©2007 LANDES BIOSCIENCE. DO NOT DISTRIBUTE. 16 RNA Biology 2006; Vol. 4 Issue 1 Research Paper Localization of the Developmental Timing Regulator Lin28 to mRNP Complexes, P-bodies and Stress Granules [RNA Biology 4:1, 16-25, January/February/March 2007]; ©2007 Landes Bioscience Erica Balzer Eric G. Moss* Department of Molecular Biology; University of Medicine and Dentistry of New Jersey; Stratford, New Jersey USA *Correspondence to: Eric G. Moss; Department of Molecular Biology; University of Medicine and Dentistry of New Jersey; 2 Medical Center Drive; Stratford, New Jersey 08084 USA; Tel.: 856.566.2896; Fax: 856.566.6291; Email: mosseg@ umdnj.edu Original manuscript submitted: 03/09/07 Manuscript accepted: 04/30/07 Previously published online as a RNA Biology E-publication: http://www.landesbioscience.com/journals/rnabiology/abstract.php?id=4364 KEY WORDS P-bodies, stress granules, cold shock domain, CCHC zinc finger, nucleocytoplasmic shut- tling, mRNPs, polyribosomes ABBREVIATIONS CSD cold shock domain CCHC cys-cys-his-cys IF immunofluorescence PABP poly(A)-binding protein TAP tandem affinity purification NLS nuclear localization signal ACKNOWLEDGEMENTS This work was supported by the National Science Foundation (IBN-0212373) and UMDNJ. We are very grateful to Kevin Kemper for technical support, Dr. Jack Keene for anti-PABP antibodies, Dr. Nancy Kedersha for Dcp1a:RFP and TIA:RFP plasmids, to Dr. Andrej Sali for homology modeling, and to Dr. Michael Henry, Dr. Anita Pepper, and anonymous reviwers for critical comments on the manuscript. ABSTRACT Lin28 is a conserved cytoplasmic protein with an unusual pairing of RNA‑binding motifs: a cold shock domain and a pair of retroviral‑type CCHC zinc fingers. In the nematode C. elegans, it is a regulator of developmental timing. In mammals, it is abundant in diverse types of undifferentiated cells. However, its molecular function is unknown. In pluripotent mammalian cells, Lin28 is observed in RNase‑sensitive complexes with poly(A)‑binding protein, and in polysomal fractions of sucrose gradients, suggesting it is associated with translating mRNAs. Upon cellular stress, Lin28 locates to stress granules, which contain non-translating mRNA complexes. However, Lin28 also localizes to cytoplasmic Processing bodies, or P‑bodies, sites of mRNA degradation and microRNA regulation, consistent with it acting to regulate mRNA translation or stability. Mutational analysis shows that Lin28’s conserved RNA binding domains cooperate to put Lin28 in mRNPs, but that only the CCHC domain is required for localization to P‑bodies. When both RNA‑binding domains are mutated, Lin28 accumulates in the nucleus, suggesting that it normally shuttles from nucleus to cytoplasm bound to RNA. These studies are consistent with a model in which Lin28 binds mRNAs in the nucleus and accompanies them to ribosomes and P‑bodies. We propose that Lin28 influences the translation or stability of specific mRNAs during differentiation. INTRODUCTION Post‑transcriptional regulation occurs widely to control the time and place of gene activity. 1 Regulatory factors act on specific mRNAs to alter their nuclear processing, cytoplasmic transport, translation, or stability. 2 In addition to proteins, microRNAs also regulate gene activity by basepairing with mRNAs and inhibiting their translation and stability. 3 The fates of the regulated mRNAs affect the amount and types of protein synthesized, and ultimately cell proliferation, differentiation, and function. As complexes of mRNAs and proteins (mRNPs) move within the cell, they are exten- sively remodeled as the constituents of the complexes change. 4,5 For example, some proteins that bind the mRNA during splicing, leave the mRNP before it exits the nucleus while others remain associated, and others join the mRNPs after nuclear export. In the cytoplasm, the complexes may be transported to different sub‑cellular compartments where they are translated, sequestered in storage granules, or processed for degradation. 6,7 At any point, gene expression may be controlled by specific regulatory proteins that alter the fate of particular mRNAs. Many mRNA‑binding proteins that act in post‑transcriptional regulation are known from studies of development. Sex‑lethal of Drosophila regulates splicing of specific mRNAs to control sex determination. 8 Staufen mediates the localization of particular mRNAs in fly embryos. 9 CPEB proteins affect poly(A) tail length and consequently mRNA transla- tion and stability. 10 Other RNA binding regulators are known from genetic studies, and often they affect the translation of their target mRNAs. 11,12 The activity of microRNAs on their mRNA targets may also be influenced by specific mRNA‑binding proteins. 13,14 However, the molecular mechanisms of many regulatory RNA‑binding proteins known from developmental studies remain unclear. The developmental timing pathway of C. elegans is comprised of several genes that are post‑transcriptionally regulated. This pathway controls the succession of patterns of divi- sion and differentiation during the animal’s larval stages. 15 MicroRNAs play a prominent role in this pathway controlling the stage‑specific repression of several key targets. One of these is lin‑28, a regulator at a major developmental transition in C. elegans development,

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

©2007 L
ANDES BIOSCI
ENCE.
DO NOT DIST
RIBUTE.
16 RNABiology 2006;Vol.4Issue1
Research Paper
Localization of the Developmental Timing Regulator Lin28 to mRNP Complexes, P-bodies and Stress Granules
[RNABiology4:1,16-25,January/February/March2007];©2007LandesBioscience
Erica BalzerEric G. Moss*
Department of Molecular Biology; University of Medicine and Dentistry of New Jersey; Stratford, New Jersey USA
*Correspondence to: Eric G. Moss; Department of Molecular Biology; University of Medicine and Dentistry of New Jersey; 2 Medical Center Drive; Stratford, New Jersey 08084 USA; Tel.: 856.566.2896; Fax: 856.566.6291; Email: [email protected]
Original manuscript submitted: 03/09/07Manuscript accepted: 04/30/07
Previously published online as a RNA Biology E-publication:http://www.landesbioscience.com/journals/rnabiology/abstract.php?id=4364
KEy words
P-bodies,stressgranules,coldshockdomain,CCHC zinc finger, nucleocytoplasmic shut-tling,mRNPs,polyribosomes
ABBrEviAtions
CSD coldshockdomainCCHC cys-cys-his-cysIF immunofluorescencePABP poly(A)-bindingproteinTAP tandemaffinitypurificationNLS nuclearlocalizationsignal
AcKnowlEdGEMEnts
This work was supported by the NationalScience Foundation (IBN-0212373) andUMDNJ. We are very grateful to KevinKemperfortechnicalsupport,Dr.JackKeeneforanti-PABPantibodies,Dr.NancyKedershafor Dcp1a:RFP and TIA:RFP plasmids, toDr.AndrejSali forhomologymodeling,andtoDr.MichaelHenry,Dr.AnitaPepper,andanonymousreviwersforcriticalcommentsonthemanuscript.
ABstrActLin28 is a conserved cytoplasmic protein with an unusual pairing of RNA‑binding
motifs: a cold shock domain and a pair of retroviral‑type CCHC zinc fingers. In the nematode C. elegans, it is a regulator of developmental timing. In mammals, it is abundant in diverse types of undifferentiated cells. However, its molecular function is unknown. In pluripotent mammalian cells, Lin28 is observed in RNase‑sensitive complexes with poly(A)‑binding protein, and in polysomal fractions of sucrose gradients, suggesting it is associated with translating mRNAs. Upon cellular stress, Lin28 locates to stress granules, which contain non-translating mRNA complexes. However, Lin28 also localizes to cytoplasmic Processing bodies, or P‑bodies, sites of mRNA degradation and microRNA regulation, consistent with it acting to regulate mRNA translation or stability. Mutational analysis shows that Lin28’s conserved RNA binding domains cooperate to put Lin28 in mRNPs, but that only the CCHC domain is required for localization to P‑bodies. When both RNA‑binding domains are mutated, Lin28 accumulates in the nucleus, suggesting that it normally shuttles from nucleus to cytoplasm bound to RNA. These studies are consistent with a model in which Lin28 binds mRNAs in the nucleus and accompanies them to ribosomes and P‑bodies. We propose that Lin28 influences the translation or stability of specific mRNAs during differentiation.
introductionPost‑transcriptional regulation occurs widely to control the time and place of gene
activity.1 Regulatory factors act on specific mRNAs to alter their nuclear processing,cytoplasmic transport, translation, or stability.2 In addition to proteins, microRNAsalso regulate gene activity bybasepairingwithmRNAs and inhibiting their translationandstability.3ThefatesoftheregulatedmRNAsaffecttheamountandtypesofproteinsynthesized,andultimatelycellproliferation,differentiation,andfunction.
