodulin. Alignment of bands between blots is often imprecise and positions are considered approximate. Molecular weight markers are indicated to the left. A: immunoreactive bands were identified by chemiluminescence and exposed to X-ray film for 5 to 10 min. Lanes 1 to 4: rat brain microsomes. Lanes 1 and 3: controls with preimmune serum. Lane 2: polyclonal antibody against a3 subunit fusion protein of Na,K-ATPase. Primary antibody diluted 1:5000. Lane 4: polyclonal anti-PMCA3,,,z_,,s, (not affinity purified). Primary antibody diluted 1:lOOO. Lane 5: HN33p neurons probed with affinity-purified anti-PMCAS,,,,_,,,,. Primary antibody diluted 1:lOOO. Lanes 6 and 7: NGlO%15 neuron-glial hybrids probed with unpurified (lane 6) and affinity-purified (lane 7) anti-PMCA3,,,,_,,s,. Primary antibody diluted 1:lOOO. B: bands binding [ ‘251]calmodulin were identified by exposing blots to X-ray film for 4 h (lane 1) or 12 h (lanes 2 and 3). Lane 1: rat brain microsomes. Lane 2: HN33p cells. Lane 3: NGlO&15 cells.
76 T.J. Eukin et al. /Molecular Brain Research 29 (1995) 71-80
and showed similar levels of reactivity on Western blots. The antiserum (anti-PMCA3,,,,_,,,,) from ani- ma1 3 was studied more extensively (Fig. 2). Im- munoblots using PMCA3 antiserum and rat brain mi- crosomal proteins showed a band of approximate molecular mass 135,000 (Fig. 2A, lane 4) as predicted [20], whereas antiserum against the Na,K-ATPase re- acted with a band at approximate molecular mass 100,000 (Fig. 2A, lane 21, in agreement with the size of the catalytic subunit of the Na,K-ATPase. Immunore- activity was also associated with bands of appropriate
size (135 kDaI from cultured HN33p neurons and NG108-15 neuron-glial hybrids. Affinity-purification of antiserum removed non-specific antibodies (com- pare lane 7 with purified antiserum to lane 6 probed with unpurified antiserum). In order to verify that a band at 135 kDa had properties consistent with a PMCA isoform, [ 1251]calmodulin binding was utilized. Calmodulin binds to PMCA on Western blots and is an established modulator of PMCA activity in cells [8,101. Western blots of rat brain microsomes and cultured HN33p and NG108-15 cells (Fig. 2B, lanes 1 to 3)
Fig. 3. Immunocytochemical localization of PMCA3 and GAP-43 in frozen tissue sections. Micrographs obtained using brightfield optics. A,B: cerebellum incubated with affinity-purified anti-PMCA3, ,42_ 1 ,s9 (A) and preimmune serum (B). Primary antibody and serum diluted 15000. Bar = 500 Frn (A) and 750 pm (B). C: PMCA3 immunoreactivity of cerebellum at higher magnification. Note high levels of immunoreactivity in granule cell layer (GCL) and molecular layer (ML). Primary antibody diluted 1:5000. Bar = 100 pm. D: GAP-43 immunoreactivity in cerebellum. Immunoreactivity was present in the ML but absent in GCL. Primary antibody diluted 1:200. Bar = 100 pm. CP, choroid plexus.

T.J. Eakin et al. /Molecular Brain Research 29 (1995) 71-80 77
showed a high level of binding of [‘2511calmodulin associated with the 135 kDa band. [‘251]calmodulin also bound to other proteins present in these unpurified membrane preparations.
3.3. Localization of PMCA3 protein in rat brain
Using affinity-purified anti-PMCA3,,,,_,,,,, im- munoreactive sites were found in the molecular and granule cell layers of cerebellum (Fig. 3A) and in choroid plexus (Fig. 4A,B). In choroid plexus, PMCA3 immunostaining was strongest on the cerebrospinal fluid (CSFI-facing microvillus surface (Fig. 4B). Im-
munostaining was low in white matter (Fig. 3A) and in medial habenula (MH) (Fig. 4A). Preimmune serum controls (Fig. 3B) and pre-absorbed serum controls (not shown) exhibited very low background reactivity. Prominent immunoreactivity was found in the granule cell and molecular layers (Fig. 3C) and appeared to be localized in neuropil. Purkinje cells (Fig. 3A,C) had very low PMCA3-like immunoreactivity. Immunoreac- tivity for PMCA3 was not found in rat liver sections (not shown).