AscomplexesofmRNAsandproteins(mRNPs)movewithinthecell,theyareexten-sively remodeled as the constituents of the complexes change.4,5 For example, someproteinsthatbindthemRNAduringsplicing,leavethemRNPbeforeitexitsthenucleuswhileothersremainassociated,andothers jointhemRNPsafternuclearexport.Inthecytoplasm, the complexes may be transported to different sub‑cellular compartmentswheretheyaretranslated,sequesteredinstoragegranules,orprocessedfordegradation.6,7Atanypoint,geneexpressionmaybecontrolledbyspecificregulatoryproteinsthatalterthefateofparticularmRNAs.
ManymRNA‑bindingproteinsthatact inpost‑transcriptionalregulationareknownfromstudiesofdevelopment.Sex‑lethalofDrosophilaregulatessplicingofspecificmRNAstocontrolsexdetermination.8StaufenmediatesthelocalizationofparticularmRNAsinflyembryos.9CPEBproteinsaffectpoly(A)taillengthandconsequentlymRNAtransla-tionandstability.10OtherRNAbindingregulatorsareknownfromgeneticstudies,andoftentheyaffectthetranslationoftheirtargetmRNAs.11,12TheactivityofmicroRNAsontheirmRNAtargetsmayalsobeinfluencedbyspecificmRNA‑bindingproteins.13,14However, themolecularmechanismsofmanyregulatoryRNA‑bindingproteinsknownfromdevelopmentalstudiesremainunclear.
ThedevelopmentaltimingpathwayofC. elegansiscomprisedofseveralgenesthatarepost‑transcriptionallyregulated.Thispathwaycontrolsthesuccessionofpatternsofdivi-sionanddifferentiationduringtheanimal’slarvalstages.15MicroRNAsplayaprominentroleinthispathwaycontrollingthestage‑specificrepressionofseveralkeytargets.Oneoftheseislin‑28,aregulatoratamajordevelopmentaltransitioninC. elegansdevelopment,

www.landesbioscience.com RNABiology 17
Lin28inmRNPsandP-bodies
whichispost‑transcriptionallyrepressedbythemicroRNAlin‑4.16,17Iflin‑28ismutant,animalsskipeventsspecificforthesecondlarvalstage and later events occur precociously, resulting in a deformedadult.18 lin‑28 appears to positively regulate two other importantmicroRNA‑regulatedgenesthatactinthispathway,buthowitdoesthisisnotknown.19,20
Mammals and other animals encode a Lin28 homolog in theirgenomes.21As inC. elegans,Lin28ofmammals isdown‑regulatedbymicroRNAsduringdifferentiation.21‑23Theproteinisabundantin undifferentiated cells and developing tissues from early‑stageembryos to adults and is absent from differentiated cells.21,24 It isexpressed specifically in undifferentiated embryonic stem cells andisoneofseveralfactorsthatdefinetheimmaturestateofthesecells,andoneof thefewthatdoesnotencodeanapparenttranscriptionfactor.25
Lin28 is a cytoplasmic protein with a unique pairing of RNAbindingdomains: a cold‑shockdomain (CSD), and retroviral‑typeCCHCzincfingers(Fig.1).16Bothofthesedomainsarefoundinwell‑characterizedRNA‑bindingproteins.TheCSDisthecharacter-isticdomainofY‑boxproteins,whichincludeFRGY2ofXenopusandYB‑1ofhumans,andresemblestheRNA‑recognitionmotif(RRM)domainthatexistsinmanyRNA‑bindingproteins.26,27TheCCHCzincfingerscloselyresemblethoseofthesmallnucleocapsidproteinNCp7ofHIVandotherretrovirusesthatbindstheviralgenomeataspecificsite.16BothofthesedomainshavebeenwellstudiedwithregardtotheirstructuresandRNA‑bindingactivities.28,29However,Lin28istheonlyanimalproteintohavethemboth.
We set out todeterminehowLin28 acts in the cell to regulategene expression. We asked where Lin28 localizes in cells, what itcomplexeswith,andwhataretherelativerolesofitstwoconserveddomains in mediating these activities. To do this, we used bothmolecularandcellbiologyapproachesinundifferentiatedembryonal
carcinoma cells, which naturally express Lin28. Mammalian tissueculture cells provide many distinct advantages over C. elegans foraddressingthesequestions,includingtheirlargesizeformicroscopy,amenability to affinity purification methods, and availability ofmarkers. Our findings provide insight into how Lin28 may havea specialized function in regulating its targets during the develop-mentaltransitionsitgoverns.
MAtEriAls And MEthodsPlasmids. All plasmids were constructed in the expression
vector pCAGI‑Puro (Ian Chambers). The open reading frames ofhumanLin28 (GenbankAccessionNM024674) andhumanYB‑1(NM004559)werefusedtoEGFP,TAP,andfireflyluciferaseattheC‑terminus.
Cell culture and cell transfection.MouseP19andhumanNT2cellswereobtainedfromtheAmericanTypeCultureCollection.P19cells were maintained in alpha minimal essential medium (Sigma)supplemented with 2.5% fetal bovine serum, 7.5% bovine serum,and50U/mlofpenicillin/streptomycin.NT2cellsweremaintainedin DMEM containing 4mM L‑glutamine, 4.5 g/L glucose and1.5g/L sodiumbicarbonate (Gibco) supplementedwith10%fetalbovine serum and 50 units/ml penicillin/streptomycin. Cell linesweretransfectedusingFugene6transfectionreagentasdescribedbythemanufacturer(Roche).Stablytransfectedcelllineswereselectedusing2mg/mlpuromycin(Sigma).
TAP purification. Approximately 500ml of packed cells werethawedoniceandresuspendedin5mlLysisBuffer(150mMNaCl,2.5 mM MgCl2, 10 mM Hepes, pH 7.5, 0.05% NP‑40) supple-mentedwith2mg/mlheparin, 0.2%vanadyl complexes, 0.2mMPMSF, 1 mM DTT and Complete EDTA‑free protease inhibitorcocktail (Roche). Cells were lysed following one round of freeze/thawandcellulardebrisandnucleiwerepelletedbycentrifugation.Absorbance at 260 nm was taken of the clarified supernatant andequivalent amounts of control and test lysates were added to 100ml packed IgG‑sepharose beads (Amersham). EDTA was addedto 20mM and binding was done at 4˚C for 2 hours. Beads wereharvestedandwashedwithLysisBuffer,3MUrea/LysisBuffer,andTEVBuffer (10mMHepes,pH7.5,150mMNaCl,0.1%NP‑40,0.5mMEDTAand1mMDTT).BeadswereresuspendedinTEVBuffer and 50 units AcTEV protease (Invitrogen). Cleavage wasdoneat4˚Cfor1hour.Beadswereharvestedbycentrifugationandthe super was removed and concentrated over aYM‑10 Microconcolumn.2XSDSloadingbufferwasaddedtosamplesbeforesepara-tionbySDS‑PAGE.
SDS‑PAGE and immunoblotting.Wholecelllysatesandeluatesfrom TAP purification were fractionated in NuPAGE 4–12%Bis‑Tris polyacrylamide gels (Invitrogen) and transferred to PVDFmembranes.Membraneswereincubatedwithblockingsolutionfor1hrandthenwithanti‑Lin2821(1:2500),anti‑TAP(1:2000;OpenBiosystems), or anti‑PABP (1:10,000; J. Keene, Duke Univeristy).BoundratorrabbitantibodiesweredetectedusingHRP‑conjugatedgoatanti‑ratorgoatanti‑rabbitantibodies(BioRad).
Sucrose Gradients. 500 ml of frozen cells were thawed on iceand resuspended in equal volume Lysis Buffer (150 mM NaCl,2.5mMMgCl210mMHepes,pH7.5,0.5%NP‑40)supplementedwith 2 mg/ml heparin, 0.2% vanadyl complexes, 0.2 mM PMSF,1mM DTT, 100mg/ml cyclohexamide and Complete EDTA‑freeprotease inhibitor cocktail (Roche). Following three freeze/thaws,lysates were clarified by centrifugation. Equivalent amounts of
Figure 1. Schematic structure of Lin28. (A) Homology model of Lin28 based on the CSD of human YB-1 and NCp7 of HIV-1. Blue, beta sheet, Red, alpha helix, Yellow, residues mutated. (B) Linear representation of Lin28. Grey boxes, CSD and CCHC domain. rnp1, rnp2, RNA-binding motifs in CSD. Residues mutated indicated by arrows. Below, three deletion forms discussed in the text.
Lin28inmRNPsandP-bodies
18 RNABiology 2007;Vol.4Issue1
cytoplasmicextracts(approximately40A260units)wereloadedonto15–45%w/wlineardensitygradientofsucrosein200mMTris,pH8.5,50mMKCl,25mMMgCl2.Intheindicatedsamples,EDTAwas added to a final 30 mM concentration before loading on thegradient.Thegradientswerecentrifugedat40,000xg for90minat4˚inaBeckmanSW41rotor.Fractions(0.5mL)werecollectedwhile reading absorbance at 254nm using an ISCO UA‑6 UVdetector.Fractionswerepooledandanalyzedbyimmunoblot.
Immunofluorescence and microscopy. Cells were grown oncoverslipsfor24hours,washedwithPBS,thenstainedwithHoechst33342 (Invitrogen). Expressionwasmonitoredwith a fluorescencemicroscope (Leica). For direct immunofluorescence, cells weregrown on coverslips and fixed with 4% formaldehyde. Fixed cellswerewashedwithPBSandpermeabilized inPBScontaining0.1%Triton‑X100for5minutes.Primaryandsecondaryantibodieswerediluted inPBSTcontaining2%bovine serumalbumin (BSA)andincubatedfor1houratroomtemperature.Thefollowingantibodiesandtheirrespectivedilutionswereused:ratanti‑Lin2821(1:500)andgoat anti‑rat Alexafluor488 (1:2000; Invitrogen). To induce stressgranule formation, transfected cells were heat‑shocked at 44°C for20min,washedwithPBSandfixedimmediately.
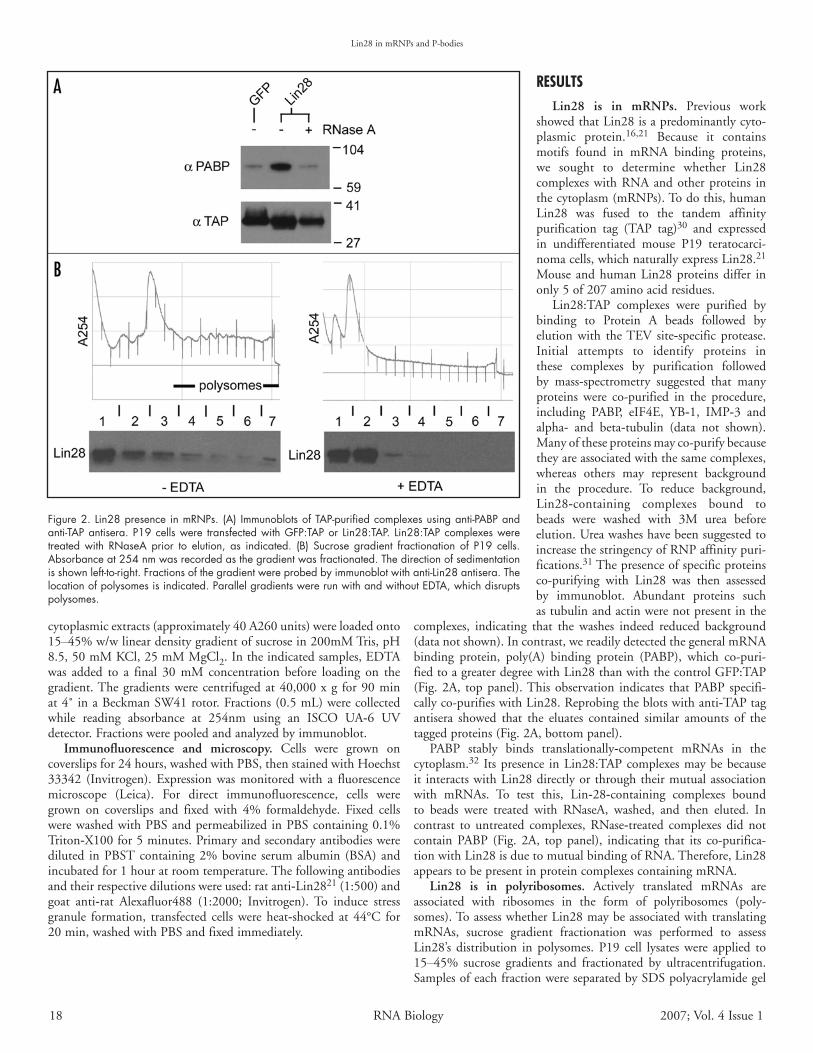
rEsultsLin28 is in mRNPs. Previous work
showedthatLin28isapredominantlycyto-plasmic protein.16,21 Because it containsmotifs found in mRNA binding proteins,we sought to determine whether Lin28complexeswithRNAandotherproteins inthecytoplasm(mRNPs).Todothis,humanLin28 was fused to the tandem affinitypurification tag (TAP tag)30 and expressedin undifferentiated mouse P19 teratocarci-nomacells,whichnaturallyexpressLin28.21Mouse andhumanLin28proteinsdiffer inonly5of207aminoacidresidues.
Lin28:TAP complexes were purified bybinding to Protein A beads followed byelution with theTEV site‑specific protease.Initial attempts to identify proteins inthese complexes by purification followedby mass‑spectrometry suggested that manyproteins were co-purified in the procedure,including PABP, eIF4E, YB‑1, IMP‑3 andalpha‑ and beta‑tubulin (data not shown).Manyoftheseproteinsmayco-purifybecausetheyareassociatedwiththesamecomplexes,whereas others may represent backgroundin the procedure. To reduce background,Lin28‑containing complexes bound tobeads were washed with 3M urea beforeelution.UreawasheshavebeensuggestedtoincreasethestringencyofRNPaffinitypuri-fications.31Thepresenceofspecificproteinsco-purifying with Lin28 was then assessedby immunoblot. Abundant proteins suchastubulinandactinwerenotpresentinthe
complexes, indicating that the washes indeed reduced background(datanotshown).Incontrast,wereadilydetectedthegeneralmRNAbinding protein, poly(A) binding protein (PABP), which co-puri-fiedtoagreaterdegreewithLin28thanwiththecontrolGFP:TAP(Fig.2A, toppanel).Thisobservation indicates thatPABP specifi-callyco-purifieswithLin28.Reprobingtheblotswithanti‑TAPtagantisera showed that the eluates contained similar amounts of thetaggedproteins(Fig.2A,bottompanel).
PABP stably binds translationally‑competent mRNAs in thecytoplasm.32 Itspresence inLin28:TAPcomplexesmaybebecauseit interactswithLin28directlyorthroughtheirmutualassociationwith mRNAs. To test this, Lin‑28‑containing complexes boundto beads were treated with RNaseA, washed, and then eluted. Incontrast to untreated complexes, RNase‑treated complexes did notcontain PABP (Fig. 2A, top panel), indicating that its co-purifica-tionwithLin28isduetomutualbindingofRNA.Therefore,Lin28appearstobepresentinproteincomplexescontainingmRNA.
Lin28 is in polyribosomes. Actively translated mRNAs areassociated with ribosomes in the form of polyribosomes (poly-somes).ToassesswhetherLin28maybeassociatedwithtranslatingmRNAs, sucrose gradient fractionation was performed to assessLin28’s distribution in polysomes. P19 cell lysates were applied to15–45% sucrose gradients and fractionated by ultracentrifugation.SamplesofeachfractionwereseparatedbySDSpolyacrylamidegel
Figure 2. Lin28 presence in mRNPs. (A) Immunoblots of TAP-purified complexes using anti-PABP and anti-TAP antisera. P19 cells were transfected with GFP:TAP or Lin28:TAP. Lin28:TAP complexes were treated with RNaseA prior to elution, as indicated. (B) Sucrose gradient fractionation of P19 cells. Absorbance at 254 nm was recorded as the gradient was fractionated. The direction of sedimentation is shown left-to-right. Fractions of the gradient were probed by immunoblot with anti-Lin28 antisera. The location of polysomes is indicated. Parallel gradients were run with and without EDTA, which disrupts polysomes.