Distribution of growth-associated protein-43 (GAP- 43) [24], a marker for synaptic terminals and cell pro- cesses, (Fig. 3 ID). I m-
Fig. 4. Immunocytochemical localization of PMCA3 in choroid plexus and hippocampus in frozen and paraffin tissue sections incubated with affinity purified antkPMCA3,,,,_,,,,. Micrographs obtained using brightfield optics. A: immunoreactivity of choroid plexus in third ventricle. Primary antibody diluted 1:500. Bar = 200 Fm. B: immunoreactivity of choroid plexus in fourth ventricle. Immunoreactive sites were associated with the CSF-facing microvillus surface (arrow). Primary antibody diluted 1:SOOO. Bar = 100 Km. C: immunoreactivity of CA1 region of hippocampus (paraffin tissue section). Primary antibody diluted 1~500. Bar = 500 pm. D: immunoreactivity of CA1 region of hippocampus at higher magnification in the region of the pyramidal cell layer. Primary antibody diluted 1:500. Bar = 100 pm. MH, medial habenula; SO, stratum oriens; SP, stratum pyramidilae; SR, stratum radiatum.

78 T.J. Eakin et al. /Molecular Brain Research 29 (1995) 71-80
munoreactive sites were associated with the molecular layer and appeared as punctate deposits in the neu- ropil. Very little immunoreactivity was found in the granule cell layer.
In hippocampus immunostaining for PMCA3 was highest in the stratum oriens and stratum radiatum (Fig. 4C,D). Cell bodies of the pyramidal layer ap- peared to have lower immunoreactivity.
4. Discussion
The purpose of the present study was to establish if PMCA3 protein is expressed in the nervous system and to determine its localization. We found PMCA3-like immunoreactive protein expressed in rat brain and cultured neuronal cells, with the highest levels of im- munoreactivity associated with choroid plexus cells and with cell processes of cerebellum and hippocampus. The immunostaining results are supported by studies with immunoblots, which showed that antibodies to the C-terminal region of PMCA3 bind to a protein band with a molecular mass of approximately 135 kDa, con- sistent with the predicted value from molecular cloning studies [20]. Affinity-purification of antiserum removed non-specific antibodies (Fig. 2, lanes 6 and 7). The 135 kDa band also binds [‘251]calmodulin, which is known to bind to and activate PMCAs [lo], while calmodulin does not bind to Ca*+-ATPase of endoplasmic reticu- lum or other P-type ATPases. Not unexpectedly, [‘25Il calmodulin bound to other proteins present in these unpurified membrane preparations [29]. In addition, the pattern of immunocytochemical localization of PMCA3 in rat cerebellum and choroid plexus coincides with the observed distribution of PMCA3 mRNA in these cells. Together, these data support the conclu- sion that the PMCA3 isoform is expressed as a protein in the nervous system, principally by neurons and choroid plexus epithelial cells. Immunoreactivity was not found in rat liver, which does not contain apprecia- ble PMCA3 mRNA [231.
The existence of translated PMCA protein in ner- vous tissue has been previously reported by Penniston and colleagues [4,14,32] using a monoclonal antibody (anti-PMCA 5FlO) raised against human erythrocyte Ca2+-ATPase holoenzyme, a preparation which con- tains isoforms PMCAl and PMCA4 [42]. Anti-PMCA 5FlO binds to amino acid residues 714 to 738 in the large cytoplasmic loop [ll] between putative trans- membrane domains M4 and M5 [l], an epitope com- mon to all PMCA isoforms. Anti-PMCA 5FlO was used to probe western blots of synaptosomes [14,32] and choroid plexus [4] and bands of predicted molecu- lar mass (130 to 140 kDa) were identified. In addition immunocytochemical localization studies in avian and mammalian brain [14] and in choroid plexus [41 were
carried out using anti-PMCA 5FlO confirming, the presence of PMCA epitopes in brain tissue. In another study, Mann et al. [271 used anti-peptide antibodies based on deduced amino acid sequence from a PMCA clone obtained from bovine brain (possibly a new iso- form, PMCAS). Immunoreactivity was associated with Purkinje and granule cells of cerebellum and pyramidal cells of hippocampus.