www.landesbioscience.com RNABiology 19
Lin28inmRNPsandP-bodies
electrophoresis,blottedandprobedwith anti‑Lin28antibody (Fig.2B, left panel).The amountof absorbance at 254nmdeterminedthelocationofpolysomalandsub‑polysomalfractions.Endogenous
Lin28proteinwasobservedthroughoutthe gradient, including the polysomalfractions. A significant portion of thetotalLin28protein,however,ispresentinsub‑polysomal fractions.Toconfirmthat Lin28 is indeed associated withpolysomes, lysates were run on gradi-ents containing EDTA to disrupt theribosomes. The RNA profile of thesegradientsdemonstrated thatpolysomeswere disrupted as expected (Fig. 2B,right panel). Lin28 protein remainednearthetopofthegradient,suggestingitssedimentationisdependentonasso-ciation with ribosomes. Therefore, atleast a portion of the total Lin28 inthe cell appears to be associated withpolysomes. A significant fraction mayalsobe freeor incomplexeswithnon-translating mRNAs. The distributionof many mRNA‑binding proteins inpolysomal and sub‑polysomal fractionshavebeensimilarlyexamined.33,34Theamount of protein in polysomes maydependontheabundanceoftheproteinrelativetoitsnumberoftargets,aswellastheprotein’sfunction.
The CSD and CCHC domains coop‑erate to associate Lin28 with mRNPs.The two conserved RNA‑bindingdomains of Lin28 may have distinctrolesintheprotein’sfunction.Toaddresswhether the association of Lin28 withRNA in mRNPs is due to the activityof one or both of these domains, twodeletion forms were generated: a dele-tion from the C‑terminus to the startof the CCHC domain (CSD‑only),andadeletionfromtheN‑terminusto
theendoftheCSDdomain(CCHC‑only;Fig.1B).ThesedeletionformswerefusedtoaGFP‑TAPfusiontoallowbothaffinitypuri-ficationandmonitoringofexpressionandlocalization,andassayedforassociationwithPABPbyaffinitypurification.PABPco-purifiedwiththeCSD‑onlyform,althoughsignificantlylessefficientlythandidthefull‑lengthform(Fig.3A,toppanel).Inthesestablytrans-fectedcells,thedeletionformswereexpressedandpurifiedatlevelssimilartothoseofthewildtypefusion(Fig.3A,bottompanel).TheCCHC‑onlyformco-purifiedPABPatlevelsequivalenttotheGFPcontrol(Fig.3A,toppanel).TheseobservationssuggestthatthetwoconserveddomainscooperatetoassociateLin28withmRNPs.
To assess whether each domain is required to associate Lin28with polysomes, the distribution of each deletion form in sucrosegradientswasdetermined(Fig.3B).Bothformsarefoundinheavyfractionsofthegradients,co-sedimentingwithpolysomes.However,thepolysomeassociationoftheCCHC‑onlyformwassignificantlyless than that of the full‑length protein. EDTA treatment of thelysatesshowsthattheCCHC‑onlyformrequiresintactribosomestosediment.However,theCSD‑onlyformsedimentsinheavyfractionsevenafterEDTAtreatment,suggestingthatitisinlargecomplexesthat are not ribosomes. It is possible that this truncated form of
Figure 3. Analysis of Lin28 deletion forms. (A) Immunoblots of TAP-purified complexes using anti-PABP and anti-TAP antisera. P19 cells were transfected with GFP:TAP, Lin28:TAP, CSD-only:GFP:TAP, or CCHC-only:GFP:TAP. (B) Sucrose gradient fractionation of P19 cells transfected with deletion forms. Absorbance at 254 nm was recorded as the gradient was fractionated. The direction of sedimentation is shown left-to-right. Fractions of the gradient were probed by immunoblot with anti-TAP antisera. The location of polysomes is indicated. Parallel gradients were run with and without EDTA, which disrupts polysomes.
Table 1 P‑bodylocalizationofLin28,YB‑1andLin28 mutantforms
Protein1 % cells with Foci (n)1 Lin28 (IF) 65 (100)2 Lin28 (GFP) 72 (104)3 YB-1 (GFP) 0 (>100)4 CSD only 8.9 (112)5 CCHC only 9.6 (125)6 CSD wt CCHC mut 1.8 (109)7 CSD mut CCHC wt 3.6 (111)8 CSD mut CCHC mut 0 (>200)
1Lin28(IF), endogenous Lin28 detected by indirect immunofluoresence. All other proteins are GFP-tagged and expressed from transfected plasmids.
Lin28inmRNPsandP-bodies
20 RNABiology 2007;Vol.4Issue1
Lin28 aggregates due to a possibly altered structure. Alternatively,theseobservationssuggestthattheCCHCdomainhelpskeepLin28outofsuchcomplexes.
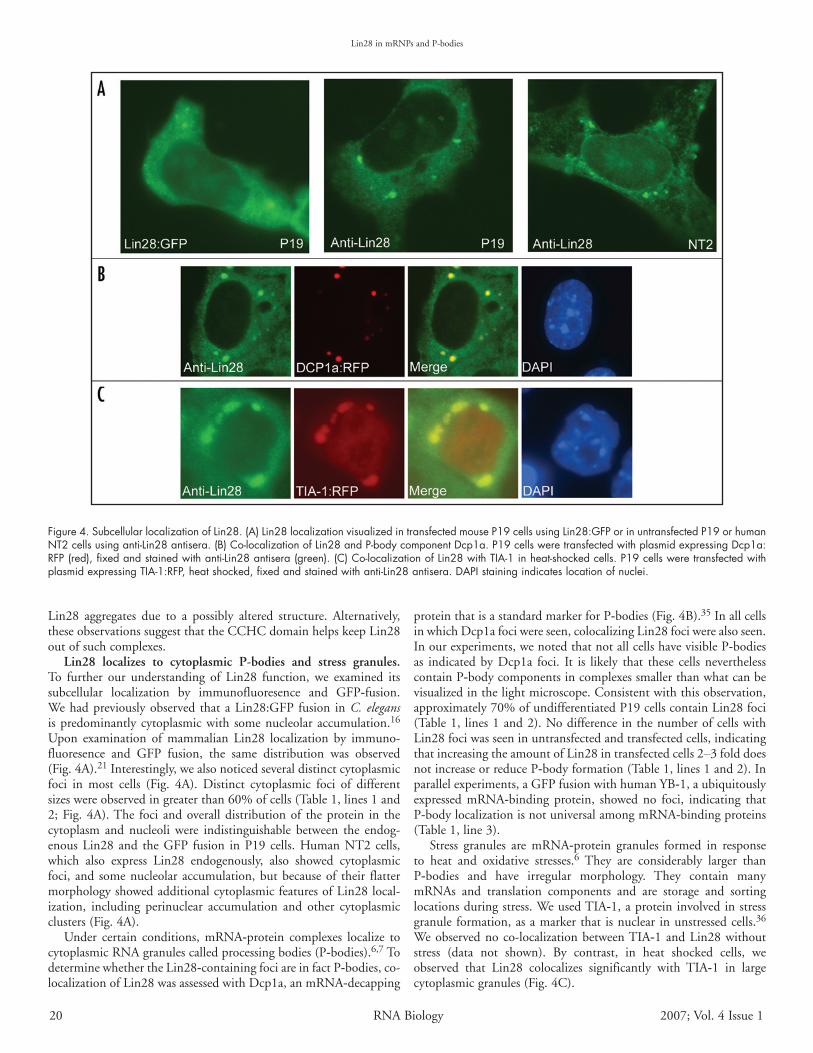
Lin28 localizes to cytoplasmic P‑bodies and stress granules. To further our understanding of Lin28 function, we examined itssubcellular localization by immunofluoresence and GFP‑fusion.Wehadpreviouslyobserved thataLin28:GFPfusion inC. elegansispredominantlycytoplasmicwith somenucleolaraccumulation.16Upon examination of mammalian Lin28 localization by immuno-fluoresence and GFP fusion, the same distribution was observed(Fig.4A).21Interestingly,wealsonoticedseveraldistinctcytoplasmicfoci in most cells (Fig. 4A). Distinct cytoplasmic foci of differentsizeswereobservedingreaterthan60%ofcells(Table1,lines1and2;Fig.4A).The foci andoveralldistributionof theprotein in thecytoplasm and nucleoli were indistinguishable between the endog-enousLin28 and theGFP fusion inP19 cells.HumanNT2cells,which also express Lin28 endogenously, also showed cytoplasmicfoci, and somenucleolar accumulation,butbecauseof their flattermorphologyshowedadditionalcytoplasmicfeaturesofLin28local-ization, including perinuclear accumulation and other cytoplasmicclusters(Fig.4A).
Under certain conditions, mRNA‑protein complexes localize tocytoplasmicRNAgranulescalledprocessingbodies(P‑bodies).6,7TodeterminewhethertheLin28‑containingfociareinfactP‑bodies,co-localizationofLin28wasassessedwithDcp1a,anmRNA‑decapping
proteinthatisastandardmarkerforP‑bodies(Fig.4B).35InallcellsinwhichDcp1afociwereseen,colocalizingLin28fociwerealsoseen.Inourexperiments,wenotedthatnotallcellshavevisibleP‑bodiesas indicatedbyDcp1a foci. It is likely that these cellsneverthelesscontainP‑bodycomponentsincomplexessmallerthanwhatcanbevisualizedinthelightmicroscope.Consistentwiththisobservation,approximately70%ofundifferentiatedP19cellscontainLin28foci(Table1, lines1and2).NodifferenceinthenumberofcellswithLin28fociwasseeninuntransfectedandtransfectedcells,indicatingthatincreasingtheamountofLin28intransfectedcells2–3folddoesnotincreaseorreduceP‑bodyformation(Table1,lines1and2).Inparallelexperiments,aGFPfusionwithhumanYB‑1,aubiquitouslyexpressed mRNA‑binding protein, showed no foci, indicating thatP‑bodylocalizationisnotuniversalamongmRNA‑bindingproteins(Table1,line3).
Stress granules are mRNA‑protein granules formed in responseto heat and oxidative stresses.6 They are considerably larger thanP‑bodies and have irregular morphology. They contain manymRNAs and translation components and are storage and sortinglocationsduringstress.WeusedTIA‑1,aproteininvolvedinstressgranuleformation,asamarkerthat isnuclear inunstressedcells.36Weobservednoco-localizationbetweenTIA‑1andLin28withoutstress (data not shown). By contrast, in heat shocked cells, weobserved that Lin28 colocalizes significantly with TIA‑1 in largecytoplasmicgranules(Fig.4C).
Figure 4. Subcellular localization of Lin28. (A) Lin28 localization visualized in transfected mouse P19 cells using Lin28:GFP or in untransfected P19 or human NT2 cells using anti-Lin28 antisera. (B) Co-localization of Lin28 and P-body component Dcp1a. P19 cells were transfected with plasmid expressing Dcp1a:RFP (red), fixed and stained with anti-Lin28 antisera (green). (C) Co-localization of Lin28 with TIA-1 in heat-shocked cells. P19 cells were transfected with plasmid expressing TIA-1:RFP, heat shocked, fixed and stained with anti-Lin28 antisera. DAPI staining indicates location of nuclei.

www.landesbioscience.com RNABiology 21
Lin28inmRNPsandP-bodies
To determine whether the RNA‑binding domains of Lin28 arerequired for localization to P‑bodies, we examined cells expressingGFPfusionstothetwodeletionformsofLin28,CSD‑onlywhichlackedtheCCHCdomainandC‑terminus,andCCHC‑onlywhichlacked the N‑terminus and CSD (Fig. 1). Although both dele-tion forms showed accumulation in foci, substantially fewer cellsshowed foci than seen with full‑length Lin28 (Table 1, lines 2, 4and 5). These observations suggest that efficient accumulation inP‑bodies requires the entire protein, possibly the activity of bothRNA‑bindingdomains.
To more specifically test the requirement of each conservedRNA‑binding domain, we generated three point‑mutant forms ofLin28: a CSD mutant, a CCHC mutant and a mutant in bothdomains (Fig. 1). A CSD mutant consisting of full‑length Lin28withthreearomaticaminoacidsoftheRNAbindingmotifsoftheCSDchangedtoalanines;aCCHCmutantconsistingoffull‑lengthLin28 with the two histidines of the CCHC motifs changed toalanines;andatwo‑domainmutantcontainingallfivepointmuta-tions(Fig.1).Similarmutationshavebeenmadepreviouslyinother
proteins and been found toaffecttheRNA‑bindingabilityof each of these domains.37,38EachmutantformwasfusedtoGFP:TAPandexpressedinP19cells. The mutants with onlyonewildtypedomainproducedsomefoci,butsubstantiallylessefficientlythanwildtypeLin28(Table 1, lines 2, 6 and 7;Fig.5A).TheCSDmutantfocididcolocalizewiththeDcp1a:RFP marker, indicating theyareP‑bodies.However,theveryrarefociformedbytheCCHCpoint‑mutant did not overlapwiththeDcp1amarker(Table1;datanot shown), therefore wedo not know the nature ofthese foci. Therefore, activityoftheCCHCdomainappearsto be required for localizationtoP‑bodies,whereas theCSDmayaidinefficientlocalizationto these granules. In contrast,bothmutantformslocalizedtostress granules in stressed cells(Fig.5B).
Surprisingly, the two‑do-mainmutantlocalizedentirelyto the nucleus (Fig. 6). Thislocalization is in strikingcontrast to that of the wild-type Lin28:GFP fusion whichis predominantly cytoplasmic(cf. Fig. 4), despite only fiveaminoaciddifferencesbetweenthewildtypeandmutantforms(Fig.1).Becauseof thisexclu-sively nuclear localization,no co-localization with cyto-
plasmicgranuleswasobserved(Table1,line8).Thetwosingle‑domainmutantswerealsoassayedfortheirability
to copurify with PABP. Neither mutant showed association withPABPtothesameextentasthewildtypeprotein(Fig.7).Therefore,localizationtoP‑bodiesappears tocorrelatewithefficientor stableincorporationintomRNPs.
Predominantly cytoplasmic Lin28 appears to shuttle through the nucleus. The nuclear localization of the two‑domain mutantsuggests thatLin28containsanuclear targeting sequence.Proteinsas small as 40 kD may readily diffuse through nuclear pores.39Although Lin28:GFP is approximately 50 kD, we observed thatGFP and GFP:TAP alone show some nuclear accumulation (datanot shown).Therefore, a larger fluorescentmarker,GFP:luciferase,which isgreater than100kD,wasused to test thepossibility thatLin28isactivelylocalizedtothenucleus.AfusionofTAP,whichissimilarinsizetoLin28,toGFP:luciferasewaspredominantlycyto-plasmic(Fig.8A).Bycontrast,theSV40nuclearlocalizationsignalcausedthefusiontobeentirelynuclear(Fig.8B).Fusingthetwo‑do-main mutant to GFP:luciferase also caused nuclear accumulation
Figure 5. Subcellular localization of Lin28 point mutants. (A) Subcellular localization of Lin28, CSD mutant (CSD mut), and CCHC mutant (CCHC mut) with Dcp1a. P19 cells were co-transfected with plasmids expressing wildtype Lin28 fused to GFP or mutant Lin28 fused to GFP:TAP (green) and Dcp1a:RFP (red), and fixed. (B) Subcellular localization of the same proteins with TIA-1. P19 cells were co-transfected with the same Lin28 plasmids and TIA-1:RFP (red), heat shocked and fixed. DAPI staining indicates location of nuclei.

Lin28inmRNPsandP-bodies
22 RNABiology 2007;Vol.4Issue1
(Fig.8C).TheseobservationsindicatethatLin28isactivelyimportedinto the nucleus. Because the single‑domain mutants localize tothe cytoplasm (Fig. 5), either of its RNA‑binding domains mustbe sufficient for a steady‑state cytoplasmic distribution.Treatmentof cells with leptomycin B, which inhibits the nuclear exporterCRM‑1, did not cause nuclear retention of wildtype Lin28 (datanot shown), suggesting that Lin28 may be actively exported via adifferentexporter,possiblythemajormRNPexporterTAP/NXF1.39Attemptstomapashortcontiguousnuclearlocalizationsignalwerenotsuccessful.Forexample,aregionbetweentheCSDandCCHCdomaincontainingbasicaminoacidresidues(thebasiclinkerregion,BLR;Fig.1)isnotcapableofcausingnuclearaccumulationofGFP:luciferase(Fig.8D).NeitherthemutantCSD‑onlynorthemutantCCHC‑onlydeletionformslocalizetothenucleus(Figs.8EandF),suggesting that nuclear localization requires sequences at multipleplacesintheprimarysequence.
discussionHerewepresentevidenceforamodelforLin28activityindevel-
opmentalregulation(Fig.9).Inthismodel,Lin28entersthenucleusbyanactiveimportmechanismandassemblesintomRNPsbyvirtueofitsRNA‑bindingdomains.ItisthenexportedtothecytoplasmaspartofthemRNPs,andremainsboundasthemRNAistranslatedor sequestered in cytoplasmic RNA granules.This model suggests,asoutlinedbelow,thatLin28isaregulatorofmRNAtranslationorstabilityinundifferentiatedcells,andtherebycontrolsthesynthesisofdevelopmentalfactorsatthelevelofthemRNA.