Cellular localization studies using ISH show that PMCA3 mRNA is expressed at moderate levels in neurons throughout rat brain [38], and particularly at high levels in choroid plexus epithelial cells, and in granule cells of cerebellum (Fig. 1). Both of these cells are advantageous for study of expression of PMCA3 protein due to their known roles in CNS function and well-defined morphological characteristics.
Choroid plexus cells, a non-neuronal cell type, func- tion in production of CSF in the CNS by transporting water and solutes, and may also be involved in regula- tion of CSF Ca*+ levels. Using anti-PMCA3,,,,_,,,,, immunoreactivity was localized to the CSF-facing mi- crovillus surface, indicating that PMCA3 is probably the major PMCA isoform in these cells. This is also consistent with our previous ISH study which showed that the levels of PMCA3 mRNA were 10 to 30 times higher than PMCAl and PMCA2 in choroid plexus. PMCA4 mRNA has not been detected in choroid plexus by ISH [39]. Borke et al. [4] used anti-PMCA 5FlO and found PMCA-immunoreactive sites on the same CSF-facing surface.
In earlier work ISH probes hybridized to PMCA3 mRNA in the medial habenula [38], but very little immunoreactivity was found in this region. One inter- pretation of this result is that little PMCA3 protein is associated with soma of MH neurons, but may be present on axons or nerve terminals which project to other sites in brain. This would be consistent with our finding that PMCA3 immunoreactivity is associated with cellular processes of cerebellar and hippocampal neurons.
The present work shows that cerebellum contains high levels of PMCA3 immunoreactivity associated with granule cells. These are small, glutaminergic neurons which serve to relay mossy fiber inputs to other cell types within the cerebellar cortex. They express a vari- ety of voltage-gated channels [12] and excitatory amino acid receptor subtypes 1171 and stimulation of cultured granule cells by glutamate causes transient increases of [Ca2+li [12]. Mechanisms of calcium homeostasis in these cells may involve operation of several homeo- static mechanisms, including PMCA-mediated trans- port of Ca *+ into the extracellular space. The present study did not detect PMCA3 immunoreactivity associ- ated with granule cell bodies and the focal reactive sites in neuropil within the granule cell layer are likely associated with the dendrites of these granule cells. In

T.J. Eakin et al. /Molecular Brain Research 29 (1995) 71-80 79
the molecular layer immunoreactive sites are most likely associated with processes or nerve terminals pro- jecting from the granule cells. Very low PMCA3-like immunoreactivity was associated with Purkinje cells, consistent with ISH results (Fig. 1A).
PMCA3 is expressed in rat brain predominantly as two mRNA splice variants, designated PMCA3a and PMCA3b [5,6,23] which are alternatively spliced within the calmodulin binding domain. A 19-amino acid pep- tide (18 amino acids of PMCA3 sequence and terminal cysteine for coupling to carrier) was used for produc- tion of antibodies in this study. The complete 18 amino acid sequence is present in PMCA3a, but only a lo- amino acid portion is found in PMCA3b. Therefore, the localization of PMCA3 reported here may not distinguish between PMCA3a and PMCA3b isoform variants.
In order to provide additional evidence in support of localization of PMCA3 at synaptic contacts in the molecular layer, we examined distribution of growth- associated protein-43 (GAP-431 [24]. This protein is known to be present in axons and nerve terminals, probably associated with presynaptic plasma mem- branes in adult cerebellum [301 but is absent from granule cell bodies and dendrites located in the gran- ule cell layer [13,30]. We found that GAP-43 im- munoreactive sites were associated with the molecular layer and appeared as punctate deposits in the neu- ropil, a pattern of localization also found using anti-
PMCA311,,-115,. As previously reported [13,30], GAP- 43 immunoreactivity was absent from the granule cell layer. The congruent pattern of localization found for PMCA3 and GAP-43 in the molecular layer suggests that at least a portion of the sites localized using anti-PMCA3,,,,_,,,, antibodies are probably presynap- tic terminals [18] of granule cells. Purkinje cell im- munoreactivity to PMCA3 was unlikely, due to the low levels of PMCA3 mRNA found in these cells and the apparent absence of a pattern of immunoreactivity consistent with Purkinje cell dendritic arborization. The results of De Talamoni et al. [14] using anti-PMCA 5FlO showed that an epitope of one or more PMCA isoforms is associated with Purkinje cells in avian and mammalian cerebellum. Moderate PMCA immunore- activity was associated with soma and distal dendrites in the molecular layer. This pattern of localization was not found in the present study. Together the evidence suggests that Purkinje cells express a PMCA isoform(s) other than PMCA3.