P‑bodiesarespecializedsitesofmRNAregulation.Originally,theyweredefinedbythefocallocalizationofseveralproteinsinvolvedinde‑capping and degradation of mRNAs, such as Dcp1a.40 Morerecently, theyhavebeenfoundtoalsocontainproteins involved inmiRNA‑andsiRNA‑mediatedregulation,includingmembersoftheArgonautefamily.41,42IthasbeenshownthatsmallRNAscausetherelocalizationoftheirmRNAtargetsfromthetranslationalmachineryto P‑bodies.42 P‑bodies do not contain ribosomes or several otherfactors essential to translation, such asPABP.6,7,35Therefore, thesegranules are intimately involved in the translational repression anddegradation of mRNAs. Whereas a number of factors have beenshowntolocalizetoP‑bodies,currentlyfewmRNAbindingproteinsthought tohave specific targetshaveyetbeenshowntobeP‑bodycomponents.7 Two developmentally important regulatory proteinsknown to be in P‑bodies are CPEB and Staufen.6,43,44 Lin28 isunlikely to be a general factor in P‑body function or formationbecauseitisexpressedinrelativelyfewcellsintheanimal.16,21,24
Unlike P‑bodies, stress granules contain a number of factorsdirectlyinvolvedintranslation,includingstalledribosomalsubunit
complexes.6,45,46 Although P‑bodies are visible in most cells (60–70%;Table1),stressgranulesappearonlyuponinductionbyheat,oxidation,orothercellularstresses.StressgranulesaresubstantiallylargerthanP‑bodiesandactastriagelocationsfromwhichmRNAsmayreturntotranslationortothemRNAdegradationmachinery.6A number of proteins associated with translating mRNAs can befound in stressgranulesduringcellular stress, includingPABPandspecific translational regulators such as HuR and Staufen.6,36,47,48ThepresenceofLin28instressgranuleslikelyreflectsmerelyitsasso-ciationwithmRNAsandthetemporaryrelocationofmanymRNAsandassociatedproteinstostressgranules.
Lin28 is not only in cytoplasmic granules but is found withtranslatingmRNAsaswell,asindicatedbyitsassociationwithPABPviaRNAanditspresenceinpolysomes.BecausePABPisnotfoundin P‑bodies, Lin28 must be present in at least two populations ofmRNPs, those thatare translatedand those thatare translationallyrepressed or destined for degradation. Therefore, the significanceofour findings is thatLin28 is likely toregulate the translationorstabilityofmRNAsbywayoftheirlocalizationtoP‑bodies.mRNAsundergoing translational regulation have been shown to movebetween polysomes and P‑bodies.49Two other conserved develop-mental regulatoryproteins thatarepresent inP‑bodies,CPEBandStaufen,arealsoboundtoactivelytranslatedmRNAs.50,51CPEBisaspecificRNAbindingproteinthatactsasatranslationalactivator,causing polyadenylation of specific messages in the cytoplasm.10Staufenisaspecificdouble‑strandedRNAbindingproteininvolvedinmRNAtransportandlocalizationandtranslationalregulation.52‑54WhetherLin28causesspecificmRNAstoleavepolysomesandgotoP‑bodiesorvisaversaisnotyetclear.OurfindingthatP‑bodylocal-izationofLin28requiresanintactCCHCdomainimpliesthatthislocalization depends on RNA‑binding by this domain. Therefore,Lin28doesnotappeartolocalizetoP‑bodiesonitsown,butdoessopassivelywhenboundtomRNAsthataredirectedtherebyotherfactors.IthasbeensuggestedthatthemRNA‑bindingproteinHuRis partly responsible for the translational activation of an mRNArepressedbymicroRNAsbyrecruitingthemawayfromP‑bodies.14WespeculatethatLin28mayactsimilarlytoenhancetranslationof
Figure 6. Nuclear localization of Lin28 two-domain mutant. P19 cells were co-transfected with Lin28 mutant in both RNA-binding domains (CSD mut CCHC mut) fused to GFP:TAP (green) and Dcp1a:RFP (red), and fixed. DAPI staining indicates location of nuclei.
Figure 7. Failure of Lin28 mutants to form mRNPs. Immunoblots of TAP-purified complexes using anti-PABP and anti-TAP antisera. P19 cells were transfected with GFP:TAP (GFP), Lin28:TAP (Lin28), CCHC mutant fused to GFP:TAP (CCHC mut:GFP), or CSD mutant fused to GFP:TAP (CSD mut:GFP). Mutant forms are larger due to the addition of GFP.

www.landesbioscience.com RNABiology 23
Lin28inmRNPsandP-bodies
specificmessages.Therefore,thepresenceofLin28inP‑bodieseitherreflectsLin28rescuingspecificmRNAsfromtranslationalrepressionor reveals its failure to overcome repressionof particularmessages,orboth.Alternatively,Lin28mayactdifferentlydependingon thespecific targets and other interacting factors, and may help repressspecific mRNAs. In light of these possibilities, we have depictedtwo‑way movement of Lin28 between P‑bodies and translation(Fig.9).
In C. elegans, genetic and molecular evidence indicate thatlin‑14 is positively regulated by lin‑28, suggesting that the Lin28proteinmayacttoenhancethetranslationorstabilityofthelin‑14mRNA.17,20,55 lin‑14 is translationally repressedbythemicroRNAencoded by lin‑4.56 lin‑4 and lin‑28 act simultaneously on lin‑14duringakeytransitioninlarvaldevelopment.20Untilnow,therehasbeennomolecularunderstandingofhowtheLin28proteinmayactinthisregulation.OurfindingssupportahypothesiswherebyLin28opposes the microRNA‑mediated repression of specific mRNAs,perhapsuntilathresholdofmicroRNAactivityisreached,triggeringa developmental switch.15 We have not observed foci of Lin28 inC. eleganslarvaelikeweseeinmammaliancells.16However,theC. elegans homologs of P‑body components Argonaute and GW182donot show fociunlessover‑expressed.57Therefore, just asnotallmammalian cells have visible P‑bodies, C. elegans larvae may haveP‑bodycomponentsingranulessmallerthancanbereadilyseeninthelightmicrocope.
Whereas both of Lin28’s RNA‑binding domains are requiredforefficientco-purificationofPABP,eitherdomainissufficientforstress granule localization and sedimentation in sucrose gradients.ThisindicateseachdomainbindsRNAinvivotosomedegree,butthat stable incorporation intomRNPs requiresbothdomains.Ourfinding that theCCHCdomain,butnot theCSD, is required forthelocalizationofLin28toP‑bodiesimpliesthatthisdomainmayconfertarget‑specificitytotheprotein.TheCCHCdomainisverysimilar to retroviral nucleocapsid proteins that interact specificallywith structures in the viral RNA genome.28,58‑60 The retroviralCCHCproteincanalsoactasachaperone,aidingchangesinRNAbasepairing.61Similarly,theCSDhasbeenshowntohavechaperoneactivity.62,63 However, the CSD is generally thought to have littlespecificity forparticularRNAsand ispresent inproteins thatbindmanymRNAs.26,29Severalproteins fromplants also containCSDand CCHC domains and may carry out similar activities duringplantdevelopment.64,65
An unexpected finding was that the full-length Lin28 proteinwith mutations in both RNA binding domains was localizedexclusively to thenucleus.By fusing themutatedLin28 to a largecytoplasmic reporter, we showed that import into the nucleus isactive,ratherthanduetodiffusion.However,nuclearaccumulationofthismutantsuggeststhatLin28’snuclearexportisdependentonitsabilitytobindmRNA.Therefore,thesteady-statelocalizationofwildtypeLin28tothecytoplasmmaybetheresultofmoreefficient
Figure 8. Nuclear localization of Lin28. P19 cells were transfected with plasmids expressing fusions to GFP:luciferase. (A) TAP, Tandem Affinity Purification tag. (B) SV40, nuclear localization sequence from SV40 Large T antigen. (C) CSD mut CCHC mut, two-domain mutant of Lin28. (D) BLR, basic linker region indicated in Figure 1. (E), CSD-only form of Lin28 with mutations in CSD indicated in Figure 1. (F) CCHC-only form Lin28 with mutations indicated in Figure 1. DAPI staining indicates location of nuclei.

Lin28inmRNPsandP-bodies
24 RNABiology 2007;Vol.4Issue1
nuclearexportrelativetoimport.Stau2,ahomologofStaufenandalsoapredominantlycytoplasmicprotein,alsoshuttlesthroughthenucleus.66,67NuclearaccumulationofStau2wassimilarlyobservedwhen it was mutated in an RNA-binding domain.67 Interestingly,YB-1, another CSD-containing protein, was found to localize tothe nucleus and lose its biological activity when mutated to losethe RNA-binding activity of the CSD.68 Although it remains tobe determined whether nuclear localization is required for Lin28function,itislikelythatLin28entersthenucleustobindtospecifictargetsbeforeexporttothecytoplasmwheretheyareacteduponbymicroRNAsandotherregulatoryproteins.