In hippocampus the stratum oriens and stratum radiatum showed high immunoreactivity to PMCA3. Lower immunoreactivity was found with soma in the pyramidal cell layer itself. This layer predominantly contains large, excitatory glutaminergic pyramidal cells which were previously shown [38] to express PMCA3 mRNA. The stratum oriens and stratum radiatum con-
tain numerous synaptic contacts, but it is unclear which type of synapse expresses PMCA3 immunoreactivity or whether glial cells may also be involved. The ISH results showed that pyramidal cells are the major sites of production of PMCA3 mRNA [38]. However, contri- butions of other cell types are not known and must await ISH studies utilizing cell-specific markers. Im- munocytochemical studies at the ultrastructural level may also be required. Based on the present results with cerebellum and hippocampus, it appears that neuronal cell processes are a major site of expression of PMCA3 protein in rat brain.
Acknowledgements
This research was supported by the Medical Re- search Service of the Department of Veterans Affairs, by grants from the National Science Foundation (W.L.S.), CONICET (M.C.A.) and by NIH Grants NS 20482 (W.L.S.) and AM 17047 (Diabetes Endocrinol- ogy Research Center, University of Washington). We thank Dr. Bruce Wainer for kindly providing HN33p pyramidal neurons and John Breininger for excellent technical assistance.
References
111
121
[31
[41
El
161
[71
k31
Adamo, H.P., Caride, A.J. and Penniston, J.T., Use of expres- sion mutants and monoclonal antibodies to map the erythrocyte Ca*+ pump, J. Biol. Chem., 267 (1992) 14244-14249. Benham, CD., Evans, M.L. and McBain, C.J., Ca2+ efflux mechanisms following depolarization evoked calcium transients in cultured rat sensory neurones, J. Physiol., 455 (1992) 567-583. Bleakman, D., Roback, J.D., Wainer, B.H., Miller, R.J. and Harrison, N.L., Calcium homeostasis in rat septal neurons in tissue culture, Brain Rex, 600 (1993) 257-267. Borke, J.L., Caride, A.J., Yaksh, T.L., Penniston, J.T. and Kumar, R., Cerebrospinal fluid calcium homeostasis: evidence for a plasma membrane Ca2+-pump in mammalian choroid plexus, Brain Rex, 489 (1989) 355-360. Brandt, P. and Neve, R.L., Expression of plasma membrane calcium-pumping ATPase mRNAs in developing rat brain and adult brain subregions: evidence for stage-specific expression, I Neurochem., 59 (1992) 1566-1569. Brandt, P., Neve, R.L., Kammesheidt, A., Rhoads, R.E. and Vanaman, T.C., Analysis of the tissue-specific distribution of mRNAs encoding the plasma membrane calcium-pumping AT- Pases and characterization of an alternately spliced form of PMCA4 at the cDNA and genomic levels, J. Biol. Chem., 267 (1992) 4376-4385. Burk, SE. and Shull, G.E., Structure of the rat plasma mem- brane Ca2+-ATPase isoform 3 gene and characterization of alternative splicing and transcription products. Skeletal muscle- specific splicing results in a plasma membrane CaZf-ATPase with a novel calmodulin-binding domain, .I. Biol. Chem., 267 (1992) 19683-19690. Carafoli, E., The plasma membrane calcium pump. Structure, function, regulation, Biochim. Biophys. Acta, 1101 (1992) 266- 267.

T.I. Eakin et al. /Molecular Brain Research 29 (1995) 71-80 80
[91
[lOI
[Ill
[121
[I31
[I41
[151
[161
[I71
[181
[I91
DO1
D11
Dl
D31
D41
Dl
Carafoli, E. and Guerini, D., Molecular and cellular biology of plasma membrane calcium ATPase, Trends Cardioc,asc. Med., 3 (1993) 177-184. Carafoli, E., Kessler, F., Falchetto, R., Heim, R., Quadroni, M., Krebs, J., Strehler, E.E. and Vorherr, T., The molecular basis of the modulation of the plasma membrane calcium pump by calmodulin, Ann. N.Y. Acad. Sci., 671 (1992) 58-69. Caride, A.J., Filoteo, A.G., Enyedi, A. and Penniston, J.T., Epitope location and isoenzyme specificity of three monoclonal antibodies against the plasma membrane calcium pump, Bio- phys. J., 66 (1994) A120. Ciardo, A. and Meldolesi, J., Regulation of intracellular calcium in cerebellar granule neurons: effects of depolarization and of glutaminergic and cholinergic stimulation, J. Neurochem., 56 (1991) 184-191. Dani, J.W., Armstrong, D.M. and Benowitz, L.I., Mapping the development of the rat brain by GAP-43 immunocytochemistry, Neuroscience, 40 (1991) 277-287. De Talamoni, N.T., Smith, C.A., Wasserman, R.H., Beltramino, C., Fullmer, C.S. and Penniston, J.T., Immunocytochemical lo- calization of the plasma membrane calcium pump, calbindin- D28k, and parvalbumin in Purkinje cells of avian and mam- malian cerebellum, Proc. Natl. Acad. Sci. USA, 90 (1993) 11949- 11953. Eakin, T.J., Stahl, W.L. and Anderson, W.R., Localization of mRNAs encoding isoforms of cation transporting ATPases by in situ hybridization, J. Histochem. Cytochem., 41 (1993) 1128. Filuk, P.E., Miller, M.A., Dorsa, D.M. and Stahl, W.L., Local- ization of messenger RNA encoding isoforms of the catalytic subunit of the Na,K-ATPase in rat brain by in situ hybridization histochemistry, Neurosci. Rex Commun., 5 (1989) 155-162. Gallo, V., Suergiu, R., Giovannini, C. and Levi, G., Glutamate receptor subtypes in cultured cerebellar neurons: modulation of glutamate and y-aminobutyric acid release, L Neurochem., 49 (1987) 1801-1809. Gispen, W.H., Leunissen, J.L.M., Oestreicher, A.B., Verkleij, A.J. and Zwiers, H., Presynaptic localization of B-50 phospho- protein: the ACTH-sensitive protein kinase substrate involved in rat brain polyphosphoinoisitide metabolism, Brain Rex, 328 (1985) 381-385. Goh, J.W., Sanches-Vives, M.V. and Pennefather, P.S., Influ- ence of Na/Ca exchange and mobilization of intracellular cal- cium on the time course of the slow afterhyper-polarization current (IAHP) in bullfrog sympathetic ganglion neurons, Neu- rosci. Lett., 138 ( 1992) 123- 127. Greeb, J. and Shull, G.E., Molecular cloning of a third isoform of the calmodulin-sensitive plasma membrane Ca*+-transporting ATPase that is expressed predominantly in brain and skeletal muscle, J. Biol. Chem., 264 (1989) 18569-18576. Grover, A.K. and Khan, I., Calcium pump isoforms: diversity, selectivity and plasticity, Cell Calcium, 13 (1992) 9-17. Hoffman, G.E., Smith, M.S. and Fitzsimmons, M.D., Detecting steroidal effects of immediate early gene expression in the hypothalamus, Neuroprotocols, 1 (1992) 52-66. Keeton, T.P., Burk, SE. and Shull, G.E., Alternative splicing of exons encoding the calmodulin-binding domains and C termini of plasma membrane CaZf-ATPase isoforms 1, 2, 3, and 4, J. Biol. Chem., 268 (1993) 2740-2748. Kruger, L., Bendotti, C., Rivolta, R. and Samanin, R., Distribu- tion of GAP-43 mRNA in the adult rat brain, J. Camp. Neural., 333 (1993) 417-434. Lee, H.J., Hammond, D.N., Large, T.H., Roback, J.D., Sim, J.A., Brown, D.A., Otten, U.H. and Wainer, B.H., Neuronal properties and trophic activities of immortalized hippocampal ceils from embryonic and young adult mice, J. Neurosci., 10 (1990) 1779-1787.
[26] Levy, S. and Tillotson, D., Effects of Na+ and Ca2+ gradients on intracellular free Ca2+ in voltage-clamped Aplysia neurons, Brain Rex, 474 (1988) 333-342.
[27] Mann, D.M., Brandt, P., Sisken, B. and Vanaman, T.C., Identi- fication of a synaptic membrane-localized isoform of the cal- cium-pumping ATPase, Adu. Exp. Med. BioL, 255 (1990) 71-78.