We set out to address how Lin28 acts in the cell to regulategeneexpression inundifferentiatedcells.Ourdatasuggestamodelin which Lin28 specifically binds certain target mRNAs that canlocalizetoP-bodies.Thesetargetsmayvaryfromtissuetotissuebutmay contain common binding sites for Lin28.The next challengeis to identify these targets. A recent study of the role of Lin28 inmuscledifferentiationindicatesthatitindeedbindstoandenhancesthe translation of at least one specific mRNA target in this celltype.69 Because of the strong requirement for the CCHC domainfor forming mRNPs and localizing to P-bodies, this domain ofLin28mayprovidetargetspecificity.WhatisthefateofthetargetsonceLin28isboundalsoremainstobedetermined.AnintriguingpossibilityisthatLin28opposestheactionofmicroRNAsonthosetargets. RNA-binding proteins such as Lin28 may be in dynamictensionwithmicroRNAstoensurestrictregulationofgeneexpres-sionatthetranslationallevel.
References 1. In:HersheyJWB,MathewsMB,SonenbergN,eds.TranslationalControlinBiologyand
Medicine.Plainview,NY:ColdSpringHarborLaboratoryPress,2007. 2. Sanchez-Diaz P, Penalva LO. Post-transcription meets post-genomic: The saga of RNA
bindingproteinsinanewera.RNABiol2006;3. 3. AmbrosV.ThefunctionsofanimalmicroRNAs.Nature2004;431:350-5.
4. MooreMJ.Frombirthtodeath:ThecomplexlivesofeukaryoticmRNAs.Science2005;309:1514-8.
5. ColeCN,ScarcelliJJ.TransportofmessengerRNAfromthenucleustothecytoplasm.CurrOpinCellBiol2006;18:299-306.
6. AndersonP,KedershaN.RNAgranules.JCellBiol2006;172:803-8. 7. EulalioA,Behm-AnsmantI,IzaurraldeE.Pbodies:Atthecrossroadsofpost-transcriptional
pathways.NatRevMolCellBiol2007;8:9-22. 8. Graveley BR. Sex, AGility, and the regulation of alternative splicing. Cell 2002;
109:409-12. 9. StJohnstonD.Movingmessages:Theintracellular localizationofmRNAs.NatRevMol
CellBiol2005;6:363-75. 10. MendezR,RichterJD.TranslationalcontrolbyCPEB:Ameanstotheend.NatRevMol
CellBiol2001;2:521-9. 11. Lee MH, Schedl T. RNA-binding proteins. In: Community TCeR, ed. WormBook:
WormBook,2006. 12. WilhelmJE,SmibertCA.MechanismsoftranslationalregulationinDrosophila.Biologyof
thecell/undertheauspicesoftheEuropeanCellBiologyOrganization2005;97:235-52. 13. GeorgeAD,TenenbaumSA.MicroRNAmodulationofRNA-bindingproteinregulatory
elements.RNABiol2006;3. 14. Bhattacharyya SN, Habermacher R, Martine U, Closs EI, Filipowicz W. Relief of
microRNA-mediatedtranslationalrepressioninhumancellssubjectedtostress.Cell2006;125:1111-24.
15. MossEG.Heterochronicgenesandthenatureofdevelopmentaltime.CurrBiol2007,(Inpress).
16. MossEG,LeeRC,AmbrosV.ThecoldshockdomainproteinLIN-28controlsdevelop-mentaltiminginC. elegansandisregulatedbythelin-4RNA.Cell1997;88:637-46.
17. Seggerson K, Tang L, Moss EG. Two genetic circuits repress the Caenorhabditis elegansheterochronicgenelin‑28aftertranslationinitiation.DevBiol2002;243:215-25.
18. AmbrosV,HorvitzHR.Heterochronicmutants of thenematodeCaenorhabditis elegans.Science1984;226:409-16.
19. AbbottAL,Alvarez-SaavedraE,MiskaEA,LauNC,BartelDP,HorvitzHR,AmbrosV.The let-7MicroRNAfamilymembersmir-48,mir-84,andmir-241functiontogethertoregulatedevelopmentaltiminginCaenorhabditis elegans.DevCell2005;9:403-14.
20. ArasuP,WightmanB,RuvkunG.Temporalregulationoflin‑14bytheantagonisticactionoftwootherheterochronicgenes,lin‑4andlin‑28.GenesDev1991;5:1825-33.
21. MossEG,TangL.ConservationoftheheterochronicregulatorLin-28,itsdevelopmentalexpressionandmicroRNAcomplementarysites.DevBiol2003;258:432-42.
22. SempereLF,FreemantleS,Pitha-Rowe I,MossE,DmitrovskyE,AmbrosV.ExpressionprofilingofmammalianmicroRNAsuncoversasubsetofbrain-expressedmicroRNAswithpossiblerolesinmurineandhumanneuronaldifferentiation.GenomeBiol2004;5:R13.
23. WuL,BelascoJG.Micro-RNAregulationofthemammalianlin‑28geneduringneuronaldifferentiationofembryonalcarcinomacells.MolCellBiol2005;25:9198-208.
24. YangDH,MossEG.Temporally regulated expressionofLin‑28 indiverse tissues of thedevelopingmouse.GeneExprPatterns2003;3:719-26.
25. RichardsM,TanSP,TanJH,ChanWK,BongsoA.ThetranscriptomeprofileofhumanembryonicstemcellsasdefinedbySAGE.StemCells(Dayton,Ohio)2004;22:51-64.
26. Graumann PL, Marahiel MA. A superfamily of proteins that contain the cold-shockdomain.TrendsBiochemSci1998;23:286-90.
27. ManivalX,Ghisolfi-NietoL,JosephG,BouvetP,ErardM.RNA-bindingstrategiescom-montocold-shockdomain-andRNArecognitionmotif-containingproteins.NucleicAcidsRes2001;29:2223-33.
28. Ramboarina S, Druillennec S, Morellet N, Bouaziz S, Roques BP. Target specificity ofhumanimmunodeficiencyvirustype1NCp7requiresanintactconformationofitsCCHCN-terminalzincfinger.JVirol2004;78:6682-7.
29. YuJ,HechtNB,SchultzRM.RequirementforRNA-bindingactivityofMSY2forcytoplas-miclocalizationandretentioninmouseoocytes.DevBiol2003;255:249-62.
30. PuigO,CasparyF,RigautG,RutzB,BouveretE,Bragado-NilssonE,WilmM,SeraphinB.Thetandemaffinitypurification(TAP)method:Ageneralprocedureofproteincomplexpurification.Methods(SanDiego,Calif )2001;24:218-29.
31. KeeneJD,KomisarowJM,FriedersdorfMB.RIP-Chip:TheisolationandidentificationofmRNAs, microRNAs and protein components of ribonucleoprotein complexes from cellextracts.NatureProtocols2006;1:302-7.
32. MangusDA,EvansMC,JacobsonA.Poly(A)-bindingproteins:Multifunctionalscaffoldsforthepost-transcriptionalcontrolofgeneexpression.GenomeBiol2003;4:223.
33. DavidovicL,HuotME,KhandjianEW.Lostonce,theFragileXMentalRetardationpro-teinisnowbackontobrainpolyribosomes.RNABiol2005;2:1-3.
34. YangJ,MedvedevS,ReddiPP,SchultzRM,HechtNB.TheDNA/RNA-bindingproteinMSY2marksspecifictranscriptsforcytoplasmicstorageinmousemalegermcells.ProcNatlAcadSciUSA2005;102:1513-8.
35. KedershaN,StoecklinG,AyodeleM,YaconoP,Lykke-AndersenJ,FritzlerMJ,ScheunerD,KaufmanRJ,GolanDE,AndersonP.StressgranulesandprocessingbodiesaredynamicallylinkedsitesofmRNPremodeling.JCellBiol2005;169:871-84.
36. Kedersha NL, Gupta M, LiW, Miller I, Anderson P. RNA-binding proteinsTIA-1 andTIARlinkthephosphorylationofeIF-2alphatotheassemblyofmammalianstressgran-ules.JCellBiol1999;147:1431-42.
37. DorfmanT, Luban J, Goff SP, Haseltine WA, Gottlinger HG. Mapping of functionallyimportantresiduesofacysteine-histidineboxinthehumanimmunodeficiencyvirustype1nucleocapsidprotein.JVirol1993;67:6159-69.