[28] Miller, R.J., The control of neuronal Ca2+ homeostasis, Prog. Neurobiol., 37 (1991) 255-285.
[29] Natsukari, N., Miwa, A. and Fujita, M., Calmodulin-binding proteins of calcium-independent type in rat brain synaptosomal membranes: their localization and properties, Biochem. Biophys. Res. Commun., 183 (1992) 725-732.
[30] Oestreicher, A.B. and Gispen, W.H., Comparison of the im- munocytochemical distribution of the phosphoprotein B-50 in the cerebellum and hippocampus of immature and adult rat brain, Brain Rex, 375 (1986) 267-279.
[31] Penniston, J.T. and Enyedi, A., Plasma membrane Ca2+ pump: recent developments, Cell. Physiol. Biochem., 4 (1994) 148-159.
[32] Sarkadi, B., Enyedi, A., Penniston, J.T., Verman, A.K., Dux, L., Molnar, E. and Gardos, G., Characterization of membrane calcium pumps by simultaneous immunoblotting and 32P radiog- raphy, Biochim. Biophys. Acta, 939 (1988) 40-46.
[33] Schellenberg, G., Pech, I.V. and Stahl, W.L., Immunoreactivity of subunits of the (Na+,K+)-ATPase: cross-reactivity of the ~y,oy( + I and p forms in different organs and species, Biochim. Biophys. Acta, 649 (1981) 691-700.
[34] Shull, G.E. and Greeb, J., Molecular cloning of two isoforms of the plasma membrane Ca*+-transporting ATPase from rat brain, J. Biol. Chem., 263 (1988) 8646-8657.
I351 Stahl, W.L. and Baskin, D.G., Histochemistry of ATPases, J. Histochem. Cytochem., 38 (1990) 1099-1122.
[36] Stahl, W.L., Eakin, T.J., Anderson, W.R., Owens Jr., J.W.M., Breininger, J.F. and Filuk, P.E., Localization of mRNA coding for plasma membrane Ca-ATPase isoforms in rat brain by in situ hybridization, Ann. N.Y. Acad. Sci., 671 (1992) 433-435.
[37] Stahl, W.L., Eakin, T.J. and Baskin, D.G., Selection of oligonu- cleotide probes for detection of mRNA isoforms, J. Histochem. Cytochem., 41 (1993) 1735-1740.
[38] Stahl, W.L., Eakin, T.J., Owens Jr., J.W.M., Breininger, J.F., Filuk, P.E. and Anderson, W.R., Plasma membrane Ca2+- ATPase isoforms: distribution of mRNAs in rat brain by in situ hybridization, Mol. Brain Res., 16 (1992) 223-231.
[39] Stahl, W.L., Keeton, T.P. and Eakin, T.J., The plasma mem- brane Ca2+-ATPase mRNA isoform, PMCA 4, is expressed at high levels in neurons of rat piriform cortex and neocortex, Neurosci. Lett., 178 (1994) 267-270.
[40] Stauffer, T.P., Hilfiker, H., Carafoli, E. and Strehler, E.E., Quantitative analysis of alternative splicing options of human plasma membrane calcium pump genes, J. Biol. Chem., 268 (1993) 25993-26003.
[41] Strehler, E.E., Plasma membrane Ca*+ pumps and Na+/Ca*+ exchangers, Seminars Cell Biol., 1 (1990) 283-295.
[42] Strehler, E.E., James, P., Fischer, R., Heim, R., Vorherr, T., Filoteo, A.G., Penniston, J.T. and Carafoli, E., Peptide se- quence analysis and molecular cloning reveal two Ca pump isoforms in the human erythrocyte membrane, J. Biol. Chem., 265 (1990) 2835-2842.
[43] Thayer, S.A. and Miller, R.J., Regulation of the intracellular free calcium concentration in single rat dorsal root ganglion neurones in vitro, J. Physiol., 425 (1990) 85-115.
[44] Wang, K.K.W., Villalobo, A. and Roufogalis, B.D., The plasma membrane calcium pump: a multiregulated transporter, Trends Cell Biol., 2 (1992) 46-52.
[45] Wuytack, F. and Raeymaekers, L., The Ca2+-transport ATPases from the plasma membrane, J. Bioenerg. Biomembr., 24 (1992) 285-300.
Related Documents