Figure 9. Model for Lin28 activity in undifferentiated cells. Lin28 enters the nucleus by an active mechanism, is incorporated into mRNPs by virtue of its RNA-binding domains, and is rapidly exported to the cytoplasm. The Lin28-containing mRNPs may go to P-bodies (PB) or ribosomes, or in stressed cells, to stress granules (SG). Lin28 likely affects the translation of the specific mRNAs to which it is bound by either recruiting them to P-bodies, or alterna-tively, by rescuing them from P-bodies, and stimulating their translation.

www.landesbioscience.com RNABiology 25
Lin28inmRNPsandP-bodies
38. SchroderK,GraumannP,SchnuchelA,HolakTA,MarahielMA.Mutationalanalysisoftheputativenucleicacid-bindingsurfaceofthecold-shockdomain,CspB,revealedanessentialroleofaromaticandbasicresiduesinbindingofsingle-strandedDNAcontainingtheY‑boxmotif.MolMicrobiol1995;16:699-708.
39. CullenBR.NuclearRNAexport.JCellSci2003;116:587-97. 40. ShethU,ParkerR.DecappinganddecayofmessengerRNAoccurincytoplasmicprocess-
ingbodies.Science2003;300:805-8. 41. SenGL,BlauHM.Argonaute2/RISCresidesinsitesofmammalianmRNAdecayknown
ascytoplasmicbodies.NatCellBiol2005;7:633-6. 42. LiuJ,Valencia-SanchezMA,HannonGJ,ParkerR.MicroRNA-dependentlocalizationof
targetedmRNAstomammalianP-bodies.NatCellBiol2005;7:719-23. 43. WilczynskaA,AigueperseC,KressM,DautryF,WeilD.ThetranslationalregulatorCPEB1
providesalinkbetweendcp1bodiesandstressgranules.JCellSci2005;118:981-92. 44. Barbee SA, Estes PS, Cziko AM, Hillebrand J, Luedeman RA, Coller JM, Johnson N,
HowlettIC,GengC,UedaR,BrandAH,NewburySF,WilhelmJE,LevineRB,NakamuraA,ParkerR,RamaswamiM.Staufen-andFMRP-containingneuronalRNPsarestructur-allyandfunctionallyrelatedtosomaticPbodies.Neuron2006;52:997-1009.
45. KimballSR,HoretskyRL,RonD,JeffersonLS,HardingHP.Mammalianstressgranulesrepresentsitesofaccumulationofstalledtranslationinitiationcomplexes.AmericanJournalofPhysiology2003;284:C273-84.
46. KedershaN,AndersonP.Stressgranules:SitesofmRNAtriagethatregulatemRNAstabilityandtranslatability.BiochemicalSocietyTransactions2002;30:963-9.
47. GallouziIE,BrennanCM,StenbergMG,SwansonMS,EversoleA,MaizelsN,SteitzJA.HuRbindingtocytoplasmicmRNAisperturbedbyheatshock.ProcNatlAcadSciUSA2000;97:3073-8.
48. ThomasMG,MartinezTosarLJ,LoschiM,PasquiniJM,CorrealeJ,KindlerS,BoccaccioGL.StaufenrecruitmentintostressgranulesdoesnotaffectearlymRNAtransportinoligo-dendrocytes.MolBiolCell2005;16:405-20.
49. BrenguesM,TeixeiraD,ParkerR.Movementof eukaryoticmRNAsbetweenpolysomesandcytoplasmicprocessingbodies.Science2005;310:486-9.
50. MarionRM,FortesP,BelosoA,DottiC,OrtinJ.AhumansequencehomologueofStaufenis an RNA-binding protein that is associated with polysomes and localizes to the roughendoplasmicreticulum.MolCellBiol1999;19:2212-9.
51. GebauerF,RichterJD.Mousecytoplasmicpolyadenylylationelementbindingprotein:Anevolutionarilyconservedproteinthatinteractswiththecytoplasmicpolyadenylylationele-mentsofc-mosmRNA.ProcNatlAcadSciUSA1996;93:14602-7.
52. Dugre-Brisson S, Elvira G, Boulay K, Chatel-Chaix L, Mouland AJ, DesGroseillers L.Interaction of Staufen1 with the 5’ end of mRNA facilitates translation of these RNAs.NucleicAcidsRes2005;33:4797-812.
53. Ferrandon D, Elphick L, Nusslein-Volhard C, St Johnston D. Staufen protein associateswiththe3’UTRofbicoidmRNAtoformparticlesthatmoveinamicrotubule-dependentmanner.Cell1994;79:1221-32.
54. Wickham L, Duchaine T, Luo M, Nabi IR, DesGroseillers L. Mammalian staufen is adouble-stranded-RNA-andtubulin-bindingproteinwhichlocalizestotheroughendoplas-micreticulum.MolCellBiol1999;19:2220-30.
55. PepperAS,McCaneJE,KemperK,YeungDA,LeeRC,AmbrosV,MossEG.TheC. elegansheterochronicgenelin‑46affectsdevelopmentaltimingattwolarvalstagesandencodesarelativeofthescaffoldingproteingephyrin.Development2004;131:2049-59.
56. Olsen PH, Ambros V. The lin-4 regulatory RNA controls developmental timing inCaenorhabditis elegansbyblockingLIN-14proteinsynthesisaftertheinitiationoftransla-tion.DevBiol1999;216:671-80.
57. DingL,SpencerA,MoritaK,HanM.ThedevelopmentaltimingregulatorAIN-1interactswithmiRISCsandmaytargettheargonauteproteinALG-1tocytoplasmicPbodiesinC. elegans.MolCell2005;19:437-47.
58. SouthTL, Blake PR, Sowder IIIrd RC, Arthur LO, Henderson LE, Summers MF.ThenucleocapsidproteinisolatedfromHIV-1particlesbindszincandformsretroviral-typezincfingers.Biochemistry1990;29:7786-9.
59. KarpelRL,HendersonLE,OroszlanS. Interactionsof retroviral structuralproteinswithsingle-strandednucleicacids.JBiolChem1987;262:4961-7.
60. GreenLM,Berg JM.A retroviralCys-Xaa2-Cys-Xaa4-His-Xaa4-Cyspeptidebindsmetalions:Spectroscopicstudiesandaproposedthree-dimensionalstructure.ProcNatlAcadSciUSA1989;86:4047-51.
61. ReinA,HendersonLE,LevinJG.Nucleic-acid-chaperoneactivityofretroviralnucleocapsidproteins:Significanceforviralreplication.TrendsBiochemSci1998;23:297-301.
62. JiangW,HouY,InouyeM.CspA,themajorcold-shockproteinofEscherichia coli, isanRNAchaperone.JBiolChem1997;272:196-202.
63. NakaminamiK,KarlsonDT, ImaiR.Functional conservationof cold shockdomains inbacteriaandhigherplants.ProcNatlAcadSciUSA2006;103:10122-7.
64. Fusaro AF, Bocca SN, Ramos RL, Barroco RM, Magioli C, Jorge VC, Coutinho TC,Rangel-LimaCM,DeRyckeR,InzeD,EnglerG,Sachetto-MartinsG.AtGRP2,acold-in-ducednucleo-cytoplasmicRNA-bindingprotein,hasaroleinflowerandseeddevelopment.Planta2006.
65. KingsleyPD,Palis J.GRP2proteins containbothCCHCzinc fingers anda cold shockdomain.PlantCell1994;6:1522-3.
66. MikiT, Yoneda Y. Alternative splicing of Staufen2 creates the nuclear export signal forCRM1(Exportin1).JBiolChem2004;279:47473-9.
67. MacchiP,BrownawellAM,GrunewaldB,DesGroseillersL,MacaraIG,KieblerMA.Thebrain-specificdouble-strandedRNA-bindingproteinStaufen2:Nucleolaraccumulationandisoform-specificexportin-5-dependentexport.JBiolChem2004;279:31440-4.
68. BaderAG,FeltsKA,JiangN,ChangHW,VogtPK.Ybox-bindingprotein1inducesresis-tancetooncogenictransformationbythephosphatidylinositol3-kinasepathway.ProcNatlAcadSciUSA2003;100:12384-9.
69. PolesskayaA,CuvellierS,NaguibnevaI,DuquetA,MossEG,Harel-BellanA.Lin‑28bindsIGF‑2mRNAandparticipates in skeletalmyogenesisby increasing translationefficiency.GenesDev2007,(inpress).
Related Documents








![Targeting ornithine decarboxylase reverses the LIN28/Let-7 ... · the LIN28/Let-7 pathway [13, 14], which is important in a number of cancers, including NB, and was recently identified](https://static.cupdf.com/doc/110x72/5f7e699b6c944249467265c5/targeting-ornithine-decarboxylase-reverses-the-lin28let-7-the-lin28let-7-pathway.jpg)


